Submitted:
17 March 2023
Posted:
20 March 2023
You are already at the latest version
Abstract
The risk of the emergence and reemergence of zoonoses is high in regions that are under anthropogenic actions, as they contribute to the risk of vector disease transmission. Yellow fever (YF) is among the main pathogenic arboviral disease in the world, and the Culicidae Aedes albopictus has been proposed to have the potential to transmit yellow fever virus (YFV). This mosquito inhabits both urban and wild environments, and under experimental conditions, it has been shown to be susceptible to infection by YFV. In this study, the vector competence of the mosquito Ae. albopictus for the YFV was investigated. Female Ae. albopictus were exposed to non-human primates (NHP) of the genus Callithrix infected with YFV for blood meal. Subsequently, on the 14th and 21st days post infection, the legs, heads, thorax/abdomen and saliva of the arthropods were collected and analyzed by viral isolation and molecular analysis techniques to verify the infection, dissemination and transmission. The presence of YFV was detected in saliva samples through viral isolation and in the head, thorax/abdomen and legs both by viral isolation and by molecular detection. The susceptibility of Ae. Albopictus to YFV confers a potential risk of reemergence of urban YF in Brazil.
Keywords:
infecção experimental
; Aedes albopictus
; febre amarela
; reemergência
; RT‒qPCR
; isolamento viral
1. Introduction
Yellow fever (YF) is among the main arboviral disease in the world and is endemic in tropical regions of Africa and in several countries of South America, including Brazil, affecting public health care in these countries [1,2,3]. It is an infectious, acute febrile disease caused by an arbovirus of the genus Flavivirus of the family Flaviviridae, i.e., Yellow fever virus (YFV), which is transmitted by the bite of mosquitoes of the genus Aedes, especially Aedes aegypti, in urban areas and the genera Haemagogus and Sabethes in wild areas in the Americas. It is estimated the occurrence of 200,000 cases of YF and 30,000 deaths each year worldwide; notably, 20% to 50% of those infected who develop severe disease die [4,5].
Environmental changes strongly affect vector ecology and epidemiology of diseases. In tropical forest regions that are under the influence of anthropogenic actions, the risk of emergence and reemergence of zoonoses is high, considering that changes such as urbanization increase the risk of transmission of etiological agents by vectors. The dispersal of anthropophilic mosquitoes may be facilitated by changes in the ecosystem that affect the interactions between vectors and hosts and favor the action of vectors in the transmission of pathogens between wild and urban cycles [6,7,8].
Vector-borne diseases account for more than 17% of all infectious diseases worldwide, causing more than 1 million deaths annually. Viruses transmitted by arthropods belong to the group of arboviruses (arthropod-borne viruses), which are related to human pathogens, most of which are zoonotic. One of the vectors that has been widely investigated in the transmission of arboviruses is the Aedes (Stegomyia) albopictus [9], a mosquito native from the forests of Southeast Asia and found in Brazil for the first time in 1986, subsequently expanding to urban areas in recent years [6,10,11,12,13,14].
Ae. albopictus has been proposed as a potential transmission vector of YFV, in addition to other viruses, because it has a wide geographical distribution and ecological plasticity, a characteristic that allows procreation both in urban and wild environments, allowing this mosquito to become a bridge vector between urban environments and rural areas [6,8,15]. Although it has not been implicated as a vector of arboviruses in Brazil, in Asia, Ae. albopictus acts as a vector of Dengue virus (DENV) and Japanese encephalitis virus (JEV) in rural and urban areas; in the Americas, it was found to be naturally infected with Venezuelan equine encephalitis virus (VEEV) and West Nile virus (WNV) and, under laboratory conditions, was found to be competent for at least 22 arboviruses, including DENV, Zika virus (ZIKV), Chikungunya virus (CHIKV) and YFV [2,16,17,18,19,20]. In studies conducted in Brazil, Ae. albopictus has also been found to be naturally infected with ZIKV [21,22] and DENV [22,23,24].
From 2000 to 2015, 339 human cases of sylvatic yellow fever (SYF) and 161 (47.4%) deaths were confirmed in Brazil. From December 2016 to May 2017, the country experienced the largest outbreak of SYF in recent decades, with 642 cases in non-human primates (NHPs) and 778 cases in humans, with 262 (34.59%) deaths reported in the states of the Midwest, Southeast, and North regions [1,25,26].
From 2018 to March 2019, 50 cases of YF were confirmed in the states of São Paulo and Paraná, 24% of which resulted in deaths. From 2019 to June 2022, there were 33 cases in the states of Pará, Tocantins, Acre and Santa Catarina, 12% of which resulted in deaths. This scenario has prioritized the organization of public health services, prevention activities, disease control and vector control and highlights the need to better understand the factors related to the dispersion of YFV [3,27].
The reemergence of YFV in endemic countries may be favored by a combination of several factors, such as the low demand and supply of the YF vaccine, the susceptibility of exposed human populations, the high density of vectors and primary hosts (i.e., NHPs), favorable climatic conditions, the emergence of a new genetic lineage and the circulation of infected people and NHPs [28,29].
Elucidating knowledge about vector competence and the transmission of mosquito-borne infections improves the possibilities of preventing and responding to emerging threats from arboviruses as well as developing new warning systems and, consequently, vector control. The aim of this study was to evaluate, using NHPs of the genus Callithrix as an experimental model, the vector competence of Ae. albopictus and its potential role as a link between the sylvatic and urban cycles in a possible urban expansion of YFV in Brazil.
2. Materials and Methods
2.1. Mosquitoes
To conduct this study, populations of mosquitoes of the species Ae. albopictus were obtained from the municipality of Ananindeua, Pará, Brazil. The mosquitoes were kept until the fifth generation in an insectary of the Laboratory of Medical Entomology, Section of Arbovirology and Hemorrhagic Fevers, Evandro Chagas Institute (SAARB/IEC), under controlled temperature and humidity conditions (26±1 °C; 80-90 % RH) and a 12-h light-dark cycle. The larvae were fed fish chow, and adult mosquitoes were fed a 10% glucose solution.
For the experiment, the mosquitoes were deprived of food for 24 hours before the blood meal. A total of 200 females were used, 5 to 7 days after emerging from the pupal stage, of which 50 were assigned to the control group (kept until the end of the experimental period).
2.2. NHPs
Ten NHPs of the genus Callithrix sp. bred and maintained at the National Primate Center (CENP- body linked to the Secretary of Health and Environment Surveillance – SVS - of the Ministry of Health, which operates in the areas of conservation, reproduction and research with non-human primates) Ananindeua, Pará, Brazil were used in this study. The animals were transported to the Animal Biosafety Level 3 (NBA3) Laboratory of the SAARB/IEC, strictly following biosafety, containment and sanitary control recommendations [31,32,33] and were housed in 10 individual stainless steel cages with retractable bottoms (0.80 cm x 0.80 cm x 0.90 cm). During the entire experiment, the nutritional protocol recommended by the CENP was adopted for animal maintenance. All NHPs were previously tested by serological methods and were negative for flavivirus. One animal was maintained as a negative control until the end of the experimental period.
2.3. Virus
The isolate BeAN838691, belonging to South American genotype I, obtained from the serum of a natural YFV infection in Callithrix penicillata, provided by SAARB/IEC, was used. The viral stock was prepared using Ae. albopictus (clone C6/36) [34]. Confirmation of the presence of YFV in the referred cells was performed using the indirect immunofluorescence assay [35], and the viral load was obtained using RT-qPCR.
2.4. Infection Procedure
Three NHPs were inoculated by intramuscular route with a single dose of YFV suspension (1 x 105 PFU/mL) following the method described by Ferreira et al. [36]. On the 3rd day post-infection (dpi), the animals were sedated with tiletamine-zolazepam (8 mg.kg-1) combined with chlorpromazine (0.5 mg.kg-1) [37]. Next, each NHP was exposed to 50 female Ae. albopictus mosquitoes (totaling 150 mosquitoes), which were retained in an adapted feeder that was placed on the abdominal region for a period of 30 minutes for blood feeding. After feeding, the engorged females were transferred to individual cages and maintained with 10% glucose up to 14 dpi, when they were placed in contact with 3 healthy and sedated NHPs (NHP4, NHP5 and NHP6) for blood feeding, following the same conditions previously described for the first feeding. Subsequently, the females were distributed into individual cages, properly identified and maintained with 10% glucose until 21 dpi. At this time, 3 healthy and sedated NHPs (NHP7, NHP8, and NHP9) were exposed to female Ae. albopictus for blood feeding following the same protocol described above (Figure 1).
The 3 NHPs inoculated with YFV viral suspension and the 6 NHPs exposed to mosquitoes were euthanized on the 7th day after exposure to the virus using an overdose of intracardiac thiopental sodium (1 g) [38].
2.5. Collection of Mosquito Samples
Female Ae. albopictus had their saliva extracted and were segmented (head, thorax, and abdomen). For saliva extraction, the proboscis of each female was inserted into a 10-μL tip containing 5 μL of fetal bovine serum (FBS). The sample was pipetted into an a 1,5 mL microtube containing 45 μL of Leibowitz L-15 medium and used for cell culture [17,39]. Segmentation was performed individually using sterile plates, microscope slides, entomological forceps and a light stereomicroscope 14 and 21 dpi. The abdomen and thorax were examined to confirm infection, the head and legs were examined to confirm dissemination, and saliva was collected to verify the infection and eventual transmission of YFV.
The samples were organized into batches of 10 specimens, placed in properly labeled tubes and ground in a Tissue Lyser II (Qiagen, Hilden, Germany) for 60 seconds at a frequency of 25 Hz in a 2 mL microtube (U-bottom) containing 1 mL of 1x D-PBS extender (Gibco) with 2% penicillin and streptomycin, 1% fungizone and 5% FBS and a tungsten bead (3 mm). After grinding, the tubes were placed in a -80 °C freezer for 12 hours.
2.6. Cell Culture, Inoculation and Indirect Immunofluorescence (IIF)
Following the protocol described by Igarashi [34], Aedes aegypti clone C6/36 cells were maintained at 28 °C using Leibowitz L-15 medium with L-glutamine (Gibco, USA) supplemented with 5% FBS (Gibco) (phosphate), tryptose phosphate (Himedia, Kennet Square,PA, USA ) (2.95%), antibiotics (penicillin 10,000 U/L and streptomycin 10,000 µg/L) (Gibco) and nonessential amino acids (10 mL/L) (Baktron Microbiology, Rio de Janeiro, Brazil ) [40]. Before inoculation, the growth medium (5% FBS) was discarded from the tubes containing the cells, followed by the addition of 100 µL of the arthropod macerate solution. Sample adsorption was conducted for 60 minutes at 28 °C, with gently shaking every 15 minutes.
After the sample was allowed to adsorb, 1.5 mL of L-15 maintenance medium was added to each tube containing the inoculated cells. The tubes were incubated again at 28 °C and observed daily for 7 days under an inverted optical microscope to visualize a possible cytopathogenic effect (CPE) [41,42]. On the 7th day after inoculation, IIF was performed following the protocol described by Gubler et al. [35].
For the IIF, the cells inoculated with the macerates were placed on slides and fixed in acetone PA (-20 °C) for 10 minutes. Subsequently, 10 μL of polyclonal antibodies against YFV provided by FIOCRUZ – Rio de Janeiro, Brazil, and diluted 1:500 in phosphate saline solution (PBS, pH 7.4) was added to each of the slides. Next, the slides were incubated for 30 minutes in an oven at 37 °C, followed by a PBS wash for 10 minutes. The slides were then allowed to dry at room temperature. Then, 10 μL of anti-mouse conjugated antibody (Cappel) diluted 1:900 in PBS pH 7.4 was added to the slides, and the incubation and washing steps previously described were repeated. The slides were then mounted (coverslip and a small drop of buffered glycerin, pH 7.4) and observed under an epifluorescence microscope.
2.7. Quantification of the YFV Genome in Mosquitoes
2.7.1. RNA Extraction
Total RNA was extracted from 140 μl of the macerate obtained during the collection stage using a QIAamp viral RNA® kit (Qiagen, Hilden, Germany) following the manufacturer’s protocol. Bacteriophage MS2 RNA (Roche Diagnostics, Basel, Switzerland) was used as a noncompetitive exogenous internal control. For this purpose, 2 ng of MS2 RNA was added to each mosquito pool during extraction [43]. The extracted RNA was stored at -80 °C until use.
2.7.2. Preparation of a YFV Standard Curve
To construct a YFV standard curve, a low-passage YFV isolate (strain BeAr843721, GenBank accession number MF370530) cultivated in Vero cells was used. Viral RNA was extracted from the cell culture supernatant using a QIAamp viral RNA® kit (Qiagen) following the manufacturer’s instructions. Viral RNA was then quantified by RT-ddPCR using the One-step RT-ddPCR Advanced Kit for Probe (Bio-Rad, Hercules, CA, USA) and a set of primers and probe specific for YFV [44]. RT-ddPCR mixtures contained 11 nmol of each primer, 4.4 nmol of probe and 5 μL of extracted viral RNA in a total volume of 22 μL. Next, the reaction mixtures were transferred to the wells of the DG8 droplet generator cartridge (Bio-Rad), to which 70 μL of droplet generator oil (Bio-Rad) was added. The reactions were emulsified in a QX-200 droplet generator (Bio-Rad). Then, 40 μL of emulsion/reaction was transferred to a 96-well reaction plate (ddPCR Plates 96-Well, Semi-Skirted, Bio-Rad). The plates were sealed in a PX1™ PCR Plate Sealer (Bio-Rad) and subjected to amplification in a C1000 Touch™ Thermal Cycler (Bio-Rad).
The cycling conditions were as follows: 95 °C for 10 min, followed by 40 cycles of denaturation at 95 °C for 30 seconds and elongation at 60 °C for 1 minute. At the end of the amplification reaction, the PCR products were read using a Quanta Soft Droplet Reader (Bio-Rad). The absolute concentration of viral RNA, in copies/µL, was determined automatically using ddPCR QuantaSoft Software V.1.7.4.0917 (Bio-Rad). After determining the absolute quantification of viral RNA in copies of the YFV genome per microliter, the RNA was used as a standard to construction a standard curve to use in the quantification of RNA extracted from mosquito pools. The extracted viral RNA was serially diluted 1:10, ranging from 3.3x104 to 3.3x100 copies/µL. The standards were stored at -80 °C until use.
2.7.3. YFV RT-qPCR
Quantification of the YFV genome from total RNA extracted from mosquito pools was performed using RT-qPCR [44] together with the standard curve prepared in the previous step. The reaction was performed using the Superscript III® Platinum® One-Step Quantitative RT-PCR System kit (Invitrogen) in a total volume of 25 μL, containing 12.5 nM of each primer (YfallF-GCT AAT TGA GGT GYA TTG GTC TGC and YfallR- CTG CTA ATC GCT CAA MGA ACG CAC), 5 nM of probe (YfallP-ATC GAG TTG CTA GGC AAT AAA, labeled with FAM and BHQ1) and 5 μL of the extracted total mosquito RNA. The runs were performed in a 7500 real-time PCR system (Applied Biosystems, Waltham, MA, USA). The cycling conditions were as follows: 50 °C for 30 minutes, followed by a 2-minute cycle at 95 °C and then 45 cycles of 15 seconds at 95 °C and 1 minute at 60 °C. Samples with CT values below 37 were considered positive. The YFV genome concentration in the samples was determined in RNA copies/µL using a standard curve in Applied Biosystems 7500 Software version 2.3. The values were then converted into copies/mg of mosquito using the following formula:
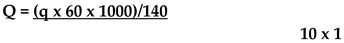 where
where
Q = quantification in copies/mg
q = quantification in copies/µl
60 = RNA elution volume
1000 = volume of PBS used in the maceration
140 = volume of macerate used in the extraction
10 = number of mosquitoes per pool
1 = mean mass of 1 mosquito in mg
2.8. Ethical Aspects
This study was approved by the directors of the CENP (concession of animals), the Committee on Ethics in the Use of Animals of the IEC (CEUA/IEC - protocol n° 0030/2019) and the Biodiversity Authorization and Information System of Instituto Chico Mendes de Biodiversity Conservation (SISBIO/ICMBio - protocol no. 61757-1).
3. Results
Ae. albopictus were infected with YFV after feeding on Callithrix experimentally infected with YFV. Engorged female Ae. albopictus were analyzed at 14 and 21 dpi. The presence of viral replication was detected by viral isolation in C6/36 cells from samples of saliva, head, thorax/abdomen and legs of the mosquitoes, and these results were confirmed by simultaneous molecular detection. Thus, the laboratory transmission of YFV by these mosquitoes to NHPs of the genus Callithrix was confirmed.
All arthropods were subjected to viral isolation. First-passage C6/36 cells were used for the detection of infection in Ae. albopictus saliva samples. Saliva samples from 60 mosquitoes were analyzed and organized into batches of 10 specimens, with 3 batches from 14 dpi (30 mosquitoes) and 3 from 21 dpi (30 mosquitoes). Of the saliva samples from 14 dpi, 66.7% (2/3) were positive for the presence of YFV. For the saliva samples from 21 dpi, 33.3% (1/3) were positive for YFV (Figure 2).
Infection is a prerequisite for the spread of the virus, and in the present study, the presence of YFV was also analyzed in batches of the head, thorax/abdomen and legs, with 10 specimens in each sample. A total of 21 lots were analyzed, 9 at 14 dpi and 12 at 21 dpi. Three negative control samples (referring to the same body parts tested) were tested for each analyzed group (Table 1). One hundred percent of the samples from the head, thorax/abdomen and legs were positive for YFV at 14 dpi. For the samples, 58% (7/12) were positive for YFV at 21 dpi (Figure 3).
For the molecular analysis, 70 female Ae. albopictus were used to make 21 head, thorax/abdomen and leg batch sample, with 9 samples (30 specimens) for 14 dpi and 12 samples (40 specimens) for 21 dpi. The RT-qPCR results indicated that all samples from 14 dpi were positive, with Ct values ranging from 23.5 to 17.6, and that for the samples from 21 dpi, 58% (7/12) were positive and presented Ct values between 34.2 and 15.4. Absolute quantification of the YFV genome was also performed for all samples (Table 1).
Blood samples from the 10 NHPs were analyzed by RT-qPCR. NHP1, NHP2 and NHP3 (infected by inoculation) were positive for YFV; NHP4, NHP5 and NHP6 were negative (blood meal on 14 dpi); and of the animals exposed to Ae. Albopictus on 21 dpi (i.e., NHP7, NHP8 and NHP9), only 1 animal (NHP9) was positive, with a CT of 8.4 (Table 2).
4. Discussion
Evaluations of the vector competence of Ae. albopictus in the transmission of YFV to NHPs of the genus Callithrix, to the best of our knowledge, have not yet been performed, makes the data obtained in the present study relevant, especially considering that risk assessments are necessary in the context of the identification of potential vectors for the virus and for the urbanization of YF in Brazil. Furthermore, the presence of YFV confirmed in one of the primates reinforces the relevance of the present study.
Urbanization increases larval habitats of Ae. albopictus and accelerates the development and survival of mosquitoes. In the last 20 years, the area inhabited by this culicid has increased significantly, which, in turn, has potentially increased the ability of the vector to transmit diseases. Reurbanization of YF has been much discussed, especially in terms of the true susceptibility and ability of Ae. albopictus to become infected and act as a link vector between the sylvatic cycle and the urban environment [28,45,46,47].
Since the reemergence of YF in 2014/2015, Brazil has recorded its largest YF epidemic in recent decades, affecting mainly the Southeast region. Abreu et al. [48] demonstrated the risk of reurbanization and the seasonal reemergence of YFV in a study conducted in the state of Rio de Janeiro, where they found a female of the genus Alouatta infected with YFV and Ae. albopictus in the environment by probing the animal’s body, thus emphasizing the need for continuous and effective surveillance, in addition to high vaccination coverage.
The transmission of arboviruses from a vector to a vertebrate host usually occurs through the saliva expelled by the mosquito during blood feeding [49]. In the present study, YFV was found in the saliva of Ae. albopictus in 100% of the samples analyzed from 14 dpi and 21 dpi. In studies conducted by Amraoui et al. [2], YFV was detected in the saliva of Ae. albopictus after 4 passages in cells; the authors suggest that the culicid represents a threat in the context of the reurbanization of YF.
The presence of YFV in the saliva of Ae. albopictus indicates the potential of this Culicidae, adapted to the periurban environment, to transmit YF in urban areas in countries where it is present. Additionally, there should be concern about the transmission of other arboviruses by this vector, such as those that cause dengue, Zika and Chikungunya, in the urban environment [20].
By viral isolation and molecular detection, YFV was found in different parts of the body of Ae. albopictus, both at 14 and 21 dpi, indicating the dispersion and infection of the mosquitoes, in addition to suggesting 14 dpi as the best period for understanding the kinetics of vector infection. Notably, the legs of Culicidae are fundamental in the spread of the virus in arthropod organisms. Our results indicated that the virus replicates in all parts of the mosquito’s body, including the legs.
Although this Culicidae has not been indicated as a YF vector in Brazil, in a study carried out by researchers from the Evandro Chagas Institute (IEC/PA) in 2017, YFV was found in samples of Ae. albopictus from the southeastern region of the country; however, more studies of natural infection need to be developed. Experimentally, this vector has already been infected with YFV [50,51].
It is necessary to combat this mosquito through a better understanding of its biology to allow the implementation of new effective control strategies. A metagenomic analysis of Ae. albopictus, for example, may allow comparative analyses and even the discovery of new genes and elements that may be useful for innovative genetic control strategies for this mosquito, which is a potential vector of important viruses that can affect public health [52,53].
The genus Callithrix was used as an experimental model due to its periurban and urban habits as well as ease of handling. In this study, one of the primates (NHP9) was positive by molecular analysis for YFV, a finding that suggests the transmission of YFV by mosquitoes via blood feeding 21 dpi. Although YFV was detected both by viral isolation and by molecular analysis in all samples from 14 dpi, only on 21 dpi was there transmission of YFV to NHP9. However, it is possible that the age of the mosquitoes at 14 dpi is related to virus transmission processes. In addition, there was a lower viral load at 14 dpi than at 21 dpi, suggesting that despite the positive results, on 14 dpi, the viral load was insufficient for transmission.
The transmission of arboviruses by mosquitoes depends on the genetic characteristics of the vectors and the virus, as well as on several factors such as incubation temperature, viral load, and stress conditions. More studies and investigations need to be performed to elucidate the roles of these factors.
The detection of epizootic diseases in NHPs by surveillance services is essential for surveillance activities and for the diagnosis and detection of cases in humans, enabling better epidemic control [47].
In recent years, entomovirological surveillance has proven to be a useful tool for the detection of medically important viruses in the event of outbreaks, the monitoring of virus circulation and characterizing vectors, which are fundamental elements for understanding the dynamics of vector-borne diseases. Vector competence studies in different mosquito species are important. Data on vectors and viruses associated with human diagnoses are essential to establish disease control strategies [2,7,17].
The risk of new YF outbreaks is high in the region of the Americas, especially considering the impacts of the COVID-19 pandemic, which led to a decrease in the proportion of the population vaccinated against YF; at national levels, YF vaccination coverage was 57% in 2021 [3].
Entomological data, continuous epidemiological surveillance and analyses of landscape composition can contribute to the prevention of susceptible areas to possible outbreaks of YF, allowing protective measures to be taken to avoid cases in humans [54].
There is a need for continuous effective surveillance and for an increase in vaccination coverage, as vaccination is the most effective preventive measure available for YF. Entomological surveillance actions should be considered for the control of Ae. albopictus, especially in urban areas. Control efforts should therefore be strengthened and maintained to prevent the urbanization of YF and new outbreaks of the disease.
5. Conclusions
The detection of YFV in the saliva, head, thorax/abdomen and legs confirms that Aedes albopictus can become infected through blood feeding on NHPs, suggesting the competence of this Culicidae as a potential vector for YFV. The positive results for the presence of YFV in NHPs of the genus Callithrix after exposure to Aedes albopictus indicates that this mosquito serves as a transmission vector of YFV, reinforcing the relevance of the present study and the need for conducting research, especially with a view to conducting natural and continuous entomological surveillance.
Author Contributions
Conceptualization, R.D.C., C.F.A., J.P.N.N. and A.C.R.C; methodology, R.D.C., M.N.O.F., M.S.F., P.H.G.C., C.F.A., P.A.S.A., D.D.D and R.C.F.B.; software, B.T.D.N.; validation, A.C.R.C, J.P.N.N, L.C.M and E.V.P.S; formal analysis, A.C.R.C and E.V.P.S.; investigation, R.D.C, C.F.A, A.C.R.C. and J.P.N.N.; resources, J.P.N.N., A.C.R.C, E.V.P.S., L.C.M. and P.F.C.V.; writing—original draft preparation: R.D.C. and A.C.R.C.; writing—review and editing, R.D.C, A.C.R.C, E.V.P.S., B.T.D.N.,C.F.A., J.P.N.N. and P.F.C.V.; funding acquisition, A.C.R.C., J.P.N.N., L.C.M. and P.F.C.V. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Instituto Evandro Chagas.
Institutional Review Board Statement
The animal study protocol was approved by the Committee for Ethics in the Use of Animals (CEUA) of the Evandro Chagas Institute (protocol code 30/2019 and approval date June 3, 2019).
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
To Dr. Ricardo Lourenço de Oliveira from the Oswaldo Cruz Foundation (IOC) for his important contribution to the realization of the project. To the PPGBAIP program (UFPA). To the employees and students of the Medical Entomology Laboratory of the Arbovirology and Hemorrhagic Fevers Section of the Evandro Chagas Institute (SAARB/IEC/SVS/MS) for their support in the creation and maintenance of the Culicidae colony.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ministério da Saúde. Plano de Contingência Para Resposta às Emergências em Saúde Pública: Febre Amarela; Ministério da Saúde: Brasil, 2016; pp. 7–8. [Google Scholar]
- Amraoui, F.; Pain, A.; Piorkowski, G.; Vazeille, M.; Couto-Lima, D.; de Lamballerie, X.; Lourenço-de-Oliveira, R.; Failloux, A.B. Experimental adaptation of the yellow fever virus to the mosquito Aedes albopictus and potential risk of urban epidemics in Brazil, South America. Sci. Rep. 2018, 8, 14337. [Google Scholar] [CrossRef] [PubMed]
- Organização Pan-Americana da Saúde; Organização Mundial da Saúde. Alerta Epidemiológico: Febre Amarela; OPAS/OMS: Washington, DC, 2022. [Google Scholar]
- Monath, T.P.; Vasconcelos, P.F.C. Yellow fever. J. Clin. Virol. 2015, 64, 160–173. [Google Scholar] [CrossRef] [PubMed]
- CDC-Febre Amarela. Saúde global-febre amarela. Available online: https://www.cdc.gov/globalhealth/newsroom/topics/yellowfever/index.html (accessed on 2 February 2022).
- Vasconcelos, P.F.C.; Azevedo, R.S.S.; Rodrigues, S.G.; Martins, L.C.; Jannifer, O.C.; Travassos-da-Rosa, A.P.A. Arboviroses. In Medicina Tropical e Infectologia na Amazônia; Leão, R.N.Q., Ed.; Editora Samaúma: Belém, 2013; pp. 481–503. [Google Scholar]
- Dos Santos, T.P.; Roiz, D.; de Abreu, F.V.S.; Luz, S.L.B.; Santalucia, M.; Jiolle, D.; Neves, M.S.A.S.; Simard, F.; Lourenço-de-Oliveira, R.; Paupy, C. Potential of Aedes albopictus as a bridge vector for enzootic pathogens at the urban-forest interface in Brazil. Emerg. Microbes Infect. 2018, 7, 191. [Google Scholar] [CrossRef] [PubMed]
- Lwande, O.W.; Obanda, V.; Lindstrom, A.; Ahlm, C.; Evander, M.; Naslund, J.; Bucht, G. Globe-Trotting Aedes aegypti and Aedes albopictus: Risk Factors for Arbovirus Pandemics. Vector Borne Zoonotic Dis. 2020, 20, 71–81. [Google Scholar] [CrossRef]
- Forattini, O.P. Identificação de aedes (stegomyia) albopictus (skuse) no Brasil. Rev. Saúde Pública 1986, 20, 244–245. [Google Scholar] [CrossRef] [PubMed]
- Forattini, O.P. Identificação de aedes (stegomyia) albopictus (skuse) no Brasil. Rev. Saúde Pública 1986, 20, 244–245. [Google Scholar] [CrossRef] [PubMed]
- Lopes, N.; Nozawa, C.; Linhares, R.E.C. Características gerais e epidemiologia dos arbovírus emergentes no Brasil. Rev. Pan-Amaz. Saúde 2014, 5, 55–64. [Google Scholar] [CrossRef]
- Vasconcelos, P.F.C.; Calisher, C.H. Emergence of Human Arboviral Diseases in the Americas, 2000–2016. Vector Borne Zoonotic Dis. 2016, 0, 1–7. [Google Scholar] [CrossRef]
- Santos, N.S.O.; Romanos, M.T.V.; Wigg, M.D. Virologia humana, 3ª ed.; Guanabara Koogan: Rio de janeiro, Brazil, 2015; pp. 1065–1084. [Google Scholar]
- Ferreira-de-Lima, V.H.; Câmara, D.C.P.; Honório, N.A.; Lima-Camara, T.N. The Asian tiger mosquito in Brazil: Observations on biology and ecological interactions since its first detection in 1986. Acta Trop. 2020, 205, 105386. [Google Scholar] [CrossRef]
- Trabulsi; Alterthum. Microbiologia, 6ª ed.; Editora Atheneu: São Paulo, Brazil, 2015; p. 912. [Google Scholar]
- Moore, C.G.; Mitchell, C.J. Aedes albopictus in the United States: Ten-year presence and public health implications. Emerg. Infect. Dis. 1997, 3, 329–334. [Google Scholar] [CrossRef]
- Castro, M.G.D.; Nogueira, R.M.R.; Schatzmayr, H.G.; Miagostovich, M.P.; Lourenço-de-Oliveira, R. Dengue virus detection by using reverse transcription-polymerase chain reaction in saliva and progeny of experimentally infected Aedes albopictus from Brazil. Mem. Inst. Oswaldo Cruz 2004, 99, 809–814. [Google Scholar] [CrossRef] [PubMed]
- Paupy, C.; Delatte, H.; Bagny, L.; Corbel, V.; Fontenille, D. Aedes albopictus, an arbovirus vector: From the darkness to the light. Microbes Infect. 2009, 11, 1177–1185. [Google Scholar] [CrossRef] [PubMed]
- Couto-Lima, D.; Madec, Y.; Bersot, M.I.; Campos, S.S.; Motta, M.D.A.; Santos, F.B.D.; Vazeille, M.; Vasconcelos, P.F.C.; Lourenço-de-Oliveira, R.; Failloux, A.B. Potential risk of re-emergence of urban transmission of Yellow Fever virus in Brazil facilitated by competent Aedes populations. Sci. Rep. 2017, 7, 4848. [Google Scholar] [CrossRef] [PubMed]
- Little, E.A.H.; Hutchinson, M.L.; Price, K.J.; Marini, A.; Shepard, J.J.; Molaei, G. Spatiotemporal distribution, abundance,.
- and host interactions of two invasive vectors of arboviruses, Aedes albopictus and Aedes japonicus, in Pennsylvania, USA. Parasit. vectors 2022, 15,36. [CrossRef]
- Smartt, C.T.; Stenn, T.M.S.; Chen, T.Y.; Teixeira, M.G.; Queiroz, E.P.; Dos Santos, L.S.; Queiroz, G.A.N.; Souza, K.R.; Silva, L.K.; Shin, D.; et al. Evidence of Zika Virus RNA Fragments in Aedes albopictus (Diptera: Culicidae) field-collected eggs from Camaçari, Bahia, Brazil. J. Med. Entomol. 2017, 54, 1085–1087. [Google Scholar] [CrossRef]
- Rezende, H.R.; Romano, C.M.; Claro, I.M.; Caleiro, G.S.; Sabino, E.C.; Felix, A.C.; Bissoli, J.; Hill, S.; Faria, N.R.; Da Silva, T.C.C.; et al. First report of Aedes albopictus infected by Dengue and Zika virus in a rural outbreak in Brazil. PloS One 2020, 15, e0229847. [Google Scholar] [CrossRef] [PubMed]
- de Figueiredo, M.L.G.; de C Gomes, A.; Amarilla, A.A.; Leandro, A.D.S.; Orrico, A.D.S.; de Araujo, R.F.; Castro, J.D.S.M.; Durigon, E.L.; Aquino, V.H.; Figueiredo, L.T.M. Mosquitoes infected with dengue viruses in Brazil. Virol. J. 2010, 7, 152. [Google Scholar] [CrossRef] [PubMed]
- Martins, V.E.P.; Alencar, C.H.; Kamimura, M.T.; de Carvalho Araújo, F.M.; De Simone, S.G.; Dutra, R.F.; Guedes, M.I.F. Occurrence of natural vertical transmission of dengue-2 and dengue-3 viruses in Aedes aegypti and Aedes albopictus in Fortaleza, Ceará, Brazil. PloS One 2012, 7, e41386. [Google Scholar] [CrossRef]
- Ministério da Saúde. Boletim Epidemiológico; Ministério da Saúde: Brasil, 2015; pp. 1–8. [Google Scholar]
- Ministério da Saúde. Centro de Operações de Emergências em Saúde Pública Sobre Febre Amarela, Informe 43/17; Ministério da Saúde: Brasil, 2017; pp. 1–5. [Google Scholar]
- Pan American Health Organization (PAHO); World Health Organization (WHO). Epidemiological Update: Yellow Fever-6 March 2019; PAHO/WHO: Washington, D.C, 2020. [Google Scholar]
- Vasconcelos, P.F.C. Yellow fever: reflections on the disease, prospects for the century and risk of re-urbanization. Rev. Bras. Epidemiol. 2002, 5, 244–258. [Google Scholar] [CrossRef]
- Vasconcelos, P.F.C. Yellow fever in Brazil: thoughts and hypotheses on the emergence in previously free areas. Rev. Saúde pública 2010, 44, 1144–1149. [Google Scholar] [CrossRef]
- Instituto Evandro Chagas. Instituto Evandro Chagas detecta vírus da febre amarela em mosquito Aedes albopictus no Brasil. Available online: https://www.iec.gov.br/descoberta/ (accessed on 14 November 2020).
- Müller, C.; Andrade, M.; Gonçalves, M.; Calzavara, N.; Santos, P.; Monteiro, R. Biossegurança. In Biologia, Manejo e Medicina de Primatas Não Humanos na Pesquisa Biomédica; Fiocruz: Rio de Janeiro, 2010; pp. 385–433. [Google Scholar]
- Washington, D.C. Guide for the Care and Use of Laboratory Animals. 8ª ed; The national academies press. United States of America. 2011.
- Ministério da Saúde. Biocontenção: O Gerenciamento do Risco em Ambientes de Alta Contenção Biológica NB3 e NBA3; Ministério da Saúde: Brasil, 2015; pp. 88–94. [Google Scholar]
- Igarashi, A. Isolation of a singh’s Aedes albopictus cell clone sensitive to dengue and chikungunya viruses. J. Gen. Virol. 1978, 40, 531–544. [Google Scholar] [CrossRef]
- Gubler, D.J.; Velez, M.; Kuno, G.; Oliver, A.; Sather, G.E. Mosquito cell cultures and specific monoclonal antibodies in surveillance for dengue viruses. Am. J. Trop. Med. Hyg. 1984, 33, 158–165. [Google Scholar] [CrossRef]
- Ferreira, M.S.; de Castro, P.H.G.; Silva, G.A.; Casseb, S.M.M.; Júnior, A.G.D.; Rodrigues, S.G.; Azevedo, R.D.S.D.S.; Silva, M.F.C.E.; Zauli, D.A.G.; Araújo, M.S.S.; et al. Callithrix penicillata: A feasible experimental model for dengue virus infection. Immunol. Lett. 2014, 158, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Soffiati, F. L. Comparação entre dois protocolos anestésicos para contenção química de sagüis híbridos Callithrix sp. Dissertação de mestrado. Universidade Estadual norte Fluminense. Rio de Janeiro. 2016.
- Fasamo, D.M. Anestesia e Controle da Dor. Biologia e Manejo e Medicina de Primatas não Humanos na Pesquisa Biomédica; Fiocruz: Rio de Janeiro, 2010. [Google Scholar]
- Nuñez, A.I.; Talavera, S.; Birnberg, L.; Rivas, R.; Pujol, N.; Verdún, M.; Aranda, C.; Berdugo, M.; Busquets, N. Evidence of Zika virus horizontal and vertical transmission in Aedes albopictus from Spain but not infectious virus in saliva of the progeny. Emerg. microbes & infect. 2020, 9, 2237–2244. [Google Scholar] [CrossRef]
- Barbosa, M.L.; Rocco, I.M.; Felippe, J.M.; Cruz, A.S. Growth and maintenance of Aedes albopictus cell line, clone C6/36, in different media. Rev. Inst. Adolfo Lutz 1993, 531, 63–70. [Google Scholar] [CrossRef]
- Tesh, R.B. A method for the isolation and identification of dengue viruses, using mosquito cell cultures Am. J. Trop. Med. Hyg. 1979, 28, 1053–1059. [Google Scholar] [CrossRef] [PubMed]
- Beaty, B.J.; Calisher, C.H.; Shope, R.E. Arboviruses. In Diagnostic Procedures for Viral Rickettsial and Chlamydial Infections, 7th ed.; Lennette, E.H., Lunette, D.A., Lenett, E.T., Eds.; American Public Health Association: Washington, DC, 1995; pp. 189–212. [Google Scholar]
- Menting, S.; Thai, K.T.D.; Nga, T.T.T.; Phuong, H.L.; Klatser, P.; Wolthers, K.C.; Binh, T.Q.; de Vries, P.J.; Beld, M. Internally controlled, generic real-time PCR for quantification and multiplex real-time PCR with serotype-specific probes for serotyping of dengue virus infections. Adv. Virol. 2011, 2011, 514681. [Google Scholar] [CrossRef] [PubMed]
- Domingo, C.; Patel, P.; Yillah, J.; Weidmann, M.; Méndez, J.A.; Nakouné, E.R.; Niedrig, M. Advanced yellow fever virus genome detection in point-of-care facilities and reference laboratories. J. Clin. Microbiol. 2012, 50, 4054–4060. [Google Scholar] [CrossRef]
- Li, Y.; Kamara, F.; Zhou, G.; Puthiyakunnon, S.; Li, C.; Liu, Y.; Zhou, Y.; Yao, L.; Yan, G.; Chen, X.G. Urbanization increases Aedes albopictus larval habitats and accelerates mosquito development and survivorship. Plos Negl. Trop. Dis. 2014, 8, e3301. [Google Scholar] [CrossRef]
- de Miranda, R.M.; Fernandes, R.S.; da Silva-Fernandes, A.T.; Ferreira-de-Brito, A.; Moreira, S.B.; Pereira, R.C.; da Silva Mendes, Y.; de Lima, S.M.B.; Pissinatti, A.; Freire, M.D.S.; et al. Neotropical sylvatic mosquitoes and aedes aegypti are not competent to transmit 17DD attenuated yellow fever virus from vaccinated viremic new world non-human primates. Viruses 2022, 14, 2231. [Google Scholar] [CrossRef]
- Gava, C.; Silva, T.C.C.D.; Lyra, D.G.P.; Ardisson, K.S.; Marques, C.S.; Almada, G.L.; Corrêa, L.M.C.; Siqueira, P.C.; Rodrigues, G.A.P.; Moura, L.D.; et al. Prevenção e controle da febre amarela: avaliação de ações de vigilância em área indene no Brasil. Cad. Saúde Pública 2022, 38. [Google Scholar] [CrossRef]
- Abreu, F.V.S.D.; Ribeiro, I.P.; Ferreira-de-Brito, A.; Santos, A.A.C.D.; Miranda, R.M.D.; Bonelly, I.D.S.; Neves, M.S.A.S.; Bersot, M.I.; Santos, T.P.D.; Gomes, M.Q.; et al. Haemagogus leucocelaenus and Haemagogus janthinomys are the primary vectors in the major yellow fever outbreak in Brazil, 2016-2018. Emerg. Microbes Infect. 2019, 8, 218–231. [Google Scholar] [CrossRef] [PubMed]
- Drubulle, M; Mousson, L.; Moutailler, S.; Vazeille, M.; Failloux, A.-B. Chikungunya virus and Aedes mosquitoes: saliva is infectious as soon as two days after oral inection. Plos one 2009, 4. [Google Scholar] [CrossRef]
- Consoli, R.; Oliveira, R.L. Principais Mosquitos de Importância Sanitária no Brasil; Fiocruz: Rio de Janeiro, 1994; p. 228. [Google Scholar]
- Caldeira, R.; Nunes-Neto, J.; Maia, A.; Freitas, M.; Dias, D.; Pereira, G.; Vinagre, S.; Araújo, P.; Ferreira, M.; Martins, L.; et al. Infecção Experimental de Aedes albopictus Pelo Vírus da Febre Amarela. Sessão E-pôster, Eixo 4, Entomologia/Controle de Vetores, 55. Congresso da Sociedade Brasileira de Medicina Tropical e XXVI Congresso Brasileiro de Parasitologia, Belo Horizonte, Minas Gerais; 2019. [Google Scholar]
- Kubacki, J.; Flacio, E.; Qi, W.; Guidi, V.; Tonolla, M.; Fraefel, C. Viral metagenomic analysis of Aedes albopictus mosquitos from southern Switzerland. Viruses 2020, 12, 929. [Google Scholar] [CrossRef] [PubMed]
- Gamez, S.; Antoshechkin, I.; Mendez-Sanchez, S. C.; Akbari, O. S. The Developmental Transcriptome of Aedes albopictus, a Major Worldwide Human Disease Vector. Genes Genom Genet. 2020, 10, 1051–1062. [Google Scholar] [CrossRef]
- Abreu, F.V.S.D.; de Andreazzi, C.S.; Neves, M.S.A.S.; Meneguete, P.S.; Ribeiro, M.S.; Dias, C.M.G.; de Albuquerque Motta, M.; Barcellos, C.; Romão, A.R.; Magalhães, M.D.A.F.M.; et al. Ecological and environmental factors affecting transmission of sylvatic yellow fever in the 2017-2019 outbreak in the Atlantic Forest, Brazil. Parasites Vectors 2022, 15, 23. [Google Scholar] [CrossRef]
Figure 1.
Experimental procedure. Legend: (A) Experimental infection of non-human primates (NHP1, NHP2 and NHP3) with YFV; (B) NHP exposure to female Ae. albopictus (AL) on 3 dpi; (C) Exposure of healthy NHPs (NHP4, NHP5 and NHP6) to female Ae. albopictus on 14 dpi; (E) Exposure of healthy NHPs (NHP7, NHP8 and NHP9) to females Ae. albopictus on 21 dpi; (D) Segmentation of females into thorax, abdomen and head and extraction of saliva; (F) NHP and 50 mosquitoes used as negative controls.
Figure 1.
Experimental procedure. Legend: (A) Experimental infection of non-human primates (NHP1, NHP2 and NHP3) with YFV; (B) NHP exposure to female Ae. albopictus (AL) on 3 dpi; (C) Exposure of healthy NHPs (NHP4, NHP5 and NHP6) to female Ae. albopictus on 14 dpi; (E) Exposure of healthy NHPs (NHP7, NHP8 and NHP9) to females Ae. albopictus on 21 dpi; (D) Segmentation of females into thorax, abdomen and head and extraction of saliva; (F) NHP and 50 mosquitoes used as negative controls.
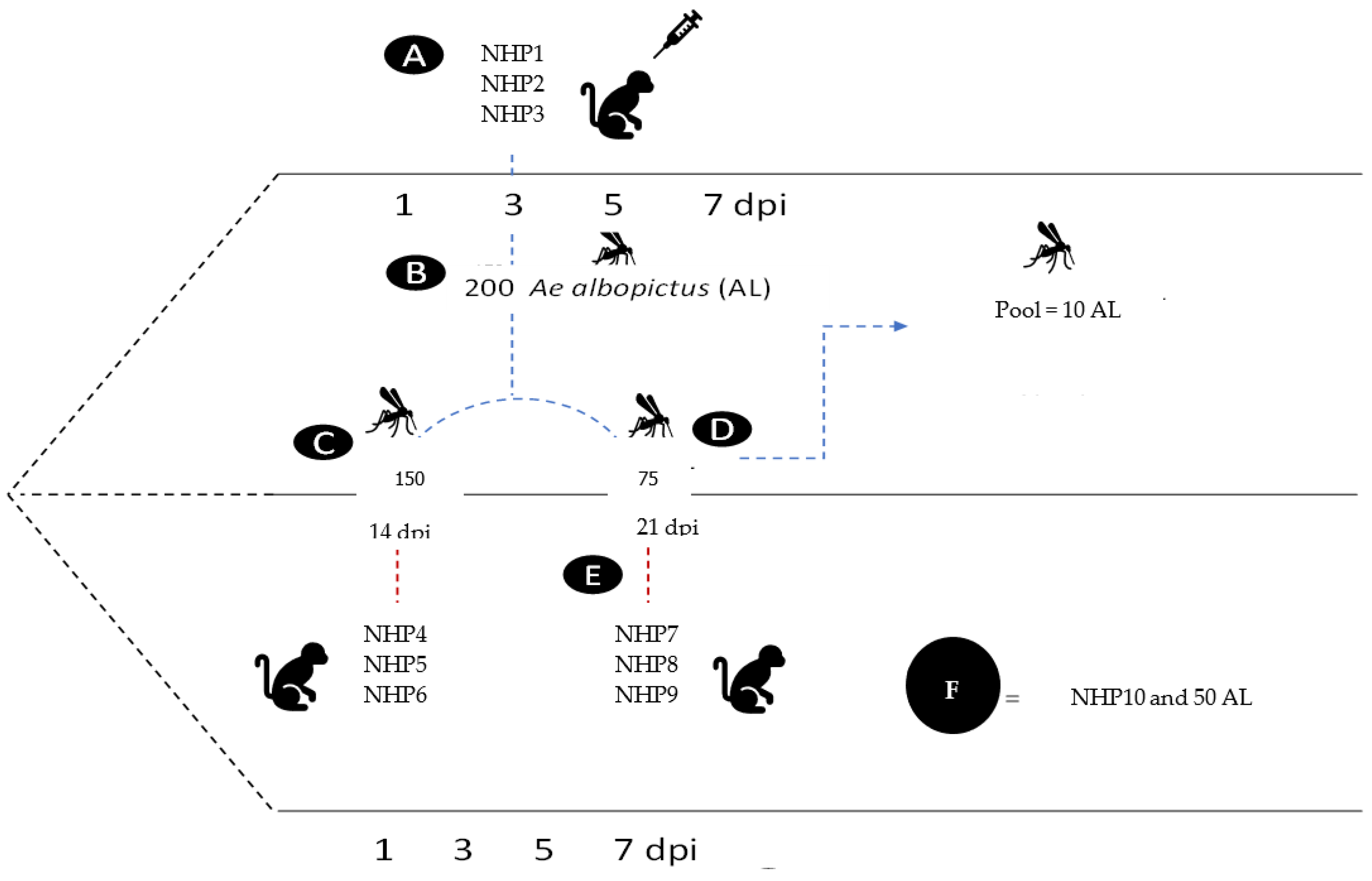
Figure 2.
Indirect immunofluorescence using polyclonal antibodies specific for YFV in C6/36 cell cultures: (A) Negative control; (B) Positive saliva pool sample (1AALS) on 14 dpi; (C) Positive saliva pool sample (3AALS) on 21 dpi. 400x magnification.
Figure 2.
Indirect immunofluorescence using polyclonal antibodies specific for YFV in C6/36 cell cultures: (A) Negative control; (B) Positive saliva pool sample (1AALS) on 14 dpi; (C) Positive saliva pool sample (3AALS) on 21 dpi. 400x magnification.
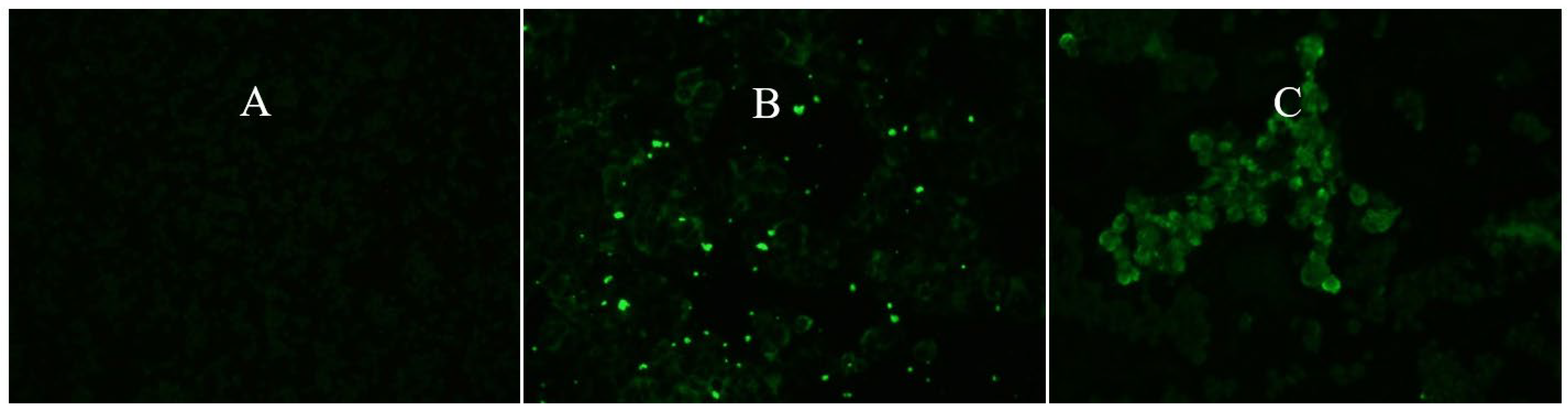
Figure 3.
Indirect immunofluorescence using monoclonal antibodies specific for YFV in C6/36 cell cultures: (A) Negative control; (B) Positive head sample (2AALC) analyzed at 14 dpi; (C) Positive head batch sample (3AALC) analyzed at 21 dpi; (D) Positive thorax/abdomen batch sample (2AALT) analyzed at 14 dpi; (E) Positive thorax/abdomen batch sample (3AALT) analyzed at 21 dpi; (F) Positive leg batch sample (2ALP) analyzed at 14 dpi; (G) Positive leg batch sample (3AALP) analyzed at 21 dpi. 40× magnification.
Figure 3.
Indirect immunofluorescence using monoclonal antibodies specific for YFV in C6/36 cell cultures: (A) Negative control; (B) Positive head sample (2AALC) analyzed at 14 dpi; (C) Positive head batch sample (3AALC) analyzed at 21 dpi; (D) Positive thorax/abdomen batch sample (2AALT) analyzed at 14 dpi; (E) Positive thorax/abdomen batch sample (3AALT) analyzed at 21 dpi; (F) Positive leg batch sample (2ALP) analyzed at 14 dpi; (G) Positive leg batch sample (3AALP) analyzed at 21 dpi. 40× magnification.

Table 1.
Ae. albopictus batches, dpi, cell culture results, and RT-qPCR quantification of YFV.
| Ae. albopictus (batch) | DPI | Isolation in cell culture | RT-qPCR | CT | Quantification (copies/mg) |
|---|---|---|---|---|---|
| 1AALT | 14 | Positive | Positive | 18.2 | >1.43 × 106 |
| 1AALC | 14 | Positive | Positive | 20.8 | 2.92 × 105 |
| 1AALP | 14 | Positive | Positive | 23.5 | 4.81 × 104 |
| 2AALT | 14 | Positive | Positive | 18.7 | 1.19 × 106 |
| 2AALC | 14 | Positive | Positive | 22.0 | 1.30 × 105 |
| 2AALP | 14 | Positive | Positive | 23.8 | 3.95 × 104 |
| 3AALT | 14 | Positive | Positive | 17.6 | >1.43 × 106 |
| 3AALC | 14 | Positive | Positive | 20.8 | 2.85 × 105 |
| 3AALP | 14 | Positive | Positive | 20.4 | 3.82 × 105 |
| 1AALT | 21 | Positive | Positive | 20.3 | 4.05 × 105 |
| 1AALC | 21 | Positive | Positive | 18.1 | 1.75 × 106 |
| 1AALP | 21 | Positive | Positive | 20.4 | 3.74 × 105 |
| 2AALT | 21 | Negative | Negative | >37 | 0 |
| 2AALC | 21 | Positive | Negative | >37 | 0 |
| 2AALP | 21 | Negative | Negative | >37 | 0 |
| 3AALT | 21 | Positive | Positive | 15.4 | >1.43 × 106 |
| 3AALC | 21 | Positive | Positive | 16.4 | >1.43 × 106 |
| 3AALP | 21 | Positive | Positive | 19.4 | 7.12 × 105 |
| 4AALT | 21 | Negative | Negative | >37 | 0 |
| 4AALC | 21 | Negative | Positive | 34.2 | <1.43 × 102 |
| 4AALP | 21 | Negative | Negative | >37 | 0 |
Legend: DPI = days post-infection; AALT = batch sample, thorax/abdomen; AALC = batch sample, head; AALP = batch sample, legs; CT = cycle threshold.
Table 2.
Non-human primates of the genus Callithrix exposed to Ae. albopictus infected with YFV.
| Primate | DPI | RT-qPCR | CT | Quantification |
|---|---|---|---|---|
| NHP1 | INOC | Positive | 28.1 | 463.4 |
| NHP2 | INOC | Positive | 22.5 | 17882.1 |
| NHP3 | INOC | Positive | 26.2 | 671.8 |
| NHP4 | 14 | Negative | NA | NA |
| NHP5 | 14 | Negative | NA | NA |
| NHP6 | 14 | Negative | NA | NA |
| NHP7 | 21 | Negative | NA | NA |
| NHP8 | 21 | Negative | NA | NA |
| NHP9 | 21 | Positive | 8.4 | 320019296.0 |
| NHP10 | NC | Negative | NA | NA |
Legend: NHP = non-human primate; DPI = days post-infection; INOC = inoculation; CT = cycle threshold; NC = negative control; NA = not applicable.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



