
Submitted:
22 April 2023
Posted:
23 April 2023
You are already at the latest version
Abstract
The potential to improve the effectiveness and efficiency of potential estrogen-based oral contraceptives (fertility control) for possums was investigated by comparing the inhibitory potential of hepatic CYP3A and UGT2B catalytic activity using a selected compound library (CYP450 inhibitor-based compounds) in possums to that of three other species (mouse, avian, and human). The results showed higher CYP3A protein levels in possum liver microsomes compared to other test species (up to a 4-fold difference). Also, possum liver microsomes had significantly higher basal p-nitrophenol glucuronidation activity than other test species (up to an 8-fold difference). However, no CYP450 inhibitor-based compounds significantly decreased the catalytic activity of possum CYP3A and UGT2B below the estimated IC50 and 2-fold IC50 values and were therefore not considered to be potent inhibitors of these enzymes. However, compounds such as isosilybin (65%), ketoconazole (72%) and fluconazole (74%) showed reduced UGT2B glucuronidation activity in possums, mainly at 2-fold IC50 values compared to control (p<0.05). Given the structural features of these compounds, these results could provide opportunities for future compound screening. More importantly, however, this study provided preliminary evidence that the basal activity and protein content of two major drug metabolising enzymes differ in possums compared to other test species, suggesting that this could be further exploited to reach the ultimate goal, a potential target-specific fertility control for possums in New Zealand.
Keywords:
Brushtail possum
; Trichosurus vulpecula
; fertility control
; and drug-metabolising enzymes (CYP3A and UGT2B)
1. Introduction
Brushtail possums (Trichosurus vulpecula) are a major pest species in New Zealand, threatening biodiversity and the economy [1]. For decades there have been continuous efforts (mainly using chemical and physical methods) to reduce the possum population, but the species remains a threat to native biodiversity and the economy, as also recently reviewed by our research group [2].
Synthetic estrogens in the form of oral contraceptives could serve as a potential tool for possum management in New Zealand. Compounds such as ethinylestradiol (EE2) and diethylstilbestrol can be used alone or in combination with progestin as an oral contraceptive, since elevated levels of such synthetic hormones impair the normal functioning of the hypothalamic-pituitary-gonadal (HPG) axis [3]. In particular, EE2 works together with the hormone progestin, which is particularly important for changes in the endometrium of the uterus, to suppress the HPG axis and thus reduce the production of follicle-stimulating hormone (FSH) and luteinising hormone (LH) [4]. The reduced FSH and LH levels thus prevent follicle development and ovulation [5]. This has significantly reduced ovulation in species such as koalas [6,7], mice [8,9], red foxes [10], and white-tailed deer [11].
Although oral contraceptives have successfully prevented ovulation in animals, they also come with the major challenge of rapid hepatic metabolism [12,13,14,15]. The rapid hepatic metabolism of oral contraceptives by drug-metabolising enzymes reduces the amount of synthetic estrogen and progestin absorbed into the body from the given formulation [15]. The reduced bioavailability of these oral contraceptives can therefore lead to low oral contraceptive efficacy [16]. Previously, as reported for the first time by our group, possum-specific differences in major drug metabolising enzymes were found compared to rodents (mice and rats). The results showed that possums had ~2-fold higher UGT catalytic activity and ~20% lower CYP3A catalytic activity compared to mice and rats [17]. Therefore, the present study aimed to further investigate possum-specific differences in the major drug-metabolising enzymes (CYP3A and UGT2B) using CYP450-based compounds in rodent, avian and human liver samples. Since estrogens are metabolised by CYP3A and UGT2B enzymes, finding possum-specific potent inhibitors of these enzymes could help improve the effectiveness of fertility control in possums, while limiting their impact on non-target species.
2. Results
2.1. CYP3A catalytic activity
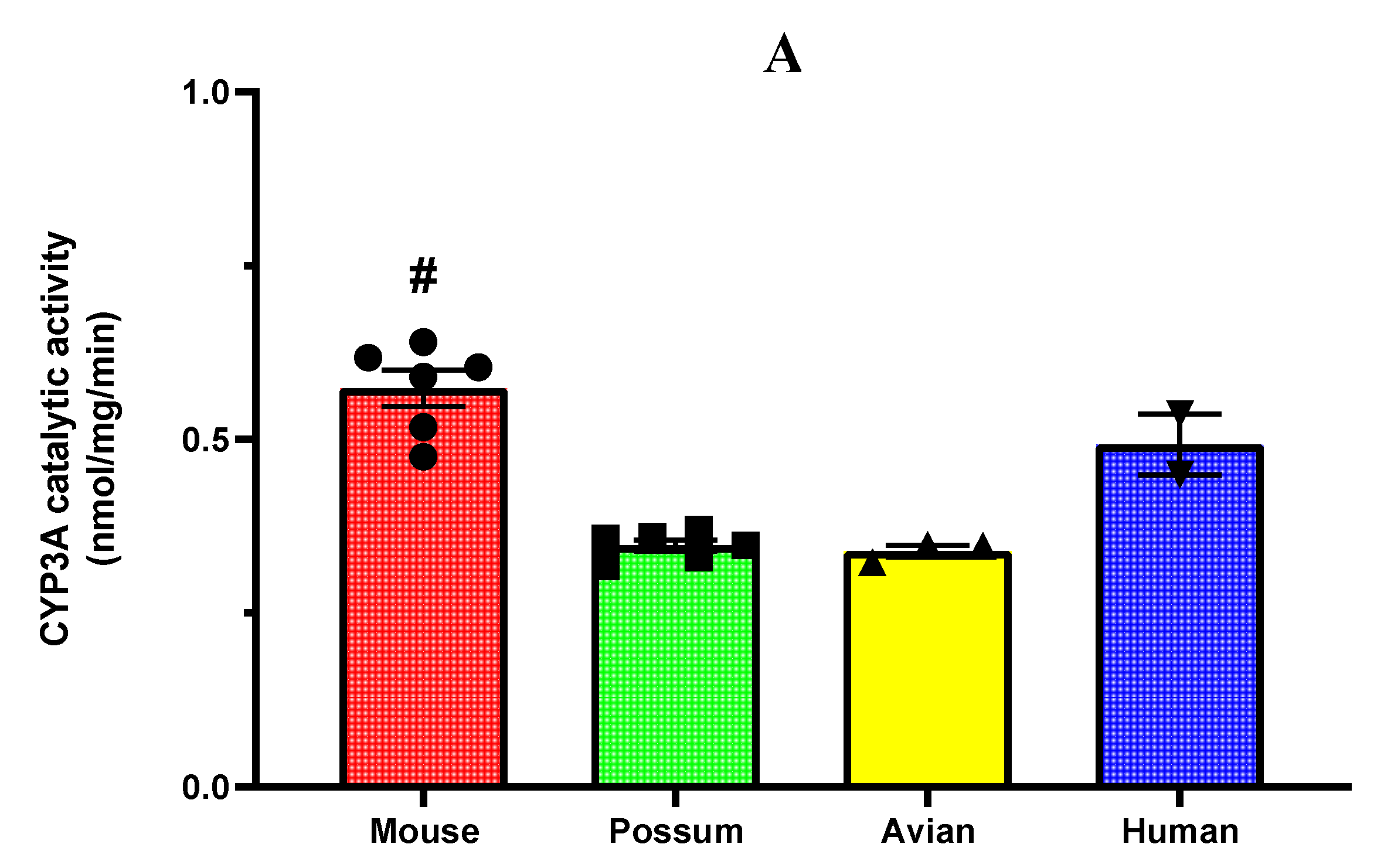
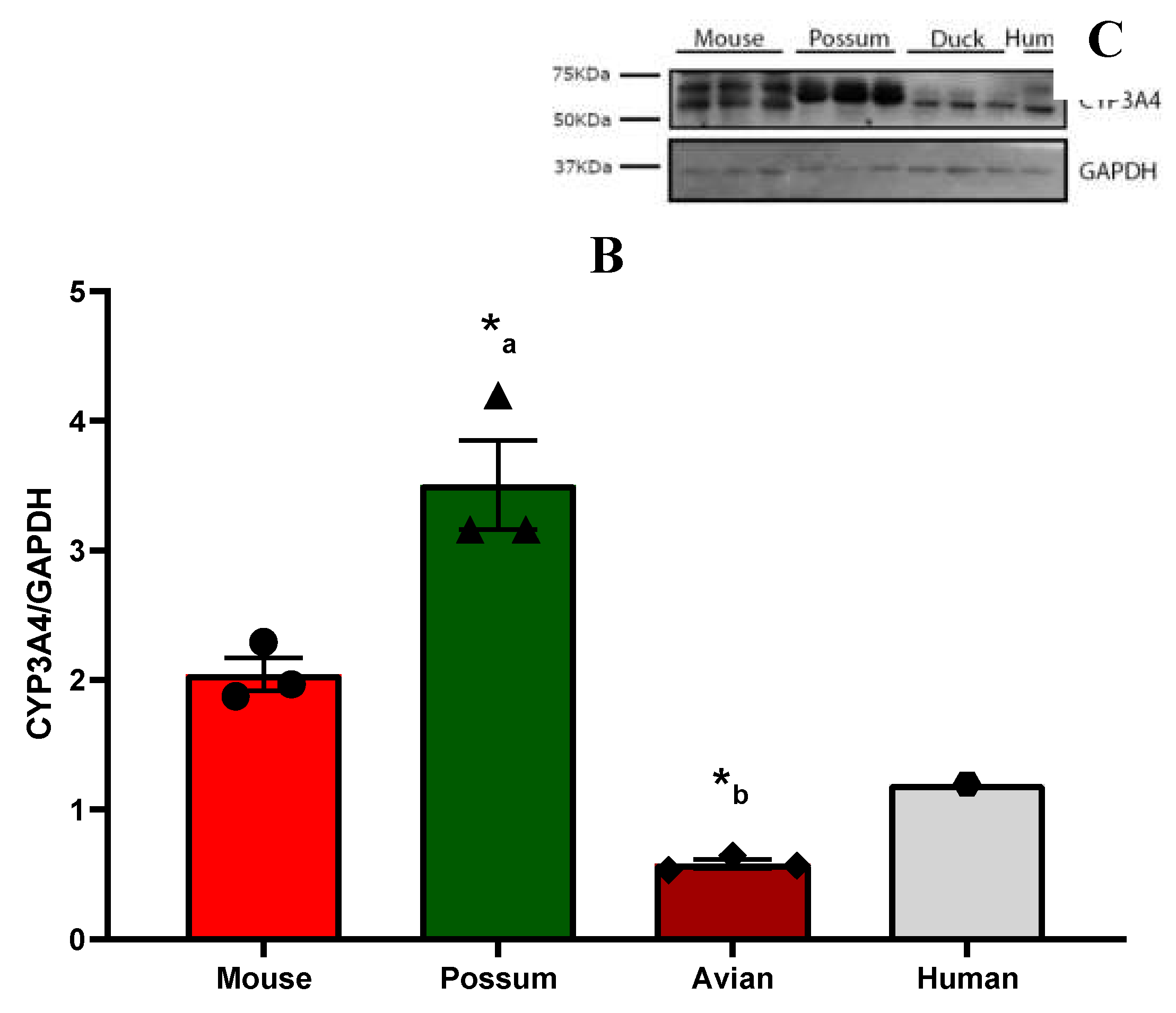
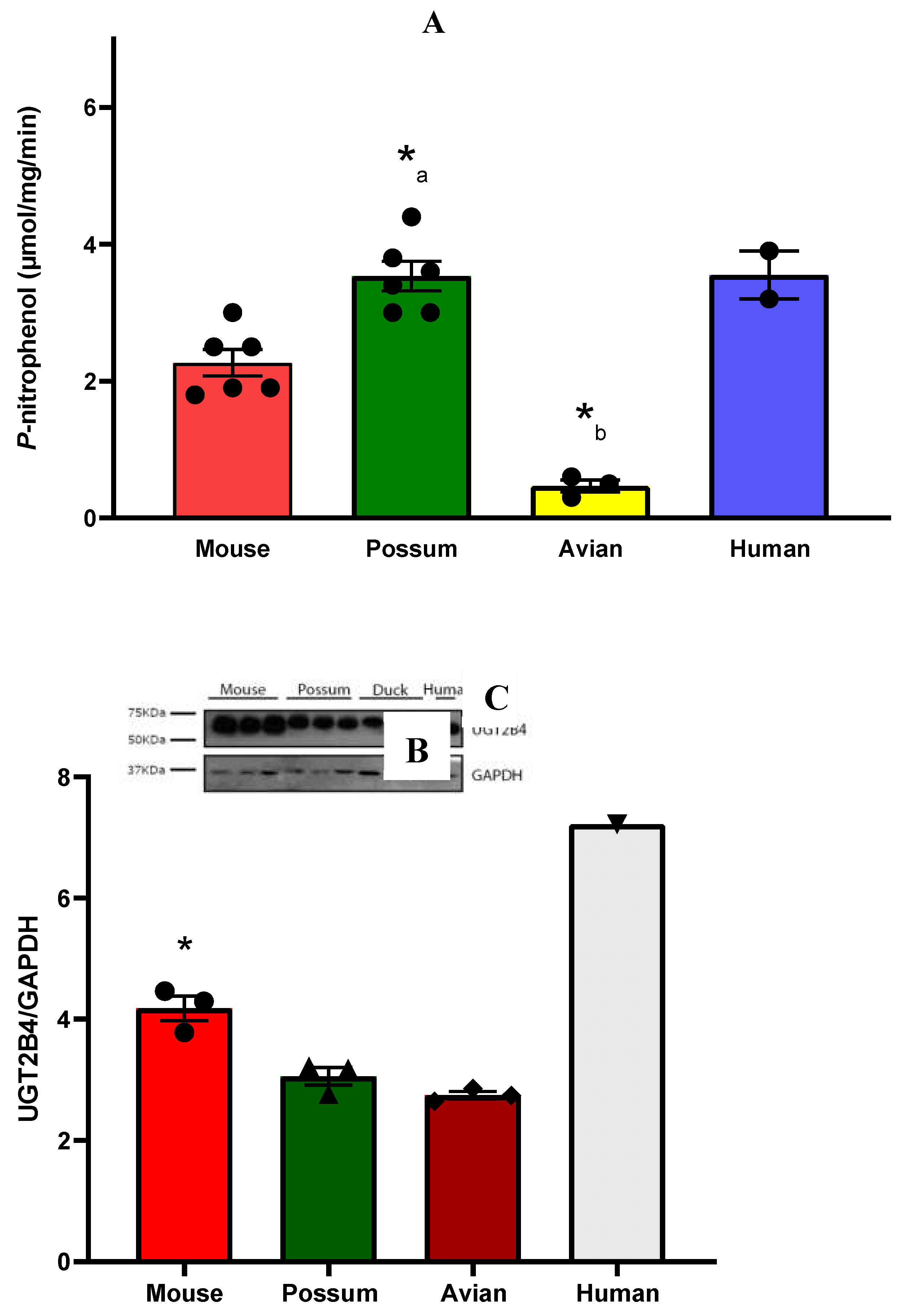
To identify possum-specific activity differences in phase I drug metabolising enzymes, the basal catalytic activities of CYP3A in mouse, possum, avian, and human were determined (Figure 1A). The basal CYP3A catalytic activity was significantly higher (~2-fold) in mouse than in possum and avian species (p<0.05). While the basal CYP3A catalytic activities between mouse and human, and possum and avian were not significantly different (p>0.05). The mean basal CYP3A catalytic activities in these species were 0.57±0.03 (mouse), 0.35±0.01 (possum), 0.34±0.01 (avian) and 0.49±0.04 (human) in nmol/mg/min. In addition, CYP3A4 protein levels also varied between these species (Figure 1: B and C). In particular, CYP3A4 protein levels were significantly higher in possum than in avian (~4-fold). While CYP3A4 protein levels were also higher in possum, but not statistically significant (p>0.05), compared to mouse (~2-fold). The mouse CYP3A4 protein levels were ~2-fold higher than the avian. To make a comparison across target and non-target species, mouse, possum, avian and human were examined together.
Figure 1.
Basal hepatic CYP3A activity and protein levels in mouse, possum, avian and human. Liver microsomes of each species were used to determine CYP3A activity via erythromycin N-demethylation. (A). Values are expressed in nmol/mg/min. Bars represent the mean ± SEM of n=6 in triplicate (mouse and possum), n=3 in triplicate (avian) and n=2 in duplicate (pooled human liver microsomes). #Significantly different compared to possum and avian, p<0.05. The protein content for CYP3A4 in liver microsomes from 4 species was determined by Western blotting (load of protein in each well was 2 µg). Hepatic microsomes were also subjected to CYP3A4 Western blotting and the scanning densitometry results were normalised to the housekeeping protein GAPDH. (B) The bars represent the mean ± SEM of optical density of n=3 (except pooled human microsomes, n=1). (*a) Significantly different compared to avian, while (*b) Significantly different compared to mouse, p<0.05. Representative Western blot (C).
Figure 1.
Basal hepatic CYP3A activity and protein levels in mouse, possum, avian and human. Liver microsomes of each species were used to determine CYP3A activity via erythromycin N-demethylation. (A). Values are expressed in nmol/mg/min. Bars represent the mean ± SEM of n=6 in triplicate (mouse and possum), n=3 in triplicate (avian) and n=2 in duplicate (pooled human liver microsomes). #Significantly different compared to possum and avian, p<0.05. The protein content for CYP3A4 in liver microsomes from 4 species was determined by Western blotting (load of protein in each well was 2 µg). Hepatic microsomes were also subjected to CYP3A4 Western blotting and the scanning densitometry results were normalised to the housekeeping protein GAPDH. (B) The bars represent the mean ± SEM of optical density of n=3 (except pooled human microsomes, n=1). (*a) Significantly different compared to avian, while (*b) Significantly different compared to mouse, p<0.05. Representative Western blot (C).
Since differences in the basal catalytic activities of CYP3A were found in the test species, a compound library on the structural features of known CYP450 inhibitors was then examined for possum-specific differences in inhibition. These 59 compounds were initially screened with estimated IC50 and 2-fold IC50 values (Supplementary Table 1). Compounds with previously unreported IC50 values were screened at concentrations of 1 and 5 μM. From this initial screen, 24 compounds that showed the greatest inhibition against possum CYP3A were selected for further examination. Of these 24 compounds, none showed significant inhibition (<50% of control) of CYP3A enzyme activity in possum liver microsomes at estimated IC50 and 2-fold IC50 values or at concentrations of 1 and 5 μM (Supplementary Table 2). However, 12 compounds reduced the catalytic activity of possum CYP3A to some extent, mainly at a concentration of 5 μM or estimated 2-fold IC50 values, but these reduced catalytic activities were not significantly different from their respective controls (Figure 2A). These compounds were WAY-325945 (65%), ketoconazole (73%), cobicistat (76%), terfenadine (79%), benzbromarone (80%), WAY-657644 (81%), ritonavir (81%), gentiopicroside (82%), WAY-325412 (83%), chlorzoxazone (83%), galeterone (83%), and alizarin (84%) of the control. In order to determine species-specific differences, these compounds were then examined in the mouse. Interestingly, ketoconazole was the only compound that significantly reduced mouse CYP3A catalytic activity at IC50 and 2-fold IC50 values (40-46% of control, p<0.05) (Figure 2B). While other compounds such as WAY-325945 (67%), cobicistat (68%), 2’-hydroxyacetophenone (75%), terfenadine (83%), ritonavir (84%) and WAY-657644 (88%) showed some level of inhibition in mouse liver microsomes at a concentration of 5 μM or an estimated 2-fold IC50 value. However, these compounds were not statistically significant from their respective controls. A further six compounds compared from the list of 12 compounds that showed some reduction in possum CYP3A catalytic activities were further examined at a higher concentration (10 μM) in possum and mouse liver microsomes. This was done to determine if the tested compounds would further decrease the catalytic activities of CYP3A, particularly in possums. The results showed that these 6 compounds (alizarin, galeterone, cobicistat, gentiopicroside, chlorzoxazone, and benzbromarone) did not show any significant inhibition of CYP3A catalytic activities in either species at 10 μM (Figure 3).
Figure 2.
Effect of CYP450 inhibitor-based compounds on CYP3A catalytic activity in possum and mouse liver microsomes. Possum (A) and mouse (B) liver microsomes were incubated with 0.1 M phosphate buffer and CYP450 inhibitor-based compounds, and erythromycin N-demethylation was determined as an indicator of CYP3A inhibitory catalytic activity at estimated IC50 and 2-fold IC50 values. The dotted line shows the 50% control of CYP3A enzymes. The bars represent the mean ± SEM of n=3 in duplicate in percent control. *Significantly different from the respective solvent control. Abbreviated compound names refer to WAY-325945 (WAY-3259), ketoconazole (Keto), cobicistat (Cobi), terfenadine (Terf), benzbromarone (Benz), WAY-657644 (WAY-6576), ritonavir (Rito), gentiopicroside (Genti), WAY-325412 (WAY-3254), chlorzoxazone (Chlor), galeterone (Gale), and alizarin (Ali).
Figure 2.
Effect of CYP450 inhibitor-based compounds on CYP3A catalytic activity in possum and mouse liver microsomes. Possum (A) and mouse (B) liver microsomes were incubated with 0.1 M phosphate buffer and CYP450 inhibitor-based compounds, and erythromycin N-demethylation was determined as an indicator of CYP3A inhibitory catalytic activity at estimated IC50 and 2-fold IC50 values. The dotted line shows the 50% control of CYP3A enzymes. The bars represent the mean ± SEM of n=3 in duplicate in percent control. *Significantly different from the respective solvent control. Abbreviated compound names refer to WAY-325945 (WAY-3259), ketoconazole (Keto), cobicistat (Cobi), terfenadine (Terf), benzbromarone (Benz), WAY-657644 (WAY-6576), ritonavir (Rito), gentiopicroside (Genti), WAY-325412 (WAY-3254), chlorzoxazone (Chlor), galeterone (Gale), and alizarin (Ali).
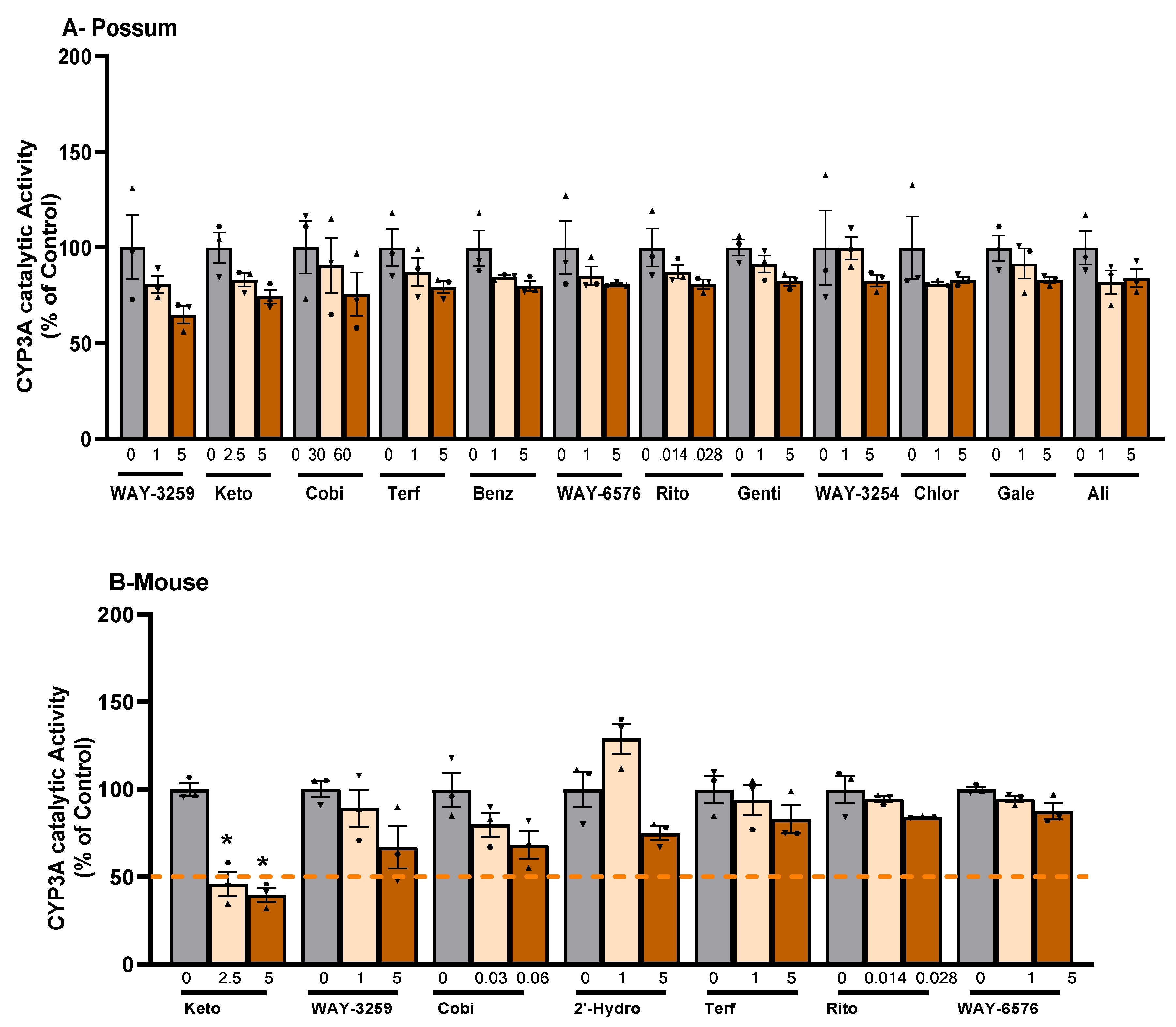
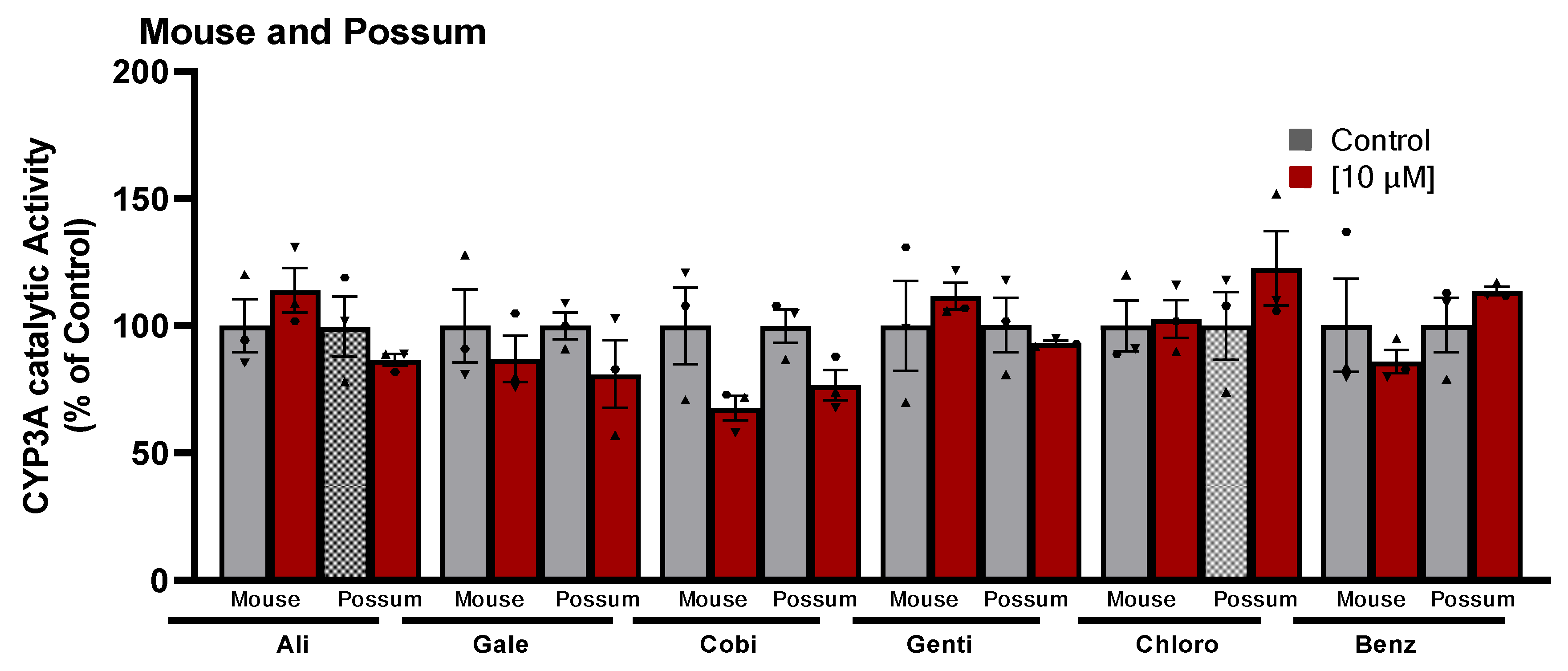
Figure 3.
Effect of CYP450 inhibitor-based compounds on CYP3A catalytic activity in mouse and possum liver microsomes. Possum and mouse liver microsomes were incubated with 0.1 M phosphate buffer and CYP450 inhibitor-based compounds, and erythromycin N-demethylation was determined at a higher concentration (10 µM) as an indicator of CYP3A inhibitory catalytic activity. The bars represent the mean ± SEM of n=3 in duplicate. Data were analysed using one-way ANOVA and none were statistically different compared to controls. Abbreviated compound names refer to alizarin (Ali), galeterone (Gale), cobicistat (Cobi), gentiopicroside (Genti), chlorzoxazone (Chlor), and benzbromarone (Benz).
Figure 3.
Effect of CYP450 inhibitor-based compounds on CYP3A catalytic activity in mouse and possum liver microsomes. Possum and mouse liver microsomes were incubated with 0.1 M phosphate buffer and CYP450 inhibitor-based compounds, and erythromycin N-demethylation was determined at a higher concentration (10 µM) as an indicator of CYP3A inhibitory catalytic activity. The bars represent the mean ± SEM of n=3 in duplicate. Data were analysed using one-way ANOVA and none were statistically different compared to controls. Abbreviated compound names refer to alizarin (Ali), galeterone (Gale), cobicistat (Cobi), gentiopicroside (Genti), chlorzoxazone (Chlor), and benzbromarone (Benz).
In addition, 19-21 compounds that showed potential differences in CYP3A inhibitory activity in possum and mouse were further examined in avian and human liver microsomes. None of the test compounds showed any significant inhibition (<50% of control) of catalytic activity in either species. Nonetheless, in avian liver microsomes, most of the compounds showed some level of inhibition at 5 μM concentrations or the estimated 2-fold IC50 values, but these did not differ significantly from their respective controls.
2.1. P-nitrophenol glucuronidation activity
To identify possum-specific differences in phase II drug metabolising enzymes, basal UGT2B glucuronidation activity was determined. Possum had significantly higher p-nitrophenol glucuronidation activity than the avian (~8-fold) and mouse (~1.5-fold) (Figure 4A). The mouse also had ~5-fold higher p-nitrophenol glucuronidation activity than the avian. The basal p-nitrophenol glucuronidation activities in these species were 3.60±0.17 (possum), 3.44±0.13 (human), mouse (2.44±0.18) and avian (0.46±0.04) in μmol/mg/min. Additionally, UGT2B4 protein levels also varied between species (Figure 4B and 4C). The UGT2B4 protein levels from pooled human liver microsomes were higher than those from avian (~3-fold), possum (~3-fold), and mouse (~2-fold). Furthermore, the mouse hepatic UGT2B4 protein levels were significantly increased from the possum and mouse.
Figure 4.
P-nitrophenol glucuronidation activity in mouse, possum, avian and human liver microsomes. Liver microsomes of each species were incubated with 0.1 M phosphate buffer and <10% DMSO, and the p-nitrophenol glucuronidation activity was determined as an indicator of UGT glucuronidation activity (A). Values are expressed as µmol/mg/min. Bars represent the mean ± SEM of n=6 in triplicate for possum and mouse, n=3 in triplicate for avian, and n=2 in duplicate for pooled human liver microsomes. (*a) Significantly different from avian and (*b) Significantly different from mouse, p<0.05. The protein content for UGT2B4 in hepatic microsomes from 4 species was determined by Western blotting (load of protein in each well was 2 µg). UGT2B4 Western blots (B) and scanning densitometry results normalized to the housekeeping protein GAPDH (C). The bars represent the mean ± SEM of optical density from n=3 (except for pooled human liver microsomes, n=1). *Significantly different compared to solvent control, p<0.05.
Figure 4.
P-nitrophenol glucuronidation activity in mouse, possum, avian and human liver microsomes. Liver microsomes of each species were incubated with 0.1 M phosphate buffer and <10% DMSO, and the p-nitrophenol glucuronidation activity was determined as an indicator of UGT glucuronidation activity (A). Values are expressed as µmol/mg/min. Bars represent the mean ± SEM of n=6 in triplicate for possum and mouse, n=3 in triplicate for avian, and n=2 in duplicate for pooled human liver microsomes. (*a) Significantly different from avian and (*b) Significantly different from mouse, p<0.05. The protein content for UGT2B4 in hepatic microsomes from 4 species was determined by Western blotting (load of protein in each well was 2 µg). UGT2B4 Western blots (B) and scanning densitometry results normalized to the housekeeping protein GAPDH (C). The bars represent the mean ± SEM of optical density from n=3 (except for pooled human liver microsomes, n=1). *Significantly different compared to solvent control, p<0.05.
To examine the possum-specific differences in UGT enzymes, 17 compounds were further screened for their potential to inhibit UGT2B enzymes in possum, mouse, avian, and human liver microsomes (Supplementary Table 3). Compounds such as isosilybin (65%), ketoconazole (72%) and fluconazole (74%) showed reduced UGT2B glucuronidation activity in possums, mainly at 2-fold IC50 values compared to their respective controls (p<0.05). Even at 2-fold IC50 values, these reduced UGT2B inhibitory activities were not less than 50% of control (Figure 5). When one of these compounds (isosilybin) was further tested for its inhibitory potential on glucuronidation activity with varying concentrations of p-nitrophenol (substrate), it also showed no significant changes compared to their respective control activities (Table 1). Similarly, in the mouse, compounds such as fluconazole (78%) and cyclosporin A (80%) reduced p-nitrophenol glucuronidation activity at 2-fold IC50 values compared to their respective controls (p<0.05), but these inhibitory activities did not fall below 50%. In addition, activity elicited by most of the test compounds in human liver microsomes was over 88% of control. Even at 2-fold IC50 values, there was no major inhibition. Specifically, the compounds screened in human liver microsomes were itraconazole (88%), cobicistat (90%) and fluconazole (94%). In addition, most of the compounds examined showed reduced p-nitrophenol glucuronidation activity (ranging from 33-87% of control) in avian species, but these did not differ significantly from their respective controls.
Figure 5.
Compound-mediated inhibition of p-nitrophenol glucuronidation activity in possum and mouse liver microsomes. Liver microsomes of possum (A) and mouse (B) were incubated with 0.1 M phosphate buffer and inhibitory compounds, and p-nitrophenol glucuronidation activity was determined as an indicator of UGT glucuronidation activity. * Significantly different from solvent control, p<0.05. The bars represent the mean ± SEM of n=3 in duplicate in percent control. Abbreviated compound names refer to isosilybin (Isosi), ketoconazole (Keto), fluconazole (Fluco), and cyclosporin A (Cyclo).
Figure 5.
Compound-mediated inhibition of p-nitrophenol glucuronidation activity in possum and mouse liver microsomes. Liver microsomes of possum (A) and mouse (B) were incubated with 0.1 M phosphate buffer and inhibitory compounds, and p-nitrophenol glucuronidation activity was determined as an indicator of UGT glucuronidation activity. * Significantly different from solvent control, p<0.05. The bars represent the mean ± SEM of n=3 in duplicate in percent control. Abbreviated compound names refer to isosilybin (Isosi), ketoconazole (Keto), fluconazole (Fluco), and cyclosporin A (Cyclo).
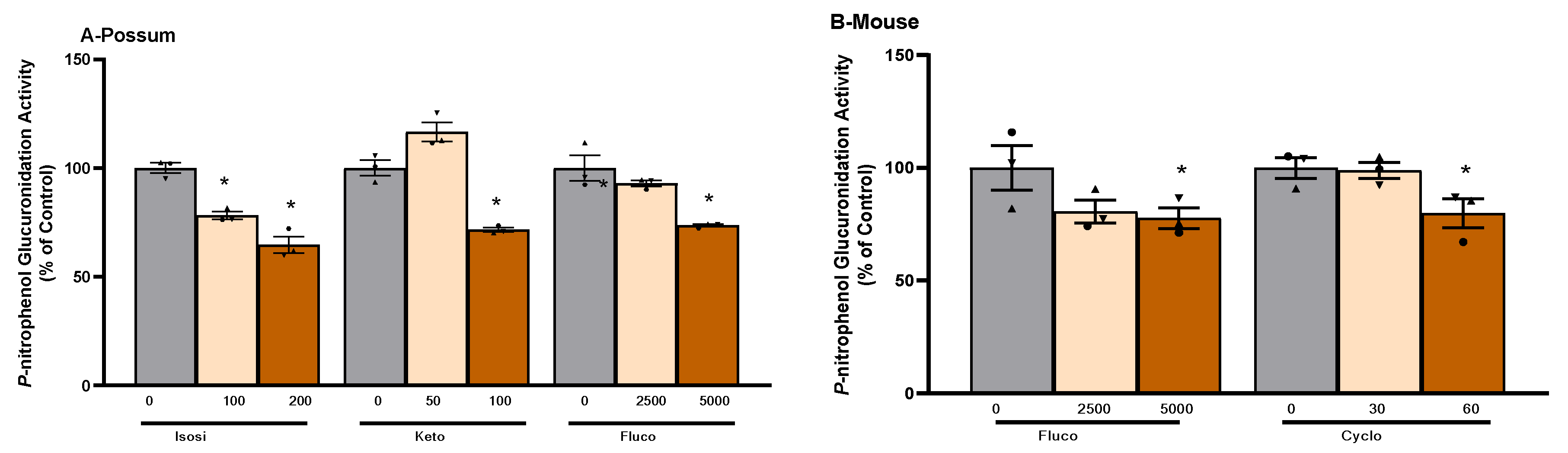

Table 1.
Substrate-dependent effect of isosilybin on p-nitrophenol glucuronidation activity in possum liver microsomes.
Table 1.
Substrate-dependent effect of isosilybin on p-nitrophenol glucuronidation activity in possum liver microsomes.
| Compound |
p-nitrophenol concentration |
Concentration (µM) | Glucuronidation activities+ | Percent of control |
|---|---|---|---|---|
| Isosilybin | 2.5 mM | 0 | 0.65±0.15 | 100 |
| 100 | 0.47±0.14 | 75 | ||
| 200 | 0.72±0.32 | 109 | ||
| Isosilybin | 1.25 mM | 0 | 0.30±0.07 | 100 |
| 100 | 0.15±0.01 | 56 | ||
| 200 | 0.17±0.02 | 70 | ||
| Isosilybin | 0.625 mM | 0 | 0.18±0.02 | 100 |
| 100 | 0.12±0.09 | 63 | ||
| 200 | 0.13±0.05 | 67 |
+P-nitrophenol glucuronidation activity expressed in µmol/mg/min.
3. Discussion
3.1. CYP3A catalytic activity
The basal catalytic activity of CYP3A was up to 2-fold lower in possum than in mouse species. However, there was no significant difference in catalytic activity between possum-avian and mouse-human. The low catalytic activity of CYP3A in possums may mean that the rate of metabolism of contraceptives in possums is likely to be slower than in rodents and humans. For comparison purposes, it should be noted that there are no previous reports in the literature of basal CYP3A catalytic activity in possum using erythromycin N-demethylation, with the exception of the study previously published by our research team [17]. Our previous study also showed that the basal hepatic CYP3A catalytic activity was approximately 20% lower in possums than in rats and mice. In addition, Hunt, et al. [18] reported similar CYP3A catalytic activity (~0.52 nmol/min/mg) in a similar age range (27-60 years) as in the present study in pooled human liver microsomes. While differences (~1.5-fold higher) in the CYP3A catalytic activity have been reported in similar species and age groups of mouse liver microsomes [19]. Furthermore, in the present study, up to 2-fold lower CYP3A catalytic activity was observed in closely related wild mallard species (10 weeks old) compared to avian species [20]. Although differences in CYP3A activity are reported in the literature, there are no studies that have specifically compared possum CYP3A catalytic activity to that of rodents and humans, making it difficult to draw conclusions about those observed in the present. However, it is clear from the present study and our previously published data that possums have a lower rate of CYP3A catalytic activity, and therefore the rate of metabolism of oral contraceptives is likely to be slower than rodents and humans.
Western blotting showed that CYP3A4 protein levels varied between the 4 species examined. Interestingly, possum liver had higher levels of hepatic CYP3A4 protein than avian (~4-fold), mouse (~2-fold), and human (~2-fold), similar to our previous study in possums and rodents [17]. The high CYP3A protein levels in possums could be due to the use of a mammalian CYP3A polyclonal antibody, which may have also detected other highly homologous isoforms of CYP3A in possums. This can be further supported by a previous study that reported that possums express at least three different CYP3A-like isoforms (CYP3A P1, P2 and P3) in their liver and duodenum [21]. However, it is unknown how similarly these CYP3A-like isoforms behave in terms of catalytic activity and subsequently contribute to the observed discrepancy in protein levels and enzyme activity.
A CYP450 inhibitor-based compound library was then screened in test species to identify possum-specific differences in inhibition of CYP3A catalytic activity. The majority of CYP450 inhibitor-based compounds failed to significantly reduce CYP3A catalytic activity in test species compared to their respective controls. Ketoconazole was the only compound that significantly reduced mouse CYP3A catalytic activity at the estimated IC50 and 2-fold IC50 values compared to their respective controls. Notably, none significantly reduced possum CYP3A catalytic activity at 1 and 5 µM or the estimated IC50 and 2-fold IC50 values. However, compounds such as ritonavir, alizarin, galeterone, cobicistat, gentiopicroside, chlorzoxazone, benzbromarone, terfenadine, 7-hydroxyflavone, WAY-325412, WAY-325945, and WAY-657644 showed the potential to reduce the catalytic activity of possum CYP3A to some degree (i.e., reduced activity to ~35% of control, mainly at 5 µM or an estimated 2-fold IC50) but these reduced catalytic activities were not significantly different. None of these compounds (except ketoconazole) have previously been evaluated for their inhibitory effect on CYP3A or UGT2B enzyme activities in possums. Ketoconazole previously inhibited cineole metabolism in wild possum liver microsomes [22]. The same study reported that the formation of all hydroxycineole metabolites (9-, 3-, 7-hydroxycineole) in possum was reduced to approximately 25% of control by 5 µM ketoconazole. In the present study, ketoconazole (5 µM) similarly reduced the catalytic activity of possum CYP3A, as determined by erythromycin N-demethylation, by approximately 27% compared to control, but this was not statistically significant, which may be mainly due to inter-individual differences. However, ketoconazole (2.5 and 5 µM) significantly reduced mouse CYP3A by <50% of control. At similar concentrations, ketoconazole showed only 80% of control in pooled human liver microsomes. Another screen of ketoconazole at a concentration of 1.25 µM showed a reduced activity of ~64% of control in pooled human liver microsomes. However, a previous study showed that ketoconazole reduced erythromycin N-demethylation by ~50% of control at a much lower concentration (0.07 µM) in pooled human liver microsomes [23].
Ritonavir, on the other hand, showed reduced possum CYP3A catalytic activity (81% of control) at 2-fold IC50 (0.28 µM), similar to that in the mouse (84% of control), but not in humans (95% of control). Ritonavir has been shown to reduce the metabolism of triazolam (a probe substrate of the CYP3A enzyme) by ~50% at a concentration of 0.014 µM in pooled human liver microsomes [24]. But when ritonavir was tested for inhibitory activity at 0.014 µM using erythromycin as substrate, the activity was 95% of the control. In addition, cobicistat also showed potential reduction in CYP3A activity in all test species: avian (63%), mouse (68%), possum (76%), and human (78%), mainly at 2-fold IC50 (or 0.06 µM). Hossain, et al. [24] showed that cobicistat reduced the activity of triazolam metabolism by ~50% at 0.032 µM in pooled human liver microsomes. However, the present study found that cobicistat at 0.032 µM reduced erythromycin N-demethylation metabolism to ~78% of control in pooled human liver microsomes. In addition, Tsujimoto, et al. [25] reported that testosterone metabolism in pooled human liver microsomes was inhibited by 7-hydroxyflavone with an IC50>100 µM. However, our study with 7-hydroxyflavone at estimated IC50 and 2-fold IC50 values (or 100 and 200 µM) showed activity that was >83% of control in possum, mouse, avian, and human.
3.2. P-nitrophenol glucuronidation activity
This study found that possum had significantly higher basal UGT2B glucuronidation activity than avian (~8-fold). It should be noted that there are no previous reports in the literature of basal UGT2B glucuronidation activity in possums using p-nitrophenol glucuronidation, except for the study previously published by our research team [17]. In that study, possums had ~2-fold higher UGT catalytic activity compared to rodents, consistent with the findings of the present study. The higher basal p-nitrophenol glucuronidation activity may indicate that possums have a greater ability to metabolise phenolic compounds (including estrogens) in their liver. This higher p-nitrophenol glucuronidation in possum liver microsomes could also explain why marsupial folivores such as the koala can rapidly excrete higher amounts (2–3 g) of glucuronic acids per day than humans (~1000-fold difference) [26,27]. Future studies could therefore focus on examining the role of the possum UGT2B enzymes and their likely association with the greater excretion of glucuronic acids observed in koalas.
In addition, the basal p-nitrophenol glucuronidation activity in mouse was within the range reported in the literature for closely related species: 2.8-2.9 µmol/min/mg in 5-6 week old female Swiss Webster mouse [28]. However, p-nitrophenol glucuronidation activity in humans and avian differed significantly from those reported in the literature. Kuhn, et al. [29] reported a lower p-nitrophenol glucuronidation activity in pooled human liver microsomes in the range of 0.025-0.38 µmol/min/mg: an approximately 9-fold difference compared to the present study. Similarly, Short, et al. [30] reported that p-nitrophenol glucuronidation in wild pekin ducks was 2.55±1.36 nmol/mg/protein/min, approximately 192-fold higher compared to the present study in avian species. No studies in the literature have compared basal UGT2B glucuronidation activity in possums and other test species. But overall, from the present study, the low catalytic activity of CYP3A and the high glucuronidation activity of UGT2B in possums need to be further investigated to understand how these differences in the enzymatic activities of CYP3A and UGT2B enzymes likely contribute to the metabolism and excretion of estrogen-based contraceptives.
Western blot analysis showed that possum UGT2B4 protein levels were ~3-fold lower than pooled human liver microsomes. The possum UGT2B4 protein levels also differed by ~15% compared to the mouse. Although UGTs are highly homologous across species, their glucuronidation activity and protein content most likely differ due to several factors, as discussed later in subsection 3.3. It is therefore important that future research also consider the assessment of such factors in order to understand the extent to which these factors can affect UGT2B activity and protein levels, particularly in possums.
In addition, the compound library used for CYP3A inhibitors was further screened to determine differences in inhibition of UGT2B activity. None of the compounds screened significantly reduced p-nitrophenol glucuronidation activity at estimated IC50 values. However, some reduction in possum p-nitrophenol glucuronidation activity was mainly observed at much higher concentrations (2-fold IC50 values) by compounds such as isosilybin (65% of control at 200 µM), ketoconazole (72% control at 100 µM), and fluconazole (74% control at 5 mM). None of these compounds have previously been assessed for their potential UGT2B inhibitory effects in possums or other marsupial species. However, these compounds have previously been studied for their UGT inhibitory activities with different enzyme sources and substrate choices in human samples. For example, Gufford, et al. [38] reported that silybin B and isosilybin B (both regioisomers of the parent silybin) reduced the glucuronidation of 4-methylumbelliferone (which is known to be metabolised by several UGT isoforms of the UGT1 and UGT2B families) by ~50% of the control in pooled human liver and intestinal microsomes at concentrations of 87 µM and 187 µM, respectively. In the present study, isosilybin showed no significant inhibition of p-nitrophenol glucuronidation in pooled human liver microsomes up to 200 µM. In addition, ketoconazole showed a broad UGT inhibitory effect on human Corning supersomes (recombinant forms produced from baculovirus-transfected insect cells) [39]. The results showed that ketoconazole reduced the 4-methylumbelliferone glucuronidation activity of UGT2B4, UGT1A10, UGT1A8, and UGT1A7 by ~50% at concentrations of 12, 20, >100, and 8 µM, respectively. Based on these findings, ketoconazole was screened in the present study for its inhibitory effect on p-nitrophenol glucuronidation at 50 and 100 µM. Ketoconazole did not significantly reduce UGT2B activity to less than 50% of control in any test species at concentrations up to 100 µM. Similarly, other potential inhibitor compounds showed no significant inhibition of p-nitrophenol glucuronidation activity at IC50 or 2-fold IC50 values and thus were not further investigated for their potential possum-specific differences in UGT2B enzymes.
3.3. Understanding discrepancies in enzyme activity: Factors affecting CYP3A and UGT2B enzymes in species
The inhibitory activity of screened compounds, as well as the basal catalytic activities and protein content in prepared microsomes of test species on CYP3A and UGT2B enzymes are important factors in drug metabolism and can have significant implications for drug discovery and development [40,41]. Nonetheless, the discrepancies observed in the inhibitory effect of the compounds examined in this study, in contrast to the literature, can be attributed to various factors.
One significant factor that could account for the observed discrepancies in the major drug metabolising enzymes observed in the current study versus the literature could be attributed to the choice of substrate. For example, in the present study, ketoconazole at a concentration of 2.5 µM significantly reduced the N-demethylation activity of erythromycin in mice by ~44%. In another study, ketoconazole reduced CYP3A catalytic activity by ~50% at a concentration of 0.0076-0.025 µM with a different set of substrates (midazolam and triazolam) [42]. The difference in substrate choice between the present study and the reported study showed a >100-fold change in inhibition of the CYP3A enzyme by ketoconazole. Similarly, Stresser, et al. [43] reported 2.1 to 195-fold differences in CYP3A inhibitory activity based on substrate choice (benzyloxyresorufin, 7-benzyloxy-4-trifluoromethylcoumarin, 7-benzyloxyquinoline, and dibenzyl fluorescein) in human liver microsomes. These examples therefore suggest that the choice of substrate used to assay inhibition of drug metabolizing enzymes can significantly affect the results observed. Furthermore, another study suggested that the intricate action of substrates on CYP3A enzymes could be attributed to their interactions within the CYP3A enzyme active site, which could vary depending on the concentration of substrates used and ultimately affect their inhibitory potential [44]. Therefore, it is recommended that future studies use multiple probe substrates at different concentrations to comprehensively assess the inhibitory potential of CYP3A enzymes. Although a single probe substrate specific for CYP3A enzymes was used in the present study to assess species differences, the use of multiple probe substrates would allow for a better understanding of how different substrates affect CYP3A inhibition in in vitro models.
Furthermore, it is worth noting that determining the enzymatic activities of UGTs can present even greater difficulties when compared to CYP3As. This is primarily due to the high homology of UGT enzymes and the overlap of their substrates [45,46]. Even the design and development of fluorescent probes for UGTs is very difficult due to the lack of a full UGT crystal structure, making it difficult to rationally design the candidate substrates [46]. Although UGT2B enzymes were specifically targeted in the present study using p-nitrophenol as the probe substrate due to their reported high expression in liver microsomes, it is important to note that p-nitrophenol is also known to be metabolized by the UGT1 subfamily [45,47,48,49]. Previous studies have also emphasised the overlapping inhibitory effects of different compounds on UGT2B enzymes. For example, the inhibitory effect of diclofenac, amitriptyline, quinidine and fluconazole on the enzymatic activities of human UGT2B7 was observed with an IC50 between 82 and 2,500 μM [37]. However, these compounds also showed inhibitory effects on other UGTs such as UGT1A1, UGT1A9, and UGT1A6. Similarly, bisphenol A (an endocrine disrupting chemical) showed both competitive (UGT2B4) and non-competitive (UGT2B7, UGT2B15, and UGT2B17) inhibition [50]. Interestingly, this was observed in a recombinant UGT-catalysed 4-methylumbelliferone (4-MU) glucuronidation reaction in vitro with inhibition kinetics (Ki) ranging from 1-20 μM. Therefore, caution should be exercised in interpreting the results of UGT enzyme activities, even in the present study, since other subfamilies of UGT enzymes may also contribute to the overall p-nitrophenol glucuronidation activity. Future research should aim to develop techniques that can assess the activity of specific UGT isoforms to better understand their role in drug metabolism.
In addition, it is important to note that the use of prepared microsomal samples for different species in the present study may also have resulted in discrepancies in the basal and inhibitory activities of the test compounds for obvious reasons. The discrepancies resulted in the present study could have been influenced by various factors, including but not limited to genetic factors [33,34,51,52,53,54,55,56,57,58], environment or exposure to xenobiotics [36,59,60,61] and, physiological [62,63,64,65,66,67,68], all of which can affect the activity of these important drug metabolising enzymes. It is therefore important to consider such factors and exercise caution when interpreting the results and drawing conclusions about the inhibitory activity of the compounds being screened. This will help to better understand the potential effects of different compounds on drug metabolism that can influence the drug discovery and development process.
4. Materials and Methods
4.1. Compound selection
Bovine serum albumin (BSA), dimethyl sulfoxide (DMSO), nicotinamide adenine dinucleotide phosphate (NADP+), and uridine 5′-diphosphoglucuronic acid trisodium salt (UDPGA) were purchased from Sigma-Aldrich (St. Louis, MO, USA). The following primary antibodies were purchased; CYP3A polyclonal antibody (PA5-14896, Invitrogen, Waltham, MA, USA), UGT2B4 (PA5-92155, Invitrogen, Waltham, MA, USA), and GAPDH (Sigma-Aldrich, Sydney, Australia; a subsidiary of Merck KGaA, Darmstadt, Germany, and SHOP, Abcam Australia Pty Ltd). Fifty-nine CYP450 inhibitor-based compounds (L2000) were obtained from SelleckChem (Houston, USA). These compounds were selected using the SelleckChem database and has focused on the key structural moieties of potent CYP3A inhibitors. In particular, the search for compounds focused on heteroatoms/heterocyclic moieties (such as imidazole, pyridine, triazole) that allow coordination to the iron in CYP3A enzymes [69]. The compound search also included carboxylate ions and aromatic rings. The aim was to improve the interactions between compound and enzyme via chemical bonds and ionic interactions. Other reagents used were commercially available in the highest purity.
4.2. Animals
Possums were collected from the Otago Peninsula (Dunedin) and the Okuti Valley, Banks Peninsula (Christchurch) in New Zealand in collaboration with the Otago Peninsula Biodiversity Group and Manaaki Whenua - Landcare Research, respectively. Possum trapping and liver harvesting were approved by the Peninsula Biodiversity Committee and Manaaki Whenua - Landcare Research. Possums were caged, humanely euthanised, and the livers harvested, snap frozen in liquid nitrogen, and stored in a freezer at -80°C until preparation of microsomal fractions. BALB/c mice (7–9 weeks old) were purchased from Hercus Taieri Resource Unit (Dunedin, New Zealand). Mice were euthanised by carbon dioxide inhalation, and the livers were removed and microsomes prepared. The use of mice was approved by University of Otago animal ethics committee #18/224. The livers from avian species (mallards and a swan) were kindly donated by a local bird hunting group based in Dunedin, New Zealand. The livers from avian species were treated similarly as mentioned above for the possum samples. Human liver microsomes (HLM) pooled from 50 donors at a protein content of 20 mg/ml were purchased from Thermo Fisher Scientific (Waltham, USA).
4.3. Preparation of mouse, possum, and avian liver microsomes
Microsomal preparation was carried out as previously described [70]. Individual livers were placed in beakers containing 1.15% KCl. All procedures were performed at 0-4°C. The liver samples were minced and homogenised in Buffer A (0.1 M Tris, 0.1 M potassium chloride, 1 mM EDTA, and 20 μM butylated hydroxytoluene, at pH=7.4) using the Teflon-glass homogeniser (5-6 vertical passes). The samples were centrifuged at 10,000 g for 20 min. The supernatant was then transferred into another set of centrifuge tubes and spun at 100,000 g for 60 min. The pellet was resuspended in Buffer B (0.1 M potassium pyrophosphate, 1 mM EDTA, and 20 μM butylated hydroxytoluene, at pH=7.4), homogenised, and recentrifuged at 100,000 g for 60 min. Finally, the pellet was homogenised in 1-2 mL of Buffer C (10 mM Tris-HCl, 1 mM EDTA, and 4% glycerol (v/v)), and stored at -80°C until use. The protein concentration of the microsomes was determined using the bicinchoninic acid (BCA) method [71].
4.4. CYP3A catalytic activity
CYP3A catalytic activity was determined using the erythromycin N-demethylation activity assay as described [72]. The reaction mixture (final volume of 1 mL and final protein concentration of 1 mg) contained 500 µL of microsomes in 0.1 M phosphate buffer (pH 7.4), 400 µL of erythromycin buffer (0.45 mM), and 50 µL of the test compound was pre-warmed at 37°C for 2 min. The reaction was initiated with 50 µL of 2.5 mM nicotinamide adenine dinucleotide phosphate (NADP+) and incubated for 30 min at 37°C. The reaction was stopped by the addition of 330 µL of 15% zinc sulphate. The tubes were vortexed and left for 5 min (at room temperature), and then 330 µL of saturated barium hydroxide was added. The tubes were further vortexed and left for 5 min (at room temperature). The tubes were then centrifuged at 1, 800 g for 10 min. To 830 µL of the supernatant, 330 µL of Nash reagent [30% (w/v) ammonium acetate, 0.4% (v/v) acetylacetone] was added and further incubated for 30 min at 60°C. The blank tube contained 330 µL of Nash reagent and 830 µL of Milli-Q water. The tubes were recentrifuged at 1, 800 g for 10 min, and the absorbance was read at 415 nm. The CYP3A activities were determined in nmol/mg/min using standard curve data. Results presented are the mean ± SEM of 3 independent experiments performed in duplicate unless otherwise stated.
4.5. Glucuronidation of p-nitrophenol
The rate of p-nitrophenol glucuronidation was determined using the optimised protocol [73,74]. Briefly, the incubation mixture (500 µL in total) contained 100 µL of 1 mg/mL microsomes, 50 µL of 1 M Tris-HCl (pH=7.4), 20 µL of 0.25% (v/v) Triton-X 100, 50 µL of 50 mM MgCl2, 130 µL of Milli-Q water, 50 µL of 5 mM p-nitrophenol, 50 µL of the test compound, and 50 µL of 30 mM uridine 5′-diphosphoglucuronic acid trisodium salt (UDPGA). The blank tubes contained all components except UDPGA and microsomes; instead, 150 µL of 0.1 M phosphate buffer was added to equal the volume as the reaction mixture. The reaction mixture and blank were incubated at 37°C for 2 min. 50 µL of 30 mM UDPGA was added to initiate the reaction, which proceeded at 37°C for 30 min. The reaction was stopped using 1 mL of 5% trichloroacetic acid (TCA). The tubes were centrifuged at 1, 800 g for 10 min and then 1 mL of supernatant was added to 250 µL of 2 M sodium hydroxide (NaOH). The absorbance was read at 405 nm. The sample value was subtracted from the blank absorbance and divided by the extinction coefficient of p-nitrophenol (18.1 x 103 cm2/mole) and the spectrophotometer light path (1 cm). Glucuronidation activities were expressed in µmol/mg/min and are the mean ± SEM of 3 independent experiments performed in duplicate unless otherwise stated.
4.6. Western blotting
The microsomes were diluted to 1 mg/mL in 4X Laemmli buffer (62.5 mM Tris-HCl, 1% sodium dodecyl sulphate, 10% glycerol, 0.005% bromophenol blue, and 355 mM β-mercaptoethanol). The samples were then heated to 95°C for 5 min and stored at -20°C. Gel electrophoresis was performed as previously described [75]. All gel casting and running were performed using a Mini PROTEAN® (BioRad). The samples were heated to 37°C before loading 1-2 µg of protein onto a 7.5% acrylamide gel (acrylamide, 1.5 M lower Tris, 50% glycerol, 10% ammonium persulfate, and tetramethylethylenediamine), and Precision Plus Protein Dual Colour Standards (Bio-Rad Laboratories, Hercules, CA, USA) were loaded as the protein ladder. Empty wells were loaded with 15 μL of 1X sample buffer. The gels were run at 80 V for protein stacking (~15 min) and then at 120 V for protein resolution (~1.5 h). After separation, proteins were transferred to a PVDF membrane (Merck Millipore Ltd., New Zealand) in a transfer buffer at 100 V for 60 min. The membranes were blocked in 5% non-fat milk and TBST (Tris-buffered saline with 1% Triton-X 100) for 1 hr. The membranes were then incubated with primary antibodies in TBS (Tris-buffered saline) and left on a shaker overnight at 4°C. The following antibodies were used: CYP3A4 polyclonal antibody (1:1000, PA5-14896, Invitrogen, USA), UGT2B4 (1:1000, PA5-92155, Invitrogen, USA), and GAPDH (1:2,000, Sigma-Aldrich, Australia). The membranes were then stripped and washed with TBST buffer (6 x 5 min). Horseradish peroxidase-conjugated secondary antibodies (anti-rabbit for CYP3A4 and UGT2B4, and anti-mouse for GAPDH) in TBS and milk powder were added to the membranes and incubated for 45 min at room temperature. The membranes were re-washed (6 x 5 min) with TBST buffer before the addition of the SuperSignal West Pico chemiluminescent substrate or Femto (ThermoFisher, Albany, New Zealand), and blots were visualised using a CL-XPosure film (ThermoFisher, Albany, New Zealand).
4.7. Statistical analysis
Statistical significance was assessed using a one-way ANOVA with Dunnett’s post hoc test, where p<0.05 was the minimum requirement for a statistically significant difference. All graphs and statistical analysis were performed using GraphPad Prism software version 9.0 for Windows (GraphPad Software, San Diego, CA, USA).
5. Conclusions
This is one of the first in vitro studies to investigate how inhibition of key major drug-metabolising enzymes (CYP3A and UGT2B) in possums could be viewed as a tool to improve the bioavailability of future oral contraceptives for an improved hormonal approach to fertility control. This study confirms that the basal catalytic activity of CYP3A and UGT2B in possums differs from other test species. Specifically, possums had higher CYP3A4 protein levels compared to other test species (up to a 4-fold difference) and significantly higher basal p-nitrophenol glucuronidation activity than other test species (up to an 8-fold difference). None of the CYP450 inhibitor-based compounds significantly decreased the catalytic activity of possum CYP3A and UGT2B below the estimated IC50 and 2-fold IC50 values. Some reduction (>65% of control) in p-nitrophenol glucuronidation activity by compounds such as isosilybin, ketoconazole, and fluconazole has been observed in possums. However, when some of these compounds were tested further, they failed to significantly reduce enzyme activity below the estimated IC50 and 2-fold IC50 values. There could be many factors that could have influenced the enzymatic activities observed in the present study, some of which may include genetics, enzyme source, preparation, selection of a probe substrate, or the design of in vitro studies. Furthermore, research could also address the development of techniques to isolate and characterise specific isoforms (even to develop recombinant enzymes) to selectively study the unique pattern of major drug-metabolising enzymes in possums. Much of the work in this area is still limited and requires further investigation to uncover the details of its contribution to the study of drug metabolism. Although the literature suggests that these enzymes are highly conserved across species, the development of recombinant forms of these enzymes could aim to provide a thorough understanding for a more effective design of possum-specific inhibitors.
Supplementary Materials
Supplementary Table 1: CYP3A catalytic activity in possum and mouse liver microsomes. (n=1). Supplementary Table 2: CYP3A catalytic activities of selected compounds in mouse, possum, avian, and human liver microsomes. Supplementary Table 3: Selected compounds screened for p-nitrophenol inhibitory activities in 4 species at estimated IC50 and 2-fold IC50 values.
Author Contributions
Conceptualization, R.R.C., R.J.R. and B.C.; methodology, R.R.C., M.N. and R.J.R.; software, R.R.C. and M.N.; investigation and analysis, R.R.C. and M.N.; writing—original draft preparation, R.R.C.; writing—review and editing; R.R.C., M.N. and R.J.R.; supervision, R.J.R.; funding acquisition, B.C. and R.R.C. R.R.C was supported by a University of Otago PhD scholarship. All authors have read and agreed to the published version of the manuscript.
Funding
This work received funding from the Operational Solutions for Primary Industries (OSPRI) (Grant number: PL.115115. 01.P. LM). This study was also supported by the Kathleen Spragg Agricultural Research Trust Grant (PCS-iManPCS.FID619804).
Institutional Review Board Statement
The animal work was conducted according to the guidelines approved by the Manaaki Whenua—Landcare Research Animal Ethics Committee and the Otago University Animal Ethics Committee.
Informed Consent Statement
Not applicable
Data Availability Statement
The data presented in this study are reported in the results and Supplementary Material.
Conflicts of Interest
The authors have no conflict of interest.
References
- Owens, B. Behind New Zealand’s wild plan to purge all pests. Nature News 2017, 541, 148. [Google Scholar] [CrossRef]
- Chand, R.R.; Cridge, B.J. Upscaling Pest Management From Parks to Countries: A New Zealand Case Study. Journal of Integrated Pest Management 2020, 11, 1–12. [Google Scholar] [CrossRef]
- D’Arpe, S.; Di Feliciantonio, M.; Candelieri, M.; Franceschetti, S.; Piccioni, M.G.; Bastianelli, C. Ovarian function during hormonal contraception assessed by endocrine and sonographic markers: a systematic review. Reproductive Biomedicine Online 2016, 33, 436–448. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Luo, R.; Wang, H.; Cao, G.; Wang, Y. Recovery of fertility in quinestrol-treated or diethylstilbestrol-treated mice: Implications for rodent management. Integrative Zoology 2017, 12, 250–259. [Google Scholar] [CrossRef] [PubMed]
- Christin-Maitre, S. History of oral contraceptive drugs and their use worldwide. Best Practice & Research Clinical Endocrinology & Metabolism 2013, 27, 3–12. [Google Scholar] [CrossRef]
- Hynes, E.F.; Handasyde, K.A.; Shaw, G.; Renfree, M.B. Levonorgestrel, not etonogestrel, provides contraception in free-ranging koalas. Reproduction, Fertility, and Development 2010, 22, 913–919. [Google Scholar] [CrossRef] [PubMed]
- Ballantyne, K.; Anderson, S.T.; Pyne, M.; Nicolson, V.; Mucci, A.; Lisle, A.; Johnston, S.D. The use of a synthetic progesterone, levonorgestrel (LNG), to control the oestrous cycle in the koala. Reproduction, Fertility, and Development 2015, 28, 1526–1532. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Nakajima, T.; Hayashi, S.; Chambon, P.; Watanabe, H.; Iguchi, T.; Sato, T. Effects of diethylstilbestrol on programmed oocyte death and induction of polyovular follicles in neonatal mouse ovaries. Biology of Reproduction 2009, 81, 1002–1009. [Google Scholar] [CrossRef]
- Meyer, N.; Santamaria, C.G.; Müller, J.E.; Schumacher, A.; Rodriguez, H.A.; Zenclussen, A.C. Exposure to 17α-ethinyl estradiol during early pregnancy affects fetal growth and survival in mice. Environmental Pollution 2019, 251, 493–501. [Google Scholar] [CrossRef]
- Linhart, S.B.; Enders, R.K. Some effects of diethylstilbestrol on reproduction in captive red foxes. The Journal of Wildlife Management 1964, 358–363. [Google Scholar] [CrossRef]
- Harder, J.D.; Peterle, T.J. Effect of diethylstilbestrol on reproductive performance of white-tailed deer. The Journal of Wildlife Management 1974, 183–196. [Google Scholar] [CrossRef]
- Stegeman, B.H.; Vos, H.L.; Helmerhorst, F.M.; Rosendaal, F.R.; Reitsma, P.H.; van Hylckama Vlieg, A. Genetic variation in the first-pass metabolism of ethinylestradiol, sex hormone binding globulin levels and venous thrombosis risk. European Journal of Internal Medicine 2017, 42, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Fotherby, K. Bioavailability of orally administered sex steroids used in oral contraception and hormone replacement therapy. Contraception 1996, 54, 59–69. [Google Scholar] [CrossRef] [PubMed]
- De Lignieres, B.; Basdevant, A.; Thomas, G.; Thalabard, J.C.; Mercier-Bodard, C.; Conard, J.; Guyene, T.T.; Mairon, N.; Corvol, P.; Guy-Grand, B.; et al. Biological effects of estradiol-17 beta in postmenopausal women: oral versus percutaneous administration. Journal of Clinical Endocrinology and Metabolism 1986, 62, 536–541. [Google Scholar] [CrossRef] [PubMed]
- Mahanur, V.; Rajge, R.; Tawar, M. A review on emerging oral dosage forms which helps to bypass the hepatic first pass metabolism. Asian Journal of Pharmacy Technology 2022, 12, 1–14. [Google Scholar] [CrossRef]
- Rhoda Lee, C. Drug interactions and hormonal contraception. Trends in Urology Gynacology and Sexual Health 2009, 14, 23–26. [Google Scholar] [CrossRef]
- Chand, R.R.; Nimick, M.; Cridge, B.; Rosengren, R.J. In vitro hepatic assessment of cineole and its derivatives in common brushtail possums (Trichosurus vulpecula) and rodents. Biology 2021, 10, 1326. [Google Scholar] [CrossRef]
- Hunt, C.M.; Westerkam, W.R.; Stave, G.M. Effect of age and gender on the activity of human hepatic CYP3A. Biochemical Pharmacology 1992, 44, 275–283. [Google Scholar] [CrossRef]
- Bland, A.R.; Shrestha, N.; Rosengren, R.J.; Ashton, J.C. Does Crizotinib Auto-Inhibit CYP3A in vivo? Pharmacology 2020, 105, 715–718. [Google Scholar] [CrossRef]
- Raynal, M.; Bailly, J.; Benard, G.; Guerre, P. Effects of fumonisin B1 present in Fusarium moniliforme culture material on drug metabolising enzyme activities in ducks. Toxicology letters 2001, 121, 179–190. [Google Scholar] [CrossRef]
- Sorensen, J.S.; Forbey, K.C.; Tanquay, R.L.; McLeod, B. Tissue distribution of cytochrome P450 3A (CYP3A) in brushtail possums (Trichosurus vulpecula) exposed to Eucalyptus terpenes. Comparative Biochemistry and Physiology Part C: Pharmacology, Toxicology and Endocrinology 2007, 145, 194–201. [Google Scholar] [CrossRef]
- Pass, G.J.; McLean, S. Inhibition of the microsomal metabolism of 1,8-cineole in the common brushtail possum (Trichosurus vulpecula) by terpenes and other chemicals. Xenobiotica 2002, 32, 1109–1126. [Google Scholar] [CrossRef] [PubMed]
- Iwata, H.; Tezuka, Y.; Kadota, S.; Hiratsuka, A.; Watabe, T. Identification and characterization of potent CYP3A4 inhibitors in Schisandra fruit extract. Drug Metabolism and Disposition 2004, 32, 1351–1358. [Google Scholar] [CrossRef]
- Hossain, M.A.; Tran, T.; Chen, T.; Mikus, G.; Greenblatt, D.J. Inhibition of human cytochromes P450 in vitro by ritonavir and cobicistat. Journal of Pharmacy and Pharmacology 2017, 69, 1786–1793. [Google Scholar] [CrossRef] [PubMed]
- Tsujimoto, M.; Horie, M.; Honda, H.; Takara, K.; Nishiguchi, K. The structure–activity correlation on the inhibitory effects of flavonoids on cytochrome P450 3A activity. Biological Pharmaceutical Bulletin 2009, 32, 671–676. [Google Scholar] [CrossRef]
- McLean, S.; Brandon, S.; Davies, N.W.; Boyle, R.; Foley, W.J.; Moore, B.; Pass, G.J. Glucuronuria in the Koala. Journal of Chemical Ecology 2003, 29, 1465–1477. [Google Scholar] [CrossRef] [PubMed]
- Lake, B.G.; Longland, R.C.; Harris, R.A.; Gangolli, S.D.; Rundle, A. The excretion of metabolites of the D-glucuronic acid pathway in human urine.: Effect of phenobarbitone administration. Xenobiotica 1982, 12, 241–247. [Google Scholar] [CrossRef]
- Valentine, S.P.; Le Nedelec, M.J.; Menzies, A.R.; Scandlyn, M.J.; Goodin, M.G.; Rosengren, R.J. Curcumin modulates drug metabolizing enzymes in the female Swiss Webster mouse. Life Sciences 2006, 78, 2391–2398. [Google Scholar] [CrossRef]
- Kuhn, U.D.; Rost, M.; Müller, D. Para-nitrophenol glucuronidation and sulfation in rat and human liver slices. Experimental and Toxicologic Pathology 2001, 53, 81–88. [Google Scholar] [CrossRef]
- Short, C.R.; Flory, W.; Hsieh, L.C.; Aranas, T.; Ou, S.P.; Weissinger, J. Comparison of hepatic drug metabolising enzyme activities in several agricultural species. Comparative Biochemistry and Physiology Part C: Comparative Pharmacology 1988, 91, 419–424. [Google Scholar] [CrossRef]
- Strassburg, C.; et al. Developmental aspects of human hepatic drug glucuronidation in young children and adults. Gut 2002, 50, 259–265. [Google Scholar] [CrossRef]
- Tanaka, E. Gender-related differences in pharmacokinetics and their clinical significance. Journal of Clinical Pharmacy and Therapeutics 1999, 24, 339–346. [Google Scholar] [CrossRef]
- Hu, D.G.; Mackenzie, P.I.; McKinnon, R.A.; Meech, R. Genetic polymorphisms of human UDP-glucuronosyltransferase (UGT) genes and cancer risk. Drug Metabolism Reviews 2016, 48, 47–69. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Meng, H. Effects of genetic polymorphism of drug-metabolizing enzymes on the plasma concentrations of antiepileptic drugs in Chinese population. Bioengineered 2022, 13, 7709–7745. [Google Scholar] [CrossRef] [PubMed]
- Toide, K.; et al. Hepatocyte nuclear factor-1alpha is a causal factor responsible for interindividual differences in the expression of UDP-glucuronosyltransferase 2B7 mRNA in human livers, (in eng). Drug Metabolism and Disposition 2002, 30, 613–615. [Google Scholar] [CrossRef]
- Abdullah, N.H.; Ismail, S. Inhibition of UGT2B7 enzyme activity in human and rat liver microsomes by herbal constituents. Molecules 2018, 23, 1–19. [Google Scholar] [CrossRef]
- Donato, M.T.; Montero, S.; Castell, J.V.; Gómez-Lechón, M.J.; Lahoz, A. Validated assay for studying activity profiles of human liver UGTs after drug exposure: inhibition and induction studies. Analytical and Bioanalytical Chemistry 2010, 396, 2251–2263. [Google Scholar] [CrossRef]
- Gufford, B.T.; Chen, G.; Lazarus, P.; Graf, T.N.; Oberlies, N.H.; Paine, M.F. Identification of diet-derived constituents as potent inhibitors of intestinal glucuronidation. Drug Metabolism and Disposition 2014, 42, 1675–1683. [Google Scholar] [CrossRef] [PubMed]
- Yerino, P.; Muranjan, S.; Ogilvie, B.W.; Buckley, D.B. The in vitro evaluation of ketoconazole and its alternative clinical CYP3a4/5 inhibitors (ritonavir, clarithromycin and itraconazole) as inhibitors of non-CYP enzymes. Available online: https://www.xenotech.com/wp-content/uploads/2020/05/ISSX_In-Vitro-Ketoconazole-Clinical-CYP3A45-Inhibitors-Ritonavir-Clarithromycin-Itraconazole-Non-CYP-Enzymes.pdf.
- Lamb, D.C.; Waterman, M.R.; Kelly, S.L.; Guengerich, F.P. Cytochromes P450 and drug discovery. Current Opinion in Biotechnology 2007, 18, 504–512. [Google Scholar] [CrossRef] [PubMed]
- Oda, S.; Fukami, T.; Yokoi, T.; Nakajima, M. A comprehensive review of UDP-glucuronosyltransferase and esterases for drug development. Drug Metabolism and Pharmacokinetics 2015, 30, 30–51. [Google Scholar] [CrossRef]
- Perloff, M.D.; von Moltke, L.L.; Kotegawa, T.; Shader, R.I.; Greenblatt, D.J. Midazolam and triazolam biotransformation in mouse and human liver microsomes: relative contribution of CYP3A and CYP2C isoforms. Journal of Pharmacology and Experimental Therapeutics 2000, 292, 618–628. [Google Scholar]
- Stresser, D.M.; Blanchard, A.P.; Turner, S.D.; Erve, J.C.; Dandeneau, A.A.; Miller, V.P.; Crespi, C.L. Substrate-dependent modulation of CYP3A4 catalytic activity: analysis of 27 test compounds with four fluorometric substrates. Drug Metabolism and Disposition 2000, 28, 1440–1448. [Google Scholar]
- Ueng, Y.F.; Kuwabara, T.; Chun, Y.J.; Guengerich, F.P. Cooperativity in oxidations catalyzed by cytochrome P450 3A4. Biochemistry 1997, 36, 370–381. [Google Scholar] [CrossRef]
- Tukey, R.H.; Strassburg, C.P. Human UDP-glucuronosyltransferases: metabolism, expression, and disease. Annual Review of Pharmacology Toxicology 2000, 40, 581–616. [Google Scholar] [CrossRef] [PubMed]
- Lv, X.; Zhang, J.-B.; Hou, J.; Dou, T.-Y.; Ge, G.-B.; Hu, W.-Z.; Yang, L. Chemical Probes for Human UDP-Glucuronosyltransferases: A Comprehensive Review. Biotechnology Journal 2019, 14, 1–47. [Google Scholar] [CrossRef]
- Izukawa, T.; Nakajima, M.; Fujiwara, R.; Yamanaka, H.; Fukami, T.; Takamiya, M.; Aoki, Y.; Ikushiro, S.-i.; Sakaki, T.; Yokoi, T. Quantitative analysis of UDP-glucuronosyltransferase (UGT) 1A and UGT2B expression levels in human livers. Drug Metabolism and Disposition 2009, 37, 1759–1768. [Google Scholar] [CrossRef] [PubMed]
- Court, M.H.; Zhang, X.; Ding, X.; Yee, K.K.; Hesse, L.M.; Finel, M. Quantitative distribution of mRNAs encoding the 19 human UDP-glucuronosyltransferase enzymes in 26 adult and 3 fetal tissues. Xenobiotica 2012, 42, 266–277. [Google Scholar] [CrossRef]
- Ohno, S.; Nakajin, S. Determination of mRNA expression of human UDP-glucuronosyltransferases and application for localization in various human tissues by real-time reverse transcriptase-polymerase chain reaction. Drug Metabolism and Disposition 2009, 37, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.-M.; Fang, Z.-Z.; Cao, Y.-F.; Hu, C.-M.; Sun, X.-Y.; Hong, M.; Yang, L.; Ge, G.-B.; Liu, Y.; Zhang, Y.-Y. , et al. New insights for the risk of bisphenol A: Inhibition of UDP-glucuronosyltransferases (UGTs). Chemosphere 2013, 93, 1189–1193. [Google Scholar] [CrossRef]
- Wang, D.; Guo, Y.; Wrighton, S.A.; Cooke, G.E.; Sadee, W. Intronic polymorphism in CYP3A4 affects hepatic expression and response to statin drugs. The Pharmacogenomics Journal 2011, 11, 274–286. [Google Scholar] [CrossRef]
- Wang, D.; Sadee, W. CYP3A4 intronic snp rs35599367 (CYP3A4* 22) alters RNA splicing. Pharmacogenetics and Genomics 2016, 26, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Özdemir, V.; Kalow, W.; Tang, B.-K.; Paterson, A.D.; Walker, S.E.; Endrenyi, L.; Kashuba, A.D.M. Evaluation of the genetic component of variability in CYP3A4 activity: a repeated drug administration method. Pharmacogenetics 2000, 10, 373–388. [Google Scholar] [CrossRef]
- Klein, K.; Zanger, U.M. Pharmacogenomics of Cytochrome P450 3A4: Recent Progress Toward the "Missing Heritability" Problem. Frontiers in Genetics 2013, 4, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Rebbeck, T.R.; Jaffe, J.M.; Walker, A.H.; Wein, A.J.; Malkowicz, S.B. Modification of Clinical Presentation of Prostate Tumors by a Novel Genetic Variant in CYP3A4. Journal of the National Cancer Institute 1998, 90, 1225–1229. [Google Scholar] [CrossRef] [PubMed]
- Werk, A.N.; Cascorbi, I. Functional Gene Variants of CYP3A4. Clinical Pharmacology & Therapeutics 2014, 96, 340–348. [Google Scholar] [CrossRef]
- Kliewer, S.A.; Goodwin, B.; Willson, T.M. The Nuclear Pregnane X Receptor: A Key Regulator of Xenobiotic Metabolism. Endocrine Reviews 2002, 23, 687–702. [Google Scholar] [CrossRef]
- Riffel, A.K.; Schuenemann, E.; Vyhlidal, C.A. Regulation of the CYP3A4 and CYP3A7 promoters by members of the nuclear factor I transcription factor family. Molecular Pharmacology 2009, 76, 1104–1114. [Google Scholar] [CrossRef]
- Bolton, R.; Ahokas, J. Mixed function oxidases in an Australian marsupial, the brushtail possum (Trichosurus vulpecula). Archives of Environmental Contamination Toxicology 1997, 33, 83–89. [Google Scholar] [CrossRef]
- Lim, Y.-P.; Kuo, S.-C.; Lai, M.-L.; Huang, J.-D. Inhibition of CYP3A4 expression by ketoconazole is mediated by the disruption of pregnane X receptor, steroid receptor coactivator-1, and hepatocyte nuclear factor 4α interaction. Pharmacogenetics and Genomics 2009, 19, 11–24. [Google Scholar] [CrossRef]
- Kocarek, T.A.; Schuetz, E.G.; Strom, S.C.; Fisher, R.A.; Guzelian, P.S. Comparative analysis of cytochrome P4503A induction in primary cultures of rat, rabbit, and human hepatocytes. Drug Metabolism Disposition 1995, 23, 415–421. [Google Scholar]
- De Wildt, S.N.; Kearns, G.L.; Leeder, J.S.; van den Anker, J.N. Cytochrome P450 3A: ontogeny and drug disposition. Clinical Pharmacokinetics 1999, 37, 485–505. [Google Scholar] [CrossRef] [PubMed]
- Warrington, J.S.; Greenblatt, D.J.; von Moltke, L.L. Age-related differences in CYP3A expression and activity in the rat liver, intestine, and kidney. Journal of Pharmacology Experimental Therapeutics 2004, 309, 720–729. [Google Scholar] [CrossRef]
- Wolbold, R.; Klein, K.; Burk, O.; Nüssler, A.K.; Neuhaus, P.; Eichelbaum, M.; Schwab, M.; Zanger, U.M. Sex is a major determinant of CYP3A4 expression in human liver. Liver Failure and Liver Disease 2003, 38, 978–988. [Google Scholar] [CrossRef]
- El-Merhibi, A.; Ngo, S.N.; Crittenden, T.A.; Marchant, C.L.; Stupans, I.; McKinnon, R.A. Cytochrome P450 CYP3A in marsupials: cloning and characterisation of the second identified CYP3A subfamily member, isoform 3A78 from koala (Phascolarctos cinereus). Comparative Biochemistry and Physiology C 2011, 154, 367–376. [Google Scholar] [CrossRef] [PubMed]
- Stern, S.T.; Tallman, M.N.; Miles, K.K.; Ritter, J.K.; Dupuis, R.E.; Smith, P.C. Gender-related differences in mycophenolate mofetil-induced gastrointestinal toxicity in rats. Drug Metabolism and Disposition 2007, 35, 449–454. [Google Scholar] [CrossRef]
- Takeuchi, T.; Tsutsumi, O.; Nakamura, N.; Ikezuki, Y.; Takai, Y.; Yano, T.; Taketani, Y. Gender difference in serum bisphenol A levels may be caused by liver UDP-glucuronosyltransferase activity in rats. Biochemical and Biophysical Research Communications 2004, 325, 549–554. [Google Scholar] [CrossRef]
- Muraca, M.; Fevery, J. Influence of sex and sex steroids on bilirubin uridine diphosphate-glucuronosyltransferase activity of rat liver. Gastroenterology 1984, 87, 308–313. [Google Scholar] [CrossRef]
- Shu, Y.-Z.; Johnson, B.M.; Yang, T.J. Role of biotransformation studies in minimizing metabolism-related liabilities in drug discovery. The AAPS journal 2008, 10, 178–192. [Google Scholar] [CrossRef]
- Lake, B.G. Preparation and characterization of microsomal fractions for studies on xenobiotic metabolism. In Biochemical Toxicology: A Practical Approach, Snell, K., Mullock, B., Eds. IRL Press: Oxford, 1987; pp. 183–215.
- Smith, P.e.; Krohn, R.I.; Hermanson, G.T.; Mallia, A.K.; Gartner, F.H.; Provenzano, M.; Fujimoto, E.K.; Goeke, N.M.; Olson, B.J.; Klenk, D. Measurement of protein using bicinchoninic acid. Analytical biochemistry 1985, 150, 76–85. [Google Scholar] [CrossRef]
- Kitada, M.; Igoshi, N.; Kamataki, T.; Itahashi, K.; Imaoka, S.; Komori, M.; Funae, Y.; Rikihisa, T.; Kanakubo, Y. Immunochemical similarity of P-450 HFLa, a form of cytochrome P-450 in human fetal livers, to a form of rat liver cytochrome P-450 inducible by macrolide antibiotics. Archives of Biochemistry and Biophysics 1988, 264, 61–66. [Google Scholar] [CrossRef]
- Fowler, B.; Kleinow, K.; Squibb, K.; Lucier, G.; Hayes, A. Organelles as tools in toxicology. In Principles of Clinical Toxicology, A.W, H., Ed. CRC Press: New York, UK, 1994; pp. 1201–1230.
- Henderson, P.T.; Kersten, K.J. Metabolism of drugs during rat liver regeneration. Biochemical Pharmacology 1970, 19, 2343–2351. [Google Scholar] [CrossRef] [PubMed]
- Towbin, H. Electrophoretic transfer of proteins from sodium dodecyl sulfate-polyacrylamide gels to unmodified nitrocellulose and radiographic detection with antibody and radioiodinated protein. Analytical Biochemistry 1979, 112, 195–203. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



