
Submitted:
01 June 2023
Posted:
01 June 2023
You are already at the latest version
Abstract
Collagen, the most abundant structural protein found in mammals, plays a vital role as a constituent of the extracellular matrix (ECM) that surrounds cells. Collagen fibrils are strengthened through the formation of covalent cross-links, which involve complex enzymatic and non-enzymatic reactions. Lysyl oxidase (LOX) is responsible for catalyzing the oxidative deamination of lysine and hydroxylysine residues, resulting in the production of aldehydes, allysine, and hydroxyallysine. These intermediates undergo spontaneous condensation reactions, leading to the formation of immature cross-links, which are the initial step in the development of mature covalent cross-links. Additionally, non-enzymatic glycation contributes to the formation of abnormal cross-linking in collagen fibrils. During glycation, specific lysine and arginine residues in collagen are modified by reducing sugars, leading to the creation of Advanced Glycation End-products (AGEs). These AGEs have been associated with changes in the mechanical properties of collagen fibers. Interestingly, various studies have reported that plant polyphenols possess amine oxidase-like activity and can act as potent inhibitors of protein glycation. This review article focuses on compiling literature describing polyphenols with amine oxidase-like activity and antiglycation properties. Specifically, we explore the molecular mechanisms by which specific flavonoids impact or protect the normal collagen cross-linking process. Furthermore, we discuss how these dual activities can be harnessed to generate properly cross-linked collagen molecules, thereby promoting the stabilization of highly organized collagen fibrils.
Keywords:
Collagen Cross-linking
; Extracellular Matrix
; Fibrosis
; Glycation
; Lysyl Oxidase
; Polyphenols
1. Introduction
Biosynthesis and maturation of collagen fibrils involves several post-translational modifications including folding, hydroxylation of proline and lysine residues, glycosylation of lysine and hydroxylysine residues, disulphide bonding, trimerization, secretion, processing, self-assembly and cross-linking [1,2,3,4,5,6,7]. The lysyl oxidase family catalyzed the final enzyme step in the biosynthetic collagen cross-linking [8,9]. This last reaction is a meticulous sequential process that occurs outside the cell, and it is classified as a physiologically controlled enzymatic process, which is essential to create a standard suprafibrillar architecture of collagen fibers, that contribute to mechanical properties and structural organization of tissues [2,7,10,11].
Lysyl oxidase-mediated collagen cross-linking is an extracellular post-translational modification crucial for collagen structure stabilization and tensile strength properties of connective tissues [5,8,9,12]. LOX catalyzes the oxidative deamination of ε-amine lateral chains of lysine and hydroxylysine in telopeptide regions in growing fibrils, which originate reactive aldehydes (α-aminoadipic- δ-semialdehydes) that can condense each other or with lysine and hydroxylysine residues in the helical domain to form inter- and intramolecular of divalent cross-links, that spontaneously condense to form a variety of mature and permanent covalent cross-links. Therefore, this final enzymatic reaction is essential for form immature and mature cross-links that stabilize tropocollagen macromolecules during the formation of collagen fibrils, and it is critically important to support normal tissue remodelling and growth [8,9,11,13].
In contrast, glycation is a non-enzymatic reaction that can modify collagen cross-linking in fibrils, producing advanced glycation end-products (AGEs), which are associated with altered mechanical properties of collagen fibers and stiffening of aged tissues [13,14]. Such uncontrolled processes occur when sugar molecules attach covalently to lateral ε- amine chains of lysine residues in collagens. The non-enzymatic collagen cross-linking by glycation is considered affect the typical functions of connective tissues negatively. Two lysine-arginine cross-links have been identified in collagen, the first is pentosidine which is a fluorescent product formed from ribose, and the second is glucosepane derived from glucose. The latter is found abundantly in senescent skin collagen [7]. The collagen turnover decreases during normal ageing and the accumulation of AGEs produces that they can easily bind and cross-links collagen, modifying the physical properties of fibers, which are deformed, altering the homeostasis between the degradation and synthesis of the extracellular matrix (ECM) [15].
On the other hand, polyphenols are a group of phytochemicals found in plants, fruits, vegetables, floral tissues, stems, bark and roots. They are widely known for their antioxidative capacity in general [16,17,18]. However, besides antioxidant activity, some polyphenols also have the potential to promote oxidative deamination of primary amines by oxidation to the corresponding o-quinone derivative [19,20,21,22]. Interestingly, through the formation of the o-quinone derivative bioflavonoids can form α-aminoadipic-5-semialdehyde in collagen and elastin by the oxidation of the ε-amine group of lysine residues [20,23,24,25,26]. This activity has been related to non-specific chemical catalysis because it is not blocked by β- Aminopropionitrile (BAPN), a competitive inhibitor of the lysyl oxidase family [27]. Noteworthy, polyphenols can also inhibit protein glycation and have been postulated as potential antiglycation agents, mainly due to their antioxidant and chelating activities, which are able to act as compounds inhibiting or breaking AGE-induced cross-links in collagens [28,29]. However, despite plentiful findings about polyphenols reported to date, the specific biological functions of such polyphenols on collagen cross-link types at their molecular level are still unclear. Also, influence of bioflavonoids on the activity of other members of the lysyl oxidase family has not been explored.
Therefore, the objective of this review is to elucidate the oxidative properties of certain polyphenols in facilitating the formation of both immature and mature cross-links, which contribute to the stabilization of tropocollagen macromolecules. Additionally, this review delves into the inhibitory and disruptive effects of polyphenols on the formation of advanced glycation end-products (AGEs) in collagen. By shedding light on these mechanisms, this research lays the foundation for the development of active compounds capable of modulating oxidative deamination and interfering with the glycation of lysine ε-amino groups within collagen fibrils.
2. Fibrillar Type I Collagen
Extracellular matrices provide essential support and create a conducive environment for the assembly of large macromolecular structures, which consist of macromolecules and peptides, including collagen precursors and mature collagen. These structures combine to form larger fibrillar arrangements [88]. The presence of collagen within the connective tissues’ fundamental framework is of utmost importance for establishing a proper tissue architecture. These intricate networks of macromolecules exert specific mechanical forces that profoundly impact cellular behavior and morphology [89]. Collagens are the most abundant proteins in the extracellular matrix (ECM), where most interacts with other ECM proteins to form the structural network of tissues [30]. Collagen interaction with cells involves several processes, including cell differentiation, mechanical strength, proliferation, cell adhesion and migration [1,7]. Based on their supramolecular assemblies, collagens are classified into four subfamilies: fibrils, beaded filaments, networks and anchoring fibrils [7,30,31,32]. Until today, twenty-eight types of collagens have been found in mammals, with type I collagen being the most common and abundant form in animals [7,33]. Type I collagen, a fibril-forming collagen, is found in several tissues, including skin, bones, tendons, vascular system and cornea [1,7]. The triple helix is a common structural motif to all collagen types, and it stabilizes by the presence of the characteristic repeating amino acid sequence Gly-X-Y, where glycine occupies every third position, and X a Y are often proline and hydroxyproline, respectively [7]. Three parallel α-chains twisted around each other build procollagen molecules with a right-handed triple helix. In particular, type I collagen is mainly comprises heterotrimers containing one α2 and two α1 chains (Figure 1) [34]. The triple helix is stabilized conformationally by interchain hydrogen bonds formed by proline and hydroxyproline. Hydroxyproline is formed by prolyl hydroxylase in a reaction that requires essential cofactors such as Fe2+, 2-oxoglutarate, O2 and vitamin C (Figure 1A) [5]. Also, other amino acids, including lysine, arginine, glutamate and aspartate, participate in forming electrostatic attractions in type I procollagen and provide conformational stability to the triple helix [1,35].
It has been proposed that lysine hydroxylation and lysine glycosylation are crucial for the formation of reducible and mature cross-links produced via the lysyl oxidase system and for stabilizing the supramolecular structures of fibrils-formed collagens [1,7,36,37]. In particular, lysine residues in procollagens are post-translationally modified by lysyl hydroxylases (LH) (Figure 1A). LH1 catalyzes lysine modifications inside the cell by hydroxylating specific peptidyl lysine residues producing hydroxylysine residues in triple-helical regions, whereas LH2 mediates the hydroxylation of lysine residues in terminal telopeptidyl regions [38]. This hydroxylation occurs almost exclusively in the Y positions of the repeating sequence Gly-X-Y in a well-ordered reaction that requires the same essential cofactors used for proline hydroxylation reactions, which leads to the release of a hydroxylated lysyl residue in the procollagen polypeptide, CO2, and succinate (Figure 1A) [16]. The hydroxylation of particular lysine residues along the procollagen α-chains is particularly important to subsequent normal collagen cross-linking [38]. Also, some of the hydroxylysine residues in the helical domain are further modified by sequential steps of O-linked glycosylation and whose structural and biological functions have been proposed for control of collagen fibrillogenesis, cross-linking and remodelling of collagen fibrils, among others [3,5,39]. An extensive review was published by Yamauchi et al., focusing on lysine post-translational modifications [5].
In the process of collagen biosynthesis, after the export of procollagen molecules to the extracellular matrix, procollagen is cleaved by matrix metalloproteinases (MMPs), generating tropocollagen molecules (trimeric collagen molecules) which self-assembly laterally and longitudinally to form fibrils [40]. Lysine and hydroxylysine residues in trimeric collagen molecules are post-translationally modified to form immature cross-links [12]. The resulting aldehydes then react with neighboring lysine residues or other aldehyde residues spontaneously to generate intra- and intermolecular covalent cross-links, leading to the formation of immature covalent connections which are converted into stable and mature cross-links that generate macroscopic fibers [41]. These fibers are observed as a part of normal collagen supramolecular structures in tissues [35] (Figure 1C–E). It is worth mentioning that the use of hydroxylysine and copper sulfate has been established to enhance the formation of collagen cross-links, specifically pyridinoline [42]. This augmentation significantly improves the biomechanical properties of neocartilage [42].
In the fibrillogenesis process, the enzyme-mediated inter- and intramolecular cross-links stabilize tropocollagen molecules within the fibril, which involves lysine, hydroxylysine and histidine residues [7]. Then, covalent cross-links reinforce the mechanical properties of the collagen supramolecular structure in a tissue-specific manner, which is why they are the most crucial post-translational modification described in detail in these revisions [43,44,45]. In addition, this review focuses on enzymatic and non-enzymatic covalent cross-links of type I collagen because, at present, the knowledge about collagen cross-linking has mostly been explored from studies of fibrillar type I collagen.
3. Enzymatic Collagen Cross-Linking Mediated by Lysyl Oxidases
It has been well established that lysyl oxidase acts upon its substrates collagen, elastin and nonpeptidyl amines following a ping-pong mechanism [45,46,47]. In particular for collagens, after the secretion of procollagen molecules outside of the cell, lysyl oxidase catalyzes the conversion of lysine and hydroxylysine residues into aldehydes in nonhelical telopeptide regions. Aldehydes reacts with lysine or hydroxylysine residues contained in the triple helical domain on an adjacent collagen molecule to form immature divalent cross-links that can also spontaneously react with another divalent cross-links to create mature trivalent cross-links [47]. Furthermore, it has been observed that when the lysine- derived cross-links are formed by hydroxylysine-derived aldehydes, it is more stable than those formed from the lysine aldehyde pathway [5].
LOX enzymes regulate several biological processes, including extracellular matrix stabilization, cellular growth, and homeostasis [48,49]. Still, the primary role of this enzyme family is to participate in the remodelling of extracellular matrices through the formation of inter and intrachain cross-links in collagen and elastin. Primarily, LOX enzymes promote the first step in the formation of covalent cross-linking to stabilize collagen fibrils [12,50,51]. Specifically, for fibrillar type I collagen it has been reported that LOX and LOXL2 can form covalent cross-link in the molecule [52]. Also, it has been demonstrated that LOX, LOXL2 and LOXL4 cross-link collagen IV, along with indications that LOXL4 cross-links IV by an increase in collagen IV deposition in vascular matrix remodeling [47,53,54,55].
It has been shown that LOX activity is vital for wound healing but also is involved in the pathogenesis of fibrotic diseases, more precisely, its dysregulated activity promote ineffective or excessive collagen cross-linking, which drives multiple diseases [49,56]. In this context, the collagen cross-linking formation has been associated with many chronic diseases including diabetes, cancer metastasis, osteoarthritis and vascular and fibrotic diseases [57,58,59]. In addition, the augmented cross-linking activity of these enzymes is responsible for large insoluble extracellular proteins resistant to proteolysis reported in several pathological conditions. Also, it has been associated with increased deposition of fibrillar collagens in fibrotic areas. Dysregulation of expression and activity of lysyl oxidases have been found to correlate with numerous diseases and adverse physiological states, including fibrosis in different organs such as the liver, lung, and kidney [60,61,62]. Very recently, it has been proposed that LOXL4 in the main LOX activity and critical determinant of collagen cross-linking in lung fibrosis [63]. Among all lysyl oxidases enzymes, the isoforms LOX and LOXL2 are widely associated with metastasis progression because it is required to produce a permissive niche to maintain metastatic tumor cell growth [64]. Lastly, it has been proposed that defining tissue-specific variance in collagen cross-linking may help to create biomarkers of pathological connective tissues [13].
Five members constitute the mammal LOX which are classified according to primary structure and functions: LOX, LOXL1, LOXL2, LOXL3 and LOXL4 [12]. Members have been divided into two subfamilies based on a phylogenetic study described by Rodriguez-Pascual group [65]. The first subfamily includes LOX and LOXL1, and the second comprises LOXL2, LOXL3 and LOXL4. These enzymes are classified as copper amine oxidases and display a conserved catalytic domain that contains the copper binding site [47]. The enzymatic reaction of lysyl oxidases isoforms requires, besides copper, the organic quinone cofactor named lysyl tyrosylquinone (LTQ) (Figure 2A) [66,67]. The copper ion is incorporated into LOX in the trans-Golgi network by ATP7A, a copper-transporting P-type ATPase 1 [47,68]. It has been described that tree histidine residues, H292, H294 and H296, coordinate the essential copper cation in LOX [47,69]. In LOXL2, the equivalent histidine residues H662, H628, and H630 form the cooper-binding site [70,71]. The LTQ is formed by specific residues within the nascent enzyme derived from tyrosine Y355 and lysine K320 in the LOX isoform. For LOXL2, Mure and colleagues described the spatial arrangement of LTQ between Y689 and K653 and their position relative to the coordination site of Cu2+ [66,70,72]. For another side, the N-terminal domain differ that the highly conserved catalytic domain: AlphaFold provides a protein structure of hLOX (Uniprot-ID: P28300) [73,74] (Figure 2B,C). The catalytic domain is modelled with quality attributes confident and very high confidence according to AlphaFold’s confidence score (blue-colored regions in the protein structure). Solely the area around K320 shows low confidence (white-colored region in the protein structure). This is likely due to the flexibility of this region to allow conformational adjustments during LTQ co-factor formation, as proposed by Meier and colleagues [70].
On the other hand, the N-terminal domain of LOX protein differ from the highly conserved catalytic domain. LOX and LOXL1 contain a propeptide region in their N-terminal part, whereas LOXL2, LOXL3 and LOXL4 contain four scavenger receptor cysteine-rich domains (SRCR) [12]. This type of domain has been involved in the proteolytic processing of the LOXL2 isoform by the proprotein convertase PACE4 [47,75]. Also, two N-glycosylation sites have been described in LOXL2 that are present in the second and fourth SRCR domains [12,76]. It has been proposed that there are important differences in the modulation of catalytic activity of LOX isoforms and their molecular mechanisms [75].
It has been proposed that the formation of immature enzymatic cross-links, generated during collagen fibrillogenesis by the enzymatic activity of lysyl oxidase, is a beneficial process in development [2]. In contrast, the formation of mature cross-links damages connective tissues over time and is particularly associated with ageing [38]. Examples of immature reducible and divalent cross-links are aldimine and keto-imine bonds that form in newly synthesized collagens [7,41]. In particular, aldimine bonds are formed between an aldehyde and an amine group through a condensation reaction, where the carbonyl group of the aldehyde reacts with the amine group to form a Schiff base [1,77]. The resulting molecule is named an aldimine or imine containing cross-link. It has been established that dehydro-lysinonorleucine (deH-LNL), dehydro-hydroxylysinonorleucine (deH-HLNL) and dehydro-dihydroxylysinonorleucine (deH-DHLNL) are immature divalent cross-links that have shown to be crucial for the formation of more complex cross-links in type I collagen. Ketoimine cross-links are another type of immature divalent cross-links when lysine-keto-norleucine (LKNL) and hydroxylysine-keto-norleucine (HLKNL) are produced via Amadori rearrangements using deH-HLNL and deH-DHLNL cross-links [1]. These bifunctional reducible cross-links undergo spontaneous maturation into nonreducible trivalent cross-links, such as pyridinoline and deoxypyridinoline (found in bone and cartilage), pyrrole cross-links (present in bone), arginoline (found in cartilage), and histidinohydroxylysinonorleucine (found in skin) [78,79]. The presence of these specific cross-links highlights their tissue specificity [7]. Interestingly, cross-links associated with histidine were reported to be artefacts found in mass spectrometry [80]. However, this type of cross-linking has been detected in vivo, suggesting that it can be susceptible to the low pH that produces cross-link degradation [1].
Significantly, aging often manifests in two contrasting scenarios: an excessive local deposition of collagen, as seen in fibrosis, or an overall gradual reduction in collagen mass [81]. As individuals age, the normal cross-linking of collagen in connective tissues diminishes due to cumulative damage from collagen fragmentation, oxidation, and glycation [82]. This progressive decline in collagen mass, observed not only in supporting tissues but also in other organs, compromises the integrity of the extracellular matrix and has implications for age-related conditions like diabetes, cancer, chronic liver disease, and cardiovascular diseases [83]. Simultaneously, the accumulation of molecular damage, chronic inflammation, or injury during aging can drive abnormal collagen deposition leading to fibrosis [36,84]. Furthermore, the deficiency of the LOX enzyme in adult skin has been associated with inadequate or abnormal collagen cross-linking, which contributes to skin aging [85]. These observations indicate a general decline in enzymatic collagen cross-linking with age. However, due to increased occurrences of damage or injury, atypical collagen accumulation, as observed in fibrosis, can transpire.
Immature reducible and divalent cross-links are aldimine and keto-imine bonds. Mature multivalent cross-links include pyrrole-containing cross-links (deoxypyrrololine (d-Prl) and pyrrololine (Prl)), pyridinium-salt containing cross-links (pyridinoline (Pyr) and deoxypyridinoline (Dpyr)) and histidine-containing cross-links (histidinohydroxylysinonorleucine (HHL) and histidinohydroxymerodesmosine (HHMD)). Glycation derived cross-links include dicarbonyl compounds and advanced glycation end products (AGEs).
4. Non-Enzymatic Collagen Glycation
Reducing sugars such as glucose, fructose, pentoses, galactose, mannose and xylulose possess a reactive carbonyl moiety that reacts non-enzymatically, predominantly, with the ε- amino group of lysine and the guanidine group of arginine residues, to form a labile compound [1,84,86,87,88]. Then the early stage in the reaction is the formation of a Schiff base between glucose and protein amino groups, followed by an Amadori rearrangement, a more stable early glycation product [15]. These Amadori products can produce a variety of reactive dicarbonyl compounds such as methylglyoxal and glyoxal, an intermediate stage that can react with other free amino groups of proteins [89,90]. Afterwards, spontaneous reactions occur in the late stage to produce a chemically related group of moieties, termed advanced glycation end products (AGEs), which remain irreversibly bound to proteins. AGE products are a series of spontaneous post-translational modifications found on long-lived proteins, including collagens, and their tissue accumulation is markedly associated with loss of structural integrity in ageing [1,13,36,77,89,91,92,93,94]. And it has been shown that protein glycation disrupts normal activities such as enzymatic activity, molecular conformation, degradation ability and receptor recognition [95].
Multiple shreds of evidence suggest that AGEs are involved in inflammation and contribute to diverse pathologies, such as cardiovascular and renal diseases, diabetes and many cancer types of extensively reviewed in [15,87,92,96]. For example, the glycation of collagen by AGE products generated through persistently elevated glucose levels has been suggested to promote the development of fibrosis produced by tissue stiffening and reduce its solubility in diabetes, a chronic disease with hyperglycemia. Various types of AGEs have been studied, and the most common is pentosidine, which exhibits fluorescent properties. Other types of AGEs are the vesperlysine-type AGEs, glyoxal lysine-dimer, methylglyoxal lysine-dimer and glucosepane. It has been demonstrated that glucosepane is elevates in elderly patients and diabetic patients [97]. Also, an amide-containing AGE crosslink called GOLA has been correlated to collagen stiffness in rat tendons. A full review was published by Gaar and colleagues, focusing on enzymatic and non-enzymatic collagen cross-links found in collagen [1].
It has been suggested that the number of lysyl-derived cross-links occurs in significant abundance in comparison with AGEs collagen cross-links in younger persons. But, as a part ageing, collagen glycation increases while lysyl oxidase-mediated cross-links are maintained, which promotes an imbalance in the cross-linking formation [98]. In particular, the collagen glycation and the formation of AGE products can alter collagen fibrils’ elasticity, thermal denaturation and morphology [98]. Also, it has been demonstrated that the target of glycation in type I collagen is the same site that uses lysyl oxidase to generate immature cross-links. Moreover, it has been shown that the glycation on lysine residues in type I collagen has the potential to block normal lysyl oxidase-mediated collagen cross-linking [14]. In fact, lysine positions are more conserved than glycine, hydroxylysine or lysine residues in collagen, and the glycation in this position leads to the loss of enzymatic collagen cross-links [94]. It has been proposed that this effect is mainly due to both sharing the same target in the helical domain producing a steric hindrance by glucose carbonyls that significantly impair the normal lysyl oxidase-mediated collagen cross-linking [1,9,14,79]. However, the authors propose the AGE product glucosepane in diabetes and quantified it as the most abundant and relevant AGE product associated with type I collagen molecules [93,97]. By combining experimental and computational analysis, it has been shown that glucosepane alters the density of collagen packing and the denaturation temperature and enhances porosity to water molecules in aged tendons [93,98]. Also, high glucosepane levels have been identified in diabetic skin samples and associated with microvascular disease in other organs [97,99]. Until recently, the focus of collagen modification by AGEs and their consequences has been explored mainly with glucose because it is the primary glycating sugar in ageing and diabetes. However, it has been shown that 5-P-Ribose (5PR) has the propensity to glycate collagen, which can modify the cancer microenvironment. A study by Bandose and colleagues describing the effect of ribose on the molecular organization of collagen fibrils found that R5P glycation impact on the enzymatic cross-links and molecular arrangement into collagen fibrils, inhibiting or breaking the formation of lysyl derived cross- links, not only in the surface of the fibrils, because LC-MS experiments showed that R5P glycation is able to reduce the number of HLNL cross-links impacting in the fibril structure more than the surface of the collagen molecules, which has a significant effects on the arrangement of protomers in the formation the collagen fibrils [94]. It has been proposed that this effect on the changes in the molecular alignment of collagen fibrils can impact cell adhesion and migration processes [94].
5. Lysyl Oxidase Activity of Polyphenols
Polyphenols, derived from plants, fruits, vegetables, floral tissues, stems, bark, and roots, serve as major sources of polyphenolic compounds, and are the most varied group of phytochemicals [100]. Extensive research has been conducted on these natural products due to their well-known health benefits and protective effects [16,23]. Among the polyphenols, flavonoids play a crucial role and are associated with various positive health outcomes, including increased longevity and reduced risk of cardiovascular diseases in populations with a diet rich in flavonoids [101,102]. These beneficial effects are primarily attributed to the potent antioxidant activity exhibited by polyphenols. However, it is important to note that under certain conditions, polyphenols can also act as prooxidants, promoting the oxidation of other compounds [20,21,22,101]. This prooxidant activity may be linked to the non-enzymatic, metal-catalyzed oxidation of polyphenols, leading to the generation of hydrogen peroxide (H2O2). This non-enzymatic system can lead oxidative modification of proteins by oxidative deamination of the ε-amine group of lysine to α-aminoadipic-5-semialdehyde, the main carbonyl product, similar to the reaction of LOX enzymes [20,21]. Therefore, the biological activities of polyphenols, including their behavior as antioxidants or prooxidants, are believed to be concentration dependent and directly proportional to the total number of hydroxyl groups, especially those present in the B-ring of flavonoid molecules [101].
Several studies have described the properties of specific flavonoids in preserving collagen stability [103,104]. For instance, anthocyanidins, natural plant pigments found in fruits, flowers, and certain vegetables, have been shown to stabilize collagens [105,106]. Extracts rich in anthocyanidins inhibit collagen degradation, reduce metalloprotease (MMP) activity, and protect against UV radiation in dermal fibroblast models, thus preventing UV-induced skin photoaging [107]. Furthermore, flavonoids can stimulate the production of fibrillar collagen in mouse fibroblast models. Another mechanism associated with collagen stability preservation involves the inhibition of collagenase and elastase activity [108].
Importantly, polyphenols and flavonoids have been found to contribute to the formation of non-covalent and covalent collagen cross-links [1,103]. It has been established that polyphenols possess amine oxidase-like activity in the presence of Cu2+, which is essential for facilitating appropriate collagen and elastin cross-linking [20]. Several polyphenols from different plant species, including chlorogenic acid, gallic acid and caffeic acid, have been associated with this amine oxidase-like activity [21]. It has been assumed that these polyphenols are converted to the o-quinones and acquire a lysyl oxidase-like activity [21,101]. In addition, the conversion of catechol-type polyphenols into o-quinone derivatives has also been observed [20]. These quinones can catalyze the oxidative deamination of primary amines by polyphenols, leading to the formation of iminoquinone and iminophenol, and ultimately resulting in the oxidation product α-aminoadipic-5-semialdehyde [20]. Notably, the catechol-type polyphenols, which includes catechin (C), epicatechin (EC), epigallocatechin (EGC), epigallocatechin gallate (EGCG), gallocatechin gallate (GCG), and epicatechin gallate (ECG), are flavonoids primarily found in green tea and grapes [16,22] (Figure 4). In particular EGCG shown a high capacity to oxidize specific lysine residues, further contributing to oxidative deamination [20]. Also, the analog of resveratrol, piceatannol, has been associated with the formation of dehydrolysinonorleucine (HLNL) [109].
It has been established that the failure to form covalent cross-links results in increased degradation of collagen molecules [40,110]. Considering the impact of both polyphenol activities on collagen cross-linking, it becomes highly intriguing to explore their influence on the regulation of fibrotic processes and wound healing. However, further studies are necessary to elucidate the effect of natural extracts on collagen cross-linking. Most scientific investigations have primarily focused on evaluating the biosynthesis and deposition of the extracellular matrix. Therefore, it is crucial to examine the levels of cross-linking and the molecular mechanisms involved in the interaction between flavonoids and lysyl oxidase activity. This will enable us to establish appropriate methods for measuring collagen cross-linking accurately. Additional research is essential to enhance our understanding of the previously undefined roles of natural extracts and their beneficial effects on collagen biosynthesis and enzymatic cross-linking.
6. Anti-Glycating Activity of Polyphenols
At present, a compelling body of evidence supports the anti-glycation activities of polyphenols. Glycation is a spontaneous and non-enzymatic reaction between reducing sugars, such as glucose and fructose, and free amino groups of proteins, DNA, and lipids, which render an unstable Schiff base and then converted to more stable structures known as Amadori products. These products may undergo a complex series of reactions and thus leading to the formation of advanced glycation end-products (AGEs). The formation of AGEs occurs at a very high rate in the presence of hyperglycemia and tissue oxidative stress [89].
The pathological implications of AGEs formation are extensively supported in many human diseases. Although initially ascribed to the mechanisms underlying the micro and macrovascular complications in Diabetes Mellitus [92,111], the contribution of advanced glycation end-products in many human diseases is well-documented [88,112]. The pathologic effects of AGEs are mainly related to their ability to promote oxidative stress and inflammation by binding to the receptor for advanced glycation end-products (RAGE) [113], or by cross-linking with proteins and thus altering their structure and function [114]. Crosslinking by advanced glycation end products increases the stiffness of the collagen network in human articular cartilage: a possible mechanism through which age is a risk factor for osteoarthritis.
Polyphenols can significantly reduce the unhealthy consequences of advanced glycation end-products by different mechanisms. These effects are achieved by interfering with either RAGE expression and signaling, or by inhibiting the cross-linking with body proteins [28]. Noteworthy, the trapping capacity of dicarbonyls compounds particularly methylglyoxal (MGO) or glyoxal (GO), have been reported for some polyphenols, and deserves special attention because these dicarbonyls are considered as one of the most efficient protein crosslinkers [90].
In this regard, several polyphenols displayed important dicarbonyls trapping activities, as reported for quercetin [115], chrysin derivatives [116], genistein [117], epigallocatechin-3-gallate [118], as well as for resveratrol and different hydroxycinnamic acids [119,120] among many other polyphenols. Of note, this activity has been reported not only for soluble polyphenols but also to bound-polyphenol-rich insoluble dietary fiber [121].
7. Conclusions
Collagen, elastin, and other extracellular matrix (ECM) proteins play a crucial role in maintaining tissue integrity, providing structural support, and facilitating cellular functions [110]. The proper regulation of their synthesis, assembly, and cross-linking is essential for maintaining tissue homeostasis [122]. In recent years, the synergistic action of polyphenols and lysyl oxidase (LOX) enzymes has emerged as a fascinating area of research, offering new insights into the modulation of ECM proteins. This essay explores the collaborative relationship between polyphenols and LOX enzymes and their impact on the homeostasis of collagen, elastin, and other ECM proteins. Polyphenols are a diverse group of natural compounds found abundantly in fruits, vegetables, and plant-based products [23]. They possess antioxidant, anti-inflammatory, and anti-glycation properties, making them potential candidates for maintaining ECM homeostasis. By exerting their antioxidative effects, polyphenols can mitigate oxidative stress-induced damage to ECM proteins, preserving their structural integrity.
LOX enzymes are key regulators of ECM cross-linking, particularly in collagen and elastin fibers [9]. These enzymes catalyze the oxidative deamination of lysine and hydroxylysine residues, leading to the formation of aldehydes that subsequently undergo spontaneous condensation reactions, resulting in the generation of covalent cross-links. LOX enzymes contribute to the maturation and stabilization of ECM proteins, imparting strength and resilience to tissues. In fact, a lysyl oxidase-like activity, resembling an amino oxidase-like activity, has been observed in a various of polyphenols, particularly those of the o-diphenolic type, in the presence of Cu2+ ions [20]. However, most studies on the oxidation of lysine residues mediated by polyphenols have been conducted using small molecules such as lysine analogs [20]. The potential of these polyphenols to modulate the levels of cross-linking (both immature and mature cross-links) of human type I collagen, remains unclear. Further research is required to fully comprehend the impact of polyphenols-mediated cross-linking on the structure and function of collagen.
Emerging evidence suggests a synergistic interaction between polyphenols and LOX enzymes in the maintenance of ECM homeostasis. Polyphenols have been shown to influence LOX enzyme activity and expression levels, thereby impacting the cross-linking process [123]. Certain polyphenols have demonstrated the ability to enhance LOX enzyme activity, promoting the formation of mature and functional cross-links in collagen and elastin fibers [27]. Additionally, polyphenols can protect LOX enzymes from degradation, ensuring their sustained activity and promoting ECM integrity. Beyond their influence on LOX enzymes, polyphenols can directly modulate ECM remodeling processes. These compounds have been shown to regulate matrix metalloproteinases (MMPs) and tissue inhibitors of metalloproteinases (TIMPs), which are involved in ECM turnover and remodeling [108]. Polyphenols can inhibit excessive MMP activity and promote the synthesis of TIMPs, thereby maintaining the balance between ECM synthesis and degradation [40]. Then, the interplay between polyphenols and LOX enzymes represents a fascinating area of research with significant implications for maintaining ECM homeostasis. Through their antioxidant properties, polyphenols protect ECM proteins from oxidative damage, while their ability to modulate LOX enzyme activity and expression influences the formation of cross-links critical for collagen and elastin fiber integrity. Furthermore, polyphenols can directly regulate ECM remodeling processes, further contributing to the overall balance of ECM synthesis and degradation. Understanding the synergistic action of polyphenols and LOX enzymes offers exciting prospects for the development of therapeutic strategies aimed at preserving tissue integrity and addressing ECM-related disorders. In addition, the development of active compounds capable of modulating the oxidative deamination properties of specific natural polyphenols and facilitating the formation of immature and mature cross-links could contribute to the stabilization of collagen macrostructures for various applications in tissue engineering and regenerative medicine, including wound healing. By modulating collagen with polyphenols, the mechanical properties of collagen-based materials can be enhanced, making them more resilient to damage and degradation. Further investigations are warranted to unravel the intricate mechanisms underlying this collaborative relationship and its full therapeutic potential.
Furthermore, the effect of catechol-type polyphenols on the generation of collagen cross-links through enzymatic modifications (LOX and LOXL2) and non-enzymatic (AGE-products) has yet to be explored. The inhibitory and disruptive effects of polyphenols on the formation of advanced glycation end-products (AGEs) in collagen have not been thoroughly investigated either. The interference of several polyphenols with the glycation of lysine ε-amino groups within collagen fibrils, leading to significant trapping of dicarbonyls, could potentially reduce the adverse consequences associated with advanced glycation end-products. However, further studies are necessary to elucidate the effects of natural polyphenols-rich extracts on the enzymatic and non-enzymatic collagen cross-linking.
Author Contributions
Conceptualization, C.A.; writing—original draft preparation, C.A. and J.R.; writing—review and editing, C.A., J.R., A.R. and L.V.M.; funding acquisition, C.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Fondo Nacional de Desarrollo Científico y Tecnológico (FONDECYT 1212026), Fondo de Apoyo a la Postulación de Proyectos Externos-FAPPE 2021 (VRID-FAPPE21-09) and Programa de Fortalecimiento al Desarrollo Científico de Centros Regionales 2020 (R20F0001). The APC was funded by Vicerrectoría de Investigación y Doctorados de la Universidad San Sebastian – Fondo VRID_APC23/05 and FONDECYT 1212026.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gaar, J.; Naffa, R.; Brimble, M. Enzymatic and non-enzymatic crosslinks found in collagen and elastin and their chemical synthesis. Org. Chem. Front. 2020, 7, 2789–2814. [Google Scholar] [CrossRef]
- Robins, S. Biochemistry and functional significance of collagen cross-linking. Biochem. Soc. Trans. 2007, 35, 849–852. [Google Scholar] [CrossRef]
- Heikkinen, J.; Risteli, M.; Wang, C.; Latvala, J.; Rossi, M.; Valtavaara, M.; Myllylä, R. Lysyl Hydroxylase 3 Is a Multifunctional Protein Possessing Collagen Glucosyltransferase Activity. J. Biol. Chem. 2000, 275, 36158–36163. [Google Scholar] [CrossRef] [PubMed]
- Kirchner, M.; Deng, H.; Xu, Y. Heterogeneity in proline hydroxylation of fibrillar collagens observed by mass spectrometry. PLOS ONE 2021, 16, e0250544. [Google Scholar] [CrossRef]
- Yamauchi, M.; Sricholpech, M. Lysine post-translational modifications of collagen. Essays Biochem. 2012, 52, 113–133. [Google Scholar] [CrossRef]
- Lucero, H.A.; Kagan, H.M. Lysyl oxidase: an oxidative enzyme and effector of cell function. Cell. Mol. Life Sci. 2006, 63, 2304–2316. [Google Scholar] [CrossRef] [PubMed]
- Ricard-Blum, S., The collagen family. Cold Spring Harb Perspect Biol, 2011. 3(1): p. a004978.
- Siegel, R.C. Biosynthesis of Collagen Crosslinks: Increased Activity of Purified Lysyl Oxidase with Reconstituted Collagen Fibrils. Proc. Natl. Acad. Sci. 1974, 71, 4826–4830. [Google Scholar] [CrossRef]
- Trackman, P.C. Enzymatic and non-enzymatic functions of the lysyl oxidase family in bone. Matrix Biol. 2016, 52-54, 7–18. [Google Scholar] [CrossRef] [PubMed]
- van der Slot-Verhoeven, A.J., et al., The type of collagen cross-link determines the reversibility of experimental skin fibrosis. Biochimica et Biophysica Acta (BBA)-Molecular Basis of Disease, 2005. 1740(1): p. 60-67.
- Nagan, N.; Kagan, H. Modulation of lysyl oxidase activity toward peptidyl lysine by vicinal dicarboxylic amino acid residues. Implications for collagen cross-linking. J. Biol. Chem. 1994, 269, 22366–22371. [Google Scholar] [CrossRef]
- Finney, J., et al., Human copper-dependent amine oxidases. Archives of Biochemistry and Biophysics, 2014. 546(0): p. 19-32.
- Hudson, D.M.; Archer, M.; Rai, J.; Weis, M.; Fernandes, R.J.; Eyre, D.R. Age-related type I collagen modifications reveal tissue-defining differences between ligament and tendon. Matrix Biol. Plus 2021, 12, 100070. [Google Scholar] [CrossRef] [PubMed]
- Hudson, D.M.; Archer, M.; King, K.B.; Eyre, D.R. Glycation of type I collagen selectively targets the same helical domain lysine sites as lysyl oxidase–mediated cross-linking. J. Biol. Chem. 2018, 293, 15620–15627. [Google Scholar] [CrossRef]
- Chen, C.-Y.; Zhang, J.-Q.; Li, L.; Guo, M.-M.; He, Y.-F.; Dong, Y.-M.; Meng, H.; Yi, F. Advanced Glycation End Products in the Skin: Molecular Mechanisms, Methods of Measurement, and Inhibitory Pathways. Front. Med. 2022, 9, 837222. [Google Scholar] [CrossRef]
- Imran, M., et al., Health Benefits of Grapes Polyphenols. J. Environ. Agric. Sci, 2017. 10: p. 40-51.
- Kim, Y.; Keogh, J.B.; Clifton, P.M. Polyphenols and Glycemic Control. Nutrients 2016, 8, 17. [Google Scholar] [CrossRef] [PubMed]
- Stagos, D. Antioxidant Activity of Polyphenolic Plant Extracts. Antioxidants 2019, 9, 19. [Google Scholar] [CrossRef] [PubMed]
- Shishido, S.; Miyano, R.; Nakashima, T.; Matsuo, H.; Iwatsuki, M.; Nakamura, K.; Kanno, T.; Egusa, H.; Niwano, Y. A novel pathway for the photooxidation of catechin in relation to its prooxidative activity. Sci. Rep. 2018, 8, 12888. [Google Scholar] [CrossRef]
- Yamaguchi, K.; Itakura, M.; Kitazawa, R.; Lim, S.-Y.; Nagata, K.; Shibata, T.; Akagawa, M.; Uchida, K. Oxidative deamination of lysine residues by polyphenols generates an equilibrium of aldehyde and 2-piperidinol products. J. Biol. Chem. 2021, 297, 101035. [Google Scholar] [CrossRef]
- Akagawa, M.; Suyama, K. Amine oxidase-like activity of polyphenols. JBIC J. Biol. Inorg. Chem. 2001, 268, 1953–1963. [Google Scholar] [CrossRef]
- Akagawa, M.; Shigemitsu, T.; Suyama, K. Oxidative Deamination of Benzylamine and Lysine Residue in Bovine Serum Albumin by Green Tea, Black Tea, and Coffee. J. Agric. Food Chem. 2005, 53, 8019–8024. [Google Scholar] [CrossRef] [PubMed]
- Bravo, L., Polyphenols: Chemistry, Dietary Sources, Metabolism, and Nutritional Significance. Nutrition Reviews, 2009. 56(11): p. 317-333.
- Prochazkova, D.; Bousova, I.; Wilhelmova, N. Antioxidant and prooxidant properties of flavonoids. Fitoterapia 2011, 82, 513–523. [Google Scholar] [CrossRef]
- Anazco, C.; Rojas, A.; Gonzalez, I.; Castro, M.A.; Robert, P.; Oyarzun-Ampuero, F. Dermal Collagen Stabilization by Polyphenols and Spray Drying as an Encapsulation Strategy. Curr. Top. Med. Chem. 2018, 18, 1242–1251. [Google Scholar] [CrossRef] [PubMed]
- Hanauske-Abel, H.M., et al., Pyrroloquinoline quinone and molecules mimicking its functional domains Modulators of connective tissue formation? FEBS Letters, 1987. 214(2): p. 236-243.
- DiSilvestro, R.A.; Harris, E.D. Evaluation of (+)-catechin action on lysyl oxidase activity in aortic tissue. Biochem. Pharmacol. 1983, 32, 343–346. [Google Scholar] [CrossRef] [PubMed]
- González, I.; Morales, M.A.; Rojas, A. Polyphenols and AGEs/RAGE axis. Trends and challenges. Food Res. Int. 2019, 129, 108843. [Google Scholar] [CrossRef]
- Odjakova, M., et al., Plant-derived agents with anti-glycation activity. Glycosylation, 2012. 10: p. 48186.
- Karamanos, N.K.; Theocharis, A.D.; Piperigkou, Z.; Manou, D.; Passi, A.; Skandalis, S.S.; Vynios, D.H.; Orian-Rousseau, V.; Ricard-Blum, S.; Schmelzer, C.E.; et al. A guide to the composition and functions of the extracellular matrix. FEBS J. 2021, 288, 6850–6912. [Google Scholar] [CrossRef] [PubMed]
- Gauza-Włodarczyk, M.; Kubisz, L.; Włodarczyk, D. Amino acid composition in determination of collagen origin and assessment of physical factors effects. Int. J. Biol. Macromol. 2017, 104, 987–991. [Google Scholar] [CrossRef]
- Gordon, M.K. and R.A. Hahn, Collagens. Cell and Tissue Research, 2010. 339(1): p. 247-257.
- Fidler, A.L.; Boudko, S.P.; Rokas, A.; Hudson, B.G. The triple helix of collagens – an ancient protein structure that enabled animal multicellularity and tissue evolution. J. Cell Sci. 2018, 131, jcs203950. [Google Scholar] [CrossRef]
- Ramachandran, G. and G. Kartha, Structure of collagen. Nature, 1955. 176: p. 593-595.
- Zhang, X.; Xu, S.; Shen, L.; Li, G. Factors affecting thermal stability of collagen from the aspects of extraction, processing and modification. J. Leather Sci. Eng. 2020, 2, 1–29. [Google Scholar] [CrossRef]
- Reiser, K.; Amigable, M.; Last, J. Nonenzymatic glycation of type I collagen. The effects of aging on preferential glycation sites. J. Biol. Chem. 1992, 267, 24207–24216. [Google Scholar] [CrossRef] [PubMed]
- Gautieri, A.; Redaelli, A.; Buehler, M.J.; Vesentini, S. Age- and diabetes-related nonenzymatic crosslinks in collagen fibrils: Candidate amino acids involved in Advanced Glycation End-products. Matrix Biol. 2014, 34, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Eekhoff, J.D.; Fang, F.; Lake, S.P. Multiscale mechanical effects of native collagen cross-linking in tendon. Connect. Tissue Res. 2018, 59, 410–422. [Google Scholar] [CrossRef]
- Sricholpech, M., et al., Lysyl hydroxylase 3-mediated glucosylation in type I collagen: molecular loci and biological significance. The Journal of biological chemistry, 2012. 287(27): p. 22998-23009.
- Cabral-Pacheco, G.A.; Garza-Veloz, I.; la Rosa, C.C.-D.; Ramirez-Acuña, J.M.; A Perez-Romero, B.; Guerrero-Rodriguez, J.F.; Martinez-Avila, N.; Martinez-Fierro, M.L. The Roles of Matrix Metalloproteinases and Their Inhibitors in Human Diseases. Int. J. Mol. Sci. 2020, 21, 9739. [Google Scholar] [CrossRef]
- Shoulders, M.D.; Raines, R.T. Collagen Structure and Stability. Annu. Rev. Biochem. 2009, 78, 929–958. [Google Scholar] [CrossRef] [PubMed]
- Makris, E.A.; MacBarb, R.F.; Responte, D.J.; Hu, J.C.; Athanasiou, K.A. A copper sulfate and hydroxylysine treatment regimen for enhancing collagen cross-linking and biomechanical properties in engineered neocartilage. FASEB J. 2013, 27, 2421–2430. [Google Scholar] [CrossRef]
- Knott, L.; Bailey, A. Collagen cross-links in mineralizing tissues: A review of their chemistry, function, and clinical relevance. Bone 1998, 22, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Reiser, K.; McCormick, R.J.; Rucker, R.B. Enzymatic and nonenzymatic cross-linking of collagen and elastin. FASEB J. 1992, 6, 2439–2449. [Google Scholar] [CrossRef] [PubMed]
- Molnar, J.; Fong, K.; He, Q.; Hayashi, K.; Kim, Y.; Fong, S.; Fogelgren, B.; Szauter, K.M.; Mink, M.; Csiszar, K. Structural and functional diversity of lysyl oxidase and the LOX-like proteins. Biochim. et Biophys. Acta (BBA) - Proteins Proteom. 2003, 1647, 220–224. [Google Scholar] [CrossRef]
- Trackman, P.; Kagan, H. Nonpeptidyl amine inhibitors are substrates of lysyl oxidase. J. Biol. Chem. 1979, 254, 7831–7836. [Google Scholar] [CrossRef]
- Vallet, S.D.; Ricard-Blum, S. Lysyl oxidases: from enzyme activity to extracellular matrix cross-links. Essays Biochem. 2019, 63, 349–364. [Google Scholar] [CrossRef]
- Kagan, H.M.; Li, W. Lysyl oxidase: Properties, specificity, and biological roles inside and outside of the cell. J. Cell. Biochem. 2003, 88, 660–672. [Google Scholar] [CrossRef]
- Liburkin-Dan, T.; Toledano, S.; Neufeld, G. Lysyl Oxidase Family Enzymes and Their Role in Tumor Progression. Int. J. Mol. Sci. 2022, 23, 6249. [Google Scholar] [CrossRef]
- Csiszar, K. Lysyl oxidases: A novel multifunctional amine oxidase family. Prog. Nucleic Acid Res. Mol. Biol. 2001, 70, 1–32. [Google Scholar] [CrossRef]
- Gacheru, S.; Trackman, P.; Shah, M.; O'Gara, C.; Spacciapoli, P.; Greenaway, F.; Kagan, H. Structural and catalytic properties of copper in lysyl oxidase. J. Biol. Chem. 1990, 265, 19022–19027. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Poe, A.; Yus, M.M.; Pak, L.; Nandakumar, K.; Santhanam, L. Lysyl oxidase-like 2 processing by factor Xa modulates its activity and substrate preference. Commun. Biol. 2023, 6, 1–12. [Google Scholar] [CrossRef]
- Añazco, C.; Lopez-Jimenez, A.J.; Rafi, M.; Vega-Montoto, L.; Zhang, M.Z.; Hudson, B.G.; Vanacore, R.M. Lysyl Oxidase-like-2 Cross-links Collagen IV of Glomerular Basement Membrane. J. Biol. Chem. 2016, 291, 25999–26012. [Google Scholar] [CrossRef] [PubMed]
- Bignon, M.; Pichol-Thievend, C.; Hardouin, J.; Malbouyres, M.; Bréchot, N.; Nasciutti, L.; Barret, A.; Teillon, J.; Guillon, E.; Etienne, E.; et al. Lysyl oxidase-like protein-2 regulates sprouting angiogenesis and type IV collagen assembly in the endothelial basement membrane. Blood 2011, 118, 3979–3989. [Google Scholar] [CrossRef] [PubMed]
- Busnadiego, O.; González-Santamaría, J.; Lagares, D.; Guinea-Viniegra, J.; Pichol-Thievend, C.; Muller, L.; Rodríguez-Pascual, F. LOXL4 Is Induced by Transforming Growth Factor β1 through Smad and JunB/Fra2 and Contributes to Vascular Matrix Remodeling. Mol. Cell. Biol. 2013, 33, 2388–2401. [Google Scholar] [CrossRef]
- Aronoff, M.R.; Hiebert, P.; Hentzen, N.B.; Werner, S.; Wennemers, H. Imaging and targeting LOX-mediated tissue remodeling with a reactive collagen peptide. Nat. Chem. Biol. 2021, 17, 865–871. [Google Scholar] [CrossRef]
- Moon, H.-J.; Finney, J.; Xu, L.; Moore, D.; Welch, D.R.; Mure, M. MCF-7 Cells Expressing Nuclear Associated Lysyl Oxidase-like 2 (LOXL2) Exhibit an Epithelial-to-Mesenchymal Transition (EMT) Phenotype and Are Highly Invasive in Vitro. J. Biol. Chem. 2013, 288, 30000–30008. [Google Scholar] [CrossRef]
- Añazco, C.; Delgado-López, F.; Araya, P.; González, I.; Morales, E.; Pérez-Castro, R.; Romero, J.; Rojas, A.; Tecer, D.; Sezgin, M.; et al. Lysyl oxidase isoforms in gastric cancer. Biomarkers Med. 2016, 10, 987–998. [Google Scholar] [CrossRef] [PubMed]
- Añazco, C.; Cerro, S.; Pereira, N.; Rojas, C.; Torres. ; Vidal-Beltrán, I. Dysregulation of Lysyl Oxidases Expression in Diabetic Nephropathy and Renal Cell Carcinoma. Curr. Drug Targets 2021, 22, 1916–1925. [Google Scholar] [CrossRef] [PubMed]
- Miner, J.H., The glomerular basement membrane. Experimental Cell Research, 2012. 318(9): p. 973-978.
- Di Donato, A.; Ghiggeri, G.M.; Di Duca, M.; Jivotenko, E.; Acinni, R.; Campolo, J.; Ginevri, F.; Gusmano, R. Lysyl Oxidase Expression and Collagen Cross-Linking during Chronic Adriamycin Nephropathy. Nephron 1997, 76, 192–200. [Google Scholar] [CrossRef]
- Neusser, M.A.; Lindenmeyer, M.T.; Moll, A.G.; Segerer, S.; Edenhofer, I.; Sen, K.; Stiehl, D.P.; Kretzler, M.; Gröne, H.-J.; Schlöndorff, D.; et al. Human Nephrosclerosis Triggers a Hypoxia-Related Glomerulopathy. Am. J. Pathol. 2010, 176, 594–607. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.-Y.; Li, Q.; Wong, W.R.; N’diaye, E.-N.; Caplazi, P.; Bender, H.; Huang, Z.; Arlantico, A.; Jeet, S.; Wong, A.; et al. LOXL4, but not LOXL2, is the critical determinant of pathological collagen cross-linking and fibrosis in the lung. Sci. Adv. 2023, 9, eadf0133. [Google Scholar] [CrossRef]
- Setargew, Y.F.; Wyllie, K.; Grant, R.D.; Chitty, J.L.; Cox, T.R. Targeting Lysyl Oxidase Family Meditated Matrix Cross-Linking as an Anti-Stromal Therapy in Solid Tumours. Cancers 2021, 13, 491. [Google Scholar] [CrossRef] [PubMed]
- Grau-Bové, X.; Ruiz-Trillo, I.; Rodriguez-Pascual, F. Origin and evolution of lysyl oxidases. Sci. Rep. 2015, 5, 10568. [Google Scholar] [CrossRef]
- Bollinger, J.A.; Brown, D.E.; Dooley, D.M. The Formation of Lysine Tyrosylquinone (LTQ) Is a Self-Processing Reaction. Expression and Characterization of a Drosophila Lysyl Oxidase. Biochemistry 2005, 44, 11708–11714. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, Q.; Wu, J.; Wang, J.; Shi, Y.; Liu, M. Crystal structure of human lysyl oxidase-like 2 (hLOXL2) in a precursor state. Proc. Natl. Acad. Sci. 2018, 115, 3828–3833. [Google Scholar] [CrossRef]
- Shanbhag, V.; Jasmer-McDonald, K.; Zhu, S.; Martin, A.L.; Gudekar, N.; Khan, A.; Ladomersky, E.; Singh, K.; Weisman, G.A.; Petris, M.J. ATP7A delivers copper to the lysyl oxidase family of enzymes and promotes tumorigenesis and metastasis. Proc. Natl. Acad. Sci. USA 2019, 116, 6836–6841. [Google Scholar] [CrossRef] [PubMed]
- Vallet, S.D.; Guéroult, M.; Belloy, N.; Dauchez, M.; Ricard-Blum, S. A Three-Dimensional Model of Human Lysyl Oxidase, a Cross-Linking Enzyme. ACS Omega 2019, 4, 8495–8505. [Google Scholar] [CrossRef] [PubMed]
- Meier, A.A., K. Kuczera, and M. Mure, A 3D–Predicted Structure of the Amine Oxidase Domain of Lysyl Oxidase–Like 2. International Journal of Molecular Sciences, 2022. 23(21): p. 13385.
- Meier, A.A.; Moon, H.-J.; Sabuncu, S.; Singh, P.; Ronnebaum, T.A.; Ou, S.; Douglas, J.T.; Jackson, T.A.; Moënne-Loccoz, P.; Mure, M. Insight into the Spatial Arrangement of the Lysine Tyrosylquinone and Cu2+ in the Active Site of Lysyl Oxidase-like 2. Int. J. Mol. Sci. 2022, 23, 13966. [Google Scholar] [CrossRef]
- Mure, M. Tyrosine-Derived Quinone Cofactors. Accounts Chem. Res. 2004, 37, 131–139. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Varadi, M.; Anyango, S.; Deshpande, M.; Nair, S.; Natassia, C.; Yordanova, G.; Yuan, D.; Stroe, O.; Wood, G.; Laydon, A.; et al. AlphaFold Protein Structure Database: massively expanding the structural coverage of protein-sequence space with high-accuracy models. Nucleic Acids Res. 2022, 50, D439–D444. [Google Scholar] [CrossRef] [PubMed]
- Okada, K.; Moon, H.-J.; Finney, J.; Meier, A.; Mure, M. Extracellular Processing of Lysyl Oxidase-like 2 and Its Effect on Amine Oxidase Activity. Biochemistry 2018, 57, 6973–6983. [Google Scholar] [CrossRef]
- Xu, L.; Go, E.P.; Finney, J.; Moon, H.; Lantz, M.; Rebecchi, K.; Desaire, H.; Mure, M. Post-translational Modifications of Recombinant Human Lysyl Oxidase-like 2 (rhLOXL2) Secreted from Drosophila S2 Cells*. J. Biol. Chem. 2013, 288, 5357–5363. [Google Scholar] [CrossRef] [PubMed]
- Sell, D.R.; Monnier, V.M. Structure Elucidation of a Senescence Cross-Link from Human Extracellular Matrix. J. Biol. Chem. 1989, 264, 21597–21602. [Google Scholar] [CrossRef] [PubMed]
- Eyre, D.R.; Weis, M.A.; Wu, J.-J. Maturation of Collagen Ketoimine Cross-links by an Alternative Mechanism to Pyridinoline Formation in Cartilage. J. Biol. Chem. 2010, 285, 16675–16682. [Google Scholar] [CrossRef]
- Hanson, D.A.; Eyre, D.R. Molecular Site Specificity of Pyridinoline and Pyrrole Cross-links in Type I Collagen of Human Bone. J. Biol. Chem. 1996, 271, 26508–26516. [Google Scholar] [CrossRef]
- Eyre, D.R.; Weis, M.; Rai, J. Analyses of lysine aldehyde cross-linking in collagen reveal that the mature cross-link histidinohydroxylysinonorleucine is an artifact. J. Biol. Chem. 2019, 294, 6578–6590. [Google Scholar] [CrossRef]
- Teuscher, A.C., et al., Assessing collagen deposition during aging in mammalian tissue and in Caenorhabditis elegans, in Collagen. 2019, Springer. p. 169-188.
- Fenske, N.A.; Lober, C.W. Structural and functional changes of normal aging skin. J. Am. Acad. Dermatol. 1986, 15, 571–585. [Google Scholar] [CrossRef]
- Fisher, G.J.; Quan, T.; Purohit, T.; Shao, Y.; Cho, M.K.; He, T.; Varani, J.; Kang, S.; Voorhees, J.J. Collagen Fragmentation Promotes Oxidative Stress and Elevates Matrix Metalloproteinase-1 in Fibroblasts in Aged Human Skin. Am. J. Pathol. 2009, 174, 101–114. [Google Scholar] [CrossRef] [PubMed]
- Reiser, K.M., Nonenzymatic glycation of collagen in aging and diabetes. Proceedings of the Society for Experimental Biology and Medicine, 1998. 218(1): p. 23-37.
- Varani, J.; Dame, M.K.; Rittie, L.; Fligiel, S.E.; Kang, S.; Fisher, G.J.; Voorhees, J.J. Decreased Collagen Production in Chronologically Aged Skin: Roles of Age-Dependent Alteration in Fibroblast Function and Defective Mechanical Stimulation. Am. J. Pathol. 2006, 168, 1861–1868. [Google Scholar] [CrossRef] [PubMed]
- Bucala, R. and A. Cerami, Advanced glycosylation: chemistry, biology, and implications for diabetes and aging. Adv pharmacol, 1992. 23(1-34): p. 13.
- Rojas, A.; Morales, M.A. Advanced glycation and endothelial functions: A link towards vascular complications in diabetes. Life Sci. 2004, 76, 715–730. [Google Scholar] [CrossRef] [PubMed]
- Fournet, M., F. Bonté, and A. Desmoulière, Glycation Damage: A Possible Hub for Major Pathophysiological Disorders and Aging. Aging Dis, 2018. 9(5): p. 880-900.
- Brings, S.; Fleming, T.; Freichel, M.; Muckenthaler, M.U.; Herzig, S.; Nawroth, P.P. Dicarbonyls and Advanced Glycation End-Products in the Development of Diabetic Complications and Targets for Intervention. Int. J. Mol. Sci. 2017, 18, 984. [Google Scholar] [CrossRef]
- Meade, S.J.; Miller, A.G.; A Gerrard, J. The role of dicarbonyl compounds in non-enzymatic crosslinking: a structure–activity study. Bioorganic Med. Chem. 2003, 11, 853–862. [Google Scholar] [CrossRef] [PubMed]
- Sell, D.R. and V.M. Monnier, Aging of long-lived proteins: Extracellular matrix (collagens, elastins, proteoglycans) and lens crystallins. Comprehensive physiology, 2010: p. 235-305.
- Wautier, J.-L. and P.-J. Guillausseau, Diabetes, advanced glycation endproducts and vascular disease. Vascular Medicine, 1998. 3(2): p. 131-137.
- Nash, A.; Notou, M.; Lopez-Clavijo, A.F.; Bozec, L.; de Leeuw, N.H.; Birch, H.L. Glucosepane is associated with changes to structural and physical properties of collagen fibrils. Matrix Biol. Plus 2019, 4, 100013. [Google Scholar] [CrossRef]
- Bansode, S.; Bashtanova, U.; Li, R.; Clark, J.; Müller, K.H.; Puszkarska, A.; Goldberga, I.; Chetwood, H.H.; Reid, D.G.; Colwell, L.J.; et al. Glycation changes molecular organization and charge distribution in type I collagen fibrils. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef]
- Fessel, G.; Li, Y.; Diederich, V.; Guizar-Sicairos, M.; Schneider, P.; Sell, D.R.; Monnier, V.M.; Snedeker, J.G. Advanced Glycation End-Products Reduce Collagen Molecular Sliding to Affect Collagen Fibril Damage Mechanisms but Not Stiffness. PLOS ONE 2014, 9, e110948. [Google Scholar] [CrossRef] [PubMed]
- Rojas, A.; Añazco, C.; González, I.; Araya, P. Extracellular matrix glycation and receptor for advanced glycation end-products activation: a missing piece in the puzzle of the association between diabetes and cancer. Carcinog. 2018, 39, 515–521. [Google Scholar] [CrossRef]
- Sell, D.R., et al., Glucosepane is a major protein cross-link of the senescent human extracellular matrix: relationship with diabetes. Journal of Biological Chemistry, 2005. 280(13): p. 12310-12315.
- Vaez, M.; Asgari, M.; Hirvonen, L.; Bakir, G.; Khattignavong, E.; Ezzo, M.; Aguayo, S.; Schuh, C.M.; Gough, K.; Bozec, L. Modulation of the biophysical and biochemical properties of collagen by glycation for tissue engineering applications. Acta Biomater. 2023, 155, 182–198. [Google Scholar] [CrossRef]
- Monnier, V.M.; Sell, D.R.; Strauch, C.; Sun, W.; Lachin, J.M.; Cleary, P.A.; Genuth, S. The association between skin collagen glucosepane and past progression of microvascular and neuropathic complications in type 1 diabetes. J. Diabetes its Complicat. 2012, 27, 141–149. [Google Scholar] [CrossRef]
- Bravo, L., Polyphenols: chemistry, dietary sources, metabolism, and nutritional significance. Nutrition reviews, 1998. 56(11): p. 317-333.
- Prochazkova, D.; Bousova, I.; Wilhelmova, N. Antioxidant and prooxidant properties of flavonoids. Fitoterapia 2011, 82, 513–523. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [PubMed]
- Lucarini, M.; Sciubba, F.; Capitani, D.; Di Cocco, M.E.; D’evoli, L.; Durazzo, A.; Delfini, M.; Boccia, G.L. Role of catechin on collagen type I stability upon oxidation: a NMR approach. Nat. Prod. Res. 2019, 34, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Porto, I.C.C.M.; Nascimento, T.G.; Oliveira, J.M.S.; Freitas, P.H.; Haimeur, A.; França, R. Use of polyphenols as a strategy to prevent bond degradation in the dentin-resin interface. Eur. J. Oral Sci. 2018, 126, 146–158. [Google Scholar] [CrossRef]
- Nazaruk, J. and A. Galicka, The influence of selected flavonoids from the leaves of Cirsium palustre (L.) Scop. on collagen expression in human skin fibroblasts. Phytotherapy Research, 2014. 28(9): p. 1399-1405.
- Zhang, Y., et al., Apigenin induces dermal collagen synthesis via smad2/3 signaling pathway. European journal of histochemistry: EJH, 2015. 59(2).
- Bae, J.; Lim, S.S.; Kim, S.J.; Choi, J.; Park, J.; Ju, S.M.; Han, S.J.; Kang, I.; Kang, Y. Bog blueberry anthocyanins alleviate photoaging in ultraviolet-B irradiation-induced human dermal fibroblasts. Mol. Nutr. Food Res. 2009, 53, 726–738. [Google Scholar] [CrossRef]
- Wittenauer, J.; Mäckle, S.; Sußmann, D.; Schweiggert-Weisz, U.; Carle, R. Inhibitory effects of polyphenols from grape pomace extract on collagenase and elastase activity. Fitoterapia 2015, 101, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Furuhashi, M.; Hatasa, Y.; Kawamura, S.; Shibata, T.; Akagawa, M.; Uchida, K. Identification of Polyphenol-Specific Innate Epitopes That Originated from a Resveratrol Analogue. Biochemistry 2017, 56, 4701–4712. [Google Scholar] [CrossRef]
- Bonnans, C.; Chou, J.; Werb, Z. Remodelling the extracellular matrix in development and disease. Nat. Rev. Mol. Cell Biol. 2014, 15, 786–801. [Google Scholar] [CrossRef] [PubMed]
- Stirban, A.; Gawlowski, T.; Roden, M. Vascular effects of advanced glycation endproducts: Clinical effects and molecular mechanisms. Mol. Metab. 2013, 3, 94–108. [Google Scholar] [CrossRef]
- Twarda-Clapa, A.; Olczak, A.; Białkowska, A.M.; Koziołkiewicz, M. Advanced Glycation End-Products (AGEs): Formation, Chemistry, Classification, Receptors, and Diseases Related to AGEs. Cells 2022, 11, 1312. [Google Scholar] [CrossRef]
- Bierhaus, A., et al., Understanding RAGE, the receptor for advanced glycation end products. J Mol Med (Berl), 2005. 83(11): p. 876-86.
- Verzijl, N.; DeGroot, J.; Ben Zaken, C.; Braun-Benjamin, O.; Maroudas, A.; Bank, R.A.; Mizrahi, J.; Schalkwijk, C.G.; Thorpe, S.R.; Baynes, J.W.; et al. Crosslinking by advanced glycation end products increases the stiffness of the collagen network in human articular cartilage: A possible mechanism through which age is a risk factor for osteoarthritis. Arthritis Rheum. 2002, 46, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Bhuiyan, M.N.I.; Mitsuhashi, S.; Sigetomi, K.; Ubukata, M. Quercetin inhibits advanced glycation end product formation via chelating metal ions, trapping methylglyoxal, and trapping reactive oxygen species. Biosci. Biotechnol. Biochem. 2017, 81, 882–890. [Google Scholar] [CrossRef]
- Hwang, S.H.; Kim, H.Y.; Zuo, G.; Wang, Z.; Lee, J.-Y.; Lim, S.S. Anti-glycation, Carbonyl Trapping and Anti-inflammatory Activities of Chrysin Derivatives. Molecules 2018, 23, 1752. [Google Scholar] [CrossRef]
- Lv, L.; Shao, X.; Chen, H.; Ho, C.-T.; Sang, S. Genistein Inhibits Advanced Glycation End Product Formation by Trapping Methylglyoxal. Chem. Res. Toxicol. 2011, 24, 579–586. [Google Scholar] [CrossRef] [PubMed]
- Sang, S.; Shao, X.; Bai, N.; Lo, C.-Y.; Yang, C.S.; Ho, C.-T. Tea Polyphenol (−)-Epigallocatechin-3-Gallate: A New Trapping Agent of Reactive Dicarbonyl Species. Chem. Res. Toxicol. 2007, 20, 1862–1870. [Google Scholar] [CrossRef] [PubMed]
- Intagliata, S.; Spadaro, A.; Lorenti, M.; Panico, A.; Siciliano, E.A.; Barbagallo, S.; Macaluso, B.; Kamble, S.H.; Modica, M.N.; Montenegro, L. In Vitro Antioxidant and Anti-Glycation Activity of Resveratrol and Its Novel Triester with Trolox. Antioxidants 2020, 10, 12. [Google Scholar] [CrossRef]
- Lee, S.M.; Zheng, L.W.; Jung, Y.; Hwang, G.-S.; Kim, Y.-S. Effects of hydroxycinnamic acids on the reduction of furan and α-dicarbonyl compounds. Food Chem. 2019, 312, 126085. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, H.; Yang, X.; Zhu, L.; Wu, G.; Qi, X.; Zhang, H. Trapping of reactive carbonyl species by fiber-bound polyphenols from whole grains under simulated physiological conditions. Food Res. Int. 2022, 156, 111142. [Google Scholar] [CrossRef] [PubMed]
- Mienaltowski, M.J., et al., Basic Structure, Physiology, and Biochemistry of Connective Tissues and Extracellular Matrix Collagens. Adv Exp Med Biol, 2021. 1348: p. 5-43.
- Wu, L.; Shao, H.; Fang, Z.; Zhao, Y.; Cao, C.Y.; Li, Q. Mechanism and Effects of Polyphenol Derivatives for Modifying Collagen. ACS Biomater. Sci. Eng. 2019, 5, 4272–4284. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Post-translational modifications involved in the formation of enzymatic collagen covalent cross-linking. Structure of general collagen-forming reaction pathway and its constituent amino acids. Procollagen molecules convert to trimeric propeptide fragments that form tropocollagen molecules. Tropocollagen molecules self-assemble via the reaction of aldehyde groups and the formation of covalent bonds that cross-link collagen molecules into fibrils and fibers. A. The intracellular lysine modification to hydroxylysine by lysyl hydroxylase. Ascorbate and iron act as co-factors for the proline and lysine hydroxylases that stabilize the tertiary structure of collagen molecules. B. The intracellular triple helix formation highlighting hydroxylysine residues. C. Extracellular lysine oxidation and deamination of lysine to allysine and hydroxylysine to hydroxyallysine. The LOX superfamily modifies lysine and hydroxylysine residues in collagen post-translationally. D and E showing mature tropocollagen cross-linked, which organize into collagen fibrils and assemble into collagen fibers.
Figure 1.
Post-translational modifications involved in the formation of enzymatic collagen covalent cross-linking. Structure of general collagen-forming reaction pathway and its constituent amino acids. Procollagen molecules convert to trimeric propeptide fragments that form tropocollagen molecules. Tropocollagen molecules self-assemble via the reaction of aldehyde groups and the formation of covalent bonds that cross-link collagen molecules into fibrils and fibers. A. The intracellular lysine modification to hydroxylysine by lysyl hydroxylase. Ascorbate and iron act as co-factors for the proline and lysine hydroxylases that stabilize the tertiary structure of collagen molecules. B. The intracellular triple helix formation highlighting hydroxylysine residues. C. Extracellular lysine oxidation and deamination of lysine to allysine and hydroxylysine to hydroxyallysine. The LOX superfamily modifies lysine and hydroxylysine residues in collagen post-translationally. D and E showing mature tropocollagen cross-linked, which organize into collagen fibrils and assemble into collagen fibers.
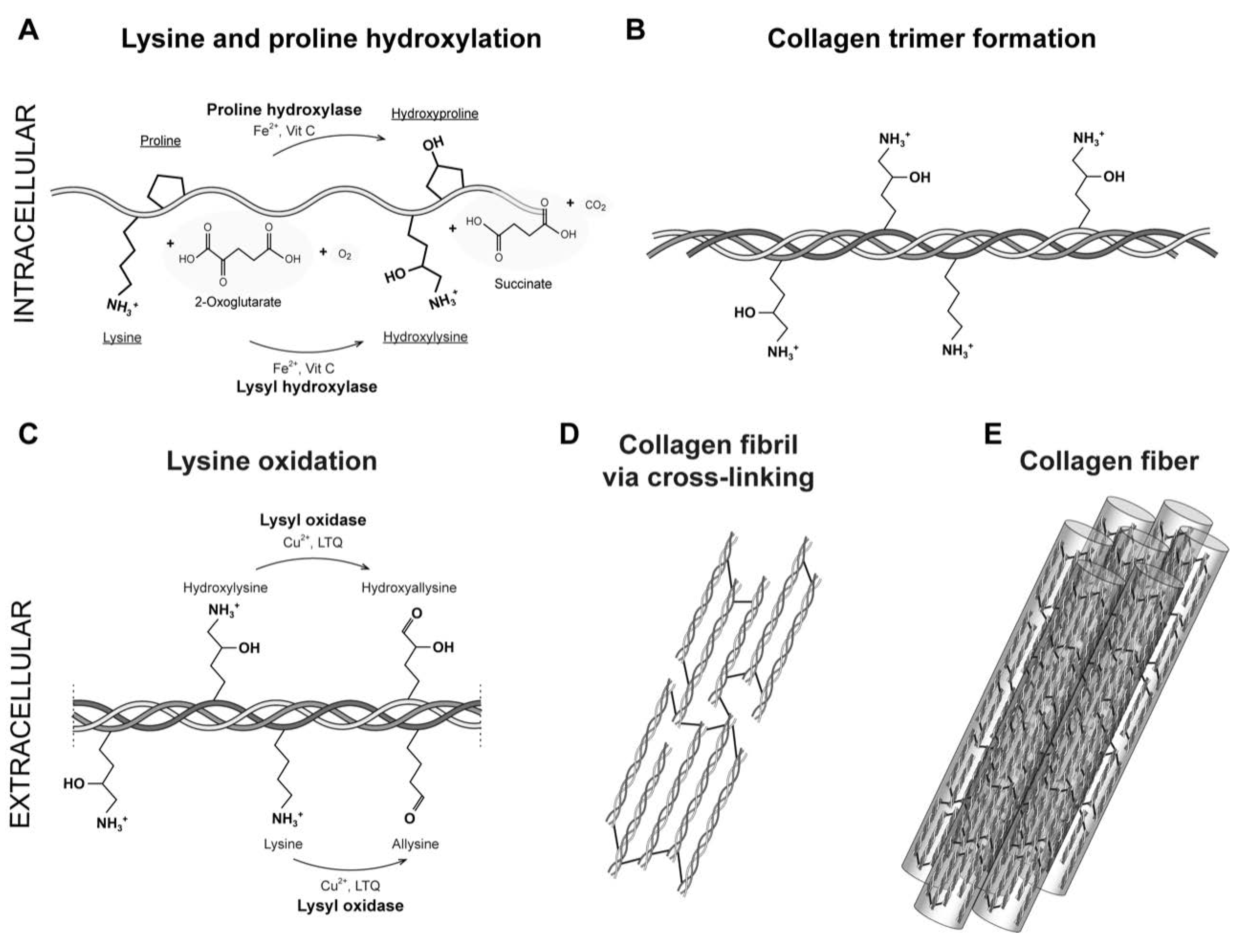
Figure 2.
The lysyl oxidase family. A. LOX isoforms contains a signal peptide (SP) in the amino-terminal region and a catalytic domain in the highly conserved carboxyl-terminal region. The catalytic domain includes a copper-binding domain (Cu), lysyl tyrosylquinone cofactor (LTQ) and cytokine receptor-like domain (CRL), which are also present in isoforms LOXL1, LOXL2, LOXL3 and LOXL4. B. AlphaFold model of hLOX lysyl oxidase-like domain represented in NewCartoon. The color illustrates the confidence of the model structure with white indicating low confidence regions and blue high confidence regions. Highlighted are the copper binding site forming residues H292, H294 and H296, and LTQ cofactor residues K320 and Y355. C. Detailed view of the residues highlighted in B. Note that the confidence of the structure around residue K320 is low which indicated uncertainty in this region (see main text for more information). .
Figure 2.
The lysyl oxidase family. A. LOX isoforms contains a signal peptide (SP) in the amino-terminal region and a catalytic domain in the highly conserved carboxyl-terminal region. The catalytic domain includes a copper-binding domain (Cu), lysyl tyrosylquinone cofactor (LTQ) and cytokine receptor-like domain (CRL), which are also present in isoforms LOXL1, LOXL2, LOXL3 and LOXL4. B. AlphaFold model of hLOX lysyl oxidase-like domain represented in NewCartoon. The color illustrates the confidence of the model structure with white indicating low confidence regions and blue high confidence regions. Highlighted are the copper binding site forming residues H292, H294 and H296, and LTQ cofactor residues K320 and Y355. C. Detailed view of the residues highlighted in B. Note that the confidence of the structure around residue K320 is low which indicated uncertainty in this region (see main text for more information). .
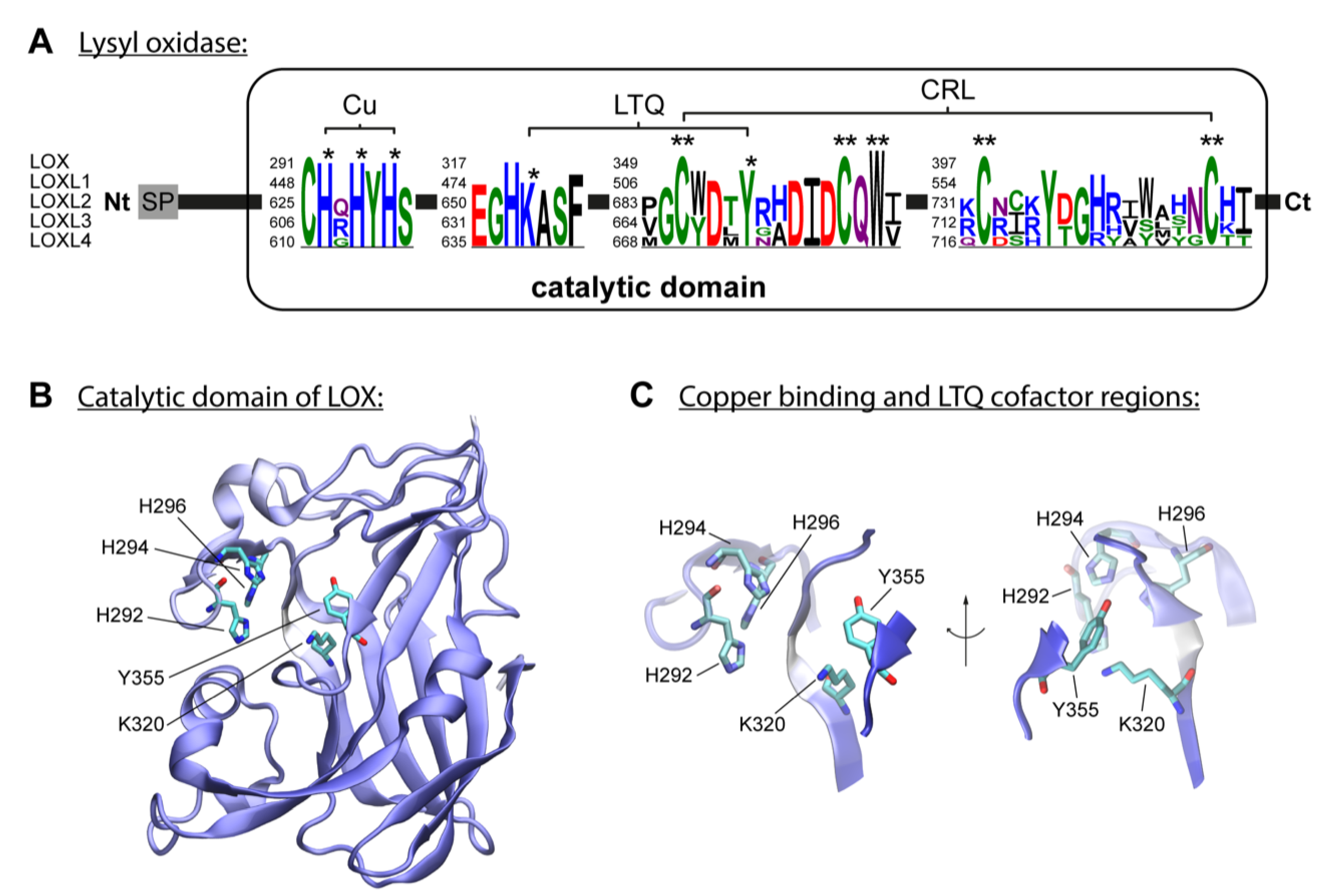
Figure 3.
Lysine Glycation of Collagen Fibers. Extracellular lysine glycation by glucose to form AGE products. A. Reducing glucose possess a reactive carbonyl moiety that reacts non-enzymatically with the ε-amino group of lysine, to form a labile compound, a Schiff base. B and C showing mature tropocollagen cross-linked by glycation, with disorganized collagen fibrils and suprafibrillar architecture. The bonds in red color represent collagen glycation.
Figure 3.
Lysine Glycation of Collagen Fibers. Extracellular lysine glycation by glucose to form AGE products. A. Reducing glucose possess a reactive carbonyl moiety that reacts non-enzymatically with the ε-amino group of lysine, to form a labile compound, a Schiff base. B and C showing mature tropocollagen cross-linked by glycation, with disorganized collagen fibrils and suprafibrillar architecture. The bonds in red color represent collagen glycation.
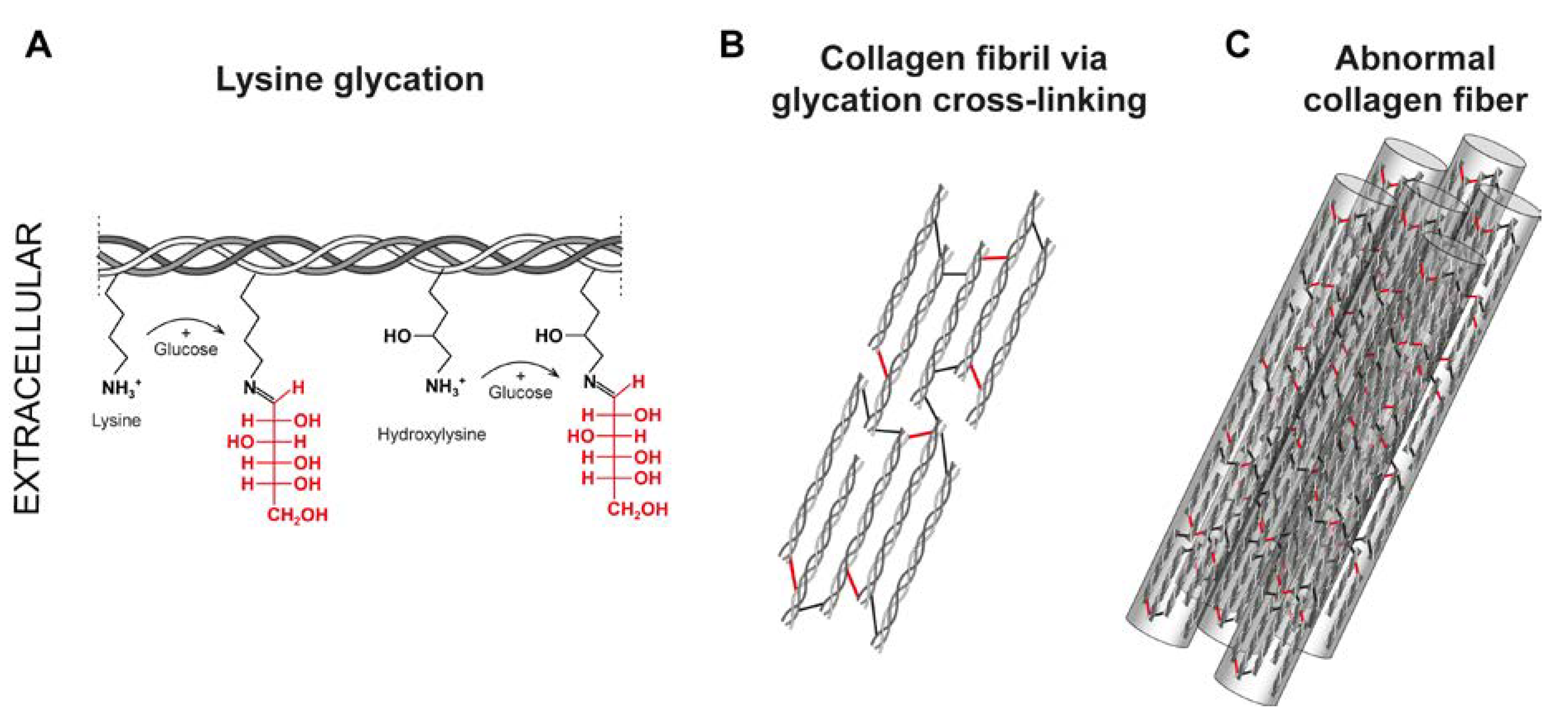
Figure 4.
Catechol- type polyphenols: Chemical structures of catechin derivatives, which are proposed as putative quinone cofactors due to their amine oxidase-like activity, which could be attributed to polyphenols, or where these polyphenols could be required. Catechin (C), epicatechin (EC), epigallocatechin (EGC), epigallocatechin (EGCG), gallocatechin gallate (GCG), epicatechin gallate (ECG).
Figure 4.
Catechol- type polyphenols: Chemical structures of catechin derivatives, which are proposed as putative quinone cofactors due to their amine oxidase-like activity, which could be attributed to polyphenols, or where these polyphenols could be required. Catechin (C), epicatechin (EC), epigallocatechin (EGC), epigallocatechin (EGCG), gallocatechin gallate (GCG), epicatechin gallate (ECG).

Table 1.
Types of covalent collagen cross-links.
| Immature cross-links | Mature cross-links | Glycation derived cross-links |
|---|---|---|
|
Aldimine cross-links 1. Dehydro-lysinonorleucine (deH-NL) 2. Dehydro-hydroxylysinonorleucine (deH-HLNL) 3. Dehydro-dihydroxylysinonorleucine (deH-DHLNL) |
Pyrrole containing cross-links 1. Deoxypyrrololine (d-Prl) 2. Pyrrololine (Prl) Pyridinium-salt containing cross-links 1. Pyridinoline (Pyr) 2. Deoxypyridinoline (Dpyr) |
Alpha-dicarbonyl compounds (α-DC) 1. Methylglyoxal (MGO) 2. Glyoxal (GO) |
|
Ketoamine cross-links 1. Lysine-keto-norleucine (LKNL) 2. Hydroxylysine-keto-norleucine (HLKNL) |
Histidine containing cross-links 1. Histidinohydroxylysinonorleucine (HHL) 2. Histidinohydroxymerodesmosine (HHMD) Arginoline Ketoimine cross-link derivate (Eyre et al. 2010) |
Fluorescent cross-linking AGEs 1. Pentosidine (PEN) Non-Fluorescent cross-linking AGEs 1. Carboxymethyl-lysine (CML) 2. Glucosepane (GSP) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



