
Submitted:
16 April 2023
Posted:
18 April 2023
You are already at the latest version
Abstract
Migraine is a neurovascular disorder that affects approximately 12% of the global population. While its exact causes are still being studied, researchers believe that nociceptors in the trigeminal ganglia play a key role in the pain signals of migraine. These nociceptors innervate the intracranial meninges and convey pain signals from the meninges to the thalamus. Targeting these nociceptors is considered promising due to their accessibility and distinct molecular profile, which includes the expression of several transient receptor potential (TRP) channels. These channels have been linked to various pain conditions, including migraine. This review discusses the role and mechanisms of nociceptors in migraine, the challenges of current antimigraine drugs, and the evidence for well-studied and emerging TRP channels, particularly TRPC4, as novel targets for migraine prevention and treatment.
Keywords:
migraine
; headache
; peripheral sensory neurons
; transient receptor potential channels
; TRPC4
; CGRP
1. Introduction
Approximately 12% of the adult global population suffer from migraine [1], a disorder characterized by recurrent, severe, one-sided throbbing headaches that can be accompanied by nausea, vomiting, photophobia, and cutaneous allodynia [2]. Although the exact causes of migraine are still being studied and debated, it is generally considered to be a neurovascular disorder with a complex interplay of both neurological and vascular factors.
The vascular component of migraine is indicated by the throbbing nature of the headache; however, this theory remains controversial, with some researchers regarding vascular changes as an epiphenomenon [3,4]. In contrast, the neurological nature of migraine is better understood, with evidence suggesting that it is influenced by factors such as anxiety, stress, and cortical spreading depression (CSD) [5]. Approximately one third of those who suffer from migraine also experience aura, which is a manifestation of transient visual and somatosensory disturbances induced by CSD, a slowly propagating wave of neuronal and glial depolarization across the cortex [6,7,8]. In addition, CSD is believed to activate sensory neurons in the trigeminal ganglia (TG), suggesting the role of the central and peripheral nervous systems in migraine [9,10].
In the peripheral nervous system, nociceptors in the TG are thought to play a key role in the pain signals of migraine [11,12], as they innervate the intracranial meninges and convey pain signals from the meninges to the thalamus (Figure 1A). Targeting these nociceptors is considered a promising approach for migraine treatment owing to their accessibility and distinct molecular profile [13], which includes the expression of several transient receptor potential (TRP) channels that are being investigated as potential therapeutic targets for migraine [14,15,16]. In this review, we discuss the role and mechanisms of nociceptors in migraine, the challenges of current antimigraine drugs, and the evidence for well-studied and emerging TRP channels as novel targets for migraine prevention and treatment.
2. Nociceptors and meningeal inflammation
Sensory neurons located in the dorsal root ganglion (DRG) and TG are responsible for detecting and transmitting sensory information from the periphery to the central nervous system (CNS) [17]. Nociceptors are specialized peripheral sensory neurons that encode painful stimuli [18]. Activation and sensitization of nociceptors innervating cephalic tissues, mainly the cranial meninges and their associated blood vessels, are the basis of migraine pain [12]. Nociceptors can release neuropeptides associated with migraine, such as calcitonin gene-related peptide (CGRP), serotonin, and pituitary adenylate cyclase activating polypeptide-38 (PACAP-38). This can lead to vasodilation and meningeal inflammation, which can trigger migraine attacks [19,20,21,22].
Meningeal inflammation, triggered by various cells and molecules, can activate and sensitize nociceptors (Figure 1B), leading to migraine attacks [23,24,25,26]. Following CSD, molecules such as ATP, glutamate, K+, H+, arachidonic acid (AA), and nitrous oxide (NO) are released locally and are thought to diffuse towards and activate meningeal nociceptors [27,28,29,30]. This occurs a few minutes after CSD, which is consistent with the time delay between the onset of aura and migraine attack [9]. Previous studies have observed that mast cell degranulation, which involves the release of ATP, histamine, and pro-inflammatory cytokines, can cause meningeal inflammation and long-term activation and sensitization of dural nociceptors [26]. In fact, injections of the mast cell degranulator compound 48/80 in mice resulted in migraine-associated behaviors, some of which were reversed by mast cell stabilizers and anti-inflammatory drugs [26,31,32].
Targeting nociceptors and meningeal inflammation represents a viable strategy for preventing and relieving migraine pain. Current migraine pharmacotherapies can be divided into two main categories: abortive therapies, which aim to end a migraine episode, and prophylactic therapies, which aim to prevent future migraine attacks. Several classes of drugs are used to prevent migraine, such as anti-epileptics, anti-hypertensive agents, tricyclic antidepressants, anti-CGRP antibodies, and botulinum toxin [27,33,34,35,36]. Abortive treatments mostly include triptans and non-steroidal anti-inflammatory drugs (NSAIDs) [37]. The following paragraphs provide a brief overview of how these therapies affect nociceptors and meningeal inflammation, without providing clinical advice or indications.
3. Current antimigraine drugs
NSAIDs are widely available as over-the-counter medications for pain relief and are often used to treat mild to moderate acute migraine [38]. These drugs possess anti-inflammatory, analgesic, and antipyretic properties, mostly by blocking the enzymes cyclooxygenase-1 and -2 (COX-1 and -2) and thus reducing prostaglandin synthesis. It is important to note that COX-1 is widely distributed and involved in homeostatic mechanisms, while COX-2 is mainly expressed in areas of inflammation and responsible for anti-inflammatory and analgesic effects [39]. NSAIDs with selective inhibition of COX-2 may be considered as a relatively safer treatment for migraine, although they are associated with more adverse cardiovascular effects [40].
Triptans targeting 5-HT receptors are a gold standard treatment for migraine attacks [36]. They block nociceptive signaling to second-order neurons in the trigeminal nucleus caudalis but have limited ability to cross the blood-brain barrier (BBB). Their primary action is in the periphery (Figure 1C). Triptans are 5-HT1B/1D receptor agonists, causing vasoconstriction via 5-HT1B and inhibiting neuropeptide release via 5-HT1D [41]. However, they increase blood pressure, and therefore, may be contraindicated in those with cardiovascular disease or hypertension and pregnant women [42,43]. Another concern with the chronic use of triptans is the development of medication overuse headache [44]. Ditans are 5-HT1F agonists that do not cause vasoconstriction due to low affinity for the 5-HT1B receptor [41]. Ditans are often used for the treatment of migraine in patients with or at risk of cardiovascular disease and in patients who respond poorly to their current treatment [45].
Despite CGRP being identified as a key molecule in migraine pathology more than 30 years ago, drugs acting on the CGRP or its receptor have only recently been approved for migraine treatment [46]. These drugs can be divided into two main groups: small-molecule CGRP receptor antagonists and monoclonal antibodies targeting either CGRP or its receptor [46,47]. Both small-molecule antagonists and antibodies do not easily cross the BBB; hence, they mainly act peripherally on the trigeminal ganglion and meninges (Figure 1D). Monoclonal antibodies are generally considered safe and effective for migraine prevention and treatment; however, long-term CGRP blockade effects remain unknown and may be a concern for patients with or at risk of cardiovascular diseases [48,49].
Botulinum neurotoxin serotype-A (BoNT/A) is emerging as a potential treatment for chronic migraine [50]. It has been well-documented that BoNT/A interferes with synaptic vesicles, blocking the release of neuropeptides (Figure 1E) [51,52]. For instance, a clinical study has shown that BoNT/A reduces interictal CGRP plasma levels in chronic migraine patients [53]. However, it is becoming increasingly clear that BoNT/A also regulates the expression and function of TRP channels in nociceptors, which may explain its analgesic effect by inhibiting CGRP release [54]. The effects of BoNT/A on TRP channels have not yet been tested directly in migraineurs. However, the reported analgesic response to BoNT/A injections in people injected with capsaicin is likely due to a decrease in expression of the TRP cation channel subfamily V member 1 (TRPV1) in nociceptors [55], and the potential analgesic response to BoNT/A treatments in a population of female patients with chronic migraine may be associated with a particular polymorphism in the TRPV1 gene [56]. The success of BoNT/A and its interactions with TRPV1 provide support for targeting TRP channels in migraine treatment.
4. TRP channels as targets for antimigraine drugs
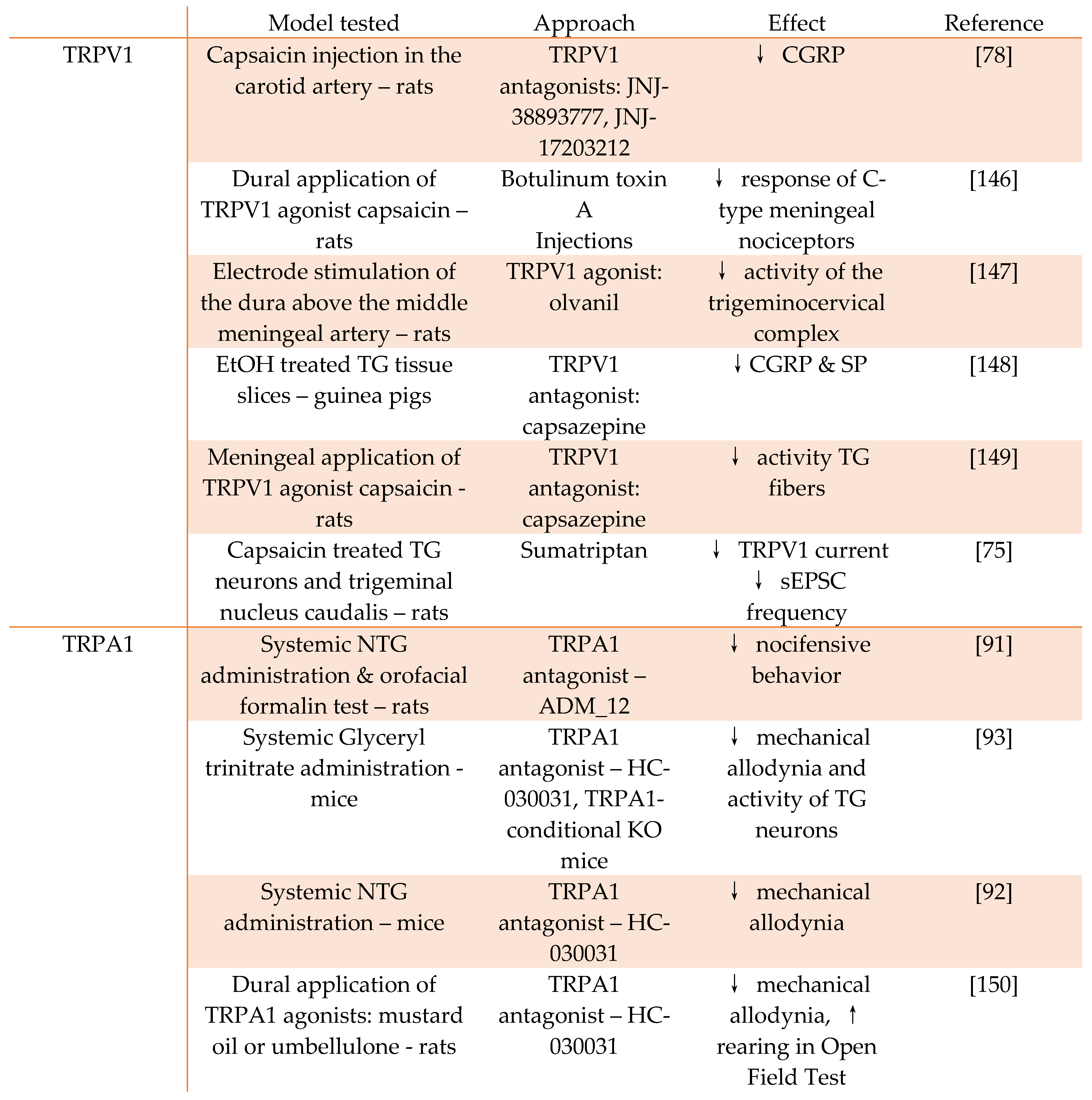
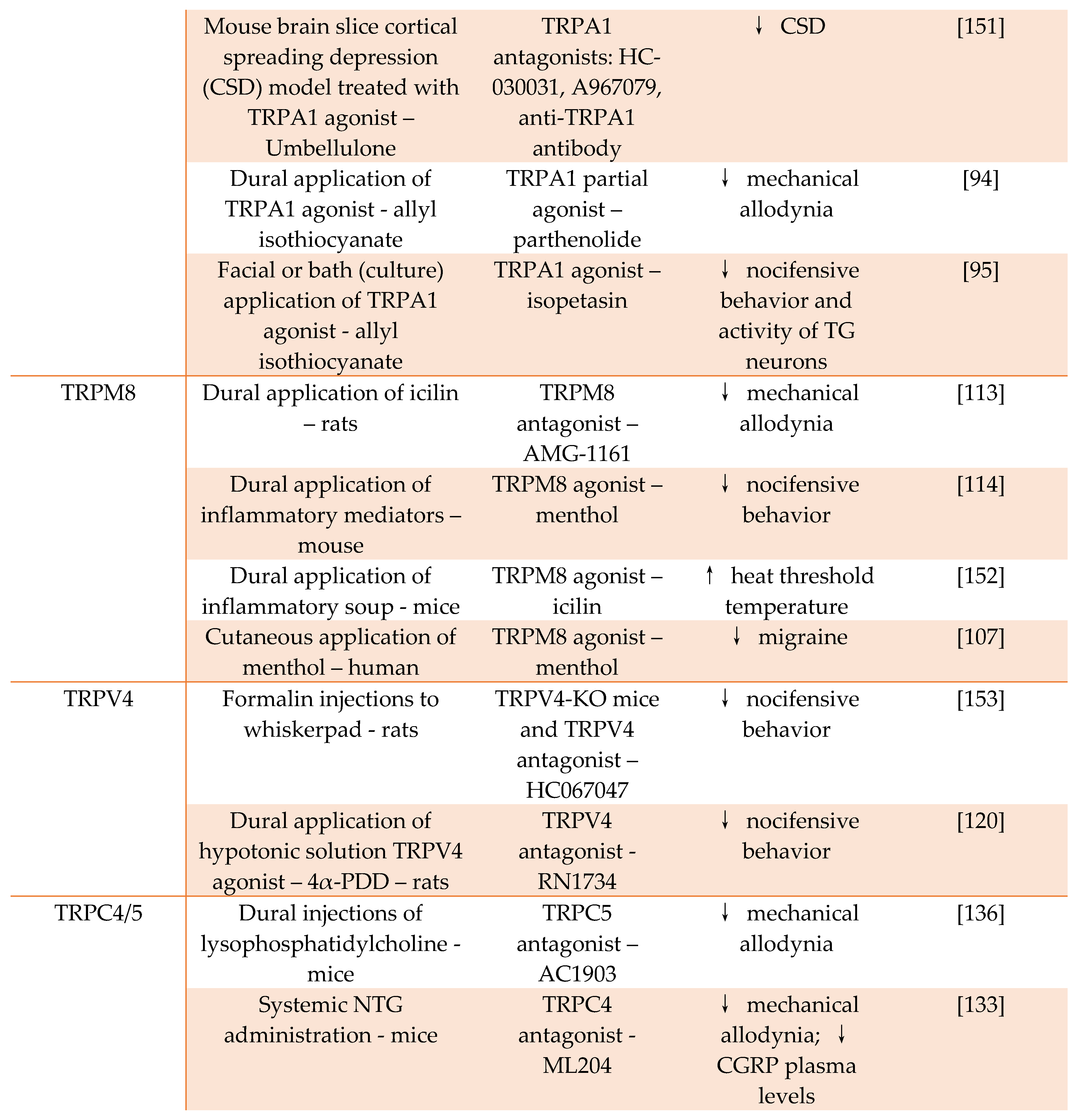
TRP channels represent a non-selective ion channel superfamily consisting of 28 subtypes. In mammals, these are classified into six subfamilies: TRPA (ankyrin), TRPV (vanilloid), TRPC (canonical), TRPM (melastatin), TRPML (mucolipin), and TRPP (polycystin) [57]. TRP channel members are structurally similar, with six transmembrane domains (S1-S6) and cytoplasmic amino and carboxy termini. However, they differ in their primary amino acid sequence [58]. The S1-S6 domains assemble into homo- or hetero-tetramers, with a hydrophilic loop between the fifth and sixth transmembranes forming a pore that is permeable to cations such as Na+ and Ca2+ ions. TRP channels can be triggered by various stimuli, such as thermal, mechanical, chemical, pH, and osmolarity, as well as external and internal ligands [57]. Consequently, they are implicated in many diseases [59]. TRP channels are particularly abundant in sensory neurons and have been linked to various pain conditions [60]. Evidence suggests that they activate meningeal nociceptors, contributing to the development and progression of migraine [14,15]. For instance, TRP channels are sensitive to environmental factors, such as pollutants and temperature changes, which are known migraine triggers [61,62,63]. Additionally, their activation induces transcriptional changes and the release of pro-inflammatory and algesic neuropeptides, such as CGRP and substance P (SP), from TG nociceptors [10]. Moreover, polymorphisms in TRP genes may also regulate the propensity for migraine and responsiveness to its treatment [56,64]. Preclinical studies have further supported the role of TRP channels in migraine and have shown that their silencing or inhibition can reduce migraine pain (Table 1).
TRPV1, the first of many thermosensitive TRP channels to be identified, functions as a noxious heat sensor that is activated by temperatures greater than 42°C and pH changes [65,66]. Additionally, it is triggered by exogenous stimulants, such as capsaicin from chili peppers, and endogenous stimulants, such as anandamide produced in inflammatory processes [65,66,67]. TRPV1 activity is further modulated by inflammatory mediators, such as prostaglandin E2 and bradykinin, which trigger the release of proinflammatory and pro-migraine neuropeptides, including CGRP and SP, within the meninges [68,69]. Therefore, TRPV1 has been thoroughly studied in migraine [70]. TRPV1 is highly expressed in small and medium-sized neurons in peripheral ganglia, with approximately 10–20% of TG neurons reported to be TRPV1-positive [71]. Additionally, 70% of CGRP-positive neurons have been shown to co-localize with TRPV1-positive neurons in the TG, and dural trigeminal fibers also exhibit co-localization [72,73]. Genetic evidence suggests the involvement of TRPV1 in migraine, as a study conducted in the Spanish population found single nucleotide polymorphisms (SNPs) in the TRPV1 gene in patients with migraine [74]. Furthermore, the anti-migraine drug sumatriptan was recently shown to block trigeminal TRPV1 channels [75]. Thus, TRPV1 agonists and antagonists have been identified as potential therapeutic agents for migraine management [76]. However, several promising preclinical drugs directly targeting TRP channels have failed to translate into clinical use [60]. Intranasal TRPV1 agonists, such as capsaicin and civamide (a synthetic isomer of capsaicin), were efficacious in relieving patients' migraine pain. However, adverse side effects, including nasal burning and lacrimation, prevented the clinical application of these drugs [68,77]. Since then, the focus of most TRPV1 drugs for migraine treatment has been on TRPV1 antagonists [68]. Unfortunately, SB-705498, a TRPV1 antagonist, failed to demonstrate superior efficacy against migraine headaches compared to placebo in a Phase II clinical trial [78]. Moreover, other studies testing TRPV1 antagonists were terminated due to serious adverse effects, notably hyperthermia [79]. Adverse side effects of TRP channel targeting may be avoided by indirect inhibition using resolvins, which have no apparent thermoregulatory effects [80]. Notably, we found that resolvin D3 significantly inhibited TRPV1 activity and CGRP release in both mouse and human DRG neurons [81] and may represent a new antimigraine drug.
TRPA1 is another calcium-permeable non-selective cation channel that was initially thought to detect noxious cold temperatures with a threshold of 17°C [82,83]. However, later studies indicated that TRPA1 is involved in detecting cold hypersensitivity, rather than physiological cold pain [84]. TRPA1 can be activated by a variety of environmental irritants, pollutants, and pungent food such as garlic, cinnamon, and mustard oil [85,86,87]. Additionally, the monoterpene umbellulone, produced by Umbellaria californica, has been linked to migraine induction in humans and reported as a TRPA1 activator [62,88]. In rodents, umbellulone administration has been shown to result in CGRP release, meningeal vasodilation, and migraine-like behaviors [62]. TRPA1 is also linked to migraine through its endogenous activators, such as reactive oxygen species (ROS), reactive prostaglandins, and nitric oxide (NO), all of which are all well-known to play a role in migraine attacks [14]. Rodent studies have shown that TRPA1 is mainly localized to small and medium-sized peptidergic neurons and is present in around 36% of trigeminal sensory neurons and dural afferents [10,71]. In addition, it has been found that TRPA1 is present in a subset of C and Aδ neuronal fibers expressing TRPV1, CGRP, SP, and protease-activated receptor 2 (PAR-2) [10,73,89]. Studies have shown that the PAR-2 receptor can induce migraine pain behavior in mice, and a PAR-2 monoclonal antibody has been recently tested in rodents as an anti-migraine drug [31,90]. Pre-clinical studies have also shown that several TRPA1 antagonists and genetic deletion are effective in reducing migraine-like symptoms such as allodynia [91,92,93]. However, it is important to note that allodynia, but not vasodilation, was prevented through the genetic deletion or pharmacological blockade of TRPA1 [93]. Further evidence establishing TRPA1 as a therapeutic target for migraine comes from in vivo and in vitro preclinical studies. These studies showed that plant extracts from the feverfew herb (Tanacetum parthenium) and butterbur (Petasites hybridus) desensitizes meningeal TRPA1 channels, resulting in decreased release of vasoactive neuropeptides and consequent pain relief [94,95]. Currently, high-affinity and selective TRPA1 antagonists are in the developmental phase and undergoing phase I and phase II clinical trials for different diseases that have a significant pain component [70].
TRPM8 is activated by mild cold temperatures ranging from 8–25°C, as well as by cooling agents such as menthol and icilin [96,97]. TRPM8 can also be activated and regulated by various endogenous molecules, including lipids, artemin, testosterone, and membrane phosphatidylinositol 4,5-bisphosphate (PI(4,5)P2) [98,99,100,101]. Single nucleotide polymorphisms (SNPs) in TRPM8 have been associated with migraine [102,103,104]; however, it is unclear whether some of these SNPs are beneficial or detrimental. For instance, SNP rs10166942 has been associated with a lower risk of migraine, but it has been linked to the development of chronic migraine and allodynia in some patients [102,103,105]. Cold temperatures are also known to trigger migraines, and several migraine patients exhibit cold allodynia [63,106]. Although cold temperature and allodynia may suggest a detrimental role for TRPM8, the TRPM8 agonist menthol offers pain relief in some migraine patients [107]. In mice, TRPM8 is expressed in approximately 12% of potentially peptidergic neurons in the trigeminal ganglion [71]; however, only a small percentage of TRPM8-containing neurons also express CGRP and TRPV1 [108,109,110]. Interestingly, TRPM8 expression was also described in supraspinal regions [111]. Although it is clear that TRPM8 mediates both innocuous and noxious cold sensations, as well as being necessary for cold allodynia and hyperalgesia in various pain models, the role of this channel in migraine-related pain is controversial [112]. For instance, it has been shown that the activation of meningeal TRPM8 by exogenous agonists can both cause and alleviate headache behaviors, depending on whether other meningeal afferents concurrently receive noxious stimuli [113,114]. A recent study using transgenic knockout TRPM8 mice suggests a protective role in promoting a faster recovery from chronic migraine-like symptoms in male mice compared to that in female mice [115]. However, another study with transgenic mice showed that TRPM8 channels or afferents are both required for the development of acute and chronic migraine-like symptoms [116]. This suggests that further investigations are needed to determine the role and therapeutic potential of TRPM8.
TRPV4 responds to various stimuli, including changes in osmolarity and mechanical forces imposed on the cell membrane, suggesting that it functions as a mechanosensory receptor [117,118]. Although little is known about the relationship between TRPV4 and migraine, its function is particularly intriguing as changes in intracranial pressure are known to affect headaches. TRPV4 is expressed in meningeal nociceptors with PAR2, and activating this channel in the dura causes headache behavioral responses, such as cephalic and extracephalic allodynia [119,120]. A TRPV4 antagonist inhibited these behavioral responses, suggesting that this channel can be a potential therapeutic target for antimigraine drugs.
5. TRPC4 channel as an emerging target for antimigraine drugs
The TRP channel subfamily known as Transient Receptor Potential Canonical (TRPC) consists of seven members, TRPC1–7, which can be further divided into four subgroups: TRPC1, TRPC4/5, TRPC3/6/7, and TRPC2, based on sequence similarity [121]. TRPC channels are expressed across species, with the exception of TRPC2, which is not expressed in humans [122]. These channels possess six transmembrane helices and assemble into cation channels as homo- and hetero-tetramers, including two or three TRPC subtypes [123]. They are permeable nonselective cation channels that are activated by G-protein-coupled receptors or receptor tyrosine kinases. Most of these channels are linked to the phospholipase C signaling and calcium signaling [124,125,126,127]. Recent studies have revealed that these channels play an integral role in neuronal functions and brain diseases through their regulation of intracellular Ca2+, which contributes to synaptic transmission and neural plasticity [128]. For instance, studies have shown that TRPC4 and TRPC5 are highly expressed in brain regions associated with anxiety and fear, and genetic silencing or pharmacological inhibition of these channels have been found to be effective in preclinical animal models of depression [129,130,131]. TRPC4 and TRPC5 are expressed not only in the brain but also in peripheral sensory neurons, where they are involved in axon guidance and sensory functions [132,133,134,135,136].
TRPC4 expression has been initially identified in sensory neurons from dorsal root ganglia as a major driver of serotonergic and psoriasiform itch. First, we found that TRPC4 has a unique function in DRG neurons, mediating itch to serotonergic antidepressants independently of TRPV1 and TRPA1 [137]. Second, we characterized the expression of TRPC4 in peptidergic DRG neurons and showed that acute itch induced by serotonin and histamine was attenuated in TRPC4 knockout mice and mice treated with ML204 [132], a specific TRPC4 antagonist [138,139]. More importantly, pharmacological inhibition of TRPC4 resulted in the reduction of psoriasiform skin inflammation and itch, potentially through a decrease of cutaneous pro-inflammatory cytokine and CGRP levels [132]. This suggests that TRPC4, via its expression in primary sensory neurons and regulation of CGRP, may represent a novel therapeutic target for migraine.
A recent study showed that TRPC4 is highly expressed in TG neurons and that pharmacological inhibition of TRPC4 significantly prevented migraine-linked cutaneous mechanical hypersensitivity and increased plasma levels of CGRP [133]. Specifically, we found that TRPC4 is highly expressed in TG tissues and neurons, with most of these neurons also expressing CGRP and innervating the skin. To investigate whether TRPC4 plays a role in migraine, we used mouse models of migraine induced by intraperitoneal administration of nitroglycerin (NTG, Figure 2A). A single injection mimicked an acute episodic migraine attack, while repeated injections mimicked chronic migraine [140]. Mice injected with NTG develop cutaneous mechanical hypersensitivity, also known as allodynia, in the periorbital area and hindpaw, which is a symptom often reported by migraineurs [141]. We observed that the TRPC4-specific inhibitor ML204 reduced cutaneous mechanical hypersensitivity following a single injection and prevented its development after repeated injections. Interestingly, our results indicate that both male and female mice experienced similar reduction and prevention of mechanical hypersensitivity, despite the fact that female mice may have different underlying mechanisms and women are generally more susceptible to migraines [142,143]. Additionally, CGRP plasma levels were reduced in both male and female mice after daily treatment with ML204. A recent report suggests that CGRP may have female-specific actions in peripheral tissues [142]; however, as a small-molecule inhibitor, ML204 can act in both peripheral and central nervous system tissues. Thus, ML204 can target the release of CGRP in the brainstem, which is important in migraine pathophysiology. However, no study has evaluated the role of male vs. female brainstem in migraine [144,145]. Previously, we showed that ML204 and small interfering RNA targeting TRPC4 can attenuate the activity of sensory neurons [137]. This suggests that targeting TRPC4 can lead to analgesia by reducing neural activity and the release of CGRP (Figure 2B). Since TRPC4 is expressed in human DRG tissue [137], and TRPC4/5 has entered clinical trials for the treatment of central nervous system disorders, it is possible that TRPC4 antagonists may soon be useful in migraine treatment.
6. Conclusions
Migraine is a complex disorder with both neurological and vascular components. The exact causes of migraine are still being studied and debated. It is thought that nociceptors in the trigeminal ganglia play a key role in the pain signals of migraine, as they innervate the intracranial meninges and convey pain signals from the meninges to the thalamus. Because these nociceptors are accessible and have a distinct molecular profile, targeting them is considered a promising approach. Among the TRP channels, TRPV1, TRPA1, and TRPM8 have been well studied and are being investigated as potential therapeutic targets for migraine prevention and treatment. Apart from these, TRPC4 is another subtype of TRP channels that has been implicated in pain and migraine. We found that TRPC4 is highly expressed in TG neurons and that inhibiting TRPC4 can prevent migraine-linked cutaneous mechanical hypersensitivity and reduce CGRP plasma levels in a mouse model of migraine. Thus, targeting TRP channels, including TRPC4, represents a promising approach for the prevention and treatment of migraine, a debilitating neurological disorder that affects millions of people worldwide.
Author Contributions
Conceptualization, C.F.C., C.-K.P. and T.B.; Formal analysis, C.F.C. and J.R.; Funding acquisition, C.-K.P. and T.B.; Resources, C.F.C. and T.B.; Supervision, C.-K.P. and T.B.; Validation, C.F.C. and T.B.; Visualization, J.R. and S.H.L.; Writing–original draft, C.F.C., S.H.L., T.B.; Writing–review and editing, C.F.C., C.-K.P., T.B.; All authors have read and agreed to the published version of the manuscript.
Acknowledgments
This work was supported by grants from the National Research Foundation of Korea (NRF-2022M3E5E8081191 to C.-K.P.) and the US National Institutes of Health-NINDS (NS113243 to T.B.). All authors approved the final manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Burch, R. C.; Buse, D. C.; Lipton, R. B. , Migraine: Epidemiology, Burden, and Comorbidity. Neurol Clin 2019, 37, 631–649. [Google Scholar] [CrossRef] [PubMed]
- Charles, A. , The pathophysiology of migraine: implications for clinical management. Lancet Neurol 2018, 17, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Charles, A. , Vasodilation out of the picture as a cause of migraine headache. Lancet Neurol 2013, 12, 419–420. [Google Scholar] [CrossRef] [PubMed]
- Mason, B. N.; Russo, A. F. , Vascular Contributions to Migraine: Time to Revisit? Front Cell Neurosci 2018, 12, 233. [Google Scholar] [CrossRef]
- Burstein, R.; Noseda, R.; Borsook, D. , Migraine: multiple processes, complex pathophysiology. J Neurosci 2015, 35, 6619–6629. [Google Scholar] [CrossRef] [PubMed]
- Noseda, R.; Burstein, R. , Migraine pathophysiology: anatomy of the trigeminovascular pathway and associated neurological symptoms, CSD, sensitization and modulation of pain. Pain, 2013, 154 Suppl 1.
- Goadsby, P. J.; Holland, P. R.; Martins-Oliveira, M.; Hoffmann, J.; Schankin, C.; Akerman, S. , Pathophysiology of Migraine: A Disorder of Sensory Processing. Physiol Rev 2017, 97, 553–622. [Google Scholar] [CrossRef]
- Dodick, D. W. , A Phase-by-Phase Review of Migraine Pathophysiology. Headache 2018, 58 Suppl 1, 4–16. [Google Scholar] [CrossRef]
- Zhang, X.; Levy, D.; Noseda, R.; Kainz, V.; Jakubowski, M.; Burstein, R. , Activation of meningeal nociceptors by cortical spreading depression: implications for migraine with aura. J Neurosci 2010, 30, 8807–8814. [Google Scholar] [CrossRef]
- Dux, M.; Rosta, J.; Messlinger, K. , TRP Channels in the Focus of Trigeminal Nociceptor Sensitization Contributing to Primary Headaches. Int J Mol Sci 2020, 21. [Google Scholar] [CrossRef]
- Olesen, J.; Burstein, R.; Ashina, M.; Tfelt-Hansen, P. , Origin of pain in migraine: evidence for peripheral sensitisation. Lancet Neurol 2009, 8, 679–690. [Google Scholar] [CrossRef]
- Levy, D. , Migraine pain and nociceptor activation--where do we stand? Headache 2010, 50, 909–916. [Google Scholar] [CrossRef] [PubMed]
- Berta, T.; Qadri, Y.; Tan, P. H.; Ji, R. R. , Targeting dorsal root ganglia and primary sensory neurons for the treatment of chronic pain. Expert Opin Ther Targets 2017, 21, 695–703. [Google Scholar] [CrossRef] [PubMed]
- Benemei, S.; Dussor, G. , TRP Channels and Migraine: Recent Developments and New Therapeutic Opportunities. Pharmaceuticals (Basel) 2019, 12. [Google Scholar] [CrossRef] [PubMed]
- Dussor, G.; Yan, J.; Xie, J. Y.; Ossipov, M. H.; Dodick, D. W.; Porreca, F. , Targeting TRP channels for novel migraine therapeutics. ACS Chem Neurosci 2014, 5, 1085–1096. [Google Scholar] [CrossRef] [PubMed]
- Artero-Morales, M.; González-Rodríguez, S.; Ferrer-Montiel, A. , TRP Channels as Potential Targets for Sex-Related Differences in Migraine Pain. Front Mol Biosci 2018, 5, 73. [Google Scholar] [CrossRef] [PubMed]
- Basbaum, A. I.; Bautista, D. M.; Scherrer, G.; Julius, D. , Cellular and molecular mechanisms of pain. Cell 2009, 139, 267–284. [Google Scholar] [CrossRef]
- Woolf, C. J.; Ma, Q. , Nociceptors--noxious stimulus detectors. Neuron 2007, 55, 353–364. [Google Scholar] [CrossRef]
- Edvinsson, L.; Goadsby, P. J. , Neuropeptides in migraine and cluster headache. Cephalalgia 1994, 14, 320–327. [Google Scholar] [CrossRef]
- Iyengar, S.; Ossipov, M. H.; Johnson, K. W. , The role of calcitonin gene-related peptide in peripheral and central pain mechanisms including migraine. Pain 2017, 158, 543–559. [Google Scholar] [CrossRef]
- Markovics, A.; Kormos, V.; Gaszner, B.; Lashgarara, A.; Szoke, E.; Sandor, K.; Szabadfi, K.; Tuka, B.; Tajti, J.; Szolcsanyi, J.; Pinter, E.; Hashimoto, H.; Kun, J.; Reglodi, D.; Helyes, Z. , Pituitary adenylate cyclase-activating polypeptide plays a key role in nitroglycerol-induced trigeminovascular activation in mice. Neurobiol Dis 2012, 45, 633–644. [Google Scholar] [CrossRef]
- Aggarwal, M.; Puri, V.; Puri, S. , Serotonin and CGRP in migraine. Ann Neurosci 2012, 19, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Burstein, R.; Levy, D. , Local action of the proinflammatory cytokines IL-1β and IL-6 on intracranial meningeal nociceptors. Cephalalgia 2012, 32, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Edvinsson, L.; Haanes, K. A.; Warfvinge, K. , Does inflammation have a role in migraine? Nat Rev Neurol 2019, 15, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Lukács, M.; Haanes, K. A.; Majláth, Z.; Tajti, J.; Vécsei, L.; Warfvinge, K.; Edvinsson, L. , Dural administration of inflammatory soup or Complete Freund's Adjuvant induces activation and inflammatory response in the rat trigeminal ganglion. J Headache Pain 2015, 16, 564. [Google Scholar] [CrossRef]
- Conti, P.; D'Ovidio, C.; Conti, C.; Gallenga, C. E.; Lauritano, D.; Caraffa, A.; Kritas, S. K.; Ronconi, G. , Progression in migraine: Role of mast cells and pro-inflammatory and anti-inflammatory cytokines. Eur J Pharmacol 2019, 844, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Pietrobon, D.; Moskowitz, M. A. , Pathophysiology of migraine. Annu Rev Physiol 2013, 75, 365–391. [Google Scholar] [CrossRef] [PubMed]
- Gursoy-Ozdemir, Y.; Qiu, J.; Matsuoka, N.; Bolay, H.; Bermpohl, D.; Jin, H.; Wang, X.; Rosenberg, G. A.; Lo, E. H.; Moskowitz, M. A. , Cortical spreading depression activates and upregulates MMP-9. J Clin Invest 2004, 113, 1447–1455. [Google Scholar] [CrossRef]
- Karatas, H.; Erdener, S. E.; Gursoy-Ozdemir, Y.; Lule, S.; Eren-Koçak, E.; Sen, Z. D.; Dalkara, T. , Spreading depression triggers headache by activating neuronal Panx1 channels. Science 2013, 339, 1092–1095. [Google Scholar] [CrossRef]
- Chen, S. P.; Qin, T.; Seidel, J. L.; Zheng, Y.; Eikermann, M.; Ferrari, M. D.; van den Maagdenberg, A.; Moskowitz, M. A.; Ayata, C.; Eikermann-Haerter, K. , Inhibition of the P2X7-PANX1 complex suppresses spreading depolarization and neuroinflammation. Brain 2017, 140, 1643–1656. [Google Scholar] [CrossRef]
- Hassler, S. N.; Ahmad, F. B.; Burgos-Vega, C. C.; Boitano, S.; Vagner, J.; Price, T. J.; Dussor, G. , Protease activated receptor 2 (PAR2) activation causes migraine-like pain behaviors in mice. Cephalalgia 2019, 39, 111–122. [Google Scholar] [CrossRef]
- Ramachandran, R.; Wang, Z.; Saavedra, C.; DiNardo, A.; Corr, M.; Powell, S. B.; Yaksh, T. L. , Role of Toll-like receptor 4 signaling in mast cell-mediated migraine pain pathway. Mol Pain 2019, 15, 1744806919867842. [Google Scholar] [CrossRef]
- Hoffmann, J.; Goadsby, P. J. , Emerging targets in migraine. CNS Drugs 2014, 28, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Belvis, R.; Mas, N.; Aceituno, A. , Migraine attack treatment : a tailor-made suit, not one size fits all. Recent Pat CNS Drug Discov 2014, 9, 26–40. [Google Scholar] [CrossRef] [PubMed]
- Tso, A. R.; Goadsby, P. J. , Anti-CGRP Monoclonal Antibodies: the Next Era of Migraine Prevention? Curr Treat Options Neurol 2017, 19, 27. [Google Scholar] [CrossRef] [PubMed]
- Diener, H. C.; Charles, A.; Goadsby, P. J.; Holle, D. , New therapeutic approaches for the prevention and treatment of migraine. Lancet Neurol 2015, 14, 1010–1022. [Google Scholar] [CrossRef] [PubMed]
- Lipton, R. B.; Silberstein, S. D. , Episodic and chronic migraine headache: breaking down barriers to optimal treatment and prevention. Headache, 2015, 55 Suppl 2, 103-22; quiz 123-6.
- Pardutz, A.; Schoenen, J. , NSAIDs in the Acute Treatment of Migraine: A Review of Clinical and Experimental Data. Pharmaceuticals (Basel) 2010, 3, 1966–1987. [Google Scholar] [CrossRef] [PubMed]
- Ong, J. J. Y.; De Felice, M. , Migraine Treatment: Current Acute Medications and Their Potential Mechanisms of Action. Neurotherapeutics 2018, 15, 274–290. [Google Scholar] [CrossRef]
- Cannon, C. P.; Cannon, P. J. , Physiology. COX-2 inhibitors and cardiovascular risk. Science 2012, 336, 1386–1387. [Google Scholar] [CrossRef] [PubMed]
- Negro, A.; Koverech, A.; Martelletti, P. , Serotonin receptor agonists in the acute treatment of migraine: a review on their therapeutic potential. J Pain Res 2018, 11, 515–526. [Google Scholar] [CrossRef]
- Diener, H. C. , The Risks or Lack Thereof of Migraine Treatments in Vascular Disease. Headache 2020, 60, 649–653. [Google Scholar] [CrossRef]
- Victor, T. W.; Hu, X.; Campbell, J. C.; Buse, D. C.; Lipton, R. B. , Migraine prevalence by age and sex in the United States: a life-span study. Cephalalgia 2010, 30, 1065–1072. [Google Scholar] [CrossRef]
- Meng, I. D.; Dodick, D.; Ossipov, M. H.; Porreca, F. , Pathophysiology of medication overuse headache: insights and hypotheses from preclinical studies. Cephalalgia 2011, 31, 851–860. [Google Scholar] [CrossRef] [PubMed]
- Kuca, B.; Silberstein, S. D.; Wietecha, L.; Berg, P. H.; Dozier, G.; Lipton, R. B. , Lasmiditan is an effective acute treatment for migraine: A phase 3 randomized study. Neurology 2018, 91, e2222–e2232. [Google Scholar] [CrossRef] [PubMed]
- Wattiez, A. S.; Sowers, L. P.; Russo, A. F. , Calcitonin gene-related peptide (CGRP): role in migraine pathophysiology and therapeutic targeting. Expert Opin Ther Targets 2020, 24, 91–100. [Google Scholar] [CrossRef]
- de Vries, T.; Villalón, C. M.; MaassenVanDenBrink, A. , Pharmacological treatment of migraine: CGRP and 5-HT beyond the triptans. Pharmacol Ther 2020, 211, 107528. [Google Scholar] [CrossRef] [PubMed]
- Saely, S.; Croteau, D.; Jawidzik, L.; Brinker, A.; Kortepeter, C. , Hypertension: A new safety risk for patients treated with erenumab. Headache 2021, 61, 202–208. [Google Scholar] [CrossRef]
- MaassenVanDenBrink, A.; Meijer, J.; Villalón, C. M.; Ferrari, M. D. , Wiping Out CGRP: Potential Cardiovascular Risks. Trends Pharmacol Sci 2016, 37, 779–788. [Google Scholar] [CrossRef] [PubMed]
- Raciti, L.; Raciti, G.; Militi, D.; Casella, C.; Calabrò, R. S. , Chronic Migraine: A Narrative Review on the Use of Botulinum Toxin with Clinical Indications and Future Directions. J Integr Neurosci 2022, 21, 141. [Google Scholar] [CrossRef]
- Rossetto, O.; Pirazzini, M.; Montecucco, C. , Botulinum neurotoxins: genetic, structural and mechanistic insights. Nat Rev Microbiol 2014, 12, 535–549. [Google Scholar] [CrossRef]
- Burstein, R.; Blumenfeld, A. M.; Silberstein, S. D.; Manack Adams, A.; Brin, M. F. , Mechanism of Action of OnabotulinumtoxinA in Chronic Migraine: A Narrative Review. Headache 2020, 60, 1259–1272. [Google Scholar] [CrossRef]
- Cernuda-Morollón, E.; Ramón, C.; Martínez-Camblor, P.; Serrano-Pertierra, E.; Larrosa, D.; Pascual, J. , OnabotulinumtoxinA decreases interictal CGRP plasma levels in patients with chronic migraine. Pain 2015, 156, 820–824. [Google Scholar] [CrossRef]
- Go, E. J.; Ji, J.; Kim, Y. H.; Berta, T.; Park, C. K. , Transient Receptor Potential Channels and Botulinum Neurotoxins in Chronic Pain. Front Mol Neurosci 2021, 14, 772719. [Google Scholar] [CrossRef]
- Gazerani, P.; Pedersen, N. S.; Staahl, C.; Drewes, A. M.; Arendt-Nielsen, L. , Subcutaneous Botulinum toxin type A reduces capsaicin-induced trigeminal pain and vasomotor reactions in human skin. Pain 2009, 141, (1–2), 60. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Mayordomo, R.; Ruiz, M.; Pascual, J.; Gallego de la Sacristana, M.; Vidriales, I.; Sobrado, M.; Cernuda-Morollon, E.; Gago-Veiga, A. B.; Garcia-Azorin, D.; Telleria, J. J.; Guerrero, A. L. , CALCA and TRPV1 genes polymorphisms are related to a good outcome in female chronic migraine patients treated with OnabotulinumtoxinA. J Headache Pain 2019, 20, 39. [Google Scholar] [CrossRef]
- Clapham, D. E.; Runnels, L. W.; Strübing, C. , The TRP ion channel family. Nat Rev Neurosci 2001, 2, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Koivisto, A. P.; Belvisi, M. G.; Gaudet, R.; Szallasi, A. , Advances in TRP channel drug discovery: from target validation to clinical studies. Nat Rev Drug Discov 2022, 21, 41–59. [Google Scholar] [CrossRef] [PubMed]
- Nilius, B.; Voets, T.; Peters, J. , TRP channels in disease. Sci STKE 2005, 2005, re8. [Google Scholar] [CrossRef] [PubMed]
- Patapoutian, A.; Tate, S.; Woolf, C. J. , Transient receptor potential channels: targeting pain at the source. Nat Rev Drug Discov 2009, 8, 55–68. [Google Scholar] [CrossRef]
- Kunkler, P. E.; Ballard, C. J.; Oxford, G. S.; Hurley, J. H. , TRPA1 receptors mediate environmental irritant-induced meningeal vasodilatation. Pain 2011, 152, 38–44. [Google Scholar] [CrossRef]
- Nassini, R.; Materazzi, S.; Vriens, J.; Prenen, J.; Benemei, S.; De Siena, G.; la Marca, G.; Andrè, E.; Preti, D.; Avonto, C.; Sadofsky, L.; Di Marzo, V.; De Petrocellis, L.; Dussor, G.; Porreca, F.; Taglialatela-Scafati, O.; Appendino, G.; Nilius, B.; Geppetti, P. , The 'headache tree' via umbellulone and TRPA1 activates the trigeminovascular system. Brain 2012, 135, 376–390. [Google Scholar] [CrossRef]
- Prince, P. B.; Rapoport, A. M.; Sheftell, F. D.; Tepper, S. J.; Bigal, M. E. , The effect of weather on headache. Headache 2004, 44, 596–602. [Google Scholar] [CrossRef] [PubMed]
- Gavva, N. R.; Sandrock, R.; Arnold, G. E.; Davis, M.; Lamas, E.; Lindvay, C.; Li, C. M.; Smith, B.; Backonja, M.; Gabriel, K.; Vargas, G. , Reduced TRPM8 expression underpins reduced migraine risk and attenuated cold pain sensation in humans. Sci Rep 2019, 9, 19655. [Google Scholar] [CrossRef] [PubMed]
- Caterina, M. J.; Rosen, T. A.; Tominaga, M.; Brake, A. J.; Julius, D. , A capsaicin-receptor homologue with a high threshold for noxious heat. Nature 1999, 398, 436–441. [Google Scholar] [CrossRef] [PubMed]
- Caterina, M. J.; Schumacher, M. A.; Tominaga, M.; Rosen, T. A.; Levine, J. D.; Julius, D. , The capsaicin receptor: a heat-activated ion channel in the pain pathway. Nature 1997, 389, 816–824. [Google Scholar] [CrossRef] [PubMed]
- Gladkikh, I. N.; Sintsova, O. V.; Leychenko, E. V.; Kozlov, S. A. , TRPV1 Ion Channel: Structural Features, Activity Modulators, and Therapeutic Potential. Biochemistry (Mosc) 2021, 86, S50–s70. [Google Scholar] [CrossRef] [PubMed]
- Takayama, Y.; Derouiche, S.; Maruyama, K.; Tominaga, M. , Emerging Perspectives on Pain Management by Modulation of TRP Channels and ANO1. Int J Mol Sci 2019, 20. [Google Scholar] [CrossRef]
- Hung, C. Y.; Tan, C. H. , TRP Channels in Nociception and Pathological Pain. Adv Exp Med Biol 2018, 1099, 13–27. [Google Scholar]
- Iannone, L. F.; De Logu, F.; Geppetti, P.; De Cesaris, F. , The role of TRP ion channels in migraine and headache. Neurosci Lett 2022, 768, 136380. [Google Scholar] [CrossRef]
- Huang, D.; Li, S.; Dhaka, A.; Story, G. M.; Cao, Y. Q. , Expression of the transient receptor potential channels TRPV1, TRPA1 and TRPM8 in mouse trigeminal primary afferent neurons innervating the dura. Mol Pain 2012, 8, 66. [Google Scholar] [CrossRef]
- Price, T. J.; Flores, C. M. , Critical evaluation of the colocalization between calcitonin gene-related peptide, substance P, transient receptor potential vanilloid subfamily type 1 immunoreactivities, and isolectin B4 binding in primary afferent neurons of the rat and mouse. J Pain 2007, 8, 263–272. [Google Scholar] [CrossRef]
- Shimizu, T.; Toriumi, H.; Sato, H.; Shibata, M.; Nagata, E.; Gotoh, K.; Suzuki, N. , Distribution and origin of TRPV1 receptor-containing nerve fibers in the dura mater of rat. Brain Res 2007, 1173, 84–91. [Google Scholar] [CrossRef]
- Carreño, O.; Corominas, R.; Fernández-Morales, J.; Camiña, M.; Sobrido, M. J.; Fernández-Fernández, J. M.; Pozo-Rosich, P.; Cormand, B.; Macaya, A. , SNP variants within the vanilloid TRPV1 and TRPV3 receptor genes are associated with migraine in the Spanish population. Am J Med Genet B Neuropsychiatr Genet 2012, (1), 94–103. [Google Scholar] [CrossRef] [PubMed]
- Evans, M. S.; Cheng, X.; Jeffry, J. A.; Disney, K. E.; Premkumar, L. S. , Sumatriptan inhibits TRPV1 channels in trigeminal neurons. Headache 2012, 52, 773–784. [Google Scholar] [CrossRef] [PubMed]
- Meents, J. E.; Neeb, L.; Reuter, U. , TRPV1 in migraine pathophysiology. Trends Mol Med 2010, 16, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Diamond, S.; Freitag, F.; Phillips, S. B.; Bernstein, J. E.; Saper, J. R. , Intranasal civamide for the acute treatment of migraine headache. Cephalalgia 2000, 20, 597–602. [Google Scholar] [CrossRef] [PubMed]
- Meents, J. E.; Hoffmann, J.; Chaplan, S. R.; Neeb, L.; Schuh-Hofer, S.; Wickenden, A.; Reuter, U. , Two TRPV1 receptor antagonists are effective in two different experimental models of migraine. The Journal of Headache and Pain 2015, 16. [Google Scholar] [CrossRef] [PubMed]
- Szallasi, A.; Sheta, M. , Targeting TRPV1 for pain relief: limits, losers and laurels. Expert Opin Investig Drugs 2012, 21, 1351–1369. [Google Scholar] [CrossRef] [PubMed]
- Roh, J.; Go, E. J.; Park, J. W.; Kim, Y. H.; Park, C. K. , Resolvins: Potent Pain Inhibiting Lipid Mediators via Transient Receptor Potential Regulation. Front Cell Dev Biol 2020, 8, 584206. [Google Scholar] [CrossRef] [PubMed]
- Lee, S. H.; Tonello, R.; Im, S.-T.; Jeon, H.; Park, J.; Ford, Z.; Davidson, S.; Kim, Y. H.; Park, C.-K.; Berta, T. , Resolvin D3 controls mouse and human TRPV1-positive neurons and preclinical progression of psoriasis. Theranostics 2020, 10, 12111–12126. [Google Scholar] [CrossRef]
- Story, G. M.; Peier, A. M.; Reeve, A. J.; Eid, S. R.; Mosbacher, J.; Hricik, T. R.; Earley, T. J.; Hergarden, A. C.; Andersson, D. A.; Hwang, S. W.; McIntyre, P.; Jegla, T.; Bevan, S.; Patapoutian, A. , ANKTM1, a TRP-like channel expressed in nociceptive neurons, is activated by cold temperatures. Cell 2003, 112, 819–829. [Google Scholar] [CrossRef]
- Jordt, S. E.; Bautista, D. M.; Chuang, H. H.; McKemy, D. D.; Zygmunt, P. M.; Högestätt, E. D.; Meng, I. D.; Julius, D. , Mustard oils and cannabinoids excite sensory nerve fibres through the TRP channel ANKTM1. Nature 2004, 427, 260–265. [Google Scholar] [CrossRef]
- del Camino, D.; Murphy, S.; Heiry, M.; Barrett, L. B.; Earley, T. J.; Cook, C. A.; Petrus, M. J.; Zhao, M.; D'Amours, M.; Deering, N.; Brenner, G. J.; Costigan, M.; Hayward, N. J.; Chong, J. A.; Fanger, C. M.; Woolf, C. J.; Patapoutian, A.; Moran, M. M. , TRPA1 contributes to cold hypersensitivity. J Neurosci 2010, 30, 15165–15174. [Google Scholar] [CrossRef]
- Bandell, M.; Story, G. M.; Hwang, S. W.; Viswanath, V.; Eid, S. R.; Petrus, M. J.; Earley, T. J.; Patapoutian, A. , Noxious cold ion channel TRPA1 is activated by pungent compounds and bradykinin. Neuron 2004, 41, 849–857. [Google Scholar] [CrossRef]
- Trevisani, M.; Siemens, J.; Materazzi, S.; Bautista, D. M.; Nassini, R.; Campi, B.; Imamachi, N.; Andrè, E.; Patacchini, R.; Cottrell, G. S.; Gatti, R.; Basbaum, A. I.; Bunnett, N. W.; Julius, D.; Geppetti, P. , 4-Hydroxynonenal, an endogenous aldehyde, causes pain and neurogenic inflammation through activation of the irritant receptor TRPA1. Proc Natl Acad Sci USA 2007, 104, 13519–13524. [Google Scholar] [CrossRef] [PubMed]
- Bautista, D. M.; Jordt, S. E.; Nikai, T.; Tsuruda, P. R.; Read, A. J.; Poblete, J.; Yamoah, E. N.; Basbaum, A. I.; Julius, D. , TRPA1 mediates the inflammatory actions of environmental irritants and proalgesic agents. Cell 2006, 124, 1269–1282. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J.; Minassi, A.; Prenen, J.; Taglialatela-Scafati, O.; Appendino, G.; Nilius, B. , Umbellulone modulates TRP channels. Pflugers Arch 2011, 462, 861–870. [Google Scholar] [CrossRef]
- Dai, Y.; Wang, S.; Tominaga, M.; Yamamoto, S.; Fukuoka, T.; Higashi, T.; Kobayashi, K.; Obata, K.; Yamanaka, H.; Noguchi, K. , Sensitization of TRPA1 by PAR2 contributes to the sensation of inflammatory pain. J Clin Invest 2007, 117, 1979–1987. [Google Scholar] [CrossRef] [PubMed]
- Kopruszinski, C. M.; Thornton, P.; Arnold, J.; Newton, P.; Lowne, D.; Navratilova, E.; Swiokla, J.; Dodick, D. W.; Dobson, C.; Gurrell, I.; Chessell, I. P.; Porreca, F. , Characterization and preclinical evaluation of a protease activated receptor 2 (PAR2) monoclonal antibody as a preventive therapy for migraine. Cephalalgia 2020, 40, 1535–1550. [Google Scholar] [CrossRef] [PubMed]
- Demartini, C.; Tassorelli, C.; Zanaboni, A. M.; Tonsi, G.; Francesconi, O.; Nativi, C.; Greco, R. , The role of the transient receptor potential ankyrin type-1 (TRPA1) channel in migraine pain: evaluation in an animal model. J Headache Pain 2017, 18, 94. [Google Scholar] [CrossRef]
- Kim, S. J.; Yeo, J. H.; Yoon, S. Y.; Roh, D. H. , Different Involvement of ASIC and TRPA1 in Facial and Hindpaw Allodynia in Nitroglycerin-Induced Peripheral Hypersensitivities in Mice. Life (Basel) 2022, 12. [Google Scholar] [CrossRef]
- Marone, I. M.; De Logu, F.; Nassini, R.; De Carvalho Goncalves, M.; Benemei, S.; Ferreira, J.; Jain, P.; Li Puma, S.; Bunnett, N. W.; Geppetti, P.; Materazzi, S. , TRPA1/NOX in the soma of trigeminal ganglion neurons mediates migraine-related pain of glyceryl trinitrate in mice. Brain 2018, 141, 2312–2328. [Google Scholar] [CrossRef] [PubMed]
- Materazzi, S.; Benemei, S.; Fusi, C.; Gualdani, R.; De Siena, G.; Vastani, N.; Andersson, D. A.; Trevisan, G.; Moncelli, M. R.; Wei, X.; Dussor, G.; Pollastro, F.; Patacchini, R.; Appendino, G.; Geppetti, P.; Nassini, R. , Parthenolide inhibits nociception and neurogenic vasodilatation in the trigeminovascular system by targeting the TRPA1 channel. Pain 2013, 154, 2750–2758. [Google Scholar] [CrossRef] [PubMed]
- Benemei, S.; De Logu, F.; Li Puma, S.; Marone, I. M.; Coppi, E.; Ugolini, F.; Liedtke, W.; Pollastro, F.; Appendino, G.; Geppetti, P.; Materazzi, S.; Nassini, R. , The anti-migraine component of butterbur extracts, isopetasin, desensitizes peptidergic nociceptors by acting on TRPA1 cation channel. Br J Pharmacol 2017, 174, 2897–2911. [Google Scholar] [CrossRef] [PubMed]
- McKemy, D. D.; Neuhausser, W. M.; Julius, D. , Identification of a cold receptor reveals a general role for TRP channels in thermosensation. Nature 2002, 416, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Peier, A. M.; Moqrich, A.; Hergarden, A. C.; Reeve, A. J.; Andersson, D. A.; Story, G. M.; Earley, T. J.; Dragoni, I.; McIntyre, P.; Bevan, S.; Patapoutian, A. , A TRP channel that senses cold stimuli and menthol. Cell 2002, 108, 705–715. [Google Scholar] [CrossRef]
- Zakharian, E.; Cao, C.; Rohacs, T. , Gating of transient receptor potential melastatin 8 (TRPM8) channels activated by cold and chemical agonists in planar lipid bilayers. J Neurosci 2010, 30, 12526–12534. [Google Scholar] [CrossRef] [PubMed]
- Lippoldt, E. K.; Elmes, R. R.; McCoy, D. D.; Knowlton, W. M.; McKemy, D. D. , Artemin, a glial cell line-derived neurotrophic factor family member, induces TRPM8-dependent cold pain. J Neurosci 2013, 33, 12543–12552. [Google Scholar] [CrossRef]
- Asuthkar, S.; Demirkhanyan, L.; Sun, X.; Elustondo, P. A.; Krishnan, V.; Baskaran, P.; Velpula, K. K.; Thyagarajan, B.; Pavlov, E. V.; Zakharian, E. , The TRPM8 protein is a testosterone receptor: II. Functional evidence for an ionotropic effect of testosterone on TRPM8. J Biol Chem 2015, 290, 2670–2688. [Google Scholar]
- Sousa-Valente, J.; Andreou, A. P.; Urban, L.; Nagy, I. , Transient receptor potential ion channels in primary sensory neurons as targets for novel analgesics. Br J Pharmacol 2014, 171, 2508–2527. [Google Scholar] [CrossRef]
- Chasman, D. I.; Schürks, M.; Anttila, V.; de Vries, B.; Schminke, U.; Launer, L. J.; Terwindt, G. M.; van den Maagdenberg, A. M.; Fendrich, K.; Völzke, H.; Ernst, F.; Griffiths, L. R.; Buring, J. E.; Kallela, M.; Freilinger, T.; Kubisch, C.; Ridker, P. M.; Palotie, A.; Ferrari, M. D.; Hoffmann, W.; Zee, R. Y.; Kurth, T. , Genome-wide association study reveals three susceptibility loci for common migraine in the general population. Nat Genet 2011, 43, 695–698. [Google Scholar] [CrossRef]
- Freilinger, T.; Anttila, V.; de Vries, B.; Malik, R.; Kallela, M.; Terwindt, G. M.; Pozo-Rosich, P.; Winsvold, B.; Nyholt, D. R.; van Oosterhout, W. P.; Artto, V.; Todt, U.; Hämäläinen, E.; Fernández-Morales, J.; Louter, M. A.; Kaunisto, M. A.; Schoenen, J.; Raitakari, O.; Lehtimäki, T.; Vila-Pueyo, M.; Göbel, H.; Wichmann, E.; Sintas, C.; Uitterlinden, A. G.; Hofman, A.; Rivadeneira, F.; Heinze, A.; Tronvik, E.; van Duijn, C. M.; Kaprio, J.; Cormand, B.; Wessman, M.; Frants, R. R.; Meitinger, T.; Müller-Myhsok, B.; Zwart, J. A.; Färkkilä, M.; Macaya, A.; Ferrari, M. D.; Kubisch, C.; Palotie, A.; Dichgans, M.; van den Maagdenberg, A. M. , Genome-wide association analysis identifies susceptibility loci for migraine without aura. Nat Genet 2012, 44, 777–782. [Google Scholar] [CrossRef] [PubMed]
- Sintas, C.; Fernández-Morales, J.; Vila-Pueyo, M.; Narberhaus, B.; Arenas, C.; Pozo-Rosich, P.; Macaya, A.; Cormand, B. , Replication study of previous migraine genome-wide association study findings in a Spanish sample of migraine with aura. Cephalalgia 2015, 35, 776–782. [Google Scholar] [CrossRef]
- Ling, Y.-H.; Chen, S.-P.; Fann, C. S.-J.; Wang, S.-J.; Wang, Y.-F. , TRPM8 genetic variant is associated with chronic migraine and allodynia. The Journal of Headache and Pain 2019, 20. [Google Scholar] [CrossRef] [PubMed]
- Burstein, R.; Yarnitsky, D.; Goor-Aryeh, I.; Ransil, B. J.; Bajwa, Z. H. , An association between migraine and cutaneous allodynia. Ann Neurol 2000, 47, 614–624. [Google Scholar] [CrossRef]
- Borhani Haghighi, A.; Motazedian, S.; Rezaii, R.; Mohammadi, F.; Salarian, L.; Pourmokhtari, M.; Khodaei, S.; Vossoughi, M.; Miri, R. , Cutaneous application of menthol 10% solution as an abortive treatment of migraine without aura: a randomised, double-blind, placebo-controlled, crossed-over study. Int J Clin Pract 2010, 64, 451–456. [Google Scholar] [CrossRef] [PubMed]
- Takashima, Y.; Daniels, R. L.; Knowlton, W.; Teng, J.; Liman, E. R.; McKemy, D. D. , Diversity in the neural circuitry of cold sensing revealed by genetic axonal labeling of transient receptor potential melastatin 8 neurons. J Neurosci 2007, 27, 14147–14157. [Google Scholar] [CrossRef] [PubMed]
- Dhaka, A.; Earley, T. J.; Watson, J.; Patapoutian, A. , Visualizing cold spots: TRPM8-expressing sensory neurons and their projections. J Neurosci 2008, 28, 566–575. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Fukuoka, T.; Obata, K.; Yamanaka, H.; Dai, Y.; Tokunaga, A.; Noguchi, K. , Distinct expression of TRPM8, TRPA1, and TRPV1 mRNAs in rat primary afferent neurons with adelta/c-fibers and colocalization with trk receptors. J Comp Neurol 2005, 493, 596–606. [Google Scholar] [CrossRef]
- Ordás, P.; Hernández-Ortego, P.; Vara, H.; Fernández-Peña, C.; Reimúndez, A.; Morenilla-Palao, C.; Guadaño-Ferraz, A.; Gomis, A.; Hoon, M.; Viana, F.; Señarís, R. , Expression of the cold thermoreceptor TRPM8 in rodent brain thermoregulatory circuits. J Comp Neurol 2021, 529, 234–256. [Google Scholar] [CrossRef]
- Dussor, G.; Cao, Y. Q. , TRPM8 and Migraine. Headache 2016, 56, 1406–1417. [Google Scholar] [CrossRef]
- Burgos-Vega, C. C.; Ahn, D. D.; Bischoff, C.; Wang, W.; Horne, D.; Wang, J.; Gavva, N.; Dussor, G. , Meningeal transient receptor potential channel M8 activation causes cutaneous facial and hindpaw allodynia in a preclinical rodent model of headache. Cephalalgia 2016, 36, 185–193. [Google Scholar] [CrossRef]
- Ren, L.; Dhaka, A.; Cao, Y. Q. , Function and postnatal changes of dural afferent fibers expressing TRPM8 channels. Mol Pain 2015, 11, 37. [Google Scholar] [CrossRef]
- Alarcón-Alarcón, D.; Cabañero, D.; De Andrés-López, J.; Nikolaeva-Koleva, M.; Giorgi, S.; Fernández-Ballester, G.; Fernández-Carvajal, A.; Ferrer-Montiel, A. , TRPM8 contributes to sex dimorphism by promoting recovery of normal sensitivity in a mouse model of chronic migraine. Nature Communications 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.; Kim, B.; McKemy, D. D. , Transient receptor potential melastatin 8 is required for nitroglycerin- and calcitonin gene-related peptide-induced migraine-like pain behaviors in mice. Pain 2022, 163, 2380–2389. [Google Scholar] [CrossRef] [PubMed]
- Liedtke, W.; Choe, Y.; Martí-Renom, M. A.; Bell, A. M.; Denis, C. S.; Sali, A.; Hudspeth, A. J.; Friedman, J. M.; Heller, S. , Vanilloid receptor-related osmotically activated channel (VR-OAC), a candidate vertebrate osmoreceptor. Cell 2000, 103, 525–535. [Google Scholar] [CrossRef] [PubMed]
- Vriens, J.; Watanabe, H.; Janssens, A.; Droogmans, G.; Voets, T.; Nilius, B. , Cell swelling, heat, and chemical agonists use distinct pathways for the activation of the cation channel TRPV4. Proc Natl Acad Sci U S A 2004, 101, 396–401. [Google Scholar] [CrossRef]
- Grant, A. D.; Cottrell, G. S.; Amadesi, S.; Trevisani, M.; Nicoletti, P.; Materazzi, S.; Altier, C.; Cenac, N.; Zamponi, G. W.; Bautista-Cruz, F.; Lopez, C. B.; Joseph, E. K.; Levine, J. D.; Liedtke, W.; Vanner, S.; Vergnolle, N.; Geppetti, P.; Bunnett, N. W. , Protease-activated receptor 2 sensitizes the transient receptor potential vanilloid 4 ion channel to cause mechanical hyperalgesia in mice. J Physiol 2007, 578, (Pt 3), 715–733. [Google Scholar] [CrossRef]
- Wei, X.; Edelmayer, R. M.; Yan, J.; Dussor, G. , Activation of TRPV4 on dural afferents produces headache-related behavior in a preclinical rat model. Cephalalgia 2011, 31, 1595–1600. [Google Scholar] [CrossRef]
- Abramowitz, J.; Birnbaumer, L. , Physiology and pathophysiology of canonical transient receptor potential channels. Faseb j 2009, 23, 297–328. [Google Scholar] [CrossRef]
- Löf, C.; Viitanen, T.; Sukumaran, P.; Törnquist, K. , TRPC2: of mice but not men. Adv Exp Med Biol 2011, 704, 125–134. [Google Scholar]
- Chen, X.; Sooch, G.; Demaree, I. S.; White, F. A.; Obukhov, A. G. , Transient Receptor Potential Canonical (TRPC) Channels: Then and Now. Cells 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Okada, T.; Inoue, R.; Yamazaki, K.; Maeda, A.; Kurosaki, T.; Yamakuni, T.; Tanaka, I.; Shimizu, S.; Ikenaka, K.; Imoto, K.; Mori, Y. , Molecular and functional characterization of a novel mouse transient receptor potential protein homologue TRP7. Ca(2+)-permeable cation channel that is constitutively activated and enhanced by stimulation of G protein-coupled receptor. J Biol Chem 1999, 274, 27359–27370. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, T.; Obukhov, A. G.; Schaefer, M.; Harteneck, C.; Gudermann, T.; Schultz, G. , Direct activation of human TRPC6 and TRPC3 channels by diacylglycerol. Nature 1999, 397, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Storch, U.; Forst, A. L.; Pardatscher, F.; Erdogmus, S.; Philipp, M.; Gregoritza, M.; Mederos, Y. S. M.; Gudermann, T. , Dynamic NHERF interaction with TRPC4/5 proteins is required for channel gating by diacylglycerol. Proc Natl Acad Sci U S A 2017, 114, E37–e46. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Li, W.; Riley, A. M.; Soliman, M.; Chakraborty, S.; Stamatkin, C. W.; Obukhov, A. G. , Molecular Determinants of the Sensitivity to Gq/11-Phospholipase C-dependent Gating, Gd3+ Potentiation, and Ca2+ Permeability in the Transient Receptor Potential Canonical Type 5 (TRPC5) Channel. J Biol Chem 2017, 292, 898–911. [Google Scholar] [CrossRef]
- Bollimuntha, S.; Selvaraj, S.; Singh, B. B. , Emerging roles of canonical TRP channels in neuronal function. Adv Exp Med Biol 2011, 704, 573–593. [Google Scholar]
- Riccio, A.; Li, Y.; Tsvetkov, E.; Gapon, S.; Yao, G. L.; Smith, K. S.; Engin, E.; Rudolph, U.; Bolshakov, V. Y.; Clapham, D. E. , Decreased anxiety-like behavior and Gαq/11-dependent responses in the amygdala of mice lacking TRPC4 channels. J Neurosci 2014, 34, 3653–3667. [Google Scholar] [CrossRef]
- Just, S.; Chenard, B. L.; Ceci, A.; Strassmaier, T.; Chong, J. A.; Blair, N. T.; Gallaschun, R. J.; Del Camino, D.; Cantin, S.; D'Amours, M.; Eickmeier, C.; Fanger, C. M.; Hecker, C.; Hessler, D. P.; Hengerer, B.; Kroker, K. S.; Malekiani, S.; Mihalek, R.; McLaughlin, J.; Rast, G.; Witek, J.; Sauer, A.; Pryce, C. R.; Moran, M. M. , Treatment with HC-070, a potent inhibitor of TRPC4 and TRPC5, leads to anxiolytic and antidepressant effects in mice. PLoS One 2018, 13, e0191225. [Google Scholar] [CrossRef]
- Riccio, A.; Li, Y.; Moon, J.; Kim, K. S.; Smith, K. S.; Rudolph, U.; Gapon, S.; Yao, G. L.; Tsvetkov, E.; Rodig, S. J.; Van't Veer, A.; Meloni, E. G.; Carlezon, W. A., Jr.; Bolshakov, V. Y.; Clapham, D. E. , Essential role for TRPC5 in amygdala function and fear-related behavior. Cell 2009, 137, 761–772. [Google Scholar] [CrossRef]
- Lee, S. H.; Tonello, R.; Choi, Y.; Jung, S. J.; Berta, T. , Sensory Neuron–Expressed TRPC4 Is a Target for the Relief of Psoriasiform Itch and Skin Inflammation in Mice. Journal of Investigative Dermatology 2020, 140, 2221–2229. [Google Scholar] [CrossRef]
- Cohen, C. F.; Prudente, A. S.; Berta, T.; Lee, S. H. , Transient Receptor Potential Channel 4 Small-Molecule Inhibition Alleviates Migraine-Like Behavior in Mice. Front Mol Neurosci 2021, 14, 765181. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Huang, W.; Richardson, P. M.; Priestley, J. V.; Liu, M. , TRPC4 in Rat Dorsal Root Ganglion Neurons Is Increased after Nerve Injury and Is Necessary for Neurite Outgrowth. Journal of Biological Chemistry 2008, 283, 416–426. [Google Scholar] [CrossRef] [PubMed]
- Westlund, K. N.; Zhang, L. P.; Ma, F.; Nesemeier, R.; Ruiz, J. C.; Ostertag, E. M.; Crawford, J. S.; Babinski, K.; Marcinkiewicz, M. M. , A rat knockout model implicates TRPC4 in visceral pain sensation. Neuroscience 2014, 262, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Sadler, K. E.; Moehring, F.; Shiers, S. I.; Laskowski, L. J.; Mikesell, A. R.; Plautz, Z. R.; Brezinski, A. N.; Mecca, C. M.; Dussor, G.; Price, T. J.; McCorvy, J. D.; Stucky, C. L. , Transient receptor potential canonical 5 mediates inflammatory mechanical and spontaneous pain in mice. Sci Transl Med 2021, 13. [Google Scholar] [CrossRef]
- Lee, S. H.; Cho, P. S.; Tonello, R.; Lee, H. K.; Jang, J. H.; Park, G. Y.; Hwang, S. W.; Park, C. K.; Jung, S. J.; Berta, T. , Peripheral serotonin receptor 2B and transient receptor potential channel 4 mediate pruritus to serotonergic antidepressants in mice. J Allergy Clin Immunol 2018, 142, 1349–1352. [Google Scholar] [CrossRef] [PubMed]
- Alom, F.; Miyakawa, M.; Matsuyama, H.; Nagano, H.; Tanahashi, Y.; Unno, T. , Possible antagonistic effects of the TRPC4 channel blocker ML204 on M<sub>2</sub> and M<sub>3</sub> muscarinic receptors in mouse ileal and detrusor smooth muscles and atrial myocardium. Journal of Veterinary Medical Science 2018, 80, 1407–1415. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.; Shi, J.; Zhu, Y.; Kustov, M.; Tian, J.-B.; Stevens, A.; Wu, M.; Xu, J.; Long, S.; Yang, P.; Zholos, A. V.; Salovich, J. M.; Weaver, C. D.; Hopkins, C. R.; Lindsley, C. W.; McManus, O.; Li, M.; Zhu, M. X. , Identification of ML204, a Novel Potent Antagonist That Selectively Modulates Native TRPC4/C5 Ion Channels. Journal of Biological Chemistry 2011, 286, 33436–33446. [Google Scholar] [CrossRef]
- Pradhan, A. A.; Smith, M. L.; McGuire, B.; Tarash, I.; Evans, C. J.; Charles, A. , Characterization of a novel model of chronic migraine. Pain 2014, 155, 269–274. [Google Scholar] [CrossRef]
- Mathew, N. T.; Kailasam, J.; Seifert, T. , Clinical recognition of allodynia in migraine. Neurology 2004, 63, 848–852. [Google Scholar] [CrossRef]
- Avona, A.; Burgos-Vega, C.; Burton, M. D.; Akopian, A. N.; Price, T. J.; Dussor, G. , Dural Calcitonin Gene-Related Peptide Produces Female-Specific Responses in Rodent Migraine Models. J Neurosci 2019, 39, 4323–4331. [Google Scholar] [CrossRef]
- Stewart, W. F.; Wood, C.; Reed, M. L.; Roy, J.; Lipton, R. B. , Cumulative lifetime migraine incidence in women and men. Cephalalgia 2008, 28, 1170–1178. [Google Scholar] [CrossRef] [PubMed]
- Eftekhari, S.; Edvinsson, L. , Calcitonin gene-related peptide (CGRP) and its receptor components in human and rat spinal trigeminal nucleus and spinal cord at C1-level. BMC Neurosci 2011, 12, 112. [Google Scholar] [CrossRef] [PubMed]
- Maleki, N.; Androulakis, X. M. , Is There Any MRI Pattern That Discriminates Female From Male Migraine Patients? Front Neurol 2019, 10, 961. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Strassman, A. M.; Novack, V.; Brin, M. F.; Burstein, R. , Extracranial injections of botulinum neurotoxin type A inhibit intracranial meningeal nociceptors’ responses to stimulation of TRPV1 and TRPA1 channels: Are we getting closer to solving this puzzle? Cephalalgia 2016, 36, 875–886. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, J.; Supronsinchai, W.; Andreou, A. P.; Summ, O.; Akerman, S.; Goadsby, P. J. , Olvanil acts on transient receptor potential vanilloid channel 1 and cannabinoid receptors to modulate neuronal transmission in the trigeminovascular system. Pain 2012, 153, 2226–2232. [Google Scholar] [CrossRef] [PubMed]
- Nicoletti, P.; Trevisani, M.; Manconi, M.; Gatti, R.; De Siena, G.; Zagli, G.; Benemei, S.; Capone, J. A.; Geppetti, P.; Pini, L. A. , Ethanol causes neurogenic vasodilation by TRPV1 activation and CGRP release in the trigeminovascular system of the guinea pig. Cephalalgia 2008, 28, 9–17. [Google Scholar] [CrossRef]
- Zakharov, A.; Vitale, C.; Kilinc, E.; Koroleva, K.; Fayuk, D.; Shelukhina, I.; Naumenko, N.; Skorinkin, A.; Khazipov, R.; Giniatullin, R. , Hunting for origins of migraine pain: cluster analysis of spontaneous and capsaicin-induced firing in meningeal trigeminal nerve fibers. Front Cell Neurosci 2015, 9, 287. [Google Scholar] [CrossRef]
- Edelmayer, R. M.; Le, L. N.; Yan, J.; Wei, X.; Nassini, R.; Materazzi, S.; Preti, D.; Appendino, G.; Geppetti, P.; Dodick, D. W.; Vanderah, T. W.; Porreca, F.; Dussor, G. , Activation of TRPA1 on dural afferents: a potential mechanism of headache pain. Pain 2012, 153, 1949–1958. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Wang, Y.; Xu, Y.; Ma, D.; Wang, M. , The Transient Receptor Potential Ankyrin Type 1 Plays a Critical Role in Cortical Spreading Depression. Neuroscience 2018, 382, 23–34. [Google Scholar] [CrossRef]
- Kayama, Y.; Shibata, M.; Takizawa, T.; Ibata, K.; Shimizu, T.; Ebine, T.; Toriumi, H.; Yuzaki, M.; Suzuki, N. , Functional interactions between transient receptor potential M8 and transient receptor potential V1 in the trigeminal system: Relevance to migraine pathophysiology. Cephalalgia 2018, 38, 833–845. [Google Scholar] [CrossRef]
- Chen, Y.; Kanju, P.; Fang, Q.; Lee, S. H.; Parekh, P. K.; Lee, W.; Moore, C.; Brenner, D.; Gereau, R. W. t.; Wang, F.; Liedtke, W. , TRPV4 is necessary for trigeminal irritant pain and functions as a cellular formalin receptor. Pain 2014, 155. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Physiology of the migraine pathogenesis and pharmacological targets. (A) Migraine pain axis comprising peripheral sites such as the trigeminal ganglia (TG) and meninges as well as central nervous system structures, including the spinal trigeminal nucleus (SpV) and thalamus. (B) Nociceptor activation or cortical spreading depression (CSD) triggers the release of inflammatory and nociceptive neuropeptides from the trigeminal terminals innervating the meninges and associated blood vessels. These neuropeptides induce vasodilation and sterile neurogenic inflammation involving mast cell degranulation and release of proinflammatory mediators. This in turn leads to sensitization of second-order neurons in the SpV, which increases nociceptive signaling to the thalamus and higher order cortical brain structures that process pain. (C-E) Also depicted is the contribution of CGRP to migraine pathophysiology and the mechanism of action of Triptans and Botox.
Figure 1.
Physiology of the migraine pathogenesis and pharmacological targets. (A) Migraine pain axis comprising peripheral sites such as the trigeminal ganglia (TG) and meninges as well as central nervous system structures, including the spinal trigeminal nucleus (SpV) and thalamus. (B) Nociceptor activation or cortical spreading depression (CSD) triggers the release of inflammatory and nociceptive neuropeptides from the trigeminal terminals innervating the meninges and associated blood vessels. These neuropeptides induce vasodilation and sterile neurogenic inflammation involving mast cell degranulation and release of proinflammatory mediators. This in turn leads to sensitization of second-order neurons in the SpV, which increases nociceptive signaling to the thalamus and higher order cortical brain structures that process pain. (C-E) Also depicted is the contribution of CGRP to migraine pathophysiology and the mechanism of action of Triptans and Botox.
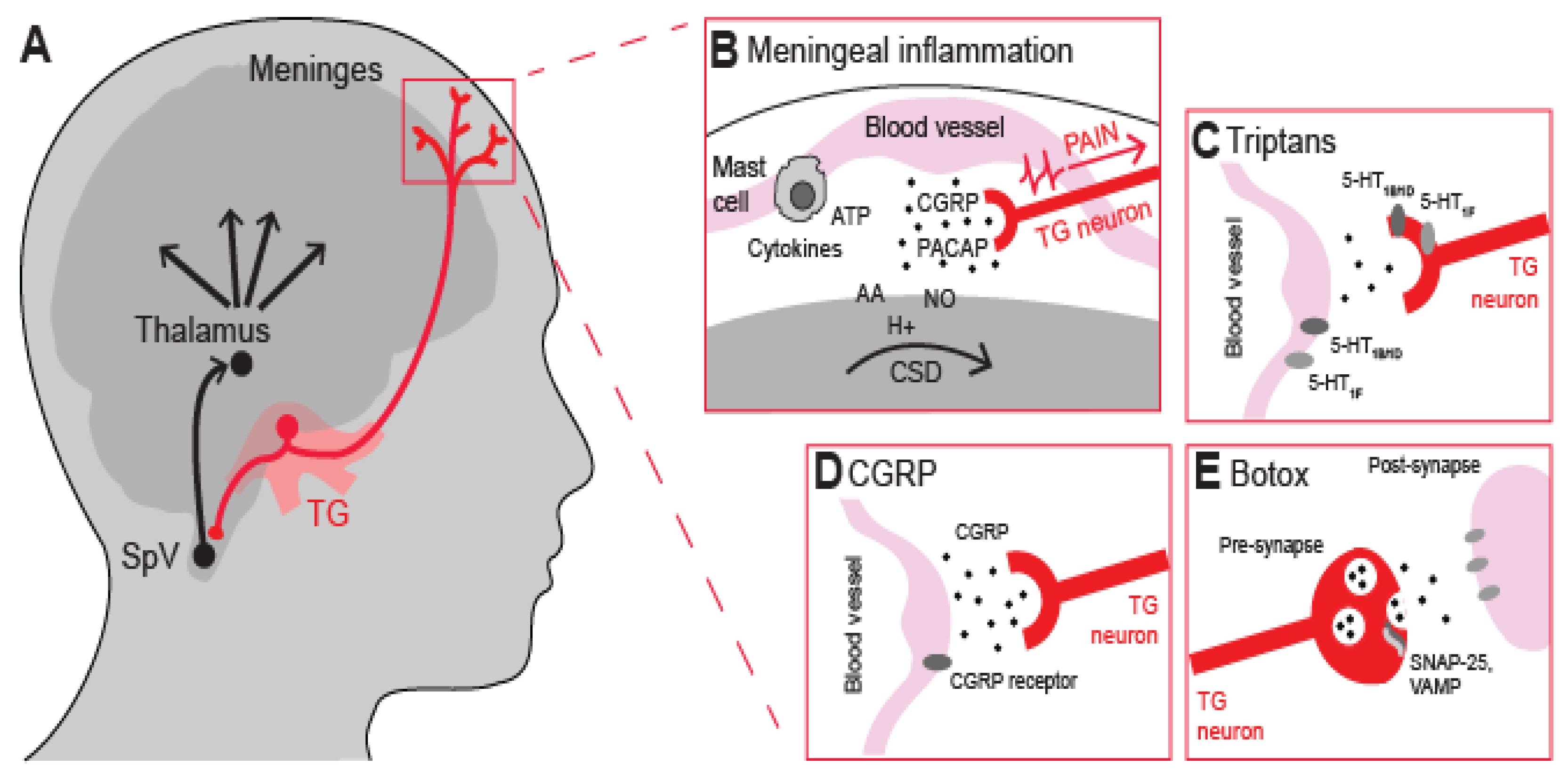
Figure 2.
TRPC4 inhibition alleviates nitroglycerin (NTG)-induced migraine in mice. (A) Illustration of the mouse models used for acute (episodic) and chronic migraine, which consist of single or repeated intraperitoneal (i.p.) injections of NTG. NTG induces mechanical allodynia that can be assessed by applying von Frey filaments to the hindpaw. (B) Inhibiting TRPC4 with the ML204 antagonist or small interfering RNA (siRNA) has the potential to exert antimigraine effects by blocking both CGRP release in peripheral tissues and pain transmission to the central nervous system (CNS).
Figure 2.
TRPC4 inhibition alleviates nitroglycerin (NTG)-induced migraine in mice. (A) Illustration of the mouse models used for acute (episodic) and chronic migraine, which consist of single or repeated intraperitoneal (i.p.) injections of NTG. NTG induces mechanical allodynia that can be assessed by applying von Frey filaments to the hindpaw. (B) Inhibiting TRPC4 with the ML204 antagonist or small interfering RNA (siRNA) has the potential to exert antimigraine effects by blocking both CGRP release in peripheral tissues and pain transmission to the central nervous system (CNS).
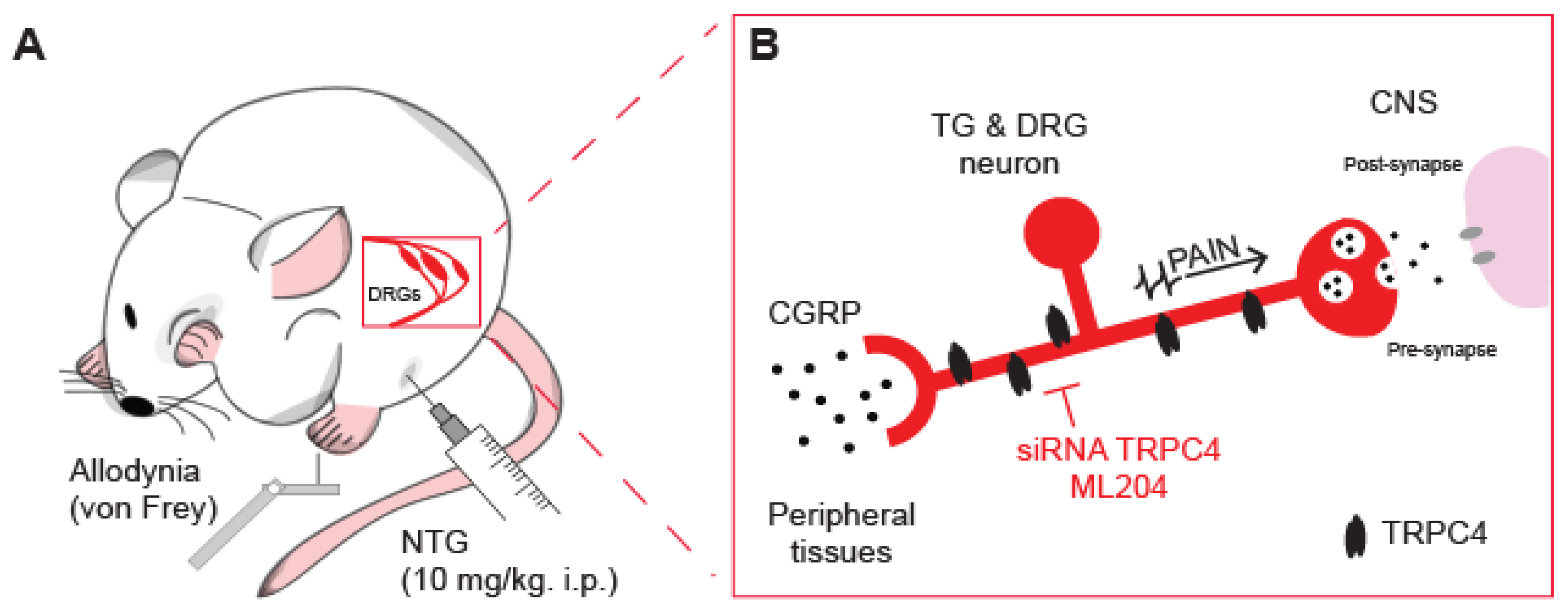

Table 1.
Studies exploring the role of TRP channels in migraine.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



