Submitted:
03 June 2023
Posted:
05 June 2023
You are already at the latest version
Abstract
Pseudomonas aeruginosa (P. aeruginosa) is an opportunistic pathogen and is the leading cause of infection in patients with cystic fibrosis (CF). The ability of P. aeruginosa to evade host responses and develop into chronic infection causes significant morbidity and mortality. Several mouse models have been developed to study chronic respiratory infections induced by P. aeruginosa, with the bead agar model being the most widely used. However, this model has several limitations, including the requirement for surgical procedures and high mortality rates. Herein, we describe novel and adapted biologically relevant models of chronic lung infection caused by P. aeruginosa. Three methods are described, a clinical isolate infection model, utilizing isolates obtained from patients with CF, an incomplete antibiotic clearance model, leading to bacterial bounce-back and the establishment of chronic infection, and an adapted water bottle chronic infection model. These models circumvent the requirement for surgical procedure and importantly, can be induced with clinical isolates of P. aeruginosa and in wild-type mice. We also demonstrate successful induction of chronic infection in the transgenic βENaC murine model of CF. We envisage that the models described will facilitate the investigations of host and microbial factors, and the efficacy of novel antimicrobials, during chronic P. aeruginosa respiratory infections.
Keywords:
Pseudomonas aeruginosa
; chronic infection
; respiratory
; murine model
; in vivo
; S. aureus
1. Introduction
Cystic fibrosis (CF) is a life-shortening genetic disease caused by a mutation in a gene that encodes the cystic fibrosis transmembrane conductance regulator (CFTR) [1]. In CF patients, chronic infections caused by P. aeruginosa cause significant morbidity and mortality [1]. Initially, P. aeruginosa infections can be cleared by antimicrobial therapy. However, in most patients with CF, a transition to chronic infection by biofilm-forming mucoid strains of P. aeruginosa occurs, which persists, eventually resulting in respiratory failure [2]. Chronic infections in patients with CF are characterized by the development of P. aeruginosa specific antibodies, chronic inflammation dominated by neutrophils, and increased release of serine proteases causing lung tissue damage, in addition to the damage caused by the bacteria [2,3].
Over the last number of decades, several animal models have been developed in attempts to fully elucidate the bacterial pathogenesis during chronic infection caused by P. aeruginosa [4]. The administration of P. aeruginosa to the murine lung either causes acute infection and subsequent sepsis or is rapidly cleared from the lung [5]. Chronic infection has been achieved by intratracheal instillation using an immobilizing agent. Cash et al. established a chronic model of respiratory infection with P. aeruginosa utilizing agar beads administered intratracheally which resulted in infection lasting up to 35 days [6]. The embedding of P. aeruginosa within agar beads facilitates bacterial retention thus, resembling bacterial biofilm formation, as evident in chronic infection. This approach is, however, technically challenging, necessitates the use of surgical procedures, injectable anesthetic agents, and has been reported to have mortality rates of up to 45% [7]. Hoffman et al. utilized a stable mucoid CF sputum isolate with hyperproduction of alginate to develop a chronic model of P. aeruginosa lung infection. The hyperproduction of alginate was induced by deletions in mucA and functional N-acylhomoserine lactone-based quorum sensing systems; when instilled in mice, they could cause infections that could last for up to 7 days [8]. In another approach, plastic tubes precoated with P. aeruginosa inserted into the bronchus were successfully used to establish chronic infection which lasted up to 120 days [9]. More recently, Bayes et al. developed and adapted a murine agar bead model, using a clinical mucoid strain to induce features of transition from transitory to chronic airway infection [10]. However, it has been reported that bead size affects the induced response in this model, causing significant differences in inflammatory responses in the lung [11]. Furthermore, it has been reported that these models can result in chronic infection in the conducting airways due to blockage of the bronchi with beads.
To date, murine models of chronic P. aeruginosa infection without the use of artificial embedding have not been extensively reported. Novel animal models that recapitulate chronic infections caused by P. aeruginosa are urgently warranted to fully understand host pathogen-interactions and to facilitate the development of new therapeutic targets. Here, we describe experimental conditions to establish biologically relevant murine models of chronic lung bacterial infection. Specifically, we report the establishment of chronic infection through the selection of clinical isolates, selection of murine strain and optimization of infection regime. Moreover, we report an incomplete antibiotic clearance model and an adapted oral challenge water bottle [12]. Importantly, our models circumvent the requirement of artificial materials, overcoming challenges associated with currently reported models.
2. Materials and Methods
Mice
Wild type mice were either purchased from Inotivco (A/J, BALB/c) or Charles River (CD-1, FVB/N, SJL, Biozzi, C57BL6), or then bred on-site in the Biological Services Unit in Queen’s University Belfast. Both male and female βENaC mice [13,14,15,16] between 8-18 weeks, and all groups were sex matched. Mice were housed in groups in individually ventilated cages with 12h light and dark cycles and fed water and diet ad libitum. All animals were randomly assigned to treatment groups, using the graph pad prism tool (https://www.graphpad.com/quickcalcs/randomize1.cfm). Mice were handled using Biosafety Level 2 practices.
Biofilm Assay
Biofilm formation of bacterial strains was quantified using crystal violet staining. Briefly, 1 ml of overnight cultures were washed, resuspended in TSB supplemented with 1% glucose, and adjusted to an OD600 of 0.1. The culture suspensions were added to the wells of a 48-welltissue culture plate (Sarstedt) and incubated overnight without agitation at 37ºC. Subsequently, all media was removed from wells, then 150µl of methanol was added and the plates were incubated for 20 min at room temperature. The methanol was then discarded, once the plate had air dried 150 µl of 20% w/v crystal violet was added to each well for 5 min. Plates were washed thoroughly with water and left to dry completely. The remaining crystal violet was eluted with 150 µl of glacial acetic acid and measured at an OD595.
Intra-nasal infection of mice
Mice were infected intranasally as previously described [17]. Mice were infected with clinical isolates of P. aeruginosa (provided by the Clinical Microbiology Laboratory in the Royal Victoria Hospital) or the commonly used laboratory strain PAO1 (20). The clinical isolates of Pseudomonas were passaged through CD1 mice three times prior to use in these experiments. The bacteria were grown to mid-log phase in nutrient broth and washed extensively using endotoxin-free sterile phosphate buffered saline (PBS). They were then resuspended in endotoxin-free sterile PBS and diluted to the appropriate optical density (OD600). The number of colony forming units (CFU) was counted by plating serial dilutions of P. aeruginosa on nutrient agar plates. Mice were anesthetised with a mixture of Rompun-ketamine injected intraperitoneally. Once anesthetised, mice were held up-right and 20µL of bacterial suspension in endotoxin-free PBS was dropped onto alternate nostrils and inhaled. Mice were held in an up-right position to allow the suspension to reach the lungs until their breathing rate to return to normal. Eye gel (Medicom Healthcare, UK) was applied, and mice were gently placed into the cages with a bedding “pillow” propped beneath their chin to maintain their airway. Animals were monitored regularly to determine if they displayed any signs of illness including ruffled fur/hunched posture, significantly abnormal respiration and mobility reduced to a few steps. If mice displayed any two of these symptoms or had weight loss greater than 20% of pre-procedure body weight, they were culled. At predetermined time-points, mice were given a terminal dose of Rompun-ketamine anaesthetic and once fully unresponsive, exsanguination was completed by cardiac puncture, lung and nasal associated lung tissue (NALT) were collected. Tissue was homogenised by passing through a cell sieve, or mechanically processed using a Precellys Evolution (Thermotech) with 1 cycle of 10 s at 4500 rpm. The homogenate was either serially diluted and plated onto selective agar to quantify the bacterial burden or centrifuged and the cell pellet used to analyse the cell populations present by flow cytometry and the supernatant was frozen for subsequent cytokine analysis.
Staphlococycuccus aureus assessment
Staphylococcal strains were cultured at 37ºC in Brain Heart Infusion (BHI) (Oxoid). To evaluate protease activity, 5μlof each strain adjusted to an OD600of 1.0 was spotted onto BHI containing 1% skimmed milk. Plates were incubated at37ºC for 48 hours and positive protease activity was recorded based on zones of clearance in the media. The haemolytic activity of strains was evaluated on sheep blood agar plates incubated at 37ºC for 48 hours. Carotenoid pigment production was assessed for each strain by pelleting stationary phase cells by centrifugation. Production of the pigment was based on colour of the pellet, where white represented negative, and orange represented positive. To assess antibiotic susceptibility the Staphylococcal strains were sub-cultured in BHI and grown to logarithmic phase (0.6-0.7 OD600) at 37ºC, washed twice and resuspended in phosphate buffered saline (PBS) (Sigma). Cell suspensions (300 μl) were spread onto BHI agar and left to dry for 10 min. Antibiotic E-test strips (Biomerieux) were overlaid and plates were incubated at 37ºC for 24 hours. Minimum inhibitory concentrations (MIC) (μg/ml) for each antibiotic were recorded in triplicate for each strain.
Antibiotic treatment
Mice were weighed and aneasthetized. Once sedated (as to not have the pedal withdrawal reflex), 20 µl of PAO1 was administered intranasally (i.n) (low dose 1x105 or high dose 1x106). The mice were left for 1 h and then tobramycin was administered by the scireqInExpose nebulisation tower. Mice were treated daily for 7 days in order to achieve complete bacterial clearance. Animals were sacrificed to assess bacterial burden at predetermined time points, lung, NALT and fecal samples were collected.
Infection via the drinking water
Mice were administered 1x107CFU’s/mL PAO1 in their water bottle, the bacterial culture was changed every day for a fresh inoculum. After 5 days, the mice were given filtered water for the remainder of the study, prior to being sacrificed.
Cytokine measurements
Cytokine levels were measured in lung homogenate supernatant using ELISAs as per manufacturer’s instructions. IL-6 and IL-1β were from R&D Systems, IFNɣ and GM-CSF were from eBiosciene/Thermotech.
Flow cytometry
Flow cytometry was performed as previously described [18,19]. Lung tissue was homogenised by passing through a cell sieve, the cells were obtained by spinning the homogenate at 600 x g for 10 min at room temperature. The red blood cells were lysed with ammonium-chloride-potassium buffer and the cells were washed and counted using the Eve automated cell counter (NanoEntek, USA). The cells were incubated for 10 mins in foetal calf serum (FCS) (100 µL) to block the Fc receptors. The cells were washed with 100 µL flow buffer (PBS, 10% FCS, 0.1% EDTA) centrifuged at 300 x g for 10 mins at room temperature, followed by extracellular and intracellular antibody staining. To identify neutrophils and macrophages, cells were stained with CD45 FITC, CD11b e660, CD11c e450, GR-1 PE and F4/80 PE Cy7 (eBiosience). After incubation, the cells were washed twice and fixed with 4% Paraformaldehyde (PFA). The fixed cells were stored at 4oC and acquired within 48 h on a FACS Canto II flow cytometer (BD Biosciences, UK), analysis was performed with FlowJo software (Tree Star Inc., USA).
Statistical analysis
All CFU data was logarithmically transformed prior to analysis. GraphPad Prism (Version 9.4.1) was used to determine statistical significance between groups of mice. To compare two groups, an unpaired t-test was carried out, to compare three or more groups, an ANOVA test, with Šídák’s correlation for post-hoc multiple comparison applied. The duration of survival was calculated by Kaplan–Meier analysis, and statistical significance was determined with the log rank (Mantel–Cox) test. Error bars represent standard error of mean (SEM). The cut-off for statistical significance between groups was p<0.05.
3. Results
3.1. Clinical isolate chronic infection model
A major challenge in establishing chronic lung infection models is finding the balance between bacterial clearance and sepsis. Several different strains of P. aeruginosa have previously been used to develop CF-like lung infections, with the laboratory strain PAO1, initially isolated from a wound infection, being the most commonly used strain [20]. However, PAO1 is more virulent than many isolates from patients with CF and has a faster replication. Moreover, the motility of PAO1 in vivo has been reported to differ from that of clinical isolates from patients with CF [21]. Accordingly, we examined several clinical isolates to determine their ability to establish chronic lung infection, without the necessity for embedding material. A range of isolates obtained from the sputum of patients with and without CF, in addition to those isolated from patients within the intensive care unit (ICU) were evaluated.
Similar to previously reported studies [21], there was considerable variability in the virulence of the isolates, with 3 isolates significantly reducing survival rates of outbred CD1 mice, in comparison to infection with PAO1 infected mice (Figure 1A). In line with previously reported studies by Kukavica et al. [21], increased virulence corresponded with increased in vitro biofilm formation (Figure 1B), except for isolates Q502 and Q511. For Q511, this appears to be largely due low levels of alginate production in vivo (Figure 1C. Isolate Q502, obtained from the sputum of a patient with CF, appeared to be distinct from the other isolates, producing high levels of biofilm, both in vitro (Figure 1B) and in vivo (Figure 1C) but not causing significant mortality in the mice. This isolate was thus, selected for use to establish a chronic infection model.
In patients with CF, the acquisition of P. aeruginosa is often preceded by S. aureus infection [22,23,24]. We therefore sought to investigate if pre-infection with S. aureus influenced the susceptibility of the lungs to the development of chronic P. aeruginosa infection. As before, a range of clinical isolates were obtained from the sputum of patients with CF, were utilised and compared to that of the commonly used Newman laboratory strain of S. aureus. Infection of CD1 mice with S. aureus, 10 days prior to infection with P. aeruginosa, resulted in a significant reduction in survival during PAO1 infection (Figure 2A), with the exception of isolate Q154 which enhanced survival (p= 0.04). Whilst the most significant effects on PAO1 infection were between pre-treatment with isolate Q154 and Q181, phenotypic characterisation of the isolates demonstrated no significant differences (Supplementary Figure 1 and Supplementary Table S1).
Whilst many murine chronic infection models utilise C57BL6 or BALBc mice, it is well established that the murine strain selected for infection can have a significant impact on the outcome of a wide variety of infection models [25,26,27,28]. We therefore aimed to determine which mouse strain would be most permissive to the establishment of chronic infection (Figure 2B) with Q502. All strains rapidly succumbed to acute infection, the exception to this was Biozzi mice, in which 80% of mice were still alive at 144h post infection (Figure 2B). When Biozzi mice were pre-inoculated with S. aureus (Q154), 10 days prior to infection with P. aeruginosa, there was a significant reduction in P. aeruginosa bacterial burden in the lungs (p=0.02) (Figure 2C), and reduced levels of the pro-inflammatory cytokine IL-6 (Figure 1D). Pre-treatment with S. aureus also resulted in significant increased levels of IFNγ and GM-CSF in the lungs, while IL-1β was unaffected (Supplementary Figure S2A-C). Moreover, there were no significant differences in the numbers of neutrophils and macrophages recruited to the lungs of mice pre-infected with S. aureus, compared to those that were not pre-infected (Supplementary Figure S3A-B).
At 10 days post infection with S. aureus and prior to infection with P. aeruginosa, we confirmed that S. aureus was still present within the lungs (Figure 2E). Co-culture of S. aureus strains Q154 and Q502 with P. aeruginosa demonstrated delayed growth of P. aeruginosa, compared to that of when P. aeruginosa was cultured alone (Figure 2E) and statistical comparison of the AUC confirmed there was a significant impairment in growth (p=0.002).
Pre-inoculation of Biozzi mice with Q154, prior to high dose P. aeruginosa infection, significantly enhanced their survival (p=0.002) with 80% of the mice surviving to day 10 (Figure 2G). In Biozzi mice that received Q502 only, there was no detectable P. aeruginosa present in the lungs of mice surviving at day 14 (data not shown), suggesting they had cleared the infection. In contrast, in the animals pre-inoculated with Q154, there was a high burden of P. aeruginosa in 5 out 6 mice (average 3.4x103 CFU ± 1.2x103) at 14 days post-infection (Figure 2H).
3.2. Incomplete antibiotic clearance leading to chronic lung infection
In patients with CF, inhaled tobramycin is the standard treatment of chronic P. aeruginosa infection [29]. While generally effective in approximately 80% of patients, failed eradication is reported [30]. Utilising a clinically relevant treatment regimen of inhaled tobramycin, in our second model we aimed to develop a model of incomplete antibibiotic clearance. Mice were infected i.n with 1x106 CFU of PAO1 and subsequenly treated with tobramycin for 5 days by nebulisation [31].
As expected, administration of nebulised tobramyicin resulted in reduced bacterial burden in the lungs of mice 1 day post treatment. Bacterial burden continued to decrease until day 5, and by day 7 mice had completely cleared the infection, as evident by the clearance of bacteria from the lung. However, by day 14, CFU counts returned to a level similar to that found at 1 day post infection, and this was maintained up to 21 days post infection (Figure 3A). Analysis of CFU in the lung and faeces of mice, demonstrated that CFU was still detectable at 14 days post-infection (Figure 3B). This data demonstrates that a clinically relevant treatment regime of tobramycin nebulised therapy can be utilised in mice to induce chronic P. aeruginosa lung infection.
3.3. Waterbottle chronic infection model
We next sought to investigate if the addition of PAO1 to drinking water could establish chronic lung infection. Animals were exposed to the bacteria for 5 days, with a two day “wash-out” period to ensure that what we were detected was established infection, particularly in the upper respiratory tract, and not simply a mouse that had recently taken a drink. As can be seen, 7 days post administration there was a significant bacterial burden within the lungs of the mice (p=. Following this, the bacterial burden reduced until day 20, after which it began to increase (Figure 4A). As expected, CFU were detected in the NALT throughout all the time points tested (Figure 4B). Levels of IL-6 in the lungs of mice were significantly increased across all time points tested. Concurrent with the bacterial burden in the lungs, the highest levels of IL-6 were detectable at 7 days post infection (Figure 4C).
As previously alluded to, the current treatment regimen for treatment of P. aeruginosa infections with patients with CF is inhaled tobramycin [29]. However, P. aeruginosa infections often persist and reoccur. Thus, in our water bottle model, we administered mice tobramycin for 7 days by nebulization to determine if the infection could be cleared. In this model antibiotic infection cleared the chronic infection as there was no CFU detectable in the lungs of mice at day 44. In this model the mice remained clear following antibiotic treatment and, unlike treatment of high dose acutely infected mice (Figure 3), the treated mice still had no detected P. aeruginosa at day 50, this was not due to natural clearance as can be seen in the untreated mice at day 50 (Figure 4D).
The βENaC mouse model accurately recapitulates many of the pulmonary features of CF and is commonly used to investigate CF pathogenesis. It was demonstrated that the water bottle model could also be utilized to induce chronic P. aeruginosa infection in βENaC mice. As before, CFU was detected in the lungs of mice by day 7, which was maintained until day 50 (Figure 4E). Both βENaC and WT mice showed stable weight over the course of infection (Figure 4F), demonstrating that whilst the mice may have chronic infection and associated lung inflammation, they are not overtly symptomatic.
4. Discussion
The establishment of clinically relevant murine models of chronic lung infection has remained a challenge for many years and has hampered our understanding of the pathogenesis of P. aeruginosa infection in patients with CF. Administration of P. aeruginosa to the murine lung either causes acute infection and subsequent sepsis or is rapidly cleared from the lung and thus, cannot be used to establish chronic infection, as found in patients with CF [5]. Accordingly, alternative approaches have been developed for example, the embedment of P. aeruginosa within agar beads, or the coating of plastic tubes with P. aeruginosa for insertion into the bronchus [6,9]. However, such approaches are technically challenging, result in high mortality rates, and the agar bead model can induce varying inflammatory responses in the lung based on the beads used and their size [11]. In the present study, we describe for the first time three models of chronic lung infection which circumvent challenges associated with previously reported chronic models of P. aeruginosa chronic respiratory infection. These models allow infection to be sustained for longer time periods and importantly, the infections could be induced in both wild type mice and in βENaC transgenic mice [32].
By careful selection of clinical isolates of S. aureus and P. aeruginosa, we were able to establish chronic infection in Biozzi mice. Interestingly, a correlation between virulence and high biofilm formation was evident across the tested strains, with the exception of Q516. This isolate, however, demonstrated a significant reduction in the levels of alginate detected in vivo, compared to that of PAO1, demonstrating that in vitro biofilm assays may not fully recapitulate the in vivo conditions. This is in line with previously reported studies [33,34]. Jordan and co-workers reported nonconformity of biofilm formation with several strains of S. aureus in vitro and in vivo. Specifically, it was reported that deletion of the accessory gene regulator (agr) resulted in increased biofilm in vitro but did not induce increased biofilm in vivo or affect mortality rates in mice [34]. To date, there have been no reported studies comparing biofilm formation of isolates of P. aeruginosa both in vitro and in vivo. A number of studies have reported that murine strain can significantly impact he severity of infection against a number of bacteria [25,26,27,28]. In this model of PAO1 infection, Biozzi mice were most resistant, with 80% surviving at 144h post infection, while all other strains rapidly succumbed.
In patients with CF, both S. aureus and P. aeruginosa are key bacterial pathogens. While chronic infection with P. aeruginosa is accepted to be the leading cause of airway infection and is associated with poorer outcomes, the impact of S. aureus colonization on subsequent outcome to P. aeruginosa remains debated [35]. Cigana and co-workers utilized the agar bead model to infect mice with S. aureus, prior to infection with P. aeruginosa [35]. However, as previously described, the agar bead model has many disadvantages. We therefore sought to determine if pre-exposure to S. aureus would render the lungs more permissive to chronic infection by P. aeruginosa. Whilst there was significant variability in the impact of S. aureus isolates on the outcome of the subsequent P. aeruginosa infection, we were able to identify an isolate, Q154, which enhanced survival of the mice and resulted in the establishment of chronic infection. S. aureus was still maintained in the lungs at 10 days post infection, prior to infection with P. aeruginosa and this prompted us to determine if there was a direct interaction between the bacterial species. Interestingly, in vitro culture of S. aureus strains with P. aeruginosa demonstrated impaired growth of P. aeruginosa. This is in line with previously reported studies by Michelson and co-worker’s, who demonstrated that S. aureus altered the growth activity and autolysis of a range of P. aeruginosa isolated in vitro [36]. Further studies are warranted to fully interrogate the interspecies interactions between isolates of S. aureus and P. aeruginosa obtained from patients with and without CF and to fully understand host adaptation in the CF lung.
Whilst the model outlined above is highly biologically relevant and recapitulates many of the features of establishment of chronic lung infection seen in patients, it has its limitations. Firstly, in requires the use of specific clinical isolates, Q502 is highly antibiotic resistant and therefore genetic manipulation of the bacteria to either make it bioluminescent to monitor infection in real time or create specific mutants to investigate virulence factor is technically demanding. Furthermore, it requires Biozzi mice; the model does not readily transfer to C57BL6 models. Although Biozzi mice are commercially available, this is not a widely utilized strain of mice and transgenic models are not available on this background, increasing the time and costs required to use transgenics to investigate host-pathogen interactions.
The antibiotic treatment model was discovered serendipitously; however, it represents a biologically relevant model. The model uses C57BL6 mice making it applicable to study how immunological manipulation of the host will impact the establishment of chronic infection. Whilst here we have used PAO1, clinical isolates can also be used and both their clearance, bounce back, and ability to establish chronic infection can be easily investigated. It has recently been reported that antibiotic exposure induces both phenotypic and genotypic alterations in P. aeruginosa [37], this model would make an excellent tool to further examine this phenomenon. It is interesting to note that in mice treated with antibiotics following either low dose infection (Supplementary Figure S3) or infection via the water bottle either don’t show any bounce back or else do have sustained levels of bacteria within their lungs. This would suggest that there is a threshold level of bacteria required for chronic infection to be maintained. Singh et al. reported that there is a bacterial threshold within the lung for induction of inflammation [38], perhaps the inflammatory response contributes to the establishment of infection, mirroring the role that inflammation plays in driving microbial dysbiosis and the establishment of chronic infection within the gut.
It has previously been reported that the water bottle method of induction of chronic lung infection is only suitable for transgenic murine models of CF [20]. However, in our adapted protocol this is not the case, and we describe the successful establishment of chronic infection in wild-type mice. This model facilitates the investigation of spontaneous infection and the investigation of pathogen virulence or host factors that modulate the initial infection or the development of chronic infection. We also demonstrate successful induction of chronic infection in βENaC mouse model of CF, thus facilitating investigations into the role of chronic infection in CF pathogenesis. Whilst P. aeruginosa is identified as a pathogen of patients with CF, it is associated with other respiratory diseases including non-CF bronchiectasis and chronic obstructive pulmonary disease [39,40,41]. Moreover, it has been reported that persistent P. aeruginosa infection is a risk factor for chronic rejection in allograft lungs [42]. Therefore, our model in wild-type mice, rather than genetically modified mice, could also be utilized for the study of non-CF related chronic infections.
5. Conclusions
P. aeruginosa remains an important cause of chronic respiratory infections in CF and non-CF lung diseases. Its ability to evade host responses results in significant morbidity and mortality. The murine models described herein present reliable and useful models to study chronic P. aeruginosa. The models present all recapitulate different aspects of clinical disease, and, when combined, they provide an essential tool set to investigate the role of both pathogen and host factors that can modulate the establishment or persistence of chronic infection. These tools are essential to increase our understanding of the biological processes involved as well as identify novel therapeutic targets to tackle chronic infection.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org., Figure S1: Phenotypic characterization of S. aureus isolates; Table S1: Characterization of divergent clinical isolates of S. aureus; Figure S2 Increased levels of IFNγ with S. aureus pre-treatment, reduced G-CSF, and no difference in IL-1β – despite changes in cytokines and CFU, there is no significant difference in macrophages or neutrophils. Figure S3 Mice were infected intranasally with high (OD1 - 106) or low (OD0.1 - 105) dose PAO1, significantly less CFU was detected in lungs of the mice infected with the low dose (A). After low dose infection, followed by 5 days of antibiotic treatment complete clearance was achieved at days 7, whilst there was bounced back at day 14 however the level was not sustained at day 21.
Author Contributions
“Conceptualization, R.J.I, J.S.E, M.B.E, M.A.M, B.J.P; methodology, J.L, A.M, A.V.D; formal analysis, R.J.I, J.L, A.V.D, A.A.F; investigation, R.J.I, J.L, A.V.D, A.A.F; resources, R.J.I.; data curation, R.J.I.; writing—original draft preparation, A.M.R.; writing—review and editing, A.M.R, R.J.I.; visualization, A.M.R, R.J.I.; supervision, R.J.I.; project administration, R.J.I.; funding acquisition, R.J.I, J.S.E, M.B.E, M.A.M, B.J.P. All authors have read and agreed to the published version of the manuscript.”
Funding
The research leading to these results has received support from the Innovative Medicines 320 Imitative Joint Undertaking under grant agreement no 115721, resources of which are 321 compared of financial contribution from the European Unions’ seventh Framework 322 Programme (FP&/2007-20130 and EFPIA companies’ in-kind contribution and from the European Union’s Seventh Framework Programme (FP7/2007-2013) under CFMATTERS grant agreement no. 603038, the German Research Foundation (SFB-TR84, sub-project B08 and SFB-1449, project 431232613, sub-project Z02) and the German Federal Ministry of Education and Research (82DZL009B1).
Institutional Review Board Statement
All animal experiments were sanctioned and approved by the UK Home Office and Queen’s University Belfast Ethical Review Committee.
Data Availability Statement
We encourage all authors of articles published in MDPI journals to share their research data. In this section, please provide details regarding where data supporting reported results can be found, including links to publicly archived datasets analyzed or generated during the study. Where no new data were created, or where data is unavailable due to privacy or ethical restrictions, a statement is still required. Suggested Data Availability Statements are available in section “MDPI Research Data Policies” at https://www.mdpi.com/ethics.
Acknowledgments
The authors would like to thank the microbiology department at The Royal Victoria Hospital, Belfast for supplying the clinical Pseudomonas strain and Queen’s University Belfast Biological Support Unit for their technical aid and expertise.
Conflicts of Interest
“The authors declare no conflict of interest.”
References
- Bhagirath, A.Y.; Li, Y.; Somayajula, D.; Dadashi, M.; Badr, S.; Duan, K. Cystic fibrosis lung environment and Pseudomonas aeruginosa infection. BMC Pulm. Med. 2016, 16, 1–22. [Google Scholar] [CrossRef]
- Oluyombo, O.; Penfold, C.N.; Diggle, S.P. Competition in Biofilms between Cystic Fibrosis Isolates of Pseudomonas aeruginosa Is Shaped by R-Pyocins. mBio 2019, 10, e01828–18. [Google Scholar] [CrossRef]
- Moser, C.; Jensen, P. .; Thomsen, K.; Kolpen, M.; Rybtke, M.; Lauland, A.S.; Trøstrup, H.; Tolker-Nielsen, T. Immune Responses to Pseudomonas aeruginosa Biofilm Infections. Front. Immunol. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Kukavica-Ibrulj, I.; Levesque, R.C. Animal models of chronic lung infection with Pseudomonas aeruginosa: useful tools for cystic fibrosis studies. Lab. Anim. 2008, 42, 389–412. [Google Scholar] [CrossRef]
- Osbourn, M.; Rodgers, A.M.; Dubois, A.V.; Small, D.M.; Humphries, F.; Delagic, N.; Moynagh, P.N.; Weldon, S.; Taggart, C.C.; Ingram, R.J. Secretory Leucoprotease Inhibitor (SLPI) Promotes Survival during Acute Pseudomonas aeruginosa Infection by Suppression of Inflammation Rather Than Microbial Killing. Biomolecules 2022, 12, 1728. [Google Scholar] [CrossRef]
- A Cash, H.; E Woods, D.; McCullough, B.; Johanson, W.G.; A Bass, J. A rat model of chronic respiratory infection with Pseudomonas aeruginosa. . 1979, 119, 453–9. [Google Scholar] [CrossRef] [PubMed]
- van Heeckeren, A.M.; Schluchter, M.D. Murine models of chronic Pseudomonas aeruginosa lung infection. Lab. Anim. 2002, 36, 291–312. [Google Scholar] [CrossRef]
- Hoffmann, N.; Rasmussen, T.B.; Jensen, P.; Stub, C.; Hentzer, M.; Molin, S.; Ciofu, O.; Givskov, M.; Johansen, H.K.; Høiby, N. Novel Mouse Model of ChronicPseudomonas aeruginosaLung Infection Mimicking Cystic Fibrosis. Infect. Immun. 2005, 73, 2504–2514. [Google Scholar] [CrossRef] [PubMed]
- Yanagihara, K.; Tomono, K.; Sawai, T.; Hirakata, Y.; Kadota, J.; Koga, H.; Tashiro, T.; Kohno, S. Effect of clarithromycin on lymphocytes in chronic respiratory Pseudomonas aeruginosa infection. Am. J. Respir. Crit. Care Med. 1997, 155, 337–342. [Google Scholar] [CrossRef]
- Bayes, H.K.; Ritchie, N.; Irvine, S.; Evans, T.J. A murine model of early Pseudomonas aeruginosa lung disease with transition to chronic infection. Sci. Rep. 2016, 6, 35838. [Google Scholar] [CrossRef]
- Christophersen, L.J.; Trøstrup, H.; Damlund, D.S.M.; Bjarnsholt, T.; Thomsen, K.; Jensen, P. .; Hougen, H.P.; Høiby, N.; Moser, C. Bead-size directed distribution of Pseudomonas aeruginosa results in distinct inflammatory response in a mouse model of chronic lung infection. Clin. Exp. Immunol. 2012, 170, 222–230. [Google Scholar] [CrossRef]
- Coleman, F.T.; Mueschenborn, S.; Meluleni, G.; Ray, C.; Carey, V.J.; Vargas, S.O.; Cannon, C.L.; Ausubel, F.M.; Pier, G.B. Hypersusceptibility of cystic fibrosis mice to chronic Pseudomonas aeruginosa oropharyngeal colonization and lung infection. Proc. Natl. Acad. Sci. 2003, 100, 1949–1954. [Google Scholar] [CrossRef]
- Johannesson, B.; Hirtz, S.; Schatterny, J.; Schultz, C.; Mall, M.A. CFTR Regulates Early Pathogenesis of Chronic Obstructive Lung Disease in βENaC-Overexpressing Mice. PLOS ONE 2012, 7, e44059. [Google Scholar] [CrossRef]
- J. R. Harkema, “Development of chronic bronchitis and emphysema in beta-epithelial na+ channel-overexpressing mice: Role of airway surface dehydration in the pathogenesis of COPD,” Int J Toxicol, vol. 29, no. 1, 2010.
- Mall, M.A.; Grubb, B.R.; Harkema, J.R.; O’Neal, W.K.; Boucher, R.C. Increased airway epithelial Na+ absorption produces cystic fibrosis-like lung disease in mice. Nat. Med. 2004, 10, 487–493. [Google Scholar] [CrossRef]
- Zhou, Z.; Duerr, J.; Johannesson, B.; Schubert, S.C.; Treis, D.; Harm, M.; Graeber, S.Y.; Dalpke, A.; Schultz, C.; Mall, M.A. The ENaC-overexpressing mouse as a model of cystic fibrosis lung disease. J. Cyst. Fibros. 2011, 10, S172–S182. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, A.M.; McCrudden, M.T.C.; Vincente-Perez, E.M.; Dubois, A.V.; Ingram, R.J.; Larrañeta, E.; Kissenpfennig, A.; Donnelly, R.F. Design and characterisation of a dissolving microneedle patch for intradermal vaccination with heat-inactivated bacteria: A proof of concept study. Int. J. Pharm. 2018, 549, 87–95. [Google Scholar] [CrossRef]
- Osbourn, M.; Rodgers, A.M.; Dubois, A.V.; Small, D.M.; Humphries, F.; Delagic, N.; Moynagh, P.N.; Weldon, S.; Taggart, C.C.; Ingram, R.J. Secretory Leucoprotease Inhibitor (SLPI) Promotes Survival during Acute Pseudomonas aeruginosa Infection by Suppression of Inflammation Rather Than Microbial Killing. Biomolecules 2022, 12, 1728. [Google Scholar] [CrossRef]
- Oleszycka, E.; Rodgers, A.M.; Xu, L.; Moynagh, P.N. Dendritic Cell–Specific Role for Pellino2 as a Mediator of TLR9 Signaling Pathway. J. Immunol. 2021, 207, 2325–2336. [Google Scholar] [CrossRef]
- Reyne, N.; McCarron, A.; Cmielewski, P.; Parsons, D.; Donnelley, M. To bead or not to bead: A review of Pseudomonas aeruginosa lung infection models for cystic fibrosis. Front. Physiol. 2023, 14, 1104856. [Google Scholar] [CrossRef]
- Kukavica-Ibrulj, I.; Bragonzi, A.; Paroni, M.; Winstanley, C.; Sanschagrin, F.; O’Toole, G.A.; Levesque, R.C. In Vivo Growth of Pseudomonas aeruginosa Strains PAO1 and PA14 and the Hypervirulent Strain LESB58 in a Rat Model of Chronic Lung Infection. J. Bacteriol. 2008, 190, 2804–2813. [Google Scholar] [CrossRef] [PubMed]
- Camus, L.; Briaud, P.; Vandenesch, F.; Moreau, K. How Bacterial Adaptation to Cystic Fibrosis Environment Shapes Interactions Between Pseudomonas aeruginosa and Staphylococcus aureus. Front. Microbiol. 2021, 12. [Google Scholar] [CrossRef]
- Jackson, L.; Waters, V. Factors influencing the acquisition and eradication of early Pseudomonas aeruginosa infection in cystic fibrosis. J. Cyst. Fibros. 2020, 20, 8–16. [Google Scholar] [CrossRef]
- Fischer, A.J.; Singh, S.B.; LaMarche, M.M.; Maakestad, L.J.; Kienenberger, Z.E.; Peña, T.A.; Stoltz, D.A.; Limoli, D.H. Sustained Coinfections withStaphylococcus aureusandPseudomonas aeruginosain Cystic Fibrosis. Am. J. Respir. Crit. Care Med. 2021, 203, 328–338. [Google Scholar] [CrossRef]
- Adam, L.; López-González, M.; Björk, A.; Pålsson, S.; Poux, C.; Wahren-Herlenius, M.; Fernández, C.; Spetz, A.-L. Early Resistance of Non-virulent Mycobacterial Infection in C57BL/6 Mice Is Associated With Rapid Up-Regulation of Antimicrobial Cathelicidin Camp. Front. Immunol. 2018, 9, 1939. [Google Scholar] [CrossRef] [PubMed]
- Fornefett, J.; Krause, J.; Klose, K.; Fingas, F.; Hassert, R.; Eisenberg, T.; Schrödl, W.; Grunwald, T.; Müller, U.; Baums, C.G. Comparative analysis of clinics, pathologies and immune responses in BALB/c and C57BL/6 mice infected with Streptobacillus moniliformis. Microbes Infect. 2018, 20, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Bernal, A.M.; Fernández-Brando, R.J.; Bruballa, A.C.; Fiorentino, G.A.; Pineda, G.E.; Zotta, E.; Vermeulen, M.; Ramos, M.V.; Rumbo, M.; Palermo, M.S. Differential Outcome between BALB/c and C57BL/6 Mice after Escherichia coli O157:H7 Infection Is Associated with a Dissimilar Tolerance Mechanism. Infect. Immun. 2021, 89. [Google Scholar] [CrossRef] [PubMed]
- Thach, D.C.; Kimura, T.; Griffin, D.E. Differences between C57BL/6 and BALB/cBy Mice in Mortality and Virus Replication after Intranasal Infection with Neuroadapted Sindbis Virus. J. Virol. 2000, 74, 6156–6161. [Google Scholar] [CrossRef] [PubMed]
- Mantero, M.; Gramegna, A.; Pizzamiglio, G.; D’adda, A.; Tarsia, P.; Blasi, F. Once daily aerosolised tobramycin in adult patients with cystic fibrosis in the management of Pseudomonas aeruginosa chronic infection. Multidiscip. Respir. Med. 2017, 12, 2. [Google Scholar] [CrossRef] [PubMed]
- Boucher, R. An overview of the pathogenesis of cystic fibrosis lung disease. Adv. Drug Deliv. Rev. 2002, 54, 1359–1371. [Google Scholar] [CrossRef]
- van Velzen, A.J.; Uges, J.W.; Heijerman, H.G.; Arets, B.G.; Nuijsink, M.; van der Wiel-Kooij, E.C.; van Maarseveen, E.M.; van Zanten, G.A.; Pullens, B.; Touw, D.J.; et al. Pharmacokinetics and safety of tobramycin nebulization with the I-neb and PARI-LC Plus in children with cystic fibrosis: A randomized, crossover study. Br. J. Clin. Pharmacol. 2019, 85, 1984–1993. [Google Scholar] [CrossRef]
- Tucker, S.L.; Sarr, D.; Rada, B. Neutrophil extracellular traps are present in the airways of ENaC-overexpressing mice with cystic fibrosis-like lung disease. BMC Immunol. 2021, 22, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Vyas, H.K.N.; Xia, B.; Mai-Prochnow, A. Clinically relevant in vitro biofilm models: A need to mimic and recapitulate the host environment. Biofilm 2022, 4, 100069. [Google Scholar] [CrossRef]
- Jordan, S.C.; Hall, P.R.; Daly, S.M. Nonconformity of biofilm formation in vivo and in vitro based on Staphylococcus aureus accessory gene regulator status. Sci. Rep. 2022, 12, 1–11. [Google Scholar] [CrossRef]
- Cigana, C.; Bianconi, I.; Baldan, R.; De Simone, M.; Riva, C.; Sipione, B.; Rossi, G.; Cirillo, D.M.; Bragonzi, A. Staphylococcus aureus Impacts Pseudomonas aeruginosa Chronic Respiratory Disease in Murine Models. J. Infect. Dis. 2017, 217, 933–942. [Google Scholar] [CrossRef]
- Michelsen, C.F.; Christensen, A.-M.J.; Bojer, M.S.; Høiby, N.; Ingmer, H.; Jelsbak, L. Staphylococcus aureus Alters Growth Activity, Autolysis, and Antibiotic Tolerance in a Human Host-Adapted Pseudomonas aeruginosa Lineage. J. Bacteriol. 2014, 196, 3903–3911. [Google Scholar] [CrossRef] [PubMed]
- Oakley, J.L.; Weiser, R.; Powell, L.C.; Forton, J.; Mahenthiralingam, E.; Rye, P.D.; Hill, K.E.; Thomas, D.W.; Pritchard, M.F. Phenotypic and Genotypic Adaptations in Pseudomonas aeruginosa Biofilms following Long-Term Exposure to an Alginate Oligomer Therapy. mSphere 2021, 6. [Google Scholar] [CrossRef]
- Singh, R.; Mackay, A.J.; Patel, A.R.; Garcha, D.S.; Kowlessar, B.S.; E Brill, S.; E Donnelly, L.; Barnes, P.J.; Donaldson, G.C.; A Wedzicha, J. Inflammatory thresholds and the species-specific effects of colonising bacteria in stable chronic obstructive pulmonary disease. Respir. Res. 2014, 15, 114. [Google Scholar] [CrossRef]
- Ghanem, M.K.; Makhlouf, H.A.; Hasan, A.A.; Rashed, H.G.; Khalifa, H.S. Bacteriological profile of critically ill patients with chronic obstructive pulmonary disease in respiratory intensive care unit in Assuit University Hospital. Egypt. J. Bronc- 2019, 13, 343–348. [Google Scholar] [CrossRef]
- Garcia-Vidal, C.; Almagro, P.; Romani, V.; Rodriguez-Carballeira, M.; Cuchi, E.; Canales, L.; Blasco, D.; Heredia, J.L.; Garau, J. Pseudomonas aeruginosa in patients hospitalised for COPD exacerbation: a prospective study. Eur. Respir. J. 2009, 34, 1072–1078. [Google Scholar] [CrossRef]
- Martínez-Solano, L.; Macia, M.D.; Fajardo, A.; Oliver, A.; Martinez, J.L. ChronicPseudomonas aeruginosaInfection in Chronic Obstructive Pulmonary Disease. Clin. Infect. Dis. 2008, 47, 1526–1533. [Google Scholar] [CrossRef]
- Botha, P.; Archer, L.; Anderson, R.L.; Lordan, J.; Dark, J.H.; Corris, P.A.; Gould, K.; Fisher, A.J. Pseudomonas aeruginosa Colonization of the Allograft After Lung Transplantation and the Risk of Bronchiolitis Obliterans Syndrome. Transplantation 2008, 85, 771–774. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
CD1 Mice were infected intranasally with clinical isolates of P. aeruginosa and compared to that of the common laboratory strain, PAO1 (1x107). Survival was monitored (n=10 per strain) (A). In vitro biofilm production of the clinical isolates and PAO1 was measured in vitro (n=8 replicates per strain) (B) and in vivo alginate production was measured (C) (n=8 per group).
Figure 1.
CD1 Mice were infected intranasally with clinical isolates of P. aeruginosa and compared to that of the common laboratory strain, PAO1 (1x107). Survival was monitored (n=10 per strain) (A). In vitro biofilm production of the clinical isolates and PAO1 was measured in vitro (n=8 replicates per strain) (B) and in vivo alginate production was measured (C) (n=8 per group).
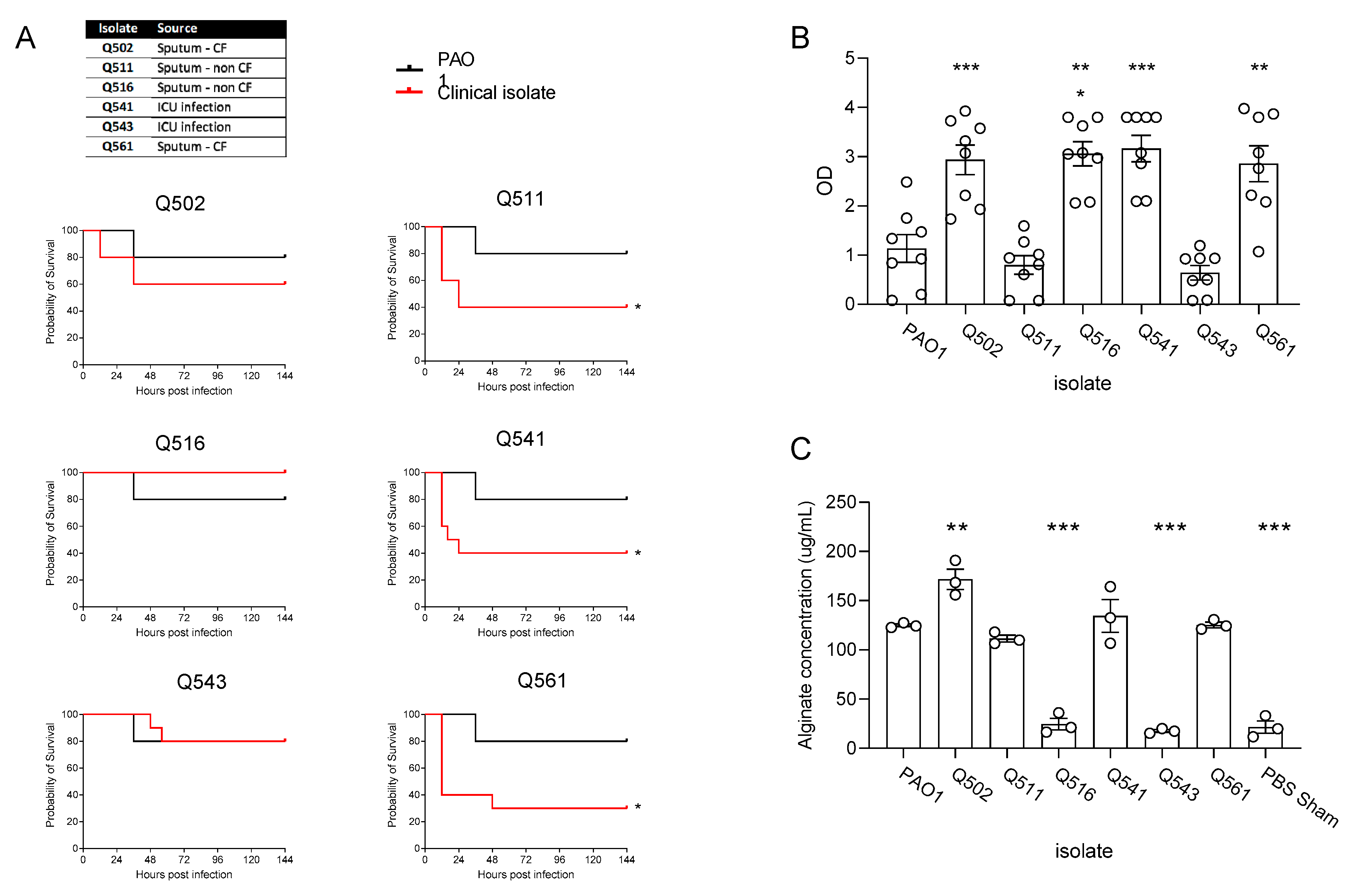
Figure 2.
Pre-treatment (at -10days) of CD1 mice with S. aureus (OD5) prior to infection – clinical isolates of S. aureus all from sputum samples from patients with CF (A). Comparison of inbred mouse strains with P. aeruginosa Q502 1x107 (B). Pre-treatment of Biozzi mice with S. aureus Q154 OD5 and OD10 (C). IL-6 with S. aureus pre-treatment (D). Detection of Q154 within the lungs 10 days after inoculation (E). In vitro co-culture of Q502 and Q154 (F). Infection of biozzi mice with high dose (1x107) Q502 – prior inoculation with Q154 significantly enhances survival (G) and there was detectable P. aeruginosa in the lungs 14 days post infection (H).
Figure 2.
Pre-treatment (at -10days) of CD1 mice with S. aureus (OD5) prior to infection – clinical isolates of S. aureus all from sputum samples from patients with CF (A). Comparison of inbred mouse strains with P. aeruginosa Q502 1x107 (B). Pre-treatment of Biozzi mice with S. aureus Q154 OD5 and OD10 (C). IL-6 with S. aureus pre-treatment (D). Detection of Q154 within the lungs 10 days after inoculation (E). In vitro co-culture of Q502 and Q154 (F). Infection of biozzi mice with high dose (1x107) Q502 – prior inoculation with Q154 significantly enhances survival (G) and there was detectable P. aeruginosa in the lungs 14 days post infection (H).
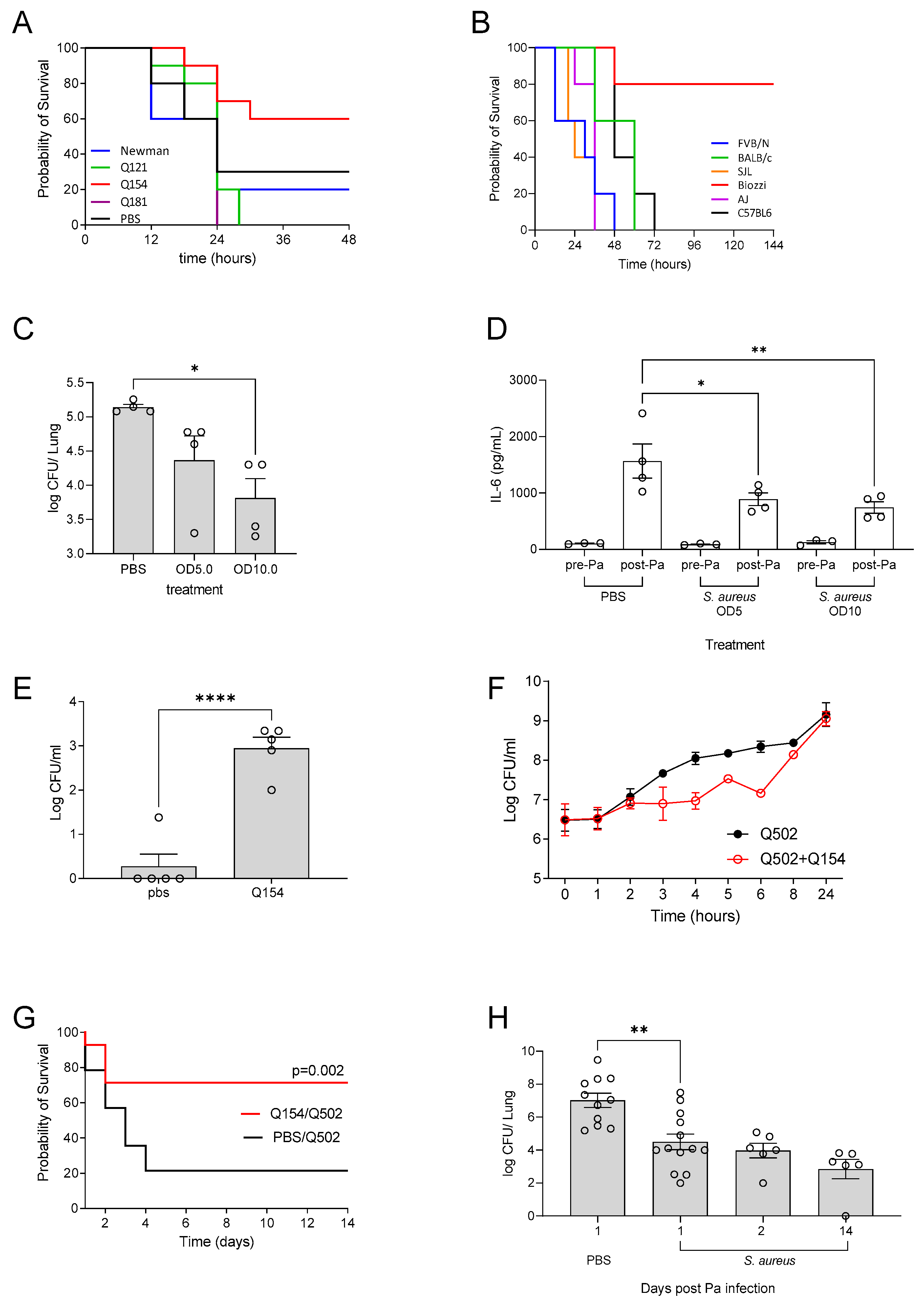
Figure 3.
Mice were infected intra-nasally with 1x106 CFU of P. aeruginosa (PAO1). At 1 h post-infection, mice were administered inhaled tobramycin via nebulization (10mg/kg) for 7 days. At each time point, mice were sacrificed for enumeration of CFU in the lungs (A). At 14 days post infection, lung and faeces were collected, there was a significant correlation between bacterial levels in the lung and the amount being shed in faeces (B). Untreated n=12, treated n=6-12 per time point, *** p ≤ 0.001, **** p ≤ 0.0001.
Figure 3.
Mice were infected intra-nasally with 1x106 CFU of P. aeruginosa (PAO1). At 1 h post-infection, mice were administered inhaled tobramycin via nebulization (10mg/kg) for 7 days. At each time point, mice were sacrificed for enumeration of CFU in the lungs (A). At 14 days post infection, lung and faeces were collected, there was a significant correlation between bacterial levels in the lung and the amount being shed in faeces (B). Untreated n=12, treated n=6-12 per time point, *** p ≤ 0.001, **** p ≤ 0.0001.
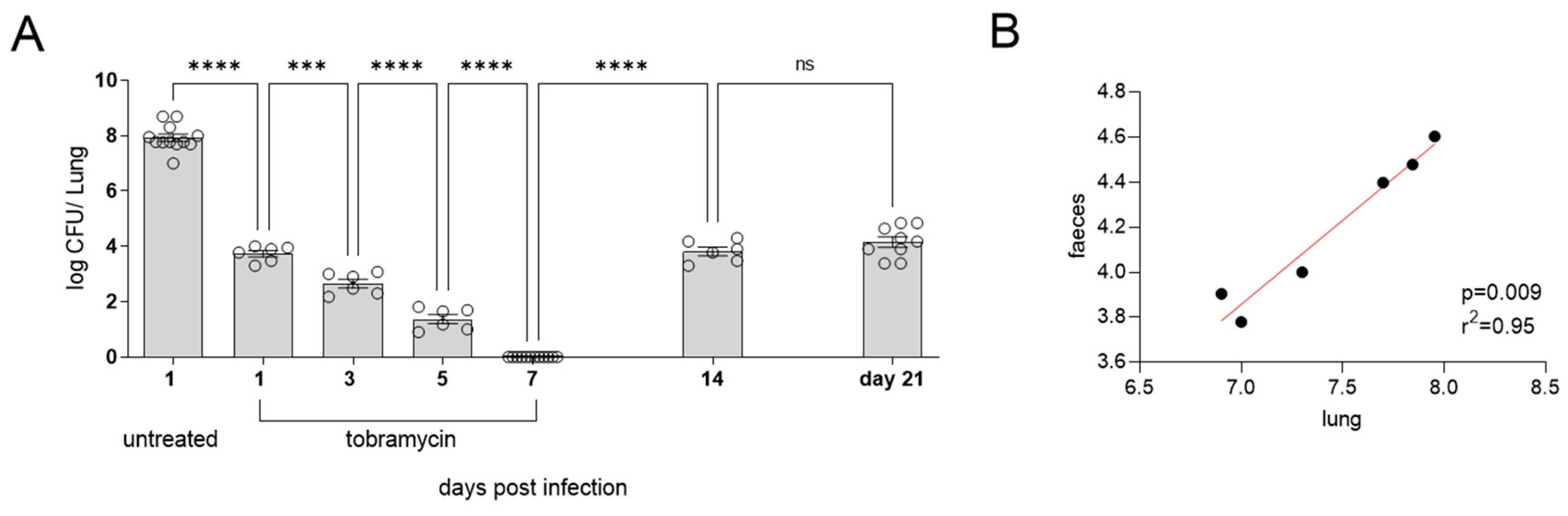
Figure 4.
C57BL6 mice infected with PAO1 in water bottle (1x107 per ml) for 5 days. Mice were then transferred back onto fresh drinking water and culled at days 7, 14, 20, 30 and 50 and the levels of CFU in the lungs (A) and NALT (B). The levels of IL-6 in the lungs were quantified (C) (n=6-9 per time point). Following treatment in this model there is sustained bacterial clearance in water bottle infected mice (n=6 per time point) (D). βENaC mice were infected with PAO1 and the CFU in the lungs was quantified (E) and weights were measured (F) (n=6 per time point).
Figure 4.
C57BL6 mice infected with PAO1 in water bottle (1x107 per ml) for 5 days. Mice were then transferred back onto fresh drinking water and culled at days 7, 14, 20, 30 and 50 and the levels of CFU in the lungs (A) and NALT (B). The levels of IL-6 in the lungs were quantified (C) (n=6-9 per time point). Following treatment in this model there is sustained bacterial clearance in water bottle infected mice (n=6 per time point) (D). βENaC mice were infected with PAO1 and the CFU in the lungs was quantified (E) and weights were measured (F) (n=6 per time point).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



