
Submitted:
29 May 2023
Posted:
30 May 2023
You are already at the latest version
Abstract
The inflammatory bowels diseases (IBD) are autoimmune diseases that deeply impact the patients’ quality of life. The IBD pathogenesis is not yet defined, but evidence demonstrated that the IBD chronic inflammation is related to an impaired intestinal barrier. Traditionally, two actors were considered for their contribution to this disfunction: the gut microbiota and intestinal epithelium. However, a third element, which is the intestinal mucus, should be considered as peer of the epithelium and microbiota. Indeed, mucus represents the biological interface between bacteria and cells, filtering molecules or toxins and preventing bacteria penetration exploiting both structural and compositional properties. The boosting effect of the mucus characterization towards IBD comprehension is far too underestimated, although some mucus-oriented studies are already reported in literature. This work reviews the intestinal barrier features, describing each component of the gut mucosa (i.e., epithelium, microbiota, and mucus) in a mucus-oriented perspective.
Keywords:
dysbiosis
; rheology
; mucosal barrier
; autoimmune
; microbiota
1. Introduction
In the last years, the prevalence of autoimmune diseases increased worldwide[1,2,3]. More than 60 different pathologies are grouped as autoimmune diseases by the World Health Organization, (WHO) without stressing the differences, but focusing on the common aspects: immune cells (or their products) react abnormally to the presence of the body self-antigens, triggering non-physiological bio-mechanisms that lead to inflammation and other dysfunctions [4,5].
The prevalence of autoimmune diseases is growing worldwide particularly considering pathologies related to the gastrointestinal tract with a net increase of ~ 6% per year [3]. The severity and the impact on patients’ life depend on the specific gastrointestinal pathology. For instance, diseases such as celiac disease can be easily controlled and managed through gluten-free diets, while the inflammatory bowel diseases (IBD), such as Crohn’s disease (CD) and ulcerative colitis (UC), maybe deeply debilitating and lacking a resolutive drug therapy heading for the remission of symptoms and the prevention of relapse [6].
In healthy condition, the gut protects the human body as a shield against pathogens and toxins, while governing the communication between cells and the overwhelming number of microorganisms, collectively called microbiota, that symbiotically grow in the human intestine[7]. Even if the microbiota and gut-tissue crosstalk are not yet completely explored, their potential in modulating the human health are leading to new therapeutic strategies, which are well-represented by the so-called pro-, pre- and post-biotic approaches [8,9,10].
Although the microbiota and gut epithelial cells are unanimously recognized to be strictly dependent [11,12], they do not directly act on each other but by means of a third, cell-free, intermediator: the intestinal mucus.
The intestinal mucus is a biological hydrogel covering the entire gastrointestinal tract [13]. It supports the microbiota growth and modulates, through specific physico-chemical and structural properties, the communication between bacteria and cells [13] by selectively filtering signal molecules or altering their absorption profile. Despite this, the mucus is rarely addressed with the dignity that it deserves [14,15,16] as a peer of microbiota and cells on the gut barrier properties in the study of IBD. Independently from the specific aim of the research, when a complex biological phenomenon is studied regarding the intestinal barrier, a mucus-including perspective, other than a purely cellular/microbiological one, could bring new insights in the etiology of these diseases by providing a “microbiota-mucus-epithelium” triad generating the final pathological scenario [17].
In this review, we present each component of the “microbiota-mucus-epithelium” triad separately. We particularly focus on how the mucus properties, under a materialistic point of view, may contribute as a cell-free intermediator of the biological actors in the pathological scenario of the IBDs.
2. General aspect and relevance of Inflammatory Bowel Disease
From 1990 to 2017, the number of patients diagnosed with autoimmune IBD doubled from ~ 4 to ~ 8 millions, with outstanding increase in respect to the past decades [18].
The diagnosis of CD and UC is not a simple process (Figure 1), as they are studied with techniques developed to highlight the shared molecular mechanisms that trigger the inflammatory response of cells. Abdominal pain, severe incontinence (more than 10 evacuations per day), colon dilation, continuous bleeding and weight loss, are specific indicators of UC but not of CD [19]. In this last case, highly variable symptomatology combined with other techniques as endoscopy, radiology and histology, makes the analysis of the patient's history the preferential approach to the disease management [1,15]. In case of CD, the inflammation can be patchy extend to different districts of the gastrointestinal (GI) tract, with frequency in the ileum and/or colon (95-98%), and, in just 2-5% of cases, in the upper GI tract [16,17]. Differently, the UC is a disease located in the colon, which not usually involve the whole thickness of mucosa (on the opposite of CD)[20]. According to the disease severity, the UC inflammation could extend until to involve the entire colon (i.e., pancolitis)[22].
CD and UC they are commonly recognized as relapsing inflammatory diseases developed in patients with genetic predisposition combined with environmental factors (e.g., diet, smoke, sanitation and socio-economic status)[23]. Intense industrial and traffic-derived pollutants (i.e., NO2 and sulphur dioxide, respectively) were associated to the IBD disease development in a still unknown age-dependent mechanism. Moreover, higher cumulative smoking (i.e. the number of cigarettes packages per year) was associated to a higher risk of CD development [24]. Interestingly, smoking cessation decreases the probability to incur in CD but increases the probability to incur in UC [25]. These different effects suggest that environmental factors should not be undervalued in the study of IBD, but instead should be considered as a contributing cause with the genetic predisposition to the IBD development.
Although the specific causes of CD and UC remain unknown [26] UC and CD have historically been considered as two distinct diseases, the hypothesis of a common origin has been proposed in recent years [27,28]. Regardless the experimental evidence that support this hypothesis or not, the scientific community is unanimous in identifying the changes in either gut microbiota or mucus or cellular barrier dysfunctions as fundamental phenomena to be considered, whether they were interpreted in terms of causes or symptoms [29,30,31]. The synergy with which the components of the “microbiota-mucus-epithelium” triad generates the final phenomenon of the leaky-gut is underestimated, independently from how the biological events are considered. It is therefore essential to optimize the traditional methods used for IBD studies - and in general mucosal-associated diseases - developing new technologies and tools that are able not only to the investigate the triad as a whole, but also to distinguish one by one the contribution that each actor performs towards the final result [11,32,33,34,35]
3. Intestinal epithelial cells in Inflammatory Bowel Disease
The epithelial barrier is a complex biological structure made of different type of highly specialized cells, which is defective in both CD and UC [36,37]. Dysbiosis and inflammation are the two main factors studied for their effect on the cellular barrier integrity, metabolism and functionality. Butyrate-producing bacteria influence several barrier-related molecular pathways (Figure 2). Colonocytes (enterocytes of colon region) metabolize butyrate through β-oxidation process, consuming free-oxygen and establishing an anaerobic environment thus promoting obligate anaerobes over facultative anaerobes proliferation (i.e. Bacteroides fragilis over Escherichia coli) [31,37]. Furthermore, butyrate regulates Treg cells by promoting the expression of Foxp3 gene [38], which is one of the key transcription factors triggering the Treg cell maturation [39]. A dysbiotic condition could lead not only to a reduction of butyrate-producing bacteria, but also to the shift of colonocyte metabolism from β-oxidation to anaerobic glycolysis. This change increases levels of oxygen and nitrates and promotes the facultative anaerobes proliferation but also pathogens colonization [40]. Similarly, the butyrate depletion leads to a decrease in Treg cell maturation promoting inflammation and pro-inflammatory molecules, such as defensins and bacteriocin [41,42] (Figure 2).
Gut dysbiosis alters also intestinal epithelium function by impairing the cellular barrier permeability. The maintenance of the epithelium integrity allows a regulated flux of nutrients, water and ions and avoids the passage of bacterial molecules or microorganisms. This regulation mechanism is mainly due to the presence of tight junctions (TJ) between epithelium cells [43]. TJs are composed by different proteins, such as claudins, occludin and others (e.g. zonula occludens-1, ZO-1 protein)[44,45], which are sensitive to inflammatory mediators and in particular the tumor necrosis factor-α (TNFα), among others [46,47,48,49]. TNFα is one of the main pro-inflammatory factors produced by T cells, macrophages and monocytes during inflammation in both CD and UC [50]. This cytokine activates different and complex signaling pathways that lead to the endocytosis of occludins and to the promotion of claudin-2 expression, which forms channels for the flux of small cations and water and is associated to leaky epithelia and intestinal barrier integrity impairment in both CD and UC [43,49,51,52].
Inflammatory mediators showed a role in epithelium integrity regulation also by triggering the nitric oxide synthase (NOS) pathway [53]. Nitric oxide (NO) is a reactive molecule synthesized by the inducible NOS (iNOS) through the oxidation of L-arginine, leading to the production of citrullin and NO. In physiological conditions, NO protects the epithelium integrity from oxidative stress-mediated by reactive oxygen species [54]. Instead, in pathological conditions, iNOS activity seems to have an opposite role as it contributes to an increase in intestinal permeability and correlates with prolonged gut inflammation [55]. Indeed, iNOS activity was found significantly higher in colonic biopsies of UC patients respect to control subjects. They showed an increasing colonic level of citrullin and thus, of NO. TNFα or the combination of IL-1α and IFN-γ are the key mediators of iNOS expression upregulation [53].
Interestingly, studies on the use of exogenous NO to treat a colonic epithelium model showed a protective role also at higher doses, in contradiction with what observed with native NO [54]. The presence of conflicting results in the literature may be indicative of the fact that cellular models, although useful tools for the decrypt the complex molecular pathways, are currently unable to model the biological phenomenon as a whole. In vitro models including non-biological components, such as advanced mucus models, could allow to understand whether the contradictions in the literature describes a real physio-pathological phenomenon or a bias, born by purely biological-oriented approaches without synergy with materialistic-oriented ones.
4. Intestinal microbiota and dysbiosis in Inflammatory Bowel Disease
More than 100 trillion of microorganisms inhabit the human intestine and compose the gut microbiota, including fungi, bacteria, and yeast. In a physiological condition, bacterial population is highly diversified and mainly characterized by the phyla Firmicutes (i.e. Lactobacillus and Clostridium), Actinobacteria (i.e. Bifidobacterium), Bacteroidetes (i.e. Bacteroides), Proteobacteria (i.e. Escherichia), Fusobacteria (i.e. Helicobacter) [56] with Firmicutes and Bacteroidetes representing the 90% of the whole microbiota composition. The gut anatomical region, the human age and the state of health profoundly influence the biodiversity and the bacterial abundance. For examples, infants of the first year of life have significant biodiversity in Firmicutes and Proteobacteria respect to adulthood and even more to old age (> 70 years old) [7], while pathologies like the irritable inflammation syndrome are associated with a reduction in aerobic bacteria – such as Lactobacillus – respect to the healthy condition [57].
The intestinal flora contributes to the intestinal barrier homeostasis mainly by inhibiting pathogen infection and regulating the nutrients extraction, synthesis and absorption [56].
The gut microbiota can interfere with pathogens adhesion and infection by physically protecting the intestinal mucosa in a prey-predator mechanism or altering intestinal physicochemical features (e.g. pH) [58]. Furthermore, the microbiota can promote the maturation of the gut-associated lymphoid tissue (GALT) and stimulate immune cells to be responsive [59]. For instance, Bacteroides fragilis produces the polysaccharide A able to stimulate the maturation of regulatory T cells (Treg), thus promoting a state of immune tolerance and avoiding uncontrolled inflammation [60]. Simultaneously, bacteria can contribute to host defense from pathogen infection by stimulating specific intestinal epithelium cells to produce antimicrobial peptides (AMPs). Among the AMPs, defensins are molecules able to impair the bacterial cell wall integrity and prevent the pathogens colonization [61,62,63].
Gut microbiota is also a key regulator of nutrients metabolism as bacteria can process non-digestible fibers into carbon dioxide, hydrogen and short-chain fatty acids (SCFA)[64,65]. SCFAs – such as acetate, propionate, and butyrate– are a fundamental energy source for the host organism since they represent metabolites for lipogenesis and gluconeogenesis [66]. Each type of SCFA specifically contributes to the host tissue and organs. For instance, butyrate produced by Firmicutes is an important energy source for colonic cells, a protective element for intestinal barrier integrity and an inhibitor of inflammation [67]. Although the process of SCFAs fermentation gained the researchers attention in different microbiota-related studies, it has not yet been included in an overall view of the gut, especially comprehensive of mucus. Indeed, it is well defined either the impact of SCFAs on cells metabolism or their effect on the mucus viscoelastic properties, especially in viscosity, which is one of the leading parameters influencing the molecules diffusion in gut mucosa [68]. However, no studies investigated the possible synergic machinery of pathological microbiota composition (dysbiosis) and variation in SCFAs fermentation with the subsequent modification of the mucus properties, varied diffusion of molecules towards epithelium, and therefore altered cellular metabolism.
The intestinal microbiota composition is influenced by a multitude of factors rather than SCFAs – such as diet, antibiotic drugs and disease development – and its alteration is commonly defined as dysbiosis. Inflammatory bowel disease is one of the pathological conditions found to affect both the gut microbiota stability and the intestinal barrier homeostasis. In particular, IBD is related to a reduced immune tolerance to the microbiota-associated molecular patterns (MAMP) and an increase in inflammation of the intestinal barrier [62].
CD and UC have a different impact on gut systems (Table 1)[69]. Each inflammatory condition alters the bacterial composition reducing the biodiversity and thus, the abundance of key regulators of gut homeostasis [69]. For instance, Fecalibacterium Praustnizii is one of the main butyrate-producing bacteria and was found to be lower in both CD and UD patients respect to healthy patients[70]. A reduction in butyrate, as in other SCFAs, causes impairment in SCFA-associated mechanisms, which regulate the immune response by promoting AMP production and by modulating inflammation through the inhibition of NF-κB signaling pathway [67]. Changes in these SCFA-associated processes lead to a decreasing level of defensins production, a lower level of GALT stimulation and an increase in gut inflammation [62,67].
Although the cause-effect relationship between IBD and dysbiosis is still not well defined [71,72,73], the importance of microbiota in IBD progression is an undeniable phenomenon [74,75]. For this reason, the gut microbiota manipulation of CD and UC patients represent a promising therapeutic approach to reduce pathological symptoms, such as diarrhea and gut inflammation. In this regard, probiotics and fecal microbiota transplantation (FMT) are promising strategies[76,77,78]. While probiotics approach appears poorly standardized and confuses in its efficacy, FMT is a clinical reality with significant dysbiosis and symptoms remission data [79].
FMT requires the transplantation of fecal microbiota of a healthy donor into an intestinal tract of a recipient [80]. The transplantation success depends on the type of dysbiosis, which can be defined as primary or secondary. The primary condition is generally acute and transient and is triggered by antibiotics, malnutrition or pathogen infection. Instead, the secondary condition is commonly chronic and recurrent due to side effects of therapies (such as chemotherapy) or complex diseases, such as IBD, irritable bowel syndrome and autism [80]. Many aspects influence the success of FMT and thus of the clinical remission: (1) mucosal inflammation; (2) fecal sample manipulation; (3) donor and recipient compatibility; (4) administration method; (5) frequency of transplants; (6) pre-treatment with antibiotics [81,82].
The percentage of clinical remission is different for CD and UC: about 25% in UC and over 55% in CD. FMT success is featured by increasing biodiversity, rescue of Treg cells at the intestinal barrier, immune tolerance enhancement, SCFAs levels increase and reduction of barrier permeability to pathogens and bacterial molecules [79,83,84].
5. Intestinal mucus Barrier in IBD
The intestinal mucus is a biological hydrogel covering the whole intestinal tract [13]. It has a complex bi-layered architecture that allows for symbiotic microbiota growth, on the one hand, the preservation of the sterility of the gut epithelium, on the other, while modulating the passages of molecules from bacteria to cells and vice versa [14]. The polymeric network is made of trimers of the gel-forming mucins (MUC2 in the intestine and MUC5A in the stomach) [13,91] and is further stabilized by a set of specific structural protein, such as FCGBP and trefoil factor 3 (TFF3) proteins among others [92,93,94]. The gel-forming mucins are composed by many amino acids that is specific for the intestinal tract considered, but maintain a very conservative structure: a linear core with highly O-glycosylated branches, which increased the molecular weight of the mucin up to 50 kDa [13,95,96,97]. Moreover, the gel-forming mucins are supported by other mucins, named transcellular mucins (e.g. MUC1, MUC3 and MUC4) that do not form the polymeric network of mucus but are involved in different biological functions, such as lubrication, bacterial adhesion and inflammation signaling [98,99].
During disease, and in particular IBD, the homeostasis of the mucus ecosystem is impaired from a chemical and/or a physical point of view [100], altering the critical equilibrium between the microbiota (which resides in the mucus), mucin-secreting cells, and the biomolecules that diffuse in the network (at the base of microbiota-host communication) (Figure 3).
5.1. Compositional variation
During UC, the quantity and quality of the mucus is reduced considering the either whole mucus produced in bulk by the entire gut tissue either the mucus secreted at the single-cell level. Indeed, the number of mucin-secreting goblet cells is depleted[87,88,101,102]. Proteomic analysis revealed alteration in the mucus protein-core of UC patients. In particular, the decrease of structural protein, both gel-forming and transmembrane, (MUC2, MUC4, MUC3, FCGBP and CLCA1) was combined with an increase of inflammatory-related protein[87,89,90,103,104,105]. Interestingly, the reduction of [89,103,104,106], the increase of these proteins suggests an abnormal mucus production concomitant to an upregulated epithelial healing process.
IBDs were associated in changes of not only mucins production, but also composition. Physiologically, mucins can be classified as negative or neutral accordingly to the charge of glycosylated branches [107,108]. The intestinal MUC2 is a negative mucin composed by a high quantity of sulfonate (R-SO3-) group, which is involved in the MUC2 resistance to the bacteria enzymatic activity[109,110]. Histological results on IBD ex vivo models showed an outstanding reduction of the sulfation degree (more than 50%) [111,112], suggesting that the pathological mucus has only a limited defensive potential concerning physiology as also demonstrated by other studies (described below).
The chronic inflammation of IBD pathologies resides in a defective immune response triggered by unbalance between pro- and anti-inflammatory signals [113]. Phospholipids play a key role in this process, as they greatly contribute, together with bacteria, in modulating the mucus function of filtering [114,115,116,117]. In physiological condition, phosphatidylcholine (PC) and lysophosphatidylcholine (LPC) are the two main phospholipid classes that are present in mucus, accounting together for more than 80% of total mucus phospholipids [114]. However, in the case of UC, a strong depletion of PC and LPC was observed. Differently, CD patients only expressed a decrease of LPC, while maintaining PC concentration similar to control (Table 2). In healthy condition, the phospholipids grant hydrophobicity to mucus [118,119], which is one of the main physical properties contrasting bacteria adhesion to substrates [120]. For this reason, the phospholipids reduction occurring during IBD favoured the bacteria penetration in the gut tissue [121,122]. Interestingly, the LPC/PC ratio was comparable to control in case of CD but increased in case of UC [123]. As LPC is commonly associated to phospholipase activity rather than synthesis [124,125,126], these results suggest two different mechanisms involved in IBD: limited availability of PC but downregulation of phospholipase (i.e. an increase of LPC/PC ratio) in case of UC and only a slight promotion of the phospholipase activity in case of CD.
Compositional variations are not only important as a cause of the changes in the physiological feature of mucus per se, but also because they can directly modify the chemical affinity of mucus with bacterial, cellular or other molecules, facilitating and impairing their diffusion towards epithelium [14,127].
5.2. Structural weakening
Acting as a selective filter, the mucus behaves like a physical barrier between human cells and the environment, preserving the sterility of the epithelium while allowing the passage of nutrients and other molecules [13]. One of the main structural parameters of this barrier is its thickness since it is closely related to the function of the specific intestinal tract. For example, in the small intestine, where the nutraceutical molecules are absorbed, the mucus thickness is minimal thus facilitating the diffusion of the molecules from the lumen to the tissue. Conversely, in the colon, where the majority of the intestinal microbiota is hosted, the thick mucus layer acts as a protection against infections (Table 3). For this reason, the colonic thickness (i.e. ~150 vs ~50 µm) is significantly higher than small intestine [68,128,129].
The IBD mucus is usually investigated after identification of the inflammation degree by standard clinical criteria, expressed by numbers from 0 to 4 in proportion to the severity. In UC patients, the mucus layer was thinner than controls (especially in the descending colon and rectum) [130]. Moreover, the thickness reduction was exacerbated with proportionality to the severity of inflammation, showing a complete absence of mucus with a severe state of inflammation [131,132].
The measurements of the CD mucus thickness are not homogeneous in the literature. Indeed, this structural parameter was found to be either higher [132], or lower [133], or comparable [134] to the healthy controls. This variability can be linked to the intrinsic heterogeneity in the distribution of the lesions in the case of CD compared to UC [20,22], but also in variations of the mucus staining protocols and preservation as well as the not-standard definition of “controls”, which often suffer from other intestinal pathologies. Interestingly, independently from the data obtained in the inflamed areas, the mucus thickness in undamaged colon portions of both UC and CD patients is not significantly different from healthy controls [135], suggesting the localized nature and impact of these pathologies.
Moreover, the mucus discontinuity, measured as the length of mucus-free mucosa normalized to the total specimen length, was significantly higher only in groups with severe active UC (25.7 %) than controls (1.0 %) [136]. No significant difference was found between controls and either quiescent UC or CD samples, as the adherent mucus layer was essentially continuous [136] similarly to the physiological condition [137].
In physiology, the mucus thickness is not the only structural factor affecting the mucus properties. Indeed, this parameter must be always coupled with the peculiar double-layered architecture, where the adherent mucus layer acts as the real barrier for the epithelium [138]. In vivo animal models with induced moderate UC showed a limited variation of the mucus thickness but a substantial modification of the mucus architecture. In particular, the adherent layer was much thicker than the controls [139,140]. These results suggest that not only the amount of mucus is a pivotal element in IBDs, but also its quality. This consideration becomes more robust if the permeability of the mucus is considered, as an index of the efficacy in maintenance of the barrier properties. Microbeads with dimension comparable to bacteria (0.5-2 µm) were able to penetrate the mucus and reach the epithelium in both ex vivo human biopsies [103] and animal models [140]. Similarly to the thickness, the mucus weakening increased with the disease severity. Indeed, the percentage of beads reaching the epithelium was 36% and 20% in the case of acute disease and remission state, but only 10% in controls [103]. As structural properties correspond to chemical composition, the impaired barrier properties were as related not only to the decrease of the gel-forming mucin MUC2 but also of the support protein FCGBP [103]. This suggests the presence of two combined effects in UC: a reduced mucus production with a structural instability of the network.
6. Barrier integrity and permeability assays
The intestinal barrier integrity and permeability are standard parameters used in clinics to identify and define the severity of IBD. The measurement of these functional aspects is currently possible using different well-established methods [144,145]:
- Active and passive permeability assays: Active permeability assays require the oral administration of sugars or polymers and the measurement of urinary concentrations at different time points from 30 min to 24 hours [146]. The higher is the molecule concentration, the leakier the epithelial barrier. A common method is based on the administration of different molecules (e.g. lactulose, mannitol, PEG (polyethylene glycol) molecules and sucrose among others) and the measurement of their concentration in urine, thus obtaining information about the permeability of the mucosa in different region of the gut, which is in dependence of the size and the nature of the administered substance [146,147,148,149,150]. The use of the radioactive 51Cr-EDTA were not only able to describe an increased permeability in CD patients (20% higher than healthy controls) but also to correlate this permeability variation with a reduction in specific bacteria abundance (i.e. Faecalibacterium prausnitzii) [151]. No consideration was proposed in terms of mucus properties, whose contribution remained clouded under the permeability characterization. The principle of passive permeability assays is similar to the active methods described above. The main difference is the nature of the detected molecules, which are usually products of the individual metabolism (e.g. bacteria-derived molecules, intestinal integrity-related or immunological biomarkers) measured in plasma, serum and biopsies [149]. Among the bacterial-derived molecules, lipopolysaccharides (LPS) and butyrate are of primary importance, as they are the major components of the outer membrane of Gram-negative bacteria and biomarker of intestinal barrier integrity, respectively [140,152,153,154,155]. Epithelial and immune biomarkers detection in plasma and urine is another effective method to define the intestinal barrier integrity (i.e. citrullin and claudin-3) [53,156,157]. The inflammatory marker mainly investigated in UC and CD is calprotectin, a product of active and infiltrated neutrophils in the intestinal mucosa. High levels of fecal calprotectin (>100 µg/g) were correlated with higher levels of intestinal permeability indicators and give an indirect measure of IBD severity [151]. Similarly to active permeability assay, the passive permeability assays have the possibility to correlate bacterial products to dysbiosis and intestinal barrier impairment, but do not provide information about the relative contribution of mucus within the epithelial barrier. The inclusion of a more detailed characterisation of mucus could provide a deeper understanding of these experimental evidence. For instance, the study of the variation of the mucus network microstructure can elucidate if the diffusion of molecules from lumen to epithelium is facilitate or impaired. Similarly, compositional changes can provide information of the dye/molecules affinity with the mucus matrix.
- Confocal laser endomicroscopy. Confocal laser endomicroscopy allows the acquisition of confocal images during endoscopic procedures [158]. It provides information about the epithelial barrier morphology. This method requires the intravenous administration of a fluorescent dye – such as fluorescein – that could be easily excited by the laser of the endomicroscope. A detector then transforms the emitted light signal into an electrical input for computational recording. This method generates accurate images giving evident information on the barrier morphology and the epithelial crypts architecture [159]. These analyses highlighted frequent morphological features of IBD epithelia, such as intra- and inter-crypt distance increase, irregular crypt organization, micro- and macro-lesions leading to cell and molecule infiltrates increase [149]. Overall, endomicroscopy not only provides critical data for the determination of the barrier impairment but also allows to limit the collection of biopsies to strictly necessary cases. Despite the epithelial barrier is well examined, the mucus barrier contribution is still poorly considered while information about gut dysbiosis is not provided at all.
- Ussing chamber. The Ussing chamber-based method is an invasive assay used to determine intestinal barrier integrity and permeability. It is performed on intestinal tissue biopsies and requires invasive procedures to collect ex vivo specimens. This technique is based on the measurement of a voltage difference (ΔVEP) and a trans-epithelial (o trans-mucosal) electric resistance (TEER o TER, respectively) between the apical and the basolateral side of the barrier. It requires the application of active ion transport through the epithelium and the measurement of the ΔVEP and TER. The ion transport is correlated to the barrier integrity. The weaker the epithelium, the higher will be the transport and the lower the TER [149,160] Like previous well-established clinical assays, the Ussing chamber-based method is a valuable approach to study the intestinal barrier function in IBD, as it was demonstrated for instance that IBD patients showed a decrease of TER near the 39% [161]. However, it cannot provide information about the relative contribution of the elements modulating barrier disruption.
Even though these methods allowed clinicians to evaluate functional aspects – such as intestinal barrier permeability, integrity and morphology – they do not discriminate the relative contribution of all the modulating elements (i.e. gut dysbiosis, epithelial barrier and mucus barrier). In particular, the assays that are commonly used for studying the IBD features rarely focused on the impact of mucus physicochemical and mechanical properties, which is usually simply considered as the stage, without active and/or substantial role, in the scene where the biological actors (i.e. epithelium and microbiota) perform their playscript. The inclusion of a more refined mechanical and compositional characterization of the mucus with the well-defined permeability assays would lead towards a synergistic comprehension of the IBD pathology from a biological and materialistic point of view, opening new horizons towards unexplored therapeutic strategies.
7. Conclusion
Autoimmune diseases of the intestinal tract are spreading worldwide. Researchers have proposed different hypotheses in their challenging task of identifying the origin of these diseases, but the pathophysiology of the inflammatory bowel disease (IBD) remains a not fully described process at the date. In this frame, the leaky intestinal barrier is unquestionable evidence, wheatear it is interpreted in the sense of trigger or symptom of the pathological states. Even if the leaky intestinal barrier in IBD is historically associated to the impaired intestinal cellular barrier and dysbiosis, it is a phenomenon derived by a complex play-script directed by another, not cellular, actor: the intestinal mucus. In this complex performance, the chemical and physical barrier of mucus properties cooperates in synergy with the bacterial and cellular metabolism, effecting the diffusion of bacterial- and cellular-derived molecules and modulating the inflammatory response. Although numerous advancements were made in the study of IBD thanks to the classic vision of these pathologies (such as the definition of standard analysis methods), the inclusion of a refined mucus characterization can open to new paths in the direction of the complete comprehension of the IBD condition. Indeed, the synergic approach between the biological- and mucus-oriented vision may boost towards new etiological and pathogenic insights, as well as alternative therapeutic strategies against IBD pathologies.
Author Contributions
Conceptualization, L.S., L.B. and P.P.; formal analysis, L.S., L.B., M.M, G.G., F.B.V. and P.P.; writing—original draft preparation, L.S. and L.B.; writing—review and editing, F.B.V. and P.P.; visualization, G.G. and M.M.; supervision, F.B.V. and P.P.;. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The data presented in this study are openly available.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lerner, P. Jeremias, and T. Matthias, “The world incidence and prevalence of autoimmune diseases is increasing,” International Journal of Celiac Disease, vol. 3, no. 4, pp. 151–155, 2015. [CrossRef]
- M. E. Sherlock and E. I. Benchimol, “Classification of inflammatory bowel disease in children,” Pediatric Inflammatory Bowel Disease: Third Edition, pp. 181–191, 2017. [CrossRef]
- F. W. Miller, “The increasing prevalence of autoimmunity and autoimmune diseases: an urgent call to action for improved understanding, diagnosis, treatment, and prevention,” Curr Opin Immunol, vol. 80, p. 102266, Feb. 2023. [CrossRef]
- R. S. Chauhan et al., “WHO task group on environmental health criteria on principles and methods for assessing autoimmunity associated with exposure to chemicals,” Environmental Health Criteria, no. 236, 2006.
- D. S. Pisetsky, “Pathogenesis of autoimmune disease,” Nature Reviews Nephrology 2023, pp. 1–16, May 2023. [CrossRef]
- M. Barreiro-de Acosta et al., “Epidemiological, Clinical, Patient-Reported and Economic Burden of Inflammatory Bowel Disease (Ulcerative colitis and Crohn’s disease) in Spain: A Systematic Review,” Adv Ther, vol. 40, no. 5, pp. 1975–2014, May 2023. [CrossRef]
- E. Thursby and N. Juge, “Introduction to the human gut microbiota,” Biochemical Journal, vol. 474, no. 11. Portland Press Ltd, pp. 1823–1836, Jun. 01, 2017. doi: 10.1042/BCJ20160510.
- R. M. Patel and P. W. Denning, “Therapeutic Use of Prebiotics, Probiotics, and Postbiotics to Prevent Necrotizing Enterocolitis. What is the Current Evidence?,” Clin Perinatol, vol. 40, no. 1, pp. 11–25, 2013. [CrossRef]
- C. Wong and M. Levy, “New Approaches to Microbiome-Based Therapies,” mSystems, vol. 4, no. 3, pp. 1–5, Jun. 2019. [CrossRef]
- S. Sekhon and S. Jairath, “Prebiotics, probiotics and synbiotics: an overview,” Journal of pharmaceutical education and research, vol. 1, no. 2, p. 13, 2010.
- S. Ahlawat, P. Kumar, H. Mohan, S. Goyal, and K. K. Sharma, “Inflammatory bowel disease: tri-directional relationship between microbiota, immune system and intestinal epithelium,” https://doi.org/10.1080/1040841X.2021.1876631, vol. 47, no. 2, pp. 254–273, 2021. [CrossRef]
- Hajj Hussein et al., “Highlights on two decades with microbiota and inflammatory bowel disease from etiology to therapy,” Transpl Immunol, vol. 78, p. 101835, Jun. 2023. [CrossRef]
- M. E. V. Johansson, H. Sjövall, and G. C. Hansson, “The gastrointestinal mucus system in health and disease,” Nat Rev Gastroenterol Hepatol, vol. 10, no. 6, pp. 352–361, 2013. [CrossRef]
- L. Sardelli et al., “ Towards bioinspired in vitro models of intestinal mucus ,” RSC Adv, vol. 9, no. 28, pp. 15887–15899, 2019. [CrossRef]
- S. K. Lai, Y.-Y. Wang, D. Wirtz, and J. Hanes, “Micro- and macrorheology of mucus,” Adv Drug Deliv Rev, vol. 61, no. 2, pp. 86–100, 2009. [CrossRef]
- M. Boegh, S. G. Baldursdóttir, A. Müllertz, and H. M. Nielsen, “Property profiling of biosimilar mucus in a novel mucus-containing in vitro model for assessment of intestinal drug absorption,” European Journal of Pharmaceutics and Biopharmaceutics, vol. 87, no. 2, pp. 227–235, 2014. [CrossRef]
- König et al., “Human Intestinal Barrier Function in Health and Disease,” Clin Transl Gastroenterol, vol. 7, no. 10, p. e196, 2016. [CrossRef]
- S. Alatab et al., “The global, regional, and national burden of inflammatory bowel disease in 195 countries and territories, 1990–2017: a systematic analysis for the Global Burden of Disease Study 2017,” Lancet Gastroenterol Hepatol, vol. 5, no. 1, pp. 17–30, Jan. 2020. [CrossRef]
- Kornbluth and D. B. Sachar, “Ulcerative colitis practice guidelines in adults: Practice Parameters Committee,” American Journal of Gastroenterology, vol. 99, no. 7, pp. 1371–1385, 2004. [CrossRef]
- Torres, S. Mehandru, J. Colombel, and L. Peyrin-Biroulet, “Crohn’s disease,” The Lancet, vol. 389, no. 10080, pp. 1741–1755, Apr. 2017. [CrossRef]
- E. Louis, A. Collard, A. F. Oger, E. Degroote, F. Aboul Nasr El Yafi, and J. Belaiche, “Behaviour of Crohn’s disease according to the Vienna classification: Changing pattern over the course of the disease,” Gut, vol. 49, no. 6, pp. 777–782, 2001. [CrossRef]
- H. Wanderås, B. A. Moum, M. L. Høivik, and Ø. Hovde, “Predictive factors for a severe clinical course in ulcerative colitis: Results from population-based studies,” World J Gastrointest Pharmacol Ther, vol. 7, no. 2, p. 235, 2016. [CrossRef]
- T. Abegunde, B. H. Muhammad, O. Bhatti, and T. Ali, “Environmental risk factors for inflammatory bowel diseases: Evidence based literature review,” World J Gastroenterol, vol. 22, no. 27, pp. 6296–6317, 2016. [CrossRef]
- S. S. Mahid, K. S. Minor, R. E. Soto, C. A. Hornung, and S. Galandiuk, “Smoking and inflammatory bowel disease: A meta-analysis,” Mayo Clin Proc, vol. 81, no. 11, pp. 1462–1471, 2006. [CrossRef]
- M. Higuchi, H. Khalili, A. T. Chan, J. M. Richter, A. Bousvaros, and C. S. Fuchs, “A prospective study of cigarette smoking and the risk of inflammatory bowel disease in women,” American Journal of Gastroenterology, vol. 107, no. 9, pp. 1399–1406, 2012. [CrossRef]
- J. Torres, S. Mehandru, J. Colombel, and L. Peyrin-Biroulet, “Crohn’s disease,” The Lancet, vol. 389, no. 10080, pp. 1741–1755, Apr. 2017. [CrossRef]
- Le Berre, A. N. Ananthakrishnan, S. Danese, S. Singh, and L. Peyrin-Biroulet, “Ulcerative Colitis and Crohn’s Disease Have Similar Burden and Goals for Treatment,” Clinical Gastroenterology and Hepatology, vol. 18, no. 1, pp. 14–23, 2020. [CrossRef]
- R. J. Kennedy, M. Hoper, K. Deodhar, P. J. Erwin, S. J. Kirk, and K. R. Gardiner, “Interleukin 10-deficient colitis: New similarities to human inflammatory bowel disease,” British Journal of Surgery, vol. 87, no. 10, pp. 1346–1351, 2000. [CrossRef]
- S. Bridger, J. C. W. Lee, I. Biarnason, J. E. Lennard Jones, and A. J. Macpherson, “In siblings with similar genetic susceptibility for inflammatory bowel disease, smokers tend to develop Crohn’s disease and non-smokers develop ulcerative colitis,” Gut, vol. 51, no. 1, pp. 21–25, 2002. [CrossRef]
- Khor, A. Gardet, and R. J. Xavier, “Genetics and pathogenesis of inflammatory bowel disease,” Nature, vol. 474, no. 7351, pp. 307–317, 2011. [CrossRef]
- G. Meng, T. M. Monaghan, N. A. Duggal, P. Tighe, and F. Peerani, “Microbial–Immune Crosstalk in Elderly-Onset Inflammatory Bowel Disease: Unchartered Territory,” J Crohns Colitis, Feb. 2023. [CrossRef]
- Oriano et al., “The Open Challenge of in vitro Modeling Complex and Multi-Microbial Communities in Three-Dimensional Niches,” Front Bioeng Biotechnol, vol. 8, no. October, pp. 1–17, 2020. [CrossRef]
- Peneda Pacheco, N. Suárez Vargas, S. Visentin, and P. Petrini, “From tissue engineering to engineering tissues: the role and application of in vitro models,” Biomater Sci, 2020. [CrossRef]
- Pacheco et al., “Heterogeneity governs 3D-cultures of clinically relevant microbial communities,” Mar. 2023. [CrossRef]
- L. Sardelli et al., “Bioinspired in vitro intestinal mucus model for 3D-dynamic culture of bacteria,” Biomaterials Advances, vol. 139, p. 213022, Aug. 2022. [CrossRef]
- F. Shroyer and S. A. Kocoshis, “Anatomy and Physiology of the Small and Large Intestines,” in Pediatric Gastrointestinal and Liver Disease, Elsevier Inc., 2011, pp. 324–336. doi: 10.1016/B978-1-4377-0774-8.10031-4.
- J. M. Allaire, S. M. Crowley, H. T. Law, S. Y. Chang, H. J. Ko, and B. A. Vallance, “The Intestinal Epithelium: Central Coordinator of Mucosal Immunity,” Trends in Immunology, vol. 39, no. 9. Elsevier Ltd, pp. 677–696, Sep. 2018. doi: 10.1016/j.it.2018.04.002.
- Y. Furusawa et al., “Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells,” Nature, vol. 504, no. 7480, pp. 446–450, Nov. 2013. [CrossRef]
- Plitas and A. Y. Rudensky, “Regulatory T cells: Differentiation and function,” Cancer Immunol Res, vol. 4, no. 9, pp. 721–725, Sep. 2016. [CrossRef]
- Rivera-Chávez et al., “Depletion of Butyrate-Producing Clostridia from the Gut Microbiota Drives an Aerobic Luminal Expansion of Salmonella,” Cell Host Microbe, vol. 19, no. 4, pp. 443–454, Apr. 2016. [CrossRef]
- M. C. Rea et al., “Effect of broad- and narrow-spectrum antimicrobials on Clostridium difficile and microbial diversity in a model of the distal colon,” Proc Natl Acad Sci U S A, vol. 108, no. SUPPL. 1, pp. 4639–4644, Mar. 2011. [CrossRef]
- J. H. Kang, “Salsolinol, a catechol neurotoxin, induces oxidative modification of cytochrome c,” BMB Rep, 2013. [CrossRef]
- C. Chelakkot, J. Ghim, and S. H. Ryu, “Mechanisms regulating intestinal barrier integrity and its pathological implications,” Experimental and Molecular Medicine, vol. 50, no. 8. Nature Publishing Group, Aug. 01, 2018. doi: 10.1038/s12276-018-0126-x.
- Zahraoui et al., “A small rab GTPase is distributed in cytoplasmic vesicles in non polarized cells but colocalizes with the tight junction marker ZO-1 in polarized epithelial cells,” Journal of Cell Biology, vol. 124, no. 1–2, pp. 101–115, Jan. 1994. [CrossRef]
- S. Tsukita, K. Oishi, T. Akiyama, Y. Yamanashi, T. Yamamoto, and S. Tsukita, “Specific proto-oncogenic tyrosine kinases of src family are enriched in cell-to-cell adherens junctions where the level of tyrosine phosphorylation is elevated,” Journal of Cell Biology, vol. 113, no. 4, pp. 867–879, May 1991. [CrossRef]
- L. L. Wu et al., “Epithelial inducible nitric oxide synthase causes bacterial translocation by impairment of enterocytic tight junctions via intracellular signals of Rho-associated kinase and protein kinase C zeta,” Crit Care Med, vol. 39, no. 9, pp. 2087–2098, Sep. 2011. [CrossRef]
- J. C. Zech, I. Pouvreau, A. Cotinet, O. Goureau, B. Le Varlet, and Y. De Kozak, “Effect of cytokines and nitric oxide on tight junctions in cultured rat retinal pigment epithelium,” Invest Ophthalmol Vis Sci, vol. 39, no. 9, pp. 1600–1608, 1998.
- C. T. Capaldo et al., “Proinflammatory cytokine-induced tight junction remodeling through dynamic self-assembly of claudins,” Mol Biol Cell, vol. 25, no. 18, pp. 2710–2719, Sep. 2014. [CrossRef]
- W. Fries, C. Muja, C. Crisafulli, S. Cuzzocrea, and E. Mazzon, “Dynamics of enterocyte tight junctions: Effect of experimental colitis and two different anti-TNF strategies,” Am J Physiol Gastrointest Liver Physiol, vol. 294, no. 4, pp. 938–947, 2008. [CrossRef]
- J. M. Reimund et al., “Increased production of tumour necrosis factor-α, interleukin-1β, and interleukin-6 by morphologically normal intestinal biopsies from patients with Crohn’s disease,” Gut, vol. 39, no. 5, pp. 684–689, 1996. [CrossRef]
- M. Marchiando et al., “Caveolin-1-dependent occludin endocytosis is required for TNF-induced tight junction regulation in vivo,” Journal of Cell Biology, vol. 189, no. 1, pp. 111–126, 2010. [CrossRef]
- Zhang, L. Cheng, J. Wang, M. Hao, and H. Che, “Antibiotic-Induced Gut Microbiota Dysbiosis Damages the Intestinal Barrier, Increasing Food Allergy in Adult Mice,” Nutrients 2021, Vol. 13, Page 3315, vol. 13, no. 10, p. 3315, Sep. 2021. [CrossRef]
- Kolios, V. Valatas, and S. G. Ward, “Nitric oxide in inflammatory bowel disease: A universal messenger in an unsolved puzzle,” Immunology, vol. 113, no. 4. Wiley-Blackwell, pp. 427–437, Dec. 2004. doi: 10.1111/j.1365-2567.2004.01984.x.
- T. Katsube, H. Tsuji, and M. Onoda, “Nitric oxide attenuates hydrogen peroxide-induced barrier disruption and protein tyrosine phosphorylation in monolayers of intestinal epithelial cell,” Biochim Biophys Acta Mol Cell Res, vol. 1773, no. 6, pp. 794–803, Jun. 2007. [CrossRef]
- G. Kolios, N. Rooney, C. T. Murphy, D. A. F. Robertson, and J. Westwick, “Expression of inducible nitric oxide synthase activity in human colon epithelial cells: Modulation by T lymphocyte derived cytokines,” Gut, vol. 43, no. 1, pp. 56–63, 1998. [CrossRef]
- E. Rinninella et al., “What is the healthy gut microbiota composition? A changing ecosystem across age, environment, diet, and diseases,” Microorganisms, vol. 7, no. 1, Jan. 2019. [CrossRef]
- M. Carroll, Y. H. Chang, J. Park, R. B. Sartor, and Y. Ringel, “Luminal and mucosal-associated intestinal microbiota in patients with diarrhea-predominant irritable bowel syndrome,” Gut Pathog, vol. 2, no. 1, 2010. [CrossRef]
- M. Pickard, M. Y. Zeng, R. Caruso, and G. Núñez, “Gut Microbiota: Role in Pathogen Colonization, Immune Responses and Inflammatory Disease”. [CrossRef]
- Spencer and M. Bemark, “Human intestinal B cells in inflammatory diseases,” Nature Reviews Gastroenterology & Hepatology 2023 20:4, vol. 20, no. 4, pp. 254–265, Feb. 2023. [CrossRef]
- S. Dasgupta, D. Erturk-Hasdemir, J. Ochoa-Reparaz, H.-C. Reinecker, and D. L. Kasper, “Plasmacytoid Dendritic Cells Mediate Anti-inflammatory Responses to a Gut Commensal Molecule via Both Innate and Adaptive Mechanisms,” Cell Host Microbe, vol. 15, pp. 413–423, 2014. [CrossRef]
- H.-Y. Cheng, M.-X. Ning, D.-K. Chen, and W.-T. Ma, “Interactions Between the Gut Microbiota and the Host Innate Immune Response Against Pathogens.,” Front Immunol, vol. 10, p. 607, 2019. [CrossRef]
- Coretti et al., “The Interplay between Defensins and Microbiota in Crohn’s Disease,” Mediators Inflamm, vol. 2017, 2017. [CrossRef]
- T. Ayabe, D. P. Satchell, C. L. Wilson, W. C. Parks, M. E. Selsted, and A. J. Ouellette, “Secretion of microbicidal α-defensins by intestinal Paneth cells in response to bacteria,” Nat Immunol, vol. 1, no. 2, pp. 113–118, Aug. 2000. [CrossRef]
- J. Fernandes, W. Su, S. Rahat-Rozenbloom, T. M. S. Wolever, and E. M. Comelli, “Adiposity, gut microbiota and faecal short chain fatty acids are linked in adult humans,” Nutr Diabetes, vol. 4, no. JUNE, 2014. [CrossRef]
- T. Wongsurawat et al., “Microbiome analysis of thai traditional fermented soybeans reveals short-chain fatty acid-associated bacterial taxa,” Scientific Reports 2023 13:1, vol. 13, no. 1, pp. 1–10, May 2023. [CrossRef]
- Y. Sun et al., “Effect of short-chain fatty acids on triacylglycerol accumulation, lipid droplet formation and lipogenic gene expression in goat mammary epithelial cells,” Animal Science Journal, vol. 87, no. 2, pp. 242–249, 2016. [CrossRef]
- W. Ratajczak, A. Rył, A. Mizerski, K. Walczakiewicz, O. Sipak, and M. Laszczyńska, “Immunomodulatory potential of gut microbiome-derived shortchain fatty acids (SCFAs),” Acta Biochimica Polonica, vol. 66, no. 1. Acta Biochimica Polonica, pp. 1–12, Mar. 04, 2019. doi: 10.18388/abp.2018_2648.
- Sardelli et al., “ Towards bioinspired in vitro models of intestinal mucus ,” RSC Adv, vol. 9, no. 28, pp. 15887–15899, 2019. [CrossRef]
- D. N. Frank, A. L. St. Amand, R. A. Feldman, E. C. Boedeker, N. Harpaz, and N. R. Pace, “Molecular-phylogenetic characterization of microbial community imbalances in human inflammatory bowel diseases,” Proc Natl Acad Sci U S A, vol. 104, no. 34, pp. 13780–13785, Aug. 2007. [CrossRef]
- J. A. Gilbert et al., “Microbiome-wide association studies link dynamic microbial consortia to disease,” Nature, vol. 535, no. 7610. Nature Publishing Group, pp. 94–103, Jul. 06, 2016. doi: 10.1038/nature18850.
- F. Fava and S. Danese, “Intestinal microbiota in inflammatory bowel disease: Friend of foe?,” World J Gastroenterol, vol. 17, no. 5, pp. 557–566, 2011. [CrossRef]
- J. Bien, V. Palagani, and P. Bozko, “The intestinal microbiota dysbiosis and Clostridium difficile infection: Is there a relationship with inflammatory bowel disease?,” Therap Adv Gastroenterol, vol. 6, no. 1, pp. 53–68, 2013. [CrossRef]
- L. Rigottier-Gois, “Dysbiosis in inflammatory bowel diseases: The oxygen hypothesis,” ISME Journal, vol. 7, no. 7, pp. 1256–1261, 2013. [CrossRef]
- L. Cook et al., “Analysis of Flagellin-Specific Adaptive Immunity Reveals Links to Dysbiosis in Patients With Inflammatory Bowel Disease,” Cmgh, vol. 9, no. 3, pp. 485–506, 2020. [CrossRef]
- C. P. Tamboli, “Dysbiosis in inflammatory bowel disease,” Gut, vol. 53, no. 1, pp. 1–4, Jan. 2004. [CrossRef]
- Hajj Hussein et al., “Highlights on two decades with microbiota and inflammatory bowel disease from etiology to therapy,” Transpl Immunol, vol. 78, p. 101835, Jun. 2023. [CrossRef]
- Zhou et al., “Programmable probiotics modulate inflammation and gut microbiota for inflammatory bowel disease treatment after effective oral delivery,” Nature Communications 2022 13:1, vol. 13, no. 1, pp. 1–14, Jun. 2022. [CrossRef]
- S. Porcari et al., “Fecal microbiota transplantation for recurrent C. difficile infection in patients with inflammatory bowel disease: A systematic review and meta-analysis,” J Autoimmun, p. 103036, Apr. 2023. [CrossRef]
- J. Basso, N. O. Saraiva Câmara, and H. Sales-Campos, “Microbial-based therapies in the treatment of inflammatory bowel disease – An overview of human studies,” Front Pharmacol, vol. 9, no. JAN, 2019. [CrossRef]
- Kellermayer, “Fecal microbiota transplantation: Great potential with many challenges,” Translational Gastroenterology and Hepatology, vol. 4, no. May. AME Publishing Company, 2019. doi: 10.21037/tgh.2019.05.10.
- X. Guo, X. Liu, and J. Hao, Gut Microbiota in Ulcerative Colitis: Insights on Pathogenesis and Treatment. 2020. doi: 10.1111/1751-2980.12849.
- Z. Shen et al., “Update on intestinal microbiota in Crohn’s disease 2017: Mechanisms, clinical application, adverse reactions, and outlook.,” J Gastroenterol Hepatol, vol. 32, no. 11, pp. 1804–1812, Nov. 2017. [CrossRef]
- Paramsothy et al., “Faecal microbiota transplantation for inflammatory bowel disease: A systematic review and meta-analysis,” J Crohns Colitis, vol. 11, no. 10, pp. 1180–1199, Oct. 2017. [CrossRef]
- B. P. Vaughn et al., “Increased Intestinal Microbial Diversity Following Fecal Microbiota Transplant for Active Crohn’s Disease,” Inflamm Bowel Dis, vol. 22, no. 9, pp. 2182–2190, Aug. 2016. [CrossRef]
- Joossens et al., “Dysbiosis of the faecal microbiota in patients with Crohn’s disease and their unaffected relatives,” Gut, vol. 60, no. 5, pp. 631–637, May 2011. [CrossRef]
- Machiels et al., “A decrease of the butyrate-producing species Roseburia hominis and Faecalibacterium prausnitzii defines dysbiosis in patients with ulcerative colitis.,” Gut, vol. 63, no. 8, pp. 1275–83, Aug. 2014. [CrossRef]
- Björkqvist et al., “Alterations in the relative abundance of Faecalibacterium prausnitzii correlate with changes in fecal calprotectin in patients with ileal Crohn’s disease: a longitudinal study,” Scand J Gastroenterol, vol. 54, no. 5, pp. 577–585, May 2019. [CrossRef]
- R. Ricciardi, J. W. Ogilvie, P. L. Roberts, P. W. Marcello, T. W. Concannon, and N. N. Baxter, “Epidemiology of Clostridium difficile Colitis in Hospitalized Patients with Inflammatory Bowel Diseases,” Dis Colon Rectum, vol. 52, no. 1, pp. 40–45, Jan. 2009. [CrossRef]
- J. WEHKAMP and E. F. STANGE, “A New Look at Crohn’s Disease: Breakdown of the Mucosal Antibacterial Defense,” Ann N Y Acad Sci, vol. 1072, no. 1, pp. 321–331, Aug. 2006. [CrossRef]
- Shoaei et al., “Clostridium difficile isolated from faecal samples in patients with ulcerative colitis,” BMC Infect Dis, vol. 19, no. 1, p. 361, Apr. 2019. [CrossRef]
- R. Bansil and B. S. Turner, “Mucin structure, aggregation, physiological functions and biomedical applications,” Curr Opin Colloid Interface Sci, vol. 11, no. 2–3, pp. 164–170, 2006. [CrossRef]
- E. V Johansson, K. A. Thomsson, and G. C. Hansson, “Proteomic analyses of the two mucus layers of the colon barrier reveal that their main component, the Muc2 mucin, is strongly bound to the fcgbp protein,” J Proteome Res, vol. 8, no. 7, pp. 3549–3557, 2009. [CrossRef]
- K. Godl et al., “The N Terminus of the MUC2 Mucin Forms Trimers That Are Held Together within a Trypsin-resistant Core Fragment,” Journal of Biological Chemistry, vol. 277, no. 49, pp. 47248–47256, Dec. 2002. [CrossRef]
- C. Butnarasu et al., “Mucosomes: Intrinsically Mucoadhesive Glycosylated Mucin Nanoparticles as Multi-Drug Delivery Platform,” Adv Healthc Mater, vol. 11, no. 15, p. 2200340, Aug. 2022. [CrossRef]
- E. V Johansson, J. M. H. Larsson, and G. C. Hansson, “The two mucus layers of colon are organized by the MUC2 mucin, whereas the outer layer is a legislator of host-microbial interactions,” Proceedings of the National Academy of Sciences, vol. 108, no. Supplement_1, pp. 4659–4665, 2011. [CrossRef]
- J. P. Pearson, P. I. Chater, and M. D. Wilcox, “The properties of the mucus barrier, a unique gel – how can nanoparticles cross it?,” Ther Deliv, vol. 7, no. 4, pp. 229–244, 2016. [CrossRef]
- C. ROBBE, C. CAPON, B. CODDEVILLE, and J.-C. MICHALSKI, “Structural diversity and specific distribution of O-glycans in normal human mucins along the intestinal tract,” Biochemical Journal, vol. 384, no. 2, pp. 307–316, 2004. [CrossRef]
- C. L. Hattrup and S. J. Gendler, “Structure and Function of the Cell Surface (Tethered) Mucins,” Annu Rev Physiol, vol. 70, no. 1, pp. 431–457, Mar. 2008. [CrossRef]
- D. Ambort, M. E. V. Johansson, J. K. Gustafsson, A. Ermund, and G. C. Hansson, “Perspectives on Mucus Properties and Formation--Lessons from the Biochemical World,” Cold Spring Harb Perspect Med, vol. 2, no. 11, pp. a014159–a014159, 2012. [CrossRef]
- J. Leal, H. D. C. Smyth, and D. Ghosh, “Physicochemical properties of mucus and their impact on transmucosal drug delivery,” Int J Pharm, vol. 532, no. 1, pp. 555–572, 2017. [CrossRef]
- E. Dorofeyev, I. V. Vasilenko, O. A. Rassokhina, and R. B. Kondratiuk, “Mucosal barrier in ulcerative colitis and crohn’s disease,” Gastroenterol Res Pract, vol. 2013, no. Cd, 2013. [CrossRef]
- E. V. Johansson et al., “Bacteria penetrate the normally impenetrable inner colon mucus layer in both murine colitis models and patients with ulcerative colitis,” Gut, vol. 63, no. 2, pp. 281–291, 2014. [CrossRef]
- Van Der Post et al., “Structural weakening of the colonic mucus barrier is an early event in ulcerative colitis pathogenesis,” pp. 1–10, 2019. [CrossRef]
- E. Dorofeyev, I. V. Vasilenko, O. A. Rassokhina, and R. B. Kondratiuk, “Mucosal barrier in ulcerative colitis and crohn’s disease,” Gastroenterol Res Pract, vol. 2013, no. Cd, 2013. [CrossRef]
- D. K. Podolsky and K. J. Isselbacher, “Composition of human colonic mucin. Selective alteration in inflammatory bowel disease,” Journal of Clinical Investigation, vol. 72, no. 1, pp. 142–153, 1983. [CrossRef]
- Xia et al., “Cell membrane-anchored MUC4 promotes tumorigenicity in epithelial carcinomas,” Oncotarget, vol. 8, no. 8, pp. 14147–14157, 2017. [CrossRef]
- R. I. Nonose, A. I. Paula Pimentel Spadari, D. Gonçalves Priolli III, F. I. Rodrigues Máximo, J. I. Aires Pereira, and C. V Augusto Real Martinez, “Tissue quantification of neutral and acid mucins in the mucosa of the colon with and without fecal stream in rats Quantificação tecidual de mucinas neutras e ácidas na mucosa do cólon com e sem trânsito intestinal em ratos,” Acta Cir Bras, vol. 24, no. 244, pp. 2009–268, 2009.
- S. Agawa, T. Muto, and Y. Morioka, “Mucin abnormality of colonic mucosa in ulcerative colitis associated with carcinoma and/or dysplasia,” Dis Colon Rectum, vol. 31, no. 5, pp. 387–389, 1988. [CrossRef]
- H. H. Tsai, A. D. Dwarakanath, C. A. Hart, J. D. Milton, and J. M. Rhodes, “Increased faecal mucin sulphatase activity in ulcerative colitis: A potential target for treatment,” Gut, vol. 36, no. 4, pp. 570–576, 1995. [CrossRef]
- N. Mian, C. E. Anderson, and P. W. Kent, “Effect of O-sulphate groups in lactose and N-acetylneuraminyl-lactose on their enzymic hydrolysis.,” Biochem J, vol. 181, no. 2, pp. 387–399, 1979. [CrossRef]
- J. M. H. Larsson et al., “Altered O-glycosylation profile of MUC2 mucin occurs in active ulcerative colitis and is associated with increased inflammation,” Inflamm Bowel Dis, vol. 17, no. 11, pp. 2299–2307, 2011. [CrossRef]
- G. Lennon et al., “Correlations between colonic crypt mucin chemotype, inflammatory grade and Desulfovibrio species in ulcerative colitis,” Colorectal Disease, vol. 16, no. 5, pp. 161–169, 2014. [CrossRef]
- R. Atreya and M. F. Neurath, “IBD pathogenesis in 2014 : Molecular pathways controlling barrier function in IBD,” Nat Rev Gastroenterol Hepatol, vol. 12, no. 1, pp. 37–38, 2015. [CrossRef]
- Braun et al., “Alterations of phospholipid concentration and species composition of the intestinal mucus barrier in ulcerative colitis: A clue to pathogenesis,” Inflamm Bowel Dis, vol. 15, no. 11, pp. 1705–1720, 2009. [CrossRef]
- R. L. Greer, A. Morgun, and N. Shulzhenko, “Bridging immunity and lipid metabolism by gut microbiota,” Journal of Allergy and Clinical Immunology, vol. 132, no. 2, pp. 253–262, 2013. [CrossRef]
- R. Caesar, V. Tremaroli, P. Kovatcheva-Datchary, P. D. Cani, and F. Bäckhed, “Crosstalk between gut microbiota and dietary lipids aggravates WAT inflammation through TLR signaling,” Cell Metab, vol. 22, no. 4, pp. 658–668, 2015. [CrossRef]
- M. Øgaard Mønsted et al., “Reduced phosphatidylcholine level in the intestinal mucus layer of prediabetic NOD mice,” APMIS, vol. 131, no. 6, pp. 237–248, Jun. 2023. [CrossRef]
- L. M. Lichtenberger, “The Hydrophobic Barrier Properties of Gastrointestinal Mucus,” Annu Rev Physiol, vol. 57, no. 1, pp. 565–583, Oct. 1995. [CrossRef]
- R. Ehehalt et al., “Phosphatidylcholine and lysophosphatidylcholine in intestinal mucus of ulcerative colitis patients. A quantitative approach by nanoElectrospray-tandem mass spectrometry.,” Scand J Gastroenterol, vol. 39, no. 8, pp. 737–42, 2004. [CrossRef]
- M. Rosenberg and S. Kjelleberg, “Hydrophobic Interactions: Role in Bacterial Adhesion,” pp. 353–393, 1986. [CrossRef]
- Braun et al., “Reduced hydrophobicity of the colonic mucosal surface in ulcerative colitis as a hint at a physicochemical barrier defect,” Int J Colorectal Dis, vol. 26, no. 8, pp. 989–998, 2011. [CrossRef]
- J. Diab et al., “Lipidomics in Ulcerative Colitis Reveal Alteration in Mucosal Lipid Composition Associated With the Disease State,” Inflamm Bowel Dis, vol. XX, no. Xx, pp. 1–8, 2019. [CrossRef]
- S. Almer, L. Franzén, G. Olaison, K. Smedh, and M. Ström, “Phospholipase a<inf>2</inf> activity of colonic mucosa in patients with ulcerative colitis,” Digestion, vol. 50, no. 3–4, pp. 135–141, 1991. [CrossRef]
- W. Stremmel, A. H. Robert, E. M. Karner, and A. Braun, “Phosphatidylcholine (Lecithin) and the mucus layer: Evidence of therapeutic efficacy in ulcerative colitis?,” Digestive Diseases, vol. 28, no. 3, pp. 490–496, 2010. [CrossRef]
- Braun et al., “Alterations of phospholipid concentration and species composition of the intestinal mucus barrier in ulcerative colitis: A clue to pathogenesis,” Inflamm Bowel Dis, vol. 15, no. 11, pp. 1705–1720, 2009. [CrossRef]
- R. Ehehalt et al., “Phosphatidylcholine and lysophosphatidylcholine in intestinal mucus of ulcerative colitis patients. A quantitative approach by nanoelectrospray-tandem mass spectrometry,” Scand J Gastroenterol, vol. 39, no. 8, pp. 737–742, 2004. [CrossRef]
- Paone and P. D. Cani, “Mucus barrier, mucins and gut microbiota: the expected slimy partners?,” Gut, vol. 69, no. 12, pp. 2232–2243, Dec. 2020. [CrossRef]
- J. K. Gustafsson, A. Ermund, M. E. V. Johansson, A. Schutte, G. C. Hansson, and H. Sjovall, “An ex vivo method for studying mucus formation, properties, and thickness in human colonic biopsies and mouse small and large intestinal explants,” AJP: Gastrointestinal and Liver Physiology, vol. 302, no. 4, pp. G430–G438, Feb. 2012. [CrossRef]
- M. E. V. Johansson, H. Sjövall, and G. C. Hansson, “The gastrointestinal mucus system in health and disease,” Nat Rev Gastroenterol Hepatol, vol. 10, no. 6, pp. 352–361, 2013. [CrossRef]
- D. Pullan et al., “Thickness of adherent mucus gel on colonic mucosa in humans and its relevance to colitis.,” Gut, vol. 35, no. 3, pp. 353–359, 1994. [CrossRef]
- Swidsinski et al., “Comparative study of the intestinal mucus barrier in normal and inflamed colon,” Gut, vol. 56, no. 3, pp. 343–350, 2007. [CrossRef]
- D. Pullan et al., “Thickness of adherent mucus gel on colonic mucosa in humans and its relevance to colitis.,” Gut, vol. 35, no. 3, pp. 353–359, 1994. [CrossRef]
- K. Fyderek et al., “Mucosal bacterial microflora and mucus layer thickness in adolescents with inflammatory bowel disease,” World J Gastroenterol, vol. 15, no. 42, pp. 5287–5294, 2009. [CrossRef]
- Strugala, P. W. Dettmar, and J. P. Pearson, “Thickness and continuity of the adherent colonic mucus barrier in active and quiescent ulcerative colitis and Crohn’s disease,” Int J Clin Pract, vol. 62, no. 5, pp. 762–769, 2008. [CrossRef]
- K. Fyderek et al., “Mucosal bacterial microflora and mucus layer thickness in adolescents with inflammatory bowel disease,” World J Gastroenterol, vol. 15, no. 42, pp. 5287–5294, 2009. [CrossRef]
- Strugala, P. W. Dettmar, and J. P. Pearson, “Thickness and continuity of the adherent colonic mucus barrier in active and quiescent ulcerative colitis and Crohn’s disease,” Int J Clin Pract, vol. 62, no. 5, pp. 762–769, 2008. [CrossRef]
- Swidsinski et al., “Comparative study of the intestinal mucus barrier in normal and inflamed colon,” Gut, vol. 56, no. 3, pp. 343–350, 2007. [CrossRef]
- L. Sardelli et al., “ Towards bioinspired in vitro models of intestinal mucus ,” RSC Adv, vol. 9, no. 28, pp. 15887–15899, 2019. [CrossRef]
- Schreiber et al., “iNOS-dependent increase in colonic mucus thickness in DSS-colitic rats.,” PLoS One, vol. 8, no. 8, 2013. [CrossRef]
- M. E. V. Johansson et al., “Bacteria penetrate the normally impenetrable inner colon mucus layer in both murine colitis models and patients with ulcerative colitis,” Gut, vol. 63, no. 2, pp. 281–291, Feb. 2014. [CrossRef]
- D. Pullan et al., “in Humans and Its Relevance To Colitis,” Gut, pp. 353–359, 1994.
- K. Fyderek et al., “Mucosal bacterial microflora and mucus layer thickness in adolescents with inflammatory bowel disease,” World J Gastroenterol, vol. 15, no. 42, pp. 5287–5294, 2009. [CrossRef]
- D. Ambort et al., “Calcium and pH-dependent packing and release of the gel-forming MUC2 mucin,” Proceedings of the National Academy of Sciences, vol. 109, no. 15, pp. 5645–5650, 2012. [CrossRef]
- C. Bischoff et al., “Intestinal permeability - a new target for disease prevention and therapy,” BMC Gastroenterol, vol. 14, no. 1, pp. 1–25, 2014. [CrossRef]
- Michielan and R. D’Incà, “Intestinal Permeability in Inflammatory Bowel Disease: Pathogenesis, Clinical Evaluation, and Therapy of Leaky Gut,” Mediators Inflamm, vol. 2015, 2015. [CrossRef]
- S. Rao et al., “Urine sugars for in vivo gut permeability: Validation and comparisons in irritable bowel syndrome-diarrhea and controls,” Am J Physiol Gastrointest Liver Physiol, vol. 301, no. 5, p. G919, Nov. 2011. [CrossRef]
- L. Halme, U. Turunen, J. Tuominen, T. Forsström, and U. Turpeinen, “Scandinavian Journal of Clinical and Laboratory Investigation Comparison of iohexol and lactulose-mannitol tests as markers of disease activity in patients with inflammatory bowel disease,” 2009. [CrossRef]
- M. Prager et al., “Myosin IXb variants and their pivotal role in maintaining the intestinal barrier: A study in Crohn’s disease,” Scand J Gastroenterol, vol. 49, no. 10, pp. 1191–1200, Oct. 2014. [CrossRef]
- S. C. Bischoff et al., “Intestinal permeability - a new target for disease prevention and therapy,” BMC Gastroenterol, vol. 14, no. 1, pp. 1–25, 2014. [CrossRef]
- S. Almer, L. Franzen, G. Olaison, K. Smedh, and M. Strom, “Increased absorption of polyethylene glycol 600 deposited in the colon in active ulcerative colitis,” Gut, vol. 34, no. 4, pp. 509–513, 1993. [CrossRef]
- J. Z. H. Von Martels, A. R. Bourgonje, H. J. M. Harmsen, K. N. Faber, and G. Dijkstra, “Assessing intestinal permeability in Crohn’s disease patients using orally administered 52 Cr-EDTA,” PLoS One, vol. 14, no. 2, Feb. 2019. [CrossRef]
- J. N. S. Gregson, A. Steel, M. Bower, B. G. Gazzard, F. M. Gotch, and M. R. Goodier, “Elevated plasma lipopolysaccharide is not sufficient to drive natural killer cell activation in HIV-1-infected individuals,” AIDS, vol. 23, no. 1, pp. 29–34, Jan. 2009. [CrossRef]
- A. Pasternak et al., “Lipopolysaccharide exposure is linked to activation of the acute phase response and growth failure in pediatric Crohn’s disease and murine colitis,” Inflamm Bowel Dis, vol. 16, no. 5, pp. 856–869, May 2010. [CrossRef]
- R. M. McQuade et al., “Gastrointestinal consequences of lipopolysaccharide-induced lung inflammation,” Inflammation Research, vol. 72, no. 1, pp. 57–74, Jan. 2023. [CrossRef]
- E. Ferrer-Picón et al., “Intestinal Inflammation Modulates the Epithelial Response to Butyrate in Patients With Inflammatory Bowel Disease,” Inflamm Bowel Dis, vol. 26, no. 1, pp. 43–55, Jan. 2020. [CrossRef]
- G. Thuijls et al., “Urine-based Detection of Intestinal Tight Junction Loss,” J Clin Gastroenterol, vol. 44, no. 1, pp. e14–e19, Jan. 2010. [CrossRef]
- G. Thuijls et al., “Urine-based Detection of Intestinal Tight Junction Loss,” J Clin Gastroenterol, vol. 44, no. 1, pp. e14–e19, Jan. 2010. [CrossRef]
- Michielan and R. D’Incà, “Intestinal Permeability in Inflammatory Bowel Disease: Pathogenesis, Clinical Evaluation, and Therapy of Leaky Gut,” Mediators Inflamm, vol. 2015, 2015. [CrossRef]
- G. D. De Palma and A. Rispo, “Confocal laser endomicroscopy in inflammatory bowel diseases: Dream or reality?,” World Journal of Gastroenterology, vol. 19, no. 34. Baishideng Publishing Group Inc, pp. 5593–5597, Sep. 2013. doi: 10.3748/wjg.v19.i34.5593.
- Thomson et al., “The Ussing chamber system for measuring intestinal permeability in health and disease,” BMC Gastroenterol, vol. 19, no. 1, p. 98, Jun. 2019. [CrossRef]
- Lautenschläger, C. Schmidt, C. M. Lehr, D. Fischer, and A. Stallmach, “PEG-functionalized microparticles selectively target inflamed mucosa in inflammatory bowel disease,” European Journal of Pharmaceutics and Biopharmaceutics, vol. 85, no. 3 PART A, pp. 578–586, Nov. 2013. [CrossRef]
Figure 1.
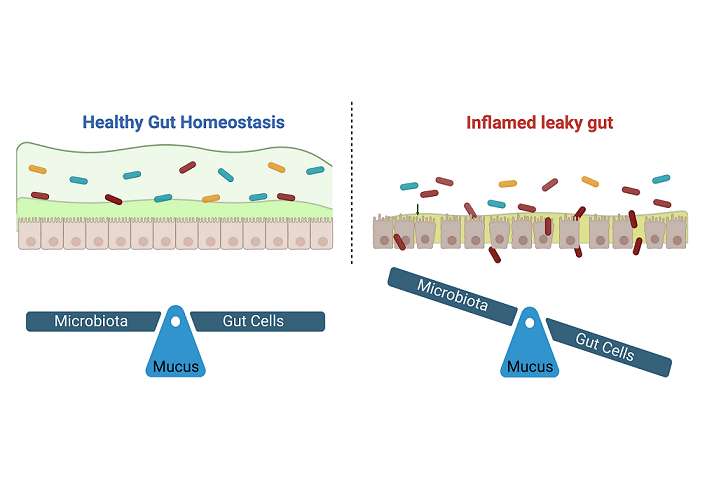
A) Graphical representation of the gut epithelial barrier. From the lumen to the lamina propria, the structure of the barrier is composed by: (1) outer mucus enriched of bacteria; (2) inner mucus; (3) cell layer; (4) lamina propria enriched of immune cells. B) Differences between the CD and UC characteristics in terms of inflammation distribution (darker areas in the images) and commonly related symptoms [15].
Figure 1.
A) Graphical representation of the gut epithelial barrier. From the lumen to the lamina propria, the structure of the barrier is composed by: (1) outer mucus enriched of bacteria; (2) inner mucus; (3) cell layer; (4) lamina propria enriched of immune cells. B) Differences between the CD and UC characteristics in terms of inflammation distribution (darker areas in the images) and commonly related symptoms [15].
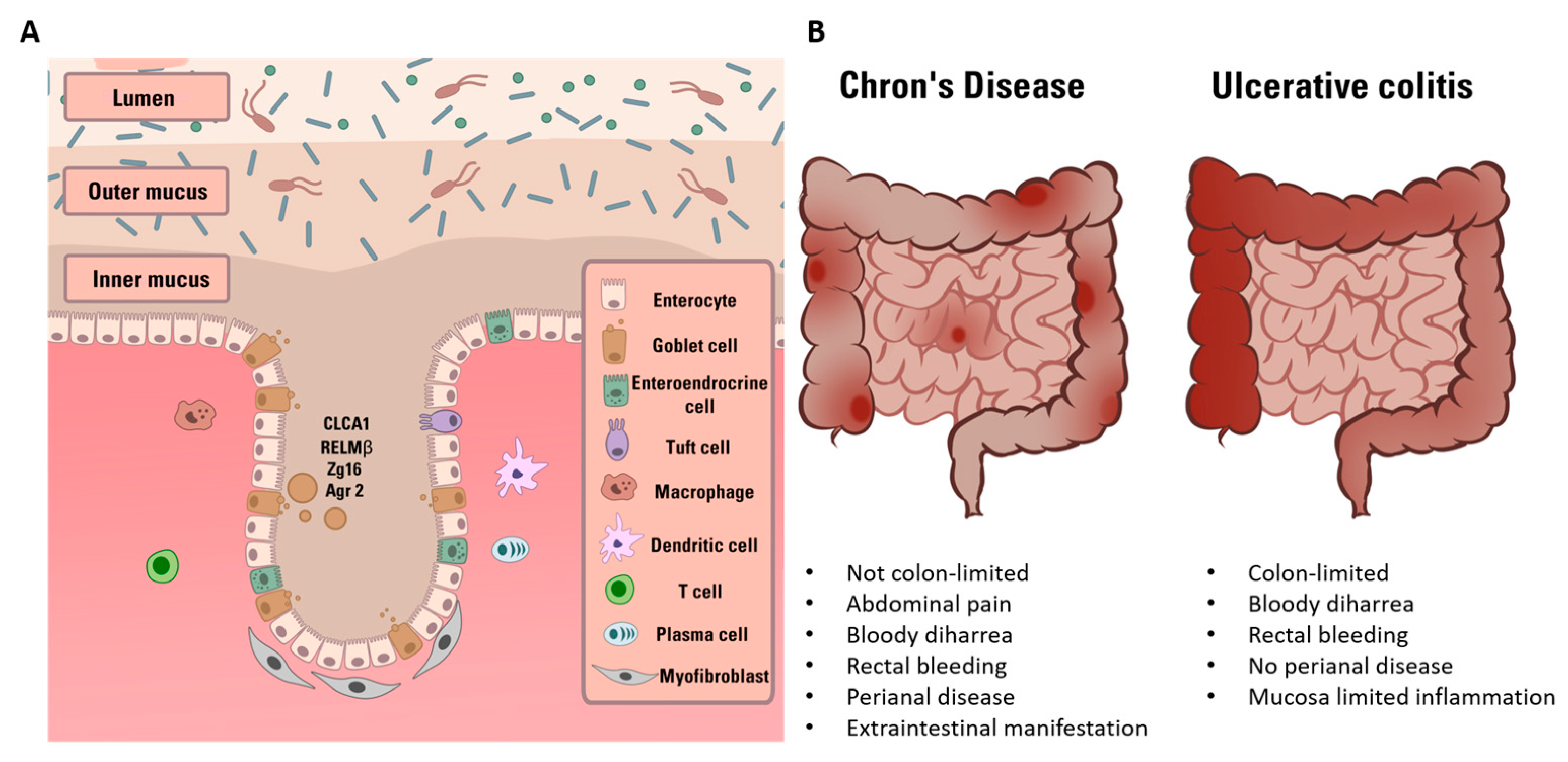
Figure 2.
Graphical representation of molecular mechanisms involved in barrier impairment in a physiological (left) or IBD condition (right). (Left) SCFA-producing bacteria are abundant and release large quantity of SCFAs (green particles). These molecules interact with epithelium cells and immune cells leading to AMPs production, immune tolerance and Treg cells maturation. (Right) in IBD pathological condition, SCFA-producing bacteria together with the anti-inflammatory effect and immune tolerance decrease. Consequently, NF-kB-mediated pathway leads to pro-inflammatory factor production and immune response. All together, these mechanisms cause TJs disruption, gut barrier impairment and increase of permeability to pro-inflammatory bacterial molecules.
Figure 2.
Graphical representation of molecular mechanisms involved in barrier impairment in a physiological (left) or IBD condition (right). (Left) SCFA-producing bacteria are abundant and release large quantity of SCFAs (green particles). These molecules interact with epithelium cells and immune cells leading to AMPs production, immune tolerance and Treg cells maturation. (Right) in IBD pathological condition, SCFA-producing bacteria together with the anti-inflammatory effect and immune tolerance decrease. Consequently, NF-kB-mediated pathway leads to pro-inflammatory factor production and immune response. All together, these mechanisms cause TJs disruption, gut barrier impairment and increase of permeability to pro-inflammatory bacterial molecules.
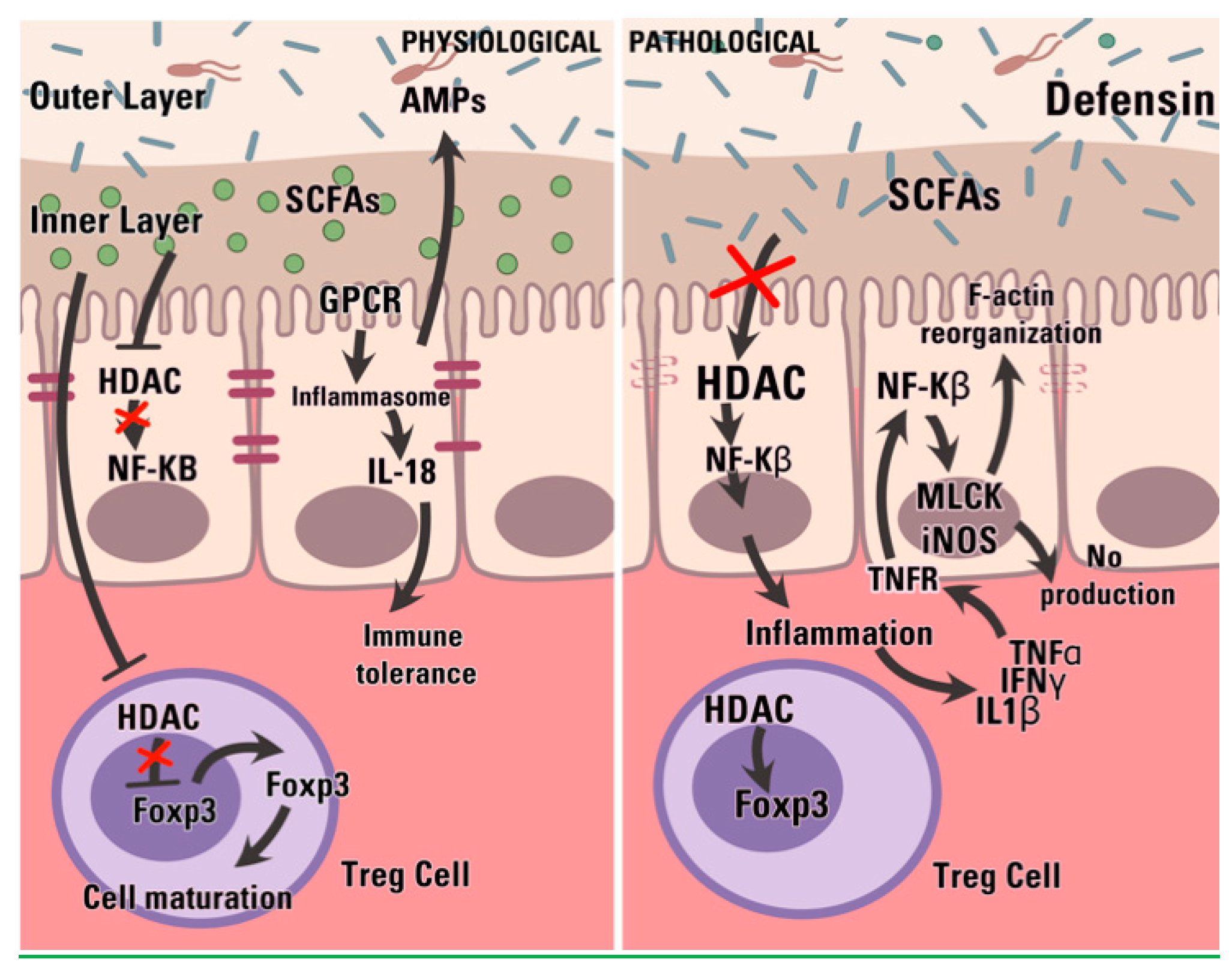
Figure 3.
In physiological condition (A), the mucus acts as a barrier for either toxins or bacteria adhesion (red arrows), modulating the immune response of the epithelial cell while filtering the nutrients and other molecules. During IBD (B) compositional changes and structural weakening impair these functions; bacteria are free to adhere to the epithelium and toxin diffusion is facilitate. These events contribute to the maintenance of the pathological state of inflammation.
Figure 3.
In physiological condition (A), the mucus acts as a barrier for either toxins or bacteria adhesion (red arrows), modulating the immune response of the epithelial cell while filtering the nutrients and other molecules. During IBD (B) compositional changes and structural weakening impair these functions; bacteria are free to adhere to the epithelium and toxin diffusion is facilitate. These events contribute to the maintenance of the pathological state of inflammation.

Table 1.
Gut microbiota composition versus CD and UC. Summary of CD and UC impact on gut microbiota composition and function respect to healthy conditions.
Table 1.
Gut microbiota composition versus CD and UC. Summary of CD and UC impact on gut microbiota composition and function respect to healthy conditions.
| Decrease | Increase | Dysbiosis impact | Ref. | |
|---|---|---|---|---|
| Chron’s disease | Actinobacteria Bifidobacterium adolescentis Bacteroidetes Bacteroides fragilis Firmicutes Fecalibacterium Praustnizii Eubacterium spp. Lachnospiraceae Clostridium prausnitzii Roseburia spp. Ruminococcus spp. Dialister invisus |
Actinobacteria Bifidobacteriaceae Coriobacteriaceae Firmicutes Ruminococcus gnavus Clostridium difficile Proteobacteria Escherichia coli |
Reduction of defensins production by Paneth cells Lower level of SCFAs Gut inflammation |
[62,69,70,82,85,86,87,88] |
| Ulcerative colitis | Firmicutes Roseburia hominis Fecalibacterium Praustnizii Clostridium Enterococcus Proteobacteria Escherichia coli |
Firmicutes Clostridium difficile |
Reduction of defensins production by Paneth cells Lower level of SCFAs Gut inflammation |
[81,86,89,90] |
Table 2.
Compositional variation in the human colonic mucus in IBD extracted from [119] and [114]. The mean values obtained were weighted with reference to the standard deviation (s.d.) and the mean s.d. obtained by the formula: 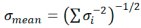 .
.
.| HEALTHY | CD | UC | |
|---|---|---|---|
|
LPC mean value [pmol/100 µg protein] |
4371 ± 487 | 2961 ± 287 | 1738 ± 288 |
|
PC mean value [pmol/100 µg protein] |
2983 ± 321 | 2508 ± 415 | 676 ± 104 |
| % of LPC / protein | 2.22% ± 0.25% | 1.50% ± 0.15% | 0.88% ± 0.15% |
| % of PC / protein | 2.30% ± 0.25% | 1.94% ± 0.32% | 0.52% ± 0.08% |
| LPC/PC | 1.45 ± 0.32 | 1.18 ± 0.31 | 2.57 ± 0.84 |
Table 3.
Human mucus thickness as indicator of the intestinal barrier weakening occurring during either Crohn’s disease (CD) or ulcerative colitis (UC). N/A = not available.
Table 3.
Human mucus thickness as indicator of the intestinal barrier weakening occurring during either Crohn’s disease (CD) or ulcerative colitis (UC). N/A = not available.
| Right colon/caecum | Left colon/ Sigmoid | Rectum | Ref. | |
|---|---|---|---|---|
| Controls | 107 ± 48 | 134 ± 68 | 155 ± 54 | [141] |
| N/A | 218 ± 81.07 | N/A | [142] | |
| N/A | 450 ± 70 | N/A | [143] | |
| UC | N/A | 83 ± 49.93 | N/A | [142] |
| 90 ± 79 | 43 ± 45 | 60 ± 86 | [141] | |
| CD | 190 ± 83 | 232 ± 40 | 294 ± 45 | [141] |
| N/A | 74 ± 40 | N/A | [142] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



