
Submitted:
14 June 2023
Posted:
14 June 2023
You are already at the latest version
Abstract
Plant endophytic fungi and rhizosphere soil fungi are often reported as biocontrol agents against plant pathogens or with plant growth promotion potential. Four treatments were performed in field and greenhouse experiments where cucumber plants were inoculated with Trichoderma harzianum and Fusarium oxysporum in 2022. The roots, stems and leaves of cucumber plants and their rhizosphere soil were collected twice individually from the field and greenhouse for isolation of cucumber endophytic and rhizosphere soil fungi. All 74 genera and 1275 fungal strains were sequenced by rDNA ITS. The PDA media yielded the highest number of genera isolated from cucumber plants, rhizosphere soil and both compared to other media. There were no significant differences among the 4 media for the isolation of cucumber endophytic fungi. The best media for isolating fungi from roots and stems were MRBA and PDA, respectively. PDA and CZA had higher isolation efficiency for the rhizosphere soil fungi than MRBA. The 28 fungal genera had high isolation efficiency, and the endophytic Trichoderma strains were best isolated by MEA. It is suggested that PDA can be used as a basic medium, and different media can be considered for specific fungal genera.
Keywords:
plant endophytic fungi
; rhizospheric soil fungi
; isolation
; cucumber
; cultural media
1. Introduction
Fungi occurring in host plants as endophytes have been demonstrated to act as biocontrol agents against plant pathogens and have plant growth-promoting potential [1]. Some soil fungi have been widely studied and commercially marketed as biopesticides, biofertilizers and soil amendments [2]. There are several reports of different isolation media being used to isolate diverse culturable fungi from plants and soil [3,4].
Different media are an important factor affecting the isolation of endophytic fungi [5,6,7]. Culturable endophytic fungal species from Artemisia thuscula have a “preference” for nutrient medium, and using different culture media to obtain a higher diversity of species was suggested [8]. Only one medium [9,10,11,12,13,14,15,16,17], two [5,18] or three [4,19] different media, four [20,21] and even seven different media [7] were reported to isolate plant endophytic fungi, while potato dextrose agar (PDA) was almost always selected among more than one isolation medium. There are various media available for the isolation of plant endophytic fungi, in which PDA is the most common medium [4,5,7,9,15,16,19,21,22,23], and PDA has been reported to have better isolation efficacy than other media [5,23]. Malt extract agar (MEA) is the second most commonly used material to isolate endophytic fungi from plants [4,5,13]. For isolating endophytic fungi, Rose Bengal agar medium (RBA) [20] and corn meal agar (CMA) [11,14] were also chosen less often than PDA and MEA, while other media, such as Czapek agar medium (CZA) [19,23], Hagem agar medium [19], and V8 juice medium, were rarely used [4].
To isolate as many soil fungi as possible, one to four different media [12,24,25,26,27,28,29,30,31] are often applied for isolation. PDA [25,27,31], CZA [27,32,33], RBA [28,34], Martin's Rose Bengal agar medium (MRBA) [26] and MEA [24,30,31,33] are common media for isolating soil fungi. Other media, such as Martin medium [25,31], water agar [27,33], and YM [24], are also used. Among PDA, CZA and WA, PDA resulted in a large number of fungal strains, and most strains were common to both PDA and CZA, while WA supported pycnidia-producing fungi [27]. When Martin medium was used, the number of total fungi in sorghum, eucalyptus and forested soils was higher than that in CZA medium [32].
Plant endophytic fungi and soil fungi have abundant populations, and the selection of suitable medium or different media is one of the key factors for successful isolation of fungal diversity. In this study, to isolate both plant endophytic fungi and soil fungi as fully as possible, a combination of multiple media was used to increase the effectiveness of fungal isolation. During two growth periods of cucumber in the field and greenhouse, potato dextrose medium (PDA), malt extract medium (MEA), Martin's Rose Bengal agar medium (MRBA) and corn powder agar medium (CMA) were selected to isolate endophytic fungi from cucumber plants, and Czapek agar medium (CZA), PDA and MRBA were used to isolate soil fungi from the cucumber rhizosphere. A total of five different media were tested to optimize isolation media to isolate various communities of cucumber plants and rhizosphere soils.
2. Materials and Methods
2.1. Plants and soil sampling
On June 20 and July 12, 2022, the cucumber field experiment and greenhouse experiment were separately carried out in the field and greenhouse at the Chinese Academy of Agricultural Sciences (39°57 '34 "N, 116°19' 19" E). Four treatments were performed: control (CC), inoculation with Trichoderma harzianum (CT), inoculation with Fusarium oxysporum (CF) and inoculation with T. harzianum and F. oxysporum (TF). The field experiment was carried out in a field with length × width =15 m × 1.2 m, which had 20 rows, and each treatment consisted of 5 rows. There were 10 cucumber plants in each row, for a total of 200 cucumber plants. Samples were collected from the field and greenhouse twice separately (Table 1), and in the field, each plant sample was collected from one row for each treatment. The soil was collected along with the whole plant, wrapped in a sterilized paper bag, and immediately transported to the laboratory. In the greenhouse experiment, each treatment contained 30 plants, and a total of 120 plants were grown. One cucumber plant was grown in each pot (10 cm×10 cm) filled with 500 g of the soil from the field. Samples were collected twice from the greenhouse experiments (Table 1). The fungi were isolated from the roots, stems and leaves of cucumber plants 48 hours after collection. The soil was stored in sterilized polythene bags at 4 ℃ and isolated within 7 days.
2.2. Cultural media
Four different cultural media were used for isolation of cucumber endophytic fungi: PDA ( potato 200 g, dextrose 20 g, agar 15~20 g, distilled water 1000 mL) , MEA ( malt extract 30 g, mycological peptone 3 g, agar 15 g, distilled water 1000 mL), CMA agar (corn meal 2 g, agar 15 g, distilled water 1000 mL), and MRBA (peptone 5 g, dextrose 10 g, KH2PO4 1 g, MgSO4 0.5 g, agar 15 g , rose bengal 0.03 g, chloram phenicol 0.1 g, distilled water 1000 mL), while three media, PDA, MRBA and CZA (NaNO3 2.0 g, K2HPO4 1 g, KCl 0.5 g, MgSO4 0.5 g, FeSO4 0.01 g, saccharose 30 g), were used for isolation of rhizosphere soil fungi. Except for MRBA, ampicillin and chloramphenicol were added to the other four culture media so that the concentration of both reached 10 μg/ml to inhibit the growth of bacteria.
2.3. Isolation of endophytic and rhizosphere soil fungi
The plant tissues were cut into 1-2 cm blocks, soaked in 70% ethanol for 30 s, transferred to 4% NaClO, immersed for 2 min, and rinsed with sterile water 3 times, and the surface moisture was absorbed by sterile filter paper. Drops of last step sterilization water were poured on medium plates as a control check for complete sterilization. After the dried tissue blocks were inoculated into various media with 4 blocks per plate, the inoculation medium was placed in a 28 °C incubator.
The soil fungi were separated by the dilution plate method. Ten grams of soil was added to a 250 ml conical flask containing 90 ml of sterile water in rotation and mixed at 160 rpm for 25 min. Subsequently, serial 10-fold dilutions were performed, and 100 µL aliquots of 10-3 and 10-4 dilutions of each sample were plated for isolation of fungi by three culture media. All plates were incubated at 28 °C for 3 days to observe and purify the fungi.
The fungi were purified by transferring a small amount of mycelium from the edge of each colony to a new PDA plate. Each pure strain was preserved on PDA media in two centrifuge tubes at room temperature.
2.4. Strain identification
The total genomic DNA of each strain was extracted using a direct PCR kit from Triumfi Plant based on the manufacturer's instructions (cat. SD311, Genes and, Chain). Using the genomic DNA as the template, the internal transcribed spacer (ITS) regions spanning from the end of the 18S rDNA to the beginning of the 28S rDNA for each strain were amplified using the primer pair V9G/ITS4 [35]. Thermal cycling conditions for PCR amplification were set as follows: 98 ℃ for 5 min, 35 cycles of 98 ℃ for 10 s, 60 ℃ for 30 s, 72 ℃ for 25 s, and a final extension at 72 ℃ for 5 min. PCR products were sent to GENEWIZ, Inc. (Suzhou, China) for sequencing. The 1560 sequences were run through the BLASTN search page using the Megablast program (National Center for Biotechnology Information; Bethesda MD, USA), where hits with more than 98% similarity with published papers and their accession numbers were obtained. The taxon names of BLAST search results were checked on the web of fungal names (https://nmdc.cn/fungalnames), and the current names were used in the resulting analysis. A total of more than 1000 Trichoderma strains were isolated, and those from the CT and TF treatments were selected for sequencing with at least one Trichoderma strain each from the leaf, stem and root of each plant and their corresponding rhizosphere soil samples.
2.4. Data analysis
The rarefaction curve was constructed using the iNEXT package, comparisons of the estimated and extrapolated species richness in plots were calculated based on 50 bootstrap replicates at a 95% confidence interval [36]. A Krona pie chart of 1275 strains was generated using the OmicShare tools at www.omicshare.com/tools, while 1584 strains of Trichoderma harzianum from the CT and TF treatments and 154 strains of Fusarium oxysproum from the CF and TF treatments were not included in the Krona pie chart. The construction of Venn diagrams and all statistical analyses of 1275 strains were performed using imageGP [37]. The diversity of fungi was analysed by the Shannon index [38], and the number of genera and Simpson index were calculated to determine the homogeneity of fungi [39]. Significance analysis of rhizosphere and endophytic fungal genera, Shannon index and Simpson index was performed by taking each period in the field and greenhouse as a replicate, with a total of 4 replicates per medium, and counting the number of genera, Shannon index and Simpson index. Then, the number of genera, Shannon index and Simpson index were compared using ANOVA and two-way ANOVA followed by Bonferroni correction (multiple comparisons) [40] and a Wilcoxon matched pairs test (pairwise comparisons) [41].
3. Results
3.1. Community composition of endophytic fungi and rhizosphere soil fungi
A total of 1275 strains isolated from cucumber plants and rhizosphere soil were analysed for fungal diversity. With the isolation depth near the saturation point of the sparse curve, the sampling effort was sufficient to achieve the diversity of the isolated fungal species (Figure 1). Through rDNA ITS sequence analysis, all 1275 strains were classified into 145 species (Table 2), representing 4 phyla, 14 classes, 27 orders, 47 families, and 74 genera. As shown in Figure 2, the percentage of Ascomycota was highest, followed by Mucoromycota, Basidiomycota and Oomycota. In Ascomycota, Sordariomycetes made up the greatest proportion of the class, followed by Dothideomycetes and Eurotiomycetes. With regard to orders, Eurotiales, Pleosporales and Hypocreales were the most abundant. On a familial level, Aspergillaceae, Didymellaceae and Cladosporiaceae were the most abundant. Among all isolated fungi, Aspergillus, Stagonosporopsis and Cladosporium were the top three genera, and they also accounted for the largest proportion of Ascomycetes. Among Mucoromycota, Mortierella and Mucor were the most isolated with 52 and 20 strains, respectively, followed by Actinomortierella and Cunninghamella with 4 strains, Rhizopus with 3 strains, Umbelopsis with 2 strains, and Linnemannia with 1 isolate. In Basidiomycota, a total of 7 genera, Malassezia (3 strains), Rhizoctonia (3 strains), Ceratobasidium (2 strains), Irpex (1 strain), Moesziomyces (1 strain), Schizophyllum (1 strain) and Trametes (1 strain), were obtained. Pythium and Globisporangium were the isolated fungal genera belonging to Oomycota.
3.2. Influence of culture medium on fungal isolation
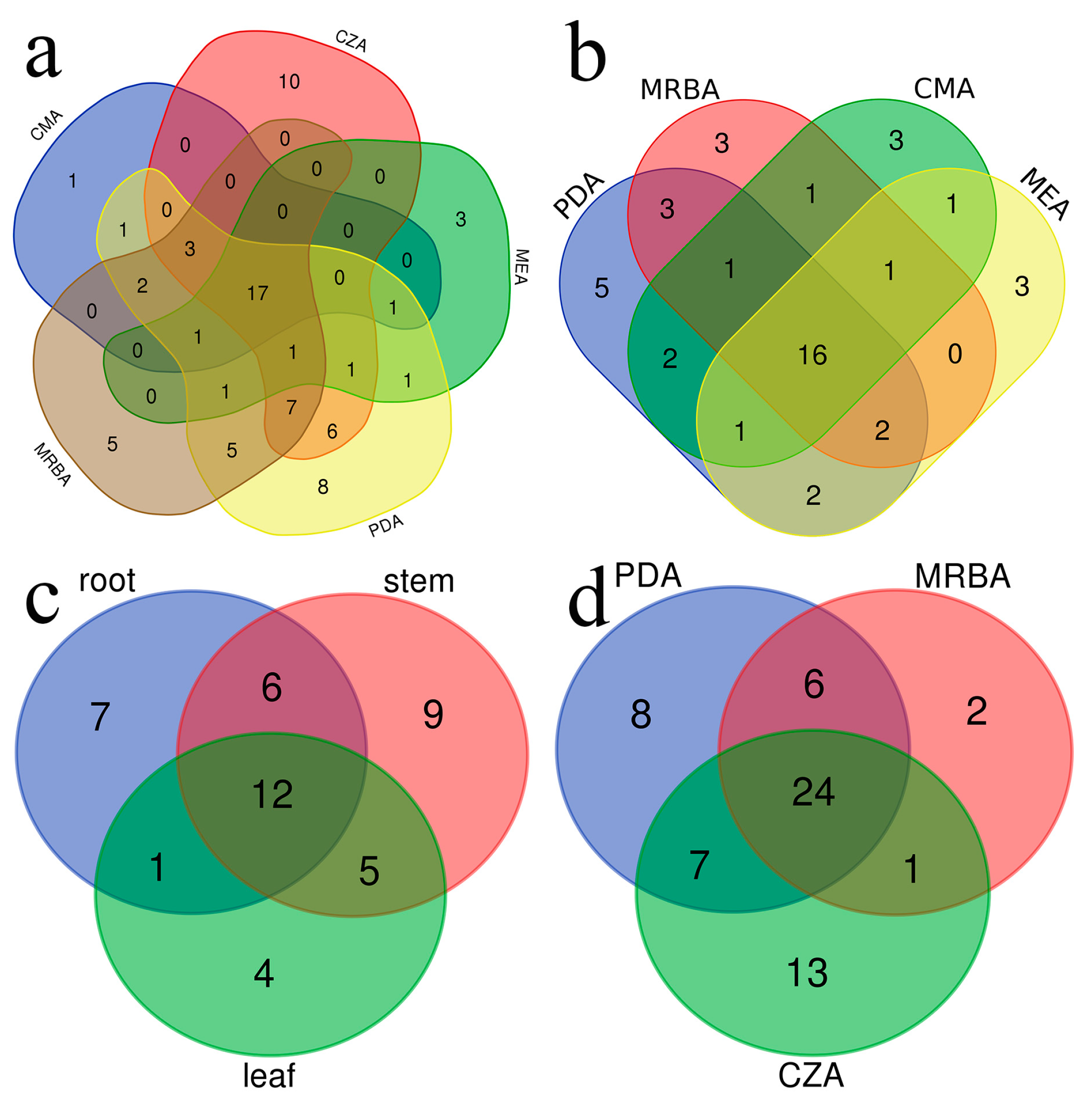
A total of five types of cultural media were utilized in this investigation, yielding a total of 74 fungal genera. Each culture medium yielded between 26 and 55 distinct fungal genera (Table 3). Furthermore, there were 28 fungal genera with high isolation efficiency. Among them, 17 fungal genera were successfully isolated using all five media, 16 and 24 genera were isolated with all 4 endophytic fungi isolation media and all 3 rhizosphere soil fungi isolation media individually, and 12 genera were isolated from roots, stems and leaves of cucumber plants (Table 4; Figure 4).
The number of genera, strains and endophytic genera of endophytic fungi and soil fungi isolated with each medium are listed in Table 3. Specifically, 10 genera of fungi were isolated only by CZA, which yielded the highest number of strains, while 8 genera were isolated only by PDA, 5 genera were isolated only by MRBA, 3 genera were isolated only by MEA and 1 genus was isolated only by CMA (Table 3; Figure 4a). In endophytic fungi, the number of genera isolated by PDA was the highest, while the number of strains isolated by PDA was the lowest. PDA yielded the highest number of genera and strains in the rhizosphere soil fungi (Figure 3).
If only one medium is selected, PDA yields the highest number of genera isolated from cucumber endophytes, rhizosphere soil fungi and both compared to other media. Therefore, it is a suitable medium for the simultaneous isolation of both cucumber endophytic and rhizosphere soil fungi. PDA alone isolated endophytic fungi of 32 genera accounting for 72.7% of all four media isolated to 44 genera, rhizosphere soil fungi of 45 genera accounting for 73.8% of all three media isolated to 61 genera, and both endophytic and rhizosphere soil fungi of 55 genera accounting for 74.3% of all five media isolated to 74 genera. There were 22 genera of endophytic fungi isolated with both PDA and MRBA, accounting for 81.5% of all endophytic fungi isolated with MRBA. Only 5 genera were isolated with MRBA but not PDA, accounting for 11.3% of all endophytic fungal genera. There were 30 genera of rhizosphere soil fungi isolated with both PDA and MRBA, accounting for 90.9% of all rhizosphere soil fungi isolated with MRBA. Only 3 were confirmed to be isolated with MRBA and not with PDA, accounting for 4.9% of all rhizosphere soil fungal genera.
Figure 4.
Venn diagram shows: (a) Genera of all isolated fungi in different media; (b) Genera of isolated endophytic fungi in different media; (c) Genera of isolated endophytic fungi in roots, stems and leaves; (d) Genera of isolated soil fungi of different media types.
Figure 4.
Venn diagram shows: (a) Genera of all isolated fungi in different media; (b) Genera of isolated endophytic fungi in different media; (c) Genera of isolated endophytic fungi in roots, stems and leaves; (d) Genera of isolated soil fungi of different media types.
3.3. Effects of culture medium on endophytic fungal diversity
In the isolation of endophytic fungi, there were no significant differences among the four media. The number of genera isolated with each medium was 11.75±2.50, 10.00±2.44, 11.75±3.30 and 10.75±2.36 (Figure 5a), and the Shannon index was 2.81±0.53, 2.57±0.29, 2.85±0.56, and 2.73±0.45 (Figure 5b), respectively; Simpson's index was 0.80±0.13, 0.79±0.14, 0.81±0.13 and 0.82±0.14 (Figure 5c), respectively. The number of endophytic genera and Shannon index of fungi isolated with PDA were larger than those of the other media.
A total of 506 strains of endophytic fungi were isolated by PDA, MRBA, MEA, and CMA media, belonging to 21 orders, 33 families and 44 genera (Figure 4b). The highest number of genera was recovered from PDA with 131 strains and 32 genera, followed by MRBA (114 strains, 27 genera), CMA (138 strains, 26 genera) and MEA (123 strains, 26 genera) (Table 3). There were 16 genera of endophytic fungi obtained in all four media (Table 4). Among the endophytic fungi, Cladosporium, Colletotrichum and Stagonosporopsis were the genera with the highest number of strains isolated from the four cultures. On the other hand, 5, 3, 3 and 3 genera were unique to the isolated endophytic fungi from the PDA, MRBA, CMA and MEA media, respectively (Figure 3b). The genera Acremonium, Aureobasidium, Cephalotheca, Humicola and Pythium were obtained only on PDA, while Globisporangium, Harknessia and Trametes were isolated only from MRBA; Boeremia, Clonostachys and Moesziomyce were isolated only from CMA; and Ceratobasidium, Curvularia and Preussia were isolated only from MEA.
There were 20 genera of endophytic fungi isolated with PDA and CMA, accounting for 76.9% of all endophytic fungal genera isolated with CMA. Only 6 genera were isolated with CMA but not PDA, accounting for 13.6% of all endophytic fungal genera. There were 17 genera of endophytic fungi isolated with PDA and MEA, accounting for 65.4% of all endophytic fungi isolated with MEA. Only 5 of them were isolated with MEA but not PDA, accounting for 11.4% of all endophytic fungal genera. Compared with PDA, 6, 5 and 5 genera of specific fungi were isolated with CMA, MEA and MRBA, respectively.
3.4. Effect of the medium type on endophytic fungal diversity in different tissues of the plant
The most genera of fungi isolated with the four media were 113 strains (32 genera) in the stems, followed by 240 strains (26 genera) in the roots and 153 strains (22 genera) in the leaves (Figure 3d). The number of endophytic fungi isolated on different parts of the plant on different media was remarkably different (two-way ANOVA test, p<0.05). When MEA was adopted, the number of genera isolated from stems (6.25) was significantly greater than that from roots (2.50) (Table 5). In the roots, the number of endophytic fungi isolated by MRBA (6.50) was significantly higher than that isolated on MEA (2.50), but in the stems, the number of fungi isolated with MRBA was significantly lower than that isolated with PDA (Table 5).
MRBA was the medium with the largest number of root fungal genera isolated among the four media, significantly more than MEA. In roots, Fusarium, Monosporascus and Stagonosporopsis were the three genera with the highest number of strains obtained. The greatest number of Monosporascus and Stagonosporopsis strains was observed in MEA and PDA media, respectively. CMA yielded the highest number of Fusarium strains. MRBA isolated the largest number of Monosporascus strains. Preussiacan was only isolated from the root using MEA, and Trametescan was only isolated from the root using MRBA.
In the stem, PDA showed significantly better separation than MRBA. In the stem, Cladosporium, Colletotrichum and Stagonosporopsis were the three genera with the highest number of strains obtained. In addition to MEA, Stagonosporopsis was the main isolated genus in the other three media. Moesziomyces was isolated from stems only by CMA. Acremonium, Cephalotheca, Humicola and Pythium were isolated from stems using only PDA. Harknessia was only isolated from the stem using MRBA.
In the leaves, the number of fungal genera isolated by the four medium differences was not significant. In the leaves, Alternaria, Cladosporium and Stagonosporopsis were the three genera with the highest number of strains obtained. Among them, Stagonosporopsis had the most strains in all four cultures. Clonostachys was isolated only from leaves using CMA; Curvularia was isolated only in leaves using MEA; and Aureobasidium was isolated from leaves only using PDA.
3.5. Effects of culture medium on soil fungal diversity
In rhizosphere soil fungi, there were significant differences in the number of genera, Shannon index and Simpson index of fungi isolated by CZA, MRBA and PDA. The number of genera, Shannon index and Simpson index of soil fungi isolated with CZA, MRBA and PDA were 22.75 ± 3.10, 11.75 ± 7.41, 24.50 ± 6.45 (Figure 5d); 3.76± 0.04, 2.88 ± 0.53, 3.85 ± 2.88 (Figure 5e) and 0.90 ± 0.01, 0.85 ± 0.03, 0.91 ± 0.01 (Figure 5f), respectively. The Simpson's index of soil fungi isolated by PDA was significantly higher than that of soil fungi isolated by MRBA (Figure 5d,f). The Shannon index of PDA and CZA was significantly higher than that of MRBA (Figure 5e). The diversity of soil fungi isolated on PDA was significantly higher than that from MRBA.
A total of 771 soil fungi were isolated by CZA, PDA and MRBA, belonging to 21 orders, 37 families and 61 genera (Figure 2). There were 24 genera of fungi that could be isolated with all three media, and the genus with the largest number of fungal strains isolated in all three media was Aspergillus (Table 4). The number of soil fungi isolated with PDA was the highest (331 strains, 45 genera), followed by CZA (290 strains, 45 genera) and MRBA (150 strains, 33 genera) (Table 3). On the other hand, there were 13 genera, 8 genera and 2 endemic genera of soil fungi isolated with CZA, MRBA and PDA media, respectively (Figure 4d). Chaetomidium and Irpex were obtained only in MRBA isolation.
There were 31 genera in the rhizosphere soil fungi isolated on PDA and CZA, accounting for 68.9% of all the fungal genera isolated with CZA. Fourteen genera were isolated with CZA but not PDA, accounting for 23% of all fungal genera isolated from rhizosphere soil.
3.6. Effects of culture medium on fungal genera with high isolation efficiency
3.6.1. Trichodera and Fusarium
All 1617 Trichoderma strains and 232 Fusarium strains isolated in this study were analysed. In endophytic Trichoderma strains, strains isolated with MEA were significantly higher than those from MRBA, and the strain number was in the order MEA>CMA>PDA>MRBA. In rhizosphere soil Trichoderma strains, there was no significant difference in the number of Trichoderma strains isolated with the three media, but the number of strains isolated with MRBA was the highest (514 strains), and CZA was the lowest (374 strains). The number of Trichoderma strains isolated with the three rhizosphere soil culture media was in the order of MRBA>PDA>CZA. There was no significant difference between Fusarium fungi isolated with each medium in endophytic and rhizosphere soil. In the endophytic fungi, Fusarium strains were the most isolated by CMA, with a total of 40 strains, and PDA was the least, with a total of 26 strains. The number of endophytic Fusarium strains was obtained in the order of CMA>MRBA>MEA>PDA. In the rhizosphere soil fungi, PDA isolated the largest number of Fusarium strains (44 strains), and MRBA isolated the least number of Fusarium strains (23 strains). The number of rhizosphere soil Fusarium strains obtained by the isolation medium was PDA>CZA>MRBA.
3.6.2. Other 26 fungal genera
There were 26 fungal genera with high isolation efficiency except Trichoderma and Fusarium (Table 4). For 16 fungal genera isolated with 4 endophytic cultural media, most genera were not significant among the 4 media except Trichoderma. In endophytic fungi, MEA obtained more strains than MRBA when only Colletotrichum, Plectosphaerella, Pseudogymnoascus and Stagonosporopsis were isolated. In the endophytic fungi, the total strain number of fungal genera with high isolation frequency obtained from the four media was in the order CMA>PDA>MEA>MRBA.
There were 22 fungal genera with high isolation efficiency, except Trichoderma and Fusarium (Table 4). In the isolation of Talaromyces, the number of strains obtained by PDA was significantly greater than that of MRBA, while in the isolation of Paramyrothecium, CZA was significantly greater than MRBA. In the rhizosphere soil fungi, except for Clonostachys, Humicola, Lectera and Malassezia, more strains were isolated with PDA than from MRBA in all genera. In the rhizosphere soil fungi, the strain number of fungal genera with high isolation frequency obtained from the three media was PDA>CZA>MRBA.
The 12 genera Alternaria, Aspergillus, Chaetomium, Cladosporium, Colletotrichum, Fusarium, Penicillium, Plectosphaerella, Pseudogymnoascus, Stagonosporopsis and Trichoderma were isolated on both 4 endophytic fungal culture media and 3 rhizosphere soil fungal culture media, which may play an important role in cucumber plants grown in this study.
4. Discussion
Our study showed 74 genera of fungi isolated from leaves, stems and roots of cucumber plants and cucumber rhizosphere soils. Among 1275 strains, the percentage of Ascomycota was highest, followed by Mucoromycota, Basidiomycota and Oomycota. Aspergillus, Stagonosporopsis and Cladosporium were the top three genera, and they also accounted for the largest proportion of Ascomycetes. Among the five media tested, PDA yielded the highest number of fungal genera from both cucumber endophytic and rhizosphere soil samples. All five media were able to isolate unique genera. It should be noted that CZA and PDA can isolate as many as 10 and 8 unique genera in total. Significant differences among the 4 media for the isolation of cucumber endophytic fungi and the 3 media for rhizosphere fungi were also tested. The 12 fungal genera were isolated both from 4 endophytic fungi culture media and 3 rhizosphere soil fungi culture media.
The success of endophytic fungi is one of the problems in the study of endophytic fungi. However, the isolation of endophytic fungi is affected by many factors, among which the type of culture medium is an important factor [6,10,18]. The nutrient difference of the culture medium is the reason for the culture medium preference of endophytic fungi [8]. In this study, we isolated endophytic fungi from cucumber at two different growth stages in the greenhouse and field by the tissue isolation method and investigated the effect of the medium type on fungal isolation. The results showed that the isolation of endophytic fungi was not affected by the type of medium, but the type of medium had a significant effect on the isolation of endophytic fungi from different parts. The best media for isolating cucumber endophytic fungi from roots and stem leaves were MRBA and PDA.
Medium nutrients affect the isolation of endophytic fungi [7,42,43]. The main carbon source of MRBA and PDA was glucose, which could be directly absorbed and utilized by fungi, while the main carbon sources of CMA and MEA were malt extract, soybean peptone and corn leaching powder, which were more complex. Therefore, more genera of endophytic fungi were isolated with PDA and MRBA than with CMA and MEA. The Simpson index and Shannon index are indicators used to describe species diversity within a habitat, with higher values indicating greater equilibrium and abundance of fungi and lower values indicating greater selectivity [44,45]. MRBA and PDA had higher Shannon‒Wiener and Simpson indices, indicating a better balance and diversity of endophytic fungi on MRBA and PDA. The Shannon‒Wiener index and Simpson index of MEA were the lowest, indicating that MEA had high selectivity for fungi and was suitable for the isolation and culture of fungi of specific species. For example, the four genera Boeremia, Didymella, Clonostachys and Pseudozyma can only be isolated by MEA. More fungal genera and fewer strains were isolated with MRBA than from MEA and CMA. This may be due to the restriction of fungal growth by the Bengal red contained in MRBA [8], and certain slow-growing fungal strains by MRBA were observed in this study. Endophytic fungal genera isolated with PDA were the most abundant. Among the endophytic fungi, 12 genera of PDA were not isolated, and the genera isolated with the other 3 media accounted for 11-13% of all endophytic fungi individually. Endophytic fungi have tissue specificity in plants. Some scholars believe that this phenomenon is due to the different substrate utilization abilities of endophytic fungi [8,42,46,47,48]. The experimental results showed that the endophytic fungi in different parts of the plant showed a certain preference for the medium. MEA was more effective in isolating stem fungi than roots, and PDA was more effective in isolating stem fungi than roots and leaves. The nutrient composition of plant parts is partially similar to that of some medium components; for example, plant roots and MRBA both contain high concentrations of inorganic salts. The results showed that endophytic fungi from different parts of cucumber plants have certain preferences for different media.
Rhizosphere soil fungi live in soil, a natural medium, and artificial media are doomed to fail to achieve soil-like conditions. Therefore, the selection of an appropriate medium or medium combination is an important factor to improve the isolation efficiency of soil fungi [49,50,51]. The type of medium had a significant effect on the isolation of soil fungi. The isolation of PDA and CZA was better than that of MRBA. The fungal colonies growing on MRBA were smaller and less numerous than those on PDA and CZA. The inhibitory effect of Bengal red and the short incubation time (3 days) may be responsible for the poor isolation of MRBA, which is also verified by the low Simpson index. Among the 3 culture media selected for isolating rhizosphere soil fungi, PDA exhibited the most effective isolation performance, with the highest number of isolated genera and strains. Except for the fungal genera Chaetomidium, Monosporascus and Irpex, there were more fungal genera isolated by PDA than by MRBA. The combination of PDA and CZA was better used for rhizosphere soil fungi isolation because CZA isolated 14 genera that PDA did not, accounting for 23% of all fungi isolated in the rhizosphere.
In this study, three fungal genera were found to prefer specific culture media. For the endophytic Trichoderma, the fungal strain number isolated on MEA was significantly higher than that from MRBA. For the rhizosphere soil fungi Talaromyces, the fungal strain number isolated with PDA was significantly higher than that with MRBA. For the rhizosphere soil fungi Paramyrothecium, the fungal strain number isolated with CZA was significantly higher than that from MRBA.
5. Conclusions
In this study, we investigated the influence of the medium type on the isolation of cucumber endophytic fungi and rhizosphere soil fungi. We found that the type of medium did not have a significant effect on the isolation of endophytic fungi, but the number of rhizosphere soil fungal genera isolated with PDA was significantly higher than that from MRBA. Based on the data obtained here, we suggest that PDA can be used as a basic medium for the isolation of both cucumber endophytic and rhizosphere soil fungi, and different media can be considered for the isolation of endophytic fungi from different parts of the plant and to emphasize specific fungal genera.
Author Contributions
Yong-chun Niu and Hui Deng conceived and designed the experiments; Chong-yang Cheng and Ming-yuan Zhang performed the experiments; Chong-yang Cheng and Hui Deng analysed the data; Yue-Hua Geng and Meng Zhang contributed materials/analysis tools; Hui Deng, Chong-yang Cheng and Yue-hua Geng wrote the paper. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Key R&D Program of China (Grant No. 2022YFD1901300), Fundamental Research Funds for Central Non-profit Scientific Institution (No. 1610132018011) and The Agricultural Science and Technology Innovation Program (ASTIP).
Acknowledgements
We thank Dr. Yinan Ma (Department of Agricultural Microbiology, Institute of Agricultural Resources and Regional Planning, Chinese Academy of Agricultural Sciences) for his suggestion of statistical analyses using imageGP.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bamisile, B.S.; Dash, C.K.; Akutse, K.S.; Keppanan, R.; Afolabi, O.G.; Hussain, M.; Qasim, M.; Wang, L. Prospects of endophytic fungal entomopathogens as biocontrol and plant growth promoting agents: An insight on how artificial inoculation methods affect endophytic colonization of host plants. Microbiol. Res. 2018, 217, 34–50. [Google Scholar] [CrossRef] [PubMed]
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, E.L.; Marra, R.; Woo, S.L.; Lorito, M. Trichoderma–plant–pathogen interactions. Soil Biology and Biochemistry 2008, 40, 1–10. [Google Scholar] [CrossRef]
- Arenz, B.E.; Held, B.W.; Jurgens, J.A.; Farrell, R.L.; Blanchette, R.A. Fungal diversity in soils and historic wood from the Ross Sea Region of Antarctica. Soil Biology and Biochemistry 2006, 38, 3057–3064. [Google Scholar] [CrossRef]
- Salazar-Cerezo, S.; Martinez-Montiel, N.; Cruz-Lopez, M.D.C.; Martinez-Contreras, R.D. Fungal diversity and community composition of culturable fungi in Stanhopea trigrina cast gibberellin producers. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Nischitha, R.; Shivanna, M.B. Antimicrobial activity and metabolite profiling of endophytic fungi in Digitaria bicornis (Lam) Roem. and Schult. and Paspalidium flavidum (Retz.) A. Camus. 3 Biotech 2021, 11. [CrossRef]
- Muggia, L.; Kopun, T.; Grube, M. Effects of growth media on the diversity of culturable fungi from lichens. Molecules 2017, 22, 824. [Google Scholar] [CrossRef]
- Prior, R.; Görges, K.; Yurkov, A.; Begerow, D. New isolation method for endophytes based on enzyme digestion. Mycol Progress 2014, 13, 849–856. [Google Scholar] [CrossRef]
- Cosoveanu, A.; Cabrera, R. Fungi as endophytes in Artemisia thuscula: Juxtaposed elements of diversity and phylogeny. Journal of Funig 2018, 4, 17. [Google Scholar] [CrossRef]
- Zeng, Z.H.; Yuan, Z.L.; Huang, X.N.; Liu, F. Isolation and diversity of mangrove endophytic fungi. Protection Forest Science and Technology 2022, 39-42, 45. [Google Scholar] [CrossRef]
- Smith, H.; Wingfield, M.J.; Petrini, O. Botryosphaeria dothidea endophytic in Eucalyptus grandis and Eucalyptus nitens in South Africa. Forest Ecol Manag 1996, 89, 189–195. [Google Scholar] [CrossRef]
- Blodgett, J.T.; Swart, W.J.; Weeks, L.W.J. Species composition of endophytic fungi in Amaranthus hybridus leaves, petioles, stems, and roots. Mycologia 2000, 92, 853–859. [Google Scholar] [CrossRef]
- Broda, D.M.; Bell, R.G.; Boerema, J.A.; Musgrave, D.R. The abattoir source of culturable psychrophilic Clostridium spp. causing 'blown pack' spoilage of vacuum-packed chilled venison. J. Appl. Microbiol. 2002, 93, 817–824. [Google Scholar] [CrossRef] [PubMed]
- Bettucci, L.; Simeto, S.; Alonso, R.; Lupo, S. Endophytic fungi of twigs and leaves of three native species of Myrtaceae in Uruguay. Sydowia 2004, 56, 8–23. [Google Scholar]
- Blodgett, J.T.; Swart, W.J.; Louw, S.V.; Weeks, W.J. Soil amendments and watering influence the incidence of endophytic fungi in Amaranthus hybridus in South Africa. Appl Soil Ecol 2007, 35, 311–318. [Google Scholar] [CrossRef]
- Xue, Q.W.; Niu, Y.C.; Deng, H. Diversity of endophytic fungi in common cucurbit plants. Mycosystema 2015, 34, 196–203. [Google Scholar] [CrossRef]
- Belfiori, B.; Rubini, A.; Riccioni, C. Diversity of endophytic and pathogenic fungi of Saffron (Crocus sativus) plants from cultivation sites in Italy. Diversity 2021, 13, 535. [Google Scholar] [CrossRef]
- Wang. Z.B.; Zhou. X.; Ma. X.B.; Wang, J.L. Isolation of endophytic fungi from medicinal plants and screening and identification of antagonism. Chinese Agricultural Science Bulletin 2022, 38, 75–81. [Google Scholar]
- Evans, H.C.; Holmes, K.A.; Thomas, S.E. Endophytes and mycoparasites associated with an indigenous forest tree, Theobroma gileri, in Ecuador and a preliminary assessment of their potential as biocontrol agents of cocoa diseases. Mycol Progress 2003, 2, 149–160. [Google Scholar] [CrossRef]
- Khan, A.R.; Waqas, M.; Ullah, I.; Khan, A.L.; Khan, M.A.; Lee, I.; Shin, J. Culturable endophytic fungal diversity in the cadmium hyperaccumulator Solanum nigrum L. and their role in enhancing phytoremediation. Environ. Exp. Bot. 2017, 135, 126–135. [Google Scholar] [CrossRef]
- Li, W.Z.; Duan, W.J.; Zhou, X.; Chen, J.; Jiang, X.Z.; Yang, Y.; Cai, L. Diversity of fungi in wheat seeds. Mycosystema 2021, 40, 487–501. [Google Scholar] [CrossRef]
- Guo, L.D.; Hyde, K.D.; Liew, E. Identification of endophytic fungi from Livistona chinensis based on morphology and rDNA sequences. New Phytol 2000, 147, 617–630. [Google Scholar] [CrossRef]
- Xiang, L.; Gong, S.; Yang, L.; Hao, J.; Xue, M.; Zeng, F.; Zhang, X.; Shi, W.; Wang, H.; Yu, D. Biocontrol potential of endophytic fungi in medicinal plants from Wuhan Botanical Garden in China. Biol. Control. 2016, 94, 47–55. [Google Scholar] [CrossRef]
- Wang, H.X.; Yu, Y.R.; Huang, B.K. Diversity of culturable endophytic fungi from Broussonetia papyrifera. Mycosystema 2020, 39, 2399–2408. [Google Scholar] [CrossRef]
- Figueredo, H.M.; Gonçalves, V.N.; Godinho, V.M.; Lopes, D.V.; Oliveira, F.S.; Rosa, L.H. Diversity and ecology of cultivable fungi isolated from the thermal soil gradients in Deception Island, Antarctica. Extremophiles 2020, 24, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.L.; Liu, M.; Huang, H.Q.; Zhu, J.; Bao, S.X. Diversity of soil culturable fungi in Bamen Bay Mangrove Forests. Chinese Journal of Tropical Crops 2013, 34, 181–187. [Google Scholar]
- Zhang, J.Z.; Chen, X.R.; Yang, C.D.; Xue, L. A study on the diversity of soil cultured fungi in the alpine grassland of Eastern Qilian Mountains. Acta Prataculturae Sinica 2010, 19, 124–132. [Google Scholar]
- Vasanthakumari, M.M.; Shivanna, M.B. Fungal assemblages in the rhizosphere and rhizoplane of grasses of the subfamily Panicoideae in the Lakkavalli region of Karnataka, India. Microbes Environ 2011, 26, 228–236. [Google Scholar] [CrossRef]
- Jiang, H.; Li, Y.C.; Jiang, Y. Seasonal variation of soil fungal diversity in the rhizosphere of Phragmites australis. Journal of Liaoning Normal University (Natural Science Edition) 2017, 40, 89–94. [Google Scholar] [CrossRef]
- Ayob, Z.; Kusai, N.A.; Ali, S.R.A. Sequence-based identification and characterisation of cultivated filamentous fungi in the Alan Bunga peat ecosystems of Sarawak, Malaysia. Mires Peat 2018, 21, 1–19. [Google Scholar] [CrossRef]
- Zhou, J.; Miao, Y.F.; Fang, K.; Chen, L.; Yang, Z.P.; Dong, X.F.; Zhang, H.B. Diversity of the endophytic and rhizosphere soil fungi of Ageratina adenophora. Ecological Science 2019, 38, 1–7. [Google Scholar] [CrossRef]
- Guo, M.J.; Zhao, X.Y.; Huang, J.H.; Wei, S.Z.; Xu, X.L. Isolation and activity analysis of fungi from the rhizosphere of mangroves in Xinglin Bay, Xiamen. Microbiology 2021, 48, 1496–1503. [Google Scholar] [CrossRef]
- Vieira, F.C.S.; Nahas, E. Comparison of microbial numbers in soils by using various culture media and temperatures. Microbiol. Res. 2005, 160, 197–202. [Google Scholar] [CrossRef]
- Grishkan, I.; Beharav, A.; Kirzhner, V.; Nevo, E. Adaptive spatiotemporal distribution of soil microfungi in 'Evolution Canyon' III, Nahal Shaharut, extreme southern Negev Desert, Israel. Biol. J. Linn. Soc. 2007, 90, 263–277. [Google Scholar] [CrossRef]
- Watrud, L.S.; Martin, K.; Donegan, K.K.; Stone, J.K.; Coleman, C.G. Comparison of taxonomic, colony morphotype and PCR-RFLP methods to characterize microfungal diversity. Mycologia 2006, 98, 384–392. [Google Scholar] [CrossRef]
- de Hoog, G.S.; van den Ende, A. Molecular diagnostics of clinical strains of filamentous Basidiomycetes. Mycoses 1998, 41, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, T.C.; Ma, K.H.; Chao, A. iNEXT: an R package for rarefaction and extrapolation of species diversity (Hill numbers). Methods Ecol. Evo. 2016, 7, 1451–1456. [Google Scholar] [CrossRef]
- Chen, T.; Liu, Y.X.; Huang, L. ImageGP: An easy-to-use data visualization web server for scientific researchers. iMeta 2022, 1, n/a-n/a. [Google Scholar] [CrossRef]
- Shannon, C.E.; Weaver, W. The mathematical theory of communication. University of Illinois Press: Urbana, 1964.
- Simpson, E.H. Measurement of Diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Armstrong, R.A. When to use the Bonferroni correction. Ophthalmic & Physiological Optics the Journal of the British College of Ophthalmic Opticians 2015, 34, 502–508. [Google Scholar]
- Gehan, E.A. A generalized Wilcoxon test for comparing arbitrarily singly-censored samples. Biometrika 1965, 52, 203–224. [Google Scholar] [CrossRef]
- Yurkov, A.M.; Kemler, M.; Begerow, D. Species Accumulation Curves and Incidence-Based Species Richness Estimators to Appraise the Diversity of Cultivable Yeasts from Beech Forest Soils. Plos One 2011, 6, e23671. [Google Scholar] [CrossRef]
- Wong Chin, J.M.; Puchooa, D.; Bahorun, T.; Jeewon, R. Antimicrobial properties of marine fungi from sponges and brown algae of Mauritius. Mycology 2021, 12, 231–244. [Google Scholar] [CrossRef]
- Jiang, X.L.; Ding, W.L.; Xing, X.K. Effects of different media on the isolation of symbiotic fungi from protocorm of medicinal orchid Gymnadenia conopsea. Mycosystema 2022, 41, 952–961. [Google Scholar] [CrossRef]
- Liu, S.Y.; Wang, Q.Z.; Xu, H.X.; Huang, S.Z.; Feng, J. Effects of common medium and optimized medium on fungal isolation and diversity in Fangchenggang Sea Area. Guangxi Sciences 2021, 28, 65–73. [Google Scholar] [CrossRef]
- White, J.F.; Morrow, A.C.; Morgan-Jones, G.; Chambless, D.A. Endophyte-host associations in forage grasses. XIV. Primary stromata formation and seed transmission in Epichloe typhina: Developmental and Regulatory Aspects. Mycologia 1991, 83, 72. [Google Scholar] [CrossRef]
- de Errasti, A.; Carmarán, C.C.; Novas, M.V. Diversity and significance of fungal endophytes from living stems of naturalized trees from Argentina. Fungal Divers. 2010, 41, 29–40. [Google Scholar] [CrossRef]
- Guo, L.D. Advances of researches on endophytic fungi. Mycosystema 2001, 20, 148–152. [Google Scholar] [CrossRef]
- Dennis, P.G.; Miller, A.J.; Hirsch, P.R. Are root exudates more important than other sources of rhizodeposits in structuring rhizosphere bacterial communities? Fems Microbiol. Ecol. 2010, 72, 313–327. [Google Scholar] [CrossRef]
- Abbott, M.J.; Brewer, J.S. Competition does not explain the absence of a carnivorous pitcher plant from a nutrient-rich marsh. Plant Soil 2016, 409, 495–504. [Google Scholar] [CrossRef]
- Zhang, W.; Xu, J.J.; Zhang, T.Y. Advancement on soil fungal research. Journal of Fungal Research 2005, 3, 52–58. [Google Scholar] [CrossRef]
Figure 1.
Rarefaction curves with 95% confidence intervals of estimated species richness of the isolated fungal community of cucumber in the field and greenhouse.
Figure 1.
Rarefaction curves with 95% confidence intervals of estimated species richness of the isolated fungal community of cucumber in the field and greenhouse.
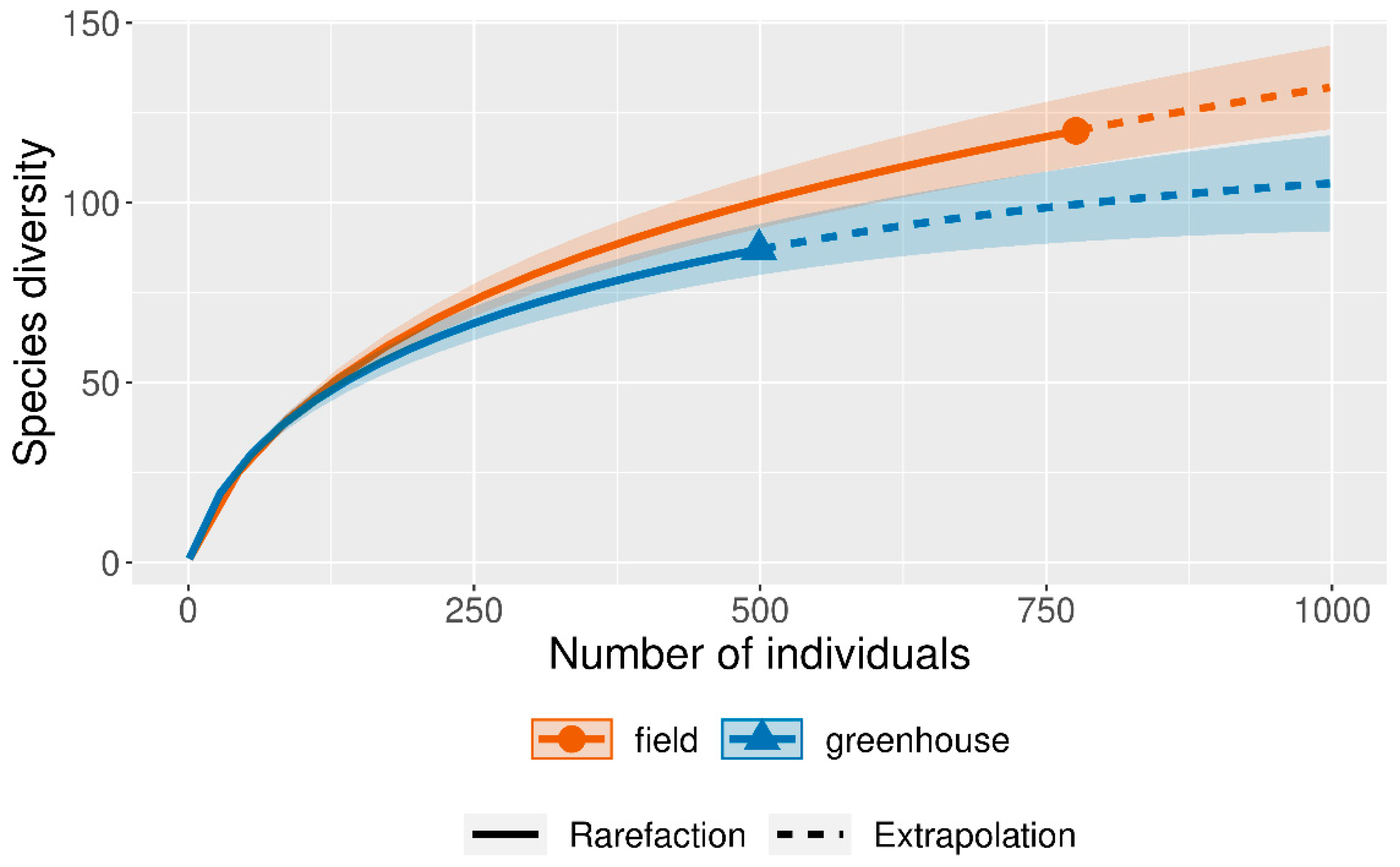
Figure 2.
Krona diagram of all isolated fungi.
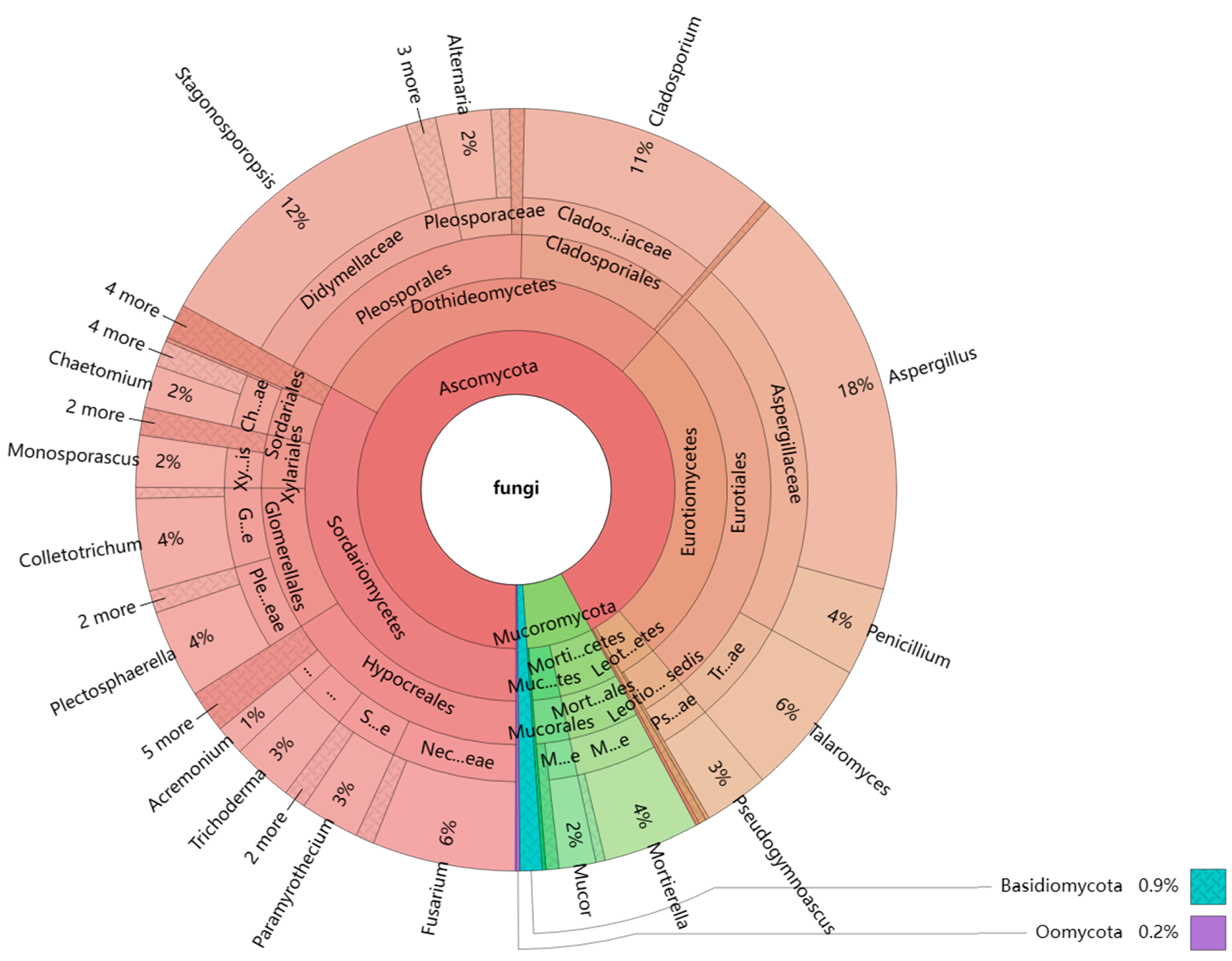
Figure 3.
Comparison among different media from cucumber plants and rhizosphere soil samples: (a) the number of fungal genera; (b) the number of fungal strains.
Figure 3.
Comparison among different media from cucumber plants and rhizosphere soil samples: (a) the number of fungal genera; (b) the number of fungal strains.
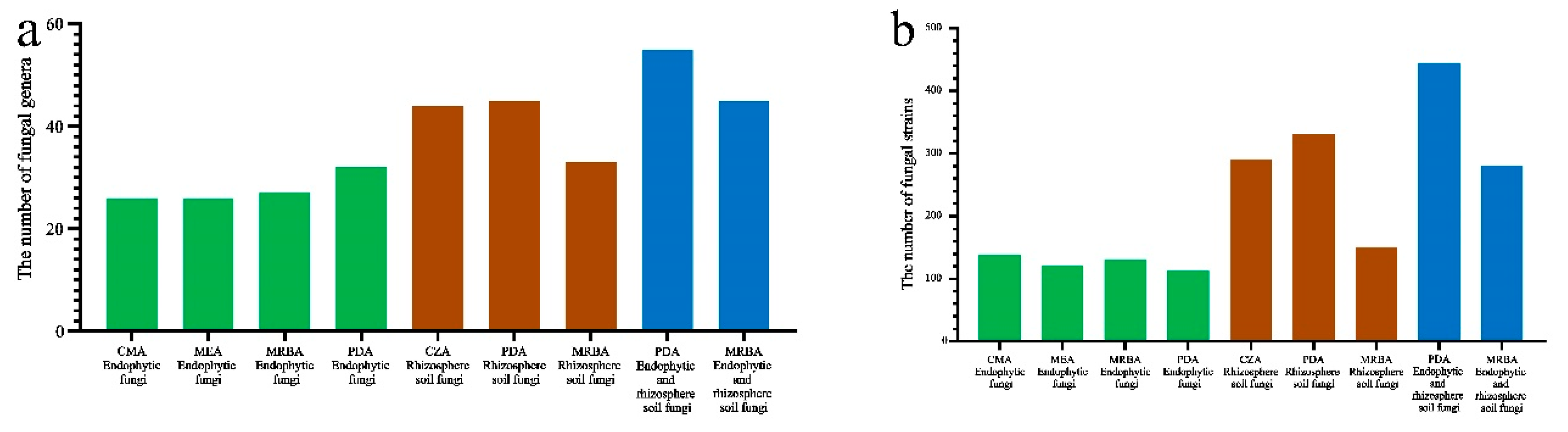
Figure 5.
Diversity indices of fungi isolated with different media. (a), (b), (c): The number of genera, Shannon index and Simpson index of endophytic fungi isolated on different media; (d), (e), (f): The number of genera, Shannon index and Simpson index of soil fungi isolated on different media; box graph data are all mean ± s.d. (n=4); a, b = same letter indicating no significant difference when p<0.05).
Figure 5.
Diversity indices of fungi isolated with different media. (a), (b), (c): The number of genera, Shannon index and Simpson index of endophytic fungi isolated on different media; (d), (e), (f): The number of genera, Shannon index and Simpson index of soil fungi isolated on different media; box graph data are all mean ± s.d. (n=4); a, b = same letter indicating no significant difference when p<0.05).
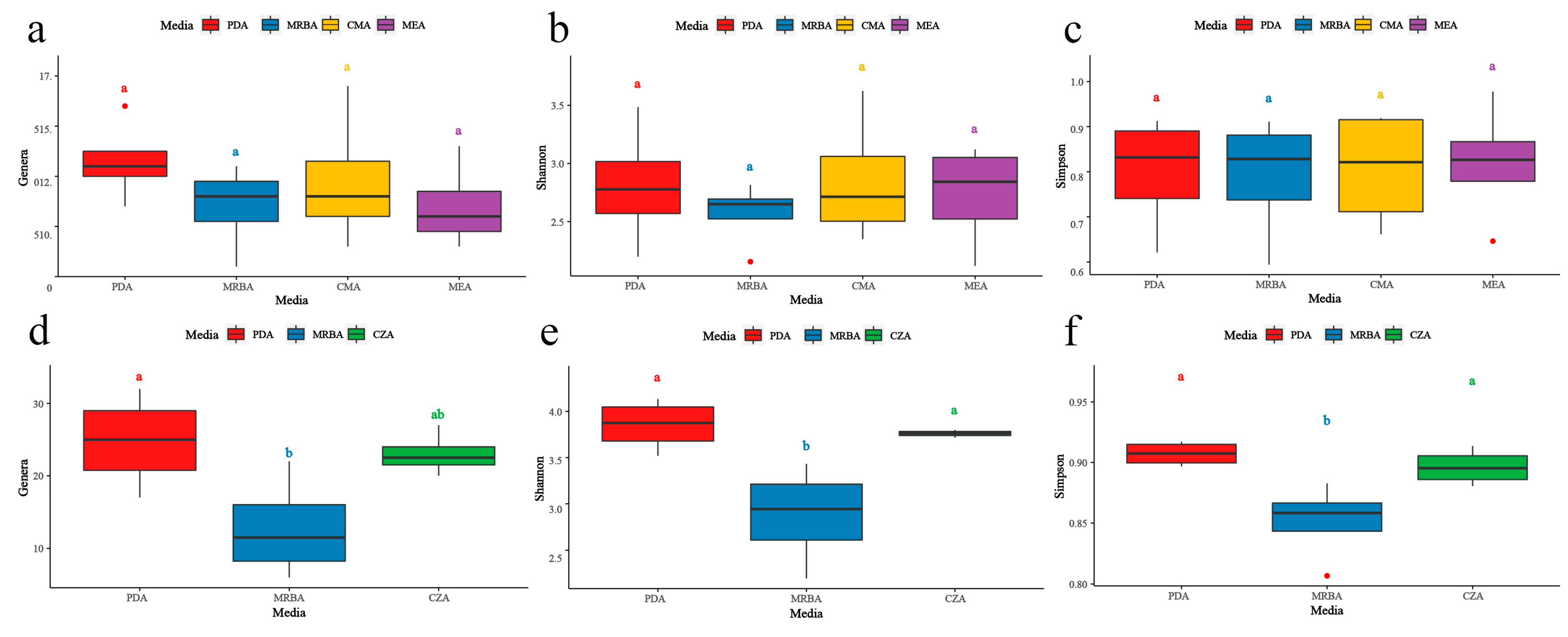

Table 1.
Details of collected cucumber plants and their corresponding rhizosphere soil
| Collection Time | Location | Growth Stage | Plants Number | Soil Number | Treatment-Row/Pot Number |
|---|---|---|---|---|---|
| Jul. 20, 2022 | field | seedling | 16 | 16 | CC-4; CT-4; CF-4; TF-4 |
| Aug. 5, 2022 | greenhouse | seedling | 16 | 16 | |
| Aug. 14, 2022 | field | flowering | 16 | 16 | |
| Aug. 21, 2022 | greenhouse | Vine growth | 16 | 16 | |
| Jul. 20, 2022 | field | seedling | 16 | 16 | |
| total | 64 | 64 |
Table 2.
A total of 74 fungal genera and 145 species identified by BLAST search in this study.
| Strains No. | Species Name | Per.Ident | Accession No. in GenBank of Reference Strain | Strains No. | Species Name | Per.Ident | Accession No. in GenBank of Reference Strain | ||
|---|---|---|---|---|---|---|---|---|---|
| GSSCF15Z4-6 | Acremonium camptosporum | 97.49% | MH859415.1 | FPSCC39RR-1 | Globisporangium recalcitrans | 100.00% | KJ716861.1 | ||
| GSVCC22P3-7 | Acremonium felinum | 100.00% | AB540563.1 | GPSCC05RR-1 | Glomerella lagenaria | 100.00% | AJ301965.1 | ||
| GSVCC22P3-8 | Acremonium persicinum | 99.85% | MH864511.1 | GSSCC05P3-4 | Graphium basitruncatum | 98.45% | EF165016.1 | ||
| GSSCF14Z3-9 | Acremonium sclerotigenum | 97.68% | KJ194115.1 | GPVTF34RS-3 | Harknessia ellipsoidea | 97.24% | MH865961.1 | ||
| GPVCC21CR-2 | Acrocalymma vagum | 99.55% | KJ188710.1 | FSSCF48R3-14 | Humicola fuscoatra var. fuscoatra | 98.81% | MH863738.1 | ||
| FSSCC42P3-11 | Actinomortierella ambigua | 98.76% | JX976067.1 | GPSCC07PS-1 | Humicola variabilis | 99.09% | MH863772.1 | ||
| GSVCC21Z3-4 | Albifimbria verrucaria | 98.51% | KU845891.1 | FSSCC41E4-1 | Irpex lacteus | 99.62% | OL685330.1 | ||
| GPVCC22EL-1 | Alternaria alternata | 100.00% | LC440583.1 | FSSCC39P3-4 | Lectera colletotrichoides | 98.04% | AJ301962.1 | ||
| FSFCC58Z3-7 | Alternaria atra | 99.70% | MH864090.1 | FSFCF64P3-3 | Linnemannia gamsii | 97.30% | MH859222.1 | ||
| FSFCF65Z3-8 | Antarctomyces psychrotrophicus | 97.19% | NR_164292.1 | GPVCT24PR-2 | Macrophomina phaseolina | 100.00% | EF570500.1 | ||
| GSVCC21P4-2 | Apiospora rasikravindrae | 99.58% | KF144915.1 | GSVCC20Z3-2 | Malassezia restricta | 99.87% | EU400587.1 | ||
| FSFCT60Z3-6 | Aspergillus alabamensis | 97.77% | MH863630 | GSSCC06Z3-9-2 | Marquandomyces marquandii | 97.24% | AB099511.1 | ||
| FSSCF50P3-6 | Aspergillus baeticus | 99.42% | MT990731.1 | FSFCT62P3-8-1 | Metarhizium anisopliae | 99.54% | FJ545329.1 | ||
| GSSCF14Z3-12 | Aspergillus clavatus | 97.24% | NR_121482.1 | GSSCC06Z3-2 | Meyerozyma guilliermondii | 99.88% | MH545918.1 | ||
| FSFCC58P3-1 | Aspergillus eucalypticola | 99.86% | OL711732.1 | FSFCF64Z3-4 | Microascus brevicaulis | 99.85% | MT732896.1 | ||
| FSSCC40Z4-15 | Aspergillus flavus | 100.00% | MH279408.1 | GPSCC04PS-1 | Microdochium musae | 97.14% | MH107898.1 | ||
| GPSCC04RS-1 | Aspergillus heldtiae | 98.71% | MK450656.1 | FPSCF50CS-1 | Moesziomyces aphidis | 99.03% | JQ743064.1 | ||
| FSFCC56Z3-7 | Aspergillus minisclerotigenes | 100.00% | MH279386.1 | GPVCC20PS-4 | Monosporascus cannonballus | 99.84% | JQ771932.1 | ||
| GPSCC05PS-1 | Aspergillus niger | 99.86% | MT990733.1 | FPFCT61CR-2 | Monosporascus ibericus | 97.53% | JQ973832.1 | ||
| FSSCT43Z4-7 | Aspergillus nomiae | 99.71% | MH279387.1 | GSSCC04Z3-9 | Mortierella alpina | 99.87% | AJ271630.1 | ||
| GSSCT09P3-5 | Aspergillus pallidofulvus | 98.81% | MK450639.1 | FPFCC58ER-3 | Mucor aseptatophorus | 97.89% | MZ433252.1 | ||
| FSSCC42Z3-7 | Aspergillus pseudodeflectus | 99.25% | MK450642.1 | FSSCC42P3-19 | Mucor circinelloides | 99.37% | MH854919.1 | ||
| FSSCC41Z4-1 | Aspergillus pseudonomiae | 97.29% | MH279417.1 | FSSCC41E4-2 | Mucor fragilis | 99.85% | GU566275.1 | ||
| GSSCC06Z3-5 | Aspergillus seifertii | 97.37% | MK450648.1 | GSVCC23P3-7 | Mucor hiemalis | 100.00% | MF782793.1 | ||
| FSFCC57Z3-3 | Aspergillus sigurros | 97.77% | MK450650.1 | GSVCF31Z3-6 | Nalanthamala vermoesenii | 97.12% | JX456473.1 | ||
| GPVCC23PS-2 | Aspergillus sydowii | 99.70% | MT990722.1 | FSFTF68Z3-3 | Ochroconis tshawytschae | 90.06% | MH858782.1 | ||
| GSSCC04P3-2 | Aspergillus uvarum | 100.00% | OL711726.1 | FSFCF64P3-4 | Paraisaria heteropoda | 100.00% | AB084157.1 | ||
| GPSCC05PL-2 | Aureobasidium melanogenum | 99.85% | MH863678.1 | GSVCC23Z3-6 | Paramyrothecium humicola | 99.71% | MH864508.1 | ||
| FSSCC39Z3-1 | Bisifusarium delphinoides | 99.82% | KP132212.1 | GSVCF31P3-8 | Paramyrothecium roridum | 99.66% | HQ115647.1 | ||
| FSSCF48P3-1 | Boeremia exigua var. exigua | 97.48% | EU343168.1 | FSFTF70P3-2 | Paramyrothecium viridisporum | 99.69% | MH864343.1 | ||
| FPFTF68CR-1 | Boeremia exigua var. populi | 97.24% | MN973161.2 | FSSCC39Z3-11 | Penicillium aurantiocandidum | 100.00% | MH861314.1 | ||
| FSSCC42P3-6 | Calonectria ciliata | 97.82% | MH864520.1 | FSFCF65R4-2 | Penicillium cerradense | 98.53% | MT006127.1 | ||
| FPFCF64PS-3 | Cephalotheca sulfurea | 97.71% | AB278194.2 | GPVCC21RS-3 | Penicillium citrinum | 100.00% | MH858380.1 | ||
| GPVCC23ER-1 | Ceratobasidium chavesanum | 97.44% | NR_164016.1 | FSSTF54Z3-5 | Penicillium commune | 97.25% | GQ458026.1 | ||
| FSSCF50R3-2 | Chaetomidium leptoderma | 97.33% | NR_164219.1 | GSVCC21Z3-1 | Penicillium copticola | 99.84% | NR_121516.1 | ||
| FPSCC40ER-2 | Chaetomium elatum | 98.65% | MH871792.1 | FSSCF49P3-11 | Penicillium daleae | 96.56% | MH854984.1 | ||
| FPSCC41RS-4 | Chaetomium globosum | 99.85% | AB449671 | FSSCC41R3-10 | Penicillium flavigenum | 99.85% | MH862182.1 | ||
| FSSCF48Z3-3 | Chaetomium perlucidum | 99.84% | MH857726.1 | FSSCC40Z4-2 | Penicillium goetzii | 99.41% | MH859954.1 | ||
| GSSCC04P3-5 | Chaetomium piluliferum | 99.85% | MH861633.1 | FSSCC41Z3-6 | Penicillium lapidosum | 98.24% | MH860150.1 | ||
| GPSCC07RR-2 | Cladosporium acalyphae | 100.00% | MH863861.1 | FSFCC58P3-4 | Penicillium nothofagi | 99.13% | MH864386.1 | ||
| FSFCC58Z3-3 | Cladosporium anthropophilum | 99.85% | MF574171.1 | FSFCF65R3-2 | Penicillium oxalicum | 98.97% | LC386216.1 | ||
| GPSCT08PL-1 | Cladosporium dominicanum | 99.85% | MF472969.1 | GSVCC20P3-3 | Penicillium senticosum | 99.41% | MH860152.1 | ||
| GSSCC04P3-4 | Cladosporium halotolerans | 99.85% | MF473103.1 | FSSCC40Z3-7 | Penicillium sizovae | 99.70% | MH858522.1 | ||
| GSSCC06P3-11 | Cladosporium parahalotolerans | 99.85% | MF473160.1 | FSSCC41R3-13 | Phoma herbarum | 98.91% | AY337712.1 | ||
| GSSCC06Z3-7 | Cladosporium sphaerospermum | 99.70% | MF473271.1 | GPSCC04ES-1 | Plectosphaerella cucumerina | 98.58% | MH858371.1 | ||
| GPSCC04RR-2 | Cladosporium tenuissimum | 99.85% | MH864840.1 | GPVCC21ER-3 | Preussia terricola | 97.78% | MH858589.1 | ||
| GSVCC21Z3-3 | Cladosporium velox | 100.00% | MF473310.1 | GSSCC04P3-6 | Pseudogymnoascus pannorum | 99.85% | MH864459.1 | ||
| GPVTF35EL-4 | Cladosporium westerdijkiae | 100.00% | MF473314.1 | FSSCC41P3-5 | Purpureocillium lilacinum | 99.58% | AB103380.1 | ||
| GPVTF33PL-1 | Cladosporium xanthochromaticum | 99.54% | MF473316.1 | GSSTF18Z4-3 | Pyricularia oryzae | 97.13% | CP034204 | ||
| GPVTF33RL-2 | Cladosporium xylophilum | 100.00% | MH863875.1 | FPSCF50PS-1 | Pythium amasculinum | 99.68% | AY598671.2 | ||
| GSSCC06Z3-4 | Clonostachys rosea | 99.41% | KX958035.1 | GPVCC22ES-2 | Retroconis fusiformis | 97.66% | EU040239.1 | ||
| FSSCC41R3-14 | Collariella gracilis | 97.45% | MH864437.1 | FPSCC42CR-2 | Rhizoctonia solani | 97.01% | MH862760.1 | ||
| FPSCC40ES-2 | Colletotrichum dematium | 99.56% | AJ301954.1 | FSSCF49P4-2 | Rhizopus arrhizus | 99.05% | AB109754.1 | ||
| GPSCC05CL-1 | Colletotrichum gloeosporioides | 98.51% | AJ301908.1 | GSSCF12Z3-15 | Sarocladium kiliense | 99.66% | MF682447.1 | ||
| GSVCC22R3-2 | Coniochaeta fasciculata | 99.07% | MH855947.1 | FSSCF49P3-15 | Schizophyllum commune | 99.23% | AF350925.1 | ||
| FSSCC41P3-7 | Coniochaeta mutabilis | 99.23% | MH856122.1 | FPSTF51PS-4 | Stagonosporopsis caricae | 100.00% | MH863092.1 | ||
| GSVCC23P3-5 | Cunninghamella echinulata | 97.36% | GQ221208.1 | FSSCC39Z3-13 | Stagonosporopsis cucurbitacearum | 98.11% | EU167573.1 | ||
| GPSCF13EL-1 | Curvularia americana | 97.03% | NR_146239.1 | GSSCF13Z3-9 | Striaticonidium brachysporum | 99.84% | MH860035.1 | ||
| FSSCF50R3-10 | Didymella pomorum | 99.06% | MN983931.1 | GSSCC04P3-7 | Talaromyces amestolkiae | 100.00% | MH856395.1 | ||
| FPSCC40CS-2 | Didymella rosea | 99.63% | KT287020.1 | FPFCC55RS-3 | Talaromyces calidominioluteus | 97.93% | NR_175199.1 | ||
| GPSCC07CR-1 | Edenia gomezpompae | 97.79% | NR_156217.1 | FSFCF65P3-11 | Talaromyces liani | 100.00% | MH858781.1 | ||
| FSFCC55P4-3 | Engyodontium album | 97.93% | HM214541.1 | GSVCC20P3-5 | Talaromyces macrosporus | 97.03% | MH860495.1 | ||
| FSSCC42Z3-18 | Fusarium anthophilum | 97.41% | MH864506.1 | GSSCC06Z3-9-1 | Talaromyces pinophilus | 100.00% | CP017345.1 | ||
| GSSCC04P3-10 | Fusarium falciforme | 99.85% | MT251174.1 | FSSCF47P3-13 | Talaromyces purpureogenus | 97.59% | KM086709.1 | ||
| FSFCC57P3-5 | Fusarium fujikuroi | 99.85% | CP023090.1 | FSSCF47R3-4 | Thelonectria olida | 98.66% | MH857841.1 | ||
| FPSCF47CS-1-2 | Fusarium graminearum | 99.69% | HG970335.2 | GPSCC07RR-1 | Trametes hirsuta | 97.64% | AF516556.1 | ||
| GSVCC22R3-5 | Fusarium napiforme | 99.00% | MH862670.1 | FSSCC40Z3-11 | Trichocladium griseum | 98.79% | KU705826.1 | ||
| GPVTF34ER-1 | Fusarium oxysporum | 99.85% | LC383471.1 | GSSCC05P3-9 | Trichoderma harzianum | 97.82% | MF782824.1 | ||
| FSFCC58Z3-6 | Fusarium solani | 100.00% | MH864517.1 | FSSCC41R3-12 | Trichoderma pleuroticola | 98.49% | MH864423 | ||
| GPSCC04ES-2 | Fusarium tricinctum | 99.80% | MH931273.1 | FSFCC58Z3-2 | Trichoderma pseudokoningii | 99.72% | FJ605099.1 | ||
| FSSCC41R3-11 | Fusicolla acetilerea | 99.82% | EU860058.1 | GPSCC07RR-1 | Trichoderma simmonsii | 97.18% | CP075868.1 | ||
| FSFCF66Z3-4 | Geomyces asperulatus | 97.13% | MH861038.1 | FSSCF50P4-5 | Umbelopsis isabellina | 97.78% | MZ078794.1 | ||
| FSSCC42Z3-15 | Gibellulopsis nigrescens | 99.85% | HE972037.1 | ||||||
Table 3.
Fungal genera isolated by each medium.
| Fungal Type | Media | Number of Genera | Strain Number | Specific Genera compared to PDA |
|---|---|---|---|---|
| endophytic fungi(total 44 genera 504 strains) | CMA | 26 | 138 | Acrocalymma; Boeremia; Clonostachys; Graphium; Moesziomyces1; Mucor; Paramyrothecium |
| MEA | 26 | 121 | Ceratobasidium; Curvularia; Mucor; Paramyrothecium; Preussia | |
| rhizosphere soil fungi(total 61genera 771strains) | CZA | 45 | 290 | Antarctomyces; Bisifusarium; Edenia; Geomyces; Marquandomyces; Meyerozyma; Microascus; Monosporascus; Nalanthamala; Ochroconis; Pyricularia; Sarocladium; Striaticonidium; Trichocladium |
| endophytic fungirhizosphere soil fungitotal fungi(74genera 1275strains) | PDA | 32 | 114 | - |
| 45 | 331 | |||
| 55 | 445 | |||
| endophytic fungi | MRBA | 27 | 131 | Globisporangium; Graphium; Harknessia; Mucor; Trametes |
| rhizosphere soil fungi | 33 | 150 | Chaetomidium; Irpex; Monosporascus | |
| total fungi | 45 | 281 | Chaetomidium; Globisporangium; Harknessia; Irpex;Trametes |
1 Bold and underline fonts indicate the specific genera isolated only by this media.
Table 4.
Fungal genera with high isolation efficiency.
| Genera | Isolated with 5 types of media | Isolated with 3 types of rhizosphere soil fungi culture media | Isolated with 4 types of endophytic fungi culture media | Isolated from roots, stems and leaves of cucumber plants | |
|---|---|---|---|---|---|
| 1 | Acremonium | + | |||
| 2 | Acrocalymma | + | |||
| 3 | Albifimbria | + | |||
| 4 | Alternaria* | + | + | + | + |
| 5 | Apiospora | + | |||
| 6 | Aspergillus | + | + | + | + |
| 7 | Chaetomium | + | + | + | |
| 8 | Cladosporium | + | + | + | + |
| 9 | Clonostachys | + | |||
| 10 | Colletotrichum | + | + | + | + |
| 11 | Coniochaeta | + | + | ||
| 12 | Edenia | + | + | ||
| 13 | Fusarium | + | + | + | + |
| 14 | Humicola | + | |||
| 15 | Lectera | + | |||
| 16 | Malassezia | + | |||
| 17 | Microdochium | + | + | ||
| 18 | Monosporascus | + | + | + | |
| 19 | Mortierella | + | |||
| 20 | Mucor | + | + | ||
| 21 | Paramyrothecium | + | + | ||
| 22 | Penicillium | + | + | + | + |
| 23 | Phoma | + | |||
| 24 | Plectosphaerella | + | + | + | + |
| 25 | Pseudogymnoascus | + | + | + | |
| 26 | Stagonosporopsis | + | + | + | + |
| 27 | Talaromyces | + | + | + | + |
| 28 | Trichoderma | + | + | + | + |
*Bold fonts indicate the fungal genera isolated on both cucumber endophytic and rhizosphere soil samples.
Table 5.
The number of genera of endophytic fungi isolated in different plant parts with each medium.
Table 5.
The number of genera of endophytic fungi isolated in different plant parts with each medium.
| Genus | CMA | MEA | PDA | MRBA |
|---|---|---|---|---|
| root | 5.50a AB1 | 2.50b B | 3.00b AB | 6.50a A |
| stem | 7.75a AB | 6.25a AB | 8.75a B | 4.50a A |
| leaf | 4.25a A | 3.75ab A | 4.75b A | 3.25a A |
1The same row marked with different lowercase letters indicates significant differences between groups, and the same row marked with different capital letters indicates significant differences between groups.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



