
Submitted:
20 September 2023
Posted:
22 September 2023
You are already at the latest version
Abstract
This review presents an updated overview of research concerning Leishmania protein structures, primarily sourced from the Protein Data Bank (PDB), that play a role in the metabolic pathways of the Leishmania parasite. Furthermore, we assess the current progress in the identification and development of bioactive chemical agents aimed at addressing this substantially overlooked tropical disease. We have analyzed experimental data obtained from in vitro, in vivo and in silico sources. This data has been categorized into four main areas: a) vector taxonomy and geographic distribution; b) parasite taxonomy and geographic distribution; c) enzymatic functions of proteins engaged in parasite/host interactions throughout various developmental stages (such as oxidoreductases, transferases, hydrolases, lyases, isomerases, ligases, and cytokines); and d) established and experimental treatments employing bioactive chemical compounds. Our objective is to establish a foundational point of reference for research efforts concentrating on the elucidation of interaction mechanisms and the processes of ligand-protein activation/inactivation, specifically linked to Leishmania infections. Consequently, we emphasize enzymes recognized for their involvement in the biochemical pathways incited during Leishmania infection episodes. This review encapsulates the current understanding, offering insights to inform and direct future explorations aimed at targeting proteins and pathways to enhance the management of Leishmania-related diseases.
Keywords:
Leishmania
; Trypanosoma
; Molecular Dynamics
; Theoretical Chemistry
1. Overview
Leishmaniasis constitutes a group of tropical and subtropical neglected zoonotic diseases, caused by various species within the Leishmania genus. [1,2,3,4] These diseases predominantly affect mammals and are transmitted via the bites of infected female sandflies. [5,6,7,8,9] The World Health Organization (WHO) classifies leishmaniasis as a Neglected Tropical Disease (NTD) due to the limited availability of effective treatments and insufficient investment in research and development. [3,10,11,12,13] Consequently, instances of the predominant forms of leishmaniasis - cutaneous (CL) and visceral (VL) - continue to escalate globally. While mucocutaneous (MCL) leishmaniasis also exists, WHO surveillance concentrates on the more widespread CL and VL variants. CL, the most prevalent clinical presentation worldwide,[14,15] is exemplified by cutaneous lesions (refer to Figure 1). On the other hand, VL profoundly affects internal organs such as the spleen and liver, rendering it more perilous and potentially lethal. In the year 2021, Brazil documented the highest global count of cases for both cutaneous leishmaniasis (CL) and visceral leishmaniasis (VL), registering 16,800 cases of CL and 1,492 cases of VL. Colombia followed with the second-highest figures, reporting 6,175 CL cases and 7 VL cases. These combined statistics constituted 71% of all leishmaniasis instances in the New World.[16] However, when comparing the disease’s prevalence, Colombia exhibited a higher prevalence rate (0.012%) in contrast to Brazil’s rate (0.0075%). Regarding incidence, Brazil disclosed 6.94 CL cases per 100,000 individuals, while Colombia documented 12.10 cases per 100,000 individuals. Based on the most recent report from the World Health Organization (WHO) encompassing the culmination of the past decade (2021), these data underscore the ongoing significance of leishmaniasis as a pertinent public health concern in both nations.
Figure 1.
Number of Visceral and Cutaneous Leishmaniasis cases reported by the World Health Organization (WHO). The figure shows the most relevant reports in the world (new and old world).
Figure 1.
Number of Visceral and Cutaneous Leishmaniasis cases reported by the World Health Organization (WHO). The figure shows the most relevant reports in the world (new and old world).
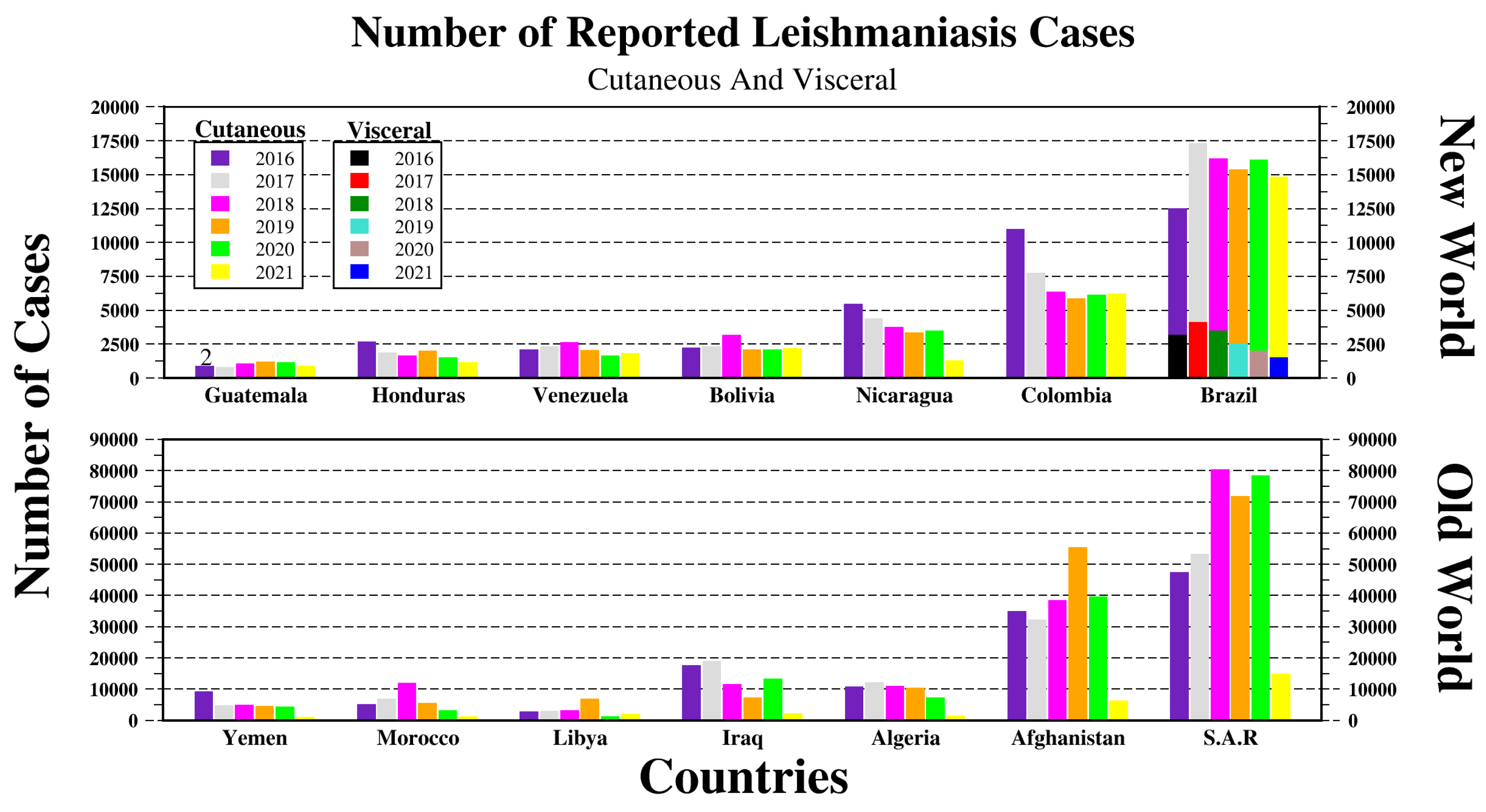
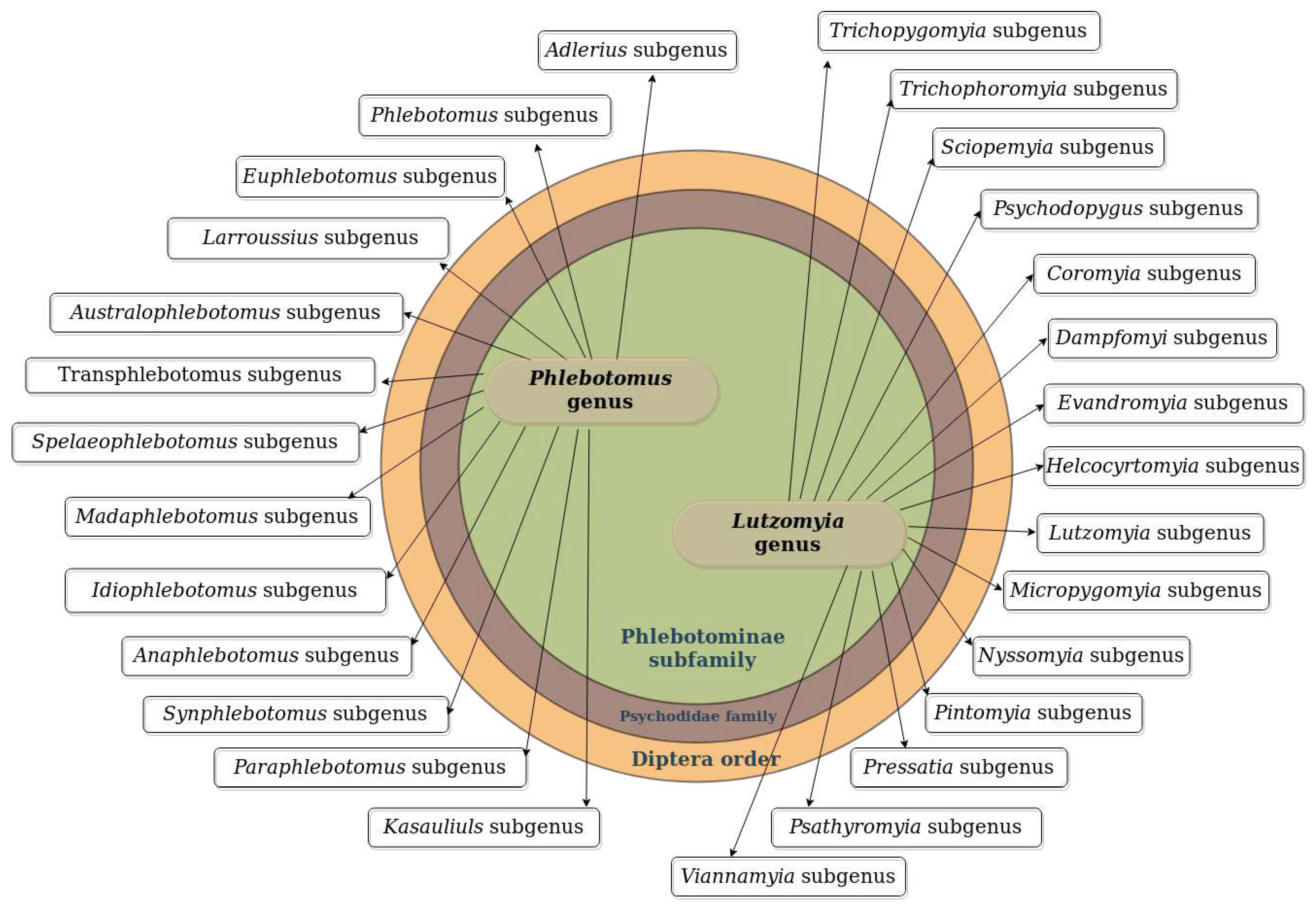
Figure 2.
Taxonomic distribution of Leishmania vector: Diptera order and Psychodidae family, phlebotomiae subfamily. Gographical distribution: Phlebotomus genus for the old world and Lutzomyia genus for the new world. [19,25]
Leishmania species are taxonomically grouped and classified into two main divisions called "sections": Euleishmania and Paraleishmania [17,18]. These divisions were proposed by Cupolillo et al. (2000) based on molecular and phylogenetic studies of Leishmania species, using the genus Endotrypanum as an external phylogenetic group to construct a dendrogram. The phylogenetic results demonstrated evolutionary divergences among several established Leishmania species (Nei’s genetic distance (D) = 1.04). Cupolillo utilized various molecular techniques including Multilocus Enzyme Electrophoresis (MLEE), Restriction Fragment Length Polymorphism (RFLP) analysis of the intergenic transcribed spacers (ITSrRNA) in the rRNA gene cluster, measurement of sialidase activity, and primary DNA sequencing of the small subunit (SSU) rRNA gene to compare and verify their results [17,19,20].
According to Cupolillo and colleagues, additional species have been included in the two proposed divisions. Currently, Euleishmania encompasses the following subgenera: L. (Viannia), L. (Leishmania), and L. (Sauroleishmania) (Figure 3) [17,21,22,23]. On the other hand, Paraleishmania includes the subgenus L. (Endotrypanum), which consists of the species E. schaudinni and E. monterogeii (See Figure 3) [24,25,26,27]. Specifically in Colombia, reports indicate the presence of L. amazonensis, L. braziliensis, L. mexicana, L. colombiensis, L. guyanensis, L. panamensis, L. chagasi, L. lainsoni, and L. equatoriensis as transmission parasites of leishmaniasis. [28,29,30,31]
Presently, the standard active compounds employed against Leishmania parasites encompass Amphotericin B, Miltefosine, Pentamidine, Antimonials, Paromomycin, Sitamaquine, Pamidronate, Azoles, and Nucleoside analogues. [32,33,34,35,36,37,38] Nonetheless, these medications frequently exhibit inefficacy and pronounced toxicity in patients undergoing clinical treatments. The efficacy and toxicity profiles of these drugs hinge on diverse factors, including the immunological status of the afflicted host and the pharmacokinetic attributes intrinsic to the drugs themselves.
For instance, drugs based on Miltefosine display teratogenic effects, while antimonial treatments can result in unfavorable outcomes such as arthralgia, nausea, abdominal discomfort, pancreatitis, and cardiotoxicity. [39,40,41]. The experimental development and assessment of efficacy for potential antileishmanial compounds have proven to be intricate and financially demanding, often yielding a modest success rate. The creation of novel drugs is undeniably a multifaceted undertaking.
As an alternative approach, computational methods like Molecular Dynamics and Molecular Docking offer a more viable avenue for investigating potential antileishmanial compounds and protein targets. These methodologies enable the examination of numerous interactions between ligands and proteins at a relatively modest computational expense, furnishing a comprehensive atomistic and molecular portrayal when compared to empirical assays.
The exploration of innovative pharmaceuticals arising from alternative chemical sources, including secondary metabolites and peptides obtained from plant and animal species, presents significant promise in propelling advancements towards more potent and efficacious treatments in contrast to traditional methodologies.
The primary aim of this study is to review the state-of-the-art protein targets concerning the Leishmania parasite. We executed a methodical inquiry across various databases, comprising the Protein Data Bank (PDB), literary works, and scholarly articles. Additionally, we scrutinize alternative compounds, encompassing those originating from minor secondary metabolites and proteins.
2. Leishmania Vectors
Sandflies play a crucial role as vectors for various dangerous parasites, including Leishmania, Bartonella bacilliformis bacteria, and certain viruses, which can have severe consequences, even leading to the death of the host [42,43,44,45,46]. These tiny insects belong to the Diptera order, Psychodidae family, and Phlebotominae subfamily. Taxonomically, sandflies are classified into five genera: Phlebotomus and Sergentomyia, prevalent in the Old World, and Lutzomyia, Brumptomyia, and Warileya, prevalent in the New World [47,48,49]. Among these genera, Phlebotomus and Lutzomyia are particularly responsible for the transmission of Leishmania parasites [47,50] (See Figure 2).
Nine species of the Leishmania genus and fourteen species belonging to the Lutzomyia genus have been reported for Colombia. [16,29] Parasite species reported for Colombia are L. amazonensis, L. braziliensis, L. mexicana, L. colombiensis, L. guyanensis L. panamensis, L. infantum, L. lainsoni and L. equatoriensis (ver Figure 3). Lutzomyia species Colombia reported are L. flaviscutellata, L. colombiana, L. spinicrassa, L. pia, L. towsendi, L. hartmanni, L. umbratilis, L. longiflocosa, L. trapidoi, L. panamensis, L. yuli yuli, L. cruciata, L. columbiana and L. gomezi.[16,51,52]
3. Leishmania Life Cycle and Host Immune Response
Leishmania parasites invade, develop, and replicate inside the host Mononuclear Phagocyte System (MPS), targeting macrophages and dendritic cells [53,54,55,56]. The incubation period for the transformation of Leishmania promastigotes to amastigotes ranges from two to three months, during which the host’s immune system response determines the outcome, whether favorable or unfavorable [54]. The parasite cycle initiates with the bite of an infected female sandfly carrying promastigote Leishmania parasites in a metacyclic state (see Figure 4) [57,58,59]. At this stage, neutrophils and macrophages are the first line of defense activated by the immune system, with neutrophils initiating the inflammatory response [56,58,60].
The Leishmania parasite employs two mechanisms to enter macrophages: a direct pathway involving the macrophage itself and an indirect pathway through the attack on neutrophils [61]. In the direct pathway, promastigotes are endocytosed by macrophages and enclosed within a phagosome, which subsequently undergoes biochemical transformation into a phagolysosome [62]. The indirect pathway involves the phagocytosis of the parasite by neutrophils, followed by subsequent phagocytosis by macrophages (see Figure 4) [61].
Within the phagolysosome vacuole, the promastigote parasite state survives by producing specific factors such as Lipophosphoglycan (LPG), gp63 protein, and glutathione transferase [57,63].
Following the initial attack on macrophages, the host immune system responds by triggering a biochemical cascade that involves the production of cytokines by specialized CD4 T-helper (T) cells, specifically T1 or T2 cells. Among the cytokines secreted by T1 cells, gamma interferon (IFN-) plays a crucial role. IFN- activates and stimulates macrophages, enhancing their antimicrobial activity. Additionally, interleukin (IL)-12 and IL-2, which are also secreted by T1 cells, promote the differentiation of CD4 T cells into the T1 phenotype. This phenotypic transformation is important for an effective response against the exogenous attack, in this case, the metacyclic promastigote form of Leishmania [64,65].
The immune T2 response produces IL-4, IL-5, IL-10, the Transforming Growth Factor Beta (TGF-) and other cytokines. These compound are perfectly suited for facing allergenic diseases or helmintic infections, therefore, is more desirable the T1 response. Moreover, the T2 response inhibits the T1, favoring the propagation and survival of the Leishmania parasite.[64,66]
4. Leishmania Proteomics and Metabolomics Analysis
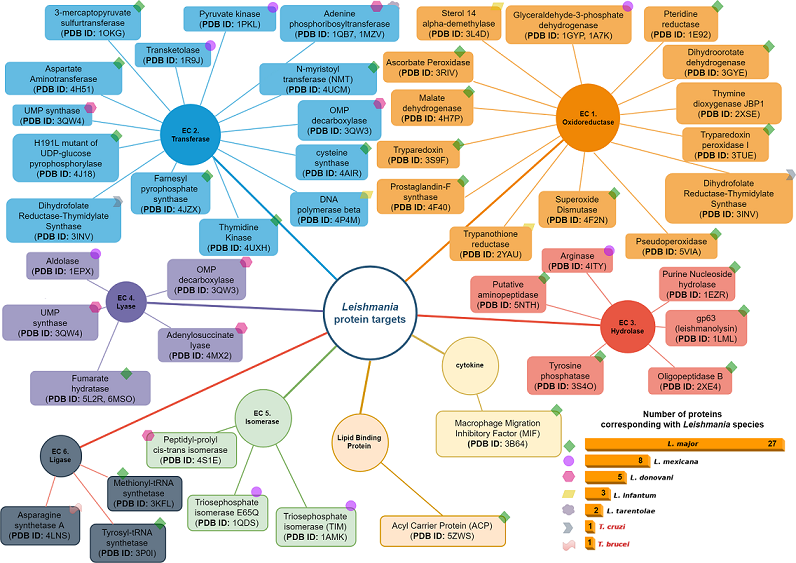
We conducted an exhaustive examination of databases and scholarly literature concerning Leishmania metabolic pathways. Utilizing the compiled data, we systematically analyzed, arranged, and interlinked the pathways and the corresponding involved proteins, as visualized in Figure 5. The proteins showcased in Figure 5 are integral constituents of crucial metabolic pathways, such as glycolysis, PPP, citric acid cycle, among others. They undertake pivotal functions in host infection and immune evasion mechanisms, thus underlining their indispensability for the survival of Leishmania.[67] Several proteins exhibit multifaceted roles across distinct metabolic pathways. For instance, Arginase engages in arginine biosynthesis, proline and arginine metabolism, as well as secondary metabolite and antibiotic biosynthesis pathways. [68]
However, it is noteworthy that proteins 1E92, 2XSE, 3VIA, 4F2N, 4F40, 4UCM, 2XE4, 3S4O, 5NTH, 4S1E, 3P0I, 3KFL, and 5ZWS were absent from the reviewed metabolic databases, thus preventing their assignment to specific pathways in Figure 5. Nevertheless, we conducted a thorough review and analysis of the functions of these proteins within this paper.
Furthermore, proteins 3B64, 1LML, and 4P4M are structural macromolecules with diverse roles within the Leishmania parasite. They do not align with any distinct metabolic pathway and are primarily considered for structural evaluations. It is vital to underscore the importance of these proteins in the context of Leishmania v, along with their potential implications for future research endeavors aimed at comprehending the parasite’s metabolism and pathogenicity. Figure 6 also displays correlations between proteins and pathways, concurrently highlighting protein structures and outlining the proteins not presented in Figure 5. In this illustration, proteins not linked to any specific pathway are denoted with the label "Not included in pathway". Additionally, their EC functions are differentiated by colors; thus, proteins featuring multiple colors denote having two or more EC functions.
The crystal structures presented in this study were obtained from the Protein Data Bank (PDB) [69]. A search was conducted in the PDB using the keywords "Leishmaniasis" and "Leishmania," resulting in a total of 00 items available until 2023. The selected proteins were then classified based on their relevance to the parasitization cycle. To ensure diversity in the protein selection, other criteria were considered. For instance, proteins with different PDB codes but identical structures were included, as well as proteins with the same structure but elucidated from different organisms. An example of such proteins is the Dihydrofolate Reductase-Thymidylate Synthase (DHFR-TS) (PDB ID: 3INV), which functions as both an oxidoreductase and a transferase.
Ultimately, the study group comprised a total of 49 proteins, as shown in Figure 5. Among these proteins, the largest groups were oxidoreductases (Enzyme Commission Number - EC 1) and transferases (EC 2), each containing 15 representatives. Additionally, there were 6 hydrolases (EC 3), 5 lyases (EC 4), 3 isomerases (EC 5), 3 ligases (EC 6), one cytokine, and one Lipid Binding Protein (Figure 6). These findings provide valuable insights into the diverse enzymatic functions and molecular characteristics of the selected proteins, shedding light on their potential roles in the biology of Leishmania parasites.
4.1. Oxidoreductases (EC 1)
The group of oxidoreductase enzymes identified in this study plays a crucial role in catalyzing oxidation-reduction reactions, where a substrate donates electrons to an electron-withdrawing species, resulting in the substrate being oxidized [70]. These enzymes are essential for the Glycolysis and Pentose Phosphate Pathways (PPP), which are key metabolic pathways in Trypanosomatids, including Leishmania parasites [71].
Glucose and other hexoses are vital nutrients for Leishmania parasites, and these parasites, whether in the promastigote or amastigote form, can acquire glucose from the extracellular environment or synthesize it through gluconeogenesis [72]. The promastigote parasites are capable of both glucose uptake and gluconeogenesis, while the amastigote parasites mainly rely on the gluconeogenic pathway for their glucose requirements [72]. These findings highlight the importance of oxidoreductase enzymes in the metabolic processes of Leishmania parasites, particularly in relation to glucose metabolism and energy production. Understanding the role of these enzymes and the specific pathways involved can provide valuable insights for developing targeted interventions against Leishmania infections.
Glyceraldehyde 3-phosphate dehydrogenase from Leishmania (major) mexicana, represented by PDB IDs 1GYP and 1A7K (see Figure 7), is an oxidoreductase enzyme that plays a role in both the Pentose Phosphate Pathway and glycolysis metabolic pathways [71]. This enzyme is a crucial component of glycolysis and is a homotetrameric protein with a molecular weight of 156 kDa. It catalyzes the conversion of glyceraldehyde 3-phosphate (G3P) to 1,3-biphosphoglycerate (1,3-BPG), simultaneously reducing NAD to NADH using the NAD cofactor [73,74]. Another important enzyme involved in the gluconeogenic process is malate dehydrogenase, represented by PDB ID 4H7P. This enzyme participates in the conversion of oxaloacetate (OAA) to malate and vice versa, utilizing the NAD/NADH coenzyme system [75,76]. It is worth noting that 4H7P has two isoforms in eukaryotes, which differ in their subcellular localization and their specificity for the coenzymes NAD (all types of malate dehydrogenases) or NADP (only malate dehydrogenases from chloroplast cells) [76]. These enzymes, glyceraldehyde 3-phosphate dehydrogenase and malate dehydrogenase, play vital roles in Leishmania’s metabolic pathways, contributing to the conversion of key substrates and the production of important intermediates in glycolysis and gluconeogenesis.
Dihydrofolate Reductase-Thymidylate Synthase (DHFR-TS), represented by PDB ID 3INV, is an enzyme found in Leishmania major. It combines the functions of two enzymes, dihydrofolate reductase (DHFR) and thymidylate synthase (TS), in a single polypeptide chain. DHFR-TS is involved in the salvage pathway of folate compounds in trypanosomatid parasites, including Leishmania. These parasites are unable to synthesize folate de novo and rely on the salvage pathway to obtain folate compounds for their metabolic processes. Folate compounds are crucial for various metabolic pathways in Leishmania, including nucleic acid and protein biosynthesis [77,78].
Pteridine reductase (PTR1), represented by PDB ID 1E92, is an enzyme found in Trypanosoma cruzi, another member of the Trypanosomatidae family. PTR1 is also involved in the salvage pathway of folate compounds. It plays a crucial role in the metabolism of folate and pteridine derivatives in trypanosomatids, including Leishmania. The salvage of folate compounds through PTR1 is essential for the normal metabolic processes of Leishmania parasites, as they are auxotrophic for folate and require these compounds for nucleic acid and protein biosynthesis [77,78].
Both DHFR-TS and PTR1 are important enzymes in the salvage pathways of folate compounds in Leishmania parasites. Their functions are critical for the synthesis of essential biomolecules and the survival of the parasites. Understanding the structure and function of these enzymes can provide valuable insights for the development of therapeutic strategies against Leishmania infections. The structure of the DHFR enzyme varies significantly between host organisms and parasites, leading to the development of specific DHFR inhibitors known as antifolates. These inhibitors have been utilized as therapeutic agents against parasites, including Leishmania infections [79].
Dihydroorotate dehydrogenase (DHODH), represented by PDB ID 3GYE, is a flavoprotein enzyme involved in the de novo biosynthesis pathway of pyrimidines. The pyrimidine biosynthesis pathway in Leishmania parasites is crucial for various cellular processes, such as DNA and RNA biosynthesis, protein glycosylation, membrane lipid biosynthesis, and DNA strand break repair [80]. DHODH enzymes are classified into two major classes: class 1 (A and B) and class 2, based on their subcellular localization. Class 1 proteins are found in the cytosol and are present in Gram-positive bacteria, anaerobic yeast Saccharomyces cerevisiae, and all trypanosomatid species, including Leishmania parasites [80,81]. The protein represented by PDB ID 3GYE belongs to the class 1 DHODH enzymes and catalyzes the oxidation of (S)-dihydroorotate to orotate in a redox reaction [81]. Understanding the structure and function of DHODH can provide insights into the development of drugs targeting this enzyme, which could be potential therapeutics for Leishmania infections.
Eukaryotic unicellular kinetoplastid flagellates, such as Trypanosoma and Leishmania species, possess a unique hypermodified base in their nuclear DNA known as J base or -D-glucosylhydroxymethyl-uracil [82,83,84]. The presence of the J base is associated with the silencing of telomeric expression sites for variant surface glycoprotein (VSG) genes, except in actively transcribed VSG genes [82,83]. The biosynthesis and maintenance of the J base in these parasites rely on the JBP1 protein, which is essential for the growth and survival of Leishmania parasites. JBP1 contains a DNA-binding JBP1 domain (DB-JBP1), represented by PDB ID 2XSE, that specifically binds to J-DNA. This interaction between DB-JBP1 and J-DNA makes JBP1 a potential target for drug development. Understanding the structure and function of DB-JBP1 and its role in J base biosynthesis and maintenance can provide valuable insights for the development of drugs aimed at disrupting this process and inhibiting the growth and survival of Leishmania parasites [85,86]. Targeting JBP1 could offer new therapeutic strategies for combating Leishmania infections.
Macrophages employ various defense mechanisms against Leishmania parasites, including the production of toxic compounds such as peroxynitrite, hydroxyl radicals, hydrogen peroxide, hydroperoxide, superoxide radicals, and others [87,88,89]. These compounds can disrupt the parasite’s metabolism and hinder its survival. However, Leishmania parasites have developed a trypanothione-mediated hydroperoxide metabolism to counteract the effects of oxidative agents, whether endogenous or exogenous [90]. Tryparedoxin (represented by PDB ID: 3S9F) and Tryparedoxin peroxidase I (represented by PDB ID: 3TUE) (TXN/TXNPx) are proteins that play a crucial role in reducing macrophage-generated hydroperoxides to water [91]. These proteins exist in a cytosolic form and are involved in the detoxification pathway, which is essential for the parasite’s survival [91]. Pseudoperoxidase L. major (LmPP) (PDB ID: 5VIA) is a heme protein expressed by Leishmania parasites as a defense mechanism against Reactive Nitrogen Species (RNS) [92,93]. Heme peroxidases, such as LmPP, utilize peroxides to oxidize various physiologically important molecules. Ascorbate peroxidase (APX) (PDB ID: 3RIV) is another example of a heme peroxidase and is considered a potential drug target [92].
Understanding the structure and function of these proteins involved in the detoxification pathways can provide insights for developing novel therapeutic approaches to disrupt the parasite’s defense mechanisms and enhance the efficacy of anti-Leishmania treatments [90,91,92,93]. Superoxide dismutase enzyme (FeSODA) (PDB ID: 4F2N) protects the Leishmania parasite against macrophage toxic radicals. When the amastigote is phagocytized, macrophage cells produce a respiratory burst generating Reactive Oxygen Species (ROS) intermediates, such as hydrogen peroxide, OH, O radicals, and peroxynitrite, as part of the macrophage mechanism to fight invasive microorganisms [94,95,96]. Thus, 4F2N acts as the first line of defense against those ROS. This fact makes 4F2N a suitable enzymatic target for Leishmania drug development [94].
Trypanothione reductase (TR) (PDB ID: 2YAU) is a homodimeric enzyme extracted from Leishmania infantum and is essential for parasite survival [97]. This enzyme catalyzes the reduction of trypanothione by NADPH, protecting the parasite against oxidative damage and toxic heavy metals [97,98,99]. TR reduces trypanothione disulfide (TS), a bis(-L-glutamyl-L-cysteinylglycine) spermidine or bis(glutathionyl) spermidine conjugate, to the di-thiol form [T(SH)] [98].
The enzyme 9,11-endoperoxide prostaglandin H2 reductase from L. major (PDB ID: 4F40) is involved in the lipid metabolic pathway, acting through an NADP cofactor [100,101]. Sterol 14-demethylase (CYP51) from L. infantum (PDB ID: 3L4D) is an enzyme that catalyzes the removal of the 14-methyl group from sterol precursors. This reaction is essential for membrane cell biosynthesis, specifically, CYP51 is involved in the ergosterol pathway and is believed to be decisive for the survival of Leishmania parasites [102]. Understanding the structures and functions of these enzymes involved in parasite defense mechanisms, oxidative stress response, and essential metabolic pathways provides valuable insights for the design of novel therapeutic strategies against Leishmania infections [94,95,96,97,100,102].
4.2. Transferases Group (EC. 2)
Transferases are enzymes that catalyze reactions involving the transfer of a chemical group from an electron/proton donor substrate to an electron/proton withdrawing substrate [70]. These proteins play crucial roles in various cellular processes across all kingdoms of life, including DNA repair, RNA editing, and activation of metabolites in catabolic pathways [104]. In the case of Leishmania parasites, the purine nucleotide salvage process is vital for their viability and growth, as these parasites are strict purine nucleotide auxotrophs [105,106]. To acquire purine nucleotides, Leishmania parasites employ a protein arsenal and molecular mechanisms [106,107]. Adenine phosphoribosyltransferase (APRT) (PDB ID: 1QB7 and 1MZV) belongs to the phosphoribosyltransferase family type I (PRTs) [108] and is involved in the purine salvage process. APRT catalyzes the conversion of adenine to adenosine-5-monophosphate (AMP) [106,107].
Protein fatty acylation is a common form of protein modification, with myristoylation being one of the most prevalent types [109]. Myristoylation plays roles in targeting proteins to membrane locations, stabilizing protein structures, mediating protein-protein interactions, and substrate activation [109,110]. Proteins destined for myristoylation typically start their primary sequence with a Methionine-Glycine (Met-Gly) dipeptide, where the initiator methionine residue is removed by methionine aminopeptidase, and a myristate molecule is linked via an amide bond [109].
N-myristoyltransferase (NMT) protein (PDB ID: 4UCM) is responsible for the co-translational transfer of myristic acid (myristate) from myristoyl-CoA to the N-terminal glycine of target proteins [110,111]. NMT plays a crucial role in Leishmania parasites and is considered a potential drug target [112]. Understanding the structures and functions of transferases, such as APRT and NMT, involved in purine salvage and protein modification processes, respectively, can provide insights for the development of targeted therapies against Leishmania infections.
The de novo pyrimidine biosynthesis pathway is a series of six enzymatic steps that lead to the synthesis of Uridine 5’-monophosphate (UMP). The final two steps of this pathway are mediated by the enzymes Orotate Phosphoribosyltransferase (OPRT) and Orotidine 5’-monophosphate decarboxylase (OMPDC) [113,114]. In Leishmania parasites, these two enzymes are expressed as a bifunctional protein known as UMP synthase [114]. Crystal structures of the bifunctional UMP synthase protein (PDB ID: 3QW4) and the OMP decarboxylase single protein (PDB ID: 3QW3) are available in the PDB database. Trypanothione plays a crucial role in maintaining intracellular redox homeostasis and providing defense against oxidative stress in Leishmania parasites [115,116,117]. The biosynthesis of trypanothione indirectly depends on the availability of cysteine, and the de novo biosynthesis pathway of cysteine relies on the enzymes Serine Acetyltransferase and Cysteine Synthase (PDB ID: 4AIR) [115,118].
Amastigote Leishmania parasites survive and proliferate within the phagolysosome vacuole, which exposes them to extreme acidic conditions and various toxic compounds [119]. These harsh host cell conditions cause high levels of DNA damage in the parasite [120]. Therefore, DNA Polymerase Beta (PDB ID: 4P4M) plays an essential role in Leishmania parasites for the maintenance, replication, and recombination of DNA [120]. This protein is particularly required in the amastigote form of the parasite [120]. Understanding the structures and functions of enzymes involved in pyrimidine biosynthesis, trypanothione metabolism, and DNA repair can provide insights into the development of targeted therapies against Leishmania infections.
In Leishmania parasites, both pyrimidine and purine nucleotides can be obtained through de novo biosynthesis and salvage pathways [121]. However, it is important to note that Leishmania species are incapable of synthesizing purines de novo [121].
Thymidine kinase (TK) (PDB ID: 4UXH) is an essential enzyme that initiates the pyrimidine salvage process in Leishmania parasites [122]. This enzyme catalyzes the transfer of the -phosphate group from ATP to 2’-deoxythymidine (dThd), forming thymidine monophosphate (dTMP). TK plays a key role in the parasitization process and is an important target for drug development [123]. Farnesyl pyrophosphate synthase (FPPS) (PDB ID: 4JZX) is involved in ergosterol synthesis and is responsible for the early steps of isoprene synthesis, as well as the maintenance of lipid bilayer integrity in Leishmania parasites [124,125]. FPPS is considered a potential enzymatic target, as it has been successfully inhibited with bisphosphonates [124].
UDP-glucose pyrophosphorylase L. major (UGP) (PDB ID: 4J18) is an enzyme involved in the glycolytic pathway of Leishmania parasites [104]. It catalyzes the reversible conversion of glucose-1-phosphate (Glc-1-P) and uridine 5’-triphosphate (UTP) to UDP-glucose (UDP-Glc) and inorganic pyrophosphate (PPi) in the presence of Mg. UGP plays a role in carbohydrate metabolism and is important for the synthesis of essential glycoconjugates in the parasite [104]. Understanding the pathways involved in nucleotide biosynthesis and the enzymes associated with these processes can provide valuable insights for the development of targeted therapies against Leishmania infections.
Aspartate aminotransferase (AAT) (PDB ID: 4H51) is an enzyme that catalyzes the reversible transfer of the -amino group between aspartate and glutamate [126]. Specifically, it converts L-aspartate and 2-oxoglutarate to oxaloacetate and L-glutamate. This enzymatic reaction is essential for amino acid metabolism and the synthesis of key molecules in the cell. AAT relies on the pyridoxal phosphate (PLP) cofactor, which plays a crucial role in the catalytic mechanism of the enzyme [126]. PLP acts as a coenzyme, facilitating the transfer of the amino group between the amino acid substrates. The dependence of AAT on PLP makes it an attractive target for drug development, as inhibiting its activity can disrupt amino acid metabolism and lead to detrimental effects on the parasite. Targeting AAT with specific inhibitors can interfere with the normal functioning of Leishmania parasites, disrupting their ability to synthesize essential molecules and causing metabolic imbalances. Consequently, AAT represents a potential drug target for the development of novel therapies against Leishmania infections [127].
Sulfurtransferases are a diverse family of enzymes that play important roles in sulfur metabolism in both prokaryotes and eukaryotes. One member of this family is 3-mercaptopyruvate sulfurtransferase (PDB ID: 1OKG), which is involved in cysteine metabolism. This enzyme catalyzes the transfer of a sulfur atom from 3-mercaptopyruvate to various sulfur acceptor molecules, polarizing the carboxyl group of 3-mercaptopyruvate through a nucleophilic attack [128,129]. Transketolase (PDB ID: 1R9J) is another important enzyme involved in carbohydrate metabolism. It plays a key role in the nonoxidative branch of the pentose phosphate pathway (PP pathway). Transketolase transfers two-carbon glycolaldehyde units from ketose donors to aldose acceptor sugars, facilitating the interconversion of different sugar phosphates [130]. Pyruvate kinase (PDB ID: 1PKL) is an enzyme involved in the glycolytic pathway, specifically in the final step of glycolysis. It catalyzes the transfer of a phosphate group from phosphoenolpyruvate to adenosine diphosphate (ADP), generating pyruvate and ATP. This reaction is crucial for energy production in cells [131].
These enzymes, sulfurtransferase, transketolase, and pyruvate kinase, have important roles in various metabolic pathways and are essential for the proper functioning of cells. Understanding their structures and functions provides insights into their mechanisms of action and their potential as targets for drug development and therapeutic interventions.
4.3. Hydrolases Group (EC. 3)
Hydrolases are a class of enzymes that catalyze hydrolysis reactions, where a bond in a substrate molecule is cleaved by the addition of a water molecule, resulting in the formation of two product fragments [70]. In the case of the Leishmania parasite, which cannot synthesize purines de novo, the Purine Nucleoside Hydrolase of L. major (PDB ID: 1EZR) plays a crucial role in salvaging nucleotides from the host [132]. This enzyme is responsible for hydrolyzing purine nucleosides to release the purine base and ribose or deoxyribose [132]. Due to its essential role in nucleotide metabolism, Purine Nucleoside Hydrolase is considered a potential drug target for Leishmania [132].
Leishmanolysin (gp63 gene) is a glycoprotein expressed on the surface of Leishmania promastigotes, and its structure is represented by PDB ID: 1LML [133]. This enzyme plays a critical role in the infection of macrophages by Leishmania parasites, mediating the degradation of host extracellular matrix proteins and evading the immune response [133]. Consequently, Leishmanolysin is considered a potential drug target for combating Leishmania infections [133]. Peptidases, a family of enzymes, have diverse functions in metabolic pathways, host invasion, and immune evasion by parasites [134]. Oligopeptidase B (OPB) of L. major (PDB ID: 2XE4) is a peptidase enzyme known to be an important virulence factor [135,136]. It plays a role in the degradation of host proteins and the processing of parasite virulence factors [135,136].
Leucyl aminopeptidase (LAP) of L. major (PDB ID: 5NTH) is another peptidase enzyme involved in the catalysis of N-terminal residues of proteins [137]. It participates in the hydrolysis of peptide bonds, contributing to the processing and turnover of proteins in Leishmania [137]. These hydrolase enzymes, including Purine Nucleoside Hydrolase, Leishmanolysin, Oligopeptidase B, and Leucyl aminopeptidase, have significant roles in the metabolism, virulence, and survival of Leishmania parasites. Understanding their structures and functions can aid in the development of drugs and interventions targeted against Leishmania infections.
Tyrosine phosphatase PRL-1 (PDB ID: 3S4O) is primarily secreted by the promastigote form of Leishmania parasites. However, during the macrophage infection process, the amastigote form produces PRL-1 more efficiently and abundantly, which is crucial for the parasite’s survival [138]. This enzyme is involved in the dephosphorylation of tyrosine residues in protein substrates, regulating various signaling pathways and cellular processes [138]. The increased production of PRL-1 by amastigotes suggests its importance in the adaptation and virulence of Leishmania during the intracellular infection of macrophages [138].
Arginase (PDB ID: 4ITY) is an enzyme that catalyzes the hydrolysis of arginine to urea and ornithine. It plays a critical role in the first step of polyamine biosynthesis, which is essential for cellular growth and proliferation [139]. Polyamines are involved in various biological processes, including DNA synthesis, cell division, and differentiation [139]. In Leishmania, arginase is crucial for the production of polyamines, which are required for the parasite’s survival and virulence [139]. Both tyrosine phosphatase PRL-1 and arginase are important enzymes for the survival and adaptation of Leishmania parasites. They contribute to key cellular processes and are potential targets for the development of interventions against Leishmania infections.
4.4. Lyase Group (EC. 4)
Adenylosuccinate lyase (ASL) (PDB ID: 4MX2) is a lyase enzyme that plays a vital role in purine salvage in Leishmania donovani [106]. It catalyzes the cleavage of adenylosuccinate into fumarate and AMP, participating in the recycling of purine nucleotides in the parasite [106]. Fumarate hydrolase (FH) is a lyase enzyme with two isoforms: a mitochondrial form (PDB ID: 6MSO) and a cytosolic form (PDB ID: 5L2R) [140]. The cytosolic isoform (5L2R) produces fumarate, which serves as a substrate for dihydroorotate dehydrogenase and participates in the de novo pyrimidine biosynthetic pathway [140]. Additionally, FH migrates to the cellular nucleus and plays a role in DNA repair processes [140].
4.5. Isomerases Group (EC. 5)
Isomerase enzymes catalyze the conversion of one substrate to its isomeric form, and Triosephosphate isomerase (TIM) is a notable example of such an enzyme. TIM (PDB ID: 1AMK) is primarily involved in the glycolysis process and is responsible for catalyzing the reversible isomerization between dihydroxyacetone phosphate (DHAP) and D-glyceraldehyde-3-phosphate (GAP) [142]. This enzymatic interconversion between DHAP and GAP is a crucial step in glycolysis, as it facilitates the transformation of DHAP, a ketose, into GAP, an aldose. The resulting GAP can then proceed through subsequent metabolic reactions to produce ATP and other essential intermediates. Studies have also focused on a mutant form of TIM known as TIM E65Q (PDB ID: 1QDS), which has shown increased stability compared to the wild-type TIM [143]. This mutant variant exhibits altered kinetic properties and enhanced resistance to unfolding, providing valuable insights into the structural and functional characteristics of TIM and its stability.
Peptidyl-prolyl cis-trans isomerase (PPIase) is an isomerase enzyme involved in protein folding processes [144]. One example is the PDB entry 4S1E, which catalyzes the cis-trans isomerization of proline imidic peptide bonds in oligopeptides [144]. PPIases play a crucial role in accelerating protein folding, aiding in the correct formation of peptide bonds and facilitating the proper conformation of proteins.
4.6. Ligases Group (EC. 6)
Ligase enzymes catalyze the formation of bonds between macromolecules through a process often associated with the hydrolysis of a small chemical molecule coupled to the macromolecules [70]. In this review, three proteins belonging to the ligase group have been identified as potential enzymatic targets: Methionyl-tRNA synthetase (PDB ID: 3KFL) and Tyrosyl-tRNA synthetase (PDB ID: 3P0I), whose structural parameters have been elucidated with MgATP as a substrate and methionine as a solvent, and are recognized as essential for biological processes such as gene translation [145,146]. The third enzyme is Asparagine synthetase A (ASNA) (PDB ID: 4LNS), an ammonium and glutamine-dependent enzyme. Experimental studies (in vivo and in vitro) have shown that the 4LNS protein causes growth delay in the parasite and has been identified as a potential drug target for the development of bioactive compounds against Leishmania [103].
4.7. Cytokines Group
The Migration Inhibitory Factor (MIF) from L. major (PDB ID: 3B64) has been identified as a potential target for drug development. This cytokine, known as Lm1740MIF, shares orthology with human MIF. 3B64 interacts with MIF receptors, such as HLA class II histocompatibility antigen gamma chain (also known as invariant chain or CD74), and exhibits antiapoptotic activity. This activity may facilitate the intracellular persistence of Leishmania within macrophages.[147]
4.8. Lipid Binding Protein Group
The Apo-Acyl Carrier Protein (PDB ID: 5ZWS) plays a crucial role in the synthesis of fatty acids, non-ribosomal polypeptides, and polyketides. The fatty acid pathway, including its more complex forms, has recently gained attention in research on Leishmania due to its involvement in the survival of protozoan parasites inside the host [148].
5. Leishmaniasis Drugs
Pentavalent antimonials (Sb(V)) were the initial bioactive chemical entities devised for managing Leishmania infections. Nonetheless, the emergence of Leishmania resistance has considerably diminished the effectiveness of (Sb(V)).[19] Consequently, there has been a growing emphasis on the exploration of novel compounds with antileishmanial properties. Presently, standard treatment drugs like Amphotericin B, Pentamidine, Miltefosine, Paromomycin, and Sitamaquine face challenges of inefficacy and elevated costs, which have contributed to the development of drug resistance in Leishmania parasites.[14,19,41]
5.1. Standard Drugs
Antimonials, including meglumine antimoniate (Glucantime) and sodium stibogluconate (Pentostam), were the first antileishmanial compounds introduced in the 1940s [19,40]. These drugs have been the standard first-line treatment for leishmaniasis. However, the emergence of resistance has significantly limited their effectiveness [41,149]. Studies have shown that L. donovani and L. braziliensis are more sensitive to sodium stibogluconate compared to L. major, Leishmania tropica, and L. mexicana [19,39].
Amphotericin B, a macrolide antibiotic isolated from Streptomyces nodosus in 1956, has been widely used since the 1980s, particularly as amphotericin B deoxycholate [19,40]. It selectively inhibits membrane synthesis in the parasite and creates pores in the membrane, leading to parasite death [40]. Amphotericin B is commonly used as a second-line treatment and is available in various formulations, including amphotericin B deoxycholate (Fungizone), amphotericin B in intralipid 20% (Amphotericin B Intralipid), liposomal amphotericin B (AmBisome), amphotericin B lipid complex (ABLC; Abelcet), and amphotericin B cholesterol dispersion (ABCD; Amphotec) [39,40]. Lipid formulations of amphotericin B, in particular, have gained significant importance and have been established as the preferred treatment for leishmaniasis by the US Food and Drug Administration (FDA) [19].
Pentamidine is an antileishmanial drug that targets the parasite’s polyamine biosynthesis and mitochondrial membrane. It is considered a second-line treatment for visceral leishmaniasis (VL) due to its toxicity, which includes adverse effects such as myalgia, nausea, headache, hypoglycemia, irreversible insulin-dependent diabetes mellitus, and even death. Additionally, its high cost makes it economically prohibitive [40].
In India and East Africa, paromomycin has been used as a cheaper alternative treatment option for VL, despite its toxicity [19]. Initially, paromomycin was neglected, but in the 1980s, topical formulations for VL were developed, renewing interest in its therapeutic potential [40]. To address the issue of accessibility and affordability, organizations such as One World Health, the Bill and Melinda Gates Foundation, Gland Pharma Limited, IDA Solutions, and WHO/TDR have partnered to develop paromomycin as a public health tool to be sold on a not-for-profit basis at a significantly reduced price [40].
Miltefosine, originally developed as an anticancer drug, has emerged as the first effective oral treatment for visceral leishmaniasis (VL) [19,40]. Its efficacy has been investigated against various species of Leishmania, including L. donovani, L. major, L. tropica, Leishmania aethiopica, L. mexicana, and L. panamensis, in both the promastigote and amastigote stages. In vitro studies have shown varying sensitivities, with L. donovani being recognized as the most sensitive species to miltefosine [39]. The effective concentration (ED) of miltefosine against L. donovani ranges from 0.12 to 1.32 M [39].
Sitamaquine is another drug used for leishmaniasis treatment. It undergoes rapid metabolism, forming desethyl and 4-CHOH derivatives, which may contribute to its activity. Sitamaquine is relatively well-tolerated, with mild methemoglobinemia reported as a potential side effect [40,150]. Pamidronate, a bisphosphonate drug, has shown significant activity against Leishmania donovani in vitro. It targets the FPPS protein (PDB ID: 4JZX) and potently inhibits it in the trypanosomatid parasite [40]. Azoles and nucleoside analogues are two other types of drugs considered for the treatment of leishmaniasis [39,151]. Within the azole group, drugs such as ketoconazole and itraconazole are commonly used. They exert their antileishmanial activity by inhibiting the enzyme C14-demethylase, which is involved in the synthesis of ergosterol, an essential component of the Leishmania cell membrane [39].
Nucleoside analogues, such as allopurinol and pyrazolopyrimidine, exert their antileishmanial effects by disrupting the enzymatic processes involved in the purine salvaging pathway of Leishmania [39]. This pathway is essential for the parasite’s ability to synthesize and utilize purine nucleotides, which are crucial for its growth and survival. By inhibiting key enzymes in this pathway, nucleoside analogues interfere with the parasite’s ability to obtain the necessary building blocks for nucleic acid synthesis and energy metabolism, ultimately leading to its demise.
5.2. Metabolomics: Non Standard Drugs
We have curated a roster of databases housing bioactive compounds that have undergone investigation for their potential in Leishmaniasis treatment, presenting alternative options to commercial pharmaceuticals (refer to Figure 9; see supplementary material). These databases encompass diverse facets, encompassing metabolome analysis, chemical tools, chemical structure data, natural products, and pertinent literature in the medical domain. Additionally, we have conducted an extensive evaluation of research articles that have compiled over 200 chemical compounds and numerous plant extracts (see supplementary material). These investigations encompass an extensive array of experimental methodologies, spanning both in vivo and in vitro approaches, and encompassing distinct Leishmania species, while considering various physical and chemical conditions.[152,153,154]
On the other hand, antimicrobial peptides (AMPs) have emerged as potential treatments for Leishmaniasis, particularly in in vitro assays. AMPs are a diverse group of compounds that are naturally produced by various organisms, including humans, and exhibit antimicrobial activity against viruses, bacteria, fungi, and parasites. In animals, AMPs are primarily found in tissues and organs and serve as a crucial component of the innate immune system’s first line of defense [155,156]. Their broad-spectrum antimicrobial properties make them attractive candidates for developing novel therapeutic strategies against Leishmania parasites.
Is important to highlight that Leishmania parasites can develop resistance against these secondary metabolites and AMPs too, therefore, the use of monotherapies is not an good option. The rationale behind combined therapies is to prevent the risk of parasite resistance, to provide increased efficacy resulting from synergistic effects, minimizing dose requirements and to reduce therapy time, eventually leading to a reduction of side toxic effects, or al least to mild toxicity and reduce mortality.[14]
5.2.1. From Venom to Medicine: Phospholipases A2 (PLA2) as Potential Antiparasitic Agents against Leishmania Parasites
Phospholipase (PL) enzymes constitute a diverse protein group that hydrolyze ester bonds within phospholipid structures. Phospholipids are located in cellular membranes and play important roles in biochemical signaling and lipid metabolism processes. PLs are classified into several families, with PLA, PLB, PLC, and PLD being the most relevant. Specifically, the PLA2 group hydrolyzes the sn-2 position of phospholipids such as phosphatidylcholines and phosphatidylethanolamines, releasing free fatty acids. PLA2 enzymes have been characterized across a wide range of organisms and play conserved roles in lipid digestion and cell signaling cascades through arachidonic acid release. Further research into the structural biology and biochemical functions of PLA2 enzymes may provide insights into their potential as therapeutic targets or tools. [157,158]
Moreover, the secretory PLA2 (sPLA2) isoforms are the most studied, playing a crucial role in cellular signaling due to their capability to generate free arachidonic acid and important eicosanoid precursors. These eicosanoids regulate inflammatory responses. However, PLs are not only essential for cellular signaling and lipid homeostasis. Some species utilize PLs for alternative functions. For instance, bees and wasps use PLs as venom components to trigger inflammatory and allergic responses in humans. Additionally, certain fungal PLAs facilitate pathogen invasion and damage of host cells during infection. Likewise, in plants, PLA2 contributes to pathogen defense and stress response, enabling plants to adapt and survive adverse conditions. [158,159,160,161,162,163,164]
It’s essential to recognize that while secondary metabolites like venoms can pose risks, they can also serve beneficial roles, such as safeguarding against pathogens, fostering symbiotic interactions, or facilitating chemical communication. Nevertheless, the primary function of venom production is often linked to defense and the capture of prey. Within this context, PLA2 enzymes play a pivotal role in snake venoms, aiding in the digestion of prey and eliciting toxic effects. The presence of PLA2 in snake and other animal venoms has ignited extensive research efforts aimed at comprehending their mechanisms of action and pharmacological attributes for potential drug development.
Due to their antiparasitic properties against various parasites, PLA2 stands as a promising reservoir of antimicrobial peptides (AMPs). For instance, PLA2 enzymes have exhibited antiparasitic efficacy against malaria, Chagas disease, and toxoplasmosis parasites. Specifically, they can impede parasite growth, disrupt cell membranes, and curtail invasiveness. These instances underscore the range of susceptible parasites to PLA2 and their capacity to exert antimicrobial effects through multiple avenues. Further exploration of PLA2 as a source of AMPs has the potential to unveil innovative therapeutic strategies against parasitic diseases.[159,160,161,165,166,167]
Research within this domain remains dynamic and centers on gaining a more comprehensive grasp of the mechanisms of action inherent to these enzymes, as well as their viability as targets for therapeutic intervention. Progress in this arena holds the potential to introduce novel approaches for addressing parasitic illnesses and enhancing worldwide health standards. The continuous endeavors in research and the formulation of therapies founded on PLA2 instill optimism for effectively countering these parasites, ultimately paving the way for more efficient and secure therapeutic alternatives in the times ahead.
6. Conclusions
The utilization of conventional pharmaceuticals in the management of leishmaniasis may entail certain limitations. These medications have the potential to give rise to unfavorable effects, resistance, and toxicity. Furthermore, their application demands consistent clinical and paraclinical monitoring, and they can incur substantial costs while also exhibiting ineffectiveness in specific instances. As a result, it becomes imperative to undertake research endeavors aimed at discerning the parasite’s metabolic pathways. Such inquiries play a pivotal role in identifying susceptible targets within the parasite, encompassing proteins engaged in immune responses and defense mechanisms. Through the comprehension of these precise elements, we can formulate more effective therapeutic approaches that ultimately bolster patient outcomes.
Author Contributions
”Conceptualization: JGS and AFC. Methodology: AFC and CAP. Validation: AFC. Data curation: CAP, EC, MJA and AFC; writing - original draft preparation: CAP, EC and AFC. Writing - review and editing: CAP, EM and AFC. Supervision: AFC. All authors have read and agreed to the published version of the manuscript.”
Funding
“This research was funded by Vicerrectoría de Investigaciones of Universidad de Sucre, Sincelejo-Sucre, Colombia; Ministerio de Ciencias - BPIN 2020000100536, Bogotá, Colombia; Instituto de Investigación Astronómico y Aeroespacial Pedro Paulet (IAAPP), Universidad Nacional de San Agustin (UNSA), Arequipa, Perú”.
Acknowledgments
Authors acknowledge Universidad Nacional de San Agustín (UNSA, Arequipa, Perú) and Instituto de Investigación Astronómico y Aeroespacial Pedro Paulet (IAAPP) for providing access to INKARI (High Performance Computer Facilities, located at the Astronomic Observatory of CHARACATO - UNSA, Arequipa, Perú).
Conflicts of Interest
“The authors declare no conflict of interest.”
References
- Ornellas-Garcia, U.; Cuervo, P.; Ribeiro-Gomes, F.L. Malaria and leishmaniasis: Updates on co-infection. Frontiers in Immunology 2023, 14, 1122411. [Google Scholar] [CrossRef] [PubMed]
- Awab, G.R. Leishmaniasis Epidemiology and Psychosocial Aspect; IntechOpen, 2023. [Google Scholar] [CrossRef]
- Grifferty, G.; Shirley, H.; McGloin, J.; Kahn, J.; Orriols, A.; Wamai, R. Vulnerabilities to and the Socioeconomic and Psychosocial Impacts of the Leishmaniases: A Review. Research and Reports in Tropical Medicine 2021, Volume 12, 135–151. [Google Scholar] [CrossRef]
- Maheshwari, K.K.; Bandyopadhyay, D. Heterocycles in the Treatment of Neglected Tropical Diseases. Current Medicinal Chemistry 2021, 28, 472–495. [Google Scholar] [CrossRef] [PubMed]
- Morales-Yuste, M.; Martín-Sánchez, J.; Corpas-Lopez, V. Canine Leishmaniasis: Update on Epidemiology, Diagnosis, Treatment, and Prevention. Veterinary Sciences 2023, 9, 387. [Google Scholar] [CrossRef]
- Gao, C.h.; Wang, J.y.; Zhang, S.; Yang, Y.t.; Wang, Y. Survey of Wild and Domestic Mammals for Infection with Leishmania infantum following an Outbreak of Desert Zoonotic Visceral Leishmaniasis in Jiashi, People’s Republic of China. PLOS ONE 2023, 10, e0132493. [Google Scholar] [CrossRef] [PubMed]
- Alcover, M.M.; Riera, M.C.; Fisa, R. Leishmaniosis in Rodents Caused by Leishmania infantum: A Review of Studies in the Mediterranean Area. Frontiers in Veterinary Science 2021, 8, 702687. [Google Scholar] [CrossRef] [PubMed]
- Rezaei, Z.; Pourabbas, B.; Asaei, S.; Sepehrpour, S.; Ahmadnia Motlagh, S.; Pourabbas, P.; Abdolahi Khasibi, S.; Alborzi, A. Livestock infected with Leishmania spp. in southern Iran. Parasites & Vectors 2023, 15, 215. [Google Scholar] [CrossRef]
- Ratzlaff, F.R.; Osmari, V.; da Silva, D.; de Paula Vasconcellos, J.S.; Pötter, L.; Fernandes, F.D.; de Mello Filho, J.A.; de Avila Botton, S.; Vogel, F.S.F.; Sangioni, L.A. Identification of infection by Leishmania spp. in wild and domestic animals in Brazil: a systematic review with meta-analysis (2001–2021). Parasitology Research 2023, 122, 1605–1619. [Google Scholar] [CrossRef]
- J, B.; M, B.M.; Chanda, K. An Overview on the Therapeutics of Neglected Infectious Diseases—Leishmaniasis and Chagas Diseases. Frontiers in Chemistry 2021, 9. [Google Scholar] [CrossRef]
- Pandian, S.R.K.; Panneerselvam, T.; Pavadai, P.; Govindaraj, S.; Ravishankar, V.; Palanisamy, P.; Sampath, M.; Sankaranarayanan, M.; Kunjiappan, S. Nano Based Approach for the Treatment of Neglected Tropical Diseases. Frontiers in Nanotechnology 2021, 3. [Google Scholar] [CrossRef]
- J, B.; M, B.M.; Chanda, K. An Overview on the Therapeutics of Neglected Infectious Diseases—Leishmaniasis and Chagas Diseases. Frontiers in Chemistry, 9.
- Choi, H.L.; Jain, S.; Postigo, J.A.R.; Borisch, B.; Dagne, D.A. The global procurement landscape of leishmaniasis medicines. PLOS Neglected Tropical Diseases, P: Publisher; 15, e0009181. [CrossRef]
- Zulfiqar, B.; Shelper, T.B.; Avery, V.M. Leishmaniasis drug discovery: recent progress and challenges in assay development. Drug Discovery Today 2017, 22, 1516–1531. [Google Scholar] [CrossRef] [PubMed]
- Reimão, J.Q.; Coser, E.M.; Lee, M.R.; Coelho, A.C. Laboratory Diagnosis of Cutaneous and Visceral Leishmaniasis: Current and Future Methods. Microorganisms, 8, 1632. [CrossRef]
- OPS/OMS, Organización Panamericana de la Salud. Leishmaniasis: Informe Epidemiológico de las Américas. Technical report, OPS, Organización Panamericana de la Salud, 2021.
- Cupolillo, E.; Medina-Acosta, E.; Noyes, H.; Momen, H.; Grimaldi, G. A revised classification for Leishmania and Endotrypanum. Parasitology Today 2000, 16, 142–144. [Google Scholar] [CrossRef] [PubMed]
- Momen, H.; Cupolillo, E. Speculations on the Origin and Evolution of the Genus Leishmania. Revista da Sociedade Brasileira de Medicina Tropical 2000, 95, 583–588. [Google Scholar] [CrossRef]
- Bruschi, F.; Gradoni, L. (Eds.) The leishmaniases: Old neglected tropical diseases; Springer International Publishing: Cham, 2018; pp. 1–245. [Google Scholar] [CrossRef]
- Kostygov, A.Y.; Yurchenko, V. Revised classification of the subfamily Leishmaniinae (Trypanosomatidae). Folia Parasitologica 2017, 64, 1–5. [Google Scholar] [CrossRef] [PubMed]
- (a)Lainson, R.; Shaw, J. Evolution, classification and geographical distribution. In The Leishmaniases in Biology and Epidemiology; Peters, W.; Killick-Kendrick, R., Eds.; Academic Press, 1987; Vol. 1, chapter 1, pp. 1 – 120.
- Lainson, R.; Shaw, J. The Role of Animals in the Epidemiology of South American Leishmaniasis. In Biology of the Kinetoplastida, 2 ed.; In WHR Lumsden, D.E., Ed.; Academic Press: London and New York, 1979; p. 116. [Google Scholar]
- Corrêa, J.R.; Brazil, R.P.; Soares, M.J. Leishmania (Viannia) lainsoni (Kinetoplastida: Trypanosomatidae), a divergent Leishmania of the Viannia subgenus - A mini review. Memorias do Instituto Oswaldo Cruz 2005, 100, 587–592. [Google Scholar] [CrossRef] [PubMed]
- Franco, A.M.; Grimaldi, G. Characterization of Endotrypanum (Kinetoplastida: Trypanosomatidae), a Unique Parasite Infecting the Neotropical Tree Sloths (Edentata). Memorias do Instituto Oswaldo Cruz 1999, 94, 261–268. [Google Scholar] [CrossRef]
- Akhoundi, M.; Kuhls, K.; Cannet, A.; Votýpka, J.; Marty, P.; Delaunay, P.; Sereno, D. A Historical Overview of the Classification, Evolution, and Dispersion of Leishmania Parasites and Sandflies. PLoS Neglected Tropical Diseases 2016, 10, 1–40. [Google Scholar] [CrossRef]
- Mesnil F, B.E. Sur un hematozoaire nouveau (Endotrypanum n.gen.) d’un edente de la Guyane. C R Soc Biol 1908, 65, 587. [Google Scholar]
- JJ, S. The Haemoflagellates of Sloths H.K. Lewis and Co. Ltd., London, In London School of Hygiene and Tropical Medicine Memoir, 13 ed.; H. K. Lewis (1969): London, 1969; p. 132. [Google Scholar]
- WHO/PAHO. Leishmaniases: Epidemiological Report in the Americas. Technical report, WHO and PAHO, 2018.
- Universidad del Rosario. Colombia, el país con más especies de parásitos de Leishmania, 2017.
- Kmetiuk, L.B.; Tirado, T.C.; Biondo, L.M.; Biondo, A.W.; Figueiredo, F.B. Leishmania spp. in indigenous populations: A mini-review. Frontiers in Public Health, 10.
- Herrera, G.; Barragán, N.; Luna, N.; Martínez, D.; De Martino, F.; Medina, J.; Niño, S.; Páez, L.; Ramírez, A.; Vega, L.; Velandia, V.; Vera, M.; Zúñiga, M.F.; Bottin, M.J.; Ramírez, J.D. An interactive database of Leishmania species distribution in the Americas. Scientific Data, 7, 110. [CrossRef]
- Frézard, F.; Aguiar, M.M.G.; Ferreira, L.A.M.; Ramos, G.S.; Santos, T.T.; Borges, G.S.M.; Vallejos, V.M.R.; De Morais, H.L.O. Liposomal Amphotericin B for Treatment of Leishmaniasis: From the Identification of Critical Physicochemical Attributes to the Design of Effective Topical and Oral Formulations. Pharmaceutics, 15, 99. [CrossRef]
- Musa, A.M.; Mbui, J.; Mohammed, R.; Olobo, J.; Ritmeijer, K.; Alcoba, G.; Muthoni Ouattara, G.; Egondi, T.; Nakanwagi, P.; Omollo, T.; Wasunna, M.; Verrest, L.; Dorlo, T.P.C.; Musa Younis, B.; Nour, A.; Taha Ahmed Elmukashfi, E.; Ismail Omer Haroun, A.; Khalil, E.A.G.; Njenga, S.; Fikre, H.; Mekonnen, T.; Mersha, D.; Sisay, K.; Sagaki, P.; Alvar, J.; Solomos, A.; Alves, F. Paromomycin and Miltefosine Combination as an Alternative to Treat Patients With Visceral Leishmaniasis in Eastern Africa: A Randomized, Controlled, Multicountry Trial. Clinical Infectious Diseases: An Official Publication of the Infectious Diseases Society of America, 76, e1177–e1185. [CrossRef]
- Román-Álamo, L.; Allaw, M.; Avalos-Padilla, Y.; Manca, M.L.; Manconi, M.; Fulgheri, F.; Fernández-Lajo, J.; Rivas, L.; Vázquez, J.A.; Peris, J.E.; Roca-Geronès, X.; Poonlaphdecha, S.; Alcover, M.M.; Fisa, R.; Riera, C.; Fernàndez-Busquets, X. In Vitro Evaluation of Aerosol Therapy with Pentamidine-Loaded Liposomes Coated with Chondroitin Sulfate or Heparin for the Treatment of Leishmaniasis. Pharmaceutics, 15, 1163. [CrossRef]
- Rojas Cabrera, E.; Verduguez-Orellana, A.; Tordoya-Titichoca, I.J.; Sejas, C.; Ledezma, R.; Álvarez, I.; Limachi-Choque, J.; Ortuño-Gutiérrez, N.; Córdova Rojas, M.; Guzman-Rivero, M. Intralesional Meglumine Antimoniate: Safe, Feasible and Effective Therapy for Cutaneous Leishmaniasis in Bolivia. Tropical Medicine and Infectious Disease, 7, 286. [CrossRef]
- Yadagiri, G.; Singh, A.; Arora, K.; Mudavath, S.L. Immunotherapy and immunochemotherapy in combating visceral leishmaniasis. Frontiers in Medicine, 10.
- Feng, M.; Jin, Y.; Yang, S.; Joachim, A.M.; Ning, Y.; Mori-Quiroz, L.M.; Fromm, J.; Perera, C.; Zhang, K.; Werbovetz, K.A.; Wang, M.Z. Sterol profiling of Leishmania parasites using a new HPLC-tandem mass spectrometry-based method and antifungal azoles as chemical probes reveals a key intermediate sterol that supports a branched ergosterol biosynthetic pathway. International Journal for Parasitology. Drugs and Drug Resistance, 20, 27–42. [CrossRef]
- Hofer, A. Targeting the nucleotide metabolism of Trypanosoma brucei and other trypanosomatids. FEMS Microbiology Reviews, 47, fuad020. [CrossRef]
- Croft, S.L.; Sundar, S.; Fairlamb, A.H. Drug Resistance in Leishmaniasis. Society 2006, 19, 111–126. [Google Scholar] [CrossRef]
- Kumar, A. Leishmania and Leishmaniasis; Vol. 3, SpringerBriefs in Immunology, Springer, 2013. [CrossRef]
- Ghorbani, M.; Farhoudi, R. Leishmaniasis in humans: Drug or vaccine therapy? Drug Design, Development and Therapy 2018, 12, 25–40. [Google Scholar] [CrossRef]
- Tsirigotakis, N.; Pavlou, C.; Christodoulou, V.; Dokianakis, E.; Kourouniotis, C.; Alten, B.; Antoniou, M. Phlebotomine sand flies (Diptera: Psychodidae) in the Greek Aegean Islands: ecological approaches. Parasites & Vectors 2018, 11, 97. [Google Scholar] [CrossRef]
- Bates, P.A. Transmission of Leishmania metacyclic promastigotes by phlebotomine sand flies. International Journal for Parasitology 2007, 37, 1097–1106. [Google Scholar] [CrossRef] [PubMed]
- ESTRADA, L.G.; APONTE, O.A.; BEJARANO, E.E. REGISTROS NUEVOS DE ESPECIES DE Lutzomyia (DIPTERA: PSYCHODIDAE) EN EL DEPARTAMENTO DE CESAR, COLOMBIA. Acta Biológica Colombiana 2015, 20, 225–228. [Google Scholar] [CrossRef]
- Yuko, E.; Sang, R.; Owino, E.A.; Ingonga, J.; Matoke-Muhia, D.; Hassaballa, I.B.; Junglen, S.; Tchouassi, D.P. Sandfly Blood-Feeding Habits and Competence in Transmitting Ntepes Virus, a Recently Discovered Member of the Genus Phlebovirus. BioMed Research International 2022, 2022, 4231978. [Google Scholar] [CrossRef] [PubMed]
- Amaro, F.; Zé-Zé, L.; Alves, M.J. Sandfly-Borne Phleboviruses in Portugal: Four and Still Counting. Viruses, M: Number: 8 Publisher; 14, 1768. [CrossRef]
- Alemayehu, B.; Alemayehu, M. Leishmaniasis: A Review on Parasite, Vector and Reservoir Host. Health Science Journal 2017, 11, 1–6. [Google Scholar] [CrossRef]
- Pérez-doria, A.; Hernández-oviedo, E.; Elías-Bejarano, E. Brumptomyia hamata ( Psychodidae ), A New Addition To The Phlebotomine Fauna Of The Colombian Caribbean. Acta Biológica Colombiana 2009, 14, 135–140. [Google Scholar]
- Ready, P.D. Biology of Phlebotomine Sand Flies as Vectors of Disease Agents. Annual Review of Entomology 2013, 58, 227–250. [Google Scholar] [CrossRef]
- Raj, S.; Sasidharan, S.; Balaji, S.N.; Dubey, V.K.; Saudagar, P. Review on natural products as an alternative to contemporary anti-leishmanial therapeutics. Journal of Proteins and Proteomics 11, 1768–158. [CrossRef]
- de Salud y Protección Social de Colombia, M. Boletín Epidemiológico Semanal, 2022.
- Mann, S.; Frasca, K.; Scherrer, S.; Henao-Martínez, A.F.; Newman, S.; Ramanan, P.; Suarez, J.A. A Review of Leishmaniasis: Current Knowledge and Future Directions. Current Tropical Medicine Reports, 8, 121–132. [CrossRef]
- Catta-Preta, C.M.C.; Mottram, J.C. Drug candidate and target for leishmaniasis. Nature 2018, 560, 171–172. [Google Scholar] [CrossRef]
- Chappuis, F.; Sundar, S.; Hailu, A.; Ghalib, H.; Rijal, S.; Peeling, R.W.; Alvar, J.; Boelaert, M. Visceral leishmaniasis: What are the needs for diagnosis, treatment and control? Nature Reviews Microbiology 2007, 5, 873–882. [Google Scholar] [CrossRef]
- Rittig, M.G.; Bogdan, C. Leishmania-host-cell interaction: Complexities and alternative views. Parasitology Today 2000, 16, 292–297. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, C.; Estaquier, J.; Silvestre, R. Immune-metabolic interactions between Leishmania and macrophage host. Current Opinion in Microbiology 2021, 63, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, A.C. Parasitic Adaptive Mechanisms in Infection by Leishmania. Experimental and Molecular Pathology 2002, 72, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Nathan, C. Neutrophils and immunity: challenges and opportunities. Nature Reviews Immunology 2006, 6, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Torres-Vélez, C. ; María Cynthia Fuentes-Lacouture.; Yurany Duarte-Torres.; Myriam Beatriz AmayaBernal.; Jair Figueroa-Emiliani. Visceral Leishmaniasis in A Patient with Multiple Myeloma: Casuality or Causality? Journal of Medical Case Reports and Case Series, 3. [CrossRef]
- Charmoy, M.; Auderset, F.; Allenbach, C.; Tacchini-Cottier, F. The Prominent Role of Neutrophils during the Initial Phase of Infection by Leishmania Parasites. Journal of Biomedicine and Biotechnology 2010, 2010, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Laskay, T.; Zandbergen, G.v.; Solbach, W. Neutrophil granulocytes as host cells and transport vehicles for intracellular pathogens : Apoptosis as infection-promoting factor. Immunology 2008, 213, 183–191. [Google Scholar] [CrossRef]
- Naderer, T.; McConville, M.J. The Leishmania-macrophage interaction: a metabolic perspective. Cellular Microbiology 2008, 10, 301–308. [Google Scholar] [CrossRef]
- Naderer, T.; Vince, J.; McConville, M. Surface Determinants of Leishmania Parasites and their Role in Infectivity in the Mammalian Host. Current Molecular Medicine 2004, 4, 649–665. [Google Scholar] [CrossRef]
- de León Fraga, J. (Ed.) INMUNOLOGÍA by Thomas J. Kindt, Richard A. Goldsby, and Barbara A. Osborne, 6 ed.; McGraw-Hill, Interamericana, 2007.
- Parham, P. The Immune System, Fourth Edition, 4 ed.; Taylor and Francis Group, 2014; p. 624.
- Kima, P.E. Leishmania molecules that mediate intracellular pathogenesis. Microbes and Infection 2014, 16, 721–726. [Google Scholar] [CrossRef]
- McConville, M.J.; Naderer, T. Metabolic Pathways Required for the Intracellular Survival of Leishmania. Annual Review of Microbiology 2011, 65, 543–561. [Google Scholar] [CrossRef]
- Kanehisa, M. From genomics to chemical genomics: new developments in KEGG. Nucleic Acids Research 2006, 34, D354–D357. [Google Scholar] [CrossRef] [PubMed]
- Berman, H.M. The Protein Data Bank. Nucleic Acids Research 2000, 28, 235–242. [Google Scholar] [CrossRef]
- Cornish-Bowden, A. Current IUBMB recommendations on enzyme nomenclature and kinetics. Perspectives in Science 2014, 1, 74–87. [Google Scholar] [CrossRef]
- De Rycker, M.; Baragaña, B.; Duce, S.L.; Gilbert, I.H. Challenges and recent progress in drug discovery for tropical diseases. Nature, 559, 498–506. [CrossRef]
- Rodriguez-Contreras, D.; Hamilton, N. Gluconeogenesis in Leishmania mexicana. Journal of Biological Chemistry 2014, 289, 32989–33000. [Google Scholar] [CrossRef] [PubMed]
- Seidler, N.W. GAPDH: Biological Properties and Diversity; Vol. 985, Advances in Experimental Medicine and Biology, Springer Netherlands: Dordrecht, 2013; pp. 1–291. [Google Scholar] [CrossRef]
- Kim, H.; Feil, I.K.; Verlinde, C.L.M.J.; Petra, P.H.; Hol, W.G.J. Crystal Structure of Glycosomal Glyceraldehyde-3-phosphate Dehydrogenase from Leishmania mexicana: Implications for Structure-Based Drug Design and a New Position for the Inorganic Phosphate Binding Site. Biochemistry, 34, 14975–14986. [CrossRef]
- Pelley, J.W. Gluconeogenesis and Glycogen Metabolism. In Elsevier’s Integrated Biochemistry; Elsevier, 2007; pp. 65–71. [CrossRef]
- Minárik, P.; Tomásková, N.; Kollárová, M.; Antalík, M. Malate dehydrogenases–structure and function. General physiology and biophysics 2002, 21, 257–65. [Google Scholar]
- Gourley, D.G.; Schüttelkopf, A.W.; Leonard, G.A.; Luba, J.; Hardy, L.W.; Beverley, S.M.; Hunter, W.N. Pteridine reductase mechanism correlates pterin metabolism with drug resistance in trypanosomatid parasites. Nature Publishing Group 2001, 8, 521–525. [Google Scholar] [CrossRef]
- Vadloori, B.; Sharath, A.K.; Prabhu, N.P.; Maurya, R. Homology modelling, molecular docking, and molecular dynamics simulations reveal the inhibition of Leishmania donovani dihydrofolate reductase-thymidylate synthase enzyme by Withaferin-A. BMC Research Notes 2018, 11, 1–7. [Google Scholar] [CrossRef]
- Chang, C.f.; Papadopoulou, B.; Wang, J.; Bray, T.; Whiteley, J.M.; Lin, S.x.; Ouellette, M.; Al, W.E.T. Pterin and Folate Reduction by the Leishmania tarentolae H Locus Short-Chain Dehydrogenase / Reductase PTR1. Archives of Biochemistry and Biophysics 1997, 342, 197–202. [Google Scholar]
- Feliciano, P.R.; Cordeiro, A.T.; Costa-Filho, A.J.; Nonato, M.C. Cloning, expression, purification, and characterization of Leishmania major dihydroorotate dehydrogenase. Protein Expression and Purification 2006, 48, 98–103. [Google Scholar] [CrossRef]
- Cordeiro, A.T.; Feliciano, P.R.; Pinheiro, M.P.; Nonato, M.C. Crystal structure of dihydroorotate dehydrogenase from Leishmania major. Biochimie 2012, 94, 1739–1748. [Google Scholar] [CrossRef]
- Gommers-Ampt, J.H.; Van Leeuwen, F.; de Beer, A.L.; Vliegenthart, J.F.; Dizdaroglu, M.; Kowalak, J.A.; Crain, P.F.; Borst, P. β-d-glucosyl-hydroxymethyluracil: A novel modified base present in the DNA of the parasitic protozoan T. brucei. Cell 1993, 75, 1129–1136. [Google Scholar] [CrossRef] [PubMed]
- van Leeuwen, F.; Taylor, M.C.; Mondragon, A.; Moreau, H.; Gibson, W.; Kieft, R.; Borst, P. -D-Glucosyl-hydroxymethyluracil is a conserved DNA modification in kinetoplastid protozoans and is abundant in their telomeres. Proceedings of the National Academy of Sciences 2002, 95, 2366–2371. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Genest, P.A.; ter Riet, B.; Sweeney, K.; DiPaolo, C.; Kieft, R.; Christodoulou, E.; Perrakis, A.; Simmons, J.M.; Hausinger, R.P.; van Luenen, H.G.; Rigden, D.J.; Sabatini, R.; Borst, P. The protein that binds to DNA base J in trypanosomatids has features of a thymidine hydroxylase. Nucleic Acids Research 2007, 35, 2107–2115. [Google Scholar] [CrossRef] [PubMed]
- Heidebrecht, T.; Christodoulou, E.; Chalmers, M.J.; Jan, S.; Ter Riet, B.; Grover, R.K.; Joosten, R.P.; Littler, D.; Van Luenen, H.; Griffin, P.R.; Wentworth, P.; Borst, P.; Perrakis, A. The structural basis for recognition of base J containing DNA by a novel DNA binding domain in JBP1, 2011. [CrossRef]
- Kryshtafovych, A.; Moult, J.; Bartual, S.G.; Bazan, J.F.; Berman, H.; Casteel, D.E.; Christodoulou, E.; Everett, J.K.; Hausmann, J.; Heidebrecht, T.; Hills, T.; Hui, R.; Hunt, J.F.; Seetharaman, J.; Joachimiak, A.; Kennedy, M.A.; Kim, C.; Lingel, A.; Michalska, K.; Montelione, G.T.; Otero, J.M.; Perrakis, A.; Pizarro, J.C.; van Raaij, M.J.; Ramelot, T.A.; Rousseau, F.; Tong, L.; Wernimont, A.K.; Young, J.; Schwede, T. Target highlights in CASP9: Experimental target structures for the critical assessment of techniques for protein structure prediction. Proteins: Structure, Function, and Bioinformatics 2011, 79, 6–20. [Google Scholar] [CrossRef]
- Barr, S.D.; Gedamu, L. Role of Peroxidoxins in Leishmania chagasi Survival. Journal of Biological Chemistry 2003, 278, 10816–10823. [Google Scholar] [CrossRef]
- Murray, H.W.; Nathan, C.F. Macrophage microbicidal mechanisms in vivo: reactive nitrogen versus oxygen intermediates in the killing of intracellular visceral Leishmania donovani. The Journal of experimental medicine 1999, 189, 741–6. [Google Scholar] [CrossRef]
- Nathan, C.; Shiloh, M.U. Reactive oxygen and nitrogen intermediates in the relationship between mammalian hosts and microbial pathogens. Proceedings of the National Academy of Sciences of the United States of America 2000, 97, 8841–8848. [Google Scholar] [CrossRef]
- Mutlu, O. In silico molecular modeling and docking studies on the leishmanial tryparedoxin peroxidase. Brazilian Archives of Biology and Technology 2014, 57, 244–252. [Google Scholar] [CrossRef]
- Fiorillo, A.; Colotti, G.; Boffi, A.; Baiocco, P.; Ilari, A. The Crystal Structures of the Tryparedoxin-Tryparedoxin Peroxidase Couple Unveil the Structural Determinants of Leishmania Detoxification Pathway. PLoS Neglected Tropical Diseases 2012, 6, e1781. [Google Scholar] [CrossRef]
- Bose, M.; Saha, R.; Sen Santara, S.; Mukherjee, S.; Roy, J.; Adak, S. Protection against peroxynitrite by pseudoperoxidase from Leishmania major. Free Radical Biology and Medicine 2012, 53, 1819–1828. [Google Scholar] [CrossRef]
- Chreifi, G.; Dejam, D.; Poulos, T.L. Crystal structure and functional analysis of Leishmania major pseudoperoxidase. Journal of Biological Inorganic Chemistry 2017. [Google Scholar] [CrossRef]
- Phan, I.Q.H.; Davies, D.R.; Moretti, N.S.; Shanmugam, D.; Cestari, I.; Anupama, A.; Fairman, J.W.; Edwards, T.E.; Stuart, K.; Schenkman, S.; Myler, P.J. Iron superoxide dismutases in eukaryotic pathogens: new insights from Apicomplexa and Trypanosoma structures. Acta Crystallographica Section F Structural Biology Communications 2015, 71, 615–621. [Google Scholar] [CrossRef]
- GHOSH, S.; GOSWAMI, S.; ADHYA, S. Role of superoxide dismutase in survival of Leishmania within the macrophage. Biochemical Journal 2003, 369, 447–452. [Google Scholar] [CrossRef] [PubMed]
- Slauch, J.M. How does the oxidative burst of macrophages kill bacteria? Still an open question. Molecular Microbiology 2011, 80, 580–583. [Google Scholar] [CrossRef] [PubMed]
- Ilari, A.; Baiocco, P.; Messori, L.; Fiorillo, A.; Boffi, A.; Gramiccia, M.; Di Muccio, T.; Colotti, G. A gold-containing drug against parasitic polyamine metabolism: the X-ray structure of trypanothione reductase from Leishmania infantum in complex with auranofin reveals a dual mechanism of enzyme inhibition. Amino Acids 2012, 42, 803–811. [Google Scholar] [CrossRef]
- Fairlamb, A.H.; Cerami, A. Metabolism and Functions of Trypanothione in the Kinetoplastida. Annual Review of Microbiology 1992, 46, 695–729. [Google Scholar] [CrossRef] [PubMed]
- Bernardes, L.; Zani, C.; Carvalho, I. Trypanosomatidae Diseases: From the Current Therapy to the Efficacious Role of Trypanothione Reductase in Drug Discovery. Current Medicinal Chemistry 2013, 20, 2673–2696. [Google Scholar] [CrossRef] [PubMed]
- Kabututu, Z.; Martin, S.K.; Nozaki, T.; Kawazu, S.i.; Okada, T.; Munday, C.J.; Duszenko, M.; Lazarus, M.; Thuita, L.W.; Urade, Y.; Kubata, B.K. Prostaglandin production from arachidonic acid and evidence for a 9,11-endoperoxide prostaglandin H2 reductase in Leishmania. International Journal for Parasitology 2002, 32, 1693–1700. [Google Scholar] [CrossRef]
- Moen, S.O.; Fairman, J.W.; Barnes, S.R.; Sullivan, A.; Nakazawa-Hewitt, S.; Van Voorhis, W.C.; Staker, B.L.; Lorimer, D.D.; Myler, P.J.; Edwards, T.E. Structures of prostaglandin F synthase from the protozoa Leishmania major and Trypanosoma cruzi with NADP. Acta Crystallographica Section F Structural Biology Communications 2015, 71, 609–614. [Google Scholar] [CrossRef]
- Hargrove, T.Y.; Wawrzak, Z.; Liu, J.; Nes, W.D.; Waterman, M.R.; Lepesheva, G.I. Substrate preferences and catalytic parameters determined by structural characteristics of sterol 14$α$-demethylase (CYP51) from Leishmania infantum. Journal of Biological Chemistry 2011. [Google Scholar] [CrossRef]
- Manhas, R.; Tripathi, P.; Khan, S.; Sethu Lakshmi, B.; Lal, S.K.; Gowri, V.S.; Sharma, A.; Madhubala, R. Identification and Functional Characterization of a Novel Bacterial Type Asparagine Synthetase A. Journal of Biological Chemistry 2014, 289, 12096–12108. [Google Scholar] [CrossRef]
- Führing, J.; Cramer, J.T.; Routier, F.H.; Lamerz, A.C.; Baruch, P.; Gerardy-Schahn, R.; Fedorov, R. Catalytic Mechanism and Allosteric Regulation of UDP-Glucose Pyrophosphorylase from Leishmania major. ACS Catalysis 2013, 3, 2976–2985. [Google Scholar] [CrossRef]
- Martin, J.L.; Yates, P.A.; Boitz, J.M.; Koop, D.R.; Fulwiler, A.L.; Cassera, M.B.; Ullman, B.; Carter, N.S. A role for adenine nucleotides in the sensing mechanism to purine starvation in Leishmania donovani. Molecular Microbiology 2016, 101, 299–313. [Google Scholar] [CrossRef]
- Boitz, J.M.; Strasser, R.; Yates, P.A.; Jardim, A.; Ullman, B. Adenylosuccinate synthetase and adenylosuccinate lyase deficiencies trigger growth and infectivity deficits in Leishmania donovani. Journal of Biological Chemistry 2013, 288, 8977–8990. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.; Silva, C.H.; Iulek, J.; Oliva, G.; Thiemann, O.H. Crystal structure of adenine phosphoribosyltransferase from Leishmania tarentolae: Potential implications for APRT catalytic mechanism. Biochimica et Biophysica Acta - Proteins and Proteomics, 2004. [Google Scholar] [CrossRef]
- Phillips, C.L.; Ullman, B.; Brennan, R.G.; Hill, C.P. Crystal structures of adenine phosphoribosyltransferase from Leishmania donovani. EMBO Journal 1999, 18, 3533–3545. [Google Scholar] [CrossRef] [PubMed]
- Resh, M.D. Fatty acylation of proteins: new insights into membrane targeting of myristoylated and palmitoylated proteins. Biochimica et Biophysica Acta (BBA) - Molecular Cell Research 1999, 1451, 1–16. [Google Scholar] [CrossRef]
- Robinson, D.A.; Wyatt, P.G. Identification and structure solution of fragment hits against kinetoplastid N -myristoyltransferase. Acta Crystallographica Section F Structural Biology Communications 2015, 71, 586–593. [Google Scholar] [CrossRef]
- Selvakumar, P.; Kumar, S.; Dimmock, J.R.; Sharma, R.K. NMT1 (N-myristoyltransferase 1). Atlas of genetics and cytogenetics in oncology and haematology, 15, 570–575. [CrossRef]
- Price, H.P.; Menon, M.R.; Panethymitaki, C.; Goulding, D.; McKean, P.G.; Smith, D.F. Myristoyl-CoA:Protein N -Myristoyltransferase, an Essential Enzyme and Potential Drug Target in Kinetoplastid Parasites. Journal of Biological Chemistry 2003, 278, 7206–7214. [Google Scholar] [CrossRef]
- French, J.B.; Yates, P.A.; Soysa, D.R.; Boitz, J.M.; Carter, N.S.; Chang, B.; Ullman, B.; Ealick, S.E. The Leishmania donovani UMP synthase is essential for promastigote viability and has an unusual tetrameric structure that exhibits substrate-controlled oligomerization. Journal of Biological Chemistry 2011. [Google Scholar] [CrossRef]
- Jones, M.E. Pyrimidine Nucleotide Biosynthesis in Animals: Genes, Enzymes, and Regulation of UMP Biosynthesis. Annual Review of Biochemistry 1980, 49, 253–279. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.A.M.; Westrop, G.D.; Coombs, G.H. Two pathways for cysteine biosynthesis in Leishmania major. Biochemical Journal 2009, 420, 451–462. [Google Scholar] [CrossRef]
- Oza, S.L.; Shaw, M.P.; Wyllie, S.; Fairlamb, A.H. Trypanothione biosynthesis in Leishmania major. Molecular and Biochemical Parasitology 2005, 139, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Krauth-Siegel, R.L.; Comini, M.A. Redox control in trypanosomatids, parasitic protozoa with trypanothione-based thiol metabolism. Biochimica et Biophysica Acta (BBA) - General Subjects 2008, 1780, 1236–1248. [Google Scholar] [CrossRef] [PubMed]
- Fyfe, P.K.; Westrop, G.D.; Ramos, T.; Müller, S.; Coombs, G.H.; Hunter, W.N. Structure of Leishmania major cysteine synthase. Acta Crystallographica Section F Structural Biology and Crystallization Communications 2012, 68, 738–743. [Google Scholar] [CrossRef]
- McConville, M.J.; de Souza, D.; Saunders, E.; Likic, V.A.; Naderer, T. Living in a phagolysosome; metabolism of Leishmania amastigotes. Trends in Parasitology 2007, 23, 368–375. [Google Scholar] [CrossRef]
- Mejia, E.; Burak, M.; Alonso, A.; Larraga, V.; Kunkel, T.A.; Bebenek, K.; Garcia-Diaz, M. Structures of the Leishmania infantum polymerase beta. DNA Repair 2014, 18, 1–9. [Google Scholar] [CrossRef]
- Hammond, D.J.; Gutteridge, W.E. Purine and pyrimidine metabolism in the trypanosomatidae. Molecular and Biochemical Parasitology 1984, 13, 243–261. [Google Scholar] [CrossRef] [PubMed]
- Al-Madhoun, A. The Role of Thymidine Kinases in the Activation of Pyrimidine Nucleoside Analogues. Mini Reviews in Medicinal Chemistry 2004, 4, 341–350. [Google Scholar] [CrossRef]
- Timm, J.; Bosch-Navarrete, C.; Recio, E.; Nettleship, J.E.; Rada, H.; González-Pacanowska, D.; Wilson, K.S. Structural and Kinetic Characterization of Thymidine Kinase from Leishmania major. PLOS Neglected Tropical Diseases 2015, 9, e0003781. [Google Scholar] [CrossRef]
- Aripirala, S.; Gonzalez-Pacanowska, D.; Oldfield, E.; Kaiser, M.; Amzel, L.M.; Gabelli, S.B. Structural and thermodynamic basis of the inhibition of Leishmania major farnesyl diphosphate synthase by nitrogen-containing bisphosphonates. Acta Crystallographica Section D: Biological Crystallography, 2014. [Google Scholar] [CrossRef]
- de Mattos Oliveira, L.; Araújo, J.S.C.; Bacelar Costa Junior, D.; Santana, I.B.; Duarte, A.A.; Leite, F.H.A.; Benevides, R.G.; Coelho dos Santos Junior, M. Pharmacophore modeling, docking and molecular dynamics to identify Leishmania major farnesyl pyrophosphate synthase inhibitors. Journal of Molecular Modeling 2018, 24, 314. [Google Scholar] [CrossRef]
- Schomburg, I.; Jeske, L.; Ulbrich, M.; Placzek, S.; Chang, A.; Schomburg, D. The BRENDA enzyme information system–From a database to an expert system. Journal of Biotechnology 2017, 261, 194–206. [Google Scholar] [CrossRef] [PubMed]
- Abendroth, J.; Choi, R.; Wall, A.; Clifton, M.C.; Lukacs, C.M.; Staker, B.L.; Van Voorhis, W.; Myler, P.; Lorimer, D.D.; Edwards, T.E. Structures of aspartate aminotransferases from Trypanosoma brucei, Leishmania major and Giardia lamblia. Acta Crystallographica Section F:Structural Biology Communications, 2015. [Google Scholar] [CrossRef]
- Alphey, M.S.; Williams, R.A.; Mottram, J.C.; Coombs, G.H.; Hunter, W.N. The crystal structure of leishmania major 3-mercaptopyruvate sulfurtransferase. A three-domain architecture with a serine protease-like triad at the active site. Journal of Biological Chemistry 2003, 278, 48219–48227. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.A.M.; Kelly, S.M.; Mottram, J.C.; Coombs, G.H. 3-Mercaptopyruvate Sulfurtransferase of Leishmania Contains an Unusual C-terminal Extension and Is Involved in Thioredoxin and Antioxidant Metabolism. Journal of Biological Chemistry 2003, 278, 1480–1486. [Google Scholar] [CrossRef] [PubMed]
- VEITCH, N.J.; MAUGERI, D.A.; CAZZULO, J.J.; LINDQVIST, Y.; BARRETT, M.P. Transketolase from Leishmania mexicana has a dual subcellular localization. Biochemical Journal 2004, 382, 759–767. [Google Scholar] [CrossRef]
- Rigden, D.J.; Phillips, S.E.; Michels, P.A.; Fothergill-Gilmore, L.A. The structure of pyruvate kinase from Leishmania mexicana reveals details of the allosteric transition and unusual effector specificity 1 1Edited by I. A. Wilson. Journal of Molecular Biology 1999, 291, 615–635. [Google Scholar] [CrossRef]
- Shi, W.; Schramm, V.L.; Almo, S.C. Nucleoside Hydrolase from Leishmania major. The Journal of Biological Chemistry 1999, 274, 21114–21120. [Google Scholar] [CrossRef]
- Schlagenhauf, E.; Etges, R.; Metcalf, P. The crystal structure of the Leishmania major surface proteinase leishmanolysin (gp63). Structure 1998, 6, 1035–1046. [Google Scholar] [CrossRef]
- McKerrow, J.H.; Caffrey, C.; Kelly, B.; Loke, P.; Sajid, M. PROTEASES IN PARASITIC DISEASES. Annual Review of Pathology: Mechanisms of Disease 2006, 1, 497–536. [Google Scholar] [CrossRef]
- McLuskey, K.; Paterson, N.G.; Bland, N.D.; Isaacs, N.W.; Mottram, J.C. Crystal Structure of Leishmania major Oligopeptidase B Gives Insight into the Enzymatic Properties of a Trypanosomatid Virulence Factor. Journal of Biological Chemistry 2010, 285, 39249–39259. [Google Scholar] [CrossRef]
- Teixeira, P.C.; Velasquez, L.G.; Lepique, A.P.; de Rezende, E.; Bonatto, J.M.C.; Barcinski, M.A.; Cunha-Neto, E.; Stolf, B.S. Regulation of Leishmania (L.) amazonensis Protein Expression by Host T Cell Dependent Responses: Differential Expression of Oligopeptidase B, Tryparedoxin Peroxidase and HSP70 Isoforms in Amastigotes Isolated from BALB/c and BALB/c Nude Mice. PLOS Neglected Tropical Diseases 2015, 9, e0003411. [Google Scholar] [CrossRef]
- Timm, J.; Valente, M.; García-Caballero, D.; Wilson, K.S.; González-Pacanowska, D. Structural Characterization of Acidic M17 Leucine Aminopeptidases from the TriTryps and Evaluation of Their Role in Nutrient Starvation in Trypanosoma brucei. mSphere 2017, 2. [Google Scholar] [CrossRef]
- Leitherer, S.; Clos, J.; Liebler-Tenorio, E.M.; Schleicher, U.; Bogdan, C.; Soulat, D. Characterization of the protein tyrosine phosphatase LmPRL-1 secreted by Leishmania major via the exosome pathway. Infection and Immunity 2017. [Google Scholar] [CrossRef] [PubMed]
- D’Antonio, E.L.; Ullman, B.; Roberts, S.C.; Dixit, U.G.; Wilson, M.E.; Hai, Y.; Christianson, D.W. Crystal structure of arginase from Leishmania mexicana and implications for the inhibition of polyamine biosynthesis in parasitic infections. Archives of Biochemistry and Biophysics 2013, 535, 163–176. [Google Scholar] [CrossRef] [PubMed]
- Feliciano, P.R.; Drennan, C.L.; Nonato, M.C. Crystal structure of an Fe-S cluster-containing fumarate hydratase enzyme from Leishmania major reveals a unique protein fold. Proceedings of the National Academy of Sciences 2016, 113, 9804–9809. [Google Scholar] [CrossRef]
- Chudzik, D.M.; Michels, P.A.; De Walque, S.; Hol, W.G. Structures of type 2 peroxisomal targeting signals in two trypanosomatid aldolases. Journal of Molecular Biology 2000, 300, 697–707. [Google Scholar] [CrossRef]
- Williams, J.C.; Zeelen, J.P.; Neubauer, G.; Vriend, G.; Backmann, J.; Michels, P.a.; Lambeir, a.M.; Wierenga, R.K. Structural and mutagenesis studies of leishmania triosephosphate isomerase: a point mutation can convert a mesophilic enzyme into a superstable enzyme without losing catalytic power. Protein engineering 1999, 12, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Lambeir, A.M.; Backmann, J.; Ruiz-Sanz, J.; Filimonov, V.; Nielsen, J.E.; Kursula, I.; Norledge, B.V.; Wierenga, R.K. The ionization of a buried glutamic acid is thermodynamically linked to the stability of Leishmania mexicana triose phosphate isomerase. European Journal of Biochemistry 2000, 267, 2516–2524. [Google Scholar] [CrossRef] [PubMed]
- Bateman, A.; Martin, M.J.; O’Donovan, C.; Magrane, M.; Alpi, E.; Antunes, R.; Bely, B.; Bingley, M.; Bonilla, C.; Britto, R.; Bursteinas, B.; Bye-AJee, H.; Cowley, A.; Da Silva, A.; De Giorgi, M.; Dogan, T.; Fazzini, F.; Castro, L.G.; Figueira, L.; Garmiri, P.; Georghiou, G.; Gonzalez, D.; Hatton-Ellis, E.; Li, W.; Liu, W.; Lopez, R.; Luo, J.; Lussi, Y.; MacDougall, A.; Nightingale, A.; Palka, B.; Pichler, K.; Poggioli, D.; Pundir, S.; Pureza, L.; Qi, G.; Rosanoff, S.; Saidi, R.; Sawford, T.; Shypitsyna, A.; Speretta, E.; Turner, E.; Tyagi, N.; Volynkin, V.; Wardell, T.; Warner, K.; Watkins, X.; Zaru, R.; Zellner, H.; Xenarios, I.; Bougueleret, L.; Bridge, A.; Poux, S.; Redaschi, N.; Aimo, L.; ArgoudPuy, G.; Auchincloss, A.; Axelsen, K.; Bansal, P.; Baratin, D.; Blatter, M.C.; Boeckmann, B.; Bolleman, J.; Boutet, E.; Breuza, L.; Casal-Casas, C.; De Castro, E.; Coudert, E.; Cuche, B.; Doche, M.; Dornevil, D.; Duvaud, S.; Estreicher, A.; Famiglietti, L.; Feuermann, M.; Gasteiger, E.; Gehant, S.; Gerritsen, V.; Gos, A.; Gruaz-Gumowski, N.; Hinz, U.; Hulo, C.; Jungo, F.; Keller, G.; Lara, V.; Lemercier, P.; Lieberherr, D.; Lombardot, T.; Martin, X.; Masson, P.; Morgat, A.; Neto, T.; Nouspikel, N.; Paesano, S.; Pedruzzi, I.; Pilbout, S.; Pozzato, M.; Pruess, M.; Rivoire, C.; Roechert, B.; Schneider, M.; Sigrist, C.; Sonesson, K.; Staehli, S.; Stutz, A.; Sundaram, S.; Tognolli, M.; Verbregue, L.; Veuthey, A.L.; Wu, C.H.; Arighi, C.N.; Arminski, L.; Chen, C.; Chen, Y.; Garavelli, J.S.; Huang, H.; Laiho, K.; McGarvey, P.; Natale, D.A.; Ross, K.; Vinayaka, C.R.; Wang, Q.; Wang, Y.; Yeh, L.S.; Zhang, J. UniProt: the universal protein knowledgebase. Nucleic Acids Research 2017, 45, D158–D169. [Google Scholar] [CrossRef]
- Larson, E.T.; Kim, J.E.; Zucker, F.H.; Kelley, A.; Mueller, N.; Napuli, A.J.; Verlinde, C.L.; Fan, E.; Buckner, F.S.; Van Voorhis, W.C.; Merritt, E.A.; Hol, W.G. Structure of Leishmania major methionyl-tRNA synthetase in complex with intermediate products methionyladenylate and pyrophosphate. Biochimie 2011, 93, 570–582. [Google Scholar] [CrossRef]
- Larson, E.T.; Kim, J.E.; Castaneda, L.J.; Napuli, A.J.; Zhang, Z.; Fan, E.; Zucker, F.H.; Verlinde, C.L.M.J.; Buckner, F.S.; Van Voorhis, W.C.; Hol, W.G.J.; Merritt, E.A. The double-length tyrosyl-tRNA synthetase from the eukaryote leishmania major forms an intrinsically asymmetric pseudo-dimer. Journal of Molecular Biology 2011. [Google Scholar] [CrossRef]
- Kamir, D.; Zierow, S.; Leng, L.; Cho, Y.; Diaz, Y.; Griffith, J.; McDonald, C.; Merk, M.; Mitchell, R.A.; Trent, J.; Chen, Y.; Kwong, Y.K.A.; Xiong, H.; Vermeire, J.; Cappello, M.; McMahon-Pratt, D.; Walker, J.; Bernhagen, J.; Lolis, E.; Bucala, R. A Leishmania Ortholog of Macrophage Migration Inhibitory Factor Modulates Host Macrophage Responses. The Journal of Immunology 2008, 180, 8250–8261. [Google Scholar] [CrossRef] [PubMed]
- Arya, R.; Sharma, B.; Dhembla, C.; Pal, R.K.; Patel, A.K.; Sundd, M.; Ghosh, B.; Makde, R.D.; Kundu, S. A conformational switch from a closed apo- to an open holo-form equips the acyl carrier protein for acyl chain accommodation. Biochimica et Biophysica Acta - Proteins and Proteomics 2019, 1867, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Chawla, B.; Madhubala, R. Drug targets in Leishmania. Journal of Parasitic Diseases 2010, 34, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Soto, J.; Soto, P. Miltefosina oral para el tratamiento de la leishmaniasis. Biomédica 2012, 26, 207. [Google Scholar] [CrossRef]
- Ramón Azanza, J.; García-Quetglas, E.; Sádaba, B. Farmacología de los azoles. Rev Iberoam Micol 2007, 24, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Tiuman, T.S.; Santos, A.O.; Ueda-Nakamura, T.; Filho, B.P.D.; Nakamura, C.V. Recent advances in leishmaniasis treatment. International Journal of Infectious Diseases 2011, 15. [Google Scholar] [CrossRef]
- Sen, R.; Chatterjee, M. Plant derived therapeutics for the treatment of Leishmaniasis. Phytomedicine 2011, 18, 1056–1069. [Google Scholar] [CrossRef]
- Gutiérrez-Rebolledo, G.A.; Drier-Jonas, S.; Jiménez-Arellanes, M.A. Natural compounds and extracts from Mexican medicinal plants with anti-leishmaniasis activity: An update. Asian Pacific Journal of Tropical Medicine 2017, 10, 1105–1110. [Google Scholar] [CrossRef]
- Zahedifard, F.; Rafati, S. Prospects for antimicrobial peptide-based immunotherapy approaches in <i>Leishmania</i> control. Expert Review of Anti-infective Therapy 2018, 16, 461–469. [Google Scholar] [CrossRef]
- Patiño-Márquez, I.A.; Manrique-Moreno, M.; Patiño-González, E.; Jemioła-Rzemińska, M.; Strzałka, K. Effect of antimicrobial peptides from Galleria mellonella on molecular models of Leishmania membrane. Thermotropic and fluorescence anisotropy study. The Journal of Antibiotics 2018, 71, 642–652. [Google Scholar] [CrossRef]
- Balboa, M.A.; Balsinde, J. Phospholipases: From Structure to Biological Function. Biomolecules, 11, 428. [CrossRef]
- Saddhe, A.A.; Potocký, M. Comparative phylogenomic and structural analysis of canonical secretory PLA2 and novel PLA2-like family in plants. Frontiers in Plant Science, 14.
- Tonello, F.; Rigoni, M. Cellular Mechanisms of Action of Snake Phospholipase A2 Toxins. In Snake Venoms; Inagaki, H.; Vogel, C.W.; Mukherjee, A.K.; Rahmy, T.R.; Gopalakrishnakone, P., Eds.; Toxinology, Springer Netherlands, 2017; pp. 49–65. [CrossRef]
- Jaiswal, A.; Rehman, R.; Dutta, J.; Singh, S.; Ray, A.; Shridhar, M.; Jaisankar, J.; Bhatt, M.; Khandelwal, D.; Sahoo, B.; Ram, A.; Mabalirajan, U. Cellular Distribution of Secreted Phospholipase A2 in Lungs of IPF Patients and Its Inhibition in Bleomycin-Induced Pulmonary Fibrosis in Mice. Cells, 12, 1044. [CrossRef]
- Castro-Amorim, J.; Novo de Oliveira, A.; Da Silva, S.L.; Soares, A.M.; Mukherjee, A.K.; Ramos, M.J.; Fernandes, P.A. Catalytically Active Snake Venom PLA2 Enzymes: An Overview of Its Elusive Mechanisms of Reaction. Journal of Medicinal Chemistry, 66, 5364–5376. [CrossRef]
- Moreira, V.; Leiguez, E.; Janovits, P.M.; Maia-Marques, R.; Fernandes, C.M.; Teixeira, C. Inflammatory Effects of Bothrops Phospholipases A2: Mechanisms Involved in Biosynthesis of Lipid Mediators and Lipid Accumulation. Toxins, 13, 868. [CrossRef]
- Lee, K.S.; Kim, B.Y.; Park, M.J.; Deng, Y.; Kim, J.M.; Kim, Y.H.; Heo, E.J.; Yoon, H.J.; Lee, K.Y.; Choi, Y.S.; Jin, B.R. Bee Venom Induces Acute Inflammation through a H2O2-Mediated System That Utilizes Superoxide Dismutase. Toxins, 14, 558. [CrossRef]
- Ma, X.; Liu, H.; Liu, Z.; Wang, Y.; Zhong, Z.; Peng, G.; Gu, Y. Trichosporon asahii PLA2 Gene Enhances Drug Resistance to Azoles by Improving Drug Efflux and Biofilm Formation. International Journal of Molecular Sciences, 24, 8855. [CrossRef]
- Soltan-Alinejad, P.; Alipour, H.; Meharabani, D.; Azizi, K. Therapeutic Potential of Bee and Scorpion Venom Phospholipase A2 (PLA2): A Narrative Review. Iranian Journal of Medical Sciences, 47, 300–313. [CrossRef]
- Rojas-Pirela, M.; Kemmerling, U.; Quiñones, W.; Michels, P.A.M.; Rojas, V. Antimicrobial Peptides (AMPs): Potential Therapeutic Strategy against Trypanosomiases? Biomolecules, M: Number: 4 Publisher; 13, 599. [CrossRef]
- Borges, I.P.; Castanheira, L.E.; Barbosa, B.F.; de Souza, D.L.N.; da Silva, R.J.; Mineo, J.R.; Tudini, K.A.Y.; Rodrigues, R.S.; Ferro, E.A.V.; de Melo Rodrigues, V. Anti-parasitic effect on Toxoplasma gondii induced by BnSP-7, a Lys49-phospholipase A2 homologue from Bothrops pauloensis venom. Toxicon: Official Journal of the International Society on Toxinology, 119, 84–91. [CrossRef]
Figure 3.
Leishmania taxonomic classification and geographical distribution. Leishmania genus: euleishmania and paraleishmania. Paraleishmania is conformed by L. (endotrypanum) subgenus and euleishmania by L. (viannia), L. (leishmania) and L. (sauroleishmania) subgenus. [17,21,22,23]
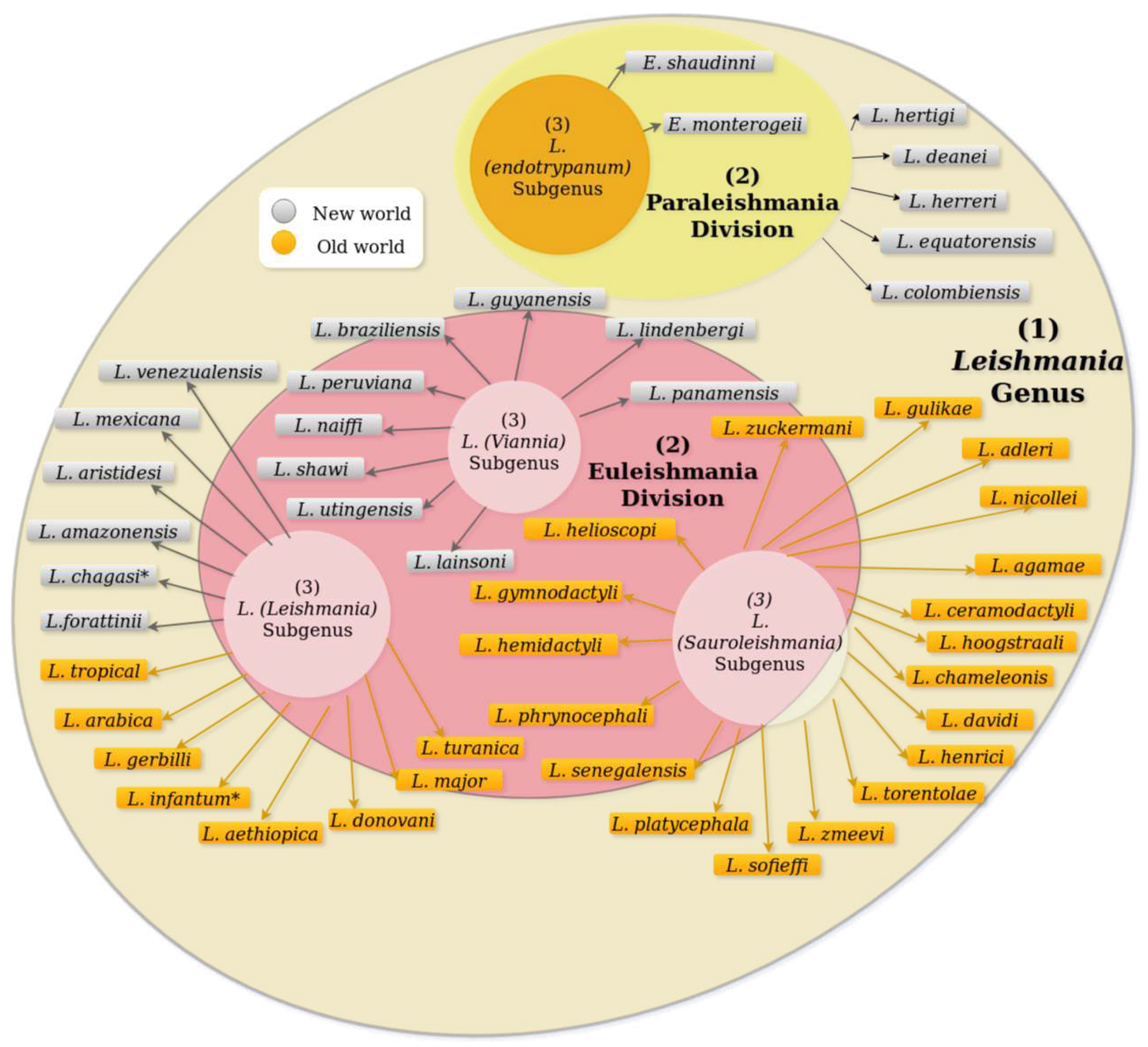
Figure 4.
Leishmania life cycle can be divided in five steps. Step 1: Virulent metacyclic promastigotes are egested when a female sandfly carrier bites a possible host. Step 2: Here there are two possible paths. Direct path: promastigotes are phagocytized by macrophage cells. Indirect path: promastigotes are phagocytized by neutrophil cells, subsequently, neutrophils are phagocytized by macrophage. At the end of step 2, promastigotes end up inside a macrophage phagolysosome. Step 3: The promastigote parasite produces glutathione transferase, a protein that protects it from the acidic conditions of the phagolysosome, then initiating the replication process . Step 4: Transformation of vector promastigote into amastigote takes place. Leishmania amastigotes are experts at exploiting host cell machinery to thrive. Step 5: Amastigotes are then taken out when a sand fly, either infected or not, bites the host, closing the cycle.
Figure 4.
Leishmania life cycle can be divided in five steps. Step 1: Virulent metacyclic promastigotes are egested when a female sandfly carrier bites a possible host. Step 2: Here there are two possible paths. Direct path: promastigotes are phagocytized by macrophage cells. Indirect path: promastigotes are phagocytized by neutrophil cells, subsequently, neutrophils are phagocytized by macrophage. At the end of step 2, promastigotes end up inside a macrophage phagolysosome. Step 3: The promastigote parasite produces glutathione transferase, a protein that protects it from the acidic conditions of the phagolysosome, then initiating the replication process . Step 4: Transformation of vector promastigote into amastigote takes place. Leishmania amastigotes are experts at exploiting host cell machinery to thrive. Step 5: Amastigotes are then taken out when a sand fly, either infected or not, bites the host, closing the cycle.
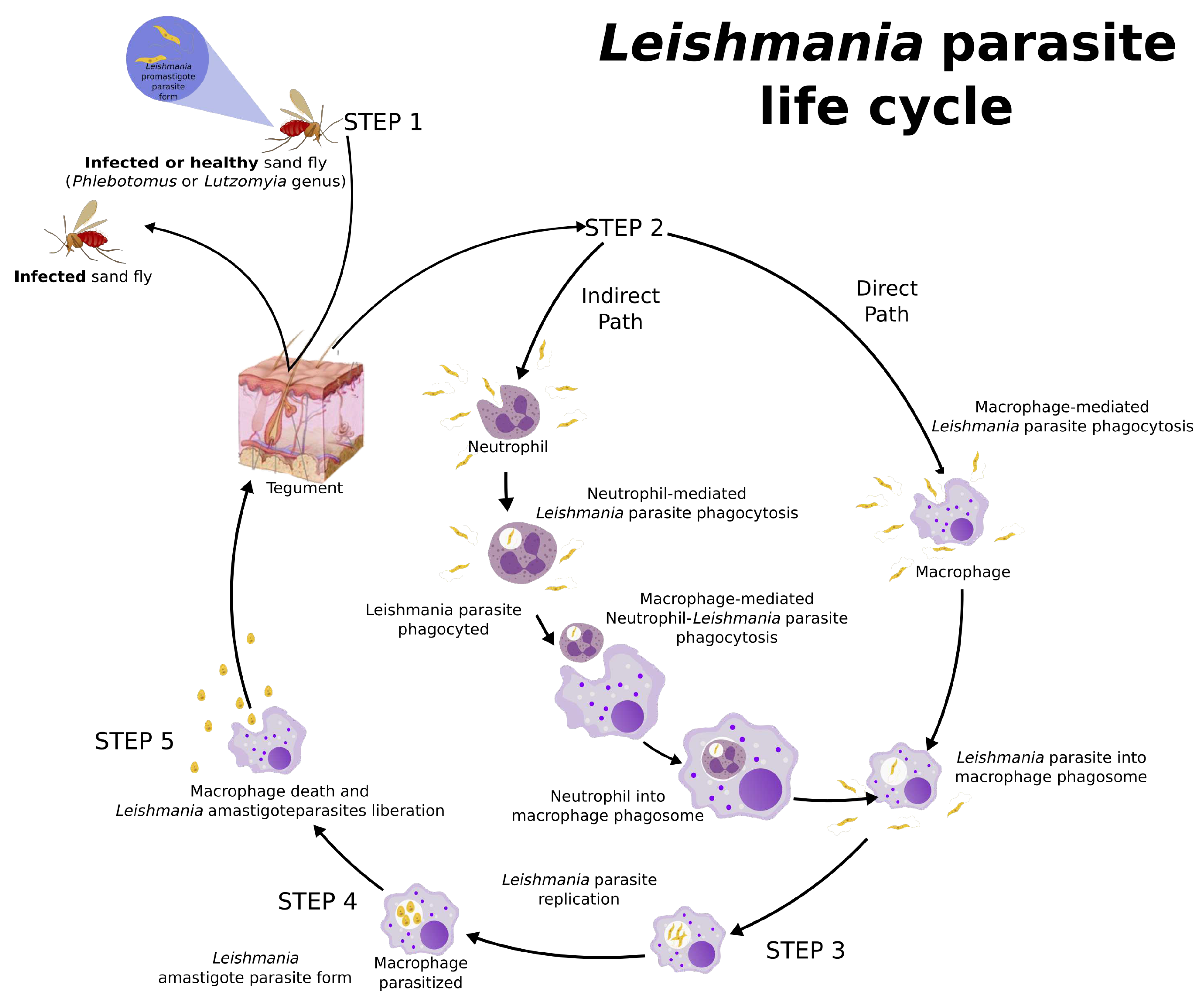
Figure 5.
This figure shows Leishmania metabolic pathways related with retrieved proteins from databases. The PDB IDs 1E92, 2XSE, 3VIA, 4F2N, 4F40, 4UCM, 2XE4, 3S4O, 5NTH, 4S1E, 3P0I, 3KFL and 5ZWS, do not appear in metabolic pathway databases
Figure 5.
This figure shows Leishmania metabolic pathways related with retrieved proteins from databases. The PDB IDs 1E92, 2XSE, 3VIA, 4F2N, 4F40, 4UCM, 2XE4, 3S4O, 5NTH, 4S1E, 3P0I, 3KFL and 5ZWS, do not appear in metabolic pathway databases
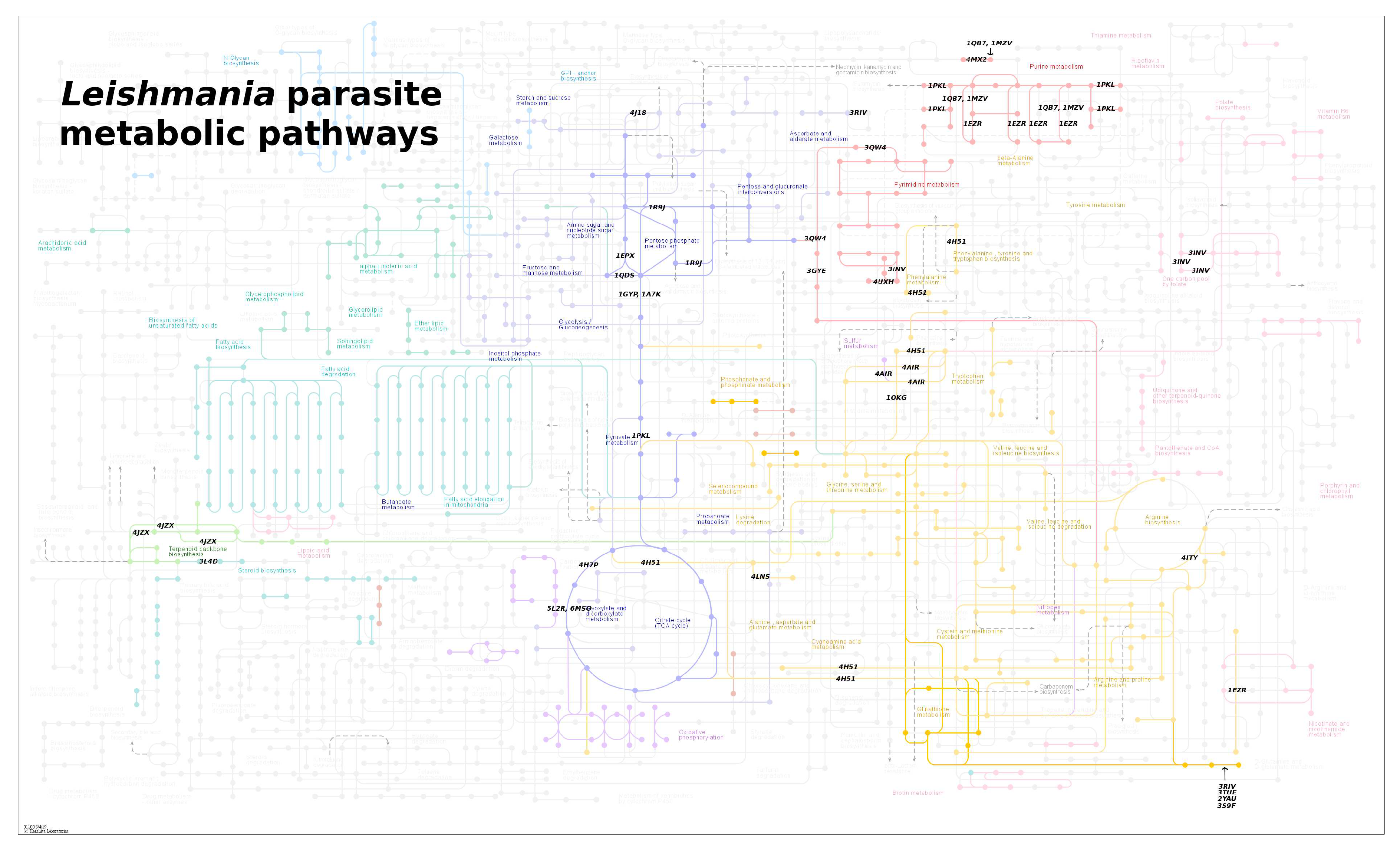
Figure 6.
Proteins linked with specific Leishmania metabolic pathways and proteins with no metabolic pathway association.
Figure 6.
Proteins linked with specific Leishmania metabolic pathways and proteins with no metabolic pathway association.
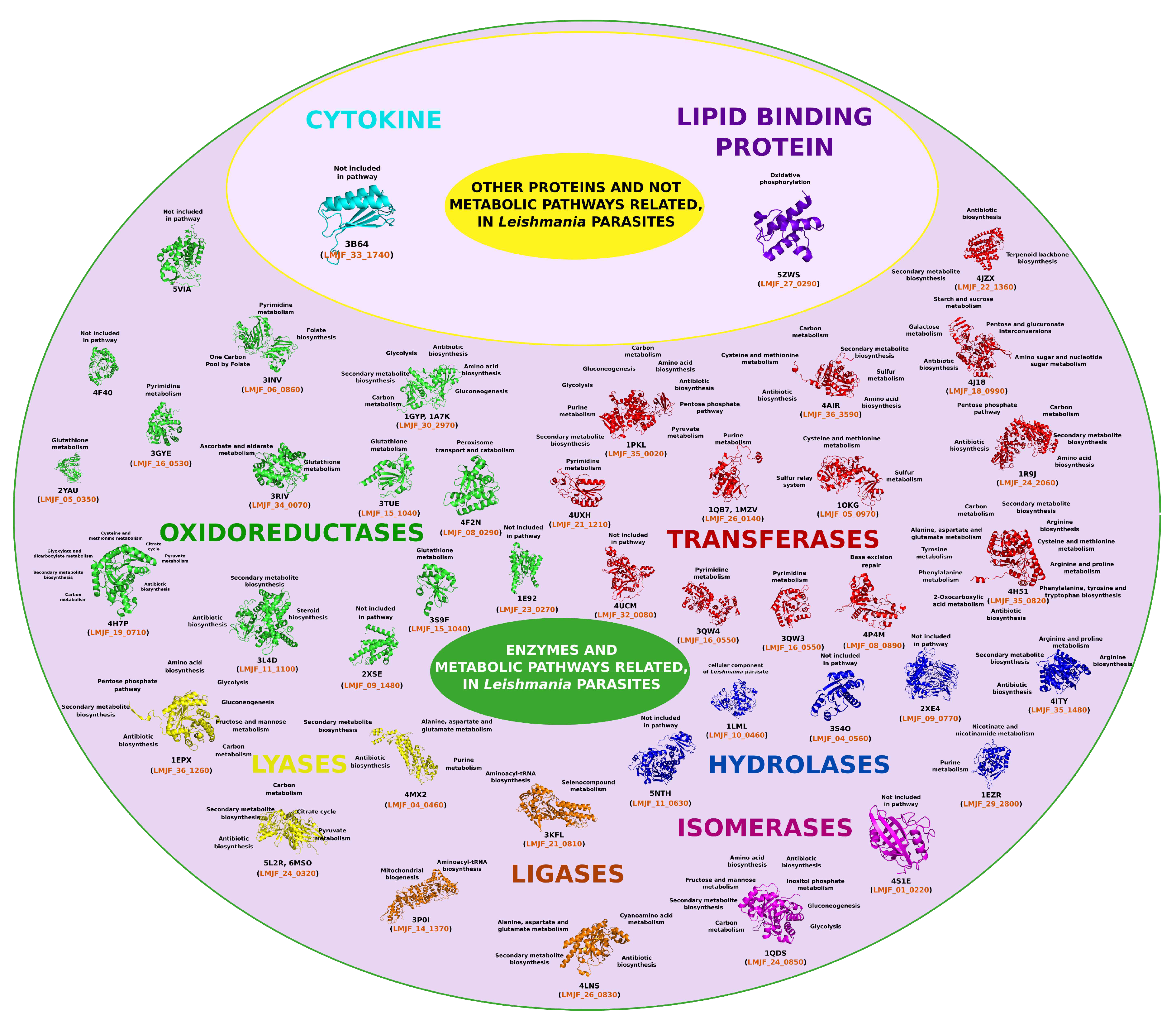
Figure 7.
Leishmania protein classification by EC group. Oxidoreductases (orange), transferases (blue), hydrolases (red), lyases (violet), isomerases (green), ligases (dark blue), cytokines (yellow) and Lipid Binding protein (light orange). The chart at the bottom right of the figure shows the number of proteins found for each species. T. cruzi and T. brucei do not belong at the Leishmania protein group, but they have been used in some studies as homologous proteins. [78,103]
Figure 7.
Leishmania protein classification by EC group. Oxidoreductases (orange), transferases (blue), hydrolases (red), lyases (violet), isomerases (green), ligases (dark blue), cytokines (yellow) and Lipid Binding protein (light orange). The chart at the bottom right of the figure shows the number of proteins found for each species. T. cruzi and T. brucei do not belong at the Leishmania protein group, but they have been used in some studies as homologous proteins. [78,103]
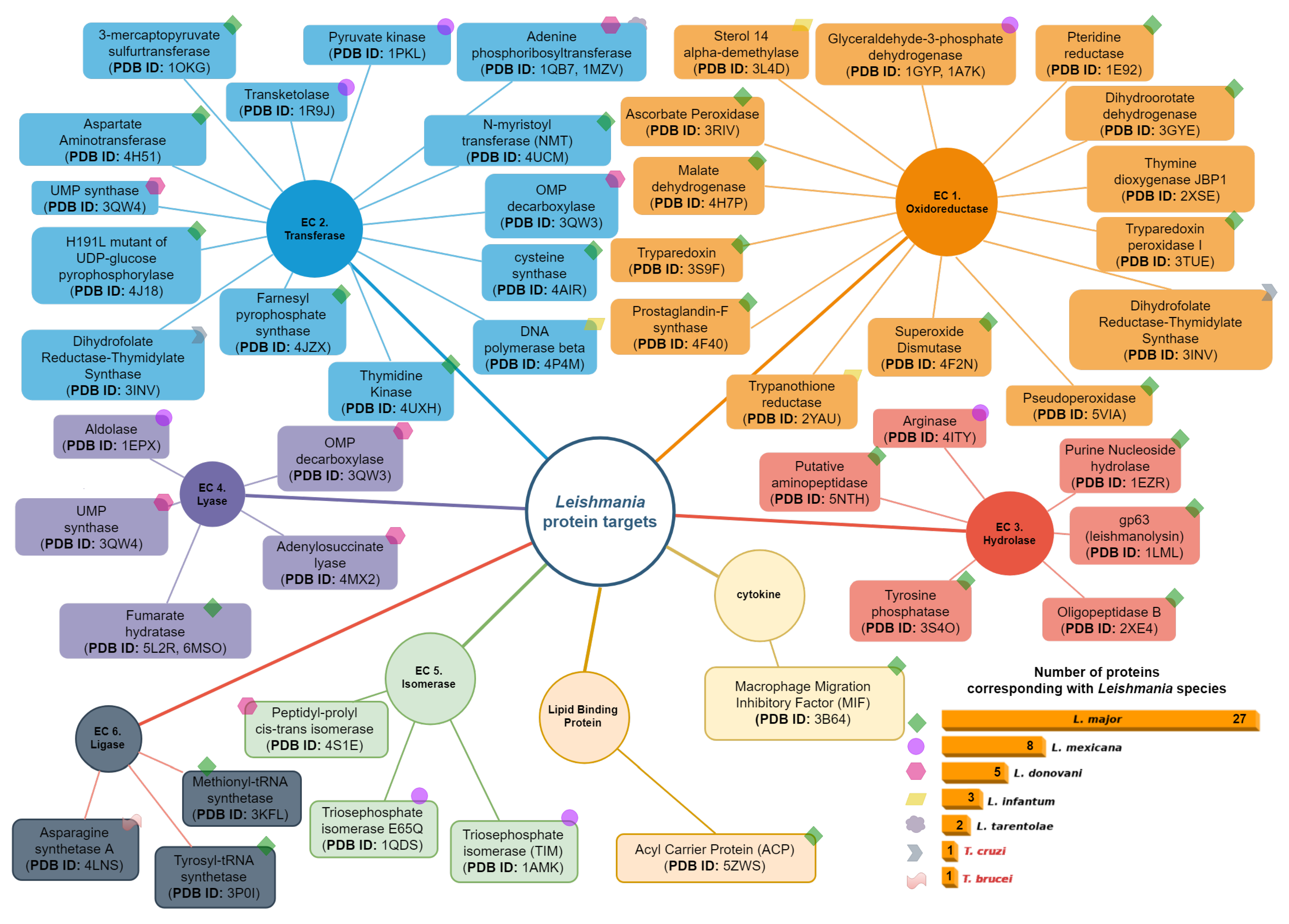
Figure 8.
Drugs used against Leishmania can be divided in two main groups: standard drugs and alternative drugs. Graphic shows molecular structures of drug active compounds. Structures were obtained from ChemSpider database (http://www.chemspider.com/).
Figure 8.
Drugs used against Leishmania can be divided in two main groups: standard drugs and alternative drugs. Graphic shows molecular structures of drug active compounds. Structures were obtained from ChemSpider database (http://www.chemspider.com/).
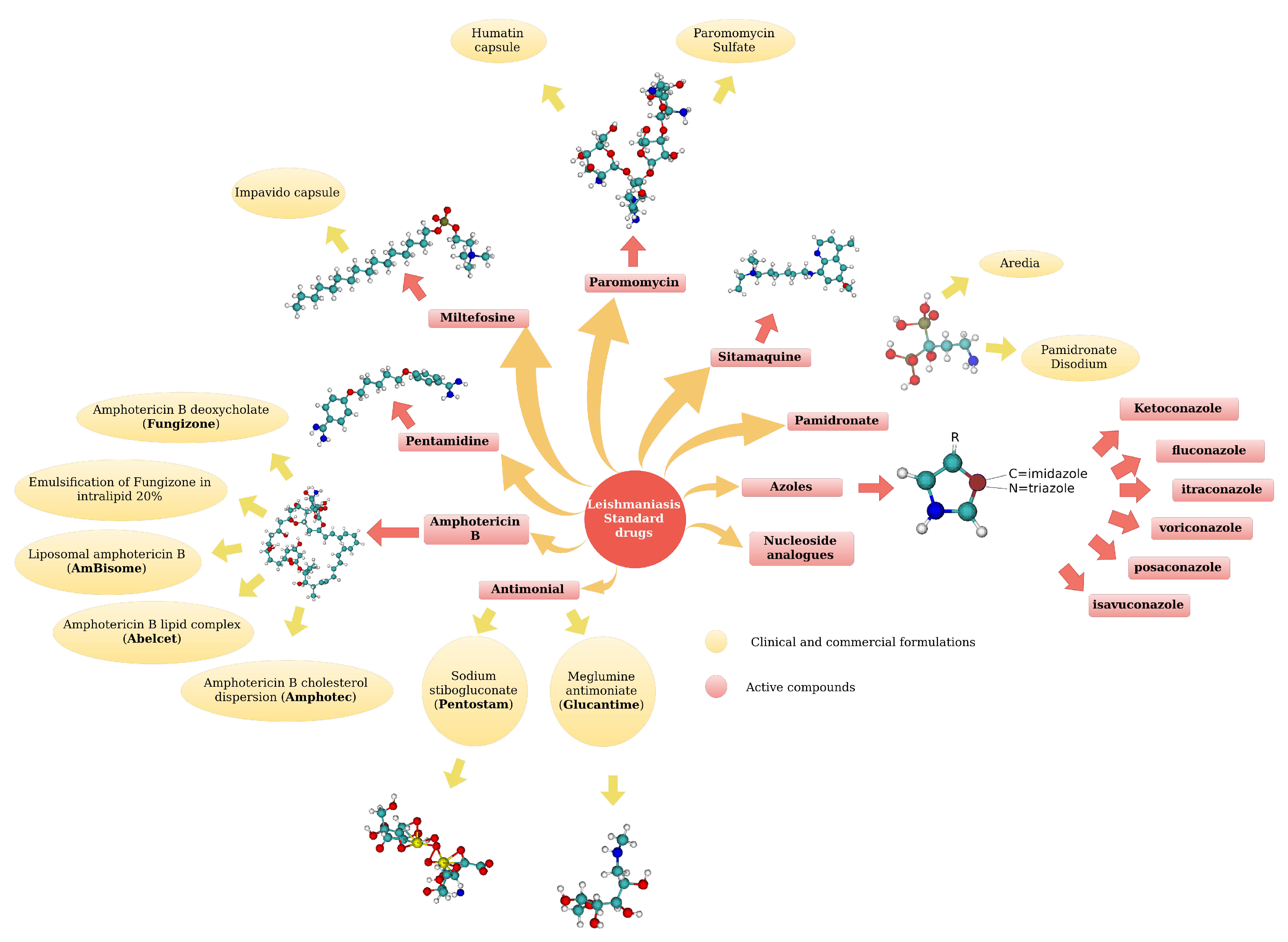
Figure 9.
Drug-database review. This figure shows useful information related to general databases and databases related with specific antileishmanial drugs
Figure 9.
Drug-database review. This figure shows useful information related to general databases and databases related with specific antileishmanial drugs
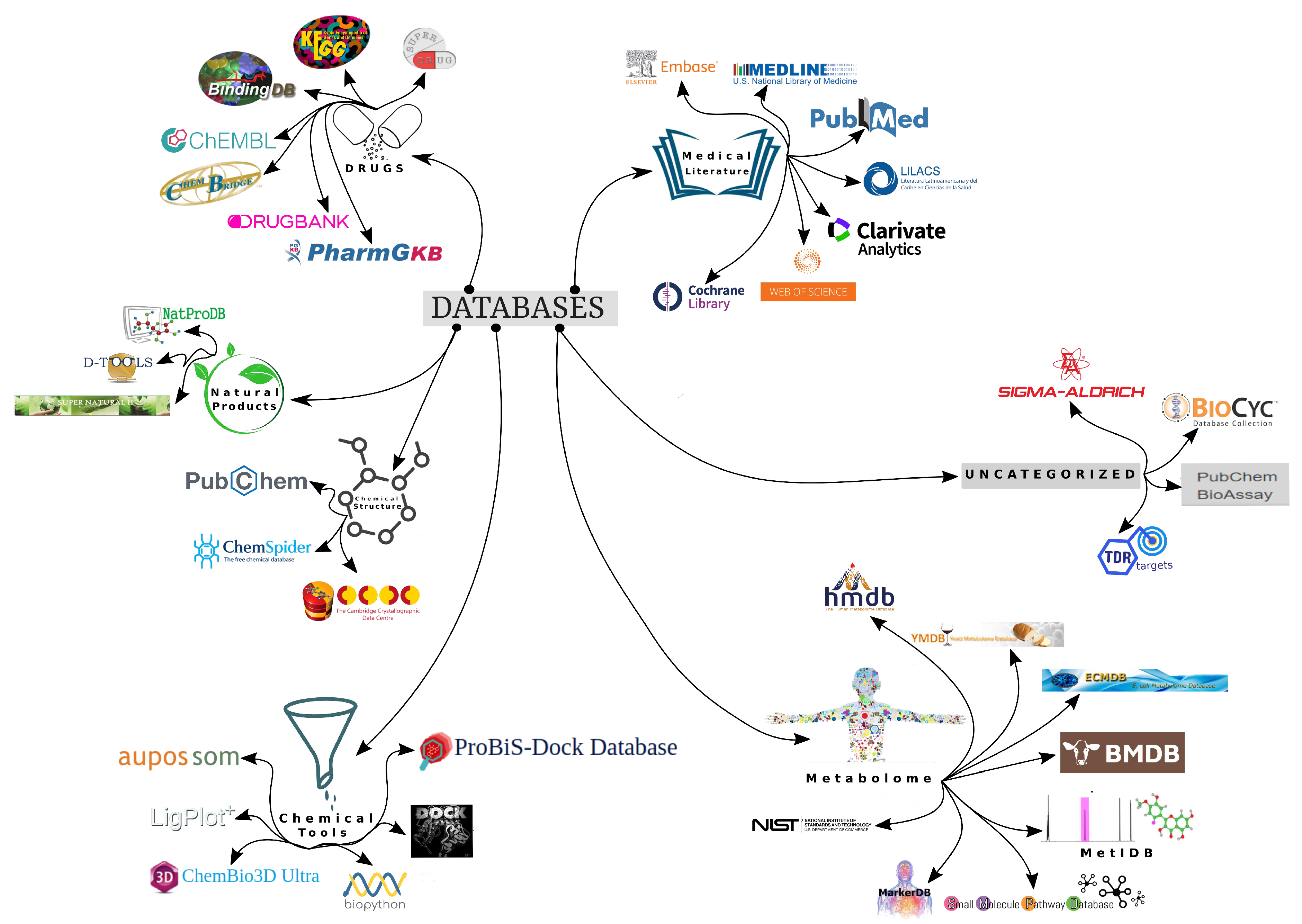
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



