
Submitted:
20 January 2023
Posted:
24 January 2023
You are already at the latest version
Abstract
The impact of cyclic heat stress (CHS) and lighting events on pig feeding behavior (FB) was investigated. The FB of 90 gilts was recorded in real time under two ambient temperatures (AT): thermoneutrality (TN, 22 °C) or CHS (22°/35 °C). The day was divided into four periods: PI (06-08 h); PII (08-18 h); PIII (18-20 h); and PIV (20-06 h). Automatic and Intelligent Precision Feeders recorded each feed event for each pig. An estimated meal criterion (49 minutes) was used to calculate the FB variables. Feed behavior in both ATs followed a circadian pattern. The CHS reduced the feed intake by 6.9%. Pigs prioritized feed intake during the coolest hours of the day; however, nocturnal cooling did not allow the pigs to compensate for the reduced meal size due to CHS. The highest meal size and most of the meals were observed during the lighting-on period. Pigs reduced their interval between meals during periods II and III. The lighting program increased the meal size when the lights were switched on and reduced the meal size when the lights were switched off. Thus, the dynamics of the FB were largely influenced by AT, while the meal size was affected by the lighting program.
Keywords:
circadian rhythm
; feed pattern
; heat stress
; light program
; meal pattern
; swine
1. Introduction
The improper thermal environment of housing facilities and lighting programs are detrimental factors that affect pig growth performance. Ambient temperature (AT) has been related to the regulation of voluntary feed intake (VFI), triggering changes in feed behavior (FB). Finishing pigs exposed to high AT in cyclic heat stress (CHS) conditions reduce their VFI by 7.3% [1] and up to 50% in constant heat stress (HS) of 35 °C [2]. Reduction of VFI is a pig´s adaptive response in an attempt to decrease the metabolic heat production inherent to the digestive and metabolic processes [3]. In addition to the impact on VFI, the AT may change the FB based on the type of heat challenge. The FB dynamics of pigs reared in constant HS differ from those of pigs reared in CHS [4]. In a CHS challenge, when the AT returns to the thermoneutrality (TN) zone, pigs increase VFI rapidly to similar levels to unchallenged pigs or can temporarily exceed the normal level [5,6]. Regarding the behaviors that underlie feed intake, meal size, ingestion time per day, and occupation time of the feeding station decrease as AT increases from 19 to 29 °C, while the daily number of meals remains constant [7]. However, the impact on VFI is smaller for daily CHS compared to constant HS. This is because pigs show specific behavioral adaptations, such as reducing VFI during the warm periods of the day and increasing it during fresh periods, especially in the nocturnal period [5,8]. Since pigs are sensitive to their environment [9], lighting programs may also alter the feeding pattern of pigs. However, to our knowledge, no data regarding the effects of light events (turning the lights on and off) on FB for pigs on CHS are available.
A better understanding of pigs’ FB during CHS and light events could provide key information for improving productivity and animal well-being, especially in tropical climate areas where pigs are usually exposed to cyclic variation in AT [10]. Accordingly, a comparison between pigs with similar body weight (BW), genetics, and sex raised in similar facilities, management systems, and diets but in a different AT appears to be a good strategy to evaluate the effects of AT and light events on FB. The aforementioned factors affect FB in different ways. Even when all these influences are kept constant, individual differences still exist [11,12], making it difficult to compare studies that do not have the same overall experimental conditions. Thus, considering that only AT differs in the current study, we hypothesized that: I) pigs raised in CHS conditions prioritize their feed intake in the coolest hours of the day; II) pigs overconsume during the TN period of the CHS challenge, thus having a higher meal size than their counterpart raised in TN conditions; and III) light events modulate the meal size of pigs according to AT. Therefore, the objectives of this study were to: (i) investigate the effects of diurnal AT variation compared to TN conditions on FB and (ii) evaluate the impact of light events on FB in both AT conditions of finishing pigs.
2. Materials and Methods
2.1. Data Source
The animals, facilities, and experimental procedures used in this study were previously described [1]. In brief, information on the feed intake (FI) of 90 crossbred (Landrace × Large White) finishing gilts (Agroceres PIC Camborough) was recorded in real time by 10 feeder stations (Automatic and Intelligent Precision Feeder - AIPF; University of Lleida, Lleida, Spain). Details on AIPF functioning, trigger, feed delivery, and calibration were previously reported [13].
Pigs were shipped at the Swine Research Facilities of São Paulo State University (Unesp), Jaboticabal, SP, Brazil with 24.5 ± 2.9 kg of BW and group-housed in two similar facilities (45 pigs/facility) on a full concrete floor, with a single 95 m2 pen each, equipped with fans and an evaporative pad cooling system (Big Dutchman, Araraquara, SP, Brazil). Pigs had free access to water with low-pressure nipple drinkers and were fed ad libitum, individually, with diets formulated to meet or exceed their nutritional requirements according to NRC [14] recommendations. The lighting program was fixed to 12 h of artificial light (06–18 h) controlled by a timer switch.
All pigs had an exclusive electronic chip (plastic button tag containing passive transponders of radio frequency identification; Allflex, Joinville, SC, Brazil) inserted in the right ear, previously registered in the AIPF system. The exclusive electronic chip allowed any pig to access any of the feeders and, at the same time, to record the individual FI during the trial. Therefore, it enabled all animals to be housed in the same pen and allowed each individual to be an experimental unit.
For feed delivery, the chip in the right ear is identified when each pig introduces its head into the feeder, and when the pig triggers a push button to demand feed, the AIPF delivers a service to each requestion. The service was set to deliver the feed according to the experimental phase (EP), with 20 g during EP1 (0–20 days, 68 to 93 kg of BW) and 23 g during EP2 (21–38 Days 93 to 111 kg of BW). The size of the service was adjusted to be less than 150 services per pig per day following the manufacturer’s recommendation. A time lag (30 seg) was imposed between services to ensure that pigs consumed each serving before requesting a new one. The visit was made up of various requests, and hence, the total feed consumed in each visit was the sum of all requests. The pigs generally empty the feeder after each visit, but sometimes leave the feeder hopper with small amounts of feed (less than the service size), which is insignificant when considering the total FI in the day.
Regardless of the facility, during the growing phase (25 to 67 kg of BW), pigs were kept in similar conditions of AT (thermoneutrality), feeding management, and diets until achieving a BW of 67.7 ± 6.2 kg when the trial started (finishing phase). Pigs remained in the experiment for 45 days, which consisted of a 7-day adaptation period to experimental diets and CHS and a subsequent 38-day experimental phase, which was divided into EP1 (Days 0 to 20) and EP 2 (Days 21 to 38).
During the experimental period, in the facility with TN conditions, the AT was maintained at 22 °C, while in the other henceforth called CHS conditions, a diurnal cyclic variation in AT was set to fluctuate between 22 °C (20–08 h) and 35 °C (8–20 h) with the aid of thermostat-controlled heaters. An air humidifier (Hidrogiro, Ribeirão Preto, SP, Brazil) was used to increase the relative humidity of the air in the CHS condition (8–20 h) to decrease the evaporation rate and exacerbate the effect of HS. These temperatures and time periods aimed to mimic diurnal variation in ambient temperature based on studies with cyclic heat stress [10,15,16]. The AT and relative humidity were recorded daily at 30 min intervals throughout the experiment using a thermohygrometer (Instrutherm Mod. HT-70, São Paulo, BR).
2.2. Data Collection and Management
The visit of each pig to the AIPF was registered by a monitoring device that continuously and automatically recorded the start and end time of each visit (day, hour, minute, and second), the number of requests made by the pig, and the total amount of feed served in each visit. Collected information was used to build the initial database, initially managed using a Microsoft Excel spreadsheet. Feeding information collected on the days that pigs were handled (i.e., blood collection, dual-energy X-ray absorptiometry scanning, as described in [1]) was removed from the dataset (3 days). In the beginning, the initial dataset had 25,704 records, which were then analyzed using R Statistical Software (version 2.14.0; R Foundation, Austria). Initially, the dataset was evaluated by searching for the presence of outliers. Using graphical analysis, only 1 observation was removed from the dataset during the experimental period (visit longer than 3 h), which was considered a technical failure of the system. Observations of zero FI (0.06%) were excluded assuming that this visit may be associated with animal-feeder interactions, in which pigs quickly introduce and remove their head of the feeder and hence are recognized and recorded by the AIPF software as a visit
2.3. Definition of the Meal Criterion
A visit represents every time a pig introduces its head into the feeder. However, it does not imply a meal. A meal may be defined as an accumulation of sequential visits that were separated by an interval below a calculated meal criterion. The interval represents short breaks mainly for feeder change and water intake [17] along with social interactions that can occur during breaks within a meal. Thus, expressing FB data in terms of meals is considered a more appropriate unit.
The meal criterion used in this study was based on the analysis of the starting probability. This method defines a meal based on statistical measures rather than behavioral measures, which were previously described and recommended for situations where neither the within- nor the between-meal interval distributions are known [18]. For this purpose, the probability of the pigs starting a new feeding event within the next minute (pstart) was calculated over the day (all dataset) and for the TN period, as well as for the HS period, and plotted against the interval between visits to the feeder. The meal criterion was estimated as the first point that minimizes the pstart curve as previously described [18] and had been recently used in this type of dataset [8,19]. To reduce the effect of random variation in the pstart function, a simple moving average over 21-minute intervals was used.
2.4. Calculations and Statistical Analysis
Using the defined meal criterion (see Pstart and meal criterion section), a new database was created to group visits that have composed the same meal, resulting in a database with 13,834 records. In the new database, intervals between meals longer than 1440 min were arbitrarily excluded and assumed to be physiologically noncompatible, but this exclusion represents 0.8% of observations from the initial database, which is a percentage that does not compromise the final analysis. Based on the data grouped into meals, the number of meals (n/pig), meal size (feed intake, g/meal), meal duration (feeder occupancy, min/meal), feed intake rate (defined as the feed intake for each minute spent feeding over a meal, g/min/meal), and the interval between meals (min) of each animal were calculated. It should be noted that pig FB changes throughout the 24-h day occur regardless of AT [8,20]. In addition, the circadian rhythm may also change FB [21]. Thus, for a more adequate approach, the day was divided into four periods: PI (06-08 h), PII (08-18 h), PIII (18-20 h), and PIV (20-06 h). PI (06-08 h) and PII (08-18 h) together represent the diurnal period of the day, while PIII (18-20 h) and PIV (20-06 h) are the nocturnal period. PII (08-18 h) and PIII (18-20 h) were defined as the HS period of the day, while PI (06-08 h) and PIV (20-06 h) were the TN period.
All the data were analyzed using the MIXED procedure in SAS software (version 9.4; SAS Inst. Inc., Cary, NC, USA). The presence of outliers was evaluated through the residual analysis of data and by daily records of anomalies. The normality of studentized residuals was verified using the Cramer‒von Mises test. When needed, the response variable was transformed into a normal distribution approximation. If the data were transformed for analysis, least squares means were back-transformed to the original scale for reporting purposes. Each pig was considered an experimental unit. The model included the fixed effects of temperature (T), period of the day (P), experimental phase (EP), and their interactions (T×P, T×EP, P×EP, and EP×T×P) as follows:
where Yijmk is the observed variable (observation for the kth pig from the (i,j,m)th cell), μ is the overall mean, αi is the effect of the ith level of the T factor, βj is the effect of the jth level of the P factor, γm is the effect of the mth level of the EP factor, and their interactions (that is, ith level of T with the jth level of P, (αβ)ij; ith level of T with the mth level of EP, (αγ)im; jth level of P with the mth level of the EP, (βγ)jm; and ith level of T with the jth level of P with the mth level of the EP, (αβγ)ijm). The eijmk was considered the random error of the kth observation from (i,j,m)th.
Yijmk = µ + αi + βj + γm + (αβ)ij + (αγ)im + (βγ)jm + (αβγ)ijm + eijmk
The repeated measurement option was used with a compound symmetry covariance structure to account for the animal effect over sampling time, and the initial body weight was included as a covariate. Differences were considered significant if p ≤ 0.05. When there was a P×EP×P interaction effect (p < 0.05), adjusted means of P were compared in each T within each EP using the Tukey‒Kramer test. When there was a T×P interaction effect (p < 0.05), adjusted means of P were compared in each T using the Tukey‒Kramer test. Likewise, when there was a P×EP interaction effect (p < 0.05), adjusted means of P were compared in each EP.
3. Results
3.1. General Observations
The average AT and relative humidity registered in TN and CHS conditions during the trial are presented in Figure 1. In the TN condition, AT averaged 21.9 ± 0.4 °C and was maintained relatively constant throughout the experiment on a 24 h basis. Alternatively, by designing and following the objectives of the study, AT in CHS conditions varied during the day (p < 0.05). During the TN period in the CHS condition, AT averaged 22.4 ± 0.2 °C, while in the HS period, AT averaged 33.8 ± 0.3 °C. Relative humidity averaged 87 ± 1.7% and 79.6 ± 2.7% under the TN and CHS conditions, respectively.
Detailed information on growth performance and carcass composition was reported in our previous study [1], and individual pig behavior profiles showing the heterogeneity in FB response are available in Supplementary Figure S1. Briefly, in the overall growth performance evaluation, pigs in the CHS condition had a lower average daily feed intake, final BW, and average daily gain than pigs in the TN condition (p ≤ 0.05).
3.2. Pstart and Meal Criterion
A total of 24,128 visit records were used to calculate the pstart function. There was no difference in determining the meal criterion using the pstart methodology for exposure to different temperatures. Thus, the probability of pigs starting a new feeding event within the next minute after the last visit – over the day, during exposure to CHS (35 °C) or TN (22 °C) – was 49 min (Figure 2).
3.3. Feeding Behavior Responses
Overall, the feed intake (meal size × number of meal) of pigs throughout the experiment on a 24 h basis was 2,416 g and 2,268 g for the TN and CHS conditions, respectively, indicating that CHS reduced the FI by 6.13%.
The meal size continuously recorded in TN and CHS conditions followed a circadian pattern with typical diurnal FB (Figure 3A). Interestingly, the meal size of pigs decreased during the first hour after the light had been switched on, independent of the AT. However, during the entire light-on period (PI (06-08 h) + PII (08-18 h)), the meal size was 24.3% and 12.5% (on average) greater in TN and CHS, respectively, compared to the light-off period (PIII (18-20 h) + PIV (20-06 h)). In the TN condition, the hourly meal size peaked twice over the diurnal period, while in the CHS condition, it peaked once. Under TN conditions, the first peak took place 5 h after the light had been switched on (11 h), while the second peak took place before the end of the diurnal period (18 h). In the CHS condition, the diurnal peak took place 3 h after the light had been switched on (9 h). When the lights were switched off, pigs in the TN condition started to drop the meal size until 23 h, while in the CHS condition, an increase in meal size was observed between 19 and 21 h. Overall, switching off the lights reduced the meal size by 25.9% and 16.4% in TN and CHS conditions, respectively. The meal duration had similar behavior to meal size (Figure 3B).
Most feed intake, 77.2% and 73.2%, and meals 71.6% and 72.2%, were observed between 6 and 18 h in the TN and CHS conditions, respectively, which corresponded to the light-on period (Figure 3C). Irrespective of AT, the interval between meals started to decrease during the diurnal period, right after the light had been switched on (Figure 3D). Overall, the feed intake rate was quite similar throughout the 24 h-day (Figure 3E).
The FB features of the pigs are summarized in Table 1. Residuals of all dependent variables were normally distributed, and initial BW, as a covariate, was significant (p < 0.01) for all FB variables. All responses were influenced by the period of the day and experimental phase, except the number of meals (p < 0.01).
Interactions between P×T were detected for feed intake rate (Table 1). In the CHS condition, pigs had the highest feed intake rate during PIII (HS, 18-20 h), the lowest feed intake rate during the diurnal period, from 06 to 18 h (PI (TN, 06-08 h) + PII (HS, 08-18 h)), and an intermediate value during PIV (TN, 20-06 h) (p < 0.01, P×T). However, there was no difference in feed intake rate in the TN condition during the periods of the day. A P×EP interaction was detected for feed intake rate (Table 1). During EP1, pigs had a greater feed intake rate during PIII (18-20 h) than during the diurnal periods, while no effect of P was observed in EP2 (p = 0.01, P×EP, data not shown). In contrast, as expected, regardless of the period of the day, pigs during EP2 had a greater feed intake rate than those during EP1 (p = 0.01, data not shown).
The interaction between EP×T×P was significant for meal size, meal duration, and the interval between meals (Table 2). In terms of period effects within each temperature, in EPI (Days 0 to 20), in TN conditions, pigs had the highest meal size and duration in PII (08-18 h), while in the other period, similar values were observed (p < 0.01). Under CHS conditions, pigs had a decrease in meal size in PIII (HS, 18-20 h), while in the remaining period, similar values were observed (p < 0.01). The highest meal duration was observed in PII (HS, 08-18 h), and the lowest meal duration was observed in PIII (HS, 18-20 h) (p < 0.01). It should be noted that for both ATs, pigs had a decrease (p < 0.05) in meal size and duration from PII (08-18 h) to PIII (18-20 h) (after turning off the light). However, only for the CHS condition did pigs show an increased (p < 0.05) meal size and duration from PIII (HS, 18-20 h) to PIV (TN, 20-06 h). In both conditions, pigs had a lower interval between meals from 08 to 20 h (PII (08-18 h) + PIII (18-20 h)) (p < 0.01). In EPII (Days 21 to 38), pigs in TN conditions had the highest and lowest meal size and duration in PII (08-18 h) and PIII (18-20 h), respectively (p < 0.01). The highest interval between meals was observed in PIV (20-06 h), the lowest in PI (06-08 h), and intermediate values from 08 to 20 h (PII (08-18 h) + PIII (18-20 h)) (p <0.01). Under CHS conditions, the lowest meal size and duration were observed in PIII (HS, 18-20 h). Meal duration in the remaining periods was similar (p > 0.05). For both AT, pigs had a decrease (p < 0.05) in meal size and duration from PII (08-18 h) to PIII (18-20 h) (after turning off the light) and an increase (p < 0.05) in meal size and duration from PIII (18-20 h) to PIV (20-06 h). The highest interval between meals was observed in PIV (20-06 h), while the lowest was observed in PII (HS, 08-18 h) (p < 0.01).
Regarding temperature effects within each period, several changes were observed. Overall, the temperature reduced the values for all analyzed variables in all periods, except for the interval between meal in PIV (20-06 h) during EP1 and in PIII (18-20 h) during EP2 (p < 0.01).
4. Discussion
The current literature evaluating the effects of CHS and pig circadian rhythm on FB traits is scarce. Understanding the impact of AT variation and light events may aid in explaining the physiology and growth response of pigs under CHS conditions. Therefore, we hypothesized that pigs reared in CHS conditions prioritize feed intake in the coolest hours of the day, while light events alter the FB of pigs under cyclic AT variation over the day. Our major and original finding was that even when adapted, pigs in CHS conditions had a different FB when compared to TN pigs, which may partially explain differences in growth performance and body composition, as previously reported [1].
Feeding behavior is affected by animal-related factors such as sex [22], breed [8], body weight [23], circadian rhythm [21], ambient temperature [7], type of heat challenge (warm constant × cyclic temperatures; [4]), origin, age [24], diet, and handling. Therefore, a single experiment with pigs with similar characteristics, differing only in the AT condition, as described in the present study, seems to be more suitable to evaluate the impact of AT on FB. In the current study, it was possible to measure the FI over different periods of the day. Period II (08-18 h) and PIV (20-06 h) represented more than 80% of the diurnal and nocturnal period of a 24-h day. Thus, we used both periods to evaluate the dynamic impact of HS on FI. Alternatively, PI (06-08 h) and PIII (18-20 h) represented the period after switching the lights on and off, respectively, and herein, these periods were used to compare changes that occurred in FB due to light events.
Feed intake rate can be interpreted as voracious eating [21]. Voracious eating increases over time since body size and oral capacity become larger, which allows pigs to take larger bites and eat more feed in a shorter time [6], as observed in the current study.
Although changes in feed intake rate were observed in EP1 among periods of the day, no differences were detected during EP2. We believe that changes in feed intake rate in EP1 may be more closely associated with HS impact once a reduced feed intake is observed in pigs during the warmer period of the day. Since pigs under CHS conditions have been in an adaptation process during EP1, the higher voracity for CHS pigs in some specific periods of the day may have contributed to the overall mean, which could trigger the interaction effects. This concept is supported by the higher feed intake rate observed during the nocturnal period (the coolest part of the day). Alternatively, as long as pigs became more adapted to the CHS condition, they normalized their feed intake rate over the day, which resulted in a lack of period effect in the EP2. Our outcomes agree with Fraga et al. [8], who also did not observe changes in feed intake rate throughout the day for pigs (22 kg to 105 kg of body weight) raised under a similar cyclic AT protocol. To maintain a constant feed intake rate throughout the day under cyclic AT, pigs increase the meal size, which implies an increase in meal duration [25].
Moreover, an interaction between P×T was observed for feed intake rate for pigs in different AT conditions. In our study, as expected for pigs reared under unrestricted conditions [25], voracious eating was kept relatively constant throughout the day for TN pigs. However, it should be noted that for pigs under CHS conditions, voracious eating increased when the lights were switched off, while for TN, no changes were observed. Such behavior highlights that the light event modulated pigs’ FB according to the AT. As widely described in the literature [26], pigs in HS conditions reduce physical activity, which probably disrupted the normal feed intake rate of pigs in the current study. The increase in voracious eating associated with the light off event seems to be an attempt of pigs to overcome the lower feed intake rate during the HS diurnal period. Considering that a light-off event would be expected to lower the feed intake (positively correlated with voracious eating, [27]), the opposite behavior observed herein might be understood as a feeding motivation for pigs to try to compensate for the HS diurnal impact. In other words, the light-off event may have triggered some competition for feeder access, since group-housed pigs in restricted conditions increase voracity as a mechanism to maintain meal size levels [6].
The current study showed that meal size, duration, and interval between meals were affected by AT and P, which varied according to the EP. These variables are expected to be correlated because when the meal size increases, the meal duration increases simultaneously since the feed intake rate is relatively constant throughout the day for pigs under unrestricted conditions [25]. Irrespective of AT, as long as pigs increase the meal size and duration while keeping a constant number of daily meals, the interval between meals decreases [7]. According to the concept of satiety, when a pig is satiated at the end of a meal, the probability of starting a new meal soon is low but increases over time [19]. This finding supports the reduction in meal size and duration and the increased interval between meals observed for CHS pigs during the nocturnal compared to the diurnal period, irrespective of EP.
Regarding the P effect within each EP, the pattern of meal size, meal duration, and interval between meals for CHS pigs changed in comparison to TN pigs. During EP1, pigs in CHS conditions had similar meal sizes in the diurnal and nocturnal periods. Interestingly, during EP2, the meal size during the nocturnal period exceeded that observed for the diurnal period, while no changes in meal duration were observed. Since the animals had a constant number of daily meals, pigs increased the meal size in the same meal duration, which implied a higher feed intake. Taken together, these points reinforce that pigs raised under CHS use mechanisms associated with changes in the FB to adapt to AT conditions and to maintain a constant number of meals throughout the day.
Irrespective of the EP, our study demonstrated that pigs raised in TN reduced their meal size during the nocturnal period. In contrast, pigs raised in CHS had a similar meal size during the diurnal and nocturnal periods in EP1, while it was greater during the nocturnal period in EP2. Thus, our data support the hypothesis that pigs tried to compensate for the impact of CHS by eating more during the TN period of challenge. These results agree with previous studies that also evaluated pigs exposed to CHS (22 °C from 18 to 10 h and 30 °C from 10 to 18 h [8]) and that evaluated growing pigs exposed to temperature that fluctuated over the day by ± 4.5 °C approximately 28 °C [5]. However, it should be noted that the compensatory response was not enough to allow a similar meal size compared to pigs under TN conditions. Although pigs exhibit adaptive FB, the time required for FI recovery after exposure to HS to reach a similar level to that of TN pigs can vary between 2 and 5 days [28, 29, 30]. However, factors related to the physical capacity of the pigs´ digestive tract and the impact of successive heat waves might not have allowed a full recovery of feed consumption in the current study. In addition, changes in hormonal levels (e.g., decreased thyroxine) due to CHS trigger changes in physiologic and metabolic responses [1]. Thus, changes in the FB of pigs under CHS observed in the current study may be associated with the adaptation process to the cyclical AT condition over the day. This adaptation response may explain the differences in growth performance, as well as body composition, which was previously reported [1].
Pigs exhibit a predominant diurnal feeding behavior, with most meals (73% on average) consumed between 06 and 18 h [21,23]. Similar values were observed in our experiment, where pigs in the TN and CHS conditions had 71.6% and 72.2% of their meals, respectively, during the diurnal period. In the TN condition, the hourly meal size peaked twice over the diurnal period (at approximately 11 and 18 h), suggesting that this response may be related to the lighting program used [6]. In contrast, the meal size of pigs under CHS conditions, in addition to being lower, ended up in just a single peak at the beginning of the HS period (at approximately 9 h). This response may be an adaptation mechanism of pigs to HS in an attempt to decrease heat production and, consequently, the amount of heat that needs to be dissipated into the environment [31, 32].
According to our results, an interesting finding is that the meal size of pigs decreased during the first hour after the light-off event and started to increase shortly thereafter. This behavior suggests a strong influence of the circadian rhythm on pigs´ FB. In contrast, when the light was switched off, a huge drop in meal size was observed for pigs under TN conditions, and this response was maintained until the end of the day. Again, this response confirms the influence of lighting events on circadian rhythm and hence on FB. Interestingly, after the first hour of darkness, the meal size increased by 2 hours straight for pigs reared in CHS. This means that although the nocturnal period has begun, pigs continued to ingest feed, which suggests that CHS conditions affected the pigs´ circadian rhythm. This response coincides also with a higher feed intake rate observed over 24 h for pigs. In addition, our data show that the lighting program changed the meal size of pigs by a similar proportion during the light on or off events. However, the changes were different in each AT condition. Pigs in TN conditions reduced/increased the meal size by an average of 25%, while pigs in CHS conditions reduced/increased the meal size by 14.4%. The explanation for the lower impact of the lighting event on meal size for pigs raised in CHS may be associated with reduced FI during the HS period, as already discussed.
5. Conclusions
The present study indicates that light events stimulated feeding activity. However, CHS affected the pig feeding pattern related to the circadian rhythm. Our results support the initial hypothesis that pigs could prioritize their FI in the coolest hours of the day. Alternatively, nocturnal cooling did not allow the pigs to fully compensate for the FI depression caused by the HS period.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Individual pig behavior profiles of pigs during the experimental period throughout 24 h-day according to ambient temperature (Thermoneutral vs. Cyclic heat stress) and periods of the day: PI (06-08 h); PII (08-18 h); PIII (18-20 h); PIV (20-06 h).
Author Contributions
Conceptualization, M.J.K.O., L.H.; methodology, M.V.; I.A.; formal analysis, M.J.K.O.; M.V.; investigation, M.J.K.O; A.D.B.M.; D.A.M.; C.A.S.; G.A.C.V.; P.R.A. and J.P.R.G.; resources, L.H.; data curation, M.J.K.O.; M.V.; writing—original draft preparation, M.J.K.O; writing—review and editing, M.J.K.O.; M.V.; A.D.B.M.; D.A.M.; C.A.S.; G.A.C.V.; P.R.A.; J.P.R.G.; I.A.; L.H.; visualization, M.J.K.O; A.D.B.M.; D.A.M.; L.H.; supervision, I.A.; L.H.; project administration, L.H.; funding acquisition, L.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by São Paulo Research Foundation (FAPESP), grant 2018/15559-7. This study was possible thanks to the scholarship granted from the Brazilian National Council for Scientific and Technological Development (CNPq), grant 142555/2019-3 in the scope of the Program CAPES- PrInt (process number 88881.310312/2018-01).
Institutional Review Board Statement
Ethical review and approval were waived for this study due to this study had been developed using only database analysis. The original data were collected in a previously published study, which has the experimental procedures reviewed and approved by the Institutional Animal Care and Use Committee at Sao Paulo State University, SP, Brazil (protocol No. 3380/20).
Data Availability Statement
The data supporting the conclusions of this article will be made available by the authors upon request.
Acknowledgments
The authors would like to thank Seara Alimentos and Evonik Nutrition and Care GmbH for the donation in kind of animals and raw materials for the production of feed at the trial used to collect the data.
Conflicts of Interest
The authors declare no conflict of interest.
References
- de Oliveira, M.J.K.; Melo, A.D.B.; Marçal, D.A.; Valini, G.A.C.; Silva, C.A.; Veira, A.M.; Fraga, A.Z.; Arnaut, P.R.; Campos, P.H.R.F.; dos Santos, L.S.; Htoo, J.K.K.; Brand, H.G.; Hauschild, L. Effects of lowering dietary protein content without or with increased protein-bound and feed-grade amino acids supply on growth performance, body composition, metabolism, and acute-phase protein of finishing pigs under daily cyclic heat stress. J. Anim. Sci. 2022, skac387. [Google Scholar] [CrossRef]
- Shi, Z.B.; Ma, X.Y.; Zheng, C.T.; Hu, Y.J.; Yang, X.F.; Gao, K.G.; Wang, L.; Jiang, Z.Y. Effects of high ambient temperature on meat quality, serum hormone concentrations, and gene expression in the longissimus dorsi muscle of finishing pigs. Anim. Prod. Sci. 2016, 57, 1031–1039. [Google Scholar] [CrossRef]
- Baumgard, L.H.; Rhoads Jr, R.P. Effects of heat stress on postabsorptive metabolism and energetics. Annu. Rev. Anim. Biosci. 2013, 1, 311–337. [Google Scholar] [CrossRef] [PubMed]
- Xin, H.; DeShazer, J.A. Feeding patterns of growing pigs at warm constant and cyclic temperatures. Trans. Am. Soc. Agric. Eng. 1992, 35, 319–323. [Google Scholar] [CrossRef]
- Quiniou, N.; Massabie, P.; Granier, R. Diurnally variation of ambient temperature around 24 or 28 C: Influence on performance and feeding behavior of growing pigs. 2000, In Swine Housing Conference (p. 1). American Society of Agricultural and Biological Engineers.
- Bus, J.D.; Boumans, I.J.M.M.; Webb, L.E.; Bokkers, E.A.M. The potential of feeding patterns to assess generic welfare in growing-finishing pigs. Appl. Anim. Behav. Sci. 2021, 241, 105383. [Google Scholar] [CrossRef]
- Quiniou, N.; Dubois, S.; Noblet, J. Voluntary feed intake and feeding behaviour of group-housed growing pigs are affected by ambient temperature and body weight. Livest. Prod. Sci. 2000, 63, 245–253. [Google Scholar] [CrossRef]
- Fraga, A.Z.; Hauschild, L.; Campos, P.H.R.F.; Valk, M.; Bello, D.Z.; Kipper, M.; Andretta, I. Genetic selection modulates feeding behavior of group-housed pigs exposed to daily cyclic high ambient temperatures. PloS one 2022, 17, 1, e0258904. [Google Scholar] [CrossRef]
- Adelowo, O.V.; Adebiyi, O.A.; Odu, O. Effects of pen colour and photoperiod interactions on the performance and respiratory rate of finisher gilts. Niger. J. Anim. Prod. 2018, 45, 58–65. [Google Scholar] [CrossRef]
- Cervantes, M.; Cota, M.; Arce, N.; Castillo, G.; Avelar, E.; Espinoza, S.; Morales, A. Effect of heat stress on performance and expression of selected amino acid and glucose transporters, HSP90, leptin and ghrelin in growing pigs. J. Therm. Biol. 2016, 59, 69–76. [Google Scholar] [CrossRef]
- Brown-Brandl, T.M.; Rohrer, G.A.; Eigenberg, R.A. Analysis of feeding behavior of group housed growing-finishing pigs. Comput. Electron. Agric. 2013, 96, 246–252. [Google Scholar] [CrossRef]
- Hessel, E.F.; Van den Weghe HF, A. Individual online-monitoring of feeding frequency and feeding duration of group-housed weaned piglets via high frequent radiofrequency identification (HF RFID). 2011. In: Lokhorst C, Berckmans D, editors. Papers presented at the 5th European Conference on Precision Livestock Farming, 210–222, Prague, Czech Republic.
- Pomar, J.; López, V.; Pomar, C. Agent-based simulation framework for virtual prototyping of advanced livestock precision feeding systems. Comput. Electron. Agric. 2011, 78, 88–97. [Google Scholar] [CrossRef]
- NRC (National Research Council). Nutrient requirements of swine. 2012, 12th ed. Washington, DC: National Academy Press.
- Morales, A.; Cota, S.E.M.; Ibarra, N.O.; Arce, N.; Htoo, J.K.; Cervantes, M. Effect of heat stress on the serum concentrations of free amino acids and some of their metabolites in growing pigs. J. Anim. Sci. 2016, 94, 2835–2842. [Google Scholar] [CrossRef]
- Lan, R.; Kim, I. Effects of feeding diets containing essential oils and betaine to heat-stressed growing-finishing pigs. Arch. Anim. Nutr. 2018, 72, 368–378. [Google Scholar] [CrossRef] [PubMed]
- Morgan, C.A.; Emmans, G.C.; Tolkamp, B.J.; Kyriazakis, I. Analysis of the feeding behavior of pigs using different models. Physiol. Behav. 2000, 68, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Howie, J.A.; Tolkamp, B.J.; Avendano, S.; Kyriazakis, I. A novel flexible method to split feeding behaviour into bouts. Appl. Anim. Behav. Sci. 2009, 116, 116,101–109. [Google Scholar] [CrossRef]
- Gomes, B.C.; Andretta, I.; Valk, M.; Pomar, C.; Hauschild, L.; Fraga, A.Z.; Kipper, M.; Trevizan, L.; Remus, A. Prandial Correlations and Structure of the Ingestive Behavior of Pigs in Precision Feeding Programs. Animals, 2021, 11, 2998. [Google Scholar] [CrossRef]
- Santos, L.S.; Pomar, C.; Campos, P.H.R.F.; da Silva, W.C.; Gobi, J.P.; Veira, A.M.; Fraga, A.Z.; Hauschild, L. Precision feeding strategy for growing pigs under heat stress conditions. J. Anim. Sci. 2018, 96, 4789–4801. [Google Scholar] [CrossRef] [PubMed]
- Andretta, I.; Pomar, C.; Kipper, M.; Hauschild, L.; Rivest, J. Feeding behavior of growing–finishing pigs reared under precision feeding strategies. J. Anim. Sci. 2016, 94, 3042–3050. [Google Scholar] [CrossRef] [PubMed]
- Renaudeau, D.; Giorgi, M.; Silou, F.; Weisbecker, J.L. Effect of breed (lean or fat pigs) and sex on performance and feeding behaviour of group housed growing pigs in a tropical climate. Asian-australas. J. Anim. Sci. 2006, 19, 4, 593–600. [Google Scholar] [CrossRef]
- Remus, A.; Hauschild, L.; Létourneau-Montminy, M.P.; Andretta, I.; Pomar, C. Feeding behavior of growing and finishing pigs fed different dietary threonine levels in a group-phase feeding and individual precision feeding system. Transl. Anim. Sci. 2020, 4, txaa177. [Google Scholar] [CrossRef]
- Nielsen, B.L. On the interpretation of feeding behaviour measures and the use of feeding rate as an indicator of social constraint. Appl. Anim. Behav. Sci., 1999, 63, 79–91. [Google Scholar] [CrossRef]
- Baumung, R.; Lercher, G.; Willam, A.; Sölkner, J. Feed intake behaviour of different pig breeds during performance testing on station. Arch. Anim. Breed. 2006, 49, 77–88. [Google Scholar] [CrossRef]
- Collin, A.; Van Milgen, J.; Dubois, S.; Noblet, J. Effect of high temperature and feeding level on energy utilization in piglets. J. Anim. Sci. 2001, 79, 1849–1857. [Google Scholar] [CrossRef] [PubMed]
- Rauw, W.M.; Soler, J.; Tibau, J.; Reixach, J.; Gomez Raya, L. Feeding time and feeding rate and its relationship with feed intake, feed efficiency, growth rate, and rate of fat deposition in growing Duroc barrows. J. Anim. Sci. 2006, 84, 3404–3409. [Google Scholar] [CrossRef]
- Renaudeau, D. Impact of single or repeated short-term heat challenges mimicking summer heat waves on thermoregulatory responses and performances in finishing pigs. Transl. Anim. Sci. 2020, 4, txaa192. [Google Scholar] [CrossRef]
- Kim, B.; Reddy, K.E.; Kim, H.R.; Kim, K.H.; Lee, Y.; Kim, M.; Ji, S.Y.; Lee, S.D.; Jeong, J.Y. Effects of recovery from short-term heat stress exposure on feed intake, plasma amino acid profiles, and metabolites in growing pigs. J. Anim. Sci. Technol. 2021, 63, 531–544. [Google Scholar] [CrossRef]
- Mayorga, E.J.; Kvidera, S.K.; Horst, E.A.; Al-Qaisi, M.; Dickson, M.J.; Seibert, J.T.; Lei, S.; Keating, A.F.; Ross, J.W.; Rhoads, R.P.; Rambo, Z.J.; Wilson, M.E.; Baumgard, L.H. Effects of zinc amino acid complex on biomarkers of gut integrity and metabolism during and following heat stress or feed restriction in pigs. J. Anim. Sci. 2018, 96, 4173–4185. [Google Scholar] [CrossRef] [PubMed]
- Quiniou, N.; Noblet, J.; van Milgen, J.; Dubois, S. Modelling heat production and energy balance in group-housed growing pigs exposed to low or high ambient temperatures. Br. J. Nutr. 2001, 85, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Collin, A.; van Milgen, J.; Le Dividich, J. Modeling the effect of high, constant temperature on food intake in young growing pigs. Anim. Sci. 2001, 72, 519–527. [Google Scholar] [CrossRef]
Figure 1.
Average ambient temperature (a) and air relative humidity (b) were measured in thermoneutral (TN) and cyclic heat stress (CHS) conditions, with the aid of a thermohygrometer, at 30-min intervals, during the experimental days. Each vertical bar represents the standard error of the mean.
Figure 1.
Average ambient temperature (a) and air relative humidity (b) were measured in thermoneutral (TN) and cyclic heat stress (CHS) conditions, with the aid of a thermohygrometer, at 30-min intervals, during the experimental days. Each vertical bar represents the standard error of the mean.
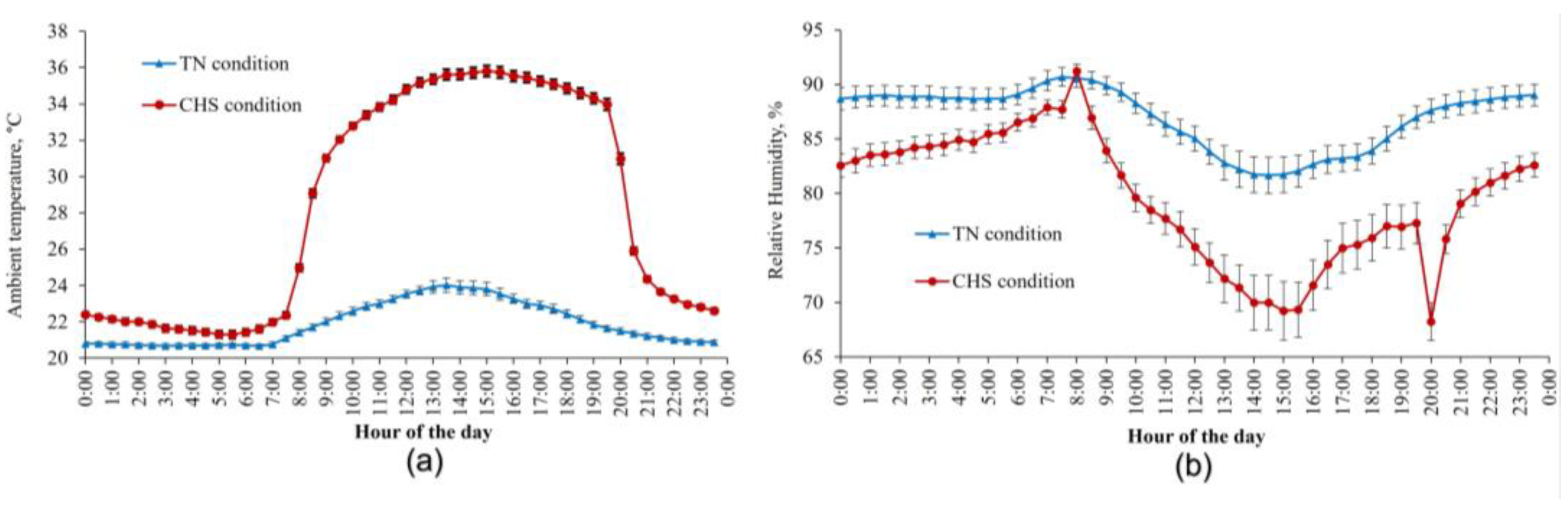
Figure 2.
Probability of pigs starting a new feeding event (pstart) within the next minute since the last visit – over the day, or during exposure to cyclic high ambient temperature (35 °C, 08-20 h) or thermoneutrality (22 °C, 21-08 h).
Figure 2.
Probability of pigs starting a new feeding event (pstart) within the next minute since the last visit – over the day, or during exposure to cyclic high ambient temperature (35 °C, 08-20 h) or thermoneutrality (22 °C, 21-08 h).
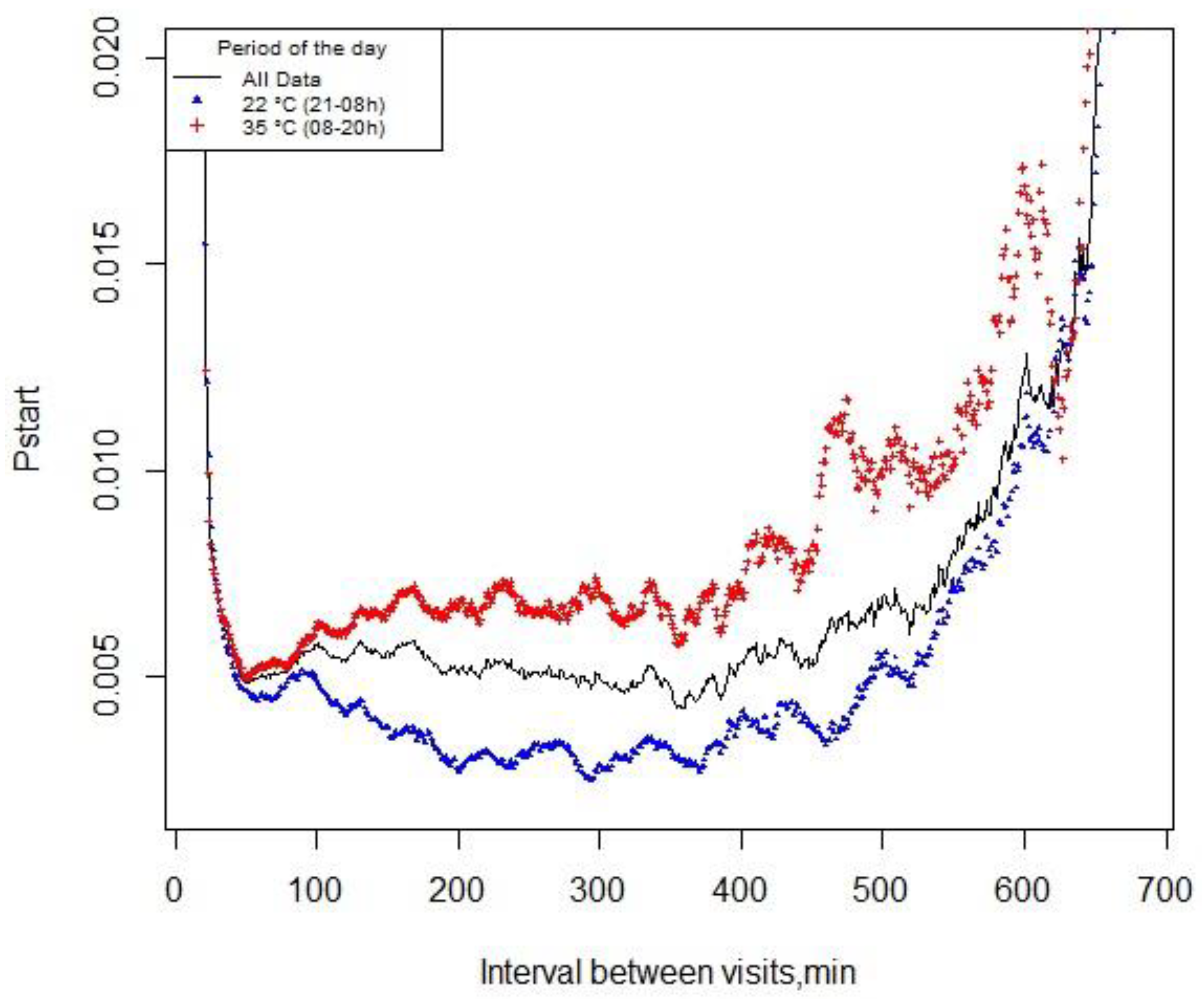
Figure 3.
Circadian variation of (a) Meal size, (b) Meal duration, (c) Number of meals, (d) Interval between meals, and (e) Feed intake rate for finishing-pigs reared in Thermoneutrality (TN, 24 h at 22 °C) or Cyclic Heat Stress (CHS, 12 h at 22 °C, 20-08 h, and 12 h at 35 °C, 8-20 h) conditions during 38-days of the experimental period. Periods of the day: PI (06-08 h); PII (08-18 h); PIII (18-20 h); PIV (20-06 h). Average data of 45 pigs per AT condition throughout 24 h-day, regardless of the experimental phase. The lighting program was fixed to 12 h of artificial light (06 to 18 h).
Figure 3.
Circadian variation of (a) Meal size, (b) Meal duration, (c) Number of meals, (d) Interval between meals, and (e) Feed intake rate for finishing-pigs reared in Thermoneutrality (TN, 24 h at 22 °C) or Cyclic Heat Stress (CHS, 12 h at 22 °C, 20-08 h, and 12 h at 35 °C, 8-20 h) conditions during 38-days of the experimental period. Periods of the day: PI (06-08 h); PII (08-18 h); PIII (18-20 h); PIV (20-06 h). Average data of 45 pigs per AT condition throughout 24 h-day, regardless of the experimental phase. The lighting program was fixed to 12 h of artificial light (06 to 18 h).
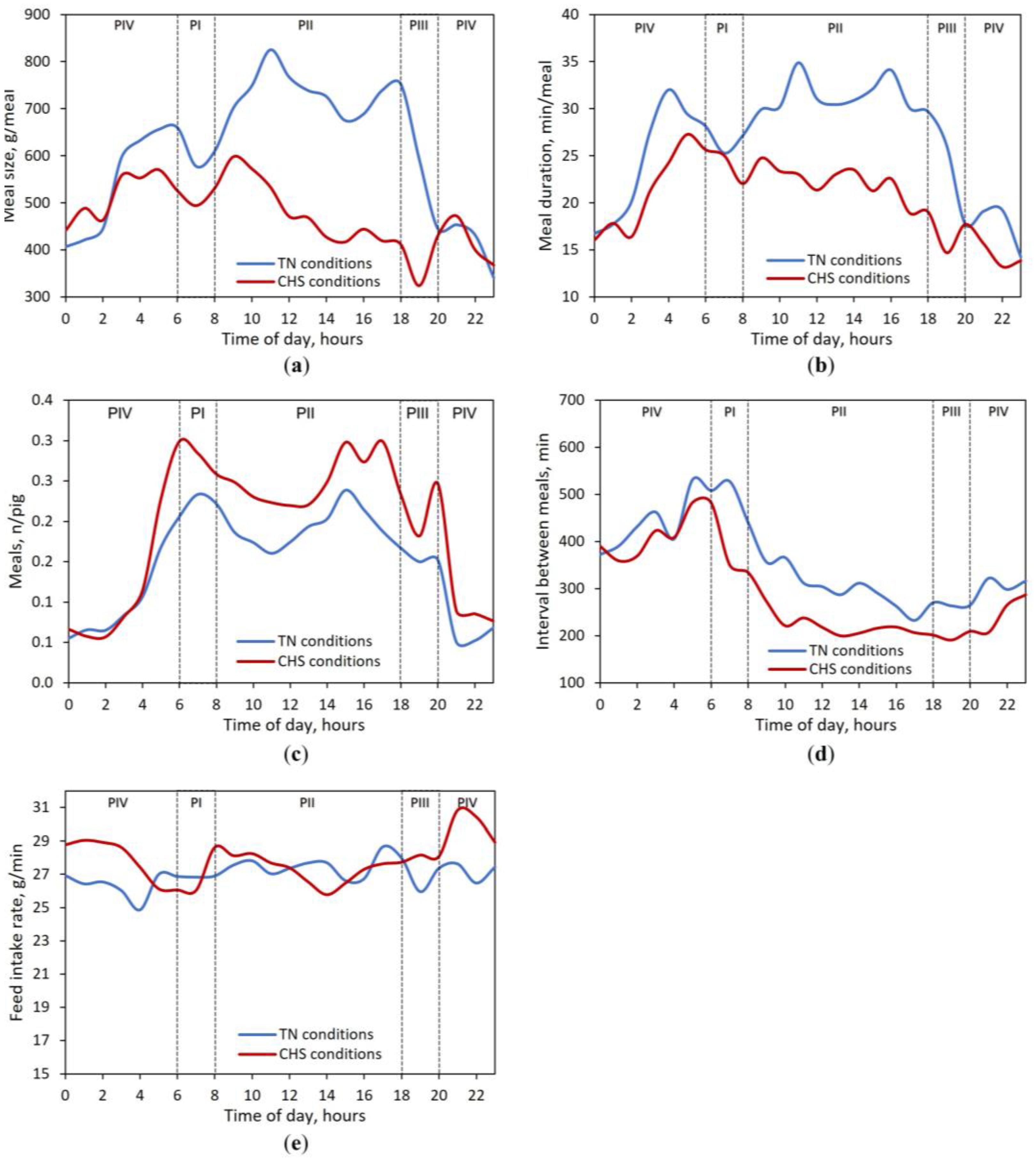

Table 1.
Feeding behavior of finishing pigs exposed to thermoneutrality or cyclic heat stress conditions, according to the period of the day1,2.
Table 1.
Feeding behavior of finishing pigs exposed to thermoneutrality or cyclic heat stress conditions, according to the period of the day1,2.
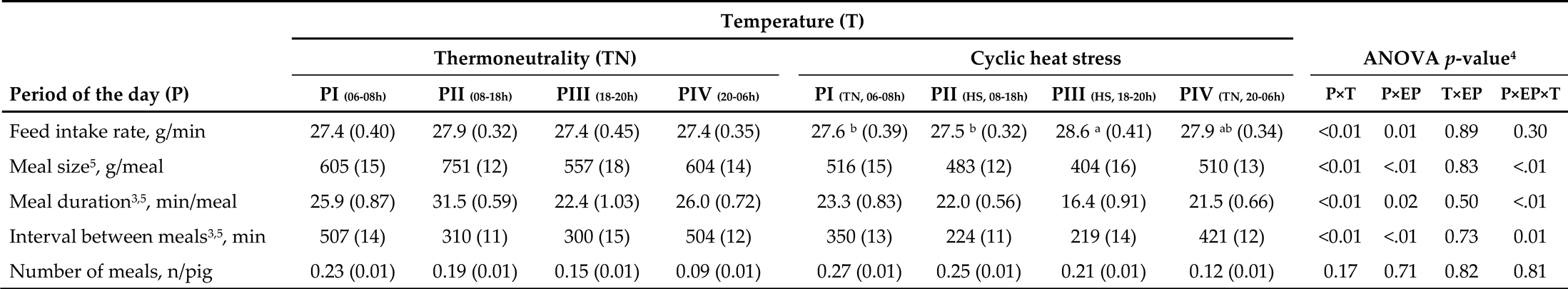 |
¹ Data after application of meal criterion defined as 49 min and expressed per hour of the day, since the periods of the day had a different duration. The thermoneutrality condition was 24 h at 22 °C, while the heat stress (HS) in cyclical conditions was 12 h at 22 °C, 20-08 h, and 12 h at 35 °C, 8-20 h. ² Least squares means and standard error (SE). ³ Meal duration and the interval between meals were log-transformed for statistical analysis and back-transformed to the original scale for reporting purposes. 4 Probability of periods of the day (n = 4; P), temperature (n = 2; T), experimental phase (n = 2; EP), and interactions between P×T, P×EP, T×EP, and P×EP×T. For variables whose interaction was significant, average values were analyzed by Tukey’s test. Initial BW as a covariate was significant for all variables; p < 0.01. 5 The P×EP×T interaction for feed intake, meal duration, and the interval between meals are presented in Table 2. a,b Values within a row and temperature, with different superscripts, differed during the period of the day (P×T, p < 0.01).
Table 2.
Feeding behavior of finishing pigs exposed to thermoneutrality conditions or cyclic heat stress conditions, according to the period of the day in each experimental phase1,2.
Table 2.
Feeding behavior of finishing pigs exposed to thermoneutrality conditions or cyclic heat stress conditions, according to the period of the day in each experimental phase1,2.
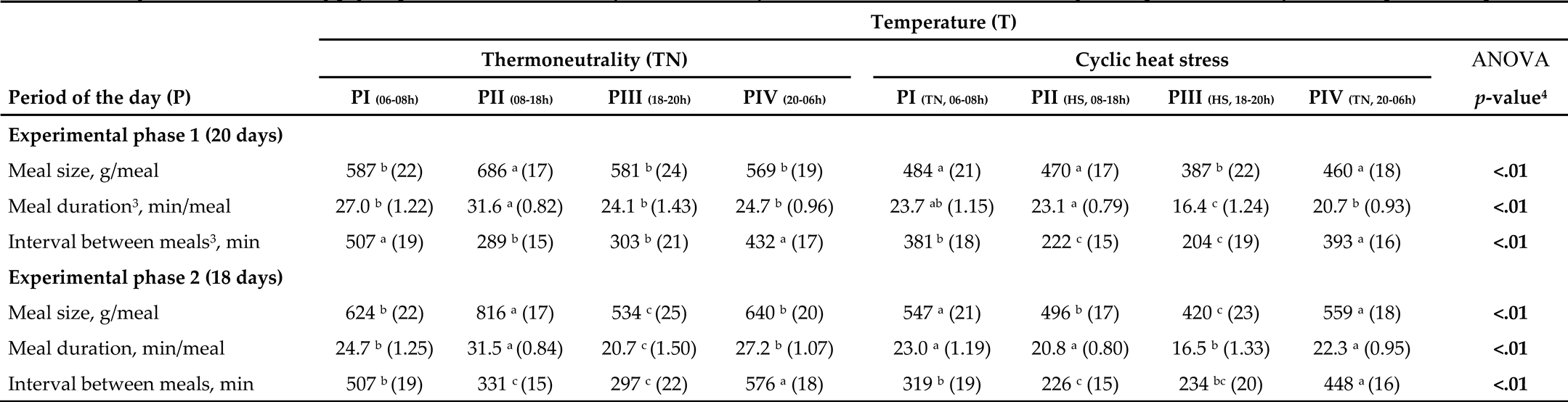 |
¹ Data after application of meal criterion defined as 49 min and expressed per hour of the day since the periods of the day had a different duration. The thermoneutrality condition was 24 h at 22 °C, while the heat stress (HS) set in cyclic condition was 12 h at 22 °C, 20-08 h, and 12 h at 35 °C, 8-20 h. ² Least squares means and standard error (SE). ³ Meal duration and the interval between meals were log-transformed for statistical analysis and back-transformed to the original scale for reporting purposes. 4 Statistical analyses between periods of the day in each experimental phase for control and challenged barn. a-e Values within a row with different superscripts differed between periods of the day in each barn (p < 0.05) according to Tukey’s test.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



