
Submitted:
18 September 2024
Posted:
20 September 2024
You are already at the latest version
Abstract
Often, in biology, we are faced with the problem of exploring relevant unknown biological hypotheses in the form of myriads of combination of factors that might be affecting the pathway under certain conditions. For example, Brancati et al.1 observe that mutations in poliovirus receptor related protein 4 (PVRL4), encoding cell adhesion molecule nectin-4, causes Ectodermal dysplasia-syndactyly syndrome. Interaction with cad- herins also implies an influence of nectin-4 on Wnt signaling, which plays a relevant role in limb development (Brancati et al.1). However, not much work has been done to explore the relation of Wnts and PVR family. In CRC cells treated with ETC-1922159, both were found up regulated. In a recent unpublished work in Open Science Framework, Sinha2, we had the opportunity to rank these unknown biological hypotheses for both up and down regulated genes at 2nd order level after drug administration. The search engine alloted high nu- merical valued rankings to some combinations of PVR-WNT, thus indicating a possibility of high combinatorial synergy also. The in-silico derived influences can be represented graphically as - PVR w.r.t WNT with PVR <- WNT9A; and WNT w.r.t PVR with WNT-7B/9A <- PVR and WNT4 <- PVRL2; In the light of the recent findings of PVR with IFN (interferon) and the known interactions between IFN and Wnts, there might be a possibilty to explore the bridge of PVR, IFN and WNTs. The 3 fold (PVR - IFN; IFN - WNT; WNT - PVR), 2 way cross family analysis might shed light on the possible combinations that might be of import. Here, we present a 2-way cross family analysis of multiple, such in-silico ranked 2nd order synergistic combinations, after ETC-1922159 treatment of CRC cells. Via this 2-way cross family analysis, we are able to discover through majority voting, the combinations that might of interest to biologists and also derive plausible influences of components of combinations among themselves. Note that these form biological hypotheses which indicate whether a particular combination and the direction of influence within the combination, exist synergistically in CRC cells. Wet lab tests will indicate the veracity of these combinations and if proven true, will lead to further study of mechanism between the components.
Keywords:
WNT
; NF-κB
; Ion Channels
; Anthrax toxin recep- tors
; Poliovirus receptors
; Porcupine inhibitor ETC-1922159
; Sensitivity analysis
; Colorectal cancer
; Unknown biological hypotheses
; Combinatorial search space
; Support vector ranking
This manuscript extends to 80 pages and is cumbersome to read. Thus the manuscript has been divided into 8 smaller articles, each focusing on a particular gene/protein of interest and its possible synergy with other genes/proteins. These 8 articles are as follows -
- Wnt related synergies in https://www.preprints.org/manuscript/202409.0453/v1
- NFκB related synergies in https://www.preprints.org/manuscript/202409.0696/v1
- TNF related synergies in https://www.preprints.org/manuscript/202409.0471/v1
- DNA repair related synergies in https://www.preprints.org/manuscript/202409.0885/v1
- ABC transporter related synergies in https://www.preprints.org/manuscript/202409.0908/v1
- Interleukin related synergies in https://www.preprints.org/manuscript/202409.1353/v1
- BCL related synergies in https://www.preprints.org/manuscript/202409.0855/v1
- ANTXR2 related synergies in https://www.preprints.org/manuscript/202409.0817/v1
1. Introduction
We reproduce a part of the manuscript [3] before we delve into the details of the current work. In [2], a frame work of a search engine is developed which can rank combinations of factors in a signaling pathway. Such combinations are of import due to the vast search space in which they exist and the difficulty to find them. The search engine facilitates in prioritizing the combinations as ranked biological hypotheses which the biologists might want to test in wet lab, to know if a synergistic combination is prevalent in a signaling pathway, in a (in)direct manner. Interested readers are advised to go through [2] for details regarding the search engine and the discoveries mentioned in there.
The research article, which is a mini encyclopaedia, contains results from this search engine. For each of the combinations of a family of genes at 2nd order level, we first present experimental confirmations of existence of combinations that are working synergistically, as cited in published literature. Next, based on the confirmed combinations, we infer plausible combinations, generated from the search engine, that might be working synergistically in CRC cells affected by ETC-1922159. The research shows ground breaking results in detail, which the biologists/oncologists can refer to for further wet lab tests. We present a 2-way cross family analysis of multiple, such in-silico ranked 2nd order synergistic combinations, after ETC-1922159 treatment of CRC cells. Via this 2-way cross family analysis, we are able to discover through majority voting, the combinations that might of interest to biologists and also derive plausible influences of components of combinations among themselves. Note that these form biological hypotheses which indicate whether a particular combination and the direction of influence within the combination, exist synergistically in CRC cells. Wet lab tests will indicate the veracity of these combinations and if proven true, will lead to further study of mechanism between the components.
2. Materials and Methods
2.1. Combinatorial Search Problem and a Possible Solution
We have already addressed the issue of combinatorial search problem and a possible solution in [3,4]. The details of the methodology of this manuscript have been explained in great detail in [4] & its application in [3]. Readers are requested to go through the same for gaining deeper insight into the working of the pipeline and its use of published data set generated after administration of ETC-1922159. In order to understand the significance of the solution proposed to the problem of combinatorial search that the biologists face in revealing unknown biological search problem, these works are of importance.
Briefly, the pipleline works by computing sensitivity indicies for each of these unique combinations and then vectorising these indices to connote and form discriminative feature vector for each combination. Since each combination is unique, the training and the test data are same. In the training data, the combinations are arranged and ranks from 1 to n are assigned. The ranking algorithm then learns the patterns from these combinations/sensitivity index vectors. Next the learned model is used to rank the test data by generating the ranking score for each of the unique combination. Sorting these shuffled scores of test data leads to prioritization of the combinations.
2.2. Wnt Signaling and Secretion
[5]’s accidental discovery of the Wingless played a pioneering role in the emergence of a widely expanding research field of the Wnt signaling pathway. A majority of the work has focused on issues related to • the discovery of genetic and epigenetic factors affecting the pathway [6,7], • implications of mutations in the pathway and its dominant role on cancer and other diseases [8], • investigation into the pathway’s contribution towards embryo development [9], homeostasis [10,11] and apoptosis [12] and • safety and feasibility of drug design for the Wnt pathway [13,14,15,16,17].
The Wnt phenomena can be roughly segregated into signaling and secretion part. The Wnt signaling pathway works when the WNT ligand gets attached to the Frizzled(FZD)/LRP coreceptor complex. FZD may interact with the Dishevelled (DVL) causing phosphorylation. It is also thought that Wnts cause phosphorylation of the LRP via casein kinase 1 (CK1) and kinase GSK3. These developments further lead to attraction of Axin which causes inhibition of the formation of the degradation complex. The degradation complex constitutes of AXIN, the β-catenin transportation complex APC, CK1 and GSK3. When the pathway is active the dissolution of the degradation complex leads to stabilization in the concentration of β-catenin in the cytoplasm. As β-catenin enters into the nucleus it displaces the GROUCHO and binds with transcription cell factor TCF thus instigating transcription of Wnt target genes. GROUCHO acts as lock on TCF and prevents the transcription of target genes which may induce cancer. In cases when the Wnt ligands are not captured by the coreceptor at the cell membrane, AXIN helps in formation of the degradation complex. The degradation complex phosphorylates β-catenin which is then recognised by F BOX/WD repeat protein β-TRCP. β-TRCP is a component of ubiquitin ligase complex that helps in ubiquitination of β-catenin thus marking it for degradation via the proteasome. A cartoon of the signaling transduction snapshot is shown in Figure 1.
Contrary to the signaling phenomena, the secretion phenomena is about the release and transportation of the WNT protein/ligand in and out of the cell, respectively. Briefly, the WNT proteins that are synthesized with the endoplasmic reticulum (ER), are known to be palmitoyleated via the Porcupine (PORCN) to form the WNT ligand, which is then ready for transportation [18]. It is believed that these ligands are then transported via the EVI/WNTLESS transmembrane complex out of the cell [19,20]. The EVI/WNTLESS themselves are known to reside in the Golgi bodies and interaction with the WNT ligands for the later’s glycosylation [21,22]. Once outside the cell, the WNTs then interact with the cell receptors, as explained in the foregoing paragraph, to induce the Wnt signaling. Of importance is the fact that the EVI/WNTLESS also need a transporter in the from of a complex termed as Retromer. A cartoon of the signaling transduction snapshot is shown in Figure 2.
2.3. PORCN-WNT Inhibitors
The regulation of the Wnt pathway is dependent on the production and secretion of the WNT proteins. Thus, the inhibition of a causal factor like PORCN which contributes to the WNT secretion has been proposed to be a way to interfere with the Wnt cascade, which might result in the growth of tumor. Several groups have been engaged in such studies and known PORCN-WNT inhibitors that have been made available till now are IWP-L6 [23,24], C59 [25], LGK974 [26] and ETC-1922159 [27]. In this study, the focus of the attention is on the implications of the ETC-1922159, after the drug has been administered. The drug is a enantiomer with a nanomolar activity and excellent bioavailability as claimed in [27].
3. Results & Discussion
3.1. WNT Related Synergies
3.1.1. WNT10B-ASCL2
WNT10B has been found to be implicated in a range of cancers. In gastric cancer, the knockdown of WNT10B showed reduced expression of cell proliferation and migration as well as inhibition of epithelial-mesenchymal transition Wu et al. [28]. On the other hand, WNT10B is also involved in the formation of bone mass and progenitor maintenance of various kinds of tissue, while deletion of the same leads to loss of bone mass and mesenchymal progenitor cells Stevens et al. [29]. Their contribution is also reported in axonal regeneration in injured CNS Tassew et al. [30]. Furthermore, like WNT10B, WNT10A and WNT6 have shown to play a major role in inhibiting adipogenesis and stimulates osteoblastogenesis while regulating the mesenchymal stem cells Cawthorn et al. [31] & Collins et al. [32]. Involvement in heptocellular carcinoma of WNT10B has been found wherein it is shown that stable silencing of WNT10B leads to significant reduction in proliferation, colony formation, migration and invasion in HepG2 HCC cell line Wu et al. [33]. Its implication in breast cancer Wend et al. [34] & Chen et al. [35] as well as endometrial cancer Chen et al. [36] has also been reported.
In colorectal cancer, WNT10B has shown to play a dual function of both oncogenesis promotion via β-catenin/TCF pathway and the inhibition of cell growth, possibly via FGF family of proteins Yoshikawa et al. [37]. Methylation of WNT10B has been found in the some of the cancer cell lines while its reversal has lead to over-expression of the WNT10B. However, the over-expression of WNT10B has lead to reduced cell growth in cancer, indicating a β-catenin independent component to be behind such a phenomena. Methylation of over-expressed WNT10B and synergistic work with FGF family of proteins later indicate the promotion of oncogenesis, as has been demonstrated in Yoshikawa et al. [37].
In a more recent work, ASCL2 has been found to play a major role in stemness in colon crypts and is implicated in colon cancer Zhu et al. [38]. Switching off the ASCL2 leads to a literal blockage of the stemness process and vice versa. At the downstream level, ASCL2 is regulated by TCF4/β-catenin via non-coding RNA target named WiNTRLINC1 Giakountis et al. [39]. Activation of ASCL2 leads to feedforward transcription of the non-coding RNA and thus a loop is formed which helps in the stemness and is highly effective in colon cancer. At the upstream level, ASCL2 is known act as a WNT/RSPONDIN switch that controls the stemness Schuijers et al. [40]. It has been shown that removal of RSPO1 lead to decrease in the Wnt signaling due to removal of the FZD receptors that led to reduced expression of ASCL2. Also, low levels of LGR5 were observed due to this phenomena. The opposite happened by increasing the RSPO1 levels. After the drug treatment, it was found that ASCL2 was highly suppressed pointing to the inhibition of stemness in the colorectal cancer cells. Also, Schuijers et al. [40] show that by genetically disrupting PORCN or inducing a PORCN inhibitor (like IWP-2), there is loss of stem cell markers like LGR5 and RNF43, which lead to disappearance of stem cells and moribund state of mice. A similar affect can be found with ETC-1922159, where there is suppression of RNF43 and LGR5 that lead to inhibition of the Wnt pathway and thus the ASCL2 regulation. These wet lab evidences are confirmed in the relatively low ranking of the combination ASCL2-RNF43 via the inhibition of PORCN-WNT that leads to blocking of the stemness that is induced by ASCL2. Since ASCL2 is directly mediated by the WNT proteins, the recorded ASCL2-WNT10B combination showed low priority ranking of 488, 497 and 321 for rbf, laplace and linear kernels, respectively, thus indicating a possible connection between WNT10B and ASCL2 activation. WNT10B might be playing a crucial role in stemness. This is further confirmed by wet lab experiments in Reddy et al. [41], which show BVES deletion results in amplified stem cell activity and Wnt signaling after radiation. WNT10B has been implicated in colorectal cancer Yoshikawa et al. [37].
3.1.2. ABC Transporters - WNT Cross Family Analysis
Hlavata et al. [42] have shown the role of ABC transporters in progression and clinical outcome of colorectal cancer. Work by Kobayashi et al. [43] show that Wnt-β catenin signaling regulates ABCC3 (MRP3) transporter expression in colorectal cancer. ABCA2 belongs to the category of ABC transporters that play an essential role in the development of resistance by the efflux of anticancer agents outside of cancer cells Hlavata et al. [42]. Hlavata et al. [42] observed that ABCA2 had no significant change/affect in colorectal cancer cases. Kobayashi et al. [43] found ABCA2 to be downregulated in colorectal cancer case. In ETC-1922159 affected CRC cells, down regulation of ABCA2 was observed, after the inhibition of proliferation in respective cells. Multiple members of ABC transporters and WNTs were found to be UP regulated after ETC-159 in CRC cells and WNTs are known to regulate ABCs. Below, we show a range of up regulated, possible unknown and unexplored synergistic 2nd order combinations that were ranked by the search engine. Note that the high numerical valued ranks (i.e nearing to 1800/2000 and above) indicate high potential of synergy that might be existing in CRC cells after the drug administration. Majority voting of rankings across the three different kernels point to the potential of the synergistic discovery. Wet labs investigations will assist in confirmation of these discoveries and if proven true, might lead to understanding of further mechanism between the components.
Table 1 and Table 2 show the rankings of ABC family w.r.t to WNT family members and WNT family w.r.t to ABC family members, respectively. From these two tables, we derive the plausible influences that might be existing in a two way format that is depicted in Table 3. In Table 1, WNT2B - ABC-C3 combination shows a majority voting of 1853 (laplace) and 2498 (rbf). Similarly, WNT7B - ABC-C13 shows a majority voting of 2245 (linear) and 2298 (rbf). These two combinations are depicted in Table 3 as ABC members influenced by WNT members (see under ABC w.r.t WNT). Reversibily, in Table 2 ABC-A5 - WNT2B shows a majority voting of 2018 (linear) and 2132 (rbf), ABC-A5 - WNT4 shows a majority voting of 2436 (linear) and 2449 (rbf), ABC-A5 - WNT9A shows a majority voting of 1989 (laplace), 2209 (linear) and 2365 (rbf), WNT2B - ABC-C5 shows a majority voting of 1970 (laplace), 2309 (linear) and 2248 (rbf), ABC-C5 - WNT9A shows a majority voting of 2183 (linear) and 2480 (rbf), WNT2B - ABC-C13 shows a majority voting of 2150 (linear) and 2048 (rbf), WNT7B - ABC-C13 shows a majority voting of 2508 (laplace) and 1830 (linear), WNT7B - ABC-D1 shows a majority voting of 2238 (laplace) and 2021 (linear), WNT7B - ABC-G1 shows a majority voting of 1808 (linear) and 1866 (rbf), WNT7B - ABC-G2 shows a majority voting of 2334 (linear) and 2145 (rbf) and WNT9A - ABC-G2 shows a majority voting of 1919 (laplace) and 2003 (rbf). These point to WNT members influenced by ABC members (see under WNT w.r.t ABC). Hypothetically, what we find is that the synergies can be bi-directional also and might contain various intermitent factors through which the factors might be working synergistically. These hypothese form present themselves as important combinations that might be of interest to biologists/oncologists.
One can also interpret the results of the Table 3 graphically, with the following influences - • ABC w.r.t WNT with WNT-2B -> ABC-C3; WNT-7B -> ABC-C13; and • WNT w.r.t ABC with ABC-A5 <- WNT-2B/4/9A; WNT-2B/9A <- ABC-C5; WNT-2B/7B <- ABC-C13; WNT-7B <- ABC-D1; WNT-7B <- ABC-G1; WNT-7B/9A <- ABC-G2. Thus, in this way, we can utilize the search engine to derive the various probable combinations between the factors of interest and their interdependent influences through the two-way cross family analysis.
3.1.3. IL - WNT Cross Family Analysis
Interleukin (IL) has been found in cross talk with WNT pathway. Kaler et al. [44] show that NFκB induced WNT signaling in colorectal cancer via interleukin-1β IL1B. Further, Zhong et al. [45] have shown that nitric oxide mediates crosstalk between interleukin 1β and Wnt signaling in primary human chondrocytes by reducing DKK1 and FRZB expression. The role of IL-17 (Interleukin-17) family is known to be controversial in CRC, however there are cases were it has been reported to be a prognostic marker for colorectal cancer Lin et al. [46] & Housseau et al. [47]. A homologue of the family, IL-17D a novel cytokine has been discovered Starnes et al. [48] and found to play a role in many of the cancers. In cells treated with ETC-1922159, IL-17D was found to be down regulated and reversibly it must have been regulated in the colorectal cancer cases. Recently, crosstalk between WNT/β-Catenin and NF-κB signaling pathway during inflammation has been reported by Ma and Hottiger [49]. Ma et al. [50] also show WNT/β-catenin negative feedback loop inhibits IL-1 induced matrix metalloproteinase expression in human articular chondrocytes. Masckauchán et al. [51] conclude that WNT/β-catenin signaling promotes angiogenesis possibly via the induction of known angiogenic regulators such as Interleukin-8. In mouse colon, Interleukin-1 signaling is shown to mediate obesity-promoted elevations in inflammatory cytokines, WNT activation, and epithelial proliferation by Pfalzer et al. [52]. In pulmonary fibrosis, Aumiller et al. [53] show that WNT/β-Catenin signaling induces IL-1β expression by alveolar epithelial cells. Chen et al. [54] show that IL-23 promotes the epithelial-mesenchymal transition of oesophageal carcinoma cells via the WNT/β-catenin pathway. Finally, Malysheva et al. [55] show that IL-6/WNT interactions in rheumatoid arthritis.
Family members belonging to each of the factors like WNT, IL etc, might be involved synergistically in pathological case or otherwise. IL and WNT members were found to be up regulated after the treatment of ETC-1922159 in colorectal cancer cells. We present here, multiple plausible and alternative synergistic combinatorial biological hypotheses for IL-WNT combination, which emerge after a cross family member analysis of the in silico revelations pertaining to the components under investigation.
Table 4 shows IL-WNT two way cross family analysis. The left side of the Table contains rankings of IL family with respect to WNTs and the right side of the Table contains rankings of WNT family with respect to ILs. Depicted in Table are the plausible combinatorial hypotheses derived from majority voting of the rankings in Table 4. On the left half, w.r.t WNT2B, IL-6ST/8/17REL show a synergy with WNT2B. These are reflected with rankings of 1797 (linear) and 2088 (rbf) for IL-6ST - WNT2B; rankings of 2107 (laplace), 1817 (linear) and 2088 (rbf) for IL-8 - WNT2B and rankings of 1824 (laplace) and 2241 (rbf) for IL-17REL - WNT2B, respectively. W.r.t WNT4, IL-1B/1RAP/15RA/17C show a synergy with WNT4. These are reflected with rankings of 1867 (laplace) and 1976 (linear) for IL-1B - WNT4; rankings of 2302 (laplace) and 1826 (linear) for IL-1RAP - WNT4; rankings of 1987 (laplace) and 2265 (linear) for IL-15RA - WNT4 and rankings of 2018 (laplace) and 1881 (linear) for IL-17C - WNT4, respectively. W.r.t WNT7B, IL-1RN/17REL show a synergy with WNT7B. These are reflected with rankings of 1882 (laplace) and 1796 (linear) for IL-1RN - WNT7B and rankings of 2053 (laplace), 2445 (linear) and 2489 (rbf) for IL-17REL - WNT4, respectively. W.r.t WNT9A, IL-1RAP/15RA show a synergy with WNT9A. These are reflected with rankings of 2273 (linear) and 2159 (rbf) for IL-1RAP - WNT9A and rankings of 1776 (laplace) and 2380 (linear) for IL-15RA - WNT9A, respectively.
On the right half, WNT2B w.r.t IL family, IL-1A/1RAP/8 show a synergy with WNT2B. These are reflected with rankings of 2290 (laplace) and 2427 (rbf) for IL-1A - WNT2B; rankings of 2488 (laplace) and 1892 (rbf) for IL-1RAP - WNT2B and rankings of 2157 1824 (laplace) and 2025 (linear) for IL-8 - WNT2B, respectively. WNT4 w.r.t IL family, IL-8/10RB show a synergy with WNT4. These are reflected with rankings of 1980 (laplace) and 2144 (linear) for IL-8 - WNT4 and rankings of 1828 (laplace), 2259 (linear) and 1993 (rbf) for IL-10RB - WNT4; respectively. WNT7B w.r.t IL family, IL-1A/1RN/6ST/17C show a synergy with WNT7B. These are reflected with rankings of 2134 (linear) and 2312 (rbf) for IL-1A - WNT7B; rankings of 1907 (laplace) and 2162 (linear) for IL-1RN - WNT7B; rankings of 1881 (linear) and 2020 (rbf) for IL-ST - WNT7B; and rankings of 1956 (laplace), 2388 (linear) and 1982 (rbf) for IL-17C - WNT7B, respectively. WNT9A w.r.t IL family, IL-1RAP/15RA/17REL show a synergy with WNT9A. These are reflected with rankings of 2003 (laplace) and 2179 (linear) for IL-1RAP - WNT9A; rankings of 2149 (laplace) and 2362 (linear) for IL-15RA - WNT9A; and rankings of 2101 (laplace) and 1940 (linear) for IL-17REL - WNT9A, respectively. One can also interpret the results of the Table 5 graphically, with the following influences - • IL w.r.t WNT with IL-6ST/8/17REL <- WNT-2B; IL-1B/1RAP/15RA/17C <- WNT-4; IL-1RN/17REL <- WNT-7B; IL-1RAP/15RA <- WNT-9A and • WNT w.r.t IL with IL-1A/1RAP/8 -> WNT-2B; IL-8/10RB -> WNT-4; IL-1A/1RN/6ST/17C -> WNT-7B and IL-1RAP/15RA/17REL -> WNT-9A.
3.1.4. UBE2 - WNT Cross Family Analysis
Mukai et al. [56] observed balanced ubiquitylation and deubiquitylation of Frizzled regulate cellular responsiveness to Wg/Wnt. Family members belonging to each of the factors like UBE2, WNT etc, might be involved synergistically in pathological case or otherwise. UBE2 and WNT members were found to be up regulated after the treatment of ETC-159 in colorectal cancer cells. However, not much is known about interation between the UBE2 family members and WNTs. Here we present a range of synergies that were ranked highly for up regulation. Table 6 presents the rankings of UBE family VS WNT family. Following this, is the Table 7 which derives the necessary influences via majority voting of rankings in Table 6.
On the left half, w.r.t WNT family, UBE2A show a synergy with WNT4. These are reflected with rankings of 2314 (linear) and 2279 (rbf) for UBE2A - WNT4; UBE2B show a synergy with WNT4/7B. These are reflected with rankings of 2260 (laplace), 2008 (linear) and 2141 (rbf) for UBE2B - WNT4 and rankings of 2116 (laplace) and 2206 (rbf) for UBE2B - WNT7B, respectively; UBE2F show a synergy with WNT4/7B. These are reflected with rankings of 2135 (laplace) and 2505 (linear) for UBE2F - WNT4 and rankings of 2423 (laplace) and 2077 (rbf) for UBE2F - WNT7B, respectively; UBE2H show a synergy with WNT2B. These are reflected with rankings of 1841 (laplace) and 2178 (linear) for UBE2H - WNT2B; UBE2J1 show a synergy with WNT-7B/9A. These are reflected with rankings of 2349 (laplace) and 2183 (rbf) for UBE2J1 - WNT7B and rankings of 1835 (laplace) and 2053 (rbf) for UBE2J1 - WNT9A, respectively. UBE2Z show a synergy with WNT-2B/4/9A. These are reflected with rankings of 1756 (linear) and 1878 (rbf) for UBE2J1 - WNT2B, rankings of 2195 (laplace) and 2468 (rbf) for UBE2J1 - WNT4, and 2343 (laplace) and 1973 (rbf) for UBE2J1 - WNT9A, respectively.

Table 7.
2nd order combinatorial hypotheses between UBE2 and WNT family members.
| Unexplored combinatorial hypotheses | |
| UBE2 w.r.t WNT | |
| WNT-4 | UBE2-A |
| WNT-4/7 | UBE2-B |
| WNT-4/7B | UBE2-F |
| WNT-2B | UBE2-H |
| WNT-7B/9B | UBE2-J1 |
| WNT-2B/4/7B | UBE2-Z |
| WNT w.r.t UBE2 | |
| WNT-7B | UBE2-A |
| WNT-7B/9A | UBE2-B |
| WNT-7B/9A | UBE2-F |
| WNT-4 | UBE2-H |
| WNT-7B/9A | UBE2-J1 |
| WNT-7B | UBE2-Z |
On the right half, w.r.t UBE2, UBE2A shows a synergy with WNT4. These are reflected with rankings of 2345 (linear) and 2151 (rbf) for UBE2A - WNT7B; UBE2B shows a synergy with WNT-7B/9A. These are reflected with rankings of 2052 (linear) and 1903 (rbf) for UBE2B - WNT7B and rankings of 2300 (laplace), 2476 (linear) and 2326 (rbf) for UBE2B - WNT9A, respectively; UBE2F shows a synergy with WNT-7B/9A. These are reflected with rankings of 2236 (laplace) and 1751 (rbf) for UBE2F - WNT7B and rankings of 2251 (linear) and 2179 (rbf) for UBE2F - WNT9A, respectively; UBE2H shows a synergy with WNT4. These are reflected with rankings of 2248 (linear) and 2155 (rbf) for UBE2H - WNT4; UBE2J1 shows a synergy with WNT-7B/9A. These are reflected with rankings of 1877 (llinear) and 1846 (rbf) for UBE2J1 - WNT7B and rankings of 2471 (laplace), 2137 (linear) and 2469 (rbf) for UBE2J1 - WNT9A, respectively. UBE2Z shows a synergy with WNT-9A. These are reflected with rankings of 1972 (laplace) and 1800 (linear) for UBE2Z - WNT7B, respectively.
One can also interpret the results of the Table 7 graphically, with the following influences - • UBE2 w.r.t WNT with WNT-4 -> UBE2-A; WNT-4/7 -> UBE2-B; WNT-4/7B -> UBE2-F; WNT-2B -> UBE2-H; WNT-7B/9B -> UBE2-J1; WNT-2B/4/7B -> UBE2-Z and • WNT w.r.t UBE2 with WNT-7B <- UBE2-A; WNT-7B/9A <- UBE2-B; WNT-7B/9A <- UBE2-F; WNT-4 <- UBE2-H; WNT-7B/9A <- UBE2-J1; WNT-7B <- UBE2-Z;
3.1.5. EXOSC - WNT10B Cross Family Analysis
Recently, emerging role of exosome (EXOSC) has been studied in WNT secretion and transportation by Zhang and Wrana [57]. It has been found that exosomes play a critical role in morphogen signaling during embryonic development and cancer progression. In injured CNS, exosomes mediate mobilization of WNT10B to promote axonal regeneration as shown by Tassew et al. [58]. Koles and Budnik [59] show the importance of exsosomes in WNT transportation. Emerging on these lines, we conducted a small two-way analysis of EXOSC components and WNT10B which were found to be down regulated in CRC cells after administration of ETC-1922159. Note that here, the interpretation of the rankings changes as the low numerical valued ranks (nearing to 1) are considered of high importance as they point to the synergistic down regulation after the drug administration. In line with the experiments, as ETC-1922159 a PORCN-WNT inhibitor block the transportation of WNTs, it might be that the affects of EXOSC components are also down regulated. These were rightly allocated with the low numerical valued in-silico ranks by the engine, thus pointing to the experimental down regulation in cells also. This confirmatory results also helps us in exploring the unknown combinations that might be prevailing synergistically when the WNT-EXOSC were up regulated before the administration of ETC-1922159 in CRC cells.
Table 8 shows the rankings of EXOSC family w.r.t WNT10B and vice versa. Followed by this is the unexplored combinatorial hypotheses in Table 9 generated from two-way analysis of the ranks in Table 8. On the left half of the Table 8, except for EXOSC7 - WNT10B, all other combinations of EXOSC family show high synergy with WNT10B. This is depicted by the low numerical valued ranks allocated by the search engine for EXOSC-2/3/5/6/8/9 with WNT10B, via majority voting across the ranking methods using laplace, linear and rbf kernels. This shows that EXOSC-2/3/5/6/8/9 had a critical role in the transport of WNT10B. On the right half of the same table, EXOSC-2/5/6/7/9 show synergistic affiliation with respect to WNT10B, via low numerical valued ranks. These are translated to graphical influences in Table 9. One can also interpret the results of the Table 9 graphically, with the following influences - • EXOSC w.r.t WNT10B with EXOSC-2/5/6/7/9 <- WNT10B and • WNT10B w.r.t EXOSC with EXOSC-2/3/5/6/8/9 -> WNT10B. Further analyses of these combinations in wet lab might help biologists explore the deeper mechanism of exosome components and WNT10B in CRC cells.
3.1.6. CASP - WNT Cross Family Analysis
Wu et al. [60] show that a caspase-dependent pathway is involved in Wnt/β-catenin signaling promoted apoptosis in Bacillus Calmette-Guerin infected RAW264.7 macrophages. Abdul-Ghani et al. [61] have shown that WNT11 promotes cardiomyocyte development by caspase-mediated suppression of canonical WNT signals. Additionally, Bisson et al. [62] show that Wnt5a and Wnt11 inhibit the canonical Wnt pathway and promote cardiac progenitor development via the Caspase-dependent degradation of AKT. These findings indicate probable interplay of Caspase and WNTs in various pathological cases. In mice, caspase-1 activation and IL-1βsecretion together have shown to contribute to inflammatory condition of acute arthritis (see Singh et al. [63]). Recently, Caspase-3 inhibition has been found to be a therapeutic approach in colorectal cancer as shown by Flanagan et al. [64]. Yao et al. [65] also show synergistic role of Caspase-8 and Caspase-3 expressions as biomarkers in colorectal cancer. Family members belonging to each of the factors like CASP, WNT etc, might be involved synergistically in pathological case or otherwise. CASP and WNT members were found to be up regulated after the treatment of colorectal cancer cells with ETC-1922159.
Table 10 shows the rankings of CASP family w.r.t WNTs and vice versa. Followed by this is the unexplored combinatorial hypotheses in Table 11 generated from two-way analysis of the ranks in Table 10. On the first three tabular rows of the Table 10 show rankings of CASP family w.r.t WNT family. Here we present the possible interdependent WNT-CASP combinations that might be working synergistically in CRC cells. Considering CASP5 w.r.t WNTs, CASP5 - WNT2B show up regulated synergy through rankings of 2171 (laplace) and 2366 (linear). Considering CASP9 w.r.t WNTs, CASP9 - WNT-4/7B/9A show up regulated synergy through rankings of 2472 (laplace) and 2200 (linear) for CASP9 - WNT4; 2196 (laplace) and 1935 (linear) for CASP9 - WNT7B; and 1863 (laplace) and 2002 (linear) for CASP9 - WNT9A, respectively. Finally, considering CASP16 w.r.t WNTs, CASP16 - WNT4 showed up regulated synergy with rankings of 2070 (laplace) and 1783 (linear).
The next three tabular rows show rankings of WNT family w.r.t CASP family. W.r.t CASP4, WNT-7B/9A show promise of up regulation. These are reflected with rankings of 2479 (linear) and 1739 (rbf) for WNT7B - CASP4 and rankings of 2278 (linear) and 1939 (rbf) for WNT9A - CASP4, respectively. W.r.t CASP5, WNT-7B shows promise of up regulation. This is reflected with rankings of 2112 (laplace), 1919 (linear) and 2440 (rbf) for WNT7B - CASP5. W.r.t CASP7, WNT-2B/4/9A show promise of up regulation. These are reflected with rankings of 2505 (laplace) and 1891 (linear) for WNT2B - CASP7; rankings of 2456 (linear) and 2455 (rbf) for WNT4 - CASP7; and rankings of 2183 (laplace) and 1941 (linear) for WNT9A - CASP7, respectively. W.r.t CASP9, WNT-9A shows promise of up regulation. This is reflected with rankings of 2378 (laplace), 2396 (linear) and 2058 (rbf) for WNT9A - CASP9. W.r.t CASP10, WNT-4/9A show promise of up regulation. These are reflected with rankings of 1830 (laplace), 2229 (linear) and 1847 (rbf) for WNT4 - CASP10; and rankings of 2185 (laplace) and 1977 (linear) for WNT9A - CASP10, respectively. Finally, w.r.t CASP16, WNT-2B/4/9A show promise of up regulation. These are reflected with rankings of 2197 (laplace), 2489 (linear) and 1775 (rbf) for WNT2B - CASP16; rankings of 2508 (laplace), 1820 (linear) and 1867 (rbf) for WNT7B - CASP16; and rankings of 1943 (laplace) and 1839 (linear) for WNT9A - CASP16, respectively.
Table 10.
2nd order interaction ranking between WNT VS CASP family members.
| Ranking CASP family VS WNT family | |||||||
| Ranking of CASP4 w.r.t WNTs family | Ranking of CASP5 w.r.t WNTs family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| CASP4 - WNT2B | 2265 | 320 | 1517 | CASP5 - WNT2B | 975 | 2171 | 2366 |
| CASP4 - WNT4 | 1050 | 1081 | 558 | CASP5 - WNT4 | 1788 | 1356 | 569 |
| CASP4 - WNT7B | 622 | 9 | 632 | CASP5 - WNT7B | 716 | 978 | 606 |
| CASP4 - WNT9A | 446 | 1413 | 583 | CASP5 - WNT9A | 383 | 808 | 147 |
| Ranking of CASP7 w.r.t WNTs family | Ranking of CASP9 w.r.t WNTs family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| CASP7 - WNT2B | 1152 | 305 | 248 | CASP9 - WNT2B | 1345 | 1501 | 1328 |
| CASP7 - WNT4 | 936 | 1260 | 1787 | CASP9 - WNT4 | 1344 | 2472 | 2200 |
| CASP7 - WNT7B | 901 | 1403 | 1303 | CASP9 - WNT7B | 2196 | 1935 | 1713 |
| CASP7 - WNT9A | 1330 | 1527 | 2436 | CASP9 - WNT9A | 1863 | 428 | 2002 |
| Ranking of CASP10 w.r.t WNTs family | Ranking of CASP16 w.r.t WNTs family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| CASP10 - WNT2B | 1607 | 1108 | 739 | CASP16 - WNT2B | 240 | 621 | 193 |
| CASP10 - WNT4 | 432 | 689 | 132 | CASP16 - WNT4 | 2070 | 1783 | 711 |
| CASP10 - WNT7B | 1906 | 1171 | 1165 | CASP16 - WNT7B | 411 | 713 | 103 |
| CASP10 - WNT9A | 1611 | 2152 | 1451 | CASP16 - WNT9A | 14 | 2512 | 181 |
| Ranking of WNTs family w.r.t CASP4 | Ranking of WNTs family w.r.t CASP5 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| CASP4 - WNT2B | 609 | 1317 | 2372 | CASP5 - WNT2B | 1849 | 1192 | 1590 |
| CASP4 - WNT4 | 105 | 711 | 1062 | CASP5 - WNT4 | 890 | 682 | 714 |
| CASP4 - WNT7B | 1093 | 2479 | 1739 | CASP5 - WNT7B | 2112 | 1919 | 2440 |
| CASP4 - WNT9A | 456 | 2278 | 1939 | CASP5 - WNT9A | 315 | 1880 | 1437 |
| Ranking of WNTs family w.r.t CASP7 | Ranking of WNTs family w.r.t CASP9 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| CASP7 - WNT2B | 2505 | 1891 | 1120 | CASP9 - WNT2B | 282 | 639 | 1414 |
| CASP7 - WNT4 | 108 | 2456 | 2455 | CASP9 - WNT4 | 572 | 1788 | 378 |
| CASP7 - WNT7B | 1380 | 1559 | 1681 | CASP9 - WNT7B | 979 | 901 | 676 |
| CASP7 - WNT9A | 2183 | 1941 | 1632 | CASP9 - WNT9A | 2378 | 2396 | 2058 |
| Ranking of WNTs family w.r.t CASP10 | Ranking of WNTs family w.r.t CASP16 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| CASP10 - WNT2B | 625 | 1471 | 81 | CASP16 - WNT2B | 2197 | 2489 | 1775 |
| CASP10 - WNT4 | 1830 | 2229 | 1847 | CASP16 - WNT4 | 1382 | 954 | 1017 |
| CASP10 - WNT7B | 1965 | 937 | 147 | CASP16 - WNT7B | 2508 | 1820 | 1867 |
| CASP10 - WNT9A | 2185 | 1977 | 1350 | CASP16 - WNT9A | 1943 | 1154 | 1839 |
One can also interpret the results of the Table 11 graphically, with the following influences - • CASP w.r.t WNT with CASP5 <- WNT2B; CASP9 <- WNT-4/7B/9A; CASP16 <- WNT4 and • WNT w.r.t CASP with. WNT-7B/9A <- CASP4; WNT7B <- CASP5; WNT-2B/4/9A <- CASP7; WNT9A <- CASP9; WNT-4/9A <- CASP10; WNT-2B/7B/9A <- CASP16.
Table 11.
2nd order combinatorial hypotheses between CASP and WNT family members.
| Unexplored combinatorial hypotheses | |
| CASP w.r.t WNT | |
| CASP5 | WNT2B |
| CASP9 | WNT4/WNT7B/WNT9A |
| CASP16 | WNT4 |
| WNT w.r.t CASP | |
| WNT7B/WNT9A | CASP4 |
| WNT7B | CASP5 |
| WNT2B/WNT4/WNT9A | CASP7 |
| WNT9A | CASP9 |
| WNT4/WNT9A | CASP10 |
| WNT2B/WNT7B/WNT9A | CASP16 |
3.1.7. TP53 - WNT Cross Family Analysis
Sadot et al. [66] have shown that down regulation of β-catenin is activated by TP53. Wnt/β-catenin signaling is known to regulate the proliferation and differentiation of mesenchymal progenitor cells through the TP53 Pathway, as shown by Peng et al. [67]. Zhukova et al. [68] show that WNT activation by lithium abrogates TP53 mutation associated radiation resistance in medulloblastoma. In mouse cochlea, Liu et al. [69] show that WNT signaling activates TP53-induced glycolysis and apoptosis regulator and protects against cisplatin-induced spiral ganglion neuron damage. These range of interactions of TP53 with WNT points towards definite synergy. Okayama et al. [70] show that TP53 protein regulates Hsp90 ATPase activity and thereby Wnt signaling by modulating Aha1 expression. Family members belonging to each of the factors like TP53, WNT etc, might be involved synergistically in pathological case or otherwise. TP53 and WNT members were found to be up regulated after the treatment of ETC-159 in colorectal cancer cells.
Table 12 contains rankings of TP53 w.r.t WNTs and vice versa. Followed by this is the unexplored combinatorial hypotheses in Table 13 generated from two-way analysis of the ranks in Table 12. On the left half of Table 12 are rankings of TP53 w.r.t WNTs and on the right half are the rankings of WNTs w.r.t TP53 family. Beginning with the left half, TP53I3 - WNT2B shows synergistic up regulation with rankings of 2056 (laplace) and 1712 (linear); TP53INP1 - WNT2B shows synergistic up regulation with rankings of 1805 (linear) and 2056 (rbf) and TP53BP2 - WNT9A shows synergistic up regulation with rankings of 2232 (linear) and 2143 (rbf). On the right half the table, TP53INP1 - WNT2B shows synergistic up regulation with rankings of 1853 (laplace) and 2089 (linear); TP53INP2 - WNT2B shows synergistic up regulation with rankings of 1723 (linear) and 2335 (rbf); TP53INP1 - WNT4 shows synergistic up regulation with rankings of 2414 (linear) and 2493 (rbf); TP53I3 - WNT7B shows synergistic up regulation with rankings of 1988 (laplace) and 2393 (rbf) and finally, TP53INP1 - WNT9A shows synergistic up regulation with rankings of 2045 (linear) and 2437 (rbf).
One can also interpret the results of the Table 11 graphically, with the following influences - • TP53 family w.r.t WNTs with TP53I3 <- WNT2B; TP53INP1 <- WNT2B and TP53BP2 <- WNT9A; and • WNT family VS TP53 with TP53INP1 -> WNT2B; TP53INP2 -> WNT2B; TP53INP1 -> WNT4; TP53I3 -> WNT7B and TP53INP1 -> WNT9A.
3.1.8. BCL - WNT Cross Family Analysis
Wang et al. [71] observed that silencing Wnt2B by siRNA interference inhibits metastasis and enhances chemotherapy sensitivity in ovarian cancer. More specifically, Wang et al. [71] show that in the presence of Wnt2B siRNA treatment, the caspase-9/B-cell lymphoma 2 (BCL2)/B-cell lymphoma-xL (BCL-xL) pathway and the epithelial-mesenchymal transition/phosphorylated protein kinase B pathway were inhibited. Takada et al. [72] show that targeted disruption of the BCL9/β-catenin complex inhibits oncogenic WNT signaling. CDK1-mediated BCL9 phosphorylation inhibits clathrin to promote mitotic Wnt signaling as shown by Chen et al. [73]. These findings point to the existing synergy of BCL family with WNTs. Family members belonging to each of the factors like BCL, WNT etc, might be involved synergistically in pathological case or otherwise. BCL and WNT members were found to be up regulated after the treatment of ETC-159 in colorectal cancer cells.
Table 14 contains rankings of BCL w.r.t WNTs and vice versa. Followed by this is the unexplored combinatorial hypotheses in Table 15 generated from two-way analysis of the ranks in Table 14. On the left half of Table 14 are rankings of BCL w.r.t WNTs. WNT4 - BCL2L2 shows high ranking with 2364 (laplace) and 2042 (linear); WNT7B - BCL2L2 shows high ranking with 1877 (laplace) and 2456 (linear); WNT9A - BCL2L2 shows high ranking with 1877 (laplace) and 2447 (linear); WNT4 - BCL2L13 shows high ranking with 1938 (laplace), 2425 (linear) and 1900 (rbf); WNT7B - BCL2L13 shows high ranking with 1993 (linear) and 2284 (rbf) and WNT2B - BCL10 shows high ranking with 2321 (laplace) and 2023 (linear).
On the right side are rankings of WNTs w.r.t BCL. WNT7B - BCL2L1 shows high ranking with 2213 (laplace) and 2266 (linear); WNT7B - BCL2L2 shows high ranking with 2456 (laplace), 2512 (linear) and 2286 (rbf); WNT9A - BCL2L2 shows high ranking with 1868 (laplace) and 2333 (rbf); WNT9A - BCL2L13 shows high ranking with 1858 (laplace), 2422 (linear) and 1934 (rbf); WNT2B - BCL3 shows high ranking with 1846 (laplace), 2056 (linear) and 1896 (rbf); WNT4 - BCL6 shows high ranking with 2483 (laplace) and 2488 (linear); WNT7B - BCL6 shows high ranking with 1893 (laplace) and 2284 (linear); WNT9A - BCL6 shows high ranking with 2098 (linear) and 1905 (rbf); WNT2B - BCL9L shows high ranking with 1918 (laplace) and 1882 (rbf) and WNT4 - BCL9L shows high ranking with 2498 (linear) and 2509 (rbf);
Table 14.
2nd order interaction ranking between WNT VS BCL family members.
| Ranking BCL family VS WNT | |||||||
| Ranking of BCL2L1 w.r.t WNT family | Ranking of WNT family w.r.t BCL2L1 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| WNT2B - BCL2L1 | 1884 | 101 | 966 | WNT2B - BCL2L1 | 1854 | 1666 | 1699 |
| WNT4 - BCL2L1 | 98 | 1162 | 719 | WNT4 - BCL2L1 | 21 | 107 | 16 |
| WNT7B - BCL2L1 | 1434 | 1891 | 620 | WNT7B - BCL2L1 | 2213 | 2266 | 1511 |
| WNT9A - BCL2L1 | 1088 | 1020 | 1318 | WNT9A - BCL2L1 | 1019 | 1462 | 1345 |
| Ranking of BCL2L2 w.r.t WNT family | Ranking of WNT family w.r.t BCL2L2 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| WNT2B - BCL2L2 | 625 | 2204 | 1677 | WNT2B - BCL2L2 | 1574 | 2206 | 955 |
| WNT4 - BCL2L2 | 2364 | 2042 | 1610 | WNT4 - BCL2L2 | 160 | 590 | 316 |
| WNT7B - BCL2L2 | 843 | 1877 | 2456 | WNT7B - BCL2L2 | 2456 | 2512 | 2286 |
| WNT9A - BCL2L2 | 1877 | 538 | 2447 | WNT9A - BCL2L2 | 1868 | 2333 | 990 |
| Ranking of BCL2L13 w.r.t WNT family | Ranking of WNT family w.r.t BCL2L13 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| WNT2B - BCL2L13 | 201 | 1862 | 1353 | WNT2B - BCL2L13 | 1256 | 1254 | 1490 |
| WNT4 - BCL2L13 | 1938 | 2425 | 1900 | WNT4 - BCL2L13 | 922 | 270 | 187 |
| WNT7B - BCL2L13 | 1105 | 1993 | 2284 | WNT7B - BCL2L13 | 1610 | 1319 | 954 |
| WNT9A - BCL2L13 | 1855 | 268 | 2387 | WNT9A - BCL2L13 | 1858 | 2422 | 1934 |
| Ranking of BCL3 w.r.t WNT family | Ranking of WNT family w.r.t BCL3 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| WNT2B - BCL3 | 950 | 1328 | 2482 | WNT2B - BCL3 | 1846 | 2056 | 1896 |
| WNT4 - BCL3 | 1228 | 1562 | 1353 | WNT4 - BCL3 | 591 | 359 | 1932 |
| WNT7B - BCL3 | 591 | 615 | 553 | WNT7B - BCL3 | 1687 | 2160 | 1428 |
| WNT9A - BCL3 | 1037 | 1410 | 1102 | WNT9A - BCL3 | 1539 | 1424 | 398 |
| Ranking of BCL6 w.r.t WNT family | Ranking of WNT family w.r.t BCL6 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| WNT2B - BCL6 | 455 | 2426 | 1529 | WNT2B - BCL6 | 52 | 107 | 170 |
| WNT4 - BCL6 | 256 | 486 | 787 | WNT4 - BCL6 | 2483 | 2488 | 1273 |
| WNT7B - BCL6 | 2147 | 1466 | 1105 | WNT7B - BCL6 | 975 | 1893 | 2284 |
| WNT9A - BCL6 | 1547 | 734 | 2012 | WNT9A - BCL6 | 1558 | 2098 | 1905 |
| Ranking of BCL9L w.r.t WNT family | Ranking of WNT family w.r.t BCL9L | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| WNT2B - BCL9L | 2348 | 804 | 1558 | WNT2B - BCL9L | 1918 | 700 | 1882 |
| WNT4 - BCL9L | 1446 | 657 | 309 | WNT4 - BCL9L | 303 | 2498 | 2509 |
| WNT7B - BCL9L | 1539 | 253 | 1279 | WNT7B - BCL9L | 1608 | 811 | 2168 |
| WNT9A - BCL9L | 1923 | 677 | 688 | WNT9A - BCL9L | 941 | 1843 | 1238 |
| Ranking of BCL10 w.r.t WNT family | Ranking of WNT family w.r.t BCL10 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| WNT2B - BCL10 | 2321 | 69 | 2023 | WNT2B - BCL10 | 1951 | 1101 | 1599 |
| WNT4 - BCL10 | 285 | 1170 | 465 | WNT4 - BCL10 | 2032 | 34 | 406 |
| WNT7B - BCL10 | 1847 | 606 | 1252 | WNT7B - BCL10 | 1297 | 74 | 2009 |
| WNT9A - BCL10 | 217 | 798 | 1649 | WNT9A - BCL10 | 1771 | 335 | 861 |
One can also interpret the results of the Table 15 graphically, with the following influences - • BCL family w.r.t WNTs with WNT4 -> BCL2L2; WNT7B -> BCL2L2; WNT9A -> BCL2L2; WNT4 -> BCL2L13; WNT7B -> BCL2L13; WNT2B -> BCL10 and • WNT family w.r.t BCL with WNT7B <- BCL2L1; WNT7B <- BCL2L2; WNT9A <- BCL2L2; WNT9A <- BCL2L13; WNT2B <- BCL3; WNT4 <- BCL6; WNT7B <- BCL6; WNT9A <- BCL6; WNT2B <- BCL9L; WNT4 <- BCL9L.
Table 15.
2nd order combinatorial hypotheses between TP53 and WNT family members.
| Unexplored combinatorial hypotheses | |
| BCL w.r.t WNT family | |
| WNT-4/7B/9A | BCL2L2 |
| WNT-4/7B | BCL2L13 |
| WNT-2B | BCL10 |
| WNT family w.r.t BCL | |
| WNT-7B | BCL2L1 |
| WNT-7B/9A | BCL2L2 |
| WNT-9A | BCL2L13 |
| WNT-2B | BCL3 |
| WNT-4/7B/9A | BCL6 |
| WNT-2B/4 | BCL9L |
3.2. NF-κB Related Synergies
3.2.1. CASP - RIPK Cross Family Analysis
The caspase - receptor interacting protein kinases (RIPK) has an intricate mechanism which has not yet been discovered and many views exist about their synergistic interaction. Green et al. [74] presents a review of RIPK-dependent necrosis and its regulation by CASPs. Furthermore, Lin et al. [75] show that cleavage of the death domain RIPK by CASP-8 prompts TNF-induced apoptosis. RIPK1 is known to promote death receptor-independent CASP-8 mediated apoptosis under unresolved ER stress conditions, as shown by Estornes et al. [76]. Weng et al. [77] show that CASP-8 and RIPK regulate bacteria-induced innate immune responses and cell death. Also, Moriwaki et al. [78] show that RIPK3-CASP8 complex mediates atypical pro-IL-1β processing. Recent work by Declercq et al. [79] shows RIPK importance in cell death and survival along with CASP influence. These interactions point to a definite synergy between the CASP - RIPK. Chaudhary et al. [80] showed activation of NF-κB pathway via Caspase-8 (CASP-8) and its homologs. Additionally, Caspase-8 was found to interact with Receptor-interacting serine/threonine-protein kinase 1 (RIPK1). Family members belonging to each of the factors like CASP, RIPK etc, might be involved synergistically in pathological case or otherwise. CASP and RIPK members were found to be up regulated after the treatment of ETC-1922159 in colorectal cancer cells.
Table 16 and Table 17 show the rankings of CASP family w.r.t RIPK and vice versa, respectively. Followed by this is the derived influences between CASP and RIPK via two way analysis of majority voting of rankings in the two foregoing tables. These influences are tabulated in Table 18. In Table 16, only CASP9 - RIPK3 combination showed up regulation with rankings of 2133 (laplace), 2030 (linear) and 2295 (rbf). In Table 17, RIPK1 showed up regulation with CASP-4/10 with rankings of 2363 (laplace) and 1805 (rbf) for CASP4 - RIPK1; and 2438 (laplace) and 1915 (linear) for CASP10 - RIPK1, respectively. RIPK2 showed up regulation with CASP-5/9/16 with rankings of 1776 (linear) and 2247 (rbf) for CASP5 - RIPK2; 2000 (laplace), 2476 (linear) and 2138 (rbf) for CASP9 - RIPK2; and 2006 (linear) and 2046 (rbf) for CASP16 - RIPK2; Finally, RIPK4 showed up regulation with CASP-16 with rankings of 2273 (laplace) and 2023 (linear) for CASP16 - RIPK4.
One can also interpret the results of the Table 18 graphically, with the following influences - • CASP w.r.t RIKP family with CASP9 <- RIPK3 and • RIPK w.r.t CASP family with RIPK1 <- CASP-4/10; RIPK2 <- CASP-5/9/16 and RIPK4 <- CASP16.
Table 16.
2nd order interaction ranking between CASP w.r.t RIPK family members.
| Ranking CASP family w.r.t RIPK family | |||||||
| Ranking of CASP4 w.r.t RIPK family | Ranking of CASP5 family w.r.t RIPK | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| CASP4 - RIPK1 | 1154 | 1259 | 147 | CASP5 - RIPK1 | 490 | 152 | 1818 |
| CASP4 - RIPK2 | 559 | 2147 | 434 | CASP5 - RIPK2 | 1274 | 2485 | 608 |
| CASP4 - RIPK3 | 111 | 131 | 41 | CASP5 - RIPK3 | 523 | 1047 | 317 |
| CASP4 - RIPK4 | 187 | 1048 | 1039 | CASP5 - RIPK4 | 1176 | 2361 | 1292 |
| Ranking of CASP7 w.r.t RIPK family | Ranking of CASP9 family w.r.t RIPK | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| CASP7 - RIPK1 | 2445 | 1289 | 1253 | CASP9 - RIPK1 | 1726 | 1304 | 1480 |
| CASP7 - RIPK2 | 1584 | 406 | 155 | CASP9 - RIPK2 | 2079 | 291 | 1647 |
| CASP7 - RIPK3 | 1406 | 1057 | 2091 | CASP9 - RIPK3 | 2133 | 2030 | 2295 |
| CASP7 - RIPK4 | 1739 | 231 | 2332 | CASP9 - RIPK4 | 2037 | 1627 | 363 |
| Ranking of CASP10 w.r.t RIPK family | Ranking of CASP16 family w.r.t RIPK | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| CASP10 - RIPK1 | 758 | 846 | 1405 | CASP16 - RIPK1 | 73 | 1046 | 1887 |
| CASP10 - RIPK2 | 1535 | 2312 | 884 | CASP16 - RIPK2 | 20 | 932 | 1189 |
| CASP10 - RIPK3 | 1530 | 250 | 2181 | CASP16 - RIPK3 | 30 | 359 | 717 |
| CASP10 - RIPK4 | 954 | 415 | 1547 | CASP16 - RIPK4 | 493 | 2507 | 519 |
Table 17.
2nd order interaction ranking between RIPK w.r.t CASP family members.
| Ranking RIPK family w.r.t CASP family | |||||||
| Ranking of RIPK family w.r.t CASP4 | Ranking of RIPK family w.r.t CASP5 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| CASP4 - RIPK1 | 2363 | 1374 | 1805 | CASP5 - RIPK1 | 7 | 82 | 131 |
| CASP4 - RIPK2 | 1713 | 2349 | 1261 | CASP5 - RIPK2 | 1577 | 1776 | 2247 |
| CASP4 - RIPK3 | 1397 | 768 | 1008 | CASP5 - RIPK3 | 574 | 14 | 30 |
| CASP4 - RIPK4 | 2215 | 1334 | 1425 | CASP5 - RIPK4 | 2448 | 1178 | 810 |
| Ranking of RIPK family w.r.t CASP7 | Ranking of RIPK family w.r.t CASP9 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| CASP7 - RIPK1 | 1341 | 2005 | 1131 | CASP9 - RIPK1 | 820 | 140 | 611 |
| CASP7 - RIPK2 | 1287 | 727 | 1143 | CASP9 - RIPK2 | 2000 | 2476 | 2138 |
| CASP7 - RIPK3 | 579 | 595 | 775 | CASP9 - RIPK3 | 1550 | 430 | 97 |
| CASP7 - RIPK4 | 852 | 1586 | 595 | CASP9 - RIPK4 | 1565 | 862 | 209 |
| Ranking of RIPK family w.r.t CASP10 | Ranking of RIPK family w.r.t CASP16 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| CASP10 - RIPK1 | 2438 | 1915 | 1039 | CASP16 - RIPK1 | 924 | 686 | 587 |
| CASP10 - RIPK2 | 1526 | 1800 | 1228 | CASP16 - RIPK2 | 1613 | 2006 | 2046 |
| CASP10 - RIPK3 | 419 | 1481 | 2001 | CASP16 - RIPK3 | 827 | 494 | 328 |
| CASP10 - RIPK4 | 1303 | 947 | 785 | CASP16 - RIPK4 | 2273 | 2023 | 1698 |
Table 18.
2nd order combinatorial hypotheses between CASP and RIPK.
| Unexplored combinatorial hypotheses | |
| CASP w.r.t RIKP family | |
| CASP9 | RIPK3 |
| RIPK w.r.t CASP family | |
| RIPK1 | CASP4/CASP10 |
| RIPK2 | CASP5/CASP9/CASP16 |
| RIPK4 | CASP16 |
3.2.2. MUC - RIPK Cross Family Analysis
In a recent work Sheng et al. [81] show that MUC13 promoted tumor necrosis factro (TNF)-induced NF-κB activation by interacting with TNFR1 and the E3 ligase, cIAP1, to increase ubiquitination of Receptor-interacting serine/threonine-protein kinase 1 (RIPK1). Family members belonging to each of the factors like MUC, RIPK etc, might be involved synergistically in pathological case or otherwise. MUC and RIPK members were found to be up regulated after the treatment of ETC-1922159 in colorectal cancer cells.
Table 19 and Table 20 show the rankings of MUC family w.r.t RIPK family and vice versa, respectively. Followed by this is the derived influences between MUC and RIPK. In Table 19, MUC1 was found to be highly upregulated with RIPK1. This is reflected in the rankings of 2027 (linear) and 2249 (rbf) for MUC1 - RIPK1. MUC3A was found to be highly upregulated with RIPK3. This is reflected in the rankings of 2208 (laplace) and 2017 (rbf) for MUC3A - RIPK3. MUC12 was found to be highly upregulated with RIPK4. This is reflected in the rankings of 2249 (linear) and 2130 (rbf), for MUC12 - RIPK4. MUC20 was found to be highly upregulated with RIPK3. This is reflected in the rankings of 2192 (laplace), 2288 (linear) and 1796 (rbf) for MUC20 - RIPK3.
In Table 20, RIPK-1/2 was found to be highly upregulated with MUC1. This is reflected in the rankings of 1839 (laplace) and 2421 (rbf) for MUC1 - RIPK1; and 1913 (laplace) and 2091 (linear) for MUC1 - RIPK2. RIPK4 was found to be highly upregulated with MUC4. This is reflected in the rankings of 1981 (laplace), 1949 (linear) and 2028 for MUC4 - RIPK4. RIPK4 was found to be highly up regulated with MUC17. This is reflected in the rankings of 2225 (linear) and 2048 (rbf) for MUC17 - RIPK4. RIPK2 was found to be highly up regulated with MUC20. This is reflected in the rankings of 1751 (linear) and 1950 (rbf) for MUC20 - RIPK2.
Table 19.
2nd order interaction ranking between MUC w.r.t RIPK family members.
| Ranking MUC family w.r.t RIPK family | |||||||
| Ranking of MUC1 w.r.t RIPK family | Ranking of MUCA3 w.r.t MUC3A | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| MUC1 - RIPK1 | 2027 | 2249 | 218 | MUC3A - RIPK1 | 945 | 186 | 1508 |
| MUC1 - RIPK2 | 248 | 1802 | 389 | MUC3A - RIPK2 | 840 | 2390 | 1653 |
| MUC1 - RIPK3 | 342 | 410 | 342 | MUC3A - RIPK3 | 2208 | 2017 | 689 |
| MUC1 - RIPK4 | 176 | 162 | 853 | MUC3A - RIPK4 | 714 | 1494 | 797 |
| Ranking of MUC4 w.r.t RIPK family | Ranking of MUC12 w.r.t RIPK family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| MUC4 - RIPK1 | 358 | 2384 | 690 | MUC12 - RIPK1 | 317 | 2437 | 167 |
| MUC4 - RIPK2 | 371 | 500 | 408 | MUC12 - RIPK2 | 286 | 2178 | 76 |
| MUC4 - RIPK3 | 809 | 371 | 1096 | MUC12 - RIPK3 | 747 | 366 | 136 |
| MUC4 - RIPK4 | 652 | 1863 | 1248 | MUC12 - RIPK4 | 176 | 2249 | 2130 |
| Ranking of MUC13 w.r.t RIPK family | Ranking of MUC17 w.r.t RIPK family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| MUC13 - RIPK1 | 379 | 2241 | 227 | MUC17 - RIPK1 | 858 | 932 | 1503 |
| MUC13 - RIPK2 | 824 | 2483 | 227 | MUC17 - RIPK2 | 248 | 934 | 37 |
| MUC13 - RIPK3 | 1687 | 19 | 24 | MUC17 - RIPK3 | 342 | 64 | 329 |
| MUC13 - RIPK4 | 562 | 532 | 184 | MUC17 - RIPK4 | 209 | 2335 | 1080 |
| Ranking of MUC20 w.r.t RIPK family | |||||||
| laplace | linear | rbf | |||||
| MUC20 - RIPK1 | 1419 | 760 | 1794 | ||||
| MUC20 - RIPK2 | 948 | 2482 | 137 | ||||
| MUC20 - RIPK3 | 2192 | 2288 | 1796 | ||||
| MUC20 - RIPK4 | 1564 | 1619 | 2179 | ||||
Table 20.
2nd order interaction ranking between RIPK w.r.t MUC family members.
| Ranking RIPK family w.r.t MUC family | |||||||
| Ranking of RIPK family w.r.t MUC1 | Ranking of RIPK family w.r.t MUC3A | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| MUC1 - RIPK1 | 1839 | 58 | 2421 | MUC3A - RIPK1 | 783 | 1668 | 1842 |
| MUC1 - RIPK2 | 1913 | 2091 | 954 | MUC3A - RIPK2 | 758 | 2301 | 459 |
| MUC1 - RIPK3 | 1038 | 268 | 295 | MUC3A - RIPK3 | 268 | 1595 | 1893 |
| MUC1 - RIPK4 | 1385 | 2246 | 1298 | MUC3A - RIPK4 | 1770 | 1109 | 1461 |
| Ranking of RIPK family w.r.t MUC4 | Ranking of RIPK family w.r.t MUC12 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| MUC4 - RIPK1 | 562 | 1621 | 2216 | MUC12 - RIPK1 | 1462 | 682 | 2351 |
| MUC4 - RIPK2 | 383 | 924 | 494 | MUC12 - RIPK2 | 989 | 597 | 1798 |
| MUC4 - RIPK3 | 541 | 43 | 129 | MUC12 - RIPK3 | 2158 | 1286 | 1636 |
| MUC4 - RIPK4 | 1981 | 1949 | 2028 | MUC12 - RIPK4 | 1577 | 975 | 976 |
| Ranking of RIPK family w.r.t MUC13 | Ranking of RIPK family w.r.t MUC17 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| MUC13 - RIPK1 | 1961 | 1535 | 32 | MUC17 - RIPK1 | 260 | 446 | 260 |
| MUC13 - RIPK2 | 784 | 494 | 1467 | MUC17 - RIPK2 | 1021 | 1114 | 2355 |
| MUC13 - RIPK3 | 860 | 1514 | 1425 | MUC17 - RIPK3 | 427 | 223 | 128 |
| MUC13 - RIPK4 | 107 | 1387 | 1972 | MUC17 - RIPK4 | 1567 | 2225 | 2048 |
| Ranking of RIPK family w.r.t MUC20 | |||||||
| laplace | linear | rbf | |||||
| MUC20 - RIPK1 | 514 | 2042 | 420 | ||||
| MUC20 - RIPK2 | 1039 | 1751 | 1950 | ||||
| MUC20 - RIPK3 | 303 | 2504 | 280 | ||||
| MUC20 - RIPK4 | 794 | 1193 | 989 | ||||
One can also interpret the results of the Table 21 graphically, with the following influences - • MUC w.r.t RIKP family with MUC1 <- RIPK1; MUC3A <- RIPK3; MUC12 <- RIPK4; MUC20 <- RIPK3 and • RIPK w.r.t MUC family with MUC1 -> RIPK-1/2; MUC4 -> RIPK4; MUC17 -> RIPK4; MUC20 -> RIPK2.
3.2.3. TNF - NF-κB-2/I Cross Family Analysis
The NF-κB family and NF-κB-Inhibitor i.e NF-κB-I play a significant role in immune response to infection. Problems in its functioning leads to cancer, infections, inflammatory and autoimmune diseases. The discovery and seminal work by Sen and Baltimore [82] on NF-κB lead to range of research on immune responses and study of related pathological cases. Tanaka and Nakano [83] have shown that NF-κB2 limits TNF-α induced osteoclastogenesis. Recently, in Japanese population, Imamura et al. [84] show that the impaired NF-κBIE gene function decreases cellular uptake of methotrexate by down-regulating SLC19A1 expression in a human rheumatoid arthritis cell line. They postulate that NF-κBIE could be closely related to NF-κB activity. Also, Lee et al. [85] show through deep study of fold-change analysis of the inter-relation between NF-κB and TNFs. However, the synergy between these members has yet not been explored completely. We found some interesting combinations that were allocated high numerical ranking (in silico) to indicate synergistic up regulation in CRC cells after ETC-1922159 treatment, apart from the individual up regulation that was observed in wet experiements.
Table 22 and Table 23 depict the rankings of TNF family w.r.t to NF-κB-2/I and vice versa, respectively. Followed by this is Table 24 that contains the derived influences via majority voting of the rankings in the tables containing two-way cross family rankings.
In Table 22 we find TNF-RSF10A/RSF12A up regulated with NFkB2. These are reflected in rankings of 2095 (laplace) and 2509 (rbf) for NFkB2 - TNFRSF10A; and 1813 (laplace) and 1893 (rbf) for NFkB2 - TNFRSF12A. TNF-AIP1/RSF10A/RSF10D/RSF14/SF10 were found to be up regulated with NFkBI-A. These are reflected in rankings of 1779 (laplace) and 1904 (linear) for NFkBI-A - TNF-AIP1; 2499 (laplace) and 2191 (rbf) for NFkBI-A - TNFRSF10A; 2498 (laplace), 2344 (linear) and 2501 (rbf) for NFkBI-A - TNFRSF10D; 1974 (laplace) and 2045 (linear) for NFkBI-A - TNFRSF14; and 2185 (laplace) and 2316 (rbf) for NFkBI-A - TNFSF10, respectively. TNF-AIP2/RSF14 were found to be up regulated with NFkBI-E. These are reflected in rankings of 2347 (laplace) and 1863 (linear) for NFkBI-E - TNFAIP2; and 1877 (laplace) and 2282 (linear) for NFkBI-E - TNFRSF14, respectively. Finally, TNF-RSF10B/RSF10D/RSF12A were found to be up regulated with NFkBI-Z. These are reflected in rankings of 2204 (laplace) and 1991 (rbf) for NFkBI-Z - TNFRSF10B; 2214 (laplace), 2033 (linear) and 2514 (rbf) for NFkBI-Z - TNFRSF10D; and 2370 (linear) and 1841 (rbf) for NFkBI-Z - TNFRSF12A, respectively. In Table 23 we find NFkB-2 to be up regulated along with TNF-AIP1/AIP2/AIP3. These are reflected in rankings of 2027 (linear) and 1807 (rbf) for NFkB2 - TNFAIP1; 2077 (laplace) and 2224 (rbf) for NFkB2 - TNFAIP2; and 2336 (linear) and 2130 (rbf) for NFkB2 - TNFAIP3, respectively. Finally, NFkBI-E was found to be up regulated with TNFRSF10D. These are reflected in rankings of 2136 (laplace) and 1811 (rbf) for NFkBI-E - TNFRSF10D.
One can also interpret the results of the Table 24 graphically, with the following influences - • TNF w.r.t NFkB family with NFkB2 -> TNF-RSF10A/RSF12A; NFkBI-A -> TNF-AIP1/RSF10A/RSF10D/RSF14/SF10; NFkBI-E -> TNF-AIP2/RSF14; NFkBI-Z -> TNF-RSF10B/RSF10D/RSF12A; and • NFkB w.r.t TNF family with NFkB-2 <- TNF-AIP1/AIP2/AIP3 and NFkBI-E <- TNF-RSF10D.
Table 24.
2nd order combinatorial hypotheses between NFkB-2/I and TNF.
| Unexplored combinatorial hypotheses | |
| TNF w.r.t NFkB-2/I | |
| NFkB2 | TNF-RSF10A/RSF12A |
| NFkBI-A | TNF-AIP1/RSF10A/RSF10D/RSF14/SF10 |
| NFkBI-E | TNF-AIP2/RSF14 |
| NFkBI-Z | TNF-RSF10B/RSF10D/RSF12A |
| NFkB-2/I w.r.t TNF | |
| NFkB-2 | TNF-AIP1/AIP2/AIP3 |
| NFkBI-E | TNF-RSF10D |
3.2.4. NFkB-2/I - STAT Cross Family Analysis
Grivennikov and Karin [86] show the potent collaboration and cross talk of STAT3 and NF-κB in cancer. In chronic lymphocytic leukemia cells, Liu et al. [87] observe that STAT-3 activates NF-κB. Co-opertion between STAT3 and NF-κB pathways has been observed in subtypes of diffuse large B Cell Lymphoma by Lam et al. [88]. Lee et al. [89] also shows a signal network involving coactivated NF-κB and STAT3 and altered p53 modulates BAX/BCL-XL expression and promotes cell survival of head and neck squamous cell carcinomas. These observations show a definite, concommitent functioning of the two pathways and we further found that some of them were up regulated synergistically in CRC cells after ETC-1922159 treatment, via in silico ranking of the combinations. Table 25 and Table 26 show ranking of STAT family w.r.t NFkB-2/I and vice versa, respectively. Followed by this is the derived influences from majority voting of rankings in the two foregoing tables, which is shown in Table 27.
Table 25 and Table 26 show the rankings of STAT family w.r.t NFkB-2/I and vice versa, respectively. Followed by this is the influence between the components in Table 27, via majority voting of the rankings. In the drug treated CRC cells, we found members of the STAT family to be up regulated with NFkB-2/I. These are reflected with rankings of 2211 (laplace) and 2402 (rbf) for NFkBIA -> STAT2; 2121 (linear) and 1862 (rbf) for NFkBIZ -> STAT2; and 1969 (linear) and 2485 (rbf) for NFkBIE -> STAT5A, respectively. One can also interpret the results of the Table 27 graphically, with the following influences - • STAT w.r.t NFkB-2/I with NFkBIA -> STAT2; NFkBIZ -> STAT2; and NFkBIE -> STAT5A;
Table 25.
2nd order interaction ranking between STAT w.r.t NFkB-2/I family members.
| Ranking STAT family w.r.t NFkB-2/I family | |||||||
| Ranking of STAT2 w.r.t NFkB-2/I family | Ranking of STAT3 w.r.t NFkB-2/I family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| NFkB2 - STAT2 | 2220 | 1068 | 1207 | NFkB2 - STAT3 | 2125 | 252 | 1453 |
| NFkBIA - STAT2 | 2211 | 1253 | 2402 | NFkBIA - STAT3 | 1614 | 702 | 1333 |
| NFkBIE - STAT2 | 1809 | 512 | 1207 | NFkBIE - STAT3 | 1493 | 211 | 1850 |
| NFkBIZ - STAT2 | 802 | 2121 | 1862 | NFkBIZ - STAT3 | 1633 | 1679 | 2122 |
| Ranking of NFkB-2/I family w.r.t STAT5A | |||||||
| laplace | linear | rbf | |||||
| NFkB2 - STAT5A | 2034 | 1321 | 1502 | ||||
| NFkBIA - STAT5A | 490 | 2215 | 283 | ||||
| NFkBIE - STAT5A | 578 | 1969 | 2485 | ||||
| NFkBIZ - STAT5A | 2286 | 473 | 1409 | ||||
Table 26.
2nd order interaction ranking between NFkB-2/I family w.r.t STAT members.
| Ranking NFkB-2/I family w.r.t STAT family | |||||||
| Ranking of NFkB-2/I family w.r.t STAT2 | Ranking of NFkB-2/I family w.r.t STAT3 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| NFkB2 - STAT2 | 935 | 952 | 86 | NFkB2 - STAT3 | 858 | 606 | 162 |
| NFkBIA - STAT2 | 543 | 36 | 1180 | NFkBIA - STAT3 | 1547 | 88 | 476 |
| NFkBIE - STAT2 | 1449 | 1861 | 1262 | NFkBIE - STAT3 | 1731 | 1063 | 509 |
| NFkBIZ - STAT2 | 483 | 1150 | 262 | NFkBIZ - STAT3 | 1262 | 489 | 1145 |
| Ranking of NFkB-2/I family w.r.t STAT3 | |||||||
| laplace | linear | rbf | |||||
| NFkB2 - STAT5A | 558 | 1070 | 670 | ||||
| NFkBIA - STAT5A | 1509 | 1020 | 81 | ||||
| NFkBIE - STAT5A | 18 | 854 | 1052 | ||||
| NFkBIZ - STAT5A | 83 | 1208 | 240 | ||||
Table 27.
2nd order combinatorial hypotheses between NFkB-2/I and TNF.
| Unexplored combinatorial hypotheses | |
| STAT w.r.t NFkB-2/I | |
| NFkBIA | STAT2 |
| NFkBIZ | STAT2 |
| NFkBIE | STAT5A |
3.2.5. IKBKE and STAT Cross Family Analysis
Ng et al. [90] show that phosphorylation of STAT1 by IκB kinase ε (IKBKE) inhibits STAT1 homodimerization, and thus assembly of GAF, but does not disrupt ISGF3 formation. Furthermore, Guo et al. [91] show that IKBKE is induced by STAT3 and tobacco carcinogen and determines chemosensitivity in non-small cell lung cancer. It has already been established in some cases that IKBKE has a confirmed role with one of the STAT members. Here we found that both IKBKE and STAT were up regulated after ETC-1922159 treatment of CRC cells. Table 28 shows ranking of STAT family vs IKBKE and vice versa. Table 29 shows the dervied influences from majority voting of the rankings. On the left half of Table 28 we find STAT2 to be up regulated w.r.t IKBKE. This is reflected with the rankings of 2033 (linear) and 1892 (rbf) for STAT2 - IKBKE. On the right half of the same Table we find IKBKE being up regulated w.r.t STAT-3/5A. These are reflected in rankings of 2179 (linear) and 1976 (rbf) for STAT3 - IKBKE; and 2085 (laplace), 2409 (linear) and 2277 (rbf) for STAT5A - IKBKE, respectively. One can also interpret the results of the Table 29 graphically, with the following influences - • STAT w.r.t IKBKE with STAT2 <- IKBKE; and • IKBKE w.r.t STAT with STAT3 -> IKBKE and STAT5A -> IKBKE;
3.2.6. IKBKE - TRAF Cross Family Analysis
Shen et al. [92] show interaction of IKBKE with TRAF2, by observing that IκB kinase ε phosphorylates TRAF2 to promote mammary epithelial cell transformation. Zhou et al. [93] observe IKKε-mediated tumorigenesis requires K63-linked polyubiquitination by a cIAP1/cIAP2/TRAF2 E3 ubiquitin ligase complex. Also, Nakanishi and Akira [94] show NF-κB activation through IKK-i-dependent I-TRAF/TANK phosphorylation. These findings suggest interaction between IKBKE - TRAF family members. IKBKE and TRAF members were found to be up regulated in CRC cells treated with ETC-1922159. Their combinations were allocated with high numerical ranks indicating synergistic up regulation. Table 30 rankings between TRAF and IKBKE, both ways. TRAF4 was found to up regulated with IKBKE and the rankings reflect the same with 2158 (linear) and 2416 (rbf). Also IKBKE was found to be up regulated with TRAF6 and the rankings reflect the same with 2105 (laplace) and 1819 (rbf). Table 31 reflects the derived influences graphically for - • TRAF w.r.t IKBKE with TRAF6 <- IKBKE and • IKBKE w.r.t TRAF with TRAF4 -> IKBKE.
3.2.7. ABC Transporters - NFkB Cross Family Analysis
Gerbod-Giannone et al. [95] observe that TNFα induces ABCA1 through NF-κB in macrophages and in phagocytes ingesting apoptotic cells. ABCA1 has also been found to be a key regulator in cholesterol related problems. Van Eck et al. [96] report leukocyte ABCA1 controls susceptibility to atherosclerosis and macrophage recruitment into tissues. The macrophage cholesterol exporter ABCA1 functions as an anti-inflammatory receptor, as shown by Tang et al. [97]. Furthermore, macrophage ABCA1 reduces MyD88-dependent Toll-like receptor trafficking to lipid rafts by reduction of lipid raft cholesterol, as shown by Zhu et al. [98]. These findings suggest the intricate role of NFκB family components play with ABC transporters. Both were up regulated in CRC cells after treatment with ETC-1922159. Our search engine allocated numerically high rank to several of the combinations in silico. These have been tabulated in Table 32 and Table 33, i.e rankings of ABC transporters w.r.t NFkB members and vice versa, respectively. Table 34 shows the un explored hypotheses between the two in the form of the derived influences after majority voting of the two-way cross family the rankings.
In Table 32, we find ABC-C13/ABC-D1 to be up regulated w.r.t. NFkBIE. These are reflected in rankings of 2048 (linear) and 1735 (rbf) for ABC-C13 - NFkBIE and 2380 (laplace) and 1795 (linear) for ABC-D1 - NFkBIE, respectively. In Table 33, we find NFkB2 to be up regulated w.r.t ABC-A5/ABC-B11. These are reflected in rankings of 2097 (laplace), 1772 (linear) and 2086 (rbf) for NFkB2 - ABC-A5; and 1916 (linear) and 1955 (rbf) for NFkB2 - ABC-B11, respectively. NFkBIE was up regulated with ABC-C13 and the rankings for the same are reflected in 2318 (laplace) and 2513 (rbf). Also, NFkBIZ was up regulated with ABC-C13 and the rankings for the same are reflected in 1799 (laplace) and 2175 (linear). NFkB2 was up regulated with ABC-G1 and the rankings for the same are reflected in 1951 (laplace), 2240 (linear) and 2215 (rbf).
Finally, Table 34 shows derived influences which can be represented graphically, with the following influences - • ABC w.r.t NFkB-2/I family with NFkIBE -> ABC-C13/ABC-D1 and • NFkB-2/I w.r.t ABC family with NFkB2 <- ABC-A5/ABC-B11; NFkBIE <- ABC-C13; NFkBIZ <- ABC-C13 and NFkB2 <- ABC-G1;
Table 32.
2nd order interaction ranking between ABC w.r.t NFkB-2/I family members.
| Ranking ABC family w.r.t NFkB-2/I family | |||||||
| Ranking of ABC family w.r.t NFkB2 | Ranking of ABC family w.r.t NFkBI-A | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| NFkB2 - ABC-A5 | 851 | 1517 | 350 | ABC-A5 - NFkBIA | 398 | 365 | 1660 |
| ABC-B11 - NFkB2 | 1684 | 400 | 412 | ABC-B11 - NFkBIA | 1079 | 566 | 104 |
| NFkB2 - ABC-C3 | 127 | 2031 | 6 | ABC-C3 - NFkBIA | 601 | 1048 | 1760 |
| NFkB2 - ABC-C5 | 1035 | 1431 | 889 | NFkBIA - ABC-C5 | 1683 | 2404 | 1341 |
| NFkB2 - ABC-C13 | 1399 | 1951 | 747 | NFkBIA - ABC-C13 | 200 | 886 | 1275 |
| NFkB2 - ABC-D1 | 1317 | 1133 | 1773 | ABC-D1 - NFkBIA | 1361 | 1361 | 1432 |
| NFkB2 - ABC-G1 | 1983 | 1343 | 1140 | ABC-G1 - NFkBIA | 21 | 313 | 461 |
| NFkB2 - ABC-G2 | 1322 | 955 | 1292 | ABC-G2 - NFkBIA | 809 | 613 | 48 |
| Ranking of ABC family w.r.t NFkBI-E | Ranking of ABC family w.r.t NFkBI-Z | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| ABC-A5 - NFkBIE | 1445 | 1662 | 679 | ABC-A5 - NFkBIZ | 699 | 1806 | 1290 |
| ABC-B11 - NFkBIE | 2285 | 1154 | 54 | ABC-B11 - NFkBIZ | 1240 | 37 | 803 |
| ABC-C3 - NFkBIE | 1547 | 2168 | 355 | ABC-C3 - NFkBIZ | 468 | 1366 | 1571 |
| NFkBIE - ABC-C5 | 876 | 2048 | 1735 | ABC-C5 - NFkBIZ | 1278 | 1714 | 1065 |
| NFkBIE - ABC-C13 | 623 | 1992 | 2351 | ABC-C13 - NFkBIZ | 1083 | 1063 | 1386 |
| ABC-D1 - NFkBIE | 2380 | 1795 | 861 | ABC-D1 - NFkBIZ | 1677 | 1688 | 794 |
| ABC-G1 - NFkBIE | 2193 | 251 | 208 | ABC-G1 - NFkBIZ | 979 | 2373 | 590 |
| ABC-G2 - NFkBIE | 2124 | 383 | 766 | ABC-G2 - NFkBIZ | 86 | 77 | 845 |
Table 33.
2nd order interaction ranking between NFkB-2/I w.r.t ABC family members.
| Ranking NFkB-2/I family w.r.t ABC family | |||||||
| Ranking of NFkB-2/I family w.r.t ABC-A5 | Ranking of NFkB-2/I family w.r.t ABC-B11 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| ABC-A5 - NFkB2 | 2097 | 1772 | 2086 | NFkB2 - ABC-B11 | 1916 | 1955 | 1020 |
| ABC-A5 - NFkBIA | 827 | 1142 | 379 | NFkBIA - ABC-B11 | 365 | 1702 | 602 |
| ABC-A5 - NFkBIE | 1276 | 1749 | 1795 | NFkBIE - ABC-B11 | 893 | 1285 | 1173 |
| ABC-A5 - NFkBIZ | 778 | 272 | 930 | NFkBIZ - ABC-B11 | 683 | 254 | 421 |
| Ranking of NFkB-2/I family w.r.t ABC-C3 | Ranking of NFkB-2/I family w.r.t ABC-C5 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| ABC-C3 - NFkB2 | 1225 | 936 | 281 | NFkB2 - ABC-C5 | 1510 | 1712 | 939 |
| ABC-C3 - NFkBIA | 782 | 271 | 1996 | NFkBIA - ABC-C5 | 2017 | 953 | 1649 |
| ABC-C3 - NFkBIE | 1071 | 1094 | 308 | NFkBIE - ABC-C5 | 567 | 615 | 1600 |
| ABC-C3 - NFkBIZ | 546 | 653 | 841 | ABC-C5 - NFkBIZ | 1978 | 943 | 160 |
| Ranking of NFkB-2/I family w.r.t ABC-C13 | Ranking of NFkB-2/I family w.r.t TNF-ABC-D1 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| NFkB2 - ABC-C13 | 618 | 1423 | 1550 | NFkB2 - ABC-D1 | 2094 | 1655 | 318 |
| NFkBIA - ABC-C13 | 1499 | 1092 | 456 | NFkBIA - ABC-D1 | 613 | 1812 | 1581 |
| NFkBIE - ABC-C13 | 2318 | 586 | 2513 | NFkBIE - ABC-D1 | 806 | 2204 | 410 |
| NFkBIZ - ABC-C13 | 1799 | 2175 | 1068 | NFkBIZ - ABC-D1 | 16 | 1723 | 955 |
| Ranking of NFkB-2/I family w.r.t ABC-G1 | Ranking of NFkB-2/I family w.r.t ABC-G2 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| NFkB2 - ABC-G1 | 1951 | 2240 | 2215 | NFkB2 - ABC-G2 | 957 | 1427 | 788 |
| NFkBIA - ABC-G1 | 1155 | 258 | 238 | NFKBIA - ABC-G2 | 508 | 417 | 686 |
| NFkBIE - ABC-G1 | 2034 | 612 | 490 | NFKBIE - ABC-G2 | 2223 | 806 | 685 |
| NFkBIZ - ABC-G1 | 1146 | 324 | 900 | NFkBIZ - ABC-G2 | 229 | 221 | 1196 |
Table 34.
2nd order combinatorial hypotheses between NFkB-2/I and ABC
| Unexplored combinatorial hypotheses | |
| ABC w.r.t NFkB-2/I family | |
| NFkIBE | ABC-C13/ABC-D1 |
| NFkB-2/I w.r.t ABC family | |
| NFkB2 | ABC-A5/ABC-B11 |
| NFkBIE | ABC-C13 |
| NFkBIZ | ABC-C13 |
| NFkB2 | ABC-G1 |
3.2.8. IKBKE - UBA/UBE Cross Family Analysis
Not much is known about IKBKE and Ubiquitination modifier enzyme and ubiquitination conjugating enzymes interaction. They were found them to be up regulated in CRC cells after ETC-1922159 treatment. Our search engine allocated high ranks to some of the combinations between IKBKE and UBA/UBE family members. These combinations might be worth exploring if it is of interest. Table 35 shows the rankings of UBE/A w.r.t to IKBKE and vice versa. We find IKBKE to be up regulated w.r.t UBA/E2 family. These are reflected with rankings of 2327 (laplace), 1807 (linear) and 2066 (rbf) for IKBKE - UBA-1; 2326 (linear) and 2456 (rbf) IKBKE - UBA-7; 2162 (laplace) and 1817 (linear) for IKBKE - UBA-P1; 2422 (laplace) and 2328 (rbf) for IKBKE - UBE2-A; 2367 (linear) and 2427 (rbf) for IKBKE - UBE2-B; and finally 2366 (laplace) and 1909 (rbf) for IKBKE - UBE2-Z; We also find UBA/E2 family to be up regulated w.r.t IKBKE also. This is reflected in rankings of 2189 (laplace) and 2271 (linear) for IKBKE - UBA-7; 2262 (laplace), 1901 (linear) and 2341 (rbf) for IKBKE - UBA-P1; 2293 (laplace), 2319 (linear) and 2396 (rbf) for IKBKE - UBE2-A; 2129 (laplace) and 1795 (linear) for IKBKE - UBE2-B; 2494 (laplace), 2233 (linear) and 1896 (rbf) for IKBKE - UBE2-F; 2016 (laplace) and 2103 (linear) for IKBKE - UBE2-Z;
Table 35.
2nd order interaction ranking between UBA/E2 family w.r.t IKBKE.
| Ranking UBA/E2 family vs IKBKE | |||||||
| Ranking of UBA/E2 family w.r.t IKBKE | Ranking of IKBKE w.r.t UBA/E2 family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IKBKE - UBA-1 | 1752 | 785 | 966 | UBA-1 - IKBKE | 2327 | 1807 | 2066 |
| IKBKE - UBA-7 | 2189 | 2271 | 1335 | IKBKE - UBA-7 | 1134 | 2326 | 2456 |
| IKBKE - UBA-P1 | 2262 | 1901 | 2341 | IKBKE - UBA-P1 | 2162 | 1817 | 1407 |
| IKBKE - UBA-LD2 | 2034 | 1773 | 1409 | IKBKE - UBA-LD2 | 1381 | 1647 | 556 |
| IKBKE - UBE2-A | 2293 | 2319 | 2396 | IKBKE - UBE2-A | 2422 | 536 | 2328 |
| IKBKE - UBE2-B | 2129 | 1516 | 1795 | IKBKE - UBE2-B | 680 | 2367 | 2427 |
| IKBKE - UBE2-F | 2494 | 2233 | 1896 | IKBKE - UBE2-F | 2309 | 181 | 24 |
| IKBKE - UBE2-H | 1265 | 1666 | 1257 | IKBKE - UBE2-H | 385 | 710 | 746 |
| IKBKE - UBE2-J1 | 905 | 1936 | 1046 | IKBKE - UBE2-J1 | 903 | 1729 | 2215 |
| IKBKE - UBE2-Z | 2016 | 2103 | 481 | IKBKE - UBE2-Z | 783 | 2366 | 1909 |
Table 36.
2nd order combinatorial hypotheses between NFkB-2/I and TNF
| Unexplored combinatorial hypotheses | |
| UBA/E2 w.r.t IKBKE | |
| IKBKE | UBA-1/7/P1 |
| IKBKE | UBE2-A/B/Z |
| IKBKE w.r.t UBE/A2 | |
| IKBKE | UBA-7/P1/LD2 |
| IKBKE | UBE2-A/B/F/Z |
Table 36 shows the derived influences which can be represented graphically, with the following influences - • UBA/E2 w.r.t IKBKE with IKBKE -> UBA-1; IKBKE -> UBA-7; IKBKE -> UBA-P1; and IKBKE -> UBE2-A; IKBKE -> UBE2-B; IKBKE -> UBE2-Z •; IKBKE w.r.t UBE/A2 with IKBKE <- UBA-7; IKBKE <- UBA-P1; IKBKE <- UBA-LD2; and IKBKE <- UBE2-A; IKBKE <- UBE2-B; IKBKE <- UBE2-F; IKBKE <- UBE2-Z;
3.2.9. REL-A/B - NF-kB Cross Family Analysis
Table 37.
2nd order interaction ranking between NFkB-2/I VS REL-A family members.
| Ranking NFkB-2/I VS REL-A | |||||||
| Ranking of NFkB-2/I family w.r.t REL-A | Ranking of REL-A w.r.t NFkB-2/I family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| NFkB2 - RELA | 664 | 420 | 271 | NFkB2 - RELA | 2454 | 794 | 2307 |
| NFKBIA - RELA | 198 | 205 | 190 | NFKBIA - RELA | 2106 | 2305 | 1153 |
| NFKBIE - RELA | 1503 | 2321 | 331 | NFKBIE - RELA | 1664 | 456 | 1926 |
| NFKBIZ - RELA | 323 | 1714 | 619 | NFKBIZ - RELA | 1924 | 1687 | 1584 |
Table 38.
2nd order interaction ranking between NFkB-2/I VS REL-B family members.
| Ranking REL-B VS NFkB-2/I family | |||||||
| Ranking of NFKB-2/I w.r.t REL-B | Ranking of REL-B w.r.t NFKB-2/I | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| NFkB2 - RELB | 503 | 2146 | 1788 | NFkB2 - RELB | 1156 | 1346 | 2184 |
| NFKBIA - RELB | 239 | 1576 | 924 | NFKBIA - RELB | 968 | 424 | 1725 |
| NFKBIE - RELB | 1203 | 714 | 2200 | NFKBIE - RELB | 1414 | 2228 | 800 |
| NFKBIZ - RELB | 1776 | 2244 | 1869 | NFKBIZ - RELB | 746 | 1281 | 1055 |
REL-A is known to be associated with NF-κB and most deeply studied member of the NF-κB. Tian et al. [99] observe that the NFkB subunit RELA is a master transcriptional regulator of the committed epithelial-mesenchymal transition in airway epithelial cells. Ke et al. [100] observe that inactivation of NF-κB p65 (RelA) in liver improves insulin sensitivity and inhibits cAMP/PKA pathway. Weichert et al. [101] observe that high expression of RelA/p65 is associated with activation of NF-κB-dependent signaling in pancreatic cancer. These findings and many others not cited here show the deep interaction between REL and NF-κB members. Table 37 shows rankings of RELA w.r.t NFkB members and vice versa. Table 38 shows rankings of RELB w.r.t NFkB members and vice versa. Finally, Table 39 shows the hypotheses generated from majority voting of the ranks. In Table 37 we find RELA to be up regulated w.r.t NFKB2. This is reflected in rankings of 2454 (laplace) and 2307 (rbf) for NFkB2 - RELA. Similarly, NFKBIA was found to be up regulated w.r.t RELA. This is reflected in rankings of 2106 (laplace) and 2305 (linear) for NFKBIA - RELA. In Table 38 we find NFkB2 to be up regulated RELB. This is reflected in 2146 (laplace) and 1788 (rbf) for NFkB2 - RELB. Similarly, we find NFKBIZ to be 1776 (laplace), 2244 (linear) and 1869 (rbf) for NFKBIZ - RELB. Table 39 shows the derived influences which can be represented graphically, with the following influences - • NFkB-2/I family w.r.t REL-B with NFkB2 <- REL-B and NFKBIZ <- RELB and • REL-A w.r.t NFkB-2/I family with NFkB2 -> RELA and NFKBIA -> RELA.
3.3. Tumor Necrosis Factor Related Synergies
3.3.1. TNF - WNT Cross Family Analysis
Brooks et al. [102] observed TNF-α induced alterations in the Wnt signaling cascade as a potential mechanism for obesity-associated colorectal tumorigenesis. Effects of TNF inhibitors on parathyroid hormone and Wnt signaling antagonists in rheumatoid arthritis have been studies in Adami et al. [103]. A complex interaction between Wnt signaling and TNF-α in nucleus pulposus cells has been studied by Hiyama et al. [104]. Ma and Hottiger [49] study the crosstalk between Wnt/β-catenin and NF-κB signaling pathway during inflammation. Roubert et al. [105] study the influence of tumor necrosis factor-α on the tumorigenic Wnt-signaling pathway in human mammary tissue from obese women. Jang et al. [106] observe that WNT/β-catenin pathway modulates the TNF-α-induced inflammatory response in bronchial epithelial cells. These studies suggest already existing synergistic roles of WNTs and TNFs. In CRC cells affected with ETC-1922159, members of TNF and WNT family were found to be up regulated. Our search engine alloted high numerical valued ranks to some of the combinations between WNTs and TNFs. Table 40 shows rankings of TNF w.r.t to WNTs on the left half and vice verse on the right half.
On the left half, we found TNF-RSF1A/RSF10A/RSF10B/SF15 to be up regulated w.r.t WNT2B. These were reflected in rankings of 2170 (laplace) and 2127 (linear) for TNFRSF1A - WNT2B; 1861 (laplace), 2367 (linear) and 1800 (rbf) for TNFRSF10A - WNT2B; 2020 (laplace) and 1881 (rbf) for TNFRSF10B - WNT2B and 2476 (laplace) and 2073 (rbf) for TNFSF15 - WNT2B. TNF-RSF10A/RSF10D/RSF12A/SF10 were found to be up regulated w.r.t WNT4. These were reflected in rankings of 2509 (laplace) and 2460 (linear) for TNFRSF10A - WNT4; 2233 (linear) and 2126 (rbf) for TNFRSF10D - WNT4; 2294 (linear), 1775 (linear) and 2384 (rbf) for TNFRSF12A - WNT4 and 2451 (linear) and 1782 (rbf) for TNFSF10 - WNT4. TNF-RSF12A/SF10 were found to be up regulated w.r.t WNT7B. These were reflected in rankings of 2100 (laplace) and 1983 (rbf) for TNFRSF12A - WNT7B and 2462 (laplace) and 2179 (rbf) for TNFSF10 - WNT7B. TNFRSF21 were found to be up regulated w.r.t WNT9A. These were reflected in rankings of 1805 (laplace) and 1999 (linear) for TNFRSF21 - WNT9A.
On the left half, we found WNT2B to be up regulated w.r.t TNF-RSF10B/RSF10D/RSF14. These were reflected in rankings of 1797 (laplace) and 2056 (rbf) for TNFRSF10B - WNT2B; 1989 (linear) and 2130 (rbf) for TNFRSF10D - WNT2B and 1932 (laplace) and 2399 (rbf) for TNFRSF14 - WNT2B. WNT4 was upregulated w.r.t TNF-AIP3/RSF10B. These are refliected in rankings of 2336 (laplace), 2511 (linear) and 2342 (rbf) for TNFAIP3 - WNT4 and 2105 (linear) and 2264 (rbf) for TNFRSF10B - WNT4. WNT7B was upregulated w.r.t TNF, TNF-RSF1A/RSF14. These are reflected in rankings of 2511 (linear) and 2210 (rbf) for TNF - WNT7B; 2084 (laplace), 1975 (linear) and 2154 (rbf) for TNFRSF1A - WNT7B and 2079 (laplace) and 1928 (rbf) for TNFRSF14 - WNT7B. WNT9A was upregulated w.r.t TNF-AIP2/AIP3/RSF10A/RSF12A/SF10. These are reflected in rankings of 2125 (laplace) and 2437 (linear) for TNFAIP2 - WNT9A; 1764 (laplace) and 2460 (linear) for TNFAIP3 - WNT9A; 2259 (laplace) and 2413 (linear) for TNFRSF10A - WNT9A; 2345 (laplace) and 2466 (rbf) for TNFRSF12A - WNT9A and 2054 (laplace) and 2338 (linear) for TNFSF10 - WNT9A.
Table 40.
2nd order combinatorial hypotheses between ABC and IL
| Ranking TNF family vs WNT family | |||||||
| Ranking of TNF family w.r.t WNT2B | Ranking of WNT2B w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| TNF - WNT2B | 503 | 893 | 1656 | TNF - WNT2B | 1341 | 808 | 1366 |
| TNFAIP1 - WNT2B | 235 | 156 | 1811 | TNFAIP1 - WNT2B | 1671 | 1434 | 1404 |
| TNFAIP2 - WNT2B | 868 | 527 | 2439 | TNFAIP2 - WNT2B | 1130 | 218 | 1105 |
| TNFAIP3 - WNT2B | 1135 | 2381 | 1688 | TNFAIP3 - WNT2B | 997 | 1280 | 1902 |
| TNFRSF1A - WNT2B | 2170 | 2127 | 1628 | TNFRSF1A - WNT2B | 1747 | 1857 | 1550 |
| TNFRSF10A - WNT2B | 1861 | 2367 | 1800 | TNFRSF10A - WNT2B | 100 | 464 | 1162 |
| TNFRSF10B - WNT2B | 2020 | 615 | 1881 | TNFRSF10B - WNT2B | 1797 | 120 | 2056 |
| TNFRSF10D - WNT2B | 29 | 2515 | 1174 | TNFRSF10D - WNT2B | 1348 | 1989 | 2130 |
| TNFRSF12A - WNT2B | 1072 | 2061 | 1109 | TNFRSF12A - WNT2B | 1595 | 298 | 1432 |
| TNFRSF14 - WNT2B | 333 | 1585 | 1247 | TNFRSF14 - WNT2B | 1932 | 277 | 2399 |
| TNFRSF21 - WNT2B | 1275 | 648 | 1114 | TNFRSF21 - WNT2B | 1396 | 620 | 2136 |
| TNFSF10 - WNT2B | 1204 | 2287 | 1396 | TNFSF10 - WNT2B | 1732 | 738 | 1751 |
| TNFSF15 - WNT2B | 2476 | 359 | 2073 | TNFSF15 - WNT2B | 402 | 128 | 1875 |
| Ranking of TNF family w.r.t WNT4 | Ranking of WNT4 w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| TNF - WNT4 | 1982 | 1301 | 928 | TNF - WNT4 | 1021 | 420 | 864 |
| TNFAIP1 - WNT4 | 1434 | 1078 | 804 | TNFAIP1 - WNT4 | 1114 | 337 | 1015 |
| TNFAIP2 - WNT4 | 1810 | 1047 | 330 | TNFAIP2 - WNT4 | 1611 | 1341 | 423 |
| TNFAIP3 - WNT4 | 646 | 1955 | 1534 | TNFAIP3 - WNT4 | 2336 | 2511 | 2342 |
| TNFRSF1A - WNT4 | 915 | 545 | 829 | TNFRSF1A - WNT4 | 132 | 333 | 1321 |
| TNFRSF10A - WNT4 | 2509 | 2460 | 897 | TNFRSF10A - WNT4 | 535 | 202 | 582 |
| TNFRSF10B - WNT4 | 517 | 875 | 1365 | TNFRSF10B - WNT4 | 320 | 2105 | 2264 |
| TNFRSF10D - WNT4 | 1719 | 2233 | 2126 | TNFRSF10D - WNT4 | 660 | 49 | 341 |
| TNFRSF12A - WNT4 | 2294 | 1775 | 2384 | TNFRSF12A - WNT4 | 649 | 1756 | 780 |
| TNFRSF14 - WNT4 | 1608 | 2284 | 1436 | TNFRSF14 - WNT4 | 61 | 519 | 1542 |
| TNFRSF21 - WNT4 | 1915 | 1596 | 93 | TNFRSF21 - WNT4 | 201 | 533 | 657 |
| TNFSF10 - WNT4 | 1747 | 2451 | 1782 | TNFSF10 - WNT4 | 904 | 1511 | 2280 |
| TNFSF15 - WNT4 | 1542 | 806 | 2439 | TNFSF15 - WNT4 | 64 | 709 | 793 |
| Ranking of TNF family w.r.t WNT7 | Ranking of WNT7 w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| TNF - WNT7B | 815 | 381 | 47 | TNF - WNT7B | 1530 | 2511 | 2210 |
| TNFAIP1 - WNT7B | 313 | 1438 | 992 | TNFAIP1 - WNT7B | 2196 | 519 | 1058 |
| TNFAIP2 - WNT7B | 1897 | 85 | 631 | TNFAIP2 - WNT7B | 2121 | 599 | 1313 |
| TNFAIP3 - WNT7B | 577 | 1807 | 1251 | TNFAIP3 - WNT7B | 1901 | 1357 | 830 |
| TNFRSF1A - WNT7B | 165 | 844 | 353 | TNFRSF1A - WNT7B | 2084 | 1975 | 2154 |
| TNFRSF10A - WNT7B | 1084 | 1341 | 2119 | TNFRSF10A - WNT7B | 1301 | 1120 | 1663 |
| TNFRSF10B - WNT7B | 1274 | 1980 | 744 | TNFRSF10B - WNT7B | 1209 | 908 | 1075 |
| TNFRSF10D - WNT7B | 1314 | 774 | 1928 | TNFRSF10D - WNT7B | 1252 | 2301 | 1250 |
| TNFRSF12A - WNT7B | 2100 | 1332 | 1983 | TNFRSF12A - WNT7B | 1104 | 22 | 1879 |
| TNFRSF14 - WNT7B | 1576 | 981 | 1811 | TNFRSF14 - WNT7B | 2079 | 1028 | 1928 |
| TNFRSF21 - WNT7B | 1565 | 798 | 720 | TNFRSF21 - WNT7B | 2114 | 1219 | 737 |
| TNFSF10 - WNT7B | 1598 | 2462 | 2179 | TNFSF10 - WNT7B | 2129 | 763 | 204 |
| TNFSF15 - WNT7B | 1026 | 756 | 621 | TNFSF15 - WNT7B | 130 | 1599 | 2504 |
| Ranking of TNF family w.r.t WNT9A | Ranking of WNT9A w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| TNF - WNT9A | 1624 | 2025 | 799 | TNF - WNT9A | 1121 | 1930 | 1400 |
| TNFAIP1 - WNT9A | 1433 | 839 | 465 | TNFAIP1 - WNT9A | 1254 | 569 | 394 |
| TNFAIP2 - WNT9A | 40 | 2167 | 397 | TNFAIP2 - WNT9A | 2125 | 2437 | 1605 |
| TNFAIP3 - WNT9A | 1427 | 1109 | 2040 | TNFAIP3 - WNT9A | 1764 | 2460 | 1032 |
| TNFRSF1A - WNT9A | 1470 | 719 | 1933 | TNFRSF1A - WNT9A | 1645 | 58 | 1419 |
| TNFRSF10A - WNT9A | 2272 | 1234 | 918 | TNFRSF10A - WNT9A | 2259 | 2413 | 1204 |
| TNFRSF10B - WNT9A | 2249 | 1222 | 1071 | TNFRSF10B - WNT9A | 882 | 566 | 813 |
| TNFRSF10D - WNT9A | 410 | 2132 | 968 | TNFRSF10D - WNT9A | 1808 | 1055 | 568 |
| TNFRSF12A - WNT9A | 1080 | 373 | 1120 | TNFRSF12A - WNT9A | 2345 | 1211 | 2466 |
| TNFRSF14 - WNT9A | 1106 | 2166 | 198 | TNFRSF14 - WNT9A | 1127 | 1147 | 1191 |
| TNFRSF21 - WNT9A | 1805 | 1999 | 986 | TNFRSF21 - WNT9A | 1265 | 832 | 1098 |
| TNFSF10 - WNT9A | 1258 | 864 | 1839 | TNFSF10 - WNT9A | 2054 | 2338 | 1523 |
| TNFSF15 - WNT9A | 1621 | 1129 | 1139 | TNFSF15 - WNT9A | 37 | 1105 | 1076 |
Table 41 shows the derived influences which can be represented graphically, with the following influences - • TNF w.r.t WNT with TNF-RSF1A/RSF10A/RSF10B/SF15 <- WNT2B; TNF-RSF10A/RSF10D/RSF12A/SF10 <- WNT4; TNF-RSF12A/SF10 <- WNT7B and TNF-RSF21 <- WNT9A; and • WNT w.r.t TNF with TNF-RSF10B/RSF10D/RSF14 -> WNT2B; TNF-AIP3/RSF10B -> WNT4; TNF, TNF-RSF1A/RSF14 -> WNT7B; and TNF-AIP2/AIP3/RSF10A/RSF12A/SF10 -> WNT9A.
3.3.2. MUC - TNF Cross Family Analysis
In a recent development in Sheng et al. [81] MUC13 promoted tumor necrosis factro (TNF)-induced NFkB activation by interacting with TNFR1 and the E3 ligase, cIAP1, to increase ubiquitination of RIPK1. Dharmani et al. [107] show that TNF-α and MUC2 (Mucin 2) play major roles in disease onset and progression in dextran sodium sulphate-induced colitis. TNF-α is also shown to induce mucin hypersecretion and MUC2 gene expression by human airway epithelial cells by Levine et al. [108]. Also, inhibition of TNF-α induced MUC5AC expression and production by wogonin through the inactivation of NF-κB signaling in airway epithelial cells, as shown by Sikder et al. [109]. Similarly, neutrophil elastase induces MUC5AC production in human airway epithelial cells via a cascade involving protein kinase-C, reactive oxygen species, and TNF-α-converting enzyme, as shown by Shao and Nadel [110]. TNF-α or transforming growth factor-α stimulation of human epithelial cells resulted in mucus secretion as measured by MUC5AC mRNA and protein (Lora et al. [111]). In earlier experiments by Fischer et al. [112], TNF-α was found to stimulate mucin secretion and cyclic GMP production by guinea pig tracheal epithelial cells in vitro. Similar earlier experiments by Lin et al. [113], induction of mucin gene expression in middle ear of rats by TNF-α was the potential cause for mucoid otitis media. Effects of TNF-α and IL-1β on mucin, lysozyme, IL-6 and IL-8 in passage-2 normal human nasal epithelial cells have been stuided by Yoon et al. [114]. Also, Mercogliano et al. [115] show that TNF-α induced MUC4 expression elicits trastuzumab resistance in HER2-+ive breast cancer. These findings suggest deep synergy between Mucin family and TNF family members. However, not all synergies might have been explored till now. A set of family members of MUC and TNFs were found to be UP regulated after ETC-1922159 treatment in CRC cells.
Table 42 and Table 43 show the additional range of TNFs and MUCs that might be engaged in CRC through the NFkB pathway, in the light of the recent findings of MUC13 and TNFRSF1A in Sheng et al. [81]. Table 42 shows the rankings of the TNF family w.r.t to MUCIN family and Table 43 shows the rankings of the MUCIN family w.r.t to TNF family. Followed by this are the derived influences from the majority votings of the rankings in the foregoing tables, which are depicted in Table 44.
Considering Table 42, TNF family w.r.t MUC1, we find TNFAIP3, TNFRSF-10D/12A/14 to be highly up regulated. These are reflected in the rankings of 2115 (laplace) and 1882 (rbf) for MUC1 - TNFAIP3; 2303 (laplace) and 2154 (linear) for MUC1 - TNFRSF10D; 2019 (laplace) and 2009 (linear) for MUC1 - TNFRSF12A; and 1955 (laplace) and 1899 (linear) for MUC1 - TNFRSF14. TNF family w.r.t MUC3A, we find TNFRSF-10A/10D to be highly up regulated. These are reflected in the rankings of 2237 (laplace) and 1910 (linear) for MUC3 - TNFRSF10A; 1678 (laplace) and 2049 (linear) for MUC3 - TNFRSF10D. TNF family w.r.t MUC4 we find TNFRSF10D/TNFSF10 to be highly up regulated. These are reflected in the rankings of 2503 (laplace), 2403 (linear) and 2356 (rbf) for MUC4 - TNFRSF10D and 2134 (laplace) and 1957 (linear) for MUC4 - TNFSF10. TNF family w.r.t MUC12 we find TNFRSF21/TNFSF10 to be highly up regulated. These are reflected in the rankings of 1795 (laplace) and 2438 (linear) for MUC12 - TNFRSF21 and 1795 (linear) 2435 (rbf) for MUC12 - TNFSF10. TNF family w.rt MUC13 we find TNFRSF10A/TNFRSF10D to be highly up regulated. These are reflected in the rankings of 2500 (laplace) and 1844 (rbf) for MUC13 - TNFRSF10A and 2263 (linear) and 2294 (rbf) for MUC13 - TNFRSF10D. TNF family w.r.t MUC17 we find TNFRSF-10A/10D/12A to be highly up regulated. These are reflected in the rankings of 2269 (laplace) 2364 (linear) and 2005 (rbf) for MUC17 - TNFRSF10A; 1798 (laplace) and 2302 (rbf) for MUC17 - TNFRSF10D and 2041 (laplace) and 2303 (linear) for MUC17 - TNFRSF12A. TNF family w.r.t MUC20 we find TNFAIP3/TNFSF15 to be highly up regulated. These are reflected in the rankings of 2205 (laplace) and 2136 (rbf) for MUC20 - TNFAIP3 and 2493 (laplace) and 2108 (rbf) for MUC20 - TNFSF15.
Considering Table 43, MUC1 w.r.t TNF family, we find TNFRSF1A to be highly up regulated. These are reflected in the rankings of 2344 (linear) and 2312 (rbf) for MUC1 - TNFRSF1A. MUC4 w.r.t TNF family, we find TNFAIP2 to be highly up regulated. These are reflected in the rankings of 1875 (laplace) and 1792 (linear) for MUC4 - TNFAIP2. MUC12 w.r.t TNF family we find TNFAIP1/TNFAIP2/TNFRSF21/TNFSF10 to be highly up regulated. These are reflected in the rankings of 2321 (laplace) and 2457 (rbf) for MUC12 - TNFAIP1; 1829 (linear) and 1913 (rbf) for MUC12 - TNFAIP2; 1975 (linear) and 1769 (rbf) for MUC12 - TNFRSF21; 2135 (linear) and 2255 (rbf) for MUC12 - TNFSF10. MUC12 w.r.t TNF family we find TNFRSF21/TNFSF10 to be highly up regulated. These are reflected in the rankings of 1795 (laplace) and 2438 (linear) for MUC12 - TNFRSF21 and 1795 (linear) and 2435 (rbf) for MUC12 - TNFSF10. MUC20 w.r.t TNF family we find TNFAIP1/TNFAIP2 to be highly up regulated. These are reflected in the rankings of 2266 (laplace) and 2057 (rbf) for MUC20 - TNFAIP1 and 2404 (linear) and 2157 (rbf) for MUC20 - TNFAIP2.
One can also interpret the results of the Table 44 graphically, with the following influences - • TNF family w.r.t MUC family with MUC1 -> TNFAIP3/TNFRSF-10D/12A/14; MUC3A -> TNFRSF-10A/10D; MUC4 -> TNFRSF10D/TNFSF10; MUC12 -> TNFRSF21/TNFSF10; MUC13 -> TNFRSF-10A/10D; MUC17 -> TNFRSF-10A/10D/12A; MUC20 -> TNFAIP3/TNFSF15 and • MUC family w.r.t TNF family with MUC1 <- TNFRSF1A; MUC4 <- TNFAIP2; MUC12 <- TNFAIP1/TNFAIP2/TNFRSF21/ TNFSF10 and MUC13 <- TNFAIP1/TNFAIP2.
3.3.3. STEAP4 - TNF Cross Family Analysis
STEAP4 or six transmembrane epithelial antigen of prostate 4, resides in the golgi apparatus and functions as a metalloreductase with the capacity to reduce insoluble ferric ions Fe3+ to soluble ferrous ions Fe2+. Emerging role of STEAP4 in metabolism and homeostasis of cellular iron and copper in metabolism and homeostasis of cellular iron and copper has been studied in Scarl et al. [116]. STEAP4 was first identified as a novel gene induced by TNF-α during adipose differentiation by Moldes et al. [117]. Zhang et al. [118] observe that STEAP4 was up-regulated by LPS at a very early time point, consistent with reports that STEAP4 could be up-regulated by tumor necrosis factor-alpha. Tanaka et al. [119] show that STEAP4 is expressed on monocytes/neutrophils, and is regulated by TNF antagonist in patients with rheumatoid arthritis. Also, Tanaka et al. [120] show STEAP4 is a tumor necrosis factor alpha-induced protein that regulates IL-6, IL-8, and cell proliferation in synovium from patients with rheumatoid arthritis. Gauss et al. [121] observe that the STEAP4 expression in adipocytes is normally induced by nutritional stress, leptin, and proinflammatory cytokines, including TNF-α, interleukin-1β, and interleukin-6. ZHANG et al. [122] show that the downregulation of STEAP4, a highly-expressed TNF-α-inducible gene in adipose tissue, is associated with obesity in humans 1. Liang et al. [123] show that STEAP comprises a novel inflammatory nexus in patients with pustular skin disorders. They show that in primary human keratinocytes STEAP4 expression was induced by TNF-α, IL-1β, IL-36α, IL-36γ, IL-17A, and IL-17A combined with TNF-α or IL-22. Gomes et al. [124] further show the TNF STEAP interactions while studying the structure of STEAP proteins and its applications to cancer therapy. Such interactions point to the existing synergy between STEAP4 and TNF-α. In CRC cells treated with ETC-1922159, both TNF members and STEAP4 were found to be up regulated. Our search engine alloted the dual combinations with numerically high ranked values thus pointing to the possible synergies that might be existing in the cells and may not have been explored. Table 45 shows the rankings of each with the other. On the left we found, TNF, TNF-AIP1/AIP2/AIP3/RSF10B/RSF10D/SF10 to be up regulated w.r.t STEAP4. These are reflected in rankings of 1914 (linear) and 2130 (rbf) for TNF - STEAP4; 2189 (laplace) and 1910 (rbf) for TNFAIP1 - STEAP4; 2002 (linear) and 1840 (rbf) for TNFAIP2 - STEAP4; 1882 (linear) and 2197 (rbf) for TNFAIP3 - STEAP4; 2210 (laplace), 1717 (linear) and 1827 (rbf) for TNFRSF10B - STEAP4; 2192 (linear) and 1797 (rbf) for TNFRSF10D - STEAP4; and 2083 (laplace) and 1773 (rbf) for TNFSF10 - STEAP4. On the right we found, STEAP4 to be up regulated w.r.t TNF-RSF10A/RSF10D/RSF14. These are reflected in rankings of 1796 (linear) and 2026 (rbf) for TNFRSF10A - STEAP4; 2339 (linear) and 2405 (rbf) for TNFRSF10D - STEAP4; and 1956 (linear) and 2256 (rbf) for TNFRSF14 - STEAP4.
One can also interpret the results of the Table 44 graphically, with the following influences - • TNF w.r.t STEAP4 with TNF, TNF-AIP1/AIP2/AIP3/RSF10B/RSF10D/SF10 <- STEAP4 and • STEAP4 w.r.t TNF with TNF-RSF10A/RSF10D/RSF14 -> STEAP4.
Table 46.
2nd order combinatorial hypotheses between TNF and STEAP4 family.
| Unexplored combinatorial hypotheses | |
| TNF w.r.t STEAP4 | |
| TNF, TNF-AIP1/AIP2/AIP3/RSF10B/RSF10D/SF10 | STEAP4 |
| STEAP4 w.r.t TNF | |
| TNF-RSF10A/RSF10D/RSF14 | STEAP4 |
3.3.4. TNF - UBE2 Cross Family Analysis
Fu et al. [125] show that the ubiquitin conjugating enzyme UBE2L3 regulates TNFα-induced linear ubiquitination. They show by western blotting of HOIL-1L immunoprecipitates demonstrates that endogenous HOIL-1L interacts with endogenous UBE2L3 in vivo and these associations are stable following TNFα stimulation. Through various hypotheses, the authors show the interaction of UBE2L3 with TNF. In conclusion, the authours state that increased UBE2L3 expression enhances NF-κB activation, and increased levels of NF-κB activity are linked to inflammatory and autoimmune diseases. Li et al. [126] show that TNF-α increases ubiquitin-conjugating activity in skeletal muscle by up-regulating UBCH2/E220k. Shembade et al. [127] show that IL-1β or TNF induce late depletion of UBE2D3 (UBCH5C) and UBE2N (UBC13) in mouse embryonic fibroblasts. These studies show a definite synergy between UBE family and TNFs. In CRC cells treated with ETC-1922159, both TNF members and UBE2 were found to be up regulated. Our search engine alloted the dual combinations with numerically high ranked values thus pointing to the possible synergies that might be existing in the cells and may not have been explored. Table 47 and Table 48 shows the rankings of each with the other.
Table 47.
2nd order combinatorial hypotheses between UBE2 and TNF
| Ranking TNF family vs UBE2 family | |||||||
| Ranking of UBE2A w.r.t TNF family | Ranking of TNF family w.r.t UBE2A | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| TNF - UBE2A | 1360 | 2307 | 1720 | TNF - UBE2A | 499 | 1379 | 750 |
| TNFAIP1 - UBE2A | 498 | 2357 | 2455 | TNFAIP1 - UBE2A | 1340 | 2494 | 578 |
| TNFAIP2 - UBE2A | 524 | 1161 | 2385 | TNFAIP2 - UBE2A | 441 | 1852 | 691 |
| TNFAIP3 - UBE2A | 855 | 1642 | 812 | TNFAIP3 - UBE2A | 1157 | 1048 | 207 |
| TNFRSF1A - UBE2A | 2457 | 1087 | 2020 | TNFRSF1A - UBE2A | 1066 | 655 | 1701 |
| TNFRSF10A - UBE2A | 2164 | 2126 | 621 | TNFRSF10A - UBE2A | 2116 | 858 | 2376 |
| TNFRSF10B - UBE2A | 2284 | 1901 | 1203 | TNFRSF10B - UBE2A | 362 | 1083 | 756 |
| TNFRSF10D - UBE2A | 1989 | 2291 | 677 | TNFRSF10D - UBE2A | 1848 | 1336 | 903 |
| TNFRSF12A - UBE2A | 2484 | 2427 | 339 | TNFRSF12A - UBE2A | 1537 | 1304 | 629 |
| TNFRSF14 - UBE2A | 2301 | 2180 | 2323 | TNFRSF14 - UBE2A | 908 | 1519 | 1945 |
| TNFRSF21 - UBE2A | 2419 | 2035 | 1169 | TNFRSF21 - UBE2A | 605 | 2245 | 60 |
| TNFSF10 - UBE2A | 832 | 2202 | 1036 | TNFSF10 - UBE2A | 1520 | 44 | 2125 |
| TNFSF15 - UBE2A | 1768 | 1184 | 1942 | TNFSF15 - UBE2A | 545 | 580 | 1448 |
| Ranking of UBE2B w.r.t TNF family | Ranking of TNF family w.r.t UBE2B | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| TNF - UBE2B | 1072 | 2046 | 1316 | TNF - UBE2B | 1719 | 218 | 346 |
| TNFAIP1 - UBE2B | 1097 | 744 | 1295 | TNFAIP1 - UBE2B | 920 | 90 | 1028 |
| TNFAIP2 - UBE2B | 669 | 1158 | 2407 | TNFAIP2 - UBE2B | 1680 | 147 | 45 |
| TNFAIP3 - UBE2B | 470 | 1528 | 1388 | TNFAIP3 - UBE2B | 2259 | 742 | 1610 |
| TNFRSF1A - UBE2B | 937 | 1473 | 2390 | TNFRSF1A - UBE2B | 1277 | 1454 | 1258 |
| TNFRSF10A - UBE2B | 2132 | 1128 | 2184 | TNFRSF10A - UBE2B | 551 | 2318 | 2265 |
| TNFRSF10B - UBE2B | 2399 | 2000 | 402 | TNFRSF10B - UBE2B | 2272 | 1268 | 1080 |
| TNFRSF10D - UBE2B | 1959 | 1562 | 2232 | TNFRSF10D - UBE2B | 1157 | 207 | 1729 |
| TNFRSF12A - UBE2B | 1632 | 12 | 2259 | TNFRSF12A - UBE2B | 1940 | 1868 | 1758 |
| TNFRSF14 - UBE2B | 1137 | 2297 | 2373 | TNFRSF14 - UBE2B | 1143 | 1657 | 1507 |
| TNFRSF21 - UBE2B | 1986 | 1439 | 1754 | TNFRSF21 - UBE2B | 1291 | 569 | 17 |
| TNFSF10 - UBE2B | 2265 | 1488 | 769 | TNFSF10 - UBE2B | 2208 | 2326 | 2470 |
| TNFSF15 - UBE2B | 1432 | 2460 | 1655 | TNFSF15 - UBE2B | 2055 | 1964 | 183 |
| Ranking of UBE2F w.r.t TNF family | Ranking of TNF family w.r.t UBE2F | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| TNF - UBE2F | 2162 | 2484 | 2500 | TNF - UBE2F | 638 | 435 | 1471 |
| TNFAIP1 - UBE2F | 1732 | 2239 | 2003 | TNFAIP1 - UBE2F | 447 | 1376 | 1357 |
| TNFAIP2 - UBE2F | 693 | 1446 | 1706 | TNFAIP2 - UBE2F | 900 | 208 | 883 |
| TNFAIP3 - UBE2F | 498 | 2265 | 1264 | TNFAIP3 - UBE2F | 1881 | 113 | 1185 |
| TNFRSF1A - UBE2F | 1980 | 2255 | 1872 | TNFRSF1A - UBE2F | 368 | 1756 | 266 |
| TNFRSF10A - UBE2F | 2085 | 2218 | 179 | TNFRSF10A - UBE2F | 1767 | 1599 | 781 |
| TNFRSF10B - UBE2F | 2432 | 2011 | 2144 | TNFRSF10B - UBE2F | 1413 | 1157 | 1510 |
| TNFRSF10D - UBE2F | 1164 | 1400 | 2150 | TNFRSF10D - UBE2F | 389 | 206 | 2481 |
| TNFRSF12A - UBE2F | 2458 | 2336 | 531 | TNFRSF12A - UBE2F | 581 | 2022 | 630 |
| TNFRSF14 - UBE2F | 1757 | 574 | 1070 | TNFRSF14 - UBE2F | 2324 | 1924 | 1954 |
| TNFRSF21 - UBE2F | 1056 | 2498 | 1418 | TNFRSF21 - UBE2F | 718 | 2123 | 1022 |
| TNFSF10 - UBE2F | 1710 | 2365 | 1691 | TNFSF10 - UBE2F | 1656 | 1584 | 810 |
| TNFSF15 - UBE2F | 1910 | 1171 | 2353 | TNFSF15 - UBE2F | 1224 | 1637 | 394 |
Table 48.
2nd order combinatorial hypotheses between UBE2 and TNF
| Ranking TNF family vs UBE2 family | |||||||
| Ranking of UBE2H w.r.t TNF family | Ranking of TNF family w.r.t UBE2H | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| TNF - UBE2H | 967 | 1966 | 1018 | TNF - UBE2H | 2277 | 770 | 640 |
| TNFAIP1 - UBE2H | 1235 | 1484 | 817 | TNFAIP1 - UBE2H | 883 | 2396 | 608 |
| TNFAIP2 - UBE2H | 1251 | 978 | 2517 | TNFAIP2 - UBE2H | 762 | 1362 | 593 |
| TNFAIP3 - UBE2H | 889 | 1055 | 1837 | TNFAIP3 - UBE2H | 1942 | 1421 | 1467 |
| TNFRSF1A - UBE2H | 589 | 1498 | 1428 | TNFRSF1A - UBE2H | 1134 | 2154 | 182 |
| TNFRSF10A - UBE2H | 1317 | 905 | 2229 | TNFRSF10A - UBE2H | 1139 | 202 | 1061 |
| TNFRSF10B - UBE2H | 33 | 2128 | 1725 | TNFRSF10B - UBE2H | 1053 | 539 | 1207 |
| TNFRSF10D - UBE2H | 1326 | 1814 | 1657 | TNFRSF10D - UBE2H | 2227 | 926 | 791 |
| TNFRSF12A - UBE2H | 1950 | 1793 | 1851 | TNFRSF12A - UBE2H | 1347 | 776 | 1899 |
| TNFRSF14 - UBE2H | 2297 | 1601 | 2385 | TNFRSF14 - UBE2H | 2244 | 703 | 1208 |
| TNFRSF21 - UBE2H | 2022 | 1131 | 2231 | TNFRSF21 - UBE2H | 827 | 880 | 672 |
| TNFSF10 - UBE2H | 2387 | 7 | 760 | TNFSF10 - UBE2H | 1313 | 1169 | 2002 |
| TNFSF15 - UBE2H | 125 | 58 | 96 | TNFSF15 - UBE2H | 350 | 2416 | 1960 |
| Ranking of UBE2J1 w.r.t TNF family | Ranking of TNF family w.r.t UBE2J1 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| TNF - UBE2J1 | 1289 | 2308 | 2336 | TNF - UBE2J1 | 1101 | 1549 | 105 |
| TNFAIP1 - UBE2J1 | 1109 | 2292 | 1756 | TNFAIP1 - UBE2J1 | 329 | 1971 | 252 |
| TNFAIP2 - UBE2J1 | 1379 | 1516 | 1696 | TNFAIP2 - UBE2J1 | 112 | 22 | 969 |
| TNFAIP3 - UBE2J1 | 187 | 1261 | 1065 | TNFAIP3 - UBE2J1 | 289 | 891 | 1202 |
| TNFRSF1A - UBE2J1 | 1992 | 326 | 2268 | TNFRSF1A - UBE2J1 | 1422 | 624 | 73 |
| TNFRSF10A - UBE2J1 | 1893 | 2090 | 2363 | TNFRSF10A - UBE2J1 | 2379 | 2213 | 2135 |
| TNFRSF10B - UBE2J1 | 1913 | 1299 | 1838 | TNFRSF10B - UBE2J1 | 807 | 1793 | 1231 |
| TNFRSF10D - UBE2J1 | 325 | 1500 | 588 | TNFRSF10D - UBE2J1 | 2393 | 1360 | 2102 |
| TNFRSF12A - UBE2J1 | 2401 | 1901 | 437 | TNFRSF12A - UBE2J1 | 380 | 1284 | 650 |
| TNFRSF14 - UBE2J1 | 2277 | 2347 | 1943 | TNFRSF14 - UBE2J1 | 1614 | 2133 | 2313 |
| TNFRSF21 - UBE2J1 | 1976 | 2333 | 1681 | TNFRSF21 - UBE2J1 | 1315 | 1266 | 1070 |
| TNFSF10 - UBE2J1 | 511 | 2508 | 506 | TNFSF10 - UBE2J1 | 1322 | 203 | 1148 |
| TNFSF15 - UBE2J1 | 2021 | 2013 | 2515 | TNFSF15 - UBE2J1 | 678 | 886 | 1128 |
| Ranking of UBE2Z w.r.t TNF family | Ranking of TNF family w.r.t UBE2Z | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| TNF - UBE2Z | 2264 | 2505 | 2479 | TNF - UBE2Z | 739 | 701 | 1241 |
| TNFAIP1 - UBE2Z | 1283 | 2055 | 2332 | TNFAIP1 - UBE2Z | 1198 | 1213 | 226 |
| TNFAIP2 - UBE2Z | 2404 | 1625 | 2139 | TNFAIP2 - UBE2Z | 1281 | 1431 | 492 |
| TNFAIP3 - UBE2Z | 1066 | 1152 | 1627 | TNFAIP3 - UBE2Z | 530 | 51 | 972 |
| TNFRSF1A - UBE2Z | 2473 | 2194 | 2405 | TNFRSF1A - UBE2Z | 692 | 43 | 1382 |
| TNFRSF10A - UBE2Z | 2234 | 2152 | 713 | TNFRSF10A - UBE2Z | 2410 | 2103 | 1513 |
| TNFRSF10B - UBE2Z | 1501 | 451 | 2081 | TNFRSF10B - UBE2Z | 948 | 1369 | 403 |
| TNFRSF10D - UBE2Z | 2264 | 2360 | 2278 | TNFRSF10D - UBE2Z | 1786 | 661 | 1746 |
| TNFRSF12A - UBE2Z | 2207 | 2149 | 353 | TNFRSF12A - UBE2Z | 1621 | 2010 | 1448 |
| TNFRSF14 - UBE2Z | 1683 | 1983 | 705 | TNFRSF14 - UBE2Z | 1779 | 1360 | 2100 |
| TNFRSF21 - UBE2Z | 994 | 604 | 219 | TNFRSF21 - UBE2Z | 459 | 1030 | 584 |
| TNFSF10 - UBE2Z | 516 | 2374 | 2235 | TNFSF10 - UBE2Z | 1100 | 2047 | 168 |
| TNFSF15 - UBE2Z | 2081 | 1037 | 2102 | TNFSF15 - UBE2Z | 1342 | 1180 | 536 |
Table 49.
2nd order combinatorial hypotheses between TNF and UBE2 family.
| Unexplored combinatorial hypotheses | |
| UBE2 w.r.t TNF | |
| TNF-AIP1/RSF1A/RSF10A/RSF10B/RSF10D/RSF12A/RSF14/RSF21/SF15 | UBE2A |
| TNF-RSF10A/RSF10B/RSF10D/RSF14/RSF21 | UBE2B |
| TNF, TNF-AIP1/RSF1A/RSF10A/RSF10B/RSF12A/SF15 | UBE2F |
| TNF-RSF12A/RSF14/RSF21 | UBE2H |
| TNF, TNF-AIP1/RSF1A/RSF10A/RSF10B/RSF12A/RSF14/RSF21/SF15 | UBE2J1 |
| TNF, TNF-AIP1/AIP2/RSF1A/RSF10A/RSF10D/RSF12A/SF10/SF15 | UBE2Z |
| TNF w.r.t UBE2 | |
| TNF-RSF10A | UBE2A |
| TNF-RSF10A/RSF12A/SF10/SF15 | UBE2B |
| TNF-RSF14 | UBE2F |
| TNF-SF15 | UBE2H |
| TNF-RSF10A/RSF10D/RSF14 | UBE2J1 |
| TNF-RSF10A/RSF14 | UBE2Z |
On the left side is the ranking of UBE2 family w.r.t TNF family. We found UBE2A to be up regulated w.r.t TNF-AIP1/RSF1A/RSF10A/RSF10B/RSF10D/RSF12A/RSF14/ RSF21/SF15. These are reflected in rankings of 2357 (linear) and 2455 (rbf) for TNFAIP1 - UBE2A; 2457 (laplace) and 2020 (rbf) for TNFRSF1A - UBE2A; 2164 (laplace) and 2126 (linear) for TNFRSF10A - UBE2A; 2284 (laplace) and 1901 (linear) for TNFRSF10B - UBE2A; 1989 (laplace) and 2291 (linear) for TNFRSF10D - UBE2A; 2484 (laplace) and 2427 (linear) for TNFRSF12A - UBE2A; 2301 (laplace), 2180 (linear) and 2323 (rbf) for TNFRSF14 - UBE2A; 2419 (laplace) and 2035 (linear) for TNFRSF21 - UBE2A; 1768 (laplace) and 1942 (rbf) for TNFSF15 - UBE2A. UBE2B to be up regulated w.r.t TNF-RSF10A/RSF10B/RSF10D/RSF14/RSF21. These are reflected in rankings of 2132 (laplace) and 2184 (rbf) for TNFRSF10A - UBE2B; 2399 (laplace) and 2000 (linear) for TNFRSF10B - UBE2B; 1959 (laplace) and 2232 (rbf) for TNFRSF10D - UBE2B; 2297 (linear) and 2373 (rbf) for TNFRSF14 - UBE2B; and 1986 (laplace) and 1754 (rbf) for TNFRSF21 - UBE2B. UBE2F to be up regulated w.r.t TNF, TNF-AIP1/RSF1A/RSF10A/RSF10B/RSF12A/SF15. These are reflected in rankings of 2162 (laplace), 2484 (linear) and 2500 (rbf) for TNF - UBE2F; 1732 (laplace), 2239 (linear) and 2003 (rbf) for TNFAIP1 - UBE2F; 1980 (laplace), 2255 (linear) and 1872 (rbf) for TNFRSF1A - UBE2F; 2085 (laplace), 2218 (linear) for TNFRSF10A - UBE2F; 2432 (laplace), 2011 (linear) and 2144 (rbf) for TNFRSF10B - UBE2F; 2458 (laplace) and 2336 (linear) for TNFRSF12A - UBE2F; 1910 (laplace) and 2353 (rbf) for TNFSF15 - UBE2F; UBE2H to be up regulated w.r.t TNF-RSF12A/RSF14/RSF21. These are reflected in rankings of 1950 (laplace), 1793 (linear) and 1851 (rbf) for TNFRSF12A - UBE2H; 2297 (laplace) and 2385 (rbf) for TNFRSF14 - UBE2H; and 2022 (laplace) and 2231 (rbf) for TNFRSF21 - UBE2H; UBE2J1 to be up regulated w.r.t TNF, TNF-AIP1/RSF1A/RSF10A/RSF10B/RSF12A/RSF14/RSF21/SF15. These are reflected in rankings of 2308 (linear) and 2336 (rbf) for TNF - UBE2J1; 2292 (linear) and 1756 (rbf) for TNFAIP1 - UBE2J1; 1992 (laplace) and 2268 (rbf) for TNFRSF1A - UBE2J1; 1893 (laplace), 2090 (linear) and 2363 (rbf) for TNFRSF10A - UBE2J1; 1913 (laplace) and 1838 (rbf) for TNFRSF10B - UBE2J1; 2401 (laplace) and 1901 (linear) for TNFRSF12A - UBE2J1; 2277 (laplace), 2347 (linear) and 1943 (rbf) for TNFRSF14 - UBE2J1; 1976 (laplace), 2333 (linear) for TNFRSF21 - UBE2J1; and 2021 (laplace), 2013 (linear) and 2515 (rbf) for TNFSF15 - UBE2J1; UBE2Z to be up regulated w.r.t TNF, TNF-AIP1/AIP2/RSF1A/RSF10A/RSF10D/RSF12A/SF10/SF15. These are reflected in rankings of 2264 (laplace), 2505 (linear) and 2479 (rbf) for TNF - UBE2Z; 2055 (linear) and 2332 (rbf) for TNFAIP1 - UBE2Z; 2404 (laplace) and 2139 (rbf) for TNFAIP2 - UBE2Z; 2473 (laplace), 2194 (linear) and 2405 (rbf) for TNFRSF1A - UBE2Z; 2234 (laplace) and 2152 (linear) for TNFRSF10A - UBE2Z; 2264 (laplace), 2360 (linear) and 2278 (rbf) for TNFRSF10D - UBE2Z; 2207 (laplace) and 2149 (linear) for TNFRSF12A - UBE2Z; 2374 (linear) and 2235 (rbf) for TNFSF10 - UBE2Z; and 2081 (laplace) and 2102 (rbf) for TNFSF15 - UBE2Z;
One the right side is the ranking of TNF family w.r.t UBE2 family. We found TNF-RSF10A to be up regulated w.r.t UBE2A. This is reflected in rankings of 2116 (laplace) and 2376 (rbf) for TNFRSF10A - UBE2A. TNF-RSF10A/RSF12A/SF10/SF15 were up regulated w.r.t UBE2B. These are reflected in rankings of 2318 (linear) and 2265 (linear) for TNFRSF10A - UBE2B; 1940 (laplace); 1868 (linear) and 1758 (linear) for TNFRSF12A - UBE2B; 2208 (laplace); 2326 (linear) and 2470 (linear) for TNFSF10 - UBE2B; and 2055 (laplace) and 1964 (linear) for TNFSF15 - UBE2B. TNF-RSF14 were up regulated w.r.t UBE2F. These is reflected in rankings of 2324 (laplace) and 1924 (linear) for TNF-RSF14 - UBE2F. TNF-SF15 were up regulated w.r.t UBE2H. These is reflected in rankings of 2416 (linear) and 1960 (rbf) for TNF-SF15 - UBE2H. TNF-RSF10A/RSF10D/RSF14 were up regulated w.r.t UBE2J1. These are reflected in rankings of 2379 (laplace), 2213 (linear) and 2135 (rbf) for TNFRSF10A - UBE2J1; 2393 (laplace) and 2102 (rbf) for TNFRSF10D - UBE2J1; and 2133 (linear) and 2313 (rbf) for TNFRSF14 - UBE2J1. TNF-RSF10A/RSF14 were up regulated w.r.t UBE2Z. These are reflected in rankings of 2410 (laplace) and 2103 (laplace) for TNFRSF10A - UBE2Z and 1779 (laplace) and 2100 (rbf) for TNFRSF14 - UBE2Z.
One can also interpret the results of the Table 49 graphically, with the following influences - • UBE2 w.r.t TNF with TNF-AIP1/RSF1A/RSF10A/RSF10B/RSF10D/RSF12A/RSF14/RSF21/SF15 -> UBE2A; TNF-RSF10A/RSF10B/RSF10D/RSF14/RSF21 -> UBE2B; TNF, TNF-AIP1/RSF1A/RSF10A/RSF10B/RSF12A/SF15 -> UBE2F; TNF-RSF12A/RSF14/RSF21 -> UBE2H; TNF, TNF-AIP1/RSF1A/RSF10A/RSF10B/RSF12A/RSF14/RSF21/SF15 -> UBE2J1; and TNF, TNF-AIP1/AIP2/RSF1A/RSF10A/RSF10D/RSF12A/SF10/SF15 -> UBE2Z • TNF w.r.t UBE2 with TNF-RSF10A <- UBE2A; TNF-RSF10A/RSF12A/SF10/SF15 <- UBE2B; TNF-RSF14 <- UBE2F; TNF-SF15 <- UBE2H; TNF-RSF10A/RSF10D/RSF14 <- UBE2J1; TNF-RSF10A/RSF14 <- UBE2Z.
3.3.5. TNF - BCL Cross Family Analysis
Tamatani et al. [128] observe that tumor necrosis factor induces BCL-2 and BCL-x expression through NFκB activation in primary hippocampal neurons. The role of Bcl-2 Expression in EGF Inhibition of TNF-α/IFN-γ-induced Villous Trophoblast Apoptosis has been studied by Ho et al. [129]. Genestier et al. [130] show that tumor necrosis factor-α up-regulates BCL-2 expression and decreases calcium-dependent apoptosis in human B cell lines. In breast carcinoma cells, Bcl-x and Bcl-2 inhibit TNF and FAS-induced apoptosis and activation of phospholipase A2 (Jäättelä et al. [131]). Kim et al. [132] show that TNF-α-induced ROS production triggering apoptosis is directly linked to Romo1 and BCL-XL. Kuwata et al. [133] showed that IL-10-inducible BCL-3 negatively regulates LPS-induced TNF-α production in macrophages. Esche et al. [134] showed that tumor necrosis factor-α-promoted expression of BCL-2 and inhibition of mitochondrial cytochrome c release mediate resistance of mature dendritic cells to melanoma-induced apoptosis. These studies show a definite synergy between BCL family and TNFs. In CRC cells treated with ETC-1922159, both TNF members and BCL were found to be up regulated. Our search engine alloted the dual combinations with numerically high ranked values thus pointing to the possible synergies that might be existing in the cells and may not have been explored. Table 50 and Table 51 show the rankings of each with the other.
On the left side is the ranking of BCL family w.r.t TNF family. We found BCL2L2 to be up regulated w.r.t TNF, TNF-AIP1/RSF1A/RSF10B/RSF10D/RSF12A/RSF14/RSF21/SF10/SF15. These are reflected in rankings of 1822 (laplace), 1926 (linear) and 2359 (rbf) for TNF - BCL2L2; 2266 (laplace), 2478 (linear) and 1847 (rbf) for TNFAIP1 - BCL2L2; 2311 (linear) and 1920 (rbf) for TNFRSF1A - BCL2L2; 2478 (laplace) and 2239 (rbf) for TNFRSF10B - BCL2L2; 2278 (linear) and 2237 (rbf) for TNFRSF10D - BCL2L2; 1945 (laplace) and 2484 (rbf) for TNFRSF12A - BCL2L2; 2358 (laplace) and 2310 (rbf) for TNFRSF14 - BCL2L2; 2292 (laplace) and 1850 (linear) for TNFRSF21 - BCL2L2; 2438 (laplace) and 2013 (rbf) for TNFSF10 - BCL2L2 and 2443 (linear) and 2350 (rbf) for TNFSF15 - BCL2L2; BCL2L3 was up regulated w.r.t TNF, TNF-AIP1/RSF1A/RSF10A/RSF10D/RSF12A/RSF14/RSF21/SF10/SF15. These are reflected in rankings of 2437 (laplace), 2482 (linear) and 2482 (rbf) for TNF - BCL2L13; 1863 (laplace) and 2386 (linear) for TNFAIP1 - BCL2L13; 1962 (linear) and 2489 (rbf) for TNFRSF1A - BCL2L13; 2055 (linear) and 2499 (rbf) for TNFRSF10A - BCL2L13; 2204 (laplace), 2159 (linear) and 2343 (rbf) for TNFRSF10D - BCL2L13; 2183 (laplace), 2509 (linear) for TNFRSF12A - BCL2L13; 1852 (laplace), 1974 (linear) and 2339 (rbf) for TNFRSF14 - BCL2L13; 2280 (laplace), 2424 (linear) and 2301 (rbf) for TNFRSF21 - BCL2L13; 2429 (linear) and 1803 (rbf) for TNFSF10 - BCL2L13; and 2438 (linear) and 2252 (rbf) for TNFSF15 - BCL2L13; BCL3 was up regulated w.r.t TNFRSF10B. This is reflected in rankings of 2427 (laplace) and 1868 (rbf). BCL6 was up regulated w.r.t TNF, TNF-AIP1/AIP2/RSF1A/RSF10A/RSF10D/RSF21. These are reflected in rankings of 2271 (laplace), 2071 (linear) and 1810 (rbf) for TNF - BCL6; 2135 (laplace) and 2158 (linear) for TNFAIP1 - BCL6; 2340 (laplace) 1808 (rbf) for TNFAIP2 - BCL6; 1771 (linear) and 2503 (rbf) for TNFRSF1A - BCL6; and 1831 (linear) and 2096 (rbf) for TNFRSF10A - BCL6; and 2213 (laplace) and 2188 (rbf) for TNFRSF10D - BCL6; and 2071 (linear) and 2335 (rbf) for TNFRSF21 - BCL6; BCL10 was up regulated w.r.t TNF-RSF10D/RSF12A. These are reflected in rankings of 1831 (laplace) and 2040 (rbf) for TNFRSF10D - BCL10; and 2015 (laplace) and 1883 (rbf) for TNFRSF12A - BCL10;
On the right side is the ranking of TNF family w.r.t BCL family. We found TNF-RSF10A/RSF10B/RSF10D/RSF12A/RSF14/SF10 to be up regulated w.r.t BCL2L1. These are reflected in rankings of 2273 (laplace) and 1928 (linear) for TNFRSF10A - BCL2L1; 2252 (linear) and 2217 (rbf) for TNFRSF10B - BCL2L1; 1868 (laplace), 2420 (linear) and 2392 (rbf) for TNFRSF10D - BCL2L1; 1923 (laplace) and 1936 (rbf) for TNFRSF12A - BCL2L1; 2350 (linear) and 2414 (rbf) for TNFRSF14 - BCL2L1 and 2115 (laplace) and 2299 (linear) for TNFSF10 - BCL2L1; TNFRSF10B was up regulated w.r.t BCL2L2. This is reflected in rankings of 2153 (laplace) and 1983 (rbf) for TNFRSF10B - BCL2L2; TNFRSF14 was up regulated w.r.t BCL2L13. This is reflected in rankings of 2459 (laplace) and 2381 (linear) for TNFRSF14 - BCL2L13; TNF-RSF10A/SF10D/SF10 were up regulated w.r.t BCL3. These are reflected in rankings of 2388 (laplace) and 2493 (linear) for TNFRSF10A - BCL3; 2213 (laplace) and 1972 (rbf) for TNFRSF10D - BCL3 and 1926 (laplace) and 2107 (rbf) for TNFSF10 - BCL3; TNFRSF12A was up regulated w.r.t BCL6. This is reflected in rankings of 2200 (linear) and 1910 (rbf) for TNFRSF12A - BCL6; TNF-AIP1/SF10/SF15 was up regulated w.r.t BCL9L. This is reflected in rankings of 2470 (linear) and 1802 (rbf) for TNFAIP1 - BCL9L; 1812 (laplace), 1796 (linear) and 2095 (rbf) for TNFSF10 - BCL9L; and 1939 (laplace), 2114 (linear) and 2405 (rbf) for TNFSF15 - BCL9L; TNFRSF14 was up regulated w.r.t BCL10. This is reflected in rankings of 1894 (linear) and 2227 (rbf) for TNFRSF14 - BCL10;
Table 52.
2nd order combinatorial hypotheses between TNF and BCL family.
| Unexplored combinatorial hypotheses | |
| BCL w.r.t TNF | |
| TNF, TNF-AIP1/RSF1A/RSF10B/RSF10D/RSF12A/RSF14/RSF21/SF10/SF15 | BCL2L2 |
| TNF, TNF-AIP1/RSF1A/RSF10A/RSF10D/RSF12A/RSF14/RSF21/SF10/SF15 | BCL2L13 |
| TNFRSF10B | BCL3 |
| TNF, TNF-AIP1/AIP2/RSF1A/RSF10A/RSF10D/RSF21 | BCL6 |
| TNF-RSF10D/RSF12A | BCL10 |
| TNF w.r.t BCL | |
| TNF-RSF10A/RSF10B/RSF10D/RSF12A/RSF14/SF10 | BCL2L1 |
| TNF-RSF10B | BCL2L2 |
| TNF-RSF14 | BCL2L13 |
| TNF-RSF10A/SF10D/SF10 | BCL3 |
| TNF-RSF12A | BCL6 |
| TNF-AIP1 | BCL9L |
| TNF-SF10/SF15 | BCL9L |
| TNF-RSF14 | BCL10 |
One can also interpret the results of the Table 111 graphically, with the following influences - • BCL w.r.t TNF with TNF, TNF-AIP1/RSF1A/RSF10B/RSF10D/RSF12A/RSF14/RSF21/SF10/SF15 -> BCL2L2; TNF, TNF-AIP1/RSF1A/RSF10A/RSF10D/RSF12A/RSF14/RSF21/SF10/SF15 -> BCL2L13; TNFRSF10B -> BCL3; TNF, TNF-AIP1/AIP2/RSF1A/RSF10A/RSF10D/RSF21 -> BCL6; TNF-RSF10D/RSF12A -> BCL10; • TNF w.r.t BCL with TNF-RSF10A/RSF10B/RSF10D/RSF12A/RSF14/SF10 <- BCL2L1; TNF-RSF10B <- BCL2L2; TNF-RSF14 <- BCL2L13; TNF-RSF10A/SF10D/SF10 <- BCL3; TNF-RSF12A <- BCL6; TNF-AIP1 <- BCL9L; TNF-SF10/SF15 <- BCL9L and TNF-RSF14 <- BCL10.
3.4. DNA Repair Related Synergies
3.4.1. XRCC - RAD Cross Family Analysis
X-ray repair cross-complementing protein (XRCC) plays major role in DNA repair process, especially in Double Strand Repair (DBS) Thacker and Zdzienicka [135] and Thacker and Zdzienicka [136]. Sultana et al. [137] observe that ataxia telangiectasia mutated and RAD3 related (ATR) protein kinase inhibition is synthetically lethal in XRCC1 deficient ovarian cancer cells. Della-Maria et al. [138] observe that human Mre11/human RAD50/Nbs1 and DNA ligase IIIα/XRCC1 protein complexes act together in an alternative nonhomologous end joining pathway. These findings along with multiple published work indicate the joint synergy of XRCC - RAD family. In colorectal cancer cell lines treated with ETC-1922159, both XRCC and RAD members were found to be down regulated. The search engine gave the 2nd order synergies between XRCC - RAD families, low numerical valued ranks to signify plausible synergistic down regulations that might not have been explored. Table 53 shows the rankings of RAD family w.r.t XRCC family and Table 54 shows the rankings of the XRCC family w.r.t RAD family. In Table 53 we found RAD-18/51/51AP1/51C/54B/54L to be down regulated w.r.t XRCC1. These are reflected with rankings of 1027 (laplace), 456 (linear) and 1355 (rbf) for RAD-18 - XRCC1; 282 (laplace), 365 (linear) and 1003 (rbf) for RAD51 - XRCC1; 753 (laplace), 5 (linear) and 27 (rbf) for RAD51AP1 - XRCC1; 337 (laplace), 111 (linear) and 968 (rbf) for RAD51C - XRCC1; 175 (laplace), 224 (linear) and 78 (rbf) for RAD54B - XRCC1; and 327 (laplace), 889 (linear) and 709 (rbf) for RAD54L - XRCC1. RAD-18/51/51AP1/51C/54B/54L were also found to be down regulated w.r.t XRCC2. These are reflected in 1388 (laplace), 847 (linear) and 765 (rbf) for XRCC2 - RAD18; 1247 (laplace), 1033 (linear) and 629 (rbf) for XRCC2 - RAD51; 302 (laplace); 247 (linear) and 42 (rbf) for XRCC2 - RAD51AP1; 1079 (laplace), 674 (linear) and 323 (rbf) for XRCC2 - RAD51C; 387 (laplace), 566 (linear) and 506 (rbf) for XRCC2 - RAD54B; and 976 (laplace), 918 (linear) and 847 (rbf) for XRCC2 - RAD54L. RAD-18/51/51AP1/51C/54B/54L were found to be down regulated with w.r.t XRCC6. These are reflected in 541 (laplace), 25 (linear) and 1068 (rbf) for RAD18 - XRCC6; 608 (laplace), 425 (linear) and 900 (rbf) for RAD51 - XRCC6; 216 (laplace), 67 (linear) and 83 (rbf) for RAD51AP1 - XRCC6; 426 (laplace), 865 (linear) and 503 (rbf) for RAD51C - XRCC6; 3 (laplace), 610 (linear) and 112 (rbf) for RAD54B - XRCC6; and 85 (laplace), 252 (linear) and 432 (rbf) for RAD54L - XRCC6. RAD-1/18/50/51/51AP1/51C/54B/54L were found to be down regulated w.r.t XRCC6BP1. These are reflected in 1167 (laplace) and 308 (rbf) for RAD1 - XRCC6BP1; 656 (linear) and 1612 (rbf) for RAD18 - XRCC6BP1; 1302 (laplace) and 328 (rbf) for XRCC6BP1 - RAD50; 435 (laplace), 495 (linear) and 1275 (rbf) for RAD51 - XRCC6BP1; 81 (laplace), 177 (linear) and 73 (rbf) for RAD51AP1 - XRCC6BP1; 645 (laplace), 1366 (linear) and 1414 (rbf) for RAD51C - XRCC6BP1; 154 (laplace), 693 (linear) and 1398 (rbf) for RAD54B - XRCC6BP1; and 420 (linear) and 1060 (rbf) for RAD54L - XRCC6BP1;
In Table 54 we found XRCC-2/6BP1 to be down regulated w.r.t RAD1. These are reflected in 62 (laplace), 498 (linear) and 1231 (rbf) for RAD1 - XRCC2; and 764 (laplace) and 1325 (rbf) for RAD1 - XRCC6BP1. XRCC-1/2/6 were found to be down regulated with w.r.t RAD18. These are reflected in 927 (laplace) and 200 (rbf) for RAD18 - XRCC1; 506 (laplace) and 1517 (rbf) for RAD18 - XRCC2; and 279 (laplace) and 804 (rbf) for RAD18 - XRCC6; XRCC-2/6BP1 were found to be down regulated w.r.t RAD50. These are reflected in rankings of 53 (laplace), 244 (linear) and 147 (rbf) for XRCC-2 - RAD50; and 1375 (linear) and 1366 (rbf) for RAD50 - XRCC6BP1. XRCC-6/6BP1 were found to be down regulated w.r.t RAD51; These are reflected in rankings of 80 (laplace) and 1244 (linear) for XRCC6 - RAD51; and 792 (laplace), 951 (linear) and 1595 (rbf) for XRCC6BP1 - RAD51. XRCC-2/6BP1 were found to be down regulated w.r.t RAD51AP1. These were reflected in 78 (laplace), 112 (linear) and 351 (rbf) for XRCC2 - RAD51AP1; and 936 (linear) and 974 (rbf) for XRCC6BP1 - RAD51AP1; XRCC2 was found to be down regulated w.r.t RAD51C. This are reflected in 1695 (laplace), 932 (linear) and 520 (rbf) for XRCC2 - RAD51C. XRCC2 was found to be down regulated w.r.t RAD54B. This is reflected in rankings of 1554 (laplace), 744 (linear) and 620 (rbf) for XRCC2 - RAD54B. XRCC-1/2/6/6BP1 were found to be down regulated w.r.t RAD54L. These are reflected in rankings of 657 (linear) and 525 (rbf) for XRCC1 - RAD54L; 167 (laplace) and 565 (rbf) for XRCC2 - RAD54L; 496 (linear) and 1247 (rbf) for XRCC6 - RAD54L; and 1389 (laplace), 1227 (linear) and 1454 (rbf) for RAD54L - XRCC6BP1;
Table 54.
2nd order interaction ranking between XRCC w.r.t RAD family members.
| Ranking XRCC family w.r.t RAD family | |||||||
| Ranking of XRCC w.r.t RAD1 | Ranking of XRCC w.r.t RAD18 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| RAD1 - XRCC1 | 1751 | 1808 | 793 | RAD18 - XRCC1 | 927 | 2669 | 200 |
| XRCC2 - RAD1 | 62 | 498 | 1231 | XRCC2 - RAD18 | 506 | 1844 | 1517 |
| XRCC6 - RAD1 | 2736 | 2511 | 1284 | RAD18 - XRCC6 | 279 | 2193 | 804 |
| RAD1 - XRCC6BP1 | 764 | 2108 | 1325 | RAD18 - XRCC6BP1 | 819 | 1954 | 1976 |
| Ranking of XRCC w.r.t RAD50 | Ranking of XRCC w.r.t RAD51 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| XRCC1 - RAD50 | 2573 | 2374 | 2497 | RAD51 - XRCC1 | 1673 | 1818 | 2611 |
| XRCC2 - RAD50 | 53 | 244 | 147 | XRCC2 - RAD51 | 472 | 2348 | 1973 |
| XRCC6 - RAD50 | 2615 | 2568 | 2582 | RAD51 - XRCC6 | 80 | 1244 | 2595 |
| RAD50 - XRCC6BP1 | 1962 | 1375 | 1366 | RAD51 - XRCC6BP1 | 792 | 951 | 1595 |
| Ranking of XRCC w.r.t RAD51AP1 | Ranking of XRCC w.r.t RAD51C | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| XRCC1 - RAD51AP1 | 1802 | 2732 | 801 | RAD51C - XRCC1 | 2282 | 1846 | 2026 |
| XRCC2 - RAD51AP1 | 78 | 112 | 351 | XRCC2 - RAD51C | 1695 | 932 | 520 |
| XRCC6 - RAD51AP1 | 2653 | 2439 | 347 | RAD51C - XRCC6 | 2545 | 1848 | 1858 |
| RAD51AP1 - XRCC6BP1 | 1790 | 936 | 974 | RAD51C - XRCC6BP1 | 2325 | 1070 | 1844 |
| Ranking of XRCC w.r.t RAD54B | Ranking of XRCC w.r.t RAD54L | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| XRCC1 - RAD54B | 2475 | 2670 | 1824 | RAD54L - XRCC1 | 1834 | 657 | 525 |
| XRCC2 - RAD54B | 1554 | 744 | 620 | XRCC2 - RAD54L | 2564 | 167 | 565 |
| XRCC6 - RAD54B | 2505 | 2709 | 2604 | RAD54L - XRCC6 | 2597 | 496 | 1247 |
| RAD54B - XRCC6BP1 | 1932 | 2504 | 2170 | RAD54L - XRCC6BP1 | 1389 | 1227 | 1454 |
Table 55 shows the derived influences which can be represented graphically, with the following influences - • RAD w.r.t XRCC with RAD-18/51/51AP1/51C/54B/54L <- XRCC1; RAD-18/51/51AP1/51C/54B/54L <- XRCC2; RAD-18/51/51AP1/51C/54B/54L <- XRCC6 and RAD-1/18/50/51/51AP1/51C/54B/54L <- XRCC6BP1; •; XRCC w.r.t RAD with RAD1 -> XRCC-2/6BP1; RAD18 -> XRCC-1/2/6; RAD50 -> XRCC-2/6BP1; RAD51 -> XRCC-6/6BP1; RAD51AP1 -> XRCC-2/6BP1; RAD51C -> XRCC-2; RAD54B -> XRCC-2; RAD54L -> XRCC-1/2/6/6BP1;
Table 55.
2nd order combinatorial hypotheses between RAD and XRCC.
| Unexplored combinatorial hypotheses | |
| RAD w.r.t XRCC family | |
| RAD-18/51/51AP1/51C/54B/54L | XRCC1 |
| RAD-18/51/51AP1/51C/54B/54L | XRCC2 |
| RAD-18/51/51AP1/51C/54B/54L | XRCC6 |
| RAD-1/18/50/51/51AP1/51C/54B/54L | XRCC6BP1 |
| XRCC w.r.t RAD family | |
| RAD1 | XRCC-2/6BP1 |
| RAD18 | XRCC-1/2/6 |
| RAD50 | XRCC-2/6BP1 |
| RAD51 | XRCC-6/6BP1 |
| RAD51AP1 | XRCC-2/6BP1 |
| RAD51C | XRCC-2 |
| RAD54B | XRCC-2 |
| RAD54L | XRCC-1/2/6/6BP1 |
3.4.2. XRN2 - RAD Cross Family Analysis
XRN2 (5’-3’ exoribonuclease 2) is involved in RNA synthesis/trafficking and termination. Morales et al. [139] observe that XRN2 links transcription termination to DNA damage and replication stress. They found an increase in the amount of RAD51 foci in shXRN2 cells compared to controls, suggesting that cells depleted of XRN2 are subjected to an increased level of basal DNA damage and show that loss of XRN2 also leads to the focal accumulation of several factors required for homologous recombination, such as ATM, BRCA1 and RAD51. This definitely shows that there is synergy between the XRN2 and RAD51. We found that both the XRN2 and RAD families were down regulated in CRC cell after ETC-1922159 treatment. The search engine gave rankings to the combinations of the XRN2 and RAD family members with low numerical valued in silico ranks. Table 56 shows the rankings of XRN2 w.r.t RAD family and vice versa. Following this is the derived influences in Table 57. We find RAD-51AP1/51/54L/51C/18/54B to be down regulated w.r.t XRN2. These are reflected in rankings of 340 (laplace), 545 (linear) and 290 (rbf) for RAD51AP1 - XRN2; 387 (laplace), 560 (linear) and 605 (rbf) for XRN2 - RAD51; 594 (laplace), 827 (linear) and 879 (rbf) for XRN2 - RAD54L; 639 (laplace), 1236 (linear) and 745 (rbf) for XRN2 - RAD51C; 794 (laplace), 688 (linear) and 804 (rbf) for XRN2 - RAD18; 255 (linear) and 122 (rbf) for XRN2 - RAD1 and 951 (laplace), 165 (linear) and 34 (rbf) for XRN2 - RAD54B; On the other hand, XRN2 was found to be down regulated w.r.t RAD family. These are reflected in rankings of 255 (laplace) and 122 (rbf) for XRN2 - RAD1; 1256 (linear) and 852 (rbf) for XRN2 - RAD51AP1; 1541 (laplace) and 1246 (linear) for XRN2 - RAD54L and 1037 (laplace) and 1777 (linear) for XRN2 - RAD51C. Graphical depiction of XRN2 and RAD family dependencies is shown as • RAD w.r.t XRN2 with XRN2 -> RAD-51AP1/51/54L/51C/18/54B and • XRN2 w.r.t RAD with XRN2 <- RAD1; XRN2 <- RAD51AP1; XRN2 <- RAD54L; XRN2 <- RAD51C;
Table 57 shows the derived influences which can be represented graphically, with the following influences - • RAD w.r.t XRN2 with XRN2 -> RAD-51AP1/51/54L/51C/18/54B; and • XRN2 w.r.t RAD with XRN2 <- RAD-1/51AP1/54L/51C.
Table 57.
2nd order combinatorial hypotheses between RAD and XRN2.
| Unexplored combinatorial hypotheses | |
| RAD w.r.t XRN2 | |
| XRN2 | RAD-51AP1/51/54L/51C/18/54B |
| XRN2 w.r.t RAD | |
| XRN2 | RAD1 |
| XRN2 | RAD51AP1 |
| XRN2 | RAD54L |
| XRN2 | RAD51C |
3.4.3. NKRF - RAD Cross Family Analysis
Not much is known about the NKRF (NF-κB-repressing factor) and RAD members. We found the combinations to be down regulated by the search engine between NKRF and RAD family. Table 58 shows the rankings of NKRF and RAD family. We found NKRF down regulated w.r.t RAD family. These are reflected in rankings of 1724 (laplace), 1642 (linear) and 649 (rbf) for RAD51AP1 <- NKRF; 982 (laplace), 1724 (linear) and 1352 (rbf) RAD51 <- NKRF; 1727 (laplace), 1387 (linear) and 1120 (rbf) for RAD54L <- NKRF; 1568 (laplace), 472 (linear) and 1505 (rbf) for RAD51C <- NKRF; 1508 (laplace), 615 (linear) and 405 (rbf) for RAD18 <- NKRF; and 1476 (laplace), 1189 (linear) and 1534 (rbf) for RAD54B <- NKRF;
Also, we found RAD family to be down regulated w.r.t NKRF. These are reflected in rankings of 157 (laplace) and 553 (linear) for RAD51AP1 - NKRF; 439 (laplace), 1441 (linear) and 1606 (rbf) for RAD51 - NKRF; 117 (laplace), 1175 (linear) and 1415 (rbf) for RAD54L - NKRF; 418 (laplace), and 1653 (rbf) for RAD51C - NKRF; 164 (laplace) and 1509 (rbf) for RAD18 - NKRF; 1391 (laplace), 1115 (linear) and 735 (rbf) NKRF - RAD1; 1354 (laplace), 851 (linear) and 824 (rbf) for NKRF - RAD50;
Table 58.
2nd order interaction ranking between RAD family vs NKRF.
| Ranking NKRF w.r.t RAD family | |||||||
| Ranking of NFRK w.r.t RAD family | Ranking of RAD family w.r.t NKRF | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| RAD51AP1 - NKRF | 1724 | 1642 | 649 | RAD51AP1 - NKRF | 157 | 553 | 2561 |
| RAD51 - NKRF | 982 | 1724 | 1352 | RAD51 - NKRF | 439 | 1441 | 1606 |
| RAD54L - NKRF | 1727 | 1387 | 1120 | RAD54L - NKRF | 117 | 1175 | 1415 |
| RAD51C - NKRF | 1568 | 472 | 1505 | RAD51C - NKRF | 418 | 2178 | 1653 |
| RAD18 - NKRF | 1508 | 615 | 405 | RAD18 - NKRF | 164 | 2306 | 1509 |
| RAD1 - NKRF | 2667 | 2222 | 1181 | NKRF - RAD1 | 1391 | 1115 | 735 |
| RAD54B - NKRF | 1476 | 1189 | 1534 | RAD54B - NKRF | 207 | 1869 | 2244 |
| RAD50 - NKRF | 2003 | 2343 | 2511 | NKRF - RAD50 | 1354 | 851 | 824 |
Table 59 shows the derived influences which can be represented graphically, with the following influences - • RAD w.r.t NKRF with RAD51AP1 <- NKRF; RAD51 <- NKRF; RAD54L <- NKRF; RAD51C <- NKRF; RAD18 <- NKRF; RAD1 <- NKRF; RAD54B <- NKRF and • NKRF w.r.t RAD with RAD51AP1 -> NKRF; RAD51 -> NKRF; RAD54L -> NKRF; RAD51C -> NKRF; RAD18 -> NKRF; NKRF -> RAD1; NKRF -> RAD50.
3.4.4. RAD - BCL Cross Family Analysis
Saintigny et al. [140] show a specific role of BCL2 in suppression of the RAD51 recombination pathway. They observe that BCL2 consistently inhibits recombination stimulated by RAD51 overexpression and alters RAD51 protein by post-translation modification. Based on the findings that CARD9 and BCL10 acted together to activate NF-κB following cytosolic DNA sensing, Meng et al. [141] demonstrated that BCL10 was recruited to the dsDNA–RAD50 complexes in a CARD9-dependent manner. These mechanisms point to a synergy between BCL and RAD family. In CRC cells treated with ETC-1922159, BCL and RAD family members were found to be down regulated. The search engine alloted the combinations of RAD and BCL low numerical valued ranks pointing to possible synergistic down regulations. Table 60 shows rankings of BCL and RAD w.r.t to each other. The left half of the Table points to rankings of BCL family w.r.t RAD family. The right half of the Table points to rankings of RAD family w.r.t BCL family.
On the left side, BCL2L12 was found to be down regulated w.r.t RAD-1/18/50/51/51C/54B/54L. These are reflected in rankings of 1530 (linear) and 1401 (rbf) for RAD1 - BCL2L12; 675 (laplace) and 1312 (rbf) for RAD18 - BCL2L12; 1151 (linear) and 929 (rbf) for RAD50 - BCL2L12; 1234 (laplace) and 1334 (linear) for RAD51 - BCL2L12; 1561 (laplace) and 1647 (rbf) for RAD51C - BCL2L12; 1329 (linear) and 1625 (rbf) for RAD54B - BCL2L12, and 821 (linear) and 210 (rbf) for RAD54L - BCL2L12; BCL6B was found to be down regulated w.r.t RAD-1/18/50/51/51AP1/51C/54B/54L. 194 (laplace), 481 (linear) and 102 (rbf) for RAD1 - BCL6B; 176 (linear) and 929 (rbf) for RAD18 - BCL6B; 860 (laplace), 87 (linear) and 74 (rbf) for RAD50 - BCL6B; 263 (linear) and 58 (rbf) for RAD51 - BCL6B; 723 (laplace), 428 (linear) and 579 (rbf) for RAD51AP1 - BCL6B; 660 (laplace), 521 (linear) and 1609 (rbf) for RAD51C - BCL6B; 708 (laplace), 596 (linear) and 647 (rbf) for RAD54B - BCL6B; and 108 (laplace) and 1326 (rbf) for RAD54L - BCL6B; BCL7A was found to be down regulated w.r.t RAD-1/18/50/51/54L. These are reflected in rankings of 690 (laplace) and 1202 (rbf) for BCL7A - RAD1; 385 (laplace) and 185 (rbf) for BCL7A - RAD18; 137 (laplace), 601 (linear) and 41 (rbf) for RAD50 - BCL7A; 514 (laplace) and 1694 (linear) for BCL7A - RAD51; 1519 (laplace), 418 (linear) and 842 (rbf) for RAD54L - BCL7A; BCL9 was found to be down regulated w.r.t RAD-18/51/51C/54L. These are reflected in rankings for 461 (laplace) and 1453 (linear) for RAD18 - BCL9; 1143 (linear) and 95 (rbf) for RAD51 - BCL9; 956 (laplace) and 376 (rbf) for RAD51C - BCL9; 1450 (laplace), 1096 (linear) and 400 (rbf) for RAD54L - BCL9; BCL11A was found to be down regulated w.r.t RAD-1/18/50/51/51AP1/51C/54B. These are reflected in rankings of 1069 (laplace), 507 (linear) and 1267 (rbf) for RAD1 - BCL11A; 1561 (laplace), 169 (linear) and 692 (rbf) for RAD18 - BCL11A; 582 (laplace), 1144 (linear) and 1047 (rbf) for RAD50 - BCL11A; 1120 (laplace), 752 (linear) and 645 (rbf) for RAD51AP1 - BCL11A; 1024 (laplace), 199 (linear) and 899 (rbf) for RAD51C - BCL11A; and 1037 (laplace), 917 (linear) and 867 (rbf) for RAD54B - BCL11A. BCL11B was found to be down regulated w.r.t RAD-50/51/51AP1/54B/54L. These are reflected in rankings of 1198 (linear) and 903 (rbf) for RAD50 - BCL11B; 449 (linear) and 971 (rbf) for RAD51 - BCL11B; 1247 (laplace), 908 (linear) and 1671 (rbf) for RAD51AP1 - BCL11B; 1193 (laplace), 1192 (linear) and 832 (rbf) for RAD54B - BCL11B and 1421 (laplace) and 1385 (linear) for RAD54L - BCL11B.
Table 60.
2nd order interaction ranking between RAD and BCL family members.
| Ranking RAD family VS BCL family | |||||||
| Ranking of BCL2L12 w.r.t RAD family | Ranking of RAD family w.r.t BCL2L12 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| RAD1 - BCL2L12 | 1797 | 1530 | 1401 | RAD1 - BCL2L12 | 1958 | 2120 | 1957 |
| RAD18 - BCL2L12 | 675 | 2437 | 1312 | RAD18 - BCL2L12 | 779 | 652 | 1388 |
| RAD50 - BCL2L12 | 2080 | 1151 | 929 | RAD50 - BCL2L12 | 1668 | 2566 | 1703 |
| RAD51 - BCL2L12 | 1234 | 1334 | 2350 | RAD51 - BCL2L12 | 1164 | 365 | 1213 |
| RAD51AP1 - BCL2L12 | 2267 | 2500 | 2265 | RAD51AP1 - BCL2L12 | 306 | 57 | 28 |
| RAD51C - BCL2L12 | 1561 | 2384 | 1647 | RAD51C - BCL2L12 | 495 | 1191 | 429 |
| RAD54B - BCL2L12 | 1979 | 1329 | 1625 | RAD54B - BCL2L12 | 678 | 432 | 787 |
| RAD54L - BCL2L12 | 2446 | 821 | 210 | RAD54L - BCL2L12 | 901 | 1128 | 263 |
| Ranking of BCL6B w.r.t RAD family | Ranking of RAD family w.r.t BCL6B | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| RAD1 - BCL6B | 194 | 481 | 102 | RAD1 - BCL6B | 2110 | 2151 | 2059 |
| RAD18 - BCL6B | 1790 | 176 | 929 | RAD18 - BCL6B | 1113 | 640 | 482 |
| RAD50 - BCL6B | 860 | 87 | 74 | RAD50 - BCL6B | 2164 | 2412 | 2581 |
| RAD51 - BCL6B | 2324 | 263 | 58 | RAD51 - BCL6B | 287 | 681 | 497 |
| RAD51AP1 - BCL6B | 723 | 428 | 579 | RAD51AP1 - BCL6B | 1607 | 1638 | 916 |
| RAD51C - BCL6B | 660 | 521 | 1609 | RAD51C - BCL6B | 43 | 871 | 999 |
| RAD54B - BCL6B | 708 | 596 | 647 | RAD54B - BCL6B | 1212 | 1392 | 1170 |
| RAD54L - BCL6B | 108 | 2684 | 1326 | RAD54L - BCL6B | 1867 | 1009 | 785 |
| Ranking of BCL7A w.r.t RAD family | Ranking of RAD family w.r.t BCL7A | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| RAD1 - BCL7A | 690 | 1791 | 1202 | RAD1 - BCL7A | 1989 | 2101 | 1804 |
| RAD18 - BCL7A | 385 | 2366 | 185 | RAD18 - BCL7A | 1514 | 1515 | 783 |
| RAD50 - BCL7A | 137 | 601 | 417 | RAD50 - BCL7A | 2123 | 1771 | 2085 |
| RAD51 - BCL7A | 514 | 1694 | 2361 | RAD51 - BCL7A | 879 | 274 | 639 |
| RAD51AP1 - BCL7A | 2440 | 2609 | 774 | RAD51AP1 - BCL7A | 412 | 416 | 4 |
| RAD51C - BCL7A | 2726 | 2448 | 983 | RAD51C - BCL7A | 215 | 394 | 461 |
| RAD54B - BCL7A | 2729 | 1830 | 2743 | RAD54B - BCL7A | 809 | 1407 | 213 |
| RAD54L - BCL7A | 1519 | 418 | 842 | RAD54L - BCL7A | 435 | 783 | 1499 |
| Ranking of BCL9 w.r.t RAD family | Ranking of RAD family w.r.t BCL9 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| RAD1 - BCL9 | 1296 | 2418 | 1775 | RAD1 - BCL9 | 1749 | 2528 | 1391 |
| RAD18 - BCL9 | 461 | 1952 | 1453 | RAD18 - BCL9 | 656 | 1194 | 482 |
| RAD50 - BCL9 | 2338 | 2653 | 2559 | RAD50 - BCL9 | 2220 | 1441 | 1098 |
| RAD51 - BCL9 | 1748 | 1143 | 952 | RAD51 - BCL9 | 622 | 929 | 860 |
| RAD51AP1 - BCL9 | 1861 | 2280 | 786 | RAD51AP1 - BCL9 | 331 | 61 | 102 |
| RAD51C - BCL9 | 956 | 2741 | 376 | RAD51C - BCL9 | 1113 | 417 | 1154 |
| RAD54B - BCL9 | 2063 | 2375 | 1050 | RAD54B - BCL9 | 1045 | 53 | 650 |
| RAD54L - BCL9 | 1450 | 1096 | 400 | RAD54L - BCL9 | 636 | 602 | 934 |
| Ranking of BCL11A w.r.t RAD family | Ranking of RAD family w.r.t BCL11A | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| RAD1 - BCL11A | 1069 | 507 | 1267 | RAD1 - BCL11A | 1430 | 1475 | 1584 |
| RAD18 - BCL11A | 1561 | 169 | 692 | RAD18 - BCL11A | 465 | 164 | 1952 |
| RAD50 - BCL11A | 582 | 1144 | 1047 | RAD50 - BCL11A | 2649 | 875 | 1226 |
| RAD51 - BCL11A | 1722 | 2073 | 339 | RAD51 - BCL11A | 255 | 2064 | 2461 |
| RAD51AP1 - BCL11A | 1120 | 752 | 645 | RAD51AP1 - BCL11A | 659 | 388 | 496 |
| RAD51C - BCL11A | 1024 | 199 | 899 | RAD51C - BCL11A | 363 | 1673 | 97 |
| RAD54B - BCL11A | 1037 | 917 | 867 | RAD54B - BCL11A | 581 | 2743 | 799 |
| RAD54L - BCL11A | 172 | 2193 | 2318 | RAD54L - BCL11A | 846 | 2733 | 209 |
| Ranking of BCL11B w.r.t RAD family | Ranking of RAD family w.r.t BCL11B | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| RAD1 - BCL11B | 2371 | 2360 | 43 | RAD1 - BCL11B | 2571 | 230 | 1373 |
| RAD18 - BCL11B | 1741 | 993 | 2677 | RAD18 - BCL11B | 1747 | 2028 | 14 |
| RAD50 - BCL11B | 2010 | 1198 | 903 | RAD50 - BCL11B | 919 | 860 | 2263 |
| RAD51 - BCL11B | 2067 | 449 | 971 | RAD51 - BCL11B | 1095 | 1238 | 2373 |
| RAD51AP1 - BCL11B | 1247 | 908 | 1671 | RAD51AP1 - BCL11B | 196 | 2646 | 987 |
| RAD51C - BCL11B | 1736 | 1234 | 2282 | RAD51C - BCL11B | 1122 | 1844 | 1161 |
| RAD54B - BCL11B | 1193 | 1192 | 832 | RAD54B - BCL11B | 363 | 2150 | 1561 |
| RAD54L - BCL11B | 1421 | 1385 | 1854 | RAD54L - BCL11B | 579 | 2543 | 159 |
On the right side, w.r.t BCL2L12, RAD-18/50/51/51AP1/51C/54B/54L were found to be down regulated. These are found in the rankings of 779 (laplace), 652 (linear) and 1388 (rbf) for RAD18 - BCL2L12; 1668 (laplace), 2566 (linear) and 1703 (rbf) for RAD50 - BCL2L12; 1164 (laplace), 365 (linear), 1213 (rbf) for RAD51 - BCL2L12; 306 (laplace), 57 (linear) and 28 (rbf) for RAD51AP1 - BCL2L12; 495 (laplace), 1191 (linear) and 429 (rbf) for RAD51C - BCL2L12; 678 (laplace), 432 (linear) and 787 (rbf) for RAD54B - BCL2L12; and 901 (laplace), 1128 (linear) and 263 (rbf) for RAD54L - BCL2L12; w.r.t BCL6B, RAD-18/51/51AP1/51C/54B/54L were found to be down regulated. These are reflected in rankings of 1113 (laplace), 640 (linear) and 482 (rbf) for RAD18 - BCL6B; 287 (laplace), 681 (linear) and 497 (rbf) for RAD51 - BCL6B; 1607 (laplace), 1638 (linear) and 916 (rbf) for RAD51AP1 - BCL6B; 43 (laplace), 871 (linear) and 999 (rbf) for RAD51C - BCL6B; 1212 (laplace), 1392 (linear) and 1170 (rbf) for RAD54B - BCL6B; and 1009 (linear) and 785 (rbf) for RAD54L - BCL6B; w.r.t BCL7A, RAD-18/51/51AP1/51C/54B/54L were found to be down regulated. These are reflected in rankings of 1514 (laplace), 1515 (linear), 783 (rbf) for RAD18 - BCL7A; 879 (laplace), 274 (linear) and 639 (rbf) for RAD51 - BCL7A; 412 (laplace), 416 (linear) and 4 (rbf) for RAD51AP1 - BCL7A; 215 (laplace), 394 (linear) and 461 (rbf) for RAD51C - BCL7A; 809 (laplace), 1407 (linear) and 213 (rbf) for RAD54B - BCL7A and 435 (laplace), 783 (linear) and 1499 (rbf) for RAD54L - BCL7A. w.r.t BCL9, RAD-18/50/51/51AP1/51C/54B/54L were found to be down regulated. These are reflected in the rankings of 656 (laplace), 1194 (linear) and 482 (rbf) for RAD18 - BCL9; 1441 (linear) and 1098 (rbf) for RAD50 - BCL9; 622 (laplace), 929 (linear), 860 (rbf) for RAD51 - BCL9; 331 (laplace), 61 (linear) and 102 (rbf) for RAD51AP1 - BCL9; 1113 (laplace), 417 (linear) and 1154 (rbf) for RAD51C - BCL9; 1045 (laplace), 53 (linear) and 650 (rbf) for RAD54B - BCL9 and 636 (laplace), 602 (linear) and 934 (rbf) for RAD54L - BCL9. w.r.t BCL11A, RAD-1/18/50/51/51AP1/51C/54B/54L were found to be down regulated. These are reflected in 1430 (laplace), 1475 (linear) and 1584 (rbf) for RAD1 - BCL11A; 465 (laplace) and 164 (linear) for RAD18 - BCL11A; 875 (linear) and 1226 (rbf) for RAD50 - BCL11A; 659 (laplace), 388 (linear) and 496 (rbf) for RAD51AP1 - BCL11A; 363 (laplace), 1673 (linear) and 97 (rbf) for RAD51C - BCL11A; 581 (laplace) and 799 (rbf) for RAD54B - BCL11A; and 846 (laplace) and 209 (rbf) for RAD54L - BCL11A; w.r.t BCL11B, RAD-1/50/51/51AP1/51C/54B/54L were found to be down regulated. These are reflected in rankings of 230 (linear) and 1373 (rbf) RAD1 - BCL11B; 919 (laplace) and 860 (linear) for RAD50 - BCL11B; 1095 (laplace) and 1238 (linear) RAD51 - BCL11B; 196 (laplace) and 987 (rbf) for RAD51AP1 - BCL11B; 1122 (laplace) and 1161 (rbf) for RAD51C - BCL11Bl; 363 (laplace) and 1561 (rbf) for RAD54B - BCL11B; 579 (laplace), 2543 (linear) and 159 (rbf) for RAD54L - BCL11B.
Table 61 shows the derived influences which can be represented graphically, with the following influences - • RAD w.r.t BCL with RAD-18/50/51/51AP1/51C/54B/54L <- BCL-2L12; RAD-18/51/51AP1/51C/54B/54L <- BCL-6B; RAD-18/51/51AP1/51C/54B/54L <- BCL-7A; RAD-18/50/51/51AP1/51C/54B/54L <- BCL-9; RAD-1/18/50/51/51AP1/51C/54B/54L <- BCL-11A; RAD-1/50/51/51AP1/51C/ 54B/54L <- BCL-11B; and • BCL w.r.t RAD with RAD-1/18/50/51/51C/54B/54L -> BCL-2L12; RAD-1/18/50/51/51AP1/51C/54B/54L -> BCL-6B; RAD-1/18/50/51/54L -> BCL-7A; RAD-18/51/51C/54L -> BCL-9; RAD-1/18/50/51/51AP1/51C/54B -> BCL-11A; and RAD-50/51/51AP1/54B/54L -> BCL-11B.
3.4.5. RAD - EXOSC Cross Family Analysis
Marin-Vicente et al. [142] show that RRP6/EXOSC10 is required for the repair of DNA double-strand breaks by homologous recombination. The authors results suggest that ribonucleolytic activity of RRP6/EXOSC10 is required for the recruitment of RAD51 to DSBs. The therapeutic potential of exosome-mediated siRNA delivery was demonstrated in vitro by the strong knockdown of RAD51, a prospective therapeutic target for cancer cells (Shtam et al. [143]). These findings point to the synergy between EXOSC and RAD family. In CRC cells treated with ETC-1922159, they were down regulated and the search engine allocated low numerical rankings for combinations, thus pointing to possible synergistic down regulation. Table 62 shows the rankings of the EXOSC and RAD family w.r.t to each other. On the left half of the Table is the rankings of EXOSC w.r.t RAD family. EXOSC2 was found to be down regulated w.r.t RAD-1/18/50/51/51AP1/51C/54B/54L. These are reflected in rankings of 1033 (laplace), 1311 (linear) and 1207 (rbf) for EXOSC2 - RAD1; 1210 (laplace) and 995 (linear) for EXOSC2 - RAD18; 1124 (laplace), 698 (linear) and 629 (rbf) for EXOSC2 - RAD50; 1754 (laplace), 191 (linear) and 633 (rbf) and for EXOSC2 - RAD51; 198 (laplace) and 1462 (linear) for EXOSC2 - RAD51AP1; 87 (laplace), 463 (linear) and 1130 (rbf) for EXOSC2 - RAD51C; 351 (laplace), 135 (linear) and 142 (rbf) for EXOSC2 - RAD54B; and 1131 (laplace), 1652 (linear) and 320 (rbf) for EXOSC2 - RAD54L. EXOSC3 was found to be down regulated w.r.t RAD-1/18/51/51AP1/54L. These are reflected in rankings of 1677 (linear) and 549 (rbf) for EXOSC3 - RAD1; 1676 (laplace) and 184 (rbf) for EXOSC3 - RAD18; 894 (laplace) and 1066 (linear) for EXOSC3 - RAD51; 1037 (linear) and 804 (rbf) for EXOSC3 - RAD51AP1, and 469 (linear) and 736 (rbf) for EXOSC3 - RAD54L. EXOSC5 was found to be down regulated w.r.t RAD-1/18/50/51/51AP1/51C/54B/54L. These are reflected in rankings of 568 (laplace), 1169 (linear) and 1699 (rbf) for EXOSC5 - RAD1; 219 (linear) and 1652 (rbf) for EXOSC5 - RAD18; 447 (laplace), 195 (linear) and 475 (rbf) for EXOSC5 - RAD50; 431 (linear) and 1121 (rbf) for EXOSC5 - RAD51; 1290 (laplace), 487 (linear) and 430 (rbf) for EXOSC5 - RAD51AP1; 1284 (laplace) and 1264 (linear) for EXOSC5 - RAD51C; 940 (laplace), 812 (linear) and 1036 (rbf) for EXOSC5 - RAD54B; and 408 (laplace) and 1407 (rbf) for EXOSC5 - RAD54L; EXOSC6 was found to be down regulated w.r.t RAD-18/51/54L. These were reflected in rankings of 1637 (laplace), 1599 (linear) and 2254 (rbf) for EXOSC6 - RAD18; 1056 (laplace), 1482 (linear) and 1007 (rbf) for EXOSC6 - RAD51; and 987 (laplace) and 1642 (rbf) for EXOSC6 - RAD54L; EXOSC7 was found to be down regulated w.r.t RAD-1/18/51C/54B/54L. These are reflected in rankings of 1735 (linear) and 1210 (rbf) for EXOSC7 - RAD1; 490 (laplace), 1688 (linear) and 1331 (rbf) for EXOSC7 - RAD18; 1113 (laplace), 1623 (linear) and 530 (rbf) for EXOSC7 - RAD51C; 1612 (linear) and 1191 (rbf) for EXOSC7 - RAD54B; and 1550 (laplace), 1754 (linear) and 1728 (rbf) for EXOSC7 - RAD54L; EXOSC8 was found to be down regulated w.r.t RAD-18/51/51AP1/54B/54L. These are reflected in 805 (laplace) and 1564 (rbf) for EXOSC8 - RAD18; 404 (laplace) and 1630 (linear) for EXOSC8 - RAD51; 1567 (linear) and 1701 (rbf) for EXOSC8 - RAD51AP1; 1562 (laplace) and 1736 (rbf) for EXOSC8 - RAD54B; and 1248 (laplace), 622 (linear) and 239 (rbf) for EXOSC8 - RAD54L; EXOSC9 was found to be down regulated w.r.t RAD-1/18/50/51/51C/54B/54L. These are reflected in rankings of 175 (linear) and 1648 (rbf) for EXOSC9 - RAD1; 1533 (laplace), 774 (linear) and 1180 (rbf) for EXOSC9 - RAD18; 545 (laplace), 183 (linear) and 467 (rbf) for EXOSC9 - RAD50; 866 (laplace), 106 (linear) and 99 (rbf) for EXOSC9 - RAD51; 110 (laplace), 742 (linear) and 200 (rbf) for EXOSC9 - RAD51C; 179 (laplace), 178 (linear) and 84 (rbf) for EXOSC9 - RAD54B and 1113 (laplace) and 22 (rbf) for EXOSC9 - RAD54L;
On the right half of the Table is the rankings of RAD family w.r.t EXOSC. RAD-18/51/51C/54B/54L was found to be down regulated w.r.t EXOSC2. These are reflected in rankings of 1115 (laplace), 979 (linear) and 654 (rbf) for EXOSC2 - RAD18; 795 (laplace), 1332 (linear) and 441 (rbf) for EXOSC2 - RAD51; 636 (laplace), 564 (linear) and 152 (rbf) for EXOSC2 - RAD51C; 278 (laplace), 132 (linear) and 282 (rbf) for EXOSC2 - RAD54B and 125 (laplace), 888 (linear) and 545 (rbf) for EXOSC2 - RAD54L. RAD-18/50/51/51AP1/51C/54B/54L was found to be down regulated w.r.t EXOSC3. These are reflected in rankings of 1468 (linear) and 767 (rbf) for EXOSC3 - RAD18; 1062 (laplace) and 596 (linear) for EXOSC3 - RAD50; 727 (laplace), 583 (linear) and 963 (rbf) for EXOSC3 - RAD51; 100 (laplace), 49 (linear) and 219 (rbf) for EXOSC3 - RAD51AP1; 663 (laplace), 869 (linear) and 887 (rbf) for EXOSC3 - RAD51C; 384 (laplace), 277 (linear) and 310 (rbf) for EXOSC3 - RAD54B and 546 (laplace), 1117 (linear) and 808 (rbf) for EXOSC3 - RAD54L; RAD-1/18/51/51AP1/51C/54B/54L was found to be down regulated w.r.t EXOSC5. These are reflected in rankings of 1716 (linear) and 1718 (rbf) for EXOSC5 - RAD1; 1026 (laplace), 550 (linear) and 253 (rbf) for EXOSC5 - RAD18; 260 (laplace), 1095 (linear) and 137 (rbf) for EXOSC5 - RAD51; 1555 (laplace) and 976 (rbf) for EXOSC5 - RAD51AP1; 233 (laplace), 1003 (linear) and 359 (rbf) for EXOSC5 - RAD51C; 834 (laplace), 1825 (linear) and 335 (rbf) for EXOSC5 - RAD54B; and 248 (laplace), 197 (linear) and 39 (rbf) for EXOSC5 - RAD54L. RAD-1/18/50/51AP1/51C/54L was found to be down regulated w.r.t EXOSC6. These are reflected in rankings of 142 (linear) and 639 (rbf) for EXOSC6 - RAD1; 1118 (laplace), 1313 (linear) and 1549 (rbf) for EXOSC6 - RAD18; 1722 (linear) and 575 (rbf) for EXOSC6 - RAD50; 149 (laplace) and 1060 (linear) for EXOSC6 - RAD51AP1; 500 (laplace) and 1628 (linear) for EXOSC6 - RAD51C; and 885 (laplace), 271 (linear) and 1224 (rbf) for EXOSC6 - RAD54L; RAD-18/51/51AP1/51C/54B/54L was found to be down regulated w.r.t EXOSC7. These were reflected in rankings of 441 (laplace), 385 (linear) and 1542 (rbf) for EXOSC7 - RAD18; 376 (laplace), 1180 (linear) and 550 (rbf) for EXOSC7 - RAD51; 35 (laplace), 97 (linear) and 786 (rbf) for EXOSC7 - RAD51AP1; 854 (laplace), 671 (linear) and 1459 (rbf) for EXOSC7 - RAD51C; 458 (laplace), 260 (linear) and 646 (rbf) for EXOSC7 - RAD54B; and 464 (laplace), 528 (linear) and 790 (rbf) for EXOSC7 - RAD54L; RAD-1/18/51/51AP1/51C/54B/54L was found to be down regulated w.r.t EXOSC8. These were reflected in rankings of 151 (linear) and 1563 (rbf) for EXOSC8 - RAD1; 764 (laplace), 523 (linear) and 29 (rbf) for EXOSC8 - RAD18; 98 (laplace), 1161 (linear) and 902 (rbf) for EXOSC8 - RAD51; 408 (laplace) and 541 (rbf) for EXOSC8 - RAD51AP1; 906 (laplace), 738 (linear) and 1052 (rbf) for EXOSC8 - RAD51C; 23 (laplace), 1578 (linear) and 130 (rbf) for EXOSC8 - RAD54B; and 651 (laplace), 1384 (linear) and 1047 (rbf) for EXOSC8 - RAD54L; RAD-1/18/50/51/51AP1/51C/54B/54L was found to be down regulated w.r.t EXOSC9. These were reflected in rankings of 1335 (laplace) and 978 (rbf) for EXOSC9 - RAD1; 54 (linear) and 540 (rbf) for EXOSC9 - RAD18; 211 (laplace) and 1377 (rbf) for EXOSC9 - RAD50; 807 (laplace), 74 (linear) and 429 (rbf) for EXOSC9 - RAD51; 103 (linear), 1210 (rbf) for EXOSC9 - RAD51AP1; 399 (laplace), 844 (linear) and 69 (rbf) for EXOSC9 - RAD51C; 466 (linear), 1286 (rbf) for EXOSC9 - RAD54B; and 536 (laplace), 724 (linear) and 414 (rbf) for EXOSC9 - RAD54L;
Table 63 shows the derived influences which can be represented graphically, with the following influences - • RAD w.r.t EXOSC with EXOSC-2 -> RAD-18/51/51C/54B/54L; EXOSC-3 -> RAD-18/50/51/51AP1/51C/54B/54L; EXOSC-5 -> RAD-1/18/51/51AP1/51C/54B/54L; EXOSC-6 -> RAD-1/18/50/51AP1/51C/54L; EXOSC-7 -> RAD-18/51/51AP1/51C/54B/54L; EXOSC-8 -> RAD-1/18/51/51AP1/51C/54B/54L; EXOSC-9 -> RAD-1/18/50/51/51AP1/51C/54B/54L; and • EXOSC w.r.t RAD with EXOSC-2 <- RAD-1/18/50/51/51AP1/51C/54B/54L; EXOSC-3 <- RAD-1/18/51/51AP1/54L; EXOSC-5 <- RAD-1/18/50/51/51AP1/51C/54B/54L; EXOSC-6 <- RAD-18/51/54L; EXOSC-7 <- RAD-1/18/51C/54B/54L; EXOSC-8 <- RAD-18/51/51AP1/54B/54L; and EXOSC-9 <- RAD-1/18/50/51/51C/54B/54L.
3.4.6. XRCC - EXOSC Cross Family Analysis
Not much is known about XRCC - EXOSC synergy, however both were found to be down regulated in CRC cells after treatment with ETC-1922159. The search engine also allocated rankings of low numerical values to several combinations thus indicating plausible synergistic down regulations. Table 64 shows the rankings of XRCC vs EXOSC family members.
Table 64.
2nd order interaction ranking between RAD and EXOSC family members.
| Ranking XRCC family VS EXOSC family | |||||||
| Ranking of EXOSC2 w.r.t XRCC family | Ranking of XRCC family w.r.t EXOSC2 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| EXOSC2 - XRCC1 | 277 | 176 | 423 | EXOSC2 - XRCC1 | 2708 | 2386 | 2634 |
| EXOSC2 - XRCC2 | 8 | 38 | 100 | EXOSC2 - XRCC2 | 166 | 417 | 56 |
| EXOSC2 - XRCC6 | 1252 | 398 | 623 | EXOSC2 - XRCC6 | 2678 | 2504 | 2576 |
| EXOSC2 - XRCC6BP1 | 935 | 905 | 1755 | EXOSC2 - XRCC6BP1 | 1740 | 1842 | 2177 |
| Ranking of EXOSC3 w.r.t XRCC family | Ranking of XRCC family w.r.t EXOSC3 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| EXOSC3 - XRCC1 | 1551 | 2256 | 1974 | EXOSC3 - XRCC1 | 2217 | 1418 | 2041 |
| EXOSC3 - XRCC2 | 2462 | 2553 | 2329 | EXOSC3 - XRCC2 | 125 | 15 | 194 |
| EXOSC3 - XRCC6 | 1720 | 1716 | 2398 | EXOSC3 - XRCC6 | 2742 | 2608 | 2193 |
| EXOSC3 - XRCC6BP1 | 2506 | 1523 | 1356 | EXOSC3 - XRCC6BP1 | 2561 | 2154 | 2406 |
| Ranking of EXOSC5 w.r.t XRCC family | Ranking of XRCC family w.r.t EXOSC5 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| EXOSC5 - XRCC1 | 741 | 291 | 8 | EXOSC5 - XRCC1 | 2578 | 2568 | 1910 |
| EXOSC5 - XRCC2 | 1244 | 791 | 702 | EXOSC5 - XRCC2 | 1559 | 1857 | 866 |
| EXOSC5 - XRCC6 | 65 | 1064 | 322 | EXOSC5 - XRCC6 | 2410 | 2465 | 2190 |
| EXOSC5 - XRCC6BP1 | 416 | 880 | 1434 | EXOSC5 - XRCC6BP1 | 1907 | 2029 | 1394 |
| Ranking of EXOSC6 w.r.t XRCC family | Ranking of XRCC family w.r.t EXOSC6 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| EXOSC6 - XRCC1 | 1890 | 985 | 1163 | EXOSC6 - XRCC1 | 509 | 2373 | 1046 |
| EXOSC6 - XRCC2 | 1512 | 648 | 1458 | EXOSC6 - XRCC2 | 486 | 2564 | 1901 |
| EXOSC6 - XRCC6 | 2304 | 1719 | 2690 | EXOSC6 - XRCC6 | 2576 | 35 | 188 |
| EXOSC6 - XRCC6BP1 | 2428 | 492 | 2112 | EXOSC6 - XRCC6BP1 | 1753 | 1295 | 366 |
| Ranking of EXOSC7 w.r.t XRCC family | Ranking of XRCC family w.r.t EXOSC7 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| EXOSC7 - XRCC1 | 1907 | 1510 | 1603 | EXOSC7 - XRCC1 | 1844 | 1229 | 987 |
| EXOSC7 - XRCC2 | 1369 | 2555 | 2124 | EXOSC7 - XRCC2 | 176 | 436 | 788 |
| EXOSC7 - XRCC6 | 584 | 1523 | 1018 | EXOSC7 - XRCC6 | 1074 | 242 | 288 |
| EXOSC7 - XRCC6BP1 | 1419 | 1944 | 876 | EXOSC7 - XRCC6BP1 | 2144 | 1577 | 2038 |
| Ranking of EXOSC8 w.r.t XRCC family | Ranking of XRCC family w.r.t EXOSC8 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| EXOSC8 - XRCC1 | 1373 | 1515 | 2103 | EXOSC8 - XRCC1 | 1769 | 2151 | 1435 |
| EXOSC8 - XRCC2 | 1086 | 2309 | 2435 | EXOSC8 - XRCC2 | 13 | 1932 | 6 |
| EXOSC8 - XRCC6 | 1820 | 2542 | 2693 | EXOSC8 - XRCC6 | 1869 | 1233 | 2625 |
| EXOSC8 - XRCC6BP1 | 2112 | 1994 | 2699 | EXOSC8 - XRCC6BP1 | 2305 | 2461 | 2319 |
| Ranking of EXOSC9 w.r.t XRCC family | Ranking of XRCC family w.r.t EXOSC9 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| EXOSC9 - XRCC1 | 44 | 1214 | 1410 | EXOSC9 - XRCC1 | 1804 | 2696 | 1629 |
| EXOSC9 - XRCC2 | 496 | 672 | 840 | EXOSC9 - XRCC2 | 1793 | 655 | 1526 |
| EXOSC9 - XRCC6 | 1121 | 151 | 689 | EXOSC9 - XRCC6 | 1882 | 2188 | 2404 |
| EXOSC9 - XRCC6BP1 | 362 | 463 | 1741 | EXOSC9 - XRCC6BP1 | 1206 | 1776 | 1626 |
Table 65.
2nd order combinatorial hypotheses between XRCC and EXOSC members.
| Unexplored combinatorial hypotheses | |
| XRCC w.r.t EXOSC | |
| EXOSC-2 | XRCC-2 |
| EXOSC-3 | XRCC-2 |
| EXOSC-5 | XRCC-2 |
| EXOSC-6 | XRCC-6 |
| EXOSC-7 | XRCC-1/2/6 |
| EXOSC-8 | XRCC-2 |
| EXOSC-9 | XRCC-2/6BP1 |
| EXOSC w.r.t XRCC | |
| EXOSC-2 | XRCC-1/2/6/6BP1 |
| EXOSC-3 | XRCC-6/6BP1 |
| EXOSC-5 | XRCC-1/2/6/6BP1 |
| EXOSC-6 | XRCC-1/2 |
| EXOSC-7 | XRCC-1/6/6BP1 |
| EXOSC-8 | XRCC-1 |
| EXOSC-9 | XRCC-1/2/6/6BP1 |
On the left half of the Table is the rankings of EXOSC w.r.t XRCC family. EXOSC2 was found to be down regulated w.r.t XRCC-1/2/6/6BP1. These are reflected in rankings of 277 (laplace), 176 (linear) and 423 (rbf) for EXOSC2 - XRCC1; 8 (laplace), 38 (linear) and 100 (rbf) for EXOSC2 - XRCC2; 1252 (laplace), 398 (linear) and 623 (rbf) for EXOSC2 - XRCC6; and 935 (laplace) and 905 (linear) for EXOSC2 - XRCC6BP1; EXOSC3 was found to be down regulated w.r.t XRCC-6BP1. These are reflected in rankings of 1523 (linear) and 1356 (rbf) for EXOSC3 - XRCC6BP1; EXOSC5 was found to be down regulated w.r.t XRCC-1/2/6/6BP1. These are reflected in rankings of 741 (laplace), 291 (linear) and 8 (rbf) for EXOSC5 - XRCC1; 1244 (laplace), 791 (linear) and 702 (rbf) for EXOSC5 - XRCC2; 65 (laplace), 1064 (linear) and 322 (rbf) for EXOSC5 - XRCC6; and 416 (laplace), 880 (linear) and 1434 (rbf) for EXOSC5 - XRCC6BP1. EXOSC6 was found to be down regulated w.r.t XRCC-1/2. These are reflected in rankings of 985 (linear) and 1163 (rbf) for EXOSC6 - XRCC1 and 1512 (laplace), 648 (linear) and 1458 (rbf) for EXOSC6 - XRCC2; EXOSC7 was found to be down regulated w.r.t XRCC-1/6/6BP1. These are reflected in rankings of 1510 (linear) and 1603 (rbf) for EXOSC7 - XRCC1; 584 (laplace), 1523 (linear) and 1018 (rbf) for EXOSC7 - XRCC6; and 1419 (laplace) and 876 (rbf) for EXOSC7 - XRCC6BP1. EXOSC8 was found to be down regulated w.r.t XRCC-1. These are reflected in rankings of 1373 (laplace) and 1515 (linear) for EXOSC8 - XRCC1; EXOSC9 was found to be down regulated w.r.t XRCC-1/2/6/6BP1. These are reflected in rankings of 44 (laplace), 1214 (linear) and 1410 (rbf) for EXOSC9 - XRCC1; 496 (laplace), 672 (linear) and 840 (rbf) for EXOSC9 - XRCC2; 1121 (laplace), 151 (linear) and 689 (rbf) for EXOSC9 - XRCC6 and 362 (laplace), 463 (linear) and 1741 (rbf) for EXOSC9 - XRCC6BP1.
On the right half of the Table is the rankings of XRCC w.r.t EXOSC family. W.r.t EXOSC2, XRCC-2 was found to be down regulated. These are reflected in rankings of 166 (laplace), 417 (linear) and 56 (rbf) for EXOSC2 - XRCC2. W.r.t W.r.t EXOSC3, XRCC-2 was found to be down regulated. These are reflected in rankings of 166 (laplace), 417 (linear) and 56 (rbf) for EXOSC3 - XRCC2. W.r.t EXOSC5, XRCC-2 was found to be down regulated. These are reflected in rankings of 1559 (laplace) and 56 (rbf) for EXOSC5 - XRCC2. W.r.t EXOSC6, XRCC-1/2/6/6BP1 were found to be down regulated. These are reflected in rankings of 509 (laplace) and 1046 (rbf) for EXOSC6 - XRCC1; 486 (laplace) and 1901 (rbf) for EXOSC6 - XRCC2; 35 (linear) and 188 (rbf) for EXOSC6 - XRCC6; 1295 (linear) and 366 (rbf) for EXOSC6 - XRCC6BP1. W.r.t EXOSC7, XRCC-6 was found to be down regulated. These are reflected in rankings of 1229 (linear) and 987 (rbf) for EXOSC7 - XRCC1; 176 (laplace), 436 (linear) and 788 (rbf) for EXOSC7 - XRCC2; and 1074 (laplace), 242 (linear) and 288 (rbf) for EXOSC7 - XRCC6. W.r.t EXOSC8, XRCC-2 was found to be down regulated. These are reflected in rankings of 13 (laplace) and 6 (rbf) for EXOSC8 - XRCC2. W.r.t EXOSC9, XRCC-2 was found to be down regulated. These are reflected in rankings of 655 (linear) and 1526 (rbf) for EXOSC9 - XRCC2 and 1206 (laplace) and 1626 (rbf) for EXOSC9 - XRCC6BP1;
Table 65 shows the derived influences which can be represented graphically, with the following influences - • XRCC w.r.t EXOSC with EXOSC-2 -> XRCC-2; EXOSC-3 -> XRCC-2; EXOSC-5 -> XRCC-2; EXOSC-6 -> XRCC-6; EXOSC-7 -> XRCC-1/2/6; EXOSC-8 -> XRCC-2; EXOSC-9 -> XRCC-2/6BP1; and • EXOSC w.r.t XRCC with EXOSC-2 <- XRCC-1/2/6/6BP1; EXOSC-3 <- XRCC-6/6BP1; EXOSC-5 <- XRCC-1/2/6/6BP1; EXOSC-6 <- XRCC-1/2; EXOSC-7 <- XRCC-1/6/6BP1; EXOSC-8 <- XRCC-1; and EXOSC-9 <- XRCC-1/2/6/6BP1.
3.4.7. RAD - FANC Cross Family Analysis
Fanconi Anemia (FA) is rare genetic disorder that happens mainly due to defects in proteins responsible for DNA repair via homologous recombination (Walden and Deans [144]). Cohn and D’Andrea [145] provides a review on the recent discoveries in the Fanconi Anemia and DNA double-strand break (DSB) repair pathways, which underscore the importance of regulated chromatin loading in the DNA damage response. Romick-Rosendale et al. [146] study the role Fanconi anemia pathway in squamous Cell Carcinoma. A review of the interplay between Fanconi anemia and homologous recombination pathways in genome integrity has been conducted by Michl et al. [147]. Liang et al. [148] observe the role of trimeric RAD51 and RAD51AP1-UAF1 complex in FANCD2. Taniguchi et al. [149] observe S-phase-specific interaction of the Fanconi anemia protein, FANCD2, with BRCA1 and RAD51. Zadorozhny et al. [150] show Fanconi anemia associated mutations destabilize RAD51 filaments and impair replication fork protection. Geng et al. [151] find RAD18-mediated ubiquitination of PCNA activates the Fanconi anemia DNA repair network. Rad18 E3 ubiquitin ligase activity mediates Fanconi anemia pathway activation and cell survival following DNA topoisomerase 1 inhibition as shown by Palle and Vaziri [152]. García-Luis and Machín [153] observe that Fanconi anaemia-like Mph1 helicase backs up RAD54 and RAD5 to circumvent replication stress-driven chromosome bridges. These findings suggest deep interactive role between the RAD and FA family. In colorectal cancer cell treated with ETC-1922159 these were found to both families were found to be down regulated. Our search engine alloted low laved numerical ranks to many of the 2nd order combinations between the RAD - FANC family. This signifies possible synergistic mechanism between the two in CRC cells. Table 67 shows the rankings of each, with respect to the other. On the left half is the rankings of RAD family w.r.t FANC family and vice versa on the right half.
On the left half, we find, RAD-18/51/51AP1/51C/54B/54L were found to be down regulated w.r.t FANCB. These are reflected in rankings of 10 (laplace), 2219 (linear) and 625 (rbf) for RAD18 - FANCB; 247 (laplace), 73 (linear) and 610 (rbf) for RAD51 - FANCB; 479 (laplace), 1667 (linear) and 663 (rbf) for RAD51AP1 - FANCB; 769 (laplace), 536 (linear) and 887 (rbf) for RAD51C - FANCB; 468 (laplace), 133 (linear) and 438 (rbf) for RAD54B - FANCB; and 583 (laplace), 2131 (linear) and 160 (rbf) for RAD54L - FANCB. RAD-18/51/51AP1/54B/54L were found to be down regulated w.r.t FANCD2. These are reflected in rankings of 1035 (laplace), 1271 (linear) and 405 (rbf) for RAD18 - FANCD2; 885 (laplace) and 1383 (rbf) for RAD51 - FANCD2; 1734 (laplace), 644 (linear) and 1291 (rbf) for RAD51AP1 - FANCD2; 275 (laplace), 2460 (linear) and 478 (rbf) for RAD54B - FANCD2; and 493 (laplace) and 203 (rbf) for RAD54L - FANCD2; RAD-1/18/50/51/51C/54B/54L were found to be down regulated w.r.t FANCD2OS. These are reflected in rankings of 693 (laplace) and 1146 (rbf) for RAD1 - FANCD2OS; 1472 (laplace), 526 (linear) and 239 (rbf) for RAD18 - FANCD2OS; 178 (laplace) and 1534 (linear) for RAD50 - FANCD2OS; 1080 (linear) and 1226 (rbf) for RAD51 - FANCD2OS; 1297 (laplace), 977 (linear) and 1237 (rbf) for RAD51C - FANCD2OS; 475 (laplace), 1367 (linear) for RAD54B - FANCD2OS; 1227 (linear) and 252 (rbf) for RAD54L - FANCD2OS; RAD-1/18/50/51/51AP1/51C/54B/54L were found to be down regulated w.r.t FANCF. These are reflected in rankings of 1582 (linear) and 285 (rbf) for RAD1 - FANCF; 770 (laplace), 1329 (linear) and 1445 (rbf) for RAD18 - FANCF; 1403 (laplace), 1684 (linear) and 803 (rbf) for RAD50 - FANCF; 209 (laplace), 1247 (linear) for RAD51 - FANCF; 1681 (laplace), 13 (linear) for RAD51AP1 - FANCF; 1493 (laplace) and 224 (linear) for RAD51C - FANCF; 401 (laplace) and 143 (linear) for RAD54B - FANCF; for 690 (laplace), 829 (linear) for RAD54L - FANCF; RAD-1/18/50/51/51AP1/51C/54B/54L were found to be down regulated w.r.t FANCG. These are reflected in rankings of 755 (laplace), 393 (linear) and 82 (rbf) for RAD18 - FANCG; 345 (laplace), 114 (linear) and 295 (rbf) for RAD51 - FANCG; 957 (laplace), 218 (linear) and 1360 (rbf) for RAD51C - FANCG; 17 (laplace), 182 (linear) and 423 (rbf) for RAD54B - FANCG; and 1058 (laplace), 701 (linear) and 581 (rbf) for RAD54L - FANCG. RAD-18/50/51/51C/54B/54L were found to be down regulated w.r.t FANCG. These are reflected in rankings of 1693 (laplace)and 436 (rbf) for RAD18 - FANCI; 1703 (laplace) and 1458 (rbf) for RAD50 - FANCI; 1038 (laplace), 1668 (linear) and 310 (rbf) for RAD51 - FANCI; 597 (laplace) and 165 (linear) for RAD51C - FANCI; 557 (laplace) and 84 (linear) for RAD54B - FANCI; and 468 (laplace), 606 (linear) for RAD54L - FANCI.
Table 66.
2nd order combinatorial hypotheses between RAD and FANC members.
| Ranking RAD family VS FANC family | |||||||
| Ranking of RAD family w.r.t FANCB | Ranking of FANCB w.r.t RAD family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| RAD1 - FANCB | 2431 | 400 | 2553 | RAD1 - FANCB | 1499 | 656 | 340 |
| RAD18 - FANCB | 10 | 2219 | 625 | RAD18 - FANCB | 2708 | 383 | 2298 |
| RAD50 - FANCB | 2419 | 915 | 2556 | RAD50 - FANCB | 133 | 234 | 73 |
| RAD51 - FANCB | 247 | 73 | 610 | RAD51 - FANCB | 2444 | 378 | 8 |
| RAD51AP1 - FANCB | 479 | 1667 | 663 | RAD51AP1 - FANCB | 89 | 562 | 2 |
| RAD51C - FANCB | 769 | 536 | 887 | RAD51C - FANCB | 460 | 187 | 86 |
| RAD54B - FANCB | 468 | 133 | 438 | RAD54B - FANCB | 486 | 891 | 568 |
| RAD54L - FANCB | 583 | 2131 | 160 | RAD54L - FANCB | 41 | 2675 | 692 |
| Ranking of RAD family w.r.t FANCD2 | Ranking of FANCD2 w.r.t RAD family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| RAD1 - FANCD2 | 1935 | 332 | 2102 | RAD1 - FANCD2 | 1451 | 1605 | 796 |
| RAD18 - FANCD2 | 1035 | 1271 | 405 | RAD18 - FANCD2 | 2356 | 403 | 1299 |
| RAD50 - FANCD2 | 2109 | 436 | 2038 | RAD50 - FANCD2 | 646 | 357 | 769 |
| RAD51 - FANCD2 | 885 | 1995 | 1383 | RAD51 - FANCD2 | 591 | 1938 | 85 |
| RAD51AP1 - FANCD2 | 1734 | 644 | 1291 | RAD51AP1 - FANCD2 | 993 | 603 | 2684 |
| RAD51C - FANCD2 | 54 | 2399 | 2566 | RAD51C - FANCD2 | 629 | 656 | 620 |
| RAD54B - FANCD2 | 275 | 2460 | 478 | RAD54B - FANCD2 | 227 | 230 | 131 |
| RAD54L - FANCD2 | 493 | 2530 | 203 | RAD54L - FANCD2 | 2457 | 1369 | 1816 |
| Ranking of RAD family w.r.t FANCD2OS | Ranking of FANCD2OS w.r.t RAD family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| RAD1 - FANCD2OS | 693 | 1926 | 1146 | RAD1 - FANCD2OS | 1455 | 2445 | 1624 |
| RAD18 - FANCD2OS | 1472 | 526 | 239 | RAD18 - FANCD2OS | 851 | 1457 | 653 |
| RAD50 - FANCD2OS | 178 | 1534 | 2141 | RAD50 - FANCD2OS | 1763 | 1477 | 1372 |
| RAD51 - FANCD2OS | 2061 | 1080 | 1226 | RAD51 - FANCD2OS | 2007 | 2336 | 1739 |
| RAD51AP1 - FANCD2OS | 637 | 2050 | 2660 | RAD51AP1 - FANCD2OS | 2209 | 2376 | 1722 |
| RAD51C - FANCD2OS | 1297 | 977 | 1237 | RAD51C - FANCD2OS | 1729 | 779 | 2596 |
| RAD54B - FANCD2OS | 475 | 1367 | 2571 | RAD54B - FANCD2OS | 2032 | 1241 | 1637 |
| RAD54L - FANCD2OS | 2557 | 1227 | 252 | RAD54L - FANCD2OS | 1671 | 1830 | 1839 |
| Ranking of RAD family w.r.t FANCF | Ranking of FANCF w.r.t RAD family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| RAD1 - FANCF | 1817 | 1582 | 285 | RAD1 - FANCF | 529 | 2198 | 1997 |
| RAD18 - FANCF | 770 | 1329 | 1445 | RAD18 - FANCF | 1063 | 2186 | 196 |
| RAD50 - FANCF | 1403 | 1684 | 803 | RAD50 - FANCF | 2205 | 1419 | 1676 |
| RAD51 - FANCF | 209 | 1247 | 2221 | RAD51 - FANCF | 1222 | 1060 | 2251 |
| RAD51AP1 - FANCF | 1681 | 13 | 2619 | RAD51AP1 - FANCF | 1963 | 2372 | 107 |
| RAD51C - FANCF | 1493 | 224 | 2051 | RAD51C - FANCF | 2062 | 1904 | 2386 |
| RAD54B - FANCF | 401 | 143 | 2359 | RAD54B - FANCF | 1903 | 1936 | 2026 |
| RAD54L - FANCF | 690 | 829 | 2120 | RAD54L - FANCF | 2529 | 716 | 1262 |
| Ranking of RAD family w.r.t FANCG | Ranking of FANCG w.r.t RAD family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| RAD1 - FANCG | 2013 | 2215 | 2328 | RAD1 - FANCG | 1938 | 825 | 843 |
| RAD18 - FANCG | 755 | 393 | 82 | RAD18 - FANCG | 2352 | 878 | 2574 |
| RAD50 - FANCG | 2652 | 2408 | 2663 | RAD50 - FANCG | 695 | 511 | 933 |
| RAD51 - FANCG | 345 | 114 | 295 | RAD51 - FANCG | 2163 | 1 | 397 |
| RAD51AP1 - FANCG | 1743 | 749 | 1984 | RAD51AP1 - FANCG | 661 | 400 | 23 |
| RAD51C - FANCG | 957 | 218 | 1360 | RAD51C - FANCG | 450 | 2319 | 1122 |
| RAD54B - FANCG | 17 | 182 | 423 | RAD54B - FANCG | 140 | 194 | 64 |
| RAD54L - FANCG | 1058 | 701 | 581 | RAD54L - FANCG | 2167 | 1968 | 2344 |
| Ranking of RAD family w.r.t FANCI | Ranking of FANCI w.r.t RAD family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| RAD1 - FANCI | 1919 | 2263 | 2286 | RAD1 - FANCI | 2496 | 897 | 664 |
| RAD18 - FANCI | 1693 | 2466 | 436 | RAD18 - FANCI | 1601 | 1161 | 1668 |
| RAD50 - FANCI | 1703 | 2074 | 1458 | RAD50 - FANCI | 1133 | 1211 | 1238 |
| RAD51 - FANCI | 1038 | 1668 | 310 | RAD51 - FANCI | 1612 | 2724 | 1187 |
| RAD51AP1 - FANCI | 2496 | 2517 | 383 | RAD51AP1 - FANCI | 1513 | 1211 | 65 |
| RAD51C - FANCI | 597 | 165 | 2447 | RAD51C - FANCI | 143 | 137 | 87 |
| RAD54B - FANCI | 557 | 84 | 2055 | RAD54B - FANCI | 178 | 350 | 76 |
| RAD54L - FANCI | 468 | 606 | 2461 | RAD54L - FANCI | 211 | 2304 | 1128 |
On the right half, we find, FANCB to be down regulated w.r.t RAD-1/50/51/51AP1/51C/54B/54L. These are reflected in rankings of 1499 (laplace), 656 (linear) and 340 (rbf) for RAD1 - FANCB; 133 (laplace), 234 (linear) and 73 (rbf) for RAD50 - FANCB; 378 (linear) and 8 (rbf) for RAD51 - FANCB; 89 (laplace), 562 (linear) and 2 (rbf) for RAD51AP1 - FANCB; 460 (laplace), 187 (linear) and 86 (rbf) for RAD51C - FANCB; 486 (laplace), 891 (linear) and 568 (rbf) for RAD54B - FANCB and 41 (laplace) and 692 (rbf) for RAD54L - FANCB; FANCD2 was found to be down regulated w.r.t RAD-1/50/51/51AP1/51C/54B/54L. These are reflected in rankings of 1451 (laplace), 1605 (linear) and 796 (rbf) for RAD1 - FANCD2; 403 (linear) and 1299 (rbf) for RAD18 - FANCD2; 646 (laplace), 357 (linear) and 769 (rbf) for RAD50 - FANCD2; 591 (laplace) and 85 (rbf) for RAD51 - FANCD2; 993 (laplace) and 603 (linear) for RAD51AP1 - FANCD2; 629 (laplace), 656 (linear) and 620 (rbf) for RAD51C - FANCD2; 227 (laplace), 230 (linear) and 131 (rbf) for RAD54B - FANCD2. FANCD2OS2 was found to be down regulated w.r.t RAD-1/18/5051C/54B. These are reflected in rankings of 1455 (laplace) and 1624 (rbf) for RAD1 - FANCD2OS; 851 (laplace), 1457 (linear) and 653 (rbf) for RAD18 - FANCD2OS; 1477 (linear) and 1372 (rbf) for RAD50 - FANCD2OS; 1729 (laplace) and 779 (linear) for RAD51C - FANCD2OS; 1241 (linear) and 1637 (rbf) for RAD54B - FANCD2OS; FANCF was found to be down regulated w.r.t RAD-1/18/50/51C/54B. These are reflected in rankings of 1063 (laplace) and 196 (rbf) for RAD18 - FANCF; 1419 (linear) and 1676 (rbf) for RAD50 - FANCF; 1222 (laplace) and 1060 (linear) for RAD51 - FANCF; and 716 (linear) and 1262 (rbf) for RAD54L - FANCF; FANCG was found to be down regulated w.r.t RAD-1/50/51/51AP1/51C/54B. These are reflected in rankings of 825 (linear) and 843 (rbf) for RAD1 - FANCG; 695 (laplace), 511 (linear) and 933 (rbf) for RAD50 - FANCG; 1 (linear) and 397 (rbf) for RAD51 - FANCG; 661 (laplace), 400 (linear) and 23 (rbf) for RAD51AP1 - FANCG; 450 (laplace) and 1122 (rbf) for RAD51C - FANCG; 140 (laplace), 194 (linear) and 64 (rbf) for RAD54B - FANCG; FANCI was found to be down regulated w.r.t RAD-1/18/50/51/51AP1/51C/54B/54L. These are reflected in 897 (linear) and 664 (rbf) for RAD1 - FANCI; 1601 (laplace), 1161 (linear) and 1668 (rbf) for RAD18 - FANCI; 1133 (laplace), 1211 (linear) and 1238 (rbf) for RAD50 - FANCI; 1612 (laplace) and 1187 (rbf) for RAD51 - FANCI; 1513 (laplace), 1211 (linear) and 65 (rbf) for RAD51AP1 - FANCI; 143 (laplace), 137 (linear) and 87 (rbf) for RAD51C - FANCI; 178 (laplace), 350 (linear) and 76 (rbf) for RAD54B - FANCI; 211 (laplace) and 1128 (rbf) for RAD54L - FANCI.
Table 67 shows the derived influences which can be represented graphically, with the following influences - • RAD w.r.t FANC with RAD-18/51/51AP1/51C/54B/54L <- FANCB; RAD-18/51/51AP1/54B/54L <- FANCD2; RAD-1/18/50/51/51C/54B/54L <- FANCD2OS; RAD-1/18/50/51/51AP1/51C/54B/54L <- FANCF; RAD-1/18/50/51/51AP1/51C/54B/54L <- FANCG; and RAD-18/50/51/51C/54B/54L <- FANCI, and • FANC w.r.t RAD with FANCB <- RAD-1/50/51/51AP1/51C/54B/54L; FANCD2 <- RAD-1/50/51/51AP1/51C/54B/54L; FANCD2OS <- RAD-1/18/5051C/54B; FANCF <- RAD-1/18/50/51C/54B; FANCG <- RAD-1/50/51/51AP1/51C/54B; FANCI <- RAD-1/18/50/51/51AP1/51C/54B/54L;
3.5. Telomerase Related Synergies
3.5.1. TERT - ABC Transporters Cross Family Analysis
TERT and ABC family members found to be down regulated after ETC-1922159 treatment in CRC cells. Not much is known about the TERT and ABC transporters and research is still ongoing regarding the synergy of TERT and ABC transporters. The most recent work on telomerase and drug resistance in cancer by Lipinska et al. [154] talks on a range of theories about the mechanism of inactivation of telomerase in cancer cells that is accompanied by relatively increased sensitivity to some drugs. These mechanism has not been fully understood. Some association with the telomerase expression and drug resistance has been shown by Wang et al. [155] while no correlation between the two has been indicated by Sakin et al. [156]. However Keshet et al. [157], show a deep correlation in melanoma cells revealing co-expression of ABC transporters, ABCB5 and ABCC2 and hTERT. Based on these little known associations the search engine was able to rank the combinations of some of the members of ABC with TERT. Table 68 shows the rankings of TERT and ABC members w.r.t to each other. On the left half, we find ABC family to be down regulated w.r.t TERT. These are reflected in rankings of 381 (laplace), 1047 (linear) and 316 (rbf) for ABCF2 - TERT; 1201 (laplace), 49 (linear) and 317 (rbf) for ABCA2 - TERT; and 1613 (laplace), 499 (linear) and 1217 (rbf) for ABCE1 - TERT. On the right side we find TERT to be down regulated w.r.t ABCE1. These are reflected in 120 (laplace), 2736 (linear) and 294 (rbf) for ABCE1 - TERT.
So if we look at the above rankings, what we find is that the ABC family is down regulated along with TERT, synergistically (directly or indirectly) with moderate and high promise (rankings nearing to 1) in the top table. Vice versa, the same affect is not shown in Table 69 for ABC-F2/A2 with TERT. If we look at the 2 way cross analysis what we find is the following combinatorial hypotheses in Table 69 which is graphicaly reflected as - • ABC family w.r.t TRET with TERT -> ABCF2; TERT -> ABCA2; and TERT -> ABCE1 and • TRET w.r.t ABC family with ABCE1 -> TERT. Consequently, it is possible that the TERT does have influence over ABC-F2/A2 but with ABCE1, directionality could not be established. Further more, these low rankings point to high promise of down regulation that is observed in CRC treated with ETC-1922159. Which might mean that in CRC cells which have not been treated with ETC-1922159, it is highly possible that TERT is highly up regulated and also bolsters/influences the functioning of ABC transporters. Wet lab study and further experiments will be needed to establish the dual role of TERT and ABC transporters.
3.6. ABC Transporter Related Synergies
3.6.1. ABC Transporters - UBE2 Cross Family Analysis
Not much is known about the interaction or any possible direct/indirect synergy of ABC transporters and the Ubiquitin-conjugating enzyme E2 family. In CRC cells treated with ETC-1922159, family members of both were found to be up regulated. The search engine also assigned numerically high valued ranks to a few of 2nd order synergies between the the two. We document here these synergies and show the possible unexplored combinations between the two familes. Table 70 and Table 71 show the rankings of ABC w.r.t UBE2 and vice versa, respectively.
In Table 70 we found ABC-C3 up regulated w.r.t UBE2-A. This is reflected in the rankings of 2137 (laplace) and 2491 (linear) for ABC-C3 - UBE2-A. ABC-C5 was up regulated w.r.t UBE2-B. This is reflected in the rankings of 2317 (laplace) and 2266 (rbf) for ABC-C5 - UBE2-B. ABC-A5/D1/G2 were up regulated w.r.t UBE2-F. These are reflected in the rankings of 2408 (linear) and 1784 (rbf) for ABC-A5 - UBE2-F, 2069 (linear) and 1959 (rbf) for ABC-D1 - UBE2-F and 1995 (linear) and 2120 (rbf) for ABC-G2 - UBE2-F. ABC-A5/C13 were up regulated w.r.t UBE2-H. These are reflected in 2068 (linear) and 2438 (rbf) for ABC-A5 - UBE2-H and 2492 (linear) and 2051 (rbf) for ABC-C13 - UBE2-H. ABC-C13 was up regulated w.r.t UBE2-J1. This is reflected in the rankings of 2095 (laplace), 2412 (linear) and 2360 (rbf). ABC-C5 was up regulated w.r.t UBE2-Z. This is reflected in rankings of 2348 (laplace) and 1859 (rbf) for ABC-C5 - UBE2-Z.
In Table 71 we found UBE2-A up regulated w.r.t ABC-C5/G2. This is reflected in the rankings of 2122 (linear) and 2297 (rbf) for ABC-C5 - UBE2-A; and 2048 (laplace) and 1829 (linear) for ABC-G2 - UBE2-A. UBE2-B up regulated w.r.t ABC-A5/C3/C13/D1/G2. This is reflected in the rankings of 1846 (laplace) and 2038 (linear) for ABC-A5 - UBE2-B; 1999 (laplace) and 2050 (rbf) for ABC-C3 - UBE2-B; 1863 (linear) and 2496 (rbf) for ABC-C13 - UBE2-B; 2322 (laplace), 1917 (linear) and 2426 (rbf) for ABC-D1 - UBE2-B and 1833 (laplace), 2445 (linear) and 2506 (rbf) for ABC-G2 - UBE2-B. UBE2-F was found up regulated w.r.t ABC-B11/C3/D1/G1. These were reflected in 2003 (laplace) and 2422 (rbf) for ABC-B11 - UBE2-F; 2132 (laplace) and 2163 (linear) for ABC-C3 - UBE2-F; 2421 (laplace) and 2176 (rbf) for ABC-D1 - UBE2-F; and 2202 (laplace) and 1953 (rbf) for ABC-G1 - UBE2-F. UBE2-H was found to be up regulated w.r.t ABC-B11/C3/C5/C13/G1. These are reflected in rankings of 1950 (laplace), 1770 (linear) and 2461 (rbf) for ABC-B11 - UBE2-H; 2439 (laplace), 1972 (linear) and 2305 (rbf) for ABC-C3 - UBE2-H; 2473 (linear) and 2355 (rbf) for ABC-C5 - UBE2-H; 2004 (laplace), 2317 (linear) and 1847 (rbf) for ABC-C13 - UBE2-H; and 1921 (linear) and 2288 (rbf) for ABC-G1 - UBE2-H; UBE2-J1 was found to be up regulated w.r.t ABC-B11/C3/C13/D1/G1/G2; 1806 (laplace) and 1935 (rbf) for ABC-B11 - UBE2-J1; 2073 (laplace) and 2291 (linear) for ABC-C3 - UBE2-J1; 2329 (laplace), 2153 (linear) and 1951 (rbf) ABC-C13 - UBE2-J1; 2263 (laplace), 1886 (linear) and 2249 (rbf) for ABC-D1 - UBE2-J1; and 2418 (linear) and 2277 (rbf) for ABC-G1 - UBE2-J1; Finally, UBE2-Z was found up regulated w.r.t ABC-C3/D1/G1/G2. These are reflected in rankings of 1978 (laplace), 1823 (linear) and 1859 (rbf) for ABC-C3 - UBE2-Z; 2292 (laplace) and 2381 (linear) for ABC-D1 - UBE2-Z; 2515 (linear) and 1858 (rbf) for ABC-G1 - UBE2-Z; 2270 (laplace), 2080 (linear) and 2448 (rbf) for ABC-G2 - UBE2-Z.
Table 72 shows the derived influences which can be represented graphically, with the following influences - • ABC w.r.t UBE2 with ABC-C3 <- UBE2-A; ABC-C5 <- UBE2-B; ABC-A5/D1/G2 <- UBE2-F; ABC-A5/C13 <- UBE2-H; ABC-C13 <- UBE2-J1; ABC-C5 <- UBE2-Z; and • UBE2 w.r.t ABC with UBEA-2 <- ABC-C5/G2; UBE2-B <- ABC-A5/C3/C13/D1/G2; UBE2-F <- ABC-B11/C3/D1/G1; UBE2-H <- ABC-B11/C3/C5/C13/G1; UBE2-J1 <- ABC-B11/C3/C13/D1/G1/G2; UBE2-Z <- ABC-C3/D1/G1/G2.
3.6.2. ABC Transporters Intra Cross Family Analysis
A range of ABC transporters were found to be up regulated in CRC cells after ETC-1922159 treatment. We checked the rankings of the ABC transporters within the ABC family and found multiple synergistic upregulation at 2nd order level that were ranked appropriately. Table 73 shows intra family rankings of ABC members among themselves. We found ABC-C13 upregulated w.r.t ABC-A5. These were reflected in rankings of 1943 (linear) and 2151 (rbf); ABC-C5/C13/G1 were up regulated w.r.t ABC-B11. These are reflected in rankings of 2226 (laplace) and 2241 (rbf) for ABC-C5 - ABC-B11; 1971 (laplace) and 2150 (rbf) for ABC-C13 - ABC-B11 and 1957 (laplace) and 1920 (linear) for ABC-G1 - ABC-B11; ABC-C3/C13 were found to be up regulated w.r.t ABC-C5. These are reflected in 2084 (laplace), 2274 (linear) and 1758 (rbf) for ABC-C3 - ABC-C5 and 2476 (linear) and 2446 (rbf) for ABC-C13 - ABC-C5. ABC-C5/C13 were found to be up regulated w.r.t ABC-D1. 2423 (laplace) and 2388 (rbf) for ABC-C5 - ABC-D1 and 2383 (laplace) and 2029 (linear) for ABC-C13 - ABC-D1. ABC-A5 was found to be up regulated w.r.t ABC-G1. This is reflected in rankings of 2488 (laplace) and 1776 (linear) for ABC-A5 - ABC-G1. ABC-A5 was found to be up regulated w.r.t ABC-G2 also. This is reflected in rankings of 2284 (laplace), 1904 (linear) and 1829 (rbf) for ABC-A5 - ABC-G2.
Table 74 shows the derived influences which can be represented graphically, with the following influences - • ABC intra family with ABC-C13 <- ABC-A5; ABC-C5/C13/G1 <- ABC-B11; ABC-C3/C13 <- ABC-C5; ABC-C5/C13 <- ABC-D1; ABC-A5 <- ABC-G1; ABC-C5 <- ABC-G2.
3.6.3. Interleukin - ABC Transporters Cross Family Analysis
Zhou et al. [158] have observed that the ABCA1 contributes to the secretion of interleukin 1β from macrophages. Haskó et al. [159] show that inhibitors of ABC transporters suppress interleukin-12 p40 production and major histocompatibility complex II up-regulation in macrophages. Park et al. [160] conclude that anti-cancer drug-induced IL-8 secretion increased the expression of ABC transporters and SP cells, promoting the growth of HCC in vitro. Marty et al. [161] show that ABC1 is required for the release of interleukin-1β by P2X7-stimulated and lipopolysaccharide-primed mouse Schwann cells. Lottaz et al. [162] observe that inhibition of ABC transporter downregulates interleukin-1β-mediated autocrine activation of human dermal fibroblasts. These findings and many more indicate the synergy between IL family and ABC transporters. In colorectal cancer cells treated with ETC-1922159, some of the members of both families were up regulated. Given the studied synergies, our search engine found multiple combinations which were ranked with high numerical values, thus indicating possible dual combinatorial role. Table 75 and Table 76, each show rankings of ABC transporters w.r.t IL family on the left half and vice versa on the right half.
On the left half we found IL-17REL up regulated w.r.t ABCA5. This is reflected in the rankings of 2405 (linear) and 2202 (rbf) for IL17REL - ABCA5. IL-2RG/6ST/15/15RA up regulated w.r.t ABCB11. This is reflected in the rankings of 2182 (laplace), 2102 (linear) and 550 (rbf) for IL2RG - ABCB11; 1793 (laplace), 2140 (linear) and 1938 (rbf) for IL6ST - ABCB11; 2438 (laplace) and 2512 (linear) for IL15 - ABCB11; and 2271 (laplace) and 1784 (rbf) for IL15RA - ABCB11. IL-8/15RA up regulated w.r.t ABCC3. This is reflected in the rankings of 1767 (laplace) and 2419 (rbf) for IL8 - ABCC3 and 2403 (linear) and 1795 (rbf) for IL15RA - ABCC3. IL-15RA/17REL up regulated w.r.t ABCC5. These are reflected in rankings of 2255 (linear) and 1861 (rbf) for IL15RA - ABCC5 and 2462 (linear) and 2509 (rbf) for IL17REL - ABCC5. IL-15RA/17REL were up regulated w.r.t ABCC13. These are reflected in 2248 (laplace), 1955 (linear) and 2456 (rbf) for IL15RA - ABCC13 and 2339 (laplace) and 2137 (linear) for IL17REL - ABCC13. IL-1A/1RAP/8/15RA were up regulated w.r.t ABCD1. These are reflected in rankings of 1932 (laplace) and 2203 (rbf) for IL1A - ABCD1; 2508 (laplace), 2006 (linear) and 1907 (rbf) for IL1RAP - ABCD1; 2010 (laplace), 2315 (linear) and 1814 (rbf) for IL8 - ABCD1; and 2097 (laplace) and 1765 (linear) for IL15RA - ABCD1. IL-1RAP was up regulated w.r.t ABCG1. This was reflected in rankings of 2205 (linear) and 2339 (rbf) for IL1RAP - ABCG1. IL-1RAP/15RA were up regulated w.r.t ABCG2. These were reflected in rankings of 2184 (laplace) and 2167 (linear) for IL1RAP - ABCG2 and 1910 (laplace), 2428 (linear) and 1921 (rbf) for IL15RA - ABCG2.
Table 75.
2nd order interaction ranking between ABC and IL family members.
| Ranking ABC family VS IL family | |||||||
| Ranking of IL family w.r.t ABCA5 | Ranking of ABCA5 family w.r.t IL | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - ABCA5 | 705 | 95 | 11 | IL1A - ABCA5 | 677 | 2069 | 871 |
| IL1B - ABCA5 | 240 | 35 | 353 | IL1B - ABCA5 | 2069 | 790 | 2301 |
| IL1RAP - ABCA5 | 1515 | 2354 | 514 | IL1RAP - ABCA5 | 1763 | 335 | 2345 |
| IL1RN - ABCA5 | 771 | 1093 | 1417 | IL1RN - ABCA5 | 892 | 2252 | 1482 |
| IL2RG - ABCA5 | 500 | 246 | 173 | IL2RG - ABCA5 | 993 | 750 | 1745 |
| IL6ST - ABCA5 | 2464 | 1564 | 1365 | IL6ST - ABCA5 | 155 | 266 | 1386 |
| IL8 - ABCA5 | 1676 | 1568 | 1111 | IL8 - ABCA5 | 104 | 1261 | 946 |
| IL10RB - ABCA5 | 492 | 146 | 643 | IL10RB - ABCA5 | 2230 | 2184 | 2240 |
| IL15 - ABCA5 | 638 | 1169 | 65 | IL15 - ABCA5 | 661 | 169 | 711 |
| IL15RA - ABCA5 | 2151 | 1672 | 740 | IL15RA - ABCA5 | 706 | 1300 | 2031 |
| IL17C - ABCA5 | 680 | 197 | 164 | IL17C - ABCA5 | 615 | 575 | 1518 |
| IL17REL - ABCA5 | 1014 | 2405 | 2202 | IL17REL - ABCA5 | 212 | 1024 | 146 |
| Ranking of IL family w.r.t ABCB11 | Ranking of ABCB11 family w.r.t IL | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - ABCB11 | 1962 | 465 | 648 | IL1A - ABCB11 | 551 | 140 | 385 |
| IL1B - ABCB11 | 1778 | 851 | 438 | IL1B - ABCB11 | 255 | 428 | 208 |
| IL1RAP - ABCB11 | 1427 | 1704 | 1318 | IL1RAP - ABCB11 | 1681 | 878 | 1709 |
| IL1RN - ABCB11 | 1832 | 539 | 297 | IL1RN - ABCB11 | 342 | 1912 | 779 |
| IL2RG - ABCB11 | 2182 | 2102 | 550 | IL2RG - ABCB11 | 814 | 67 | 584 |
| IL6ST - ABCB11 | 1793 | 2140 | 1938 | IL6ST - ABCB11 | 1347 | 1504 | 385 |
| IL8 - ABCB11 | 1607 | 2441 | 1028 | IL8 - ABCB11 | 349 | 846 | 1786 |
| IL10RB - ABCB11 | 341 | 1119 | 449 | IL10RB - ABCB11 | 2101 | 2419 | 1352 |
| IL15 - ABCB11 | 2438 | 2512 | 576 | IL15 - ABCB11 | 344 | 224 | 256 |
| IL15RA - ABCB11 | 2271 | 1288 | 1784 | IL15RA - ABCB11 | 1052 | 48 | 719 |
| IL17C - ABCB11 | 1262 | 69 | 706 | IL17C - ABCB11 | 653 | 316 | 437 |
| IL17REL - ABCB11 | 50 | 305 | 783 | IL17REL - ABCB11 | 1004 | 736 | 896 |
| Ranking of IL family w.r.t ABCC3 | Ranking of ABCC3 family w.r.t IL | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - ABCC3 | 1860 | 758 | 1538 | IL1A - ABCC3 | 1343 | 1798 | 2459 |
| IL1B - ABCC3 | 1764 | 749 | 896 | IL1B - ABCC3 | 1647 | 1369 | 569 |
| IL1RAP - ABCC3 | 1514 | 2294 | 1989 | IL1RAP - ABCC3 | 2074 | 1377 | 303 |
| IL1RN - ABCC3 | 647 | 607 | 1252 | IL1RN - ABCC3 | 1366 | 975 | 1354 |
| IL2RG - ABCC3 | 990 | 444 | 40 | IL2RG - ABCC3 | 1229 | 379 | 844 |
| IL6ST - ABCC3 | 98 | 1589 | 339 | IL6ST - ABCC3 | 970 | 712 | 1342 |
| IL8 - ABCC3 | 1767 | 1046 | 2419 | IL8 - ABCC3 | 937 | 1033 | 430 |
| IL10RB - ABCC3 | 1354 | 78 | 359 | IL10RB - ABCC3 | 1609 | 29 | 1830 |
| IL15 - ABCC3 | 1580 | 602 | 1560 | IL15 - ABCC3 | 1087 | 1191 | 1084 |
| IL15RA - ABCC3 | 189 | 2403 | 1795 | IL15RA - ABCC3 | 2153 | 163 | 1324 |
| IL17C - ABCC3 | 1587 | 778 | 2425 | IL17C - ABCC3 | 466 | 631 | 2237 |
| IL17REL - ABCC3 | 1135 | 403 | 54 | IL17REL - ABCC3 | 2089 | 2388 | 1618 |
| Ranking of IL family w.r.t ABCC5 | Ranking of ABCC5 family w.r.t IL | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - ABCC5 | 2004 | 681 | 60 | IL1A - ABCC5 | 2217 | 2022 | 1512 |
| IL1B - ABCC5 | 1948 | 112 | 251 | IL1B - ABCC5 | 1223 | 2137 | 942 |
| IL1RAP - ABCC5 | 1038 | 355 | 2023 | IL1RAP - ABCC5 | 1982 | 1892 | 2296 |
| IL1RN - ABCC5 | 709 | 430 | 1087 | IL1RN - ABCC5 | 1668 | 816 | 2142 |
| IL2RG - ABCC5 | 1421 | 264 | 601 | IL2RG - ABCC5 | 500 | 2018 | 1691 |
| IL6ST - ABCC5 | 1569 | 2010 | 845 | IL6ST - ABCC5 | 754 | 2326 | 874 |
| IL8 - ABCC5 | 1869 | 143 | 1589 | IL8 - ABCC5 | 855 | 1211 | 2434 |
| IL10RB - ABCC5 | 1162 | 70 | 434 | IL10RB - ABCC5 | 1337 | 736 | 958 |
| IL15 - ABCC5 | 1262 | 147 | 389 | IL15 - ABCC5 | 1947 | 1991 | 1584 |
| IL15RA - ABCC5 | 1083 | 2255 | 1861 | IL15RA - ABCC5 | 2457 | 1444 | 534 |
| IL17C - ABCC5 | 2447 | 96 | 116 | IL17C - ABCC5 | 1836 | 845 | 1802 |
| IL17REL - ABCC5 | 54 | 2462 | 2509 | IL17REL - ABCC5 | 1247 | 2149 | 1031 |
On the right half we found ABCA5 up regulated w.r.t IL-1B/1RAP/10RB. These are reflected in the rankings of 2069 (laplace) and 2301 (rbf) for IL1B - ABCA5; 1763 (laplace) and 2345 (rbf) for IL1RAP - ABCA5; and 2230 (laplace), 2184 (linear) and 2240 (rbf) for IL10RB - ABCA5; ABCB11 was up regulated w.r.t IL-10RB. This is reflected in the rankings of 2101 (laplace) and 2419 (linear) for IL10RB - ABCB11. ABCC3 was up regulated w.r.t IL-1A/17REL. This is reflected in the rankings of 1798 (linear) and 2459 (rbf) for IL1A - ABCC3 and 2089 (laplace) and 2388 (linear) for IL17REL - ABCC3. ABCC5 was up regulated w.r.t IL-1A/1RAP/15/17C. This are reflected in the rankings of 2217 (laplace) and 2022 (linear) for IL1A - ABCC5; 1982 (laplace), 1892 (linear) and 2296 (rbf) for IL1RAP - ABCC5; 1947 (laplace) and 1991 (linear) for IL15 - ABCC5 and 1836 (laplace) and 1802 (rbf) for IL17C - ABCC5. ABCC13 was up regulated w.r.t IL-1RAP/15RA. This are reflected in the rankings of 2136 (laplace) and 2392 (linear) for IL1RAP - ABCC13 and 2397 (laplace) and 2485 (linear) for IL15RA - ABCC13; ABCD1 was up regulated w.r.t IL-8/10RB. This are reflected in the rankings of 2501 (laplace) and 2154 (linear) for IL8 - ABCD1 and 1795 (laplace) and 2325 (rbf) for IL10RB - ABCD1. ABCG2 was up regulated w.r.t IL-10RB. This is reflected in the rankings of 2144 (laplace), 2335 (linear) and 2434 (rbf) for IL10RB - ABCG2.
Table 76.
2nd order interaction ranking between ABC and IL family members.
| Ranking ABC family VS IL family | |||||||
| Ranking of IL family w.r.t ABCC13 | Ranking of ABCC13 family w.r.t IL | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - ABCC13 | 512 | 135 | 1207 | IL1A - ABCC13 | 464 | 352 | 201 |
| IL1B - ABCC13 | 152 | 103 | 1553 | IL1B - ABCC13 | 635 | 974 | 60 |
| IL1RAP - ABCC13 | 1092 | 502 | 2442 | IL1RAP - ABCC13 | 2136 | 2392 | 33 |
| IL1RN - ABCC13 | 1753 | 559 | 323 | IL1RN - ABCC13 | 114 | 1016 | 1839 |
| IL2RG - ABCC13 | 2064 | 674 | 1076 | IL2RG - ABCC13 | 807 | 1079 | 938 |
| IL6ST - ABCC13 | 332 | 1416 | 2112 | IL6ST - ABCC13 | 119 | 1098 | 2323 |
| IL8 - ABCC13 | 551 | 1200 | 1680 | IL8 - ABCC13 | 592 | 984 | 907 |
| IL10RB - ABCC13 | 631 | 621 | 561 | IL10RB - ABCC13 | 2011 | 1272 | 1297 |
| IL15 - ABCC13 | 502 | 296 | 373 | IL15 - ABCC13 | 612 | 968 | 170 |
| IL15RA - ABCC13 | 2248 | 1955 | 2456 | IL15RA - ABCC13 | 2397 | 2485 | 790 |
| IL17C - ABCC13 | 25 | 140 | 123 | IL17C - ABCC13 | 924 | 308 | 711 |
| IL17REL - ABCC13 | 2339 | 2137 | 1497 | IL17REL - ABCC13 | 462 | 376 | 461 |
| Ranking of IL family w.r.t ABCC5 | Ranking of ABCC5 family w.r.t IL | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - ABCD1 | 1932 | 30 | 2203 | IL1A - ABCD1 | 530 | 2046 | 1196 |
| IL1B - ABCD1 | 569 | 109 | 1778 | IL1B - ABCD1 | 1400 | 605 | 453 |
| IL1RAP - ABCD1 | 2508 | 2006 | 1907 | IL1RAP - ABCD1 | 399 | 840 | 1548 |
| IL1RN - ABCD1 | 606 | 2003 | 789 | IL1RN - ABCD1 | 551 | 2025 | 60 |
| IL2RG - ABCD1 | 1064 | 284 | 2374 | IL2RG - ABCD1 | 311 | 1233 | 1322 |
| IL6ST - ABCD1 | 1347 | 1237 | 1220 | IL6ST - ABCD1 | 1581 | 507 | 612 |
| IL8 - ABCD1 | 2010 | 2315 | 1814 | IL8 - ABCD1 | 2501 | 2154 | 539 |
| IL10RB - ABCD1 | 631 | 825 | 85 | IL10RB - ABCD1 | 1795 | 1028 | 2325 |
| IL15 - ABCD1 | 890 | 325 | 1578 | IL15 - ABCD1 | 1795 | 302 | 1258 |
| IL15RA - ABCD1 | 2097 | 1765 | 1629 | IL15RA - ABCD1 | 580 | 1240 | 2342 |
| IL17C - ABCD1 | 1372 | 56 | 2509 | IL17C - ABCD1 | 687 | 1753 | 851 |
| IL17REL - ABCD1 | 5 | 2388 | 237 | IL17REL - ABCD1 | 1423 | 642 | 2164 |
| Ranking of IL family w.r.t ABCG1 | Ranking of ABCG1 family w.r.t IL | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - ABCG1 | 724 | 67 | 80 | IL1A - ABCG1 | 699 | 824 | 600 |
| IL1B - ABCG1 | 938 | 178 | 533 | IL1B - ABCG1 | 70 | 783 | 81 |
| IL1RAP - ABCG1 | 1263 | 2205 | 2339 | IL1RAP - ABCG1 | 2298 | 394 | 612 |
| IL1RN - ABCG1 | 1240 | 688 | 1396 | IL1RN - ABCG1 | 2465 | 834 | 1051 |
| IL2RG - ABCG1 | 1396 | 7 | 112 | IL2RG - ABCG1 | 587 | 24 | 21 |
| IL6ST - ABCG1 | 357 | 845 | 520 | IL6ST - ABCG1 | 1723 | 1345 | 177 |
| IL8 - ABCG1 | 977 | 1835 | 1099 | IL8 - ABCG1 | 1730 | 1748 | 382 |
| IL10RB - ABCG1 | 2244 | 349 | 840 | IL10RB - ABCG1 | 167 | 1315 | 61 |
| IL15 - ABCG1 | 1960 | 613 | 1279 | IL15 - ABCG1 | 2212 | 734 | 326 |
| IL15RA - ABCG1 | 785 | 651 | 2191 | IL15RA - ABCG1 | 1195 | 862 | 1876 |
| IL17C - ABCG1 | 2516 | 486 | 51 | IL17C - ABCG1 | 80 | 95 | 177 |
| IL17REL - ABCG1 | 2229 | 732 | 150 | IL17REL - ABCG1 | 1579 | 1025 | 452 |
| Ranking of IL family w.r.t ABCG2 | Ranking of ABCG2 family w.r.t IL | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - ABCG2 | 745 | 716 | 1299 | IL1A - ABCG2 | 238 | 89 | 659 |
| IL1B - ABCG2 | 354 | 232 | 668 | IL1B - ABCG2 | 31 | 197 | 439 |
| IL1RAP - ABCG2 | 2184 | 2167 | 1384 | IL1RAP - ABCG2 | 1314 | 253 | 2434 |
| IL1RN - ABCG2 | 783 | 228 | 11 | IL1RN - ABCG2 | 552 | 1692 | 827 |
| IL2RG - ABCG2 | 444 | 463 | 1024 | IL2RG - ABCG2 | 261 | 87 | 1275 |
| IL6ST - ABCG2 | 1647 | 1827 | 55 | IL6ST - ABCG2 | 1792 | 1477 | 1222 |
| IL8 - ABCG2 | 2212 | 1362 | 563 | IL8 - ABCG2 | 448 | 441 | 1423 |
| IL10RB - ABCG2 | 31 | 80 | 667 | IL10RB - ABCG2 | 2144 | 2335 | 2434 |
| IL15 - ABCG2 | 76 | 312 | 187 | IL15 - ABCG2 | 247 | 590 | 832 |
| IL15RA - ABCG2 | 1910 | 2428 | 1921 | IL15RA - ABCG2 | 1116 | 1005 | 1059 |
| IL17C - ABCG2 | 649 | 692 | 61 | IL17C - ABCG2 | 784 | 462 | 775 |
| IL17REL - ABCG2 | 883 | 1435 | 35 | IL17REL - ABCG2 | 852 | 1606 | 1597 |
Table 77 shows the derived influences which can be represented graphically, with the following influences - • ABC w.r.t IL with IL-1B/1RAP/10RB -> ABCA5; IL-10RB -> ABCB11; IL-1A/17REL -> ABCC3; IL-1A/1RAP/15/17C -> ABCC5; IL-1RAP/15RA -> ABCC13; IL-8/10RB -> ABCD1 and IL-10RB -> ABCG2; • IL w.r.t ABC with IL-17REL <- ABCA5; IL-2RG/6ST/15/15RA <- ABCB11; IL-8/15RA <- ABCC3; IL-15RA/17REL <- ABCC5; IL-15RA/17REL <- ABCC13; IL-1A/1RAP/8/15RA <- ABCD1; IL-1RAP <- ABCG1 and IL-1RAP/15RA <- ABCG2;
3.6.4. BCL - ABC Transporters Cross Family Analysis
Ruzickova et al. [163] show clinically relevant interactions of anti-apoptotic Bcl-2 protein inhibitors with ABC transporters. Alla et al. [164] observe that E2F1 confers anticancer drug resistance by targeting ABC transporter family members and Bcl-2 via the p73/DNp73-miR-205 circuitry. Yasui et al. [165] show a range of ABC family members along with BCL member to be overexpressed while studying the alteration in copy numbers of genes as a mechanism for acquired drug resistance. These point to the possible synergistic workings of BCL with ABC. In colorectal cancer cells treated with ETC-1922159, these were found to be up regulated. The search engine pointed to some of these 2nd order combinations and alloted rankings of high numerical value, thus indicating possible synergy. Table 78 and Table 79 show rankings of BCL family w.r.t ABC members on the left half and vice versa on the right half.
On the left half we found BCL2L1 up regulated w.r.t ABCC5. This is reflected in the rankings of 2239 (laplace) and 1845 (linear). BCL2L2 was up regulated w.r.t ABC-B11/C5/C13/D1. These are reflected in the rankings of 2097 (laplace) and 2311 (rbf) for ABCB11 - BCL2L2; 2195 (laplace), 2359 (linear) and 2322 (rbf) for ABCC5 - BCL2L2; 2438 (laplace) and 2494 (linear) for ABCC13 - BCL2L2 and 2477 (laplace) and 2156 (rbf) for ABCD1 - BCL2L2. BCL2L13 was up regulated w.r.t ABC-B11/C5/C13/D1/G1. These are reflected in the rankings of 2505 (laplace) and 1855 (rbf) for ABCB11 - BCL2L13; 1835 (linear) and 2178 (rbf) for ABCC5 - BCL2L13; 2484 (laplace), 2184 (linear) and 2410 (rbf) for ABCC13 - BCL2L13; 2472 (laplace) and 2201 (rbf) for ABCD1 - BCL2L13 and 2276 (linear) and 2095 (rbf) for ABCG1 - BCL2L13. BCL3 was up regulated w.r.t ABC-D1/G1. These are reflected in the rankings of 2194 (linear) and 2106 (rbf) for ABCD1 - BCL3 and 2014 (laplace) and 2253 (rbf) for ABCG1 - BCL3. BCL6 was up regulated w.r.t ABC-B11. These are reflected in the rankings of 2010 (linear) and 2350 (rbf) for ABC-B11 - BCL6. BCL10 was up regulated w.r.t ABC-B11. These are reflected in the rankings of 2234 (laplace) and 2382 (rbf) for ABC-B11 - BCL10.
Table 79.
2nd order interaction ranking between ABC and BCL family members.
| Ranking BCL family VS ABC family | |||||||
| Ranking of BCL6 w.r.t ABC family | Ranking of ABC family w.r.t BCL6 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| ABCA5 - BCL6 | 2045 | 557 | 1384 | ABCA5 - BCL6 | 211 | 283 | 1615 |
| ABCB11 - BCL6 | 1611 | 2010 | 2350 | ABCB11 - BCL6 | 841 | 427 | 2320 |
| ABCC3 - BCL6 | 1895 | 983 | 958 | ABCC3 - BCL6 | 1084 | 570 | 594 |
| ABCC5 - BCL6 | 615 | 597 | 567 | ABCC5 - BCL6 | 1370 | 1841 | 2389 |
| ABCC13 - BCL6 | 1097 | 2431 | 1731 | ABCC13 - BCL6 | 2172 | 2456 | 1063 |
| ABCD1 - BCL6 | 1446 | 1139 | 1953 | ABCD1 - BCL6 | 1097 | 1297 | 827 |
| ABCG1 - BCL6 | 1462 | 1688 | 1918 | ABCG1 - BCL6 | 192 | 27 | 1111 |
| ABCG2 - BCL6 | 947 | 1503 | 978 | ABCG2 - BCL6 | 129 | 745 | 719 |
| Ranking of BCL9L w.r.t ABC family | Ranking of ABC family w.r.t BCL9L | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| ABCA5 - BCL9L | 67 | 1008 | 94 | ABCA5 - BCL9L | 1753 | 1167 | 2312 |
| ABCB11 - BCL9L | 1989 | 158 | 1705 | ABCB11 - BCL9L | 1033 | 494 | 48 |
| ABCC3 - BCL9L | 1307 | 2249 | 1357 | ABCC3 - BCL9L | 457 | 2296 | 971 |
| ABCC5 - BCL9L | 1694 | 432 | 477 | ABCC5 - BCL9L | 1775 | 1551 | 2073 |
| ABCC13 - BCL9L | 1724 | 1410 | 862 | ABCC13 - BCL9L | 110 | 2475 | 2325 |
| ABCD1 - BCL9L | 1366 | 2344 | 1666 | ABCD1 - BCL9L | 1016 | 2440 | 2411 |
| ABCG1 - BCL9L | 1248 | 1680 | 536 | ABCG1 - BCL9L | 1146 | 676 | 16 |
| ABCG2 - BCL9L | 2451 | 1119 | 224 | ABCG2 - BCL9L | 1263 | 1421 | 218 |
| Ranking of BCL10 w.r.t ABC family | Ranking of ABC family w.r.t BCL10 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| ABCA5 - BCL10 | 687 | 176 | 808 | ABCA5 - BCL10 | 1753 | 1167 | 2312 |
| ABCB11 - BCL10 | 2234 | 2382 | 322 | ABCB11 - BCL10 | 1033 | 494 | 48 |
| ABCC3 - BCL10 | 589 | 379 | 492 | ABCC3 - BCL10 | 457 | 2296 | 971 |
| ABCC5 - BCL10 | 1489 | 397 | 1643 | ABCC5 - BCL10 | 1775 | 1551 | 2073 |
| ABCC13 - BCL10 | 956 | 538 | 1491 | ABCC13 - BCL10 | 110 | 2475 | 2325 |
| ABCD1 - BCL10 | 1009 | 470 | 1597 | ABCD1 - BCL10 | 1016 | 2440 | 2411 |
| ABCG1 - BCL10 | 1613 | 310 | 1115 | ABCG1 - BCL10 | 1146 | 676 | 16 |
| ABCG2 - BCL10 | 361 | 676 | 2020 | ABCG2 - BCL10 | 1263 | 1421 | 218 |
On the right half we found ABCC3 up regulated w.r.t BCL2L1. This is reflected in the rankings of 2085 (laplace) and 2309 (linear) for ABCC3 - BCL2L1. ABC-C5/C13 were up regulated w.r.t BCL2L13. These was reflected in the rankings of 1975 (laplace) and 2421 (linear) for ABCC5 - BCL2L13; and 1894 (laplace), 2335 (linear) and 2475 (rbf) for ABCC13 - BCL2L13. ABC-C3 was up regulated w.r.t BCL3. This is reflected in the rankings of 1782 (linear) and 2186 (rbf) for ABCC3 - BCL3. ABC-C5/C13 were up regulated w.r.t BCL6. This is reflected in the rankings of 1841 (linear) and 2389 (rbf) for ABCC5 - BCL6 and 2172 (laplace) and 2456 (linear) for ABCC13 - BCL6. ABC-C5/C13/D1 were up regulated w.r.t BCL9L. This is reflected in the rankings of 1775 (laplace) and 2073 (rbf) for ABCC5 - BCL9L; 2475 (linear) and 2325 (rbf) for ABCC13 - BCL9L and 2440 (linear) and 2411 (rbf) for ABCD1 - BCL9L; ABC-A5/C5/C13/D1 were up regulated w.r.t BCL10. These were reflected in the rankings of 1753 (laplace) and 2312 (rbf) for ABCA5 - BCL10; 1775 (laplace) and 2073 (rbf) for ABCC5 - BCL10; 2475 (linear) and 2325 (rbf) for ABCC13 - BCL10 and 2440 (linear) and 2411 (rbf) for ABCD1 - BCL10.
Table 80 shows the derived influences which can be represented graphically, with the following influences - • BCL w.r.t ABC with ABC-C5 -> BCL2L1; ABC-B11/C5/C13/D1 -> BCL2L2; ABC-B11/C5/C13/D1/G1 -> BCL2L13; ABC-D1/G1 -> BCL3; ABC-B11 -> BCL6; ABC-B11 -> BCL10; and • ABC w.r.t BCL with ABC-C3 <- BCL2L1; ABC-C5/C13 <- BCL2L13; ABC-C3 <- BCL3; ABC-C5/C13 <- BCL6; ABC-C5/C13/D1 <- BCL9L; ABC-A5/C5/C13/D1 <- BCL10.
3.6.5. CASPASE - ABC Transporters Cross Family Analysis
Hu et al. [166] observe that the loss of ABCB4 attenuates the caspase-dependent apoptosis regulating resistance to 5-Fu in colorectal cancer. Ihlefeld et al. [167] analyze whether the observed upregulation of the multidrug transporters contributed to the resistance of Sgpl1/-MEFs against chemotherapy-induced apoptosis by measuring the influence of ABC transporter inhibitors on cell viability and caspase-3 cleavage. Though recent developements, they point to the synergy between the transporters and the CASP family. In CRC cells, treated with ETC-1922159, these were found to be UP regulated. The engine alotted high numerical valued ranks to some of the 2nd order combinations between the members of the two families. Table 81 and Table 82 show the rankings of ABC transporters w.r.t CASP and vice versa.
In Table 81, we found ABC-C5 to be up regulated w.r.t CASP4. These are reflected in rankings of 2495 (laplace) and 2257 (rbf) for CASP4 - ABC-C5. ABC-C5 was up regulated w.r.t CASP5. These are reflected in rankings of 2475 (laplace) and 2234 (rbf) for CASP5 - ABC-C5. ABC-A5/C13/D1 were up regulated w.r.t CASP7. These are reflected in rankings of 2515 (laplace) and 1742 (linear) for CASP7 - ABC-C5; 2489 (laplace) and 2418 (linear) for CASP7 - ABC-C13; and 2323 (laplace) and 2004 (linear) for CASP7 - ABC-D1. ABC-B11/C5/D1/G1 were up regulated w.r.t CASP9. These are reflected in rankings of 2001 (linear) and 2051 (rbf) for CASP9 - ABC-B11; 2180 (laplace) and 2343 (linear) for CASP9 - ABC-C5; 2267 (laplace) and 2382 (rbf) for CASP9 - ABC-C13; 1890 (linear) and 2286 (rbf) for CASP9 - ABC-G1; ABC-A5/C13 were up regulated w.r.t CASP10. These are reflected in rankings of 2292 (laplace), 2311 (linear) and 1108 (rbf) for CASP10 - ABC-A5; 2139 (laplace) and 2203 (linear) for CASP10 - ABC-C13;
In Table 82, we found ABC-C5 to be up regulated w.r.t CASP4. These are reflected in rankings of 2495 (laplace) and 2257 (rbf) for CASP4 - ABC-C5. ABC-C5 was up regulated w.r.t CASP5. These are reflected in rankings of 2475 (laplace) and 2234 (rbf) for CASP5 - ABC-C5. ABC-A5/C13/D1 were up regulated w.r.t CASP7. These are reflected in rankings of 2515 (laplace) and 1742 (linear) for CASP7 - ABC-C5; 2489 (laplace) and 2418 (linear) for CASP7 - ABC-C13; and 2323 (laplace) and 2004 (linear) for CASP7 - ABC-D1. ABC-B11/C5/D1/G1 were up regulated w.r.t CASP9. These are reflected in rankings of 2001 (linear) and 2051 (rbf) for CASP9 - ABC-B11; 2180 (laplace) and 2343 (linear) for CASP9 - ABC-C5; 2267 (laplace) and 2382 (rbf) for CASP9 - ABC-C13; 1890 (linear) and 2286 (rbf) for CASP9 - ABC-G1; ABC-A5/C13 were up regulated w.r.t CASP10. These are reflected in rankings of 2292 (laplace), 2311 (linear) and 1108 (rbf) for CASP10 - ABC-A5; 2139 (laplace) and 2203 (linear) for CASP10 - ABC-C13;
In Table 82, we found CASP4 to be up regulated w.r.t ABC-D1. These are reflected in rankings of 1791 (laplace) and 1954 (rbf) for CASP4 - ABC-D1. CASP5 was up regulated w.r.t ABC-C13. These are reflected in rankings of 2286 (laplace) and 1905 (rbf) for CASP5 - ABC-C13. CASP7 was up regulated w.r.t ABC-C5. This is reflected in rankings of 2168 (laplace), 1881 (linear) and 2016 (rbf) for CASP7 - ABC-C5. CASP9 were up regulated w.r.t ABC-C5/C13/D1/G1. These are reflected in rankings of 2404 (laplace) and 2374 (linear) for CASP9 - ABC-A5; 2449 (laplace) and 2506 (rbf) for CASP9 - ABC-C13; 1858 (laplace) and 2430 (rbf) for CASP9 - ABC-D1; and 2342 (linear) and 2468 (rbf) for CASP9 - ABC-G1; CASP16 were up regulated w.r.t ABC-A5. This is reflected in rankings of 2477 (linear) and 2315 (rbf) for CASP16 - ABC-A5.
Table 83 shows the derived influences which can be represented graphically, with the following influences - • ABC w.r.t CASP with CASP-4 -> ABC-C5; CASP-5 -> ABC-C5; CASP-7 -> ABC-A5/C13/D1; CASP-9 -> ABC-B11/C5/D1/G1; CASP-10 -> ABC-A5/C13; and • CASP w.r.t ABC with CASP-4 <- ABC-D1; CASP-5 <- ABC-C13; CASP-7 <- ABC-C5; CASP-9 <- ABC-C5/C13/D1/G1; CASP-16 <- ABC-A5;
Table 81.
2nd order interaction ranking between ABC and CASP family members.
| Ranking ABC family w.r.t CASP family | |||||||
| Ranking of ABC family w.r.t CASP4 | Ranking of ABC family w.r.t CASP5 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| CASP4 - ABC-A5 | 957 | 682 | 991 | CASP5 - ABC-A5 | 733 | 1986 | 421 |
| CASP4 - ABC-B11 | 19 | 727 | 158 | CASP5 - ABC-B11 | 513 | 406 | 355 |
| CASP4 - ABC-C3 | 1242 | 857 | 1848 | CASP5 - ABC-C3 | 685 | 1694 | 1558 |
| CASP4 - ABC-C5 | 2495 | 1316 | 2257 | CASP5 - ABC-C5 | 2475 | 1038 | 2234 |
| CASP4 - ABC-C13 | 154 | 1537 | 1206 | CASP5 - ABC-C13 | 1660 | 1581 | 853 |
| CASP4 - ABC-D1 | 1494 | 964 | 999 | CASP5 - ABC-D1 | 354 | 725 | 1304 |
| CASP4 - ABC-G1 | 1405 | 70 | 326 | CASP5 - ABC-G1 | 298 | 485 | 382 |
| CASP4 - ABC-G2 | 157 | 176 | 523 | CASP5 - ABC-G2 | 706 | 846 | 1598 |
| Ranking of ABC family w.r.t CASP7 | Ranking of ABC family w.r.t CASP9 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| CASP7 - ABC-A5 | 2515 | 1742 | 25 | CASP9 - ABC-A5 | 1125 | 1863 | 694 |
| CASP7 - ABC-B11 | 1299 | 207 | 348 | CASP9 - ABC-B11 | 729 | 2001 | 2051 |
| CASP7 - ABC-C3 | 992 | 511 | 2222 | CASP9 - ABC-C3 | 1108 | 1470 | 1465 |
| CASP7 - ABC-C5 | 1232 | 1449 | 2154 | CASP9 - ABC-C5 | 2180 | 2343 | 1732 |
| CASP7 - ABC-C13 | 2489 | 2418 | 1623 | CASP9 - ABC-C13 | 2267 | 1472 | 2382 |
| CASP7 - ABC-D1 | 1544 | 2323 | 2004 | CASP9 - ABC-D1 | 1011 | 1086 | 174 |
| CASP7 - ABC-G1 | 665 | 382 | 670 | CASP9 - ABC-G1 | 580 | 1890 | 2286 |
| CASP7 - ABC-G2 | 1930 | 23 | 963 | CASP9 - ABC-G2 | 647 | 2374 | 310 |
| Ranking of ABC family w.r.t CASP10 | Ranking of ABC family w.r.t CASP16 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| CASP10 - ABC-A5 | 2292 | 2311 | 1108 | CASP16 - ABC-A5 | 165 | 408 | 113 |
| CASP10 - ABC-B11 | 2245 | 1467 | 1182 | CASP16 - ABC-B11 | 495 | 949 | 1417 |
| CASP10 - ABC-C3 | 760 | 2479 | 923 | CASP16 - ABC-C3 | 50 | 4 | 556 |
| CASP10 - ABC-C5 | 326 | 485 | 1429 | CASP16 - ABC-C5 | 1635 | 2487 | 1309 |
| CASP10 - ABC-C13 | 2139 | 2203 | 1524 | CASP16 - ABC-C13 | 1517 | 936 | 1236 |
| CASP10 - ABC-D1 | 2210 | 475 | 1655 | CASP16 - ABC-D1 | 1029 | 1210 | 1285 |
| CASP10 - ABC-G1 | 2337 | 128 | 71 | CASP16 - ABC-G1 | 350 | 756 | 109 |
| CASP10 - ABC-G2 | 2075 | 1693 | 1306 | CASP16 - ABC-G2 | 318 | 476 | 515 |
Table 82.
2nd order interaction ranking between CASP and ABC family members.
| Ranking CASP family w.r.t ABC family | |||||||
| Ranking of CASP4 w.r.t ABC family | Ranking of CASP5 w.r.t ABC family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| CASP4 - ABC-A5 | 791 | 586 | 753 | CASP5 - ABC-A5 | 696 | 427 | 48 |
| CASP4 - ABC-B11 | 462 | 263 | 427 | CASP5 - ABC-B11 | 1470 | 1300 | 242 |
| CASP4 - ABC-C3 | 1013 | 54 | 1140 | CASP5 - ABC-C3 | 821 | 286 | 459 |
| CASP4 - ABC-C5 | 2396 | 26 | 209 | CASP5 - ABC-C5 | 2368 | 665 | 171 |
| CASP4 - ABC-C13 | 1305 | 775 | 2193 | CASP5 - ABC-C13 | 2286 | 739 | 1905 |
| CASP4 - ABC-D1 | 1791 | 591 | 1954 | CASP5 - ABC-D1 | 653 | 440 | 972 |
| CASP4 - ABC-G1 | 593 | 99 | 173 | CASP5 - ABC-G1 | 2176 | 446 | 317 |
| CASP4 - ABC-G2 | 423 | 109 | 1364 | CASP5 - ABC-G2 | 332 | 122 | 533 |
| Ranking of CASP7 w.r.t ABC family | Ranking of CASP9 w.r.t ABC family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| CASP7 - ABC-A5 | 1726 | 697 | 1874 | CASP9 - ABC-A5 | 2404 | 2374 | 1265 |
| CASP7 - ABC-B11 | 1549 | 189 | 1692 | CASP9 - ABC-B11 | 998 | 1258 | 2046 |
| CASP7 - ABC-C3 | 2331 | 1572 | 69 | CASP9 - ABC-C3 | 1398 | 2358 | 1445 |
| CASP7 - ABC-C5 | 2168 | 1881 | 2016 | CASP9 - ABC-C5 | 1023 | 965 | 1080 |
| CASP7 - ABC-C13 | 1822 | 865 | 1239 | CASP9 - ABC-C13 | 2449 | 1545 | 2506 |
| CASP7 - ABC-D1 | 111 | 813 | 2230 | CASP9 - ABC-D1 | 1858 | 2430 | 412 |
| CASP7 - ABC-G1 | 1609 | 983 | 1994 | CASP9 - ABC-G1 | 305 | 2342 | 2468 |
| CASP7 - ABC-G2 | 1094 | 952 | 102 | CASP9 - ABC-G2 | 1868 | 1621 | 1154 |
| Ranking of CASP10 w.r.t ABC family | Ranking of CASP16 w.r.t ABC family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| CASP10 - ABC-A5 | 683 | 1453 | 1437 | CASP16 - ABC-A5 | 960 | 2477 | 2315 |
| CASP10 - ABC-B11 | 1301 | 774 | 558 | CASP16 - ABC-B11 | 402 | 1860 | 794 |
| CASP10 - ABC-C3 | 369 | 683 | 1453 | CASP16 - ABC-C3 | 171 | 825 | 23 |
| CASP10 - ABC-C5 | 1823 | 346 | 761 | CASP16 - ABC-C5 | 2467 | 585 | 258 |
| CASP10 - ABC-C13 | 1320 | 832 | 868 | CASP16 - ABC-C13 | 428 | 177 | 64 |
| CASP10 - ABC-D1 | 249 | 1440 | 387 | CASP16 - ABC-D1 | 651 | 153 | 2010 |
| CASP10 - ABC-G1 | 1687 | 1232 | 156 | CASP16 - ABC-G1 | 2398 | 421 | 1120 |
| CASP10 - ABC-G2 | 1151 | 651 | 464 | CASP16 - ABC-G2 | 1193 | 734 | 479 |
Table 83.
2nd order combinatorial hypotheses between BCL and ABC family members.
| Unexplored combinatorial hypotheses | |
| ABC w.r.t CASP | |
| CASP-4 | ABC-C5 |
| CASP-5 | ABC-C5 |
| CASP-7 | ABC-A5/C13/D1 |
| CASP-9 | ABC-B11/C5/D1/G1 |
| CASP-10 | ABC-A5/C13 |
| CASP w.r.t ABC | |
| CASP-4 | ABC-D1 |
| CASP-5 | ABC-C13 |
| CASP-7 | ABC-C5 |
| CASP-9 | ABC-C5/C13/D1/G1 |
| CASP-16 | ABC-A5 |
3.7. Interleukin Related Synergies
3.7.1. NFkB-2/I - Interleukin Cross Family Analysis
Hörber et al. [168] show that the atypical inhibitor of NF-κB, IκBζ, controls macrophage interleukin-10 expression. Yamazaki et al. [169] observe that stimulus-specific induction of a novel nuclear factor-κB regulator, IκB-ζ, via Toll/Interleukin-1 receptor is mediated by mRNA stabilization. Kurzrock et al. [170] show that Interleukin-1 increases expression of the LYT-10 (NFκB2) proto-oncogene/transcription factor in renal cell carcinoma lines. These studies and many others not indicated here, show the connection between Interleukin and NFkB-2 and NFkBI family. In CRC cells treated with ETC-1922159, members of these families were UP regulated. Table 84 shows the rankings of each family with the other.
On the left side, rankings of IL w.r.t NFkB-2/I has been indicated. We found IL-15RA/17C to be up regulated w.r.t NFkB2. These are reflected in rankings of 1787 (rbf) and 1957 (rbf) IL15RA - NFkB2 and 2288 (linear) and 2018 (rbf) IL17C - NFkB2. IL-1RN/6ST/15RA to be up regulated w.r.t NFkB2. These are reflected in rankings of 1753 (laplace) and 1906 (linear) for IL1RN - NFkBIA; 2400 (linear) and 2094 (rbf) for IL6ST - NFkBIA and 2251 (laplace) and 2390 (linear) for IL15RA - NFkBIA. IL-1RAP/6ST/8/17REL to be up regulated w.r.t NFkB2. These are reflected in rankings of 2221 (linear) and 1807 (rbf) IL1RAP - NFkBIE; 2381 (linear) and 2277 (rbf) for IL6ST - NFkBIE; 2198 (linear) and 2133 (rbf) for IL8 - NFkBIE and 2216 (linear) and 2168 (rbf) for IL17REL - NFkBIE. IL-1A/6ST/15 to be up regulated w.r.t NFkB2. These are reflected in rankings of 2381 (laplace) and 2049 (linear) for IL1A - NFkBIZ; 2279 (laplace) and 2431 (linear) for IL6ST - NFkBIZ and 1780 (laplace) and 2098 (linear) for IL15 - NFkBIZ;
On the right side, rankings of NFkB-2/I w.r.t IL has been indicated. We found NFkB-2 to be up regulated w.r.t IL10RB. This is reflected in rankings of 2282 (laplace), 2381 (linear) and 1897 (rbf) for NFkB2 - IL10RB. NFkBIZ to be up regulated w.r.t IL-10RB/17REL. These were reflected in rankings of 2271 (laplace) and 2082 (rbf) for IL10RB - NFkBIZ and 1883 (linear) and 1830 (rbf) for IL17REL - NFkBIZ.
Table 84.
2nd order combinatorial hypotheses between NFkB-2/I and IL.
| Ranking Interleukin family vs NFkB-2 family | |||||||
| Ranking of IL family w.r.t NFkB-2 | Ranking of NFkB-2/I w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - NFkB2 | 1485 | 6 | 2494 | IL1A - NFkB2 | 616 | 276 | 1358 |
| IL1B - NFkB2 | 1852 | 638 | 1587 | IL1B - NFkB2 | 283 | 284 | 1088 |
| IL1RAP - NFkB2 | 1369 | 1849 | 1463 | IL1RAP - NFkB2 | 967 | 377 | 161 |
| IL1RN - NFkB2 | 1285 | 1963 | 1604 | IL1RN - NFkB2 | 1386 | 2086 | 52 |
| IL2RG - NFkB2 | 486 | 1077 | 1300 | IL2RG - NFkB2 | 1436 | 1123 | 2163 |
| IL6ST - NFkB2 | 493 | 814 | 283 | IL6ST - NFkB2 | 2177 | 343 | 2255 |
| IL8 - NFkB2 | 1907 | 865 | 335 | IL8 - NFkB2 | 303 | 2355 | 1152 |
| IL10RB - NFkB2 | 707 | 1607 | 595 | IL10RB - NFkB2 | 2282 | 2381 | 1897 |
| IL15 - NFkB2 | 792 | 1113 | 1434 | IL15 - NFkB2 | 2112 | 1214 | 1217 |
| IL15RA - NFkB2 | 1787 | 233 | 1957 | IL15RA - NFkB2 | 1289 | 1235 | 1913 |
| IL17C - NFkB2 | 2288 | 305 | 2018 | IL17C - NFkB2 | 380 | 529 | 1492 |
| IL17REL - NFkB2 | 9 | 2464 | 167 | IL17REL - NFkB2 | 115 | 1540 | 308 |
| Ranking of IL family w.r.t NFkBI-A | Ranking of NFkB-2/I w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - NFkBIA | 116 | 46 | 1885 | IL1A - NFkBIA | 989 | 1179 | 705 |
| IL1B - NFkBIA | 328 | 56 | 1228 | IL1B - NFkBIA | 611 | 397 | 1378 |
| IL1RAP - NFkBIA | 1376 | 778 | 359 | IL1RAP - NFkBIA | 1131 | 515 | 1887 |
| IL1RN - NFkBIA | 1753 | 1906 | 267 | IL1RN - NFkBIA | 2357 | 578 | 382 |
| IL2RG - NFkBIA | 32 | 6 | 898 | IL2RG - NFkBIA | 132 | 684 | 784 |
| IL6ST - NFkBIA | 1011 | 2400 | 2094 | IL6ST - NFkBIA | 2008 | 533 | 90 |
| IL8 - NFkBIA | 1988 | 1234 | 1232 | IL8 - NFkBIA | 183 | 993 | 1109 |
| IL10RB - NFkBIA | 864 | 2239 | 8 | IL10RB - NFkBIA | 616 | 1251 | 107 |
| IL15 - NFkBIA | 1181 | 453 | 462 | IL15 - NFkBIA | 2227 | 958 | 165 |
| IL15RA - NFkBIA | 2251 | 2390 | 1652 | IL15RA - NFkBIA | 765 | 291 | 2301 |
| IL17C - NFkBIA | 538 | 229 | 330 | IL17C - NFkBIA | 450 | 178 | 19 |
| IL17REL - NFkBIA | 643 | 16 | 4 | IL17REL - NFkBIA | 1275 | 403 | 2190 |
| Ranking of IL family w.r.t NFkBI-E | Ranking of NFkBI-E w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - NFkBIE | 2486 | 27 | 76 | IL1A - NFkBIE | 433 | 1574 | 953 |
| IL1B - NFkBIE | 2089 | 39 | 311 | IL1B - NFkBIE | 1103 | 507 | 1931 |
| IL1RAP - NFkBIE | 201 | 2221 | 1807 | IL1RAP - NFkBIE | 474 | 1404 | 875 |
| IL1RN - NFkBIE | 2025 | 610 | 1153 | IL1RN - NFkBIE | 2051 | 381 | 468 |
| IL2RG - NFkBIE | 1141 | 986 | 654 | IL2RG - NFkBIE | 1327 | 1464 | 983 |
| IL6ST - NFkBIE | 1155 | 2381 | 2277 | IL6ST - NFkBIE | 309 | 143 | 939 |
| IL8 - NFkBIE | 259 | 2198 | 2133 | IL8 - NFkBIE | 1507 | 911 | 67 |
| IL10RB - NFkBIE | 1730 | 191 | 310 | IL10RB - NFkBIE | 305 | 478 | 1960 |
| IL15 - NFkBIE | 1922 | 365 | 117 | IL15 - NFkBIE | 2476 | 783 | 1302 |
| IL15RA - NFkBIE | 1912 | 839 | 1385 | IL15RA - NFkBIE | 424 | 526 | 1423 |
| IL17C - NFkBIE | 2179 | 105 | 404 | IL17C - NFkBIE | 2231 | 1205 | 321 |
| IL17REL - NFkBIE | 13 | 2216 | 2168 | IL17REL - NFkBIE | 333 | 831 | 949 |
| Ranking of IL family w.r.t NFkBI-Z | Ranking of NFkBI-Z w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - NFkBIZ | 2381 | 2049 | 1578 | IL1A - NFkBIZ | 157 | 792 | 1460 |
| IL1B - NFkBIZ | 1241 | 2210 | 463 | IL1B - NFkBIZ | 586 | 217 | 1617 |
| IL1RAP - NFkBIZ | 694 | 1077 | 936 | IL1RAP - NFkBIZ | 1326 | 240 | 1080 |
| IL1RN - NFkBIZ | 860 | 2151 | 231 | IL1RN - NFkBIZ | 2463 | 739 | 579 |
| IL2RG - NFkBIZ | 1362 | 2054 | 68 | IL2RG - NFkBIZ | 68 | 829 | 1212 |
| IL6ST - NFkBIZ | 2279 | 980 | 2431 | IL6ST - NFkBIZ | 996 | 1223 | 140 |
| IL8 - NFkBIZ | 992 | 1732 | 966 | IL8 - NFkBIZ | 816 | 1510 | 119 |
| IL10RB - NFkBIZ | 717 | 2275 | 571 | IL10RB - NFkBIZ | 2271 | 42 | 2082 |
| IL15 - NFkBIZ | 1780 | 2098 | 626 | IL15 - NFkBIZ | 2155 | 200 | 245 |
| IL15RA - NFkBIZ | 633 | 1726 | 2422 | IL15RA - NFkBIZ | 834 | 1284 | 1785 |
| IL17C - NFkBIZ | 1716 | 2430 | 1098 | IL17C - NFkBIZ | 848 | 1282 | 1391 |
| IL17REL - NFkBIZ | 14 | 75 | 314 | IL17REL - NFkBIZ | 289 | 1883 | 1830 |
Table 85 shows the derived influences which can be represented graphically, with the following influences - • IL w.r.t NFkB with IL w.r.t NFkB-2/I with IL15RA <- NFkB2; IL17C <- NFkB2; IL1RN <- NFkBIA; IL6ST <- NFkBIA; IL15RA <- NFkBIA; IL1RAP <- NFkBIE; IL6ST <- NFkBIE; IL8 <- NFkBIE; IL17REL <- NFkBIE; IL1A <- NFkBIZ; IL6ST <- NFkBIZ; IL15 <- NFkBIZ; and • NFkB-2/I w.r.t IL with IL10RB -> NFkB2; IL10RB -> NFKBIZ; IL17REL -> NFkBIZ;
3.7.2. Potassium Channel - Interleukin Cross Family Analysis
In 1986, Lee et al. [171] showed that increased voltage-gated potassium conductance during interleukin 2-stimulated proliferation of a mouse helper T lymphocyte clone. Martin et al. [172] show that interleukin-4 activates large-conductance, calcium-activated potassium (BKCa) channels in human airway smooth muscle cells. However, the author is not aware of deep studies between the Potassium ion channel subfamily members (KCN) and interleukin. In CRC cells treated with ETC-1922159, these were found to be UP regulated. The search engine found alotted multiple combinations between the members of these two families. These were reflected in ranking of the each with the other in the following Table 86 and Table 87. On the left is rankings of IL family with respect to the KCN family member and on the right, vice versa.
Beginning on the left side we found IL-1A/1B/15RA/17C to be up regulated w.r.t KCND3. These are reflected in rankings of 1995 (laplace) and 2255 (linear) for IL1A - KCND3; 2083 (laplace) and 1897 (linear) for IL1B - KCND3; 2074 (laplace) and 2495 (rbf) for IL15RA - KCND3; and 1881 (laplace) and 2139 (linear) for IL17C - KCND3. IL-1A/1B to be up regulated w.r.t KCNH2. These are reflected in rankings of 2103 (laplace) and 1832 (linear) for IL1A - KCNH2 and 2447 (laplace) and 2068 (linear) for IL1B - KCNH2; IL-1A/1B/17C to be up regulated w.r.t KCNH8. These are reflected in rankings of 2268 (laplace), 2507 (linear) and 1877 (rbf) for IL1A - KCNH8; 2223 (laplace), 2013 (linear) and 2204 (rbf) for IL1B - KCNH8; and 1847 (laplace), and 2354 (rbf) for IL17C - KCNH8. IL-1A/1B/1RN/15 to be up regulated w.r.t KCNK1. These are reflected in rankings of 2290 (laplace) and 2066 (linear) for IL1A - KCNK1; 1941 (laplace) and 2452 (linear) and 1905 (rbf) for IL1B - KCNK1; 2468 (laplace) and 1897 (linear) for IL1RN - KCNK1; 2280 (laplace) and 2009 (rbf) for IL15 - KCNK1. IL-1RN/10RB/17REL to be up regulated w.r.t KCNK5. These are reflected in rankings of 1930 (linear) and 2136 (rbf) for IL1RN-KCNK5; 1879 (laplace), 2298 (linear) and 1903 (rbf) for IL10RB-KCKK5; and 2118 (laplace) and 1873 (rbf) for IL17REL - KCNK5; IL-8/17REL to be up regulated w.r.t KCNK5. These are reflected in rankings of 2168 (laplace) and 2442 (linear) for IL8 - KCNK6; and 2066 (laplace) and 2159 (linear) for IL17REL - KCNK6.
Beginning on the right side we found KCND3 to be up regulated w.r.t IL-1A/1B/15RA/17C. These are reflected in rankings of 2495 (laplace), 2390 (linear) for IL1RAP - KCND3; 2048 (laplace), 2306 (linear) and 2197 (rbf) for IL10RB - KCND3 and 2511 (laplace) and 2517 (linear) for IL15RA - KCND3; KCNH2 to be up regulated w.r.t IL-1A/1RAP. These are reflected in rankings of 1897 (laplace), 2152 (linear) and 2179 (rbf) for IL1A - KCNH2; and 2451 (laplace), 1805 (linear) and 2002 (rbf) for IL1RAP - KCNH2; KCNH8 to be up regulated w.r.t IL-1B/10RB. These are reflected in rankings of 2060 (laplace) and 2177 (rbf) for IL1B - KCNH8; and 2381 (laplace) and 2008 (linear) for IL10RB - KCNH8; KCNK1 to be up regulated w.r.t IL-1A/6ST/8. These are reflected in rankings of 1818 (linear) and 2362 (rbf) for IL1A - KCNK1; 2226 (laplace) and 2283 (rbf) for IL6ST - KCNK1; and 1872 (laplace) and 1978 (linear) for IL8 - KCNK1; KCNK5 to be up regulated w.r.t IL-10RB. This is reflected in rankings of 1769 (linear) and 2206 (rbf) for IL10RB - KCNK5; KCNK6 to be up regulated w.r.t IL-1RAP/10RB/15. These are reflected in rankings of 2386 (laplace) and 2053 (rbf) for IL1RAP - KCNK6; 1903 (linear) and 2156 (rbf) for IL10RB - KCNK6; and 1944 (laplace) and 2047 (rbf) for IL15 - KCNK6;
Finally, Table 88 shows the derived influences which can be represented graphically, with the following influences - • IL w.r.t KCN with IL-1A/1B/15RA/17C <- KCND3; IL-1A/1B <- KCNH2; IL-1A/1B/17C <- KCNH8; IL-1A/1B/1RN/15 <- KCNK1; IL-1RN/10RB/17REL <- KCNK5; IL-8/17REL <- KCNK6; and • KCN w.r.t IL family with IL-1A/1B/15RA/17C -> KCND3; IL-1A/1RAP -> KCNH2; IL-1B/10RB -> KCNH8; IL-1A/6ST/8 -> KCNK1; IL-10RB -> KCNK5; and IL-1RAP/10RB/15 -> KCNK6;
3.7.3. Mucin - Interleukin Cross Family Analysis
Kerschner et al. [173] have observed that middle ear epithelial mucin production in response to interleukin-6 exposure in vitro. Chen et al. [174] observe that stimulation of airway mucin gene expression by interleukin (IL)-17 through IL-6 paracrine/autocrine loop. Suppression of mucin 2 promotes interleukin-6 secretion and tumor growth in an orthotopic immune-competent colon cancer animal model was observed by Shan et al. [175]. Yokoigawa et al. [176] show enhanced production of interleukin 6 in peripheral blood monocytes stimulated with mucins secreted into the bloodstream. Gray et al. [177] show that interleukin-1β-induced mucin production in human airway epithelium is mediated by cyclooxygenase-2, prostaglandin E2 receptors, and cyclic AMP-protein kinase A signaling. Finally, in colorectal cancer, Hsu et al. [178] mucin 2 silencing promotes metastasis through interleukin-6 signaling. In CRC cells treated with ETC-1922159, both were found to be up regulated. Table 89 and Table 90 show the rankings of IL family w.r.t MUC family on the left side and vice versa on the right side.
On the left side, we found IL-1B/17C to be up regulated with respect to MUC1. These are reflected in rankings of 2218 (laplace) 1757 (linear) for IL1B - MUC1; and 1841 (linear) and 2003 (rbf) for IL17C - MUC1; IL-1A/1B/1RN/2RG/15/17C were up regulated with respect to MUC3A. These are reflected in rankings of 2513 (laplace) and 2480 (linear) for IL1A - MUC3A; 1820 (laplace) and 2308 (linear) for IL1B - MUC3A; 2138 (laplace) and 2270 (linear) for IL1RN - MUC3A; 1816 (laplace), 2115 (linear) and 1900 (rbf) for IL2RG - MUC3A; 2391 (laplace) and 2288 (linear) for IL15 - MUC3A; and 2443 (laplace) and 2512 (linear) for IL17C - MUC3A; IL-1RN/6ST/15RA were up regulated with respect to MUC4. These are reflected in rankings of 2010 (laplace) and 1960 (rbf) for IL1RN - MUC4; 2204 (laplace) and 1765 (rbf) for IL6ST - MUC4; and 2190 (laplace), 1814 (linear) and 2061 (rbf) for IL15RA - MUC4; IL-1A/2RG/8/15/17C were up regulated with respect to MUC12. These are reflected in rankings of 1806 (laplace) and 2396 (rbf) for IL1A - MUC12; 2195 (laplace) and 2089 (rbf) for IL2RG - MUC12; 1814 (laplace) and 2497 (rbf) for IL8 - MUC12; 2408 (laplace) and 2340 (rbf) for IL15 - MUC12; and 2436 (laplace) and 2416 (rbf) for IL17C - MUC12; IL-15RA were up regulated with respect to MUC17. These are reflected in rankings of 2265 (laplace) and 2064 (linear) for IL15RA - MUC17. IL-1RAP/8/17REL were up regulated with respect to MUC20. These are reflected in rankings of 2025 (linear) and 2251 (rbf) for IL1RAP - MUC20; 1820 (laplace) and 2303 (rbf) for IL8 - MUC20; and 2121 (laplace) and 2267 (rbf) for IL17REL - MUC20.
On the left side, we found MUC1 to be up regulated with respect to IL-1B. These are reflected in rankings of 1847 (laplace) and 2049 (rbf) for IL1B - MUC1. MUC12 to be up regulated with respect to IL-1RN/2RG/6ST. These are reflected in rankings of 2505 (laplace) and 1891 (linear) for IL1RN - MUC12; 1913 (laplace) and 1833 (linear) for IL2RG - MUC12; and 2100 (laplace) and 1759 (linear) for IL6ST - MUC12. MUC13 to be up regulated with respect to IL-1RAP/15RA. These are reflected in rankings of 1887 (laplace) and 2263 (rbf) for IL1RAP - MUC13; and 2109 (laplace) and 2402 (rbf) for IL15RA - MUC13; MUC20 to be up regulated with respect to IL-1A/10RB/17C. These are reflected in rankings of 2218 (laplace) and 2260 (rbf) for IL1A - MUC20; 1883 (linear) and 1947 (rbf) for IL10RB - MUC20; and 2212 (laplace) and 1843 (linear) for IL17C - MUC20.
Finally, Table 91 shows the derived influences which can be represented graphically, with the following influences - • IL w.r.t MUC with IL-1B/17C <- MUC1; IL-1A/1B/1RN/2RG/15/17C and MUC3A; IL-1RN/6ST/15RA <- MUC4; IL-1A/2RG/8/15/17C <- MUC12; IL-15RA <- MUC17; and IL-1RAP/8/17REL <- MUC20; and • MUC w.r.t IL with IL-1B <- MUC1; IL-1RN/2RG/6ST <- MUC12; IL-1RAP/15RA <- MUC13; and IL-1A/10RB/17C <- MUC20;
Table 89.
2nd order combinatorial hypotheses between MUC and IL.
| Ranking Interleukin family vs MUC family | |||||||
| Ranking of IL family w.r.t MUC1 | Ranking of MUC1 w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - MUC1 | 1961 | 1711 | 107 | IL1A - MUC1 | 111 | 879 | 535 |
| IL1B - MUC1 | 2218 | 1757 | 228 | IL1B - MUC1 | 1847 | 520 | 2049 |
| IL1RAP - MUC1 | 837 | 604 | 146 | IL1RAP - MUC1 | 1968 | 589 | 439 |
| IL1RN - MUC1 | 1084 | 918 | 1859 | IL1RN - MUC1 | 1752 | 353 | 507 |
| IL2RG - MUC1 | 1872 | 272 | 1281 | IL2RG - MUC1 | 1769 | 1009 | 285 |
| IL6ST - MUC1 | 2415 | 1115 | 1633 | IL6ST - MUC1 | 296 | 801 | 245 |
| IL8 - MUC1 | 1276 | 544 | 1055 | IL8 - MUC1 | 2079 | 1320 | 82 |
| IL10RB - MUC1 | 291 | 1638 | 1710 | IL10RB - MUC1 | 973 | 1691 | 924 |
| IL15 - MUC1 | 212 | 1003 | 1060 | IL15 - MUC1 | 160 | 205 | 942 |
| IL15RA - MUC1 | 213 | 1346 | 1067 | IL15RA - MUC1 | 1127 | 1057 | 1521 |
| IL17C - MUC1 | 1215 | 1841 | 2003 | IL17C - MUC1 | 3 | 236 | 7 |
| IL17REL - MUC1 | 19 | 44 | 2069 | IL17REL - MUC1 | 1142 | 541 | 1464 |
| Ranking of IL family w.r.t MUC3A | Ranking of MUC3A w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - MUC3A | 2513 | 2480 | 194 | IL1A - MUC3A | 1426 | 1017 | 1484 |
| IL1B - MUC3A | 1820 | 2308 | 1086 | IL1B - MUC3A | 816 | 1157 | 908 |
| IL1RAP - MUC3A | 753 | 1270 | 526 | IL1RAP - MUC3A | 1403 | 1402 | 102 |
| IL1RN - MUC3A | 2138 | 2270 | 313 | IL1RN - MUC3A | 1123 | 360 | 1333 |
| IL2RG - MUC3A | 1816 | 2115 | 1900 | IL2RG - MUC3A | 480 | 1560 | 514 |
| IL6ST - MUC3A | 283 | 1126 | 1229 | IL6ST - MUC3A | 1601 | 908 | 889 |
| IL8 - MUC3A | 356 | 760 | 1517 | IL8 - MUC3A | 2350 | 587 | 80 |
| IL10RB - MUC3A | 1401 | 729 | 157 | IL10RB - MUC3A | 520 | 458 | 2324 |
| IL15 - MUC3A | 850 | 2391 | 2288 | IL15 - MUC3A | 1385 | 1351 | 959 |
| IL15RA - MUC3A | 1304 | 1949 | 959 | IL15RA - MUC3A | 1538 | 1685 | 584 |
| IL17C - MUC3A | 2443 | 2512 | 647 | IL17C - MUC3A | 2153 | 623 | 1349 |
| IL17REL - MUC3A | 200 | 243 | 2048 | IL17REL - MUC3A | 1274 | 1250 | 1387 |
| Ranking of IL family w.r.t MUC4 | Ranking of MUC4 w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - MUC4 | 1268 | 489 | 112 | IL1A - MUC4 | 42 | 1449 | 331 |
| IL1B - MUC4 | 779 | 1142 | 393 | IL1B - MUC4 | 780 | 301 | 393 |
| IL1RAP - MUC4 | 1672 | 1203 | 926 | IL1RAP - MUC4 | 460 | 358 | 883 |
| IL1RN - MUC4 | 2010 | 438 | 1960 | IL1RN - MUC4 | 1681 | 1164 | 51 |
| IL2RG - MUC4 | 161 | 292 | 36 | IL2RG - MUC4 | 581 | 659 | 1056 |
| IL6ST - MUC4 | 2204 | 1116 | 1765 | IL6ST - MUC4 | 977 | 1555 | 873 |
| IL8 - MUC4 | 619 | 741 | 1030 | IL8 - MUC4 | 222 | 1341 | 1552 |
| IL10RB - MUC4 | 1818 | 1343 | 599 | IL10RB - MUC4 | 87 | 1511 | 95 |
| IL15 - MUC4 | 434 | 1268 | 602 | IL15 - MUC4 | 440 | 806 | 276 |
| IL15RA - MUC4 | 2190 | 1814 | 2061 | IL15RA - MUC4 | 427 | 1145 | 305 |
| IL17C - MUC4 | 255 | 60 | 558 | IL17C - MUC4 | 167 | 152 | 159 |
| IL17REL - MUC4 | 222 | 482 | 52 | IL17REL - MUC4 | 2266 | 419 | 160 |
| Ranking of IL family w.r.t MUC12 | Ranking of MUC12 w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - MUC12 | 1806 | 166 | 2396 | IL1A - MUC12 | 706 | 84 | 570 |
| IL1B - MUC12 | 1004 | 113 | 2086 | IL1B - MUC12 | 1352 | 167 | 445 |
| IL1RAP - MUC12 | 1906 | 1588 | 517 | IL1RAP - MUC12 | 52 | 272 | 1955 |
| IL1RN - MUC12 | 2209 | 669 | 235 | IL1RN - MUC12 | 2505 | 1891 | 567 |
| IL2RG - MUC12 | 2195 | 751 | 2089 | IL2RG - MUC12 | 1913 | 1833 | 939 |
| IL6ST - MUC12 | 1115 | 1522 | 1031 | IL6ST - MUC12 | 2100 | 1759 | 1508 |
| IL8 - MUC12 | 1814 | 1554 | 2497 | IL8 - MUC12 | 439 | 121 | 1635 |
| IL10RB - MUC12 | 2467 | 1114 | 1044 | IL10RB - MUC12 | 381 | 1863 | 12 |
| IL15 - MUC12 | 2408 | 192 | 2340 | IL15 - MUC12 | 2400 | 1307 | 1408 |
| IL15RA - MUC12 | 612 | 1636 | 203 | IL15RA - MUC12 | 137 | 127 | 468 |
| IL17C - MUC12 | 2436 | 484 | 2416 | IL17C - MUC12 | 411 | 182 | 283 |
| IL17REL - MUC12 | 2421 | 331 | 611 | IL17REL - MUC12 | 1452 | 678 | 651 |
Table 90.
2nd order combinatorial hypotheses between MUC and IL.
| Ranking Interleukin family vs MUC family contd. | |||||||
| Ranking of IL family w.r.t MUC13 | Ranking of MUC13 w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - MUC13 | 655 | 2323 | 826 | IL1A - MUC13 | 1176 | 148 | 803 |
| IL1B - MUC13 | 2250 | 298 | 185 | IL1B - MUC13 | 833 | 30 | 8 |
| IL1RAP - MUC13 | 386 | 490 | 360 | IL1RAP - MUC13 | 1887 | 1142 | 2263 |
| IL1RN - MUC13 | 904 | 1614 | 698 | IL1RN - MUC13 | 1749 | 1607 | 313 |
| IL2RG - MUC13 | 1043 | 59 | 27 | IL2RG - MUC13 | 434 | 852 | 1140 |
| IL6ST - MUC13 | 635 | 1774 | 730 | IL6ST - MUC13 | 1901 | 535 | 163 |
| IL8 - MUC13 | 225 | 510 | 1130 | IL8 - MUC13 | 2328 | 722 | 555 |
| IL10RB - MUC13 | 944 | 491 | 1631 | IL10RB - MUC13 | 1459 | 1841 | 342 |
| IL15 - MUC13 | 1773 | 609 | 1047 | IL15 - MUC13 | 315 | 465 | 302 |
| IL15RA - MUC13 | 1884 | 1360 | 1067 | IL15RA - MUC13 | 2109 | 158 | 2402 |
| IL17C - MUC13 | 562 | 106 | 149 | IL17C - MUC13 | 73 | 4 | 84 |
| IL17REL - MUC13 | 1808 | 83 | 59 | IL17REL - MUC13 | 694 | 676 | 586 |
| Ranking of IL family w.r.t MUC17 | Ranking of MUC17 w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - MUC17 | 1573 | 2431 | 1622 | IL1A - MUC17 | 881 | 311 | 254 |
| IL1B - MUC17 | 1122 | 514 | 1035 | IL1B - MUC17 | 676 | 1243 | 174 |
| IL1RAP - MUC17 | 1634 | 1148 | 1469 | IL1RAP - MUC17 | 136 | 369 | 2512 |
| IL1RN - MUC17 | 38 | 260 | 911 | IL1RN - MUC17 | 361 | 22 | 690 |
| IL2RG - MUC17 | 754 | 218 | 403 | IL2RG - MUC17 | 1379 | 530 | 177 |
| IL6ST - MUC17 | 1616 | 554 | 1381 | IL6ST - MUC17 | 1782 | 668 | 270 |
| IL8 - MUC17 | 241 | 583 | 402 | IL8 - MUC17 | 1612 | 436 | 1984 |
| IL10RB - MUC17 | 401 | 464 | 51 | IL10RB - MUC17 | 1707 | 1305 | 1857 |
| IL15 - MUC17 | 307 | 438 | 878 | IL15 - MUC17 | 466 | 366 | 596 |
| IL15RA - MUC17 | 2265 | 2064 | 1458 | IL15RA - MUC17 | 63 | 376 | 849 |
| IL17C - MUC17 | 1045 | 581 | 2291 | IL17C - MUC17 | 1530 | 285 | 1449 |
| IL17REL - MUC17 | 656 | 657 | 456 | IL17REL - MUC17 | 380 | 580 | 1306 |
| Ranking of IL family w.r.t MUC20 | Ranking of MUC20 w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - MUC20 | 103 | 1729 | 18 | IL1A - MUC20 | 2218 | 1499 | 2260 |
| IL1B - MUC20 | 85 | 1810 | 30 | IL1B - MUC20 | 1313 | 1719 | 735 |
| IL1RAP - MUC20 | 974 | 2025 | 2251 | IL1RAP - MUC20 | 1784 | 859 | 1705 |
| IL1RN - MUC20 | 1176 | 2264 | 246 | IL1RN - MUC20 | 1265 | 726 | 823 |
| IL2RG - MUC20 | 405 | 2168 | 335 | IL2RG - MUC20 | 2152 | 165 | 1400 |
| IL6ST - MUC20 | 1475 | 1093 | 2233 | IL6ST - MUC20 | 1743 | 203 | 1643 |
| IL8 - MUC20 | 1820 | 538 | 2303 | IL8 - MUC20 | 1875 | 883 | 488 |
| IL10RB - MUC20 | 394 | 1884 | 312 | IL10RB - MUC20 | 889 | 1883 | 1947 |
| IL15 - MUC20 | 244 | 2241 | 166 | IL15 - MUC20 | 1412 | 2057 | 1669 |
| IL15RA - MUC20 | 589 | 1406 | 1406 | IL15RA - MUC20 | 1450 | 1902 | 1570 |
| IL17C - MUC20 | 228 | 2278 | 46 | IL17C - MUC20 | 2212 | 1843 | 255 |
| IL17REL - MUC20 | 2121 | 962 | 2267 | IL17REL - MUC20 | 1130 | 1000 | 1868 |
Table 91.
2nd order combinatorial hypotheses between IL and NFkB-2/I family.
| Unexplored combinatorial hypotheses | |
| IL w.r.t MUC | |
| IL-1B/17C | MUC1 |
| IL-1A/1B/1RN/2RG/15/17C | MUC3A |
| IL-1RN/6ST/15RA | MUC4 |
| IL-1A/2RG/8/15/17C | MUC12 |
| IL-15RA | MUC17 |
| IL-1RAP/8/17REL | MUC20 |
| MUC w.r.t IL | |
| IL-1B | MUC1 |
| IL-1RN/2RG/6ST | MUC12 |
| IL-1RAP/15RA | MUC13 |
| IL-1A/10RB/17C | MUC20 |
3.7.4. Interleukin - TP53 Cross Family Analysis
In a new pathway connecting inflammation to cancer, Brighenti et al. [179] show that interleukin 6 downregulates p53 expression and activity by stimulating ribosome biogenesis. Tan et al. [180] show that loss of p53 attenuates the contribution of IL-6 deletion on suppressed tumor progression and extended survival in Kras-driven murine lung cancer. Pützer et al. [181] show that combination therapy with interleukin-2 and wild-type p53 expressed by adenoviral vectors potentiates tumor regression in a murine model of breast cancer. A critical role for p53 in the control of NF-κB-dependent gene expression in TLR4-stimulated dendritic cells exposed to genistein has been shown by Dijsselbloem et al. [182]. The authors previously demonstrated that genistein suppresses TNF-α induced NF-κB-dependent IL-6 gene expression in cancer cells by interfering with the mitogen- and stress-activated protein kinase 1 activation pathway. Schauer et al. [183] show that interleukin-1β promotes ovarian tumorigenesis through a p53/NF-κB-mediated inflammatory response in stromal fibroblasts. These findings indicate connection between IL and TP53 family. Table 92 shows the rankings of IL family w.r.t TP53 family on the left and vice versa on the right.
On the left side, we found IL-17REL to be up regulated with respect to TP53BP2. These are reflected in rankings of 1873 (linear) and 2403 (rbf). IL-15RA was up regulated with respect to TP53I3. These are reflected in rankings of 2069 (laplace), 2079 (linear) and 2228 (rbf) for IL15RA - TP53I3. IL-1RN/2RG/8/10RB/17REL was up regulated with respect to TP53INP1. These are reflected in rankings of 2482 (laplace) and 1911 (linear) for IL1RN - TP53INP1; 2152 (laplace) and 1798 (linear) for IL2RG - TP53INP1; 2388 (linear) and 2343 (rbf) for IL8 - TP53INP1; 2510 (laplace), 2293 (linear) for IL10RB - TP53INP1; and 2505 (linear) and 2509 (rbf) for IL17REL - TP53INP1.
On the right side, we found TP53BP2 to be up regulated with respect to IL-1A/1B/2RG/6ST/8/15/15RA. These are reflected in rankings of 2306 (linear) and 2483 (rbf) for IL1A - TP53BP2; 2003 (laplace) and 2317 (rbf) for IL1B - TP53BP2; 1842 (laplace), 1888 (linear) and 1791 (rbf) for IL2RG - TP53BP2; 1862 (laplace) and 2234 (rbf) for IL6ST - TP53BP2; 2356 (laplace), 2336 (linear) for IL8 - TP53BP2; 2029 (linear) and 1896 (rbf) for IL15 - TP53BP2; 2086 (laplace), 2287 (linear) and 2198 (rbf) for IL15RA - TP53BP2; TP53I3 was up regulated with respect to IL-17REL. This is reflected in rankings of 2268 (laplace) and 2220 (rbf) for IL17REL - TP53I3. TP53INP1 was up regulated with respect to IL2RG. This is reflected in rankings of 2063 (laplace) and 1864 (linear) and 1956 (rbf) IL2RG - TP53INP1. TP53INP2 was up regulated with respect to IL6ST. This is reflected in rankings of 2512 (laplace) and 1952 (linear).
Finally, Table 93 shows the derived influences which can be represented graphically, with the following influences - • IL w.r.t TP53 with IL17REL <- TP53BP2; IL15RA <- TP53I3; IL-1RN/2RG/8/10RB/17REL <- TP53INP1; and • TP53 w.r.t IL with IL-1A/1B/2RG/6ST/8/15/15RA -> TP53BP2; IL17REL -> TP53I3; IL2RG -> TP53INP1; and IL6ST -> TP53INP2.
Table 92.
2nd order combinatorial hypotheses between TP53 and IL.
| Ranking Interleukin family vs TP53 family | |||||||
| Ranking of IL family w.r.t TP53BP2 | Ranking of TP53BP2 w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - TP53BP2 | 2396 | 1377 | 302 | IL1A - TP53BP2 | 390 | 2306 | 2483 |
| IL1B - TP53BP2 | 1868 | 1606 | 16 | IL1B - TP53BP2 | 2003 | 1319 | 2317 |
| IL1RAP - TP53BP2 | 154 | 1863 | 1166 | IL1RAP - TP53BP2 | 1565 | 1196 | 133 |
| IL1RN - TP53BP2 | 320 | 1676 | 1920 | IL1RN - TP53BP2 | 1559 | 1149 | 2489 |
| IL2RG - TP53BP2 | 755 | 377 | 644 | IL2RG - TP53BP2 | 1842 | 1888 | 1791 |
| IL6ST - TP53BP2 | 2237 | 581 | 1526 | IL6ST - TP53BP2 | 1862 | 1530 | 2234 |
| IL8 - TP53BP2 | 1135 | 1279 | 2250 | IL8 - TP53BP2 | 2356 | 2336 | 325 |
| IL10RB - TP53BP2 | 645 | 977 | 289 | IL10RB - TP53BP2 | 420 | 705 | 2040 |
| IL15 - TP53BP2 | 1715 | 281 | 973 | IL15 - TP53BP2 | 879 | 2029 | 1896 |
| IL15RA - TP53BP2 | 1225 | 727 | 567 | IL15RA - TP53BP2 | 2086 | 2287 | 2198 |
| IL17C - TP53BP2 | 2286 | 1214 | 617 | IL17C - TP53BP2 | 1158 | 1243 | 2313 |
| IL17REL - TP53BP2 | 76 | 1873 | 2403 | IL17REL - TP53BP2 | 1526 | 1463 | 1600 |
| Ranking of IL family w.r.t TP53I3 | Ranking of TP53I3 w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - TP53I3 | 1140 | 1547 | 1558 | IL1A - TP53I3 | 283 | 157 | 341 |
| IL1B - TP53I3 | 759 | 333 | 1392 | IL1B - TP53I3 | 156 | 164 | 463 |
| IL1RAP - TP53I3 | 1521 | 885 | 1978 | IL1RAP - TP53I3 | 432 | 605 | 818 |
| IL1RN - TP53I3 | 737 | 340 | 1797 | IL1RN - TP53I3 | 1504 | 1674 | 16 |
| IL2RG - TP53I3 | 7 | 3 | 328 | IL2RG - TP53I3 | 836 | 637 | 134 |
| IL6ST - TP53I3 | 524 | 363 | 981 | IL6ST - TP53I3 | 2157 | 897 | 778 |
| IL8 - TP53I3 | 579 | 485 | 697 | IL8 - TP53I3 | 1921 | 290 | 1265 |
| IL10RB - TP53I3 | 185 | 137 | 758 | IL10RB - TP53I3 | 345 | 1080 | 326 |
| IL15 - TP53I3 | 240 | 244 | 428 | IL15 - TP53I3 | 353 | 1153 | 456 |
| IL15RA - TP53I3 | 2069 | 2079 | 2228 | IL15RA - TP53I3 | 106 | 644 | 1794 |
| IL17C - TP53I3 | 74 | 114 | 647 | IL17C - TP53I3 | 49 | 75 | 37 |
| IL17REL - TP53I3 | 597 | 326 | 1290 | IL17REL - TP53I3 | 2268 | 429 | 2220 |
| Ranking of IL family w.r.t TP53INP1 | Ranking of TP53INP1 w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - TP53INP1 | 2309 | 746 | 7 | IL1A - TP53INP1 | 1049 | 1135 | 1138 |
| IL1B - TP53INP1 | 2281 | 21 | 461 | IL1B - TP53INP1 | 1395 | 1370 | 1684 |
| IL1RAP - TP53INP1 | 531 | 1274 | 2407 | IL1RAP - TP53INP1 | 2223 | 1460 | 680 |
| IL1RN - TP53INP1 | 2482 | 1911 | 891 | IL1RN - TP53INP1 | 1473 | 1252 | 2399 |
| IL2RG - TP53INP1 | 2152 | 1798 | 932 | IL2RG - TP53INP1 | 2063 | 1864 | 1956 |
| IL6ST - TP53INP1 | 591 | 790 | 1740 | IL6ST - TP53INP1 | 537 | 404 | 2042 |
| IL8 - TP53INP1 | 573 | 2388 | 2343 | IL8 - TP53INP1 | 1671 | 1787 | 1014 |
| IL10RB - TP53INP1 | 2510 | 2293 | 1664 | IL10RB - TP53INP1 | 1000 | 2339 | 218 |
| IL15 - TP53INP1 | 663 | 878 | 1116 | IL15 - TP53INP1 | 2147 | 588 | 429 |
| IL15RA - TP53INP1 | 663 | 149 | 169 | IL15RA - TP53INP1 | 1266 | 2264 | 1636 |
| IL17C - TP53INP1 | 2455 | 220 | 435 | IL17C - TP53INP1 | 823 | 523 | 438 |
| IL17REL - TP53INP1 | 83 | 2505 | 2509 | IL17REL - TP53INP1 | 1085 | 1476 | 1393 |
| Ranking of IL family w.r.t TP53INP2 | Ranking of TP53INP2 w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - TP53INP2 | 1481 | 41 | 2490 | IL1A - TP53INP2 | 952 | 505 | 487 |
| IL1B - TP53INP2 | 489 | 310 | 267 | IL1B - TP53INP2 | 200 | 26 | 146 |
| IL1RAP - TP53INP2 | 1159 | 684 | 1263 | IL1RAP - TP53INP2 | 1168 | 757 | 1827 |
| IL1RN - TP53INP2 | 2374 | 779 | 110 | IL1RN - TP53INP2 | 1735 | 1927 | 264 |
| IL2RG - TP53INP2 | 2118 | 103 | 995 | IL2RG - TP53INP2 | 1151 | 539 | 380 |
| IL6ST - TP53INP2 | 261 | 1459 | 333 | IL6ST - TP53INP2 | 2512 | 1952 | 113 |
| IL8 - TP53INP2 | 82 | 679 | 779 | IL8 - TP53INP2 | 2349 | 85 | 1561 |
| IL10RB - TP53INP2 | 865 | 1991 | 67 | IL10RB - TP53INP2 | 653 | 2479 | 236 |
| IL15 - TP53INP2 | 1354 | 989 | 161 | IL15 - TP53INP2 | 1105 | 449 | 1506 |
| IL15RA - TP53INP2 | 1574 | 1545 | 2295 | IL15RA - TP53INP2 | 345 | 488 | 825 |
| IL17C - TP53INP2 | 449 | 56 | 221 | IL17C - TP53INP2 | 1065 | 260 | 116 |
| IL17REL - TP53INP2 | 1325 | 93 | 593 | IL17REL - TP53INP2 | 1251 | 643 | 1832 |
Table 93.
2nd order combinatorial hypotheses between IL and NFkB-2/I family.
| Unexplored combinatorial hypotheses | |
| IL w.r.t TP53 | |
| IL17REL | TP53BP2 |
| IL15RA | TP53I3 |
| IL-1RN/2RG/8/10RB/17REL | TP53INP1 |
| TP53 w.r.t IL | |
| IL-1A/1B/2RG/6ST/8/15/15RA | TP53BP2 |
| IL17REL | TP53I3 |
| IL2RG | TP53INP1 |
| IL6ST | TP53INP2 |
3.7.5. Interleukin - STAT Cross Family Analysis
Jones et al. [184] study the roles of interleukin-6 in activation of STAT proteins and recruitment of neutrophils during Escherichia coli pneumonia. Characterization of the interleukin-4 nuclear activated factor/STAT and its activation independent of the insulin receptor substrate proteins have been studied by Kotanides et al. [185]. Adam et al. [186] have unraveled viral interleukin-6 binding to gp130 and activation of STAT-signaling pathways independently of the interleukin-6 receptor. Frank et al. [187] report the involvement of interleukin 2 signaling in phosphorylation of Stat proteins. Boyd et al. [188] show that interleukin-10 receptor signaling through STAT-3 regulates the apoptosis of retinal ganglion cells in response to stress. Essential role of endocytosis for interleukin-4-receptor-mediated JAK/STAT signalling has been studied in Kurgonaite et al. [189]. Contribution of the interleukin-6/STAT-3 signaling pathway to chondrogenic differentiation of human mesenchymal stem cells has been studied in Kondo et al. [190]. Tanaka et al. [191] show interleukin-10 induces inhibitory C/EBPβ through STAT-3 and represses HIV-1 transcription in macrophages. Jobst et al. [192] show that inhibition of interleukin-3-and interferon-α-induced JAK/STAT signaling by the synthetic α-X-2’,3,4,4’-tetramethoxychalcones α-Br-TMC and α-CF3-TMC. These indicate significant interaction between interleukin family and the STAT family. In CRC cells, treated with ETC-1922159 both were found to be up regulated. The search engine alotted high numerical ranked values to some of the 2nd order combinations between the two. Table 94 indicates the rankings of IL family w.r.t STAT2 family on the left and vice versa on the right.
On the left side, we found IL-1RAP/6ST/17REL to be up regulated with respect to STAT2. These are reflected in rankings of 2111 (laplace), 2258 (linear) and 2012 (rbf) for IL1RAP - STAT2; 2167 (laplace) and 2313 (linear) for IL6ST - STAT2; and 2508 (laplace), 2488 (linear) and 2172 (rbf) for IL17REL - STAT2. IL-1RAP/17REL were up regulated with respect to STAT3. These are reflected in rankings of 2252 (linear) and 2211 (rbf) for IL1RAP - STAT3; and 2282 (linear) and 2517 (rbf) for IL17REL - STAT3; IL-1RAP/15RA were up regulated with respect to STAT5A. These are reflected in rankings of 1768 (laplace) and 2149 (linear) for IL1RAP - STAT5A; and 2342 (laplace) and 2350 (linear) for IL15RA - STAT5A.
On the right side, we found STAT2 to be up regulated with respect to IL-1RAP/1RN/2RG/15RA/ 17C/17REL. These are reflected in rankings of 1826 (laplace) and 2005 (linear) for IL1RAP - STAT2; 2050 (laplace) 2082 (linear) for IL1RN - STAT2; 1986 (laplace) 2021 (linear) and 2031 (rbf) for IL2RG - STAT2; 1988 (linear) and 1863 (rbf) for IL15RA - STAT2; 2473 (linear) and 1883 (rbf) for IL17C - STAT2; 1890 (linear) and 1885 (rbf) for IL17REL - STAT2. STAT3 was up regulated with respect to IL-1RN/2RG. These are reflected in rankings of 2090 (laplace) and 2312 (linear) for IL1RN - STAT3; and 2233 (laplace) and 2146 (linear) IL2RG - STAT3. STAT5A was up regulated with respect to IL-2RG/8/17C. These are reflected in rankings of 1832 (linear) and 2149 (rbf) for IL2RG - STAT5A; 2000 (laplace) and 2386 (linear) for IL8 - STAT5A; and 1760 (laplace), 2060 (linear) and 2201 (rbf) for IL17C - STAT5A.
Finally, Table 95 shows the derived influences which can be represented graphically, with the following influences - • IL w.r.t STAT with IL-1RAP/6ST/17REL <- STAT2; IL-1RAP/17REL <- STAT3 and IL-1RAP/15RA <- STAT5A; and • STAT w.r.t IL with IL-1RN/2RG -> STAT2; IL-1A/1RN/2RG/6ST/15 -> STAT3 and IL-2RG/8/17C -> STAT5A;
Table 94.
2nd order combinatorial hypotheses between STAT and IL.
| Ranking Interleukin family vs STAT family | |||||||
| Ranking of IL family w.r.t STAT2 | Ranking of STAT2 w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - STAT2 | 171 | 207 | 709 | IL1A - STAT2 | 1000 | 687 | 1941 |
| IL1B - STAT2 | 347 | 559 | 188 | IL1B - STAT2 | 1629 | 1019 | 2351 |
| IL1RAP - STAT2 | 2111 | 2258 | 2012 | IL1RAP - STAT2 | 1826 | 2005 | 70 |
| IL1RN - STAT2 | 828 | 1942 | 1226 | IL1RN - STAT2 | 2050 | 2082 | 1030 |
| IL2RG - STAT2 | 939 | 1424 | 272 | IL2RG - STAT2 | 1986 | 2021 | 2031 |
| IL6ST - STAT2 | 2167 | 2313 | 1042 | IL6ST - STAT2 | 1532 | 1766 | 696 |
| IL8 - STAT2 | 806 | 1012 | 69 | IL8 - STAT2 | 397 | 1015 | 2349 |
| IL10RB - STAT2 | 1093 | 2401 | 1260 | IL10RB - STAT2 | 1566 | 1241 | 467 |
| IL15 - STAT2 | 929 | 197 | 446 | IL15 - STAT2 | 1875 | 1724 | 940 |
| IL15RA - STAT2 | 537 | 415 | 1916 | IL15RA - STAT2 | 1406 | 1988 | 1863 |
| IL17C - STAT2 | 175 | 78 | 514 | IL17C - STAT2 | 1199 | 2473 | 1883 |
| IL17REL - STAT2 | 2508 | 2488 | 2172 | IL17REL - STAT2 | 244 | 1890 | 1885 |
| Ranking of IL family w.r.t STAT3 | Ranking of STAT3 w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - STAT3 | 2516 | 173 | 7 | IL1A - STAT3 | 1872 | 1289 | 2350 |
| IL1B - STAT3 | 1628 | 127 | 613 | IL1B - STAT3 | 1367 | 2391 | 901 |
| IL1RAP - STAT3 | 23 | 2252 | 2211 | IL1RAP - STAT3 | 2169 | 1483 | 179 |
| IL1RN - STAT3 | 2309 | 300 | 488 | IL1RN - STAT3 | 2090 | 2312 | 1440 |
| IL2RG - STAT3 | 1168 | 397 | 611 | IL2RG - STAT3 | 2233 | 2146 | 1387 |
| IL6ST - STAT3 | 1355 | 1217 | 381 | IL6ST - STAT3 | 2400 | 2491 | 1953 |
| IL8 - STAT3 | 2353 | 740 | 1176 | IL8 - STAT3 | 1371 | 942 | 2018 |
| IL10RB - STAT3 | 2494 | 1257 | 1320 | IL10RB - STAT3 | 1118 | 406 | 1299 |
| IL15 - STAT3 | 2164 | 903 | 62 | IL15 - STAT3 | 2015 | 2412 | 1356 |
| IL15RA - STAT3 | 1140 | 1572 | 1618 | IL15RA - STAT3 | 1724 | 1638 | 1963 |
| IL17C - STAT3 | 2437 | 30 | 20 | IL17C - STAT3 | 554 | 1446 | 1428 |
| IL17REL - STAT3 | 339 | 2282 | 2517 | IL17REL - STAT3 | 573 | 2181 | 521 |
| Ranking of IL family w.r.t STAT5A | Ranking of STAT5A w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - STAT5A | 1631 | 1110 | 2210 | IL1A - STAT5A | 275 | 464 | 1645 |
| IL1B - STAT5A | 1194 | 1561 | 2223 | IL1B - STAT5A | 416 | 240 | 1659 |
| IL1RAP - STAT5A | 1768 | 1680 | 2149 | IL1RAP - STAT5A | 1852 | 391 | 432 |
| IL1RN - STAT5A | 119 | 285 | 908 | IL1RN - STAT5A | 86 | 2026 | 960 |
| IL2RG - STAT5A | 1136 | 1088 | 1435 | IL2RG - STAT5A | 1367 | 1832 | 2149 |
| IL6ST - STAT5A | 1441 | 2022 | 1697 | IL6ST - STAT5A | 1903 | 436 | 317 |
| IL8 - STAT5A | 1932 | 1543 | 1069 | IL8 - STAT5A | 2000 | 2386 | 4 |
| IL10RB - STAT5A | 897 | 87 | 2033 | IL10RB - STAT5A | 2103 | 1292 | 1326 |
| IL15 - STAT5A | 1116 | 801 | 1653 | IL15 - STAT5A | 436 | 2139 | 1041 |
| IL15RA - STAT5A | 2342 | 2350 | 788 | IL15RA - STAT5A | 621 | 1185 | 1537 |
| IL17C - STAT5A | 984 | 1386 | 2045 | IL17C - STAT5A | 1760 | 2060 | 2201 |
| IL17REL - STAT5A | 1308 | 755 | 3 | IL17REL - STAT5A | 477 | 369 | 992 |
Table 95.
2nd order combinatorial hypotheses between IL and STAT family.
| Unexplored combinatorial hypotheses | |
| IL w.r.t STAT | |
| IL-1RAP/6ST/17REL | STAT2 |
| IL-1RAP/17REL | STAT3 |
| IL-1RAP/15RA | STAT5A |
| STAT w.r.t IL | |
| IL-1RN/2RG | STAT2 |
| IL-1A/1RN/2RG/6ST/15 | STAT3 |
| IL-2RG/8/17C | STAT5A |
3.7.6. Interleukin - TRAF Cross Family Analysis
Greene and O’Neill [193] show that interleukin-1 receptor-associated kinase and TRAF-6 mediate the transcriptional regulation of interleukin-2 by interleukin-1 via NFκB but unlike interleukin-1 are unable to stabilise interleukin-2 mRNA. Cao et al. [194] observe that TRAF6 is a signal transducer for interleukin-1. Schwandner et al. [195] show the requirement of tumor necrosis factor receptor-associated factor (TRAF) 6 in interleukin 17 signal transduction. Lomaga et al. [196] show that TRAF6 deficiency results in osteopetrosis and defective interleukin-1, CD40, and LPS signaling. Jefferies et al. [197] observe that transactivation by the p65 subunit of NF-κB in response to interleukin-1 (IL-1) involves MyD88, IL-1 receptor-associated kinase 1, TRAF-6, and Rac1. Wu and Arron [198] study the role of TRAF6 as a molecular bridge spanning adaptive immunity, innate immunity and osteoimmunology and find relation with the interleukin-1 receptor family. These findings indicate the range of interaction between IL family and TRAF family. In CRC cells treated with ETC-1922159, these were found to be UP regulated. Table 96 show the rankings of IL family w.r.t TRAF family on the left side and vice versa on the right side.
On the left we found, we found IL-1RAP/15RA/17REL to be up regulated with respect to TRAF3IP2. These are reflected in rankings of 2482 (linear) and 2385 (rbf) for IL1RAP - TRAF3IP2; 2024 (laplace), 2162 (linear) and 1800 (rbf) for IL15RA - TRAF3IP2; and 2515 (linear) and 2057 (rbf) for IL17REL - TRAF3IP2. IL-6ST/17REL were up regulated with respect to TRAF4. These are reflected in rankings of 2333 (laplace) and 1914 (rbf) for IL6ST - TRAF4; and (laplace) and 2487 (rbf) for IL17REL - TRAF4 2422; IL-8/17REL were up regulated with respect to TRAF6. These are reflected in rankings of 2088 (laplace), 1883 (linear) and 2089 (rbf) for IL8 - TRAF6; and 2454 (laplace) and 2517 (linear) for IL17REL - TRAF6; IL-6ST were up regulated with respect to TRAFD1. These are reflected in rankings of 1835 (laplace) and 1824 (linear) for IL6ST - TRAFD1.
On the right we found, we found TRAF3IP2 was up regulated with respect to IL-1B/2RG/6ST/8/17REL. These are reflected in rankings of 1953 (laplace) and 2359 (rbf) for IL1B - TRAF3IP2; 1767 (laplace), 2385 (linear) and 2059 (rbf) for IL2RG - TRAF3IP2; 1991 (linear) and 1871 (rbf) for IL6ST - TRAF3IP2; 2192 (linear) and 2289 (rbf) for IL8 - TRAF3IP2; and 1836 (laplace) and 2042 (linear) for IL17REL - TRAF3IP2. TRAF4 was up regulated with respect to IL-10RB/15/15RA. These are reflected in rankings of 2407 (laplace) and 1781 (linear) for IL10RB - TRAF4; 2408 (linear) and 1759 (rbf) for IL15 - TRAF4; and 2408 (linear) and 1759 (rbf) for IL15RA - TRAF4; TRAF6 was up regulated with respect to IL-1RAP/8/15/17C. These are reflected in rankings of 2219 (laplace), 1984 (linear) and 1766 (rbf) for IL1RAP - TRAF6; 2457 (laplace) and 2139 (linear) for IL8 - TRAF6; 2071 (lapalce) and 2475 (linear) for IL15 - TRAF6; and 2469 (laplace) and 2309 (linear) for IL17C - TRAF6;
Finally, Table 97 shows the derived influences which can be represented graphically, with the following influences - • IL w.r.t TRAF with IL-1RAP/15RA/17REL <- TRAF3IP2; IL-6ST/17REL <- TRAF4; IL-8/17REL <- TRAF6; and IL-6ST <- TRAFD1; and • TRAF w.r.t IL with IL-1B/2RG/6ST/8/17REL -> TRAF3IP2; IL-10RB/15/15RA -> TRAF4 and IL-1RAP/8/15/17C -> TRAF6.
3.7.7. Interleukin - Metalloreductase STEAP4 Cross Family Analysis
Ramadoss et al. [199] show that C/EBPα also regulates hepatic expression of STEAP4 during feeding, whereas both C/EBPα and STAT3 regulate expression of steap4 in the presence of high levels of IL-6. Also, Tanaka et al. [120] show STEAP4 is a tumor necrosis factor alpha-induced protein that regulates IL-6, IL-8, and cell proliferation in synovium from patients with rheumatoid arthritis. Gauss et al. [121] observe that the STEAP4 expression in adipocytes is normally induced by nutritional stress, leptin, and proinflammatory cytokines, including TNF-α, interleukin-1β, and interleukin-6. These were found to be up regulated in CRC cells treated with ETC-1922159. Table 98 shows the interaction between the IL family and STEAP4. We found that IL-8/10RB/17C/17REL was up regulated w.r.t STEAP4. These are reflected in rankings of 2204 (laplace) and 1987 (linear) for IL8 - STEAP4; 2422 (laplace) and 2310 (linear) for IL10RB - STEAP4; 2103 (linear) and 1889 (rbf) for IL17C - STEAP4; and 1965 (linear) and 2297 (rbf) for IL17REL - STEAP4; Also STEAP4 was up regulated w.r.t IL-1A/1RAP/1RN/15. These are reflected in rankings of 2358 (linear) and 2223 (rbf) for IL1A - STEAP4; 1871 (laplace), 1898 (linear) and 2077 (rbf) for IL1RAP - STEAP4; 2043 (linear) and 1763 (rbf) for IL1RN - STEAP4; and 1965 (linear) and 2283 (rbf) for IL15 - STEAP4;
Finally, Table 99 shows the derived influences which can be represented graphically, with the following influences - • IL w.r.t STEAP4 with IL-8/10RB/17C/17REL <- STEAP4 • STEAP4 w.r.t IL with IL-1A/1RAP/1RN/15 -> STEAP4.
Table 98.
2nd order combinatorial hypotheses between STEAP4 and IL.
| Ranking Interleukin family vs STEAP4 family | |||||||
| Ranking of IL family w.r.t STEAP4 | Ranking of STEAP4 w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - STEAP4 | 422 | 482 | 992 | IL1A - STEAP4 | 71 | 2358 | 2223 |
| IL1B - STEAP4 | 423 | 814 | 982 | IL1B - STEAP4 | 240 | 1570 | 1863 |
| IL1RAP - STEAP4 | 2092 | 262 | 661 | IL1RAP - STEAP4 | 1871 | 1898 | 2077 |
| IL1RN - STEAP4 | 404 | 1602 | 370 | IL1RN - STEAP4 | 195 | 2043 | 1763 |
| IL2RG - STEAP4 | 1293 | 1458 | 1323 | IL2RG - STEAP4 | 299 | 1562 | 1284 |
| IL6ST - STEAP4 | 920 | 1641 | 2424 | IL6ST - STEAP4 | 1374 | 504 | 1628 |
| IL8 - STEAP4 | 2204 | 1987 | 1558 | IL8 - STEAP4 | 794 | 1049 | 1615 |
| IL10RB - STEAP4 | 2422 | 2310 | 1179 | IL10RB - STEAP4 | 476 | 254 | 906 |
| IL15 - STEAP4 | 700 | 1154 | 2320 | IL15 - STEAP4 | 288 | 1965 | 2283 |
| IL15RA - STEAP4 | 2277 | 1114 | 1528 | IL15RA - STEAP4 | 1170 | 1334 | 1347 |
| IL17C - STEAP4 | 433 | 2103 | 1889 | IL17C - STEAP4 | 17 | 2426 | 1108 |
| IL17REL - STEAP4 | 33 | 1965 | 2297 | IL17REL - STEAP4 | 2439 | 715 | 100 |
Table 99.
2nd order combinatorial hypotheses between IL and STEAP4 family.
| Unexplored combinatorial hypotheses | |
| IL w.r.t STEAP4 | |
| IL-8/10RB/17C/17REL | STEAP4 |
| STEAP4 w.r.t IL | |
| IL-1A/1RAP/1RN/15 | STEAP4 |
3.7.8. Interleukin - Metalloreductase STEAP3 Cross Family Analysis
Based on the interactions of STEAP4 and interleukin, we also generated rankings for STEAP3 and interleukin family. It was found that STEAP3 and interleukin family were down regulated. Table 100 shows the rankings of IL family w.r.t STEAP3 and vice versa. We found IL-1RL2/17D/17RB/17RD/33/F2/F3.AS1 to be down regulated w.r.t STEAP3. These are reflected in rankings of 619 (laplace) and 1471 (linear) for IL1RL2 - STEAP3; 1338 (laplace), 1275 (linear) and 458 (rbf) for IL17D - STEAP3; 1101 (laplace) and 239 (rbf) for IL17RB - STEAP3; 1323 (laplace) and 810 (linear) for IL17RD - STEAP3; 1589 (laplace) and 781 (linear) and 1210 (rbf) for IL33 - STEAP3; 1571 (laplace) and 811 (linear) and 579 (rbf) for ILF2 - STEAP3; and 947 (laplace) and 926 (rbf) for ILF3.AS1 - STEAP3. STEAP3 to be down regulated w.r.t IL-1RL2/17D/17RB/33/F3/F3.AS1. These are reflected in rankings of 835 (laplace) and 1733 (rbf) for IL1RL2 - STEAP3; 596 (laplace) and 705 (linear) for IL17D - STEAP3; 208 (laplace) and 404 (rbf) for IL17RB - STEAP3; 1070 (laplace) and 57 (linear) for IL33 - STEAP3; 121 (laplace) and 926 (rbf) for ILF3 - STEAP3 and 1592 (laplace), 678 (linear) and 1094 (rbf) for ILF3.AS1 - STEAP3.
Finally, Table 101 shows the derived influences which can be represented graphically, with the following influences - • IL w.r.t STEAP3 with IL-1RL2/17D/17RB/17RD/33/F2/F3.AS1 <- STEAP3; and • STEAP3 w.r.t IL with IL-1RL2/17D/17RB/33/F3/F3.AS1 -> STEAP3.
Table 100.
2nd order combinatorial hypotheses between STEAP3 and IL.
| Ranking Interleukin family vs STEAP3 family | |||||||
| Ranking of IL family w.r.t STEAP3 | Ranking of STEAP3 w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1RL2 - STEAP3 | 619 | 1471 | 2246 | IL1RL2 - STEAP3 | 835 | 2234 | 1733 |
| IL17D - STEAP3 | 1338 | 1275 | 458 | IL17D - STEAP3 | 596 | 705 | 2273 |
| IL17RB - STEAP3 | 1101 | 2302 | 239 | IL17RB - STEAP3 | 208 | 2462 | 404 |
| IL17RD - STEAP3 | 1323 | 810 | 1834 | IL17RD - STEAP3 | 2352 | 589 | 2233 |
| IL33 - STEAP3 | 1589 | 781 | 1210 | IL33 - STEAP3 | 1070 | 57 | 2098 |
| ILF2 - STEAP3 | 1571 | 811 | 579 | ILF2 - STEAP3 | 1986 | 1029 | 2474 |
| ILF3 - STEAP3 | 261 | 1866 | 1953 | ILF3 - STEAP3 | 121 | 2314 | 926 |
| ILF3.AS1 - STEAP3 | 947 | 2255 | 926 | ILF3.AS1 - STEAP3 | 1592 | 678 | 1094 |
Table 101.
2nd order combinatorial hypotheses between IL and STEAP3 family.
| Unexplored combinatorial hypotheses | |
| IL w.r.t STEAP3 | |
| IL-1RL2/17D/17RB/17RD/33/F2/F3.AS1 - STEAP3 | |
| STEAP3 w.r.t IL | |
| IL-1RL2/17D/17RB/33/F3/F3.AS1 - STEAP3 | |
3.7.9. Interleukin - ATP-Binding Cassette Transporters
Haskó et al. [159] show that the inhibitors of ATP-binding cassette transporters suppress interleukin-12 p40 production and major histocompatibility complex II up-regulation in macrophages. Marty et al. [161] observe that ATP binding cassette transporter ABC1 is required for the release of interleukin-1β by P2X7-stimulated and lipopolysaccharide-primed mouse Schwann cells. Hamon et al. [200] observe that interleukin-1β secretion is impaired by inhibitors of the ATP binding cassette transporter, ABC1. Lottaz et al. [162] show that inhibition of ATP-binding cassette transporter downregulates interleukin-1β-mediated autocrine activation of human dermal fibroblasts. These findings indicate the interaction of ABC transporters with Interleukin family. In CRC cells, treated with ETC-1922159 these were found to be down regulated. Table 102 shows rankings of IL family with respect to a few ABC members on the left and vice versa on the right.
Table 102.
2nd order combinatorial hypotheses between ABC and IL.
| Ranking Interleukin family vs ABC family | |||||||
| Ranking of IL family w.r.t ABCA2 | Ranking of ABCA2 w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1RL2 - ABCA2 | 2055 | 2097 | 405 | IL1RL2 - ABCA2 | 2022 | 2490 | 1234 |
| IL17D - ABCA2 | 1778 | 2160 | 1120 | IL17D - ABCA2 | 540 | 227 | 1006 |
| IL17RB - ABCA2 | 2419 | 1404 | 1727 | IL17RB - ABCA2 | 2146 | 1543 | 1991 |
| IL17RD - ABCA2 | 2202 | 1799 | 358 | IL17RD - ABCA2 | 1717 | 1671 | 517 |
| IL33 - ABCA2 | 1076 | 1707 | 1854 | IL33 - ABCA2 | 1507 | 497 | 743 |
| ILF2 - ABCA2 | 944 | 1054 | 2607 | ILF2 - ABCA2 | 831 | 822 | 752 |
| ILF3 - ABCA2 | 1380 | 1369 | 1702 | ILF3 - ABCA2 | 1691 | 2094 | 2275 |
| ILF3.AS1 - ABCA2 | 2243 | 1006 | 1924 | ILF3.AS1 - ABCA2 | 2058 | 1664 | 2165 |
| Ranking of IL family w.r.t ABCE1 | Ranking of ABCE1 w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1RL2 - ABCE1 | 906 | 1403 | 2365 | IL1RL2 - ABCE1 | 525 | 2034 | 723 |
| IL17D - ABCE1 | 1531 | 636 | 753 | IL17D - ABCE1 | 1432 | 2146 | 1401 |
| IL17RB - ABCE1 | 459 | 2056 | 1993 | IL17RB - ABCE1 | 1090 | 2618 | 263 |
| IL17RD - ABCE1 | 1030 | 1332 | 1565 | IL17RD - ABCE1 | 1523 | 727 | 2185 |
| IL33 - ABCE1 | 1649 | 719 | 937 | IL33 - ABCE1 | 2619 | 808 | 2025 |
| ILF2 - ABCE1 | 20 | 310 | 560 | ILF2 - ABCE1 | 2650 | 331 | 2103 |
| ILF3 - ABCE1 | 2410 | 2409 | 1826 | ILF3 - ABCE1 | 1767 | 2674 | 19 |
| ILF3.AS1 - ABCE1 | 1154 | 2222 | 786 | ILF3.AS1 - ABCE1 | 1788 | 1948 | 820 |
| Ranking of IL family w.r.t ABCF2 | Ranking of ABCF2 w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1RL2 - ABCF2 | 1031 | 1806 | 2002 | IL1RL2 - ABCF2 | 2257 | 818 | 1274 |
| IL17D - ABCF2 | 2481 | 2016 | 1006 | IL17D - ABCF2 | 796 | 2104 | 568 |
| IL17RB - ABCF2 | 509 | 1294 | 2302 | IL17RB - ABCF2 | 1271 | 621 | 1631 |
| IL17RD - ABCF2 | 610 | 1935 | 1084 | IL17RD - ABCF2 | 957 | 2276 | 1431 |
| IL33 - ABCF2 | 735 | 2050 | 1855 | IL33 - ABCF2 | 421 | 1781 | 252 |
| ILF2 - ABCF2 | 2093 | 1104 | 2073 | ILF2 - ABCF2 | 683 | 2304 | 529 |
| ILF3 - ABCF2 | 812 | 1686 | 1080 | ILF3 - ABCF2 | 1243 | 585 | 1452 |
| ILF3.AS1 - ABCF2 | 430 | 2416 | 1983 | ILF3.AS1 - ABCF2 | 2272 | 1169 | 862 |
Table 103.
2nd order combinatorial hypotheses between IL and ABC family.
| Unexplored combinatorial hypotheses | |
| IL w.r.t ABC | |
| IL-1RB/33/F2/F3 | ABCA2 |
| IL-1RL2/17D/17RD/33/F2/F3.AS1 | ABCE1 |
| IL-17RB/17RD/F3 | ABCF2 |
| ABC w.r.t IL | |
| IL-17D/17RD/33/F2 | ABCA2 |
| IL-1RL2/17D/17RB/17RD | ABCE1 |
| IL-1RL2/17D/17RB/17RD/33/F2/F3/F3.AS1 | ABCF2 |
On the left we found IL-1RB/33/F2/F3 were down regulated w.r.t ABCA2. These are reflected in rankings of 1404 (linear) and 1727 (rbf) for IL17RB - ABCA2; 1076 (laplace), 1707 (linear) for IL33 - ABCA2; 944 (laplace) and 1054 (linear) for ILF2 - ABCA2; 1380 (laplace), 1369 (linear) and 1702 (rbf) for ILF3 - ABCA2; IL-1RL2/17D/17RD/33/F2/F3.AS1 were up regulated w.r.t ABCE1. These are reflected in rankings of 906 (laplace) and 1403 (linear) for IL1RL2 - ABCE1; 1531 (laplace), 636 (linear) and 753 (rbf) for IL17D - ABCE1; 1030 (laplace), 1332 (linear) and 1565 (rbf) for IL17RD - ABCE1; 1649 (laplace), 719 (linear) and 937 (rbf) for IL33 - ABCE1; 20 (laplace), 310 (linear) and 560 (rbf) for ILF2 - ABCE1; and 1154 (laplace) and 786 (rbf) for ILF3.AS1 - ABCE1. IL-17RB/17RD/F3 were up regulated w.r.t ABCF2. These are reflected in rankings of 509 (laplace) and 1294 (laplace) for IL17RB - ABCF2; 610 (laplace) and 1084 (rbf) for IL17RD - ABCF2; and 812 (laplace), 1686 (laplace) and 1080 (rbf) for ILF3 - ABCF2.
On the right, we found ABCA2 were up regulated w.r.t IL-17D/17RD/33/F2. These are reflected in rankings of 540 (laplace), 227 (linear) and 1006 (rbf) for IL17D - ABCA2; 1717 (laplace), 1671 (linear) and 517 (rbf) for IL17RD - ABCA2; 1507 (laplace), 497 (linear) and 743 (rbf) for IL33 - ABCA2; and 831 (laplace), 822 (linear) and 752 (rbf) for ILF2 - ABCA2; ABCE1 were up regulated w.r.t IL-1RL2/17D/17RB/17RD. These are reflected in rankings of 525 (laplace) and 723 (rbf) for IL1RL2 - ABCE1; 1432 (laplace) and 1401 (rbf) for IL17D - ABCE1; 1090 (laplace) and 263 (rbf) for IL17RB - ABCE1; and 1523 (laplace) and 727 (linear) for IL17RD - ABCE1; ABCF2 were up regulated w.r.t IL-1RL2/17D/17RB/17RD/33/F2/F3/F3.AS1. These are reflected in rankings of 818 (rbf) and 1274 (rbf) for IL1RL2 - ABCF2; 796 (laplace) and 568 (rbf) for IL17D - ABCF2; 1271 (laplace), 621 (linear) and 1631 (rbf) for IL17RB - ABCF2; 957 (laplace) and 1431 (rbf) for IL17RD - ABCF2; 421 (laplace) and 252 (rbf) for IL33 - ABCF2; 683 (laplace) and 529 (rbf) for ILF2 - ABCF2; 1243 (laplace), 585 (linear) and 1452 (rbf) for ILF3 - ABCF2 and 1169 (linear) and 862 (rbf) for ILF3.AS1 - ABCF2.
Finally, Table 103 shows the derived influences which can be represented graphically, with the following influences - • IL w.r.t ABC with IL-1RB/33/F2/F3 <- ABCA2; IL-1RL2/17D/17RD/33/F2/ F3.AS1 <- ABCE1 and IL-17RB/17RD/F3 <- ABCF2. • ABC w.r.t IL with IL-17D/17RD/33/F2 -> ABCA2; IL-1RL2/17D/17RB/17RD -> ABCE1 and IL-1RL2/17D/17RB/17RD/33/F2/F3/F3.AS1 -> ABCF2.
3.7.10. Interleukin - TNF Cross Family Analysis
Neta et al. [201] study the relationship of TNF to interleukins way back in 1992. The review by Rieckmann et al. [202] studies role of TNF-α and IL-6 in normal and pathophysiological conditions of B-cell function. Bethea et al. [203] demonstrate that IL-1β induces TNF-α gene expression in CH235-MG cells in a protein kinase C-dependent manner. Tumor necrosis factor (TNF)-α and interleukin (IL)-1β down-regulate intercellular adhesion molecule (ICAM)-2 expression on the endothelium as shown by McLaughlin et al. [204]. Zhai et al. [205] suggest that serum levels of tumor necrosis factor-α receptors and interleukin 6 (IL-6) are associated with the fibrotic process of coal workers’ pneumoconiosis (CWP) and serum cytokine levels may be correlated with the severity of CWP. However, in arthritic conditions, Koenders: 2006 interleukin show that Interleukin-17 acts independently of TNF-α. Serum interleukin-6 (IL-6), IL-10, tumor necrosis factor (TNF) alpha, soluble type II TNF receptor, and transforming growth factor beta levels in human immunodeficiency virus type 1-infected individuals with Mycobacterium avium complex disease have been studied by Havlir et al. [206]. Tissi et al. [207] study the role of tumor necrosis factor alpha, interleukin-1β, and interleukin-6 in a mouse model of group B streptococcal arthritis. They conclude that their results account for a strong involvement of IL-1β and IL-6, but not of TNF-α, in the pathogenesis of GBS arthritis. Ismail et al. [208] study the role of tumor necrosis factor alpha (TNF-α) and interleukin-10 in the pathogenesis of severe murine monocytotropic ehrlichiosis. Their data suggest that the balance between TNF-α and IL-10 produced by either macrophages or T cells in response to infection with Ehrlichia may modulate the induction of apoptosis during the infection. Yap et al. [209] observe that Tumor necrosis factor (TNF) inhibits interleukin (IL)-1 and/or IL-6 stimulated synthesis of C-reactive protein (CRP) and serum amyloid A (SAA) in primary cultures of human hepatocytes. These findings suggest interactive role of IL and TNF family in a synergistic way. In CRC cells treated with ETC-1922159, both were found to be up regulated. The search engine assigned high valued numerical ranks to 2nd order combinations of IL and TNF family members. These are tabulated in Table 104, Table 105, Table 106 and Table 107. The left side contains rankings of IL w.r.t TNF family and the right side contains rankings of TNF family w.r.t IL.
Table 104.
2nd order combinatorial hypotheses between TNF and IL.
| Ranking Interleukin family vs TNF family | |||||||
| Ranking of IL family w.r.t TNF | Ranking of TNF w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - TNF | 1382 | 727 | 725 | IL1A - TNF | 172 | 660 | 230 |
| IL1B - TNF | 519 | 539 | 187 | IL1B - TNF | 443 | 458 | 244 |
| IL1RAP - TNF | 1475 | 1995 | 2255 | IL1RAP - TNF | 564 | 550 | 1500 |
| IL1RN - TNF | 163 | 106 | 609 | IL1RN - TNF | 292 | 462 | 276 |
| IL2RG - TNF | 276 | 820 | 340 | IL2RG - TNF | 419 | 708 | 1035 |
| IL6ST - TNF | 2374 | 2037 | 2003 | IL6ST - TNF | 2410 | 1901 | 666 |
| IL8 - TNF | 921 | 1325 | 1148 | IL8 - TNF | 1072 | 206 | 118 |
| IL10RB - TNF | 346 | 595 | 339 | IL10RB - TNF | 2065 | 2120 | 2296 |
| IL15 - TNF | 242 | 944 | 616 | IL15 - TNF | 265 | 828 | 279 |
| IL15RA - TNF | 2341 | 1843 | 2195 | IL15RA - TNF | 131 | 914 | 1488 |
| IL17C - TNF | 906 | 1573 | 776 | IL17C - TNF | 2148 | 568 | 280 |
| IL17REL - TNF | 296 | 804 | 677 | IL17REL - TNF | 1223 | 1901 | 11 |
| Ranking of IL family w.r.t TNFAIP1 | Ranking of TNFAIP1 w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - TNFAIP1 | 2515 | 549 | 1534 | IL1A - TNFAIP1 | 533 | 1901 | 1548 |
| IL1B - TNFAIP1 | 2398 | 440 | 2449 | IL1B - TNFAIP1 | 1324 | 756 | 1062 |
| IL1RAP - TNFAIP1 | 326 | 866 | 2226 | IL1RAP - TNFAIP1 | 1555 | 1284 | 1291 |
| IL1RN - TNFAIP1 | 1952 | 649 | 1453 | IL1RN - TNFAIP1 | 1567 | 307 | 979 |
| IL2RG - TNFAIP1 | 1791 | 104 | 2482 | IL2RG - TNFAIP1 | 421 | 973 | 1169 |
| IL6ST - TNFAIP1 | 156 | 1415 | 1062 | IL6ST - TNFAIP1 | 1281 | 104 | 2086 |
| IL8 - TNFAIP1 | 456 | 682 | 1389 | IL8 - TNFAIP1 | 2293 | 2126 | 752 |
| IL10RB - TNFAIP1 | 97 | 425 | 2020 | IL10RB - TNFAIP1 | 716 | 2092 | 569 |
| IL15 - TNFAIP1 | 367 | 1392 | 159 | IL15 - TNFAIP1 | 24 | 436 | 324 |
| IL15RA - TNFAIP1 | 1860 | 1979 | 611 | IL15RA - TNFAIP1 | 873 | 2141 | 1853 |
| IL17C - TNFAIP1 | 2382 | 1072 | 2446 | IL17C - TNFAIP1 | 961 | 2143 | 791 |
| IL17REL - TNFAIP1 | 307 | 79 | 161 | IL17REL - TNFAIP1 | 1603 | 1462 | 1764 |
| Ranking of IL family w.r.t TNFAIP2 | Ranking of TNFAIP2 w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - TNFAIP2 | 219 | 1815 | 790 | IL1A - TNFAIP2 | 450 | 1041 | 465 |
| IL1B - TNFAIP2 | 210 | 1123 | 538 | IL1B - TNFAIP2 | 1923 | 557 | 944 |
| IL1RAP - TNFAIP2 | 1535 | 660 | 1525 | IL1RAP - TNFAIP2 | 105 | 229 | 845 |
| IL1RN - TNFAIP2 | 1769 | 2475 | 683 | IL1RN - TNFAIP2 | 957 | 868 | 839 |
| IL2RG - TNFAIP2 | 1358 | 576 | 188 | IL2RG - TNFAIP2 | 415 | 1132 | 613 |
| IL6ST - TNFAIP2 | 2007 | 633 | 1704 | IL6ST - TNFAIP2 | 1649 | 929 | 1558 |
| IL8 - TNFAIP2 | 769 | 331 | 368 | IL8 - TNFAIP2 | 1262 | 1412 | 1595 |
| IL10RB - TNFAIP2 | 2319 | 2497 | 719 | IL10RB - TNFAIP2 | 93 | 1583 | 204 |
| IL15 - TNFAIP2 | 1362 | 2383 | 795 | IL15 - TNFAIP2 | 537 | 749 | 120 |
| IL15RA - TNFAIP2 | 2032 | 821 | 1502 | IL15RA - TNFAIP2 | 519 | 737 | 1146 |
| IL17C - TNFAIP2 | 868 | 1684 | 1770 | IL17C - TNFAIP2 | 199 | 424 | 687 |
| IL17REL - TNFAIP2 | 279 | 563 | 299 | IL17REL - TNFAIP2 | 2057 | 437 | 2008 |
Table 105.
2nd order combinatorial hypotheses between TNF and IL.
| Ranking Interleukin family vs TNF family | |||||||
| Ranking of IL family w.r.t TNFAIP3 | Ranking of TNFAIP3 w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - TNFAIP3 | 2307 | 319 | 108 | IL1A - TNFAIP3 | 78 | 51 | 2058 |
| IL1B - TNFAIP3 | 495 | 98 | 339 | IL1B - TNFAIP3 | 140 | 146 | 520 |
| IL1RAP - TNFAIP3 | 30 | 2428 | 1376 | IL1RAP - TNFAIP3 | 1802 | 1610 | 903 |
| IL1RN - TNFAIP3 | 579 | 277 | 299 | IL1RN - TNFAIP3 | 60 | 1610 | 1320 |
| IL2RG - TNFAIP3 | 1705 | 330 | 125 | IL2RG - TNFAIP3 | 1056 | 1608 | 2333 |
| IL6ST - TNFAIP3 | 2068 | 2432 | 2282 | IL6ST - TNFAIP3 | 1652 | 1470 | 1507 |
| IL8 - TNFAIP3 | 1918 | 2255 | 1587 | IL8 - TNFAIP3 | 2224 | 1717 | 118 |
| IL10RB - TNFAIP3 | 1576 | 666 | 1377 | IL10RB - TNFAIP3 | 1073 | 417 | 943 |
| IL15 - TNFAIP3 | 732 | 254 | 273 | IL15 - TNFAIP3 | 907 | 628 | 684 |
| IL15RA - TNFAIP3 | 727 | 1547 | 1476 | IL15RA - TNFAIP3 | 1340 | 445 | 1031 |
| IL17C - TNFAIP3 | 1675 | 222 | 138 | IL17C - TNFAIP3 | 1105 | 1887 | 866 |
| IL17REL - TNFAIP3 | 2364 | 2503 | 2283 | IL17REL - TNFAIP3 | 2040 | 1143 | 1486 |
| Ranking of IL family w.r.t TNFRSF1A | Ranking of TNFRSF1A w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - TNFRSF1A | 1556 | 2184 | 1375 | IL1A - TNFRSF1A | 2028 | 113 | 226 |
| IL1B - TNFRSF1A | 1621 | 1917 | 446 | IL1B - TNFRSF1A | 147 | 2027 | 2247 |
| IL1RAP - TNFRSF1A | 1236 | 2500 | 2293 | IL1RAP - TNFRSF1A | 1339 | 1003 | 2062 |
| IL1RN - TNFRSF1A | 411 | 1571 | 755 | IL1RN - TNFRSF1A | 1713 | 387 | 102 |
| IL2RG - TNFRSF1A | 565 | 2350 | 574 | IL2RG - TNFRSF1A | 1191 | 597 | 1479 |
| IL6ST - TNFRSF1A | 2221 | 1465 | 561 | IL6ST - TNFRSF1A | 1143 | 291 | 225 |
| IL8 - TNFRSF1A | 1536 | 750 | 304 | IL8 - TNFRSF1A | 1483 | 669 | 673 |
| IL10RB - TNFRSF1A | 620 | 35 | 1791 | IL10RB - TNFRSF1A | 230 | 1510 | 385 |
| IL15 - TNFRSF1A | 345 | 489 | 384 | IL15 - TNFRSF1A | 157 | 838 | 425 |
| IL15RA - TNFRSF1A | 442 | 1155 | 697 | IL15RA - TNFRSF1A | 682 | 322 | 1575 |
| IL17C - TNFRSF1A | 1113 | 284 | 149 | IL17C - TNFRSF1A | 5 | 169 | 122 |
| IL17REL - TNFRSF1A | 766 | 336 | 249 | IL17REL - TNFRSF1A | 1547 | 452 | 22 |
| Ranking of IL family w.r.t TNFRSF10A | Ranking of TNFRSF10A w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - TNFRSF10A | 366 | 73 | 48 | IL1A - TNFRSF10A | 1972 | 1805 | 2504 |
| IL1B - TNFRSF10A | 317 | 45 | 367 | IL1B - TNFRSF10A | 2375 | 2373 | 2320 |
| IL1RAP - TNFRSF10A | 2104 | 1342 | 2027 | IL1RAP - TNFRSF10A | 981 | 1665 | 2504 |
| IL1RN - TNFRSF10A | 1739 | 346 | 173 | IL1RN - TNFRSF10A | 1261 | 2287 | 2469 |
| IL2RG - TNFRSF10A | 645 | 1448 | 1009 | IL2RG - TNFRSF10A | 1244 | 2246 | 2467 |
| IL6ST - TNFRSF10A | 1307 | 823 | 1778 | IL6ST - TNFRSF10A | 2128 | 2320 | 1738 |
| IL8 - TNFRSF10A | 402 | 1615 | 1908 | IL8 - TNFRSF10A | 566 | 733 | 2117 |
| IL10RB - TNFRSF10A | 1243 | 689 | 2119 | IL10RB - TNFRSF10A | 389 | 532 | 723 |
| IL15 - TNFRSF10A | 321 | 1602 | 358 | IL15 - TNFRSF10A | 2414 | 2260 | 1705 |
| IL15RA - TNFRSF10A | 2126 | 2342 | 148 | IL15RA - TNFRSF10A | 2398 | 1970 | 2088 |
| IL17C - TNFRSF10A | 981 | 269 | 1027 | IL17C - TNFRSF10A | 1831 | 2025 | 1718 |
| IL17REL - TNFRSF10A | 2497 | 2470 | 2109 | IL17REL - TNFRSF10A | 1034 | 1482 | 2068 |
On the left side, we found IL-1RAP/6ST/15RA to be up regulated w.r.t TNF. These are reflected in the rankings of 1995 (linear) and 2255 (rbf) for IL1RAP - TNF; 2374 (laplace), 2037 (linear) and 2003 (rbf) for IL6ST - TNF; 2341 (laplace), 1843 (linear) and 2195 (rbf) for IL15RA - TNF; IL-1B/2RG/15RA/17C were up regulated w.r.t TNFAIP1. These are reflected in the rankings of 2398 (laplace) and 2449 (rbf) for IL1B - TNFAIP1; 1791 (laplace) and 2482 (rbf) for IL2RG - TNFAIP1; 1860 (laplace) and 1979 (linear) for IL15RA - TNFAIP1; 2382 (laplace) and 2446 (rbf) for IL17C - TNFAIP1. IL-1RN/10RB were up regulated w.r.t TNFAIP2. These are reflected in the rankings of 1769 (laplace) and 2475 (rbf) for IL1RN - TNFAIP2; and 2319 (laplace) and 2497 (rbf) for IL10RB - TNFAIP2; IL-6ST/8/17REL were up regulated w.r.t TNFAIP3. These are reflected in the rankings of 2068 (laplace), 2432 (linear) and 2282 (rbf) for IL6ST - TNFAIP3; 1918 (laplace) and 2255 (linear) for IL8 - TNFAIP3; and 2364 (laplace), 2503 (linear) and 2283 (rbf) for IL17REL - TNFAIP3; IL-1RAP was up regulated w.r.t TNFRSF1A. This is reflected in the rankings of 2500 (linear) and 2293 (rbf) for IL1RAP - TNFRSF1A; IL-1RAP/15RA/17REL were up regulated w.r.t TNFRSF10A. These are reflected in the rankings of 2104 (laplace) and 2027 (rbf) for IL1RAP - TNFRSF10A; 2126 (laplace), 2342 (linear) for IL15RA - TNFRSF10A; 2497 (laplace), 2470 (linear) and 2109 (rbf) for IL17REL - TNFRSF10A; IL-15RA was up regulated w.r.t TNFRSF10B. This is reflected in the rankings of 2330 (laplace) and 1932 (rbf) for IL15RA - TNFRSF10B; IL-15RA was up regulated w.r.t TNFRSF10D. This is reflected in the rankings of 2197 (laplace) and 2126 (rbf) for IL-15RA - TNFRSF10D; IL-8/15RA/17REL were up regulated w.r.t TNFRSF12A. These are reflected in the rankings of 1827 (linear) and 2355 (rbf) for IL8 - TNFRSF12A; 2138 (laplace), 2090 (linear) and 1981 (rbf) for IL15RA - TNFRSF12A; 2475 (laplace) and 2496 (rbf) for IL17REL - TNFRSF12A. IL-15RA was up regulated w.r.t TNFRSF14. This is reflected in the rankings of 2378 (laplace) and 1929 (rbf) for IL-15RA - TNFRSF14; IL-1B/1RAP/2RG were up regulated w.r.t TNFRSF21. These are reflected in the rankings of 1862 (laplace), 2164 (linear), 2305 (rbf) for IL1B - TNFRSF21; 1762 (linear) and 2163 (rbf) for IL1RAP - TNFRSF21; and 2297 (linear) and 2351 (rbf) for IL2RG - TNFRSF21; IL-1B/15RA/17C were up regulated w.r.t TNFRSF10. These are reflected in the rankings of 2448 (linear) and 1993 (rbf) for IL1B - TNFSF10; 2163 (linear) and 2059 (rbf) for IL15RA - TNFSF10; and 2337 (linear) and 2431 (rbf) for IL17C - TNFSF10. IL-15RA/17C to be up regulated w.r.t TNFSF15. This is reflected in the rankings of 2222 (laplace) and 2328 (linear) for IL-17C - TNFSF15; and 2124 (laplace) and 2365 (rbf) for IL15RA - TNFSF15;
Table 106.
2nd order combinatorial hypotheses between IL and TNF.
| Ranking Interleukin family vs TNF family | |||||||
| Ranking of IL family w.r.t TNFRSF10B | Ranking of TNFRSF10B w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - TNFRSF10B | 771 | 190 | 110 | IL1A - TNFRSF10B | 294 | 1870 | 1471 |
| IL1B - TNFRSF10B | 2301 | 109 | 19 | IL1B - TNFRSF10B | 829 | 626 | 1465 |
| IL1RAP - TNFRSF10B | 752 | 2148 | 1579 | IL1RAP - TNFRSF10B | 2102 | 1685 | 405 |
| IL1RN - TNFRSF10B | 840 | 2005 | 443 | IL1RN - TNFRSF10B | 2087 | 1403 | 1966 |
| IL2RG - TNFRSF10B | 1868 | 1485 | 57 | IL2RG - TNFRSF10B | 1616 | 2134 | 1376 |
| IL6ST - TNFRSF10B | 788 | 1851 | 1038 | IL6ST - TNFRSF10B | 1149 | 510 | 1603 |
| IL8 - TNFRSF10B | 1494 | 1467 | 2312 | IL8 - TNFRSF10B | 1769 | 1763 | 196 |
| IL10RB - TNFRSF10B | 461 | 1770 | 1497 | IL10RB - TNFRSF10B | 1212 | 994 | 1542 |
| IL15 - TNFRSF10B | 360 | 1028 | 620 | IL15 - TNFRSF10B | 1712 | 815 | 2039 |
| IL15RA - TNFRSF10B | 2330 | 932 | 1932 | IL15RA - TNFRSF10B | 1640 | 1375 | 2210 |
| IL17C - TNFRSF10B | 557 | 1911 | 91 | IL17C - TNFRSF10B | 1594 | 969 | 1624 |
| IL17REL - TNFRSF10B | 457 | 1701 | 2422 | IL17REL - TNFRSF10B | 1074 | 2117 | 347 |
| Ranking of IL family w.r.t TNFRSF10D | Ranking of TNFRSF10D w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - TNFRSF10D | 143 | 625 | 21 | IL1A - TNFRSF10D | 2415 | 2517 | 1894 |
| IL1B - TNFRSF10D | 185 | 142 | 191 | IL1B - TNFRSF10D | 2513 | 2300 | 2430 |
| IL1RAP - TNFRSF10D | 1106 | 1750 | 1376 | IL1RAP - TNFRSF10D | 811 | 1241 | 1946 |
| IL1RN - TNFRSF10D | 881 | 520 | 337 | IL1RN - TNFRSF10D | 2512 | 1658 | 857 |
| IL2RG - TNFRSF10D | 713 | 413 | 905 | IL2RG - TNFRSF10D | 2514 | 2419 | 2043 |
| IL6ST - TNFRSF10D | 752 | 2009 | 1617 | IL6ST - TNFRSF10D | 2324 | 2515 | 460 |
| IL8 - TNFRSF10D | 1267 | 903 | 629 | IL8 - TNFRSF10D | 463 | 446 | 2468 |
| IL10RB - TNFRSF10D | 1072 | 1050 | 1031 | IL10RB - TNFRSF10D | 1822 | 1959 | 982 |
| IL15 - TNFRSF10D | 108 | 842 | 333 | IL15 - TNFRSF10D | 2490 | 2234 | 2019 |
| IL15RA - TNFRSF10D | 2197 | 943 | 2126 | IL15RA - TNFRSF10D | 1895 | 1048 | 24 |
| IL17C - TNFRSF10D | 11 | 268 | 7 | IL17C - TNFRSF10D | 2493 | 2062 | 2488 |
| IL17REL - TNFRSF10D | 54 | 638 | 278 | IL17REL - TNFRSF10D | 2514 | 100 | 2452 |
| Ranking of IL family w.r.t TNFRSF12A | Ranking of TNFRSF12A w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - TNFRSF12A | 52 | 2189 | 374 | IL1A - TNFRSF12A | 239 | 2080 | 1330 |
| IL1B - TNFRSF12A | 709 | 1592 | 1066 | IL1B - TNFRSF12A | 1422 | 516 | 1025 |
| IL1RAP - TNFRSF12A | 606 | 1030 | 1639 | IL1RAP - TNFRSF12A | 165 | 1595 | 1273 |
| IL1RN - TNFRSF12A | 122 | 1173 | 1182 | IL1RN - TNFRSF12A | 2176 | 529 | 1135 |
| IL2RG - TNFRSF12A | 206 | 1875 | 756 | IL2RG - TNFRSF12A | 1705 | 1060 | 2416 |
| IL6ST - TNFRSF12A | 2128 | 898 | 1092 | IL6ST - TNFRSF12A | 707 | 2213 | 2187 |
| IL8 - TNFRSF12A | 1132 | 1827 | 2355 | IL8 - TNFRSF12A | 461 | 1199 | 1587 |
| IL10RB - TNFRSF12A | 51 | 37 | 238 | IL10RB - TNFRSF12A | 852 | 781 | 910 |
| IL15 - TNFRSF12A | 281 | 1535 | 686 | IL15 - TNFRSF12A | 1984 | 1469 | 530 |
| IL15RA - TNFRSF12A | 2138 | 2090 | 1981 | IL15RA - TNFRSF12A | 1065 | 576 | 1568 |
| IL17C - TNFRSF12A | 326 | 2512 | 52 | IL17C - TNFRSF12A | 1497 | 1898 | 2209 |
| IL17REL - TNFRSF12A | 2475 | 587 | 2496 | IL17REL - TNFRSF12A | 148 | 1299 | 410 |
| Ranking of IL family w.r.t TNFRSF14 | Ranking of TNFRSF14 w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - TNFRSF14 | 208 | 29 | 683 | IL1A - TNFRSF14 | 2061 | 1969 | 693 |
| IL1B - TNFRSF14 | 70 | 664 | 924 | IL1B - TNFRSF14 | 592 | 1647 | 1743 |
| IL1RAP - TNFRSF14 | 1356 | 2249 | 756 | IL1RAP - TNFRSF14 | 2103 | 1414 | 1691 |
| IL1RN - TNFRSF14 | 1001 | 794 | 745 | IL1RN - TNFRSF14 | 1898 | 2414 | 975 |
| IL2RG - TNFRSF14 | 1619 | 1780 | 1158 | IL2RG - TNFRSF14 | 2009 | 1949 | 1367 |
| IL6ST - TNFRSF14 | 2248 | 221 | 619 | IL6ST - TNFRSF14 | 1033 | 1923 | 2175 |
| IL8 - TNFRSF14 | 517 | 299 | 1301 | IL8 - TNFRSF14 | 1776 | 578 | 2205 |
| IL10RB - TNFRSF14 | 1595 | 156 | 943 | IL10RB - TNFRSF14 | 763 | 1457 | 834 |
| IL15 - TNFRSF14 | 1265 | 550 | 1692 | IL15 - TNFRSF14 | 2039 | 954 | 1230 |
| IL15RA - TNFRSF14 | 2378 | 1929 | 1577 | IL15RA - TNFRSF14 | 2440 | 2031 | 253 |
| IL17C - TNFRSF14 | 11 | 40 | 605 | IL17C - TNFRSF14 | 1856 | 1836 | 671 |
| IL17REL - TNFRSF14 | 46 | 306 | 293 | IL17REL - TNFRSF14 | 2312 | 72 | 1623 |
Table 107.
2nd order combinatorial hypotheses between IL and TNF.
| Ranking Interleukin family vs TNF family | |||||||
| Ranking of IL family w.r.t TNFRSF21 | Ranking of TNFRSF21 w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - TNFRSF21 | 904 | 2313 | 1127 | IL1A - TNFRSF21 | 322 | 1745 | 688 |
| IL1B - TNFRSF21 | 1862 | 2164 | 2305 | IL1B - TNFRSF21 | 1336 | 157 | 829 |
| IL1RAP - TNFRSF21 | 1446 | 1762 | 2163 | IL1RAP - TNFRSF21 | 563 | 22 | 497 |
| IL1RN - TNFRSF21 | 1593 | 2373 | 627 | IL1RN - TNFRSF21 | 1626 | 1341 | 320 |
| IL2RG - TNFRSF21 | 403 | 2297 | 2351 | IL2RG - TNFRSF21 | 618 | 719 | 981 |
| IL6ST - TNFRSF21 | 1372 | 1894 | 753 | IL6ST - TNFRSF21 | 2019 | 1123 | 1143 |
| IL8 - TNFRSF21 | 1204 | 1944 | 1585 | IL8 - TNFRSF21 | 2493 | 999 | 1513 |
| IL10RB - TNFRSF21 | 238 | 845 | 1081 | IL10RB - TNFRSF21 | 2502 | 842 | 1641 |
| IL15 - TNFRSF21 | 1591 | 1905 | 1740 | IL15 - TNFRSF21 | 65 | 1459 | 96 |
| IL15RA - TNFRSF21 | 421 | 1934 | 1269 | IL15RA - TNFRSF21 | 98 | 1109 | 1259 |
| IL17C - TNFRSF21 | 2130 | 1039 | 1676 | IL17C - TNFRSF21 | 2272 | 1163 | 266 |
| IL17REL - TNFRSF21 | 557 | 765 | 61 | IL17REL - TNFRSF21 | 1846 | 704 | 2381 |
| Ranking of IL family w.r.t TNFRS10 | Ranking of TNFRS10 w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - TNFSF10 | 120 | 1575 | 2499 | IL1A - TNFSF10 | 2369 | 1086 | 1034 |
| IL1B - TNFSF10 | 972 | 2448 | 1993 | IL1B - TNFSF10 | 2348 | 1544 | 1076 |
| IL1RAP - TNFSF10 | 754 | 1045 | 2015 | IL1RAP - TNFSF10 | 1613 | 2470 | 966 |
| IL1RN - TNFSF10 | 740 | 1535 | 570 | IL1RN - TNFSF10 | 1035 | 75 | 1074 |
| IL2RG - TNFSF10 | 2272 | 1447 | 1285 | IL2RG - TNFSF10 | 1032 | 882 | 1271 |
| IL6ST - TNFSF10 | 1978 | 227 | 778 | IL6ST - TNFSF10 | 1647 | 1602 | 2369 |
| IL8 - TNFSF10 | 818 | 1702 | 791 | IL8 - TNFSF10 | 1161 | 790 | 2265 |
| IL10RB - TNFSF10 | 744 | 1146 | 2257 | IL10RB - TNFSF10 | 1496 | 2252 | 1864 |
| IL15 - TNFSF10 | 967 | 1382 | 1910 | IL15 - TNFSF10 | 1400 | 1383 | 486 |
| IL15RA - TNFSF10 | 346 | 2163 | 2059 | IL15RA - TNFSF10 | 1458 | 790 | 1428 |
| IL17C - TNFSF10 | 460 | 2337 | 2431 | IL17C - TNFSF10 | 558 | 1004 | 942 |
| IL17REL - TNFSF10 | 1728 | 145 | 989 | IL17REL - TNFSF10 | 1664 | 718 | 250 |
| Ranking of IL family w.r.t TNFRS15 | Ranking of TNFRS15 w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - TNFSF15 | 1177 | 2494 | 979 | IL1A - TNFSF15 | 1014 | 613 | 1449 |
| IL1B - TNFSF15 | 1435 | 1529 | 1571 | IL1B - TNFSF15 | 1898 | 1032 | 767 |
| IL1RAP - TNFSF15 | 271 | 1665 | 2368 | IL1RAP - TNFSF15 | 890 | 843 | 793 |
| IL1RN - TNFSF15 | 2319 | 377 | 566 | IL1RN - TNFSF15 | 414 | 1457 | 1704 |
| IL2RG - TNFSF15 | 316 | 874 | 487 | IL2RG - TNFSF15 | 2332 | 1362 | 1632 |
| IL6ST - TNFSF15 | 1834 | 1004 | 1471 | IL6ST - TNFSF15 | 771 | 1171 | 1445 |
| IL8 - TNFSF15 | 1266 | 1571 | 1141 | IL8 - TNFSF15 | 2422 | 515 | 966 |
| IL10RB - TNFSF15 | 1488 | 326 | 1367 | IL10RB - TNFSF15 | 1611 | 2041 | 1635 |
| IL15 - TNFSF15 | 1356 | 1508 | 737 | IL15 - TNFSF15 | 201 | 1922 | 1756 |
| IL15RA - TNFSF15 | 2124 | 956 | 2365 | IL15RA - TNFSF15 | 1551 | 668 | 864 |
| IL17C - TNFSF15 | 2222 | 2328 | 954 | IL17C - TNFSF15 | 2403 | 1049 | 1338 |
| IL17REL - TNFSF15 | 1214 | 177 | 208 | IL17REL - TNFSF15 | 513 | 1515 | 1943 |
On the right side, we found TNF was up regulated w.r.t IL-6ST/10RB. These are reflected in the rankings of 2410 (laplace) and 1901 (linear) for IL6ST - TNF; and 2065 (laplace), 2120 (linear) and 2296 (rbf) for IL10RB - TNF; TNFAIP1 was up regulated w.r.t IL-8/15RA. These are reflected in the rankings of 2293 (laplace) and 2126 (linear) for IL8 - TNFAIP1; and 2141 (linear) and 1853 (rbf) for IL15RA - TNFAIP1; TNFRSF1A was up regulated w.r.t IL-1B. This is reflected in the rankings of 2027 (linear) and 2247 (rbf) for IL1B - TNFRSF1A; TNFRSF10A was up regulated w.r.t IL-1A/1B/1RN/2RG/6ST/15/15RA/17C. These are reflected in the rankings of 1972 (laplace), 1805 (linear) and 2504 (rbf) for IL1A - TNFRSF10A; 2375 (laplace), 2373 (linear) and 2320 (rbf) for IL1B - TNFRSF10A; 2287 (linear) and 2469 (rbf) for IL1RN - TNFRSF10A; 2246 (linear) and 2467 (rbf) for IL2RG - TNFRSF10A; 2128 (laplace) and 2320 (linear) for IL6ST - TNFRSF10A; 2414 (laplace) and 2260 (linear) for IL15 - TNFRSF10A; 2398 (laplace) and 1970 (linear) and 2088 (rbf) for IL15RA - TNFRSF10A; and 1831 (laplace) and 2025 (linear) for IL17C - TNFRSF10A; TNFRSF10B was up regulated w.r.t IL-1RN. This is reflected in the rankings of 2087 (laplace) and 1966 (rbf) for IL1RN - TNFRSF10B; TNFRSF10D was up regulated w.r.t IL-1A/1B/2RG/6ST/10RB/15/17C/17REL. These are reflected in the rankings of 2415 (laplace), 2517 (linear) and 1894 (rbf) for IL1A - TNFRSF10D; 2513 (laplace), 2300 (linear) and 2430 (rbf) for IL1B - TNFRSF10D; 2514 (laplace), 2419 (linear) and 2043 (rbf) for IL2RG - TNFRSF10D; 2324 (laplace), 2515 (linear) for IL6ST - TNFRSF10D; 1822 (laplace), 1959 (linear) for IL10RB - TNFRSF10D; 2490 (laplace), 2234 (linear) and 2019(rbf) for IL15 - TNFRSF10D; 2493 (laplace), 2062 (linear) and 2488 (rbf) for IL17C - TNFRSF10D; and 2514 (laplace) and 2452 (rbf) for IL17REL - TNFRSF10D. TNFRSF12A was up regulated w.r.t IL-16ST/17C. These are reflected in the rankings of 2213 (linear) and 2187 (rbf) for IL6ST - TNFRSF12A; and 1898 (linear) and 2209 (rbf) for IL17C - TNFRSF12A; TNFRSF14 was up regulated w.r.t IL-1A/1RN/2RG/6ST/8/15RA/17C. These are reflected in the rankings of 2061 (laplace) and 1969 (linear) for IL1A - TNFRSF14; 1898 (laplace) and 2414 (linear) for IL1RN - TNFRSF14; 2009 (laplace) and 1949 (linear) for IL2RG - TNFRSF14; 1923 (linear) and 2175 (rbf) for IL6ST - TNFRSF14; 1776 (laplace) and 2205 (rbf) for IL8 - TNFRSF14; 2440 (laplace) and 2031 (linear) for IL15RA - TNFRSF14; and 1856 (laplace) and 1836 (linear) for IL17C - TNFRSF14. TNFRSF21 was up regulated w.r.t IL-17REL. This is reflected in the rankings of 1846 (laplace) and 2381 (rbf) for IL17REL - TNFRSF21; TNFRSF10 was up regulated w.r.t IL-10RB. This is reflected in the rankings 2252 (linear) and 1864 (rbf) of IL10RB - TNFSF10; TNFRSF15 was up regulated w.r.t IL-15. This is reflected in the rankings of 1922 (linear) and 1756 (rbf) for IL15 - TNFSF15.
Finally, Table 108 shows the derived influences which can be represented graphically, with the following influences - • IL w.r.t TNF with IL-1RAP/6ST/15RA <- TNF; IL-1B/2RG/15RA/17C <- TNFAIP1; IL-1RN/10RB <- TNFAIP2; IL-6ST/8/17REL <- TNFAIP3; IL-1RAP <- TNFRSF1A; IL-1RAP/15RA/17REL <- TNFRSF10A; IL-15RA <- TNFRSF10B; IL-15RA <- TNFRSF10D; IL-8/15RA/17REL <- TNFRSF12A; IL-15RA <- TNFRSF14; IL-1B/1RAP/2RG <- TNFRSF21; IL-1B/15RA/17C <- TNFSF10 and IL-17C <- TNFSF15; and • TNF w.r.t IL with IL-6ST/10RB -> TNF; IL-8/15RA -> TNFAIP1; IL-1B -> TNFRSF1A; IL-1A/1B/1RN/2RG/6ST/15/15RA/17C -> TNFRSF10A; IL-1RN -> TNFRSF10B; IL-1A/1B/2RG/6ST/10RB/15/17C/17REL -> TNFRSF10D; IL-6ST/17C -> TNFRSF12A; IL-1A/1RN/2RG/6ST/8/15RA/17C/17REL -> TNFRSF14; IL-17REL -> TNFRSF14; IL10RB -> TNFSF10; and IL15 -> TNFSF15;
3.8. BCL Related Synergies
3.8.1. Interleukin - BCL Cross Family Analysis
Qin et al. [210] observe that IL-6 inhibits starvation-induced autophagy via the STAT3/Bcl-2 signaling pathway. Gabellini et al. [211] observed that interleukin 8 mediates bcl-xL-induced enhancement of human melanoma cell dissemination and angiogenesis in a zebrafish xenograft model. Guruprasath et al. [212] show taht interleukin-4 receptor-targeted delivery of Bcl-xL siRNA sensitizes tumors to chemotherapy and inhibits tumor growth. Maraskovsky et al. [213] indicate that Bcl-2 can rescue T lymphocyte development in interleukin-7 receptor-deficient mice but not in mutant rag-1−/− mice. Akashi et al. [214] show that Bcl-2 rescues T lymphopoiesis in interleukin-7 receptor-deficient mice. Interleukin-10 increases Bcl-2 expression and survival in primary human CD34+ hematopoietic progenitor cells as shown by Weber-Nordt et al. [215]. Interleukin-7 and interleukin-15 regulate the expression of thebcl-2 and c-myb genes in cutaneous T-cell lymphoma cells as shown by Qin et al. [216]. Bcl-2 is a negative regulator of interleukin-1β secretion in murine macrophages in pharmacological-induced apoptosis as shown by Escandell et al. [217]. Alas et al. [218] observe that inhibition of interleukin 10 by rituximab results in down-regulation of bcl-2 and sensitization of B-cell non-Hodgkin’s lymphoma to apoptosis. These findings indicate the synergy between BCL and Interleukin in different pathological cases. In CRC cells treated with ETC-1922159, these were found to be up regulated. Table 109 and Table 110 indicate the rankings of the IL and BCL family.
On the left side is the rankings of IL w.r.t BCL family and the right side, the vice versa. We found IL-1A/1B/17C up regulated w.r.t BCL2L1. These are reflected in rankings of 2482 (laplace) and 1834 (rbf) for IL1A - BCL2L1; 2252 (laplace), 1920 (linear) for IL1B - BCL2L1; and 2481 (laplace), 2410 (linear) and 2512 (rbf) for IL17C - BCL2L1; IL-6ST/17REL were up regulated w.r.t BCL2L2. These are reflected in rankings of 2239 (laplace), 1927 (linear) and 2085 (rbf) for IL6ST - BCL2L2; and 2454 (laplace), 2510 (linear) and 2482 (rbf) for IL17REL - BCL2L2. IL-17REL were up regulated w.r.t BCL2L13. These are reflected in rankings of 2420 (laplace), 2419 (linear) and 2464 (rbf) for IL17REL - BCL2L13; IL-6ST/15RA were up regulated w.r.t BCL3. These are reflected in rankings of 1928 (laplace) and 2344 (rbf) for IL6ST - BCL3; and 2478 (laplace), 1820 (linear) and 2500 (rbf) for IL15RA - BCL3; IL-1RAP/6ST/8/17REL were up regulated w.r.t BCL6. These are reflected in rankings of 2360 (linear) and 1813 (rbf) for IL1RAP - BCL6; 2419 (laplace) and 1962 (rbf) for IL6ST - BCL6; 2363 (laplace) and 2233 (linear) for IL8 - BCL6; and 2253 (laplace) and 2396 (linear) for IL17REL - BCL6; IL-1A/6ST/8/17REL were up regulated w.r.t BCL9L. These are reflected in rankings of 1932 (laplace) and 1942 (linear) for IL1A - BCL9L; 2249 (laplace) and 1960 (linear) for IL6ST - BCL9L; 2197 (linear) and 2162 (rbf) for IL8 - BCL9L; and 2308 (linear) and 1926 (rbf) for IL17REL - BCL9L; IL-6ST/15RA were up regulated w.r.t BCL10. These are reflected in rankings of 2008 (laplace) and 1816 (rbf) for IL6ST - BCL10; and 2064 (linear) and 1789 (rbf) for IL15RA - BCL10;
On the right side is the rankings of BCL w.r.t IL family. We found BCL2L1 up regulated IL-1B/2RG/10RB. These are reflected in rankings of 1838 (laplace) and 2132 (rbf) for IL1B - BCL2L1; 2048 (laplace) and 1949 (rbf) for IL2RG - BCL2L1; and 1965 (linear) and 2024 (rbf) for IL10RB - BCL2L1; BCL2L2 was up regulated IL-1A/1B/1RN/6ST/8/15/17C. These are reflected in rankings of 2407 (laplace), 2362 (linear) and 2464 (rbf) for IL1A - BCL2L2; 1807 (laplace), 2462 (linear) and 2344 (rbf) for IL1B - BCL2L2; 2298 (laplace) and 2092 (rbf) for IL1RN - BCL2L2; 2046 (linear) and 1859 (rbf) for IL6ST - BCL2L2; 1803 (laplace) and 2024 (rbf) for IL8 - BCL2L2; 2474 (laplace), 2142 (linear) and 2416 (rbf) for IL15 - BCL2L2; and 2512 (linear) and 2447 (rbf) for IL17C - BCL2L2; BCL2L13 was up regulated IL-1RAP/1RN/2RG/6ST/8/10RB/15/15RA/17C. These are reflected in rankings of 2450 (linear) and 2510 (rbf) for IL1RAP - BCL2L13; 2503 (laplace) and 2378 (rbf) for IL1RN - BCL2L13; 2483 (laplace) and 2248 (rbf) for IL2RG - BCL2L13; 1899 (laplace), 2473 (linear) and 2046 (rbf) for IL6ST - BCL2L13; 2099 (laplace) and 2294 (rbf) for IL8 - BCL2L13; 2120 (laplace) and 1895 (linear) for IL10RB - BCL2L13; 2515 (laplace), 2160 (linear) and 2420 (rbf) for IL15 - BCL2L13; 1844 (linear) and 2318 (rbf) for IL15RA - BCL2L13; and 2004 (laplace), 2434 (linear) and 2500 (rbf) for IL17C - BCL2L13; BCL3 was up regulated IL-8/10RB. These are reflected in rankings of 2266 (laplace) and 1983 (rbf) for IL8 - BCL3; and 2187 (laplace) and 2170 (rbf) for IL10RB - BCL3; 2298 (laplace); 2423 (linear) and 2294 (rbf) for IL1B - BCL6; 1919 (laplace) and 2301 (linear) for IL1RN - BCL6; 2106 (linear) and 2478 (rbf) for IL2RG - BCL6; 2123 (laplace), 2068 (linear) for IL8 - BCL6; 2084 (laplace), 1791 (linear) and 2203 (rbf) for IL15RA - BCL6; and for 1949 (linear) and 1930 (rbf) for IL17REL - BCL6; BCL10 was up regulated IL-1A/1RAP/1RN/2RG/10RB/15RA. These are reflected in rankings of 2405 (linear) and 1889 (rbf) for IL1A - BCL10; 1929 (laplace) and 2112 (rbf) for IL1RAP - BCL10; 1846 (laplace) and 1823 (linear) for IL1RN - BCL10; 1885 (laplace) and 1803 (linear) for IL2RG - BCL10; 2244 (laplace) and 2150 (linear) for IL10RB - BCL10; and 1810 (laplace) and 1835 (rbf) for IL15RA - BCL10;
Finally, Table 111 shows the derived influences which can be represented graphically, with the following influences - • IL w.r.t BCL with IL-1A/1B/17C <- BCL2L1; IL-6ST/17REL <- BCL2L2; IL-17REL <- BCL2L13; IL-6ST/15RA <- BCL3; IL-1RAP/6ST/8/17REL <- BCL6; IL-1A/6ST/8/17REL <- BCL9L; and IL-6ST/15RA <- BCL10; • BCL w.r.t IL with IL-1B/2RG/10RB -> BCL2L1; IL-1A/1B/1RN/6ST/8/15/17C -> BCL2L2; IL-1RAP/1RN/2RG/6ST/8/10RB/15/15RA/17C -> BCL2L13; IL-8/10RB -> BCL3; IL-1B/1RN/2RG/8/15RA/17REL -> BCL6; and IL-1A/1RAP/1RN/2RG/10RB/15RA -> BCL10;
3.8.2. Selenbp1 - BCL Cross Family Analysis
Deng et al. [219] study the effects of selenium on lead-induced alterations in Aβ production and Bcl-2 family proteins. Yaming et al. [220] studied the effects of selenium dioxide on apoptosis, Bcl-2 and p53 expression, intracellular reactive oxygen species and calcium level in three human lung cancer cell lines. Activity of selenium on cell proliferation, cytotoxicity, and apoptosis and on the expression of CASP9, BCL-XL and APC in intestinal adenocarcinoma cells has been studied by Mauro et al. [221]. These studies suggest the synergy between BCL and Selenium based genes. In CRC cells treated with ETC-1922159, these were found to be down regulated. Table 112 shows the rankings of BCL family w.r.t to SELENBP1 and vice versa.
On the right side, we found BCL-6B/11A to be up regulated with respect to SELENBP1. These were reflected in the rankings of 182 (laplace), 110 (linear) and 494 (rbf) for SELENBP1 - BCL6B; and 905 (laplace), 931 (linear) and 401 (rbf) for SELENBP1 - BCL11A. On the left side SELENBP1 was up regulated w.r.t BCL-9/11B. These are reflected in rankings of 1568 (linear) and 1738 (rbf) for SELENBP1 - BCL9; and 299 (linear) and 1385 (rbf) for SELENBP1 - BCL11B; Finally, Table 113 shows the derived influences which can be represented graphically, with the following influences - • SELENBP1 w.r.t BCL with SELENBP1 <- BCL-9/11B; and • BCL w.r.t SELENBP1 with SELENBP1 -> BCL-6B/11A;
Table 112.
2nd order combinatorial hypotheses between BCL and SELENBP1.
| Ranking SELENBP1 vs BCL family | |||||||
| Ranking of BCL family w.r.t SELENBP1 | Ranking of SELENBP1 w.r.t BCL | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| SELENBP1 - BCL2L12 | 2426 | 2033 | 2629 | SELENBP1 - BCL2L12 | 2589 | 2195 | 2082 |
| SELENBP1 - BCL6B | 2446 | 2575 | 1956 | SELENBP1 - BCL6B | 182 | 110 | 494 |
| SELENBP1 - BCL7A | 2620 | 1326 | 2006 | SELENBP1 - BCL7A | 2015 | 1799 | 767 |
| SELENBP1 - BCL9 | 2453 | 1568 | 1738 | SELENBP1 - BCL9 | 2538 | 1916 | 1793 |
| SELENBP1 - BCL11A | 1921 | 2463 | 1566 | SELENBP1 - BCL11A | 905 | 931 | 401 |
| SELENBP1 - BCL11B | 1896 | 299 | 1385 | SELENBP1 - BCL11B | 2496 | 2636 | 2510 |
Table 113.
2nd order combinatorial hypotheses between SELENBP1 and BCL family.
| Unexplored combinatorial hypotheses | |
| SELENBP1 w.r.t BCL | |
| SELENBP1 | BCL-9/11B |
| BCL w.r.t SELENBP1 | |
| SELENBP1 | BCL-6B/11A |
3.8.3. TP53 - BCL Cross Family Analysis
The p53-Bcl-2 connection has been studied by Hemann and Lowe [222]. Tomita et al. [223] show wild type p53, but not tumor-derived mutants, bind to Bcl2 via the DNA binding domain and induce mitochondrial permeabilization. Bcl-2 constitutively suppresses p53-dependent apoptosis in colorectal cancer cells as shown by Jiang and Milner [224]. The tissue dependent interactions between p53 and Bcl-2 in vivo has been studied by Li et al. [225]. Synthetic lethality of combined Bcl-2 inhibition and p53 activation in AML has been studied by Pan et al. [226]. Zaidi et al. [227] observe that the chloroquine-induced neuronal cell death is p53 and Bcl-2 family-dependent but caspase-independent. Relationship of p53, bcl-2, and tumor proliferation to clinical drug resistance in non-Hodgkin’s lymphomas has been studied in Wilson et al. [228]. TP53 and BCL family members were found to be up regulated in CRC cells treated with ETC-1922159. Table 114 show rankings of BCL and TP53 family w.r.t to each other.
On the left side, we found BCL2L2 to be up regulated w.r.t TP53-I3/INP2. These are reflected in the rankings of 2423 (laplace), 2377 (linear) and 2452 (rbf) for TP53I3 - BCL2L2; 1827 (linear) and 2035 (rbf) for TP53INP2 - BCL2L2. BCL2L13 to be up regulated w.r.t TP53-INP2. These are reflected in the rankings of 2427 (linear) and 2008 (rbf) for TP53INP2 - BCL2L13; BCL6 to be up regulated w.r.t TP53-I3/INP2. These are reflected in the rankings of 2275 (laplace), 2312 (linear) and 2146 (rbf) for TP53I3 - BCL6; and 2329 (linear) and 2352 (rbf) for TP53INP2 - BCL6; BCL9L to be up regulated w.r.t TP53-BP2. These are reflected in the rankings of 2320 (linear) and 2197 (rbf) for TP53BP2 - BCL9L; BCL10 to be up regulated w.r.t TP53-BP2/INP2. These are reflected in the rankings of 2230 (laplace) and 2418 (linear) for TP53BP2 - BCL10 and 1910 (linear) and 2087 (rbf) for TP53INP2 - BCL10;
On the right side, we found TP53-BP2/I3 to be up regulated w.r.t BCL2L1. These are reflected in the rankings of 1786 (laplace) and 1961 (linear) for TP53BP2 - BCL2L1; 1980 (laplace) and 1752 (linear) for TP53I3 - BCL2L1; TP53-INP1 were up regulated w.r.t BCL3. These are reflected in the rankings for 2259 (linear) and 2043 (rbf) for TP53INP1 - BCL3; TP53-BP2/INP2 were up regulated w.r.t BCL9L. These are reflected in the rankings for 2093 (laplace) and 2217 (linear) for TP53BP2 - BCL9L; and 2222 (laplace) and 1900 (linear) for TP53INP2 - BCL9L;
Finally, Table 115 shows the derived influences which can be represented graphically, with the following influences - • BCL w.r.t TP53 with TP53-I3/INP2 <- BCL2L2; TP53-INP2 <- BCL2L13; TP53-I3/INP2 <- BCL6; TP53-BP2 <- BCL9L; and TP53-BP2/INP2 <- BCL10; • TP53 w.r.t BCL with TP53-BP2/I3 <- BCL2L1; TP53-INP1 <- BCL3 and TP53-BP2/INP2 <- BCL9L.
Table 114.
2nd order combinatorial hypotheses between BCL and SELENBP1.
| Ranking TP53 family vs BCL family | |||||||
| Ranking of BCL2L1 w.r.t TP53 family | Ranking of TP53 family w.r.t BCL2L1 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| TP53BP2 - BCL2L1 | 2431 | 1529 | 1728 | TP53BP2 - BCL2L1 | 1786 | 1961 | 1225 |
| TP53I3 - BCL2L1 | 799 | 554 | 728 | TP53I3 - BCL2L1 | 1980 | 1752 | 756 |
| TP53INP1 - BCL2L1 | 1064 | 1154 | 1414 | TP53INP1 - BCL2L1 | 1193 | 258 | 1850 |
| TP53INP2 - BCL2L1 | 282 | 2371 | 851 | TP53INP2 - BCL2L1 | 830 | 1477 | 1512 |
| Ranking of BCL2L2 w.r.t TP53 family | Ranking of TP53 family w.r.t BCL2L2 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| TP53BP2 - BCL2L2 | 1471 | 34 | 1367 | TP53BP2 - BCL2L2 | 1076 | 2168 | 1658 |
| TP53I3 - BCL2L2 | 2423 | 2377 | 2452 | TP53I3 - BCL2L2 | 1911 | 245 | 378 |
| TP53INP1 - BCL2L2 | 1693 | 180 | 987 | TP53INP1 - BCL2L2 | 482 | 1653 | 1130 |
| TP53INP2 - BCL2L2 | 1688 | 1827 | 2035 | TP53INP2 - BCL2L2 | 85 | 376 | 1146 |
| Ranking of BCL2L13 w.r.t TP53 family | Ranking of TP53 family w.r.t BCL2L13 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| TP53BP2 - BCL2L13 | 1515 | 1261 | 1842 | TP53BP2 - BCL2L13 | 1128 | 1827 | 1613 |
| TP53I3 - BCL2L13 | 1264 | 1501 | 1963 | TP53I3 - BCL2L13 | 419 | 1088 | 959 |
| TP53INP1 - BCL2L13 | 759 | 387 | 205 | TP53INP1 - BCL2L13 | 1550 | 1616 | 1245 |
| TP53INP2 - BCL2L13 | 507 | 2427 | 2008 | TP53INP2 - BCL2L13 | 1190 | 573 | 513 |
| Ranking of BCL3 w.r.t TP53 family | Ranking of TP53 family w.r.t BCL3 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| TP53BP2 - BCL3 | 1754 | 335 | 226 | TP53BP2 - BCL3 | 1177 | 1625 | 423 |
| TP53I3 - BCL3 | 388 | 392 | 25 | TP53I3 - BCL3 | 921 | 1151 | 233 |
| TP53INP1 - BCL3 | 2350 | 766 | 472 | TP53INP1 - BCL3 | 1126 | 2259 | 2043 |
| TP53INP2 - BCL3 | 266 | 1184 | 379 | TP53INP2 - BCL3 | 325 | 609 | 726 |
| Ranking of BCL6 w.r.t TP53 family | Ranking of TP53 family w.r.t BCL6 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| TP53BP2 - BCL6 | 1172 | 1783 | 1120 | TP53BP2 - BCL6 | 1667 | 1140 | 185 |
| TP53I3 - BCL6 | 2275 | 2312 | 2146 | TP53I3 - BCL6 | 979 | 71 | 859 |
| TP53INP1 - BCL6 | 201 | 1818 | 1572 | TP53INP1 - BCL6 | 1458 | 1200 | 2503 |
| TP53INP2 - BCL6 | 1681 | 2329 | 2352 | TP53INP2 - BCL6 | 346 | 833 | 1557 |
| Ranking of BCL9L w.r.t TP53 family | Ranking of TP53 family w.r.t BCL9L | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| TP53BP2 - BCL9L | 263 | 2320 | 2197 | TP53BP2 - BCL9L | 2093 | 2217 | 1010 |
| TP53I3 - BCL9L | 819 | 635 | 789 | TP53I3 - BCL9L | 1249 | 927 | 107 |
| TP53INP1 - BCL9L | 2090 | 1740 | 1179 | TP53INP1 - BCL9L | 2113 | 854 | 1711 |
| TP53INP2 - BCL9L | 640 | 951 | 316 | TP53INP2 - BCL9L | 2222 | 1900 | 151 |
| Ranking of BCL10 w.r.t TP53 family | Ranking of TP53 family w.r.t BCL10 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| TP53BP2 - BCL10 | 2230 | 2418 | 73 | TP53BP2 - BCL10 | 493 | 1999 | 351 |
| TP53I3 - BCL10 | 727 | 1159 | 1301 | TP53I3 - BCL10 | 519 | 1572 | 446 |
| TP53INP1 - BCL10 | 543 | 1223 | 1275 | TP53INP1 - BCL10 | 1094 | 1120 | 1848 |
| TP53INP2 - BCL10 | 632 | 1910 | 2087 | TP53INP2 - BCL10 | 789 | 1566 | 848 |
Table 115.
2nd order combinatorial hypotheses between SELENBP1 and BCL family.
| Unexplored combinatorial hypotheses | |
| BCL w.r.t TP53 | |
| TP53-I3/INP2 | BCL2L2 |
| TP53-INP2 | BCL2L13 |
| TP53-I3/INP2 | BCL6 |
| TP53-BP2 | BCL9L |
| TP53-BP2/INP2 | BCL10 |
| TP53 w.r.t BCL | |
| TP53-BP2/I3 | BCL2L1 |
| TP53-INP1 | BCL3 |
| TP53-BP2/INP2 | BCL9L |
3.8.4. CASP - BCL Cross Family Analysis
Expression of caspase and BCL-2 apoptotic family members in mouse preimplantation embryos have been studied by Exley et al. [229]. Swanton et al. [230] observed that Bcl-2 regulates a caspase-3/caspase-2 apoptotic cascade in cytosolic extracts. Their role in the regulation of the immune response of Caspases, Bcl-2 family proteins and other components of the death machinery has been observed in Pellegrini and Strasser [231]. Moriishi et al. [232] show that Bcl-2 family members do not inhibit apoptosis by binding the caspase activator Apaf-1. In CRC cells treated with ETC-1922159, these families were found to be UP regulated. Table 116 shows rankings of CASP and BCL family.
On the left side, we found BCL2L2 to be up regulated w.r.t CASP-10/16. These are reflected in the rankings of 2043 (linear) and 1809 (rbf) for CASP10 - BCL2L2; and 2263 (laplace) and 1863 (rbf) for CASP16 - BCL2L2; BCL2L13 to be up regulated w.r.t CASP-4/5/16. These are reflected in the rankings of 1873 (laplace) and 2415 (rbf) for CASP4 - BCL2L13; 1962 (laplace), 2514 (linear) and 2493 (rbf) for CASP5 - BCL2L13; and 1762 (laplace), 2492 (linear) and 2166 (rbf) for CASP16 - BCL2L13; BCL3 to be up regulated w.r.t CASP-10. These are reflected in the rankings of 2409 (laplace) and 2011 (linear) for CASP10 - BCL3; BCL6 to be up regulated w.r.t CASP-5/16. These are reflected in the rankings of 1787 (laplace), 2124 (linear) and 2309 (rbf) for CASP5 - BCL6; and 2397 (laplace), 2166 (linear) and 2387 (rbf) for CASP16 - BCL6.
On the right side, we found CASP-5/7 to be up regulated w.r.t BCL2L1. These are reflected in the rankings of 1992 (laplace) and 2053 (linear) for CASP5 - BCL2L1; and 2203 (linear) and 1750 (rbf) for CASP7 - BCL2L1. CASP-4/7 to be up regulated w.r.t BCL2L1. These are reflected in the rankings of 1902 (linear) and 1979 (rbf) for CASP4 - BCL2L13 and 1877 (laplace) and 2216 (rbf) for CASP7 - BCL2L13; CASP-7/16 to be up regulated w.r.t BCL9L. These are reflected in the rankings of 1813 (laplace) and 1980 (rbf) for CASP7 - BCL9L; and 2499 (linear) and 2027 (rbf) for CASP16 - BCL9L; CASP-7 to be up regulated w.r.t BCL10. These are reflected in the rankings of 2489 (laplace) and 1945 (rbf) for CASP7 - BCL10.
Table 116.
2nd order combinatorial hypotheses between BCL and SELENBP1.
| Ranking CASP family vs BCL family | |||||||
| Ranking of BCL2L1 w.r.t CASP family | Ranking of CASP family w.r.t BCL2L1 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| CASP4 - BCL2L1 | 170 | 1441 | 1555 | CASP4 - BCL2L1 | 355 | 1603 | 202 |
| CASP5 - BCL2L1 | 1236 | 766 | 1261 | CASP5 - BCL2L1 | 1992 | 2053 | 291 |
| CASP7 - BCL2L1 | 2235 | 1161 | 1252 | CASP7 - BCL2L1 | 657 | 2203 | 1750 |
| CASP9 - BCL2L1 | 291 | 984 | 692 | CASP9 - BCL2L1 | 833 | 1386 | 1855 |
| CASP10 - BCL2L1 | 1162 | 2043 | 218 | CASP10 - BCL2L1 | 721 | 2088 | 101 |
| CASP16 - BCL2L1 | 239 | 34 | 305 | CASP16 - BCL2L1 | 43 | 489 | 351 |
| Ranking of BCL2L2 w.r.t CASP family | Ranking of CASP family w.r.t BCL2L2 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| CASP4 - BCL2L2 | 1144 | 1441 | 2348 | CASP4 - BCL2L2 | 988 | 966 | 1400 |
| CASP5 - BCL2L2 | 1896 | 766 | 914 | CASP5 - BCL2L2 | 401 | 174 | 1136 |
| CASP7 - BCL2L2 | 895 | 1161 | 1604 | CASP7 - BCL2L2 | 2371 | 1352 | 1312 |
| CASP9 - BCL2L2 | 1414 | 984 | 1933 | CASP9 - BCL2L2 | 863 | 720 | 102 |
| CASP10 - BCL2L2 | 1335 | 2043 | 1809 | CASP10 - BCL2L2 | 1630 | 1912 | 884 |
| CASP16 - BCL2L2 | 2263 | 34 | 1863 | CASP16 - BCL2L2 | 2 | 151 | 114 |
| Ranking of BCL2L13 w.r.t CASP family | Ranking of CASP family w.r.t BCL2L13 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| CASP4 - BCL2L13 | 1873 | 1096 | 2415 | CASP4 - BCL2L13 | 1257 | 1902 | 1979 |
| CASP5 - BCL2L13 | 1962 | 2514 | 2493 | CASP5 - BCL2L13 | 1438 | 1376 | 664 |
| CASP7 - BCL2L13 | 601 | 1195 | 756 | CASP7 - BCL2L13 | 1877 | 1646 | 2216 |
| CASP9 - BCL2L13 | 1592 | 2371 | 1376 | CASP9 - BCL2L13 | 447 | 1618 | 844 |
| CASP10 - BCL2L13 | 489 | 384 | 987 | CASP10 - BCL2L13 | 1403 | 1048 | 354 |
| CASP16 - BCL2L13 | 1762 | 2492 | 2166 | CASP16 - BCL2L13 | 1927 | 376 | 510 |
| Ranking of BCL3 w.r.t CASP family | Ranking of CASP family w.r.t BCL3 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| CASP4 - BCL3 | 18 | 844 | 1229 | CASP4 - BCL3 | 335 | 172 | 1629 |
| CASP5 - BCL3 | 728 | 953 | 1616 | CASP5 - BCL3 | 343 | 498 | 628 |
| CASP7 - BCL3 | 737 | 574 | 580 | CASP7 - BCL3 | 1313 | 1804 | 1556 |
| CASP9 - BCL3 | 1478 | 284 | 242 | CASP9 - BCL3 | 2392 | 1123 | 1394 |
| CASP10 - BCL3 | 2409 | 2011 | 1425 | CASP10 - BCL3 | 156 | 838 | 1678 |
| CASP16 - BCL3 | 868 | 103 | 715 | CASP16 - BCL3 | 361 | 162 | 2505 |
| Ranking of BCL6 w.r.t CASP family | Ranking of CASP family w.r.t BCL6 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| CASP4 - BCL6 | 1311 | 2266 | 1297 | CASP4 - BCL6 | 27 | 507 | 944 |
| CASP5 - BCL6 | 1787 | 2124 | 2309 | CASP5 - BCL6 | 760 | 10 | 770 |
| CASP7 - BCL6 | 996 | 1314 | 2322 | CASP7 - BCL6 | 1478 | 1230 | 2366 |
| CASP9 - BCL6 | 1022 | 824 | 2021 | CASP9 - BCL6 | 1855 | 903 | 1296 |
| CASP10 - BCL6 | 469 | 1559 | 1085 | CASP10 - BCL6 | 591 | 787 | 1410 |
| CASP16 - BCL6 | 2397 | 2166 | 2387 | CASP16 - BCL6 | 1514 | 54 | 1881 |
| Ranking of BCL9L w.r.t CASP family | Ranking of CASP family w.r.t BCL9L | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| CASP4 - BCL9L | 578 | 897 | 325 | CASP4 - BCL9L | 1758 | 1346 | 1584 |
| CASP5 - BCL9L | 1075 | 791 | 1134 | CASP5 - BCL9L | 363 | 1731 | 632 |
| CASP7 - BCL9L | 2279 | 1347 | 632 | CASP7 - BCL9L | 1813 | 853 | 1980 |
| CASP9 - BCL9L | 98 | 1126 | 455 | CASP9 - BCL9L | 1472 | 717 | 940 |
| CASP10 - BCL9L | 24 | 841 | 2358 | CASP10 - BCL9L | 675 | 1449 | 699 |
| CASP16 - BCL9L | 591 | 666 | 233 | CASP16 - BCL9L | 12 | 2499 | 2027 |
| Ranking of BCL10 w.r.t CASP family | Ranking of CASP family w.r.t BCL10 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| CASP4 - BCL10 | 1272 | 1457 | 619 | CASP4 - BCL10 | 244 | 1637 | 426 |
| CASP5 - BCL10 | 1732 | 1092 | 1293 | CASP5 - BCL10 | 667 | 2488 | 522 |
| CASP7 - BCL10 | 1448 | 1028 | 681 | CASP7 - BCL10 | 2489 | 1516 | 1945 |
| CASP9 - BCL10 | 612 | 553 | 205 | CASP9 - BCL10 | 1644 | 1117 | 956 |
| CASP10 - BCL10 | 2289 | 1694 | 1401 | CASP10 - BCL10 | 664 | 917 | 84 |
| CASP16 - BCL10 | 27 | 102 | 301 | CASP16 - BCL10 | 2192 | 3 | 387 |
Table 117.
2nd order combinatorial hypotheses between CASP and BCL family.
| Unexplored combinatorial hypotheses | |
| BCL w.r.t CASP | |
| CASP-10/16 | BCL2L2 |
| CASP-4/5/16 | BCL2L13 |
| CASP-10 | BCL3 |
| CASP-5/16 | BCL6 |
| CASP w.r.t BCL | |
| CASP-5/7 | BCL2L1 |
| CASP-4/7 | BCL2L13 |
| CASP-7/16 | BCL9L |
| CASP-7 | BCL10 |
3.8.5. MUC - BCL Cross Family Analysis
MUC1 and bcl-2 expression in preinvasive lesions and adenosquamous carcinoma of the lung have been studied by Demirag et al. [233]. Sheng et al. [234] report that MUC13 prevents colorectal cancer cell death by promoting two distinct pathways of NF-kB activation, consequently upregulating BCL-XL. In CRC cells treated with ETC-1922159, family members of BCL and MUC were found up regulated. The search engine assigned high valued numerical ranks to some of the 2nd order combinations of BCL-MUC family members. Table 118 show the rankings of the members with respect to each other.
On the left side, we found BCL2L1 to be up regulated w.r.t MUC-1/13. These are reflected in the rankings of 2055 (laplace), 2297 (linear) and 1854 (rbf) for MUC1 - BCL2L1; and 1927 (laplace) and 2108 (rbf) for MUC13 - BCL2L1; BCL2L2 was up regulated w.r.t MUC-4/13/17. These are reflected in the rankings of 2506 (linear) and 1988 (rbf) for MUC4 - BCL2L2; 2084 (laplace) and 2402 (linear) for MUC13 - BCL2L2; and 2283 (laplace) and 2212 (linear) for MUC17 - BCL2L2; BCL2L13 was up regulated w.r.t MUC-1/12. These are reflected in the rankings of 2029 (laplace) and 2347 (linear) for MUC1 - BCL2L13; and 2353 (linear) and 1997 (rbf) for MUC12 - BCL2L13; BCL3 was up regulated w.r.t MUC-20. These are reflected in the rankings of 2512 (laplace) and 2440 (rbf) for MUC20 - BCL3; BCL6 was up regulated w.r.t MUC-17. These are reflected in the rankings of 2411(laplace), 2153 (linear) and 1808 (rbf) for MUC17 - BCL6; BCL9L was up regulated w.r.t MUC-17. These are reflected in the rankings of 2101 (laplace) and 2408 (rbf) for MUC20 - BCL9L.
On the right side, we found MUC3A to be up regulated w.r.t BCL2L2. These are reflected in the rankings of 2099 (laplace) and 2397 (rbf) for MUC3A - BCL2L2; MUC3A to be up regulated w.r.t BCL9L. These are reflected in the rankings of 2180 (linear) and 2106 (rbf) for MUC3A - BCL9L;
Table 118.
2nd order combinatorial hypotheses between BCL and SELENBP1.
| Ranking MUC family vs BCL family | |||||||
| Ranking of BCL2L1 w.r.t MUC family | Ranking of MUC family w.r.t BCL2L1 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| MUC1 - BCL2L1 | 2055 | 2297 | 1854 | MUC1 - BCL2L1 | 1226 | 1681 | 986 |
| MUC3A - BCL2L1 | 603 | 2089 | 1637 | MUC3A - BCL2L1 | 759 | 1107 | 678 |
| MUC4 - BCL2L1 | 531 | 1137 | 711 | MUC4 - BCL2L1 | 1758 | 999 | 487 |
| MUC12 - BCL2L1 | 882 | 810 | 1305 | MUC12 - BCL2L1 | 1591 | 900 | 272 |
| MUC13 - BCL2L1 | 1927 | 1201 | 2108 | MUC13 - BCL2L1 | 98 | 2160 | 1099 |
| MUC17 - BCL2L1 | 1170 | 917 | 743 | MUC17 - BCL2L1 | 2500 | 93 | 148 |
| MUC20 - BCL2L1 | 1810 | 700 | 1627 | MUC20 - BCL2L1 | 270 | 343 | 423 |
| Ranking of BCL2L2 w.r.t MUC family | Ranking of MUC family w.r.t BCL2L2 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| MUC1 - BCL2L2 | 1578 | 1425 | 1826 | MUC1 - BCL2L2 | 2476 | 903 | 739 |
| MUC3A - BCL2L2 | 1542 | 370 | 159 | MUC3A - BCL2L2 | 2099 | 241 | 2397 |
| MUC4 - BCL2L2 | 1323 | 2506 | 1988 | MUC4 - BCL2L2 | 797 | 727 | 851 |
| MUC12 - BCL2L2 | 602 | 2504 | 815 | MUC12 - BCL2L2 | 516 | 38 | 1688 |
| MUC13 - BCL2L2 | 2084 | 2402 | 1200 | MUC13 - BCL2L2 | 2201 | 717 | 233 |
| MUC17 - BCL2L2 | 2283 | 2212 | 1279 | MUC17 - BCL2L2 | 903 | 295 | 913 |
| MUC20 - BCL2L2 | 890 | 1886 | 480 | MUC20 - BCL2L2 | 1892 | 569 | 1040 |
| Ranking of BCL2L13 w.r.t MUC family | Ranking of MUC family w.r.t BCL2L13 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| MUC1 - BCL2L13 | 2029 | 2347 | 550 | MUC1 - BCL2L13 | 1838 | 903 | 739 |
| MUC3A - BCL2L13 | 2140 | 1123 | 1100 | MUC3A - BCL2L13 | 173 | 241 | 2397 |
| MUC4 - BCL2L13 | 1497 | 1918 | 1579 | MUC4 - BCL2L13 | 1906 | 727 | 851 |
| MUC12 - BCL2L13 | 581 | 2353 | 1997 | MUC12 - BCL2L13 | 2096 | 38 | 1688 |
| MUC13 - BCL2L13 | 1210 | 2185 | 1658 | MUC13 - BCL2L13 | 1688 | 717 | 233 |
| MUC17 - BCL2L13 | 1079 | 1270 | 1254 | MUC17 - BCL2L13 | 1167 | 295 | 913 |
| MUC20 - BCL2L13 | 187 | 2081 | 535 | MUC20 - BCL2L13 | 1653 | 569 | 1040 |
| Ranking of BCL3 w.r.t MUC family | Ranking of MUC family w.r.t BCL3 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| MUC1 - BCL3 | 458 | 1016 | 1881 | MUC1 - BCL3 | 273 | 360 | 1683 |
| MUC3A - BCL3 | 1642 | 668 | 588 | MUC3A - BCL3 | 1044 | 860 | 1452 |
| MUC4 - BCL3 | 427 | 321 | 457 | MUC4 - BCL3 | 624 | 1360 | 585 |
| MUC12 - BCL3 | 1813 | 311 | 1623 | MUC12 - BCL3 | 1193 | 1092 | 132 |
| MUC13 - BCL3 | 2151 | 641 | 1407 | MUC13 - BCL3 | 279 | 65 | 603 |
| MUC17 - BCL3 | 1106 | 531 | 2310 | MUC17 - BCL3 | 305 | 1285 | 257 |
| MUC20 - BCL3 | 2512 | 63 | 2440 | MUC20 - BCL3 | 16 | 539 | 2198 |
| Ranking of BCL6 w.r.t MUC family | Ranking of MUC family w.r.t BCL6 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| MUC1 - BCL6 | 1652 | 2294 | 173 | MUC1 - BCL6 | 1550 | 595 | 788 |
| MUC3A - BCL6 | 2323 | 1435 | 187 | MUC3A - BCL6 | 407 | 809 | 318 |
| MUC4 - BCL6 | 723 | 711 | 1403 | MUC4 - BCL6 | 176 | 203 | 1963 |
| MUC12 - BCL6 | 184 | 1024 | 1267 | MUC12 - BCL6 | 1126 | 26 | 229 |
| MUC13 - BCL6 | 158 | 1083 | 2198 | MUC13 - BCL6 | 1633 | 1052 | 603 |
| MUC17 - BCL6 | 2411 | 2153 | 1808 | MUC17 - BCL6 | 242 | 719 | 1026 |
| MUC20 - BCL6 | 925 | 840 | 2153 | MUC20 - BCL6 | 1132 | 1669 | 652 |
| Ranking of BCL9L w.r.t MUC family | Ranking of MUC family w.r.t BCL9L | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| MUC1 - BCL9L | 2194 | 744 | 1112 | MUC1 - BCL9L | 1144 | 1999 | 896 |
| MUC3A - BCL9L | 2114 | 1441 | 1359 | MUC3A - BCL9L | 901 | 2180 | 2106 |
| MUC4 - BCL9L | 882 | 466 | 1526 | MUC4 - BCL9L | 658 | 1152 | 781 |
| MUC12 - BCL9L | 1547 | 526 | 2391 | MUC12 - BCL9L | 1733 | 1510 | 366 |
| MUC13 - BCL9L | 1545 | 1891 | 796 | MUC13 - BCL9L | 1529 | 502 | 602 |
| MUC17 - BCL9L | 1282 | 1160 | 1362 | MUC17 - BCL9L | 955 | 1788 | 99 |
| MUC20 - BCL9L | 2101 | 116 | 2408 | MUC20 - BCL9L | 307 | 1516 | 1042 |
| Ranking of BCL10 w.r.t MUC family | Ranking of MUC family w.r.t BCL10 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| MUC1 - BCL10 | 1325 | 1524 | 1900 | MUC1 - BCL10 | 547 | 1319 | 284 |
| MUC3A - BCL10 | 1298 | 1004 | 1509 | MUC3A - BCL10 | 1681 | 751 | 2250 |
| MUC4 - BCL10 | 304 | 1632 | 1050 | MUC4 - BCL10 | 591 | 570 | 151 |
| MUC12 - BCL10 | 1019 | 1093 | 2239 | MUC12 - BCL10 | 38 | 1155 | 817 |
| MUC13 - BCL10 | 358 | 1687 | 2004 | MUC13 - BCL10 | 517 | 2229 | 455 |
| MUC17 - BCL10 | 524 | 2038 | 1579 | MUC17 - BCL10 | 216 | 803 | 132 |
| MUC20 - BCL10 | 1380 | 619 | 2081 | MUC20 - BCL10 | 97 | 465 | 239 |
Table 119.
2nd order combinatorial hypotheses between MUC and BCL family.
| Unexplored combinatorial hypotheses | |
| MUC w.r.t BCL | |
| MUC-3A | BCL2L2 |
| MUC-3A | BCL9L |
| BCL w.r.t MUC | |
| MUC-1/13 | BCL2L1 |
| MUC-4/13/17 | BCL2L2 |
| MUC-1/12 | BCL2L13 |
| MUC-20 | BCL3 |
| MUC-17 | BCL6 |
| MUC-20 | BCL9L |
3.8.6. EXOSC - BCL Cross Family Analysis
The exosome complex is involved in the degradation of various kinds of RNA. Recently, Deng et al. [235] observe that Exosome-transmitted LINC00461 promotes multiple myeloma cell proliferation and suppresses apoptosis by modulating microRNA/BCL-2 expression. Xu et al. [236] show that Exosome-derived microRNA-29c induces apoptosis of BIU-87 cells by down regulating BCL-2 and MCL-1. Exosomes were demonstrated to upregulate the expression of Bcl-2 and Cyclin D1 proteins, but reduce the levels of Bax and caspase-3 proteins in these cells in work of Yang et al. [237]. In western blot analysis results showed that exosomes can block the significant reduction of BCL-2, full-length caspase-3 and full-length PARP, while preventing the increase of BAX, cleaved caspase-3 and cleaved PARP induced by VP16, as studied by Wang et al. [238]. These findings point to the definite synergistic role of exosome with BCL family. In CRC cells, both exosome components EXOSC and BCL family members were found to be down regulated, after ETC-1922159 drug treatment. The search engine allocated low numerical valued ranks for many of the EXOSC and BCL combinations which might suggest greater role of EXOSC along with BCL. However, the nature of the mechanism between the two families yet needs to be explored, despite the generated hypothesis of possible synergy.
Table 120 shows rankings of EXOSC and BCL family with respect to each other. Left half of the Table shows rankings of EXOSC w.r.t BCL and right half shows the vice versa. On the left, we find EXOSC2 to be down regulated w.r.t BCL-2L12/6B/7A/9/11A/11B. These are shown in the rankings of 723 (laplace), 355 (linear) and 1211 (rbf) for EXOSC2 - BCL2L12; 1092 (laplace), 1033 (linear) and 638 (rbf) for EXOSC2 - BCL6; 1633 (laplace), 1047 (linear) and 317 (rbf) for EXOSC2 - BCL7A; 699 (laplace), 559 (linear) and 425 (rbf) for EXOSC2 - BCL9; 338 (laplace), 319 (linear) and 1598 (rbf) for EXOSC2 - BCL11A; and 1285 (laplace), 1440 (linear) and 812 (rbf) for EXOSC2 - BCL11B; EXOSC3 was found to down regulated w.r.t BCL11B. This is reflected in rankigns of 1677 (laplace), 199 (linear) and 267 (rbf) for EXOSC3 - BCL11B. EXOSC5 was found to be down regulated w.r.t BCL family. These are reflected in the rankings of 498 (laplace), 1342 (linear) and 436 (rbf) for EXOSC5 - BCL2L12; 786 (laplace), 1272 (linear) and 1194 (rbf) for EXOSC5 - BCL6B; 374 (laplace), 1338 (linear) and 874 (rbf) for EXOSC5 - BCL7A; 613 (laplace), 946 (linear) and 772 (rbf) for EXOSC5 - BCL9; 459 (laplace), 90 (linear) and 1034 (rbf) for EXOSC5 - BCL11A; and 1404 (laplace) and 1558 (linear) for EXOSC5 - BCL11B; EXOSC6 was found to be down regulated w.r.t BCL family. These are reflected in rankings of 1676 (laplace), 787 (linear) and 944 (rbf) for EXOSC6 - BCL7A; 1059 (linear) and 1091 (rbf) for EXOSC6 - BCL9; 1677 (laplace) and 1573 (linear) for EXOSC6 - BCL11A; EXOSC7 was found to be down regulated w.r.t BCL family. These are reflected in rankings of 666 (laplace); 98 (linear) and 743 (rbf) EXOSC7 - BCL6B; 1501 (linear) and 1513 (rbf) for EXOSC7 - BCL7A; and 1477 (laplace) and 1217 (rbf) for EXOSC7 - BCL11A; EXOSC8 was found to be down regulated w.r.t BCL family. Thesea reflected in 1175 (laplace), 1504 (linear) and 1743 (rbf) for EXOSC8 - BCL7A; 906 (linear) and 1130 (rbf) EXOSC8 - BCL11A; and 605 (linear) and 374 (rbf) for EXOSC8 - BCL11B; EXOSC9 found to be down regulate w.r.t BCL family. These are reflected in rankings of 1179 (laplace); 1018 (linear) and 687 (rbf) for EXOSC9 - BCL2L12; 437 (laplace), 852 (linear) and 1358 (rbf) EXOSC9 - BCL6B; 821 (laplace), 346 (linear) and 727 (rbf) for EXOSC9 - BCL7A; 1305 (laplace) and 299 (rbf) EXOSC9 - BCL9; 1569 (laplace), 549 (linear) and 1456 (rbf) for EXOSC9 - BCL11B.
On the right, we find BCL-6B/11A/11B to be down regulated w.r.t EXOSC2. These are reflected in the rankings of 202 (laplace), 81 (linear) and 194 (rbf) for EXOSC2 - BCL6B; 574 (laplace), 834 (linear) and 1055 (rbf) for EXOSC2 - BCL11A; and 1368 (laplace), 1353 (linear) and 1455 (rbf) for EXOSC2 - BCL11B. BCL-6B/7A/11A was found to be down regulated w.r.t EXOSC3. These are reflected in rankings of 571 (laplace), 335 (linear) and 307 (rbf) for EXOSC3 - BCL6B; 1739 (laplace) and 1700 (rbf) for EXOSC3 - BCL7A; and 1018 (laplace), 1345 (linear) and 483 (rbf) for EXOSC3 - BCL11A; BCL-6B/11A/11B was found to be down regulated w.r.t EXOSC5. These were reflected in rankings of 571 (laplace), 335 (linear) and 307 (rbf) for EXOSC5 - BCL6B; 756 (laplace), 389 (linear) and 1183 (rbf) for EXOSC5 - BCL11A; and 1368 (laplace), 1353 (linear) and 1455 (rbf) for EXOSC5 - BCL11B. BCL-9 was found to be down regulated w.r.t EXOSC6. These are reflected in rankigns of 851 (linear) and 1564 (rbf) for EXOSC6 - BCL9. BCL-2L12/7A/9/11A/11B was found to be down regulated w.r.t EXOSC7. These are reflected in rankings of 1551 (linear) and 1099 (rbf) for EXOSC7 - BCL2L12; 1282 (laplace), 831 (linear) and 1218 (rbf) for EXOSC7 - BCL7A; 1234 (linear) and 328 (rbf) for EXOSC7 - BCL9; 520 (laplace), 117 (linear) and 686 (rbf) for EXOSC7 - BCL11A; and 1529 (laplace) and 1418 (rbf) for EXOSC7 - BCL11B; BCL-6B/11A was found to be down regulated with EXOSC8. These are reflected in rankings of 190 (laplace), 1630 (linear) and 472 (rbf) for EXOSC8 - BCL6B; and 944 (laplace) and 532 (rbf) for EXOSC8 - BCL11A. Finally, BCL-6B/9/11A/11B was found to be down regulated with EXOSC9. These are reflected in rankings of 634 (laplace), 304 (linear) and 146 (rbf) for EXOSC9 - BCL6B; 1197 (laplace) and 1279 (rbf) for EXOSC9 - BCL9; 481 (laplace), 441 (linear) and 1372 (rbf) for EXOSC9 - BCL11A; and 1454 (linear) and 133 (rbf) for EXOSC9 - BCL11B.
Table 121 shows the derived influences which can be represented graphically, with the following influences - • EXOSC w.r.t BCL with EXOSC2 <- BCL-2L12/6B/7A/9/11A/11B; EXOSC3 <- BCL-11B; EXOSC5 <- BCL-2L12/6B/7A/9/11A/11B; EXOSC6 <- BCL-7A/9/11A; EXOSC7 <- BCL-6B/7A/11A; EXOSC8 <- BCL-7A/11A/11B and EXOSC9 <- BCL-2L12/6B/7A/9/11B; and • BCL w.r.t EXOSC with EXOSC2 -> BCL-6B/11A/11B; EXOSC3 -> BCL-6B/7A/11A; EXOSC5 -> BCL-6B/11A/11B; EXOSC6 -> BCL-2L12/9; EXOSC7 -> BCL-2L12/7A/9/11A/11B; EXOSC8 -> BCL-6B/11A and EXOSC9 -> BCL-6B/9/11A/11B.
3.9. Poliovirus-Receptor Related Synergies
3.9.1. PVR - Interferon Cross Family Analysis
Brown et al. [239] show that cancer immunotherapy with recombinant poliovirus induces IFN-dominant activation of dendritic cells and tumor antigen-specific CTLs. Stamm et al. [240] show that immune checkpoints PVR and PVRL2 are prognostic markers in AML and their blockade represents a new therapeutic option. Stamm et al. [241] observe that expression of novel immune checkpoint molecules PVR and PVRL2 confers a negative prognosis to patients with acute myeloid leukemia and their blockade augments T-Cell mediated lysis of AML cells alone or in combination with the BiTE® antibody construct AMG 330. In a latest development, Whelan et al. [242] observe that poliovirus receptor related immunoglobulin domain containing (PVRIG) and poliovirus receptor-related 2 (PVRL2) are induced in cancer and inhibit CD8+ T-cell function. For colorectal cancer, the were highest percentage of PVR+PVRL2− cells. In CRC cells treated with ETC-1922159, it was found that PVR, PVRL2 and PVRL4 were up regulated. Whelan et al. [242] report that when they compared the combination of receptor blockade (i.e., anti-PVRIG and anti-TIGIT) with the combination of ligand blockade (i.e., anti-PVR and anti-PVRL2), similar increases in IFN-γ were observed, suggesting no additional functional interactions are present among these proteins. This might also suggest that expression of PVRL2 blocks the production of IFN-γ. Using the search engine, rankings at 2nd order indicate similar patterns of combinatorial synergy. These ranks are tabulated in Table 122. Note that high numerical valued ranks indicate a synergy between PVR and IFN family. However, low numercial valued ranks possibly indicate the negative role, i.e PVR family up regulation leads to blocking of IFN family production.
Here we depict the possible synergy of up regulation of PVR family with IFN family in CRC cells treated with ETC-1922159. However, of low numerical valued ranks might indicate the similar behaviour as found by Whelan et al. [242]. On the left we find PVR ranks w.r.t IFN and on the right vice versa. We found PVR to be up regulated w.r.t IFN-E/GR2/LR1. These are reflected in rankings of 2044 (linear) and 1975 (rbf) for PVR - IFNE; 1911 (laplace) and 1871 (linear) for PVR - IFNGR2; and 2212 (linear) and 1884 (rbf) for PVR - IFNLR1; PVRL2 was up regulated w.r.t IFN-E. This was reflected in rankings of 1851 (laplace) and 2120 (rbf) for PVRL2 - IFNE; Reversibly, in context of findings by Whelan et al. [242] all low numerical valued ranks point to the fact that PVR/PVRL2/PVRL4 up regulation might be blocking the production of IFN family members. On the right side, we found IFN-GR1/LR1 to be up regulated w.r.t PVR. These are reflected in rankings of 2268 (laplace), 2040 (linear) and 2235 (rbf) for PVR - IFNGR1 and 2119 (laplace) and 1918 (linear) for PVR - IFNLR1; IFN-GR2 was up regulated w.r.t PVRL2. These are reflected in rankings of 2049 (laplace) and 2056 (rbf) for PVRL2 - IFNGR2; IFN-E was up regulated w.r.t PVRL4. These are reflected in rankings of 2026 (laplace) and 1908 (linear). Again, reversibly, in context of findings by Whelan et al. [242] all low numerical valued ranks point to the fact that IFN family up regulation might indicate bloackage of PVR family members.
Table 123 shows the derived influences which can be represented graphically, with the following influences - • PVR w.r.t IFN with PVR <- IFN-E/GR2/LR1; PVRL2 <- IFN-E; and • IFN w.r.t PVR with IFN-GR1/LR1 -> PVR; IFN-GR2 -> PVRL2; IFN-E -> PVRL4;
Table 122.
2nd order combinatorial hypotheses between IFN and PVR family.
| Ranking PVR family vs IFN family | |||||||
| Ranking of PVR w.r.t IFN family | Ranking of IFN family w.r.t PVR | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| PVR - IFNAR2 | 1378 | 1651 | 1539 | PVR - IFNAR2 | 1630 | 161 | 930 |
| PVR - IFNE | 1305 | 2044 | 1975 | PVR - IFNE | 1071 | 1486 | 362 |
| PVR - IFNGR1 | 1331 | 268 | 1000 | PVR - IFNGR1 | 2268 | 2040 | 2235 |
| PVR - IFNGR2 | 1911 | 1871 | 1426 | PVR - IFNGR2 | 598 | 1059 | 832 |
| PVR - IFNLR1 | 717 | 2212 | 1884 | PVR - IFNLR1 | 2119 | 1918 | 1499 |
| PVR - IFNWP19 | 1648 | 1438 | 1547 | PVR - IFNWP19 | 1699 | 168 | 2218 |
| Ranking of PVRL2 w.r.t IFN family | Ranking of IFN family w.r.t PVRL2 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| PVRL2 - IFNAR2 | 1877 | 1085 | 492 | PVRL2 - IFNAR2 | 623 | 1259 | 2073 |
| PVRL2 - IFNE | 1851 | 990 | 2120 | PVRL2 - IFNE | 683 | 328 | 1416 |
| PVRL2 - IFNGR1 | 1174 | 510 | 526 | PVRL2 - IFNGR1 | 1352 | 1885 | 1433 |
| PVRL2 - IFNGR2 | 1608 | 173 | 1036 | PVRL2 - IFNGR2 | 2049 | 490 | 2056 |
| PVRL2 - IFNLR1 | 1577 | 701 | 1333 | PVRL2 - IFNLR1 | 535 | 1258 | 295 |
| PVRL2 - IFNWP19 | 1455 | 954 | 182 | PVRL2 - IFNWP19 | 870 | 1803 | 1394 |
| Ranking of PVRL4 w.r.t IFN family | Ranking of IFN family w.r.t PVRL4 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| PVRL4 - IFNAR2 | 1555 | 227 | 2433 | PVRL4 - IFNAR2 | 490 | 2303 | 1701 |
| PVRL4 - IFNE | 64 | 781 | 1466 | PVRL4 - IFNE | 2026 | 1908 | 465 |
| PVRL4 - IFNGR1 | 2218 | 651 | 188 | PVRL4 - IFNGR1 | 560 | 793 | 889 |
| PVRL4 - IFNGR2 | 220 | 31 | 873 | PVRL4 - IFNGR2 | 213 | 2079 | 31 |
| PVRL4 - IFNLR1 | 284 | 958 | 683 | PVRL4 - IFNLR1 | 766 | 1432 | 2153 |
| PVRL4 - IFNWP19 | 138 | 2271 | 384 | PVRL4 - IFNWP19 | 788 | 1046 | 921 |
Table 123.
2nd order combinatorial hypotheses between PVR and IFN family.
| Unexplored combinatorial hypotheses | |
| PVR w.r.t IFN | |
| PVR | IFN-E/GR2/LR1 |
| PVRL2 | IFN-E |
| IFN w.r.t PVR | |
| IFN-GR1/LR1 | PVR |
| IFN-GR2 | PVRL2 |
| IFN-E | PVRL4 |
3.9.2. Interferon - Wnt Cross Family Analysis
The crosstalk between β-catenin signaling and type I, type II and type III interferons in lung cancer cells has been observed by Bai et al. [243]. Hillesheim et al. [244] show that β-catenin promotes the type I IFN synthesis and the IFN-dependent signaling response but is suppressed by influenza A virus-induced RIG-I/NF-κB signaling. Ohsugi et al. [245] show that decreased expression of interferon-induced protein 2 (IFIT2) by Wnt/β-catenin signaling confers anti-apoptotic properties to colorectal cancer cells. In CRC cells treated with ETC-1922159, members of Wnt and IFN family were up regulated. The search engine assigned high numerical valued ranks to a few of the 2nd order combinations. These are depicted in Table 124.
On the left, is the rankings of IFN family w.r.t Wnt family. On the right are the rankings of Wnt family w.r.t IFN family. On the left we found IFNE to be up regulated w.r.t WNT7B. These are depicted in rankings of 1872 (laplace) and 2341 (rbf) for IFNE - WNT7B. On the right, we found WNT-2B/4/7B to be up regulated w.r.t IFNGR1. These are reflected in ranking of 1849 (linear) and 2282 (rbf) for IFNGR1 - WNT2B; 2428 (laplace) and 2479 (rbf) for IFNGR1 - WNT4 and 2278 (linear) and 2164 (rbf) for IFNGR1 - WNT7B. WNT-2B was up regulated w.r.t IFNLR1. This is reflected in ranking of 2263 (laplace) and 2216 (rbf) for IFNLR1 - WNT2B; Finally, WNT4 was up regulated w.r.t IFNWP19. This is reflected in ranking of 2423 (linear) and 2330 (rbf) for IFNWP19 - WNT4. Table 125 shows the derived influences which can be represented graphically, with the following influences - • IFN w.r.t WNT with IFNE <- WNT7B; • WNT w.r.t IFN with IFNGR1 -> WNT2B; IFNGR1 -> WNT-2B/4/7B; IFNLR1 -> WNT2B; and IFNWP19 -> WNT4.
Table 124.
2nd order combinatorial hypotheses between WNT and IFN family.
| Ranking IFN family vs WNT family | |||||||
| Ranking of IFNAR2 w.r.t WNT family | Ranking of WNT family w.r.t IFNAR2 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IFNAR2 - WNT2B | 826 | 829 | 1463 | IFNAR2 - WNT2B | 785 | 1146 | 1642 |
| IFNAR2 - WNT4 | 680 | 658 | 802 | IFNAR2 - WNT4 | 1969 | 130 | 126 |
| IFNAR2 - WNT7B | 1009 | 1252 | 581 | IFNAR2 - WNT7B | 2208 | 1635 | 1647 |
| IFNAR2 - WNT9A | 532 | 180 | 1737 | IFNAR2 - WNT9A | 1223 | 1422 | 1632 |
| Ranking of IFNE w.r.t WNT family | Ranking of WNT family w.r.t IFNE | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IFNE - WNT2B | 1612 | 1973 | 519 | IFNE - WNT2B | 1057 | 1146 | 239 |
| IFNE - WNT4 | 1609 | 2262 | 1320 | IFNE - WNT4 | 585 | 440 | 6 |
| IFNE - WNT7B | 1872 | 1240 | 2341 | IFNE - WNT7B | 2055 | 941 | 936 |
| IFNE - WNT9A | 2114 | 1029 | 267 | IFNE - WNT9A | 124 | 458 | 708 |
| Ranking of IFNGR1 w.r.t WNT family | Ranking of WNT family w.r.t IFNGR1 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IFNGR1 - WNT2B | 1623 | 144 | 361 | IFNGR1 - WNT2B | 1057 | 1849 | 2282 |
| IFNGR1 - WNT4 | 225 | 455 | 1773 | IFNGR1 - WNT4 | 2428 | 1226 | 2479 |
| IFNGR1 - WNT7B | 1004 | 1259 | 1135 | IFNGR1 - WNT7B | 710 | 2278 | 2164 |
| IFNGR1 - WNT9A | 601 | 958 | 1864 | IFNGR1 - WNT9A | 1668 | 1725 | 1462 |
| Ranking of IFNGR2 w.r.t WNT family | Ranking of WNT family w.r.t IFNGR2 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IFNGR2 - WNT2B | 1224 | 1322 | 2156 | IFNGR2 - WNT2B | 828 | 1599 | 400 |
| IFNGR2 - WNT4 | 584 | 1117 | 59 | IFNGR2 - WNT4 | 498 | 33 | 168 |
| IFNGR2 - WNT7B | 1185 | 745 | 242 | IFNGR2 - WNT7B | 1964 | 1020 | 638 |
| IFNGR2 - WNT9A | 754 | 501 | 676 | IFNGR2 - WNT9A | 261 | 1711 | 654 |
| Ranking of IFNLR1 w.r.t WNT family | Ranking of WNT family w.r.t IFNLR1 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IFNLR1 - WNT2B | 1621 | 851 | 510 | IFNLR1 - WNT2B | 2263 | 1683 | 2216 |
| IFNLR1 - WNT4 | 1538 | 250 | 220 | IFNLR1 - WNT4 | 2364 | 231 | 503 |
| IFNLR1 - WNT7B | 1012 | 173 | 506 | IFNLR1 - WNT7B | 406 | 573 | 446 |
| IFNLR1 - WNT9A | 347 | 134 | 2160 | IFNLR1 - WNT9A | 1815 | 1709 | 106 |
| Ranking of IFNWP19 w.r.t WNT family | Ranking of WNT family w.r.t IFNWP19 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IFNWP19 - WNT2B | 826 | 176 | 787 | IFNWP19 - WNT2B | 1600 | 2096 | 1650 |
| IFNWP19 - WNT4 | 680 | 1507 | 391 | IFNWP19 - WNT4 | 1441 | 2423 | 2330 |
| IFNWP19 - WNT7B | 1009 | 1101 | 1327 | IFNWP19 - WNT7B | 1838 | 966 | 1478 |
| IFNWP19 - WNT9A | 532 | 1404 | 431 | IFNWP19 - WNT9A | 1354 | 1968 | 703 |
Table 125.
2nd order combinatorial hypotheses between IFN and WNT family.
| Unexplored combinatorial hypotheses | |
| IFN w.r.t WNT | |
| IFNE | WNT7B |
| WNT w.r.t IFN | |
| IFNGR1 | WNT2B |
| IFNGR1 | WNT4/WNT7B |
| IFNLR1 | WNT2B |
| IFNWP19 | WNT4 |
3.9.3. PVR - WNT Cross Family Analysis
Mutations in PVRL4, encoding cell adhesion molecule nectin-4, causes Ectodermal dysplasia-syndactyly syndrome, Brancati et al. [1]. Interaction with cadherins also implies an influence of nectin-4 on Wnt signaling, which plays a relevant role in limb development (Brancati et al. [1]). However, not much work has been done to explore the relation of Wnts and PVR family. In CRC cells treated with ETC-1922159, both were found up regulated. The search engine alloted high numerical valued rankings to some combinations thus indicating a possibility of high combinatorial synergy also. Table 126 shows the rankings of PVR family w.r.t to Wnts on the left and vice versa on the right. We found, PVR up regulated w.r.t WNT9A and this is reflected in rankings of 2322 (laplace) and 2202 (rbf). On the right, we found WNT-7B/9A to be up regulated w.r.t PVR. These are reflected in rankings of 2216 (laplace), 1844 (linear) and 2096 (rbf) for PVR - WNT7B; and 2152 (laplace) and 2120 (rbf) for PVR - WNT9A. Also, WNT-4 was up regulated w.r.t PVRL2. This is reflected in rankings of 2324 (laplace) and 2462 (linear) for PVRL2 - WNT4.
Table 127 shows the derived influences which can be represented graphically, with the following influences - • PVR w.r.t WNT with PVR <- WNT9A; and • WNT w.r.t PVR with WNT-7B/9A <- PVR and WNT4 <- PVRL2; In the light of the recent findings of PVR with IFN and the known interactions between IFN and Wnts, there might be a possibilty to explore the bridge of PVR, IFN and WNTs. The above 3 fold (PVR - IFN; IFN - WNT; WNT - PVR), 2 way cross family analysis might shed light on the possible combinations that might be of import.
Table 126.
2nd order combinatorial hypotheses between WNT and PVR family.
| Ranking PVR family vs WNT family | |||||||
| Ranking of PVR w.r.t WNT family | Ranking of WNT family w.r.t PVR | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| PVR - WNT2B | 2204 | 733 | 1 | PVR - WNT2B | 1205 | 2257 | 607 |
| PVR - WNT4 | 1295 | 970 | 878 | PVR - WNT4 | 38 | 2470 | 1094 |
| PVR - WNT7B | 1237 | 770 | 1887 | PVR - WNT7B | 2216 | 1844 | 2096 |
| PVR - WNT9A | 2322 | 649 | 2202 | PVR - WNT9A | 2152 | 2120 | 1131 |
| Ranking of PVRL2 w.r.t WNT family | Ranking of WNT family w.r.t PVRL2 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| PVRL2 - WNT2B | 616 | 375 | 2381 | PVRL2 - WNT2B | 1901 | 1044 | 621 |
| PVRL2 - WNT4 | 1110 | 1584 | 1391 | PVRL2 - WNT4 | 2324 | 216 | 2462 |
| PVRL2 - WNT7B | 2186 | 1122 | 349 | PVRL2 - WNT7B | 560 | 560 | 953 |
| PVRL2 - WNT9A | 110 | 1367 | 1858 | PVRL2 - WNT9A | 1044 | 1502 | 794 |
| Ranking of PVRL4 w.r.t WNT family | Ranking of WNT family w.r.t PVRL4 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| PVRL4 - WNT2B | 949 | 565 | 95 | PVRL4 - WNT2B | 78 | 966 | 1938 |
| PVRL4 - WNT4 | 885 | 1672 | 2149 | PVRL4 - WNT4 | 611 | 1922 | 1488 |
| PVRL4 - WNT7B | 299 | 241 | 798 | PVRL4 - WNT7B | 1192 | 1159 | 2505 |
| PVRL4 - WNT9A | 1375 | 1306 | 492 | PVRL4 - WNT9A | 1383 | 634 | 224 |
Table 127.
2nd order combinatorial hypotheses between PVR and WNT family.
| Unexplored combinatorial hypotheses | |
| PVR w.r.t WNT | |
| PVR | WNT9A |
| WNT w.r.t PVR | |
| WNT-7B/9A | PVR |
| WNT4 | PVRL2 |
3.9.4. PVR - Integrin Cross Family Analysis
PVRL4 promotes anchorage-independence by driving cell-to-cell attachment and matrix-independent integrin β4/SHP-2/c-Src activation, as observed by Pavlova et al. [246]. Integrins are the major metazoan receptors for cell adhesion to extracellular matrix proteins and, in vertebrates, also play important roles in certain cell-cell adhesions Hynes [247]. It has been recently shown that human NK cells recognize PVR through the receptor DNAM-1, which triggers NK cell stimulation in association with 2 integrin. Fuchs et al. [248] additionally show that NK cells recognize PVR through an additional receptor, CD96, or T cell-activated increased late expression (Tactile). Ferroptosis is a type of programmed cell death dependent on iron and characterized by the accumulation of lipid peroxides, and is genetically and biochemically distinct from other forms of regulated cell death such as apoptosis (Wikipedia contributors [249]). Cell clustering mediated by the adhesion protein PVRL4 is necessary for α6β4 integrin–promoted ferroptosis resistance in matrix-detached cells, as observed by Brown et al. [250]. These findings suggest the possibility to synergy between PVR and Integrin family. In CRC cells treated with ETC-1922159, PVR and integrin families were up regulated. The search engine alloted high numerical valued ranks to some of the 2nd order combinations of PVR and integrin family members thus pointing to possible synergy in CRC cells. Table 128 shows the rankings of PVR along with integrin family members.
On the left side, we found PVRL2 to be up regulated w.r.t ITG-B4. This is reflected in the rankings of 1857 (laplace) and 1750 (rbf) for ITGB4-PVRL2. PVRL4 was up regulated w.r.t ITGB5. This is reflected in rankings of 2418 (linear) and 1802 (rbf) for ITGB5-PVRL4. On the right side, ITG-A2/B8 were found up regulated w.r.t PVR. These are reflected in rankings of 2062 (linear) and 2106 (rbf) for ITGA2-PVR; and 2498 (linear) and 2366 (rbf) for ITGB8-PVR. ITG-B1BP1/B6/B8 were found up regulated w.r.t PVRL2. These are reflected in rankings of 2470 (linear) and 2408 (rbf) for ITGB1BP1-PVRL2; 2428 (linear) and 2364 (rbf) for ITGB6-PVRL2; and 2046 (laplace), 2385 (linear) and 2110 (rbf) for ITGB8-PVRL2. ITG-B1BP1/B5 were found up regulated w.r.t PVRL4. These are reflected in rankings of 1815 (laplace) and 2362 (rbf) for ITGB1BP1-PVRL4; and 1818 (laplace) and 2256 (rbf) for ITGB5-PVRL4.
Table 129 shows the derived influences which can be represented graphically, with the following influences - • PVR w.r.t ITG with PVRL2 <- ITGB4; and PVRL4 <- ITGB5 and • ITG w.r.t PVR with ITG-A2/B8 <- PVR; ITG-B1BP1/B6/B8 <- PVRL2; and ITG-B1BP1/B5 <- PVRL4.
Table 128.
2nd order combinatorial hypotheses between ITG and PVR family.
| Ranking PVR family vs ITG family | |||||||
| Ranking of PVR w.r.t ITG family | Ranking of ITG family w.r.t PVR | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| ITGA2-PVR | 294 | 564 | 1996 | ITGA2-PVR | 627 | 2062 | 2106 |
| ITGA3-PVR | 1739 | 117 | 1420 | ITGA3-PVR | 2172 | 99 | 827 |
| ITGA6-PVR | 214 | 1328 | 435 | ITGA6-PVR | 576 | 2199 | 817 |
| ITGB1-PVR | 1896 | 136 | 1121 | ITGB1-PVR | 1506 | 1093 | 2203 |
| ITGB1BP1-PVR | 1876 | 1724 | 1505 | ITGB1BP1-PVR | 1241 | 1108 | 535 |
| ITGB4-PVR | 783 | 1495 | 1044 | ITGB4-PVR | 1499 | 120 | 873 |
| ITGB5-PVR | 1719 | 981 | 490 | ITGB5-PVR | 1269 | 1433 | 914 |
| ITGB6-PVR | 1457 | 664 | 1744 | ITGB6-PVR | 1686 | 988 | 879 |
| ITGB8-PVR | 283 | 290 | 334 | ITGB8-PVR | 1407 | 2498 | 2366 |
| Ranking of PVRL2 w.r.t ITG family | Ranking of ITG family w.r.t PVRL2 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| ITGA2-PVRL2 | 1072 | 501 | 851 | ITGA2-PVRL2 | 1883 | 327 | 1141 |
| ITGA3-PVRL2 | 960 | 1905 | 1160 | ITGA3-PVRL2 | 1199 | 1937 | 39 |
| ITGA6-PVRL2 | 352 | 993 | 212 | ITGA6-PVRL2 | 2102 | 709 | 1337 |
| ITGB1-PVRL2 | 720 | 1751 | 836 | ITGB1-PVRL2 | 922 | 568 | 1546 |
| ITGB1BP1-PVRL2 | 1436 | 1313 | 88 | ITGB1BP1-PVRL2 | 168 | 2470 | 2408 |
| ITGB4-PVRL2 | 1857 | 1269 | 1750 | ITGB4-PVRL2 | 565 | 440 | 1197 |
| ITGB5-PVRL2 | 238 | 100 | 1314 | ITGB5-PVRL2 | 543 | 1738 | 1605 |
| ITGB6-PVRL2 | 1873 | 582 | 1492 | ITGB6-PVRL2 | 1052 | 2428 | 2364 |
| ITGB8-PVRL2 | 695 | 612 | 1500 | ITGB8-PVRL2 | 2046 | 2385 | 2110 |
| Ranking of PVRL4 w.r.t ITG family | Ranking of ITG family w.r.t PVRL4 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| ITGA2-PVRL4 | 69 | 951 | 917 | ITGA2-PVRL4 | 2154 | 666 | 1266 |
| ITGA3-PVRL4 | 66 | 1648 | 825 | ITGA3-PVRL4 | 2355 | 357 | 801 |
| ITGA6-PVRL4 | 994 | 528 | 109 | ITGA6-PVRL4 | 2359 | 299 | 157 |
| ITGB1-PVRL4 | 1631 | 1724 | 917 | ITGB1-PVRL4 | 2100 | 1282 | 526 |
| ITGB1BP1-PVRL4 | 1369 | 90 | 462 | ITGB1BP1-PVRL4 | 1815 | 1287 | 2362 |
| ITGB4-PVRL4 | 743 | 1602 | 2443 | ITGB4-PVRL4 | 126 | 1844 | 703 |
| ITGB5-PVRL4 | 2418 | 1802 | 119 | ITGB5-PVRL4 | 1818 | 834 | 2256 |
| ITGB6-PVRL4 | 500 | 1187 | 122 | ITGB6-PVRL4 | 1618 | 2425 | 402 |
| ITGB8-PVRL4 | 861 | 699 | 780 | ITGB8-PVRL4 | 1641 | 394 | 1282 |
Table 129.
2nd order combinatorial hypotheses between PVR and ITG family.
| Unexplored combinatorial hypotheses | |
| PVR w.r.t ITG | |
| PVRL2 | ITGB4 |
| PVRL4 | ITGB5 |
| ITG w.r.t PVR | |
| ITG-A2/B8 | PVR |
| ITG-B1BP1/B6/B8 | PVRL2 |
| ITG-B1BP1/B5 | PVRL4 |
3.9.5. PVR - TNF Cross Family Analysis
Abdullah et al. [251] show that wild-type measles virus infection upregulates poliovirus receptor-related 4 and causes apoptosis in brain endothelial cells by induction of Tumor Necrosis Factor-related apoptosis-inducing ligand. Fabre-Lafay et al. [252] show that Nectin-4 (PVRL4), a new serological breast cancer marker, is a substrate for tumor necrosis factor-α-converting enzyme (TACE)/ADAM-17. These and other findings indicate the role of poliovirus receptor along with TNF family members. In CRC cells treated with ETC-1922159, PVR and TNF families were up regulated. The search engine alloted high numerical valued ranks to some of the 2nd order combinations of PVR and TNF family members thus pointing to possible synergy in CRC cells. Table 130 shows the rankings of PVR along with TNF family members.
On the left side, we found PVR to be up regulated w.r.t TNF and TNF-AIP1/AIP2/RSF1A/RSF10A. This is reflected in the rankings of 1963 (laplace), 2422 (linear) and 1822 (rbf) for TNF-PVR; 2210 (linear) and 2243 (rbf) for TNFAIP1-PVR; 2028 (laplace) and 2451 (rbf) for TNFAIP2-PVR; 2029 (laplace) and 2078 (rbf) for TNFRSF1A-PVR and 1978 (linear) and 1942 (rbf) for TNFRSF10A-PVR. PVRL2 to be up regulated w.r.t TNF-AIP2. This is reflected in the rankings of 2515 (laplace) and 2423 (linear) for TNFAIP2-PVRL2. On the right side, TNFRSF14 to be up regulated w.r.t PVR. This is reflected in rankigns of 2351 (laplace) and 2289 (linear) for TNFRSF14-PVR. TNF-AIP1/AIP2/RSF1A/RSF10B/RSF21 to be up regulated w.r.t PVRL2. These are reflected in rankings of 2244 (laplace) and 1932 (rbf) for TNFAIP1-PVRL2; 2337 (laplace), 2483 (linear) and 2401 (rbf) for TNFAIP2-PVRL2; 2355 (laplace) and 1810 (rbf) for TNFRSF1A-PVRL2; and 2120 (laplace) and 1782 (rbf) for TNFRSF21-PVRL2. TNF-AIP2/RSF10D/RSF12A/RSF21 to be up regulated w.r.t PVRL4. These are reflected in rankings of 2270 (laplace) and 2429 (linear) for TNFAIP2-PVRL4; 1799 (laplace) and 2430 (rbf) for TNFRSF10D-PVRL4; 2386 (laplace) and 2064 (rbf) for TNFRSF12A-PVRL4; and 2441 (laplace) and 1917 (linear) for TNFRSF21-PVRL4.
Table 131 shows the derived influences which can be represented graphically, with the following influences - • PVR w.r.t TNF with PVR <- TNF, TNF-AIP1/AIP2/RSF1A/RSF10A; and PVRL2 <- TNF-AIP2; and • TNF w.r.t PVR with TNFRSF14 <- PVR; TNF-AIP1/AIP2/RSF1A/RSF10B/RSF21 <- PVRL2; and TNF-AIP2/RSF10D/RSF12A/RSF21 <- PVRL4.
Table 130.
2nd order combinatorial hypotheses between PVR and TNF family.
| Ranking PVR vs TNF family | |||||||
| Ranking of PVR w.r.t TNF family | Ranking of TNF family w.r.t PVR | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| TNF-PVR | 1963 | 2422 | 1822 | TNF-PVR | 451 | 54 | 209 |
| TNFAIP1-PVR | 88 | 2210 | 2243 | TNFAIP1-PVR | 527 | 474 | 743 |
| TNFAIP2-PVR | 2028 | 300 | 2451 | TNFAIP2-PVR | 1422 | 632 | 1486 |
| TNFAIP3-PVR | 2454 | 1065 | 1293 | TNFAIP3-PVR | 517 | 1476 | 1611 |
| TNFRSF1A-PVR | 2029 | 500 | 2078 | TNFRSF1A-PVR | 1530 | 778 | 1865 |
| TNFRSF10A-PVR | 1140 | 1978 | 1942 | TNFRSF10A-PVR | 2124 | 1648 | 1420 |
| TNFRSF10B-PVR | 1529 | 1608 | 463 | TNFRSF10B-PVR | 151 | 1266 | 649 |
| TNFRSF10D-PVR | 1321 | 2136 | 1561 | TNFRSF10D-PVR | 1997 | 732 | 1614 |
| TNFRSF12A-PVR | 507 | 93 | 1816 | TNFRSF12A-PVR | 1149 | 1358 | 2417 |
| TNFRSF14-PVR | 983 | 1419 | 409 | TNFRSF14-PVR | 2351 | 2289 | 1577 |
| TNFRSF21-PVR | 485 | 541 | 1910 | TNFRSF21-PVR | 1414 | 969 | 1247 |
| TNFSF10-PVR | 1482 | 317 | 297 | TNFSF10-PVR | 681 | 1150 | 1983 |
| TNFSF15-PVR | 210 | 194 | 56 | TNFSF15-PVR | 635 | 2086 | 1054 |
| Ranking of PVRL2 w.r.t TNF family | Ranking of TNF family w.r.t PVRL2 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| TNF-PVRL2 | 831 | 367 | 526 | TNF-PVRL2 | 2494 | 652 | 966 |
| TNFAIP1-PVRL2 | 867 | 1568 | 1962 | TNFAIP1-PVRL2 | 2244 | 27 | 1932 |
| TNFAIP2-PVRL2 | 2515 | 2423 | 1062 | TNFAIP2-PVRL2 | 2337 | 2483 | 2401 |
| TNFAIP3-PVRL2 | 973 | 595 | 988 | TNFAIP3-PVRL2 | 1334 | 777 | 896 |
| TNFRSF1A-PVRL2 | 742 | 1326 | 1798 | TNFRSF1A-PVRL2 | 1741 | 2355 | 1810 |
| TNFRSF10A-PVRL2 | 830 | 1008 | 478 | TNFRSF10A-PVRL2 | 1027 | 1672 | 3 |
| TNFRSF10B-PVRL2 | 27 | 2160 | 1210 | TNFRSF10B-PVRL2 | 253 | 1794 | 168 |
| TNFRSF10D-PVRL2 | 312 | 1154 | 1229 | TNFRSF10D-PVRL2 | 564 | 1719 | 170 |
| TNFRSF12A-PVRL2 | 1282 | 382 | 1056 | TNFRSF12A-PVRL2 | 594 | 1870 | 1376 |
| TNFRSF14-PVRL2 | 288 | 922 | 264 | TNFRSF14-PVRL2 | 2148 | 1496 | 232 |
| TNFRSF21-PVRL2 | 1590 | 771 | 1034 | TNFRSF21-PVRL2 | 2120 | 734 | 1782 |
| TNFSF10-PVRL2 | 472 | 1160 | 1056 | TNFSF10-PVRL2 | 98 | 695 | 714 |
| TNFSF15-PVRL2 | 373 | 2154 | 420 | TNFSF15-PVRL2 | 768 | 296 | 2448 |
| Ranking of PVRL4 w.r.t TNF family | Ranking of TNF family w.r.t PVRL4 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| TNF-PVRL4 | 540 | 680 | 284 | TNF-PVRL4 | 983 | 2010 | 1345 |
| TNFAIP1-PVRL4 | 1885 | 5 | 131 | TNFAIP1-PVRL4 | 965 | 1896 | 1196 |
| TNFAIP2-PVRL4 | 72 | 2496 | 120 | TNFAIP2-PVRL4 | 2270 | 2429 | 239 |
| TNFAIP3-PVRL4 | 1926 | 599 | 794 | TNFAIP3-PVRL4 | 1442 | 1809 | 1269 |
| TNFRSF1A-PVRL4 | 615 | 1904 | 641 | TNFRSF1A-PVRL4 | 1328 | 2481 | 1316 |
| TNFRSF10A-PVRL4 | 337 | 974 | 1802 | TNFRSF10A-PVRL4 | 1375 | 324 | 1822 |
| TNFRSF10B-PVRL4 | 1762 | 1126 | 483 | TNFRSF10B-PVRL4 | 76 | 40 | 1225 |
| TNFRSF10D-PVRL4 | 2232 | 1056 | 92 | TNFRSF10D-PVRL4 | 1799 | 1327 | 2430 |
| TNFRSF12A-PVRL4 | 601 | 1722 | 1566 | TNFRSF12A-PVRL4 | 2386 | 1670 | 2064 |
| TNFRSF14-PVRL4 | 444 | 1007 | 193 | TNFRSF14-PVRL4 | 1514 | 402 | 447 |
| TNFRSF21-PVRL4 | 12 | 552 | 1875 | TNFRSF21-PVRL4 | 2441 | 1917 | 1689 |
| TNFSF10-PVRL4 | 1149 | 1554 | 341 | TNFSF10-PVRL4 | 150 | 1907 | 743 |
| TNFSF15-PVRL4 | 936 | 1057 | 1160 | TNFSF15-PVRL4 | 2019 | 1452 | 760 |
Table 131.
2nd order combinatorial hypotheses between PVR and TNF family.
| Unexplored combinatorial hypotheses | |
| PVR w.r.t TNF | |
| PVR | TNF, TNF-AIP1/AIP2/RSF1A/RSF10A |
| PVRL2 | TNF-AIP2 |
| TNF w.r.t PVR | |
| TNFRSF14 | PVR |
| TNF-AIP1/AIP2/RSF1A/RSF10B/RSF21 | PVRL2 |
| TNF-AIP2/RSF10D/RSF12A/RSF21 | PVRL4 |
3.9.6. PVR - IL Cross Family Analysis
Fabre-Lafay et al. [252] show that Nectin-4 (PVRL4), is a new serological breast cancer marker and a substrate for tumor necrosis factor-α-converting enzyme (TACE)/ADAM-17. Among the 24 ADAMs, tumor necrosis factor-α-converting enzyme (TACE)/ADAM-17 is involved in various biological processes and cleaves numerous substrates including tumor necrosis factor (TNF)-α, TNF receptor, epidermal growth factor receptor L, c-fms, c-kit, p75NTR, growth hormone receptor, interleukin-6 receptor, interleukin-1 receptor, vascular cell adhesion molecule-1, L-selectin, collagen VII, MUC1, Notch, CX3CL-1, CD40, β-amyloid precursor, and prion protein. Thus there exists an indirect synergy between PVRL4 and interleukin. In CRC cells treated with ETC-1922159, members of PVR family and interleukin family were found up regulated. Our search engine alloted high numerical valued ranks to some of the combinations of both families, thereby indicating possible synergy.
Table 132, shows the rankings of PVR and IL family with respect to each other. On the left we found, PVR to be up regulated w.r.t IL-2RG/8/10RB/15/17C/17REL. These are reflected in rankings of 2007 (laplace) and 2476 (rbf) for IL2RG-PVR, 2429 (laplace) and 2507 (linear) for IL8-PVR, 2310 (laplace) and 2190 (linear) for IL10RB-PVR, 2065 (laplace), 2008 (linear) and 2385 (rbf) for IL15-PVR, 2114 (laplace) and 2301 (linear) for IL17C-PVR and 2317 (linear) and 1971 (rbf) for IL17REL-PVR. On the right side we found, IL-1RAP/6ST/15RA/17REL to be up regulated w.r.t PVR. These are reflected in the rankings of 2194 (linear) and 2026 (rbf), for IL1RAP-PVR; 2434 (linear) and 1767 (rbf) for IL6ST-PVR; 1865 (linear) and 2405 (rbf) for IL15RA-PVR; and 2408 (linear) and 2028 (rbf) for IL17REL-PVR. IL-1A/1B/2RG/6ST was up regulated w.r.t PVRL2. These are reflected in rankings of 2168 (laplace) and 2431 (linear) for IL1A-PVRL2; 2390 (linear) and 2067 (rbf) for IL1B-PVRL2; 1908 (laplace) and 1959 (rbf) for IL2RG-PVRL2; 2349 (laplace) and 2158 (rbf) for IL6ST-PVRL2; 1968 (laplace) and 2084 (rbf) for IL10RB-PVRL2; and 2137 (laplace) and 2004 (rbf) for IL17REL-PVRL2. IL-15RA/17C/17REL was up regulated w.r.t PVRL4. These are reflected in rankings of 2429 (laplace) and 1758 (rbf) for IL15RA-PVRL4; 1782 (laplace) and 2245 (linear) for IL17C-PVRL4 and 1934 (laplace) and 2140 (linear) for IL17REL-PVRL4.
Table 133 shows the derived influences which can be represented graphically, with the following influences - • PVR w.r.t IL with PVR <- IL-2RG/8/10RB/15/17C/17REL; and IL w.r.t PVR with IL-1RAP/6ST/15RA/17REL <- PVR; IL-1A/1B/2RG/6ST <- PVRL2; and IL-15RA/17C/17REL <- PVRL4.
Table 132.
2nd order combinatorial hypotheses between IL and PVR family.
| Ranking PVR vs IL family | |||||||
| Ranking of PVR w.r.t IL family | Ranking of IL family w.r.t PVR | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A-PVR | 295 | 854 | 2060 | IL1A-PVR | 2419 | 822 | 1099 |
| IL1B-PVR | 708 | 1282 | 1092 | IL1B-PVR | 1739 | 405 | 155 |
| IL1RAP-PVR | 980 | 714 | 197 | IL1RAP-PVR | 370 | 2194 | 2026 |
| IL1RN-PVR | 1273 | 25 | 2065 | IL1RN-PVR | 2025 | 266 | 796 |
| IL2RG-PVR | 2007 | 704 | 2476 | IL2RG-PVR | 250 | 740 | 988 |
| IL6ST-PVR | 1125 | 356 | 1368 | IL6ST-PVR | 706 | 2434 | 1767 |
| IL8-PVR | 2429 | 2507 | 257 | IL8-PVR | 92 | 1468 | 812 |
| IL10RB-PVR | 2310 | 2190 | 1140 | IL10RB-PVR | 1362 | 798 | 881 |
| IL15-PVR | 2065 | 2008 | 2385 | IL15-PVR | 1612 | 134 | 658 |
| IL15RA-PVR | 376 | 1621 | 1736 | IL15RA-PVR | 752 | 1865 | 2405 |
| IL17C-PVR | 2114 | 2301 | 1160 | IL17C-PVR | 894 | 26 | 350 |
| IL17REL-PVR | 482 | 2317 | 1971 | IL17REL-PVR | 1010 | 2408 | 2028 |
| Ranking of PVRL2 w.r.t IL family | Ranking of IL family w.r.t PVRL2 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A-PVRL2 | 493 | 849 | 1517 | IL1A-PVRL2 | 2168 | 2431 | 774 |
| IL1B-PVRL2 | 1234 | 180 | 1177 | IL1B-PVRL2 | 1425 | 2390 | 2067 |
| IL1RAP-PVRL2 | 2029 | 1039 | 943 | IL1RAP-PVRL2 | 1393 | 1104 | 670 |
| IL1RN-PVRL2 | 1168 | 295 | 347 | IL1RN-PVRL2 | 959 | 2382 | 481 |
| IL2RG-PVRL2 | 356 | 1752 | 1964 | IL2RG-PVRL2 | 1908 | 990 | 1959 |
| IL6ST-PVRL2 | 243 | 770 | 513 | IL6ST-PVRL2 | 2349 | 384 | 2158 |
| IL8-PVRL2 | 1138 | 641 | 532 | IL8-PVRL2 | 612 | 489 | 1686 |
| IL10RB-PVRL2 | 1746 | 639 | 2502 | IL10RB-PVRL2 | 1968 | 839 | 2084 |
| IL15-PVRL2 | 1107 | 1263 | 759 | IL15-PVRL2 | 1841 | 1579 | 399 |
| IL15RA-PVRL2 | 1216 | 1194 | 330 | IL15RA-PVRL2 | 473 | 811 | 1472 |
| IL17C-PVRL2 | 507 | 920 | 402 | IL17C-PVRL2 | 1167 | 1469 | 43 |
| IL17REL-PVRL2 | 79 | 1306 | 2210 | IL17REL-PVRL2 | 2137 | 962 | 2004 |
| Ranking of PVRL4 w.r.t IL family | Ranking of IL family w.r.t PVRL4 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A-PVRL4 | 338 | 1487 | 666 | IL1A-PVRL4 | 1409 | 1219 | 2410 |
| IL1B-PVRL4 | 1713 | 110 | 481 | IL1B-PVRL4 | 908 | 305 | 399 |
| IL1RAP-PVRL4 | 864 | 269 | 523 | IL1RAP-PVRL4 | 1704 | 2070 | 309 |
| IL1RN-PVRL4 | 2496 | 643 | 144 | IL1RN-PVRL4 | 964 | 1264 | 697 |
| IL2RG-PVRL4 | 858 | 1229 | 2101 | IL2RG-PVRL4 | 18 | 1504 | 727 |
| IL6ST-PVRL4 | 117 | 605 | 744 | IL6ST-PVRL4 | 1954 | 976 | 204 |
| IL8-PVRL4 | 1041 | 1291 | 975 | IL8-PVRL4 | 1441 | 234 | 1826 |
| IL10RB-PVRL4 | 73 | 523 | 278 | IL10RB-PVRL4 | 2137 | 1296 | 506 |
| IL15-PVRL4 | 2062 | 549 | 556 | IL15-PVRL4 | 1029 | 1377 | 281 |
| IL15RA-PVRL4 | 302 | 1519 | 2186 | IL15RA-PVRL4 | 2429 | 1246 | 1758 |
| IL17C-PVRL4 | 25 | 110 | 14 | IL17C-PVRL4 | 1782 | 2245 | 54 |
| IL17REL-PVRL4 | 1487 | 1107 | 1148 | IL17REL-PVRL4 | 1934 | 2140 | 107 |
Table 133.
2nd order combinatorial hypotheses between PVR and IL family.
| Unexplored combinatorial hypotheses | |
| PVR w.r.t IL | |
| PVR | IL-2RG/8/10RB/15/17C/17REL |
| IL w.r.t PVR | |
| IL-1RAP/6ST/15RA/17REL | PVR |
| IL-1A/1B/2RG/6ST | PVRL2 |
| IL-15RA/17C/17REL | PVRL4 |
3.9.7. PVR - Collagen Cross Family Analysis
Fabre-Lafay et al. [252] show that Nectin-4 (PVRL4), is a new serological breast cancer marker and a substrate for tumor necrosis factor-α-converting enzyme (TACE)/ADAM-17. Among the 24 ADAMs, tumor necrosis factor-α-converting enzyme (TACE)/ADAM-17 is involved in various biological processes and cleaves numerous substrates including tumor necrosis factor (TNF)-α, TNF receptor, epidermal growth factor receptor L, c-fms, c-kit, p75NTR, growth hormone receptor, interleukin-6 receptor, interleukin-1 receptor, vascular cell adhesion molecule-1, L-selectin, collagen VII, MUC1, Notch, CX3CL-1, CD40, β-amyloid precursor, and prion protein. Thus there exists an indirect synergy between PVRL4 and collagen. In CRC cells treated with ETC-1922159, members of PVR family and collagen family were found up regulated. Our search engine alloted high numerical valued ranks to some of the combinations of both families, thereby indicating possible synergy.
Table 134, shows the rankings of PVR and COL family with respect to each other. On the left we found, PVR to be up regulated w.r.t COL6A1. This is reflected in rankings of 2259 (laplace) and 2385 (rbf) for COL6A1-PVR. On the right side, we found COL5A3 up regulated w.r.t PVR. This is reflected in rankings of 2341 (laplace) and 2472 (rbf) for COL5A3-PVR. COL9A2 was up regulated w.r.t PVRL2. This is reflected in rankings of 2483 (laplace) and 2363 (rbf) for COL9A2-PVRL2.
Table 135 shows the derived influences which can be represented graphically, with the following influences - • PVR w.r.t COL with PVR <- COL6A1; and • COL w.r.t PVR with COL5A3 <- PVR; and COL9A2 <- PVRL2.
Table 134.
2nd order combinatorial hypotheses between COL and PVR family.
| Ranking PVR vs COL family | |||||||
| Ranking of PVR w.r.t COL family | Ranking of COL family w.r.t PVR | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| COL5A3-PVR | 193 | 790 | 616 | COL5A3-PVR | 2341 | 1172 | 2472 |
| COL6A1-PVR | 2259 | 1740 | 2385 | COL6A1-PVR | 2213 | 70 | 1280 |
| COL7A1-PVR | 1448 | 1166 | 424 | COL7A1-PVR | 144 | 1008 | 1701 |
| COL9A2-PVR | 218 | 166 | 1375 | COL9A2-PVR | 244 | 2501 | 351 |
| COL17A1-PVR | 1800 | 1167 | 1528 | COL17A1-PVR | 1145 | 559 | 685 |
| COL28A1-PVR | 263 | 1273 | 177 | COL28A1-PVR | 1255 | 2266 | 1034 |
| Ranking of PVRL2 w.r.t COL family | Ranking of COL family w.r.t PVRL2 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| COL5A3-PVRL2 | 1275 | 1132 | 515 | COL5A3-PVRL2 | 962 | 1588 | 640 |
| COL6A1-PVRL2 | 533 | 1954 | 826 | COL6A1-PVRL2 | 2372 | 850 | 1193 |
| COL7A1-PVRL2 | 594 | 2111 | 1299 | COL7A1-PVRL2 | 22 | 662 | 2168 |
| COL9A2-PVRL2 | 1336 | 939 | 970 | COL9A2-PVRL2 | 2483 | 1548 | 2363 |
| COL17A1-PVRL2 | 1157 | 1080 | 1232 | COL17A1-PVRL2 | 173 | 1103 | 728 |
| COL28A1-PVRL2 | 991 | 348 | 1618 | COL28A1-PVRL2 | 955 | 864 | 1981 |
| Ranking of PVRL4 w.r.t COL family | Ranking of COL family w.r.t PVRL4 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| COL5A3-PVRL4 | 692 | 1155 | 1446 | COL5A3-PVRL4 | 499 | 405 | 331 |
| COL6A1-PVRL4 | 221 | 1906 | 571 | COL6A1-PVRL4 | 1701 | 2059 | 1315 |
| COL7A1-PVRL4 | 859 | 1320 | 1088 | COL7A1-PVRL4 | 1364 | 2205 | 65 |
| COL9A2-PVRL4 | 1893 | 754 | 1155 | COL9A2-PVRL4 | 1397 | 1797 | 1053 |
| COL17A1-PVRL4 | 1124 | 1647 | 431 | COL17A1-PVRL4 | 1401 | 1174 | 596 |
| COL28A1-PVRL4 | 417 | 1536 | 433 | COL28A1-PVRL4 | 642 | 2446 | 1540 |
Table 135.
2nd order combinatorial hypotheses between PVR and MUC family.
| Unexplored combinatorial hypotheses | |
| PVR w.r.t COL | |
| PVR | COL6A1 |
| COL w.r.t PVR | |
| COL5A3 | PVR |
| COL9A2 | PVRL2 |
3.9.8. PVR - MUCIN Cross Family Analysis
[252] show that Nectin-4 (PVRL4), is a new serological breast cancer marker and a substrate for tumor necrosis factor-α-converting enzyme (TACE)/ADAM-17. Among the 24 ADAMs, tumor necrosis factor-α-converting enzyme (TACE)/ADAM-17 is involved in various biological processes and cleaves numerous substrates including tumor necrosis factor (TNF)-α, TNF receptor, epidermal growth factor receptor L, c-fms, c-kit, p75NTR, growth hormone receptor, interleukin-6 receptor, interleukin-1 receptor, vascular cell adhesion molecule-1, L-selectin, collagen VII, MUC1, Notch, CX3CL-1, CD40, β-amyloid precursor, and prion protein. Thus there exists an indirect synergy between PVRL4 and Mucin. In CRC cells treated with ETC-1922159, members of PVR family and Mucin family were found up regulated. Our search engine alloted high numerical valued ranks to some of the combinations of both families, thereby indicating possible synergy.
Table 136, shows the rankings of PVR and MUC family with respect to each other. On the left we found, PVR to be up regulated w.r.t 1772 (laplace) and 2085 (rbf) for MUC20-PVR. PVRL2 was up regulated w.r.t MUC17. This is reflected in rankings of 2098 (linear) and 1869 (rbf) for MUC17-PVRL2. PVRL4 was up regulated w.r.t MUC13. This is reflected in rankings of 2160 (laplace) and 1937 (rbf) for MUC13-PVRL4. On the right side, MUC-1/3A were found up regulated w.r.t PVRL4. These are reflected in rankings of 2272 (laplace) and 1827 (linear) for MUC1-PVRL4; and 2103 (linear) and 1835 (rbf) for MUC3A-PVRL4.
Table 137 shows the derived influences which can be represented graphically, with the following influences - • PVR w.r.t MUC with PVR <- MUC20; PVRL2 <- MUC17 and PVRL4 <- MUC13 • MUC w.r.t PVR with MUC1 <- PVRL4; and MUC3A <- PVRL4.
Table 136.
2nd order combinatorial hypotheses between MUC and PVR family.
| Ranking PVR vs MUC family | |||||||
| Ranking of PVR w.r.t MUC family | Ranking of MUC family w.r.t PVR | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| MUC1-PVR | 2173 | 1184 | 1021 | MUC1-PVR | 2106 | 377 | 426 |
| MUC3A-PVR | 64 | 130 | 1828 | MUC3A-PVR | 136 | 1217 | 1004 |
| MUC4-PVR | 2000 | 804 | 1026 | MUC4-PVR | 2494 | 366 | 586 |
| MUC12-PVR | 1449 | 1589 | 1899 | MUC12-PVR | 2370 | 224 | 18 |
| MUC13-PVR | 1701 | 1292 | 1226 | MUC13-PVR | 1230 | 144 | 59 |
| MUC17-PVR | 209 | 684 | 881 | MUC17-PVR | 2388 | 320 | 1048 |
| MUC20-PVR | 1772 | 1242 | 2085 | MUC20-PVR | 1380 | 188 | 1039 |
| Ranking of PVRL2 w.r.t MUC family | Ranking of MUC family w.r.t PVRL2 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| MUC1-PVRL2 | 828 | 454 | 2362 | MUC1-PVRL2 | 1690 | 923 | 1060 |
| MUC3A-PVRL2 | 584 | 708 | 2332 | MUC3A-PVRL2 | 2031 | 164 | 546 |
| MUC4-PVRL2 | 91 | 1214 | 478 | MUC4-PVRL2 | 1315 | 1150 | 230 |
| MUC12-PVRL2 | 1129 | 1655 | 1439 | MUC12-PVRL2 | 1820 | 1417 | 94 |
| MUC13-PVRL2 | 1052 | 179 | 329 | MUC13-PVRL2 | 471 | 1760 | 240 |
| MUC17-PVRL2 | 328 | 2098 | 1869 | MUC17-PVRL2 | 873 | 2111 | 1462 |
| MUC20-PVRL2 | 1407 | 1350 | 1612 | MUC20-PVRL2 | 1348 | 325 | 133 |
| Ranking of PVRL4 w.r.t MUC family | Ranking of MUC family w.r.t PVRL4 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| MUC1-PVRL4 | 569 | 99 | 155 | MUC1-PVRL4 | 2272 | 1827 | 1457 |
| MUC3A-PVRL4 | 1088 | 1214 | 2498 | MUC3A-PVRL4 | 1698 | 2103 | 1835 |
| MUC4-PVRL4 | 18 | 13 | 714 | MUC4-PVRL4 | 1518 | 1251 | 789 |
| MUC12-PVRL4 | 526 | 449 | 1420 | MUC12-PVRL4 | 311 | 1059 | 2065 |
| MUC13-PVRL4 | 2160 | 1404 | 1937 | MUC13-PVRL4 | 1540 | 45 | 1570 |
| MUC17-PVRL4 | 731 | 221 | 234 | MUC17-PVRL4 | 1736 | 2206 | 219 |
| MUC20-PVRL4 | 22 | 1820 | 361 | MUC20-PVRL4 | 626 | 722 | 2232 |
Table 137.
2nd order combinatorial hypotheses between PVR and MUC family.
| Unexplored combinatorial hypotheses | |
| PVR w.r.t MUC | |
| PVR | MUC20 |
| PVRL2 | MUC17 |
| PVRL4 | MUC13 |
| MUC w.r.t PVR | |
| MUC1 | PVRL4 |
| MUC3A | PVRL4 |
3.10. Anthrax Toxin Receptor Related Synergies
3.10.1. ANTXR2 - Collagen Cross Family Analysis
Anthrax toxin receptor ANTRX is known to capture the Bacillus anthracis toxin and form the cause of the anthrax disease. Regulatory mechanism of the ANTXR1 has been demostrated essential component in the fibrosis processes in fibroproliferative diseases. Loss of ANTXR1 (a.k.a TEM8) in fibroblasts leads to increased rates of synthesis of fiber-forming collagens, resulting in progressive fibrosis in skin and other organs Besschetnova et al. [253]. TEM8 interacts with the cleaved C5 domain of collagen 3 (VI) Nanda et al. [254]. Hotchkiss et al. [255] also indicate the interaction of TEM8 and collagens. Bell et al. [256] indicate that a recombinant portion of ANTXR2 (a.k.a CMG2) was found to bind collagen type IV and laminin, suggesting a potential role in basement membrane matrix synthesis and assembly. Bürgi et al. [257] show that CMG2/ANTXR2 regulates extracellular collagen VI which accumulates in hyaline fibromatosis syndrome. A distinctive early childhood-onset disorder, systemic hyalinosis, is characterized by mutations in the anthrax toxin receptor 2 gene (ANTRX2) as shown by Shieh et al. [258]. Not much is known about the behaviour of ANTRX2 with collagens in colorectal cancer. In CRC cells treated with ETC-1922159 these were up regulated. Our search engine was able to rank the 2nd order combinations between these two to see if there is a possible existing synergy based on the already explored pathological functionality in the above works. At in silico level we found possible high numerical valued ranks pointed to some of the combinations of ANTXR2 and collagen family.
On the left side of Table 138 we found ANTXR2 to be up regulated w.r.t COL-5A3/28A1 (probably COL-7A1 also). These are reflected in rankings of 2006 (laplace) and 2217 (rbf) for COL5A3-ANTXR2; 2259 (linear) and 2296 (rbf) for COL28A1-ANTXR2 and probably 2119 (linear) and 1690 (rbf) for COL7A1-ANTXR2. On the right side, we found, COL-7A1/28A1 up regulated w.r.t ANTXR2. These are reflected in rankings of 1722 (laplace) and 2121 (rbf) for COL7A1-ANTXR2 and 1731 (linear) and 2362 (rbf) for COL28A1-ANTXR2. Table 139 shows the derived influences which can be represented graphically, with the following influences - • ANTXR2 w.r.t COL with COL5A3 -> ANTXR2 and COL28A1 -> ANTXR2; and • COL w.r.t ANTXR2 with COL7A1 <- ANTXR2 and COL28A1 <- ANTXR2.
3.10.2. ANTXR2 - Integrin Cross Family Analysis
To define whether there is a possible functional cooperation between TEM8/ANTXR1 and integrins to accomplish cell spreading, Werner et al. [259] sought to disrupt the function of collagen binding integrins, which mostly belong to β1 family of integrins, with β1 integrin-blocking antibodies. Their experiments suggest that the participation of β1 integrins can be excluded in TEM8-mediated cell spreading on collagen in primary fibroblasts. However, collectively, they indicate that the participation of these integrins in TEM8 spreading on collagen vary with cell type. The cytoplasmic domain of ANTXR1 affects binding of the protective antigen which is similar to integrin I domains that convert between open and closed conformations that bind ligand with high and low affinities, respectively. These findings are observed by Go et al. [260]. Scobie et al. [261] observe similar behaviour. Abnormal clustering of TEM8/ANTXR1 with integrin β1 and vascular endothelial growth factor receptor 2 (VEGFR2) occurs in endothelial cells within cutaneous infantile hemangiomas, the most common vascular anomaly in childhood Besschetnova et al. [253]. These findings suggest the possibility to synergy between ANTXR2 and Integrin family. In CRC cells treated with ETC-1922159, ANTXR2 and integrin families were up regulated. The search engine alloted high numerical valued ranks to some of the 2nd order combinations of ANTXR2 and integrin family members thus pointing to possible synergy in CRC cells. Table 140 shows the rankings of ANTRX2 along with integrin family members.
On the left side, we found ANTRX2 to be up regulated w.r.t ITG-A2/A3/B1BP1/B5/B8. These are reflected in the rankings of 2261 (laplace) and 2444 (rbf) for ITGA2-ANTXR2; 2027 (laplace) and 2134 (linear) for ITGA3-ANTXR2; 2444 (laplace) and 2128 (rbf) for ITGB1BP1-ANTXR2; 1860 (linear) and 2315 (rbf) for ITGB5-ANTXR2; and 2354 (linear)and 2136 (rbf) for ITGB8-ANTXR2. On the right side, we found ITGB6 to be up regulated w.r.t ANTRX2. These are reflected in the rankings of 2000 (linear) and 1896 (rbf) for ITGB6-ANTXR2; Table 141 shows the derived influences which can be represented graphically, with the following influences - • ANTXR2 w.r.t ITG with ITG-A2/A3 -> ANTXR2 and ITG-B1BP1/B5/B8 -> ANTXR2; and • ITG w.r.t ANTXR2 with ITGB6 <- ANTXR2.
Table 140.
2nd order combinatorial hypotheses between ANTRX2 and ITG family.
| Ranking ANTRX2 vs ITG family | |||||||
| Ranking of ANTRX2 w.r.t ITG family | Ranking of ITG family w.r.t ANTXR2 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| ITGA2-ANTXR2 | 2261 | 1129 | 2444 | ITGA2-ANTXR2 | 657 | 1662 | 215 |
| ITGA3-ANTXR2 | 2027 | 2134 | 179 | ITGA3-ANTXR2 | 305 | 1402 | 278 |
| ITGA6-ANTXR2 | 1065 | 1850 | 1660 | ITGA6-ANTXR2 | 352 | 2029 | 583 |
| ITGB1-ANTXR2 | 2192 | 1273 | 1431 | ITGB1-ANTXR2 | 1538 | 987 | 1593 |
| ITGB1BP1-ANTXR2 | 2444 | 498 | 2128 | ITGB1BP1-ANTXR2 | 1743 | 152 | 534 |
| ITGB4-ANTXR2 | 1484 | 699 | 249 | ITGB4-ANTXR2 | 123 | 1420 | 2116 |
| ITGB5-ANTXR2 | 1318 | 1860 | 2315 | ITGB5-ANTXR2 | 2216 | 718 | 1182 |
| ITGB6-ANTXR2 | 1205 | 1262 | 1244 | ITGB6-ANTXR2 | 1200 | 2000 | 1896 |
| ITGB8-ANTXR2 | 1710 | 2354 | 2136 | ITGB8-ANTXR2 | 296 | 1724 | 1485 |
Table 141.
2nd order combinatorial hypotheses between ANTRX2 and ITG family.
| Unexplored combinatorial hypotheses | |
| ANTXR2 w.r.t ITG | |
| ANTXR2 | ITG-A2/A3 |
| ANTXR2 | ITG-B1BP1/B5/B8 |
| ITG w.r.t ANTXR2 | |
| ITGB6 | ANTXR2 |
3.10.3. ANTXR2 - MMP Cross Family Analysis
Compromised interactions between TEM8/ANTXR1-deficient endothelial and fibroblastic cells cause dramatic reduction in the activity of the matrix-degrading enzyme MMP2 Besschetnova et al. [253]. They observe experimentally that loss of MMP2 activity requires loss of TEM8/ANTXR1 function in both endothelial and fibroblastic cells. Matrix metalloproteinases (MMP) are members of the metzincin group of proteases which share the conserved zinc-binding motif in their catalytic active site Löffek et al. [262]. these enzymes are capable of degrading all kinds of extracellular matrix proteins, but also can process a number of bioactive molecules as well as play a major role in cell behaviors such as proliferation, migration, differention, apoptosis and host defense, Wikipedia contributors [263]. In CRC cells treated with ETC-1922159, ANTXR2 and integrin families were up regulated. The search engine alloted high numerical valued ranks to some of the 2nd order combinations of ANTXR2 and integrin family members thus pointing to possible synergy in CRC cells. Table 142 shows the rankings of ANTRX2 along with MMP family members.
On the left side, we found ANTXR2 to be upregulated w.r.t MMP28. This is reflected in rankings of 2468 (laplace) and 1765 (linear) for MMP28-ANTXR2. On the right side we found MMP-1/15/28 up regulated w.r.t ANTXR2. These are reflected in rankings of 2009 (linear) and 2142 (rbf) for MMP1-ANTXR2; 2219 (linear) and 1926 (rbf) for MMP15-ANTXR2; and 1857 (linear) and 2092 (rbf) for MMP28-ANTXR2. Table 143 shows the derived influences which can be represented graphically, with the following influences - • ANTXR2 w.r.t MMP with ANTXR2 <- MMP28 and • MMP w.r.t ANTXR2 with MMP-1/15/28 <- ANTXR2.
Table 142.
2nd order combinatorial hypotheses between ANTRX2 and MMP family.
| Ranking ANTRX2 vs MMP family | |||||||
| Ranking of ANTRX2 w.r.t MMP family | Ranking of MMP family w.r.t ANTXR2 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| MMP1-ANTXR2 | 1428 | 1407 | 1620 | MMP1-ANTXR2 | 244 | 2009 | 2142 |
| MMP14-ANTXR2 | 1067 | 1141 | 900 | MMP14-ANTXR2 | 866 | 971 | 443 |
| MMP15-ANTXR2 | 1457 | 740 | 1881 | MMP15-ANTXR2 | 121 | 2219 | 1926 |
| MMP28-ANTXR2 | 2468 | 1765 | 1202 | MMP28-ANTXR2 | 11 | 1857 | 2092 |
Table 143.
2nd order combinatorial hypotheses between ANTRX2 and MMP family.
| Unexplored combinatorial hypotheses | |
| ANTXR2 w.r.t MMP | |
| ANTXR2 | MMP28 |
| MMP w.r.t ANTXR2 | |
| MMP-1/15/28 | ANTXR2 |
3.10.4. ANTXR2 - WNT Cross Family Analysis
Abrami et al. [264] show that LRP6 can indeed form a complex with ATRs (anthrax toxin receptors), and that this interaction plays a role both in Wnt signalling and in anthrax toxin endocytosis. Through the ATR-LRP6 interaction, adhesion to the extracellular matrix could locally control Wnt signalling. The authors demonstrated that physical and functional interaction between CMG2/ANTXR2 and LRP6 also raised the possibility that the complex clinical manifestation of Systemic Hyalinosis might be due in part to defects in Wnt signalling. Fluorescence microscopy and biochemical analyses showed that LRP6 enables toxin internalization by interacting at the cell surface with PA receptors TEM8/ATR and/or CMG2/ANTXR2 to form a multi-component complex that enters cells upon PA binding (Wei et al. [265]). Verma et al. [266] postulate that the developmentally controlled expression of TEM8 modulates endothelial cell response to canonical Wnt signaling to regulate vessel patterning and density. These findings definitely indicate the synergy of ANTRX with Wnts. In CRC cells treated with ETC-1922159, ANTXR2 and WNT families were up regulated. The search engine alloted high numerical valued ranks to some of the 2nd order combinations of ANTXR2 and WNT family members thus pointing to possible synergy in CRC cells. Table 144 shows the rankings of ANTRX2 along with WNT family members.
On the left side, we found ANTXR2 to be upregulated w.r.t WNT4. This is reflected in rankings of 1833 (linear) and 2341 (rbf) for WNT4-ANTXR2. Table 145 shows the derived influences which can be represented graphically, with the following influences - • ANTXR2 w.r.t WNT with ANTXR2 <- WNT4. This synergistic upregulation of the WNT4 with ANTXR2 might indicate possible control over the signalling in CRC cells treated with ETC-1922159.
Table 144.
2nd order combinatorial hypotheses between ANTRX2 and WNT family.
| Ranking ANTRX2 vs WNT family | |||||||
| Ranking of ANTRX2 w.r.t WNT family | Ranking of WNT family w.r.t ANTXR2 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| WNT2B-ANTXR2 | 1160 | 1013 | 2286 | WNT2B-ANTXR2 | 1577 | 1367 | 944 |
| WNT4-ANTXR2 | 1735 | 1833 | 2341 | WNT4-ANTXR2 | 175 | 1643 | 97 |
| WNT7B-ANTXR2 | 2453 | 304 | 1196 | WNT7B-ANTXR2 | 2106 | 242 | 1144 |
| WNT9A-ANTXR2 | 1618 | 487 | 1766 | WNT9A-ANTXR2 | 2317 | 162 | 845 |
Table 145.
2nd order combinatorial hypotheses between ANTRX2 and WNT family.
| Unexplored combinatorial hypotheses | |
| ANTXR2 w.r.t WNT | |
| ANTXR2 | WNT4 |
3.10.5. ANTXR2 - TNF Cross Family Analysis
The author could not find much about TNF-ANTXR2 combinations in pathological cases in existing literature, however, Lee et al. [267] report the "both LeTx and EdTx markedly inhibited LPS-induced transcription of tumour necrosis factor alpha (TNF-α), interleukin (IL)-1β, and IL-6 in J774A.1 cells. In contrast, EdTx synergised with LPS to increase the transcription of IL-6 and IL-8 in HAECs. We showed that HAECs are suitable for anthrax toxin research and express higher levels of the two anthrax toxin receptors - tumour endothelial marker 8 (TEM8/ANTXR1) and capillary morphogenesis protein 2 (CMG2/ANTXR2) - than do J774A.1 cells". The high expression of the ANTXR-1/2 is shown, however, the possible synergy between ANTXR and TNFs in not shown. Our search engine pointed to some of the combinations in CRC cells treated with ETC-1922159 treatment. In Table 146, on the left we found ANTXR2 to be up regulated w.r.t TNF-AIP1/RSF10B. These are reflected in rankings of 1769 (linear) and 1946 (rbf) for TNFAIP1-ANTXR2 and 2278 (linear) and 2218 (rbf) for TNFRSF10B-ANTXR2. On the right we found, TNF-RSF10A/RSF10D/RSF12A/RSF14 was up regulated w.r.t ANTXR2. These are reflected in rankings of 2260 (laplace) and 2377 (rbf) for TNFRSF10A-ANTXR2, 2258 (laplace) and 2363 (rbf) for TNFRSF10D-ANTXR2, 2190 (laplace) and 2061 (rbf) for TNFRSF12A-ANTXR2 and 2370(laplace) and 1777 (linear) for TNFRSF14-ANTXR2.
Table 147 shows the derived influences which can be represented graphically, with the following influences - • ANTXR2 w.r.t TNF with ANTXR2 <- TNFAIP1 and ANTXR2 <- TNFRSF10B and • TNF w.r.t ANTXR2 with TNFRSF10A <- ANTXR2; TNFRSF10D <- ANTXR2; TNFRSF12A <- ANTXR2 and TNFRSF14 <- ANTXR2. This synergistic upregulation of the TNF with ANTXR2 might indicate possible control over the signalling in CRC cells treated with ETC-1922159.
Table 146.
2nd order combinatorial hypotheses between ANTRX2 and TNF family.
| Ranking ANTRX2 vs TNF family | |||||||
| Ranking of ANTRX2 w.r.t TNF family | Ranking of TNF family w.r.t ANTXR2 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| TNF-ANTXR2 | 1439 | 1568 | 1285 | TNF-ANTXR2 | 709 | 1758 | 1479 |
| TNFAIP1-ANTXR2 | 1552 | 1769 | 1946 | TNFAIP1-ANTXR2 | 1252 | 2177 | 218 |
| TNFAIP2-ANTXR2 | 125 | 962 | 2134 | TNFAIP2-ANTXR2 | 659 | 1156 | 2109 |
| TNFAIP3-ANTXR2 | 1184 | 1253 | 1558 | TNFAIP3-ANTXR2 | 1429 | 2485 | 1731 |
| TNFRSF1A-ANTXR2 | 1063 | 310 | 2145 | TNFRSF1A-ANTXR2 | 1557 | 2471 | 935 |
| TNFRSF10A-ANTXR2 | 351 | 1358 | 1280 | TNFRSF10A-ANTXR2 | 2260 | 32 | 2377 |
| TNFRSF10B-ANTXR2 | 2278 | 2218 | 982 | TNFRSF10B-ANTXR2 | 852 | 715 | 216 |
| TNFRSF10D-ANTXR2 | 1352 | 891 | 1685 | TNFRSF10D-ANTXR2 | 2258 | 454 | 2363 |
| TNFRSF12A-ANTXR2 | 551 | 1283 | 1794 | TNFRSF12A-ANTXR2 | 2190 | 1150 | 2061 |
| TNFRSF14-ANTXR2 | 999 | 442 | 498 | TNFRSF14-ANTXR2 | 2370 | 1777 | 1014 |
| TNFRSF21-ANTXR2 | 897 | 997 | 298 | TNFRSF21-ANTXR2 | 1474 | 343 | 510 |
| TNFRSF10-ANTXR2 | 2151 | 966 | 324 | TNFRSF10-ANTXR2 | 2065 | 112 | 339 |
| TNFRSF15-ANTXR2 | 868 | 967 | 1590 | TNFRSF15-ANTXR2 | 664 | 1211 | 1669 |
Table 147.
2nd order combinatorial hypotheses between ANTRX2 and TNF family.
| Unexplored combinatorial hypotheses | |
| ANTXR2 w.r.t TNF | |
| ANTXR2 | TNFAIP1 |
| ANTXR2 | TNFRSF10B |
| TNF w.r.t ANTXR2 | |
| TNFRSF10A | ANTXR2 |
| TNFRSF10D | ANTXR2 |
| TNFRSF12A | ANTXR2 |
| TNFRSF14 | ANTXR2 |
3.10.6. ANTXR2 - IL Cross Family Analysis
The author could not find much about TNF-ANTXR2 combinations in pathological cases in existing literature, however, Lee et al. [267] report the "both LeTx and EdTx markedly inhibited LPS-induced transcription of tumour necrosis factor alpha (TNF-α), interleukin (IL)-1β, and IL-6 in J774A.1 cells. In contrast, EdTx synergised with LPS to increase the transcription of IL-6 and IL-8 in HAECs. We showed that HAECs are suitable for anthrax toxin research and express higher levels of the two anthrax toxin receptors - tumour endothelial marker 8 (TEM8/ANTXR1) and capillary morphogenesis protein 2 (CMG2/ANTXR2) – than do J774A.1 cells". The high expression of the ANTXR-1/2 is shown, however, the possible synergy between ANTXR and IL in not shown. Our search engine pointed to some of the combinations in CRC cells treated with ETC-1922159 treatment. In Table 148, on the left we found ANTXR2 to be up regulated w.r.t IL-1RN/6ST/17C/17REL. These are reflected in rankings of 1914 (linear) and 1894 (rbf) for IL1RN-ANTXR2; 1944 (laplace), 2219 (linear) and 1914 (rbf) for IL6ST-ANTXR2; 1832 (laplace) and 2334 (linear) for IL17C-ANTXR2 and 1889 (linear) and 2303 (rbf) for IL17REL-ANTXR2. On the right we found, IL-1A/1B/6ST/17C was up regulated w.r.t ANTXR2. These are reflected in rankings of 2356 (linear) and 1859 (rbf) for IL1A-ANTXR2; 1780 (linear) and 1865 (rbf) for IL6ST-ANTXR2; 1924 (laplace) and 1901 (rbf) for IL15RA-ANTXR2; and 2121 (linear) and 2437 (rbf) for IL17C-ANTXR2.
Table 149 shows the derived influences which can be represented graphically, with the following influences - • ANTXR2 w.r.t IL with ANTXR2 <- IL1RN; ANTXR2 <- IL6ST; ANTXR2 <- IL17C and ANTXR2 <- IL17REL; and • IL w.r.t ANTXR2 with IL1A <- ANTXR2; IL1B <- ANTXR2; IL6ST <- ANTXR2; and IL17C <- ANTXR2.
Table 148.
2nd order combinatorial hypotheses between ANTRX2 and IL family.
| Ranking ANTRX2 vs IL family | |||||||
| Ranking of ANTRX2 w.r.t IL family | Ranking of IL family w.r.t ANTXR2 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A-ANTXR2 | 1733 | 454 | 2253 | IL1A-ANTXR2 | 275 | 2356 | 1859 |
| IL1B-ANTXR2 | 1222 | 1302 | 714 | IL1B-ANTXR2 | 330 | 2011 | 1762 |
| IL1RAP-ANTXR2 | 1288 | 367 | 80 | IL1RAP-ANTXR2 | 2339 | 442 | 747 |
| IL1RN-ANTXR2 | 1389 | 1914 | 1894 | IL1RN-ANTXR2 | 349 | 1031 | 1919 |
| IL2RG-ANTXR2 | 1897 | 25 | 432 | IL2RG-ANTXR2 | 368 | 1867 | 450 |
| IL6ST-ANTXR2 | 1944 | 2219 | 1914 | IL6ST-ANTXR2 | 46 | 1780 | 1865 |
| IL8-ANTXR2 | 1169 | 1281 | 1398 | IL8-ANTXR2 | 1343 | 2002 | 434 |
| IL10RB-ANTXR2 | 1737 | 496 | 1545 | IL10RB-ANTXR2 | 1403 | 800 | 754 |
| IL15-ANTXR2 | 787 | 1812 | 927 | IL15-ANTXR2 | 1002 | 1340 | 481 |
| IL15RA-ANTXR2 | 840 | 800 | 1695 | IL15RA-ANTXR2 | 1924 | 636 | 1901 |
| IL17C-ANTXR2 | 1832 | 2334 | 1191 | IL17C-ANTXR2 | 339 | 2121 | 2437 |
| IL17REL-ANTXR2 | 29 | 1889 | 2303 | IL17REL-ANTXR2 | 2406 | 111 | 960 |
Table 149.
2nd order combinatorial hypotheses between ANTRX2 and IL family.
| Unexplored combinatorial hypotheses | |
| ANTXR2 w.r.t IL | |
| ANTXR2 | IL1RN |
| ANTXR2 | IL6ST |
| ANTXR2 | IL17C |
| ANTXR2 | IL17REL |
| IL w.r.t ANTXR2 | |
| IL1A | ANTXR2 |
| IL1B | ANTXR2 |
| IL6ST | ANTXR2 |
| IL17C | ANTXR2 |
Conclusion
We present here a range of multiple synergistic 2nd combinations that were ranked via a search engine and later conduct two-cross family analysis between components of these combinations. Via majority voting across the ranking methods, we were able to find plausible unexplored synergistic combinations that might be prevalent in CRC cells after treatment with ETC-1922159 drug. The two-way cross family analysis also assists in deriving influences between components which serve as hypotheses for further tests. In short, we are now able to locate possible synergies via this ranking search engine and two-way cross family analysis for 2nd order combinations in CRC cells treated with ETC-1922159. Further wet lab tests on these combinations for verification is needed. Also, if found true, it paves way for biologists/oncologists to further investigate and understand the mechanism behind the synergy through wet experiments.
Author Contributions
Concept, design, in silico implementation - SS. Analysis and interpretation of results - SS. Manuscript writing - SS. Manuscript revision - SS. Approval of manuscript - SS
Data Availability Statement
Data used in this research work was released in a publication in Madan et al. [268]. The ETC-1922159 was released in Singapore in July 2015 under the flagship of the Agency for Science, Technology and Research (A*STAR) and Duke-National University of Singapore Graduate Medical School (Duke-NUS).
Acknowledgments
Special thanks to Mrs. Rita Sinha and Mr. Prabhat Sinha for supporting the author financially, without which this work could not have been made possible. Marco Wiering and Silja Renooij for continued support during the years of independent research work.
Conflicts of Interest
There are no conflicts to declare.
Appendix A
Appendix A.1. Choice of Sensitivity Indices
The sensitivity package (Faivre et al. [269] and Iooss and Lemaıtre [270]) in R langauge provides a range of functions to compute the indices and the following indices will be taken into account for addressing the posed questions in this manuscript.
- sensiFdiv - conducts a density-based sensitivity analysis where the impact of an input variable is defined in terms of dissimilarity between the original output density function and the output density function when the input variable is fixed. The dissimilarity between density functions is measured with Csiszar f-divergences. Estimation is performed through kernel density estimation and the function kde of the package ks. Borgonovo [271] and Da Veiga [272]
- sensiHSIC - conducts a sensitivity analysis where the impact of an input variable is defined in terms of the distance between the input/output joint probability distribution and the product of their marginals when they are embedded in a Reproducing Kernel Hilbert Space (RKHS). This distance corresponds to HSIC proposed by Gretton et al. [273] and serves as a dependence measure between random variables.
- sobol2002 - implements the Monte Carlo estimation of the Sobol indices for both first-order and total indices at the same time (all together 2p indices), at a total cost of (p+2) × n model evaluations. These are called the Saltelli estimators. This estimator suffers from a conditioning problem when estimating the variances behind the indices computations. This can seriously affect the Sobol indices estimates in case of largely non-centered output. To avoid this effect, you have to center the model output before applying "sobol2002". Functions ”soboljansen" and "sobolmartinez" do not suffer from this problem. Saltelli [276]
- sobol2007 - implements the Monte Carlo estimation of the Sobol indices for both first-order and total indices at the same time (all together 2p indices), at a total cost of (p+2) × n model evaluations. These are called the Mauntz estimators. Saltelli and Annoni [277]
- sobolmartinez - implements the Monte Carlo estimation of the Sobol indices for both first-order and total indices using correlation coefficients-based formulas, at a total cost of (p + 2) × n model evaluations. These are called the Martinez estimators.
- sobol - implements the Monte Carlo estimation of the Sobol sensitivity indices. Allows the estimation of the indices of the variance decomposition up to a given order, at a total cost of (N + 1) × n where N is the number of indices to estimate. Sobol’ [278]
References
- Brancati, F.; Fortugno, P.; Bottillo, I.; Lopez, M.; Josselin, E.; Boudghene-Stambouli, O.; Agolini, E.; Bernardini, L.; Bellacchio, E.; Iannicelli, M.; et al. Mutations in PVRL4, encoding cell adhesion molecule nectin-4, cause ectodermal dysplasia-syndactyly syndrome. The American Journal of Human Genetics 2010, 87, 265–273. [Google Scholar] [CrossRef]
- Sinha, S. Inchoative Discovery of Plausible (Un)explored Synergistic Combinatorial Biological Hypotheses for Static/Time Series Wnt Measurements via Ranking Search Engine: BioSearch Engine Design. Preprints 2018. [Google Scholar]
- Sinha, S. Sensitivity analysis based ranking reveals unknown biological hypotheses for down regulated genes in time buffer during administration of PORCN-WNT inhibitor ETC-1922159 in CRC. bioRxiv 2017, 180927. [Google Scholar]
- Sinha, S. Prioritizing 2nd order interactions via support vector ranking using sensitivity indices on time series Wnt measurements. bioRxiv 2017, 060228. [Google Scholar]
- Sharma, R. Wingless a new mutant in Drosophila melanogaster. Drosophila information service 1973, 50, 134–134. [Google Scholar]
- Thorstensen, L.; Lind, G.E.; Løvig, T.; Diep, C.B.; Meling, G.I.; Rognum, T.O.; Lothe, R.A. Genetic and epigenetic changes of components affecting the WNT pathway in colorectal carcinomas stratified by microsatellite instability. Neoplasia 2005, 7, 99–108. [Google Scholar] [CrossRef]
- Baron, R.; Kneissel, M. WNT signaling in bone homeostasis and disease: from human mutations to treatments. Nature medicine 2013, 19, 179–192. [Google Scholar] [CrossRef]
- Clevers, H. Wnt/[β]-catenin signaling in development and disease. Cell 2006, 127, 469–480. [Google Scholar] [CrossRef]
- Sokol, S. Wnt Signaling in Embryonic Development; Elsevier, 2011; Volume 17. [Google Scholar]
- Pinto, D.; Gregorieff, A.; Begthel, H.; Clevers, H. Canonical Wnt signals are essential for homeostasis of the intestinal epithelium. Genes & development 2003, 17, 1709–1713. [Google Scholar]
- Zhong, Z.; Ethen, N.J.; Williams, B.O. WNT signaling in bone development and homeostasis. Wiley Interdisciplinary Reviews: Developmental Biology 2014, 3, 489–500. [Google Scholar] [CrossRef]
- Pećina-Šlaus, N. Wnt signal transduction pathway and apoptosis: a review. Cancer Cell International 2010, 10, 1–5. [Google Scholar] [CrossRef]
- Kahn, M. Can we safely target the WNT pathway? Nature Reviews Drug Discovery 2014, 13, 513–532. [Google Scholar] [CrossRef] [PubMed]
- Garber, K. Drugging the Wnt pathway: problems and progress. Journal of the National Cancer Institute 2009, 101, 548–550. [Google Scholar] [CrossRef]
- Voronkov, A.; Krauss, S. Wnt/beta-catenin signaling and small molecule inhibitors. Current pharmaceutical design 2012, 19, 634. [Google Scholar] [CrossRef]
- Blagodatski, A.; Poteryaev, D.; Katanaev, V. Targeting the Wnt pathways for therapies. Mol Cell Ther 2014, 2, 28. [Google Scholar] [CrossRef]
- Curtin, J.C.; Lorenzi, M.V. Drug discovery approaches to target Wnt signaling in cancer stem cells. Oncotarget 2010, 1, 552. [Google Scholar] [CrossRef]
- Tanaka, K.; Okabayashi, K.; Asashima, M.; Perrimon, N.; Kadowaki, T. The evolutionarily conserved porcupine gene family is involved in the processing of the Wnt family. The FEBS Journal 2000, 267, 4300–4311. [Google Scholar] [CrossRef]
- Bänziger, C.; Soldini, D.; Schütt, C.; Zipperlen, P.; Hausmann, G.; Basler, K. Wntless, a conserved membrane protein dedicated to the secretion of Wnt proteins from signaling cells. Cell 2006, 125, 509–522. [Google Scholar] [CrossRef]
- Bartscherer, K.; Pelte, N.; Ingelfinger, D.; Boutros, M. Secretion of Wnt ligands requires Evi, a conserved transmembrane protein. Cell 2006, 125, 523–533. [Google Scholar] [CrossRef]
- Kurayoshi, M.; Yamamoto, H.; Izumi, S.; Kikuchi, A. Post-translational palmitoylation and glycosylation of Wnt-5a are necessary for its signalling. Biochemical Journal 2007, 402, 515–523. [Google Scholar] [CrossRef]
- Gao, X.; Hannoush, R.N. Single-cell imaging of Wnt palmitoylation by the acyltransferase porcupine. Nature chemical biology 2014, 10, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Dodge, M.E.; Tang, W.; Lu, J.; Ma, Z.; Fan, C.W.; Wei, S.; Hao, W.; Kilgore, J.; Williams, N.S.; et al. Small molecule–mediated disruption of Wnt-dependent signaling in tissue regeneration and cancer. Nature chemical biology 2009, 5, 100–107. [Google Scholar] [CrossRef]
- Wang, X.; Moon, J.; Dodge, M.E.; Pan, X.; Zhang, L.; Hanson, J.M.; Tuladhar, R.; Ma, Z.; Shi, H.; Williams, N.S.; et al. The development of highly potent inhibitors for porcupine. Journal of medicinal chemistry 2013, 56, 2700–2704. [Google Scholar] [CrossRef]
- Proffitt, K.D.; Madan, B.; Ke, Z.; Pendharkar, V.; Ding, L.; Lee, M.A.; Hannoush, R.N.; Virshup, D.M. Pharmacological inhibition of the Wnt acyltransferase PORCN prevents growth of WNT-driven mammary cancer. Cancer research 2013, 73, 502–507. [Google Scholar] [CrossRef]
- Liu, J.; Pan, S.; Hsieh, M.H.; Ng, N.; Sun, F.; Wang, T.; Kasibhatla, S.; Schuller, A.G.; Li, A.G.; Cheng, D.; et al. Targeting Wnt-driven cancer through the inhibition of Porcupine by LGK974. Proceedings of the National Academy of Sciences 2013, 110, 20224–20229. [Google Scholar] [CrossRef] [PubMed]
- Duraiswamy, A.J.; Lee, M.A.; Madan, B.; Ang, S.H.; Tan, E.S.W.; Cheong, W.W.V.; Ke, Z.; Pendharkar, V.; Ding, L.J.; Chew, Y.S.; et al. Discovery and optimization of a porcupine inhibitor. Journal of medicinal chemistry 2015, 58, 5889–5899. [Google Scholar] [CrossRef]
- Wu, X.D.; Bie, Q.L.; Zhang, B.; Yan, Z.H.; Han, Z.J. Wnt10B is critical for the progression of gastric cancer. Oncology Letters 2017, 13, 4231–4237. [Google Scholar] [CrossRef]
- Stevens, J.R.; Miranda-Carboni, G.A.; Singer, M.A.; Brugger, S.M.; Lyons, K.M.; Lane, T.F. Wnt10b deficiency results in age-dependent loss of bone mass and progressive reduction of mesenchymal progenitor cells. Journal of Bone and Mineral Research 2010, 25, 2138–2147. [Google Scholar] [CrossRef]
- Tassew, N.G.; Charish, J.; Shabanzadeh, A.P.; Luga, V.; Harada, H.; Farhani, N.; D’Onofrio, P.; Choi, B.; Ellabban, A.; Nickerson, P.E.; et al. Exosomes Mediate Mobilization of Autocrine Wnt10b to Promote Axonal Regeneration in the Injured CNS. Cell reports 2017, 20, 99–111. [Google Scholar] [CrossRef]
- Cawthorn, W.P.; Bree, A.J.; Yao, Y.; Du, B.; Hemati, N.; Martinez-Santibañez, G.; MacDougald, O.A. Wnt6, Wnt10a and Wnt10b inhibit adipogenesis and stimulate osteoblastogenesis through a β-catenin-dependent mechanism. Bone 2012, 50, 477–489. [Google Scholar] [CrossRef]
- Collins, F.L.; Rios-Arce, N.D.; McCabe, L.R.; Parameswaran, N. Cytokine and hormonal regulation of bone marrow immune cell Wnt10b expression. PloS one 2017, 12, e0181979. [Google Scholar] [CrossRef]
- Wu, G.; Fan, X.; Sun, L. Silencing of Wnt10B reduces viability of heptocellular carcinoma HepG2 cells. American journal of cancer research 2015, 5, 1911. [Google Scholar]
- Wend, P.; Runke, S.; Wend, K.; Anchondo, B.; Yesayan, M.; Jardon, M.; Hardie, N.; Loddenkemper, C.; Ulasov, I.; Lesniak, M.S.; et al. WNT10B/β-catenin signalling induces HMGA2 and proliferation in metastatic triple-negative breast cancer. EMBO molecular medicine 2013, 5, 264–279. [Google Scholar] [CrossRef]
- Chen, Y.; Zeng, C.; Zhan, Y.; Wang, H.; Jiang, X.; Li, W. Aberrant low expression of p85α in stromal fibroblasts promotes breast cancer cell metastasis through exosome-mediated paracrine Wnt10b. Oncogene 2017, 36, 4692. [Google Scholar] [CrossRef]
- Chen, H.; Wang, Y.; Xue, F. Expression and the clinical significance of Wnt10a and Wnt10b in endometrial cancer are associated with the Wnt/β-catenin pathway. Oncology reports 2013, 29, 507–514. [Google Scholar] [CrossRef]
- Yoshikawa, H.; Matsubara, K.; Zhou, X.; Okamura, S.; Kubo, T.; Murase, Y.; Shikauchi, Y.; Esteller, M.; Herman, J.G.; Wang, X.W.; et al. WNT10B functional dualism: β-catenin/Tcf-dependent growth promotion or independent suppression with deregulated expression in cancer. Molecular biology of the cell 2007, 18, 4292–4303. [Google Scholar] [CrossRef]
- Zhu, R.; Yang, Y.; Tian, Y.; Bai, J.; Zhang, X.; Li, X.; Peng, Z.; He, Y.; Chen, L.; Pan, Q.; et al. Ascl2 knockdown results in tumor growth arrest by miRNA-302b-related inhibition of colon cancer progenitor cells. PloS one 2012, 7, e32170. [Google Scholar] [CrossRef]
- Giakountis, A.; Moulos, P.; Zarkou, V.; Oikonomou, C.; Harokopos, V.; Hatzigeorgiou, A.G.; Reczko, M.; Hatzis, P. A Positive regulatory loop between a Wnt-Regulated non-coding RNA and ASCL2 controls intestinal stem cell fate. Cell reports 2016, 15, 2588–2596. [Google Scholar] [CrossRef]
- Schuijers, J.; Junker, J.P.; Mokry, M.; Hatzis, P.; Koo, B.K.; Sasselli, V.; Van Der Flier, L.G.; Cuppen, E.; van Oudenaarden, A.; Clevers, H. Ascl2 acts as an R-spondin/Wnt-responsive switch to control stemness in intestinal crypts. Cell stem cell 2015, 16, 158–170. [Google Scholar] [CrossRef]
- Reddy, V.K.; Short, S.P.; Barrett, C.W.; Mittal, M.K.; Keating, C.E.; Thompson, J.J.; Harris, E.I.; Revetta, F.; Bader, D.M.; Brand, T.; et al. BVES regulates intestinal stem cell programs and intestinal crypt viability after radiation. Stem Cells 2016, 34, 1626–1636. [Google Scholar] [CrossRef]
- Hlavata, I.; Mohelnikova-Duchonova, B.; Vaclavikova, R.; Liska, V.; Pitule, P.; Novak, P.; Bruha, J.; Vycital, O.; Holubec, L.; Treska, V.; et al. The role of ABC transporters in progression and clinical outcome of colorectal cancer. Mutagenesis 2012, 27, 187–196. [Google Scholar] [CrossRef]
- Kobayashi, M.; Funayama, R.; Ohnuma, S.; Unno, M.; Nakayama, K. Wnt-β-catenin signaling regulates ABCC3 (MRP3) transporter expression in colorectal cancer. Cancer science 2016, 107, 1776–1784. [Google Scholar] [CrossRef]
- Kaler, P.; Godasi, B.N.; Augenlicht, L.; Klampfer, L. The NF-κB/AKT-dependent induction of Wnt signaling in colon cancer cells by macrophages and IL-1β. Cancer Microenvironment 2009, 2, 69. [Google Scholar] [CrossRef]
- Zhong, L.; Schivo, S.; Huang, X.; Leijten, J.; Karperien, M.; Post, J.N. Nitric Oxide Mediates Crosstalk between Interleukin 1β and WNT Signaling in Primary Human Chondrocytes by Reducing DKK1 and FRZB Expression. International journal of molecular sciences 2017, 18, 2491. [Google Scholar] [CrossRef]
- Lin, Y.; Xu, J.; Su, H.; Zhong, W.; Yuan, Y.; Yu, Z.; Fang, Y.; Zhou, H.; Li, C.; Huang, K. Interleukin-17 is a favorable prognostic marker for colorectal cancer. Clinical and Translational Oncology 2015, 17, 50–56. [Google Scholar] [CrossRef]
- Housseau, F.; Wu, S.; Wick, E.C.; Fan, H.; Wu, X.; Llosa, N.J.; Smith, K.N.; Tam, A.; Ganguly, S.; Wanyiri, J.W.; et al. Redundant innate and adaptive sources of IL17 production drive colon tumorigenesis. Cancer research 2016, 76, 2115–2124. [Google Scholar] [CrossRef]
- Starnes, T.; Broxmeyer, H.E.; Robertson, M.J.; Hromas, R. Cutting edge: IL-17D, a novel member of the IL-17 family, stimulates cytokine production and inhibits hemopoiesis. The Journal of Immunology 2002, 169, 642–646. [Google Scholar] [CrossRef]
- Ma, B.; Hottiger, M.O. Crosstalk between Wnt/β-catenin and NF-κB signaling pathway during inflammation. Frontiers in immunology 2016, 7, 378. [Google Scholar] [CrossRef]
- Ma, B.; van Blitterswijk, C.A.; Karperien, M. A Wnt/β-catenin negative feedback loop inhibits interleukin-1–induced matrix metalloproteinase expression in human articular chondrocytes. Arthritis & Rheumatism 2012, 64, 2589–2600. [Google Scholar]
- Masckauchán, T.N.H.; Shawber, C.J.; Funahashi, Y.; Li, C.M.; Kitajewski, J. Wnt/β-catenin signaling induces proliferation, survival and interleukin-8 in human endothelial cells. Angiogenesis 2005, 8, 43–51. [Google Scholar] [CrossRef]
- Pfalzer, A.C.; Crott, J.W.; Koh, G.Y.; Smith, D.E.; Garcia, P.E.; Mason, J.B. Interleukin-1 Signaling Mediates Obesity-Promoted Elevations in Inflammatory Cytokines, Wnt Activation, and Epithelial Proliferation in the Mouse Colon. Journal of Interferon & Cytokine Research 2018, 38, 445–451. [Google Scholar]
- Aumiller, V.; Balsara, N.; Wilhelm, J.; Günther, A.; Königshoff, M. WNT/β-catenin signaling induces IL-1β expression by alveolar epithelial cells in pulmonary fibrosis. American journal of respiratory cell and molecular biology 2013, 49, 96–104. [Google Scholar] [CrossRef]
- Chen, D.; Li, W.; Liu, S.; Su, Y.; Han, G.; Xu, C.; Liu, H.; Zheng, T.; Zhou, Y.; Mao, C. Interleukin-23 promotes the epithelial-mesenchymal transition of oesophageal carcinoma cells via the Wnt/β-catenin pathway. Scientific reports 2015, 5, 8604. [Google Scholar] [CrossRef]
- Malysheva, K.; Rooij, K.d.; WGM Löwik, C.; L Baeten, D.; Rose-John, S. Interleukin 6/Wnt interactions in rheumatoid arthritis: interleukin 6 inhibits Wnt signaling in synovial fibroblasts and osteoblasts. Croatian medical journal 2016, 57, 89–98. [Google Scholar] [CrossRef]
- Mukai, A.; Yamamoto-Hino, M.; Awano, W.; Watanabe, W.; Komada, M.; Goto, S. Balanced ubiquitylation and deubiquitylation of Frizzled regulate cellular responsiveness to Wg/Wnt. The EMBO journal 2010, 29, 2114–2125. [Google Scholar] [CrossRef]
- Zhang, L.; Wrana, J.L. The emerging role of exosomes in Wnt secretion and transport. Current opinion in genetics & development 2014, 27, 14–19. [Google Scholar]
- Tassew, N.G.; Charish, J.; Shabanzadeh, A.P.; Luga, V.; Harada, H.; Farhani, N.; D’Onofrio, P.; Choi, B.; Ellabban, A.; Nickerson, P.E.; et al. Exosomes mediate mobilization of autocrine Wnt10b to promote axonal regeneration in the injured CNS. Cell reports 2017, 20, 99–111. [Google Scholar] [CrossRef]
- Koles, K.; Budnik, V. Exosomes go with the Wnt. Cellular logistics 2012, 2, 169–173. [Google Scholar] [CrossRef]
- Wu, X.; Deng, G.; Hao, X.; Li, Y.; Zeng, J.; Ma, C.; He, Y.; Liu, X.; Wang, Y. A caspase-dependent pathway is involved in Wnt/β-catenin signaling promoted apoptosis in Bacillus Calmette-Guerin infected RAW264. 7 macrophages. International journal of molecular sciences 2014, 15, 5045–5062. [Google Scholar] [CrossRef]
- Abdul-Ghani, M.; Dufort, D.; Stiles, R.; De Repentigny, Y.; Kothary, R.; Megeney, L.A. Wnt11 promotes cardiomyocyte development by caspase-mediated suppression of canonical Wnt signals. Molecular and cellular biology 2011, 31, 163–178. [Google Scholar] [CrossRef]
- Bisson, J.A.; Mills, B.; Helt, J.C.P.; Zwaka, T.P.; Cohen, E.D. Wnt5a and Wnt11 inhibit the canonical Wnt pathway and promote cardiac progenitor development via the Caspase-dependent degradation of AKT. Developmental biology 2015, 398, 80–96. [Google Scholar] [CrossRef]
- Singh, V.; Holla, S.; Ramachandra, S.G.; Balaji, K.N. WNT-inflammasome signaling mediates NOD2-induced development of acute arthritis in mice. The Journal of Immunology 2015, 194, 3351–3360. [Google Scholar] [CrossRef]
- Flanagan, L.; Meyer, M.; Fay, J.; Curry, S.; Bacon, O.; Duessmann, H.; John, K.; Boland, K.C.; McNamara, D.A.; Kay, E.W.; et al. Low levels of Caspase-3 predict favourable response to 5FU-based chemotherapy in advanced colorectal cancer: Caspase-3 inhibition as a therapeutic approach. Cell death & disease 2016, 7, e2087. [Google Scholar]
- Yao, Q.; Wang, W.; Jin, J.; Min, K.; Yang, J.; Zhong, Y.; Xu, C.; Deng, J.; Zhou, Y. Synergistic role of Caspase-8 and Caspase-3 expressions: Prognostic and predictive biomarkers in colorectal cancer. Cancer Biomarkers 2018, 1–10. [Google Scholar] [CrossRef]
- Sadot, E.; Geiger, B.; Oren, M.; Ben-Ze’ev, A. Down-regulation of β-catenin by activated p53. Molecular and cellular biology 2001, 21, 6768–6781. [Google Scholar] [CrossRef]
- Peng, X.; Yang, L.; Chang, H.; Dai, G.; Wang, F.; Duan, X.; Guo, L.; Zhang, Y.; Chen, G. Wnt/β-catenin signaling regulates the proliferation and differentiation of mesenchymal progenitor cells through the p53 pathway. PloS one 2014, 9, e97283. [Google Scholar] [CrossRef]
- Zhukova, N.; Ramaswamy, V.; Remke, M.; Martin, D.C.; Castelo-Branco, P.; Zhang, C.H.; Fraser, M.; Tse, K.; Poon, R.; Shih, D.J.; et al. WNT activation by lithium abrogates TP53 mutation associated radiation resistance in medulloblastoma. Acta neuropathologica communications 2014, 2, 174. [Google Scholar] [CrossRef]
- Liu, W.; Xu, X.; Fan, Z.; Sun, G.; Han, Y.; Zhang, D.; Xu, L.; Wang, M.; Wang, X.; Zhang, S.; et al. Wnt Signaling Activates TP53-Induced Glycolysis and Apoptosis Regulator and Protects Against Cisplatin-Induced Spiral Ganglion Neuron Damage in the Mouse Cochlea. Antioxidants & redox signaling 2018. [Google Scholar]
- Okayama, S.; Kopelovich, L.; Balmus, G.; Weiss, R.S.; Herbert, B.S.; Dannenberg, A.J.; Subbaramaiah, K. p53 protein regulates Hsp90 ATPase activity and thereby Wnt signaling by modulating Aha1 expression. Journal of Biological Chemistry 2014, 289, 6513–6525. [Google Scholar] [CrossRef]
- Wang, H.; Fan, L.; Xia, X.; Rao, Y.; Ma, Q.; Yang, J.; Lu, Y.; Wang, C.; Ma, D.; Huang, X. Silencing Wnt2B by siRNA interference inhibits metastasis and enhances chemotherapy sensitivity in ovarian cancer. International Journal of Gynecological Cancer 2012, 22, 755–761. [Google Scholar] [CrossRef]
- Takada, K.; Zhu, D.; Bird, G.H.; Sukhdeo, K.; Zhao, J.J.; Mani, M.; Lemieux, M.; Carrasco, D.E.; Ryan, J.; Horst, D.; et al. Targeted disruption of the BCL9/β-catenin complex inhibits oncogenic Wnt signaling. Science translational medicine 2012, 4, 148ra117. [Google Scholar] [CrossRef]
- Chen, J.; Rajasekaran, M.; Xia, H.; Kong, S.N.; Deivasigamani, A.; Sekar, K.; Gao, H.; Swa, H.L.; Gunaratne, J.; Ooi, L.L.; et al. CDK1-mediated BCL9 phosphorylation inhibits clathrin to promote mitotic Wnt signalling. The EMBO journal 2018, 37, e99395. [Google Scholar] [CrossRef]
- Green, D.R.; Oberst, A.; Dillon, C.P.; Weinlich, R.; Salvesen, G.S. RIPK-dependent necrosis and its regulation by caspases: a mystery in five acts. Molecular cell 2011, 44, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Devin, A.; Rodriguez, Y.; Liu, Z.g. Cleavage of the death domain kinase RIP by caspase-8 prompts TNF-induced apoptosis. Genes & development 1999, 13, 2514–2526. [Google Scholar]
- Estornes, Y.; Aguileta, M.; Dubuisson, C.; De Keyser, J.; Goossens, V.; Kersse, K.; Samali, A.; Vandenabeele, P.; Bertrand, M. RIPK1 promotes death receptor-independent caspase-8-mediated apoptosis under unresolved ER stress conditions. Cell death & disease 2014, 5, e1555. [Google Scholar]
- Weng, D.; Marty-Roix, R.; Ganesan, S.; Proulx, M.K.; Vladimer, G.I.; Kaiser, W.J.; Mocarski, E.S.; Pouliot, K.; Chan, F.K.M.; Kelliher, M.A.; et al. Caspase-8 and RIP kinases regulate bacteria-induced innate immune responses and cell death. Proceedings of the National Academy of Sciences 2014, 111, 7391–7396. [Google Scholar] [CrossRef]
- Moriwaki, K.; Bertin, J.; Gough, P.J.; Chan, F.K.M. A RIPK3–caspase 8 complex mediates atypical pro–IL-1β processing. The Journal of Immunology 2015, 194, 1938–1944. [Google Scholar] [CrossRef]
- Declercq, W.; Berghe, T.V.; Vandenabeele, P. RIP kinases at the crossroads of cell death and survival. Cell 2009, 138, 229–232. [Google Scholar] [CrossRef]
- Chaudhary, P.M.; Eby, M.T.; Jasmin, A.; Kumar, A.; Liu, L.; Hood, L. Activation of the NF-κB pathway by caspase 8 and its homologs. Oncogene 2000, 19, 4451. [Google Scholar] [CrossRef]
- Sheng, Y.H.; He, Y.; Hasnain, S.Z.; Wang, R.; Tong, H.; Clarke, D.T.; Lourie, R.; Oancea, I.; Wong, K.; Lumley, J.W.; et al. MUC13 protects colorectal cancer cells from death by activating the NF-κB pathway and is a potential therapeutic target. Oncogene 2017, 36, 700. [Google Scholar] [CrossRef]
- Sen, R.; Baltimore, D. Multiple nuclear factors interact with the immunoglobulin enhancer sequences. cell 1986, 46, 705–716. [Google Scholar] [CrossRef]
- Tanaka, S.; Nakano, H. NF-κB2 (p100) limits TNF-α–induced osteoclastogenesis. The Journal of clinical investigation 2009, 119, 2879–2881. [Google Scholar] [CrossRef] [PubMed]
- Imamura, H.; Yoshina, S.; Ikari, K.; Miyazawa, K.; Momohara, S.; Mitani, S. Impaired NFKBIE gene function decreases cellular uptake of methotrexate by down-regulating SLC19A1 expression in a human rheumatoid arthritis cell line. Modern rheumatology 2016, 26, 507–516. [Google Scholar] [CrossRef]
- Lee, R.E.; Walker, S.R.; Savery, K.; Frank, D.A.; Gaudet, S. Fold change of nuclear NF-κB determines TNF-induced transcription in single cells. Molecular cell 2014, 53, 867–879. [Google Scholar] [CrossRef]
- Grivennikov, S.I.; Karin, M. Dangerous liaisons: STAT3 and NF-κB collaboration and crosstalk in cancer. Cytokine & growth factor reviews 2010, 21, 11–19. [Google Scholar]
- Liu, Z.; Hazan-Halevy, I.; Harris, D.M.; Li, P.; Ferrajoli, A.; Faderl, S.; Keating, M.J.; Estrov, Z. STAT-3 activates NF-κB in chronic lymphocytic leukemia cells. Molecular Cancer Research 2011, 9, 507–515. [Google Scholar] [CrossRef]
- Lam, L.T.; Wright, G.; Davis, R.E.; Lenz, G.; Farinha, P.; Dang, L.; Chan, J.W.; Rosenwald, A.; Gascoyne, R.D.; Staudt, L.M. Cooperative signaling through the STAT3 and NF-κB pathways in subtypes of diffuse large B cell lymphoma. Blood 2007. [Google Scholar]
- Lee, T.L.; Yeh, J.; Friedman, J.; Yan, B.; Yang, X.; Yeh, N.T.; Van Waes, C.; Chen, Z. A signal network involving coactivated NF-κB and STAT3 and altered p53 modulates BAX/BCL-XL expression and promotes cell survival of head and neck squamous cell carcinomas. International journal of cancer 2008, 122, 1987–1998. [Google Scholar] [CrossRef]
- Ng, S.L.; Friedman, B.A.; Schmid, S.; Gertz, J.; Myers, R.M.; Maniatis, T.; et al. IκB kinase ε (IKKε) regulates the balance between type I and type II interferon responses. Proceedings of the National Academy of Sciences 2011, 108, 21170–21175. [Google Scholar] [CrossRef]
- Guo, J.; Kim, D.; Gao, J.; Kurtyka, C.; Chen, H.; Yu, C.; Wu, D.; Mittal, A.; Beg, A.; Chellappan, S.; et al. IKBKE is induced by STAT3 and tobacco carcinogen and determines chemosensitivity in non-small cell lung cancer. Oncogene 2013, 32, 151. [Google Scholar] [CrossRef]
- Shen, R.R.; Zhou, A.Y.; Kim, E.; Lim, E.; Habelhah, H.; Hahn, W.C. IκB kinase ε phosphorylates TRAF2 to promote mammary epithelial cell transformation. Molecular and cellular biology 2012, 32, 4756–4768. [Google Scholar] [CrossRef] [PubMed]
- Zhou, A.Y.; Shen, R.R.; Kim, E.; Lock, Y.J.; Xu, M.; Chen, Z.J.; Hahn, W.C. IKKε-mediated tumorigenesis requires K63-linked polyubiquitination by a cIAP1/cIAP2/TRAF2 E3 ubiquitin ligase complex. Cell reports 2013, 3, 724–733. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, K.; Akira, S. NF-κB activation through IKK-i-dependent I-TRAF/TANK phosphorylation. Genes to Cells 2000, 5, 191–202. [Google Scholar]
- Gerbod-Giannone, M.C.; Li, Y.; Holleboom, A.; Han, S.; Hsu, L.C.; Tabas, I.; Tall, A.R. TNFα induces ABCA1 through NF-κB in macrophages and in phagocytes ingesting apoptotic cells. Proceedings of the National Academy of Sciences 2006, 103, 3112–3117. [Google Scholar] [CrossRef]
- Van Eck, M.; Bos, I.S.T.; Kaminski, W.E.; Orsó, E.; Rothe, G.; Twisk, J.; Böttcher, A.; Van Amersfoort, E.S.; Christiansen-Weber, T.A.; Fung-Leung, W.P.; et al. Leukocyte ABCA1 controls susceptibility to atherosclerosis and macrophage recruitment into tissues. Proceedings of the National Academy of Sciences 2002, 99, 6298–6303. [Google Scholar] [CrossRef]
- Tang, C.; Liu, Y.; Kessler, P.S.; Vaughan, A.M.; Oram, J.F. The macrophage cholesterol exporter ABCA1 functions as an anti-inflammatory receptor. Journal of Biological Chemistry 2009, 284, 32336–32343. [Google Scholar] [CrossRef]
- Zhu, X.; Owen, J.S.; Wilson, M.D.; Li, H.; Griffiths, G.L.; Thomas, M.J.; Hiltbold, E.M.; Fessler, M.B.; Parks, J.S. Macrophage ABCA1 reduces MyD88-dependent Toll-like receptor trafficking to lipid rafts by reduction of lipid raft cholesterol. Journal of lipid research 2010, 51, 3196–3206. [Google Scholar] [CrossRef]
- Tian, B.; Widen, S.G.; Yang, J.; Wood, T.G.; Kudlicki, A.; Zhao, Y.; Brasier, A.R. The NFκB subunit RELA is a master transcriptional regulator of the committed epithelial-mesenchymal transition in airway epithelial cells. Journal of Biological Chemistry 2018, 293, 16528–16545. [Google Scholar] [CrossRef]
- Ke, B.; Zhao, Z.; Ye, X.; Gao, Z.; Manganiello, V.; Wu, B.; Ye, J. Inactivation of NF-κB p65 (RelA) in liver improves insulin sensitivity and inhibits cAMP/PKA pathway. Diabetes 2015, 64, 3355–3362. [Google Scholar] [CrossRef]
- Weichert, W.; Boehm, M.; Gekeler, V.; Bahra, M.; Langrehr, J.; Neuhaus, P.; Denkert, C.; Imre, G.; Weller, C.; Hofmann, H.; et al. High expression of RelA/p65 is associated with activation of nuclear factor-κB-dependent signaling in pancreatic cancer and marks a patient population with poor prognosis. British journal of cancer 2007, 97, 523. [Google Scholar] [CrossRef]
- Brooks, R.S.; Ciappio, E.D.; Bennett, G.; Crott, J.W.; Mason, J.B.; Liu, Z. TNF-α induced alterations in the Wnt signaling cascade: a potential mechanism for obesity-associated colorectal tumorigenesis. The FASEB Journal 2010, 24, lb384. [Google Scholar] [CrossRef]
- Adami, G.; Orsolini, G.; Adami, S.; Viapiana, O.; Idolazzi, L.; Gatti, D.; Rossini, M. Effects of TNF inhibitors on parathyroid hormone and Wnt signaling antagonists in rheumatoid arthritis. Calcified tissue international 2016, 99, 360–364. [Google Scholar] [CrossRef] [PubMed]
- Hiyama, A.; Yokoyama, K.; Nukaga, T.; Sakai, D.; Mochida, J. A complex interaction between Wnt signaling and TNF-α in nucleus pulposus cells. Arthritis research & therapy 2013, 15, R189. [Google Scholar]
- Roubert, A.; Gregory, K.; Li, Y.; Pfalzer, A.C.; Li, J.; Schneider, S.S.; Wood, R.J.; Liu, Z. The influence of tumor necrosis factor-α on the tumorigenic Wnt-signaling pathway in human mammary tissue from obese women. Oncotarget 2017, 8, 36127. [Google Scholar] [CrossRef]
- Jang, J.; Jung, Y.; Chae, S.; Chung, S.I.; Kim, S.M.; Yoon, Y. WNT/β-catenin pathway modulates the TNF-α-induced inflammatory response in bronchial epithelial cells. Biochemical and biophysical research communications 2017, 484, 442–449. [Google Scholar] [CrossRef] [PubMed]
- Dharmani, P.; Leung, P.; Chadee, K. Tumor necrosis factor-α and Muc2 mucin play major roles in disease onset and progression in dextran sodium sulphate-induced colitis. PloS one 2011, 6, e25058. [Google Scholar] [CrossRef]
- Levine, S.J.; Larivee, P.; Logun, C.; Angus, C.W.; Ognibene, F.P.; Shelhamer, J.H. Tumor necrosis factor-alpha induces mucin hypersecretion and MUC-2 gene expression by human airway epithelial cells. American Journal of Respiratory Cell and Molecular Biology 1995, 12, 196–204. [Google Scholar] [CrossRef]
- Sikder, M.A.; Lee, H.J.; Mia, M.Z.; Park, S.H.; Ryu, J.; Kim, J.H.; Min, S.Y.; Hong, J.H.; Seok, J.H.; Lee, C.J. Inhibition of TNF-α-Induced MUC5AC Mucin Gene Expression and Production by Wogonin Through the Inactivation of NF-κB Signaling in Airway Epithelial Cells. Phytotherapy Research 2014, 28, 62–68. [Google Scholar] [CrossRef]
- Shao, M.X.; Nadel, J.A. Neutrophil elastase induces MUC5AC mucin production in human airway epithelial cells via a cascade involving protein kinase C, reactive oxygen species, and TNF-α-converting enzyme. The Journal of Immunology 2005, 175, 4009–4016. [Google Scholar] [CrossRef]
- Lora, J.M.; Zhang, D.M.; Liao, S.M.; Burwell, T.; King, A.M.; Barker, P.A.; Singh, L.; Keaveney, M.; Morgenstern, J.; Gutiérrez-Ramos, J.C.; et al. Tumor necrosis factor-α triggers mucus production in airway epithelium through an IκB kinase β-dependent mechanism. Journal of Biological Chemistry 2005, 280, 36510–36517. [Google Scholar] [CrossRef]
- Fischer, B.M.; Rochelle, L.G.; Voynow, J.A.; Akley, N.J.; Adler, K.B. Tumor necrosis factor-α stimulates mucin secretion and cyclic GMP production by guinea pig tracheal epithelial cells in vitro. American Journal of Respiratory Cell and Molecular Biology 1999, 20, 413–422. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Haruta, A.; Kawano, H.; Ho, S.B.; Adams, G.L.; Juhn, S.K.; Kim, Y. Induction of Mucin Gene Expression in Middle Ear of Rats by Tumor Necrosis Factor—α: Potential Cause for Mucoid Otitis Media. The Journal of infectious diseases 2000, 182, 882–887. [Google Scholar] [CrossRef]
- Yoon, J.H.; Kim, K.S.; Kim, H.U.; Linton, J.A.; Lee, J.G. Effects of TNF-a and IL-1 ß on Mucin, Lysozyme, IL-6 and IL-8 in passage-2 Normal human nasal epithelial cells. Acta oto-laryngologica 1999, 119, 905–910. [Google Scholar] [PubMed]
- Mercogliano, M.F.; De Martino, M.; Venturutti, L.; Rivas, M.A.; Proietti, C.J.; Inurrigarro, G.; Frahm, I.; Allemand, D.H.; Deza, E.G.; Ares, S.; et al. TNFα-induced mucin 4 expression elicits trastuzumab resistance in HER2-positive breast cancer. Clinical Cancer Research 2017, 23, 636–648. [Google Scholar] [CrossRef]
- Scarl, R.T.; Lawrence, C.M.; Gordon, H.M.; Nunemaker, C.S. STEAP4: its emerging role in metabolism and homeostasis of cellular iron and copper. Journal of Endocrinology 2017, 234, R123–R134. [Google Scholar] [CrossRef]
- Moldes, M.; Lasnier, F.; Gauthereau, X.; Klein, C.; Pairault, J.; Fève, B.; Chambaut-Guérin, A.M. Tumor necrosis factor-α-induced adipose-related protein (TIARP), a cell-surface protein that is highly induced by tumor necrosis factor-α and adipose conversion. Journal of Biological Chemistry 2001, 276, 33938–33946. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Tao, Y.; Zhang, Z.; Guo, X.; An, P.; Shen, Y.; Wu, Q.; Yu, Y.; Wang, F. Metalloreductase Steap3 coordinates the regulation of iron homeostasis and inflammatory responses. Haematologica 2012, 97, 1826–1835. [Google Scholar] [CrossRef]
- Tanaka, Y.; Matsumoto, I.; Iwanami, K.; Inoue, A.; Umeda, N.; Sugihara, M.; Hayashi, T.; Ito, S.; Sumida, T. Six-transmembrane epithelial antigen of prostate 4 (STEAP4) is expressed on monocytes/neutrophils, and is regulated by TNF antagonist in patients with rheumatoid arthritis. Clinical and experimental rheumatology 2012, 30, 99–102. [Google Scholar]
- Tanaka, Y.; Matsumoto, I.; Iwanami, K.; Inoue, A.; Minami, R.; Umeda, N.; Kanamori, A.; Ochiai, N.; Miyazawa, K.; Sugihara, M. Six-transmembrane epithelial antigen of prostate4 (STEAP4) is a tumor necrosis factor alpha-induced protein that regulates IL-6, IL-8, and cell proliferation in synovium from patients with rheumatoid arthritis. Modern rheumatology 2012, 22, 128–136. [Google Scholar] [CrossRef]
- Gauss, G.H.; Kleven, M.D.; Sendamarai, A.K.; Fleming, M.D.; Lawrence, C.M. The crystal structure of six-transmembrane epithelial antigen of the prostate 4 (Steap4), a ferri/cuprireductase, suggests a novel interdomain flavin-binding site. Journal of Biological Chemistry 2013, 288, 20668–20682. [Google Scholar] [CrossRef]
- ZHANG, C.m.; Chi, X.; Wang, B.; Zhang, M.; NI, Y.h.; CHEN, R.h.; LI, X.n.; GUO, X.r. Downregulation of STEAP4, a highly-expressed TNF-α-inducible gene in adipose tissue, is associated with obesity in humans 1. Acta Pharmacologica Sinica 2008, 29, 587–592. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Xing, X.; Beamer, M.A.; Swindell, W.R.; Sarkar, M.K.; Roberts, L.W.; Voorhees, J.J.; Kahlenberg, J.M.; Harms, P.W.; Johnston, A.; et al. Six-transmembrane epithelial antigens of the prostate comprise a novel inflammatory nexus in patients with pustular skin disorders. Journal of Allergy and Clinical Immunology 2017, 139, 1217–1227. [Google Scholar] [CrossRef] [PubMed]
- Gomes, I.M.; Maia, C.J.; Santos, C.R. STEAP proteins: from structure to applications in cancer therapy. Molecular Cancer Research 2012, 10, 573–587. [Google Scholar] [CrossRef] [PubMed]
- Fu, B.; Li, S.; Wang, L.; Berman, M.A.; Dorf, M.E. The ubiquitin conjugating enzyme UBE2L3 regulates TNFα-induced linear ubiquitination. Cell research 2014, 24, 376. [Google Scholar] [CrossRef]
- Li, Y.P.; Lecker, S.H.; Chen, Y.; Waddell, I.D.; Goldberg, A.L.; Reid, M.B. TNF-α increases ubiquitin-conjugating activity in skeletal muscle by up-regulating UbcH2/E220k. The FASEB Journal 2003, 17, 1048–1057. [Google Scholar] [CrossRef]
- Shembade, N.; Ma, A.; Harhaj, E.W. Inhibition of NF-κB signaling by A20 through disruption of ubiquitin enzyme complexes. Science 2010, 327, 1135–1139. [Google Scholar] [CrossRef]
- Tamatani, M.; Che, Y.H.; Matsuzaki, H.; Ogawa, S.; Okado, H.; Miyake, S.i.; Mizuno, T.; Tohyama, M. Tumor necrosis factor induces Bcl-2 and Bcl-x expression through NFκB activation in primary hippocampal neurons. Journal of Biological Chemistry 1999, 274, 8531–8538. [Google Scholar] [CrossRef] [PubMed]
- Ho, S.; Winkler-Lowen, B.; Morrish, D.; Dakour, J.; Li, H.; Guilbert, L. The role of Bcl-2 expression in EGF inhibition of TNF-α/IFN-γ-induced villous trophoblast apoptosis. Placenta 1999, 20, 423–430. [Google Scholar] [CrossRef]
- Genestier, L.; Bonnefoy-Berard, N.; Rouault, J.P.; Flacher, M.; Revillard, J.P. Tumor necrosis factor-α up-regulates Bcl-2 expression and decreases calcium-dependent apoptosis in human B cell lines. International immunology 1995, 7, 533–540. [Google Scholar] [CrossRef]
- Jäättelä, M.; Benedict, M.; Tewari, M.; Shayman, J.; Dixit, V. Bcl-x and Bcl-2 inhibit TNF and Fas-induced apoptosis and activation of phospholipase A2 in breast carcinoma cells. Oncogene 1995, 10, 2297–2305. [Google Scholar]
- Kim, J.; Lee, S.; Park, J.; Yoo, Y. TNF-α-induced ROS production triggering apoptosis is directly linked to Romo1 and Bcl-X L. Cell death and differentiation 2010, 17, 1420. [Google Scholar] [CrossRef] [PubMed]
- Kuwata, H.; Watanabe, Y.; Miyoshi, H.; Yamamoto, M.; Kaisho, T.; Takeda, K.; Akira, S. IL-10-inducible Bcl-3 negatively regulates LPS-induced TNF-α production in macrophages. Blood 2003, 102, 4123–4129. [Google Scholar] [CrossRef] [PubMed]
- Esche, C.; Shurin, G.V.; Kirkwood, J.M.; Wang, G.Q.; Rabinowich, H.; Pirtskhalaishvili, G.; Shurin, M.R. Tumor necrosis factor-α-promoted expression of Bcl-2 and inhibition of mitochondrial cytochrome c release mediate resistance of mature dendritic cells to melanoma-induced apoptosis. Clinical cancer research 2001, 7, 974s–979s. [Google Scholar] [PubMed]
- Thacker, J.; Zdzienicka, M.Z. The mammalian XRCC genes: their roles in DNA repair and genetic stability. DNA repair 2003, 2, 655–672. [Google Scholar] [CrossRef]
- Thacker, J.; Zdzienicka, M.Z. The XRCC genes: expanding roles in DNA double-strand break repair. DNA repair 2004, 3, 1081–1090. [Google Scholar] [CrossRef]
- Sultana, R.; Abdel-Fatah, T.; Perry, C.; Moseley, P.; Albarakti, N.; Mohan, V.; Seedhouse, C.; Chan, S.; Madhusudan, S. Ataxia telangiectasia mutated and Rad3 related (ATR) protein kinase inhibition is synthetically lethal in XRCC1 deficient ovarian cancer cells. PloS one 2013, 8, e57098. [Google Scholar] [CrossRef]
- Della-Maria, J.; Zhou, Y.; Tsai, M.S.; Kuhnlein, J.; Carney, J.P.; Paull, T.T.; Tomkinson, A.E. Human Mre11/human Rad50/Nbs1 and DNA ligase IIIα/XRCC1 protein complexes act together in an alternative nonhomologous end joining pathway. Journal of Biological Chemistry 2011, 286, 33845–33853. [Google Scholar] [CrossRef]
- Morales, J.C.; Richard, P.; Patidar, P.L.; Motea, E.A.; Dang, T.T.; Manley, J.L.; Boothman, D.A. XRN2 links transcription termination to DNA damage and replication stress. PLoS genetics 2016, 12, e1006107. [Google Scholar] [CrossRef]
- Saintigny, Y.; Dumay, A.; Lambert, S.; Lopez, B.S. A novel role for the Bcl-2 protein family: specific suppression of the RAD51 recombination pathway. The EMBO journal 2001, 20, 2596–2607. [Google Scholar] [CrossRef]
- Meng, J.; Liu, X.; Cao, X. A new cytosolic DNA-recognition pathway for DNA-induced inflammatory responses. Cellular & Molecular Immunology 2014, 11, 506. [Google Scholar]
- Marin-Vicente, C.; Domingo-Prim, J.; Eberle, A.B.; Visa, N. RRP6/EXOSC10 is required for the repair of DNA double-strand breaks by homologous recombination. J Cell Sci 2015, 128, 1097–1107. [Google Scholar] [CrossRef] [PubMed]
- Shtam, T.A.; Kovalev, R.A.; Varfolomeeva, E.Y.; Makarov, E.M.; Kil, Y.V.; Filatov, M.V. Exosomes are natural carriers of exogenous siRNA to human cells in vitro. Cell Communication and Signaling 2013, 11, 88. [Google Scholar] [CrossRef] [PubMed]
- Walden, H.; Deans, A.J. The Fanconi anemia DNA repair pathway: structural and functional insights into a complex disorder. Annual review of biophysics 2014, 43, 257–278. [Google Scholar] [CrossRef]
- Cohn, M.A.; D’Andrea, A.D. Chromatin recruitment of DNA repair proteins: lessons from the fanconi anemia and double-strand break repair pathways. Molecular cell 2008, 32, 306–312. [Google Scholar] [CrossRef] [PubMed]
- Romick-Rosendale, L.E.; Lui, V.W.; Grandis, J.R.; Wells, S.I. The Fanconi anemia pathway: repairing the link between DNA damage and squamous cell carcinoma. Mutation Research/Fundamental and Molecular Mechanisms of Mutagenesis 2013, 743, 78–88. [Google Scholar] [CrossRef]
- Michl, J.; Zimmer, J.; Tarsounas, M. Interplay between Fanconi anemia and homologous recombination pathways in genome integrity. The EMBO journal 2016, 35, 909–923. [Google Scholar] [CrossRef]
- Liang, F.; Longerich, S.; Miller, A.S.; Tang, C.; Buzovetsky, O.; Xiong, Y.; Maranon, D.G.; Wiese, C.; Kupfer, G.M.; Sung, P. Promotion of RAD51-mediated homologous DNA pairing by the RAD51AP1-UAF1 complex. Cell reports 2016, 15, 2118–2126. [Google Scholar] [CrossRef]
- Taniguchi, T.; Garcia-Higuera, I.; Andreassen, P.R.; Gregory, R.C.; Grompe, M.; D’Andrea, A.D. S-phase–specific interaction of the Fanconi anemia protein, FANCD2, with BRCA1 and RAD51. Blood 2002, 100, 2414–2420. [Google Scholar] [CrossRef]
- Zadorozhny, K.; Sannino, V.; Beláň, O.; Mlčoušková, J.; Špírek, M.; Costanzo, V.; Krejčí, L. Fanconi-anemia-associated mutations destabilize RAD51 filaments and impair replication fork protection. Cell reports 2017, 21, 333–340. [Google Scholar] [CrossRef]
- Geng, L.; Huntoon, C.J.; Karnitz, L.M. RAD18-mediated ubiquitination of PCNA activates the Fanconi anemia DNA repair network. The Journal of cell biology 2010, 191, 249–257. [Google Scholar] [CrossRef]
- Palle, K.; Vaziri, C. Rad18 E3 ubiquitin ligase activity mediates Fanconi anemia pathway activation and cell survival following DNA Topoisomerase 1 inhibition. Cell Cycle 2011, 10, 1625–1638. [Google Scholar] [CrossRef]
- García-Luis, J.; Machín, F. Fanconi Anaemia-Like Mph1 Helicase Backs up Rad54 and Rad5 to Circumvent Replication Stress-Driven Chromosome Bridges. Genes 2018, 9, 558. [Google Scholar] [CrossRef]
- Lipinska, N.; Romaniuk, A.; Paszel-Jaworska, A.; Toton, E.; Kopczynski, P.; Rubis, B. Telomerase and drug resistance in cancer. Cellular and Molecular Life Sciences 2017, 74, 4121–4132. [Google Scholar] [CrossRef]
- Wang, J.; Liu, X.; Fang, J. Expression and clinical significance of telomerase catalytic subunit gene in lung cancer and its correlations with genes related to drug resistance and apoptosis. Zhonghua zhong liu za zhi [Chinese journal of oncology] 1999, 21, 350–353. [Google Scholar]
- Sakin, V.; Eskiocak, U.; Kars, M.; Iseri, Ö.; Gunduz, U. hTERT gene expression levels and telomerase activity in drug resistant MCF-7 cells. Experimental oncology 2008. [Google Scholar]
- Keshet, G.I.; Goldstein, I.; Itzhaki, O.; Cesarkas, K.; Shenhav, L.; Yakirevitch, A.; Treves, A.J.; Schachter, J.; Amariglio, N.; Rechavi, G. MDR1 expression identifies human melanoma stem cells. Biochemical and biophysical research communications 2008, 368, 930–936. [Google Scholar] [CrossRef]
- Zhou, X.; Engel, T.; Goepfert, C.; Erren, M.; Assmann, G.; von Eckardstein, A. The ATP binding cassette transporter A1 contributes to the secretion of interleukin 1β from macrophages but not from monocytes. Biochemical and biophysical research communications 2002, 291, 598–604. [Google Scholar] [CrossRef]
- Haskó, G.; Deitch, E.A.; Németh, Z.H.; Kuhel, D.G.; Szabó, C. Inhibitors of ATP-binding cassette transporters suppress interleukin-12 p40 production and major histocompatibility complex II up-regulation in macrophages. Journal of Pharmacology and Experimental Therapeutics 2002, 301, 103–110. [Google Scholar] [CrossRef]
- Park, S.Y.; Han, J.; Kim, J.B.; Yang, M.G.; Kim, Y.J.; Lim, H.J.; An, S.Y.; Kim, J.H. Interleukin-8 is related to poor chemotherapeutic response and tumourigenicity in hepatocellular carcinoma. European Journal of Cancer 2014, 50, 341–350. [Google Scholar] [CrossRef]
- Marty, V.; Médina, C.; Combe, C.; Parnet, P.; Amédée, T. ATP binding cassette transporter ABC1 is required for the release of interleukin-1β by P2X7-stimulated and lipopolysaccharide-primed mouse Schwann cells. Glia 2005, 49, 511–519. [Google Scholar] [CrossRef]
- Lottaz, D.; Beleznay, Z.; Bickel, M. Inhibition of ATP-binding cassette transporter downregulates interleukin-1β-mediated autocrine activation of human dermal fibroblasts. Journal of investigative dermatology 2001, 117, 871–876. [Google Scholar] [CrossRef] [PubMed]
- Ruzickova, E.; Janska, R.; Dolezel, P.; Mlejnek, P. Clinically relevant interactions of anti-apoptotic Bcl-2 protein inhibitors with ABC transporters. Die Pharmazie-An International Journal of Pharmaceutical Sciences 2017, 72, 751–758. [Google Scholar]
- Alla, V.; Kowtharapu, B.S.; Engelmann, D.; Emmrich, S.; Schmitz, U.; Steder, M.; Pützer, B.M. E2F1 confers anticancer drug resistance by targeting ABC transporter family members and Bcl-2 via the p73/DNp73-miR-205 circuitry. Cell cycle 2012, 11, 3067–3078. [Google Scholar] [CrossRef] [PubMed]
- Yasui, K.; Mihara, S.; Zhao, C.; Okamoto, H.; Saito-Ohara, F.; Tomida, A.; Funato, T.; Yokomizo, A.; Naito, S.; Imoto, I.; et al. Alteration in copy numbers of genes as a mechanism for acquired drug resistance. Cancer research 2004, 64, 1403–1410. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Wang, M.; Guan, X.; Yuan, Z.; Liu, Z.; Zou, C.; Wang, G.; Gao, X.; Wang, X. Loss of ABCB4 attenuates the caspase-dependent apoptosis regulating resistance to 5-Fu in colorectal cancer. Bioscience reports 2018, 38, BSR20171428. [Google Scholar] [CrossRef]
- Ihlefeld, K.; Vienken, H.; Claas, R.F.; Blankenbach, K.; Rudowski, A.; Ter Braak, M.; Koch, A.; Van Veldhoven, P.P.; Pfeilschifter, J.; zu Heringdorf, D.M. Upregulation of ABC transporters contributes to chemoresistance of sphingosine 1-phosphate lyase-deficient fibroblasts. Journal of lipid research 2015, 56, 60–69. [Google Scholar] [CrossRef]
- Hörber, S.; Hildebrand, D.G.; Lieb, W.S.; Lorscheid, S.; Hailfinger, S.; Schulze-Osthoff, K.; Essmann, F. The atypical inhibitor of NF-κB, IκBζ, controls macrophage interleukin-10 expression. Journal of Biological Chemistry 2016, 291, 12851–12861. [Google Scholar] [CrossRef]
- Yamazaki, S.; Muta, T.; Matsuo, S.; Takeshige, K. Stimulus-specific induction of a novel nuclear factor-κB regulator, IκB-ζ, via Toll/Interleukin-1 receptor is mediated by mRNA stabilization. Journal of Biological Chemistry 2005, 280, 1678–1687. [Google Scholar] [CrossRef]
- Kurzrock, R.; Estrov, Z.; Ku, S.; Leonard, M.; Talpaz, M. Interleukin-1 increases expression of the LYT-10 (NFκB2) proto-oncogene/transcription factor in renal cell carcinoma lines. Journal of Laboratory and Clinical Medicine 1998, 131, 261–268. [Google Scholar] [CrossRef]
- Lee, S.; Sabath, D.; Deutsch, C.; Prystowsky, M.B. Increased voltage-gated potassium conductance during interleukin 2-stimulated proliferation of a mouse helper T lymphocyte clone. The Journal of cell biology 1986, 102, 1200–1208. [Google Scholar] [CrossRef]
- Martin, G.; O’connell, R.J.; Pietrzykowski, A.Z.; Treistman, S.N.; Ethier, M.F.; Madison, J.M. Interleukin-4 activates large-conductance, calcium-activated potassium (BKCa) channels in human airway smooth muscle cells. Experimental physiology 2008, 93, 908–918. [Google Scholar] [CrossRef] [PubMed]
- Kerschner, J.E.; Meyer, T.K.; Yang, C.; Burrows, A. Middle ear epithelial mucin production in response to interleukin-6 exposure in vitro. Cytokine 2004, 26, 30–36. [Google Scholar] [CrossRef]
- Chen, Y.; Thai, P.; Zhao, Y.H.; Ho, Y.S.; DeSouza, M.M.; Wu, R. Stimulation of airway mucin gene expression by interleukin (IL)-17 through IL-6 paracrine/autocrine loop. Journal of Biological Chemistry 2003, 278, 17036–17043. [Google Scholar] [CrossRef] [PubMed]
- Shan, Y.S.; Hsu, H.P.; Lai, M.D.; Yen, M.C.; FANg, J.H.; Weng, T.Y.; Chen, Y.L. Suppression of mucin 2 promotes interleukin-6 secretion and tumor growth in an orthotopic immune-competent colon cancer animal model. Oncology reports 2014, 32, 2335–2342. [Google Scholar] [CrossRef] [PubMed]
- Yokoigawa, N.; Takeuchi, N.; Toda, M.; Inoue, M.; Kaibori, M.; Yanagida, H.; Tanaka, H.; Ogura, T.; Takada, H.; Okumura, T.; et al. Enhanced production of interleukin 6 in peripheral blood monocytes stimulated with mucins secreted into the bloodstream. Clinical cancer research 2005, 11, 6127–6132. [Google Scholar] [CrossRef]
- Gray, T.; Nettesheim, P.; Loftin, C.; Koo, J.S.; Bonner, J.; Peddada, S.; Langenbach, R. Interleukin-1β–Induced Mucin Production in Human Airway Epithelium Is Mediated by Cyclooxygenase-2, Prostaglandin E2 Receptors, and Cyclic AMP-Protein Kinase A Signaling. Molecular Pharmacology 2004, 66, 337–346. [Google Scholar] [CrossRef] [PubMed]
- Hsu, H.P.; Lai, M.D.; Lee, J.C.; Yen, M.C.; Weng, T.Y.; Chen, W.C.; Fang, J.H.; Chen, Y.L. Mucin 2 silencing promotes colon cancer metastasis through interleukin-6 signaling. Scientific reports 2017, 7, 5823. [Google Scholar] [CrossRef]
- Brighenti, E.; Calabrese, C.; Liguori, G.; Giannone, F.; Trere, D.; Montanaro, L.; Derenzini, M. Interleukin 6 downregulates p53 expression and activity by stimulating ribosome biogenesis: a new pathway connecting inflammation to cancer. Oncogene 2014, 33, 4396. [Google Scholar] [CrossRef]
- Tan, X.; Carretero, J.; Chen, Z.; Zhang, J.; Wang, Y.; Chen, J.; Li, X.; Ye, H.; Tang, C.; Cheng, X.; et al. Loss of p53 attenuates the contribution of IL-6 deletion on suppressed tumor progression and extended survival in Kras-driven murine lung cancer. PloS one 2013, 8, e80885. [Google Scholar] [CrossRef]
- Pützer, B.; Bramson, J.; Addison, C.; Hitt, M.; Siegel, P.; Muller, W.; Graham, F. Combination therapy with interleukin-2 and wild-type p53 expressed by adenoviral vectors potentiates tumor regression in a murine model of breast cancer. Human gene therapy 1998, 9, 707–718. [Google Scholar] [CrossRef]
- Dijsselbloem, N.; Goriely, S.; Albarani, V.; Gerlo, S.; Francoz, S.; Marine, J.C.; Goldman, M.; Haegeman, G.; Berghe, W.V. A critical role for p53 in the control of NF-κB-dependent gene expression in TLR4-stimulated dendritic cells exposed to genistein. The Journal of immunology 2007, 178, 5048–5057. [Google Scholar] [CrossRef] [PubMed]
- Schauer, I.G.; Zhang, J.; Xing, Z.; Guo, X.; Mercado-Uribe, I.; Sood, A.K.; Huang, P.; Liu, J. Interleukin-1β promotes ovarian tumorigenesis through a p53/NF-κB-mediated inflammatory response in stromal fibroblasts. Neoplasia 2013, 15, 409–IN18. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.R.; Quinton, L.J.; Simms, B.T.; Lupa, M.M.; Kogan, M.S.; Mizgerd, J.P. Roles of interleukin-6 in activation of STAT proteins and recruitment of neutrophils during Escherichia coli pneumonia. The Journal of infectious diseases 2006, 193, 360–369. [Google Scholar] [CrossRef] [PubMed]
- Kotanides, H.; Moczygemba, M.; White, M.F.; Reich, N.C. Characterization of the interleukin-4 nuclear activated factor/STAT and its activation independent of the insulin receptor substrate proteins. Journal of Biological Chemistry 1995, 270, 19481–19486. [Google Scholar] [CrossRef]
- Adam, N.; Rabe, B.; Suthaus, J.; Grötzinger, J.; Rose-John, S.; Scheller, J. Unraveling viral interleukin-6 binding to gp130 and activation of STAT-signaling pathways independently of the interleukin-6 receptor. Journal of virology 2009, 83, 5117–5126. [Google Scholar] [CrossRef]
- Frank, D.A.; Robertson, M.J.; Bonni, A.; Ritz, J.; Greenberg, M.E. Interleukin 2 signaling involves the phosphorylation of Stat proteins. Proceedings of the National Academy of Sciences 1995, 92, 7779–7783. [Google Scholar] [CrossRef]
- Boyd, Z.S.; Kriatchko, A.; Yang, J.; Agarwal, N.; Wax, M.B.; Patil, R.V. Interleukin-10 receptor signaling through STAT-3 regulates the apoptosis of retinal ganglion cells in response to stress. Investigative ophthalmology & visual science 2003, 44, 5206–5211. [Google Scholar]
- Kurgonaite, K.; Gandhi, H.; Kurth, T.; Pautot, S.; Schwille, P.; Weidemann, T.; Bökel, C. Essential role of endocytosis for interleukin-4-receptor-mediated JAK/STAT signalling. J Cell Sci 2015, 128, 3781–3795. [Google Scholar] [CrossRef]
- Kondo, M.; Yamaoka, K.; Sakata, K.; Sonomoto, K.; Lin, L.; Nakano, K.; Tanaka, Y. Contribution of the interleukin-6/STAT-3 signaling pathway to chondrogenic differentiation of human mesenchymal stem cells. Arthritis & rheumatology 2015, 67, 1250–1260. [Google Scholar]
- Tanaka, N.; Hoshino, Y.; Gold, J.; Hoshino, S.; Martiniuk, F.; Kurata, T.; Pine, R.; Levy, D.; Rom, W.N.; Weiden, M. Interleukin-10 induces inhibitory C/EBPβ through STAT-3 and represses HIV-1 transcription in macrophages. American journal of respiratory cell and molecular biology 2005, 33, 406–411. [Google Scholar] [CrossRef]
- Jobst, B.; Weigl, J.; Michl, C.; Vivarelli, F.; Pinz, S.; Amslinger, S.; Rascle, A. Inhibition of interleukin-3-and interferon-α-induced JAK/STAT signaling by the synthetic α-X-2’, 3, 4, 4’-tetramethoxychalcones α-Br-TMC and α-CF3-TMC. Biological chemistry 2016, 397, 1187–1204. [Google Scholar] [CrossRef] [PubMed]
- Greene, C.; O’Neill, L. Interleukin-1 receptor-associated kinase and TRAF-6 mediate the transcriptional regulation of interleukin-2 by interleukin-1 via NFκB but unlike interleukin-1 are unable to stabilise interleukin-2 mRNA. Biochimica et Biophysica Acta (BBA)-Molecular Cell Research 1999, 1451, 109–121. [Google Scholar] [CrossRef]
- Cao, Z.; Xiong, J.; Takeuchi, M.; Kurama, T.; Goeddel, D.V. TRAF6 is a signal transducer for interleukin-1. Nature 1996, 383, 443. [Google Scholar] [CrossRef] [PubMed]
- Schwandner, R.; Yamaguchi, K.; Cao, Z. Requirement of tumor necrosis factor receptor–associated factor (TRAF) 6 in interleukin 17 signal transduction. Journal of Experimental Medicine 2000, 191, 1233–1240. [Google Scholar] [CrossRef] [PubMed]
- Lomaga, M.A.; Yeh, W.C.; Sarosi, I.; Duncan, G.S.; Furlonger, C.; Ho, A.; Morony, S.; Capparelli, C.; Van, G.; Kaufman, S.; et al. TRAF6 deficiency results in osteopetrosis and defective interleukin-1, CD40, and LPS signaling. Genes & development 1999, 13, 1015–1024. [Google Scholar]
- Jefferies, C.; Bowie, A.; Brady, G.; Cooke, E.L.; Li, X.; O’Neill, L.A. Transactivation by the p65 subunit of NF-κB in response to interleukin-1 (IL-1) involves MyD88, IL-1 receptor-associated kinase 1, TRAF-6, and Rac1. Molecular and cellular biology 2001, 21, 4544–4552. [Google Scholar] [CrossRef]
- Wu, H.; Arron, J.R. TRAF6, a molecular bridge spanning adaptive immunity, innate immunity and osteoimmunology. Bioessays 2003, 25, 1096–1105. [Google Scholar] [CrossRef]
- Ramadoss, P.; Chiappini, F.; Bilban, M.; Hollenberg, A.N. Regulation of hepatic six transmembrane epithelial antigen of prostate 4 (STEAP4) expression by STAT3 and CCAAT/enhancer-binding protein α. Journal of Biological Chemistry 2010, 285, 16453–16466. [Google Scholar] [CrossRef]
- Hamon, Y.; Luciani, M.F.; Becq, F.; Verrier, B.; Rubartelli, A.; Chimini, G. Interleukin-1β secretion is impaired by inhibitors of the ATP binding cassette transporter, ABC1. Blood 1997, 90, 2911–2915. [Google Scholar] [CrossRef]
- Neta, R.; Sayers, T.; Oppenheim, J. Relationship of TNF to interleukins. Immunology series 1992, 56, 499–566. [Google Scholar]
- Rieckmann, P.; Tuscano, J.; Kehrl, J. Tumor necrosis factor-α (TNF-α) and interleukin-6 (IL-6) in B-lymphocyte function. Methods 1997, 11, 128–132. [Google Scholar] [CrossRef] [PubMed]
- Bethea, J.R.; Gillespie, G.Y.; Benveniste, E.N. Interleukin-1β induction of TNF-α gene expression: Involvement of protein kinase C. Journal of cellular physiology 1992, 152, 264–273. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, F.; Hayes, B.P.; Horgan, C.M.; Beesley, J.E.; Campbell, C.J.; Randi, A.M. Tumor necrosis factor (TNF)-α and interleukin (IL)-1β down-regulate intercellular adhesion molecule (ICAM)-2 expression on the endothelium. Cell adhesion and communication 1998, 6, 381–400. [Google Scholar] [CrossRef] [PubMed]
- Zhai, R.; Liu, G.; Ge, X.; Bao, W.; Wu, C.; Yang, C.; Liang, D. Serum levels of tumor necrosis factor-α (TNF-α), interleukin 6 (IL-6), and their soluble receptors in coal workers’ pneumoconiosis. Respiratory medicine 2002, 96, 829–834. [Google Scholar] [CrossRef]
- Havlir, D.V.; Torriani, F.J.; Schrier, R.D.; Huang, J.Y.; Lederman, M.M.; Chervenak, K.A.; Boom, W.H. Serum interleukin-6 (IL-6), IL-10, tumor necrosis factor (TNF) alpha, soluble type II TNF receptor, and transforming growth factor beta levels in human immunodeficiency virus type 1-infected individuals with Mycobacterium avium complex disease. Journal of clinical microbiology 2001, 39, 298–303. [Google Scholar] [CrossRef]
- Tissi, L.; Puliti, M.; Barluzzi, R.; Orefici, G.; von Hunolstein, C.; Bistoni, F. Role of tumor necrosis factor alpha, interleukin-1β, and interleukin-6 in a mouse model of group B streptococcal arthritis. Infection and immunity 1999, 67, 4545–4550. [Google Scholar] [CrossRef]
- Ismail, N.; Stevenson, H.L.; Walker, D.H. Role of tumor necrosis factor alpha (TNF-α) and interleukin-10 in the pathogenesis of severe murine monocytotropic ehrlichiosis: increased resistance of TNF receptor p55-and p75-deficient mice to fatal ehrlichial infection. Infection and immunity 2006, 74, 1846–1856. [Google Scholar] [CrossRef] [PubMed]
- Yap, S.; Moshage, H.; Hazenberg, B.; Roelofs, H.; Bijzet, J.; Limburg, P.; Aarden, L.; Van Rijswijk, M. Tumor necrosis factor (TNF) inhibits interleukin (IL)-1 and/or IL-6 stimulated synthesis of C-reactive protein (CRP) and serum amyloid A (SAA) in primary cultures of human hepatocytes. Biochimica et Biophysica Acta (BBA)-Molecular Cell Research 1991, 1091, 405–408. [Google Scholar] [CrossRef]
- Qin, B.; Zhou, Z.; He, J.; Yan, C.; Ding, S. IL-6 inhibits starvation-induced autophagy via the STAT3/Bcl-2 signaling pathway. Scientific reports 2015, 5, 15701. [Google Scholar] [CrossRef]
- Gabellini, C.; Gómez-Abenza, E.; Ibáñez-Molero, S.; Tupone, M.G.; Pérez-Oliva, A.B.; de Oliveira, S.; Del Bufalo, D.; Mulero, V. Interleukin 8 mediates bcl-xL-induced enhancement of human melanoma cell dissemination and angiogenesis in a zebrafish xenograft model. International journal of cancer 2018, 142, 584–596. [Google Scholar] [CrossRef]
- Guruprasath, P.; Kim, J.; Gunassekaran, G.R.; Chi, L.; Kim, S.; Park, R.W.; Kim, S.H.; Baek, M.C.; Bae, S.M.; Kim, S.Y.; et al. Interleukin-4 receptor-targeted delivery of Bcl-xL siRNA sensitizes tumors to chemotherapy and inhibits tumor growth. Biomaterials 2017, 142, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Maraskovsky, E.; O’Reilly, L.A.; Teepe, M.; Corcoran, L.M.; Peschon, J.J.; Strasser, A. Bcl-2 can rescue t lymphocyte development in interleukin-7 receptor–deficient mice but not in mutant rag-1-/- mice. Cell 1997, 89, 1011–1019. [Google Scholar] [CrossRef] [PubMed]
- Akashi, K.; Kondo, M.; von Freeden-Jeffry, U.; Murray, R.; Weissman, I.L. Bcl-2 rescues T lymphopoiesis in interleukin-7 receptor–deficient mice. Cell 1997, 89, 1033–1041. [Google Scholar] [CrossRef] [PubMed]
- Weber-Nordt, R.; Henschler, R.; Schott, E.; Wehinger, J.; Behringer, D.; Mertelsmann, R.; Finke, J. Interleukin-10 increases Bcl-2 expression and survival in primary human CD34+ hematopoietic progenitor cells. Blood 1996, 88, 2549–2558. [Google Scholar] [CrossRef]
- Qin, J.Z.; Zhang, C.L.; Kamarashev, J.; Dummer, R.; Burg, G.; Döbbeling, U. Interleukin-7 and interleukin-15 regulate the expression of thebcl-2 and c-myb genes in cutaneous T-cell lymphoma cells. Blood 2001, 98, 2778–2783. [Google Scholar] [CrossRef]
- Escandell, J.; Recio, M.; Giner, R.; Máñez, S.; Ríos, J. Bcl-2 is a negative regulator of interleukin-1β secretion in murine macrophages in pharmacological-induced apoptosis. British journal of pharmacology 2010, 160, 1844–1856. [Google Scholar] [CrossRef]
- Alas, S.; Emmanouilides, C.; Bonavida, B. Inhibition of interleukin 10 by rituximab results in down-regulation of bcl-2 and sensitization of B-cell non-Hodgkin’s lymphoma to apoptosis. Clinical Cancer Research 2001, 7, 709–723. [Google Scholar]
- Deng, Z.; Fu, H.; Xiao, Y.; Zhang, B.; Sun, G.; Wei, Q.; Ai, B.; Hu, Q. Effects of selenium on lead-induced alterations in Aβ production and Bcl-2 family proteins. Environmental toxicology and pharmacology 2015, 39, 221–228. [Google Scholar] [CrossRef]
- Yaming, W.; Hai, B.; Haijian, Y.; Xiyan, Z. Effects of selenium dioxide on apoptosis, Bcl-2 and p53 expression, intracellular reactive oxygen species and calcium level in three human lung cancer cell lines. The Chinese-German Journal of Clinical Oncology 2004, 3, 141–146. [Google Scholar]
- Mauro, M.; Sartori, D.; Oliveira, R.J.; Ishii, P.L.; Mantovani, M.S.; Ribeiro, L.R. Activity of selenium on cell proliferation, cytotoxicity, and apoptosis and on the expression of CASP9, BCL-XL and APC in intestinal adenocarcinoma cells. Mutation Research/Fundamental and Molecular Mechanisms of Mutagenesis 2011, 715, 7–12. [Google Scholar] [CrossRef]
- Hemann, M.; Lowe, S. The p53–Bcl-2 connection. 2006. [Google Scholar] [CrossRef] [PubMed]
- Tomita, Y.; Marchenko, N.; Erster, S.; Nemajerova, A.; Dehner, A.; Klein, C.; Pan, H.; Kessler, H.; Pancoska, P.; Moll, U.M. WT p53, but not tumor-derived mutants, bind to Bcl2 via the DNA binding domain and induce mitochondrial permeabilization. Journal of Biological Chemistry 2006, 281, 8600–8606. [Google Scholar] [CrossRef]
- Jiang, M.; Milner, J. Bcl-2 constitutively suppresses p53-dependent apoptosis in colorectal cancer cells. Genes & development 2003, 17, 832–837. [Google Scholar]
- Li, X.; Miao, X.; Wang, H.; Xu, Z.; Li, B. The tissue dependent interactions between p53 and Bcl-2 in vivo. Oncotarget 2015, 6, 35699. [Google Scholar] [CrossRef] [PubMed]
- Pan, R.; Ruvolo, V.; Mu, H.; Leverson, J.D.; Nichols, G.; Reed, J.C.; Konopleva, M.; Andreeff, M. Synthetic lethality of combined Bcl-2 inhibition and p53 activation in AML: mechanisms and superior antileukemic efficacy. Cancer Cell 2017, 32, 748–760. [Google Scholar] [CrossRef]
- Zaidi, A.U.; McDonough, J.S.; Klocke, B.J.; Latham, C.B.; Korsmeyer, S.J.; Flavell, R.A.; Schmidt, R.E.; Roth, K.A. Chloroquine-induced neuronal cell death is p53 and Bcl-2 family-dependent but caspase-independent. Journal of Neuropathology & Experimental Neurology 2001, 60, 937–945. [Google Scholar]
- Wilson, W.H.; Teruya-Feldstein, J.; Fest, T.; Harris, C.; Steinberg, S.M.; Jaffe, E.S.; Raffeld, M. Relationship of p53, bcl-2, and tumor proliferation to clinical drug resistance in non-Hodgkin’s lymphomas. Blood 1997, 89, 601–609. [Google Scholar] [CrossRef]
- Exley, G.E.; Tang, C.; McElhinny, A.S.; Warner, C.M. Expression of caspase and BCL-2 apoptotic family members in mouse preimplantation embryos. Biology of reproduction 1999, 61, 231–239. [Google Scholar] [CrossRef]
- Swanton, E.; Savory, P.; Cosulich, S.; Clarke, P.; Woodman, P. Bcl-2 regulates a caspase-3/caspase-2 apoptotic cascade in cytosolic extracts. Oncogene 1999, 18, 1781. [Google Scholar] [CrossRef]
- Pellegrini, M.; Strasser, A. Caspases, Bcl-2 family proteins and other components of the death machinery: their role in the regulation of the immune response. In Madame Curie Bioscience Database [Internet]; Landes Bioscience, 2013. [Google Scholar]
- Moriishi, K.; Huang, D.C.; Cory, S.; Adams, J.M. Bcl-2 family members do not inhibit apoptosis by binding the caspase activator Apaf-1. Proceedings of the National Academy of Sciences 1999, 96, 9683–9688. [Google Scholar] [CrossRef]
- Demirag, F.; Cakir, E.; Bayiz, H.; Eren Yazici, U. MUC1 and bcl-2 expression in preinvasive lesions and adenosquamous carcinoma of the lung. Acta Chirurgica Belgica 2013, 113, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Sheng, Y.H.; He, Y.; Hasnain, S.Z.; Wang, R.; Tong, H.; Clarke, D.T.; Lourie, R.; Oancea, I.; Wong, K.; Lumley, J.W.; et al. MUC13 protects colorectal cancer cells from death by activating the NF-κB pathway and is a potential therapeutic target. Oncogene 2017, 36, 700. [Google Scholar] [CrossRef]
- Deng, M.; Yuan, H.; Liu, S.; Hu, Z.; Xiao, H. Exosome-transmitted LINC00461 promotes multiple myeloma cell proliferation and suppresses apoptosis by modulating microRNA/BCL-2 expression. Cytotherapy 2019, 21, 96–106. [Google Scholar] [CrossRef]
- Xu, X.D.; Wu, X.H.; Fan, Y.R.; Tan, B.; Quan, Z.; Luo, C.L. Exosome-derived microRNA-29c induces apoptosis of BIU-87 cells by down regulating BCL-2 and MCL-1. Asian Pacific Journal of Cancer Prevention 2014, 15, 3471–3476. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Wu, X.H.; Wang, D.; Luo, C.L.; Chen, L.X. Bladder cancer cell-derived exosomes inhibit tumor cell apoptosis and induce cell proliferation in vitro. Molecular medicine reports 2013, 8, 1272–1278. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, D.; Zhuang, Y.; Fu, J.; Li, X.; Shi, Q.; Ju, X. Exosomes derived from bone marrow stromal cells decrease the sensitivity of leukemic cells to etoposide. Oncology letters 2017, 14, 3082–3088. [Google Scholar] [CrossRef]
- Brown, M.C.; Holl, E.K.; Boczkowski, D.; Dobrikova, E.; Mosaheb, M.; Chandramohan, V.; Bigner, D.D.; Gromeier, M.; Nair, S.K. Cancer immunotherapy with recombinant poliovirus induces IFN-dominant activation of dendritic cells and tumor antigen–specific CTLs. Science translational medicine 2017, 9, eaan4220. [Google Scholar] [CrossRef]
- Stamm, H.; Klingler, F.; Grossjohann, E.M.; Muschhammer, J.; Vettorazzi, E.; Heuser, M.; Mock, U.; Thol, F.; Vohwinkel, G.; Latuske, E.; et al. Immune checkpoints PVR and PVRL2 are prognostic markers in AML and their blockade represents a new therapeutic option. Oncogene 2018, 37, 5269. [Google Scholar] [CrossRef]
- Stamm, H.; Klingler, F.; Pende, D.; Vettorazzi, E.; Heuser, M.; Mock, U.; Bokemeyer, C.; Kischel, R.; Stienen, S.; Friedrich, M.; et al. Expression of Novel Immune Checkpoint Molecules PVR and PVRL2 Confers a Negative Prognosis to Patients with Acute Myeloid Leukemia and Their Blockade Augments T-Cell Mediated Lysis of AML Cells Alone or in Combination with the BiTE® Antibody Construct AMG 330. Blood 2015, 789. [Google Scholar]
- Whelan, S.; Ophir, E.; Kotturi, M.F.; Levy, O.; Ganguly, S.; Leung, L.; Vaknin, I.; Kumar, S.; Dassa, L.; Hansen, K.; et al. PVRIG and PVRL2 Are Induced in Cancer and Inhibit CD8+ T-cell Function. Cancer immunology research 2019, 7, 257–268. [Google Scholar] [CrossRef]
- Bai, M.; Li, W.; Yu, N.; Zhang, H.; Long, F.; Zeng, A. The crosstalk between β-catenin signaling and type I, type II and type III interferons in lung cancer cells. American journal of translational research 2017, 9, 2788. [Google Scholar] [PubMed]
- Hillesheim, A.; Nordhoff, C.; Boergeling, Y.; Ludwig, S.; Wixler, V. β-catenin promotes the type I IFN synthesis and the IFN-dependent signaling response but is suppressed by influenza A virus-induced RIG-I/NF-κB signaling. Cell communication and signaling 2014, 12, 29. [Google Scholar] [CrossRef] [PubMed]
- Ohsugi, T.; Yamaguchi, K.; Zhu, C.; Ikenoue, T.; Furukawa, Y. Decreased expression of interferon-induced protein 2 (IFIT2) by Wnt/β-catenin signaling confers anti-apoptotic properties to colorectal cancer cells. Oncotarget 2017, 8, 100176. [Google Scholar] [CrossRef] [PubMed]
- Pavlova, N.N.; Pallasch, C.; Elia, A.E.; Braun, C.J.; Westbrook, T.F.; Hemann, M.; Elledge, S.J. A role for PVRL4-driven cell–cell interactions in tumorigenesis. Elife 2013, 2, e00358. [Google Scholar] [CrossRef]
- Hynes, R.O. Integrins: bidirectional, allosteric signaling machines. cell 2002, 110, 673–687. [Google Scholar] [CrossRef]
- Fuchs, A.; Cella, M.; Giurisato, E.; Shaw, A.S.; Colonna, M. Cutting edge: CD96 (tactile) promotes NK cell-target cell adhesion by interacting with the poliovirus receptor (CD155). The Journal of Immunology 2004, 172, 3994–3998. [Google Scholar] [CrossRef]
- Wikipedia contributors. Ferroptosis — Wikipedia, The Free Encyclopedia. 2019. Available online: https://en.wikipedia.org/w/index.php?title=Ferroptosis&oldid=882553314 (accessed on 11 May 2019).
- Brown, C.W.; Amante, J.J.; Mercurio, A.M. Cell clustering mediated by the adhesion protein PVRL4 is necessary for α6β4 integrin–promoted ferroptosis resistance in matrix-detached cells. Journal of Biological Chemistry 2018, 293, 12741–12748. [Google Scholar] [CrossRef]
- Abdullah, H.; Brankin, B.; Brady, C.; Cosby, S.L. Wild-type Measles Virus Infection Upregulates Poliovirus Receptor-Related 4 and Causes Apoptosis in Brain Endothelial Cells by Induction of Tumor Necrosis Factor-Related Apoptosis-lnducing Ligand. Journal of Neuropathology & Experimental Neurology 2013, 72, 681–696. [Google Scholar]
- Fabre-Lafay, S.; Garrido-Urbani, S.; Reymond, N.; Gonçalves, A.; Dubreuil, P.; Lopez, M. Nectin-4, a new serological breast cancer marker, is a substrate for tumor necrosis factor-α-converting enzyme (TACE)/ADAM-17. Journal of Biological Chemistry 2005, 280, 19543–19550. [Google Scholar] [CrossRef]
- Besschetnova, T.Y.; Ichimura, T.; Katebi, N.; Croix, B.S.; Bonventre, J.V.; Olsen, B.R. Regulatory mechanisms of anthrax toxin receptor 1-dependent vascular and connective tissue homeostasis. Matrix Biology 2015, 42, 56–73. [Google Scholar] [CrossRef]
- Nanda, A.; Carson-Walter, E.B.; Seaman, S.; Barber, T.D.; Stampfl, J.; Singh, S.; Vogelstein, B.; Kinzler, K.W.; Croix, B.S. TEM8 interacts with the cleaved C5 domain of collagen α3 (VI). Cancer Research 2004, 64, 817–820. [Google Scholar] [CrossRef] [PubMed]
- Hotchkiss, K.A.; Basile, C.M.; Spring, S.C.; Bonuccelli, G.; Lisanti, M.P.; Terman, B.I. TEM8 expression stimulates endothelial cell adhesion and migration by regulating cell–matrix interactions on collagen. Experimental cell research 2005, 305, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Bell, S.E.; Mavila, A.; Salazar, R.; Bayless, K.J.; Kanagala, S.; Maxwell, S.A.; Davis, G.E. Differential gene expression during capillary morphogenesis in 3D collagen matrices: regulated expression of genes involved in basement membrane matrix assembly, cell cycle progression, cellular differentiation and G-protein signaling. Journal of cell science 2001, 114, 2755–2773. [Google Scholar] [CrossRef] [PubMed]
- Bürgi, J.; Kunz, B.; Abrami, L.; Deuquet, J.; Piersigilli, A.; Scholl-Bürgi, S.; Lausch, E.; Unger, S.; Superti-Furga, A.; Bonaldo, P.; et al. CMG2/ANTXR2 regulates extracellular collagen VI which accumulates in hyaline fibromatosis syndrome. Nature communications 2017, 8, 15861. [Google Scholar] [CrossRef]
- Shieh, J.T.; Swidler, P.; Martignetti, J.A.; Ramirez, M.C.M.; Balboni, I.; Kaplan, J.; Kennedy, J.; Abdul-Rahman, O.; Enns, G.M.; Sandborg, C.; et al. Systemic hyalinosis: a distinctive early childhood–onset disorder characterized by mutations in the anthrax toxin receptor 2 gene (ANTRX2). Pediatrics 2006, 118, e1485–e1492. [Google Scholar] [CrossRef]
- Werner, E.; Kowalczyk, A.P.; Faundez, V. Anthrax toxin receptor 1/tumor endothelium marker 8 mediates cell spreading by coupling extracellular ligands to the actin cytoskeleton. Journal of Biological Chemistry 2006, 281, 23227–23236. [Google Scholar] [CrossRef]
- Go, M.Y.; Chow, E.M.; Mogridge, J. The cytoplasmic domain of anthrax toxin receptor 1 affects binding of the protective antigen. Infection and immunity 2009, 77, 52–59. [Google Scholar] [CrossRef]
- Scobie, H.M.; Rainey, G.J.A.; Bradley, K.A.; Young, J.A. Human capillary morphogenesis protein 2 functions as an anthrax toxin receptor. Proceedings of the National Academy of Sciences 2003, 100, 5170–5174. [Google Scholar] [CrossRef]
- Löffek, S.; Schilling, O.; Franzke, C.W. Biological role of matrix metalloproteinases: a critical balance, 2011. European Respiratory Journal 2011, 38, 191–208. [Google Scholar] [CrossRef]
- Wikipedia contributors. Matrix metallopeptidase — Wikipedia, The Free Encyclopedia. 2019. Available online: https://en.wikipedia.org/w/index.php?title=Matrix_metallopeptidase&oldid=893034699 (accessed on 11 May 2019).
- Abrami, L.; Kunz, B.; Deuquet, J.; Bafico, A.; Davidson, G.; Van Der Goot, F.G. Functional interactions between anthrax toxin receptors and the WNT signalling protein LRP6. Cellular microbiology 2008, 10, 2509–2519. [Google Scholar] [CrossRef]
- Wei, W.; Lu, Q.; Chaudry, G.J.; Leppla, S.H.; Cohen, S.N. The LDL receptor-related protein LRP6 mediates internalization and lethality of anthrax toxin. Cell 2006, 124, 1141–1154. [Google Scholar] [CrossRef] [PubMed]
- Verma, K.; Gu, J.; Werner, E. Tumor endothelial marker 8 amplifies canonical Wnt signaling in blood vessels. PloS one 2011, 6, e22334. [Google Scholar] [CrossRef]
- Lee, H.S.; Lee, S.Y.; Rajasekaran, N.; Joe, H.E.; Shin, Y.K.; Kim, S.M.; Park, S.G.; Kang, T.J.; Kim, J.C. Human aortic endothelial cells compare favourably with macrophages for the study of anthrax toxins. International Journal of Nanotechnology 2013, 10. [Google Scholar] [CrossRef]
- Madan, B.; Ke, Z.; Harmston, N.; Ho, S.Y.; Frois, A.; Alam, J.; Jeyaraj, D.A.; Pendharkar, V.; Ghosh, K.; Virshup, I.H.; et al. Wnt addiction of genetically defined cancers reversed by PORCN inhibition. Oncogene 2016, 35, 2197. [Google Scholar] [CrossRef]
- Faivre, R.; Iooss, B.; Mahévas, S.; Makowski, D.; Monod, H. Analyse de sensibilité et exploration de modèles: application aux sciences de la nature et de l’environnement; Editions Quae; 2013. [Google Scholar]
- Iooss, B.; Lemaître, P. A review on global sensitivity analysis methods. arXiv 2014, arXiv:1404.2405. [Google Scholar]
- Borgonovo, E. A new uncertainty importance measure. Reliability Engineering & System Safety 2007, 92, 771–784. [Google Scholar]
- Da Veiga, S. Global sensitivity analysis with dependence measures. Journal of Statistical Computation and Simulation 2015, 85, 1283–1305. [Google Scholar] [CrossRef]
- Gretton, A.; Bousquet, O.; Smola, A.; Schölkopf, B. Measuring statistical dependence with Hilbert-Schmidt norms. Algorithmic learning theory. Springer, 2005; pp. 63–77. [Google Scholar]
- Jansen, M.J. Analysis of variance designs for model output. Computer Physics Communications 1999, 117, 35–43. [Google Scholar] [CrossRef]
- Saltelli, A.; Annoni, P.; Azzini, I.; Campolongo, F.; Ratto, M.; Tarantola, S. Variance based sensitivity analysis of model output. Design and estimator for the total sensitivity index. Computer Physics Communications 2010, 181, 259–270. [Google Scholar] [CrossRef]
- Saltelli, A. Making best use of model evaluations to compute sensitivity indices. Computer Physics Communications 2002, 145, 280–297. [Google Scholar] [CrossRef]
- Saltelli, A.; Annoni, P. How to avoid a perfunctory sensitivity analysis. Environmental Modelling & Software 2010, 25, 1508–1517. [Google Scholar]
- Sobol’, I.M. On sensitivity estimation for nonlinear mathematical models. Matematicheskoe Modelirovanie 1990, 2, 112–118. [Google Scholar]
Figure 1.
Cartoon of Wnt Signaling from [3].
Figure 1.
Cartoon of Wnt Signaling from [3].
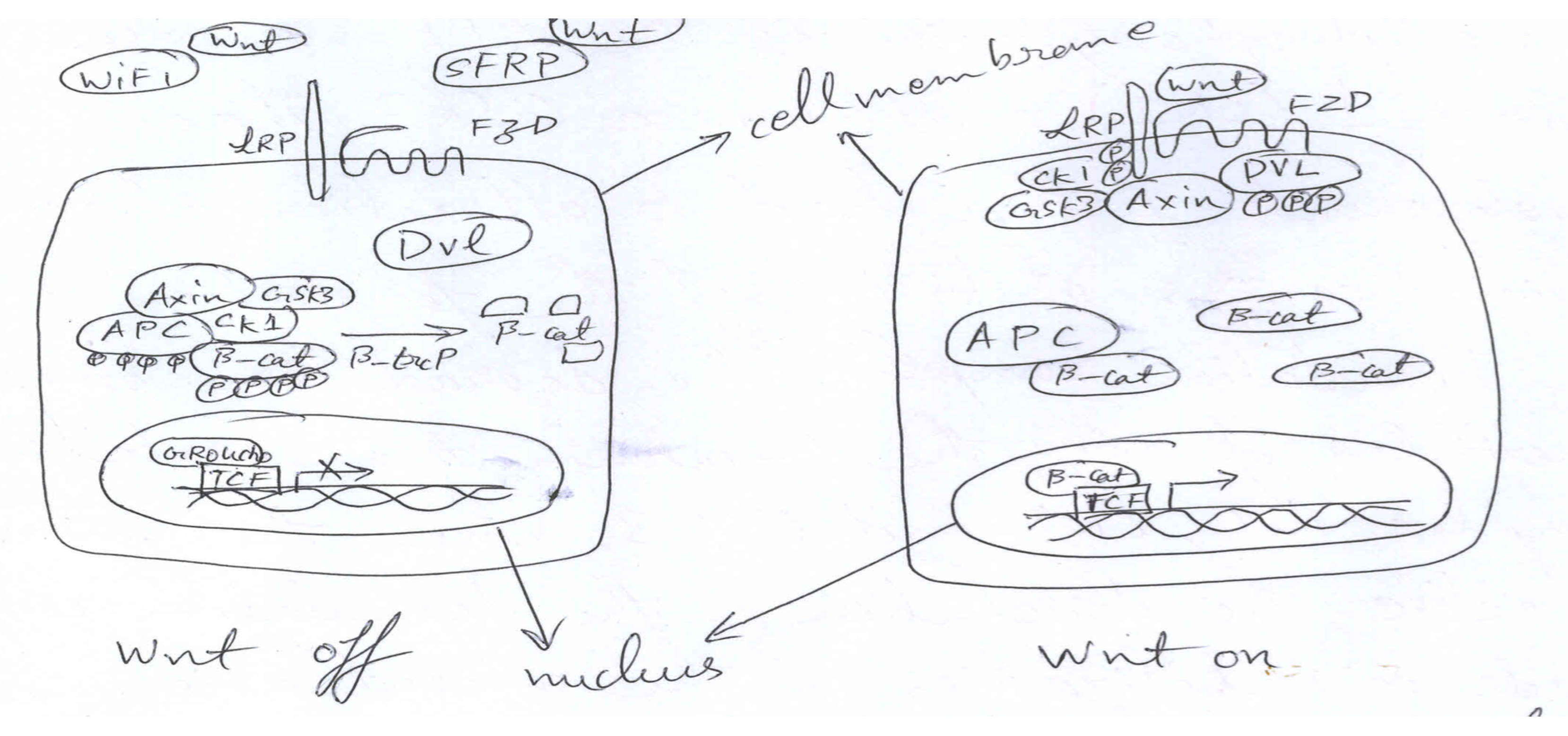
Figure 2.
Cartoon of Wnt Secretion from [3].
Figure 2.
Cartoon of Wnt Secretion from [3].
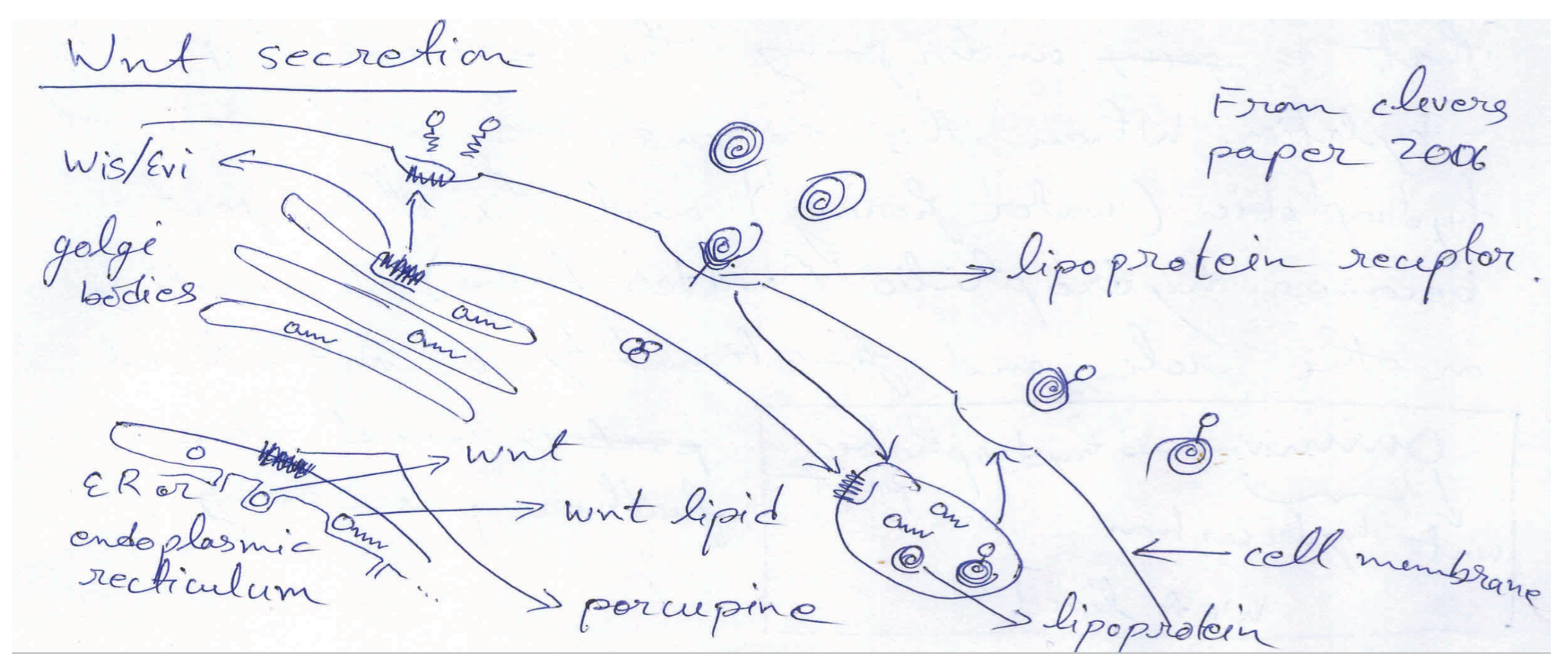
Table 1.
2nd order interaction ranking between ABC w.r.t WNT family members.
| Ranking ABC family w.r.t WNT family | |||||||
| Ranking of ABC family w.r.t WNT-2B | Ranking of ABC family w.r.t WNT4 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| WNT2B - ABC-A5 | 2108 | 310 | 72 | ABC-A5 - WNT4 | 359 | 1285 | 433 |
| ABC-B11 - WNT2B | 319 | 2132 | 18 | ABC-B11 - WNT4 | 872 | 1284 | 867 |
| WNT2B - ABC-C3 | 1853 | 262 | 2498 | ABC-C3 - WNT4 | 10 | 617 | 296 |
| WNT2B - ABC-C5 | 2213 | 1685 | 840 | WNT4 - ABC-C5 | 1383 | 2119 | 215 |
| WNT2B - ABC-C13 | 1149 | 1191 | 2175 | WNT4 - ABC-C13 | 1649 | 1814 | 542 |
| WNT2B - ABC-D1 | 1119 | 177 | 2163 | ABC-D1 - WNT4 | 1041 | 1171 | 1740 |
| WNT2B - ABC-G1 | 1068 | 1583 | 214 | ABC-G1 - WNT4 | 1020 | 1146 | 2025 |
| WNT2B - ABC-G2 | 1500 | 1533 | 172 | ABC-G2 - WNT4 | 784 | 1431 | 435 |
| Ranking of ABC family w.r.t WNT-7B | Ranking of ABC family w.r.t WNT-9A | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| ABC-A5 - WNT7B | 1550 | 516 | 995 | ABC-A5 - WNT9A | 735 | 349 | 1479 |
| ABC-B11 - WNT7B | 968 | 599 | 324 | ABC-B11 - WNT9A | 843 | 1647 | 689 |
| ABC-C3 - WNT7B | 694 | 1668 | 695 | ABC-C3 - WNT9A | 1590 | 359 | 2136 |
| WNT7B - ABC-C5 | 979 | 1715 | 2268 | ABC-C5 - WNT9A | 1295 | 368 | 2265 |
| WNT7B - ABC-C13 | 950 | 2245 | 2298 | ABC-C13 - WNT9A | 1394 | 2294 | 1134 |
| ABC-D1 - WNT7B | 252 | 850 | 1215 | ABC-D1 - WNT9A | 910 | 2367 | 675 |
| ABC-G1 - WNT7B | 269 | 733 | 1160 | ABC-G1 - WNT9A | 426 | 2457 | 1074 |
| ABC-G2 - WNT7B | 1717 | 224 | 264 | ABC-G2 - WNT9A | 1108 | 2350 | 960 |
Table 2.
2nd order interaction ranking between WNT w.r.t ABC family members.
| Ranking WNT family w.r.t ABC family | |||||||
| Ranking of WNT family w.r.t ABC-A5 | Ranking of WNT family w.r.t ABC-B11 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| ABC-A5 - WNT2B | 1549 | 2018 | 2132 | WNT2B - ABC-B11 | 1083 | 703 | 1887 |
| ABC-A5 - WNT4 | 1375 | 2436 | 2449 | WNT4 - ABC-B11 | 156 | 298 | 1517 |
| ABC-A5 - WNT7B | 2420 | 1527 | 460 | WNT7B - ABC-B11 | 1134 | 204 | 2323 |
| ABC-A5 - WNT9A | 1989 | 2209 | 2365 | WNT9A - ABC-B11 | 226 | 2134 | 1480 |
| Ranking of WNT family w.r.t ABC-C3 | Ranking of WNT family w.r.t ABC-C5 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| ABC-C3 - WNT2B | 1127 | 1482 | 1905 | WNT2B - ABC-C5 | 1970 | 2309 | 2248 |
| ABC-C3 - WNT4 | 897 | 1454 | 489 | WNT4 - ABC-C5 | 2129 | 229 | 230 |
| ABC-C3 - WNT7B | 656 | 2080 | 772 | WNT7B - ABC-C5 | 1539 | 756 | 1258 |
| ABC-C3 - WNT9A | 2339 | 1616 | 814 | ABC-C5 - WNT9A | 213 | 2183 | 2480 |
| Ranking of WNT family w.r.t ABC-C13 | Ranking of WNT family w.r.t ABC-D1 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| WNT2B - ABC-C13 | 950 | 2150 | 2048 | WNT2B - ABC-D1 | 1751 | 1370 | 1174 |
| WNT4 - ABC-C13 | 538 | 326 | 2242 | WNT4 - ABC-D1 | 45 | 1784 | 101 |
| WNT7B - ABC-C13 | 2508 | 1830 | 1219 | WNT7B - ABC-D1 | 2238 | 2021 | 1121 |
| WNT9A - ABC-C13 | 738 | 2501 | 634 | WNT9A - ABC-D1 | 732 | 1526 | 1759 |
| Ranking of WNT family w.r.t ABC-G1 | Ranking of WNT family w.r.t ABC-G2 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| WNT2B - ABC-G1 | 318 | 775 | 2040 | WNT2B - ABC-G2 | 1342 | 1987 | 1230 |
| WNT4 - ABC-G1 | 2169 | 157 | 39 | WNT4 - ABC-G2 | 862 | 1352 | 1985 |
| WNT7B - ABC-G1 | 587 | 1808 | 1866 | WNT7B - ABC-G2 | 2334 | 2145 | 1526 |
| WNT9A - ABC-G1 | 856 | 2350 | 920 | WNT9A - ABC-G2 | 1919 | 1284 | 2003 |
Table 3.
2nd order combinatorial hypotheses between ABC and WNT family members.
| Unexplored combinatorial hypotheses | |
| ABC w.r.t WNT | |
| WNT-2B | ABC-C3 |
| WNT-7B | ABC-C13 |
| WNT w.r.t ABC | |
| ABC-A5 | WNT-2B/4/9A |
| WNT-2B/9A | ABC-C5 |
| WNT-2B/7B | ABC-C13 |
| WNT-7B | ABC-D1 |
| WNT-7B | ABC-G1 |
| WNT-7B/9A | ABC-G2 |
Table 4.
2nd order interaction ranking between ABC w.r.t IL family members.
| Ranking IL family VS WNT family | |||||||
| Ranking of IL family w.r.t WNT-2B | Ranking of WNT-2B w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - WNT2B | 6 | 2363 | 924 | IL1A - WNT2B | 2290 | 1360 | 2427 |
| IL1B - WNT2B | 1015 | 1278 | 794 | IL1B - WNT2B | 847 | 2168 | 1369 |
| IL1RAP - WNT2B | 1481 | 1391 | 799 | IL1RAP - WNT2B | 2488 | 35 | 1892 |
| IL1RN - WNT2B | 1229 | 1967 | 1582 | IL1RN - WNT2B | 1307 | 43 | 2514 |
| IL2RG - WNT2B | 1434 | 1100 | 2335 | IL2RG - WNT2B | 1384 | 1255 | 1283 |
| IL6ST - WNT2B | 1157 | 1797 | 2088 | IL6ST - WNT2B | 776 | 242 | 1481 |
| IL8 - WNT2B | 2107 | 1817 | 2251 | IL8 - WNT2B | 2157 | 2025 | 593 |
| IL10RB - WNT2B | 961 | 2494 | 512 | IL10RB - WNT2B | 2419 | 856 | 1419 |
| IL15 - WNT2B | 1008 | 1214 | 1714 | IL15 - WNT2B | 1171 | 625 | 1215 |
| IL15RA - WNT2B | 728 | 1782 | 1382 | IL15RA - WNT2B | 2262 | 1021 | 657 |
| IL17C - WNT2B | 477 | 2357 | 1483 | IL17C - WNT2B | 1947 | 1304 | 1331 |
| IL17REL - WNT2B | 1824 | 12 | 2241 | IL17REL - WNT2B | 1980 | 919 | 1617 |
| Ranking of IL family w.r.t WNT-4 | Ranking of WNT-4 w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - WNT4 | 2500 | 1346 | 955 | IL1A - WNT4 | 507 | 221 | 91 |
| IL1B - WNT4 | 1867 | 1976 | 1682 | IL1B - WNT4 | 129 | 250 | 291 |
| IL1RAP - WNT4 | 2302 | 1826 | 803 | IL1RAP - WNT4 | 74 | 19 | 1553 |
| IL1RN - WNT4 | 1314 | 856 | 104 | IL1RN - WNT4 | 851 | 1218 | 2029 |
| IL2RG - WNT4 | 1289 | 590 | 319 | IL2RG - WNT4 | 520 | 920 | 424 |
| IL6ST - WNT4 | 1315 | 273 | 2422 | IL6ST - WNT4 | 991 | 1443 | 2454 |
| IL8 - WNT4 | 1722 | 549 | 11 | IL8 - WNT4 | 1980 | 2144 | 1267 |
| IL10RB - WNT4 | 1700 | 153 | 1055 | IL10RB - WNT4 | 1828 | 2259 | 1993 |
| IL15 - WNT4 | 1012 | 871 | 1658 | IL15 - WNT4 | 959 | 553 | 448 |
| IL15RA - WNT4 | 1987 | 2265 | 819 | IL15RA - WNT4 | 788 | 139 | 645 |
| IL17C - WNT4 | 2018 | 1639 | 1881 | IL17C - WNT4 | 406 | 276 | 232 |
| IL17REL - WNT4 | 1019 | 425 | 893 | IL17REL - WNT4 | 955 | 595 | 1689 |
| Ranking of IL family w.r.t WNT-7B | Ranking of WNT-7B w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - WNT7B | 662 | 950 | 149 | IL1A - WNT7B | 1058 | 2134 | 2312 |
| IL1B - WNT7B | 290 | 167 | 502 | IL1B - WNT7B | 1683 | 1871 | 1575 |
| IL1RAP - WNT7B | 872 | 1976 | 789 | IL1RAP - WNT7B | 381 | 1728 | 1517 |
| IL1RN - WNT7B | 1882 | 1796 | 503 | IL1RN - WNT7B | 1907 | 2162 | 1605 |
| IL2RG - WNT7B | 1381 | 446 | 482 | IL2RG - WNT7B | 1070 | 1695 | 2245 |
| IL6ST - WNT7B | 819 | 1284 | 1528 | IL6ST - WNT7B | 1268 | 1881 | 2020 |
| IL8 - WNT7B | 2232 | 220 | 701 | IL8 - WNT7B | 1551 | 58 | 2149 |
| IL10RB - WNT7B | 1318 | 1198 | 656 | IL10RB - WNT7B | 375 | 2145 | 803 |
| IL15 - WNT7B | 1000 | 290 | 245 | IL15 - WNT7B | 2307 | 1524 | 1687 |
| IL15RA - WNT7B | 1535 | 1054 | 2204 | IL15RA - WNT7B | 1575 | 191 | 1949 |
| IL17C - WNT7B | 515 | 263 | 113 | IL17C - WNT7B | 1956 | 2388 | 1982 |
| IL17REL - WNT7B | 2053 | 2445 | 2489 | IL17REL - WNT7B | 322 | 859 | 1631 |
| Ranking of IL family w.r.t WNT-9A | Ranking of WNT-9A w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - WNT9A | 199 | 2228 | 1270 | IL1A - WNT9A | 597 | 1322 | 469 |
| IL1B - WNT9A | 305 | 2266 | 466 | IL1B - WNT9A | 776 | 652 | 1010 |
| IL1RAP - WNT9A | 1773 | 2273 | 2159 | IL1RAP - WNT9A | 2003 | 2179 | 964 |
| IL1RN - WNT9A | 2479 | 1506 | 1503 | IL1RN - WNT9A | 1363 | 1829 | 1632 |
| IL2RG - WNT9A | 1489 | 598 | 865 | IL2RG - WNT9A | 186 | 260 | 1276 |
| IL6ST - WNT9A | 2229 | 761 | 1103 | IL6ST - WNT9A | 2099 | 1416 | 1674 |
| IL8 - WNT9A | 346 | 1103 | 1910 | IL8 - WNT9A | 589 | 1751 | 1529 |
| IL10RB - WNT9A | 1836 | 1556 | 1006 | IL10RB - WNT9A | 1021 | 2127 | 1534 |
| IL15 - WNT9A | 168 | 1445 | 855 | IL15 - WNT9A | 1357 | 1025 | 1709 |
| IL15RA - WNT9A | 1776 | 206 | 2380 | IL15RA - WNT9A | 2149 | 2362 | 737 |
| IL17C - WNT9A | 72 | 2442 | 569 | IL17C - WNT9A | 1532 | 2465 | 1607 |
| IL17REL - WNT9A | 2512 | 24 | 580 | IL17REL - WNT9A | 2101 | 1940 | 313 |
Table 5.
2nd order combinatorial hypotheses between IL and WNT family members.
| Unexplored combinatorial hypotheses | |
| IL w.r.t WNT | |
| IL-6ST/8/17REL | WNT-2B |
| IL-1B/1RAP/15RA/17C | WNT-4 |
| IL-1RN/17REL | WNT-7B |
| IL-1RAP/15RA | WNT-9A |
| WNT w.r.t IL | |
| IL-1A/1RAP/8 | WNT-2B |
| IL-8/10RB | WNT-4 |
| IL-1A/1RN/6ST/17C | WNT-7B |
| IL-1RAP/15RA/17REL | WNT-9A |
Table 6.
2nd order interaction ranking between WNT w.r.t UBE2 family members.
| Ranking UBE2 family VS WNT family | |||||||
| Ranking of UBE2-A w.r.t WNT family | Ranking of WNT family w.r.t UBE2-A | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| WNT2B - UBE2A | 1608 | 203 | 181 | WNT2B - UBE2A | 1677 | 899 | 1671 |
| WNT4 - UBE2A | 1293 | 2314 | 2279 | WNT4 - UBE2A | 424 | 1062 | 545 |
| WNT7B - UBE2A | 1139 | 1217 | 1961 | WNT7B - UBE2A | 392 | 2345 | 2151 |
| WNT9A - UBE2A | 443 | 1705 | 287 | WNT9A - UBE2A | 806 | 1581 | 1098 |
| Ranking of UBE2-B w.r.t WNT family | Ranking of WNT family w.r.t UBE2-B | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| WNT2B - UBE2B | 1473 | 2220 | 599 | WNT2B - UBE2B | 2020 | 553 | 73 |
| WNT4 - UBE2B | 2260 | 2008 | 2141 | WNT4 - UBE2B | 301 | 334 | 47 |
| WNT7B - UBE2B | 2116 | 2206 | 1454 | WNT7B - UBE2B | 1336 | 2052 | 1903 |
| WNT9A - UBE2B | 2291 | 79 | 1381 | WNT9A - UBE2B | 2300 | 2476 | 2326 |
| Ranking of UBE2-F w.r.t WNT family | Ranking of WNT family w.r.t UBE2-F | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| WNT2B - UBE2F | 1246 | 833 | 2387 | WNT2B - UBE2F | 1006 | 1917 | 49 |
| WNT4 - UBE2F | 2135 | 2505 | 1762 | WNT4 - UBE2F | 63 | 1109 | 664 |
| WNT7B - UBE2F | 2423 | 1673 | 2077 | WNT7B - UBE2F | 2236 | 1660 | 1751 |
| WNT9A - UBE2F | 2032 | 1165 | 128 | WNT9A - UBE2F | 1014 | 2251 | 2179 |
| Ranking of UBE2-H w.r.t WNT family | Ranking of WNT family w.r.t UBE2-H | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| WNT2B - UBE2H | 1841 | 351 | 2178 | WNT2B - UBE2H | 2015 | 1019 | 1331 |
| WNT4 - UBE2H | 1090 | 778 | 1224 | WNT4 - UBE2H | 218 | 2248 | 2155 |
| WNT7B - UBE2H | 1505 | 1215 | 527 | WNT7B - UBE2H | 2294 | 1209 | 1367 |
| WNT9A - UBE2H | 605 | 332 | 2479 | WNT9A - UBE2H | 437 | 1202 | 2379 |
| Ranking of UBE2-J1 w.r.t WNT family | Ranking of WNT family w.r.t UBE2-J1 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| WNT2B - UBE2J1 | 1539 | 1251 | 1814 | WNT2B - UBE2J1 | 1500 | 1562 | 1255 |
| WNT4 - UBE2J1 | 1583 | 2478 | 1604 | WNT4 - UBE2J1 | 292 | 62 | 65 |
| WNT7B - UBE2J1 | 2349 | 1207 | 2183 | WNT7B - UBE2J1 | 552 | 1877 | 1846 |
| WNT9A - UBE2J1 | 1835 | 2053 | 1652 | WNT9A - UBE2J1 | 2471 | 2137 | 2469 |
| Ranking of UBE2-Z w.r.t WNT family | Ranking of WNT family w.r.t UBE2-Z | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| WNT2B - UBE2Z | 58 | 1756 | 1878 | WNT2B - UBE2Z | 1576 | 1171 | 1543 |
| WNT4 - UBE2Z | 2195 | 2468 | 938 | WNT4 - UBE2Z | 896 | 132 | 186 |
| WNT7B - UBE2Z | 2343 | 1973 | 723 | WNT7B - UBE2Z | 1972 | 1800 | 1399 |
| WNT9A - UBE2Z | 136 | 1986 | 4 | WNT9A - UBE2Z | 1149 | 865 | 813 |
Table 8.
2nd order interaction ranking between WNT w.r.t EXOSC family members.
| Ranking EXOSC family VS WNT10B | |||||||
| Ranking of WNT10B w.r.t EXOSC family | Ranking of EXOSC family w.r.t WNT10B | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| EXOSC2 - WNT10B | 221 | 433 | 699 | EXOSC2 - WNT10B | 1695 | 1077 | 992 |
| EXOSC3 - WNT10B | 906 | 1292 | 860 | EXOSC3 - WNT10B | 610 | 2496 | 2428 |
| EXOSC5 - WNT10B | 919 | 484 | 997 | EXOSC5 - WNT10B | 832 | 1445 | 1589 |
| EXOSC6 - WNT10B | 407 | 1195 | 1747 | EXOSC6 - WNT10B | 1319 | 1738 | 1689 |
| EXOSC7 - WNT10B | 2599 | 2571 | 2584 | EXOSC7 - WNT10B | 2710 | 13 | 4 |
| EXOSC8 - WNT10B | 336 | 1437 | 391 | EXOSC8 - WNT10B | 451 | 2284 | 2493 |
| EXOSC9 - WNT10B | 222 | 701 | 732 | EXOSC9 - WNT10B | 1378 | 1501 | 1651 |
Table 9.
2nd order combinatorial hypotheses between EXOSC and WNT10B family members.
| Unexplored combinatorial hypotheses | |
| EXOSC w.r.t WNT10B | |
| EXOSC-2/5/6/7/9 | WNT10B |
| WNT10B w.r.t EXOSC | |
| EXOSC-2/3/5/6/8/9 | WNT10B |
Table 12.
2nd order interaction ranking between WNT VS TP53 family members.
| Ranking TP53 family VS WNT | |||||||
| Ranking of TP53 family w.r.t WNT2B | Ranking of WNT2B w.r.t TP53 family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| TP53BP2 - WNT2B | 2286 | 234 | 1550 | TP53BP2 - WNT2B | 313 | 908 | 2457 |
| TP53I3 - WNT2B | 2056 | 1712 | 1461 | TP53I3 - WNT2B | 713 | 1223 | 1720 |
| TP53INP1 - WNT2B | 945 | 1805 | 2056 | TP53INP1 - WNT2B | 1853 | 2089 | 762 |
| TP53INP2 - WNT2B | 369 | 1277 | 453 | TP53INP2 - WNT2B | 754 | 1723 | 2335 |
| Ranking of TP53 family w.r.t WNT4 | Ranking of WNT4 w.r.t TP53 family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| TP53BP2 - WNT4 | 1034 | 315 | 1734 | TP53BP2 - WNT4 | 678 | 1464 | 2500 |
| TP53I3 - WNT4 | 1738 | 1631 | 232 | TP53I3 - WNT4 | 297 | 319 | 493 |
| TP53INP1 - WNT4 | 645 | 498 | 450 | TP53INP1 - WNT4 | 131 | 2414 | 2493 |
| TP53INP2 - WNT4 | 671 | 1440 | 405 | TP53INP2 - WNT4 | 529 | 467 | 154 |
| Ranking of TP53 family w.r.t WNT7B | Ranking of WNT7B w.r.t TP53 family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| TP53BP2 - WNT7B | 2333 | 1282 | 1673 | TP53BP2 - WNT7B | 1442 | 2217 | 1068 |
| TP53I3 - WNT7B | 324 | 712 | 284 | TP53I3 - WNT7B | 1712 | 1988 | 2393 |
| TP53INP1 - WNT7B | 1227 | 1585 | 1019 | TP53INP1 - WNT7B | 1226 | 1685 | 1497 |
| TP53INP2 - WNT7B | 845 | 1004 | 470 | TP53INP2 - WNT7B | 1017 | 1746 | 1925 |
| Ranking of TP53 family w.r.t WNT9A | Ranking of WNT9A w.r.t TP53 family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| TP53BP2 - WNT9A | 908 | 2232 | 2143 | TP53BP2 - WNT9A | 1035 | 371 | 1218 |
| TP53I3 - WNT9A | 1707 | 2297 | 1018 | TP53I3 - WNT9A | 1351 | 1281 | 1695 |
| TP53INP1 - WNT9A | 447 | 243 | 1245 | TP53INP1 - WNT9A | 295 | 2045 | 2437 |
| TP53INP2 - WNT9A | 22 | 2497 | 1138 | TP53INP2 - WNT9A | 421 | 1765 | 1121 |
Table 13.
2nd order combinatorial hypotheses between TP53 and WNT family members.
| Unexplored combinatorial hypotheses | |
| TP53 family w.r.t WNT | |
| TP53I3 | WNT2B |
| TP53INP1 | WNT2B |
| TP53BP2 | WNT9A |
| WNT family w.r.t TP53 | |
| TP53INP1 | WNT2B |
| TP53INP2 | WNT2B |
| TP53INP1 | WNT4 |
| TP53I3 | WNT7B |
| TP53INP1 | WNT9A |
Table 21.
2nd order combinatorial hypotheses between MUC and RIPK.
| Unexplored combinatorial hypotheses | |
| MUC w.r.t RIKP family | |
| MUC1 | RIPK1 |
| MUC3A | RIPK3 |
| MUC12 | RIPK4 |
| MUC20 | RIPK3 |
| RIPK w.r.t MUC family | |
| MUC1 | RIPK1/RIPK2 |
| MUC4 | RIPK4 |
| MUC17 | RIPK4 |
| MUC20 | RIPK2 |
Table 22.
2nd order interaction ranking between TNF w.r.t NFkB-2/I family members.
| Ranking TNF family w.r.t NFkB-2/I family | |||||||
| Ranking of TNF family w.r.t NFkB2 | Ranking of TNF family w.r.t NFkBI-A | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| NFkB2 - TNF | 1620 | 615 | 1897 | NFkBI-A - TNF | 820 | 1495 | 1109 |
| NFkB2 - TNF-AIP1 | 324 | 649 | 1387 | NFkBI-A - TNF-AIP1 | 1779 | 1904 | 1400 |
| NFkB2 - TNF-AIP2 | 1437 | 715 | 1986 | NFkBI-A - TNF-AIP2 | 1247 | 217 | 766 |
| NFkB2 - TNF-AIP3 | 1272 | 1574 | 441 | NFkBI-A - TNF-AIP3 | 776 | 981 | 212 |
| NFkB2 - TNF-RSF1A | 30 | 2465 | 575 | NFkBI-A - TNF-RSF1A | 1580 | 1422 | 43 |
| NFkB2 - TNF-RSF10A | 2095 | 817 | 2509 | NFkBI-A - TNF-RSF10A | 2499 | 1438 | 2191 |
| NFkB2 - TNF-RSF10B | 37 | 1411 | 250 | NFkBI-A - TNF-RSF10B | 2075 | 1555 | 1401 |
| NFkB2 - TNF-RSF10D | 2473 | 12 | 1499 | NFkBI-A - TNF-RSF10D | 2498 | 2344 | 2501 |
| NFkB2 - TNF-RSF12A | 1813 | 824 | 1893 | NFkBI-A - TNF-RSF12A | 2337 | 1101 | 1491 |
| NFkB2 - TNF-RSF14 | 1799 | 834 | 302 | NFkBI-A - TNF-RSF14 | 1974 | 2045 | 1136 |
| NFkB2 - TNF-RSF21 | 332 | 1973 | 1719 | NFkBI-A - TNF-RSF21 | 1119 | 951 | 903 |
| NFkB2 - TNF-SF10 | 1627 | 1614 | 1299 | NFkBI-A - TNF-SF10 | 2185 | 499 | 2316 |
| NFkB2 - TNF-SF15 | 564 | 2437 | 1064 | NFkBI-A - TNF-SF15 | 564 | 1684 | 1473 |
| Ranking of TNF family w.r.t NFkBI-E | Ranking of TNF family w.r.t NFkBI-Z | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| NFkBI-E - TNF | 2443 | 925 | 228 | NFkBI-Z - TNF | 851 | 776 | 850 |
| NFkBI-E - TNF-AIP1 | 1720 | 685 | 971 | NFkBI-Z - TNF-AIP1 | 153 | 397 | 621 |
| NFkBI-E - TNF-AIP2 | 2347 | 1863 | 964 | NFkBI-Z - TNF-AIP2 | 2188 | 432 | 566 |
| NFkBI-E - TNF-AIP3 | 559 | 1663 | 280 | NFkBI-Z - TNF-AIP3 | 775 | 10 | 2362 |
| NFkBI-E - TNF-RSF1A | 846 | 1624 | 176 | NFkBI-Z - TNF-RSF1A | 399 | 2006 | 93 |
| NFkBI-E - TNF-RSF10A | 840 | 359 | 952 | NFkBI-Z - TNF-RSF10A | 1380 | 2004 | 1540 |
| NFkBI-E - TNF-RSF10B | 835 | 2257 | 1294 | NFkBI-Z - TNF-RSF10B | 2204 | 1438 | 1991 |
| NFkBI-E - TNF-RSF10D | 2454 | 1018 | 1566 | NFkBI-Z - TNF-RSF10D | 2214 | 2033 | 2514 |
| NFkBI-E - TNF-RSF12A | 383 | 166 | 1464 | NFkBI-Z - TNF-RSF12A | 1638 | 2370 | 1841 |
| NFkBI-E - TNF-RSF14 | 1877 | 2282 | 1426 | NFkBI-Z - TNF-RSF14 | 1120 | 1505 | 1899 |
| NFkBI-E - TNF-RSF21 | 2129 | 1293 | 831 | NFkBI-Z - TNF-RSF21 | 207 | 804 | 344 |
| NFkBI-E - TNF-SF10 | 890 | 1096 | 1816 | NFkBI-Z - TNF-SF10 | 609 | 1088 | 1344 |
| NFkBI-E - TNF-SF15 | 523 | 1957 | 32 | NFkBI-Z - TNF-SF15 | 1237 | 1375 | 2196 |
Table 23.
2nd order interaction ranking between NFkB-2/I family w.r.t TNF family members.
| Ranking NFkB-2/I family w.r.t TNF family | |||||||
| Ranking of NFkB-2/I family w.r.t TNF | Ranking of NFkB-2/I family w.r.t TNF-AIP1 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| NFkB-2 - TNF | 1632 | 989 | 1453 | NFkB-2 - TNF-AIP1 | 2027 | 1807 | 1140 |
| NFkBI-A - TNF | 904 | 561 | 658 | NFkBI-A - TNF-AIP1 | 2072 | 349 | 1218 |
| NFkBI-E - TNF | 2116 | 1247 | 803 | NFkBI-E - TNF-AIP1 | 56 | 420 | 1551 |
| NFkBI-Z - TNF | 691 | 51 | 265 | NFkBI-Z - TNF-AIP1 | 499 | 1648 | 646 |
| Ranking of NFkB-2/I family w.r.t TNF-AIP2 | Ranking of NFkB-2/I family w.r.t TNF-AIP3 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| NFkB-2 - TNF-AIP2 | 2077 | 1027 | 2224 | NFkB-2 - TNF-AIP3 | 1042 | 2336 | 2130 |
| NFkBI-A - TNF-AIP2 | 499 | 22 | 1192 | NFkBI-A - TNF-AIP3 | 1452 | 411 | 637 |
| NFkBI-E - TNF-AIP2 | 526 | 1755 | 338 | NFkBI-E - TNF-AIP3 | 711 | 1686 | 2041 |
| NFkBI-Z - TNF-AIP2 | 452 | 988 | 1617 | NFkBI-Z - TNF-AIP3 | 1979 | 886 | 278 |
| Ranking of NFkB-2/I family w.r.t TNF-RSF1A | Ranking of NFkB-2/I family w.r.t TNF-RSF10A | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| NFkB-2 - TNF-RSF1A | 648 | 164 | 990 | NFkB-2 - TNF-RSF10A | 611 | 1007 | 454 |
| NFkBI-A - TNF-RSF1A | 435 | 1454 | 130 | NFkBI-A - TNF-RSF10A | 458 | 190 | 1412 |
| NFkBI-E - TNF-RSF1A | 431 | 980 | 1417 | NFkBI-E - TNF-RSF10A | 1719 | 263 | 374 |
| NFkBI-Z - TNF-RSF1A | 550 | 2213 | 1447 | NFkBI-Z - TNF-RSF10A | 342 | 742 | 732 |
| Ranking of NFkB-2/I w.r.t TNF-RSF10B | Ranking of NFkB-2/I w.r.t TNF-RSF10D | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| NFkB-2 - TNF-RSF10B | 713 | 1408 | 2397 | NFkB-2 - TNF-RSF10D | 123 | 1939 | 543 |
| NFkBI-A - TNF-RSF10B | 1237 | 1054 | 562 | NFkBI-A - TNF-RSF10D | 371 | 948 | 584 |
| NFkBI-E - TNF-RSF10B | 1352 | 931 | 2142 | NFkBI-E - TNF-RSF10D | 2136 | 621 | 1811 |
| NFkBI-Z - TNF-RSF10B | 165 | 2407 | 361 | NFkBI-Z - TNF-RSF10D | 259 | 400 | 1341 |
| Ranking of NFkB-2/I family w.r.t TNF-RSF12A | Ranking of NFkB-2/I family w.r.t TNF-RSF14 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| NFkB-2 - TNF-RSF12A | 250 | 341 | 1232 | NFkB-2 - TNF-RSF14 | 299 | 1253 | 543 |
| NFkBI-A - TNF-RSF12A | 689 | 2225 | 17 | NFkBI-A - TNF-RSF14 | 280 | 1126 | 277 |
| NFkBI-E - TNF-RSF12A | 1188 | 1133 | 765 | NFkBI-E - TNF-RSF14 | 278 | 2025 | 1557 |
| NFkBI-Z - TNF-RSF12A | 973 | 1590 | 2298 | NFkBI-Z - TNF-RSF14 | 131 | 893 | 1953 |
| Ranking of NFkB-2/I family w.r.t TNF-RSF21 | Ranking of NFkB-2/I family w.r.t TNF-SF10 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| NFkB-2 - TNF-RSF21 | 250 | 341 | 1232 | NFkB-2 - TNF-SF10 | 1643 | 496 | 743 |
| NFkBI-A - TNF-RSF21 | 689 | 2225 | 17 | NFkBI-A - TNF-SF10 | 262 | 1238 | 1352 |
| NFkBI-E - TNF-RSF21 | 1188 | 1133 | 765 | NFkBI-E - TNF-SF10 | 985 | 1090 | 158 |
| NFkBI-Z - TNF-RSF21 | 973 | 1590 | 2298 | NFkBI-Z - TNF-SF10 | 537 | 1557 | 2104 |
| Ranking of NFkB-2/I family w.r.t TNF-SF15 | |||||||
| laplace | linear | rbf | |||||
| NFkB-2 - TNF-SF15 | 1521 | 786 | 1211 | ||||
| NFkBI-A - TNF-SF15 | 2367 | 325 | 1079 | ||||
| NFkBI-E - TNF-SF15 | 97 | 1868 | 1195 | ||||
| NFkBI-Z - TNF-SF15 | 774 | 407 | 372 | ||||
Table 28.
2nd order interaction ranking between STAT family w.r.t IKBKE.
| Ranking STAT family vs IKBKE | |||||||
| Ranking of STAT family w.r.t IKBKE family | Ranking of IKBKE w.r.t STAT family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| STAT2 - IKBKE | 1267 | 2033 | 1892 | STAT2 - IKBKE | 1604 | 554 | 2108 |
| STAT3 - IKBKE | 1055 | 2144 | 1672 | STAT3 - IKBKE | 1442 | 2179 | 1976 |
| STAT5A - IKBKE | 178 | 1687 | 1183 | STAT5A - IKBKE | 2085 | 2409 | 2277 |
Table 29.
2nd order combinatorial hypotheses between NFkB-2/I and TNF
| Unexplored combinatorial hypotheses | |
| STAT w.r.t IKBKE | |
| STAT2 | IKBKE |
| IKBKE w.r.t STAT | |
| STAT3 | IKBKE |
| STAT5A | IKBKE |
Table 30.
2nd order interaction ranking between STAT family w.r.t IKBKE.
| Ranking TRAF family vs IKBKE | |||||||
| Ranking of IKBKE w.r.t TRAF family | Ranking of TRAF family w.r.t IKBKE | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| TRAF4 - IKBKE | 1235 | 2158 | 2416 | TRAF4 - IKBKE | 1606 | 461 | 1330 |
| TRAF6 - IKBKE | 1694 | 389 | 1554 | TRAF6 - IKBKE | 2105 | 1376 | 1819 |
| TRAFD1 - IKBKE | 1687 | 532 | 1793 | TRAFD1 - IKBKE | 866 | 733 | 496 |
| TRAF3IP2 - IKBKE | 1349 | 738 | 1987 | TRAF3IP2 - IKBKE | 924 | 1966 | 334 |
Table 31.
2nd order combinatorial hypotheses between NFkB-2/I and TNF
| Unexplored combinatorial hypotheses | |
| TRAF w.r.t IKBKE | |
| TRAF6 | IKBKE |
| IKBKE w.r.t TRAF | |
| TRAF4 | IKBKE |
Table 39.
2nd order combinatorial hypotheses between NFkB-2/I and ABC
| Unexplored combinatorial hypotheses | |
| NFkB-2/I family w.r.t REL-B | |
| NFkB2 | RELB |
| NFKBIZ | RELB |
| REL-A w.r.t NFkB-2/I family | |
| NFkB2 | RELA |
| NFKBIA | RELA |
Table 41.
2nd order combinatorial hypotheses between TNF and WNT family.
| Unexplored combinatorial hypotheses | |
| TNF w.r.t WNT | |
| TNF-RSF1A/RSF10A/RSF10B/SF15 | WNT2B |
| TNF-RSF10A/RSF10D/RSF12A/SF10 | WNT4 |
| TNF-RSF12A/SF10 | WNT7B |
| TNF-RSF21 | WNT9A |
| WNT w.r.t TNF | |
| TNF-RSF10B/RSF10D/RSF14 | WNT2B |
| TNF-AIP3/RSF10B | WNT4 |
| TNF, TNF-RSF1A/RSF14 | WNT7B |
| TNF-AIP2/AIP3/RSF10A/RSF12A/SF10 | WNT9A |
Table 42.
2nd order interaction ranking between TNF w.r.t MUC family members.
| Ranking TNF family w.r.t MUC family | |||||||
| Ranking of TNF family w.r.t MUC1 | Ranking of TNF family w.r.t MUC3A | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| MUC1 - TNF | 112 | 72 | 88 | MUC3A - TNF | 1353 | 1659 | 1479 |
| MUC1 - TNFAIP1 | 1193 | 1603 | 997 | MUC3A - TNFAIP1 | 2178 | 1209 | 1347 |
| MUC1 - TNFAIP2 | 716 | 405 | 2340 | MUC3A - TNFAIP2 | 1075 | 1614 | 1158 |
| MUC1 - TNFAIP3 | 2115 | 1636 | 1882 | MUC3A - TNFAIP3 | 962 | 1020 | 2491 |
| MUC1 - TNFRSF1A | 1380 | 422 | 1390 | MUC3A - TNFRSF1A | 461 | 1708 | 189 |
| MUC1 - TNFRSF10A | 1009 | 2180 | 1095 | MUC3A - TNFRSF10A | 2237 | 1910 | 335 |
| MUC1 - TNFRSF10B | 1923 | 732 | 88 | MUC3A - TNFRSF10B | 450 | 1443 | 2040 |
| MUC1 - TNFRSF10D | 2303 | 2154 | 376 | MUC3A - TNFRSF10D | 1678 | 2049 | 102 |
| MUC1 - TNFRSF12A | 2019 | 2009 | 1700 | MUC3A - TNFRSF12A | 2349 | 1315 | 382 |
| MUC1 - TNFRSF14 | 1955 | 1899 | 1429 | MUC3A - TNFRSF14 | 956 | 1442 | 1953 |
| MUC1 - TNFRSF21 | 337 | 477 | 968 | MUC3A - TNFRSF21 | 1297 | 1492 | 1959 |
| MUC1 - TNFSF10 | 1111 | 1592 | 1198 | MUC3A - TNFSF10 | 891 | 257 | 798 |
| MUC1 - TNFSF15 | 936 | 986 | 2391 | MUC3A - TNFSF15 | 2285 | 795 | 1164 |
| Ranking of TNF family w.r.t MUC4 | Ranking of TNF family w.r.t MUC12 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| MUC4 - TNF | 1896 | 231 | 1355 | MUC12 - TNF | 1862 | 102 | 135 |
| MUC4 - TNFAIP1 | 864 | 397 | 987 | MUC12 - TNFAIP1 | 1386 | 479 | 942 |
| MUC4 - TNFAIP2 | 73 | 1011 | 1087 | MUC12 - TNFAIP2 | 1056 | 303 | 1587 |
| MUC4 - TNFAIP3 | 1159 | 1751 | 179 | MUC12 - TNFAIP3 | 2493 | 1259 | 1330 |
| MUC4 - TNFRSF1A | 179 | 71 | 16 | MUC12 - TNFRSF1A | 1709 | 1440 | 837 |
| MUC4 - TNFRSF10A | 1668 | 1892 | 1652 | MUC12 - TNFRSF10A | 598 | 531 | 363 |
| MUC4 - TNFRSF10B | 2024 | 1396 | 331 | MUC12 - TNFRSF10B | 409 | 1572 | 1297 |
| MUC4 - TNFRSF10D | 2503 | 2403 | 2356 | MUC12 - TNFRSF10D | 30 | 102 | 149 |
| MUC4 - TNFRSF12A | 1684 | 700 | 745 | MUC12 - TNFRSF12A | 298 | 882 | 153 |
| MUC4 - TNFRSF14 | 1675 | 2029 | 1146 | MUC12 - TNFRSF14 | 1749 | 2237 | 135 |
| MUC4 - TNFRSF21 | 647 | 326 | 323 | MUC12 - TNFRSF21 | 1795 | 607 | 2438 |
| MUC4 - TNFSF10 | 936 | 2134 | 1957 | MUC12 - TNFSF10 | 801 | 1795 | 2435 |
| MUC4 - TNFSF15 | 1440 | 1180 | 1627 | MUC12 - TNFSF15 | 1741 | 889 | 1098 |
| Ranking of TNF family w.r.t MUC13 | Ranking of TNF family w.r.t MUC17 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| MUC13 - TNF | 2282 | 220 | 127 | MUC17 - TNF | 683 | 362 | 515 |
| MUC13 - TNFAIP1 | 378 | 230 | 1935 | MUC17 - TNFAIP1 | 117 | 188 | 272 |
| MUC13 - TNFAIP2 | 2464 | 220 | 697 | MUC17 - TNFAIP2 | 1311 | 414 | 351 |
| MUC13 - TNFAIP3 | 2274 | 1233 | 1446 | MUC17 - TNFAIP3 | 1589 | 1547 | 1539 |
| MUC13 - TNFRSF1A | 274 | 2152 | 514 | MUC17 - TNFRSF1A | 428 | 205 | 329 |
| MUC13 - TNFRSF10A | 2500 | 938 | 1844 | MUC17 - TNFRSF10A | 2269 | 2364 | 2005 |
| MUC13 - TNFRSF10B | 1891 | 1497 | 225 | MUC17 - TNFRSF10B | 1199 | 1323 | 2120 |
| MUC13 - TNFRSF10D | 1191 | 2263 | 2294 | MUC17 - TNFRSF10D | 1798 | 1378 | 2302 |
| MUC13 - TNFRSF12A | 460 | 1753 | 1704 | MUC17 - TNFRSF12A | 2041 | 2303 | 1049 |
| MUC13 - TNFRSF14 | 2220 | 1602 | 1359 | MUC17 - TNFRSF14 | 2043 | 825 | 1700 |
| MUC13 - TNFRSF21 | 1612 | 1673 | 127 | MUC17 - TNFRSF21 | 2013 | 393 | 119 |
| MUC13 - TNFSF10 | 2236 | 1598 | 1495 | MUC17 - TNFSF10 | 280 | 1025 | 817 |
| MUC13 - TNFSF15 | 2423 | 1488 | 1292 | MUC17 - TNFSF15 | 833 | 967 | 950 |
| Ranking of TNF family w.r.t MUC20 | |||||||
| laplace | linear | rbf | |||||
| MUC20 - TNF | 2267 | 262 | 145 | ||||
| MUC20 - TNFAIP1 | 1273 | 2296 | 178 | ||||
| MUC20 - TNFAIP2 | 1062 | 598 | 339 | ||||
| MUC20 - TNFAIP3 | 2205 | 435 | 2136 | ||||
| MUC20 - TNFRSF1A | 483 | 2346 | 145 | ||||
| MUC20 - TNFRSF10A | 100 | 2305 | 917 | ||||
| MUC20 - TNFRSF10B | 775 | 1578 | 1556 | ||||
| MUC20 - TNFRSF10D | 200 | 1487 | 799 | ||||
| MUC20 - TNFRSF12A | 318 | 1607 | 2258 | ||||
| MUC20 - TNFRSF14 | 410 | 1832 | 745 | ||||
| MUC20 - TNFRSF21 | 1686 | 2259 | 164 | ||||
| MUC20 - TNFSF10 | 1005 | 2139 | 1548 | ||||
| MUC20 - TNFSF15 | 2493 | 387 | 2108 | ||||
Table 43.
2nd order interaction ranking between MUC w.r.t TNF family members.
| Ranking MUC family w.r.t TNF family | |||||||
| Ranking of MUC1 w.r.t TNF family | Ranking of MUC3A w.r.t TNF family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| MUC1 - TNF | 368 | 142 | 21 | MUC3A - TNF | 1478 | 985 | 2373 |
| MUC1 - TNFAIP1 | 692 | 91 | 1591 | MUC3A - TNFAIP1 | 1485 | 536 | 1698 |
| MUC1 - TNFAIP2 | 2290 | 476 | 398 | MUC3A - TNFAIP2 | 1254 | 1265 | 75 |
| MUC1 - TNFAIP3 | 810 | 492 | 748 | MUC3A - TNFAIP3 | 1844 | 960 | 243 |
| MUC1 - TNFRSF1A | 1089 | 2344 | 2312 | MUC3A - TNFRSF1A | 496 | 574 | 792 |
| MUC1 - TNFRSF10A | 1263 | 351 | 826 | MUC3A - TNFRSF10A | 1315 | 1525 | 1815 |
| MUC1 - TNFRSF10B | 1630 | 1604 | 2103 | MUC3A - TNFRSF10B | 351 | 1920 | 1489 |
| MUC1 - TNFRSF10D | 975 | 1026 | 984 | MUC3A - TNFRSF10D | 596 | 950 | 1016 |
| MUC1 - TNFRSF12A | 1597 | 1811 | 1078 | MUC3A - TNFRSF12A | 436 | 595 | 2124 |
| MUC1 - TNFRSF14 | 739 | 2119 | 938 | MUC3A - TNFRSF14 | 1612 | 1383 | 329 |
| MUC1 - TNFRSF21 | 766 | 1495 | 2322 | MUC3A - TNFRSF21 | 1254 | 1357 | 1162 |
| MUC1 - TNFSF10 | 1360 | 1969 | 477 | MUC3A - TNFSF10 | 774 | 980 | 2053 |
| MUC1 - TNFSF15 | 424 | 1183 | 542 | MUC3A - TNFSF15 | 75 | 1261 | 624 |
| Ranking of MUC4 w.r.t TNF family | Ranking of MUC12 w.r.t TNF family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| MUC4 - TNF | 1656 | 777 | 565 | MUC12 - TNF | 266 | 1223 | 628 |
| MUC4 - TNFAIP1 | 2483 | 895 | 390 | MUC12 - TNFAIP1 | 2321 | 668 | 2457 |
| MUC4 - TNFAIP2 | 1875 | 1792 | 180 | MUC12 - TNFAIP2 | 281 | 1829 | 1913 |
| MUC4 - TNFAIP3 | 54 | 498 | 464 | MUC12 - TNFAIP3 | 2353 | 153 | 576 |
| MUC4 - TNFRSF1A | 1074 | 753 | 68 | MUC12 - TNFRSF1A | 1481 | 1952 | 1406 |
| MUC4 - TNFRSF10A | 683 | 311 | 997 | MUC12 - TNFRSF10A | 445 | 337 | 888 |
| MUC4 - TNFRSF10B | 98 | 1413 | 704 | MUC12 - TNFRSF10B | 792 | 164 | 133 |
| MUC4 - TNFRSF10D | 1916 | 230 | 80 | MUC12 - TNFRSF10D | 167 | 193 | 521 |
| MUC4 - TNFRSF12A | 1321 | 2190 | 150 | MUC12 - TNFRSF12A | 216 | 2093 | 302 |
| MUC4 - TNFRSF14 | 606 | 704 | 1493 | MUC12 - TNFRSF14 | 105 | 59 | 69 |
| MUC4 - TNFRSF21 | 1225 | 1967 | 1093 | MUC12 - TNFRSF21 | 1471 | 1975 | 1769 |
| MUC4 - TNFSF10 | 815 | 1108 | 1906 | MUC12 - TNFSF10 | 662 | 2135 | 2255 |
| MUC4 - TNFSF15 | 1141 | 1841 | 920 | MUC12 - TNFSF15 | 1619 | 2204 | 1257 |
| Ranking of MUC13 w.r.t TNF family | Ranking of MUC17 w.r.t TNF family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| MUC13 - TNF | 623 | 292 | 295 | MUC17 - TNF | 203 | 811 | 57 |
| MUC13 - TNFAIP1 | 823 | 755 | 81 | MUC17 - TNFAIP1 | 381 | 193 | 118 |
| MUC13 - TNFAIP2 | 1118 | 2464 | 116 | MUC17 - TNFAIP2 | 1069 | 822 | 136 |
| MUC13 - TNFAIP3 | 1189 | 546 | 541 | MUC17 - TNFAIP3 | 2132 | 47 | 937 |
| MUC13 - TNFRSF1A | 978 | 1506 | 490 | MUC17 - TNFRSF1A | 120 | 497 | 864 |
| MUC13 - TNFRSF10A | 1180 | 540 | 1926 | MUC17 - TNFRSF10A | 852 | 218 | 346 |
| MUC13 - TNFRSF10B | 280 | 1105 | 190 | MUC17 - TNFRSF10B | 933 | 1667 | 1166 |
| MUC13 - TNFRSF10D | 655 | 725 | 1668 | MUC17 - TNFRSF10D | 546 | 133 | 304 |
| MUC13 - TNFRSF12A | 401 | 1242 | 999 | MUC17 - TNFRSF12A | 18 | 1675 | 86 |
| MUC13 - TNFRSF14 | 1324 | 374 | 389 | MUC17 - TNFRSF14 | 819 | 296 | 1014 |
| MUC13 - TNFRSF21 | 690 | 2337 | 107 | MUC17 - TNFRSF21 | 1659 | 814 | 889 |
| MUC13 - TNFSF10 | 1146 | 1208 | 2159 | MUC17 - TNFSF10 | 387 | 1542 | 156 |
| MUC13 - TNFSF15 | 1633 | 314 | 155 | MUC17 - TNFSF15 | 1207 | 1040 | 522 |
| Ranking of MUC20 w.r.t TNF family | |||||||
| laplace | linear | rbf | |||||
| MUC20 - TNF | 57 | 216 | 903 | ||||
| MUC20 - TNFAIP1 | 1265 | 2266 | 2057 | ||||
| MUC20 - TNFAIP2 | 241 | 2404 | 2157 | ||||
| MUC20 - TNFAIP3 | 484 | 1012 | 513 | ||||
| MUC20 - TNFRSF1A | 748 | 173 | 2193 | ||||
| MUC20 - TNFRSF10A | 620 | 427 | 1054 | ||||
| MUC20 - TNFRSF10B | 765 | 1563 | 790 | ||||
| MUC20 - TNFRSF10D | 509 | 2185 | 794 | ||||
| MUC20 - TNFRSF12A | 216 | 2093 | 302 | ||||
| MUC20 - TNFRSF14 | 2298 | 651 | 1368 | ||||
| MUC20 - TNFRSF21 | 2374 | 1611 | 1140 | ||||
| MUC20 - TNFSF10 | 1257 | 1088 | 1031 | ||||
| MUC20 - TNFSF15 | 142 | 2159 | 7 | ||||
Table 44.
2nd order combinatorial hypotheses between MUC and TNF.
| Unexplored combinatorial hypotheses | |
| TNF w.r.t MUC | |
| MUC1 | TNFAIP3/TNFRSF10D/TNFRSF12A/TNFRSF14 |
| MUC3A | TNFRSF10A/TNFRSF10D |
| MUC4 | TNFRSF10D/TNFSF10 |
| MUC12 | TNFRSF21/TNFSF10 |
| MUC13 | TNFRSF10A/TNFRSF10D |
| MUC17 | TNFRSF10A/TNFRSF10D/TNFRSF12A |
| MUC20 | TNFAIP3/TNFSF15 |
| MUC w.r.t TNF | |
| MUC1 | TNFRSF1A |
| MUC4 | TNFAIP2 |
| MUC12 | TNFAIP1/TNFAIP2/TNFRSF21/TNFSF10 |
| MUC13 | TNFAIP1/TNFAIP2 |
Table 45.
2nd order combinatorial hypotheses between STEAP4 and TNF.
| Ranking TNF family vs STEAP4 family | |||||||
| Ranking of TNF family w.r.t STEAP4 | Ranking of STEAP4 w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| TNF - STEAP4 | 1579 | 1914 | 2130 | TNF - STEAP4 | 1116 | 1482 | 999 |
| TNFAIP1 - STEAP4 | 2189 | 1293 | 1910 | TNFAIP1 - STEAP4 | 691 | 611 | 1105 |
| TNFAIP2 - STEAP4 | 1172 | 2002 | 1840 | TNFAIP2 - STEAP4 | 228 | 1747 | 2463 |
| TNFAIP3 - STEAP4 | 1458 | 1882 | 2197 | TNFAIP3 - STEAP4 | 159 | 727 | 219 |
| TNFRSF1A - STEAP4 | 803 | 75 | 1086 | TNFRSF1A - STEAP4 | 1483 | 408 | 1966 |
| TNFRSF10A - STEAP4 | 239 | 1949 | 339 | TNFRSF10A - STEAP4 | 1512 | 1796 | 2026 |
| TNFRSF10B - STEAP4 | 2210 | 1717 | 1827 | TNFRSF10B - STEAP4 | 565 | 571 | 248 |
| TNFRSF10D - STEAP4 | 510 | 2192 | 1797 | TNFRSF10D - STEAP4 | 1018 | 2339 | 2405 |
| TNFRSF12A - STEAP4 | 757 | 338 | 1497 | TNFRSF12A - STEAP4 | 1495 | 1430 | 581 |
| TNFRSF14 - STEAP4 | 1323 | 1512 | 792 | TNFRSF14 - STEAP4 | 1363 | 1956 | 2256 |
| TNFRSF21 - STEAP4 | 1643 | 1920 | 165 | TNFRSF21 - STEAP4 | 1646 | 802 | 160 |
| TNFSF10 - STEAP4 | 2083 | 544 | 1773 | TNFSF10 - STEAP4 | 845 | 675 | 2468 |
| TNFSF15 - STEAP4 | 631 | 1296 | 1020 | TNFSF15 - STEAP4 | 1558 | 600 | 784 |
Table 50.
2nd order combinatorial hypotheses between BCL and TNF.
| Ranking TNF family vs BCL family | |||||||
| Ranking of BCL2L1 w.r.t TNF family | Ranking of TNF family w.r.t BCL2L1 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| TNF - BCL2L1 | 174 | 14 | 235 | TNF - BCL2L1 | 56 | 1101 | 294 |
| TNFAIP1 - BCL2L1 | 1527 | 435 | 791 | TNFAIP1 - BCL2L1 | 1497 | 1150 | 74 |
| TNFAIP2 - BCL2L1 | 2142 | 1735 | 798 | TNFAIP2 - BCL2L1 | 1485 | 1735 | 400 |
| TNFAIP3 - BCL2L1 | 2467 | 842 | 867 | TNFAIP3 - BCL2L1 | 1109 | 1939 | 1553 |
| TNFRSF1A - BCL2L1 | 1004 | 1558 | 383 | TNFRSF1A - BCL2L1 | 492 | 376 | 1016 |
| TNFRSF10A - BCL2L1 | 1906 | 1270 | 1222 | TNFRSF10A - BCL2L1 | 2273 | 1928 | 508 |
| TNFRSF10B - BCL2L1 | 1506 | 2235 | 589 | TNFRSF10B - BCL2L1 | 1003 | 2252 | 2217 |
| TNFRSF10D - BCL2L1 | 1920 | 1555 | 1787 | TNFRSF10D - BCL2L1 | 1868 | 2420 | 2392 |
| TNFRSF12A - BCL2L1 | 1254 | 1388 | 1319 | TNFRSF12A - BCL2L1 | 1923 | 53 | 1936 |
| TNFRSF14 - BCL2L1 | 688 | 237 | 2009 | TNFRSF14 - BCL2L1 | 340 | 2350 | 2414 |
| TNFRSF21 - BCL2L1 | 1465 | 1269 | 100 | TNFRSF21 - BCL2L1 | 2139 | 718 | 289 |
| TNFSF10 - BCL2L1 | 532 | 560 | 2332 | TNFSF10 - BCL2L1 | 2115 | 2299 | 1307 |
| TNFSF15 - BCL2L1 | 1026 | 1551 | 1134 | TNFSF15 - BCL2L1 | 453 | 423 | 25 |
| Ranking of BCL2L2 w.r.t TNF family | Ranking of TNF family w.r.t BCL2L2 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| TNF - BCL2L2 | 1822 | 1926 | 2359 | TNF - BCL2L2 | 2140 | 109 | 1062 |
| TNFAIP1 - BCL2L2 | 2266 | 2478 | 1847 | TNFAIP1 - BCL2L2 | 2235 | 1607 | 712 |
| TNFAIP2 - BCL2L2 | 823 | 535 | 1117 | TNFAIP2 - BCL2L2 | 109 | 1002 | 54 |
| TNFAIP3 - BCL2L2 | 1201 | 1103 | 1511 | TNFAIP3 - BCL2L2 | 1470 | 1696 | 1276 |
| TNFRSF1A - BCL2L2 | 1124 | 2311 | 1920 | TNFRSF1A - BCL2L2 | 1912 | 169 | 1531 |
| TNFRSF10A - BCL2L2 | 1063 | 1532 | 2458 | TNFRSF10A - BCL2L2 | 1643 | 1095 | 953 |
| TNFRSF10B - BCL2L2 | 2478 | 739 | 2239 | TNFRSF10B - BCL2L2 | 2153 | 1164 | 1983 |
| TNFRSF10D - BCL2L2 | 910 | 2278 | 2237 | TNFRSF10D - BCL2L2 | 35 | 1012 | 1905 |
| TNFRSF12A - BCL2L2 | 1945 | 240 | 2484 | TNFRSF12A - BCL2L2 | 1971 | 1633 | 975 |
| TNFRSF14 - BCL2L2 | 2358 | 1648 | 2310 | TNFRSF14 - BCL2L2 | 1027 | 825 | 1228 |
| TNFRSF21 - BCL2L2 | 2292 | 1850 | 1014 | TNFRSF21 - BCL2L2 | 1138 | 486 | 554 |
| TNFSF10 - BCL2L2 | 2438 | 547 | 2013 | TNFSF10 - BCL2L2 | 2212 | 902 | 169 |
| TNFSF15 - BCL2L2 | 1196 | 2443 | 2350 | TNFSF15 - BCL2L2 | 2285 | 165 | 1330 |
| Ranking of BCL2L13 w.r.t TNF family | Ranking of TNF family w.r.t BCL2L13 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| TNF - BCL2L13 | 2437 | 2482 | 2482 | TNF - BCL2L13 | 1162 | 103 | 462 |
| TNFAIP1 - BCL2L13 | 1863 | 2386 | 989 | TNFAIP1 - BCL2L13 | 852 | 606 | 787 |
| TNFAIP2 - BCL2L13 | 793 | 293 | 1846 | TNFAIP2 - BCL2L13 | 438 | 479 | 742 |
| TNFAIP3 - BCL2L13 | 1350 | 1030 | 2129 | TNFAIP3 - BCL2L13 | 1804 | 879 | 626 |
| TNFRSF1A - BCL2L13 | 1173 | 1962 | 2489 | TNFRSF1A - BCL2L13 | 1577 | 1512 | 476 |
| TNFRSF10A - BCL2L13 | 737 | 2055 | 2499 | TNFRSF10A - BCL2L13 | 1534 | 2360 | 1105 |
| TNFRSF10B - BCL2L13 | 1992 | 885 | 906 | TNFRSF10B - BCL2L13 | 2177 | 960 | 1053 |
| TNFRSF10D - BCL2L13 | 2204 | 2159 | 2343 | TNFRSF10D - BCL2L13 | 171 | 1983 | 960 |
| TNFRSF12A - BCL2L13 | 2183 | 2509 | 241 | TNFRSF12A - BCL2L13 | 59 | 1706 | 2046 |
| TNFRSF14 - BCL2L13 | 1852 | 1974 | 2339 | TNFRSF14 - BCL2L13 | 2459 | 2381 | 1187 |
| TNFRSF21 - BCL2L13 | 2280 | 2424 | 2301 | TNFRSF21 - BCL2L13 | 52 | 1054 | 394 |
| TNFSF10 - BCL2L13 | 1088 | 2429 | 1803 | TNFSF10 - BCL2L13 | 1764 | 1186 | 1227 |
| TNFSF15 - BCL2L13 | 1286 | 2438 | 2252 | TNFSF15 - BCL2L13 | 638 | 1962 | 814 |
Table 51.
2nd order combinatorial hypotheses between BCL and TNF.
| Ranking TNF family vs BCL family | |||||||
| Ranking of BCL3 w.r.t TNF family | Ranking of TNF family w.r.t BCL3 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| TNF - BCL3 | 652 | 370 | 642 | TNF - BCL3 | 598 | 311 | 2473 |
| TNFAIP1 - BCL3 | 168 | 723 | 124 | TNFAIP1 - BCL3 | 596 | 500 | 158 |
| TNFAIP2 - BCL3 | 642 | 856 | 1098 | TNFAIP2 - BCL3 | 59 | 776 | 323 |
| TNFAIP3 - BCL3 | 2377 | 534 | 567 | TNFAIP3 - BCL3 | 300 | 940 | 1527 |
| TNFRSF1A - BCL3 | 163 | 206 | 740 | TNFRSF1A - BCL3 | 83 | 476 | 1355 |
| TNFRSF10A - BCL3 | 799 | 865 | 1044 | TNFRSF10A - BCL3 | 2388 | 2493 | 88 |
| TNFRSF10B - BCL3 | 1632 | 2427 | 1868 | TNFRSF10B - BCL3 | 757 | 1508 | 1062 |
| TNFRSF10D - BCL3 | 1110 | 858 | 714 | TNFRSF10D - BCL3 | 2213 | 1091 | 1972 |
| TNFRSF12A - BCL3 | 273 | 931 | 623 | TNFRSF12A - BCL3 | 671 | 1869 | 1286 |
| TNFRSF14 - BCL3 | 232 | 85 | 1422 | TNFRSF14 - BCL3 | 2149 | 1311 | 1650 |
| TNFRSF21 - BCL3 | 340 | 1384 | 2474 | TNFRSF21 - BCL3 | 411 | 729 | 998 |
| TNFSF10 - BCL3 | 1537 | 1753 | 1638 | TNFSF10 - BCL3 | 1926 | 1523 | 2107 |
| TNFSF15 - BCL3 | 129 | 284 | 729 | TNFSF15 - BCL3 | 1649 | 1032 | 2122 |
| Ranking of BCL6 w.r.t TNF family | Ranking of TNF family w.r.t BCL6 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| TNF - BCL6 | 2271 | 2071 | 1810 | TNF - BCL6 | 806 | 437 | 1411 |
| TNFAIP1 - BCL6 | 2135 | 2158 | 1330 | TNFAIP1 - BCL6 | 1089 | 850 | 372 |
| TNFAIP2 - BCL6 | 2340 | 1428 | 1808 | TNFAIP2 - BCL6 | 152 | 334 | 703 |
| TNFAIP3 - BCL6 | 267 | 1336 | 1219 | TNFAIP3 - BCL6 | 1884 | 1935 | 855 |
| TNFRSF1A - BCL6 | 1598 | 1771 | 2503 | TNFRSF1A - BCL6 | 788 | 741 | 1130 |
| TNFRSF10A - BCL6 | 1327 | 1831 | 2096 | TNFRSF10A - BCL6 | 607 | 1249 | 2360 |
| TNFRSF10B - BCL6 | 1373 | 1873 | 1264 | TNFRSF10B - BCL6 | 1746 | 1282 | 361 |
| TNFRSF10D - BCL6 | 2213 | 2188 | 788 | TNFRSF10D - BCL6 | 1540 | 1301 | 2008 |
| TNFRSF12A - BCL6 | 1867 | 99 | 2261 | TNFRSF12A - BCL6 | 545 | 2200 | 1910 |
| TNFRSF14 - BCL6 | 1409 | 1337 | 2028 | TNFRSF14 - BCL6 | 731 | 1302 | 1902 |
| TNFRSF21 - BCL6 | 645 | 2071 | 2335 | TNFRSF21 - BCL6 | 40 | 1850 | 50 |
| TNFSF10 - BCL6 | 919 | 445 | 99 | TNFSF10 - BCL6 | 2119 | 1102 | 1626 |
| TNFSF15 - BCL6 | 2106 | 1692 | 1451 | TNFSF15 - BCL6 | 969 | 1475 | 226 |
| Ranking of BCL9L w.r.t TNF family | Ranking of TNF family w.r.t BCL9L | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| TNF - BCL9L | 1964 | 1172 | 1478 | TNF - BCL9L | 2218 | 98 | 425 |
| TNFAIP1 - BCL9L | 439 | 1445 | 264 | TNFAIP1 - BCL9L | 766 | 2470 | 1802 |
| TNFAIP2 - BCL9L | 1250 | 1473 | 696 | TNFAIP2 - BCL9L | 646 | 567 | 85 |
| TNFAIP3 - BCL9L | 534 | 630 | 618 | TNFAIP3 - BCL9L | 1046 | 1223 | 2296 |
| TNFRSF1A - BCL9L | 2050 | 1096 | 978 | TNFRSF1A - BCL9L | 863 | 500 | 825 |
| TNFRSF10A - BCL9L | 212 | 1682 | 980 | TNFRSF10A - BCL9L | 2140 | 241 | 1547 |
| TNFRSF10B - BCL9L | 952 | 698 | 685 | TNFRSF10B - BCL9L | 286 | 414 | 2046 |
| TNFRSF10D - BCL9L | 1315 | 181 | 1423 | TNFRSF10D - BCL9L | 1956 | 112 | 990 |
| TNFRSF12A - BCL9L | 430 | 1167 | 1470 | TNFRSF12A - BCL9L | 1797 | 1280 | 1699 |
| TNFRSF14 - BCL9L | 1433 | 635 | 1497 | TNFRSF14 - BCL9L | 670 | 1055 | 1540 |
| TNFRSF21 - BCL9L | 495 | 2326 | 468 | TNFRSF21 - BCL9L | 1291 | 1378 | 246 |
| TNFSF10 - BCL9L | 1889 | 974 | 183 | TNFSF10 - BCL9L | 1812 | 1796 | 2095 |
| TNFSF15 - BCL9L | 878 | 2389 | 71 | TNFSF15 - BCL9L | 1939 | 2114 | 2405 |
| Ranking of BCL10 w.r.t TNF family | Ranking of TNF family w.r.t BCL10 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| TNF - BCL10 | 708 | 79 | 979 | TNF - BCL10 | 1931 | 114 | 1573 |
| TNFAIP1 - BCL10 | 1657 | 805 | 1298 | TNFAIP1 - BCL10 | 690 | 1941 | 7 |
| TNFAIP2 - BCL10 | 1101 | 2197 | 312 | TNFAIP2 - BCL10 | 523 | 2099 | 339 |
| TNFAIP3 - BCL10 | 985 | 813 | 767 | TNFAIP3 - BCL10 | 935 | 595 | 1870 |
| TNFRSF1A - BCL10 | 745 | 1191 | 1288 | TNFRSF1A - BCL10 | 362 | 173 | 448 |
| TNFRSF10A - BCL10 | 451 | 819 | 954 | TNFRSF10A - BCL10 | 1547 | 415 | 2426 |
| TNFRSF10B - BCL10 | 791 | 537 | 1446 | TNFRSF10B - BCL10 | 582 | 658 | 1464 |
| TNFRSF10D - BCL10 | 1831 | 1694 | 2040 | TNFRSF10D - BCL10 | 302 | 19 | 2497 |
| TNFRSF12A - BCL10 | 2015 | 1072 | 1883 | TNFRSF12A - BCL10 | 1865 | 1234 | 1540 |
| TNFRSF14 - BCL10 | 254 | 1400 | 847 | TNFRSF14 - BCL10 | 1175 | 1894 | 2227 |
| TNFRSF21 - BCL10 | 1912 | 571 | 958 | TNFRSF21 - BCL10 | 848 | 1943 | 418 |
| TNFSF10 - BCL10 | 1743 | 931 | 1657 | TNFSF10 - BCL10 | 2020 | 1522 | 1054 |
| TNFSF15 - BCL10 | 254 | 1469 | 577 | TNFSF15 - BCL10 | 1256 | 188 | 1074 |
Table 53.
2nd order interaction ranking between RAD w.r.t XRCC family members.
| Ranking RAD family w.r.t XRCC family | |||||||
| Ranking of RAD family w.r.t XRCC1 | Ranking of RAD family w.r.t XRCC2 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| RAD1 - XRCC1 | 1922 | 1658 | 1771 | XRCC2 - RAD1 | 1921 | 893 | 1774 |
| RAD18 - XRCC1 | 1027 | 456 | 1355 | XRCC2 - RAD18 | 1388 | 847 | 765 |
| XRCC1 - RAD50 | 2459 | 2254 | 2082 | XRCC2 - RAD50 | 1877 | 2185 | 2546 |
| RAD51 - XRCC1 | 282 | 365 | 1003 | XRCC2 - RAD51 | 1247 | 1033 | 629 |
| RAD51AP1 - XRCC1 | 753 | 5 | 275 | XRCC2 - RAD51AP1 | 302 | 247 | 42 |
| RAD51C - XRCC1 | 337 | 111 | 968 | XRCC2 - RAD51C | 1079 | 674 | 323 |
| RAD54B - XRCC1 | 175 | 224 | 782 | XRCC2 - RAD54B | 387 | 566 | 506 |
| RAD54L - XRCC1 | 327 | 889 | 709 | XRCC2 - RAD54L | 976 | 918 | 847 |
| Ranking of RAD family w.r.t XRCC6 | Ranking of RAD family w.r.t XRCC6BP1 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| XRCC6 - RAD1 | 1929 | 2029 | 2627 | RAD1 - XRCC6BP1 | 1167 | 2417 | 308 |
| RAD18 - XRCC6 | 541 | 25 | 1068 | RAD18 - XRCC6BP1 | 656 | 1612 | 2271 |
| XRCC6 - RAD50 | 2434 | 2043 | 2603 | XRCC6BP1 - RAD50 | 1302 | 2263 | 328 |
| RAD51 - XRCC6 | 608 | 425 | 900 | RAD51 - XRCC6BP1 | 435 | 495 | 1275 |
| RAD51AP1 - XRCC6 | 216 | 67 | 83 | RAD51AP1 - XRCC6BP1 | 81 | 177 | 73 |
| RAD51C - XRCC6 | 426 | 865 | 503 | RAD51C - XRCC6BP1 | 645 | 1366 | 1414 |
| RAD54B - XRCC6 | 3 | 610 | 112 | RAD54B - XRCC6BP1 | 154 | 693 | 1398 |
| RAD54L - XRCC6 | 85 | 252 | 432 | RAD54L - XRCC6BP1 | 420 | 1060 | 2542 |
Table 56.
2nd order interaction ranking between RAD family vs XRN2.
| Ranking XRN2 w.r.t RAD family | |||||||
| Ranking of RAD family w.r.t XRN2 | Ranking of XRN2 w.r.t RAD family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| XRN2 - RAD51AP1 | 340 | 545 | 290 | XRN2 - RAD51AP1 | 1905 | 1256 | 852 |
| XRN2 - RAD51 | 387 | 560 | 605 | XRN2 - RAD51 | 786 | 2647 | 1995 |
| XRN2 - RAD54L | 594 | 827 | 879 | XRN2 - RAD54L | 1541 | 1246 | 1819 |
| XRN2 - RAD51C | 639 | 1236 | 745 | XRN2 - RAD51C | 1037 | 1777 | 2228 |
| XRN2 - RAD18 | 794 | 688 | 804 | XRN2 - RAD18 | 904 | 2403 | 1801 |
| XRN2 - RAD1 | 898 | 1955 | 2506 | XRN2 - RAD1 | 255 | 122 | 2557 |
| XRN2 - RAD54B | 951 | 165 | 343 | XRN2 - RAD54B | 1818 | 2381 | 2603 |
| XRN2 - RAD50 | 1330 | 2312 | 2295 | XRN2 - RAD50 | 504 | 2100 | 1842 |
Table 59.
2nd order combinatorial hypotheses between RAD and XRN2.
| Unexplored combinatorial hypotheses | |
| RAD w.r.t NKRF | |
| RAD51AP1 | NKRF |
| RAD51 | NKRF |
| RAD54L | NKRF |
| RAD51C | NKRF |
| RAD18 | NKRF |
| RAD1 | NKRF |
| RAD54B | NKRF |
| NKRF w.r.t RAD | |
| RAD51AP1 | NKRF |
| RAD51 | NKRF |
| RAD54L | NKRF |
| RAD51C | NKRF |
| RAD18 | NKRF |
| NKRF | RAD1 |
| NKRF | RAD50 |
Table 61.
2nd order combinatorial hypotheses between RAD and BCL members.
| Unexplored combinatorial hypotheses | |
| RAD w.r.t BCL | |
| RAD-18/50/51/51AP1/51C/54B/54L | BCL-2L12 |
| RAD-18/51/51AP1/51C/54B/54L | BCL-6B |
| RAD-18/51/51AP1/51C/54B/54L | BCL-7A |
| RAD-18/50/51/51AP1/51C/54B/54L | BCL-9 |
| RAD-1/18/50/51/51AP1/51C/54B/54L | BCL-11A |
| RAD-1/50/51/51AP1/51C/54B/54L | BCL-11B |
| BCL w.r.t RAD | |
| RAD-1/18/50/51/51C/54B/54L | BCL-2L12 |
| RAD-1/18/50/51/51AP1/51C/54B/54L | BCL-6B |
| RAD-1/18/50/51/54L | BCL-7A |
| RAD-18/51/51C/54L | BCL-9 |
| RAD-1/18/50/51/51AP1/51C/54B | BCL-11A |
| RAD-50/51/51AP1/54B/54L | BCL-11B |
Table 62.
2nd order interaction ranking between RAD and EXOSC family members.
| Ranking RAD family VS EXOSC family | |||||||
| Ranking of EXOSC2 w.r.t RAD family | Ranking of RAD family w.r.t EXOSC2 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| EXOSC2 - RAD1 | 1033 | 1311 | 1207 | EXOSC2 - RAD1 | 2456 | 1368 | 2292 |
| EXOSC2 - RAD18 | 1210 | 995 | 1906 | EXOSC2 - RAD18 | 1115 | 979 | 654 |
| EXOSC2 - RAD50 | 1124 | 698 | 629 | EXOSC2 - RAD50 | 1647 | 2495 | 2375 |
| EXOSC2 - RAD51 | 1754 | 191 | 633 | EXOSC2 - RAD51 | 795 | 1332 | 441 |
| EXOSC2 - RAD51AP1 | 198 | 1462 | 2718 | EXOSC2 - RAD51AP1 | 2320 | 1316 | 2127 |
| EXOSC2 - RAD51C | 87 | 463 | 1130 | EXOSC2 - RAD51C | 636 | 564 | 152 |
| EXOSC2 - RAD54B | 351 | 135 | 142 | EXOSC2 - RAD54B | 278 | 132 | 282 |
| EXOSC2 - RAD54L | 1131 | 1652 | 320 | EXOSC2 - RAD54L | 125 | 888 | 545 |
| Ranking of EXOSC3 w.r.t RAD family | Ranking of RAD family w.r.t EXOSC3 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| EXOSC3 - RAD1 | 2492 | 1677 | 549 | EXOSC3 - RAD1 | 2200 | 1243 | 2711 |
| EXOSC3 - RAD18 | 1676 | 2516 | 184 | EXOSC3 - RAD18 | 2024 | 1468 | 767 |
| EXOSC3 - RAD50 | 2368 | 1892 | 2204 | EXOSC3 - RAD50 | 1062 | 596 | 2346 |
| EXOSC3 - RAD51 | 894 | 1066 | 2463 | EXOSC3 - RAD51 | 727 | 583 | 963 |
| EXOSC3 - RAD51AP1 | 1884 | 1037 | 804 | EXOSC3 - RAD51AP1 | 100 | 49 | 219 |
| EXOSC3 - RAD51C | 2499 | 2356 | 1248 | EXOSC3 - RAD51C | 663 | 869 | 887 |
| EXOSC3 - RAD54B | 2183 | 2518 | 2360 | EXOSC3 - RAD54B | 384 | 277 | 310 |
| EXOSC3 - RAD54L | 1735 | 469 | 736 | EXOSC3 - RAD54L | 546 | 1117 | 808 |
| Ranking of EXOSC5 w.r.t RAD family | Ranking of RAD family w.r.t EXOSC5 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| EXOSC5 - RAD1 | 568 | 1169 | 1699 | EXOSC5 - RAD1 | 2405 | 1716 | 1718 |
| EXOSC5 - RAD18 | 2481 | 219 | 1652 | EXOSC5 - RAD18 | 1026 | 550 | 253 |
| EXOSC5 - RAD50 | 447 | 195 | 475 | EXOSC5 - RAD50 | 1596 | 1952 | 2271 |
| EXOSC5 - RAD51 | 2548 | 431 | 1121 | EXOSC5 - RAD51 | 260 | 1095 | 137 |
| EXOSC5 - RAD51AP1 | 1290 | 487 | 430 | EXOSC5 - RAD51AP1 | 1555 | 1860 | 976 |
| EXOSC5 - RAD51C | 1284 | 1264 | 1790 | EXOSC5 - RAD51C | 233 | 1003 | 359 |
| EXOSC5 - RAD54B | 940 | 812 | 1036 | EXOSC5 - RAD54B | 834 | 1825 | 335 |
| EXOSC5 - RAD54L | 408 | 2539 | 1407 | EXOSC5 - RAD54L | 248 | 197 | 39 |
| Ranking of EXOSC6 w.r.t RAD family | Ranking of RAD family w.r.t EXOSC6 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| EXOSC6 - RAD1 | 2283 | 2490 | 1228 | EXOSC6 - RAD1 | 2405 | 142 | 639 |
| EXOSC6 - RAD18 | 1637 | 1599 | 2254 | EXOSC6 - RAD18 | 1118 | 1313 | 1549 |
| EXOSC6 - RAD50 | 2289 | 1969 | 1797 | EXOSC6 - RAD50 | 2309 | 1722 | 575 |
| EXOSC6 - RAD51 | 1056 | 1482 | 1007 | EXOSC6 - RAD51 | 998 | 2297 | 2219 |
| EXOSC6 - RAD51AP1 | 1854 | 2480 | 1827 | EXOSC6 - RAD51AP1 | 149 | 1060 | 2731 |
| EXOSC6 - RAD51C | 1996 | 940 | 1842 | EXOSC6 - RAD51C | 500 | 1628 | 2409 |
| EXOSC6 - RAD54B | 2289 | 2312 | 2005 | EXOSC6 - RAD54B | 262 | 2703 | 2465 |
| EXOSC6 - RAD54L | 987 | 2240 | 1642 | EXOSC6 - RAD54L | 885 | 271 | 1224 |
| Ranking of EXOSC7 w.r.t RAD family | Ranking of RAD family w.r.t EXOSC7 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| EXOSC7 - RAD1 | 2559 | 1735 | 1210 | EXOSC7 - RAD1 | 2079 | 2308 | 1604 |
| EXOSC7 - RAD18 | 490 | 1688 | 1331 | EXOSC7 - RAD18 | 441 | 385 | 1542 |
| EXOSC7 - RAD50 | 2661 | 1939 | 2021 | EXOSC7 - RAD50 | 1840 | 406 | 2100 |
| EXOSC7 - RAD51 | 842 | 1900 | 1876 | EXOSC7 - RAD51 | 376 | 1180 | 550 |
| EXOSC7 - RAD51AP1 | 2446 | 349 | 2374 | EXOSC7 - RAD51AP1 | 35 | 97 | 786 |
| EXOSC7 - RAD51C | 1113 | 1623 | 530 | EXOSC7 - RAD51C | 854 | 671 | 1459 |
| EXOSC7 - RAD54B | 2431 | 1612 | 1191 | EXOSC7 - RAD54B | 458 | 260 | 646 |
| EXOSC7 - RAD54L | 1550 | 1754 | 1728 | EXOSC7 - RAD54L | 464 | 528 | 790 |
| Ranking of EXOSC8 w.r.t RAD family | Ranking of RAD family w.r.t EXOSC8 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| EXOSC8 - RAD1 | 2380 | 2442 | 2630 | EXOSC8 - RAD1 | 1928 | 151 | 1563 |
| EXOSC8 - RAD18 | 805 | 2287 | 1564 | EXOSC8 - RAD18 | 764 | 523 | 29 |
| EXOSC8 - RAD50 | 1798 | 1830 | 1893 | EXOSC8 - RAD50 | 2103 | 2649 | 1822 |
| EXOSC8 - RAD51 | 404 | 1630 | 2092 | EXOSC8 - RAD51 | 98 | 1161 | 902 |
| EXOSC8 - RAD51AP1 | 1932 | 1567 | 1701 | EXOSC8 - RAD51AP1 | 408 | 1824 | 541 |
| EXOSC8 - RAD51C | 2439 | 1576 | 2554 | EXOSC8 - RAD51C | 906 | 738 | 1052 |
| EXOSC8 - RAD54B | 1562 | 2542 | 1736 | EXOSC8 - RAD54B | 23 | 1578 | 130 |
| EXOSC8 - RAD54L | 1248 | 622 | 239 | EXOSC8 - RAD54L | 651 | 1384 | 1047 |
| Ranking of EXOSC9 w.r.t RAD family | Ranking of RAD family w.r.t EXOSC9 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| EXOSC9 - RAD1 | 2240 | 175 | 1648 | EXOSC9 - RAD1 | 1335 | 1799 | 978 |
| EXOSC9 - RAD18 | 1533 | 774 | 1180 | EXOSC9 - RAD18 | 2529 | 54 | 540 |
| EXOSC9 - RAD50 | 545 | 183 | 467 | EXOSC9 - RAD50 | 211 | 2217 | 1377 |
| EXOSC9 - RAD51 | 866 | 106 | 99 | EXOSC9 - RAD51 | 807 | 74 | 429 |
| EXOSC9 - RAD51AP1 | 1570 | 1819 | 1807 | EXOSC9 - RAD51AP1 | 2480 | 103 | 1210 |
| EXOSC9 - RAD51C | 110 | 742 | 200 | EXOSC9 - RAD51C | 399 | 844 | 69 |
| EXOSC9 - RAD54B | 179 | 178 | 84 | EXOSC9 - RAD54B | 2385 | 466 | 1286 |
| EXOSC9 - RAD54L | 1113 | 2436 | 22 | EXOSC9 - RAD54L | 536 | 724 | 414 |
Table 63.
2nd order combinatorial hypotheses between RAD and EXOSC members.
| Unexplored combinatorial hypotheses | |
| RAD w.r.t EXOSC | |
| EXOSC-2 | RAD-18/51/51C/54B/54L |
| EXOSC-3 | RAD-18/50/51/51AP1/51C/54B/54L |
| EXOSC-5 | RAD-1/18/51/51AP1/51C/54B/54L |
| EXOSC-6 | RAD-1/18/50/51AP1/51C/54L |
| EXOSC-7 | RAD-18/51/51AP1/51C/54B/54L |
| EXOSC-8 | RAD-1/18/51/51AP1/51C/54B/54L |
| EXOSC-9 | RAD-1/18/50/51/51AP1/51C/54B/54L |
| EXOSC w.r.t RAD | |
| EXOSC-2 | RAD-1/18/50/51/51AP1/51C/54B/54L |
| EXOSC-3 | RAD-1/18/51/51AP1/54L |
| EXOSC-5 | RAD-1/18/50/51/51AP1/51C/54B/54L |
| EXOSC-6 | RAD-18/51/54L |
| EXOSC-7 | RAD-1/18/51C/54B/54L |
| EXOSC-8 | RAD-18/51/51AP1/54B/54L |
| EXOSC-9 | RAD-1/18/50/51/51C/54B/54L |
Table 67.
2nd order combinatorial hypotheses between RAD and FANC family.
| Unexplored combinatorial hypotheses | |
| RAD w.r.t FANC | |
| RAD-18/51/51AP1/51C/54B/54L | FANCB |
| RAD-18/51/51AP1/54B/54L | FANCD2 |
| RAD-1/18/50/51/51C/54B/54L | FANCD2OS |
| RAD-1/18/50/51/51AP1/51C/54B/54L | FANCF |
| RAD-1/18/50/51/51AP1/51C/54B/54L | FANCG |
| RAD-18/50/51/51C/54B/54L | FANCI |
| FANC w.r.t RAD | |
| FANCB | RAD-1/50/51/51AP1/51C/54B/54L |
| FANCD2 | RAD-1/50/51/51AP1/51C/54B/54L |
| FANCD2OS | RAD-1/18/5051C/54B |
| FANCF | RAD-1/18/50/51C/54B |
| FANCG | RAD-1/50/51/51AP1/51C/54B |
| FANCI | RAD-1/18/50/51/51AP1/51C/54B/54L |
Table 68.
2nd order interaction ranking between TRET vs ABC.
| Ranking TRET vs ABC family | |||||||
| Ranking of ABC family w.r.t TRET | Ranking of TRET w.r.t ABC family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| ABCF2 - TERT | 381 | 1047 | 316 | ABCF2 - TERT | 2069 | 238 | 2712 |
| ABCA2 - TERT | 1201 | 49 | 317 | ABCA2 - TERT | 1693 | 2739 | 1997 |
| ABCE1 - TERT | 1613 | 499 | 1217 | ABCE1 - TERT | 120 | 2736 | 294 |
Table 69.
2nd order combinatorial hypotheses between TRET and ABC family.
| Unexplored combinatorial hypotheses | |
| ABC family w.r.t TRET | |
| TERT | ABCF2 |
| TERT | ABCA2 |
| TERT | ABCE1 |
| TRET w.r.t ABC family | |
| ABCE1 | TERT |
Table 70.
2nd order interaction ranking between ABC w.r.t UBE2 family members.
| Ranking ABC family w.r.t UBE2 family | |||||||
| Ranking of ABC family w.r.t UBE2-A | Ranking of ABC family w.r.t UBE2-B | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| ABC-A5 - UBE2-A | 2101 | 185 | 382 | ABC-A5 - UBE2-B | 1223 | 1193 | 194 |
| ABC-B11 - UBE2-A | 129 | 2487 | 304 | ABC-B11 - UBE2-B | 125 | 103 | 571 |
| ABC-C3 - UBE2-A | 2137 | 2491 | 1023 | ABC-C3 - UBE2-B | 606 | 791 | 1411 |
| ABC-C5 - UBE2-A | 1630 | 490 | 2408 | ABC-C5 - UBE2-B | 1515 | 2317 | 2266 |
| ABC-C13 - UBE2-A | 742 | 1604 | 475 | ABC-C13 - UBE2-B | 2199 | 2254 | 2362 |
| ABC-D1 - UBE2-A | 316 | 620 | 596 | ABC-D1 - UBE2-B | 1082 | 374 | 1057 |
| ABC-G1 - UBE2-A | 46 | 819 | 533 | ABC-G1 - UBE2-B | 48 | 843 | 551 |
| ABC-G2 - UBE2-A | 398 | 259 | 261 | ABC-G2 - UBE2-B | 189 | 189 | 41 |
| Ranking of ABC family w.r.t UBE2-F | Ranking of ABC family w.r.t UBE2-H | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| ABC-A5 - UBE2-F | 997 | 2408 | 1784 | ABC-A5 - UBE2-H | 1247 | 2068 | 2438 |
| ABC-B11 - UBE2-F | 141 | 1122 | 578 | ABC-B11 - UBE2-H | 932 | 429 | 409 |
| ABC-C3 - UBE2-F | 931 | 2420 | 681 | ABC-C3 - UBE2-H | 540 | 1962 | 563 |
| ABC-C5 - UBE2-F | 628 | 1373 | 217 | ABC-C5 - UBE2-H | 1551 | 865 | 1450 |
| ABC-C13 - UBE2-F | 403 | 2464 | 1307 | ABC-C13 - UBE2-H | 1192 | 2492 | 2051 |
| ABC-D1 - UBE2-F | 2069 | 1959 | 1235 | ABC-D1 - UBE2-H | 1094 | 1016 | 1474 |
| ABC-G1 - UBE2-F | 209 | 1216 | 1450 | ABC-G1 - UBE2-H | 683 | 173 | 18 |
| ABC-G2 - UBE2-F | 690 | 1995 | 2120 | ABC-G2 - UBE2-H | 1328 | 1374 | 78 |
| Ranking of ABC family w.r.t UBE2-J1 | Ranking of ABC family w.r.t UBE2-Z | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| ABC-A5 - UBE2-J1 | 634 | 222 | 711 | ABC-A5 - UBE2-Z | 454 | 1059 | 1287 |
| ABC-B11 - UBE2-J1 | 1182 | 1075 | 403 | ABC-B11 - UBE2-Z | 134 | 503 | 436 |
| ABC-C3 - UBE2-J1 | 1232 | 719 | 1285 | ABC-C3 - UBE2-Z | 975 | 1722 | 2095 |
| ABC-C5 - UBE2-J1 | 964 | 1342 | 2373 | ABC-C5 - UBE2-Z | 2348 | 845 | 1859 |
| ABC-C13 - UBE2-J1 | 2095 | 2412 | 2360 | ABC-C13 - UBE2-Z | 1157 | 651 | 1335 |
| ABC-D1 - UBE2-J1 | 542 | 1198 | 704 | ABC-D1 - UBE2-Z | 392 | 1660 | 943 |
| ABC-G1 - UBE2-J1 | 306 | 97 | 122 | ABC-G1 - UBE2-Z | 545 | 142 | 354 |
| ABC-G2 - UBE2-J1 | 335 | 668 | 591 | ABC-G2 - UBE2-Z | 747 | 285 | 530 |
Table 71.
2nd order interaction ranking between UBE2 w.r.t ABC family members.
| Ranking UBE2 family w.r.t ABC family | |||||||
| Ranking of UBE2-A w.r.t ABC | Ranking of UBE2-B w.r.t ABC family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| ABC-A5 - UBE2-A | 1037 | 253 | 2091 | ABC-A5 - UBE2-B | 1846 | 2038 | 936 |
| ABC-B11 - UBE2-A | 1491 | 1269 | 2179 | ABC-B11 - UBE2-B | 1623 | 1304 | 1995 |
| ABC-C3 - UBE2-A | 1726 | 1906 | 1390 | ABC-C3 - UBE2-B | 1999 | 832 | 2050 |
| ABC-C5 - UBE2-A | 880 | 2122 | 2297 | ABC-C5 - UBE2-B | 612 | 2276 | 1681 |
| ABC-C13 - UBE2-A | 412 | 234 | 670 | ABC-C13 - UBE2-B | 467 | 1863 | 2496 |
| ABC-D1 - UBE2-A | 2507 | 237 | 1319 | ABC-D1 - UBE2-B | 2322 | 1917 | 2426 |
| ABC-G1 - UBE2-A | 907 | 2291 | 1573 | ABC-G1 - UBE2-B | 1194 | 1592 | 1239 |
| ABC-G2 - UBE2-A | 2048 | 1829 | 1376 | ABC-G2 - UBE2-B | 1833 | 2445 | 2506 |
| Ranking of UNE2-F w.r.t ABC family | Ranking of UBE2-H w.r.t ABC family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| ABC-A5 - UBE2-F | 2485 | 406 | 66 | ABC-A5 - UBE2-H | 508 | 2339 | 1110 |
| ABC-B11 - UBE2-F | 2003 | 1203 | 2422 | ABC-B11 - UBE2-H | 1950 | 1770 | 2461 |
| ABC-C3 - UBE2-F | 2132 | 2163 | 861 | ABC-C3 - UBE2-H | 2439 | 1972 | 2305 |
| ABC-C5 - UBE2-F | 406 | 1651 | 1838 | ABC-C5 - UBE2-H | 398 | 2473 | 2355 |
| ABC-C13 - UBE2-F | 821 | 959 | 1196 | ABC-C13 - UBE2-H | 2004 | 2317 | 1847 |
| ABC-D1 - UBE2-F | 2421 | 686 | 2176 | ABC-D1 - UBE2-H | 164 | 1641 | 648 |
| ABC-G1 - UBE2-F | 115 | 2202 | 1953 | ABC-G1 - UBE2-H | 201 | 1921 | 2288 |
| ABC-G2 - UBE2-F | 983 | 883 | 1012 | ABC-G2 - UBE2-H | 2063 | 1631 | 1354 |
| Ranking of UBE2-J1 w.r.t ABC family | Ranking of UBE2-Z w.r.t ABC family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| ABC-A5 - UBE2-J1 | 1740 | 1467 | 1244 | ABC-A5 - UBE2-Z | 2336 | 1710 | 35 |
| ABC-B11 - UBE2-J1 | 1806 | 991 | 1935 | ABC-B11 - UBE2-Z | 521 | 645 | 2168 |
| ABC-C3 - UBE2-J1 | 2073 | 2291 | 631 | ABC-C3 - UBE2-Z | 1978 | 1823 | 1859 |
| ABC-C5 - UBE2-J1 | 126 | 525 | 1409 | ABC-C5 - UBE2-Z | 1237 | 148 | 1928 |
| ABC-C13 - UBE2-J1 | 2329 | 2153 | 1951 | ABC-C13 - UBE2-Z | 1185 | 137 | 2475 |
| ABC-D1 - UBE2-J1 | 2263 | 1886 | 2249 | ABC-D1 - UBE2-Z | 2292 | 21 | 2381 |
| ABC-G1 - UBE2-J1 | 1262 | 2418 | 2277 | ABC-G1 - UBE2-Z | 426 | 2515 | 1858 |
| ABC-G2 - UBE2-J1 | 1558 | 2408 | 1304 | ABC-G2 - UBE2-Z | 2270 | 2080 | 2448 |
Table 72.
2nd order combinatorial hypotheses between ABC and UBE2.
| Unexplored combinatorial hypotheses | |
| ABC w.r.t UBE2 | |
| ABC-C3 | UBE2-A |
| ABC-C5 | UBE2-B |
| ABC-A5/D1/G2 | UBE2-F |
| ABC-A5/C13 | UBE2-H |
| ABC-C13 | UBE2-J1 |
| ABC-C5 | UBE2-Z |
| UBE2 w.r.t ABC | |
| UBEA-2 | ABC-C5/G2 |
| UBE2-B | ABC-A5/C3/C13/D1/G2 |
| UBE2-F | ABC-B11/C3/D1/G1 |
| UBE2-H | ABC-B11/C3/C5/C13/G1 |
| UBE2-J1 | ABC-B11/C3/C13/D1/G1/G2 |
| UBE2-Z | ABC-C3/D1/G1/G2 |
Table 73.
2nd order interaction ranking between ABC family members.
| Ranking ABC family w.r.t ABC family | |||||||
| Ranking of ABC family w.r.t ABC-A5 | Ranking of ABC family w.r.t ABC-B11 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| ABC-B11 - ABC-A5 | 733 | 471 | 26 | ABC-A5 - ABC-B11 | 1148 | 1443 | 1782 |
| ABC-C3 - ABC-A5 | 111 | 493 | 2264 | ABC-C3 - ABC-B11 | 845 | 527 | 1257 |
| ABC-C5 - ABC-A5 | 1717 | 519 | 1921 | ABC-C5 - ABC-B11 | 2226 | 1644 | 2241 |
| ABC-C13 - ABC-A5 | 1243 | 1943 | 2151 | ABC-C13 - ABC-B11 | 1971 | 609 | 2150 |
| ABC-D1 - ABC-A5 | 1262 | 2387 | 1573 | ABC-D1 - ABC-B11 | 891 | 217 | 854 |
| ABC-G1 - ABC-A5 | 657 | 991 | 533 | ABC-G1 - ABC-B11 | 1957 | 1920 | 669 |
| ABC-G2 - ABC-A5 | 587 | 397 | 104 | ABC-G2 - ABC-B11 | 685 | 1978 | 226 |
| Ranking of ABC family w.r.t ABC-C3 | Ranking of ABC family w.r.t ABC-C5 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| ABC-A5 - ABC-C3 | 163 | 861 | 1672 | ABC-A5 - ABC-C5 | 2086 | 411 | 1243 |
| ABC-B11 - ABC-C3 | 410 | 613 | 1501 | ABC-B11 - ABC-C5 | 2398 | 272 | 464 |
| ABC-C5 - ABC-C3 | 1591 | 2435 | 927 | ABC-C3 - ABC-C5 | 2084 | 2274 | 1758 |
| ABC-C13 - ABC-C3 | 405 | 880 | 1282 | ABC-C13 - ABC-C5 | 226 | 2476 | 2446 |
| ABC-D1 - ABC-C3 | 18 | 1145 | 2187 | ABC-D1 - ABC-C5 | 2010 | 891 | 1257 |
| ABC-G1 - ABC-C3 | 1858 | 173 | 842 | ABC-G1 - ABC-C5 | 2402 | 894 | 741 |
| ABC-G2 - ABC-C3 | 1462 | 275 | 1373 | ABC-G2 - ABC-C5 | 2463 | 736 | 661 |
| Ranking of ABC family w.r.t ABC-C13 | Ranking of ABC family w.r.t ABC-D1 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| ABC-A5 - ABC-C13 | 2251 | 1219 | 1614 | ABC-A5 - ABC-D1 | 163 | 1068 | 291 |
| ABC-B11 - ABC-C13 | 1106 | 56 | 1171 | ABC-B11 - ABC-D1 | 1273 | 130 | 1655 |
| ABC-C3 - ABC-C13 | 2279 | 1431 | 365 | ABC-C3 - ABC-D1 | 568 | 251 | 149 |
| ABC-C5 - ABC-C13 | 1537 | 2178 | 690 | ABC-C5 - ABC-D1 | 2423 | 538 | 2388 |
| ABC-D1 - ABC-C13 | 2370 | 171 | 362 | ABC-C13 - ABC-D1 | 2383 | 2029 | 425 |
| ABC-G1 - ABC-C13 | 833 | 1544 | 1343 | ABC-G1 - ABC-D1 | 1462 | 1175 | 827 |
| ABC-G2 - ABC-C13 | 329 | 1323 | 1755 | ABC-G2 - ABC-D1 | 467 | 670 | 2491 |
| Ranking of ABC family w.r.t ABC-G1 | Ranking of ABC family w.r.t ABC-G2 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| ABC-A5 - ABC-G1 | 2488 | 1776 | 1078 | ABC-A5 - ABC-G2 | 1011 | 1640 | 1705 |
| ABC-B11 - ABC-G1 | 2312 | 253 | 52 | ABC-B11 - ABC-G2 | 988 | 481 | 1849 |
| ABC-C3 - ABC-G1 | 273 | 1415 | 1139 | ABC-C3 - ABC-G2 | 1102 | 1082 | 1563 |
| ABC-C5 - ABC-G1 | 220 | 1988 | 437 | ABC-C5 - ABC-G2 | 2284 | 1904 | 1829 |
| ABC-C13 - ABC-G1 | 2389 | 427 | 1125 | ABC-C13 - ABC-G2 | 929 | 1238 | 222 |
| ABC-D1 - ABC-G1 | 1836 | 485 | 597 | ABC-D1 - ABC-G2 | 814 | 995 | 1152 |
| ABC-G2 - ABC-G1 | 2506 | 692 | 1143 | ABC-G1 - ABC-G2 | 596 | 460 | 848 |
Table 74.
2nd order combinatorial hypotheses between ABC family members.
| Unexplored combinatorial hypotheses | |
| ABC intra family | |
| ABC-C13 | ABC-A5 |
| ABC-C5/C13/G1 | ABC-B11 |
| ABC-C3/C13 | ABC-C5 |
| ABC-C5/C13 | ABC-D1 |
| ABC-A5 | ABC-G1 |
| ABC-C5 | ABC-G2 |
Table 77.
2nd order combinatorial hypotheses between ABC and IL family members.
| Unexplored combinatorial hypotheses | |
| ABC w.r.t IL | |
| IL-1B/1RAP/10RB | ABCA5 |
| IL-10RB | ABCB11 |
| IL-1A/17REL | ABCC3 |
| IL-1A/1RAP/15/17C | ABCC5 |
| IL-1RAP/15RA | ABCC13 |
| IL-8/10RB | ABCD1 |
| IL-10RB | ABCG2 |
| IL w.r.t ABC | |
| IL-17REL | ABCA5 |
| IL-2RG/6ST/15/15RA | ABCB11 |
| IL-8/15RA | ABCC3 |
| IL-15RA/17REL | ABCC5 |
| IL-15RA/17REL | ABCC13 |
| IL-1A/1RAP/8/15RA | ABCD1 |
| IL-1RAP | ABCG1 |
| IL-1RAP/15RA | ABCG2 |
Table 78.
2nd order interaction ranking between BCL and ABC family members.
| Ranking BCL family VS ABC family | |||||||
| Ranking of BCL2L1 w.r.t ABC family | Ranking of ABC family w.r.t BCL2L1 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| ABCA5 - BCL2L1 | 18 | 560 | 1715 | ABCA5 - BCL2L1 | 1522 | 220 | 1818 |
| ABCB11 - BCL2L1 | 1124 | 2418 | 552 | ABCB11 - BCL2L1 | 2002 | 234 | 10 |
| ABCC3 - BCL2L1 | 564 | 394 | 64 | ABCC3 - BCL2L1 | 2085 | 2309 | 929 |
| ABCC5 - BCL2L1 | 2239 | 1845 | 823 | ABCC5 - BCL2L1 | 599 | 847 | 1282 |
| ABCC13 - BCL2L1 | 805 | 1590 | 2407 | ABCC13 - BCL2L1 | 744 | 616 | 614 |
| ABCD1 - BCL2L1 | 356 | 202 | 930 | ABCD1 - BCL2L1 | 839 | 352 | 195 |
| ABCG1 - BCL2L1 | 793 | 2005 | 885 | ABCG1 - BCL2L1 | 1249 | 265 | 1165 |
| ABCG2 - BCL2L1 | 199 | 99 | 906 | ABCG2 - BCL2L1 | 401 | 620 | 277 |
| Ranking of BCL2L2 w.r.t ABC family | Ranking of ABC family w.r.t BCL2L2 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| ABCA5 - BCL2L2 | 1476 | 324 | 1792 | ABCA5 - BCL2L2 | 174 | 482 | 501 |
| ABCB11 - BCL2L2 | 2097 | 1311 | 2311 | ABCB11 - BCL2L2 | 148 | 380 | 1204 |
| ABCC3 - BCL2L2 | 1091 | 1569 | 259 | ABCC3 - BCL2L2 | 890 | 949 | 1398 |
| ABCC5 - BCL2L2 | 2195 | 2359 | 2322 | ABCC5 - BCL2L2 | 765 | 1875 | 736 |
| ABCC13 - BCL2L2 | 2438 | 2494 | 898 | ABCC13 - BCL2L2 | 2271 | 1436 | 1665 |
| ABCD1 - BCL2L2 | 2477 | 831 | 2156 | ABCD1 - BCL2L2 | 1432 | 1291 | 64 |
| ABCG1 - BCL2L2 | 352 | 1653 | 2234 | ABCG1 - BCL2L2 | 406 | 1206 | 966 |
| ABCG2 - BCL2L2 | 1515 | 2409 | 1496 | ABCG2 - BCL2L2 | 404 | 314 | 55 |
| Ranking of BCL2L13 w.r.t ABC family | Ranking of ABC family w.r.t BCL2L13 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| ABCA5 - BCL2L13 | 1398 | 202 | 2292 | ABCA5 - BCL2L13 | 655 | 951 | 1380 |
| ABCB11 - BCL2L13 | 2505 | 261 | 1855 | ABCB11 - BCL2L13 | 1579 | 53 | 224 |
| ABCC3 - BCL2L13 | 1642 | 1769 | 1334 | ABCC3 - BCL2L13 | 265 | 588 | 459 |
| ABCC5 - BCL2L13 | 1427 | 1835 | 2178 | ABCC5 - BCL2L13 | 1975 | 2421 | 927 |
| ABCC13 - BCL2L13 | 2484 | 2184 | 2410 | ABCC13 - BCL2L13 | 1894 | 2335 | 2475 |
| ABCD1 - BCL2L13 | 2472 | 1579 | 2201 | ABCD1 - BCL2L13 | 912 | 511 | 1041 |
| ABCG1 - BCL2L13 | 3 | 2276 | 2095 | ABCG1 - BCL2L13 | 957 | 649 | 488 |
| ABCG2 - BCL2L13 | 2172 | 1723 | 1502 | ABCG2 - BCL2L13 | 2142 | 392 | 1206 |
| Ranking of BCL3 w.r.t ABC family | Ranking of ABC family w.r.t BCL3 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| ABCA5 - BCL3 | 940 | 45 | 777 | ABCA5 - BCL3 | 960 | 1354 | 2477 |
| ABCB11 - BCL3 | 260 | 1002 | 483 | ABCB11 - BCL3 | 731 | 1483 | 2028 |
| ABCC3 - BCL3 | 2101 | 214 | 304 | ABCC3 - BCL3 | 1782 | 2186 | 1251 |
| ABCC5 - BCL3 | 1155 | 775 | 1176 | ABCC5 - BCL3 | 2192 | 957 | 280 |
| ABCC13 - BCL3 | 270 | 1116 | 1619 | ABCC13 - BCL3 | 1725 | 1407 | 1747 |
| ABCD1 - BCL3 | 759 | 2194 | 2106 | ABCD1 - BCL3 | 836 | 811 | 1359 |
| ABCG1 - BCL3 | 2014 | 1559 | 2253 | ABCG1 - BCL3 | 550 | 247 | 247 |
| ABCG2 - BCL3 | 480 | 465 | 1949 | ABCG2 - BCL3 | 792 | 798 | 1418 |
Table 80.
2nd order combinatorial hypotheses between BCL and ABC family members.
| Unexplored combinatorial hypotheses | |
| BCL w.r.t ABC | |
| ABC-C5 | BCL2L1 |
| ABC-B11/C5/C13/D1 | BCL2L2 |
| ABC-B11/C5/C13/D1/G1 | BCL2L13 |
| ABC-D1/G1 | BCL3 |
| ABC-B11 | BCL6 |
| ABC-B11 | BCL10 |
| ABC w.r.t BCL | |
| ABC-C3 | BCL2L1 |
| ABC-C5/C13 | BCL2L13 |
| ABC-C3 | BCL3 |
| ABC-C5/C13 | BCL6 |
| ABC-C5/C13/D1 | BCL9L |
| ABC-A5/C5/C13/D1 | BCL10 |
Table 85.
2nd order combinatorial hypotheses between IL and NFkB-2/I family.
| Unexplored combinatorial hypotheses | |
| IL w.r.t NFkB-2/I | |
| IL15RA | NFkB2 |
| IL17C | NFkB2 |
| IL1RN | NFkBIA |
| IL6ST | NFkBIA |
| IL15RA | NFkBIA |
| IL1RAP | NFkBIE |
| IL6ST | NFkBIE |
| IL8 | NFkBIE |
| IL17REL | NFkBIE |
| IL1A | NFkBIZ |
| IL6ST | NFkBIZ |
| IL15 | NFkBIZ |
| NFkB-2/I w.r.t IL | |
| IL10RB | NFkB2 |
| IL10RB | NFKBIZ |
| IL17REL | NFkBIZ |
Table 86.
2nd order combinatorial hypotheses between KCN and IL
| Ranking Interleukin family vs KCN family | |||||||
| Ranking of IL family w.r.t KCND3 | Ranking of KCND3 w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - KCND3 | 1995 | 2255 | 718 | IL1A - KCND3 | 707 | 118 | 11 |
| IL1B - KCND3 | 2083 | 1897 | 691 | IL1B - KCND3 | 1064 | 411 | 133 |
| IL1RAP - KCND3 | 212 | 1086 | 1690 | IL1RAP - KCND3 | 2495 | 2390 | 114 |
| IL1RN - KCND3 | 1091 | 1875 | 1551 | IL1RN - KCND3 | 459 | 743 | 300 |
| IL2RG - KCND3 | 2027 | 1557 | 403 | IL2RG - KCND3 | 588 | 248 | 58 |
| IL6ST - KCND3 | 28 | 24 | 2501 | IL6ST - KCND3 | 1 | 1127 | 2482 |
| IL8 - KCND3 | 46 | 1098 | 1426 | IL8 - KCND3 | 1134 | 1639 | 890 |
| IL10RB - KCND3 | 1573 | 2172 | 1302 | IL10RB - KCND3 | 2048 | 2306 | 2197 |
| IL15 - KCND3 | 1905 | 1606 | 716 | IL15 - KCND3 | 296 | 68 | 240 |
| IL15RA - KCND3 | 2074 | 483 | 2495 | IL15RA - KCND3 | 2511 | 2517 | 1606 |
| IL17C - KCND3 | 1881 | 2139 | 368 | IL17C - KCND3 | 588 | 1383 | 277 |
| IL17REL - KCND3 | 1715 | 2242 | 359 | IL17REL - KCND3 | 1361 | 748 | 1905 |
| Ranking of IL family w.r.t KCNH2 | Ranking of KCNH2 w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - KCNH2 | 2103 | 1832 | 356 | IL1A - KCNH2 | 1897 | 2152 | 2179 |
| IL1B - KCNH2 | 2447 | 2068 | 930 | IL1B - KCNH2 | 1599 | 2025 | 653 |
| IL1RAP - KCNH2 | 423 | 1275 | 2487 | IL1RAP - KCNH2 | 2451 | 1805 | 2002 |
| IL1RN - KCNH2 | 1600 | 828 | 779 | IL1RN - KCNH2 | 233 | 2304 | 305 |
| IL2RG - KCNH2 | 1501 | 903 | 929 | IL2RG - KCNH2 | 823 | 701 | 1820 |
| IL6ST - KCNH2 | 1016 | 1565 | 1929 | IL6ST - KCNH2 | 435 | 1665 | 2142 |
| IL8 - KCNH2 | 863 | 258 | 1395 | IL8 - KCNH2 | 1103 | 1062 | 2255 |
| IL10RB - KCNH2 | 1238 | 1335 | 1441 | IL10RB - KCNH2 | 648 | 1445 | 1684 |
| IL15 - KCNH2 | 2295 | 1419 | 1038 | IL15 - KCNH2 | 389 | 1247 | 1033 |
| IL15RA - KCNH2 | 1738 | 2263 | 296 | IL15RA - KCNH2 | 515 | 1572 | 2265 |
| IL17C - KCNH2 | 2084 | 1399 | 49 | IL17C - KCNH2 | 1388 | 1021 | 1079 |
| IL17REL - KCNH2 | 90 | 1956 | 1491 | IL17REL - KCNH2 | 727 | 2338 | 524 |
| Ranking of IL family w.r.t KCNH8 | Ranking of KCNH8 w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - KCNH8 | 2268 | 2507 | 1877 | IL1A - KCNH8 | 29 | 1939 | 1438 |
| IL1B - KCNH8 | 2223 | 2013 | 2204 | IL1B - KCNH8 | 2060 | 472 | 2177 |
| IL1RAP - KCNH8 | 1238 | 479 | 1717 | IL1RAP - KCNH8 | 1950 | 651 | 150 |
| IL1RN - KCNH8 | 1653 | 819 | 2040 | IL1RN - KCNH8 | 1094 | 329 | 988 |
| IL2RG - KCNH8 | 57 | 1530 | 651 | IL2RG - KCNH8 | 1853 | 1224 | 390 |
| IL6ST - KCNH8 | 2067 | 979 | 1640 | IL6ST - KCNH8 | 607 | 368 | 800 |
| IL8 - KCNH8 | 1558 | 439 | 1250 | IL8 - KCNH8 | 2484 | 269 | 1048 |
| IL10RB - KCNH8 | 937 | 448 | 416 | IL10RB - KCNH8 | 2381 | 2008 | 726 |
| IL15 - KCNH8 | 1575 | 1789 | 580 | IL15 - KCNH8 | 1365 | 1649 | 2187 |
| IL15RA - KCNH8 | 2082 | 1524 | 1550 | IL15RA - KCNH8 | 1667 | 638 | 1648 |
| IL17C - KCNH8 | 1847 | 1700 | 2354 | IL17C - KCNH8 | 1232 | 1825 | 1519 |
| IL17REL - KCNH8 | 1542 | 2 | 1803 | IL17REL - KCNH8 | 1120 | 681 | 2060 |
Table 87.
2nd order combinatorial hypotheses between KCN and IL.
| Ranking Interleukin family vs KCN family | |||||||
| Ranking of IL family w.r.t KCNK1 | Ranking of KCNK1 w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - KCNK1 | 2290 | 2066 | 1071 | IL1A - KCNK1 | 1644 | 1818 | 2362 |
| IL1B - KCNK1 | 1941 | 2452 | 1905 | IL1B - KCNK1 | 813 | 966 | 1554 |
| IL1RAP - KCNK1 | 171 | 595 | 2490 | IL1RAP - KCNK1 | 1103 | 1318 | 1803 |
| IL1RN - KCNK1 | 2468 | 1897 | 391 | IL1RN - KCNK1 | 2130 | 73 | 1326 |
| IL2RG - KCNK1 | 2384 | 1028 | 755 | IL2RG - KCNK1 | 650 | 2324 | 1413 |
| IL6ST - KCNK1 | 862 | 131 | 807 | IL6ST - KCNK1 | 2226 | 688 | 2283 |
| IL8 - KCNK1 | 722 | 22 | 2147 | IL8 - KCNK1 | 1872 | 1978 | 1201 |
| IL10RB - KCNK1 | 1965 | 125 | 1204 | IL10RB - KCNK1 | 1087 | 1633 | 1021 |
| IL15 - KCNK1 | 2280 | 2009 | 502 | IL15 - KCNK1 | 1639 | 506 | 2369 |
| IL15RA - KCNK1 | 1567 | 1546 | 895 | IL15RA - KCNK1 | 1126 | 1499 | 784 |
| IL17C - KCNK1 | 2451 | 122 | 931 | IL17C - KCNK1 | 1331 | 2472 | 611 |
| IL17REL - KCNK1 | 1515 | 659 | 2391 | IL17REL - KCNK1 | 979 | 2329 | 329 |
| Ranking of IL family w.r.t KCNK5 | Ranking of KCNK5 w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - KCNK5 | 609 | 608 | 796 | IL1A - KCNK5 | 1840 | 1512 | 576 |
| IL1B - KCNK5 | 1297 | 110 | 543 | IL1B - KCNK5 | 510 | 1379 | 347 |
| IL1RAP - KCNK5 | 1137 | 2237 | 1314 | IL1RAP - KCNK5 | 730 | 911 | 1003 |
| IL1RN - KCNK5 | 1583 | 1930 | 2136 | IL1RN - KCNK5 | 815 | 405 | 1431 |
| IL2RG - KCNK5 | 223 | 601 | 14 | IL2RG - KCNK5 | 1841 | 782 | 806 |
| IL6ST - KCNK5 | 1038 | 867 | 295 | IL6ST - KCNK5 | 455 | 848 | 1275 |
| IL8 - KCNK5 | 819 | 1737 | 105 | IL8 - KCNK5 | 272 | 479 | 1215 |
| IL10RB - KCNK5 | 1879 | 2298 | 1903 | IL10RB - KCNK5 | 457 | 1769 | 2206 |
| IL15 - KCNK5 | 981 | 1630 | 1669 | IL15 - KCNK5 | 1060 | 244 | 580 |
| IL15RA - KCNK5 | 791 | 124 | 555 | IL15RA - KCNK5 | 361 | 332 | 1662 |
| IL17C - KCNK5 | 2345 | 449 | 919 | IL17C - KCNK5 | 388 | 713 | 509 |
| IL17REL - KCNK5 | 18 | 2118 | 1873 | IL17REL - KCNK5 | 1377 | 2108 | 1634 |
| Ranking of IL family w.r.t KCNK6 | Ranking of KCNK6 w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - KCNK6 | 140 | 180 | 615 | IL1A - KCNK6 | 1683 | 944 | 860 |
| IL1B - KCNK6 | 525 | 57 | 369 | IL1B - KCNK6 | 620 | 1047 | 903 |
| IL1RAP - KCNK6 | 1878 | 830 | 902 | IL1RAP - KCNK6 | 2386 | 1248 | 2053 |
| IL1RN - KCNK6 | 1834 | 1564 | 90 | IL1RN - KCNK6 | 2047 | 970 | 1311 |
| IL2RG - KCNK6 | 2181 | 974 | 7 | IL2RG - KCNK6 | 691 | 2454 | 1443 |
| IL6ST - KCNK6 | 728 | 1895 | 1270 | IL6ST - KCNK6 | 440 | 125 | 1682 |
| IL8 - KCNK6 | 2168 | 2442 | 869 | IL8 - KCNK6 | 774 | 98 | 654 |
| IL10RB - KCNK6 | 1821 | 560 | 85 | IL10RB - KCNK6 | 802 | 1903 | 2156 |
| IL15 - KCNK6 | 1589 | 304 | 2447 | IL15 - KCNK6 | 1944 | 130 | 2047 |
| IL15RA - KCNK6 | 87 | 436 | 2447 | IL15RA - KCNK6 | 1835 | 1531 | 1529 |
| IL17C - KCNK6 | 103 | 200 | 538 | IL17C - KCNK6 | 1056 | 1350 | 1408 |
| IL17REL - KCNK6 | 2066 | 2159 | 1735 | IL17REL - KCNK6 | 1044 | 1263 | 947 |
Table 88.
2nd order combinatorial hypotheses between IL family w.r.t KCN family.
| Unexplored combinatorial hypotheses | |
| IL w.r.t KCN | |
| IL-1A/1B/15RA/17C | KCND3 |
| IL-1A/1B | KCNH2 |
| IL-1A/1B/17C | KCNH8 |
| IL-1A/1B/1RN/15 | KCNK1 |
| IL-1RN/10RB/17REL | KCNK5 |
| IL-8/17REL | KCNK6 |
| KCN w.r.t IL family | |
| IL-1A/1B/15RA/17C | KCND3 |
| IL-1A/1RAP | KCNH2 |
| IL-1B/10RB | KCNH8 |
| IL-1A/6ST/8 | KCNK1 |
| IL-10RB | KCNK5 |
| IL-1RAP/10RB/15 | KCNK6 |
Table 96.
2nd order combinatorial hypotheses between TRAF and IL
| Ranking Interleukin family vs TRAF family | |||||||
| Ranking of IL family w.r.t TRAF3IP2 | Ranking of TRAF3IP2 w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - TRAF3IP2 | 2142 | 100 | 666 | IL1A - TRAF3IP2 | 1518 | 2265 | 1107 |
| IL1B - TRAF3IP2 | 1155 | 110 | 1193 | IL1B - TRAF3IP2 | 1953 | 1294 | 2359 |
| IL1RAP - TRAF3IP2 | 704 | 2482 | 2385 | IL1RAP - TRAF3IP2 | 913 | 2034 | 38 |
| IL1RN - TRAF3IP2 | 272 | 497 | 133 | IL1RN - TRAF3IP2 | 1044 | 538 | 1173 |
| IL2RG - TRAF3IP2 | 1948 | 1043 | 942 | IL2RG - TRAF3IP2 | 1767 | 2385 | 2059 |
| IL6ST - TRAF3IP2 | 49 | 1244 | 1098 | IL6ST - TRAF3IP2 | 257 | 1991 | 1871 |
| IL8 - TRAF3IP2 | 1165 | 598 | 344 | IL8 - TRAF3IP2 | 796 | 2192 | 2289 |
| IL10RB - TRAF3IP2 | 1252 | 1426 | 552 | IL10RB - TRAF3IP2 | 840 | 237 | 2096 |
| IL15 - TRAF3IP2 | 1550 | 433 | 163 | IL15 - TRAF3IP2 | 1428 | 1183 | 2219 |
| IL15RA - TRAF3IP2 | 2024 | 2162 | 1800 | IL15RA - TRAF3IP2 | 906 | 1995 | 1717 |
| IL17C - TRAF3IP2 | 2253 | 61 | 98 | IL17C - TRAF3IP2 | 1290 | 1587 | 1839 |
| IL17REL - TRAF3IP2 | 18 | 2515 | 2057 | IL17REL - TRAF3IP2 | 1836 | 2042 | 1568 |
| Ranking of IL family w.r.t TRAF4 | Ranking of TRAF4 w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - TRAF4 | 26 | 2316 | 707 | IL1A - TRAF4 | 1806 | 439 | 1465 |
| IL1B - TRAF4 | 582 | 2136 | 175 | IL1B - TRAF4 | 1026 | 746 | 378 |
| IL1RAP - TRAF4 | 1180 | 1714 | 961 | IL1RAP - TRAF4 | 909 | 2225 | 1546 |
| IL1RN - TRAF4 | 494 | 2347 | 590 | IL1RN - TRAF4 | 625 | 1031 | 1939 |
| IL2RG - TRAF4 | 1092 | 1860 | 275 | IL2RG - TRAF4 | 1130 | 339 | 826 |
| IL6ST - TRAF4 | 2333 | 344 | 1914 | IL6ST - TRAF4 | 676 | 1966 | 1556 |
| IL8 - TRAF4 | 749 | 604 | 950 | IL8 - TRAF4 | 406 | 450 | 1531 |
| IL10RB - TRAF4 | 580 | 2512 | 424 | IL10RB - TRAF4 | 2407 | 1781 | 1136 |
| IL15 - TRAF4 | 1131 | 2078 | 227 | IL15 - TRAF4 | 905 | 2408 | 1759 |
| IL15RA - TRAF4 | 551 | 1628 | 2237 | IL15RA - TRAF4 | 1197 | 2125 | 2073 |
| IL17C - TRAF4 | 236 | 2464 | 19 | IL17C - TRAF4 | 1538 | 914 | 1515 |
| IL17REL - TRAF4 | 2422 | 381 | 2487 | IL17REL - TRAF4 | 575 | 1394 | 320 |
| Ranking of IL family w.r.t TRAF6 | Ranking of TRAF6 w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - TRAF6 | 1 | 343 | 2237 | IL1A - TRAF6 | 1637 | 455 | 2334 |
| IL1B - TRAF6 | 224 | 143 | 2107 | IL1B - TRAF6 | 861 | 1386 | 1342 |
| IL1RAP - TRAF6 | 1875 | 1483 | 1433 | IL1RAP - TRAF6 | 2219 | 1984 | 1766 |
| IL1RN - TRAF6 | 107 | 706 | 988 | IL1RN - TRAF6 | 1334 | 1067 | 1301 |
| IL2RG - TRAF6 | 790 | 1706 | 1028 | IL2RG - TRAF6 | 695 | 1717 | 1986 |
| IL6ST - TRAF6 | 1508 | 928 | 930 | IL6ST - TRAF6 | 54 | 762 | 1130 |
| IL8 - TRAF6 | 2088 | 1883 | 2089 | IL8 - TRAF6 | 2457 | 2139 | 1218 |
| IL10RB - TRAF6 | 17 | 786 | 1211 | IL10RB - TRAF6 | 303 | 1825 | 1709 |
| IL15 - TRAF6 | 320 | 1692 | 2045 | IL15 - TRAF6 | 2071 | 2475 | 1500 |
| IL15RA - TRAF6 | 1560 | 303 | 2392 | IL15RA - TRAF6 | 1688 | 1189 | 1344 |
| IL17C - TRAF6 | 42 | 227 | 1457 | IL17C - TRAF6 | 2469 | 2309 | 1503 |
| IL17REL - TRAF6 | 2454 | 2517 | 412 | IL17REL - TRAF6 | 124 | 2067 | 823 |
| Ranking of IL family w.r.t TRAFD1 | Ranking of TRAFD1 w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - TRAFD1 | 2408 | 1040 | 1579 | IL1A - TRAFD1 | 2121 | 699 | 1587 |
| IL1B - TRAFD1 | 1478 | 2046 | 1321 | IL1B - TRAFD1 | 756 | 2435 | 571 |
| IL1RAP - TRAFD1 | 491 | 1639 | 447 | IL1RAP - TRAFD1 | 528 | 857 | 2043 |
| IL1RN - TRAFD1 | 895 | 1149 | 266 | IL1RN - TRAFD1 | 1033 | 848 | 1374 |
| IL2RG - TRAFD1 | 1025 | 1948 | 43 | IL2RG - TRAFD1 | 1243 | 492 | 1579 |
| IL6ST - TRAFD1 | 1835 | 1824 | 809 | IL6ST - TRAFD1 | 1064 | 868 | 699 |
| IL8 - TRAFD1 | 1318 | 896 | 663 | IL8 - TRAFD1 | 650 | 671 | 1088 |
| IL10RB - TRAFD1 | 329 | 2371 | 355 | IL10RB - TRAFD1 | 2403 | 556 | 800 |
| IL15 - TRAFD1 | 1165 | 1934 | 769 | IL15 - TRAFD1 | 339 | 623 | 634 |
| IL15RA - TRAFD1 | 351 | 260 | 2385 | IL15RA - TRAFD1 | 265 | 1369 | 386 |
| IL17C - TRAFD1 | 1191 | 1389 | 1486 | IL17C - TRAFD1 | 756 | 1068 | 1390 |
| IL17REL - TRAFD1 | 704 | 2222 | 788 | IL17REL - TRAFD1 | 370 | 640 | 137 |
Table 97.
2nd order combinatorial hypotheses between IL and TRAF family.
| Unexplored combinatorial hypotheses | |
| IL w.r.t TRAF | |
| IL-1RAP/15RA/17REL | TRAF3IP2 |
| IL-6ST/17REL | TRAF4 |
| IL-8/17REL | TRAF6 |
| IL-6ST | TRAFD1 |
| TRAF w.r.t IL | |
| IL-1B/2RG/6ST/8/17REL | TRAF3IP2 |
| IL-10RB/15/15RA | TRAF4 |
| IL-1RAP/8/15/17C | TRAF6 |
Table 108.
2nd order combinatorial hypotheses between IL and TNF family.
| Unexplored combinatorial hypotheses | |
| IL w.r.t TNF | |
| IL-1RAP/6ST/15RA | TNF |
| IL-1B/2RG/15RA/17C | TNFAIP1 |
| IL-1RN/10RB | TNFAIP2 |
| IL-6ST/8/17REL | TNFAIP3 |
| IL-1RAP | TNFRSF1A |
| IL-1RAP/15RA/17REL | TNFRSF10A |
| IL-15RA | TNFRSF10B |
| IL-15RA | TNFRSF10D |
| IL-8/15RA/17REL | TNFRSF12A |
| IL-15RA | TNFRSF14 |
| IL-1B/1RAP/2RG | TNFRSF21 |
| IL-1B/15RA/17C | TNFSF10 |
| IL-17C | TNFSF15 |
| TNF w.r.t IL | |
| IL-6ST/10RB | TNF |
| IL-8/15RA | TNFAIP1 |
| IL-1B | TNFRSF1A |
| IL-1A/1B/1RN/2RG/6ST/15/15RA/17C | TNFRSF10A |
| IL-1RN | TNFRSF10B |
| IL-1A/1B/2RG/6ST/10RB/15/17C/17REL | TNFRSF10D |
| IL-6ST/17C | TNFRSF12A |
| IL-1A/1RN/2RG/6ST/8/15RA/17C/17REL | TNFRSF14 |
| IL-17REL | TNFRSF14 |
| IL10RB | TNFSF10 |
| IL15 | TNFSF15 |
Table 109.
2nd order combinatorial hypotheses between BCL and IL.
| Ranking IL family vs BCL family | |||||||
| Ranking of IL family w.r.t BCL2L1 | Ranking of BCL2L1 w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - BCL2L1 | 2482 | 859 | 1834 | IL1A - BCL2L1 | 780 | 1156 | 1712 |
| IL1B - BCL2L1 | 2252 | 1920 | 1482 | IL1B - BCL2L1 | 1838 | 954 | 2132 |
| IL1RAP - BCL2L1 | 1128 | 815 | 1935 | IL1RAP - BCL2L1 | 870 | 1777 | 1262 |
| IL1RN - BCL2L1 | 648 | 2504 | 650 | IL1RN - BCL2L1 | 973 | 385 | 1297 |
| IL2RG - BCL2L1 | 1542 | 1439 | 700 | IL2RG - BCL2L1 | 2048 | 486 | 1949 |
| IL6ST - BCL2L1 | 663 | 553 | 1432 | IL6ST - BCL2L1 | 284 | 674 | 468 |
| IL8 - BCL2L1 | 260 | 202 | 2070 | IL8 - BCL2L1 | 1430 | 1343 | 1417 |
| IL10RB - BCL2L1 | 1867 | 347 | 17 | IL10RB - BCL2L1 | 1659 | 1965 | 2024 |
| IL15 - BCL2L1 | 1558 | 775 | 381 | IL15 - BCL2L1 | 690 | 542 | 1277 |
| IL15RA - BCL2L1 | 2136 | 1177 | 1533 | IL15RA - BCL2L1 | 581 | 1107 | 972 |
| IL17C - BCL2L1 | 2481 | 2410 | 2512 | IL17C - BCL2L1 | 695 | 1739 | 1775 |
| IL17REL - BCL2L1 | 815 | 657 | 374 | IL17REL - BCL2L1 | 981 | 1225 | 509 |
| Ranking of IL family w.r.t BCL2L2 | Ranking of BCL2L2 w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - BCL2L2 | 138 | 361 | 86 | IL1A - BCL2L2 | 2407 | 2362 | 2464 |
| IL1B - BCL2L2 | 165 | 389 | 108 | IL1B - BCL2L2 | 1807 | 2462 | 2344 |
| IL1RAP - BCL2L2 | 623 | 1523 | 861 | IL1RAP - BCL2L2 | 77 | 1897 | 1711 |
| IL1RN - BCL2L2 | 2324 | 530 | 984 | IL1RN - BCL2L2 | 2298 | 1620 | 2092 |
| IL2RG - BCL2L2 | 2137 | 285 | 347 | IL2RG - BCL2L2 | 2429 | 850 | 1744 |
| IL6ST - BCL2L2 | 2239 | 1927 | 2085 | IL6ST - BCL2L2 | 477 | 2046 | 1859 |
| IL8 - BCL2L2 | 894 | 1418 | 1346 | IL8 - BCL2L2 | 1803 | 1072 | 2024 |
| IL10RB - BCL2L2 | 2243 | 738 | 1020 | IL10RB - BCL2L2 | 1041 | 145 | 843 |
| IL15 - BCL2L2 | 110 | 650 | 1347 | IL15 - BCL2L2 | 2474 | 2142 | 2416 |
| IL15RA - BCL2L2 | 258 | 1715 | 361 | IL15RA - BCL2L2 | 1377 | 1211 | 2298 |
| IL17C - BCL2L2 | 554 | 12 | 147 | IL17C - BCL2L2 | 1168 | 2512 | 2447 |
| IL17REL - BCL2L2 | 2454 | 2510 | 2482 | IL17REL - BCL2L2 | 539 | 1875 | 1442 |
| Ranking of IL family w.r.t BCL2L13 | Ranking of BCL2L13 w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - BCL2L13 | 1572 | 458 | 174 | IL1A - BCL2L13 | 1456 | 811 | 2403 |
| IL1B - BCL2L13 | 927 | 227 | 424 | IL1B - BCL2L13 | 1286 | 1446 | 2348 |
| IL1RAP - BCL2L13 | 278 | 718 | 1941 | IL1RAP - BCL2L13 | 823 | 2450 | 2510 |
| IL1RN - BCL2L13 | 608 | 1277 | 881 | IL1RN - BCL2L13 | 2503 | 623 | 2378 |
| IL2RG - BCL2L13 | 507 | 1182 | 5 | IL2RG - BCL2L13 | 2483 | 1648 | 2248 |
| IL6ST - BCL2L13 | 1778 | 1403 | 246 | IL6ST - BCL2L13 | 1899 | 2473 | 2046 |
| IL8 - BCL2L13 | 178 | 468 | 1606 | IL8 - BCL2L13 | 2099 | 910 | 2294 |
| IL10RB - BCL2L13 | 991 | 1211 | 804 | IL10RB - BCL2L13 | 2120 | 1895 | 194 |
| IL15 - BCL2L13 | 1868 | 432 | 15 | IL15 - BCL2L13 | 2515 | 2160 | 2420 |
| IL15RA - BCL2L13 | 1629 | 2134 | 685 | IL15RA - BCL2L13 | 933 | 1844 | 2318 |
| IL17C - BCL2L13 | 995 | 84 | 20 | IL17C - BCL2L13 | 2004 | 2434 | 2500 |
| IL17REL - BCL2L13 | 2420 | 2419 | 2464 | IL17REL - BCL2L13 | 1490 | 760 | 442 |
| Ranking of IL family w.r.t BCL3 | Ranking of BCL3 w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - BCL3 | 880 | 2462 | 396 | IL1A - BCL3 | 474 | 436 | 1045 |
| IL1B - BCL3 | 975 | 1507 | 40 | IL1B - BCL3 | 799 | 303 | 926 |
| IL1RAP - BCL3 | 1425 | 821 | 1129 | IL1RAP - BCL3 | 44 | 164 | 1115 |
| IL1RN - BCL3 | 149 | 471 | 311 | IL1RN - BCL3 | 37 | 1784 | 477 |
| IL2RG - BCL3 | 454 | 365 | 505 | IL2RG - BCL3 | 524 | 2060 | 335 |
| IL6ST - BCL3 | 1928 | 755 | 2344 | IL6ST - BCL3 | 316 | 1457 | 607 |
| IL8 - BCL3 | 1052 | 743 | 2044 | IL8 - BCL3 | 2266 | 1236 | 1983 |
| IL10RB - BCL3 | 95 | 800 | 1625 | IL10RB - BCL3 | 2187 | 1600 | 2170 |
| IL15 - BCL3 | 1041 | 820 | 214 | IL15 - BCL3 | 17 | 966 | 182 |
| IL15RA - BCL3 | 2478 | 1820 | 2500 | IL15RA - BCL3 | 462 | 1476 | 1100 |
| IL17C - BCL3 | 737 | 1682 | 8 | IL17C - BCL3 | 1069 | 923 | 1926 |
| IL17REL - BCL3 | 218 | 424 | 2019 | IL17REL - BCL3 | 692 | 1897 | 1274 |
Table 110.
2nd order combinatorial hypotheses between BCL and IL.
| Ranking IL family vs BCL family | |||||||
| Ranking of IL family w.r.t BCL6 | Ranking of BCL6 w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - BCL6 | 157 | 5 | 1029 | IL1A - BCL6 | 1034 | 2503 | 1669 |
| IL1B - BCL6 | 274 | 767 | 1904 | IL1B - BCL6 | 2298 | 2423 | 2294 |
| IL1RAP - BCL6 | 1021 | 2360 | 1813 | IL1RAP - BCL6 | 2403 | 1289 | 777 |
| IL1RN - BCL6 | 2015 | 366 | 506 | IL1RN - BCL6 | 1919 | 2301 | 1680 |
| IL2RG - BCL6 | 425 | 553 | 480 | IL2RG - BCL6 | 1389 | 2106 | 2478 |
| IL6ST - BCL6 | 2419 | 1589 | 1962 | IL6ST - BCL6 | 92 | 184 | 1752 |
| IL8 - BCL6 | 2363 | 2233 | 1343 | IL8 - BCL6 | 2123 | 2068 | 181 |
| IL10RB - BCL6 | 853 | 383 | 1983 | IL10RB - BCL6 | 847 | 1980 | 1186 |
| IL15 - BCL6 | 500 | 397 | 1767 | IL15 - BCL6 | 1297 | 1925 | 1014 |
| IL15RA - BCL6 | 1686 | 1432 | 2269 | IL15RA - BCL6 | 2084 | 1791 | 2203 |
| IL17C - BCL6 | 227 | 255 | 2412 | IL17C - BCL6 | 1349 | 1499 | 1321 |
| IL17REL - BCL6 | 2253 | 2396 | 63 | IL17REL - BCL6 | 38 | 1949 | 1930 |
| Ranking of IL family w.r.t BCL9L | Ranking of BCL9L w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - BCL9L | 1932 | 1942 | 210 | IL1A - BCL9L | 1620 | 1559 | 986 |
| IL1B - BCL9L | 1966 | 88 | 79 | IL1B - BCL9L | 361 | 1449 | 2484 |
| IL1RAP - BCL9L | 218 | 957 | 881 | IL1RAP - BCL9L | 984 | 623 | 1689 |
| IL1RN - BCL9L | 1629 | 937 | 132 | IL1RN - BCL9L | 689 | 55 | 1593 |
| IL2RG - BCL9L | 415 | 104 | 92 | IL2RG - BCL9L | 2113 | 892 | 567 |
| IL6ST - BCL9L | 2249 | 1960 | 1142 | IL6ST - BCL9L | 1718 | 1210 | 737 |
| IL8 - BCL9L | 814 | 2197 | 2162 | IL8 - BCL9L | 1679 | 1920 | 933 |
| IL10RB - BCL9L | 743 | 632 | 660 | IL10RB - BCL9L | 1631 | 717 | 1236 |
| IL15 - BCL9L | 1343 | 279 | 280 | IL15 - BCL9L | 568 | 1068 | 1794 |
| IL15RA - BCL9L | 1714 | 111 | 1279 | IL15RA - BCL9L | 206 | 951 | 251 |
| IL17C - BCL9L | 2029 | 196 | 94 | IL17C - BCL9L | 1031 | 573 | 1870 |
| IL17REL - BCL9L | 128 | 2308 | 1926 | IL17REL - BCL9L | 1214 | 1341 | 839 |
| Ranking of IL family w.r.t BCL10 | Ranking of BCL10 w.r.t IL family | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| IL1A - BCL10 | 5 | 1720 | 506 | IL1A - BCL10 | 513 | 2405 | 1889 |
| IL1B - BCL10 | 201 | 2404 | 803 | IL1B - BCL10 | 2100 | 432 | 1251 |
| IL1RAP - BCL10 | 1597 | 598 | 1869 | IL1RAP - BCL10 | 1929 | 499 | 2112 |
| IL1RN - BCL10 | 107 | 724 | 126 | IL1RN - BCL10 | 1846 | 1823 | 209 |
| IL2RG - BCL10 | 232 | 665 | 650 | IL2RG - BCL10 | 1885 | 1803 | 1577 |
| IL6ST - BCL10 | 2008 | 1698 | 1816 | IL6ST - BCL10 | 451 | 71 | 337 |
| IL8 - BCL10 | 1614 | 719 | 1555 | IL8 - BCL10 | 204 | 1653 | 544 |
| IL10RB - BCL10 | 2009 | 466 | 1053 | IL10RB - BCL10 | 2244 | 2150 | 1578 |
| IL15 - BCL10 | 35 | 2072 | 580 | IL15 - BCL10 | 2174 | 1618 | 1375 |
| IL15RA - BCL10 | 1477 | 2064 | 1789 | IL15RA - BCL10 | 1810 | 1656 | 1835 |
| IL17C - BCL10 | 8 | 2009 | 1232 | IL17C - BCL10 | 705 | 1777 | 207 |
| IL17REL - BCL10 | 2397 | 89 | 550 | IL17REL - BCL10 | 839 | 1214 | 377 |
Table 111.
2nd order combinatorial hypotheses between IL and BCL family.
| Unexplored combinatorial hypotheses | |
| IL w.r.t BCL | |
| IL-1A/1B/17C | BCL2L1 |
| IL-6ST/17REL | BCL2L2 |
| IL-17REL | BCL2L13 |
| IL-6ST/15RA | BCL3 |
| IL-1RAP/6ST/8/17REL | BCL6 |
| IL-1A/6ST/8/17REL | BCL9L |
| IL-6ST/15RA | BCL10 |
| BCL w.r.t IL | |
| IL-1B/2RG/10RB | BCL2L1 |
| IL-1A/1B/1RN/6ST/8/15/17C | BCL2L2 |
| IL-1RAP/1RN/2RG/6ST/8/10RB/15/15RA/17C | BCL2L13 |
| IL-8/10RB | BCL3 |
| IL-1B/1RN/2RG/8/15RA/17REL | BCL6 |
| IL-1A/1RAP/1RN/2RG/10RB/15RA | BCL10 |
Table 120.
2nd order combinatorial hypotheses between BCL and EXOSC.
| Ranking EXOSC family vs BCL family | |||||||
| Ranking of EXOSC2 w.r.t BCL family | Ranking of BCL family w.r.t EXOSC2 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| EXOSC2 - BCL2L12 | 723 | 355 | 1211 | EXOSC2 - BCL2L12 | 1498 | 1889 | 1856 |
| EXOSC2 - BCL6B | 1092 | 1033 | 638 | EXOSC2 - BCL6B | 202 | 81 | 194 |
| EXOSC2 - BCL7A | 1633 | 1047 | 317 | EXOSC2 - BCL7A | 2403 | 2531 | 2405 |
| EXOSC2 - BCL9 | 699 | 559 | 425 | EXOSC2 - BCL9 | 2552 | 2230 | 1755 |
| EXOSC2 - BCL11A | 338 | 319 | 1598 | EXOSC2 - BCL11A | 574 | 834 | 1055 |
| EXOSC2 - BCL11B | 1285 | 1440 | 812 | EXOSC2 - BCL11B | 1067 | 1574 | 730 |
| Ranking of EXOSC3 w.r.t BCL family | Ranking of BCL family w.r.t EXOSC3 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| EXOSC3 - BCL2L12 | 2280 | 1640 | 1955 | EXOSC3 - BCL2L12 | 1976 | 1482 | 2399 |
| EXOSC3 - BCL6B | 2429 | 2273 | 2407 | EXOSC3 - BCL6B | 571 | 335 | 307 |
| EXOSC3 - BCL7A | 2100 | 1374 | 2674 | EXOSC3 - BCL7A | 1739 | 1882 | 1700 |
| EXOSC3 - BCL9 | 2437 | 2223 | 2245 | EXOSC3 - BCL9 | 2380 | 1912 | 2321 |
| EXOSC3 - BCL11A | 2212 | 2090 | 116 | EXOSC3 - BCL11A | 1018 | 1345 | 483 |
| EXOSC3 - BCL11B | 1677 | 199 | 267 | EXOSC3 - BCL11B | 2572 | 1876 | 2395 |
| Ranking of EXOSC5 w.r.t BCL family | Ranking of BCL family w.r.t EXOSC5 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| EXOSC5 - BCL2L12 | 498 | 1342 | 436 | EXOSC5 - BCL2L12 | 2174 | 1635 | 1824 |
| EXOSC5 - BCL6B | 786 | 1272 | 1194 | EXOSC5 - BCL6B | 330 | 193 | 107 |
| EXOSC5 - BCL7A | 374 | 1338 | 874 | EXOSC5 - BCL7A | 2582 | 2701 | 2415 |
| EXOSC5 - BCL9 | 613 | 946 | 772 | EXOSC5 - BCL9 | 1777 | 1511 | 2011 |
| EXOSC5 - BCL11A | 459 | 90 | 1034 | EXOSC5 - BCL11A | 756 | 389 | 1183 |
| EXOSC5 - BCL11B | 1404 | 2520 | 1558 | EXOSC5 - BCL11B | 1368 | 1353 | 1455 |
| Ranking of EXOSC6 w.r.t BCL family | Ranking of BCL family w.r.t EXOSC6 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| EXOSC6 - BCL2L12 | 1327 | 1857 | 2063 | EXOSC6 - BCL2L12 | 2268 | 1527 | 478 |
| EXOSC6 - BCL6B | 1965 | 2525 | 1825 | EXOSC6 - BCL6B | 18 | 2334 | 2512 |
| EXOSC6 - BCL7A | 1676 | 787 | 944 | EXOSC6 - BCL7A | 593 | 2653 | 2037 |
| EXOSC6 - BCL9 | 1838 | 1059 | 1091 | EXOSC6 - BCL9 | 1846 | 851 | 1564 |
| EXOSC6 - BCL11A | 1677 | 1573 | 2217 | EXOSC6 - BCL11A | 596 | 2307 | 2547 |
| EXOSC6 - BCL11B | 1897 | 1736 | 1126 | EXOSC6 - BCL11B | 2094 | 2223 | 81 |
| Ranking of EXOSC7 w.r.t BCL family | Ranking of BCL family w.r.t EXOSC7 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| EXOSC7 - BCL2L12 | 1899 | 1755 | 974 | EXOSC7 - BCL2L12 | 1721 | 1551 | 1099 |
| EXOSC7 - BCL6B | 666 | 98 | 743 | EXOSC7 - BCL6B | 2730 | 2690 | 2689 |
| EXOSC7 - BCL7A | 2290 | 1501 | 1513 | EXOSC7 - BCL7A | 1282 | 831 | 1218 |
| EXOSC7 - BCL9 | 2363 | 1134 | 2219 | EXOSC7 - BCL9 | 1845 | 1234 | 328 |
| EXOSC7 - BCL11A | 1477 | 2239 | 1217 | EXOSC7 - BCL11A | 520 | 117 | 686 |
| EXOSC7 - BCL11B | 2396 | 1524 | 2037 | EXOSC7 - BCL11B | 1529 | 2720 | 1418 |
| Ranking of EXOSC8 w.r.t BCL family | Ranking of BCL family w.r.t EXOSC8 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| EXOSC8 - BCL2L12 | 2042 | 2152 | 506 | EXOSC8 - BCL2L12 | 1967 | 1525 | 2275 |
| EXOSC8 - BCL6B | 2469 | 2134 | 2224 | EXOSC8 - BCL6B | 190 | 1630 | 472 |
| EXOSC8 - BCL7A | 1175 | 1504 | 1743 | EXOSC8 - BCL7A | 2065 | 2351 | 1069 |
| EXOSC8 - BCL9 | 1733 | 2452 | 1164 | EXOSC8 - BCL9 | 2640 | 1895 | 1747 |
| EXOSC8 - BCL11A | 1864 | 906 | 1130 | EXOSC8 - BCL11A | 944 | 2303 | 532 |
| EXOSC8 - BCL11B | 1547 | 605 | 374 | EXOSC8 - BCL11B | 2581 | 2728 | 2359 |
| Ranking of EXOSC9 w.r.t BCL family | Ranking of BCL family w.r.t EXOSC9 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| EXOSC9 - BCL2L12 | 1179 | 1018 | 687 | EXOSC9 - BCL2L12 | 2105 | 1762 | 1453 |
| EXOSC9 - BCL6B | 437 | 852 | 1358 | EXOSC9 - BCL6B | 634 | 304 | 146 |
| EXOSC9 - BCL7A | 821 | 346 | 727 | EXOSC9 - BCL7A | 985 | 2017 | 2207 |
| EXOSC9 - BCL9 | 1305 | 1849 | 299 | EXOSC9 - BCL9 | 1197 | 1279 | 2154 |
| EXOSC9 - BCL11A | 892 | 2426 | 2011 | EXOSC9 - BCL11A | 481 | 441 | 1372 |
| EXOSC9 - BCL11B | 1569 | 549 | 1456 | EXOSC9 - BCL11B | 2606 | 1454 | 133 |
Table 121.
2nd order combinatorial hypotheses between EXOSC and BCL family.
| Unexplored combinatorial hypotheses | |
| EXOSC w.r.t BCL | |
| EXOSC2 | BCL-2L12/6B/7A/9/11A/11B |
| EXOSC3 | BCL-11B |
| EXOSC5 | BCL-2L12/6B/7A/9/11A/11B |
| EXOSC6 | BCL-7A/9/11A |
| EXOSC7 | BCL-6B/7A/11A |
| EXOSC8 | BCL-7A/11A/11B |
| EXOSC9 | BCL-2L12/6B/7A/9/11B |
| BCL w.r.t EXOSC | |
| EXOSC2 | BCL-6B/11A/11B |
| EXOSC3 | BCL-6B/7A/11A |
| EXOSC5 | BCL-6B/11A/11B |
| EXOSC6 | BCL-2L12/9 |
| EXOSC7 | BCL-2L12/7A/9/11A/11B |
| EXOSC8 | BCL-6B/11A |
| EXOSC9 | BCL-6B/9/11A/11B |
Table 138.
2nd order combinatorial hypotheses between ANTRX2 and COL family.
| Ranking ANTRX2 vs COL family | |||||||
| Ranking of ANTRX2 w.r.t COL family | Ranking of COL family w.r.t ANTXR2 | ||||||
| laplace | linear | rbf | laplace | linear | rbf | ||
| COL5A3-ANTXR2 | 2006 | 1473 | 2217 | COL5A3-ANTXR2 | 984 | 1782 | 933 |
| COL6A1-ANTXR2 | 1935 | 1061 | 1366 | COL6A1-ANTXR2 | 324 | 2211 | 398 |
| COL7A1-ANTXR2 | 1002 | 2119 | 1690 | COL7A1-ANTXR2 | 1722 | 956 | 2121 |
| COL9A2-ANTXR2 | 1498 | 552 | 1361 | COL9A2-ANTXR2 | 2391 | 80 | 135 |
| COL17A1-ANTXR2 | 1906 | 1086 | 780 | COL17A1-ANTXR2 | 576 | 2504 | 229 |
| COL28A1-ANTXR2 | 1409 | 2259 | 2296 | COL28A1-ANTXR2 | 478 | 1731 | 2362 |
Table 139.
2nd order combinatorial hypotheses between ANTRX2 and COL family.
| Unexplored combinatorial hypotheses | |
| ANTXR2 w.r.t COL | |
| COL5A3 | ANTXR2 |
| COL28A1 | ANTXR2 |
| COL w.r.t ANTXR2 | |
| COL7A1 | ANTXR2 |
| COL28A1 | ANTXR2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



