
Submitted:
16 June 2023
Posted:
20 June 2023
You are already at the latest version
Abstract
Torulaspora delbrueckii showed renewed interest in recent years in the fermentation of wine, for its biotechnological potential linked to the ability to enhance flavor and aroma and it probably is the non-Saccharomyces yeast currently widely used in winemaking. On the base of this, sequential fermentations with a selected native strain of T. delbrueckii (DiSVA 130) and low sulfite native strain of Saccharomyces cerevisiae (DiSVA 709) were carried out to establish their contribution in biocontrol and aroma profile. A first set trials, carried out in winery, were set up to establish the effect of the sulfur dioxide addition on pure and T. debrueckii/S. cerevisiae sequential fermentations. A second set of sequential fermentations without SO2 addition were conducted in the same conditions, to evaluate the biocontrol and aromatic effectiveness of the T. delbrueckii native strain and a commercial one. The effective biocontrol action of native T. delbrueckii inoculated in sequential fermentation was shown, indeed without SO2 addition the presence of native T. delbrueckii revealed an effective fungistatic action in the first two days of fermentation. Moreover, the native T. delbrueckii strain seems to have fermentative performances comparable to those of T. delbrueckii commercial strain showing a more evident biocontrol action (wild yeasts reduced by c.a. 1 Log at 2nd day) and its presence did not negatively affect S. cerevisiae fermentation activity. Finally, the combination of both native and commercial T. delbrueckii/S. cerevisiae trials led distinctive aromatic profile of wines with a significant enhancement of isoamyl acetate, phenyl ethyl acetate, supported by positive appreciations, from the tasters, for ripe and tropical fruits, citrus and balance. The whole results indicate that the proposed strain could be a potential biocontrol tool toward wild yeasts in the first phase of fermentation also contributing to improve and differentiate the final aroma wine.
Keywords:
wine yeasts
; Torulaspora delbrueckii
; low sulfites wines
; biocontrol
1. Introduction
In winemaking the use of selected cultures is a suitable strategy to control the fermentation process and improve organoleptic profiles and specific aroma compounds for the production of distinctive wines [1,2]. In this regard, the use of selected non-Saccharomyces yeasts under suitable conditions has widened the opportunities for enhancing the specific contribution of yeasts in winemaking. Indeed, their use in mixed and sequential fermentations with the starter Saccharomyces cerevisiae determined an enhancement of the organoleptic qualities of wine and the complexity of aromatic notes [3,4,5]. During the last few decades, many studies have been focused on the use of non-Saccharomyces yeasts during alcoholic fermentation for several variations of specific wine features as increase of glycerol [6] reduction of volatile acidity [7] enhancement of total acidity, production of polysaccharides [8] while others focused on the enhancement of flavor and aroma complexity [9,10,11] or ethanol reduction [12]. In addition to these features the use of non-Saccharomyces yeast has been proposed for biocontrol in winemaking. During the last few years there has been a trend in modern enology to decrease sulfites because of the affect the human health. Although the World Health Organization has recommended a daily dose of SO2 (RDA) of 0.7 mg SO2/kg of body weight, the European law has set the maximum concentrations allowed at 150 mg/L and 200 mg/L respectively in red and white wines (EU Regulation No. 606/2009). Moreover, environmental concerns led consumers to prefer “healthy” products and choose wines with lower levels of sulfites. In this perspective, the attention of winemakers was focused on the research based on new strategies to reduce the use of SO2 as chemical additive with a broad spectrum and widely used the winemaking process [13]. In this regard, in addition to chemical and physical strategies, the use of non-Saccharomyces yeasts could be a suitable and innovative strategy to achieve this goal with an improvement of the aroma profile of wine. Several studies have reported the bioprotectant activity of non-Saccharomyces which were found to be effective against spoilage wild microorganisms [14,15,16,17,18]. In particular, the presence of T. delbrueckii strain grape juice determined to a decrease in wild yeasts biodiversity if compared to the addition of sulphites [19].
Among the different non-Saccharomyces wine yeasts used in mixed fermentation with S. cerevisiae in winemaking, T. delbrueckii showed several features that positively affect the wine quality [20] and others that concern microbial interactions such as the production of active compounds (killer toxin and hydroxytyrosol). Effectively, with respect to the attributes required to perform industrial alcoholic fermentation, among the non-Saccharomyces yeasts, T. delbrueckii is the closest species to S. cerevisiae. This affinity could be probably the main reason why T. delbrueckii was the first non-Saccharomyces yeast suggested for winemaking use at industrial level.
For these reasons in this study it was evaluated the use of a selected strain of T. delbrueckii in sequential fermentation with native S. cerevisiae strain already selected [21] and tested [22] for low sulfite wine production. The aim was to evaluate the biocontrol and aroma-enhancing features of T. delbrueckii in organic wines using a low sulfite producer S. cerevisiae strain.
2. Materials and Methods
2.1. Yeast Strains
The native improved strain DiSVA 709, (Yeast Collection of the Department of Life and Environmental Sciences) [21] and the commercial starter strain, Lalvin ICV OKAY® (Lallemand Inc., Toulouse, France) were used as S. cerevisiae starter strain. Both yeast strains are characterized by the absence of H2S production and reduced production of SO2. The yeast strains used in the trials were cultivated and maintained on yeast extract–peptone–dextrose (YPD) agar medium (Oxoid, Basingstoke, U.K.) at 4 ◦C for short-term storage while for long-term storage YPD broth supplemented with 80% (w/v) glycerol at −80 ◦C was used.
2.2. Pilot fermentation Trials
The fermentation trials were carried out at Terre Cortesi Moncaro S.r.c.l. in steel vessels of 60 L containing 40 L of organic Verdicchio grape juice in duplicate under static conditions. The temperature was maintained at 18 °C. The grapes were the processed using the following procedures: soft hydraulic pressing and cold clarification at 8 ◦C for 2 days). The analytical characters of the grape juice were pH 3.22; initial sugar content 242 g/L; total acidity 4.48 g/L; malic acid 2.3 g/L; nitrogen content YAN (60 mg/L) and total SO2 14 mg/L. Yeast assimilable nitrogen adjusted to 250 mg N/L using diammonium phosphate and yeast derivative (Genesis Lift® Oenofrance, Bordeaux, France). The non-Saccharomyces strains, T. delbrueckii DiSVA 130, and T. delbrueckii commercial strain ALPHA® were used in the sequential fermentations after two days of the inoculum of S. cerevisiae starter strains (S. cerevisiae DiSVA 709 and Lalvin ICV OKAY®) in two sets of pilot fermentation trials carried out at winery level. The first set of trials were carried out with and without the addition 30 mg/l of SO2. before the inoculum of the starter strain. The other set of fermentation trials were conducted without SO2 added, evaluating pure and sequential fermentations using S. cerevisiae DiSVA 709 in sequential fermentation with T. delbrueckii DiSVA 130, and T. delbrueckii commercial strain ALPHA® (Laffort, Bordeaux, Cedex). The fermentations were monitored by measuring the sugar consumption.
Biomass production for the inoculation of the pilot fermentation trials was carried out as follows: the yeast strains preculture was grown under agitation for 48 h at 25 ◦C (150 rpm) in modified YPD medium (0.5% yeast extract, 0.1% peptone and 2% glucose). 5% (vol/vol) of this preculture was inoculated in a 30-L bioreactor (Biostat® C; B. Braun Biotech Int., Goettingen, Germany) containing 25 L of the same modified YPD medium using the following conditions: 400 rpm/min; air flow of 1 vvm (L/L/min). Yeast biomass production was in feed batch modality and the biomass was collected by centrifugation and washed three times with sterile distilled water. The inoculum of grape juice was carried out in cream form (80% humidity) at a concentration of approximately 1 × 106 cell/mL. The tracking of biomass was carried out using WL nutrient Agar medium (Oxoid, Hampshire, U.K.) and Lysine Agar medium (Oxoid, Hampshire, U.K.) [23] The sugar consumption, measured by Baumé (◦Bé) densimeter, was used to monitor the fermentation process.
2.3. Monitoring of yeast population
Biomass evolution was evaluated during the fermentation using viable cell count method. Lysine agar medium (Oxoid, Hampshire, UK) as selective medium for S. cerevisiae strains and WL nutrient agar medium (Oxoid, Hampshire, UK) as differential medium used for appreciation of form, color and diversity of wine yeast colonies. The detection of inoculated and wild yeasts in plates were performed after incubation at 25 °C for four days. The distinction between inoculated and wild yeasts was performed using lysine agar enumeration and macro- and micro-morphological estimation of colonies in WL nutrient agar medium. The presumptive identities of the yeasts were confirmed by sequencing using ITS 1 and 4 as target region. The primer pair ITS1 (50-TCCGTAGGTGAACCTCGCG-30) and ITS4 (50-TCCTCCGCTTTATTGATATGC-30) were used to amplify ITS1-5.8S rRNA-ITS2 region by PCR (Polymerase Chain Reaction) following the instructions of White and co-workers [24]. The sequences obtained were compared with that provided in GenBank database (http://www.ncbi.nlm.nih.gov/BLAST) . The inoculated S. cerevisiae strains and the presence of possible S. cerevisiae contaminant wild strains were assessed using intraspecies characterization of isolates with primer pairs δ 12/21 as described by Legras and Karst [25]. The length of the PCR products was estimated by comparing them with 100-bp marker DNA standards (GeneRuler 100-bp DNA Ladder; AB Fermentas).
2.4. Analytical Procedures
Total acidity, volatile acidity, pH, ethanol and total SO2 were analyzed following the procedures of the Official European Union Methods (EC Regulation No. 2870/00) [26]. Glucose and fructose (K-FRUGL), glycerol (K-GCROL), and malic acid (K-DMAL) were quantified using enzymatic kits (Megazyme International Ireland) according to the manufacturer instructions. A specific enzymatic kit (kit no. 112732; Roche Diagnostics, Germany) was used to determine the ammonium content. The free α-amino acids were evaluated following Dukes and Butzke protocol [27]. Ethyl acetate, acetaldehyde and higher alcohols were determined using a gas chromatograph system (GC-2014; Shimadzu, Kjoto, Japan) by direct injection. In the wines samples, set up following the procedures of Canonico et al. [26] , were determined the main volatile compounds using the solid-phase microextraction (HS-SPME) method [28]. The compounds were desorbed by inserting the fiber Divinylbenzene/Carboxen/Polydimethylsiloxane (DVB/CAR/PDMS) (Sigma-Aldrich, St. Louis, MO, USA) into a gas chromatograph (GC) injector.
2.5. Sensory Analysis
At the end of the fermentation, the wines after stabilization were bottled ( 750 mL ) with the crown cap and maintained at 4 ◦C until sensory analysis. After a storage of 3 months, wines were subjected to sensory analysis based on smell and taste. Sensory analysis was conducted by ten testers (80% expert and 20% non-expert), using a score scale from 1 to 9, for several descriptors (smell and taste) of each wine tested where 9 was the score of the best judgment while 1 was the score to be attribute in case of very poor satisfaction. The results were used to compare the wines and providing information regarding the organoleptic quality and feasible consumers’ satisfactoriness of the wines. The sensory analysis was carried out as follow: thirty milliliters of each wine were served at 22 ± 1 °C (room temperature) in glasses labeled with code and covered to prevent volatile loss. The presentation order was randomized among judges.
2.6. Statistical Analysis
Statistical Analysis of the analytical characters was conducted by analysis of variance (ANOVA) data of wine. The data were analyzed using the statistical software package JMP® 11. Duncan tests were used to detect the significant differences where the significant was associated with p-values < 0.05.
3. Results
3.1. First fermentation trial: evaluation of SO2 addition in T. delbrueckii/S. cerevisiae sequential fermentation
To evaluate the biocontrol effectiveness with and without the addition of SO2, sequential fermentation T. delbrueckii/S. cerevisiae was compared with the S. cerevisiae pure fermentations evaluating the biomass evolution, analytical and aromatic profile.
3.1.1. Biomass evolution and biocontrol activity
In Figure 1 are reported the results of the yeasts viable population during the inoculated fermentations with (on the left) and without (on the right) the addition of 30 mg/L of SO2. The S. cerevisiae commercial strain OKAY® (Fig. 1a) and selected strain DiSVA 709 (Fig. b) showed that the SO2 addition determined a completed control of wild yeast, while with the sequential inoculation (T. delbrueckii/S. cerevisiae) showed a limited growth (105 CFU/mL) of wild yeast at 2nd day that disappearing at 9th days. Without the addition of SO2 S. cerevisiae OKAY® controls wild yeasts that slightly growth (2nd days) and disappear at 9th day of fermentation. In the trial inoculated with S. cerevisiae DiSVA 709, wild yeasts exhibited a significant growth in the in the first two days compared with the S. cerevisiae OKAY® for disappearing only at the end of fermentation (Fig. 2a). The evolution of wild yeasts in sequential trial (T. delbrueckii / S. cerevisiae) (Fig.3b) exhibited a similar trend of S. cerevisiae OKAY® with a constant presence at 2nd day and disappearance at 9th day. The inoculated fermentation trials with S. cerevisiae starter strains (DiSVA 709 and OKAY®) with or without the addition of SO2 did not show relevant differences with a range of occurrence of 70-90% indicating an overall dominance.
3.1.2. Main analytical characters
In Table 1 are reported the main oenological characters of the pure and sequential fermentation trials with and without SO2addition. As expected, the addition of 30 mg/L did not exert influence on the main analytical characters of wines. Indeed, there were no relevant differences in the main analytical compounds with or without the addition of SO2. Among the trials with different inoculated starter strain, there was a reduction of final ethanol amount in the sequential fermentation T. delbrueckii DiSVA 130/S. cerevisiae DISVA 709 strain, particularly in comparison to that OKAY® commercial strain.
3.1.3. Main volatile compounds
The results of the influence sequential fermentation using T. delbrueckii/ S. cerevisiae DiSVA 709 in comparison with pure S. cerevisiae DiSVA 709 and commercial strain OKAY® fermentations on the volatile profile of wines are showed in Table 2.
The addition of SO2 did not show relevant influence on the production of volatile compounds, with the exception of ethyl acetate. Indeed, In both pure fermentation of S. cerevisiae (OKAY® and DiSVA 709) the absence of SO2 induce a production of ethyl acetate double or more if compared with the trial with SO2 added, while the presence of T. delbrueckii significantly reduced this increase Moreover, the sequential fermentation of T. delbrueckii/S. cerevisiae DiSVA 709 showed a significant enhancement of ethyl butyrate, β-phenyl ethanol and geraniol in the trials without SO2 added while, in the same condition, a significant increase in ethyl acetate was found in pure cultures of S. cerevisiae (DiSVA 709 and OKAY®). The sequential fermentation trials showed the highest amounts of higher alcohols (except for n-propanol) and isomyl acetate while there were no substantial differences in terpens production. The enological features of OKAY® strain were confirmed by Agarbati and collegues [21] with the highest production of ethyl butyrate and n-propanol and lower production of phenylethyl acetate.
3.2. Second fermentation trial: evaluation and comparison of native T. delbrueckii DiSVA 130 and commercial strain ALPHA® in sequential fermentation with S. cerevisiae DiSVA 709 (no SO2 added)
Results from the first set of fermentations established that sequential fermentations ensured effective wild yeast control in absence of sulfite. In this trials with no sulfite added, the native strain T.delbrueckii DiSVA 130 was compared with a commercial strain T. delbrueckii ALPHA® in combination with native S. cerevisiae DiSVA 709.
3.2.1. Biomass evolution and biocontrol action
In the Figure 2 are showed the the growth kinetics of the fermentation trials in a comparative assesment. The most relevant differences were found at 2nd day where the combination T. delbrueckii DiSVA 130/S.cerevisiae DiSVA 709 showed a slight reduction of initial wild yeast population of about 104 CFU/ml, while the other two fermentation trials showed an increase of a one log order. However, at 9th day the wild yeasts were completely disappeared in all trials. In the second fermentation set the S. cerevisiae inoculated strain showed a dominance toward wild S. cerevisiae strains with 70%, 75 %and 80% of occurrence for T. delbrueckii ALPHA® / S. cerevisiae DiSVA 709, T. delbrueckii DiSVA 130/ S. cerevisiae DiSVA 709 and S. cerevisiae DiSVA 709 pure fermentation respectively.
3.2.2. Main oenological characters
As showed in Table 3 sequential fermentation trials did not differ among them while pure fermentation of S.cerevisiae DiSVA 709 showed a significant higher ethanol production.
3.2.3. Main oenological volatile compounds
Regarding to the main volatile compounds (Table 4), significant differences were shown. Both sequential fermentations showed significant higher amounts of esters and higher alcohols (with the exclusion of isoamylic alcohol) and terpens, in comparison with pure fermentation. Sequential fermentation of T. delbrueckii DiSVA 130 / S. cerevisiae DiSVA 709 was characterized for ethyl acetate, isoamyl acetate, isobutanol amyl alcohol, linalool and nerol while T. delbrueckii ALPHA® / S. cerevisiae DiSVA 709 exhibited significant higher amounts of ethyl butyrate, ethyl hexanoate isoamyl acetate, β-phenyl ethanol, phenyl ethyl acetate linalool and geraniol. However, the level of terpens were lower the threshold values in wines for these compounds.
3.3. Sensory analysis of Verdicchio wines inoculated with S. cerevisiae DiSVA 709, T. delbrueckii DiSVA 130/S. cerevisiae DiSVA 709 and T. delbrueckii ALPHA® /S. cerevisiae DiSVA 709
Wines obtained by the second fermentation trials (without SO2 added) were evaluated by sensory analysis to establish the role and the influence of T. delbrueckii in aroma features and complexity. Results reported in Figure 3 showed a general appreciation by the tasters, each distinguished by distinctive aromatic notes and without defects. Wines obtained with T. delbrueckii/S. cerevisiae DiSVA709 were perceived more balanced with relevant fruitiness (ripe tropical and citrus). With the same trend but with the lower score were perceived T. delbrueckii/S. cerevisiae ALPHA® and S. cerevisiae DiSVA 709, respectively. These results fit well with the outcomes of some volatile compounds as acetate esters.
4. Discussion
The renewed interest of non-Saccharomyces yeasts has led to the industrial production of selected cultures for winemaking. Currently, T. delbrueckii was first non-Saccharomyces species produced for this purpose, and the most currently commercially available active dry yeasts.
The ability of S. cerevisiae to overcome other non-Saccharomyces yeasts and to dominate wine fermentation is undoubted. Moreover, T. delbrueckii has a general less fermentation vigor and a slower growth rate than S. cerevisiae under usual wine fermentation conditions [10,12], and this behavior may suggest a difficulty in dominating must fermentation in the presence of inoculated S. cerevisiae yeasts [31]. However, under the conditions tested (two days sequential inoculation), the T. delbrueckii DiSVA 130 strain did not seem to be affect from the presence of S. cerevisiae DiSVA 709.
The results of the fermentation kinetics agreed with previous studies [7,32], which reported a lower ethanol production in the trials where the musts were inoculated with T. delbrueckii yeast, although no statistical differences were seen between commercial T. delbrueckii and selected native strain.
Regarding to the biocontrol action Simonin et al. [19] reported the results of bio-protectant and antioxidant effects of T.delbrueckii, inoculated at the beginning of the white winemaking process while Chacon-Rodriguez, et al. [15] showed a biocontrol action of a blend of T. delbrueckii and Metschnikowia pulcherrima applied to a machine harvester as compared to standard addition of SO2 in Cabernet Sauvignon variety. In agreement with Simonin et al. [19] the addition of T. delbrueckii DiSVA 130 showed a control of wild yeasts during the first two days of fermentation, also if slightly lower if compared with sulfites fermentation control trial. However, T. delbrueckii DiSVA 130 effectively limited the development of wild yeasts demonstrating its effectiveness to protect must.
The impact of T. delbrueckii on the fermentation and aroma enhancement has been documented during the years [33,34]. A lot of studies showed the positive contribution of T. delbrueckii strains and their relative positive impact on wine quality [9,20,35]. This non-Saccharomyces yeast is recommended for the fermentation of both dry and high sugar grapes for the low production of acetic acid. Azzolini and coworkers [36] already demonstrated that multi-starter fermentation with T. delbrueckii greatly affected the content of several important volatile compounds, including ß-phenyl ethanol, isoamyl acetate, fatty acid esters, C4–C10 fatty acids and vinyl phenols. Ramirez and Velazquez [31] analyzed the variable behavior of T. delbrueckii considering the strain’s differences, and wine varieties with special emphasis on the proposals for industrial uses of this species.
The production of esters by T. delbrueckii might be strain-dependent and it has extensively demonstrated that this production is further modified in the presence of S. cerevisiae during multiple fermentations [9,37]. This could explain some results obtained in this work, concerning the volatile compounds phenyl ethyl acetate and ß-phenyl ethyl ethanol which typically increase in the presence of T. delbrueckii. In the first set of trials conducted with and without sulfur dioxide, T. delbrueckii DiSVA 130/ S. cerevisiae DiSVA 709 trials showed phenyl ethyl acetate only a slight increase, while isoamyl acetate and phenyl ethyl ethanol increased significantly only without sulfur dioxide. In the second set of the trials, both T. delbrueckii DiSVA 130 and the commercial strain ALPHA® in sequential fermentation with S. cerevisiae DiSVA 709 determined a 10-fold increase in phenyl ethyl acetate compared with pure S. cerevisiae fermentation. On the other hand, in the condition tested, β-phenyl ethyl ethanol increased only slightly in the T. delbrueckii ALPHA®/S.cerevisiae DiSVA 709 fermentation while the presence of T. delbrueckii DiSVA 130 did not cause any increase. On the other hand, in agreement with Sun et al. [29], both sequential fermentations using T. delbrueckii DiSVA 130 and ALPHA® revealed significant enhancement of ethyl acetate, phenyl ethyl acetate while the amounts of terpens were lower the thresholds values.
The overall analytical profile of wines did not show any defect in presence of T. delbrueckii showing, on the contrary, some differences in esters and higher alcohols and the sensory evaluation highlighted the effective positive contribution of this non-Saccharomyces yeasts, particularly the native strain T. delbrueckii DiSVA 130, which imparts notes of tropical fruit, citrus and ripe fruit and greater balance to the wine.
The overall results indicated the multiple roles of T. delbrueckii in winemaking since the selected DiSVA 130 strain showed an effective biocontrol action in sequential fermentation of Verdicchio wine in absence of SO2 addition. At the same time, this fermentation modality gave a distinctive and aromatic imprint to the wine as corroborated by the sensory analysis.
A negative perception developed by consumers towards sulfites in wine, because of health and environmental concerns is the new trend in winemaking market. For this, there are increasing demand for health benefits wines, and with low SO2 content, that push winemakers toward strains with tailored characteristics.
In this research, the efficacy of T. delbrueckii DiSVA 130 in sequential fermentation with native S. cerevisiae DiSVA 709 evidenced a biocontrol activity in the absence of SO2, revealing a synergistic effect of two native strains to impart distinctive aromatic notes.
Author Contributions
Laura Canonico, Alice Agarbati, Edoardo Galli, Francesca Comitini and Maurizio Ciani contributed equally to the manuscript. All the authors participated in the design and discussion of the research. Laura Canonico, Edoardo Galli and Alice Agarbati carried out the experimental part of the work and carried out the analysis of the data. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by PSR measure 16.1.A.2; ID project 28779"(2019-2021) "Innovative strategies in the wine production chain to protect the environment and consumer health” “Vitinnova"
Data Availability Statement
Not applicable.
Acknowledgments
The authors wish to thank the winery Terre Cortesi Moncaro s.r.c.l. for availability and support in the winery trials G. D’Ignazi, G. Mazzoni V. Durastanti, and T. Duca for their technical assistance.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pretorius, I.S. Tailoring wine yeast for the new millennium: novel approaches to the ancient art of winemaking. Yeast 2000, 16, 675–729. [Google Scholar] [CrossRef] [PubMed]
- Comitini, F.; Capece, A.; Ciani, M.; Romano, P. New insights on the use of wine Yeasts. Current Opinion in Food Science 2017, 13, 44–49. [Google Scholar] [CrossRef]
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not your ordinary yeast: non-Saccharomyces yeasts in wine production uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef] [PubMed]
- Del Fresno, J.M.; Morata, A.; Loira, I.; Bañuelos, M.A.; Escott, C.; Benito, S.; González Chamorro, C.; Suárez-Lepe, J.A. Use of non-Saccharomyces in single-culture, mixed and sequential fermentation to improve red wine quality. Eur. Food Res. Technol. 2017, 243, 2175–2185. [Google Scholar] [CrossRef]
- Varela, C. The Impact of non-Saccharomyces yeasts in the production of alcoholic beverages. Appl Microbiol Biotechnol 2016, 100, 9861–9874. [Google Scholar] [CrossRef]
- Ciani, M.; Ferraro, L. Combined use of immobilized Candida stellata cells and Saccharomyces cerevisiae to improve the quality of wines. Journal of Applied Microbiology 1998, 85, 247–254. [Google Scholar] [CrossRef]
- Bely, M.; Stoeckle, P.; Masneuf-Pomarède, I.; Dubourdieu, D. Impact of mixed Torulaspora delbrueckii–Saccharomyces cerevisiae culture on high-sugar fermentation. International Journal of Food Microbiology 2008, 122, 312–320. [Google Scholar] [CrossRef]
- Gobbi, M.; Comitini, F.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Lachancea thermotolerans and Saccharomyces cerevisiae in simultaneous and sequential co-fermentation: a strategy to enhance acidity and improve the overall quality of wine. Food Microbiology 2013, 33, 271–281. [Google Scholar] [CrossRef]
- Renault, P.; Coulon, J.; Moine, V.; Thibon, C.; Bely, M. Enhanced 3-sulfanylhexan-1-ol production in sequential mixed fermentation with Torulaspora delbrueckii/Saccharomyces cerevisiae reveals a situation of synergistic interaction between two industrial strains. Frontiers in Microbiology 2016, 7. [Google Scholar] [CrossRef]
- Comitini, F.; Gobbi, M.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Selected non-Saccharomyces wine yeasts in controlled multistarter fermentations with Saccharomyces cerevisiae. Food Microbiology 2011, 28, 873–882. [Google Scholar] [CrossRef]
- Zhang, B.; Tang, C.; Yang, D.; Liu, H.; Xue, J.; Duan, C.; Yan, G. Effects of three indigenous non-Saccharomyces yeasts and their pairwise combinations in co-fermentation with Saccharomyces cerevisiae on volatile compounds of petit manseng wines. Food Chemistry 2022, 368, 130807. [Google Scholar] [CrossRef]
- Ciani, M.; Morales, P.; Comitini, F.; Tronchoni, J.; Canonico, L.; Curiel, J.A.; Oro, L.; Rodrigues, A.J.; Gonzalez, R. Non-conventional yeast species for lowering ethanol content of wines. Frontiers in Microbiology 2016, 7. [Google Scholar] [CrossRef]
- Divol, B.; du Toit, M.; Duckitt, E. Surviving in the presence of sulphur dioxide: strategies developed by wine yeasts. Appl Microbiol Biotechnol 2012, 95, 601–613. [Google Scholar] [CrossRef] [PubMed]
- Escribano-Viana, R.; González-Arenzana, L.; Garijo, P.; Fernández, L.; López, R.; Santamaría, P.; Gutiérrez, A.R. Bioprotective Effect of a Torulaspora delbrueckii/Lachancea thermotolerans-mixed inoculum in red winemaking. Fermentation 2022, 8, 337. [Google Scholar] [CrossRef]
- Chacon-Rodriguez, L.; Joseph, C.M.L.; Nazaris, B.; Coulon, J.; Richardson, S.; Dycus, D.A. Innovative use of non- Saccharomyces in bio-protection: T. Delbrueckii and M. Pulcherrima Applied to a Machine Harvester. Catalyst: Discovery into Practice 2020, 4, 82–90. [Google Scholar] [CrossRef]
- Morata, A.; Loira, I.; González, C.; Escott, C. Non-Saccharomyces as biotools to control the production of off-flavors in wines. Molecules 2021, 26, 4571. [Google Scholar] [CrossRef]
- Agarbati, A.; Canonico, L.; Ciani, M.; Comitini, F. Metschnikowia pulcherrima in cold clarification: biocontrol activity and aroma enhancement in Verdicchio wine. Fermentation 2023, 9, 302. [Google Scholar] [CrossRef]
- Canonico, L.; Agarbati, A.; Galli, E.; Comitini, F.; Ciani, M. Metschnikowia pulcherrima as biocontrol agent and wine aroma enhancer in combination with a native Saccharomyces cerevisiae. LWT 2023, 181, 114758. [Google Scholar] [CrossRef]
- Simonin, S.; Alexandre, H.; Nikolantonaki, M.; Coelho, C.; Tourdot-Maréchal, R. Inoculation of Torulaspora delbrueckii as a bio-protection agent in winemaking. Food Research International 2018, 107, 451–461. [Google Scholar] [CrossRef]
- Benito, S. The impact of Torulaspora delbrueckii yeast in winemaking. Appl Microbiol Biotechnol 2018, 102, 3081–3094. [Google Scholar] [CrossRef]
- Agarbati, A.; Canonico, L.; Comitini, F.; Ciani, M. Improved Saccharomyces cerevisiae strain in pure and sequential fermentation with Torulaspora delbrueckii for the production of verdicchio wine with reduced sulfites. Applied Sciences 2020, 10, 6722. [Google Scholar] [CrossRef]
- Comitini, F.; Agarbati, A.; Canonico, L.; Ciani, M. Yeast interactions and molecular mechanisms in wine fermentation: a comprehensive review. IJMS 2021, 22, 7754. [Google Scholar] [CrossRef] [PubMed]
- Oro, L.; Canonico, L.; Marinelli, V.; Ciani, M.; Comitini, F. Occurrence of Brettanomyces bruxellensis on grape berries and in related winemaking cellar. Frontiers in Microbiology 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- White, C. i.; Haber, J. e. Intermediates of recombination during mating type switching in Saccharomyces cerevisiae. The EMBO Journal 1990, 9, 663–673. [Google Scholar] [CrossRef]
- Legras, J.-L.; Karst, F. Optimisation of Interdelta Analysis for Saccharomyces cerevisiae strain characterisation. FEMS Microbiology Letters 2003, 221, 249–255. [Google Scholar] [CrossRef]
- Canonico, L.; Comitini, F.; Ciani, M. Influence of vintage and selected starter on Torulaspora delbrueckii/Saccharomyces cerevisiae Sequential Fermentation. Eur Food Res Technol 2015, 241, 827–833. [Google Scholar] [CrossRef]
- Dukes, B.C.; Butzke, C.E. Rapid Determination of primary amino acids in grape juice using an o-phthaldialdehyde/n-acetyl-l-cysteine spectrophotometric assay. Am J Enol Vitic. 1998, 49, 125–134. [Google Scholar] [CrossRef]
- Canonico, L.; Solomon, M.; Comitini, F.; Ciani, M.; Varela, C. Volatile profile of reduced alcohol wines fermented with selected non-Saccharomyces yeasts under different aeration conditions. Food Microbiology 2019, 84, 103247. [Google Scholar] [CrossRef]
- Sun, S.Y.; Jiang, W.G.; Zhao, Y.P. Evaluation of different Saccharomyces cerevisiae strains on the profile of volatile compounds and polyphenols in cherry wines. food chemistry 2011, 127, 547–555. [Google Scholar] [CrossRef]
- Gambetta, J.M.; Bastian, S.E.P.; Cozzolino, D.; Jeffery, D.W. Factors influencing the aroma composition of chardonnay wines. J. Agric. Food Chem. 2014, 62, 6512–6534. [Google Scholar] [CrossRef]
- Ramírez, M.; Velázquez, R. The yeast Torulaspora delbrueckii: an interesting but difficult-to-use tool for winemaking. Fermentation 2018, 4, 94. [Google Scholar] [CrossRef]
- Cabrera, M.J.; Moreno, J.; Ortega, J.M.; Medina, M. Formation of ethanol, higher alcohols, esters, and terpenes by five yeast strains in musts from Pedro Ximénez grapes in various degrees of ripeness. Am J Enol Vitic. 1988, 39, 283–287. [Google Scholar] [CrossRef]
- Takush, D. g.; Osborne, J. p. Impact of yeast on the aroma and flavour of Oregon pinot noir wine. Australian Journal of Grape and Wine Research 2012, 18, 131–137. [Google Scholar] [CrossRef]
- van Breda, V.; Jolly, N.; van Wyk, J. Characterisation of commercial and natural Torulaspora delbrueckii wine yeast strains. International Journal of Food Microbiology 2013, 163, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Liu, H.; Xue, J.; Tang, C.; Duan, C.; Yan, G. Use of Torulaspora delbrueckii and Hanseniaspora vineae co-fermentation with Saccharomyces cerevisiae to Improve aroma profiles and safety quality of Petit Manseng wines. LWT 2022, 161, 113360. [Google Scholar] [CrossRef]
- Azzolini, M.; Tosi, E.; Lorenzini, M.; Finato, F.; Zapparoli, G. Contribution to the aroma of white wines by controlled Torulaspora delbrueckii cultures in association with Saccharomyces cerevisiae. World J Microbiol Biotechnol 2015, 31, 277–293. [Google Scholar] [CrossRef]
- Renault, P.; Coulon, J.; de Revel, G.; Barbe, J.-C.; Bely, M. Increase of fruity aroma during mixed T. Delbrueckii/S. cerevisiae Wine fermentation is linked to specific esters enhancement. International Journal of Food Microbiology 2015, 207, 40–48. [Google Scholar] [CrossRef]
Figure 1.
Growth kinetics in sequential fermentation trials of T. delbrueckii DiSVA 130 ( )/ S. cerevisiae DiSVA 709 (
)/ S. cerevisiae DiSVA 709 ( ) and wild yeasts (
) and wild yeasts ( ) on natural grape juice with (a) and without (b) SO2.
) on natural grape juice with (a) and without (b) SO2.
)/ S. cerevisiae DiSVA 709 () and wild yeasts () on natural grape juice with (a) and without (b) SO2.
Figure 1.
Growth kinetics in sequential fermentation trials of T. delbrueckii DiSVA 130 ()/ S. cerevisiae DiSVA 709 () and wild yeasts () on natural grape juice with (a) and without (b) SO2.
)/ S. cerevisiae DiSVA 709 () and wild yeasts () on natural grape juice with (a) and without (b) SO2.
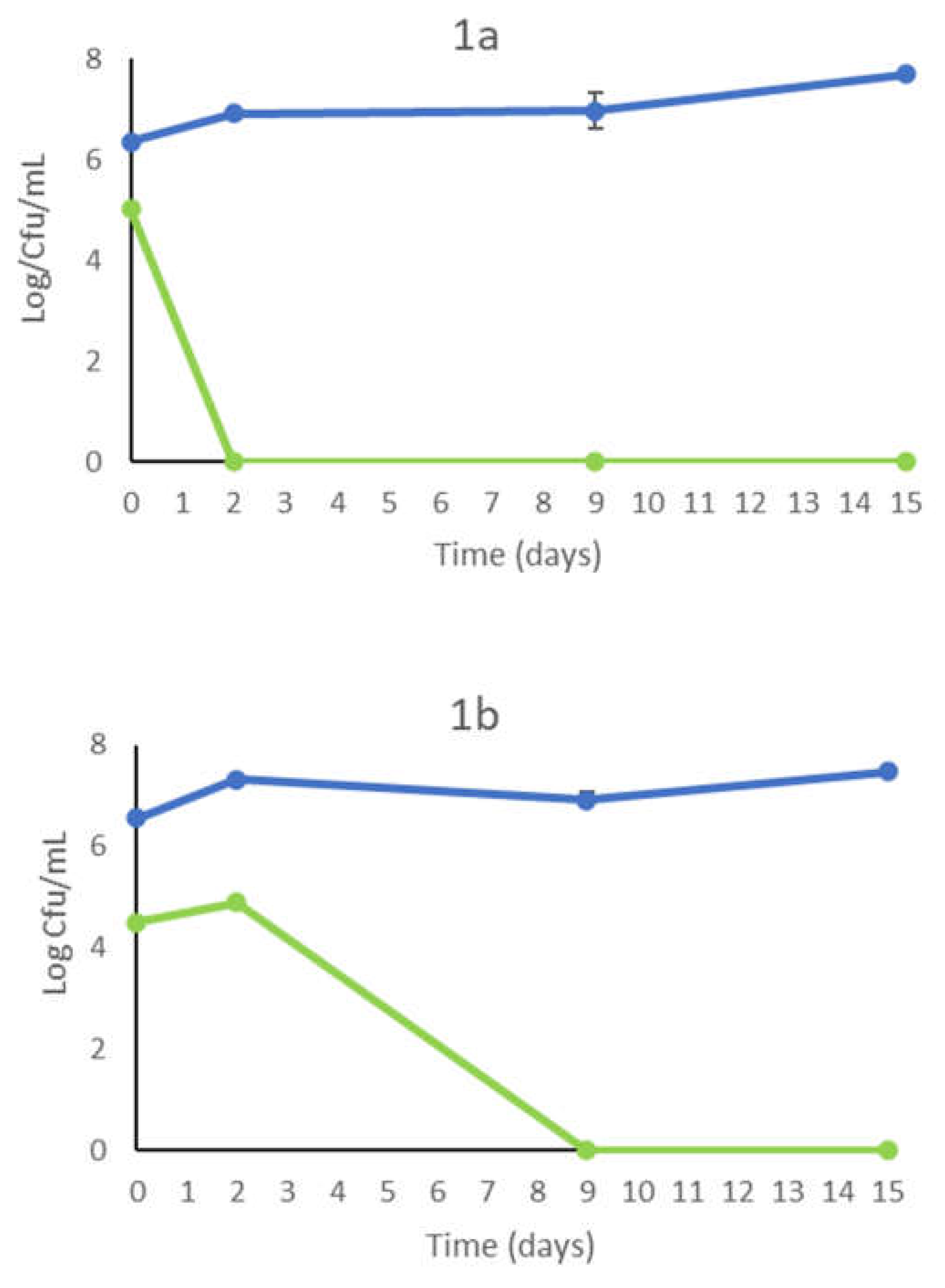
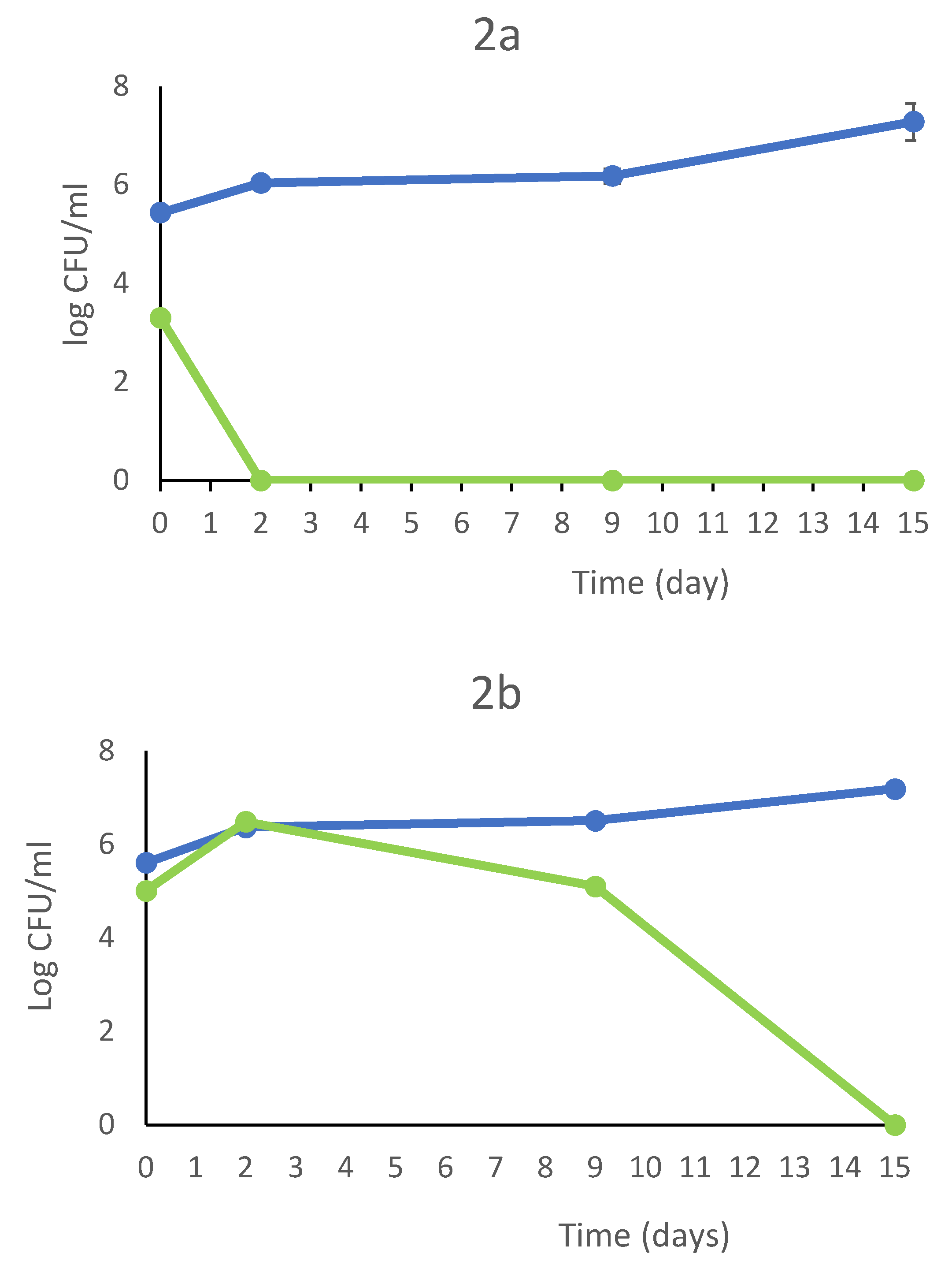
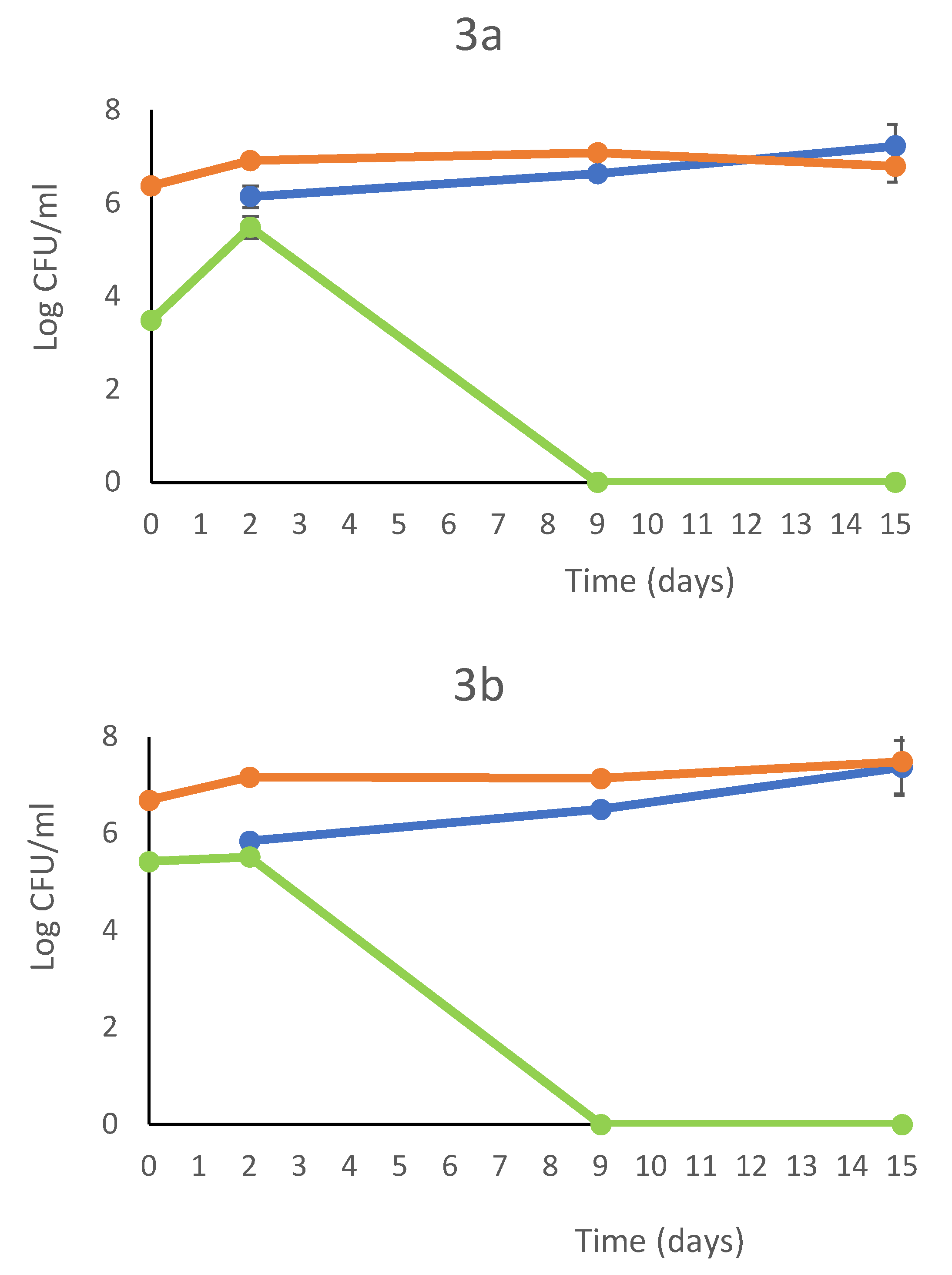
Figure 2.
Growth kinetics and bicontrol action of S. cerevisiae DiSVA 709 in pure culture (A), in sequential fermentation with T. delbrueckii strain DiSVA 130 (B) e commercial strain ALPHA® (C) (without SO2 addition). S. cerevisiae DiSVA 709 ( ), wild yeasts (
), wild yeasts ( )T. delbrueckii (
)T. delbrueckii ( ) on Verdicchio grape juice
) on Verdicchio grape juice
), wild yeasts ()T. delbrueckii () on Verdicchio grape juice
Figure 2.
Growth kinetics and bicontrol action of S. cerevisiae DiSVA 709 in pure culture (A), in sequential fermentation with T. delbrueckii strain DiSVA 130 (B) e commercial strain ALPHA® (C) (without SO2 addition). S. cerevisiae DiSVA 709 (), wild yeasts ()T. delbrueckii () on Verdicchio grape juice
), wild yeasts ()T. delbrueckii () on Verdicchio grape juice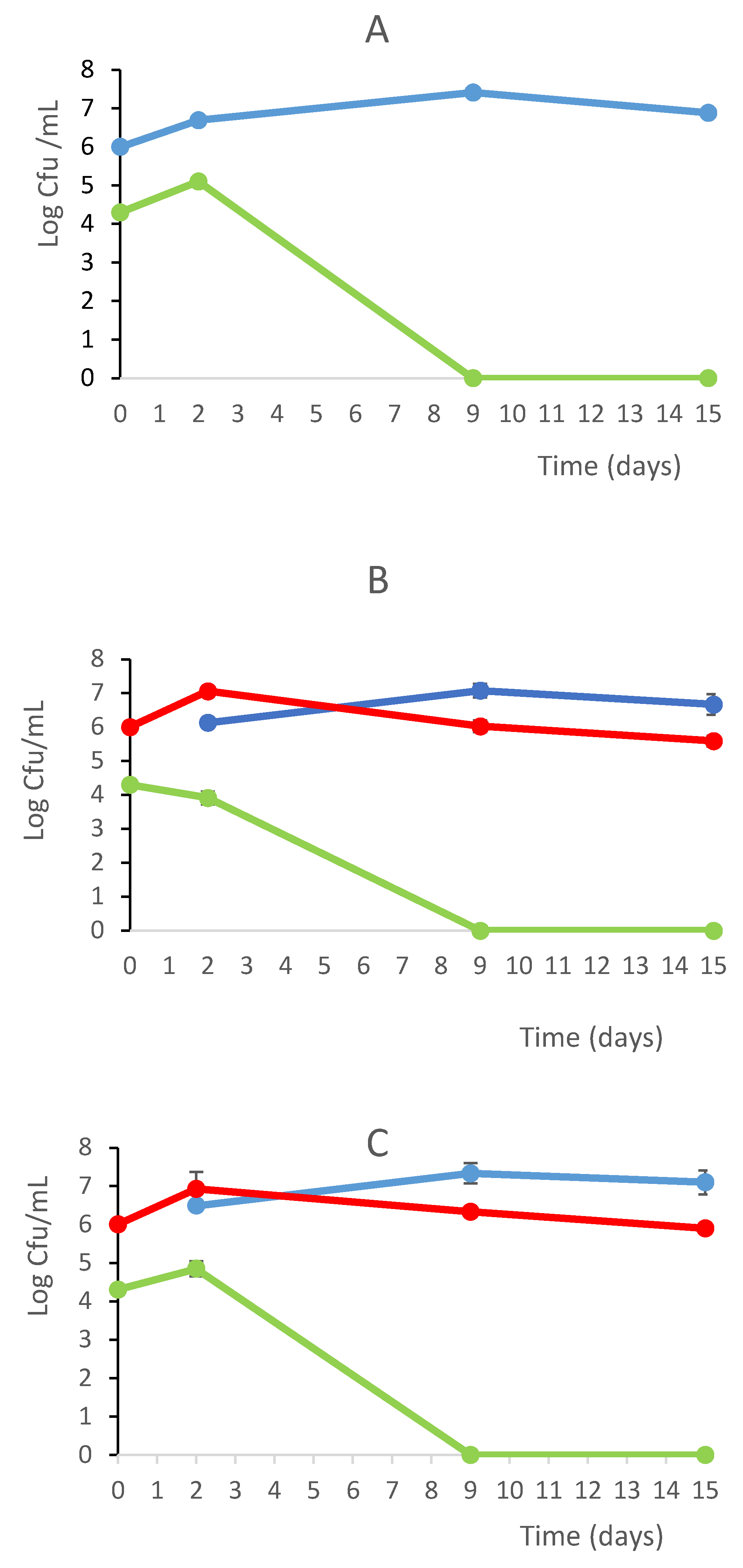
Figure 3.
Sensory analysis of Verdicchio wines inoculated with: S. cerevisiae DiSVA 709 ( ); T. delbrueckii DiSVA 130 / S. cerevisiae DiSVA 709 (
); T. delbrueckii DiSVA 130 / S. cerevisiae DiSVA 709 ( ); T. delbrueckii ALPHA® / S. cerevisiae DiSVA 709 (
); T. delbrueckii ALPHA® / S. cerevisiae DiSVA 709 ( ); * Significant difference at P=0.05.
); * Significant difference at P=0.05.
); T. delbrueckii DiSVA 130 / S. cerevisiae DiSVA 709 (); T. delbrueckii ALPHA® / S. cerevisiae DiSVA 709 (); * Significant difference at P=0.05.
Figure 3.
Sensory analysis of Verdicchio wines inoculated with: S. cerevisiae DiSVA 709 (); T. delbrueckii DiSVA 130 / S. cerevisiae DiSVA 709 (); T. delbrueckii ALPHA® / S. cerevisiae DiSVA 709 (); * Significant difference at P=0.05.
); T. delbrueckii DiSVA 130 / S. cerevisiae DiSVA 709 (); T. delbrueckii ALPHA® / S. cerevisiae DiSVA 709 (); * Significant difference at P=0.05.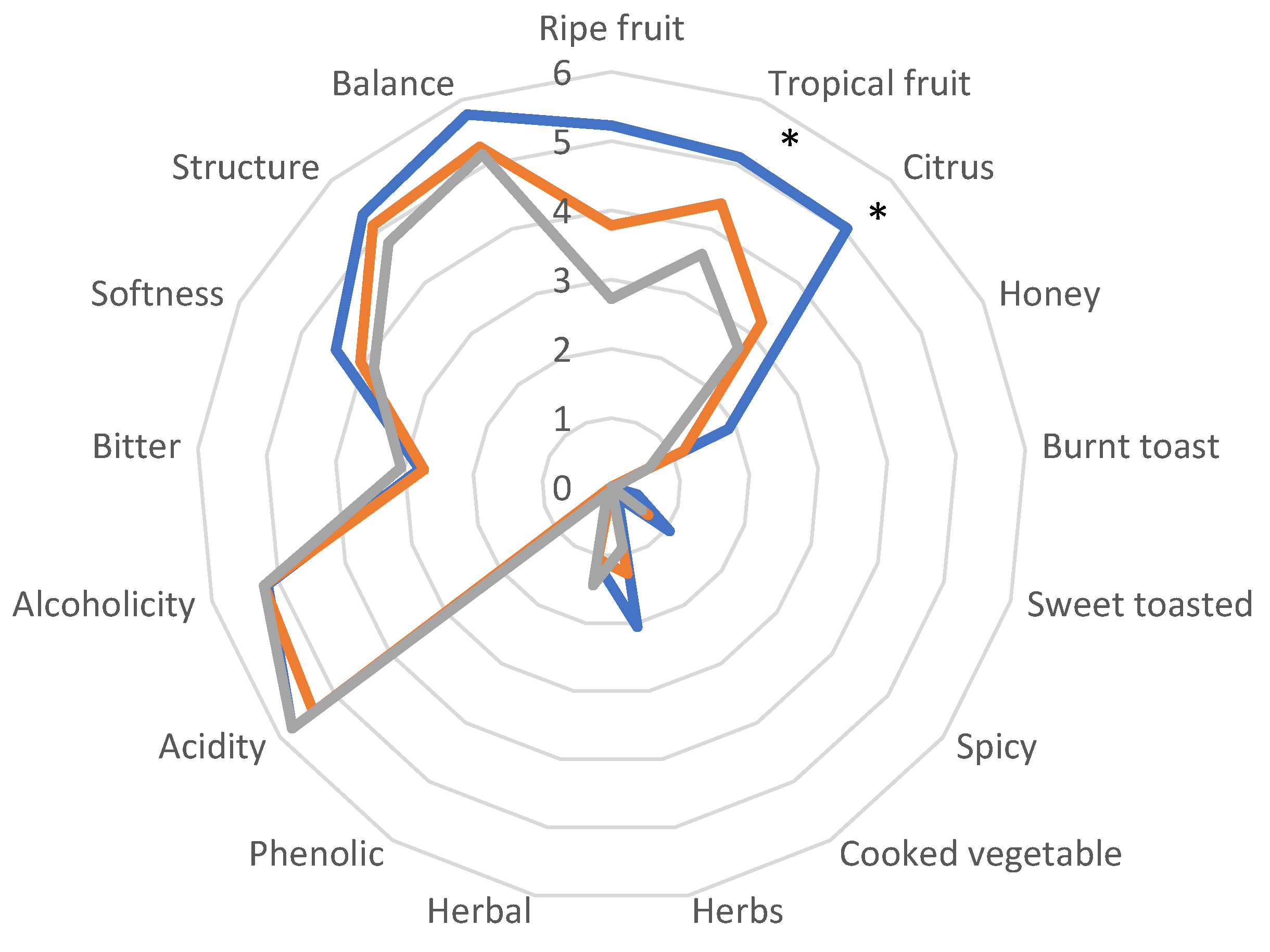

Table 1.
Main oenological characters of T. delbrueckii in sequential fermentation with and without SO2 added as compared with pure S. cerevisiae starter strains.
Table 1.
Main oenological characters of T. delbrueckii in sequential fermentation with and without SO2 added as compared with pure S. cerevisiae starter strains.
| Inoculated strains | Ethanol (%v/v) |
Total acidity (g/L) |
Volatile acidity (g /L) |
Malic acid (g /L) |
|---|---|---|---|---|
| S. cerevisiae OKAY® +SO2 | 14.69±0.01a | 5.685±0.02c | 0.30±0.00a | 0.9±0.00b |
| S. cerevisiae OKAY® | 14.88± 0.02a | 5.27±0,01c | 0.31±0.014a | 0.7±0.00b |
| S. cerevisiae DiSVA 709 +SO2 | 14.05±0.12b | 6.29±0.05a | 0.225±0.007a | 1.25±0.07a |
| S. cerevisiae DiSVA 709 | 14.35±0.1b | 5.77±0.00ab | 0.305±0.007a | 1± 0.00ab |
| T. delbrueckii DiSVA 130 / S. cerevisiae DiSVA 709 +SO2 | 13.9±0.02c | 6.14±0.14a | 0.25±0.00a | 1.35±0.07b |
| T. delbrueckii DiSVA 130 / S. cerevisiae DiSVA 709 | 13.86±0.09c | 5.355±0.03c | 0.29±0.00a | 0.45±0.07b |
Table 2.
The main volatile compounds of T. delbrueckii in sequential fermentation as compared with pure fermentation of S. cerevisiae starter strain (mg/L). In braked are reported the thresholds value (mg/L)*.
Table 2.
The main volatile compounds of T. delbrueckii in sequential fermentation as compared with pure fermentation of S. cerevisiae starter strain (mg/L). In braked are reported the thresholds value (mg/L)*.
| OKAY® + SO2 |
OKAY® | S. cerevisiae DiSVA 709 +SO2 | S. cerevisiae DiSVA 709 | T. delbrueckii DiSVA 130 /S. cerevisiae DiSVA 709 + SO2 | T. delbrueckii DiSVA 130 / S. cerevisiae DiSVA 709 | |
| ESTERS | ||||||
| Ethyl butyrate (0.02) |
1.214±0.021b | 1.491±0.075a | 0.653±0.223c | 0.628±0.035c | 0.208±0.011d | 0.441±0.084cd |
| Ethyl acetate (7.50 |
19.71±2.17b | 36.35±1.61a | 10.439±0.68b | 38.201±0.86a | 14.18±1.06b | 19.75±0.70b |
| Ethyl exanoate (0.014) |
1.063±0.2354a | 0.16±0.0017b | 0.253±0.0974b | 0.191±0.0110b | 0.081±0.0149b | 0.161±0.0464b |
| Isoamyl acetate (0.03) |
0.947±0.042b | 0.852±0.034b | 1.357±0.462b | 1.095±0.026b | 1.425±0.134b | 3.331±0.375a |
| Phenyl ethyl acetate (0.25) |
0.31±0.01 b | 0.27±0.04 b | 0.64±0.01 a | 0.76±0.04 a | 0.63±0.13 a | 0.79±0.19 a |
| ALCOHOLS | ||||||
| n-propanol (9.0) |
86.630±0.94a | 94.148±1.51a | 39.655± 0.26b | 37.032±2.64b | 27.904±0.71b | 35.734±2.10b |
| Isobutanol (40.0) |
17.51± 0.18b | 12.56±0.63b | 10.957± 2.02b | 19.211±0.52b | 32.634± 0.04a | 25.211± 0.85a |
| Amyl alcohol 1(2.2) |
12.601±2.27b | 12.245±1.51c | 19.211±0.51c | 14.909± 0.08b | 20.74±1.50a | 25.690±0.92a |
| Isoamilic alcohol (30.0) |
132.53±2.18b | 145.105±1.57 b | 137.156±0.99 b | 125.50± 0.13b | 171.56± 2.71a | 192.248±1.68a |
| β-Phenyl ethanol (14.0) |
13.93±0.091bcd | 10.05±0.174cd | 18.90±0.027ab | 15.83±0.212bc | 8.02±0.198d | 25.42±0.649a |
| CARBONYL COMPOUNDS | ||||||
| Acetaldehyde (0.5) |
6.40±2.832ab | 3.23±0.146b | 8.22±0.315a | 7.70±1.855a | 4.79±0.506ab | 3.02±0.678b |
| TERPENS | ||||||
| Linalool (0.025) |
0.197±0.0792a | 0.125±0.0636a | 0.153±0.1189a | 0.186±0.1320a | 0.078±0.0144a | 0.128±0.0119a |
| Geraniol (0.030) |
0.009±0.0004abc | 0.007±0.0028bc | 0.016±0.0029a | 0.013±0.0039ab | 0.003±0.0036c | 0.013±0.0046ab |
| Nerol (0.025) |
0.006±0.0036ab | 0.004±0.0059ab | ND** | 0.009±0.0019 ab. | 0.011±0.0045a | 0.008±0.0022ab |
Table 3.
The main analytical characters of pure fermentation of S. cerevisiae DiSVA 709 and sequential fermentation with T. delbrueckii DiSVA 130 and ALPHA® commercial strain.
Table 3.
The main analytical characters of pure fermentation of S. cerevisiae DiSVA 709 and sequential fermentation with T. delbrueckii DiSVA 130 and ALPHA® commercial strain.
| Ethanol (%v/v) |
Total acidity (g /L) |
Volatile acidity (g/ L) |
Malic acid (g/ L) |
|
|---|---|---|---|---|
| S. cerevisiae DiSVA 709 | 14.43±0.00a | 5.52±0.02a | 0.25±0.063a | 1.2±0.00a |
| T. delbrueckii DiSVA 130 /S. cerevisiae DiSVA 709 | 13.77± 0.10b | 5.55±0.14a | 0.23±0.007a | 1.2±0.14a |
| T. delbrueckii ALPHA®/ S. cerevisiae DiSVA 709 | 13.71±0.02b | 5.52±0.06a | 0.25±0.028a | 1.1±0.00a |
Table 4.
The main volatile compounds of pure fermentation of S. cerevisiae DiSVA 709 and sequential fermentation with T. delbrueckii DiSVA 130 and ALPHA® commercial strain (mg/L). In braked thresholds are reported (mg/L).
Table 4.
The main volatile compounds of pure fermentation of S. cerevisiae DiSVA 709 and sequential fermentation with T. delbrueckii DiSVA 130 and ALPHA® commercial strain (mg/L). In braked thresholds are reported (mg/L).
|
S. cerevisiae DiSVA 709 mg/L |
T. delbrueckii DiSVA 130 / S. cerevisiae DiSVA 709 mg/L | T. delbrueckii ALPHA® / S. cerevisiae DiSVA 709 mg/L | |
| ESTERS | |||
| Ethyl butyrate (0.02) |
0.40±0.10b | 0.31 ± 0.01c | 0.52±0.19a |
| Ethyl acetate (7.50) |
26.42±4.29b | 59.88±2.14a | 33.67±6.71b |
| Ethyl exanoate (0.014) |
2.76±0.33b | 2.90±0.41ab | 3.39±0.35a |
| Isoamyl acetate (0.03) |
0.90±0.01b | 0.95±0.08a | 1.03±0.04a |
| Phenylethyl acetate (0.25) |
0.08±0.01c | 0.74±0.16b | 0.98±0.08a |
| ALCOHOLS | |||
| n-propanol (9.0) |
37.01±3.09a | 39.25±1.40a | 38.73±0.79a |
| Isobutanol (40.0) |
15.38±1.72b | 26.67±5.20a | 11.92±3.30c |
| Amyl alcohol (12.2) |
12.99±0.26b | 39.76±8.28a | 14.33±3.77b |
| Isoamylic alcohol (30.0) |
123.34±8.0a | 67.11±5.45b | 126.30±2.7a |
| β-Phenyl ethanol (14.0) |
7.45±0.01b | 7.40±0.16b | 9.10±0.02a |
| CARBONYL COMPOUNDS | |||
| Acetaldehyde (0.50) |
19.23±0.50a | 14.25±0.27b | 14.23±2.87b |
| MONOTERPENS | |||
| Linalool (0.025) |
0.03±000b | 0.20±0.07a | 0.22±0.14a |
| Geraniol (0.030) |
0±0.000c | 0.006±0.00a | 0.003±0.00b |
| Nerol (0.025) |
0.003±0.001b | 0.004±0.00b | 0.006±0.00a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



