
Submitted:
22 June 2023
Posted:
22 June 2023
You are already at the latest version
Abstract
Transitional waters are fragile ecosystems with high ecological values, representing the breeding and resting sites for rare and threatened species. They deserve particular concern for protection, undergoing numerous threats from anthropogenic nature. The present review aims to analyze the recent literature on Aphanius fasciatus, nowadays considered one of the most strictly estuarine-dependent fish species, and as such affected by the degradation of lagoon habitats, and to discuss its suitability as sentinel species of the quality of transitional water environments.
The analysis and discussion highlight the potential applicability of the molecular, cellular, and physiological responses of this species as diagnostic tools for detecting the subtle effects induced by environmental pollution on the biota in transitional water environments. Moreover, the suitability of the responses of this species is suggested in the wider framework of the One Health Perspective which considers human and animal health and the environmental state highly interconnected, sharing common aspects.
To date, omics technologies show very great potential in reacquiring novel knowledge on the responses of the organisms to environmental changes and to the alterations of the environmental health status. Therefore, considering the relevant potential of this species as sentinel species, many efforts are needed in the next future to improve the quantity and quality of the -omics tools that refer to A. fasciatus.
Keywords:
Aphanius fasciatus
; Transitional waters
; lagoon
; biomarker
; genomic
1. Introduction
Transitional waters are fragile ecosystems with high ecological values, representing the breeding and resting sites for rare and threatened species. According to the European Directive 92/43/EEC (Habitat Directive), transitional waters deserve particular concern for protection, undergoing numerous threats, such as impairment of water quality due to the river inputs enriched with nutrients and pollutants, destruction or reduction of habitats due to the construction of infrastructures (such as ports), urbanization, intensive aquaculture, the introduction of alien species, and so on.
Facca et al. [1] in their recent review proposed six fish species (namely Aphanius fasciatus, A. iberus, Knipowitschia panizzae, Ninnigobius canestrinii, Valencia hispanica and V. letourneuxi), more strictly related to salt marshes and wetlands and belonging to the guild of estuarine resident fish, as ecological bioindicators of lagoon environmental conditions, providing information on their presence and abundance in these ecosystems and thus on these ecosystems health (see also, e.g., [2].
This work focuses on one of these six species, i.e., A. fasciatus (Valenciennes, 1821), a small dimorphic fish found in large-sized populations in coastal brackish waters of the central-eastern Mediterranean Sea. Aphanius fasciatus is nowadays considered one of the most strictly estuarine-dependent fish species, and as such it may be affected by the degradation of lagoon habitats [3].
The species is of conservation interest; it is included in the Habitat Directive annex II (species requiring conservation measures) and spends its entire life cycle in the Mediterranean priority habitat 1150* “Coastal lagoons”. The International Union for Conservation of Nature (IUCN) Red List categories and criteria [4] includes A. fasciatus within the “Least Concern” category.
It is known that anthropogenic alterations of the environment may significantly contribute to population decline. In recent years, the study of the molecular and cellular responses of the organisms to anthropogenic environmental stressors (biomarkers) has known a very great impulse as it is seen as a very robust tool that contributes to supporting a variety of biodiversity conservation strategies. Biomarkers have recently been applied to several research areas of biodiversity conservation, including environmental quality monitoring of protected areas and assessment of the health status of species at risk (for review see, e.g., [5]). Moreover, in the framework of biodiversity conservation, the research on endangered species and the detection of their responses to environmental changes can widely benefit from the application of such emerging technologies, as genomic, proteomic, metabolomic, and bioinformatic, which show great potential in acquiring novel knowledge on the responses of the organisms to environmental changes and their impact on the organisms’ health status [6].
The aim of this review is to discuss the suitability of A. fasciatus as a sentinel species for the quality of transitional water environments. The use of its cellular and molecular responses as diagnostic tools for detecting the subtle effects induced by the environmental changes before more dramatic effects become evident on the distribution of the species (in terms of, e.g., abundance, reduction, or absence) could provide significant insights and help drawing novel perspectives in various biodiversity conservation fields.
This study was carried out in the framework of the Project “Monitoring Natura 2000 Sites, Species and Habitats in the Apulian Region (MoSSHa)”, POR Puglia FESR-FSE, included in the Natura 2000 network.
2. Aphanius fasciatus: biology, habitat preference, distribution, and conservation status
Aphanius fasciatus (class Actinopterygii, order Cyprinodontiformes, family Aphanidae, genus Aphanius) is a small dimorphic fish found in large-sized populations in coastal brackish waters of the central-eastern Mediterranean area except for the westernmost, where it is replaced by Aphanius iberus and Aphanius baeticus, and possibly the Aegean Sea, where Aphanius almirensis may replace it. In the south-eastern Mediterranean region, A. fasciatus partially overlaps Aphanius dispar. It also colonizes the Suez Canal and Red Sea. The distribution area of the species, as reported by the International Union for Conservation of Nature (IUCN), includes all the Mediterranean region countries except the Iberian Peninsula, limited to the coastal areas. It is also found in several Mediterranean islands [7].
The genus is defined ante-lessepsian because considered thriving in the ancient Tethys gulf before its closure at the end of Miocene (Messinian, 7 My ago) and survived the consequent drought of the Mediterranean basin [8]. Probably just a highly adjustable physiology allowed the ancient species to survive intense habitat variations of the Messinian period.
Aphanius fasciatus shows several specific morphological characteristics (Figure 1) when compared to the other species of the genus Aphanius found in Europe, such as the presence of light yellow to dark yellow pale in males, with a broad dark submarginal bar in some populations, 8-15 dark blue to gray bars on a silvery background, usually of regular shape, females with 11-17 dark brown short bars on the sides, a faint greyish mediolateral stripe, 24-29 scales in a mediolateral series on the body and pectoral fin with 14-15 rays [4].
Aphanius fasciatus tolerates harsh environmental conditions, including high temperatures and salinity, and low oxygen levels. Reproduction takes place from April to September with life history traits including external fertilization, demersal eggs deposited on benthic vegetation, short generation time (~3.5 yrs), high reproductive rate, and rapid population turnover. Diet is made of invertebrates, mainly crustaceans and insect larvae. A. fasciatus is typified by habitat fidelity in its adult phase, and hatchlings resemble adults’ lifestyles [1].
Despite its tolerance to wide variations in environmental physicochemical parameters, several aspects of the biology of the species confer vulnerability to environmental changes. For example, the intra-population component of the genetic variation of the species is much smaller than the inter-population component [9], a characteristic that determines a lower evolutionary potential in natural populations [10].
Despite the sporadic presence in coastal marine waters, possibly related to floods or dystrophic crises, the most common habitats of this species are transitional waters and a variety of fresh and brackish waters including small weedy ponds and ditches. This contributes to a very restricted gene flow that may occur only between adjacent populations. Mainly because of the virtually complete isolation of brackish-water habitats and the highly selective environmental features, A. fasciatus populations are highly divergent (see, e.g., [11] and literature cited therein). In Italy, two main clusters were identified with a similar genetic structure, the North Tyrrhenian populations above about latitude 41 °N and the southern Tyrrhenian and the Adriatic populations [12].
In recent years the interest in the conservation of this species has grown, due to the documented decline and in some cases even extinction of many A. fasciatus populations at several sites. The causes of the decline have to be found in the alteration and deterioration of natural biotopes and loss of habitats due to the increased pressure of human activities on coastal areas. In addition, competition with alien species such as Gambusia holbrooki Girard, 1859, a poecilid fish introduced in southern Europe to control the proliferation of malaria-carrying mosquitoes, caused a strong reduction of A. fasciatus over time [11]. This decline of the species has stimulated conservation actions, represented by, e.g., the inclusion of A. fasciatus in the lists of protected species. This species is now included in Annex II of the Habitats Directive of the European Union (Protocol ASPIM 92/43/EEC) and in Appendices II and III of the Bern Convention (Bern/Berne, 19.IX.1979). Currently, the conservation status of this species in the Mediterranean region is defined as “unfavourable-inadequate” [13].
In Italy, the historical analysis of the distribution of A. fasciatus along the Italian coasts has been carried out by Valdesalici et al. [11] based on the census of natural populations in the Italian coasts and bibliographic and museum material. From the historical analysis of the distribution of the species, the authors concluded that A. fasciatus has shown over time a reduction of its distribution along the Italian coasts around the 1990s, with a recovery in the following years. They outlined the presence of stable populations along the central Tyrrhenian, northern Adriatic, Sardinian, and Apulian coasts [11]. In particular, along Apulian coasts the presence of populations of A. fasciatus has been documented in the Pond of Acquatina (LE) [14; Denitto, 2002, personal communication (p.c.); Zonno, 2012, p.c.], Cesine (LE) [Denitto, 2002, p.c.], Alimini (LE) [Zonno, 2012, p.c.], Palude del Capitano - S. Isidoro (LE) [Bianchi, 1994, p.c.; Salvagiani, 2001, p.c.; Denitto, 2002, p.c.], Mar Piccolo of Taranto (TA) [15; 16; Baldacconi, 2013, p.c.], salt marshes of Margherita di Savoia (FG) [15; Bufo, 2000, p.c.], Lago di Lesina (FG) [17; 9; 18; 18; 19; 20; Cepollaro, 2006, p.c.; Valdesalici, 2008, p.c.], Lago di Varano (FG) [15; Piccolo, 2002, p.c.; Kärst, 2004, p.c.; Valdesalici, 2008, p.c.]. The presence of the species has been confirmed in November 2022 in Acquatina (LE) (IT9150003 Natura 2000 site) and Cesine (IT9150032 Natura 2000 site) during the scientific activities of the above-mentioned project.
3. Aphanius fasciatus as sentinel species for ecotoxicological risk of coastal lagoons
The scientific interest in A. fasciatus is documented by the increasing number of publications produced in the last decades, with 121 papers in Scopus reporting “Aphanius fasciatus” in the “Article title”, “Abstract” or “Keywords” fields starting in 1962, and 32 papers in PubMed reporting the same query in “All Fields” starting in 1979 [Last database consultation carried out on June 13, 2023] (see Figure 2). A bibliometric analysis, carried out in a comparative way by using the two data sources PubMed (Figure 2A) and Scopus (Figure 2B), and the mapping tool VOS viewer [21], highlights the state-of-the-art of the research on A. fasciatus and finds the emerging research trends on this species; this in the larger context of Cyprinodontiformes biology (for comparison, see Figure 2C and Figure 2D). It is rather evident that the information present in the database Scopus, which includes a wider variety of disciplines, is much more consistent and detailed with respect to that reported in a biomedicine-oriented database such as PubMed. In any case, the analysis carried out on these databases clearly indicates that over the years research on A. fasciatus species has changed, moving from the general analysis of the biology and genetics of the organism to studying more the biochemical, biomolecular, toxicological, and environmental facets of this species. The occurrence in more recent years of keywords such as “environmental monitoring” and “water pollutants, chemical” (see Figure 2A) or “environmental monitoring”, “phylogeography”, “coastal zone” and “lagoon” (Figure 2B) suggests the recent interest in this species as a bioindicator of coastal lagoons. Moreover, many keywords refer to Tunisia (because of the activity of a strong research group focusing on A. fasciatus in that country) and to a variety of Italian geographic areas, suggesting that there is a specific interest in this species in our country, too. Last but not least, “Aphanius fasciatus” links to certain keywords such as “fish diseases” and “spine” (Figure 2A) or “osteology”, “spine”, “animal experiment”, “controlled study”, “comparative study” and “physiology” among others (Figure 2B), showing its increasing interest as an animal model in the study of certain pathophysiological states. Therefore, the following discussion strictly reflects the available information on selected major research topics A. fasciatus has been studied in relation to its potential use as sentinel species for ecotoxicological risk of coastal lagoons.
3.1. A. fasciatus as sentinel species for metal pollution
Coastal lagoons are important areas for life between the land and the sea, representing highly productive environments and delivering several ecosystem services as well as ecological, cultural, and socioeconomic benefits [22,23]. However, due to their location between land and sea, they are subject to several anthropogenic pressures due to tourism, intensive aquaculture, and the input from their catchment areas receiving urban, agricultural, and/or industrial effluents.
Pollution remains one of the major environmental issues in coastal lagoons, and its contribution is exacerbated by global warming and more frequent torrential rains due to climate changes. Common pollutants found in Mediterranean coastal lagoons are trace metals from mining and industrial activities [24,25], polycyclic aromatic hydrocarbons (PAHs), polychlorinated biphenyls (PCBs), and organochlorinated pesticides (OCPs) from industrial and agricultural effluents [26,27].
Pollutant-induced alterations at the molecular, cellular, and physiological levels in the organisms (biomarkers) have extensively been used to detect the relationships between chemical contamination of the environmental matrices, internal levels of tissue contamination, and effects in the organisms [28,29]. Alterations measured at the molecular or cellular level have thus been proposed as sensitive “early warning” tools in environmental quality assessment as they allow detecting effects in target biota prior to more integrated effects can be “real” at higher levels of biological organization [5,30]. In this context, it is worth noting that: i) “exposure biomarkers” are early biochemical responses related to the exposure of the organism to pollutants, often represented by the detoxification responses of the organism to pollutant exposure; ii) “effect biomarkers” are represented by the toxicological effects exerted on the organisms by the exposure to pollutants and are directly related to possible health impairment or disease; iii) “susceptibility biomarkers” are intrinsic or acquired abilities of an organism that account for increased sensitivity to the effects of an environmental pollutant [5].
Fish species are recognized as key elements for the assessment of the quality of aquatic ecosystems thanks to their high ecological relevance due to their influence on food web structure, nutrient cycling, and energy transfer. Indeed, fish species are widely used for aquatic health assessment through the biomarker approach but the choice of fish bioindicator species needs to take into account several issues including the potential pathways of exposure to the contaminant of concern, and the detectable biological responses that they are able to develop [31].
In the case of A. fasciatus some characteristics of its biology and ecology, such as the benthivore nature of the adults and their preference for terminal sections of rivers and closed zones, make this species susceptible to exposure to chemical pollutants. The benthivorous diet exposes the animals to high doses of chemical pollutants because benthic invertebrates are known to concentrate contaminants [32] from sediments, which are the sink of contamination present in the water column. Moreover, estuarine environments are known to receive pollutants through industrial discharges, yard runoff, streets, agricultural lands, and storm drains; also, closed zones are more polluted than the surrounding areas due to low hydrodynamism. In this regard, the sedentary behavior and the high territoriality of A. fasciatus makes this species very adequate for pollution monitoring in transitional water environments.
Regarding the biomarker responses of A. fasciatus, some studies are already available in the literature and are mainly related to metal contamination. For example, Kessabi et al. [33] in environments contaminated by heavy metals have found a relationship between metal concentrations in water, sediment, and tissues of A. fasciatus natural populations from the Gulf of Gabes (Tunisia) – in particular for Cd, Zn, and Cu – thus suggesting its ability to bioaccumulate trace metals from the environment. Moreover, metallothionein induction has been observed in the liver of metal-exposed animals. Metallothioneins are cysteine-rich metal-binding proteins involved in the regulation of the metabolism of trace metals and protection against heavy metal toxicity and oxidative stress in a wide range of organisms [34,35,36]. Therefore, the results reported by Kessabi et al. [33] indicate the ability of the species to develop a detoxification response after metal exposure. The A. fasciatus natural populations from the Gulf of Gabes (Tunisia) exposed to metal contamination also show liver histological alterations and deformities, such as compressed vertebral discs and general distortion of the vertebral column accompanied by vertebral lesions [33]. It is known that metals can interfere with calcium homeostasis [37]; thus, the alteration reported by the authors could be attributed to the toxicological mechanism(s) directly or indirectly affecting calcium homeostasis. This study suggests that the species is responsive to metal exposure showing bioaccumulation and the activation of exposure and effect biomarker responses. Indeed, it can accumulate heavy metals in its tissues at higher levels than the toxic concentration present in its environment, presumably due to active absorption along the gill and gut epithelium. Also, the species shows metallothionein induction, which represents a widely used metal exposure biomarker. Moreover, A. fasciatus shows effect biomarkers such as liver histological alterations and skeletal deformities when the contamination exposure exceeds the detoxification capacity of the organism.
Spinal deformities had already been detected by Messaoudi et al. [38] in the A. fasciatus natural populations from the Gulf of Gabes, with kyphosis, scoliosis, and lordosis frequently co-occurring at varying degrees of severity. This condition paralleled the high frequencies of spinal deformities in wild specimens of other teleost fish species in polluted environments [39,40,41]. As found by Kessabi et al. [33], the skeletal deformities observed in natural populations are related to the ability of A. fasciatus to accumulate large amount of Cd. The occurrences of skeletal deformity in fish have thus been proposed by Messaoudi et al. [38] as a good and practical response to assessing environmental quality, i.e., as an effect biomarker. The responses observed in A. fasciatus are similar to the symptoms described in the human species in the case of the Hitai Hitai disease caused by cadmium accumulation [42]. This suggests the suitability of the responses of this species to be considered in the wider framework of the One Health Concept which considers human and animal health and the environmental state highly interconnected, sharing common aspects that can be applied globally in these three components. In this perspective, a broader vision of the use of biomarkers of risk assessment is developing bridging environmental health and human health in a more global vision [43,44]. Therefore, the suitability of biomarker responses for both environmental and human biomonitoring is of great usefulness for the development of this emerging approach.
Annabi et al. [45] detected an accumulation of trace metals in the liver and gonads in natural populations of A. fasciatus exposed to environments characterized by trace metal pollution. The bioaccumulation resulted in parallel disturbance of the reproductive status in these natural populations, in terms of, e.g., sex ratio unbalance and alteration of sexual hormone blood concentrations. Thus, A. fasciatus is also able to express effect biomarkers related to alterations in reproduction.
Mosesso et al. [36] assessed the suitability of Aphanius fasciatus as a sentinel organism to detect complex genotoxic mixtures in a coastal lagoon ecosystem through the comet assay, which represents one of the most used assays for the evaluation of genotoxic damage. Following optimization and validation of the assay under laboratory conditions, fish were collected in the Orbetello lagoon (Tuscany, Italy), which is considered a significantly polluted site. The results showed statistically significant increases for tail DNA (%) compared to the values observed in erythrocytes of fish caught in the unpolluted reference site “Saline di Tarquinia”. The physico-chemical parameters of the water (i.e., salinity, pH and oxygen content) did not significantly influence the induction of DNA damage. These results indicate that the comet assay provides a reliable parameter and that A. fasciatus is a promising sentinel organism to detect the genotoxic impact of complex mixtures in coastal lagoon ecosystems.
Sebbio et al. [47] also found accumulation of metals in A. fasciatus natural populations from Civitavecchia area and, in parallel, genotoxic damage as assessed by comet assay. The results obtained in the natural populations of A. fasciatus mirrored a trend of increasing environmental pollution across time and space, which was evident by accumulation profiles consistent with DNA damage profiles. These results confirm A. fasciatus as a highly sensitive species in detecting heavy metal pollution.
4. Perspectives
The requirements for a species to be considered an optimal animal sentinel are represented by the sensitivity to environmental stressors and the ability to develop measurable responses, a home range overlapping the area to be monitored, and easy cataloging and capture. The review of the literature available on A. fasciatus reported in the previous section demonstrates that A. fasciatus meets these requirements and, in a perspective, outlines the usefulness of this species as sentinel species of the quality of transitional water environments. In this respect, some aspects deserve to be considered. The species shows high sensitivity to exposure to contamination and also exhibits the ability to develop detectable responses. However, the data available mainly focus on heavy metal contamination while other pollutants known to be widely present in coastal lagoons including PAHs, PCBs and OCPs have not been considered yet. Moreover, considering that coastal lagoons are particularly vulnerable to global climate change, the investigation of the pollutant responses of A. fasciatus under multiple stress conditions, e.g., related to climate change such as high temperatures and salinity and low oxygen levels, would be worth to be studied in order to exalt the role of this species as sentinel organism.
To date, genomics, transcriptomics, proteomics, metabolomics, bioinformatics, and all the other emerging -omics technologies show very great potential in acquiring novel knowledge on the responses of the organisms to environmental changes and to the alterations of the environmental health status [6]. Modern genomics offers high-throughput and information-rich approaches for characterizing the biological responses to environmental stressors and for assessing the impacts of pollutants on living organisms. Not only, such technologies also offer very useful tools to discover the consequences of exposure to environmental pollution at the population level and for understanding the bases of deployment in population-level long-term monitoring programs [48]. These approaches can thus be of relevance for environmental monitoring purposes as well as in the framework of biodiversity conservation.
Aphanius fasciatus belongs to the order Cyprinodontiformes, which expresses as many as 66 genomes per 47 species to date (NCBI database last queried in May 2023) (Table 1). Of the various families of Cyprinodontiformes, at least 5 (i.e., Fluviphylacidae, Profundulidae, Pantanodontiidae, Procatopodidae and Valencidae) still lack a sequenced genome, but all the other families include at least one species for which a reference genome is available (Table 1).
Notably, all the Cyprinodontiformes are characterized by a natural ability to adapt to extreme environments. Emblematic is the case of Nothobranchius furzeri, the African turquoise killifish (Table 1) typical of mud pools in the African savannah, which completes its life cycle in ~3 months and today represents an irreplaceable model for the study of aging and aging-related diseases (see, e.g., [49,50]). This suggests also the perspective of the possible use of A. fasciatus – like other Cyprinodontiformes – as a suitable animal model, to understand how aquatic animals tolerate harsh environmental conditions, including high temperatures and salinity, and low oxygen levels. The knowledge in this field is of great relevance also in the framework of the global climate changes that can exacerbate pollution conditions in vulnerable environments such as coastal lagoons.
Therefore, it is worth underlining that in the next future many efforts are needed to improve quantity and quality of the -omics tools that refer to A. fasciatus. In fact, its genome is not yet available, as shown in Table 1, where the list of the genomes of Cyprinodontiformes catalogued at National Center for Biotechnology Information (NCBI) is reported. However, the complete mitochondrial genome of the very close species Aphanius iberus (the Spanish toothcarp) has recently been published [51] (see Table 1). In addition, while lacking of any annotated and/or simply sequenced genome, only a few hundred sequences – the large majority of which referring to mitochondrial and not to nuclear DNA, RNA and protein – associate to A. fasciatus. This piece of information is briefly summarized in Table 2.
5. Conclusions
In conclusion, the analyses and discussion of the literature on A. fasciatus highlight the suitability of this species as a sentinel organism of the quality of transitional water environments and the use of its molecular, cellular, and physiological responses as diagnostic tools for detecting the subtle effects induced by environmental pollution on the biota in these vulnerable environments. The peculiar physiological traits of this species also suggest its usefulness as a suitable animal model, to understand tolerance to harsh environmental conditions, including high temperatures and salinity, and low oxygen levels, with relevant implications for the research on global climate change adaptation. Moreover, the increasing interest on A. fasciatus as an animal model in the study of certain pathophysiological states mainly related to metal exposure also suggests the suitability of the responses of this species in the wider framework of the One Health perspectives which considers human and animal health and the environmental state highly interconnected, sharing common aspects.
To date, omics technologies show very great potential in reacquiring novel knowledge on the responses of the organisms to environmental changes and to the alterations of the environmental health status. Therefore, considering the relevance of this species as sentinel species, many efforts are needed in the next future to improve the quantity and quality of the -omics tools that refer to A. fasciatus.
Acknowledgments
This research was funded by Project “Monitoring Natura 2000 Sites, Species and Habitats in the Apulian Region (MoSSHa)”, POR Puglia FESR-FSE, CUP: F85F20000820002.
References
- Facca, C.; Cavraro, F.; Franzoi, P.; Malavasi, S. Lagoon Resident Fish Species of Conservation Interest According to the Habitat Directive (92/43/CEE): A Review on Their Potential Use as Ecological Indicator Species. Water 2020, 12, 2059. [Google Scholar]
- Gilby, B.L.; Olds, A.D.; Connolly, R.M.; Yabsley, N.A.; Maxwell, P.S.; Tibbetts, I.; Schoeman, D.S.; Schlacher, T.A. Umbrellas can work under water: Using threatened species as indicator and management surrogates can improve coastal conservation. Estuar. Coast. Shelf Sci. 2017, 199, 132–140. [Google Scholar]
- Andolina, C.; Franzoi, P.; Cavraro, F.; Jackson, A.L.; Mazzola, A.; Vizzini, S. Trophic adaptability shapes isotopic niche of the resident fish Aphanius fasciatus across lagoon habitats. Estuar. Coast. Shelf Sci. 2022, 264, 107685. [Google Scholar]
- Kottelat, M.; Freyhof, J. Handbook of European Freshwater Fishes; Kottelat, Cornol and Freyhof: Berlin, Germany, 2007.
- Lionetto, M.G.; Caricato, R.; Giordano, M.E. Pollution biomarkers in the framework of marine biodiversity conservation: State of art and perspectives. Water 2021, 13, 1847. [Google Scholar]
- Gouveia, D; Almunia, C.; Cogne, Y.; Pible, O.; Degli-Esposti, D.; Salvador, A.; Cristobal, S.; Sheehan, D.; Chaumot, A.; Geffard, O.; et al. Ecotoxicoproteomics: A decade of progress in our understanding of anthropogenic impact on the environment. J. Proteomics 2019, 198, 66–77. [Google Scholar]
- Bianco, P.G.; Ahnelt, H.; Economidis, P.S. The freshwater fishes from eastern and large Mediterranean islands with comments on their safety status. Acta Universitatis Carolinae Biologica 1996, 40, 45–60. [Google Scholar]
- Por, F.D. The legacy of Tethys. An aquatic biogeography of the Levant. Kluwer Academic Publishers, Dordrecht, Netherlands, 1989.
- Maltagliati, F. 1999. Genetic divergence in natural populations of the Mediterranean brackish-water killifish Aphanius fasciatus. Mar. Ecol. Prog. Ser. 1999, 179, 155–162. [Google Scholar]
- Toro, M.A.; Caballero, A. 2005. Characterization and conservation of genetic diversity in subdivided populations. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2005, 360, 1367–1378. [Google Scholar]
- Valdesalici, S.; Langeneck, J.; Barbieri, M.; Castelli, A.; Maltagliati, F. Distribution of natural populations of the killifish Aphanius fasciatus (Valenciennes, 1821) (Teleostei: Cyprinodontidae) in Italy: past and current status, and future trends. It. J. Zool. 2015, 82, 212–223. [Google Scholar]
- Cimmaruta, R.; Scialanca, F.; Luccioli, F.; Nascetti, G. Genetic diversity and environmental stress in Italian populations of the cyprinodont fish Aphanius fasciatus. Oceanol. Acta 2003, 26, 101–110. [Google Scholar]
- European Environmental Agency (EEA). Report under the Article 17 of the Habitats Directive, Period 2007-2012.
- De Mitri, R. Pesci e crostacei decapodi del bacino di Acquatina (Lecce). Thalassia Salentina 2004, 27, 21–32. [Google Scholar]
- Costa, O.G. Genere Lebia. In: Fauna del Regno di Napoli. Pesci. Parte prima. Stabilimento tipografico Fr. Azzolino: Napoli, 1850, pp. 32–38.
- Cavicchioli, G. Considerazioni bio-ecologiche sopra Aphanius fasciatus Valenciennes (Cyprinodontidae). Bollettino di Zoologia 1962, 29, 713–719. [Google Scholar] [CrossRef]
- Tigano, C.; Ferrito, V. Studio biometrico e morfologico in popolazioni di Aphanius fasciatus (Nardo) dell’Adriatico e di Sicilia (Pisces, Cyprinodontidae). Nova Thalassia 1984, 6, 679–680. [Google Scholar]
- Ferrito V, Maltagliati F, Maceri A, Adorno A, Tigano C (2003) Morphological and genetic variation in four Italian populations of Lebias fasciata (Teleostei, Cypridontidae). Ital. Journal of Zoology 2003, 70, 115–121. [CrossRef]
- Ferrito, V.; Mannino, M.C.; Pappalardo, A.M.; Tigano, C. Morphological variation among populations of Aphanius fasciatus Nardo, 1827 (Teleostei, Cyprinodontidae) from the Mediterranean. J. Fish Biol. 2007, 70, 1–20. [Google Scholar]
- Manzo, C. Fish assemblages in three Mediterranean coastal lagoons: structure, functioning and spatio-temporal dynamics. PhD Thesis, Università di Roma “Tor Vergata”, Rome, Italy, 2010. [Google Scholar]
- van Eck, N.J.; Waltman, L. Software survey: VOSviewer, a computer program for bibliometric mapping. Scientometrics 2010, 84, 523–538. [Google Scholar] [PubMed]
- Gönenç, I.E.; Wolflin, J.P. Coastal Lagoons Ecosystem Processes and Modeling for Sustainable Use and Development. CRC Press: Boca Raton, Florida, 2005.
- Newton, A.; Brito, A.C.; Icely, J.D.; Derolez, V.; Clara, I.; Angus, S.; Schernewski, G.; Inacio, M.; Lillebø, A.I.; Sousa, A.I.; Bejaoui, B.; Solidoro, C. Assessing, quantifying and valuing the ecosystem services of coastal lagoons. J. Nat. Conserv. 2018, 44, 50–65. [Google Scholar]
- Martins, M.V.; Helali, M.A.; Zaaboub, N.; Boukef-BenOmrane, I.; Frontalini, F.; Reis, D.; Portela, H.; Clemente, I.M.; Nogueira, L.; Pereira, E.; Miranda, P.; El Bour, M.; Aleya, L. Organic matter quantity and quality, metals availability and foraminiferal assemblages as environmental proxy applied to the Bizerte Lagoon (Tunisia). Mar. Pollut. Bull. 2016, 105, 161–179. [Google Scholar] [PubMed]
- Lacoste, E.; Jones, A.; Callier, M.; Klein, J.; Lagarde, F.; Derolez, V. A Review of Knowledge on the Impacts of Multiple Anthropogenic Pressures on the Soft-Bottom Benthic Ecosystem in Mediterranean Coastal Lagoons. Estuaries Coast. 2023. [Google Scholar]
- LeónLeon, V.M.; R. Moreno-GonzálezGonzalez, R.; V. GarcíaGarcia, V.; J.A. Campillo, J.A. 2017. Impact of flash flood events on the distribution of organic pollutants in surface sediments from a Mediterranean coastal lagoon (Mar Menor, SE Spain). Environ. Sci. Pollut. Res. Int. 2017, 24, 4284–4300. [Google Scholar]
- Ben Salem, F.; Ben Said, O.; Cravo-Laureau, C.; Mahmoudi, E.; Bru, N.; Monperrus, M.; Duran, R. Bacterial community assemblages in sediments under high anthropogenic pressure at Ichkeul Lake/Bizerte Lagoon hydrological system, Tunisia. Environ. Pollut. 2019, 252, 644–656. [Google Scholar]
- Chambers, J.E.; Boone, J.S.; Carr, R.L.; Chambers, H.W.; Straus, D.L. Biomarkers as predictors in health and ecological risk assessment. Hum. Ecol. Risk Assess. Manag. 2002, 8, 165–176. [Google Scholar]
- Connon RE, Geist J, Werner I. Effect-based tools for monitoring and predicting the ecotoxicological effects of chemicals in the aquatic environment. Sensors (Basel) 2012, 12, 12741–12771. [Google Scholar]
- Hook, S.E.; Gallagher, E.P.; Batley, G.E. The role of biomarkers in the assessment of aquatic ecosystem health. Integr. Environ. Assess. Manag. 2014, 10, 327–341. [Google Scholar] [PubMed]
- Kroon, F.; Streten, C.; Harries, S. A protocol for identifying suitable biomarkers to assess fish health: A systematic review. PLoS One, 2017, 12, e0174762. [Google Scholar]
- Varanasi, U.; Stein, J.E. Disposition of xenobiotic chemicals and metabolites in marine organisms. Environ. Health Perspect. 1991, 90, 93–100. [Google Scholar]
- Kessabi, K.; Hwas, Z.; Sassi, A.; Said, K.; Messaoudi, I. Heavy metal accumulation and histomorphological alterations in Aphanius fasciatus (Pisces, Cyprinodontidae) from the Gulf of Gabes (Tunisia). Environ. Sci. Pollut. Res. Int. 2014, 21, 14099–14109. [Google Scholar]
- Lionetto, M. G, Giordano M.E, Caricato R, Pascariello M.F, Marinosci L, Schettino T. Biomonitoring of heavy metal contamination along the salento Coast (Italy) by metallothionein evaluation in Mytilus galloprovincialis and Mullus barbatus. Aquat. Conserv. 2001, 11, 305–310. [Google Scholar]
- Calisi, A.; Lionetto, M.G.; Lemanni, A.; De Lorenzis, E.; Schettino, T. Metallothionein induction in the coelomic fluid of the earthworm Lumbricus terrestris following heavy metal exposure: a short report. Biomed. Res. Int. 2014, Article ID 109386, pp.1-6.
- Melvin, S. Samuel, Saptashwa Datta, Rahul Sampat Khandge, Ethiraj Selvarajan. A state of the art review on characterization of heavy metal binding metallothioneins proteins and their widespread applications. Science of The Total Environment, Volume 775, 2021, 145829.
- Suzuki, N.; Yamamoto, M.; Watanabe, K.; Kambegawa, A.; Hattori, A. Both mercury and cadmium directly influence calcium homeostasis resulting from the suppression of scale bone cells: the scale is a good model for the evaluation of heavy metals in bone metabolism. J. Bone Miner. Metab. 2004, 22, 439–446. [Google Scholar]
- Messaoudi, I.; Kessabi, K.; Kacem, A.; Said, K. Incidence of spinal deformities in natural populations of Aphanius fasciatus Nardo, 1827 from the Gulf of Gabes, Tunisia. Afr. J. Ecol. 2009, 3, 360–366. [Google Scholar]
- Slooff, W. Skeletal anomalies in fish from polluted surface waters. Aquat. Toxicol. 1982, 2, 157–173. [Google Scholar]
- Loganathan, B.G.,Tanabe, S.,Tatsukawa, R.,Ogawa,K.&Miyako,G. Temporal changes of morphological abnormalitiesand parasitic infestation in fishes from the river Nagaragawa.Jpn. Nippon Suisan Gakkaishi 1989, 55,769–774.
- Whittle, D.M.; Sergeant, D.B.; Huestis, S.Y.; Hyatt, W.H. Food chain accumulation of PCDF isomers in the Great Lakesaquatic community. Chemosphere 1992, 25, 181–184. [Google Scholar] [CrossRef]
- Inaba, T.; Kobayashi, E.; Suwazono, Y. Mirei Uetani, Mitsuhiro Oishi, Hideaki Nakagawa, Koji Nogawa. Estimation of cumulative cadmium intake causing Itai–itai disease. Toxicol. Lett. 2005, 159, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Lionetto, M.G.; Caricato, R.; Giordano, M.E. . Pollution biomarkers in environmental and human biomonitoring. Open Biomark. J. 2019, 9, 1–9. [Google Scholar]
- Franco-Martínez, L.; Martínez-Subiela, S.; Cerón, J.J.; Tecles, F.; Eckersall, P.D.; Oravcova, K.; Tvarijonaviciute, A. Biomarkers of health and welfare: A One Health perspective from the laboratory side. Res. Vet. Sci. 2020, 128, 299–307. [Google Scholar] [PubMed]
- Annabi, A.; Said, K.; Messaoudi, I. Cadmium: Bioaccumulation, histopathology and detoxifying mechanisms in Fish. Am. J. Res. Comm. 2013, 1, 60–79. [Google Scholar]
- Mosesso, P.; Angeletti, D.; Pepe, G.; Pretti, C.; Nascetti, G.; Bellacima, R.; Cimmaruta, R.; Jha, A.N. The use of cyprinodont fish, Aphanius fasciatus, as a sentinel organism to detect complex genotoxic mixtures in the coastal lagoon ecosystem. Mutat. Res. 2012, 742, 31–36. [Google Scholar]
- Sebbio, C.; Carere, C.; Nascetti, G.; Bellisario, B.; Mosesso, P.; Cimmaruta, R.; Angeletti, D. Interspecies variation in DNA damage induced by pollution. Curr. Zool. 2014, 60, 308–321. [Google Scholar] [CrossRef]
- Reid, N.M.; Whitehead, A. Functional genomics to assess biological responses to marine pollution at physiological and evolutionary timescales: Toward a vision of predictive ecotoxicology. Brief. Funct. Genomics 2016, 15, 358–364. [Google Scholar]
- Terzibasi-Tozzini, E.; Cellerino, A. Nothobranchius annual killifishes. Evodevo. 2020, 11, 25. [Google Scholar]
- Holtze, S.; Gorshkova, E.; Braude, S.; Cellerino, A.; Dammann, P.; Hildebrandt, T.B.; Hoeflich, A.; Hoffmann, S.; Koch, P.; Terzibasi-Tozzini, E.; Skulachev, M.; Skulachev, V.P.; Sahm, A. Alternative Animal Models of Aging Research. Front. Mol. Biosci. 2021, 8, 660959. [Google Scholar] [PubMed]
- Lopez-Solano, A.; Nester, T.L.; Perea, S.; Doadrio, I. Complete mitochondrial genome of the Spanish toothcarp, Aphanius iberus (Valenciennes, 1846) (Actinopterygii, Aphaniidae) and its phylogenetic position within the Cyprinodontiformes order. Mol. Biol. Rep. 2023, 50, 2953–2962. [Google Scholar] [PubMed]
Figure 1.
Representative image of a male (A) and a female (B) specimen of Aphanius fasciatus.

Figure 2.
(A, B) Visualization of the main terms contained in “All Fields” after querying PubMed (A) and in the “Title”, “Abstract” and “Keywords” fields after querying Scopus (B) literature collections, as obtained by using the two (combined) keywords “Aphanius fasciatus” and the bibliometric mapping tool VOSviewer [Please note: for A, Number of papers retrieved in PubMed: 32; Minimum number of occurrences of a (MeSH) keyword: 5; of the 163 (MeSH) keywords, 12 met the threshold; for B, Number of papers retrieved in Scopus: 121, Minimum number of occurrences of a keyword: 5; of the 1096 keywords, 80 met the threshold]. (C, D) Visualization of the main terms contained in “All Fields” after querying PubMed (C) and in the “Title”, “Abstract” and “Keywords” fields after querying Scopus (D) literature collections, as obtained by using the keyword “Cyprinodontiformes” filtered for “Review” and VOSviewer [Please note: for C, Number of papers retrieved in PubMed: 127; Minimum number of occurrences of a (MeSH) keyword: 5; of the 504 (MeSH) keywords, 54 met the threshold; for D, Number of papers retrieved in Scopus: 127, Minimum number of occurrences of a keyword: 5; of the 1741 keywords, 104 met the threshold]. The range of colors used from blue to yellow indicates the temporal range of keywords, from those associated with older to those associated with more recent texts [Last database consultation carried out on June 13, 2023].
Figure 2.
(A, B) Visualization of the main terms contained in “All Fields” after querying PubMed (A) and in the “Title”, “Abstract” and “Keywords” fields after querying Scopus (B) literature collections, as obtained by using the two (combined) keywords “Aphanius fasciatus” and the bibliometric mapping tool VOSviewer [Please note: for A, Number of papers retrieved in PubMed: 32; Minimum number of occurrences of a (MeSH) keyword: 5; of the 163 (MeSH) keywords, 12 met the threshold; for B, Number of papers retrieved in Scopus: 121, Minimum number of occurrences of a keyword: 5; of the 1096 keywords, 80 met the threshold]. (C, D) Visualization of the main terms contained in “All Fields” after querying PubMed (C) and in the “Title”, “Abstract” and “Keywords” fields after querying Scopus (D) literature collections, as obtained by using the keyword “Cyprinodontiformes” filtered for “Review” and VOSviewer [Please note: for C, Number of papers retrieved in PubMed: 127; Minimum number of occurrences of a (MeSH) keyword: 5; of the 504 (MeSH) keywords, 54 met the threshold; for D, Number of papers retrieved in Scopus: 127, Minimum number of occurrences of a keyword: 5; of the 1741 keywords, 104 met the threshold]. The range of colors used from blue to yellow indicates the temporal range of keywords, from those associated with older to those associated with more recent texts [Last database consultation carried out on June 13, 2023].
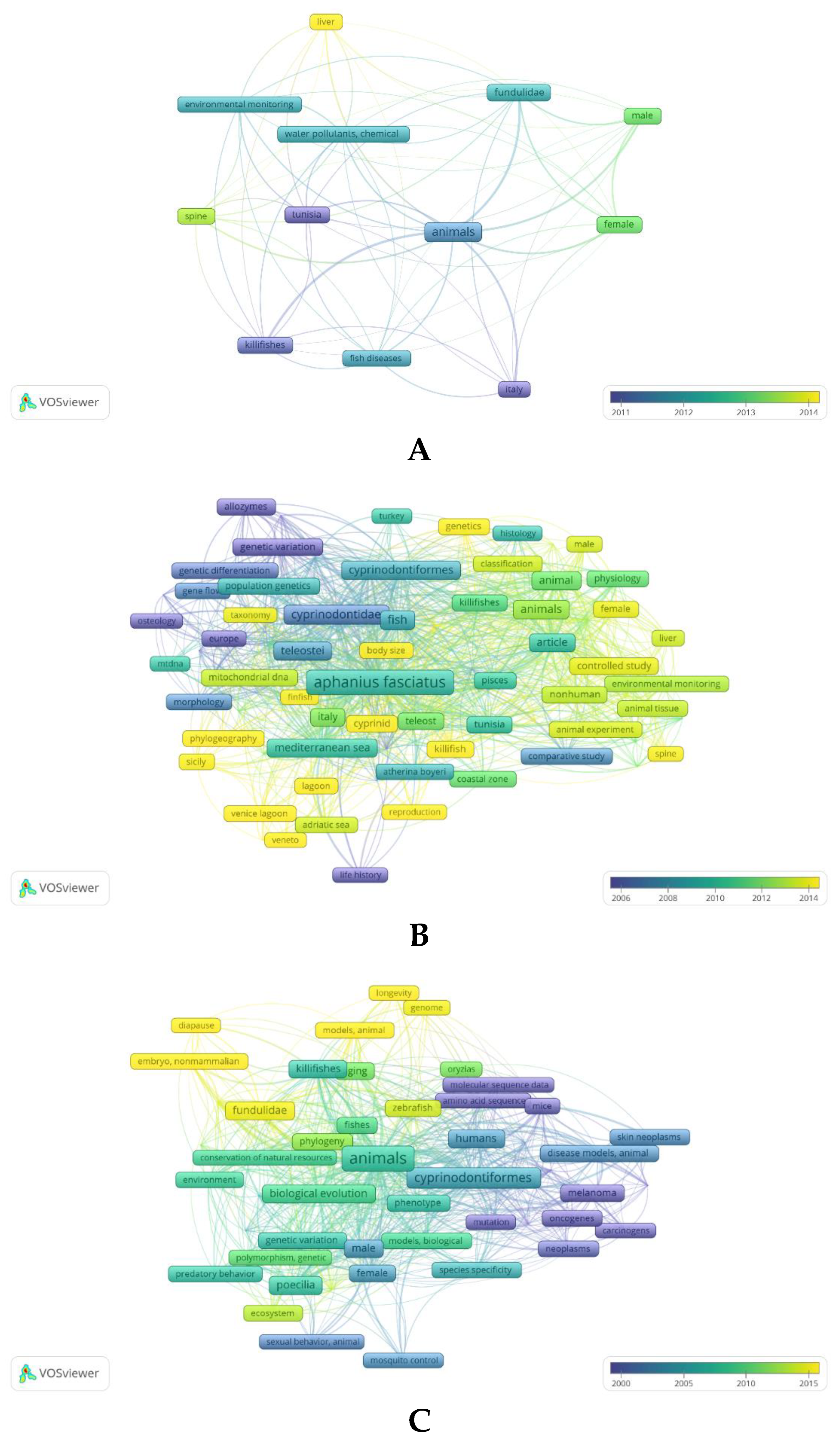
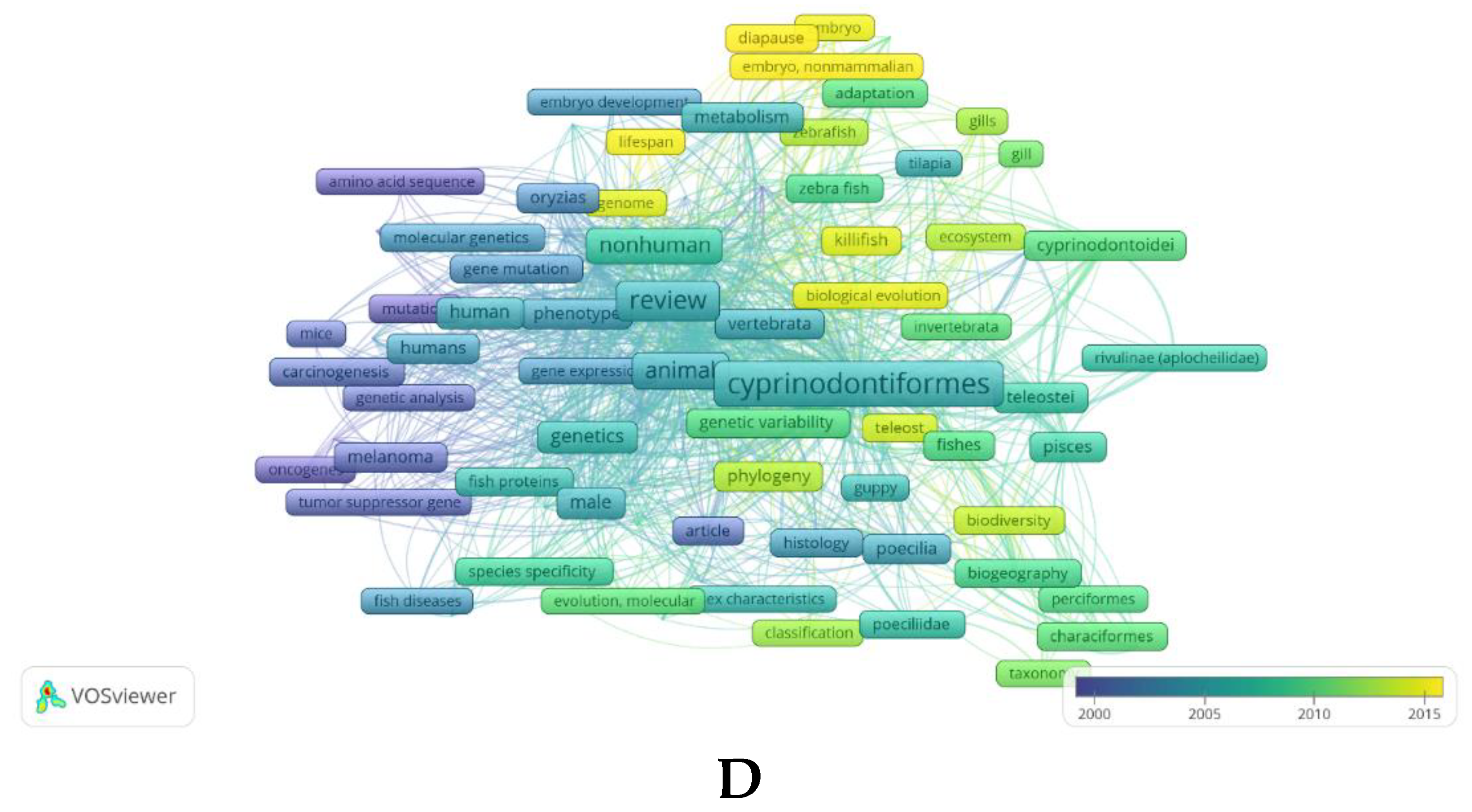

Table 1.
List of Cyprinodontiformes genomes available at NCBI (source: https://www.ncbi.nlm.nih.gov/datasets/taxonomy/28738/).
Table 1.
List of Cyprinodontiformes genomes available at NCBI (source: https://www.ncbi.nlm.nih.gov/datasets/taxonomy/28738/).
| Order (Suborder) |
Family | Genus | Species | Sequenced genomes |
|---|---|---|---|---|
| Cyprinodontiformes (killifishes and others) | 66 | |||
| (Aplocheiloidei) | Rivulidae (New World rivulines) | Austrofundulus Kryptolebias Nematolebias |
Austrofundulus limnaeus Kryptolebias brasiliensis Kryptolebias gracilis Kryptolebias hermaphroditus Kryptolebias marmoratus (mangrove rivulus) Kryptolebias ocellatus (sardinita) Nematolebias whitei (Rio pearlfish) |
1£ 1£ 1£ 1$ 2$ 1£ 1$ |
| Nothobranchiidae | Aphyosemion Callopanchax Nothobranchius |
Aphyosemion australe (lyretail killifish) Callopanchax toddi Nothobranchius furzeri (turquoise killifish) Nothobranchius kuhntae (Beira killifish) |
1£ 1£ 6£ 2£ |
|
| Aplocheilidae (rivulines) | Pachypanchax | Pachypanchax playfairii (golden panchax) | 1£ | |
| (Cyprinodontoidei) | Anablepidae | Anableps | Anableps anableps (largescale foureyes) | 2$ |
| Aphaniidae | Aphanius | Aphanius iberus (Spanish toothcarp) | 1£ | |
| Fluviphylacidae | - | - | - | |
| Goodeidae (goodeids) | Girardinichthys | Girardinichthys multiradiatus | 1$ | |
| Cyprinodontidae (killifishes) | Cyprinodon |
Cyprinodon brontotheroides Cyprinodon variegatus (sheepshead minnow) Cyprinodon nevadensis (Amargosa pupfish) Cyprinodon tularosa |
1£ 1£ 1£ 1& |
|
| Poeciliidae (livebearers) | Alfaro Brachyrhaphis Gambusia Girardinus Micropoecilia Phalloptychus Poecilia Poeciliopsis Xiphophorus |
Alfaro cultratus Brachyrhaphis roseni Brachyrhaphis terrabensis Gambusia holbrooki (eastern mosquitofish) Gambusia affinis (western mosquitofish) Girardinus metallicus (metallic livebearer) Micropoecilia bifurca Phalloptychus januarius Poecilia mexicana (shortfin molly) Poecilia formosa (Amazon molly) Poecilia latipinna (sailfin molly) Poecilia gillii (Gill’s molly) Poecilia reticulata (guppy) Poecilia wingei Poecilia picta (swamp guppy) Poecilopsis turrubanensis Poecilopsis paucimaculata Poecilopsis occidentalis (Gila topminnow) Poecilopsis infans (Lerma livebearer) Poecilopsis prolifica (blackstripe livebearer) Poecilopsis gracilis (portohole livebearer) Poecilopsis retropinna Poecilopsis turneri (blackspotted livebearer) Poecilopsis presidionis (Sinaloa livebearer) Xiphophorus hellerii (green swordtail) Xiphophorus couchianus (Monterrey platyfish) Xiphophorus maculatus (southern platyfish) |
1£ 1£ 1£ 1£ 2$ 1£ 1£ 1& 1£ 3£ 1£ 1£ 2$ 1£ 2& 1£ 1£ 1£ 1£ 2£ 2& 1£ 2& 1£ 2$ 1$ 2$ |
|
| Profundulidae (Middle American killifishes) | - | - | - | |
| Pandanodontidae | - | - | - | |
| Procatopodidae | - | - | - | |
| Fundulidae (topminnows) | Fundulus | Fundulus heteroclitus (mummichog) | 2$ | |
| Valenciidae | - | - | - |
$Reference genome available, chromosome level; £Reference genome available, scaffold level; &Reference genome available, contig level (Database last query: May 2023).
Table 2.
Aphanius fasciatus sequence and literature data from the database Taxonomy (https://www.ncbi.nlm.nih.gov/Taxonomy/Browser/wwwtax.cgi?id=30736).
Table 2.
Aphanius fasciatus sequence and literature data from the database Taxonomy (https://www.ncbi.nlm.nih.gov/Taxonomy/Browser/wwwtax.cgi?id=30736).
| Entrez records | |
|---|---|
| Database name | Number of relevant data available |
| Nucleotide | 453 |
| Protein | 211 |
| Popset | 29 |
| PubMed Central | 31 |
| Identical Protein Groups | 79 |
| Taxonomy | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



