Submitted:
04 May 2023
Posted:
08 May 2023
You are already at the latest version
Abstract
mTORC1 regulates mammalian cell metabolism and growth, in response to diverse environmental stimuli. Nutrient signals control the localization of mTORC1 onto lysosome-surface scaffolds that are critically implicated in its amino acid-dependent activation. Arginine, Leucine and S-Adenosyl-Methionine (SAM) can serve as major mTORC1-signaling activators, with SAM binding to SAMTOR (SAM + TOR), a fundamental SAM sensor, abolishing protein’s (SAMTOR’s) inhibitory action(s) against mTORC1, thereby releasing its (mTORC1) kinase activity. Given the lack of knowledge regarding the role of SAMTOR in invertebrates, we have, in silico, identified the Drosophila SAMTOR homologue (dSAMTOR) and have, herein, genetically targeted it, through utilization of the GAL4/UAS transgenic tool. Survival profiles and negative geotaxis patterns were examined both in control and dSAMTOR-downregulated adult flies, during aging. One of the two gene-targeted schemes resulted in lethal phenotypes, whereas the other one caused rather moderate pathologies in most tissues. Screening of head-specific kinase activities, via PamGene technology application, unveiled the significant upregulation of several kinases, including the dTORC1 characteristic substrate, dp70S6K, in dSAMTOR-downregulated flies, thus strongly supporting the inhibitory dSAMTOR function(s) upon dTORC1/dp70S6K signaling axis in Drosophila-brain settings. Importantly, genetic targeting of the Drosophila BHMT -bioinformatics- counterpart (dBHMT), an enzyme that catabolizes Betaine to produce Methionine (the SAM precursor), led to severe compromises in fly longevities, with glia-, motor neuron- and muscle-specific dBHMT downregulations exhibiting the strongest effects. Abnormalities in wing-vein architectures were also detected in dBHMT-targeted flies, thereby justifying their notably reduced negative geotaxis capacities, herein, observed, mainly in the brain-(mid)gut axis. In vivo adult-fly exposure to clinically relevant doses of Methionine revealed the mechanistic synergism of decreased dSAMTOR and increased Methionine levels in pathogenic longevity, thus rendering (d)SAMTOR an important component in Methionine-associated disorders, including Homocystinuria(s).
Keywords:
Aging
; Betaine
; dBHMT
; Drosophila
; dSAMTOR
; dTORC1
; Kinase
; Methionine
; SAM
1. Introduction
The mechanisms that control organismal growth and its coordination with the availability of nutrients in cellular micro-environments were unknown until a few decades ago. It is now well appreciated that the mechanistic Target Of Rapamycin (mTOR) protein kinase (formerly “mammalian” TOR) represents the major nutrient-sensitive regulator of animal growth [1,2]. The mTOR pathway responds to diverse environmental signals, coordinates cell growth and proliferation [2,3], and its deregulation has been strongly associated with aging and a number of human disorders, such as diabetes, neurodegenerative diseases and cancer [4,5].
mTOR is an evolutionary conserved Serine/Threonine protein kinase that belongs to the PI3K-relate kinases (PIKK) family and is the catalytic core of two large multi-protein complexes designated mTORC1 (mTOR complex 1) and mTORC2 (mTOR complex 2) [2,6]. Their function, substrate specificity, upstream inputs, downstream outputs and sensitivity to Rapamycin differ notably due to the distinct composition of their accessory proteins [5,6]. mTORC1 is better characterized and reviewed between the two mTOR complexes. It controls cell growth and metabolism, by regulating anabolic and catabolic processes, including protein, lipid and nucleotide synthesis, while blocking autophagy at the post-translational and transcriptional levels, in response to various environmental cues, such as growth factors, energy status, oxygen, stress and especially amino acids [3,6,7].
Amino acids, which are not only essential components for protein synthesis, but also serve as source of energy and carbon for many metabolic pathways, are also key regulators of mTORC1 [2,4]. Amino acids derive either from lysosome-mediated protein degradation or are exogenously sensed by the GATOR–Rag Guanosine Triphosphatase (GTPase) pathway, which activates the mTORC1 cascade [8]. Rag GTPases interact with Raptor, a major component of mTORC1, to translocate mTORC1 to the lysosomal membrane, whereat it is, then, activated by Rheb. Translocation of mTORC1 requires the interaction of Rag GTPases with Ragulator, which is a complex located on the lysosome surface [9].
mTORC1 is capable of sensing intra-lysosomal and cytosolic amino acid concentrations through distinct mechanisms and sensors [2,10]. Cytosolic Arginine and Leucine can signal to mTORC1 via different pathways controlled by the GATOR2 and GATOR1 complexes. GATOR1 is a negative regulator of mTORC1, when bound to the lysosome, via recruitment by KICSTOR -protein- complex, whereas GATOR2 acts as positive regulator [9]. Leucine and Arginine bind to the cytosolic amino acid sensor proteins Sestrin1/2 and CASTOR1, respectively, and dissociate them from GATOR2, which is, then, activated and triggers the activation of mTORC1, through the inactivation of GATOR1 [11,12]. Remarkably, mTORC1 cannot sense Methionine directly, but only indirectly, via a previously uncharacterized protein, called SAMTOR. SAMTOR is an S-Adenosyl-Methionine (SAM) cytoplasmic sensor, which interacts with KICSTOR and GATOR1, to release mTORC1 from the lysosomal membrane, and, thereby, acts as negative regulator of mTORC1. SAM is biochemically converted and produced from Methionine, and, under Methionine sufficiency, SAM binds to SAMTOR, which, then, dissociates from GATOR1. The disruption of SAMTOR–GATOR1 complex causes inactivation of GATOR1, which results in mTORC1 activation [2,7,13].
To in vivo investigate the role of SAMTOR in model biological environments, we have, herein, targeted the Drosophila homologous gene, dSAMTOR, in diverse fly tissues during aging, by combined employment of the GAL4/UAS and RNAi transgenic technologies. Besides dSAMTOR, dBHMT, the gene controlling the Betaine to Methionine conversion, was also downregulated, in tissue-specific manner, throughout lifetime. System pathologies derived from gene targeting revealed the in vivo importance of Methionine-SAM axis in cellular integrity, tissue architecture and organ functionality in elderly individuals.
2. Results
2.1. RNAi-mediated downregulation of the evolutionary conserved dSAMTOR protein causes either lethal or viable phenotypes: a gene-targeting efficiency dependence
Cells have a complex system to sense the availability of amino acids, which are crucial activators of mTORC1. SAMTOR, a recently identified protein that serves as an indirect sensor of Methionine, herein proves to exhibit strong amino acid sequence homology among human (Homo sapiens), mouse (Mus musculus), zebrafish (Danio rerio) and fly (Drosophila melanogaster) species, according to the Clustal Omega-mediated alignment (Figure 1), thereby indicating its (SAMTOR) structural and functional conservation during evolution.
To investigate the in vivo role of Drosophila homologue, dSAMTOR (Figure 1), in fly’s systemic pathophysiology, we have genetically targeted the dSAMTOR cognate gene in all organ tissues, through implementation of the appropriate crosses of whole body or tissue-specific, respective, GAL4 drivers, and two UAS-SAMTOR_RNAi strains, via combined employment of the GAL4/UAS and RNAi transgenic technologies. Surprisingly, two distinct phenotypes were obtained, with the first one being typified by embryonic lethality in the majority of dSAMTOR-targeted tissues (Table 1) and the second one being characterized by moderate pathologies of age-dependent profiles. It seems that neuronal system (elav.L-GAL4), glia cell populations (repo-GAL4), motor neurons (D42-GAL4) and muscles (mef2-GAL4) cannot tolerate potent reductions in dSAMTOR protein levels, which are mechanistically associated with systemic lethality (Table 1). In contrast, midgut (NP1-GAL4), eye (ninaE.GMR-GAL4) and wing (bxMS1096-GAL4) tissues carrying strong downregulation of dSAMTOR gene produced viable offsprings (Table 1). As expected, whole body (Act5C-GAL4) dSAMTOR silencing also generated two different phenotypes; a lethal and viable one (Table 1). This indicates the dependence of pathology severity on dSAMTOR targeting efficiency, with the “Strong” and “Moderate” strains being named as SAMTOR_RNAi(S) and SAMTOR_RNAi(M), respectively (Table 1). In accordance, moderately reduced expression of dSAMTOR gene in all body tissues (Act5C>SAMTOR_RNAi(M)) could not affect (in a statistically significant manner) the survival profiles in both male and female transgenic flies, compared to control settings (Act5C-GAL4/+) (Figure S1). Taken together, SAMTOR has proved as an evolutionary conserved protein, with tissue-specific downregulation of dSAMTOR resulting in either systemic mortality or systemic viability, phenotypes that are likely coupled with the targeting efficiency level of dSAMTOR gene.
2.2. Differential contribution of neuronal tissue-, glia cell-, motor neuron- and muscle tissue-specific moderate silencing of dSAMTOR gene to fly survival and kinetic capacity, during aging
Τhe downstream metabolite of Methionine, S-Adenosyl-Methionine (SAM), serves as the main methyl-group donor for numerous biochemical reactions, including the ones forming Mono-Amine Neuro-Transmitters, which are molecules used by the nervous system to transmit messages between neurons, or from neurons to other body parts. Hence, to examine if dSAMTOR’s moderate downregulation alters the nervous system development and function, neuronal cell-specific targeting of dSAMTOR gene was carried out, via employment of the elav.L-GAL4 genetic driver. Different effects in Drosophila longevity were observed after neuronal lowering of dSAMTOR contents, following sex-dependent profiles. The dSAMTOR gene’s moderate suppression significantly increases mortality of male flies (elav.L>SAMTOR_RNAi(M)), compared to control conditions (elav.L-GAL4/+), whereas dSAMTOR-targeted female flies present slightly increased viability (Figure 2A, Β). Climbing profiles, reflecting kinetic potency, of female and male transgenic flies, characterized by RNAi-mediated targeting of the dSAMTOR gene, specifically in neuronal tissues, are not notably affected, as compared to control fly populations. Nevertheless, the examined transgenic female and male flies (elav.L>SAMTOR_RNAi(M)) show reduced kinetic capacities till the 10th day for the female and at the 20th day for the male populations, compared to control settings (elav.L-GAL4/+), with all double-targeted flies ultimately recovering at the 30th experimentation day (Figure 2C, D).
In the same context, after dSAMTOR gene’s moderate silencing, specifically in glia cells, life expectancy proves to notably differ in both sexes (repo>SAMTOR_RNAi(M)) from control flies (repo-GAL4/+), and is markedly reduced after the 60th day of life (Figure 2E, F). Furthermore, our results demonstrate that kinetic capacities are rather similar in between dSAMTOR-targeted flies (repo>SAMTOR_RNAi(M)) and control populations (repo-GAL4/+), except the 10th experimentation day that is characterized by decreased mobility of dSAMTOR-downregulated flies for both sexes (repo>SAMTOR_RNAi(M)), as compared to control flies (repo-GAL4/+) (Figure 2G, H). Like neuronal targeting, glial-specific (moderate) downregulation of the dSAMTOR gene causes kinetic pathologies that can be alleviated during aging (e.g. at the 30th day of life, since birth), thereby indicating the critical contribution of dSAMTOR signaling activity to neuronal and glial tissues that control Drosophila’s kinetic functionality during early lifespan.
To further investigate the role of dSAMTOR protein in neuro-muscular systemic integrity, the dSAMTOR gene was moderately downregulated, specifically either in motor neurons (D42) or in muscles (mef2), via engagement of the appropriate GAL4 genetic drivers. Different survival profiles were obtained from female and male flies, carrying modestly suppressed dSAMTOR gene expression, specifically in motor neurons. Longevity of transgenic male flies (D42>SAMTOR_RNAi(M)) declined till (approximately) the 60th day of life (since birth), but was elevated after that time (Figure 3A), as compared to control populations (D42-GAL4/+). dSAMTOR-targeted (D42>SAMTOR_RNAi(M)) females’ viability was not affected (in a statistically significant manner), albeit a tendency towards an improved survival was observed (Figure 3B). Climbing (negative geotaxis) activities of female and male transgenic flies proved generally similar to each other in dSAMTOR moderately downregulated in vivo settings (D42>SAMTOR_RNAi(M)), with the obtained phenotypes featuring significantly compromised mobilities of age-dependent pathologies that could not be rescued at the 30th day of lifetime (Figure 3C, D).
Moderate silencing of dSAMTOR gene expression, specifically in muscles, does not seem to notably alter neither males’ longevity (Figure 3F) nor kinetic capacities of both sexes, except day 20 in males (mef2>SAMTOR_RNAi(M)), compared to control transgenic animals (mef2-GAL4/+) (Figure 3G, H). Only transgenic females bearing modest downregulation of dSAMTOR protein levels (mef2>SAMTOR_RNAi(M)) show increased viability (in a statistically significant manner), compared to control fly populations (mef2-GAL4/+) (Figure 3E), albeit a tendency for improved longevity was detected in transgenic dSAMTOR-targeted males (Figure 3F). Altogether, it seems that moderate reductions of dSAMTOR protein contents in motor neurons or muscles can benefit Drosophila longevity, but can harm climbing (locomotor) functions in age-dependent and tissue-specific fashions.
2.3. dSAMTOR midgut-specific suppression alters Drosophila’s longevity
D. melanogaster’s viability after moderate dSAMTOR gene silencing, specifically in midgut tissues, is affected in a sex-dependent manner. Male transgenic flies (NP1>SAMTOR_RNAi(M)) show increased mortality till day 40, compared to control groups (NP1-GAL4/+), with their viability being intriguingly improved after day 40 (Figure 4B). On the contrary, lifespan profiles of moderately dSAMTOR-targeted female flies do not differ significantly from control animals (Figure 4A). Nevertheless, climbing capacities of both female and male transgenic flies carrying modestly dSAMTOR-downregulated activity levels (NP1>SAMTOR_RNAi(M)) seem to statistically resemble the ones of control populations (NP1-GAL4/+), although a notable reduction can be observed at the 20th day of males‘ life (Figure 4C, D). Remarkably, strong RNAi-mediated suppression of dSAMTOR gene expression, specifically in midgut tissues, causes strikingly reduced longevities in transgenic flies of both sexes (NP1>SAMTOR_RNAi(S)), always compared to control settings (NP1-GAL4/+) (Figure 4E, F). Due to increased mortality of transgenic flies, the collection of a statistically significant number of transgenic flies for the reliable negative geotaxis assay proved impossible, and therefore, the series of climbing activity experiments were not performed. Taken together, it is the different efficiency of dSAMTOR gene targeting and/or downregulation process that directs the two distinct pathologies in midgut tissues, herein detected, with the (comparatively) more severe phenotype, of highly compromised longevity, markedly tolerating embryonic lethality, in contrast to other tissue type-specific (e.g. nervous system and muscles) strong silencing schemes of dSAMTOR gene expression, described in Table 1. Since midgut serves as the first nutritional entrance and food-management hub in fly body, it has to resist against any amino acid-dependent imbalance in basic metabolism, biosynthesis and signaling, likely induced by dSAMTOR lack, thereby providing strong support to nourishment and feeding of secondary organic systems, such as brain, glia, motor neurons and muscles.
2.4. Functional kinome profiling of mildly dSAMTOR-downregulated transgenic flies, specifically in neuronal tissues
Global profiling of the functional (active/ated) kinome is a recently developed, novel, versatile, multi-faceted and powerful molecular platform that offers the opportunity to screen and map several hundreds of protein kinase activities in a given biological specimen, in vitro. Kinase-mediated phosphorylation plays a critical regulatory role in many cellular processes, and protein kinases are decisively involved in multiple and diverse cellular functions, including, among others, cell proliferation, differentiation, and migration. Kinases represent a special and unique class of drug targets, capable of opening new windows for the development of novel targeted therapies for various diseases. PamGene platform is a well-established micro-array technology, for simultaneous multiplex kinase-activity profiling for several hundred (>430) of protein kinases in complex biological samples, and, thus, can help in bridging the gap between animals, or in vitro models, and human diseases.
Hence, in our study, we used the PamGene Ser/Thr Kinase (STK) chips, which are composed of a grid of reporter peptides, derived from literature or computational predictions, that can be phosphorylated by Ser/Thr protein kinases present in total-protein lysates of control (elav.L-GAL4/+) and moderately dSAMTOR-downregulated (elav.L>SAMTOR_RNAi(M)), manually dissected, Drosophila heads. Phosphorylated peptides were recognized by phospho-specific and FITC-labeled antibodies, while signal detection was performed in multiple cycles at different exposure times and captured by a high-resolution CCD camera. In mildly dSAMTOR-targeted transgenic flies, specifically in neuronal tissues (elav.L>SAMTOR_RNAi(M)), there is a differential substrate phosphorylation pattern detected, compared to control populations (elav.L-GAL4/+), as shown in the captured CCD images of PamChip microarrays (Figure 5A). Peptides with significant differences in signal intensities are visualized after normalization in log-transformed heatmaps (Figure 5B), which demonstrate the degree of phosphorylation for each peptide per genetic condition (control, or dSAMTOR modest suppression). Signals are sorted from high to low intensity/phosphorylation for control animals and flies carrying moderately silenced dSAMTOR expression contents. Figure 5C describes the Median Final Score plots, indicating putative Serine/Threonine (Ser/Thr) kinases ranked by their specificity score. Importantly, p70S6K (a bona fide substrate of activated mTOR kinase) is clustered in the top ranked (Ser/Thr) functional (active/ated) kinases, in the dSAMTOR mildly downregulated fly heads, as compared to control settings. Of note, x-axis indicates the values for normalized kinase activity statistics (e.g. positive value = activity of the corresponding Ser/Thr protein kinase is increased in modestly dSAMTOR-silenced transgenic flies). Altogether, an active/ated kinome-wide mapping of Drosophila head, in wild-type and mutant environments, unveils the negative role of dSAMTOR determinant in the dTOR-dp70S6K signaling axis, in vivo. The contribution of other functionally upregulated (e.g. ERK5, ROCK1 and IKKβ) or downregulated (e.g. TBK1 and IKKε) Ser/Thr protein kinases, to the obtained pathologies, in dSAMTOR modestly suppressed settings (Figure 5C), is a novel and interesting mechanistic issue that necessitates further exploration.
2.5. Strong downregulation of dSAMTOR gene expression causes eye and wing dysmorphic phenotypes in Drosophila
To reveal the role of dSAMTOR protein in eye and wing morphogenesis, during Drosophila aging, we have, herein, targeted dSAMTOR gene expression, specifically in the compound eye and wing disc, respectively, via crossing of the appropriate GAL4-genetic drivers with the UAS-SAMTOR_RNAi(S) or UAS-SAMTOR_RNAi(M) transgenic strains. Our results show that moderate downregulation of dSAMTOR gene in the eye disc (ninaE.GMR>SAMTOR_RNAi(M)) (Figure 6A, d-f) does not detectably affect the normal structure and organization of compound eye, during aging, whereas strong dSAMTOR gene silencing (ninaE.GMR>SAMTOR_RNAi(S)) (Figure 6A, g-i) significantly alters eyes’ morphology and architecture, with cilia loss following an age-dependent dysmorphic pattern, compared to control conditions (ninaE.GMR-GAL4/+) (Figure 6A, a-c).
Control (bxMS1096-GAL4/+) fly wings are characterized by the typical morphogenetic structure being comprised of five main veins (L1-L5), a posterior cross-vein (P-CV) and an anterior cross-vein (A-CV), both in female (Figure 6B, a) and male (Figure 6B, d) flies. Moderately suppressed dSAMTOR gene activity, in female (Figure 6B, b) and male (Figure 6B, e) flies (bxMS1096>SAMTOR_RNAi(M)), seems unable to induce any detectable alteration in wing organization and architecture. However, strongly reduced levels of dSAMTOR protein, specifically in the wing disc, for both sexes (bxMS1096>SAMTOR_RNAi(S)), causes severe pathologies, since wings' structural architecture is totally disrupted, carrying highly dysmorphic features, both in transgenic female (Figure 6B, c) and male (Figure 6B, f) flies. Like midgut (Figure 4), eye- and wing-disc tissues can, also, resist embryonic lethality upon dSAMTOR lack (Table 1), thus indicating their ability to compensate for SAM-dSAMTOR-dTOR axis signaling disruption, to obviating system’s mortality.
2.6. Sensing and metabolism of Methionine are evolutionary conserved pathways
Methionine, an essential amino acid required for organisms’ normal growth and development, is converted to S-Adenosyl-Methionine (SAM), a major methyl donor in cells, by an enzyme called Methionine S-Adenosyl-Transferase (MAT). SAM binds to SAMTOR, de-repressing mTORC1 signaling (target-phosphorylation) activities. Methionine is supplied by nutrition or is endogenously produced from Homocysteine via Methionine Synthase (MTR), or Betaine Homocysteine Methyl-Transferase (BHMT) involvement (Figure 7A). Through engagement of the Clustal Omega bioinformatics tool, we, herein, prove that BHMT protein is evolutionary conserved, in its amino acid sequence, among human (Homo sapiens), mouse (Mus musculus), zebrafish (Danio rerio) and fly (Drosophila melanogaster) species (Figure 7Β), thereby providing a new and powerful genetic frame for the mechanistic association of dBHMT gene targeting with age-dependent pathologies in Drosophila tissues and organs.
2.7. Methionine and Betaine administration induce reduced viabilities in transgenic flies, bearing moderately suppressed dSAMTOR gene expression, specifically in neuronal tissues
Besides the RNAi-mediated, genetic, downregulation of dSAMTOR protein, we, also, attempted the exogenous administration (in the food) of Methionine or Betaine (Trimethylglycine) amino acids, and, subsequent, monitoring of transgenic flies’ viability, in modest dSAMTOR gene silencing settings, specifically in neuronal tissues. Female and male flies, characterized by moderately suppressed expression of dSAMTOR gene (elav.L>SAMTOR_RNAi(M)) (specifically in neuronal system), after their exposure to Methionine (5 mM; administered in the food), for up to 60 consecutive days, present a remarkably increased mortality, compared to control animals (elav.L-GAL4/+) (Figure 8A, B).
Likewise, supplementation of food with Betaine (10 mM) results in prominent reduction of longevity in male transgenic flies (elav.L>SAMTOR_RNAi(M)) (Figure 8D), compared to control populations (elav.L-GAL4/+), although transgenic female lifespan profiles do not seem to differ (in a statistically significant manner) from control crosses-derived ones (Figure 8C). The synergism of Methionine food enrichment with the neuronal-specific mild downregulation of dSAMTOR protein contents, in lifespan decrease, indicates the in vivo ability of our, herein developed, genetic scheme to, indeed, target and suppress the dSAMTOR gene expression in detectable and productive, albeit moderate, levels. Similarly, Betaine can synergize with, neural system-specific, dSAMTOR modest downregulation, in diminishing lifespan, exclusively in male, but not female, transgenic flies. This sex-dependent, surprising, response may be mechanistically related to the absence (or compromise) of genomic imprinting process (e.g. Betaine metabolism) in Drosophila adult females, as previously reported [14]. Presumable duplication of allele-expression dose, in females, likely attenuates the RNAi machinery efficacy, thereby justifying the lack of pathology in transgenic female flies, in Betaine-enriched environments.
2.8. Lack of dBHMT gene expression, specifically in neuronal-, glial-, motor neuron- muscle- and midgut-tissues shortens Drosophila’s longevity
RNAi-mediated dBHMT gene silencing, specifically in neuronal tissues, significantly increases mortality of male flies (elav.L>BHMT_RNAi), compared to control settings (elav.L-GAL4/+) (Figure 9B), whereas dBHMT-targeted female flies are presented with elevated viability in older ages (after day 45) (Figure 9A). Most importantly, suppression of dBHMT gene expression, specifically in glia cells (repo>BHMT_RNAi) (Figure 9C, D), motor neurons (D42>BHMT_RNAi) (Figure 9E, F), muscle tissues (mef2>BHMT_RNAi) (Figure 9G, H) and midgut (NP1>BHMT_RNAi) (Figure 9I, J), leads to remarkably reduced longevities of both female and male flies, always compared to control populations (repo-GAL4/+), (D42-GAL4/+), (mef2-GAL4/+) and (NP1-GAL4/+), respectively. Taken together, it seems that dBHMT enzymatic activity plays an indispensable role in diverse tissue- and organ-dependent survival programs in Drosophila, during aging.
2.9. Differential contribution of neuronal, glial, motor neuron, muscle and midgut tissue-specific silencing of dBHMT gene expression to Drosophila locomotion, during aging
Lack of dBHMT protein synthesis, specifically in neuronal tissues, causes sex-dependent kinetic pathologies, with climbing (negative geotaxis) activities being reduced in male populations (elav.L>BHMT_RNAi) (Figure 10B), but remaining unaffected in female flies (Figure 10A), compared to control groups (elav.L-GAL4/+). Locomotor functions of female and male dBHMT-downregulated flies, specifically in glia cells (repo>BHMT_RNAi) (Figure 10C, D) or muscle tissues (mef2>BHMT_RNAi) (Figure 10G, H) do not seem to differ (in a statistically significant manner) from control genetic environments (repo-GAL4/+) and (mef2-GAL4/+), respectively. Remarkably, motor neuron-specific silencing of dBHMT gene D42-Gal4 (D42>BHMT_RNAi) causes notable reduction of motility for both Drosophila sexes, during aging, with females exhibiting (comparatively) more severe pathogenic features in kinetic potency (Figure 10E, F). Similar climbing-activity profiles are, also, obtained, in response to midgut-specific downregulation of dBHMT gene expression (NP1>BHMT_RNAi), revealing sex-dependent kinetic pathologies. Intriguingly, dBHMT-targeted female flies, specifically in their midgut tissues (NP1>BHMT_RNAi), are presented with elevated kinetic capacity (Figure 10I), whereas transgenic males’ locomotor performance proves prominently deteriorated, during aging, compared to control populations (NP1-GAL4/+) (Figure 10J). Altogether, it seems that different tissues and organs can unequally contribute to kinetic pathologies derived from dBHMT genetic targeting and silencing.
2.10. dBHMT gene downregulation critically affects Drosophila eye and wing development
Suppression of dBHMT gene expression in Drosophila’s compound eye-disc tissues (ninaE.GMR>BHMT_RNAi) proves to markedly disrupt eye cilia normal formation and distribution, during aging, since from the 10th day of lifetime there is a progressive loss of eye cilia observed, while at day 50 an absence of cilia is detected in a comparatively greater extent (Figure 11D-F), in dBHMT-targeted (male) flies compared to control settings (ninaE.GMR-GAL4/+) (Figure 11A-C).
Regarding wing morphogenesis, transgenic flies with reduced dBHMT protein contents, specifically in the wing disc (bxMS1096>BHMT_RNAi), were collected at day 1, 10, 20 and 30 of lifetime and (adult) wings' structural architecture was examined via a stereo-microscopic approach. Obtained phenotypes seem to vary, with abnormal vein formation being the most frequent pathogenic feature detected (Figure 12A, b-d), compared to normally developed (typical) fly wings (bxMS1096-GAL4/+) (Figure 12A, a). The most striking morphogenetic defects are being identified in the L4 wing vein, followed by architectural pathologies in the L2 and L5 veins, during aging (1-30 days, post-birth), compared to control populations (Figure 12B, a-d). Of note, compared to control settings, dBHMT-targeted flies, specifically in the wing disc, are characterized by slightly smaller average wing surface at days 1 and 10 of lifetime, with average wing-surface area being detectably increased (~5%) at the older adult-fly ages of 20 and 30 days (Figure 12C). Given the relatively mild pathology developed, the dBHMT-mediated conversion of Betaine (and Homocysteine) to Methionine does not seem to play a major role in both eye and wing formation, and structural integrity, during Drosophila aging.
3. Discussion
Nutrients are directly involved in the regulation of lifespan, aging and metabolic health of organisms, with most metabolic-related cellular activities being mediated by the mTORC1 complex, which functions as the central regulator of cell growth and metabolism in response to changes in nutrient signals, such as specific amino acid(s) absence or (excessive) presence [15]. The essential amino acid Methionine is presented as a key metabolite in many aspects of animal physiology, with its metabolism being regulated in an age-dependent manner [16]. Methionine’s primary metabolite, SAM, serves as a universal methyl donor required for most methylation reactions, including those of DNA and histones [17,18,19]. The mTORC1 signaling pathway senses the cellular levels of Methionine in an indirect fashion, through the SAM-specific sensor, SAMTOR [7]. SAMTOR protein is a recently described cytoplasmic amino acid sensor, able to negatively regulate mTORC1 signaling by inducing its (mTORC1) dissociation from the lysosomal membrane. When SAM binds to SAMTOR, the SAMTOR-containing complex can be disrupted and inhibition of mTORC1 signaling activity can be abolished [7]. SAMTOR is highly conserved among metazoans (Figure 1), and in Drosophila S2R+ cells lacking dSAMTOR, the dTOR pathway is resistant to Methionine starvation [7], indicating that dSAMTOR plays a similar regulatory role in dTORC1 signaling as SAMTOR does in human cells. Hitherto, the in vivo role of dSAMTOR protein in fly’s systemic pathophysiology has not been investigated.
Therefore, to reveal the importance of tissue-specific dSAMTOR loss to Drosophila health, lifespan and well-being, we have genetically targeted the dSAMTOR cognate gene in all major organ tissues, through employment of the appropriate GAL4 drivers and two UAS-SAMTOR_RNAi strains. Strong dSAMTOR downregulation resulted in embryonic lethality in the majority of dSAMTOR-targeted tissues, suggesting that threshold levels of dSAMTOR activity are essential for diverse cell-type functionalities, and especially for normal neuromuscular development and performance. Interestingly, moderate downregulation of dSAMTOR gene in the neuromuscular system is not detrimental and is characterized by distinct phenotypes of sex- and age-dependent pathologies, with predominantly negative impact on Drosophila health and lifespan. Disruption of the SAMTOR-containing complex leads to inactivation of GATOR1, and subsequent activation of mTORC1 and, thus, suppression of autophagy [7]. It may also be the accumulation of SAM metabolite, presumably induced by the lack of SAM-binding partner, SAMTOR, that likely contributes to dSAMTOR loss-derived pathology in fly’s tissue systems. The activation of dTORC1, in dSAMTOR-targeted environments, is strongly supported by the increased phosphorylation contents of (d)p70S6K (a bona fide dTORC1 substrate) and dTOR kinases herein identified by our active kinome profiling in fly brain (Figure 5). The mTORC1 complex acts as a central regulator of diverse cellular functions, committed to promote anabolism and cell growth, and its aberrant over-activation is often detected in up to 80% of human cancers [20,21,22]. mTORC1 exerts neuronal-specific activities during brain development, shaping both the signaling and physical landscapes of the brain, with these activities temporally spanning every stage of brain development [5,23]. Most importantly, mTORC1 suppresses the catabolic pathway, through inhibition of autophagy and lysosome biogenesis [5]. Hyperactive mTOR signaling in the brain is associated with characteristic lesions, such as focal cortical dysplasia, seizures, macrocephaly and benign brain tumors [5]. In mutant mice that are unable to suppress mTORC1 signaling and, therefore, fail to limit their energy expenditure or activate autophagy to supply tissue cytoplasms with free amino acids, perinatal lethality occurs, thus demonstrating that mTORC1 activity must be tightly controlled to maintain -normal- cellular homeostasis, in vivo [5,24,25]. Furthermore, failure in autophagic clearance in the brain results in progressive and permanent destruction of neurons, causing impairment in cognition and motor control, while inhibition of autophagy has emerged as a hallmark of neurotoxic cell death [5,26,27].
Regarding the SAM metabolite, it has long been established that it serves as a universal donor of methyl groups, for almost all methyl-transfer reactions in vivo, to produce a methylated substrate and SAH (S-Adenosyl-L-Homocysteine) moiety [18,28]. In a SAMTOR-deficient background, cellular SAM levels increase and the elevated SAM:SAH ratio gives rise to imbalanced cellular methylation potential, resulting in aberrant increase in the methylation load of proteins, nucleic acids and metabolites [29]. These types of aberrant methylation events seem to critically contribute to diverse cellular pathologies, with changes in methylation patterns of histones and nucleic acids decisively determining the epigenetic states of cells that are mechanistically associated with global alterations in gene expression profiles [18,30]. Although alterations in the methylation status do not likely account for all the, herein observed, detrimental effects in fly’s physiology, the functional synergism of hypermethylation with the high energy levels required by dTORC1 hyperactivation (to promote anabolism and cell growth), together with the accumulation of misfolded or excessive bio-macro-molecules, due to (dTORC1-dependent) suppression of autophagy (which could supply cells with energy and building blocks for proliferation), no longer seems to be compatible with early development in a dSAMTOR-deficient genetic background.
The compromised health and reduced lifespan profiles, herein described for the first time, in response to tissue-specific moderate silencing of dSAMTOR gene may result either from moderate dTORC1-complex activation or moderate SAM-metabolite accumulation. Interestingly, GNMT (Glycine N-Methyltransferase: uses SAM -as methyl donor- to produce N-Methyl-Glycine) downregulation (via RNAi-specific targeting) or SAMS (SAM Synthase) over-expression could not affect lifespan, thereby indicating that moderate increases in SAM cellular contents cannot cause baneful outcomes in fly’s longevity [31]. In contrast, (m)TORC1 suppression has been shown to extend lifespan in different organisms, including Drosophila [32], strongly suggesting the dTORC1 capacity to tightly regulate fly’s longevity and lifespan [33]. Of note, dietary Methionine restriction has proved to inhibit (m)TORC1 and extend lifespan, in both rodents and flies [13,34,35]. Most importantly, genetic or pharmacological manipulation of SAM metabolism suffices for expansion of Drosophila lifespan [31,36], thus implying that SAM, but not Methionine per se, is critical for tissue and organism homeostasis, in vivo. The mechanism via which suppression of mTORC1 leads to lifespan extension includes, likely among others, the induction of autophagy [15]. The functional regulation of mTORC1 is of major importance in defining the morphology of developing brain, by promoting both the building of new proteins at some synapses and the pruning of obsolete synapses through autophagy at others [5]. Hence, the different tissue-specific phenotypes, herein observed, following moderate downregulation of dSAMTOR gene, may be attributed to distinct degrees of balance between inhibition or activation of dTORC1 signaling, in different tissue, gender and age fly settings. Taken together, the quantitative regulation of (d)SAMTOR and SAM levels in the cytoplasm may act as a key factor in lifespan regulation. Their quantitative control is likely mediated by the tissue-dependent induction or suppression of (d)TORC1 signaling activities, which become presumably more complex at the multi-tissue and organism levels.
Methionine administration and Betaine-induced elevation of Methionine cellular contents in dSAMTOR-targeted flies, specifically in their neuronal tissues, resulted in reduced viability profiles. A high proportion of dietary protein or amino acid imbalance has been shown to reduce lifespan in Drosophila [37]. Furthermore, Methionine, when administered at high (effective) levels, acts as toxic compound and leads to decreased longevity [38]. Regarding Drosophila, the toxic level of Methionine has proved to be 10 mM, for female flies. Nevertheless, it must be the imbalance caused by the excess of Methionine over other amino acids that leads to longevity reduction [34]. Remarkably, in our in vivo experimental setting, although Methionine supplementation (5 mM in the food) remains far below the toxic dose (10 mM), the dSAMTOR (moderately) deficient background seems to produce novel complexity levels to the functional interaction of Methionine with other critical amino acids. This new metabolic sub-routine, among others, could cause pathogenic accumulations of unutilized amino acids, due to the inability of dTORC1 to productively sense Methionine actual levels, thereby resulting in severe imbalances in protein synthesis and in detrimentally decreased lifespan. SAMTOR serves as the major mTORC1 sensor for SAM, which directly reflects Methionine reservoir, in tissue cytoplasms. Therefore, in the absence, or even modest downregulation, of (d)SAMTOR protein, SAM, and subsequently Methionine, cannot be efficiently sensed and properly quantified by (d)TORC1 and its interacting components, leading to excessive Methionine- and SAM-derived disequilibrium of amino acid-network metabolism that ultimately compels fly’s organic systems to collapse, in an age-dependent fashion, in vivo. Surprisingly, as demonstrated by (real-time) quantitative PCR, the (mean) reduction of neuronal-targeted (endogenous) dSAMTOR-transcript levels was measured at 23% for the modest downregulation-carrying flies (Figure S2), thereby indicating the importance of Methionine-SAM-dSAMTOR integrity, with even small or moderate decreases in dSAMTOR gene expression (e.g. 23%) strongly compromising and lowering survival and longevity rates in enriched Methionine environments, in vivo. This unique genetic and nutritional synergism of (moderately) reduced dSAMTOR activity and excessive Methionine (or, Betaine) quantity, herein unveiled, paves the way for the development of novel and powerful nutrigenomics (nutritional genomics) platforms able to successfully treat human pathologies, via combined management of both genetic and nutritional abnormal profiles.
Betaine is a methyl derivative of Glycine and participates as methyl donor in re-synthesis of Methionine by Betaine-Homocysteine Methyl-Transferase (BHMT), utilizing Homocysteine as critical substrate [39]. To further explore the role of Methionine-SAM axis in lifespan control, we performed RNAi-mediated downregulation of dBHMT gene, in tissue-specific manner, throughout Drosophila lifetime. Genetic targeting of dBHMT resulted in notably reduced lifespan and slightly impaired climbing activity (indicative of neuromuscular function), during aging, in all tissues examined, a (negative geotaxis) phenotype with comparatively more pronounced pathogenic features in male populations. The only exemption has been observed in neuronal tissue-specific targeted female flies, which are presented with a moderately increased viability profile, especially at older ages. Gender-dependent differences in the nutritional requirements of reproduction in Drosophila have already been reported, and it is possible that dietary control is only successful in one sex, with negative or no effect in the other sex [40,41]. Likewise, Methionine restriction results in different responses in (mean) lifespan between female and male flies [34]. Blockade of Methionine re-synthesis leads to reduced levels of SAM metabolite and, thus, to inhibition of mTORC1 signaling by SAMTOR de-repression. Furthermore, Methionine depletion in human cells causes SAM exhaustion and promotes alterations in histone methylation being mechanistically associated with widespread induction of stress-response pathways, and cell death sub-routines [18,42]. It seems that, although Methionine restriction leads to longevity and metabolic health through suppression of (m)TORC1 activity, which is directly coupled with autophagy induction and improved insulin resistance, in dBHMT-targeted flies, the amount of dietary Methionine intake is not sufficient for cellular homeostasis and physiological methylation events. Potential compensatory adaptations are not able to maintain the essential processes involved in Methionine metabolism, and, therefore, a critical threshold of (d)BHMT-regulated Methionine re-synthesis is necessary for the efficient control of protein synthesis (and methylome architecture), particularly during (fly) aging.
Lifespan represents an overall sum of both positive and negative effects of different signaling pathways related to cell/tissue type, gender, age, nutritional scheme, mutational load, genetic background and reproductive state. Hence, it is difficult to precisely determine the specific role(s) of dSAMTOR-derived signaling in lifespan control. Since the association between SAM and survival/lifespan does not function in a bidirectional fashion, a threshold in SAM levels that regulate organism lifespan is strongly suggested to exist, in vivo. A similar threshold may likely act for dSAMTOR, as well, with loss or strong downregulation of dSAMTOR gene expression resulting in embryonic lethal phenotypes for most targeted tissues (Table 1), and modest reduction of dSAMTOR levels causing comparatively moderate pathologies. Further investigation of the precise molecular mechanisms that describe dSAMTOR critical involvement in lifespan control, neuromuscular performance, tissue development and organ architecture is essential and must be promptly conducted.
4. Materials and Methods
4.1. Fly stocks and culturing conditions (Drosophila husbandry)
Drosophila melanogaster transgenic fly strains were obtained from Bloomington Drosophila Stock Center (NIH P40OD018537) (Indiana, USA): y[1] v[1]; P{y[+t7.7]v[+t1.8]=TRiP.HMJ21611}attP40 (BL: 52944), y[1] v[1]; P{y[+t7.7] v[+t1.8]=TRiP.HMJ21433}attP40 (BL: 54010), y[1] v[1]; P{y[+t7.7]v[+t1.8]=TRiP.HMC02683}attP40 (BL: 43986), w[1118]; P{w[+m*]=GAL4}repo/TM3, Sb[1] (BL: 7415), w[1118]; w[*]; P{w[+mW.hs]=GawB}D42 (BL: 8816), w[*]; P{w[+mC]=GAL4-elav.L}3 (BL: 8760), y[1] w[*]; P{w[+mC]=GAL4-Mef2.R}3 (BL: 27390), w[1118]; P{w[+m*]=GAL4}repo/TM3, Sb[1] (BL: 7415), y[1] w[*]; P{w[+mC]=Act5C-GAL4}25FO1/CyO (BL: 4414), w[1118]; P{w[+mW.hs]=GawB}Bx[MS1096] (BL: 8860), w[*]; P{w[+mC]=GAL4-ninaE.GMR}12 (BL: 1104). The D. melanogaster transgenic NP1-GAL4 fly strain was kindly provided by Eric H. Baehrecke (Department of Cancer Biology, University of Massachusetts Medical School, Worcester, Massachusetts 01605, USA).
All fly stocks were maintained at 25°C, 55-65% relative humidity, on a photoperiod of 12 h light/dark, and a laboratory standard Drosophila medium (6.4% rice flour, 5% tomato paste, 3.2% sugar, 0.8% yeast, 0.8% agar, 0.4% ethanol and 0.4% propionic acid).
4.2. Chemicals and reagents
Methionine (CAS 63-68-3) was provided by Sigma-Aldrich (Missouri, USA) and Betaine (CAS 107-43-7) was obtained from Santa Cruz Biotechnology, Inc. (Heidelberg, Germany). The protein extraction buffer M-PER, Phosphatase Inhibitor Cocktail and Halt Protease Inhibitor Cocktail EDTA-free were purchased from Thermo Fisher Scientific (Darmstadt, Germany). Serine/Threonine Kinase (STK) PamChip and STK reagent kit were provided by PamGene International B.V. (BJ‘s-Hertogenbosch, Netherlands).
4.3. Longevity measurement
Newly emerged (~24 h) female and male flies from each fly cross were collected and kept separate in vials (approximately 20-25 flies per vial). Flies from each vial were transferred into a fresh vial of the same medium twice a week. Survival curves were generated by counting daily deceased flies. For each viability experiment, the sample size was set (at least) at 100 flies per sex and cross (based on the literature), for statistically significant results. Methionine and Betaine were employed to pharmacologically induce dTORC1 activity, and were supplemented to the fly food at concentrations of 5 and 10 mM, respectively. All viability experiments were performed at the same time for control and RNAi-downregulated strains. Experiments were carried out three different times, using independent genetic crosses.
4.4. Negative geotaxis assay
Negative geotaxis (climbing) assay has been established as an in vivo reliable indicator for the estimation of locomotor performance in Drosophila. The climbing assay was performed every 10 days. Flies of both sexes were kept together. Before the experimental procedure, flies were anesthetized and divided into males and females (groups of 20-25 flies). Each experimental group was, then, placed in an empty 100 ml cylinder, with a borderline drawn at the 60 ml mark (10 cm height). To climb (against gravity) simultaneously, flies were gently tapped to the bottom of the cylinder. After time interval of 20 sec, the number of flies that reached or exceeded the 60 ml mark was counted, and five repeats were performed for each group. The same populations were tested at different ages, excluding flies that died or flew away. Control and RNAi-targeted fly groups were examined simultaneously. The total sample size for each fly cross and gender was set (at least) at 100 flies. Experiments were performed three different times, using independent genetic crosses.
4.5. Statistical analysis
The Statistical Package for Social Sciences (IBM SPSS v23.0 for Windows IBM Corp., New York, USA) was used for statistical analysis of the results. Data from longevity experiments were analyzed with the Kaplan-Meier survival test, using log rank and Breslow test statistics. Climbing graphs were drawn, as an average pass rate per genotype/time-point with Sample Standard Deviation (± SSD) value. Differences between compared genotypes were evaluated by the independent t-test analysis. Significance was accepted at p < 0.05 (*), p < 0.01 (**) and p < 0.001 (***).
4.6. Light microscopy (LM)
D. melanogaster wings were visualized using Nikon Digital Eclipse C1 microscope (Nikon; Tokyo, Japan). Every 10 days, for a period of 50 days, flies were anaesthetized with CO2, and their wings were carefully dissected and mounted with DePeX (Serva Electrophoresis GmbH, Heidelberg, Germany) onto glass microscope slides. Morphology and structure of the wings were observed separately for each gender. Wing surface area was measured using the ImageJ software.
4.7. Scanning electron microscopy (SEM)
Surface structural architecture of D. melanogaster compound eyes was visualized through a Phillips 515 scanning electron microscope. Female and male transgenic flies, at the age of 10, 30 and 50 days, were collected and immediately attached to aluminum stubs. Next, flies were air-dried at room temperature, and coated with gold-palladium (60–40%) on Tousimis Samsputter 2a.
4.8. Kinase activity profiling
The PamStation platform and STK (Serine/Threonine: Ser/Thr) PamChip peptide arrays, manufactured by PamGene International B.V. (‘s-Hertogenbosch, The Netherlands), were used. A typical STK PamChip contains 4 (micro-)arrays with 140 Ser/Thr peptides and 4 positive control immobilized peptides each that are covalently attached to a porous material. The peptides harbor phosphorylation sites derived from literature or computational predictions, and are correlated with one or multiple upstream kinases. Kinases in the specimens actively phosphorylate substrates on the PamChip, in the presence of ATP. Phosphorylation is detected by a (phospho-)specific primary antibody and the signal is quantified by a second FITC-conjugated antibody. Protein lysates were prepared from frozen control and RNAi-downregulated Drosophila heads, with M-PER (cell-lysing) buffer being enriched with 1:100 Phosphatase Inhibitor Cocktail and 1:100 Halt Protease Inhibitor Cocktail EDTA-free, and quantified by Bradford assay. 1 µg of total protein from each biological sample was used for Ser/Thr kinase activity profiling, according to the standard protocol provided by the manufacturer. The STK basic mix was composed of the protein lysate, 4 µL 10x PK, 0.4 µL of 100x BSA, 4.0 µL of 4 mM ATP and 0.5 µL STK antibody mix. Total volume of the STK basic mix was adjusted to 40 µL by adding distilled water. The detection mix consisted of 3 µL of 10x antibody buffer (AB), 0.4 µL STK antibody FITC-labeled and 26.6 µL distilled water. After a pre-processing step of 30 cycles (blocking of peptide micro-arrays with 2% BSA), the STK basic mix was applied to each PamChip micro-array. Next, micro-arrays were incubated and washed for 60 cycles (30-90c). Subsequently, the detection mix was added onto the PamChips, and incubation with FITC-labeled STK antibody and image recording by a CCD camera (after 10, 50 and 200 msec) were operated for the next 30 cycles, and a final read was carried out after the end-cycle 124. Data normalization and visualization were performed using the BioNavigator Analysis Software Tool (BNAST).
4.9. RNA extraction and RT-qPCR
Total RNA was isolated from dSAMTOR-targeted fly heads (manually dissected), using TRIzol™ monophasic solution of Phenol and Guanidine Iso-thio-cyanate (Invitrogen, Thermo Fischer Scientific, Waltham, MA, USA), according to manufacturer’s instructions. Concentration and quality of isolated RNA were determined by NanoDrop One UV-Vis Spectro-photometer (Thermo Fischer Scientific, Waltham, MA, USA). RNA samples were reverse transcribed into complementary DNA (cDNA), using SuperScript™ IV First-Strand Synthesis System (Invitrogen, Thermo Fischer Scientific, Waltham, MA, USA), following manufacturer’s protocol.
Relative expression of dSAMTOR gene was investigated by Reverse Transcription, real-time, quantitative Polymerase Chain Reaction (RT-qPCR), using dSAMTOR-specific oligonucleotide TaqMan™ probes (Dm01831401_s1, Thermo Fischer Scientific, Waltham, MA, USA), the TaqMan™ Universal PCR Master Mix (Applied Biosystems, Thermo Fischer Scientific, Waltham, MA, USA) and the Applied Biosystems StepOne™ real-time PCR System (Thermo Fischer Scientific, Waltham, MA, USA), according to manufacturer's guidelines. As internal control for normalization of gene-expression data, the housekeeping gene Actin 5C (Dm02361909_s1, Thermo Fischer Scientific, Waltham, MA, USA) was accordingly used. To ensure reproducibility, each assay was performed with technical triplicates, while three negative controls were also included in the analysis. Relative mRNA levels were calculated using the comparative 2−ΔΔCt method [43] that calculates changes in gene expression as a relative fold difference between the gene of interest (dSAMTOR) and the reference gene (Actin 5C). Results were presented as a percentage of the relative gene reduction in dSAMTOR-targeted (specifically in neuronal tissues) flies compared to control populations.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Moderate silencing of dSAMTOR gene, in all Drosophila tissues, does not affect life expectancy; Figure S2: Relative change in dSAMTOR gene expression, via RT-qPCR technology engagement.
Author Contributions
Conceptualization, Dimitrios Stravopodis; Data curation, Athanassios Velentzas and Dimitrios Stravopodis; Funding acquisition, Dimitrios Stravopodis; Methodology, Stamatia Katarachia and Sophia Markaki; Project administration, Dimitrios Stravopodis; Resources, Dimitrios Stravopodis; Supervision, Athanassios Velentzas and Dimitrios Stravopodis; Validation, Stamatia Katarachia, Sophia Markaki and Athanassios Velentzas; Visualization, Dimitrios Stravopodis; Writing – original draft, Stamatia Katarachia and Athanassios Velentzas; Writing – review & editing, Dimitrios Stravopodis.
Data Availability Statement
All data are contained within the article, or supplementary material.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sabatini, D.M. , Twenty-five years of mTOR: Uncovering the link from nutrients to growth. Proc Natl Acad Sci U S A 2017, 114, 11818–11825. [Google Scholar] [CrossRef] [PubMed]
- Saxton, R.A.; Sabatini, D.M. , mTOR Signaling in Growth, Metabolism, and Disease. Cell 2017, 168, 960–976. [Google Scholar] [CrossRef] [PubMed]
- Laplante, M.; Sabatini, D.M. , mTOR signaling in growth control and disease. Cell 2012, 149, 274–293. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.C.; Guan, K.L. , mTOR: a pharmacologic target for autophagy regulation. J Clin Invest 2015, 125, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.Y.; Sabatini, D.M. , mTOR at the nexus of nutrition, growth, ageing and disease. Nat Rev Mol Cell Biol 2020, 21, 183–203. [Google Scholar] [CrossRef] [PubMed]
- Deleyto-Seldas, N.; Efeyan, A. , The mTOR-Autophagy Axis and the Control of Metabolism. Front Cell Dev Biol 2021, 9, 655731. [Google Scholar] [CrossRef]
- Gu, X.; Orozco, J.M.; Saxton, R.A.; Condon, K.J.; Liu, G.Y.; Krawczyk, P.A.; Scaria, S.M.; Harper, J.W.; Gygi, S.P.; Sabatini, D.M. , SAMTOR is an S-adenosylmethionine sensor for the mTORC1 pathway. Science 2017, 358, 813–818. [Google Scholar] [CrossRef]
- Hesketh, G.G.; Papazotos, F.; Pawling, J.; Rajendran, D.; Knight, J.D.R.; Martinez, S.; Taipale, M.; Schramek, D.; Dennis, J.W.; Gingras, A.C. , The GATOR-Rag GTPase pathway inhibits mTORC1 activation by lysosome-derived amino acids. Science 2020, 370, 351–356. [Google Scholar] [CrossRef]
- Bar-Peled, L.; Sabatini, D.M. , Regulation of mTORC1 by amino acids. Trends Cell Biol 2014, 24, 400–406. [Google Scholar] [CrossRef]
- Anandapadamanaban, M.; Masson, G.R.; Perisic, O.; Berndt, A.; Kaufman, J.; Johnson, C.M.; Santhanam, B.; Rogala, K.B.; Sabatini, D.M.; Williams, R.L. , Architecture of human Rag GTPase heterodimers and their complex with mTORC1. Science 2019, 366, 203–210. [Google Scholar] [CrossRef]
- Wolfson, R.L.; Chantranupong, L.; Saxton, R.A.; Shen, K.; Scaria, S.M.; Cantor, J.R.; Sabatini, D.M. , Sestrin2 is a leucine sensor for the mTORC1 pathway. Science 2016, 351, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Wolfson, R.L.; Sabatini, D.M. , The Dawn of the Age of Amino Acid Sensors for the mTORC1 Pathway. Cell Metab 2017, 26, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Kitada, M.; Xu, J.; Ogura, Y.; Monno, I.; Koya, D. , Mechanism of Activation of Mechanistic Target of Rapamycin Complex 1 by Methionine. Front Cell Dev Biol 2020, 8, 715. [Google Scholar] [CrossRef] [PubMed]
- Coolon, J.D.; Stevenson, K.R.; McManus, C.J.; Graveley, B.R.; Wittkopp, P.J. , Genomic imprinting absent in Drosophila melanogaster adult females. Cell Rep 2012, 2, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Guan, K.L. , mTOR as a central hub of nutrient signalling and cell growth. Nat Cell Biol 2019, 21, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Lauinger, L.; Kaiser, P. , Sensing and Signaling of Methionine Metabolism. Metabolites.
- Law, J.A.; Jacobsen, S.E. , Establishing, maintaining and modifying DNA methylation patterns in plants and animals. Nat Rev Genet 2010, 11, 204–220. [Google Scholar] [CrossRef] [PubMed]
- Mentch, S.J.; Mehrmohamadi, M.; Huang, L.; Liu, X.; Gupta, D.; Mattocks, D.; Gomez Padilla, P.; Ables, G.; Bamman, M.M.; Thalacker-Mercer, A.E.; Nichenametla, S.N.; Locasale, J.W. , Histone Methylation Dynamics and Gene Regulation Occur through the Sensing of One-Carbon Metabolism. Cell Metab 2015, 22, 861–873. [Google Scholar] [CrossRef]
- Locasale, J.W. , Serine, glycine and one-carbon units: cancer metabolism in full circle. Nat Rev Cancer 2013, 13, 572–583. [Google Scholar] [CrossRef]
- Mossmann, D.; Park, S.; Hall, M.N. , mTOR signalling and cellular metabolism are mutual determinants in cancer. Nat Rev Cancer 2018, 18, 744–757. [Google Scholar] [CrossRef]
- Zeng, J.D.; Wu, W.K.K.; Wang, H.Y.; Li, X.X. , Serine and one-carbon metabolism, a bridge that links mTOR signaling and DNA methylation in cancer. Pharmacol Res 2019, 149, 104352. [Google Scholar] [CrossRef]
- Menon, S.; Manning, B.D. , Common corruption of the mTOR signaling network in human tumors. Oncogene 2008, (0 2), S43–51. [Google Scholar] [CrossRef]
- Lipton, J.O.; Sahin, M. , The neurology of mTOR. Neuron 2014, 84, 275–291. [Google Scholar] [CrossRef] [PubMed]
- Efeyan, A.; Zoncu, R.; Chang, S.; Gumper, I.; Snitkin, H.; Wolfson, R.L.; Kirak, O.; Sabatini, D.D.; Sabatini, D.M. , Regulation of mTORC1 by the Rag GTPases is necessary for neonatal autophagy and survival. Nature 2013, 493, 679–683. [Google Scholar] [CrossRef] [PubMed]
- Peng, M.; Yin, N.; Li, M.O. , Sestrins function as guanine nucleotide dissociation inhibitors for Rag GTPases to control mTORC1 signaling. Cell 2014, 159, 122–133. [Google Scholar] [CrossRef] [PubMed]
- Nixon, R.A. , The role of autophagy in neurodegenerative disease. Nat Med 2013, 19, 983–997. [Google Scholar] [CrossRef] [PubMed]
- Hara, T.; Nakamura, K.; Matsui, M.; Yamamoto, A.; Nakahara, Y.; Suzuki-Migishima, R.; Yokoyama, M.; Mishima, K.; Saito, I.; Okano, H.; Mizushima, N. , Suppression of basal autophagy in neural cells causes neurodegenerative disease in mice. Nature 2006, 441, 885–889. [Google Scholar] [CrossRef] [PubMed]
- Finkelstein, J.D. , Methionine metabolism in mammals. J Nutr Biochem 1990, 1, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Ye, C.; Sutter, B.M.; Wang, Y.; Kuang, Z.; Tu, B.P. , A Metabolic Function for Phospholipid and Histone Methylation. Mol Cell 2017, 66, 180–193. [Google Scholar] [CrossRef]
- Barth, T.K.; Imhof, A. , Fast signals and slow marks: the dynamics of histone modifications. Trends Biochem. Sci. 2010, 35, 618–626. [Google Scholar] [CrossRef]
- Obata, F.; Miura, M. , Enhancing S-adenosyl-methionine catabolism extends Drosophila lifespan. Nature communications 2015, 6, 8332. [Google Scholar] [CrossRef]
- Bjedov, I.; Toivonen, J.M.; Kerr, F.; Slack, C.; Jacobson, J.; Foley, A.; Partridge, L. , Mechanisms of life span extension by rapamycin in the fruit fly Drosophila melanogaster. Cell Metab 2010, 11, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Kitada, M.; Ogura, Y.; Monno, I.; Koya, D. , The impact of dietary protein intake on longevity and metabolic health. EBioMedicine 2019, 43, 632–640. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.C.; Kaya, A.; Ma, S.; Kim, G.; Gerashchenko, M.V.; Yim, S.H.; Hu, Z.; Harshman, L.G.; Gladyshev, V.N. , Methionine restriction extends lifespan of Drosophila melanogaster under conditions of low amino-acid status. Nature communications 2014, 5, 3592. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.A.; Buehner, G.; Chang, Y.; Harper, J.M.; Sigler, R.; Smith-Wheelock, M. , Methionine-deficient diet extends mouse lifespan, slows immune and lens aging, alters glucose, T4, IGF-I and insulin levels, and increases hepatocyte MIF levels and stress resistance. Aging Cell 2005, 4, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Parkhitko, A.A.; Binari, R.; Zhang, N.; Asara, J.M.; Demontis, F.; Perrimon, N. , Tissue-specific down-regulation of S-adenosyl-homocysteine via suppression of dAhcyL1/dAhcyL2 extends health span and life span in Drosophila. Genes Dev 2016, 30, 1409–1422. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.P.; Simpson, S.J.; Clissold, F.J.; Brooks, R.; Ballard, J.W.; Taylor, P.W.; Soran, N.; Raubenheimer, D. , Lifespan and reproduction in Drosophila: New insights from nutritional geometry. Proc Natl Acad Sci U S A 2008, 105, 2498–2503. [Google Scholar] [CrossRef] [PubMed]
- Harper, A.E.; Benevenga, N.J.; Wohlhueter, R.M. , Effects of ingestion of disproportionate amounts of amino acids. Physiol Rev 1970, 50, 428–558. [Google Scholar] [CrossRef]
- Pajares, M.A.; Perez-Sala, D. , Betaine homocysteine S-methyltransferase: just a regulator of homocysteine metabolism? Cell Mol Life Sci 2006, 63, 2792–2803. [Google Scholar] [CrossRef]
- Jensen, K.; McClure, C.; Priest, N.K.; Hunt, J. , Sex-specific effects of protein and carbohydrate intake on reproduction but not lifespan in Drosophila melanogaster. Aging Cell 2015, 14, 605–615. [Google Scholar] [CrossRef]
- Zajitschek, F.; Zajitschek, S.R.; Friberg, U.; Maklakov, A.A. , Interactive effects of sex, social environment, dietary restriction, and methionine on survival and reproduction in fruit flies. Age (Dordr) 2013, 35, 1193–1204. [Google Scholar] [CrossRef]
- Shiraki, N.; Shiraki, Y.; Tsuyama, T.; Obata, F.; Miura, M.; Nagae, G.; Aburatani, H.; Kume, K.; Endo, F.; Kume, S. , Methionine metabolism regulates maintenance and differentiation of human pluripotent stem cells. Cell Metab 2014, 19, 780–794. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. , Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Strong evolutionary conservation of SAMTOR protein among species. Amino acid sequence alignment among human (Homo sapiens), mouse (Mus musculus), zebrafish (Danio rerio) and fly (Drosophila melanogaster) SAMTOR proteins, via employment of the Clustal Omega bioinformatics tool.
Figure 1.
Strong evolutionary conservation of SAMTOR protein among species. Amino acid sequence alignment among human (Homo sapiens), mouse (Mus musculus), zebrafish (Danio rerio) and fly (Drosophila melanogaster) SAMTOR proteins, via employment of the Clustal Omega bioinformatics tool.
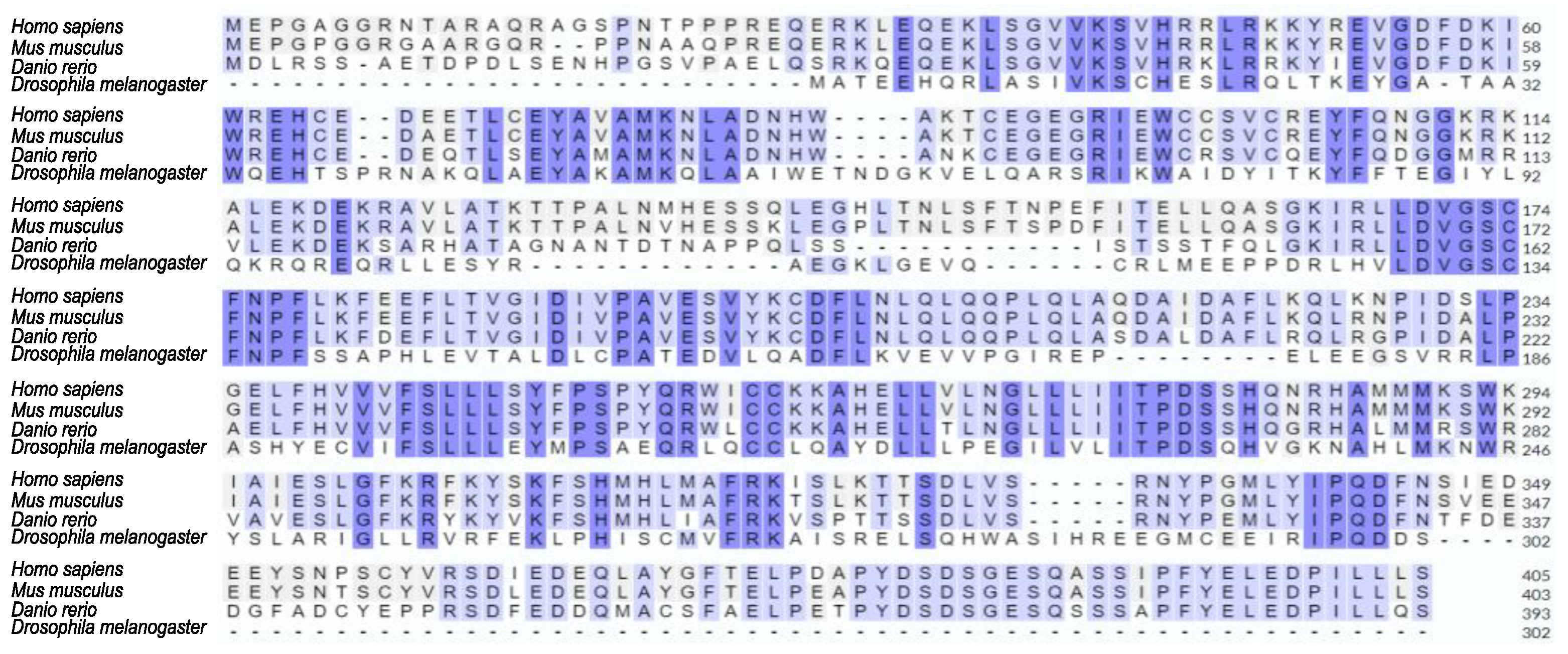
Figure 2.
Neuronal tissue- and glial cell-specific mild proficiency, targeting of dSAMTOR gene. Curves (A, B) and bar charts (C, D) showing the survival rates and (%) climbing capacities, respectively, of female (left panel) and male (right panel) transgenic flies being characterized by RNAi-mediated modest targeting of the dSAMTOR gene, specifically in neuronal tissues (elav.L>SAMTOR_RNAi(M)) (red lines/bars), as compared to control fly populations (elav.L-GAL4/+) (green lines/bars). Curves (E, F) and bar charts (G, H) presenting longevity profiles and (%) climbing patterns, respectively, of female (left panel) and male (right panel) flies with moderate downregulation of the dSAMTOR gene, specifically in glial cells (repo>SAMTOR_RNAi(M)) (red lines/bars), as compared to control flies (repo-GAL4/+) (green lines/bars). *p < 0.05, **p < 0.01 and ***p < 0.001.
Figure 2.
Neuronal tissue- and glial cell-specific mild proficiency, targeting of dSAMTOR gene. Curves (A, B) and bar charts (C, D) showing the survival rates and (%) climbing capacities, respectively, of female (left panel) and male (right panel) transgenic flies being characterized by RNAi-mediated modest targeting of the dSAMTOR gene, specifically in neuronal tissues (elav.L>SAMTOR_RNAi(M)) (red lines/bars), as compared to control fly populations (elav.L-GAL4/+) (green lines/bars). Curves (E, F) and bar charts (G, H) presenting longevity profiles and (%) climbing patterns, respectively, of female (left panel) and male (right panel) flies with moderate downregulation of the dSAMTOR gene, specifically in glial cells (repo>SAMTOR_RNAi(M)) (red lines/bars), as compared to control flies (repo-GAL4/+) (green lines/bars). *p < 0.05, **p < 0.01 and ***p < 0.001.
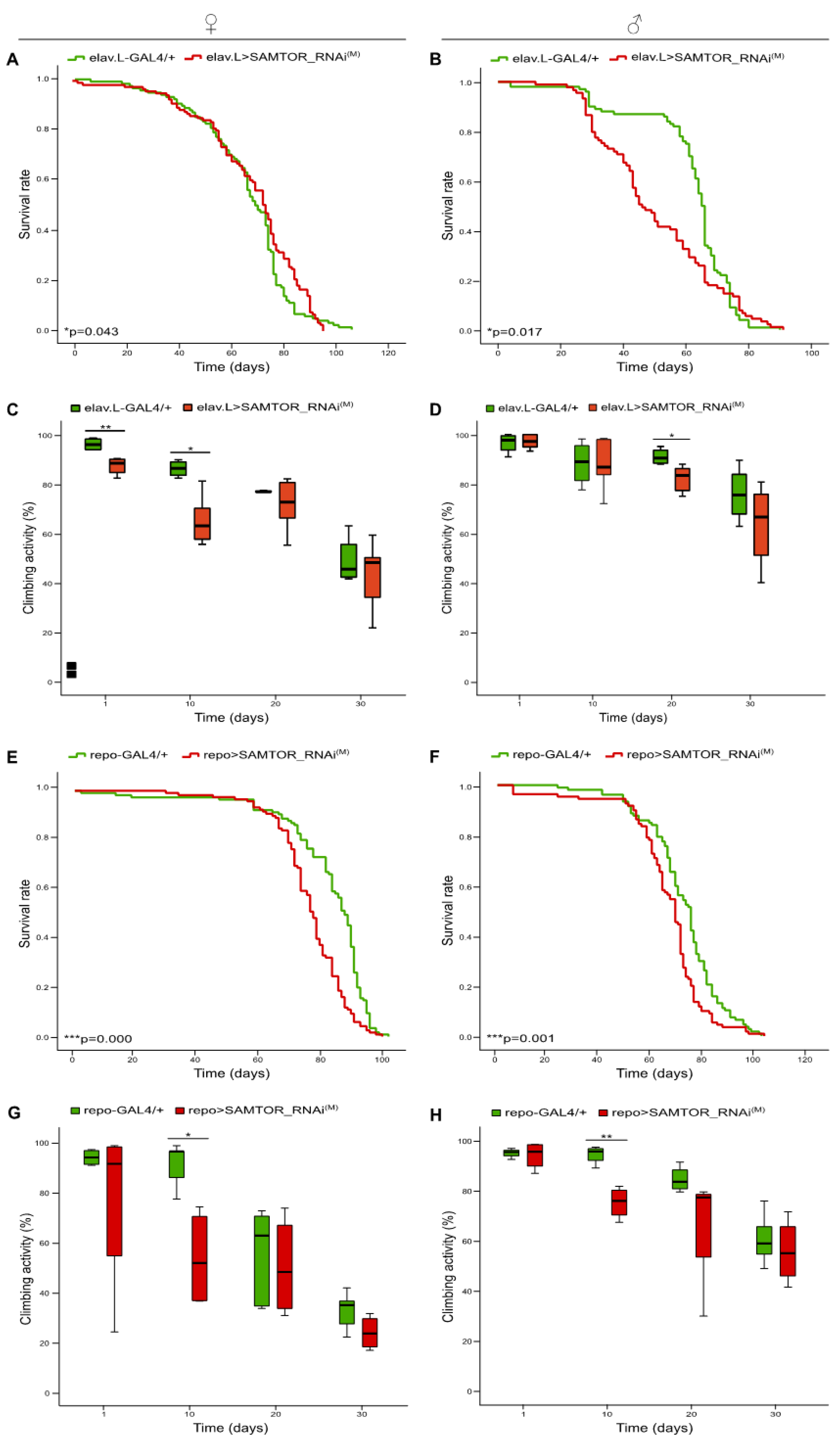
Figure 3.
dSAMTOR modest suppression in motor neurons and muscles. Lifespan curves (A, B) and (%) climbing bar charts (C, D) of female (left panel) and male (right panel) transgenic flies, carrying moderately downregulated dSAMTOR gene expression, specifically in motor neurons (D42>SAMTOR_RNAi(M)) (red lines/bars), as compared to control fly populations (D42-GAL4/+) (green lines/bars). Lifespan profiles (E, F) and climbing activity patterns (G, H) of female (left panel) and male (right panel) flies with mild downregulation of dSAMTOR gene activity, specifically in muscles (mef2>SAMTOR_RNAi(M)) (red lines/bars), as compared to control flies (mef2-GAL4/+) (green lines/bars). *p < 0.05, **p < 0.01 and ***p < 0.001.
Figure 3.
dSAMTOR modest suppression in motor neurons and muscles. Lifespan curves (A, B) and (%) climbing bar charts (C, D) of female (left panel) and male (right panel) transgenic flies, carrying moderately downregulated dSAMTOR gene expression, specifically in motor neurons (D42>SAMTOR_RNAi(M)) (red lines/bars), as compared to control fly populations (D42-GAL4/+) (green lines/bars). Lifespan profiles (E, F) and climbing activity patterns (G, H) of female (left panel) and male (right panel) flies with mild downregulation of dSAMTOR gene activity, specifically in muscles (mef2>SAMTOR_RNAi(M)) (red lines/bars), as compared to control flies (mef2-GAL4/+) (green lines/bars). *p < 0.05, **p < 0.01 and ***p < 0.001.
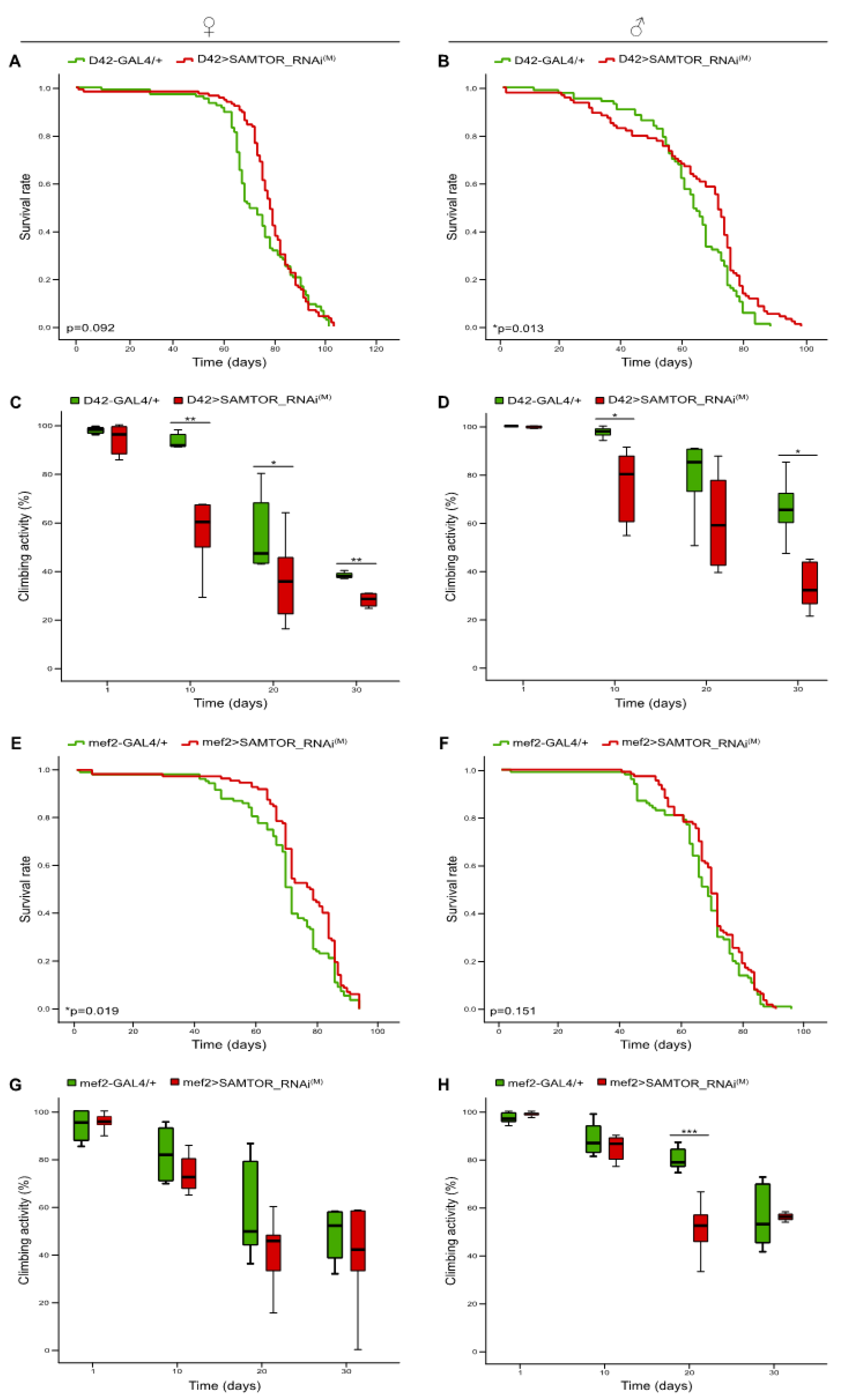
Figure 4.
Differential downregulation of dSAMTOR gene expression in midgut tissues. Lifespan profiles (A, B) and bar charts showing (%) kinetic activities (C, D) of female (left panel) and male (right panel) transgenic flies, with moderate dSAMTOR silencing capacity, specifically in midgut tissues (NP1>SAMTOR_RNAi(M)) (red lines/bars), as compared to control fly populations (NP1-GAL4/+) (green lines/bars). Curves (E, F) presenting the survival rates of female (left panel) and male (right panel) transgenic flies, with strong RNAi-mediated targeting of dSAMTOR gene, specifically in midgut tissues (NP1>SAMTOR_RNAi(S)) (red lines/bars), as compared to control flies (NP1-GAL4/+) (green lines/bars). SAMTOR_RNAi(M): RNAi strain with moderate (M) dSAMTOR silencing capacity. SAMTOR_RNAi(S): RNAi strain with strong (S) dSAMTOR silencing potency. **p < 0.01 and ***p < 0.001.
Figure 4.
Differential downregulation of dSAMTOR gene expression in midgut tissues. Lifespan profiles (A, B) and bar charts showing (%) kinetic activities (C, D) of female (left panel) and male (right panel) transgenic flies, with moderate dSAMTOR silencing capacity, specifically in midgut tissues (NP1>SAMTOR_RNAi(M)) (red lines/bars), as compared to control fly populations (NP1-GAL4/+) (green lines/bars). Curves (E, F) presenting the survival rates of female (left panel) and male (right panel) transgenic flies, with strong RNAi-mediated targeting of dSAMTOR gene, specifically in midgut tissues (NP1>SAMTOR_RNAi(S)) (red lines/bars), as compared to control flies (NP1-GAL4/+) (green lines/bars). SAMTOR_RNAi(M): RNAi strain with moderate (M) dSAMTOR silencing capacity. SAMTOR_RNAi(S): RNAi strain with strong (S) dSAMTOR silencing potency. **p < 0.01 and ***p < 0.001.
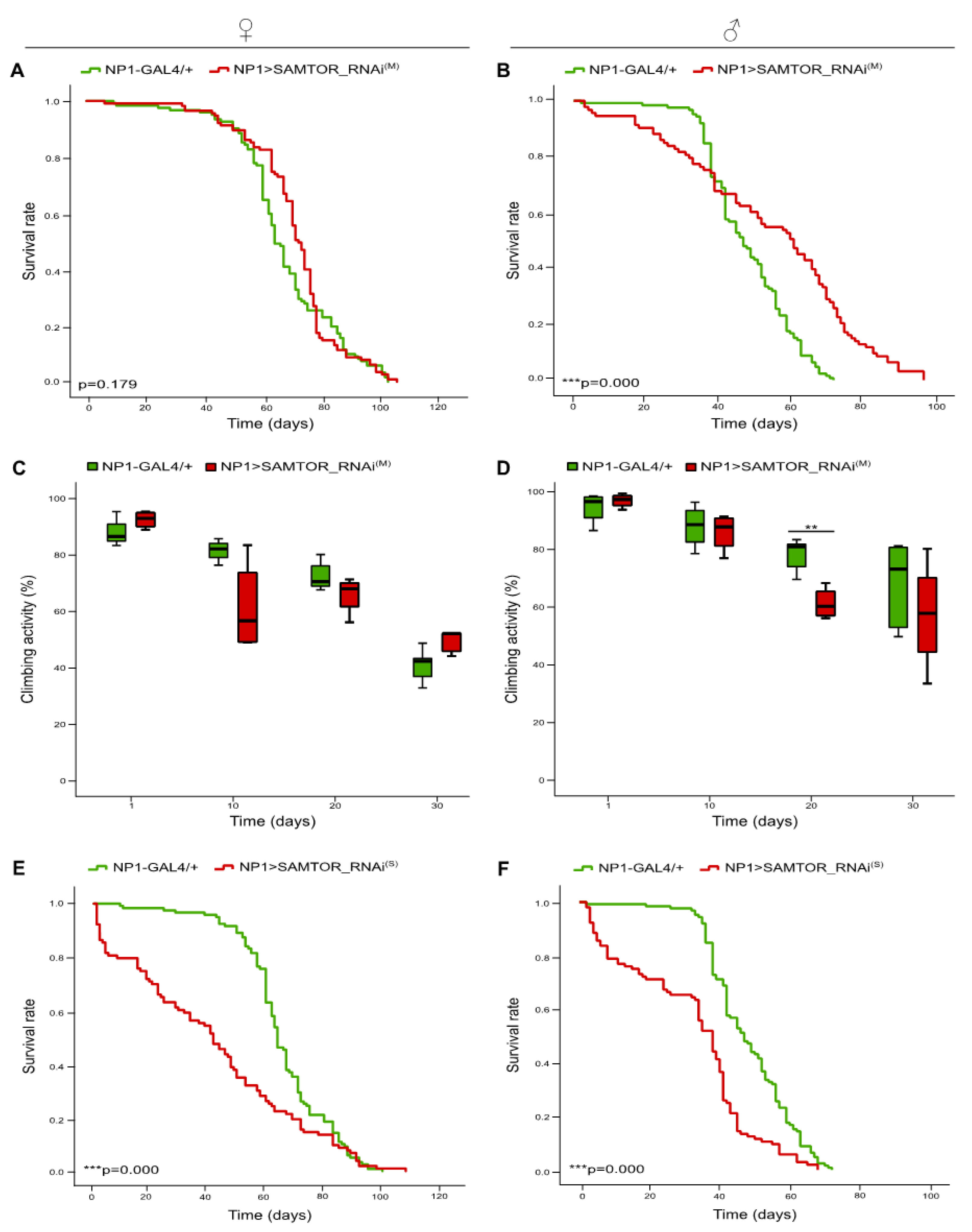
Figure 5.
Active kinome analysis of dSAMTOR-targeted D. melanogaster brains. (A) CCD images of PamChip arrays. Differential substrate phosphorylation comparing transgenic flies characterized by RNAi-mediated, modestly efficiency, targeting of the dSAMTOR gene, specifically in neuronal tissues (elav.L>SAMTOR_RNAi(M)) and control flies (elav.L-GAL4/+), showing representative Serine/Threonine (Ser/Thr) kinase activities. (B) Heatmaps visualizing Ser/Thr-peptide phosphorylation intensities (log2) of control and dSAMTOR-downregulated flies, specifically in fly brain (neuronal system). (C) Median Final Score Plots of the Ser/Thr kinase activity profiling in dSAMTOR-targeted fly brain (neuronal system) setting. Normalized kinase activity statistics is a mathematical-based algorithm indicating the estimated relative kinase activity, while the specificity score reflects the reliability and accuracy of the prediction. In particular, the x-axis indicates the values for the normalized kinase activity statistics (e.g., negative value [s < 0] = “the activity of the corresponding Ser/Thr kinase is decreased”). The specificity score (Qsp) is indicated by the colour of the points. Qsp logarithmic values > 1.3 (white [Qsp = 1.3] to red colour [Qsp ≥ 2]) were considered as statistically relevant.
Figure 5.
Active kinome analysis of dSAMTOR-targeted D. melanogaster brains. (A) CCD images of PamChip arrays. Differential substrate phosphorylation comparing transgenic flies characterized by RNAi-mediated, modestly efficiency, targeting of the dSAMTOR gene, specifically in neuronal tissues (elav.L>SAMTOR_RNAi(M)) and control flies (elav.L-GAL4/+), showing representative Serine/Threonine (Ser/Thr) kinase activities. (B) Heatmaps visualizing Ser/Thr-peptide phosphorylation intensities (log2) of control and dSAMTOR-downregulated flies, specifically in fly brain (neuronal system). (C) Median Final Score Plots of the Ser/Thr kinase activity profiling in dSAMTOR-targeted fly brain (neuronal system) setting. Normalized kinase activity statistics is a mathematical-based algorithm indicating the estimated relative kinase activity, while the specificity score reflects the reliability and accuracy of the prediction. In particular, the x-axis indicates the values for the normalized kinase activity statistics (e.g., negative value [s < 0] = “the activity of the corresponding Ser/Thr kinase is decreased”). The specificity score (Qsp) is indicated by the colour of the points. Qsp logarithmic values > 1.3 (white [Qsp = 1.3] to red colour [Qsp ≥ 2]) were considered as statistically relevant.
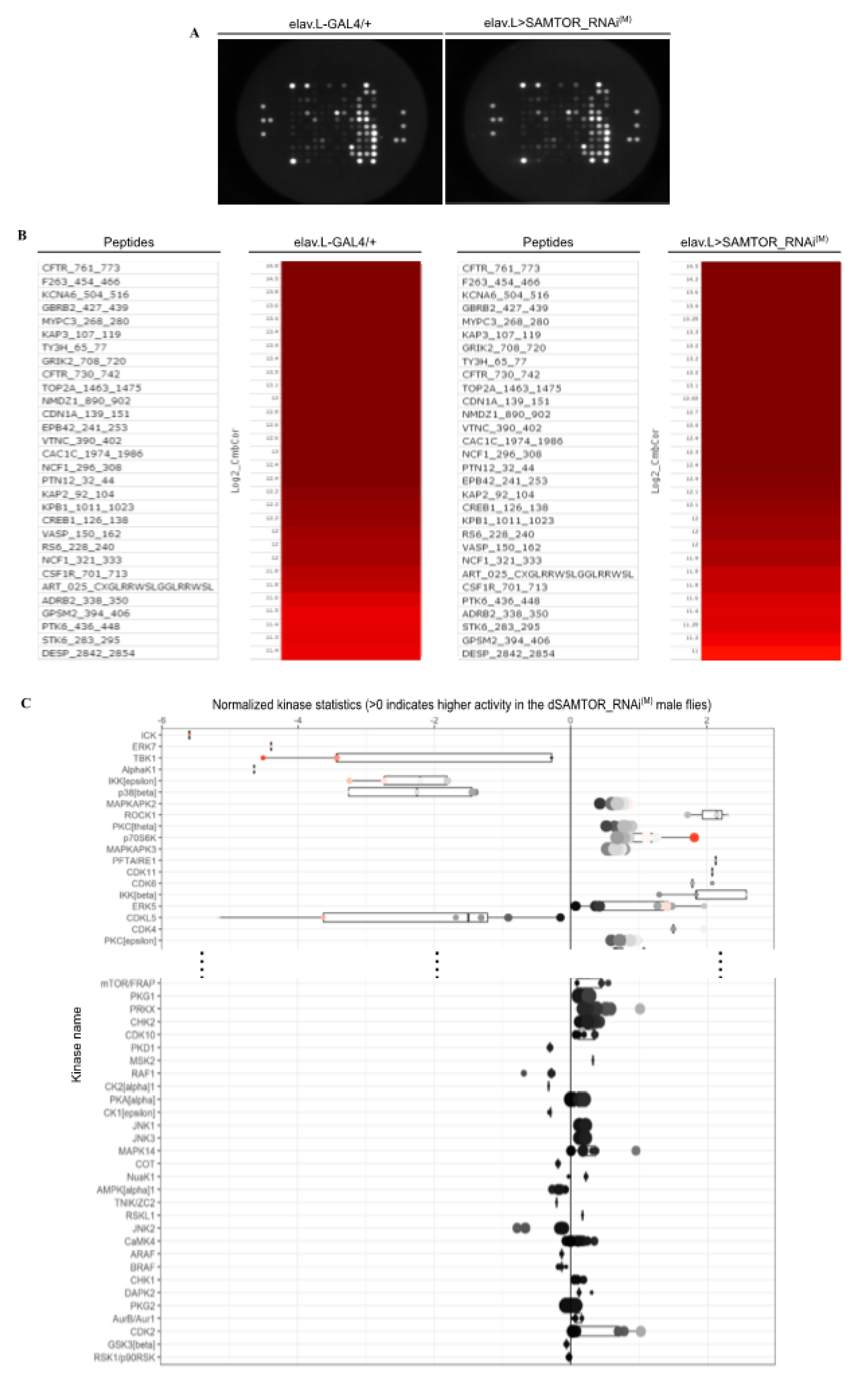
Figure 6.
Different levels of dSAMTOR gene downregulation result in distinct patterns of eye and wing pathology in Drosophila. (A) Scanning electron microscopy (SEM) images of compound eyes derived from male transgenic flies at the age of 10, 30 and 50 days, with moderate (M) (ninaE.GMR>SAMTOR_RNAi(M)) (d-f) and strong (S) (ninaE.GMR>SAMTOR_RNAi(S)) (g-i), respective, RNAi-mediated targeting of the dSAMTOR gene, as compared to control settings (ninaE.GMR-GAL4/+) (a-c). Scale Bars: 100 μm. (B) Wing images of female mutant (top panel) flies carrying moderate (M) (bxMS1096>SAMTOR_RNAi(M)) (b) and strong (S) (bxMS1096>SAMTOR_RNAi(S)) (c) dSAMTOR, respective, gene silencing, as compared to control fly populations (bxMS1096-GAL4/+) (a). Wing images of male mutant (bottom panel) flies, with moderate (M) (bxMS1096>SAMTOR_RNAi(M)) (e) and strong (S) (bxMS1096>SAMTOR_RNAi(S)) (f) dSAMTOR gene downregulation, as compared to control fly wings (bxMS1096-GAL4/+) (d). Scale Bars: (a), (b), (d) and (e): 200 µm; (c) and (f): 100 μm. SAMTOR_RNAi(M): RNAi strain with moderate (M) silencing capacity. SAMTOR_RNAi(S): RNAi strain with strong (S) silencing potency. Naming of normal (control) wing veins is indicated (L1-L6, a-cv and p-cv) (a) and (d).
Figure 6.
Different levels of dSAMTOR gene downregulation result in distinct patterns of eye and wing pathology in Drosophila. (A) Scanning electron microscopy (SEM) images of compound eyes derived from male transgenic flies at the age of 10, 30 and 50 days, with moderate (M) (ninaE.GMR>SAMTOR_RNAi(M)) (d-f) and strong (S) (ninaE.GMR>SAMTOR_RNAi(S)) (g-i), respective, RNAi-mediated targeting of the dSAMTOR gene, as compared to control settings (ninaE.GMR-GAL4/+) (a-c). Scale Bars: 100 μm. (B) Wing images of female mutant (top panel) flies carrying moderate (M) (bxMS1096>SAMTOR_RNAi(M)) (b) and strong (S) (bxMS1096>SAMTOR_RNAi(S)) (c) dSAMTOR, respective, gene silencing, as compared to control fly populations (bxMS1096-GAL4/+) (a). Wing images of male mutant (bottom panel) flies, with moderate (M) (bxMS1096>SAMTOR_RNAi(M)) (e) and strong (S) (bxMS1096>SAMTOR_RNAi(S)) (f) dSAMTOR gene downregulation, as compared to control fly wings (bxMS1096-GAL4/+) (d). Scale Bars: (a), (b), (d) and (e): 200 µm; (c) and (f): 100 μm. SAMTOR_RNAi(M): RNAi strain with moderate (M) silencing capacity. SAMTOR_RNAi(S): RNAi strain with strong (S) silencing potency. Naming of normal (control) wing veins is indicated (L1-L6, a-cv and p-cv) (a) and (d).
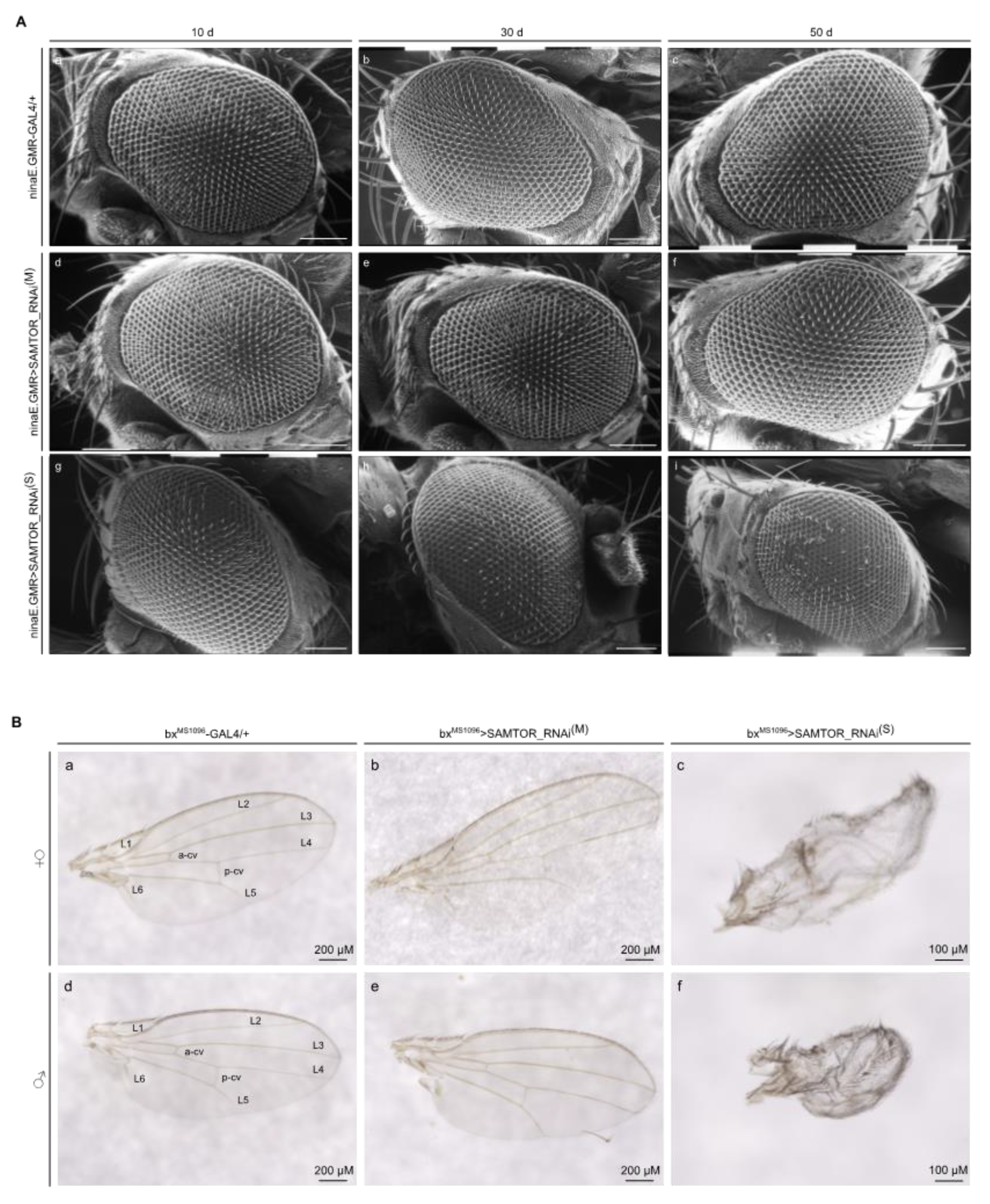
Figure 7.
(A) Schematic illustration of lysosome-associated components being critically involved in the Methionine-sensing pathway, upstream of mTORC1. BHMT uses Betaine to methylate Homocysteine directly into Methionine, which is, next, converted into SAM. SAMTOR has the ability to sense Methionine in the form of SAM. When SAM binds to SAMTOR, it (SAMTOR) dissociates from GATOR1 and leads to mTORC1 activation. (B) Strong evolutionary conservation of BHMT enzyme among species. Amino acid sequence alignment among human (Homo sapiens), mouse (Mus musculus), zebrafish (Danio rerio) and fly (Drosophila melanogaster) BHMT proteins, via employment of the Clustal Omega bioinformatics tool. BHMT: Betaine-Homocysteine S-Methyl-Transferase; MAT: Methionine Adenosyl-Transferase; ΜΤS: Methyltransferases; SAHH: S-Adenosyl-L-Homocysteine Hydrolase; SAM: S-Adenosyl-Methionine.
Figure 7.
(A) Schematic illustration of lysosome-associated components being critically involved in the Methionine-sensing pathway, upstream of mTORC1. BHMT uses Betaine to methylate Homocysteine directly into Methionine, which is, next, converted into SAM. SAMTOR has the ability to sense Methionine in the form of SAM. When SAM binds to SAMTOR, it (SAMTOR) dissociates from GATOR1 and leads to mTORC1 activation. (B) Strong evolutionary conservation of BHMT enzyme among species. Amino acid sequence alignment among human (Homo sapiens), mouse (Mus musculus), zebrafish (Danio rerio) and fly (Drosophila melanogaster) BHMT proteins, via employment of the Clustal Omega bioinformatics tool. BHMT: Betaine-Homocysteine S-Methyl-Transferase; MAT: Methionine Adenosyl-Transferase; ΜΤS: Methyltransferases; SAHH: S-Adenosyl-L-Homocysteine Hydrolase; SAM: S-Adenosyl-Methionine.
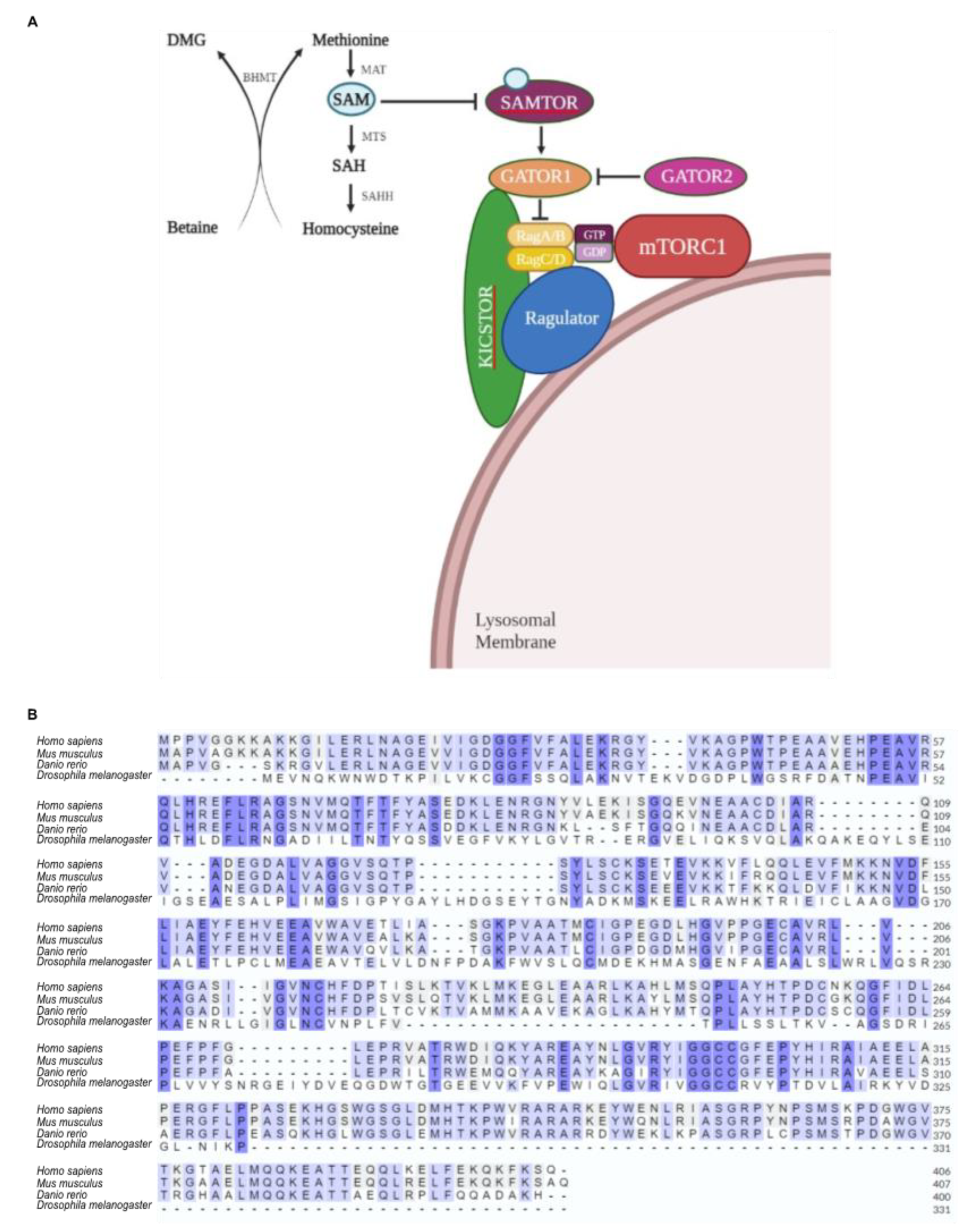
Figure 8.
Methionine and Betaine treatment of dSAMTOR, mildly, targeted flies, specifically in neuronal tissues, results in reduced viability. (A, B) Survival curves of female (left panel) and male (right panel) dSAMTOR, moderately, downregulated flies, specifically in neuronal tissues (elav.L>SAMTOR_RNAi(M)), in the presence of Methionine (5 mM) (supplemented into the food) for ~60 consecutive days (red lines), as compared to Methionine-exposed control (elav.L-GAL4/+) populations (green lines). (C, D) Survival rates of female (left panel) and male (right panel) dSAMTOR, modestly, targeted flies, specifically in neuronal tissues (elav.L>SAMTOR_RNAi(M)), after Betaine administration (10 mM) (in the food) for ~70 consecutive days (red lines), as compared to control (elav.L-GAL4/+) flies treated with the same dose of Betaine (green lines). ***p < 0.001.
Figure 8.
Methionine and Betaine treatment of dSAMTOR, mildly, targeted flies, specifically in neuronal tissues, results in reduced viability. (A, B) Survival curves of female (left panel) and male (right panel) dSAMTOR, moderately, downregulated flies, specifically in neuronal tissues (elav.L>SAMTOR_RNAi(M)), in the presence of Methionine (5 mM) (supplemented into the food) for ~60 consecutive days (red lines), as compared to Methionine-exposed control (elav.L-GAL4/+) populations (green lines). (C, D) Survival rates of female (left panel) and male (right panel) dSAMTOR, modestly, targeted flies, specifically in neuronal tissues (elav.L>SAMTOR_RNAi(M)), after Betaine administration (10 mM) (in the food) for ~70 consecutive days (red lines), as compared to control (elav.L-GAL4/+) flies treated with the same dose of Betaine (green lines). ***p < 0.001.
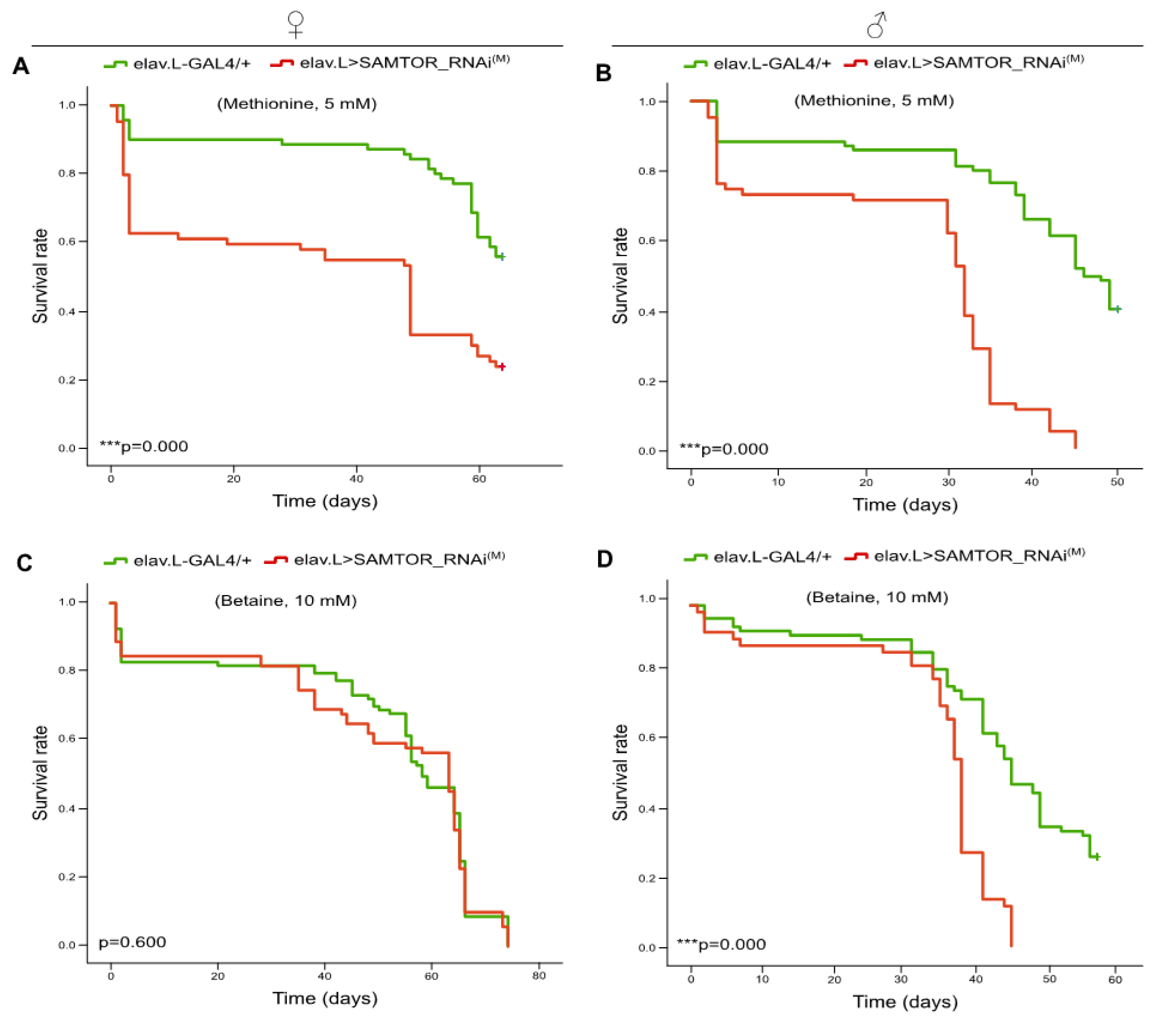
Figure 9.
dBHMT gene suppression reduces Drosophila’s viability in diverse type of tissues. Lifespan profiles of Drosophila female (left panels) and male (right panels) transgenic flies, being characterized by RNAi-mediated and tissue-specific targeting of the dBHMT gene in: (A, B) neuronal tissues (elav.L>BHMT_RNAi), (C, D) glia cells (repo>BHMT_RNAi), (E, F) motor neurons (D42>BHMT_RNAi), (G, H) muscles (mef2>BHMT_RNAi) and (I, J) intestine (midgut) tissues (NP1>BHMT_RNAi). ***p < 0.001.
Figure 9.
dBHMT gene suppression reduces Drosophila’s viability in diverse type of tissues. Lifespan profiles of Drosophila female (left panels) and male (right panels) transgenic flies, being characterized by RNAi-mediated and tissue-specific targeting of the dBHMT gene in: (A, B) neuronal tissues (elav.L>BHMT_RNAi), (C, D) glia cells (repo>BHMT_RNAi), (E, F) motor neurons (D42>BHMT_RNAi), (G, H) muscles (mef2>BHMT_RNAi) and (I, J) intestine (midgut) tissues (NP1>BHMT_RNAi). ***p < 0.001.
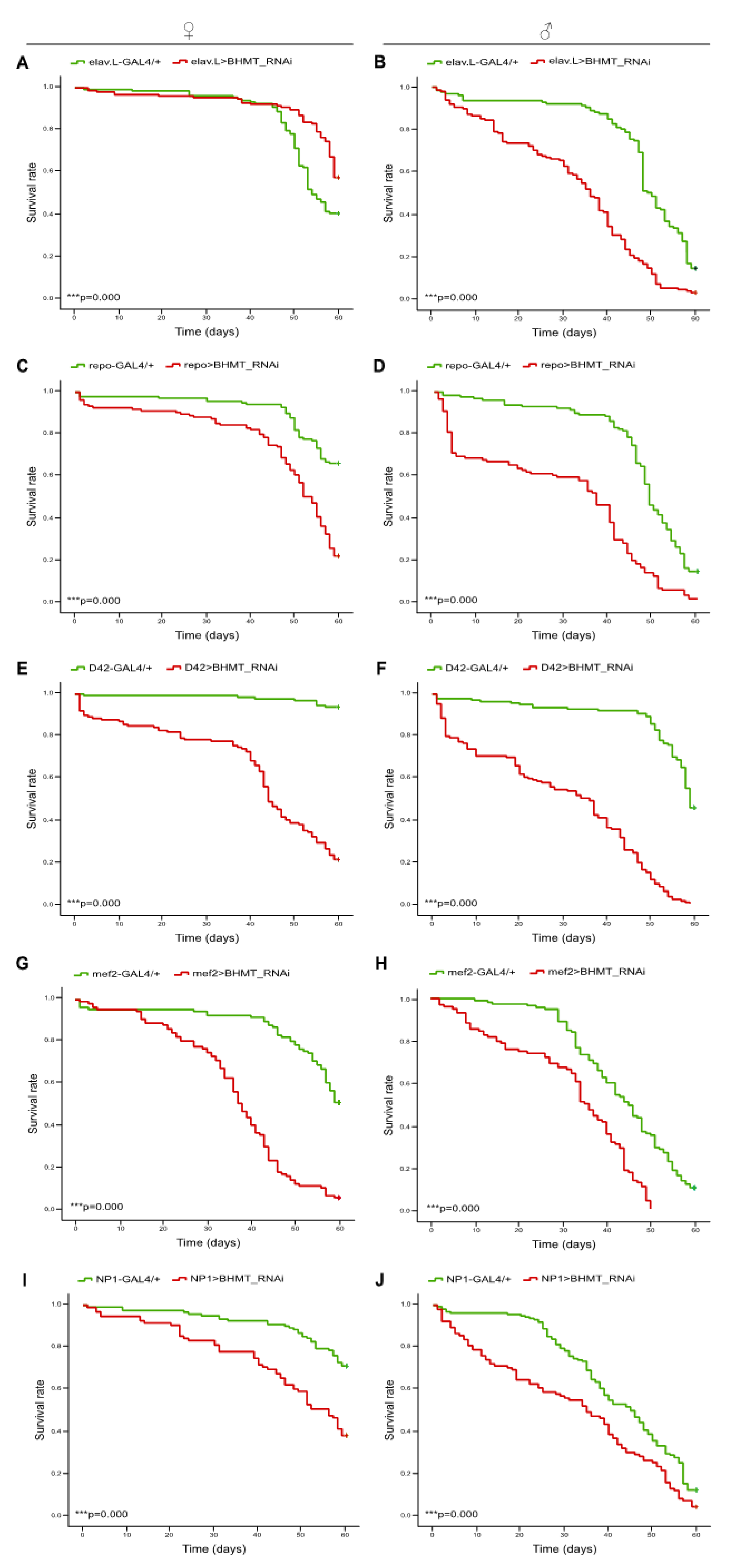
Figure 10.
Downregulation of dΒHΜΤ gene expression differentially impairs climbing capacity during Drosophila aging, revealing tissue-specific signatures. Climbing activity (negative geotaxis) patterns of female (left panels) and male (right panels) transgenic flies, being characterized by tissue-specific dΒHΜΤ gene targeting in: (A, B) neuronal tissues (elav.L>BHMT_RNAi), (C, D) glia cells (repo>BHMT_RNAi), (E, F) motor neurons (D42>BHMT_RNAi), (G, H) muscles (mef2>BHMT_RNAi) and (I, J) intestine (midgut) tissues (NP1>BHMT_RNAi). *p < 0.05 and ***p < 0.001.
Figure 10.
Downregulation of dΒHΜΤ gene expression differentially impairs climbing capacity during Drosophila aging, revealing tissue-specific signatures. Climbing activity (negative geotaxis) patterns of female (left panels) and male (right panels) transgenic flies, being characterized by tissue-specific dΒHΜΤ gene targeting in: (A, B) neuronal tissues (elav.L>BHMT_RNAi), (C, D) glia cells (repo>BHMT_RNAi), (E, F) motor neurons (D42>BHMT_RNAi), (G, H) muscles (mef2>BHMT_RNAi) and (I, J) intestine (midgut) tissues (NP1>BHMT_RNAi). *p < 0.05 and ***p < 0.001.
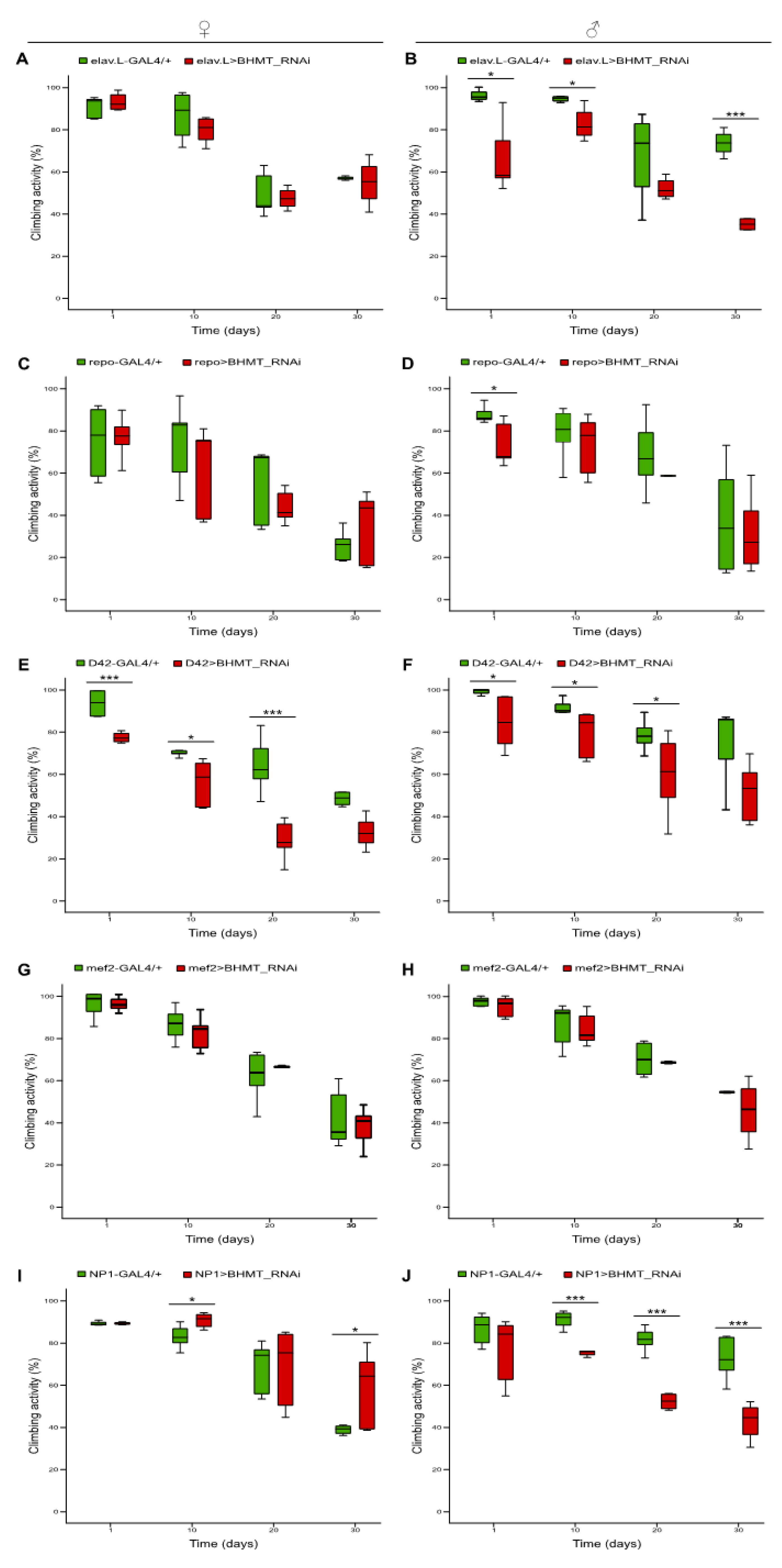
Figure 11.
dBHMT gene downregulation causes eye dysmorphia in Drosophila: an age-dependent pathology. Scanning electron microscopy (SEM) images of compound eyes derived from control (ninaE.GMR-GAL4/+) (A-C) and dBHMT-targeted, specifically in the eye (ninaE.GMR>BHMT_RNAi), (D-F) male transgenic flies, at the age of 10, 30 and 50 days (d). Scale Bars: 100 μm.
Figure 11.
dBHMT gene downregulation causes eye dysmorphia in Drosophila: an age-dependent pathology. Scanning electron microscopy (SEM) images of compound eyes derived from control (ninaE.GMR-GAL4/+) (A-C) and dBHMT-targeted, specifically in the eye (ninaE.GMR>BHMT_RNAi), (D-F) male transgenic flies, at the age of 10, 30 and 50 days (d). Scale Bars: 100 μm.

Figure 12.
Reduced levels of dBHMT gene expression disrupts wing morphology and architecture in Drosophila: age-dependent profiles. (A) Wing images with differential vein pathology of male mutant flies being subjected to dBHMT gene silencing (bxMS1096>BHMT_RNAi) (b-d), as compared to control fly populations (bxMS1096-GAL4/+) (a). Scale Bars: 200 µm. (B) Bar charts presenting abnormal vein phenotypes of male transgenic flies (bxMS1096>BHMT_RNAi), as compared to control settings (bxMS1096-GAL4/+), during aging. (C) Quantification of wing areas of dBHMT mutant (bxMS1096>BHMT_RNAi) and control (bxMS1096-GAL4/+) flies, during aging. Naming of normal (control) wing veins is indicated (L1-L6, a-cv and p-cv) (a). Manually dissected wings from at least 50 male flies of each genotype were thoroughly examined and carefully quantified, as indicated.
Figure 12.
Reduced levels of dBHMT gene expression disrupts wing morphology and architecture in Drosophila: age-dependent profiles. (A) Wing images with differential vein pathology of male mutant flies being subjected to dBHMT gene silencing (bxMS1096>BHMT_RNAi) (b-d), as compared to control fly populations (bxMS1096-GAL4/+) (a). Scale Bars: 200 µm. (B) Bar charts presenting abnormal vein phenotypes of male transgenic flies (bxMS1096>BHMT_RNAi), as compared to control settings (bxMS1096-GAL4/+), during aging. (C) Quantification of wing areas of dBHMT mutant (bxMS1096>BHMT_RNAi) and control (bxMS1096-GAL4/+) flies, during aging. Naming of normal (control) wing veins is indicated (L1-L6, a-cv and p-cv) (a). Manually dissected wings from at least 50 male flies of each genotype were thoroughly examined and carefully quantified, as indicated.
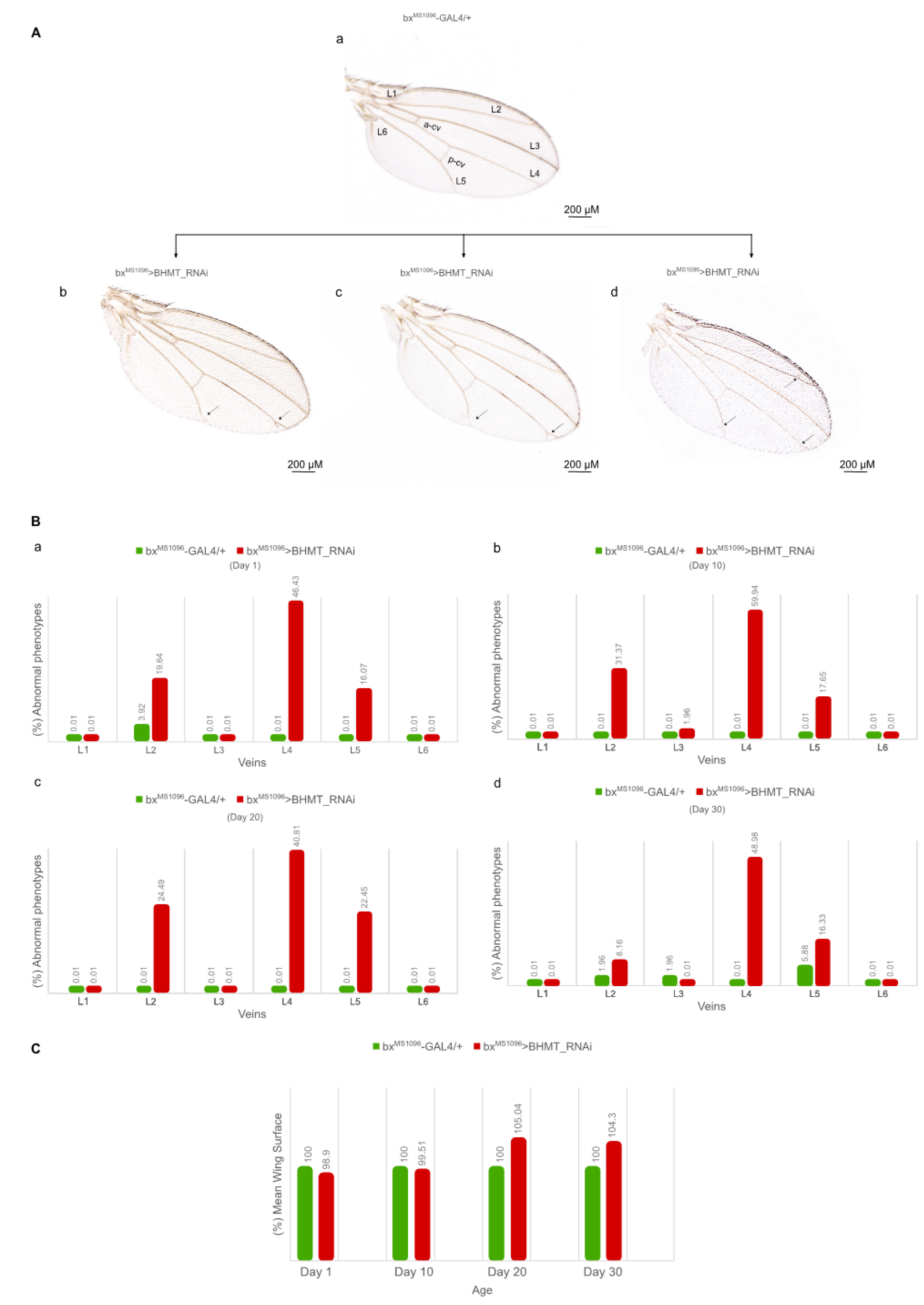

Table 1.
Pathology of dSAMTOR gene, tissue-specific, downregulation, categorized in lethal and viable, systemic, phenotypes. S: Strong downregulation; M: Moderate downregulation.
Table 1.
Pathology of dSAMTOR gene, tissue-specific, downregulation, categorized in lethal and viable, systemic, phenotypes. S: Strong downregulation; M: Moderate downregulation.
| DRIVER | dSAMTOR_RNAi(S) | dSAMTOR_RNAi(M) | |
| STRAIN | |||
| Act5C-GAL4 (whole body) | Lethal | Viable | |
| elav.L-GAL4 (nervous system) | Lethal | Viable | |
| repo-GAL4 (glial cells) | Lethal | Viable | |
| D42-GAL4 (motor neurons) | Lethal | Viable | |
| Mef2.R-GAL4 (muscles) | Lethal | Viable | |
| NP1-GAL4 (midgut) | Viable | Viable | |
| ninaE.GMR-GAL4 (eyes) | Viable | Viable | |
| bxMS1096-GAL4 (wings) | Viable | Viable | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions, or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



