Submitted:
23 June 2023
Posted:
23 June 2023
You are already at the latest version
Abstract
Wheat has a large and diverse repertoire of NLRs involved in disease resistance, with over 1,500 NLRs detected in some studies. These NLR genes occur as singletons or clusters containing copies of NLRs from different phylogenetic clades. The number of NLRs and cluster size can differ drastically among ecotypes and cultivars. Primarily, duplication has led to the evolution and diversification of NLR genes. Among the various mechanisms, whole genome duplication (WGD) is the most intense and leading cause, contributing to the complex evolutionary history and abundant gene set of hexaploid wheat. Tandem duplication or recombination is another major mechanism of NLR gene expansion in wheat. The diversity and divergence of duplicate NLR genes are responsible for the broad-spectrum resistance of most plant species with limited R genes. Understanding the mechanisms underlying the rapid evolution and diversification of wheat NLR genes will help improve disease resistance in crops. The present review focuses on the diversity and divergence of duplicate NLR genes and their contribution to wheat disease resistance. Moreover, we provide an overview of disease resistance-associated gene duplication and the underlying strategies in wheat.
Keywords:
Wheat
; Nucleotide-binding leucine-rich repeat
; NLR genes
; Gene duplication
; Disease resistance
1. Introduction
Polyploid crops, such as wheat, triticale, oats, sweet potato, and peanuts, play a vital role in ensuring food security. Among these, the tetraploid durum wheat (pasta wheat, Triticum durum; 2n = 4x = 28), and the hexaploid bread wheat (Triticum aestivum; 2n = 6x = 42) are adaptable to various environmental conditions and, therefore, the most widely cultivated [1]. The production of wheat worldwide in 2021 was recorded to be as high as 781 million tons (FaoStat; http://www.fao.org/faostat/en/#home). However, in spite of the high production, wheat and other crops are often threatened by abiotic and biotic stressors that significantly reduce their yield and quality. Specifically, diseases caused by pathogenic bacteria, viruses, fungi, and oomycetes have resulted in significant crop losses, affecting food security worldwide [2]. As in other plants, wheat resists pathogens via their multilayered innate immune responses: one mediated by pattern recognition receptor (PRR) on the cell surface and the other mediated by nucleotide-binding leucine-rich repeat (NLR) intracellularly [3,4]. Typically, plants sense pathogens via the immune receptors that detect the pathogen-derived molecules and initiate diverse defense responses interconnected to form a signaling network [5]. These PRRs, which are in the form of receptor-like proteins (RLPs) or receptor-like kinases (RKs), detect conserved pathogen-associated molecular patterns (PAMPs) and activate pattern-triggered immunity (PTI) to instigate defensive responses against non-adapted pathogens. Besides, the residual PTI provides basal resistance to the adapted pathogens [4]. Therefore, PTI is categorized as a non-race-specific resistance. Researchers have found new PRR candidates in Triticeae but are less studied.
In order to overcome the impenetrable blockade of the first layer of the immune system, pathogenic organisms secrete a class of small molecules into plant cells called effectors. Effectors of these pathogens subsequently interfere with PTI by hindering PRR translation, inhibiting the activity of PRRs and their complexes, affecting the transmission of MAPK and its downstream signals, and impacting vesicle transport and callose deposition [6,7,8]. In this way, pathogenic bacteria can successfully infect. However, plant cells will not surrender easily. Faced with this situation, plants will urgently activate the next immune layer. The NLR receptor within the cell can indirectly or directly spot the effector, causing the plant to initiate a second immune response, which is called ETI (effector-triggered immunity), the effector recognized by the NLR protein is called avirulent (Avr) protein [9]. The ETI is more intense than PTI and is usually accompanied by programmed cell death (PCD). This hypersensitive response (HR) is crucial for plant resistance to biotrophic pathogens. Besides, the downstream immune response activated by ETI is similar to PTI but with greater intensity and longer duration [4]. Previous studies have found significant differences in recognition mechanisms and early signal transduction between PTI and ETI. Therefore, traditionally, people believe that PTI and ETI are independent systems [10,11]. However, recent researchers have found that multiple mutants of PRRs and their co-receptors in Arabidopsis have significantly lost plant ETI immune resistance triggered by multiple effector factors, suggesting that PRRs and their co-receptors are critical in activating NLRs [12]. PRRs and NLRs work together to enable plants to resist infection by most pathogens. However, pathogenic organisms will never surrender easily. They will continue to invent "new weapons" (effectors) to overcome the existing immune system and successfully infect plant cells. Similarly, plants will evolve new immune receptors to deal with the continuous and varied attacks of pathogenic bacteria.
2. NLR gene structure and categories
Typically, the structure of NLR consists of three parts, including a variable N-terminal domain, a conserved, central nucleotide-binding adapter domain (NB-ARC) domain, and a C-terminal leucine-rich repeat (LRR) domain. Here, the central adaptor domain is shared among the NLRs, apoptotic protease activating factor 1 (Apaf1), cerium R genes, and Caenorhabditis elegans cell death protein 4 (CED4). In contrast, the variable N-terminal domain is divided into two categories based on their protein sequence and similarity to known proteins: TIR (Toll interleukin-1 receiver) and CC (cooled coil). The NLR genes are classified into sub-groups depending on their domain architecture, including NBS, TN, TNL, CN, NL, and CNL (Figure 1) [13]. Some studies have indicated that NLRs with a deletion of the domain at the N-terminus cannot elicit plant immunity, while its expression alone is sufficient to induce HR responses [14]. Therefore, this domain of NLR is believed to stimulate and transmit immune signals downstream. In addition, both TIR and CC domains, which can oligomerize, are considered essential for NLR activation. In fact, when the effector is recognized, the entire NLR protein can oligomerize, such as RPP1 in Arabidopsis [15] and N protein in tobacco [16]. The N-terminal TIR and CC domains may be crucial for NLR oligomerization and activation.
NB-ARC, the relatively large domain of NLR, includes three subdomains: the NB, the ARC1, and the ARC2 subdomains. Among these, NB represents nucleotide binding, and ARC is named because this domain appears in Apaf-1, R, and CED-4 proteins. The NB-ARC domain is considered an ATPase domain due to its ability to bind with, exchange, or hydrolyze ADP or ATP nucleotides [17]. The NLR protein changes its state/form depending on the binding of the NB-ARC domain to different nucleotides. Generally, the NLR protein combined with ADP is in a closed or inactive state, and the combination with ATP promotes a conformational change of the NLR protein, turning it into an open or activated state. After NLR activation, it will transmit signals downstream to activate the plant's immune system. Effector recognition by NLR (directly or indirectly) will promote NB-ARC to release ADP and bind ATP. Besides, this binding feature acts as a "molecular switch" model of NLR in plant immunity [18]. The properties of the NB-ARC domain and its binding with different nucleotides form the structural basis of this "molecular switch" model. LRR is named for the presence of multiple tandem leucine-rich repeats in this domain. LRR domains regulate the activity of NLRs through intramolecular or intermolecular interactions. Research has demonstrated that LRR domains negatively influence the activity of NLRs. The absence of the LRR domains due to deletion in the NLR proteins RPS2, RPS5, and RPP1A activated the immune response in Arabidopsis. Similarly, truncating LRRs in the potato R-protein ‘Rx’ enhanced the HR response. Further studies found that the LRR of Rx may inhibit the activity of Rx by interacting with NB-ARC [19]. However, other studies have shown that LRR can also positively regulate the activity of NLR. Self-activating mutations in potato Rx lead to HR responses without pathogenic bacteria, whereas deletion of its LRRs suppressed HR responses [20].
In wheat, several NLR genes have been genetically defined, supporting disease-resistance breeding [13]. Several studies have recently highlighted the identified NLR and avirulence (Avr) genes, specifically the architecture of novel ones, revealing the genetic basis and mechanism underlying the evolution of disease resistance in wheat [13,21]. These discoveries are significant because they provide valuable insights into the mechanisms underlying plant-pathogen coevolution and the evolution of resistance. Identifying NLR and Avr genes offers great potential in developing effective strategies for breeding disease resistance in wheat. Using these genes in breeding programs will enhance disease resistance, reduce pesticide use, and increase crop productivity. Moreover, understanding the genetic basis and mechanisms underlying disease resistance can offer important information on the evolution of plant-pathogen interactions.
3. Gene duplication for crop improvement
The genome of bread wheat is one of the largest and most complex ones of all cultivated plant species’ this complexity originated as a result of two rounds of historical whole genome duplication events [22] and recent small-scale duplications [14]. Gene duplication provides “raw genetic material” for the evolution of genes and gene functions to improve crops; it is also a key factor driving the evolution, domestication, and diversification of species [15]. Additionally, gene duplication can also provide a buffer against mutations and genetic drift, resulting in the loss of important functional genes. Scientists have deciphered numerous large, complex, and highly repetitive genomes of various species of the Triticeae tribe [1,16,23,24,25,26,27]. Comparing the available genome sequences and functional genomic data of plants, we have gained immense knowledge of how genes are duplicated, how these duplicated genes gain novel roles, and their ultimate impact on genome evolution [28,29]. Among the various events or mechanisms, the duplication of the whole genome or an entire chromosome is the most extensive form of gene duplication [29]. Among these, whole genome duplication (WGD or polyploidization) is an extreme mechanism that benefited from the complex evolutionary history; hexaploid wheat has suddenly acquired a huge genome and abundant gene set due to the hybridization event it experienced [30,31]. Then, hexaploid wheat took over its tetraploid progenitor and spread worldwide (Figure 2A). About 55% of the bread wheat homologous genes exhibit 1:1:1 correspondence across the three homologous sub-genomes, and another 15% possess a minimum of one gene copy in at least one of the sub-genomes [8]. Tandem duplication or recombination is the event when two or more genes after duplication are positioned adjacent to each other on the same chromosome [29,32,33]. And it is likely a major mechanism of NLR gene expansion to form NLR gene clusters in wheat (Figure 2B). Additionally, researchers found a recent burst of gene duplications in all sequenced species of the Triticeae tribe. Detailed analysis of the features of the gene duplication and their flanking sequences suggested the role of transposable elements (TE) in causing this recent event [14]. In contrast to local tandem duplication, replicative transposition by TEs forms dispersed duplicates [34] (Figure 2C).
4. Diversity and divergence of NLR genes for disease resistance
Earlier research suggested great variation in the number of NLRs among the closely associated species [35], and the number is not related to the size of the genome or the level of ploidy [36]. However, more than 1,500 NLRs have been detected by analyzing the transcriptional and physical organization of the intracellular immune receptor repertory in bread wheat [37]. More than 2,000 NLRs have been identified using the fully annotated reference genome of bread wheat [25]. Several studies have identified 5,905 (98% identity) to 7,780 (100% identity) unique NLR signatures in the genomes of different wheat lines, emphasizing the complexity and size of immune receptor repertory guiding disease resistance [1]. This may also demonstrate the relationship between NLR number and genome size and between NLR number and ploidy level. Generally, the NLR loci exist as singletons (isolated genes) or clusters (tightly linked, related genes) [38]. Occasionally, the gene clusters include NBS-LRR gene copies from different phylogenetic clades [39]. NLRs usually exhibit consistent clustering [40]. In bread wheat, numerous NLRs are found grouped at the distal end of all chromosome arms and often co-localize with known disease-resistance loci [25]. In such an arrangement, the gene products act together to trigger immunity, providing coregulatory benefits.
In plants, the NLR proteins are abundant and old [41]. Gene duplications, followed by point mutation-induced DNA-sequence divergence and deletion and duplication of intragenic DNA repeats encoding blocks of leucine-rich elements, led to the origin of R-gene polymorphisms. Further, recombination uses this variation between related genes to alter the gene sequences [42]. Due to the diversity and abundance of pathogens in different environments, plants face dynamic selection pressure from habitat pathogens during their evolution, which results in the inability to maintain stable distribution of NLR genes between and even within species [10]. It has been found in research on different angiosperms that significant pathogen infection pressure has driven the expansion of NLR genes [43,44,45]. In the study of wild emmer wheat, it was found that changes in the NLR gene appeared to be rapid within the species [46]. Due to differences in the abundance of pathogenic bacteria in habitats, the population of wild wheat differentiated, with wild wheat growing in areas where powdery mildew was prevalent, evolving resistance to powdery mildew. As stated earlier, the clusters (medium and large) may vary greatly in their size among the ecotypes and cultivated varieties, indicating potential local adaptability [47,48]. For clusters containing highly homologous NLRs from a single family with few inversions, direct duplication of gene probably resulted in the original clustering; subsequently, increased rates of unequal crossing-over (UCO) during meiosis probably provided the material for rapid evolution and increased diversity in immune sensors, thereby expanding the cluster. Under different pressures, these clusters rapidly contract or expand, which explains the large variations in cluster patterns between ecotypes. However, the abundance of NLR genes is not solely beneficial; the more NLR genes plants maintain, the more health costs they incur [49]. Under no or less pathogen selection pressure, various plant species have exhibited contraction of NLR genes [50,51], leading to significant variations in the number and diversity of NLR genes between or within species. Gene recombination is crucial for NLR diversification. Current evidence shows that the unconventional recombination (illegitimate recombination) between NLR genes is the main way to change the number of repeats of the LRR domain, which can cause a rapid increase or decrease in the repeat number of the LRR, further increasing the potential of NLR to recognize different effectors [24]. In addition, if the NLR gene recombines with other genes (such as WRKY, NAC), it can cause the fusion of the domains of other genes to NLR, further enriching the diversity of NLR structure and function (Figure 2B). For example, a WRKY domain is attached to the Arabidopsis NLR gene RRS1 at the C-terminus. Further research found that this gene fusion phenomenon is very common in plant NLR genes. In addition, some gene recombination events can also cause the loss of the NLR domain, resulting in truncated NLR (truncated NLR). Truncated NLRs are also ubiquitous in various plants and also play very important roles in plant immunity. Interestingly, although the domains of truncated NLRs are incomplete, they can also perform similar functions to intact NLRs: they can directly or indirectly recognize effectors and induce plant immunity [14]. Taken together, gene duplication of NLRs provided diversity to cope with the infection of thousands of evolving pathogenic bacteria.
5. Duplicated NLR genes are important for wheat disease resistance
The duplication of NLR genes has several evolutionary implications. First, duplication can result in new functions of the existing genes by accumulating mutations. This change can lead to the evolution of new features and specificities of resistance, allowing plants to respond to a wider range of pathogen types [52]. Second, duplication can result in redundancy, where multiple copies of the same gene perform the same function. This redundancy can provide a buffer against loss-of-function mutations, ensuring the maintenance of a functional immune response [53]. Third, duplication can lead to sub-functionalization, where the duplicated genes acquire distinct functions through the partitioning of the original gene's functions [54]. This process can result in the evolution of more complex immune responses, allowing plants to respond to a wider range of pathogen types.
Many studies have proven that the duplicated NLR genes contributed to wheat disease resistance. Researchers have isolated three major genes associated with yellow rust resistance in bread wheat (Yr5, Yr7, and YrSP), each with distinct recognition specificity. These resistance genes belong to an R gene cluster on the wheat chromosome 2B encoding NLR proteins [55]. Previously, the Lr42 leaf rust R gene was transferred into bread wheat from the wild relative Ae. tauschii, where the lr42 allele, four homologs of, and four partial fragments of the gene were grouped as a cluster within an 871 kb region. We speculate that Lr42 could have originated from an early locus, which expanded or maintained the copy number in other wheat species and barley. Typically, Lr42 has an NLR structure and the “MAEAVVGQLVVTLGEALAKEA’ homologous domain, similar to the MADA “MA(D/E)AxVSFxVxKLxxLLxxEx” motif of the known NLRs associated with rust resistance in wheat. These features suggest that Lr42 is a singleton or a helper NLR but not a sensor [56]. The wheat progenitor Ae. tauschii has the NLR YrAS2388 associated with Pst resistance on the 4DS chromosome. YrAS2388R, the Pst-resistant allele, has duplicate 3′-untranslated regions [57]. In synthetic hexaploid wheat, a mutation in the YrAS2388R allele disrupted Pst resistance, while the YrAS2388R-overexpressing transgenics were resistant to 11 races in common wheat and one in barley [57]. Additionally, the Lr21 leaf rust resistance gene was introgressed from Ae. tauschii using a synthetic wheat species. Detailed analysis of the nucleotide sequence showed that the functional Lr21 allele is a chimera of H1 and H2, two nonfunctional haplotypes of lr21 [38,58]. On the other hand, the bread wheat has only inactive lr21 alleles; nevertheless, an active R gene could be reconstructed in vitro by intramolecular recombination of the two haplotypes.
Head-to-head or bidirectional gene pairs are common in eukaryotes and often include genes that are transcribed at similar rates; this specific property indicates the presence of common regulatory areas in the gene pairs [59]. Although there are few reports on wheat, research on other species can provide insights. In Arabidopsis thaliana, the head-to-head cluster SOC3-CHS1-TN2 NLR has been observed to have co-expression and gene product interaction, and microarray data for other bidirectional gene pairs of NLR in A. thaliana supports the proposal of shared regulatory regions [60]. Further experimental evidence is needed to establish this as a common trend for bidirectionally arranged NLRs; however, this justifies the genomic pattern. The bidirectional gene pairs encoding NLRs with integrated decoy domains may exhibit distinct behavior compared to clusters derived from closely associated NLRs. Furthermore, this pattern accounts for the broad-spectrum resistance observed in numerous plant species with limited NLR genes [40]. The origin of these paired NLRs, which are often highly differentiated or not from the same phylogenetic cluster, where they initially came together through random evolutionary accidents, letting them resist certain pathogen pressures. The selection for joint regulation could also justify the persistence of these evolutionarily differentiated pairs in plant genomes [61].
6. Mechanism of NLR gene duplication for disease resistance
Meta-analysis of 314 cloned resistance (R) genes revealed that most genes encode intracellular or cell surface receptors. These R genes encode proteins to induce or increase disease resistance (R) via nine distinct mechanisms. The mechanisms involve either direct or indirect recognition of pathogenic molecules on the cell surface with receptor-like proteins, leucine-rich repeat receptors, nucleotide-binding, or via integrated domains; they also perceive transcription activator-like effectors (TALEs) by activating the executor genes or lose susceptibility through passive or active mechanisms or host reprogramming [62]. Researchers have elucidated the molecular mechanisms underlying the functions of a few R genes. However, elucidating the mechanisms in detail is necessary for engineering and adopting novel R genes [62]. The recognition of pathogen-derived Avr proteins by NLR proteins is a critical component of gene-for-gene or active defense, mostly occurring through the leucine-rich repeat (LRR) domain of the NLR proteins. In this process, the LRR domain specifically recognizes pathogen effectors, allowing the plant to initiate an effective defense response against the attacking pathogens. However, an interruption in disease resistance may occur due to mutations in Avr proteins, which consequently allows the pathogens to escape R protein-mediated molecular recognition [63]. This evasion mechanism enables pathogens to overcome the defense response of the plant, resulting in infection and disease incidenceand further spread. Understanding the basis of this phenomenon is essential for designing durable resistance strategies in crop plants.
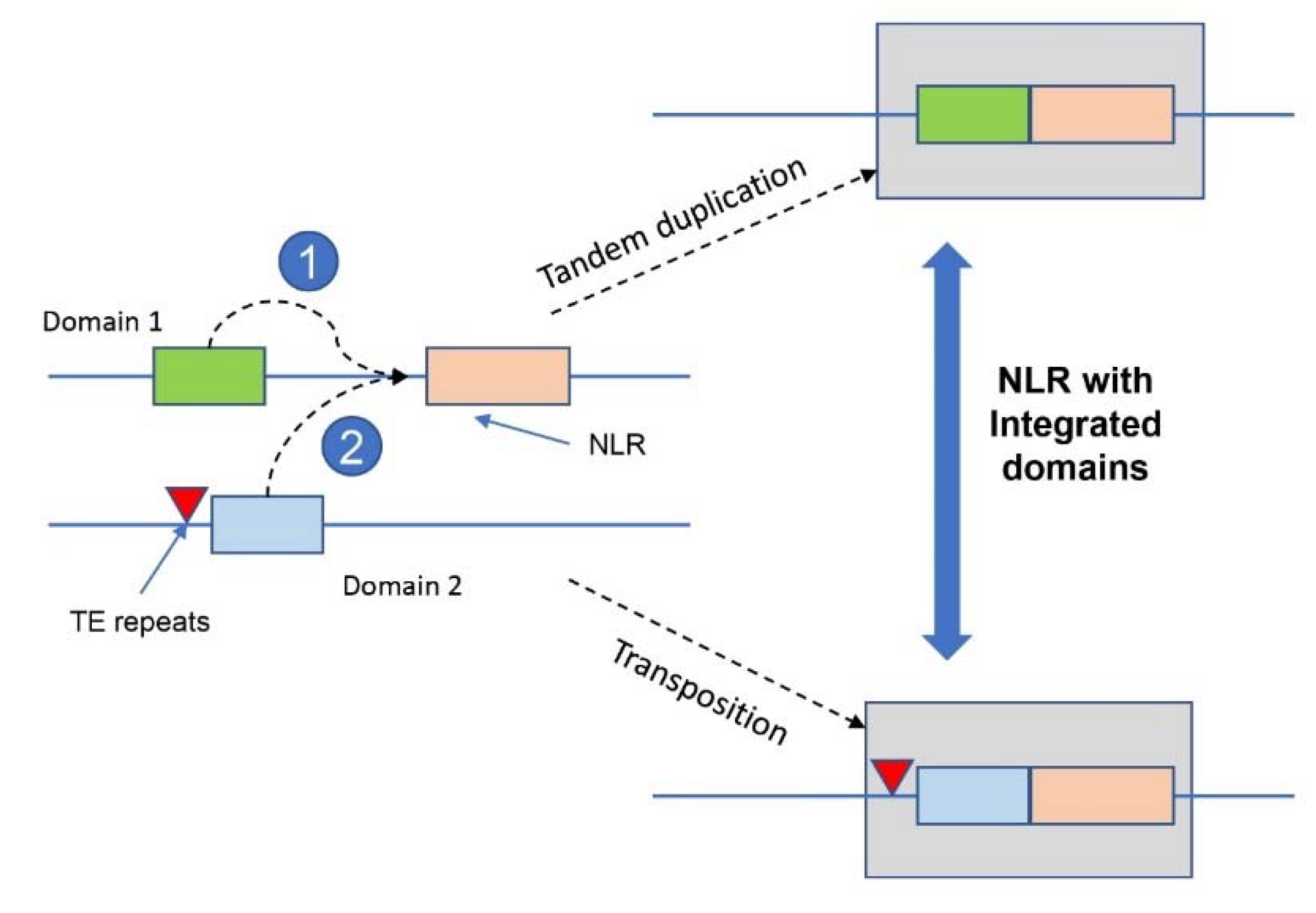
Previously reported NLR resistance proteins to have various additional domains, called integrated domains (IDs), that may confer novel functions or modulate NLR activity [64]. IDs can be derived from other immune-related proteins, such as kinases or transcription factors, and regulate the activation of receptors and signaling of the downstream components. Understanding the origin, evolution, and function of NLRs and IDs is crucial for elucidating plant immunity's molecular mechanisms and developing novel crop improvement strategies [21,65,66]. For example, the Arabidopsis resistance protein RRS1 has a WRKY domain, and the rice blast resistance protein RGA5 has a RATX1 domain, both at the C-terminus; the wheat Yr7, Yr5, and YrSP stripe rust resistance proteins have BED domains at the N-terminus. Besides, the YrU1 protein contains the ANK domain at the N-terminus and WRKY domain at the C-terminus [55,67,68]. Genes with additional domains seem occasionally. We propose a hypothesis that such genes are likely to originate from duplication events in the genome, such as tandem duplication and transposon-mediated duplication; this will improve our understanding of the evolution of the NLR genes with integrated domains (Figure 3). NLR proteins can function together to trigger plant immunity and are classified into two: NLR pairs and sensor/helper NLRs [61]. NLR pairs are encoded next to each other in the same orientation and share a promoter. Paired NLRs tend to form complexes needed for immunity. Typically, one NLR has a decoy domain and acts as the sensor, interacting with pathogens. The other is a canonical NLR that transduces signals to activate immunity. NLRs with decoy domains are common in flowering plants, likely formed through ectopic recombination and transposition [11]. The other way NLRs interact is as a sensor or a helper. Among these NLRs, the helper presumably acts downstream of the sensor in immunity. Unlike the typical adjacent NLR pairs, the helper-sensor NRLs do not physically interact. On the contrary, they are usually not encoded at the same locus. However, the mechanism via which the sensors transmit signals to the helpers remains elusive [61].
Studies have almost established that the products of larger NLR gene clusters act together in inducing immunity [40]. A good example is Rps11 in soybean, providing broad-spectrum resistance to the oomycete pathogen Phytophthora sojae. In the genome, the giant NLR gene is positioned in a region carrying a large NLR gene cluster resulting from numerous unequal recombinations [69]. In wheat, several studies also reported that NLR clusters contributed to disease resistance. In the hexaploid wheat germplasm, Pm3a–j, a large multi-allelic series of ten resistance alleles, each with distinct race specificity, accounted for the Pm3 powdery mildew resistance [21]. Scientists have identified Pm3 alleles in cultivars worldwide. After cloning Pm3b, the first allele [70], the remaining alleles were isolated by PCR and identified as a single gene’s true alleles in a cluster of Pm3-like genes [6,71]. Here, the alleles Pm3j, Pm3h, and Pm3i were identical to Pm3b, Pm3d, and Pm3c, respectively. These results enrich our understanding of the resistance mechanism of NLR gene clusters induced by gene replication in wheat. Moreover, NLR clusters could lead to improved pathogen recognition via homo- and heterodimerization/oligomerization of the NLRs encoded by the same cluster [40].
Most NLRs occur as singletons. A study showed that the Sr35 gene encodes an NLR receptor that, after recognizing AVRSr35, an Avr protein, conferred near-immunity to Ug99, the causal agent of wheat blast and stem rust [74,75]. This report supports that interaction between an effector and NLR confers immunity, based on the Avr-R gene-for-gene interaction model, and helps rapidly control pandemics in bread wheat [76]. One study using a cryo-electron microscopic structure demonstrated the complex interaction between Sr35 and the AvrSr35 effector of the stem rust pathogen of wheat [77]. The effector directly binds to the Sr35’s LRRs and forms a pentameric complex called the Sr35 resistosome. These novel insights into the complex structural interaction helped us generate novel variants of NLRs of barley and wheat that identify AvrSr35 [77].
7. Future perspectives
Over the years, our knowledge about the molecular mechanisms governing the function of NLR genes has improved. This progress has been largely driven by the advancement of molecular biology techniques and the availability of high-quality genome sequences for both plants and pathogens. The elucidation of these molecular mechanisms will be critical for the rational design and deployment of novel R genes that can impart long-lasting and broad-spectrum resistance against diverse pathogens [78]. One of the key challenges in engineering and deploying novel R genes is to identify the most effective R gene combinations that can provide durable resistance against various pathogens. For this, we need to have a complete idea of the molecular basis of R gene function and the co-evolutionary dynamics between pathogens and host plants [62]. Furthermore, the development of R gene-based resistance strategies should be complemented by implementing integrated pest management approaches that can minimize the selection pressure on pathogens and reduce the likelihood of resistance breakdown. Understanding the molecular underpinnings of R gene function is critical for modifying and deploying novel R genes that can offer long-lasting and broad-spectrum resistance against various plant pathogens [79]. As our knowledge of these mechanisms continues to grow, it holds great promise for the development of innovative strategies to enhance crop protection and improve global food security.
In bread wheat, speciation and domestication occurred remarkably compared to other crops, as reported in [80]. Bread wheat evolved due to the effect of duplications in genes, particularly via WGD, during both speciation and domestication processes. Despite the diversity bottleneck that newly formed allopolyploids experienced, bread wheat differs significantly from its diploid progenitors, which formed much earlier than bread wheat [81]. Compared with the diploid progenitors, bread wheat exhibits broad morphological variation, occupies more diverse ecological habitats, and is spread over a larger area [82]. Recent studies have delivered new insights into the occurrence and consequences of genomic changes induced by gene duplication, particularly whole genome duplication. Despite these advances, there remain many opportunities for novel discoveries in gene duplication and wheat disease resistance research. Addressing the following challenges will be crucial to advancing this field: (1) identifying novel R genes; (2) the role and mechanism via which abiotic and/or biotic factors regulate and manipulate these R genes; (3) comprehending the various mechanisms of gene replication; (4) understanding how gene duplication affects NLR cluster size and NLR number; (5) understanding how gene duplication regulates the abundance and sequence variability of NLRs, and (6) exploring the relationship between gene duplication and potentially neo-functionalized genes.
Author Contributions
Conceptualization, Z.Y.; methodology, H.Y. and P.Y.; software, H.Y.; investigation, C.W., L.M., C.N. and D.X.; writing—original draft preparation, H.Y.; writing—review and editing, Z.Y. M.A.R.R.; data visualization, H.Y. All authors have read and agreed to the manuscript’s published version.
Funding
This research work was funded by the Project of the Central Government Guides the Development of Local Science and Technology (Guike ZY20198015), the Natural Science Foundation of Shandong Province (ZR2020MC096 and ZR2021ZD31), and the Agricultural Variety Improvement Project of Shandong Province (2022LZGC002).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Walkowiak, S.; Gao, L.; Monat, C.; Haberer, G.; Kassa, M.T.; Brinton, J.; Ramirez-Gonzalez, R.H.; Kolodziej, M.C.; Delorean, E.; Thambugala, D.; et al. Multiple wheat genomes reveal global variation in modern breeding. Nature 2020, 588, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Keller, B.; Wicker, T.; Krattinger, S.G. Advances in Wheat and Pathogen Genomics: Implications for Disease Control. Annu Rev Phytopathol 2018, 56, 67–87. [Google Scholar] [CrossRef]
- Lacaze, A.; Joly, D.L. Structural specificity in plant-filamentous pathogen interactions. Mol Plant Pathol 2020, 21, 1513–1525. [Google Scholar] [CrossRef]
- Ngou, B.P.M.; Ding, P.; Jones, J.D.G. Thirty years of resistance: Zig-zag through the plant immune system. Plant Cell 2022, 34, 1447–1478. [Google Scholar] [CrossRef]
- Lu, Y.; Tsuda, K. Intimate Association of PRR- and NLR-Mediated Signaling in Plant Immunity. Mol Plant Microbe Interact 2021, 34, 3–14. [Google Scholar] [CrossRef]
- Yahiaoui, N.; Brunner, S.; Keller, B. Rapid generation of new powdery mildew resistance genes after wheat domestication. Plant J 2006, 47, 85–98. [Google Scholar] [CrossRef] [PubMed]
- Toruno, T.Y.; Stergiopoulos, I.; Coaker, G. Plant-Pathogen Effectors: Cellular Probes Interfering with Plant Defenses in Spatial and Temporal Manners. Annu Rev Phytopathol 2016, 54, 419–441. [Google Scholar] [CrossRef]
- Yu, X.; Feng, B.; He, P.; Shan, L. From Chaos to Harmony: Responses and Signaling upon Microbial Pattern Recognition. Annu Rev Phytopathol 2017, 55, 109–137. [Google Scholar] [CrossRef]
- Plissonneau, C.; Benevenuto, J.; Mohd-Assaad, N.; Fouche, S.; Hartmann, F.E.; Croll, D. Using Population and Comparative Genomics to Understand the Genetic Basis of Effector-Driven Fungal Pathogen Evolution. Front Plant Sci 2017, 8, 119. [Google Scholar] [CrossRef] [PubMed]
- Casadevall, A. Fungal virulence, vertebrate endothermy, and dinosaur extinction: is there a connection? Fungal Genet Biol 2005, 42, 98–106. [Google Scholar] [CrossRef]
- Sun, Y.; Zhu, Y.X.; Balint-Kurti, P.J.; Wang, G.F. Fine-Tuning Immunity: Players and Regulators for Plant NLRs. Trends Plant Sci 2020, 25, 695–713. [Google Scholar] [CrossRef]
- Yuan, M.; Jiang, Z.; Bi, G.; Nomura, K.; Liu, M.; Wang, Y.; Cai, B.; Zhou, J.M.; He, S.Y.; Xin, X.F. Pattern-recognition receptors are required for NLR-mediated plant immunity. Nature 2021, 592, 105–109. [Google Scholar] [CrossRef]
- Sanchez-Martin, J.; Keller, B. NLR immune receptors and diverse types of non-NLR proteins control race-specific resistance in Triticeae. Curr Opin Plant Biol 2021, 62, 102053. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yan, X.; Hu, Y.; Qin, L.; Wang, D.; Jia, J.; Jiao, Y. A recent burst of gene duplications in Triticeae. Plant Commun 2022, 3, 100268. [Google Scholar] [CrossRef]
- Juery, C.; Concia, L.; De Oliveira, R.; Papon, N.; Ramirez-Gonzalez, R.; Benhamed, M.; Uauy, C.; Choulet, F.; Paux, E. New insights into homoeologous copy number variations in the hexaploid wheat genome. Plant Genome 2021, 14, e20069. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.C.; Gu, Y.Q.; Puiu, D.; Wang, H.; Twardziok, S.O.; Deal, K.R.; Huo, N.; Zhu, T.; Wang, L.; Wang, Y.; et al. Genome sequence of the progenitor of the wheat D genome Aegilops tauschii. Nature 2017, 551, 498–502. [Google Scholar] [CrossRef] [PubMed]
- Danot, O.; Marquenet, E.; Vidal-Ingigliardi, D.; Richet, E. Wheel of Life, Wheel of Death: A Mechanistic Insight into Signaling by STAND Proteins. Structure 2009, 17, 172–182. [Google Scholar] [CrossRef]
- Bonardi, V.; Cherkis, K.; Nishimura, M.T.; Dangl, J.L. A new eye on NLR proteins: focused on clarity or diffused by complexity? Curr Opin Immunol 2012, 24, 41–50. [Google Scholar] [CrossRef]
- Lukasik, E.; Takken, F.L. STANDing strong, resistance proteins instigators of plant defence. Curr Opin Plant Biol 2009, 12, 427–436. [Google Scholar] [CrossRef]
- Moffett, P.; Farnham, G.; Peart, J.; Baulcombe, D.C. Interaction between domains of a plant NBS-LRR protein in disease resistance-related cell death. EMBO J 2002, 21, 4511–4519. [Google Scholar] [CrossRef]
- Ayliffe, M.A.; Lagudah, E.S. Molecular genetics of disease resistance in cereals. Ann Bot 2004, 94, 765–773. [Google Scholar] [CrossRef]
- Salamini, F.; Ozkan, H.; Brandolini, A.; Schafer-Pregl, R.; Martin, W. Genetics and geography of wild cereal domestication in the near east. Nat Rev Genet 2002, 3, 429–441. [Google Scholar] [CrossRef] [PubMed]
- Mascher, M.; Gundlach, H.; Himmelbach, A.; Beier, S.; Twardziok, S.O.; Wicker, T.; Radchuk, V.; Dockter, C.; Hedley, P.E.; Russell, J.; et al. A chromosome conformation capture ordered sequence of the barley genome. Nature 2017, 544, 427–433. [Google Scholar] [CrossRef]
- Zhao, G.; Zou, C.; Li, K.; Wang, K.; Li, T.; Gao, L.; Zhang, X.; Wang, H.; Yang, Z.; Liu, X.; et al. The Aegilops tauschii genome reveals multiple impacts of transposons. Nat Plants 2017, 3, 946–955. [Google Scholar] [CrossRef]
- International Wheat Genome Sequencing, C.; investigators, I.R.p.; Appels, R.; Eversole, K.; Feuillet, C.; Keller, B.; Rogers, J.; Stein, N.; investigators, I.w.-g.a.p.; Pozniak, C.J.; et al. Shifting the limits in wheat research and breeding using a fully annotated reference genome. Science 2018, 361. [Google Scholar] [CrossRef]
- Ling, H.Q.; Ma, B.; Shi, X.; Liu, H.; Dong, L.; Sun, H.; Cao, Y.; Gao, Q.; Zheng, S.; Li, Y.; et al. Genome sequence of the progenitor of wheat A subgenome Triticum urartu. Nature 2018, 557, 424–428. [Google Scholar] [CrossRef]
- Zhou, Y.; Bai, S.; Li, H.; Sun, G.; Zhang, D.; Ma, F.; Zhao, X.; Nie, F.; Li, J.; Chen, L.; et al. Introgressing the Aegilops tauschii genome into wheat as a basis for cereal improvement. Nat Plants 2021, 7, 774–786. [Google Scholar] [CrossRef] [PubMed]
- Bomblies, K. When everything changes at once: finding a new normal after genome duplication. Proc Biol Sci 2020, 287, 20202154. [Google Scholar] [CrossRef] [PubMed]
- Panchy, N.; Lehti-Shiu, M.; Shiu, S.H. Evolution of Gene Duplication in Plants. Plant Physiol 2016, 171, 2294–2316. [Google Scholar] [CrossRef]
- Matsuoka, Y. Evolution of polyploid triticum wheats under cultivation: the role of domestication, natural hybridization and allopolyploid speciation in their diversification. Plant Cell Physiol 2011, 52, 750–764. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.C.; Yang, Z.L.; You, F.M.; Kawahara, T.; Waines, J.G.; Dvorak, J. The structure of wild and domesticated emmer wheat populations, gene flow between them, and the site of emmer domestication. Theor Appl Genet 2007, 114, 947–959. [Google Scholar] [CrossRef]
- Hao, Y.; Xu, S.; lyu, Z.; Wang, H.; Kong, L.; Sun, S. Comparative Analysis of the Glutathione S-Transferase Gene Family of Four Triticeae Species and Transcriptome Analysis of GST Genes in Common Wheat Responding to Salt Stress. International Journal of Genomics 2021, 2021, 1–11. [Google Scholar] [CrossRef]
- Hao, Y.; Hao, M.; Cui, Y.; Kong, L.; Wang, H. Genome-wide survey of the dehydrin genes in bread wheat (Triticum aestivum L.) and its relatives: identification, evolution and expression profiling under various abiotic stresses. {BMC} Genomics 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Kapitonov, V.V.; Jurka, J. Helitrons on a roll: eukaryotic rolling-circle transposons. Trends Genet 2007, 23, 521–529. [Google Scholar] [CrossRef] [PubMed]
- Seo, E.; Kim, S.; Yeom, S.I.; Choi, D. Genome-Wide Comparative Analyses Reveal the Dynamic Evolution of Nucleotide-Binding Leucine-Rich Repeat Gene Family among Solanaceae Plants. Front Plant Sci 2016, 7, 1205. [Google Scholar] [CrossRef]
- Borrelli, G.M.; Mazzucotelli, E.; Marone, D.; Crosatti, C.; Michelotti, V.; Vale, G.; Mastrangelo, A.M. Regulation and Evolution of NLR Genes: A Close Interconnection for Plant Immunity. Int J Mol Sci 2018, 19. [Google Scholar] [CrossRef]
- Sappl, P.G.; Carroll, A.J.; Clifton, R.; Lister, R.; Whelan, J.; Harvey Millar, A.; Singh, K.B. The Arabidopsis glutathione transferase gene family displays complex stress regulation and co-silencing multiple genes results in altered metabolic sensitivity to oxidative stress. Plant J 2009, 58, 53–68. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Brooks, S.; Li, W.; Fellers, J.; Nelson, J.C.; Gill, B. Evolution of new disease specificity at a simple resistance locus in a crop-weed complex: reconstitution of the Lr21 gene in wheat. Genetics 2009, 182, 595–602. [Google Scholar] [CrossRef] [PubMed]
- Wei, F.; Wing, R.A.; Wise, R.P. Genome dynamics and evolution of the Mla (powdery mildew) resistance locus in barley. Plant Cell 2002, 14, 1903–1917. [Google Scholar] [CrossRef]
- van Wersch, S.; Li, X. Stronger When Together: Clustering of Plant NLR Disease resistance Genes. Trends Plant Sci 2019, 24, 688–699. [Google Scholar] [CrossRef]
- McHale, L.; Tan, X.; Koehl, P.; Michelmore, R.W. Plant NBS-LRR proteins: adaptable guards. Genome Biol 2006, 7, 212. [Google Scholar] [CrossRef]
- Ellis, J.; Dodds, P.; Pryor, T. Structure, function and evolution of plant disease resistance genes. Current Opinion in Plant Biology 2000, 3, 278–284. [Google Scholar] [CrossRef]
- Shao, Z.Q.; Xue, J.Y.; Wu, P.; Zhang, Y.M.; Wu, Y.; Hang, Y.Y.; Wang, B.; Chen, J.Q. Large-Scale Analyses of Angiosperm Nucleotide-Binding Site-Leucine-Rich Repeat Genes Reveal Three Anciently Diverged Classes with Distinct Evolutionary Patterns. Plant Physiol 2016, 170, 2095–2109. [Google Scholar] [CrossRef] [PubMed]
- Shao, Z.Q.; Zhang, Y.M.; Hang, Y.Y.; Xue, J.Y.; Zhou, G.C.; Wu, P.; Wu, X.Y.; Wu, X.Z.; Wang, Q.; Wang, B.; et al. Long-term evolution of nucleotide-binding site-leucine-rich repeat genes: understanding gained from and beyond the legume family. Plant Physiol 2014, 166, 217–234. [Google Scholar] [CrossRef]
- Glover, N.M.; Daron, J.; Pingault, L.; Vandepoele, K.; Paux, E.; Feuillet, C.; Choulet, F. Small-scale gene duplications played a major role in the recent evolution of wheat chromosome 3B. Genome Biology 2015, 16. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yin, H.; Jiao, C.; Fang, X.; Wang, G.; Li, G.; Ni, F.; Li, P.; Su, P.; Ge, W.; et al. Sympatric speciation of wild emmer wheat driven by ecology and chromosomal rearrangements. Proceedings of the National Academy of Sciences 2020, 117, 5955–5963. [Google Scholar] [CrossRef] [PubMed]
- Bahram, M.; Netherway, T.; Frioux, C.; Ferretti, P.; Coelho, L.P.; Geisen, S.; Bork, P.; Hildebrand, F. Metagenomic assessment of the global diversity and distribution of bacteria and fungi. Environ Microbiol 2021, 23, 316–326. [Google Scholar] [CrossRef]
- Liu, Y.; Zeng, Z.; Zhang, Y.M.; Li, Q.; Jiang, X.M.; Jiang, Z.; Tang, J.H.; Chen, D.; Wang, Q.; Chen, J.Q.; et al. An angiosperm NLR Atlas reveals that NLR gene reduction is associated with ecological specialization and signal transduction component deletion. Mol Plant 2021, 14, 2015–2031. [Google Scholar] [CrossRef]
- Karasov, T.L.; Kniskern, J.M.; Gao, L.; DeYoung, B.J.; Ding, J.; Dubiella, U.; Lastra, R.O.; Nallu, S.; Roux, F.; Innes, R.W.; et al. The long-term maintenance of a resistance polymorphism through diffuse interactions. Nature 2014, 512, 436–440. [Google Scholar] [CrossRef]
- Xue, J.Y.; Zhao, T.; Liu, Y.; Liu, Y.; Zhang, Y.X.; Zhang, G.Q.; Chen, H.; Zhou, G.C.; Zhang, S.Z.; Shao, Z.Q. Genome- Wide Analysis of the Nucleotide Binding Site Leucine-Rich Repeat Genes of Four Orchids Revealed Extremely Low Numbers of Disease Resistance Genes. Front Genet 2019, 10, 1286. [Google Scholar] [CrossRef]
- Lin, X.; Zhang, Y.; Kuang, H.; Chen, J. Frequent loss of lineages and deficient duplications accounted for low copy number of disease resistance genes in Cucurbitaceae. BMC Genomics 2013, 14, 335. [Google Scholar] [CrossRef]
- Hanada, K.; Zou, C.; Lehti-Shiu, M.D.; Shinozaki, K.; Shiu, S.H. Importance of lineage-specific expansion of plant tandem duplicates in the adaptive response to environmental stimuli. Plant Physiol 2008, 148, 993–1003. [Google Scholar] [CrossRef] [PubMed]
- Hanada, K.; Kuromori, T.; Myouga, F.; Toyoda, T.; Li, W.H.; Shinozaki, K. Evolutionary persistence of functional compensation by duplicate genes in Arabidopsis. Genome Biol Evol 2009, 1, 409–414. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Zhang, J. Rapid subfunctionalization accompanied by prolonged and substantial neofunctionalization in duplicate gene evolution. Genetics 2005, 169, 1157–1164. [Google Scholar] [CrossRef]
- Marchal, C.; Zhang, J.; Zhang, P.; Fenwick, P.; Steuernagel, B.; Adamski, N.M.; Boyd, L.; McIntosh, R.; Wulff, B.B.H.; Berry, S.; et al. BED-domain-containing immune receptors confer diverse resistance spectra to yellow rust. Nat Plants 2018, 4, 662–668. [Google Scholar] [CrossRef]
- Lin, G.; Chen, H.; Tian, B.; Sehgal, S.K.; Singh, L.; Xie, J.; Rawat, N.; Juliana, P.; Singh, N.; Shrestha, S.; et al. Cloning of the broadly effective wheat leaf rust resistance gene Lr42 transferred from Aegilops tauschii. Nat Commun 2022, 13, 3044. [Google Scholar] [CrossRef]
- Zhang, C.; Huang, L.; Zhang, H.; Hao, Q.; Lyu, B.; Wang, M.; Epstein, L.; Liu, M.; Kou, C.; Qi, J.; et al. An ancestral NB-LRR with duplicated 3'UTRs confers stripe rust resistance in wheat and barley. Nat Commun 2019, 10, 4023. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Shen, X.; Hao, Y.; Cai, J.; Ohm, H.W.; Kong, L. A genetic map of Lophopyrum ponticum chromosome 7E, harboring resistance genes to Fusarium head blight and leaf rust. Theor Appl Genet 2011, 122, 263–270. [Google Scholar] [CrossRef]
- Chen, Y.Q.; Yu, H.; Li, Y.X.; Li, Y.Y. Sorting out inherent features of head-to-head gene pairs by evolutionary conservation. BMC Bioinformatics 2010, 11, S16. [Google Scholar] [CrossRef]
- Liang, W.; van Wersch, S.; Tong, M.; Li, X. TIR-NB-LRR immune receptor SOC3 pairs with truncated TIR-NB protein CHS1 or TN2 to monitor the homeostasis of E3 ligase SAUL1. New Phytol 2019, 221, 2054–2066. [Google Scholar] [CrossRef]
- Adachi, H.; Derevnina, L.; Kamoun, S. NLR singletons, pairs, and networks: evolution, assembly, and regulation of the intracellular immunoreceptor circuitry of plants. Curr Opin Plant Biol 2019, 50, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Kourelis, J.; van der Hoorn, R.A.L. Defended to the Nines: 25 Years of Resistance Gene Cloning Identifies Nine Mechanisms for R Protein Function. Plant Cell 2018, 30, 285–299. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, N.; Inoue, H.; Kato, T.; Funao, T.; Shirota, M.; Shimizu, T.; Kanamori, H.; Yamane, H.; Hayano-Saito, Y.; Matsumoto, T.; et al. Durable panicle blast-resistance gene Pb1 encodes an atypical CC-NBS-LRR protein and was generated by acquiring a promoter through local genome duplication. Plant J 2010, 64, 498–510. [Google Scholar] [CrossRef] [PubMed]
- Marchal, C.; Michalopoulou, V.A.; Zou, Z.; Cevik, V.; Sarris, P.F. Show me your ID: NLR immune receptors with integrated domains in plants. Essays Biochem 2022, 66, 527–539. [Google Scholar] [CrossRef]
- Sarris, P.F.; Duxbury, Z.; Huh, S.U.; Ma, Y.; Segonzac, C.; Sklenar, J.; Derbyshire, P.; Cevik, V.; Rallapalli, G.; Saucet, S.B.; et al. A Plant Immune Receptor Detects Pathogen Effectors that Target WRKY Transcription Factors. Cell 2015, 161, 1089–1100. [Google Scholar] [CrossRef]
- Le Roux, C.; Huet, G.; Jauneau, A.; Camborde, L.; Tremousaygue, D.; Kraut, A.; Zhou, B.; Levaillant, M.; Adachi, H.; Yoshioka, H.; et al. A receptor pair with an integrated decoy converts pathogen disabling of transcription factors to immunity. Cell 2015, 161, 1074–1088. [Google Scholar] [CrossRef]
- Chen, X.; Zhu, M.; Jiang, L.; Zhao, W.; Li, J.; Wu, J.; Li, C.; Bai, B.; Lu, G.; Chen, H.; et al. A multilayered regulatory mechanism for the autoinhibition and activation of a plant CC-NB-LRR resistance protein with an extra N-terminal domain. New Phytol 2016, 212, 161–175. [Google Scholar] [CrossRef]
- Wang, H.; Zou, S.; Li, Y.; Lin, F.; Tang, D. An ankyrin-repeat and WRKY-domain-containing immune receptor confers stripe rust resistance in wheat. Nat Commun 2020, 11, 1353. [Google Scholar] [CrossRef]
- Wang, W.; Chen, L.; Fengler, K.; Bolar, J.; Llaca, V.; Wang, X.; Clark, C.B.; Fleury, T.J.; Myrvold, J.; Oneal, D.; et al. A giant NLR gene confers broad-spectrum resistance to Phytophthora sojae in soybean. Nat Commun 2021, 12, 6263. [Google Scholar] [CrossRef]
- Yahiaoui, N.; Srichumpa, P.; Dudler, R.; Keller, B. Genome analysis at different ploidy levels allows cloning of the powdery mildew resistance gene Pm3b from hexaploid wheat. Plant J 2004, 37, 528–538. [Google Scholar] [CrossRef]
- Srichumpa, P.; Brunner, S.; Keller, B.; Yahiaoui, N. Allelic series of four powdery mildew resistance genes at the Pm3 locus in hexaploid bread wheat. Plant Physiol 2005, 139, 885–895. [Google Scholar] [CrossRef] [PubMed]
- Cruz, C.D.; Valent, B. Wheat blast disease: danger on the move. Tropical Plant Pathology 2017, 42, 210–222. [Google Scholar] [CrossRef]
- Singh, R.P.; Hodson, D.P.; Huerta-Espino, J.; Jin, Y.; Bhavani, S.; Njau, P.; Herrera-Foessel, S.; Singh, P.K.; Singh, S.; Govindan, V. The emergence of Ug99 races of the stem rust fungus is a threat to world wheat production. Annu Rev Phytopathol 2011, 49, 465–481. [Google Scholar] [CrossRef] [PubMed]
- Saintenac, C.; Zhang, W.; Salcedo, A.; Rouse, M.N.; Trick, H.N.; Akhunov, E.; Dubcovsky, J. Identification of Wheat Gene Sr35 That Confers Resistance to Ug99 Stem Rust Race Group. Science 2013, 341, 783–786. [Google Scholar] [CrossRef]
- Salcedo, A.; Rutter, W.; Wang, S.; Akhunova, A.; Bolus, S.; Chao, S.; Anderson, N.; Soto, M.F.D.; Rouse, M.; Szabo, L.; et al. Variation in the AvrSr35 gene determines Sr35 resistance against wheat stem rust race Ug99. Science 2017, 358, 1604–1606. [Google Scholar] [CrossRef] [PubMed]
- Bourras, S.; McNally, K.E.; Muller, M.C.; Wicker, T.; Keller, B. Avirulence Genes in Cereal Powdery Mildews: The Gene-for-Gene Hypothesis 2.0. Front Plant Sci 2016, 7, 241. [Google Scholar] [CrossRef]
- Forderer, A.; Li, E.; Lawson, A.W.; Deng, Y.N.; Sun, Y.; Logemann, E.; Zhang, X.; Wen, J.; Han, Z.; Chang, J.; et al. A wheat resistosome defines common principles of immune receptor channels. Nature 2022, 610, 532–539. [Google Scholar] [CrossRef]
- Bailey-Serres, J.; Parker, J.E.; Ainsworth, E.A.; Oldroyd, G.E.D.; Schroeder, J.I. Genetic strategies for improving crop yields. Nature 2019, 575, 109–118. [Google Scholar] [CrossRef]
- Deng, Y.; Ning, Y.; Yang, D.L.; Zhai, K.; Wang, G.L.; He, Z. Molecular Basis of Disease Resistance and Perspectives on Breeding Strategies for Resistance Improvement in Crops. Mol Plant 2020, 13, 1402–1419. [Google Scholar] [CrossRef]
- Tanno, K.; Willcox, G. How fast was wild wheat domesticated? Science 2006, 311, 1886. [Google Scholar] [CrossRef]
- Cheng, H.; Liu, J.; Wen, J.; Nie, X.; Xu, L.; Chen, N.; Li, Z.; Wang, Q.; Zheng, Z.; Li, M.; et al. Frequent intra- and inter-species introgression shapes the landscape of genetic variation in bread wheat. Genome Biol 2019, 20, 136. [Google Scholar] [CrossRef] [PubMed]
- Feldman, M.; Levy, A.A. Genome evolution due to allopolyploidization in wheat. Genetics 2012, 192, 763–774. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Sub-groups of NLR genes based on domain structures.
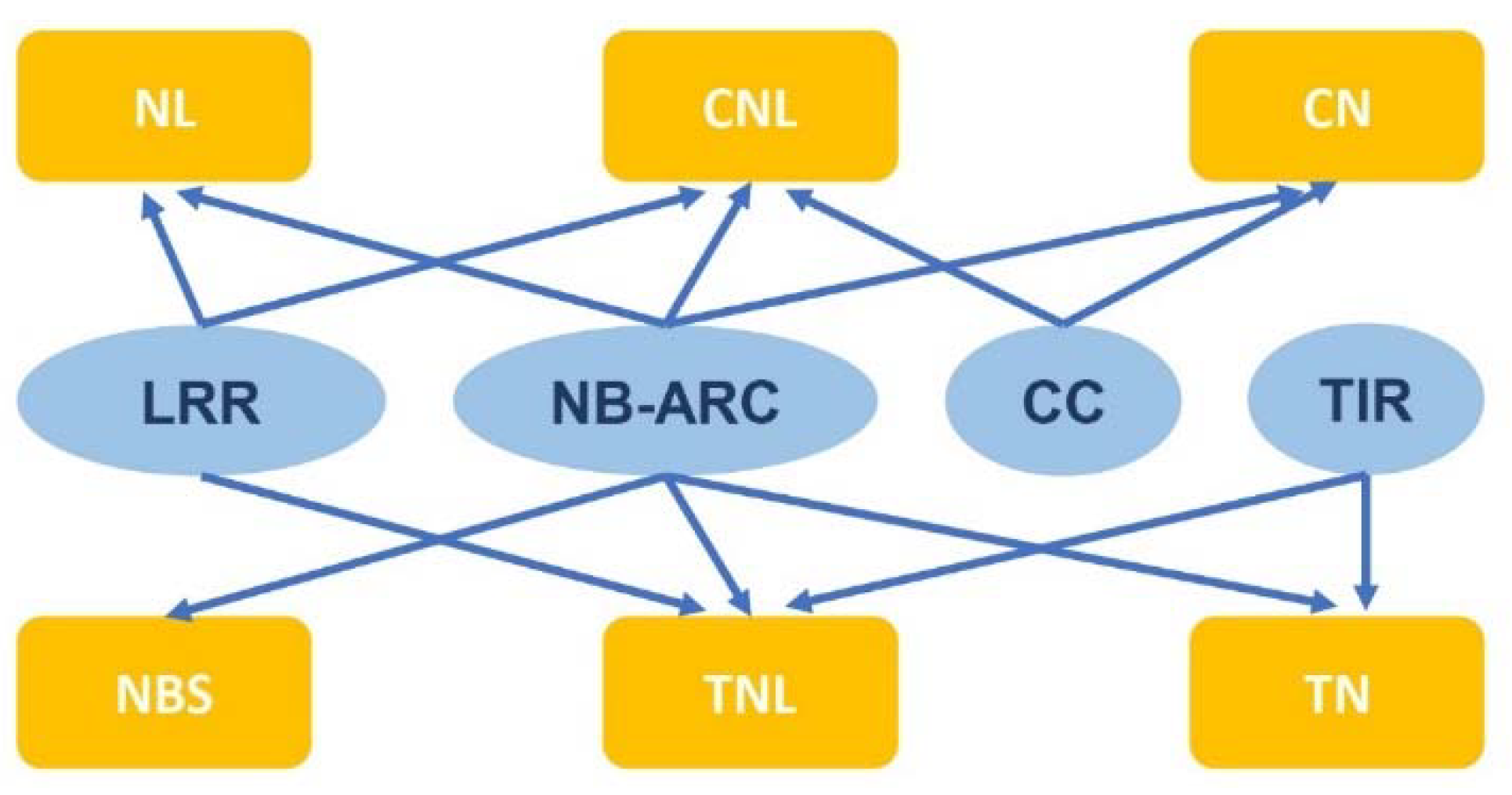
Figure 2.
NLR duplication in wheat. (A) Whole-genome duplication (WGD) through an increase in ploidy. (B) Tandem duplication through unequal recombination between similar alleles to form a gene cluster. (C) Transposon-mediated duplication of a gene associated with transposable elements (TE repeats) via replicative transposition to form dispersed duplicates.
Figure 2.
NLR duplication in wheat. (A) Whole-genome duplication (WGD) through an increase in ploidy. (B) Tandem duplication through unequal recombination between similar alleles to form a gene cluster. (C) Transposon-mediated duplication of a gene associated with transposable elements (TE repeats) via replicative transposition to form dispersed duplicates.
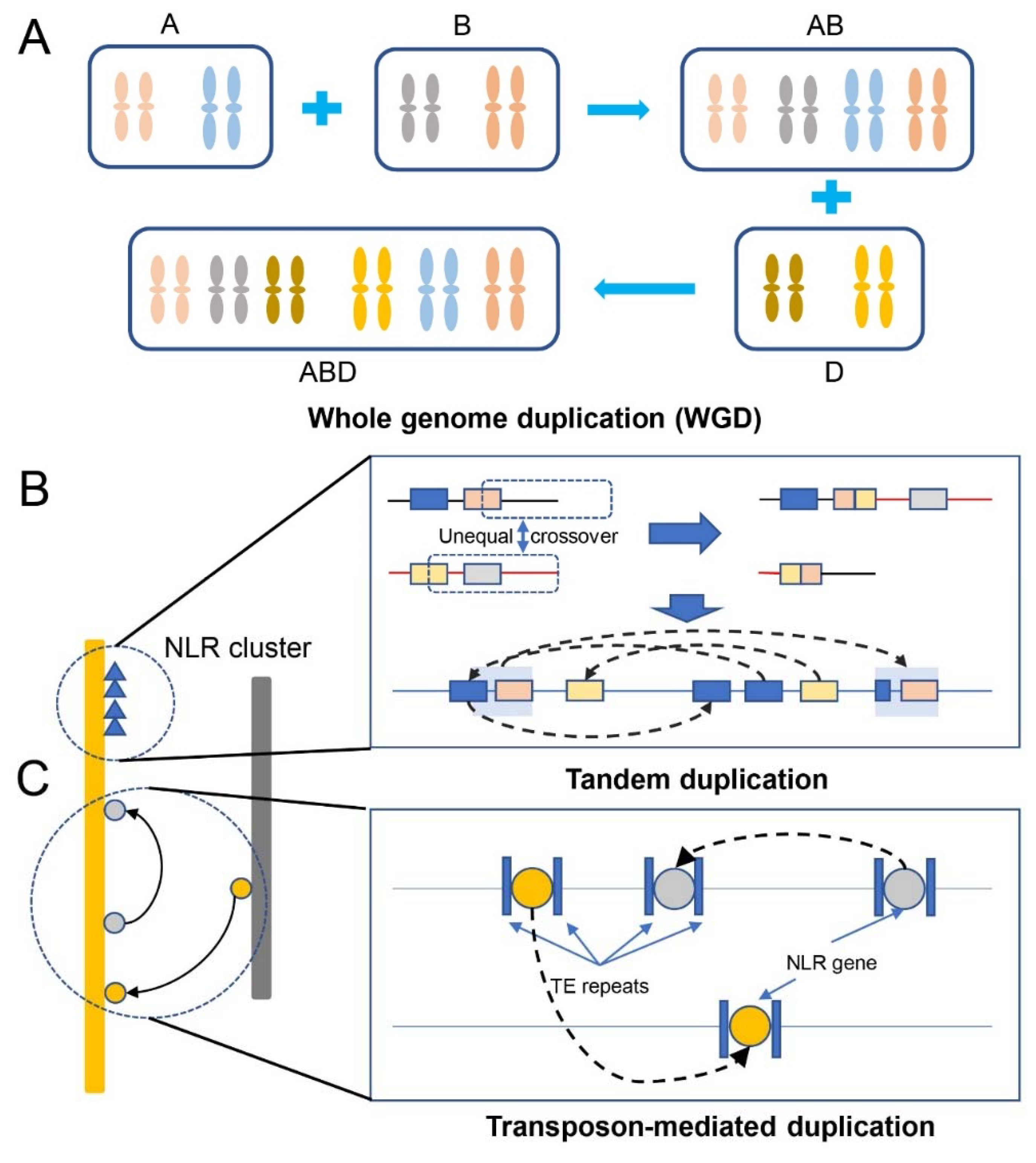
Figure 3.
Origination models of NLR gene with integrated domains.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



