Submitted:
06 April 2023
Posted:
10 April 2023
You are already at the latest version
Abstract
The term ‘Hallmarks of Cancer’ was coined by Hanahan and Weinberg in their influential reviews and they described genome instability as a property of cells enabling cancer development [1, 2]. Accurate DNA replication of genomes is central to diminish genome instability. Here, the understanding of the initiation of DNA synthesis in origins of DNA replication to start leading strand synthesis and the initiation of Okazaki fragment on the lagging strand are crucial to control genome instability. Recent findings have provided new insights into the mechanism of the remodelling of the prime initiation enzyme, DNA polymerase α-primase, during primer synthesis, how the enzyme complex achieves lagging strand synthesis, and how it is linked to replication forks to achieve optimal initiation of Okazaki fragments. Moreover, the central roles of RNA primer synthesis by Pol-prim in multiple genome stability pathways such as replication fork restart and protection of DNA against degradation by exonucleases during double-strand break repair is discussed.
Keywords:
Genome stability
; DNA replication
; DNA repair
; lagging strand DNA synthesis
; Okazaki fragments
; initiation
; DNA polymerase α
; DNA primase
; CTC1-STN1-TEN1 complex
; SV40 T antigen
; CMG complex
1. Introduction
DNA replication is essential for all living organisms and accurate DNA duplication is at the heart of preventing genome instability and associated diseases such as cancer and other genetic diseases [3,4,5]. Eukaryotic DNA replication is a highly conserved and tightly regulated process, which occurs once and only once per cell cycle in the synthesis phase or short S phase of the eukaryotic cell cycle [6]. The accurate, semiconservative synthesis of chromosomal DNA is central for the duplication of eukaryotic cells and essential for the maintenance of their genetic information. The DNA replication can be divided into multiple steps the pre-initiation, the initiation, the elongation and the termination step. The linear nature of eukaryotic chromosomes creates an additional problem for their maintenance – the incomplete duplication of the telomeric ends on the lagging strand [3,4,5,7].
The understanding of DNA replication processes requires mechanistic knowledge about the coordination of the various multiprotein complexes involved in cellular DNA replication, from the origin activation, to double-stranded DNA (dsDNA) unwinding to form single-stranded DNA (ssDNA) templates, and the establishment of replication forks to the final DNA replication products. Generally, the DNA sequences, at which DNA replication is initiated first on a chromosome and at which replication fork(s) are established, are called origins of DNA replication or ‘oris’ for short [3,4,5,8,9]. After the separation of the two strands of the parental dsDNA, these newly established ssDNAs serve as templates for DNA polymerases, which synthesize their complementary strands. The polarities of these ssDNAs (5’-3’ and 3’-5’ when looking at the residues of the deoxyribose moiety) and the requirement of all DNA polymerase to synthesize nucleic acids in 5’ to 3’ direction (template direction 3’ to 5’) requires that at a given replication fork one strand, the leading strand, is synthesized in a continuous fashion whereas the other strand, called lagging strand, is synthesized discontinuously in form of Okazaki fragments [3,4,5]. Here it is important to note that eukaryotic DNA polymerases lack the ability to start DNA synthesis de novo and require a starter molecule, an RNA primer, for synthesising DNA. Whereas the continuous nature of leading DNA synthesis requires only primer per replisome, the lagging strand synthesis requires an initiation event every one to two hundred nucleotides [3,4,5,10,11]. The latter needs further treatment and maturation to finally yield the second continuous newly synthesized strand. Thus, understanding the mechanism and regulation of the initiation of DNA replication is central for the understanding of the DNA replication process in the eukaryotic cell cycle [3,4,5].
2. Initiation of DNA Replication at Origins
Each eukaryotic genome consists of multiple chromosomes, which in turn contain numerous origins of DNA replication, which regulate the unwinding of dsDNA and the start of semiconservative DNA replication e.g., that replication of a given DNA is replicated once and only once per cell cycle [4,12]. They serve as functional organizer and each replicon or replication unit in a given chromosome contains one origin. Prior to the start of DNA replication in S phase numerous coordinated early events occur. The pre-replicative complex is formed and involves origin recognition processes by the protein complex ORC (origin recognition complex, consisting of proteins ORC1 to ORC6), the loading of the MCM2 to MCM6 (minichromosome maintenance 2 to 7) proteins, the core of eukaryotic replicative helicases, with the help of CDC6 and CDT1/TAH11, their activation, and the formation of the pre-initiation complex [8]. These processes have been recently reviewed in detail [8] and therefore, are not the focus of the current review.
3. Replication Forks
The unwinding of dsDNA at eukaryotic DNA replication origin by the CDC45/MCM2-6/GINS (CMG) complex establishes two replication forks [3,4,5,8,9]. Each reapplication fork forms the junction between the chromosomal dsDNA and the two single-stranded template DNAs. One strand serves as template for continuous DNA synthesis called leading strand, whereas the other is the template for fragmented, discontinuous DNA synthesis known as lagging strand. Each of the discontinuous replication products is initiated by DNA Polymerase α-DNA primase (Pol-prim) producing a RNA-DNA which are then extended by DNA polymerase δ (Pol δ). In contrast, DNA polymerase ε (Pol ε) synthesises DNA on the leading strand in a continuous manner [3,4,5,9,13].
- a. CDC45/MCM2-6/GINS complex
The CMG complex is a very large protein complex and consists of the replication factor CDC45 plus two protein sub-complexes the MCM2-7 and GINS complex (having SLD5-PSF1-3) with MCM2-7 forming a hexameric ring around unwound ssDNA. CMG is the eukaryotic replicative helicase, which moves on the leading strand in 3’-5’ direction ahead of and in association with Pol ε. The enzyme complex unwinds dsDNA to provide the ssDNA templates for the replicative DNA polymerases [3,4,5,8,9]. Additionally, several proteins including AND-1/CTF4, Claspin/MRC3, Timeless/TOF1, and Tipin (Timeles interacting protein)/CMS3 associate with CMG and stabilise the replication fork including linking them to replicative DNA polymerases [3,4,5,8,9,13,14].
- b. The replicative DNA polymerases
- b1. DNA polymerase α-DNA primase
Pol-prim consists of four subunits: the large catalytic DNA polymerase subunit, PolA1 or p180, PolA2/p68/p70/B subunit, PRI2/Prim2/PriL/p58, and PRI1/Prim1/PriS/p48/p49, the later two forming the DNA primase complex (Figure 1, [15,16,17,18,19]). The N terminus of PolA1/p180 consists of an intrinsic disordered region (IDR, Figure 1, residue 1 to 368 [20], these residue numbers using the sequence of human PolA1) contains SV40 T antigen (Tag) and AND-1/CTF4-binding activity plus multiple CDK recognition sites [21,22]. This is followed by the polymerase domain (aa 369 to 1220, PolA1Pol), containing an inactive Exo domain, ssDNA binding and nucleotide binding activity, and C-terminal domain (1271 to 1462, PolA1CT). The latter functions as the platform for the binding of the smaller subunits PolA2 and PRI2, the large subunit of primase (Figure 1). The polymerase and the C-terminal domain are connected by a hinge region (aa ~1221 to 1270, Figure 1 [15,16,17,18]).
The second largest subunit PolA2 also called p68/p70 binds to the C-terminus of PolA1 and has no catalytic but regulatory function e.g., it is phosphorylated by Cdk2 in cell-cycle-dependent manner at its N-terminus, which also interacts with SV40 T antigen (Tag) and the human AND-1/CTF4 protein (Figure 1, [12,21,22,24,25,26]). PolA1 and PolA2 form the Pol α core complex [12]. In contrast, the two smaller primase subunits, PRI1 or small primase subunit p49, which carries the catalytic function, and PRI2 or large primase subunit p58, which is responsible for interaction with the large PolA1 subunit and the primer length, carry the initiation functions [15,16,27]. PRI2 is also required for stabilising the enzyme activity of primase subunit of PRI1 in vitro [27]. PRI2 consists of an N-terminal, aa 39 – 252, a C-terminal domain, aa 274 – 460, and a hinge region, aa 253 – 273, connecting the two domains (Figure 1; [15,16]).
- b2. DNA polymerase δ
DNA polymerase δ (Pol δ) is a heterotetrameric protein complex [3,4,13,28]. The largest subunit, p125, contains two enzymatic activities, a DNA polymerase and a highly active proofreading 3’ to 5’ exonuclease. Pol δ synthesises DNA with low processivity but PCNA (Proliferating Cell Nuclear Antigen), the processivity factor and replication clamp of Pol ε and δ, confers Pol δ to a highly processive enzyme synthesising in turn the main part of Okazaki fragments on the lagging strand. Pol δ also possesses efficient strand displacement activity and thus synthesises DNA beyond the RNA primer produced by DNA primase and may even remove most of the DNA primer polymerised by Pol α with the help of FEN1 and DNA2 protein [3,4,13,28].
- b3. DNA polymerase ε
DNA polymerase ε (Pol ε) also a four-subunit protein complex with the largest subunit, PolE/Pol2/p260, containing two enzymatic activities, a highly processive DNA polymerase, and a proofreading 3’ to 5’ exonuclease activity [3,4,13]. Although the Pol ε complex alone synthesises DNA with high processivity, PCNA enhances this processivity allowing synthesis of a whole replicon size of DNA. At the replication fork, the leading strand replicase, Pol ε, physically and functionally interacts with CMG [13,29,30,31,32]. Pol ε binds to CMG and stimulates its helicase activity. Thus, PCNA, Pol ε, and CMG form a sandwich structure on the leading strand with Pol ε in the middle [13,29,32].
- c. CMG-associated replication factors – members of the replication protection complex
CMG has not only enzymatic function e.g., helicase activity, but is also a central organiser of the proteins at the replication fork [13,29,31,33,34]. It forms the logistic center for association of other replication factor e.g., the fork protection complex [29]. Here, AND-1/CTF4 forms homotrimers and recruits Pol-prim to CMG at the replication fork to increase Pol-prim concentration for enhancing the initiation of Okazaki fragment synthesis [22]. AND-1/CTF4 binds via its Sept B domain and HMG-box region to the N-termini of the two largest subunits of Pol-prim (Figure 1). Interestingly, the AND-1/CTF4 Sept B domain binds to the intrinsic disordered region of PolA1, aa 1-368, at position aa 148-171, which contains a CTF4 binding motif and is localised in close vicinity to a Tag binding site, aa 195-313 [20,22,33,35,36]. A second AND-1/CTF4 binding site for Pol-prim is localised in the N-terminus of PolA2 aa1-78 and overlaps with the Tag-binding site of PolA2. Thus, these AND-1/CTF4 bindings size in the N-termini of PolA1 and PolA2 either are close to or overlap the T antigen binding sites, respectively, and may serve similar function. The AND-1/CTF4-Pol-prim interactions could be important for the recruitment and loading of Pol-prim to the lagging strand for initiation [22,36,37]. Additionally Mcm10, an essential replication factor, binds to the MCM2-7 complex and supports loading and stabilising Pol-prim at replication forks via its interaction with the N-terminus of PolA1 [3,4,5,13,38,39]. MCM10 also enhances the binding of Pol-prim to primers and thus may stimulate the hand over of newly synthesised primers [40].
During unperturbed DNA replication, Claspin/MRC1 is necessary to maintain normal rates of replication fork progression [41]. To achieve this function Claspin physically binds to DNA especially to branched or fork-like DNA structures and connects the MCM complex with Pol ε, the leading DNA polymerase. Additionally, CMG interacts with a variety of proteins at the replication forks including Timeless/TOF1 and Tipin/CSM3 [13,31,33,41]. To stabilise replication forks during unperturbed and perturbed DNA replication conditions, Timeless/TOF1 and Tipin/CSM3 form a complex with CMG modulating the intra-S phase checkpoint and CHK1 phosphorylation.
- d. Additional factors supporting eukaryotic DNA replication
- d1. Replication Protein A
Replication protein A (RPA) is heterotrimeric protein complex consisting of RPA70, RPA32, and RPA14, which forms tight complexes with ssDNA [4,42,43,44,45]. RPA’s binding stabilises ssDNA and prevents the formation of secondary structures within ssDNA sequences allowing smooth DNA synthesis by replicative DNA polymerase. Beyond DNA replication, RPA is involved in all branches of eukaryotic DNA metabolism and DNA signalling after replication fork stalling towards ATR and at DNA lesion towards ATM and DNA-dependent protein kinase [4,42,43,44,45].
- d2. Proliferating Cell Nuclear Antigen and Replication factor C
PCNA forms homotrimeric ring structures around DNA and is also known as DNA sliding clamp [3,4]. As such, PCNA enhances the interaction of the replicative Pols δ and ε with the template primer allowing processive DNA synthesis. However, due to its ring shape PCNA needs a helper function for loading on DNA. The replication factor C (RFC), the clamp loader, opens the PCNA ring in an ATP-dependent manner and loads it onto DNA. Additionally, RFC binds to DNA and the replicative DNA polymerases δ and ε [3,4].
3.1. Leading Strand Synthesis
At replication forks, on the leading strand, DNA primase synthesised the RNA primer which Pol α takes over and elongates to form an RNA-DNA primer. The latter, in turn, is taken over and extended further by Pol δ, before Pol ε starts the continuous DNA synthesis for the full replicon length in an unperturbed situation. Here, interactions of Pol ε with the CMG complex is responsible for an enhanced unwinding rate [29,46].
3.2. Lagging Strand Synthesis and the Initiation of Okazaki Fragment Synthesis
The synthesis of the lagging strand has major influences on the maintenance of eukaryotic genomes similar to the leading strand DNA synthesis. Failures during the process of lagging strand synthesis can lead to increase in genome instability [47]. During the replication of a human genome, the replication machine must initiate 20 to 30 million Okazaki fragments. Despite its importance, relatively little is known about initiation process of Okazaki fragments in eukaryotes. Recently, three eukaryotic model systems have shed light on this central process: The lagging strand synthesis of telomere sequences using Pol-prim and CTC1-STN1-TEN1 (CST) complex (Figure 2; N.B in mouse CTC1 and STN1 are equivalent to the AAF132 and AAF44 subunits of alpha-Accessory factor (AAF) [48]). The SV40 system using Tag, RPA plus Pol-prim, and the cellular replication system with purified yeast and human proteins have advanced our understanding [17,18,25,29,32,46,49,50,51,52,53,54,55,56,57,58].
- a. Biochemical model systems for the initiation of Okazaki fragment synthesis
- a1. Lagging strand synthesis initiation at telomeres
The recent publication of four-landmark articles has produced major advances in our understanding of the initiation of Okazaki fragment synthesis at telomere sequences and beyond on the mechanistic level and in general ([17,18,49,58] reviewed in [51]). Telomere sequences consist of repetitive DNA sequences at the end of linear chromosomes of eukaryotes and form specific protein-DNA complexes, the sheltering complex, to stabilise these ends during normal cell metabolism to avoid DNA damage signalling [7,60]. During chromosomal DNA replication, due to the removal of RNA primers on the lagging strand, telomere sequences become shorter during each round of chromosomal replication. Thus, special telomere replication processes exist in eukaryotic cells to extend these chromosomal end sequences [7,60]. The process consists of a telomerase-dependent G-strand synthesis forming an ssDNA extension with the repetitive sequence TTAGG followed by the formation of dsDNA by the synthesis of a complementary sequence, the second strand or C-strand. The latter requires the initiation of Okazaki fragments, which is carried out by Pol-prim in cooperation with the CST complex (Figure 2) as an essential auxiliary remodelling factor [17,18,48,49,58]. In human cells, CST tightly associates with Pol-prim, and both protein complexes co-purify from human cell extracts [58]. During the replication of telomeric ssDNA, the CST complex binds to the single-stranded G strand, modulates telomerase activity, and recruits Pol-prim to these telomeric sequences. Additionally, CST stimulates Pol-prim to initiate Okazaki fragment synthesis via its primase activity and synthesise the C strand together with PCNA/Pol δ [58,61].
- a2. CST remodels DNA polymerase α and DNA primase to initiate Okazaki fragment synthesis
Biochemical and CryoEM studies of CST-Pol-prim complexes revealed an intricate mechanism of recruitment and activation of Pol-prim [17,18,58,62,63]. These findings suggest that in the first step, CST binds to the single-stranded telomeric G strand via its large CTC1 subunit and recruits Pol-prim via the C-terminal domain of PolA1 and the both the N- and C-terminal domains of PRI2 to telomeric ssDNA (Figure 3A, [17]). At this stage, Pol-prim is in a closed, inactive form also called APO state, similar to previously published structures of the free Pol-prim complex [17,18,64]. In this closed APO form the catalytic domain of PolA1 binds to PolA2 and PRI1 subunit (Figure 3A, regions A1 and A2, highlighted in red [17]). In this complex, the PRI2 C-terminal domain, PRI2C, additionally binds to the C-terminus of PolA1 (Figure 3A, binding site A3, highlighted in red), but not to PRI1, whereas the N-terminal PRI2N domain, D16-H255, independently binds to PolA1CT and PRI1 (Figure 3A). These three protein-protein interactions A1 to A3 may contribute to inactivity of the Apo form of Pol-prim as discussed later.
In the following, to establish primer synthesis, Pol-prim and CST form a pre-initiation complex (PIC). To this end, an extended re-arrangement of the CST/Pol-prim complex and its interaction mode with ssDNA takes place (Figure 3B, [18]). The C-terminus of the CTC1 subunit of CST forms an elongated complex on the G strand (Figure 3B, ssDNA2) and the C-terminal domain of second largest CST subunit STN1, STN1C, (P196-F368) binds to CTC1 and PolA1Pol (Pol domain consisting of Exo, aa 369-697, and catalytic domain aa 698-1220 with the catalytic aspartates, D1002 and D1004, Figure 1 and Figure 3). In contrast, Stn1N, the N-terminal domain of STN1 (E10-Q180, Figure 2), interacts with telomeric ssDNA, PRI1, and PolA1CT, the C terminal domain of PolA1 (aa 1270 to 1462), with both STN1 domains being connected via the flexible hinge region F182-N195 (Figure 2 and Figure 3B). Additionally, in PIC, the smallest CST subunit, TEN1, contacts the primase subunits PRI1, PRI2, or more precise its C-terminal domain PRI2C (I271-S506, Figure 1 and Figure 2), and the subunit PolA2. Here, the PRI1 loop, Y84 to A101 in human PRI1 (Figure 4), which binds to a pocket formed in PolA1Pol, the PolA1 DNA polymerase domain consisting of the Exo domain (aa 369 to 697) and polymerase domain (aa 698 – 1220; Figure 1), in the loading complex/Pol-prim Apo state, is located in the pocket formed by STN1N and TEN1 (compare Figure 3A, red labelled A1 region, with Figure 3B, green label A1 in PRI1). Interestingly, the PRI1 loop Y84-A101 shows low degrees of sequence conservation from mammalian to yeast PRI1, whereas the loop structure is well conserved between the PRI1 proteins in these organisms (Figure 4A and 4B, respectively). It is also important to mention that in the CST-Pol-prim loading complex, Apo state, large regions of CTC1 as a whole bind to Pol-prim via PolA1CT, PolA2, and PRI2 but neither STN1 nor TEN1 is involved in the complex formation. In contrast, in PIC, the smaller CST subunits closely engage with Pol-prim and ssDNA but only ~300 amino acids of the C-terminal end of CTC1 bind Pol-prim and the ssDNA (Figure 2, compare structure 7U5C with 8D0K, summarised in Figure 3A and 3B). In PIC, the two C-terminal OB fold domains of CTC1, OB-F and G, and STN1N bind to telomeric ssDNA in an oriented manner to recruit
In the diagram, PolA1 is shown in yellow (the PolA1 catalytic domain, PolA1cat, is depicted as a half-moon with catalytic aspartates (DTD) highlighted, is linked to the C-terminal domain PolA1CT, top right in all panels, which represents the binding site for the smaller subunits PolA2, in grey, and the large primase subunit PRI2 by a flexible hinge region). The large primase subunit, PRI2, shown in light green, consists of an N- and a C-terminal domain (PRI2N and PRI2c, respectively), whereas the catalytic RNA polymerase subunit PRI1 is presented as blue half-moon with the catalytic DID111 and D306 shown in grey (presenting that DID111 plus D306 are positioned away from the viewer). The red discs shown in panel A highlight the special interaction sites of PolA1cat with PolA2 and PRI1 in the closed Apo complex, and also mark the APO state-specific PolA1CT-PRI2C interaction, which are lost after the remodelling of Pol-prim into the open complex. To attenuate the distribution of these contact point residues in the rearranged structure they are again highlighted but in green or blue as indicated in panels B and C. Interestingly, the binding site of PolA2 in PolA1 overlaps with a PolA1 DNA binding site in the open complex (see panels B and C). In PIC, the CTC1, STN1 (both CST subunits) and PolA1Pol bind to telomeric ssDNA (with CTC1 and PolA1Pol surrounding the ssDNA and forming a tunnel). Additionally, the C-terminus of the large primase subunit, PRI2C, binds to the ssDNA by flipping from its interaction with PolA1CT in the Apo state to an association with telomeric ssDNA in PIC. PRI1, TEN1 and the N-terminal domain of STN1, both subunits of CST to form a passageway for the ssDNA template from the catalytic center of PRI1 to the catalytic site in PolA1Pol. The two domains of the large primase subunit form together with PRI1 a small tunnel that allows directing the ssDNA template towards the catalytic triple aspartates D109, D111 and D306. (Diagram assembled using the published structures 5exr, 7u5c & 8d0k, and information ([17,18,49,58] reviewed in [51]) and remodel together with TEN1 the Pol-prim complex, arranging the latter in the right orientation on the template, and providing a connection between the two catalytic polymerase centers of Pol-prim (connecting the primase catalytic site in PRI1 with the DNA polymerase domain in PolA1Pol). ssDNA stabilises the link and the proteins plus ssDNA form together a pre-initiation complex, PIC (Figure 3B; [18]). Here, the mobility of the PRI2C domain seen in these two structures is discussed by He et al. (2022) [18]. This supports and highlights the existing knowledge that the PRI2C domain is very mobile and involved in the handing over primer-ssDNA complex to PolA2Pol. It has been previously hypothesised that multiple interactions of PRI2C with PRI1 support dinucleotide synthesis, whereas its association with PolA1 and PolA2 might be important for the hand-over of the newly synthesised primers to the catalytic center of PolA1.
This extensive remodelling of the Pol-prim complex during the transition from the CST-Pol-prim APO state to PIC, is highlighted by the fact that the interactions of PolA1Pol with PRI1 and PolA2 are abolished and instead the region of PolA1Pol interacting with PolA2 in the Apo state binds at least in part to telomeric ssDNA (Figure 3A and 3B, respectively, the red spot A2 in panel A is transferred into the green triangle A2 in panel B). In the APO state, the PolA1 amino acids, H553-N555 and L645-Q649, interact with PolA2, whereas in PIC, the amino acids N552, Q554, K590, R616, Q649, R650, and N652-K655 physically bind to telomeric ssDNA in the CryoEM structure [17,18]. Here, Q554 and Q649 seemed to be crucial amino acids for both binding events in the PolA1-A2 and PolA1-ssDNA complex (compare structure 7U5C with 8D0K). The structure of Pol-prim in the Apo state [17] also suggests a possible resolution for activation of Apo state Pol-prim in the absence of remodellers such as CST since the residues N652-K655 of the PolA1-ssDNA complex are relatively freely available in solution and binding of Pol-prim to ssDNA could initiate a transition of the Apo state into PIC by ssDNA itself. However, such a hypothesised binding of Pol-prim to ssDNA would most likely be less efficient than Pol-prim/CST/ssDNA PIC including the remodelling to initiate priming on telomeric ssDNA, natural ssDNA, or polydT templates as seen in biochemical assays [58,62,63].
Additionally to its binding to PolA2 and CTC1, PolA1CT interacts with the N- and C-terminal domain of the large primase subunit PRI2, PRI2N and PRI2C, respectively, in the APO complex (Figure 3A). However, PolA1CT stops interacting with PRI2C when PIC is formed (Figure 3B and 3C). In contrast, PIC-PRI2C establishes new physical interactions with PRI1 and PolA1Pol, which did not exist in the APO enzyme state (Figure 3). Moreover, in PIC, PRI1, PRI2C and PRI2N form a tunnel, which directs ssDNA towards the primase catalytic center located in PRI1 (Figure 3B and 3C). The PRI1 loop, Y84-A101, which binds to PolA1 in the Apo state and associates with STN1N and TEN1 in PIC, is part of this tunnel formed by PRI1 and both PRI2 domains. In PIC, two tunnels surround ssDNA which are formed by PolA1Pol plus CTC1, and PRI1 plus the two PRI2 domains together with STN1N. They perform a tight grip on the telomeric G strand in PIC, and direct the 3’-end of the ssDNA to the catalyctic center of PRI1 marked by the three aspartates DID111 and D306, which bind Mg2+ essential for nucleotide binding and catalysis of primase. In the following, the coordinated binding of STN1N, PRI1, PRI2N, and PRI2C allow for the first di-nucleotide synthesis by primase. The importance of the rearrangement of PRI2 is underlined by the finding that mutations of the PRI2 linker region aa 256-270 result in a 5-fold decrease of the di-nucleotide formation whereas the ratio of di-nucleotide formation and longer primase products remains constant [15,16,64]. Interestingly, in PIC, the catalytic center of the DNA polymerase subunit PolA1 marked by the catalytic aspartates DID1002 does not contact the ssDNA (Figure 3B). However, in PIC the DTD1002 motif resides just opposite of the ssDNA bound by STN1N which forms a link and an opening between the two tunnels (Figure3B) and opens towards the catalytic center of PolA1 (marked as DTD1002) suggesting a mechanism by which PRI2C and STN1N hand over the primed G strand to the PolA1 for primer extension and DNA synthesis. Thus, during the initiation of Okazaki fragment synthesis on telomere sequences CST has recruitment and remodelling functions towards Pol-prim allowing the latter to initiate and synthesise the telomere C strand. Interestingly, not all interactions seen in PIC are equally important. Adding STEN1 alone to biochemical assays is sufficient to stimulate the initiation activity of Pol-prim [62,65]. However, physical and functional interactions of Pol-prim with CST and the remodelling of Pol-prim by CST are not only important for telomere C-strand DNA synthesis but also for genome-wide replication restart of stalled replication forks e.g., in GC-rich sequences, which depends on CST [66,67]. Additionally, CST and Pol-prim cooperate to regulate the processing of double-strand breaks (DSBs), which are a major threat to genome stability in eukaryotes [68]. Here, 53BP1 modulates 5’-end resection at DSBs via fill-in synthesis performed by Pol-prim in complex with CST avoiding long 3’-overhangs and increasing fidelity of the DSB repair pathway, since long 3’-overhangs could be repaired by the single strand annealing pathway which may cause sequence deletions [69,70,71,72]. The Pol-prim-dependent fill-in synthesis also plays an important role in the efficiency of PARP inhibitors in BRCA1-deficient cells and breast cancer treatments [69].
The CryoEM structures discussed above together with biochemical data explain a mechanism for Okazaki fragment synthesis at telomere sequences (summarised as a diagram in Figure 3) and raise the questions whether this mechanism can be extended towards the initiation process of Okazaki fragment synthesis at eukaryotic replication forks, and which proteins and protein complexes substitute for CST at replication. It is generally accepted that the heterotrimeric CST complex resembles structures and functions of heterotrimeric RPA, and CST even replaces RPA in replication fork restart after replication fork stalling to allow Pol-prim to initiate DNA synthesis [48,66,73]. Additionally, both CST and RPA interact with Pol-prim, which would support the hypothesis that they recruit Pol-prim to ssDNA for the RNA-DNA primer synthesis. In contrast to CST, however, RPA does not stimulate Pol-prim on ssDNA templates and even diminishes the enzyme activities of the latter [65,74,75,76,77]. Thus, RPA might be a part of the Pol-prim remodelling activity on lagging strands at replication forks but it alone is not sufficient to remodel Pol-prim in the Apo state to an enzyme equivalent to PIC-Pol-prim. Different activities and proteins independent of RPA or in collaboration with RPA have to substitute for CST function to remodel Pol-prim for the initiation of Okazaki fragments on the lagging strand. This view is supported by recent findings that the second largest RPA subunit RPA32 stimulates Pol-prim on ssDNA templates whereas heterotrimeric RPA inhibits Pol-prim’s activity [65]. Interestingly, RPA32 exist as a free protein or in complex with other proteins in human cells and not only in association with RPA70 and RPA14 forming the heterotrimeric RPA complex, since, after depleting HeLa cell extracts of RPA70 together with associated RPA32, still ~50% of RPA32 remained in these depleted extracts whereas no RPA70 was detected in these extracts suggesting that in mammalian cells RPA32 may have additional functions beyond the RPA complex (Rehmet & Nasheuer, unpublished results).
- a3. SV40 T antigen, RPA, and DNA polymerase α-primase collaborate during the initiation of Okazaki fragments
These studies of the lagging strand DNA synthesis initiation at telomeres give a deep insight into the mechanism of the initiation reaction of eukaryotic DNA replication and stimulate a fresh look at the initiation of Okazaki fragments at replication forks using established model systems. The SV40 DNA replication system has served as an excellent model for human DNA replication including lagging DNA synthesis for a long time [78,79,80,81,82]. These studies have advanced our knowledge about mechanisms of the initiation of DNA replication including the Okazaki fragment synthesis at replication forks e.g., by using SV40 T antigen (Tag), RPA, Pol-prim, and ssDNA as an assay system. Findings for the initiation of Okazaki fragment synthesis in the SV40 DNA replication system reveal similarities with those found for the C strand synthesis at telomere sequences (Figure 5, [37,50,52,74,75,76,83]).
In vivo and in vitro studies have revealed the multiple activities required to perform in SV40 DNA replication [80]. During the replication of dsDNA, SV40 Tag double-hexamers bind to and destabilise viral origin DNA sequences [78,82,85]. Then the two SV40 Tag helicase hexamers move in 3’ to 5’ direction on the leading strands of the replication bubble similar to CMG helicase [78,80,82,85], produce stretches of ssDNA, and load RPA onto ssDNA, which prevents hairpin formation of ssDNA and protects ssDNA against nuclease degradation [80,83]. Next, SV40 Tag recruits Pol-prim to the replication fork to initiate DNA replication in the SV40 origin on the leading strand. Although RPA stimulates DNA polymerase activity of Pol-prim and acts as a fidelity clamp of Pol α DNA polymerase activity [86,87], it is important to note that during the initiation of DNA replication eukaryotic primases cannot synthesise primers on RPA-bound ssDNA and require additional helper functions to be active [52,74,75,76]. Here, Tag stimulates the primase and DNA polymerase activity of Pol-prim similar to human AAF/CST [48,52,58,62]. Thus, it is hypothesized that following the RPA loading onto ssDNA [80,83], the RPA-Tag complex directs Pol-prim, the priming enzyme complex, to ssDNA of the lagging strand (summarised in Figure 5; [43,44,52,74,75,76,83,88]). After producing an RNA-DNA primer, Pol-prim stops DNA synthesis and RFC transfers the primer with the help of PCNA to Pol δ to synthesise the leading strand DNA [80,82]. Interestingly, monomeric but not the hexameric form of SV40 Tag plays a role in stimulating the initiation reaction of Okazaki fragments during replication of lagging strands (Figure 5; [52]). In the light of recent findings with telomere C strand DNA synthesis (Figure 3), it is hypothesised that monomeric Tag forms a complex with RPA to direct Pol-prim to the template, and then Tag or the Tag-RPA complex acts as a Pol-prim remodeller opening the Pol-prim Apo complex and forming a pre-initiation complex consisting of Tag, RPA and Pol-prim. This hypothesis suggests that the monomeric Tag-RPA complex would be the better functional equivalent for the Okazaki fragment synthesis in SV40 lagging strand DNA synthesis to the CST than RPA alone (compare Figure 3A & B with Figure 5). Additionally, one has to keep in mind that RPA has a high affinity to ssDNA, which is a barrier for Pol-prim to start primer synthesis. To this end, Tag interacts with the C-terminus of the second largest RPA, RPA32, to remodel the binding of the neighbouring DNA binding domain OB-D and thus destabilise the RPA-ssDNA complex, which has been thought to be Tag’s main auxiliary function in the Okazaki fragment synthesis [58,74,75,76]. However, at eukaryotic replication forks, two simultaneous or consecutive remodelling activities must occur during the initiation reaction. In the presence of RPA, the binding activity of RPA to ssDNA must be reduced e. g., via an interaction of Tag with RPA32 C-terminus, and Pol-prim must be transferred from its inactive APO form to PIC similar as seen at telomeric ssDNA (compare Figure 3 and Figure 5). In the SV40 system, on the lagging strand, monomeric Tag takes over this function as a re-modeller of Pol-prim and RPA.
However, in the model presented here, hexameric Tag has also important roles beyond its helicase activity at replication forks e. g., functioning as recruitment and loading factor of Pol-prim similar as previously described for Tag in RPA loading during SV40 DNA replication [83]. Here, the interaction of Tag with the N-termini of PolA1 and PolA2 are important. Interestingly, Tag binds to the same regions in these two Pol-prim subunits as AND-1/CTF4, which associates with CMG during eukaryotic DNA replication and recruits Pol-prim to the replisome progression complex in cell based replication systems, and CMG-AND-1/CTF4 complexes have similar supporting loading functions as hexameric Tag interactions in the SV40 system [22,29,33,36,80]. This interpretation is consistent structural biology findings showing that hexameric Tag binds via its ATPase domain to PolA2 N-terminus [25]. However, monomeric Tag is sufficient for the stimulation/activation activity on free and RPA-bound ssDNA suggesting that, at least under the conditions used, hexameric Tag is not required to stimulate Pol-prim [52,54]. The different requirements for the Pol-prim loading via the interaction of Tag with the N-termini of PolA1 and A2 and the remodelling during PIC formation and primer synthesis is supported by previous biochemical findings that species-specificity of initiation reactions of both, the origin-dependent initiation and Okazaki fragment synthesis of SV40 DNA replication, are regulated by the C-terminus of PolA1, residues K1149-S1462, which can in part be rescued by the presence of mouse instead of human PolA2 subunit [89]. The region contains binding sites for the small Pol-prim subunits including PolA2 and the hinge region between the PolA1Pol and PolA1CT domain, but none of the known large Tag binding sites suggesting that the interaction between PolA1 and the smaller subunits or the remodelling of the Pol-prim complex during the initiation reaction is the cause for the species specificity of SV40 DNA replication [25,35,89,90]. Thus, after loading Pol-prim onto RPA-monomeric Tag complexes on the lagging strand ssDNA, the Tag-helicase may stay associated with initiator Pol-prim since both N-termini flanking the Tag binding sites consists of IDRs [91] which are relatively flexible and adoptable to various conformations and having intrinsic flexibilities regarding distances of the interacting partners. Additionally, the hexameric Tag helicase may move on the leading strand whereas Pol-prim initiates Okazaki fragment synthesis and elongates the primer on the lagging strand. This view is supported data from multiple CryoEM experiments in which Pol-prim is not easily detectable at replication fork structures [92]. Similarly, He et al. describe multiple structures for Pol-prim in their CST-Pol-prim-ssDNA CryoEM data and have only modelled selected data sets [18].
It is important to note that monomeric Tag stimulates the initiation activity of Pol-prim on ssDNA and RPA-bound ssDNA [52,74,75,76] suggesting that monomeric Tag has Pol-prim remodelling function. These data also indicate that Pol-prim bound to free ssDNA can change from the APO structure to a PIC, and in the following forming an initiation complex (IC, summarised in [15]) but this ssDNA-driven rearrangements of Pol-prim are inefficient. Here, the binding of the PolA1Pol, PRI1, and PRI2 to ssDNA allow to opening the Pol-prim complex e.g., by ssDNA competing with PolA2 binding to PolA1, and a basic enzyme activity is determined. However, the remodelling activity of monomeric Tag stimulates these key transitions of Pol-prim to an active enzyme by a factor 6 to 8 [52,54,75,76] similar to CST on telomeric ssDNA [58,62,65]. Thus, similar to CST, Tag may disrupt the PolA1Pol interactions with PolA2 and PRI1 plus diminishes the PRI2C’s physical binding to PolA1CT allowing PRI2C to form a small tunnel together with PRI2N and PRI1 to stimulate the di-nucleotide synthesis by PRI1. PRI1 would then synthesise an oligoribonucleotide and, when the size of ~10 ribonucleotide is reached, PRI2C would hand over the primer to PolA1Pol, which extends the primer and synthesise the RNA-DNA pimer with a length of ~30 nucleotide. Then Pol-prim stops DNA synthesis and disintegrate from the template-primer system [74,76]. Next, RFC will take over the RNA-DNA primer and load PCNA onto the primed ssDNA, which recruits Pol δ to processively synthesise a complete Okazaki fragment [80,82]. In parallel, since Pol-prim is still attached to the replication fork via its link to the hexameric Tag helicase Pol-pim will move forward ~150 nucleotides towards the replication fork and start the synthesis of the next Okazaki fragment as described above.
Since it is known that RPA and Pol-prim form direct physical interactions, alternatively, RPA recruits the Pol-prim in its Apo form to the lagging but the latter cannot start RNA primer synthesis in this complex with RPA and remains inactive (Figure 5, top part). Taking recent findings for the telomeric ssDNA into account, it is hypothesized that in next step, monomeric Tag remodels this Apo complex into PIC and then IC. The binding of RPA to ssDNA would also be remodelled by Tag’s interaction with RPA32C and interfering with the OB-D ssDNA binding activity [74]. Furthermore, upon Tag’s remodelling activity, interactions within the Pol-prim complex e.g., PolA1Pol with PolA2 and PRI1 are diminished and PRI2C forms a tunnel structure with PRI1 and PRI2N to allow the synthesis of the first di-nucleotide. Finally, PRI2C would hand over the primed-lagging strand ssDNA to PolA1Pol as described above.
- a4. Elaborated activities of Okazaki fragment synthesis in yeast and human
The understanding of the replication of eukaryotic chromosomes has recently advanced with the establishment of biochemical replication systems using purified yeast and human proteins [46,53,57,92,93,94,95]. The yeast systems use origin-dependent replication whereas in human systems replication forks are established and fork-dependent DNA synthesis with the CMG complex and purified proteins is carried out. Both systems have been used to study leading strand DNA synthesis and they have also given new insights into the Okazaki fragment synthesis in eukaryotes [46,53,57,92,93,94,95]. Additionally, these purified proteins have been utilised for structural biological studies and single molecule studies [29,53,92,96]. Biochemical and structural biological experiments have shown that the eukaryotic replicative helicase, the CMG complex, plays a central role in the organisation of eukaryotic replication forks [29,46,95,97]. The biochemical assays showed the dependence and loading on the origin of yeast replication, and the leading strand synthesis with purified proteins at in vivo rates (reviewed in [13,97]) but the current review will focus on recent developments in the understanding the initiation of Okazaki fragment synthesis.
- a5. Initiation of lagging strand synthesis at replication forks by Pol-prim
The initiation process of Okazaki fragment synthesis is still not fully understood but data based on multiple replication systems suggest that the CMG complex is at the center for recruiting and organising the protein activities at replication forks [57,93,94,96,97,98]. Here, Pol-prim is linked to CMG through the AND-1/CTF4 complex, a homotrimer associated with CMG and Pol-prim [36]. As recently shown, CDC45 of CMG is important for the loading of RPA on ssDNA. Thus, the CMG complex and associated proteins resemble multiple functions first described in SV40 T antigen-dependent DNA replication [52,80,99]. In recent single molecule studies, Lewis et al. shed light on another angle on the organisation of the eukaryotic replication forks [53]. They showed that a limited number of DNA polymerases interact with replication forks and that their exchange rate and numbers depend on the concentration of the polymerase. Low, 5 nM, concentration of DNA polymerases showed a ratio of Pol ε-Pol δ-Pol α of 1-1-1 per replication fork, whereas at high concentration of ~20 nM the distribution was 1-1-2, respectively [53]. Taken into account that in vivo concentrations of these DNA polymerases in yeast culture is even above the higher concentration [100], two Pol-prim molecules per replication fork would exist in yeast cells. These findings raise the question how these multiple molecules of Pol-prim at replication forks are organised. Each subunit of AND-1/CTF4 homotrimer interacts with the IDRs of PolA1 and PolA2 N-termini (Figure 1, [20,91]. IDRs are relatively flexible and adoptable to various conformation allowing for flexibility in the distance of the interacting partners. Thus, via AND-1/CTF4 homotrimers up to three Pol-prim complexes and their catalytic functions can attach to the CMG complex moving on the leading strand. This solves the conundrum, at least in part, that Pol-prim initiates Okazaki fragment synthesis and its DNA polymerase function elongates the RNA primer on lagging strands moving away from replication forks but remaining attached to CMG moving with the fork. Here, IDRs may function like ‘rubber bands’ allowing Pol-prim to associate with ssDNA substrate to synthesize the short RNA-DNA primer for Okazaki fragment synthesis moving away from the fork before releasing from the template and moving back to the fork for a new initiation event. Here, it could be envisioned that one Pol-prim initiates the RNA primer synthesis including di-nucleotide formation, the rate limiting step, close to the replication fork, whereas the second Pol-prim, slightly away from the fork, extends the dinucleotide to short RNA and then RNA-DNA primers. This flexibility of the Pol-prim complex and its association with the replication fork is supported by the CryoEM data that Pol-prim is difficult to localise at the replication forks [92] and that multiple Pol-prim complexes associate with CST at telomeric replication sites [18]. Here, two scenarios are envisioned: first the CMG complex or associated proteins such as AND-1/CTF4 provide this Pol-prim remodelling activity and all happens at the replication fork simultaneously with loading of RPA and Pol-prim. Alternatively, the loading of RPA via the CMG complex is followed by the association of additional replication proteins, which would serve as remodelling factors for RPA decreasing its affinity to ssDNA and for Pol-prim to form an open complex capable of efficient primer synthesis. The latter opens the question which replication protein(s) serve these functions during cellular DNA replication at replication forks.
Regarding the initiation of Okazaki fragment synthesis at replication forks, a clear picture about the mechanism has not yet emerged and multiple scenarios are possible. In the first hypothesis, the CMG complex not only recruits and loads Pol-prim and RPA to the replication fork and onto the newly produced ssDNA, but also stimulates and remodels the Pol-prim complex to allow primer synthesis on the lagging strand as seen in the primer synthesis on telomeric ssDNA. Having CMG as the major player also on the lagging with its movement on leading strand requires a pleiotropic amount of steps that need to be coordinated e.g., having a major coordinated interaction between CMG, Pol-prim and RPA allowing the primer synthesis by the two primase subunits, and then support the handing over of the primed ssDNA to the catalytic center of Pol α. Here, Mcm10 and AND-1/CTF4 are involved in loading and stabilising of Pol-prim at replication forks but AND-1/CTF4 is not the remodelling cofactor stimulating Pol-prim activities since it does not enhance Pol-prim activity on unprimed M13 DNA [101]. Nevertheless, replication system lacking additional factors are able to initiation leading and lagging strand synthesis in the presence of RPA. However, they use relatively high amounts of Pol-prim and the lagging strand synthesis products in these partially reconstituted systems are far longer than in vivo products [10,11,46]. The addition of chromatin remodellers and histone chaperones e. g., FACT and Nhp6, enhanced the lagging strand DNA synthesis [93,94] and yields shorter Pol-prim primer products but they still do not reach the size of in vivo products as found in SV40-infected human cells and yeast cells [10,11]. These findings suggest that additional factors are most likely needed to supplement these replication systems.
In an alternative view, the CST complex or variations thereof e.g., the AAF sub-complex, not only carries out the remodelling of Pol-prim to initiate Okazaki fragment synthesis at telomere sequences, and restart replication synthesis after replication fork staling, but CST or sub-structures thereof also stimulate Pol-prim during the initiation of Okazaki fragment synthesis on the lagging strand at replication forks. Such a hypothesis is supported by the co-purification Pol-prim with CST from human cells, and the characterisation of Pol-prim stimulating factor AAF independently from the CST telomere function [48,58,102]. The latter is identical to a CTC1-STEN1 complex [103,104,105]. The characterisation of AAF showed that it stimulates both DNA primase and DNA polymerase activity of Pol-prim. It is important to note that mouse AAF (AAF132/AAF44 equivalent to CTC1/STN1 heterodimer) and human CST efficiently stimulate Pol-prim on poly(dT)/dT72 templates [58,62,102]. Thus, these protein complexes are not only restricted to telomere G-strand or GC-rich sequences for Pol-prim stimulation [58]. Furthermore, the yeast and human STN1 protein have been sufficient to stimulate Pol-prim functions as well as the heterotrimeric CST complex [62,65]. Here, the N-termini of human and yeast STN1 are sufficient to stimulate Pol-prim on poly(dT) and telomere ssDNA sequences. Interestingly, in the Apo complex and the PIC no interactions between STN1 and PolA2 were determined but there are several additional unpublished Pol-prim complexes described, which have not been analysed in detail, and a crucial yet undescribed rate limiting step in the stimulation may require STN1-PolA2 interactions [17,18,62,65]. Importantly, the interactions with PolA2 but not with ssDNA seem to correlate best with the stimulation of Pol-prim. Additionally, in human cells knocking down STN1/AAF44 also reduces the cellular DNA replication by ~50%. STN1/AAF44 also colocalises with PCNA in S phase cells [48]. However, genetic and cell-based assays have directed functions of Cdc13/CTC1/AAF132 and STN1/AAF44 towards telomere stability and replication-restart after replication fork stalling [7,66,106]. This apparent contradiction can be explained by the assumption that CST function is important for optimal replication but that other proteins, the CMG complex or CMG-associated proteins such as MCM10, can in part substitute for CST function at replication forks whereas CST is essential for telomere C-strand synthesis. This view is supported by the finding that AND-1/CT-4 is not essential in yeast [33]. Importantly, omitting AND-1/CTF4 minimally increases the Okazaki fragment length [46]. Furthermore, increasing the Pol-prim concentration in the assay can rescue the omission of AND-1/CTF4 in the cell-free replication [46]. Such a redundancy in DNA replication functions has been previously reported for the Pol δ takes-over of leading strand synthesis in yeast carrying a catalytically inactive gene coding for Pol ε [107].
In a third hypothesis, other factors stimulate primase during Okazaki fragment synthesis at replication forks. Such proteins include free MCM2-MCM7, GINS, MCM4,6,7 complex, and RNase H, which have been described as factors that stimulate Pol-prim activity in biochemical assays and may allow optimal Okazaki fragment synthesis [108,109,110,111,112,113]. Additionally, RPA32 and STN1 have structural and functional similarities [48,73]. This is also true for the primase stimulation of STN1/AAF44 and it was shown that RPA32 stimulates Pol-prim primer synthesis with similar capacities as STN1 whereas the full RPA complex inhibits Pol-prim [65].
4. Outlook
The central roles of RNA primer synthesis in multiple genome stability pathways such as leading and lagging strand replication, replication fork restart, and protection of DNA against degradation by exonucleases have raised the interest into the mechanism of primer synthesis by Pol-prim. The stimulation of Pol-prim and Okazaki fragment synthesis in model systems using CST, STN1, RPA, and Tag are well characterised and understood using biochemical, molecular and structural biological methods. Further studies to delineate the multiple steps to describe the process on a mechanistic level are still to come and will give new insights into central process for the prevention of genome instability. Additionally, the additional roles of CST and Pol-prim in genome stability such as replication fork restarting and DSB pathways are under intense analysis and exciting new insights are expected for the cooperation of GST and Pol-prim. On the other hand, the initiation of Okazaki fragments at cellular replication forks, an important process which is involved in the replication of ~50% of the genome due to the contribution of lagging strand DNA synthesis is partially understood and additional biochemical, genetic, molecular and structural biological approaches will contribute its understanding in the near future. Some new angles on this central process have been discussed above may contribute to solving the enigma of primase function on lagging strand synthesis at replication forks and beyond.
Acknowledgements
We thank Dr Kevin Sullivan for stimulatory discussions during writing the review. We thank our families for their continuous support and stimulation. This work was supported by a grant from the Else-Kröner-Fresenius Foundation.
References
- Hanahan, D. and R.A. Weinberg. The hallmarks of cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Bleichert, F.; Botchan, M.R.; Berger, J.M. Mechanisms for initiating cellular DNA replication. Science 2017, 355. [Google Scholar] [CrossRef]
- Burgers, P.M.; Kunkel, T.A. Eukaryotic DNA Replication Fork. Annu. Rev. Biochem. 2017, 86, 417–438. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D. and M. O'Donnell. The Eukaryotic Replication Machine. Enzymes 2016, 39, 191–229. [Google Scholar] [CrossRef]
- Blow, J.J.; Laskey, R.A. A role for the nuclear envelope in controlling DNA replication within the cell cycle. Nature 1988, 332, 546–548. [Google Scholar] [CrossRef]
- Lim, C.J.; Cech, T.R. Shaping human telomeres: from shelterin and CST complexes to telomeric chromatin organization. Nat. Rev. Mol. Cell Biol. 2021, 22, 283–298. [Google Scholar] [CrossRef]
- Costa, A.; Diffley, J.F. The Initiation of Eukaryotic DNA Replication. Annu. Rev. Biochem. 2022, 91, 107–131. [Google Scholar] [CrossRef]
- Hu, Y.; Stillman, B. Origins of DNA replication in eukaryotes. Mol. Cell 2023, 83, 352–372. [Google Scholar] [CrossRef]
- Anderson, S.; DePamphilis, M. Metabolism of Okazaki fragments during simian virus 40 DNA replication. J. Biol. Chem. 1979, 254, 11495–11504. [Google Scholar] [CrossRef]
- Smith, D.J.; Whitehouse, I. Intrinsic coupling of lagging-strand synthesis to chromatin assembly. Nature 2012, 483, 434–438. [Google Scholar] [CrossRef] [PubMed]
- Nasheuer, H.P., R. Smith, C. Bauerschmidt, F. Grosse, and K. Weisshart. Initiation of eukaryotic DNA replication: regulation and mechanisms. Prog Nucleic Acid Res Mol Biol 2002, 72, 41–94. [Google Scholar] [PubMed]
- Guilliam, T.A. Mechanisms for Maintaining Eukaryotic Replisome Progression in the Presence of DNA Damage. Front. Mol. Biosci. 2021, 8. [Google Scholar] [CrossRef] [PubMed]
- Nasheuer, H.P., H. Pospiech, and J. Syväoja, Progress towards the anatomy of the eukaryotic DNA replication fork, in Genome Integrity: Facets and Perspectives,, D.H. Lankenau, Editor. [: 2007, Springer, 2007; Genome Dynamics & Stability, Vol. 1, Berlin-Heidelberg-NewYork. p. 27 - 68. [Google Scholar] [CrossRef]
- Baranovskiy, A.G.; Tahirov, T.H. Elaborated Action of the Human Primosome. Genes 2017, 8, 62. [Google Scholar] [CrossRef]
- Baranovskiy, A.G.; Zhang, Y.; Suwa, Y.; Babayeva, N.D.; Gu, J.; Pavlov, Y.I.; Tahirov, T.H. Crystal Structure of the Human Primase. J. Biol. Chem. 2015, 290, 5635–5646. [Google Scholar] [CrossRef] [PubMed]
- Cai, S.W.; Zinder, J.C.; Svetlov, V.; Bush, M.W.; Nudler, E.; Walz, T.; de Lange, T. Cryo-EM structure of the human CST–Polα/primase complex in a recruitment state. Nat. Struct. Mol. Biol. 2022, 29, 813–819. [Google Scholar] [CrossRef]
- He, Q.; Lin, X.; Chavez, B.L.; Agrawal, S.; Lusk, B.L.; Lim, C.J. Structures of the human CST-Polα–primase complex bound to telomere templates. Nature 2022, 608, 826–832. [Google Scholar] [CrossRef] [PubMed]
- Stadlbauer, F., A. Brueckner, C. Rehfuess, C. Eckerskorn, F. Lottspeich, V. Förster, B.Y. Tseng, and H.P. Nasheuer. DNA replication in vitro by recombinant DNA-polymerase-α-primase. Eur. J. Biochem. 1994, 222, 781–793. [Google Scholar] [CrossRef]
- Núñez-Ramírez, R.; Klinge, S.; Sauguet, L.; Melero, R.; Recuero-Checa, M.A.; Kilkenny, M.; Perera, R.L.; García-Alvarez, B.; Hall, R.J.; Nogales, E.; et al. Flexible tethering of primase and DNA Pol α in the eukaryotic primosome. Nucleic Acids Res. 2011, 39, 8187–8199. [Google Scholar] [CrossRef]
- Schub, O.; Rohaly, G.; Smith, R.W.; Schneider, A.; Dehde, S.; Dornreiter, I.; Nasheuer, H.-P. Multiple Phosphorylation Sites of DNA Polymerase α-Primase Cooperate to Regulate the Initiation of DNA Replication in Vitro. J. Biol. Chem. 2001, 276, 38076–38083. [Google Scholar] [CrossRef]
- Kilkenny, M.L.; Simon, A.C.; Mainwaring, J.; Wirthensohn, D.; Holzer, S.; Pellegrini, L. The human CTF4-orthologue AND-1 interacts with DNA polymerase α/primase via its unique C-terminal HMG box. Open Biol. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Voitenleitner, C.; Rehfuess, C.; Hilmes, M.; O’rear, L.; Liao, P.-C.; Gage, D.A.; Ott, R.; Nasheuer, H.-P.; Fanning, E. Cell Cycle-Dependent Regulation of Human DNA Polymerase α-Primase Activity by Phosphorylation. Mol. Cell. Biol. 1999, 19, 646–656. [Google Scholar] [CrossRef] [PubMed]
- Voitenleitner, C.; Fanning, E.; Nasheuer, H.-P. Phosphorylation of DNA polymerase α-primase by Cyclin A-dependent kinases regulates initiation of DNA replication in vitro. Oncogene 1997, 14, 1611–1615. [Google Scholar] [CrossRef]
- Zhou, B.; Arnett, D.R.; Yu, X.; Brewster, A.; Sowd, G.A.; Xie, C.L.; Vila, S.; Gai, D.; Fanning, E.; Chen, X.S. Structural Basis for the Interaction of a Hexameric Replicative Helicase with the Regulatory Subunit of Human DNA Polymerase α-Primase. J. Biol. Chem. 2012, 287, 26854–26866. [Google Scholar] [CrossRef] [PubMed]
- Nasheuer, H.P.; Moore, A.; Wahl, A.F.; Wang, T.S. Cell cycle-dependent phosphorylation of human DNA polymerase alpha. J. Biol. Chem. 1991, 266, 7893–7903. [Google Scholar] [CrossRef]
- Schneider, A., R. W.P. Smith, A.R. Kautz, K. Weisshart, F. Grosse, and H.P. Nasheuer. Primase activity of human DNA polymerase α-primase. Divalent cations stabilize the enzyme activity of the p48 subunit. J Biol Chem. 1998, 273, 21608–21615. [Google Scholar] [CrossRef] [PubMed]
- Donnianni, R.A.; Zhou, Z.-X.; Lujan, S.A.; Al-Zain, A.; Garcia, V.; Glancy, E.; Burkholder, A.B.; Kunkel, T.A.; Symington, L.S. DNA Polymerase Delta Synthesizes Both Strands during Break-Induced Replication. Mol. Cell 2019, 76, 371–381. [Google Scholar] [CrossRef]
- Baretić, D.; Jenkyn-Bedford, M.; Aria, V.; Cannone, G.; Skehel, M.; Yeeles, J.T. Cryo-EM Structure of the Fork Protection Complex Bound to CMG at a Replication Fork. Mol. Cell 2020, 78, 926–940. [Google Scholar] [CrossRef]
- Bauerschmidt, C., S. Pollok, E. Kremmer, H.P. Nasheuer, and F. Grosse. Interactions of human Cdc45 with the Mcm2-7 complex, the GINS complex, and DNA polymerases delta and epsilon during S phase. Genes Cells 2007, 12, 745–758. [Google Scholar] [CrossRef]
- Gambus, A.; Jones, R.C.; Sanchez-Diaz, A.; Kanemaki, M.; van Deursen, F.; Edmondson, R.D.; Labib, K. GINS maintains association of Cdc45 with MCM in replisome progression complexes at eukaryotic DNA replication forks. Nature 2006, 8, 358–366. [Google Scholar] [CrossRef]
- E Georgescu, R.; Langston, L.; Yao, N.Y.; Yurieva, O.; Zhang, D.; Finkelstein, J.; Agarwal, T.; E O'Donnell, M. Mechanism of asymmetric polymerase assembly at the eukaryotic replication fork. Nat. Struct. Mol. Biol. 2014, 21, 664–670. [Google Scholar] [CrossRef] [PubMed]
- Gambus, A.; van Deursen, F.; Polychronopoulos, D.; Foltman, M.; Jones, R.C.; Edmondson, R.D.; Calzada, A.; Labib, K. A key role for Ctf4 in coupling the MCM2-7 helicase to DNA polymerase α within the eukaryotic replisome. EMBO J. 2009, 28, 2992–3004. [Google Scholar] [CrossRef]
- Broderick, R.; Nasheuer, H.-P. Regulation of Cdc45 in the cell cycle and after DNA damage. Biochem Soc Trans 2009, 37, 926–930. [Google Scholar] [CrossRef]
- Dornreiter, I.; Copeland, W.C.; Wang, T.S.-F. Initiation of Simian Virus 40 DNA Replication Requires the Interaction of a Specific Domain of Human DNA Polymerase α with large T Antigen. Mol. Cell. Biol. 1993, 13. [Google Scholar] [CrossRef]
- Villa, F.; Simon, A.C.; Bazan, M.A.O.; Kilkenny, M.L.; Wirthensohn, D.; Wightman, M.; Matak-Vinkovíc, D.; Pellegrini, L.; Labib, K. Ctf4 Is a Hub in the Eukaryotic Replisome that Links Multiple CIP-Box Proteins to the CMG Helicase. Mol. Cell 2016, 63, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Ning, B.; Feldkamp, M.D.; Cortez, D.; Chazin, W.J.; Friedman, K.L.; Fanning, E. Simian Virus Large T Antigen Interacts with the N-Terminal Domain of the 70 kD Subunit of Replication Protein A in the Same Mode as Multiple DNA Damage Response Factors. PLOS ONE 2015, 10, e0116093–e0116093. [Google Scholar] [CrossRef]
- Robertson, P.D.; Warren, E.M.; Zhang, H.; Friedman, D.B.; Lary, J.W.; Cole, J.L.; Tutter, A.V.; Walter, J.C.; Fanning, E.; Eichman, B.F. Domain Architecture and Biochemical Characterization of Vertebrate Mcm10. J. Biol. Chem. 2008, 283, 3338–3348. [Google Scholar] [CrossRef]
- Ricke, R.M.; Bielinsky, A.-K. Mcm10 Regulates the Stability and Chromatin Association of DNA Polymerase-α. Mol. Cell 2004, 16, 173–185. [Google Scholar] [CrossRef]
- Fien, K.; Cho, Y.-S.; Lee, J.-K.; Raychaudhuri, S.; Tappin, I.; Hurwitz, J. Primer Utilization by DNA Polymerase α-Primase Is Influenced by Its Interaction with Mcm10p. J. Biol. Chem. 2004, 279, 16144–16153. [Google Scholar] [CrossRef]
- Broderick, R., M. D. Rainey, C. Santocanale, and H.P. Nasheuer. Cell cycle-dependent formation of Cdc45-Claspin complexes in human cells are compromized by UV-mediated DNA damage. FEBS J 2013, 280, 4888–4902. [Google Scholar] [CrossRef]
- Broderick, S., K. Rehmet, C. Concannon, and H.P. Nasheuer. Eukaryotic single-stranded DNA binding proteins: central factors in genome stability. Subcell Biochem 2010, 50, 143–163. [Google Scholar] [PubMed]
- Chen, R.; Wold, M.S. Replication protein A: Single-stranded DNA's first responder. BioEssays 2014, 36, 1156–1161. [Google Scholar] [CrossRef] [PubMed]
- Wold, M.S. REPLICATION PROTEIN A: A Heterotrimeric, Single-Stranded DNA-Binding Protein Required for Eukaryotic DNA Metabolism. Annu. Rev. Biochem. 1997, 66, 61–92. [Google Scholar] [CrossRef] [PubMed]
- Maréchal, A.; Zou, L. RPA-coated single-stranded DNA as a platform for post-translational modifications in the DNA damage response. Cell Res. 2014, 25, 9–23. [Google Scholar] [CrossRef]
- Baris, Y.; Taylor, M.R.G.; Aria, V.; Yeeles, J.T.P. Fast and efficient DNA replication with purified human proteins. Nature 2022, 606, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.S.; Tumbale, P.P.; Arana, M.E.; Rana, J.A.; Williams, R.S.; Kunkel, T.A. High-fidelity DNA ligation enforces accurate Okazaki fragment maturation during DNA replication. Nat. Commun. 2021, 12, 1–11. [Google Scholar] [CrossRef]
- Casteel, D.E., S. Zhuang, Y. Zeng, F.W. Perrino, G.R. Boss, M. Goulian, and R.B. Pilz. A DNA Polymerase-{alpha}{middle dot}Primase Cofactor with Homology to Replication Protein A-32 Regulates DNA Replication in Mammalian Cells. J Biol Chem 2009, 284, 5807–5818. [Google Scholar] [CrossRef]
- He, Y.; Song, H.; Chan, H.; Liu, B.; Wang, Y.; Sušac, L.; Zhou, Z.H.; Feigon, J. Structure of Tetrahymena telomerase-bound CST with polymerase α-primase. Nature 2022, 608, 813–818. [Google Scholar] [CrossRef]
- Huang, H.; Weiner, B.E.; Zhang, H.; Fuller, B.E.; Gao, Y.; Wile, B.M.; Zhao, K.; Arnett, D.R.; Chazin, W.J.; Fanning, E. Structure of a DNA Polymerase α-Primase Domain That Docks on the SV40 Helicase and Activates the Viral Primosome. J. Biol. Chem. 2010, 285, 17112–17122. [Google Scholar] [CrossRef]
- Olson, C.L.; Barbour, A.T.; Wuttke, D.S. Filling in the blanks: how the C-strand catches up to the G-strand at replicating telomeres using CST. Nat. Struct. Mol. Biol. 2022, 29, 730–733. [Google Scholar] [CrossRef]
- Onwubiko, N.; Borst, A.; A Diaz, S.; Passkowski, K.; Scheffel, F.; Tessmer, I.; Nasheuer, H.P. SV40 T antigen interactions with ssDNA and replication protein A: a regulatory role of T antigen monomers in lagging strand DNA replication. Nucleic Acids Res. 2020, 48, 3657–3677. [Google Scholar] [CrossRef] [PubMed]
- Lewis, J.S.; Spenkelink, L.M.; Schauer, G.D.; Yurieva, O.; Mueller, S.H.; Natarajan, V.; Kaur, G.; Maher, C.; Kay, C.; O’donnell, M.E.; et al. Tunability of DNA Polymerase Stability during Eukaryotic DNA Replication. Mol. Cell 2019, 77, 17–25. [Google Scholar] [CrossRef]
- Onwubiko, N.O.; Scheffel, F.; Tessmer, I.; Nasheuer, H.P. SV40 T antigen helicase domain regions responsible for oligomerisation regulate Okazaki fragment synthesis initiation. FEBS Open Bio 2022, 12, 649–663. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Yuan, Z.; Georgescu, R.; Li, H.; O'Donnell, M. The eukaryotic CMG helicase pumpjack and integration into the replisome. Nucleus 2016, 7, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Vaithiyalingam, S.; Arnett, D.R.; Aggarwal, A.; Eichman, B.F.; Fanning, E.; Chazin, W.J. Insights into Eukaryotic Primer Synthesis from Structures of the p48 Subunit of Human DNA Primase. J. Mol. Biol. 2013, 426, 558–569. [Google Scholar] [CrossRef]
- Yeeles, J.T.; Janska, A.; Early, A.; Diffley, J.F. How the Eukaryotic Replisome Achieves Rapid and Efficient DNA Replication. Mol. Cell 2016, 65, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Zaug, A.J.; Goodrich, K.J.; Song, J.J.; Sullivan, A.E.; Cech, T.R. Reconstitution of a telomeric replicon organized by CST. Nature 2022, 608, 819–825. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.J.; Barbour, A.T.; Zaug, A.J.; Goodrich, K.J.; McKay, A.E.; Wuttke, D.S.; Cech, T.R. The structure of human CST reveals a decameric assembly bound to telomeric DNA. Science 2020, 368, 1081–1085. [Google Scholar] [CrossRef]
- de Lange, T. Shelterin-Mediated Telomere Protection. Annu. Rev. Genet. 2018, 52, 223–247. [Google Scholar] [CrossRef]
- Diede, S.J.; E Gottschling, D. Telomerase-Mediated Telomere Addition In Vivo Requires DNA Primase and DNA Polymerases α and δ. Cell 1999, 99, 723–733. [Google Scholar] [CrossRef]
- Lue, N.F.; Chan, J.; Wright, W.E.; Hurwitz, J. The CDC13-STN1-TEN1 complex stimulates Pol α activity by promoting RNA priming and primase-to-polymerase switch. Nat. Commun. 2014, 5, 5762–5762. [Google Scholar] [CrossRef]
- Nakaoka, H.; Nishiyama, A.; Saito, M.; Ishikawa, F. Xenopus laevis Ctc1-Stn1-Ten1 (xCST) Protein Complex Is Involved in Priming DNA Synthesis on Single-stranded DNA Template in Xenopus Egg Extract. J. Biol. Chem. 2012, 287, 619–627. [Google Scholar] [CrossRef]
- Baranovskiy, A.G.; Babayeva, N.D.; Zhang, Y.; Gu, J.; Suwa, Y.; Pavlov, Y.I.; Tahirov, T.H. Mechanism of Concerted RNA-DNA Primer Synthesis by the Human Primosome. J. Biol. Chem. 2016, 291, 10006–10020. [Google Scholar] [CrossRef] [PubMed]
- Ganduri, S.; Lue, N.F. STN1–POLA2 interaction provides a basis for primase-pol α stimulation by human STN1. Nucleic Acids Res. 2017, 45, 9455–9466. [Google Scholar] [CrossRef] [PubMed]
- Stewart, J.A.; Wang, F.; Chaiken, M.F.; Kasbek, C.; Chastain, P.D., II; Wright, W.E.; Price, C.M. Human CST promotes telomere duplex replication and general replication restart after fork stalling. EMBO J. 2012, 31, 3537–3549. [Google Scholar] [CrossRef] [PubMed]
- Kasbek, C.; Wang, F.; Price, C.M. Human TEN1 Maintains Telomere Integrity and Functions in Genome-wide Replication Restart. J. Biol. Chem. 2013, 288, 30139–30150. [Google Scholar] [CrossRef] [PubMed]
- Nieminuszczy, J.; Broderick, R.; Niedzwiedz, W. EXD2 - a new player joins the DSB resection team. Cell Cycle 2016, 15, 1519–1520. [Google Scholar] [CrossRef]
- Mirman, Z.; Cai, S.; de Lange, T. CST/Polα/primase-mediated fill-in synthesis at DSBs. Cell Cycle 2022, 22, 379–389. [Google Scholar] [CrossRef]
- Mirman, Z.; Lottersberger, F.; Takai, H.; Kibe, T.; Gong, Y.; Takai, K.; Bianchi, A.; Zimmermann, M.; Durocher, D.; de Lange, T. 53BP1–RIF1–shieldin counteracts DSB resection through CST- and Polα-dependent fill-in. Nature 2018, 560, 112–116. [Google Scholar] [CrossRef]
- Mirman, Z.; Sasi, N.K.; King, A.; Chapman, J.R.; de Lange, T. 53BP1–shieldin-dependent DSB processing in BRCA1-deficient cells requires CST–Polα–primase fill-in synthesis. Nature 2022, 24, 51–61. [Google Scholar] [CrossRef]
- Noordermeer, S.M.; Adam, S.; Setiaputra, D.; Barazas, M.; Pettitt, S.J.; Ling, A.K.; Olivieri, M.; Álvarez-Quilón, A.; Moatti, N.; Zimmermann, M.; et al. The shieldin complex mediates 53BP1-dependent DNA repair. Nature 2018, 560, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Miyake, Y.; Nakamura, M.; Nabetani, A.; Shimamura, S.; Tamura, M.; Yonehara, S.; Saito, M.; Ishikawa, F. RPA-like Mammalian Ctc1-Stn1-Ten1 Complex Binds to Single-Stranded DNA and Protects Telomeres Independently of the Pot1 Pathway. Mol. Cell 2009, 36, 193–206. [Google Scholar] [CrossRef] [PubMed]
- I Arunkumar, A.; Klimovich, V.; Jiang, X.; Ott, R.D.; Mizoue, L.; Fanning, E.; Chazin, W.J. Insights into hRPA32 C-terminal domain–mediated assembly of the simian virus 40 replisome. Nat. Struct. Mol. Biol. 2005, 12, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Weisshart, K.; Förster, H.; Kremmer, E.; Schlott, B.; Grosse, F.; Nasheuer, H.-P. Protein-Protein Interactions of the Primase Subunits p58 and p48 with Simian Virus 40 T Antigen Are Required for Efficient Primer Synthesis in a Cell-free System. J. Biol. Chem. 2000, 275, 17328–17337. [Google Scholar] [CrossRef] [PubMed]
- Melendy, T.; Stillman, B. An interaction between replication protein A and SV40 T antigen appears essential for primosome assembly during SV40 DNA replication. J. Biol. Chem. 1993, 268, 3389–3395. [Google Scholar] [CrossRef]
- Collins, K.L. and T.J. Kelly. The effects of T antigen and replication protein A on the initiation of DNA synthesis by DNA polymerase a-primase. Mol. Cell. Biol. 1991, 11, 2108–2115. [Google Scholar]
- Challberg, M.D.; Kelly, T.J. Animal Virus DNA Replication. Annu. Rev. Biochem. 1989, 58, 671–717. [Google Scholar] [CrossRef]
- Li, J.J.; Kelly, T.J. Simian virus 40 DNA replication in vitro. Proc. Natl. Acad. Sci. 1984, 81, 6973–6977. [Google Scholar] [CrossRef]
- Sowd, G.A.; Fanning, E. A Wolf in Sheep's Clothing: SV40 Co-opts Host Genome Maintenance Proteins to Replicate Viral DNA. PLOS Pathog. 2012, 8, e1002994. [Google Scholar] [CrossRef]
- Waga, S.; Bauer, G.; Stillman, B. Reconstitution of complete SV40 DNA replication with purified replication factors. J. Biol. Chem. 1994, 269, 10923–10934. [Google Scholar] [CrossRef]
- Waga, S.; Stillman, B. THE DNA REPLICATION FORK IN EUKARYOTIC CELLS. Annu. Rev. Biochem. 1998, 67, 721–751. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Klimovich, V.; I Arunkumar, A.; Hysinger, E.B.; Wang, Y.; Ott, R.D.; Guler, G.D.; Weiner, B.; Chazin, W.J.; Fanning, E. Structural mechanism of RPA loading on DNA during activation of a simple pre-replication complex. EMBO J. 2006, 25, 5516–5526. [Google Scholar] [CrossRef] [PubMed]
- Fanning, E.; Klimovich, V.; Nager, A.R. A dynamic model for replication protein A (RPA) function in DNA processing pathways. Nucleic Acids Res. 2006, 34, 4126–4137. [Google Scholar] [CrossRef] [PubMed]
- Fanning, E.; Knippers, R. STRUCTURE AND FUNCTION OF SIMIAN VIRUS 40 LARGE TUMOR ANTIGEN. Annu. Rev. Biochem. 1992, 61, 55–85. [Google Scholar] [CrossRef]
- Braun, K.A.; Lao, Y.; He, Z.; Ingles, C.J.; Wold, M.S. Role of Protein−Protein Interactions in the Function of Replication Protein A (RPA): RPA Modulates the Activity of DNA Polymerase α by Multiple Mechanisms. Biochemistry 1997, 36, 8443–8454. [Google Scholar] [CrossRef] [PubMed]
- Maga, G., I. Frouin, S. Spadari, and U. Hubscher. Replication protein A as a "fidelity clamp" for DNA polymerase alpha. J Biol Chem 2001, 276, 18235–18242. [Google Scholar] [CrossRef] [PubMed]
- An, P.; Robles, M.T.S.; Pipas, J.M. Large T Antigens of Polyomaviruses: Amazing Molecular Machines. Annu. Rev. Microbiol. 2012, 66, 213–236. [Google Scholar] [CrossRef]
- Smith, R.W.; Steffen, C.; Grosse, F.; Nasheuer, H.-P. Species Specificity of Simian Virus 40 DNA Replication in Vitro Requires Multiple Functions of Human DNA Polymerase α. J. Biol. Chem. 2002, 277, 20541–20548. [Google Scholar] [CrossRef]
- Stadlbauer, F.; Voitenleitner, C.; Brückner, A.; Fanning, E.; Nasheuer, H.-P. Species-Specific Replication of Simian Virus 40 DNA In Vitro Requires the p180 Subunit of Human DNA Polymerase α-Primase. Mol. Cell. Biol. 1996, 16, 94–104. [Google Scholar] [CrossRef]
- Mizuno, T.; Hirabayashi, K.; Miyazawa, S.; Kobayashi, Y.; Shoji, K.; Kobayashi, M.; Hanaoka, F.; Imamoto, N.; Torigoe, H. The intrinsically disordered N-terminal region of mouse DNA polymerase alpha mediates its interaction with POT1a/b at telomeres. Genes Cells 2021, 26, 360–380. [Google Scholar] [CrossRef]
- E Georgescu, R.; Schauer, G.D.; Yao, N.Y.; Langston, L.D.; Yurieva, O.; Zhang, D.; Finkelstein, J.; E O'Donnell, M. Reconstitution of a eukaryotic replisome reveals suppression mechanisms that define leading/lagging strand operation. eLife 2015, 4, e04988–e04988. [Google Scholar] [CrossRef] [PubMed]
- Devbhandari, S.; Jiang, J.; Kumar, C.; Whitehouse, I.; Remus, D. Chromatin Constrains the Initiation and Elongation of DNA Replication. Mol. Cell 2016, 65, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Kurat, C.F.; Yeeles, J.T.; Patel, H.; Early, A.; Diffley, J.F. Chromatin Controls DNA Replication Origin Selection, Lagging-Strand Synthesis, and Replication Fork Rates. Mol. Cell 2016, 65, 117–130. [Google Scholar] [CrossRef] [PubMed]
- Yeeles, J.T.P.; Deegan, T.D.; Janska, A.; Early, A.; Diffley, J.F.X. Regulated eukaryotic DNA replication origin firing with purified proteins. Nature 2015, 519, 431–435. [Google Scholar] [CrossRef]
- Jones, M.L.; Baris, Y.; Taylor, M.R.G.; Yeeles, J.T.P. Structure of a human replisome shows the organisation and interactions of a DNA replication machine. EMBO J. 2021, 40, e108819. [Google Scholar] [CrossRef]
- Guilliam, T.A.; Yeeles, J.T.P. An updated perspective on the polymerase division of labor during eukaryotic DNA replication. Crit. Rev. Biochem. Mol. Biol. 2020, 55, 469–481. [Google Scholar] [CrossRef]
- Taylor, M.R.; Yeeles, J.T. Dynamics of Replication Fork Progression Following Helicase–Polymerase Uncoupling in Eukaryotes. J. Mol. Biol. 2019, 431, 2040–2049. [Google Scholar] [CrossRef]
- Szambowska, A.; Tessmer, I.; Prus, P.; Schlott, B.; Pospiech, H.; Grosse, F. Cdc45-induced loading of human RPA onto single-stranded DNA. Nucleic Acids Res. 2017, 45, 3217–3230. [Google Scholar] [CrossRef]
- Ho, B.; Baryshnikova, A.; Brown, G.W. Unification of Protein Abundance Datasets Yields a Quantitative Saccharomyces cerevisiae Proteome. Cell Syst. 2018, 6, 192–205. [Google Scholar] [CrossRef]
- Zhu, W.; Ukomadu, C.; Jha, S.; Senga, T.; Dhar, S.K.; Wohlschlegel, J.A.; Nutt, L.K.; Kornbluth, S.; Dutta, A. Mcm10 and And-1/CTF4 recruit DNA polymerase α to chromatin for initiation of DNA replication. Minerva Anestesiol. 2007, 21, 2288–2299. [Google Scholar] [CrossRef]
- Goulian, M. and C.J. Heard. Intact DNA Polymerase a/Primase from Mouse Cells. J. Biol. Chem. 1989, 264, 19407–19415. [Google Scholar] [CrossRef] [PubMed]
- Miyake, Y.; Nakamura, M.; Nabetani, A.; Shimamura, S.; Tamura, M.; Yonehara, S.; Saito, M.; Ishikawa, F. RPA-like Mammalian Ctc1-Stn1-Ten1 Complex Binds to Single-Stranded DNA and Protects Telomeres Independently of the Pot1 Pathway. Mol. Cell 2009, 36, 193–206. [Google Scholar] [CrossRef] [PubMed]
- Surovtseva, Y.V.; Churikov, D.; Boltz, K.A.; Song, X.; Lamb, J.C.; Warrington, R.; Leehy, K.; Heacock, M.; Price, C.M.; Shippen, D.E. Conserved Telomere Maintenance Component 1 Interacts with STN1 and Maintains Chromosome Ends in Higher Eukaryotes. Mol. Cell 2009, 36, 207–218. [Google Scholar] [CrossRef] [PubMed]
- Wan, M.; Qin, J.; Songyang, Z.; Liu, D. OB Fold-containing Protein 1 (OBFC1), a Human Homolog of Yeast Stn1, Associates with TPP1 and Is Implicated in Telomere Length Regulation. J. Biol. Chem. 2009, 284, 26725–26731. [Google Scholar] [CrossRef] [PubMed]
- Chandra, A.; Hughes, T.R.; Nugent, C.I.; Lundblad, V. Cdc13 both positively and negatively regulates telomere replication. Minerva Anestesiol. 2001, 15, 404–414. [Google Scholar] [CrossRef]
- Kesti, T., K. Flick, S. Keränen, J.E. Syväoja, and C. Wittenberg. DNA polymerase e catalytic domains are dispensable for DNA replication, DNA repair, and cell viability. Mol Cell 1999, 3, 679–685. [Google Scholar] [CrossRef] [PubMed]
- A DiFrancesco, R.; Lehman, I.R. Interaction of ribonuclease H from Drosophila melanogaster embryos with DNA polymerase-primase. J. Biol. Chem. 1985, 260, 14764–14770. [Google Scholar] [CrossRef]
- Frank, P.; Braunshofer-Reiter, C.; Karwan, A.; Grimm, R.; Wintersberger, U. Purification of Saccharomyces cerevisiae RNase H(70) and identification of the corresponding gene. FEBS Lett. 1999, 450, 251–256. [Google Scholar] [CrossRef]
- Karwan, R., H. Blutsch, and U. Wintersberger. A ribonuclease H from yeast stimulates DNA polymerase in vitro. Adv Exp Med Biol 1984, 179, 513–518. [Google Scholar] [CrossRef]
- Hagemeier, A. and F. Grosse. A distinct form of ribonuclease H from calf thymus stimulates its homologous DNA-polymerase-alpha-primase complex. Eur J Biochem 1989, 185, 621–628. [Google Scholar] [CrossRef]
- De Falco, M.; Ferrari, E.; De Felice, M.; Rossi, M.; Hübscher, U.; Pisani, F.M. The human GINS complex binds to and specifically stimulates human DNA polymerase α-primase. EMBO Rep 2006, 8, 99–103. [Google Scholar] [CrossRef] [PubMed]
- You, Z.; De Falco, M.; Kamada, K.; Pisani, F.M.; Masai, H. The Mini-Chromosome Maintenance (Mcm) Complexes Interact with DNA Polymerase α-Primase and Stimulate Its Ability to Synthesize RNA Primers. PLOS ONE 2013, 8, e72408. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The four subunit DNA polymerarse α-primase complex. The diagram shows the domain structure of all four subunits of DNA polymerarse α-primase (Pol-prim, [15,16,17,18]). The largest subunit PolA1 of Pol α (large subunit p180) contains an inactive exonuclease domain (Exo), the catalytic domains of DNA polymerase (Palm, Finger and Thumb, PolA1Cat). Exo and PolA1Cat are the PolA1Pol domain. PolA1CT is the C-terminal domain of PolA1 and functions as an interaction site for the small subunits PolA2 and PRI2. The C-terminal domain and PolA1Pol are connected via a linker, which can form different structures depending on the state of the enzyme complex. The N-terminus of PolA1 is an intrinsically disordered region (IDR, [20]), which interacts with other replication factors e.g., AND-1/CTF4 and SV40 T antigen (Tag). PolA2 (B subunit, p68/p70) consists of an N-terminal protein-protein interaction and phosphorylation domain (ND) [21,23], followed by an IDR, an inactive phosphodiesterase and an OB-fold domain (oligonucleotide/oligosaccheride binding, [15,20]). The largest primase subunit PRI2 (p58, PRIL, Prim2) is composed of an N-terminal and C-terminal domain, which are connected via a flexible linker. PRI2C also contains four Fe-S clusters. PRI1 (p48/9, PRIS, PRIM1) is the catalytic active primase subunit and contains three aspartates important to bind divalent cations (D109, D111 and D306) [15].
Figure 1.
The four subunit DNA polymerarse α-primase complex. The diagram shows the domain structure of all four subunits of DNA polymerarse α-primase (Pol-prim, [15,16,17,18]). The largest subunit PolA1 of Pol α (large subunit p180) contains an inactive exonuclease domain (Exo), the catalytic domains of DNA polymerase (Palm, Finger and Thumb, PolA1Cat). Exo and PolA1Cat are the PolA1Pol domain. PolA1CT is the C-terminal domain of PolA1 and functions as an interaction site for the small subunits PolA2 and PRI2. The C-terminal domain and PolA1Pol are connected via a linker, which can form different structures depending on the state of the enzyme complex. The N-terminus of PolA1 is an intrinsically disordered region (IDR, [20]), which interacts with other replication factors e.g., AND-1/CTF4 and SV40 T antigen (Tag). PolA2 (B subunit, p68/p70) consists of an N-terminal protein-protein interaction and phosphorylation domain (ND) [21,23], followed by an IDR, an inactive phosphodiesterase and an OB-fold domain (oligonucleotide/oligosaccheride binding, [15,20]). The largest primase subunit PRI2 (p58, PRIL, Prim2) is composed of an N-terminal and C-terminal domain, which are connected via a flexible linker. PRI2C also contains four Fe-S clusters. PRI1 (p48/9, PRIS, PRIM1) is the catalytic active primase subunit and contains three aspartates important to bind divalent cations (D109, D111 and D306) [15].
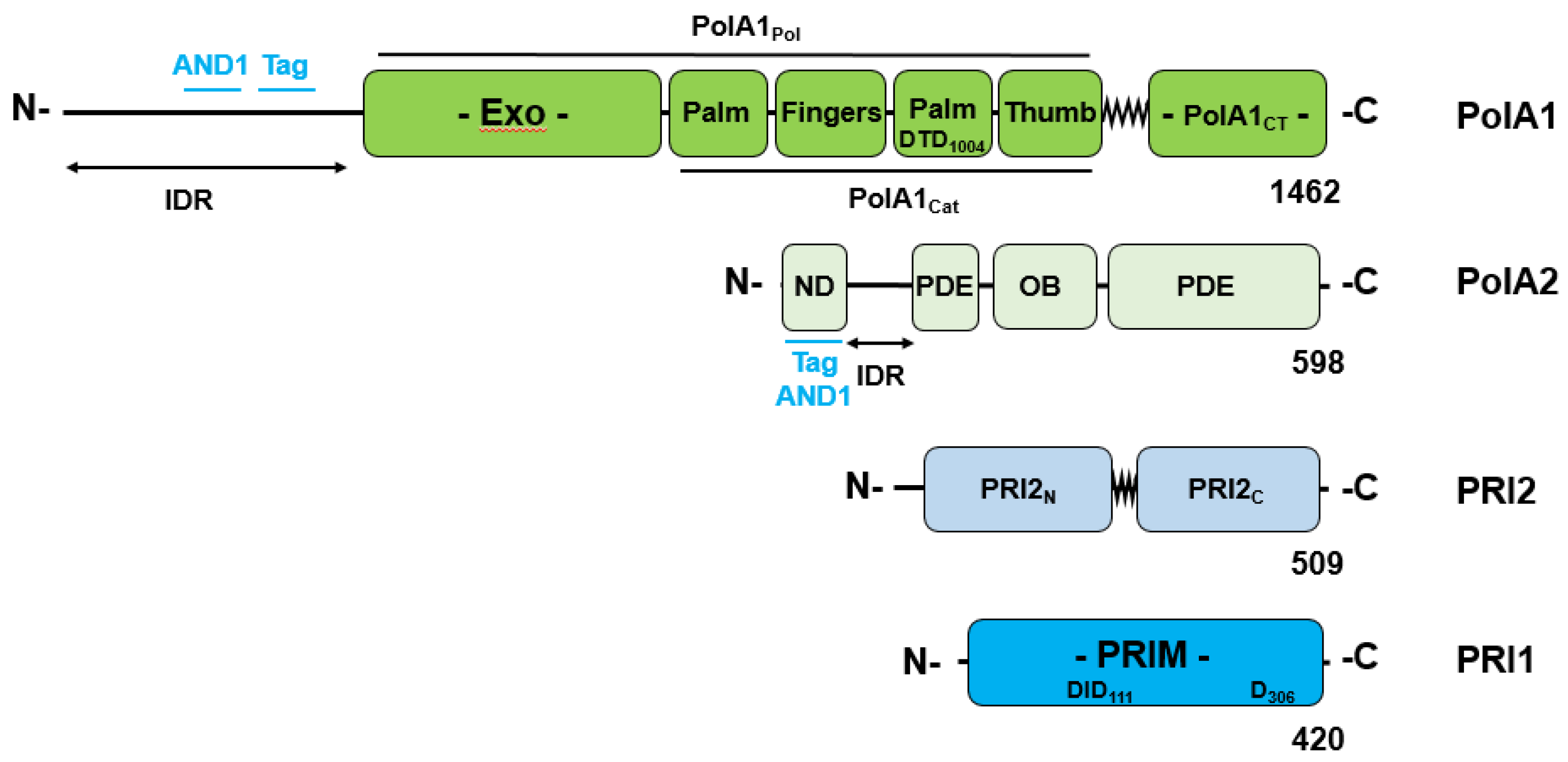
Figure 2.
The domain structure of the heterotrimeric CST complex. The largest subunit of the CST complex, CTC1 also known as CDC13 and AAF132 consists of seven OB-fold domains and a hinge region [48,59]. The three helix bundle (THB) domain forms a hinge region between the three N-terminal OB-fold (OB-A to OB-C) and the four C-terminal OB-fold domains (OB-D to OB-G) with the two OB-fold domains OB-F and G (CTC1CT) being responsible for CST ssDNA binding and protein interactions within the Pre-Initiation complex (PIC). The second largest CST subunit, STN1 (AAF44), consists of an N-terminal domain (E10 to Q180) mainly formed by an OB-fold domain (aa 58 – 152) and a C-terminal domain (P196-F368) connected via a flexible linker. The C-terminal domain of STN1 contains two winged Helix-turn-helix domains, wHTH1 and wWTH2. The smallest subunit TEN1/TEN1L comprises an OB-fold domain [59].
Figure 2.
The domain structure of the heterotrimeric CST complex. The largest subunit of the CST complex, CTC1 also known as CDC13 and AAF132 consists of seven OB-fold domains and a hinge region [48,59]. The three helix bundle (THB) domain forms a hinge region between the three N-terminal OB-fold (OB-A to OB-C) and the four C-terminal OB-fold domains (OB-D to OB-G) with the two OB-fold domains OB-F and G (CTC1CT) being responsible for CST ssDNA binding and protein interactions within the Pre-Initiation complex (PIC). The second largest CST subunit, STN1 (AAF44), consists of an N-terminal domain (E10 to Q180) mainly formed by an OB-fold domain (aa 58 – 152) and a C-terminal domain (P196-F368) connected via a flexible linker. The C-terminal domain of STN1 contains two winged Helix-turn-helix domains, wHTH1 and wWTH2. The smallest subunit TEN1/TEN1L comprises an OB-fold domain [59].
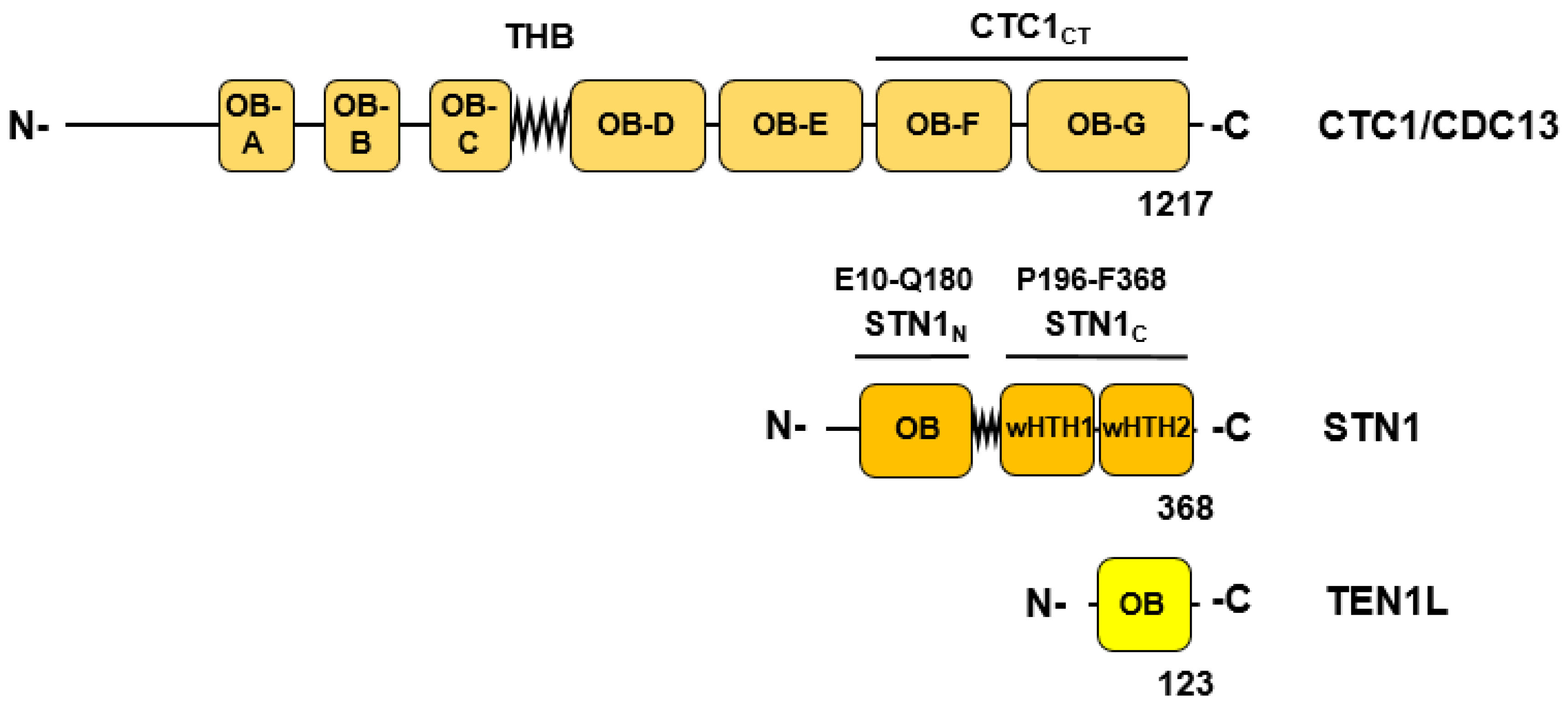
Figure 3.
Rearrangements of the Pol-prim complex during activation of a closed APO form to an initiation-active enzyme. The diagram schematically summarises the remodelling of the Pol-primase complex architecture from a compact closed shape (inactive APO-state, panel A [17]) to an open form, the pre-initiation complex (PIC-state, panel B and C, [18]) ready to synthesise RNA on ssDNA to initiate DNA synthesis. The panels A and B show the Pol-prim complex associated with CST complex. To simplify the presentation and for clarity reasons the CST complex (panel B) was omitted in panel C. The PIC Pol-prim complex is shown in the presence of telomeric ssDNA (the latter is include in the Pol-rim complex, panel C). It is important to note that especially the PolA1cat and PRI2C domains of the Pol-prim complex show a high degree of movement in CryoEM studies [18]. The depicted structure is one of several states of Pol-prim in the active form suggesting that multiple structures are associated with primase activity. Panel A only presents the STN1 N-terminus whereas the C terminus was not included in the structure 7u5c.
Figure 3.
Rearrangements of the Pol-prim complex during activation of a closed APO form to an initiation-active enzyme. The diagram schematically summarises the remodelling of the Pol-primase complex architecture from a compact closed shape (inactive APO-state, panel A [17]) to an open form, the pre-initiation complex (PIC-state, panel B and C, [18]) ready to synthesise RNA on ssDNA to initiate DNA synthesis. The panels A and B show the Pol-prim complex associated with CST complex. To simplify the presentation and for clarity reasons the CST complex (panel B) was omitted in panel C. The PIC Pol-prim complex is shown in the presence of telomeric ssDNA (the latter is include in the Pol-rim complex, panel C). It is important to note that especially the PolA1cat and PRI2C domains of the Pol-prim complex show a high degree of movement in CryoEM studies [18]. The depicted structure is one of several states of Pol-prim in the active form suggesting that multiple structures are associated with primase activity. Panel A only presents the STN1 N-terminus whereas the C terminus was not included in the structure 7u5c.
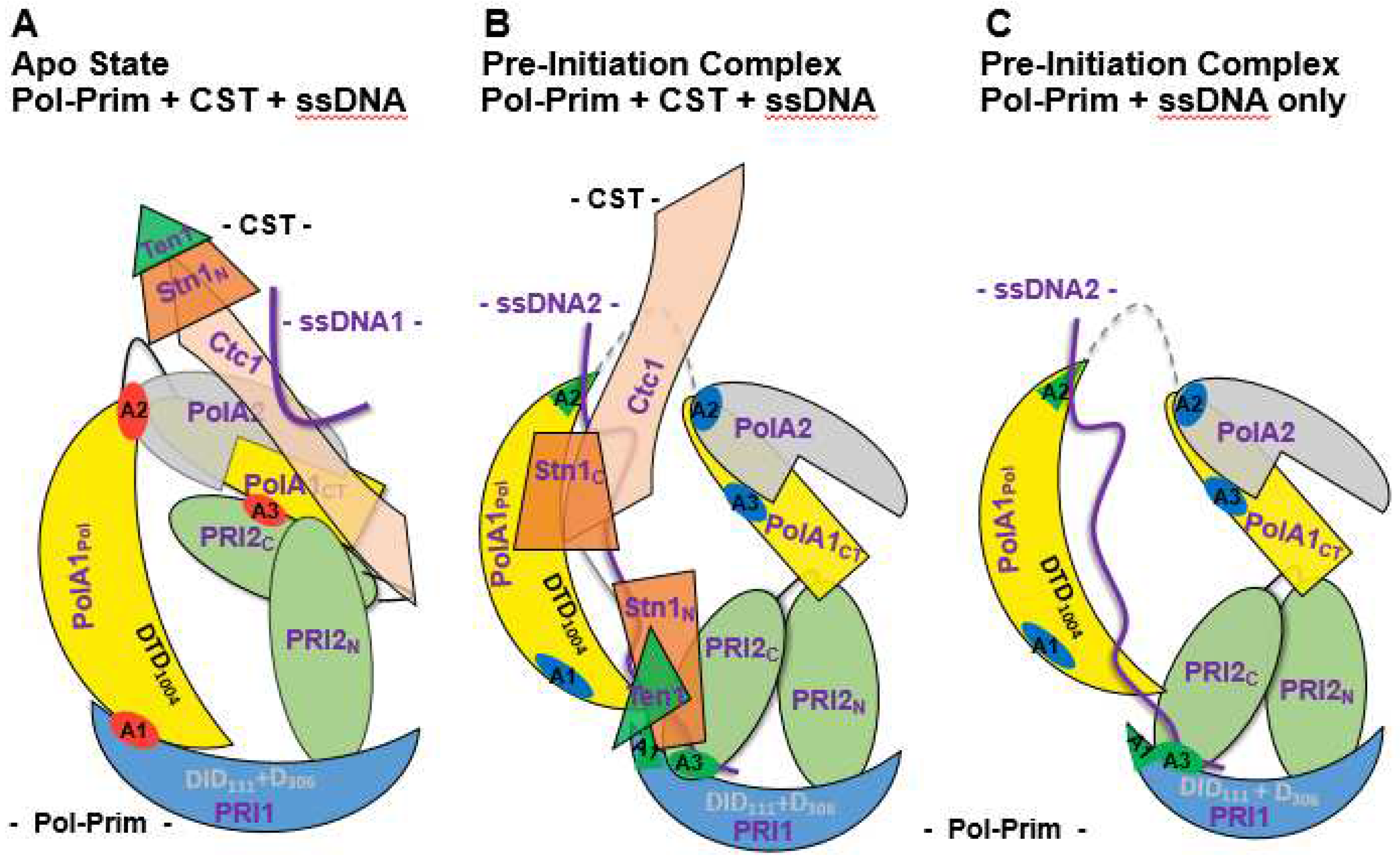
Figure 4.
Conservation of the PRI1 loop binding to the large Pol α subunit PolA1. In the APO form the catalytic primase subunit PRI binds to the large subunit PolA1 with sequences human and mouse Y84-A101, yeast Y86-A103, and Drosophila melanogastor H103–V120 (panel A), which form a loop (panel B). These residues are not well conserved from human to yeast on the sequences site (panel A shows a Clustal W alignment) but the 3 dimensional structure is well conserved (panel B, AlphaFold predicted structures were aligned using PyMol). The arrows indicate the start and end residues of the PRI1 loop. Human PRI1 is in green, mouse PRI1 in light blue, yeast PRI1 in yellow, and Drosophila melanogaster PRI1-pink (Alpha Fold structures).
Figure 4.
Conservation of the PRI1 loop binding to the large Pol α subunit PolA1. In the APO form the catalytic primase subunit PRI binds to the large subunit PolA1 with sequences human and mouse Y84-A101, yeast Y86-A103, and Drosophila melanogastor H103–V120 (panel A), which form a loop (panel B). These residues are not well conserved from human to yeast on the sequences site (panel A shows a Clustal W alignment) but the 3 dimensional structure is well conserved (panel B, AlphaFold predicted structures were aligned using PyMol). The arrows indicate the start and end residues of the PRI1 loop. Human PRI1 is in green, mouse PRI1 in light blue, yeast PRI1 in yellow, and Drosophila melanogaster PRI1-pink (Alpha Fold structures).
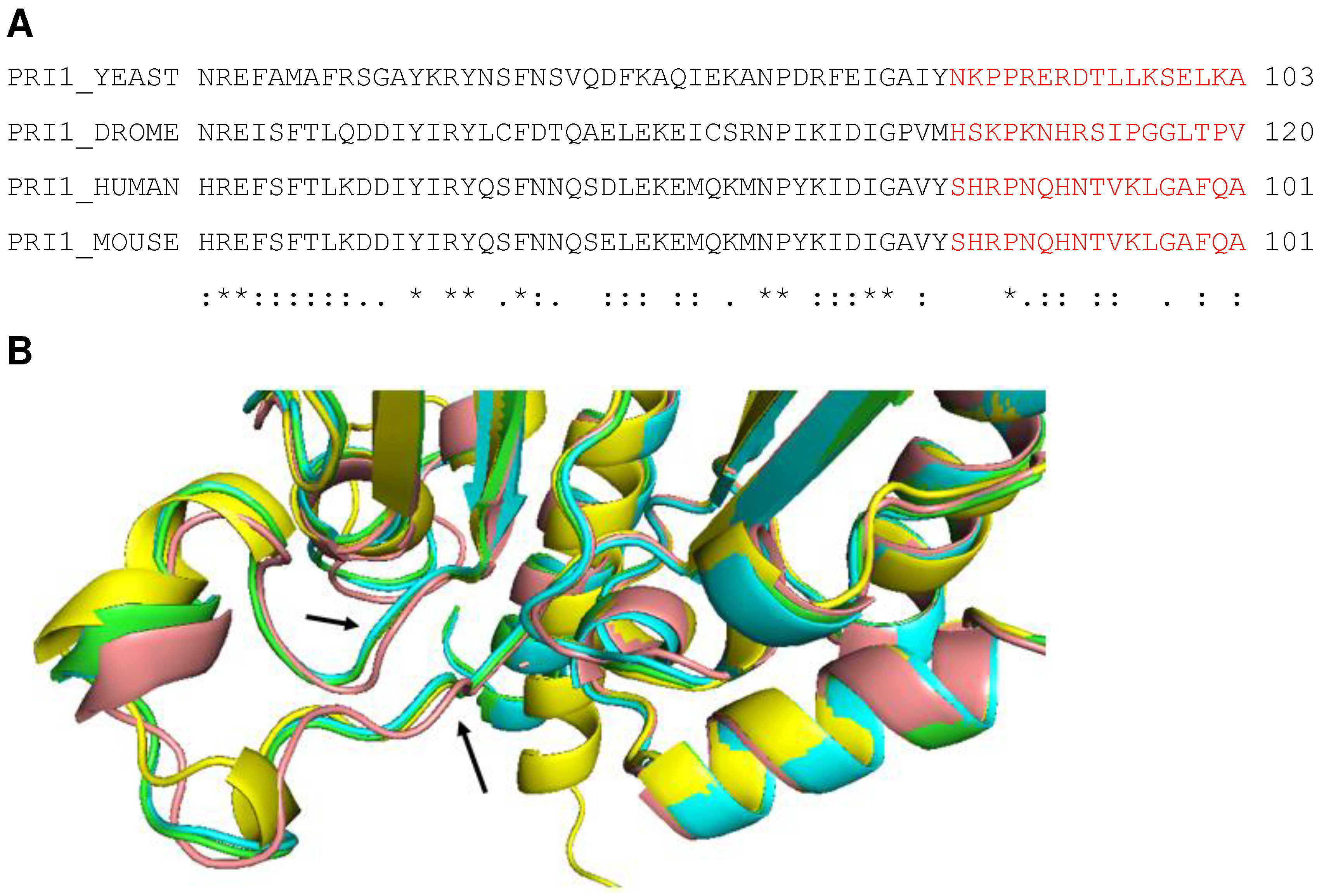
Figure 5.
Functional interactions of SV40 T antigen, RPA and Pol-prim during Okazaki fragment initiation on lagging strands of SV40 DNA replication. DNA primase does not start primer synthesis on RPA-bound ssDNA and requires the remodelling factor Tag to diminish RPA-binding to ssDNA and enabling Pol-prim to synthesise RNA and DNA on ssDNA. The components of the SV40 lagging strand system are: ssDNA (purple line); RPA (consisting of the large subunit RPA70 with the OB-fold domains A, B, C, and F shown in brown; middle subunit RPA32 containing N-terminal phosphorylation sites, the central OB-fold D and the protein interaction region RPA32C coloured in beige, plus RPA14 subunit, OB-fold domain E, in orange [43,84]); the four subunit complex Pol-Prim (DNA polymerase subunit PolA1 in dark blue consisting of N-terminal Tag interaction region (PolA1N) and the catalytic DNA polymerase domain (PolA1Pol), and the C-terminal domain (PolA1CT) having multiple protein interaction sites; the regulatory subunit PolA2 in blue, consisting of the PolA2 N-terminal region (PolA2N) highlighted with an N that interacts with SV40 Tag and the remaining PolA2 residues necessary for DNA replication; and in light blue the two primase subunits, PRI1 containing the catalytic site, and the large primase subunit PRI2 that interacts with the large subunit PolA1 [14,15,25]), and the remodelling factor Tag (green, showing its three domains, the N-terminal DNA-J domain, the Origin-Binding Domain (OBD), and the helicase domain with V350 and P417 necessary for the stimulation shown as V and P, and highlighted in red-brown).
Figure 5.
Functional interactions of SV40 T antigen, RPA and Pol-prim during Okazaki fragment initiation on lagging strands of SV40 DNA replication. DNA primase does not start primer synthesis on RPA-bound ssDNA and requires the remodelling factor Tag to diminish RPA-binding to ssDNA and enabling Pol-prim to synthesise RNA and DNA on ssDNA. The components of the SV40 lagging strand system are: ssDNA (purple line); RPA (consisting of the large subunit RPA70 with the OB-fold domains A, B, C, and F shown in brown; middle subunit RPA32 containing N-terminal phosphorylation sites, the central OB-fold D and the protein interaction region RPA32C coloured in beige, plus RPA14 subunit, OB-fold domain E, in orange [43,84]); the four subunit complex Pol-Prim (DNA polymerase subunit PolA1 in dark blue consisting of N-terminal Tag interaction region (PolA1N) and the catalytic DNA polymerase domain (PolA1Pol), and the C-terminal domain (PolA1CT) having multiple protein interaction sites; the regulatory subunit PolA2 in blue, consisting of the PolA2 N-terminal region (PolA2N) highlighted with an N that interacts with SV40 Tag and the remaining PolA2 residues necessary for DNA replication; and in light blue the two primase subunits, PRI1 containing the catalytic site, and the large primase subunit PRI2 that interacts with the large subunit PolA1 [14,15,25]), and the remodelling factor Tag (green, showing its three domains, the N-terminal DNA-J domain, the Origin-Binding Domain (OBD), and the helicase domain with V350 and P417 necessary for the stimulation shown as V and P, and highlighted in red-brown).
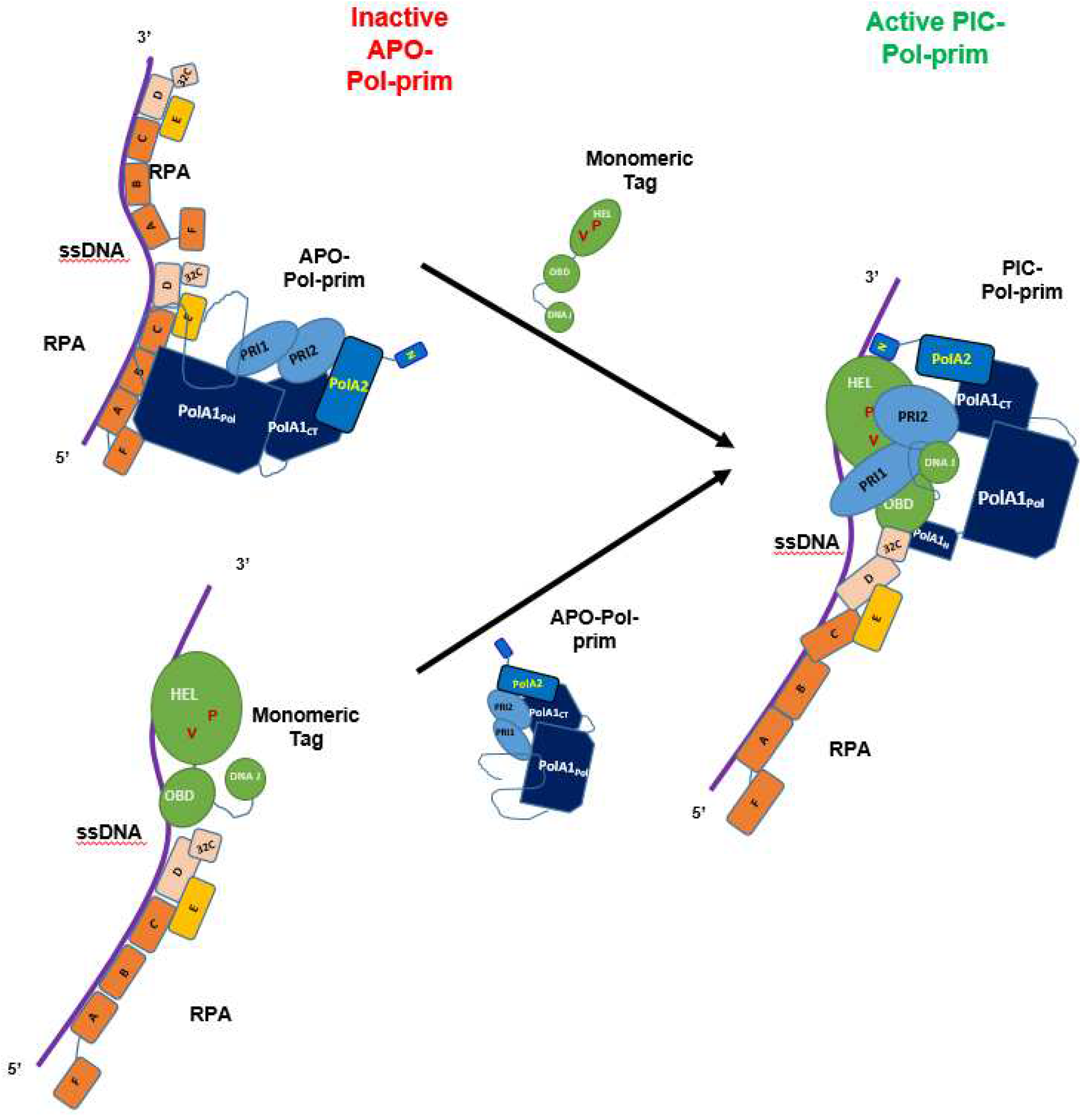
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



