Submitted:
30 June 2026
Posted:
02 July 2026
You are already at the latest version
Abstract
Induced pluripotent stem cells (iPSCs) have revolutionized the research landscape for Alzheimer's disease (AD) and related dementias by providing human, patient-specific neuronal models for studying the genetic and phenotypic diversity of these disorders. This review aims to integrate the latest advances in iPSC-based modeling of AD and dementia across multiple levels of biological organization. After describing the principles of pluripotency, iPSC generation, and quality control (genetic integrity, pluripotency assessment, long-term stability) the review will compare 2D monolayer culture with 3D organoid-based systems regarding their ability to model amyloid-beta, tau, and neuroinflammatory aspects of AD and dementia. The development of global stem cell biobanks and large iPSC collections with genetic diversity will be highlighted as an essential tool for dissecting the complex interplay of multiple genetic risk factors, ethnicity-based differences, and genotype-phenotype correlations. The review will discuss the use of genome editing technology, such as CRISPR/Cas9, for studying the effect of specific AD risk and protective variants in neurons and glial cells, and their implications for cell type-specific roles in neurodegeneration and neuroinflammation. We will highlight the challenges and limitations of iPSCs, such as cellular immaturity, loss of aging signatures, limited vascularization, and batch-to-batch differences, while noting the latest advances in partial/direct reprogramming, biomaterials science, and AI-based phenotypic analysis. Finally, the review will cover the expanding use of iPSC-based systems for high-content screening, functional therapeutic target validation, biomarker discovery, and development, and provide an overview of the future prospects for “clinical-trials-in-a-dish” and precision medicine for AD and other dementia disorders.
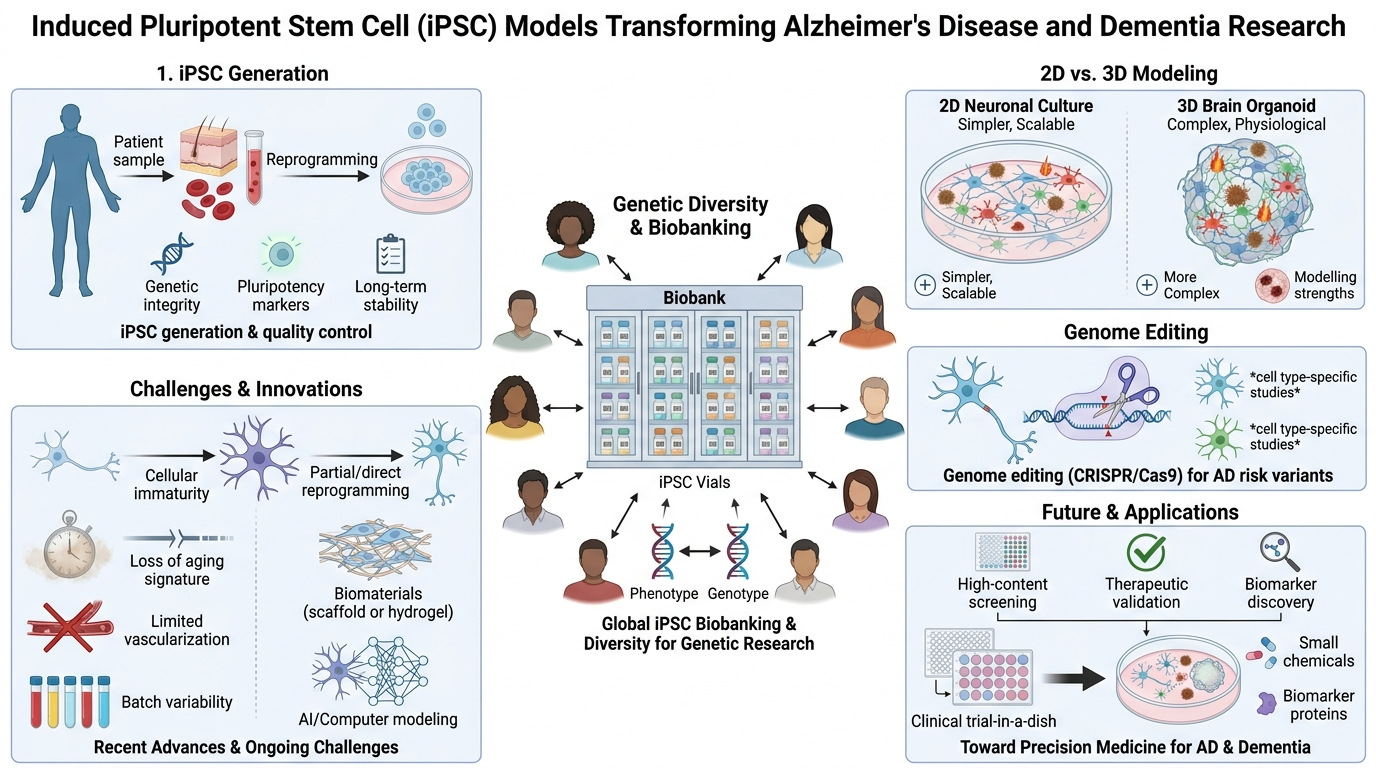
Keywords:
iPSC
; Alzheimer’s disease
; dementia
; brain
; neuroscience
; neurodegeneration
; neurons
1. Introduction
Alzheimer’s disease (AD) is the most prevalent form of dementia, which is characterized by cognitive decline, memory loss, and neuronal degeneration [1]. The neuropathological changes observed in AD are characterized by the presence of amyloid-beta (Aβ) plaques and neurofibrillary tangles consisting of hyperphosphorylated tau protein, and significant neuroinflammatory changes [2]. Despite decades of extensive research and significant investment in the development of therapeutic interventions, current pharmacological therapies are ineffective and provide symptomatic relief but do not arrest the progression of the disease [3,4,5]. This has become increasingly evident with the failure of promising preclinical animal model-based research and clinical trials [6]. This has brought into sharp focus the urgent need for more human-relevant cell-based disease models.
Recently, the development of induced pluripotent stem cell (iPSC) technology has revolutionized neuroscience research, providing unprecedented opportunities for disease modeling, mechanism-based research, and drug development [7]. Human iPSCs, derived from easily obtainable somatic cells such as human skin fibroblasts and peripheral blood mononuclear cells, possess the advantage of carrying the genetic and epigenetic memory of the patient from which they were derived, with the ability for infinite self-renewal and differentiation into any cell type desired [8]. This unique ability of iPSCs allows for the generation of patient-specific cellular models of AD, which can mimic the pathology of the disease while retaining the genetic heterogeneity of each patient, which is not seen with animal models.
Beyond the familial AD models, iPSC technology also allows the exploration of the entire spectrum of dementias, including sporadic AD, frontotemporal dementia, dementia with Lewy bodies, and vascular cognitive impairment and dementia, under the same human-relevant umbrella. By using iPSC cells from diverse patient populations and well-phenotyped clinical cohorts, scientists may begin to unravel the mysteries of how ethnicity, sex, polygenic risk scores, and comorbidities affect phenotypes. By using advanced differentiation methods and three-dimensional systems, iPSC models may recapitulate the intercellular communication among neurons, astrocytes, microglia, and oligodendrocytes to gain a more complete understanding of how AD pathology develops and propagates in the human brain [9]. Thus, iPSC models may be the key to bridging the gaps among molecular mechanisms, systems pathology, and clinical heterogeneity of the AD/dementia spectrum.
iPSC models may revolutionize the future of AD/dementia research by allowing ‘clinical trials in a dish,’ precise stratification of patients, and ultimately the development of individualized therapeutic regimens. As the iPSC biobanks are developed with the inclusion of single-cell multi-omics, spatial transcriptomics, high-content imaging, and phenotypic analysis using machine learning tools, it is conceivable to develop predictive models of individual diseases based on the ‘avatars’ of each patient. Additionally, the integration of iPSC-derived organoids, microfluidic organ-on-chip technologies, and genome editing tools may enable the testing of gene- and cell-based therapeutic approaches, combinatorial treatments, and network-based interventions in human neural cells before clinical trials. Thus, the iPSC models may be the key to understanding the fundamental mechanisms of AD/dementia in the future, but more importantly, the iPSC models may be the central decision engine to guide the diagnosis, drug development, biomarker identification, and individualized therapies of diverse patients worldwide.
2. Pluripotent Stem Cells: Foundation of iPSC Technology
2.1. Characteristics and Generation of PSCs
Pluripotent stem cells (PSC) can be categorized into two types: Embryonic stem cells (ESCs) and induced pluripotent stem cells (iPSCs) [10]. ESCs are naturally pluripotent, but the use of ESCs is often accompanied by ethical problems and the risk of immune rejection upon transplantation [11,12]. iPSCs, on the other hand, are produced through the reprogramming of differentiated somatic cells through the overexpression of specific transcription factors, which include Oct4, Sox2, Klf4, and c-Myc, also known as the Yamanaka factors [13]. This groundbreaking technology eliminates the ethical concern related to embryo destruction and also permits the derivation of patient-specific cells or even mass-produced hypoimmunogenic iPS cell lines [14], without the risk of immune rejection.
Several reprogramming techniques have been discovered, which include the use of viruses, small molecules, and non-integrative techniques, which include mRNA and protein reprogramming [15]. The type of reprogramming technique used is critical for the quality, efficacy, and safety of the stem cells produced, and the non-integrative approach is becoming the preferred choice for reprogramming. Modern iPSC derivation techniques have been successful in achieving high reprogramming efficiencies, up to 10-15%, and are free of the risk of genomic integration, making the technology ready for clinical application [16].
Even though nowadays culturing PSCs in feeder-free conditions [17] is a normal occurrence, PSCs can and used to be cultured on a feeder layer of fibroblasts. These feeder cells (often mouse embryonic fibroblasts (MEF) or human dermal fibroblasts (HDF)) provide PSCs with essential growth factors. Interestingly, feeder-free culture of pluripotent stem cells has been linked to increased acquisition of genomic alterations [18]. Despite this, it is usually seen that feeder-free cultures are advantageous due to the prospects of xeno-free, GMP-compliant iPSC culture [19].
The generation of pluripotent stem cell cultures is summarized in Figure 1.
2.2. Quality Control of iPSCs
Quality control of iPSCs is critical for their reliability and safety in disease modeling and therapeutic use [20], summarized in Figure 2. Quality control of iPSCs involves the following crucial aspects: (1) the presence of pluripotent markers such as OCT4, SOX2, NANOG, and SSEA4; (2) the capacity of the stem cells to differentiate along all three germ layers using spontaneous (or directed) differentiation assays; (3) karyotypic analysis of the stem cells to rule out any abnormalities; and (4) mycoplasma and viral clearance testing of the cells [21].
Recent advances in molecular biology have introduced techniques such as RNA sequencing and whole-genome sequencing for the comprehensive analysis of the pluripotent state of stem cells and the early detection of any abnormalities [22].
2.2.1. Genetic Integrity and Safety Testing
Genetic screening of iPSCs involves the evaluation of copy number variations, detection of reprogramming vector sequences, and spontaneous mutations that may arise during reprogramming or long culture periods [22]. Studies have shown that although most iPSC lines exhibit karyotypic stability, some lines exhibit recurrent copy number variations and single nucleotide variations, particularly in cell proliferation and differentiation-related genes [23].
2.2.2. Genetic Stability During Extended Culture
As the culture of iPSCs progresses with increasing passage numbers, the assessment of genetic stability becomes crucial. Recent investigations have indicated that the passage number is an important factor that can affect the genetic stability of the iPSCs, with the latter showing an increased tendency towards CNVs and spontaneous mutations compared to the former [22]. It is also known that higher-density culture of PSCs can lead to genomic instability [24]. Therefore, regular quality control tests, including cytogenetic tests and genome sequencing, are recommended to ensure the consistency and reliability of the iPSCs.
2.2.3. Undifferentiated State and Pluripotency
Though some may still assume that expression of “pluripotency genes” alone proves cell pluripotency, expression of genes like OCT4, NANOG etc. can only be evidence of an undifferentiated state. Indeed, some cells can have high levels of OCT4, NANOG etc. and still have impaired differentiation potential [25] or even be nullipotent [26]. Despite this, surveillance of the undifferentiated state is still an important aspect of PSC culture. This can be done by quantitative reverse transcription polymerase chain reaction (qRT-PCR), flow cytometry or immunohistochemistry [21].
The differentiation potential of PSCs can be evaluated using directed differentiation or spontaneous differentiation assays that test their capacity to form representatives of all three germ layers [27], utilizing the same methodologies as mentioned above.
3. iPSCs in Neuroscience: Monolayer Versus Organoid Culture Systems
3.1. Monolayer Culture Systems for Neural Differentiation
Monolayer cultures of iPSC-derived neurons have become the norm in neuroscience research due to their simplicity, relative ease of maintenance, and compatibility with high-content screening assays [28]. Directed differentiation protocols have been used to produce specific neuronal subtypes, such as glutamatergic, GABAergic, and dopaminergic neurons, with a reproducibility rate of over 70% [29]. The use of a transgene-based rapid induction approach, such as doxycycline-inducible NGN2, can produce mature, postmitotic neurons within 2-3 weeks [28,30,31].
Monolayer cultures have several significant advantages over other types of cultures, such as the ability to manipulate environmental conditions and analyze individual cell properties, such as electrophysiology and transcriptomics [32]. Most importantly, maturation protocols that include the right cocktail of neurotrophic factors, electrophysiological stimulation, and co-culture with glial cells have been shown to produce mature neurons that resemble those found in the brain [33].
3.2. Three-Dimensional Organoid Systems
Brain organoids mark a revolutionary step forward in the field of iPSC-based disease modeling, as they have the capability to form self-organized three-dimensional structures of the developing human brain regions, including the cellular composition, spatial organization, and functional connectivity of the brain [34,35,36,37]. Unlike two-dimensional culture systems, brain organoids have the capability to form different types of neural cells, including neurons, astrocytes, oligodendrocytes, and microglia (in the presence of added mesodermal progenitors), and study the effects of cell-cell interactions and glial contributions to pathology [38].
Cerebral organoids generated using iPSC have successfully mimicked the major AD-related pathological hallmarks, including Aβ deposits, tau hyperphosphorylation, endosomal dysfunction, and neuroinflammatory responses [39,40,41]. Organoids generated from APOE4/4 individuals, including AD patients, have also demonstrated the effects of APOE4 on AD-related pathology, as they showed increased neuronal apoptosis, compromised synaptic integrity, and impaired RNA metabolism compared to APOE3/3 organoids [42].
3.3. Comparative Advantages and Limitations
Monolayer systems have high reproducibility, scalability, and compatibility with automated high-throughput screening platforms and are therefore considered suitable for drug discovery assays for specific neuronal populations [32]. Monolayer systems, however, lack the 3D structure, cell type diversity, and cell-cell interactions present in the natural brain. Organoid systems have the advantage of better recapitulation of the complex structure of the brain and can be used to explore multi-cellular pathological mechanisms, but there are challenges associated with batch variability, incomplete neuronal maturation, limited vascularization, and the absence of a functional blood-brain barrier [35]. New assembloid technology, which combines region-specific organoids, has the potential to overcome the disadvantages associated with organoid technology and can generate more physiologically relevant neural circuits [38,43].
Figure 3 provides a schematic summary of this chapter.
4. Expanding Stem Cell Biobanks: Global Resources and Genetic Diversity
4.1. Stem Cell Biobanking Infrastructure
Large-scale stem cell biobanks have been recognized as essential resources in the study of neurodegenerative diseases, which can facilitate the study of the human population and the discovery of genetic modifiers in the pathogenesis of the diseases [44]. The National Centralized Repository for Alzheimer’s Disease and Related Dementias (NCRAD) is an example of the best practice in biobanking, which currently stores biological samples from over 135,000 well-characterized participants in 91 different studies, with stringent quality control measures in place to ensure the integrity of the biological materials [44]. Other biobanks, such as the European Bank of iPSCs (EBiSC), have developed standardized operating procedures for the generation, characterization, and distribution of iPSCs [45].
4.2. Genetic Diversity and Population Representation
A major limitation of current research into iPSCs is that there is significant underrepresentation of non-European populations, which could impact our understanding of population-specific disease mechanisms and therapeutics [46]. Recent initiatives have been undertaken for establishing African iPSC banks and characterizing iPSC lines from people of various ethnic backgrounds. This research has shown population-specific disease vulnerabilities and protective effects [47].
The IPMAR Resource represents an extensive effort in understanding genetic heterogeneity in AD by deriving iPSC lines from more than 100 donors with extremes of polygenic risk [48]. Such resources are useful for studying how genetic variations impact disease phenotypes and for finding therapeutics for genetically distinct populations with AD.
4.3. Quality Control and Standardization in Biobanks
Rigorous quality control procedures are critical for sustaining value in biobanks and ensuring reproducibility of results from different applications of biobank samples [27]. Standardized procedures for sample acquisition, processing, storage, and distribution reduce pre-analytical variability and maximize scientific output. The use of AI-driven quality control procedures also appears promising for optimizing efficiency and reproducibility of biospecimen assessment [20]. Centralized data management platforms and sample tracking can facilitate efficient collaborative research among different institutions and support multi-omics research.
5. Modeling the Full Complexity of AD Pathology: 3D and Multicellular Systems
5.1. Recapitulating Aβ and Tau Pathologies in 3D Systems
Three-dimensional systems generated from iPSCs have been shown to recapitulate simultaneously various aspects of AD pathology, making them more physiologically relevant models than conventional 2D cultures [49]. iPSC-derived cerebral organoids from AD patients with familial AD mutations, such as APP duplications or PSEN1/PSEN2 mutations, have been shown to spontaneously produce Aβ oligomers and amyloid plaques without the addition of exogenous Aβ, indicating that mutations alone are sufficient to recapitulate pathological features [40].
Notably, ferroptosis, a type of iron-dependent programmed cell death, has emerged as a critical mechanism underlying Aβ-induced neurodegeneration in iPSC-derived organoids from AD patients [50]. AD iPSC-derived organoids have been shown to exhibit increased lipid peroxidation, alterations in iron metabolism, and ferroptotic pathway activation, which can be rescued with ferroptosis inhibitors.
5.2. Modeling Neuronal and Glial Cell Interactions
The multicellular nature of 3D organoid systems also allows for the examination of the role of glial cells in the pathogenesis of AD. In a recent proteomics and transcriptomics study of AD organoids, several key protein dysregulation networks, including those related to the interaction of glial cells and neurons, have been identified, and several key driver proteins have been found to be part of these networks [2].
Microglial-derived organoid models have provided insights into the intrinsic dysfunction of microglial cells in tau-associated dementias, including altered cytoskeletal organization, phagocytic dysfunction, and TREM2 signaling dysregulation [51].
The integration of patient-derived iPSC-derived microglial cells into neural organoids has provided important insights into the role of microglial dysfunction in the pathogenesis of AD, including altered inflammatory and neuroprotective signaling pathways [52].
5.3. Modeling Mixed Pathologies and Comorbidities
In many cases, AD is associated with other pathology types, such as vascular dementia, and thus results in a mixed form of dementia. New organoid models that include neural and vascular cells have great potential for studying complex and clinically relevant disease states, such as mixed dementia, which is a combination of AD and vascular dementia [53]. New models of multiple organ assembloids that include neural, vascular, and immune cells are currently being developed to study AD pathology.
6. Gene Editing, Engineering, and Isogenic Controls
6.1. CRISPR/Cas9-Mediated Genome Editing
CRISPR/Cas9 technology has significantly advanced iPSC-based disease modeling as it allows precise introduction of disease-associated mutations or correction of disease-causing mutations within patient-derived cells [54]. Isogenic iPSC lines, where disease-causing mutations are introduced into or corrected within a particular genetic background, have also proved useful as experimental controls that can distinguish between mutation and genetic background effects [55,56].
In the case of AD, CRISPR-mediated genome editing has been used to introduce pathological mutations in APP and PSEN1 genes, to model protective mutations, and to correct disease-causing mutations [57]. The generation of isogenic lines with either the wild-type or the mutated allele of the genes of interest allowed the study of the mechanisms of AD caused by the mutations, showing that changes in excitability, in the handling of calcium, and in the metabolism occur only in the neurons with the pathological mutations [58].
6.2. Ablate-and-Replace Gene Editing Strategies
In addition, there have been new approaches in gene editing that extend beyond the simple introduction of mutations to include “ablate and replace” techniques in which the mutations that cause the disease are corrected and replaced with either wild-type sequences or protective variants [59]. These techniques can help in the assessment of whether protective variants can help in the alleviation of the disease phenotype.
6.3. Quality Control of Edited iPSCs
Comprehensive quality control of CRISPR-edited iPSCs is necessary to guarantee that phenotypes are due to intended, rather than off-target, mutations [60]. Genome sequencing has shown that, although off-target mutations can be caused by CRISPR/Cas9 gene editing, the major contributor to genetic variability is actually cell culture and cloning itself [60]. A well-designed study, with comparison of multiple clones and inclusion of unedited controls from the same cloning study, is necessary to distinguish between edits-specific and control effects.
7. Neuroinflammation and Glial Contributions in iPSC Models
7.1. iPSC-Derived Microglia and Their Role in AD
Microglia, being the key immune cells of the CNS, have established functions in AD pathogenesis [61]. Microglia have been derived from iPSCs through differentiation of iPSCs into mesodermal progenitors and subsequent specification into microglia or derivation from iPSC formed Embryoid Bodies (EB) [62,63,64]. These microglia have been shown to have functional characteristics similar to those of primary microglia, including phagocytic activity and morpho-dynamics [65].
In AD, iPSC-derived microglia have been used to study AD risk variants and have provided insight into the mechanism of action of genetic variants on microglia function and phenotype [63]. Microglia derived from AD-risk PLCG2 M28L carriers have been shown to have reduced TREM2 expression, reduced inflammation, and altered survival, indicating that these cells could be responsible for AD pathogenesis [66]. In contrast, AD-protective CR11 microglia have been shown to have enhanced phagocytic ability compared to CR12 carriers [65].
7.2. Astrocyte Contributions to AD Pathology
Astrocytes, the most numerous type of glial cell, are involved in modulating neuronal activity by providing trophic support, regulating synaptic activity, and immune responses [67]. As with other cell types, astrocytes have derived from iPSCs through a neural stem cell intermediary [68,69] and transcription factor overexpression [70]. iPSC-derived astrocytes have also been reported to be generated, with these cells showing evidence of glutamate uptake, cytokine release, and supporting neuronal maturation [71]. Recent studies have also indicated that AD patient-derived astrocytes show changes in their metabolic and transcriptomic profiles, including lipid metabolism and mitochondrial function [72].
7.3. Oligodendrocyte Dysfunction in AD
Although less researched than neurons and microglia in AD, oligodendrocytes are known for their contribution to the maintenance of axonal health [73] and thus may be involved in the disease. Differentiation protocols exist for iPSC differentiation to oligodendrocytes with [74,75] and without [76] transcription factor overexpression. A recent study using iPSC-derived oligodendrocytes found differences in gene expressions related to AD between ancestries, and carriers of APOE4/4 across ancestries were found to upregulate genes involved in cholesterol synthesis and downregulate myelination genes [47]. This study indicates a potential role for oligodendrocytes in AD pathology in APOE4 carriers.
8. iPSC Models in Personalized and Precision Medicine
8.1. Patient-Specific Disease Modeling
The ability of iPSCs to be derived from patients offers the prospect of developing personalized models of disease, which can be tailored to the specific genetic and epigenetic profile of the individual patient [77]. This can be particularly valuable in the study of the effect of individual genetic variants upon the manifestation of disease and drug response, which is a central tenet of the concept of precision medicine. iPSC models of familial AD have been developed and have been shown to replicate the specific phenotypes of the disease, including the overproduction of Aβ and abnormal tau protein metabolism, with responses to drug treatment varying depending upon the specific genetic background of the patient [78].
8.2. Genotype-Phenotype Relationships and Drug Responses
Systematic studies comparing iPSC-derived neurons from AD patients with different genetic backgrounds have demonstrated significant variability in AD phenotypes, emphasizing the need to use multiple patient-derived iPSC lines instead of relying on a single representative model for AD [8]. Such variability in iPSC lines is probably due to the complex genetics of sporadic AD, in which multiple genetic factors with small individual impact contribute to disease pathogenesis as well as drug responses.
Pharmacogenomic studies using iPSC models from large cohorts of genetically characterized donors have indicated that drug responses are genotype-specific, suggesting that pharmacogenomic strategies could be used to improve drug responses in AD patient populations with common genetic backgrounds [48].
8.3. Integration with Biomarker Development
The iPSC-derived cellular models have also helped in the identification and validation of peripheral biomarkers of AD, which can be used for the non-invasive monitoring of disease progression [79]. Extracellular vesicles (EVs) extracted from AD human brain samples were shown to contain disease state and cell type specific payloads [80]. Isolation and study of EVs from iPSC-derived neural cells could thus provide opportunities to elucidate biomarkers to be detected in patient blood, as it is known that EVs have the ability to cross the blood-brain barrier [81,82].
9. Addressing Aging and Epigenetic Memory in iPSC Reprogramming
9.1. Epigenetic Rejuvenation During Reprogramming
One of the most intriguing features of the reprogramming process mediated by iPSCs is the reversal of age-related epigenetic changes, which is indicated by the significant reduction in epigenetic age based on DNA methylation patterns [83]. Complete reprogramming to the pluripotent state resets epigenetic clocks to near-zero in cells, which can be considered to “erase” the chronological and biological aging signatures of the donor cells [84]. This epigenetic rejuvenation is also associated with the reversal of other hallmarks of aging, including telomere elongation [85] and reduction in oxidative stress [86,87], which can potentially allow the derivation of young neuron cell types from elderly patients.
9.2. Epigenetic Memory and Source Cell Lineage Bias
It is known that despite the described epigenetic ‘rejuvenation’, iPSCs can (and often do) possess epigenetic memory that is characteristic of the tissue of origin, affecting their differentiation tendencies and characteristics [88]. For example, iPSCs that are derived from pancreatic beta cells exhibit improved redifferentiation potential into insulin-producing cells compared to those derived from fibroblasts, possibly due to epigenetic programming that is carried over from the parent tissue [89]. Similarly, iPSCs that are derived from neural cells exhibit improved redifferentiation potential into neural cells compared to those derived from non-neural cells [90].
Epigenetic memory can be partially removed by serial reprogramming or using drugs that affect chromatin structure, but complete removal of epigenetic memory is difficult [91]. This has significant implications in disease modeling because of its potential to affect the iPSC-derived neural cells’ tendency to display disease characteristics that are uniquely present in the source tissue.
9.3. Strategies for Addressing Aging in iPSC-Based Models
The ‘rejuvenation’ aspect of the reprogramming process can be a double-edged sword. In terms of age-related disease models (including AD), reprogramming imposes an important limitation, since the cells derived from iPSCs can fail to properly mimic age-related disease mechanisms that require the presence of aging signatures [92], Figure 4. This aspect has led to the development of alternative reprogramming methods, including partial reprogramming and direct reprogramming [87,93,94,95,96].
Partial reprogramming utilizes transient expression of reprogramming factors to induce rejuvenation of the cell’s epigenetic state without the loss of cell identity [97]. This method holds promise for the study of the biology of aging and the generation of rejuvenated cells for regenerative medicine, although its potential for the modeling of age-related diseases awaits full evaluation [98].
Direct cellular reprogramming, which circumvents the pluripotent state and directly reprograms one cell type to another, holds promise for the maintenance of the hallmarks of aging and the generation of relevant cell types for the study of neurodegenerative diseases in which the biology of aging plays a central role in the pathogenesis of the condition [92,93,95].
10. Limitations and Innovations in Disease Modeling
10.1. Cellular Immaturity and Incomplete Recapitulation of Adult Brain Features
One of the major disadvantages of iPSC-derived neural cells is their incomplete maturation when compared to adult brain cells [99]. Even after extended periods of culture, the derived neuronal cells are fetal-like rather than adult-like, which may affect their use for modeling adult-onset disorders such as AD. Although the use of co-culture with astrocytes, extended periods of culture, and electrophysiological stimulations aids in the maturation of the cells, complete functional equivalence to adult cells is difficult to achieve [33].
10.2. Limited Vascularization and Blood-Brain Barrier Function
The lack of functional vascularization and blood-brain barrier in iPSC-derived organoids also hinders the study of vascular contributions to AD pathology, as well as predicting drug efficacy based on blood-brain barrier permeability [35]. New technologies that include endothelial cells in organoids are being developed, but fully functional vascular networks in these systems are difficult to achieve [100].
10.3. Batch-to-Batch Variability and Inter-Clonal Differences
Substantial heterogeneity is present between different iPSC cell lines from different individuals, as well as between different clones of cells from the same individuals, leading to variability that can mask disease-specific phenotypes [101]. This is due to several factors such as reprogramming efficiency, passage number effects, and culture conditions, thus underlining the need to use multiple cell lines in studies using iPSC cells.
10.4. Innovations Addressing Model Limitations
Recent innovations such as the microfluidic integration of iPSC technology, the use of automated culture systems, and the integration of artificial intelligence for phenotypic analysis hold promise for improving the reproducibility and efficiency of the technology [102].
Recent developments in biomaterials technology, such as the use of extracellular matrix and synthetic peptides, have improved the microenvironment of the brain and the functional properties of the neurons [103].
The integration of spatial transcriptomics and single-cell RNA sequencing with organoid technology has provided unprecedented insights into the mechanisms of disease through the high-resolution analysis of cell-type-specific gene expression changes [104].
11. Drug Screening, Functional Validation, and Clinical Translation
11.1. High-Throughput Drug Screening Platforms
iPSC-derived neural cells have also enabled the development of new high-throughput screening (HTS) strategies that offer better human relevance compared to traditional cell-line-based or animal models [105]. Multielectrode array recordings of iPSC-derived neuronal cultures allow for the real-time measurement of electrophysiological activity in networks, which can be used as functional readouts responsive to compound activity on neuronal excitability and connectivity [106]. Calcium imaging using genetically encoded indicators also offers additional functional readouts that are appropriate for high-throughput screening strategies [107].
Drug screening studies using patient iPSC-derived neurons have been able to identify compounds that fail to produce clinical trial results, validating the model, as well as novel therapeutic combinations with potential for clinical development [108]. These studies show that iPSC-derived models can better recapitulate aspects of human disease compared to traditional model systems.
11.2. Phenotypic Screening and Network-Level Assays
Phenotypic screening strategies that focus on measuring complex cellular or network functions, as opposed to particular molecular pathways, have also led to new therapeutic candidates for AD and other neurodegenerative diseases [109]. New innovations that have been made include brain region-specific organoid systems that have functional assays, which have enabled the development of a system that can be used for high-throughput screening while still maintaining multicellular complexity and architecture [110]. One-step drug screening systems that have microfluidic control of multiple organoids and have microelectrode arrays have enabled the screening of compound libraries in organoid models, closing the gap between 2D screening and in-vivo pharmacology. This system has been successful in identifying both novel and known AD therapeutics, including compounds that target tau phosphorylation, amyloid processing, and neuroinflammation [111,112].
11.3. Biomarker Discovery and Functional Validation
iPSC-derived models have played a significant role in the identification of novel AD biomarkers by systematically characterizing molecular signatures that are altered in the context of the disease. Small molecule screens in AD neurons have led to the identification of small molecules that target disease-relevant microRNAs, such as miR-132, which is implicated in AD tauopathies as a neuroprotective miRNA [113]. Such small molecule screens have provided useful resource datasets for drug discovery for various neurodegenerative diseases.
11.4. Translating iPSC Models Toward Clinical Application
In order to translate the iPSC-based cell therapies and discoveries into the clinic, several hurdles need to be overcome, such as large-scale GMP-compliant production of iPSCs, safety and long-term engraftment of the cells, and the establishment of standard protocols for quality control and regulatory procedures [102,114]. The first wave of iPSC-based cell therapies being tested in clinical trials for retinal diseases and other conditions serves as a valuable lesson for the development of translational strategies for various neurological conditions [102].
In spite of the hurdles and challenges associated with the development of iPSC-based cell therapies and discoveries, several promising strategies have shown significant promise for translation into the clinic. First, the regenerative capacity of iPSC-derived neural progenitor cells and neurons in preclinical models of various neurodegenerative conditions is a significant area of interest [10]. Second, cell-based therapies using iPSC-derived cells targeting neuroinflammation through the production of anti-inflammatory cytokines hold significant promise for slowing the progression of various neurodegenerative conditions [79]. Third, iPSC-based disease models have provided several novel therapeutic targets and drug candidates that could potentially benefit various cell-based and traditional drug-based cell therapies and treatments [115].
5. Conclusions
The combination of iPSC technology, advanced cell culture systems, gene editing technologies, and high-throughput screening technologies has revolutionized AD research, enabling the generation of human disease models with physiological relevance and genetic heterogeneity and complexity, which cannot be accomplished with conventional methods. iPSC-based models have successfully replicated many features of AD, such as Aβ deposition, tau pathology, neuroinflammation, and mitochondrial dysfunction, and have revealed cell type-specific and genotype-specific disease mechanisms.
The availability of diverse iPSC biobanks with well-characterized genetic heterogeneity and variability from global populations addresses historical issues with research representation bias and enables the discovery of disease mechanisms unique to different populations worldwide. Recent advances in three-dimensional organoid systems and multi-cellular assembloids have dramatically improved our ability to study complex cell-cell interactions within the human brain and have opened up new avenues for AD research. The use of gene editing technologies such as CRISPR/Cas9 enables the generation of specific mutation models, thus expanding our understanding of disease mechanisms.
Although there are still challenges to be addressed, such as difficulties achieving desired cell maturity, incomplete recapitulation of age-related signatures, incomplete/lack of vascularization, and batch-to-batch variability, recent advances in biomaterials, single-cell analysis, artificial intelligence, and organoid engineering are helping to address these issues. Recent success in iPSC-based models for identifying new therapeutic leads and biomarkers, coupled with initial clinical success with iPSC-derived cell therapies, underscores the potential for iPSC-based technologies to play an increasingly important role in AD drug discovery, patient stratification, and therapeutic development in the near future.
It is thus essential to continue to focus on quality control standards, standardization among research institutions, and inclusion of diverse populations to maximize the potential for iPSC technology to combat the global AD pandemic and extend this technology to the broader range of age-related neurodegenerative diseases and dementias affecting millions of people worldwide.
Author Contributions
Writing – Original Draft Preparation, Review and Editing were carried out by D.S. and J.S. Visualizations were created by D.S. Overall supervision J.S.
Funding
This review received no external funding.
Acknowledgments
The authors have re-viewed and edited the output and take full responsibility for the content of this publication. We thank the Department of Molecular Biology, Cell Biology and Biochemistry of Brown university and all the affiliated organizations for continued support.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
A: Amyloid-beta., AD: Alzheimer’s disease., AI: Artificial intelligence., APOE4: Apolipoprotein E4. cGMP: Current Good Manufacturing Practice., CNS: Central nervous system., CRISPR-Cas9 / CRISPRCas9: Clustered regularly interspaced short palindromic repeats-associated protein 9. EB: Embryoid body / embryoid bodies., ESC: Embryonic stem cell., ESCs: Embryonic stem cells. GMP: Good Manufacturing Practice. HDF: Human dermal fibroblasts., hiPSC: Human induced pluripotent stem cell., hiPSCs: Human induced pluripotent stem cells., HTS: High-throughput screening. iPSC: Induced pluripotent stem cell., iPSCs: Induced pluripotent stem cells. Klf4: Kruppel-like factor 4. LINE-1: Long interspersed nuclear element-1. MEF: Mouse embryonic fibroblast(s)., mRNA: Messenger RNA. NANOG: NANOG homeobox., NGN2: Neurogenin 2. Oct4: Octamer-binding transcription factor 4., OCT4: Octamer-binding transcription factor 4., ORF1p: Open reading frame 1 protein. PBMCs: Peripheral blood mononuclear cells., PSC: Pluripotent stem cell., PSCs: Pluripotent stem cells. RNA: Ribonucleic acid., RNA-seq: RNA sequencing., SSEA4: Stage-specific embryonic antigen-4., SOX2: SRY-box transcription factor 2. TREM2: Triggering receptor expressed on myeloid cells 2. 2D: Two-dimensional., 3D: Three-dimensional.
References
- Ifediora, N.; Canoll, P.; Hargus, G. Human Stem Cell Transplantation Models of Alzheimer’s Disease. Front. Aging Neurosci. 2024, 16, 1354164. [CrossRef]
- Wang, E.; Yu, K.; Cao, J.; Wang, M.; Katsel, P.; Song, W.; Wang, Z.; Li, Y.; Wang, X.; Wang, Q.; et al. Multiscale Proteomic Modeling Reveals Protein Networks Driving Alzheimer’s Disease Pathogenesis. Cell 2025, 188, 6186-6204.e13. [CrossRef]
- Fan, X.; Sun, D.; Tang, X.; Cai, Y.; Yin, Z.Q.; Xu, H. Stem-Cell Challenges in the Treatment of Alzheimer’s Disease: A Long Way from Bench to Bedside. Med. Res. Rev. 2014, 34, 957–978. [CrossRef]
- Kruk-Słomka, M.; Kuceł, D.; Małysz, M.; Machnikowska, A.; Orzelska-Górka, J.; Biała, G. New Approaches to the Treatment of Alzheimer’s Disease. Pharmaceuticals 2025, 18, 1117. [CrossRef]
- Passeri, E.; Elkhoury, K.; Morsink, M.; Broersen, K.; Linder, M.; Tamayol, A.; Malaplate, C.; Yen, F.T.; Arab-Tehrany, E. Alzheimer’s Disease: Treatment Strategies and Their Limitations. Int. J. Mol. Sci. 2022, 23, 13954. [CrossRef]
- Marshall, L.J.; Bailey, J.; Cassotta, M.; Herrmann, K.; Pistollato, F. Poor Translatability of Biomedical Research Using Animals — A Narrative Review. Altern. Lab. Anim. 2023, 51, 102–135. [CrossRef]
- Penney, J.; Ralvenius, W.T.; Tsai, L.-H. Modeling Alzheimer’s Disease with iPSC-Derived Brain Cells. Mol. Psychiatry 2020, 25, 148–167. [CrossRef]
- Barak, M.; Fedorova, V.; Pospisilova, V.; Raska, J.; Vochyanova, S.; Sedmik, J.; Hribkova, H.; Klimova, H.; Vanova, T.; Bohaciakova, D. Human iPSC-Derived Neural Models for Studying Alzheimer’s Disease: From Neural Stem Cells to Cerebral Organoids. Stem Cell Rev. Rep. 2022, 18, 792–820. [CrossRef]
- Cerneckis, J.; Cai, H.; Shi, Y. Induced Pluripotent Stem Cells (iPSCs): Molecular Mechanisms of Induction and Applications. Signal Transduct. Target. Ther. 2024, 9, 112. [CrossRef]
- Verma, I.; Seshagiri, P.B. Current Applications of Human Pluripotent Stem Cells in Neuroscience Research and Cell Transplantation Therapy for Neurological Disorders. Stem Cell Rev. Rep. 2025, 21, 964–987. [CrossRef]
- Rong, Z.; Wang, M.; Hu, Z.; Stradner, M.; Zhu, S.; Kong, H.; Yi, H.; Goldrath, A.; Yang, Y.-G.; Xu, Y.; et al. An Effective Approach to Prevent Immune Rejection of Human ESC-Derived Allografts. Cell Stem Cell 2014, 14, 121–130. [CrossRef]
- Volarevic, V.; Markovic, B.S.; Gazdic, M.; Volarevic, A.; Jovicic, N.; Arsenijevic, N.; Armstrong, L.; Djonov, V.; Lako, M.; Stojkovic, M. Ethical and Safety Issues of Stem Cell-Based Therapy. Int. J. Med. Sci. 2018, 15, 36–45. [CrossRef]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of Pluripotent Stem Cells from Adult Human Fibroblasts by Defined Factors. Cell 2007, 131, 861–872. [CrossRef]
- Deuse, T.; Hu, X.; Gravina, A.; Wang, D.; Tediashvili, G.; De, C.; Thayer, W.O.; Wahl, A.; Garcia, J.V.; Reichenspurner, H.; et al. Hypoimmunogenic Derivatives of Induced Pluripotent Stem Cells Evade Immune Rejection in Fully Immunocompetent Allogeneic Recipients. Nat. Biotechnol. 2019, 37, 252–258. [CrossRef]
- Amponsah, A.E.; Guo, R.; Kong, D.; Feng, B.; He, J.; Zhang, W.; Liu, X.; Du, X.; Ma, Z.; Liu, B.; et al. Patient-Derived iPSCs, a Reliable in Vitro Model for the Investigation of Alzheimer’s Disease. Rev. Neurosci. 2021, 32, 379–402. [CrossRef]
- Ross, C.A.; Akimov, S.S. Human-Induced Pluripotent Stem Cells: Potential for Neurodegenerative Diseases. Hum. Mol. Genet. 2014, 23, R17-26. [CrossRef]
- Nakagawa, M.; Taniguchi, Y.; Senda, S.; Takizawa, N.; Ichisaka, T.; Asano, K.; Morizane, A.; Doi, D.; Takahashi, J.; Nishizawa, M.; et al. A Novel Efficient Feeder-Free Culture System for the Derivation of Human Induced Pluripotent Stem Cells. Sci. Rep. 2014, 4, 3594. [CrossRef]
- Stavish, D.; Price, C.J.; Gelezauskaite, G.; Alsehli, H.; Leonhard, K.A.; Taapken, S.M.; McIntire, E.M.; Laing, O.; James, B.M.; Riley, J.J.; et al. Feeder-Free Culture of Human Pluripotent Stem Cells Drives MDM4-Mediated Gain of Chromosome 1q. Stem Cell Rep. 2024, 19, 1217–1232. [CrossRef]
- Rivera, T.; Zhao, Y.; Ni, Y.; Wang, J. Human-Induced Pluripotent Stem Cell Culture Methods Under cGMP Conditions. Curr. Protoc. Stem Cell Biol. 2020, 54, e117. [CrossRef]
- Gramatiuk, S.; Kryvoruchko, I.A.; Ivanova, Y.V.; Hubbard, E.; Noebauer-Babenko, M.; Sargsyan, K. Artificial Intelligence-Based Quality Control of Cell Lines. Biopreservation Biobanking 2025. [CrossRef]
- Shibamiya, A.; Schulze, E.; Krauß, D.; Augustin, C.; Reinsch, M.; Schulze, M.L.; Steuck, S.; Mearini, G.; Mannhardt, I.; Schulze, T.; et al. Cell Banking of hiPSCs: A Practical Guide to Cryopreservation and Quality Control in Basic Research. Curr. Protoc. Stem Cell Biol. 2020, 55, e127. [CrossRef]
- Jo, H.-Y.; Han, H.-W.; Jung, I.; Ju, J.H.; Park, S.-J.; Moon, S.; Geum, D.; Kim, H.; Park, H.-J.; Kim, S.; et al. Development of Genetic Quality Tests for Good Manufacturing Practice-Compliant Induced Pluripotent Stem Cells and Their Derivatives. Sci. Rep. 2020, 10, 3939. [CrossRef]
- Melnikova, E.V.; Rachinskaya, O.A.; Semenova, I.S.; Merkulov, V.A. Quality control programmes for induced pluripotent stem cell-derived medicinal products. Biol. Prod. Prev. Diagn. Treat. 2025, 25, 127–140. [CrossRef]
- Jacobs, K.; Zambelli, F.; Mertzanidou, A.; Smolders, I.; Geens, M.; Nguyen, H.T.; Barbé, L.; Sermon, K.; Spits, C. Higher-Density Culture in Human Embryonic Stem Cells Results in DNA Damage and Genome Instability. Stem Cell Rep. 2016, 6, 330–341. [CrossRef]
- Zhang, M.; Cheng, L.; Jia, Y.; Liu, G.; Li, C.; Song, S.; Bradley, A.; Huang, Y. Aneuploid Embryonic Stem Cells Exhibit Impaired Differentiation and Increased Neoplastic Potential. EMBO J. 2016, 35, 2285–2300. [CrossRef]
- Rosenstraus, M.J.; Spadoro, J.P. Autonomy of “Nullipotent” and Pluripotent Embryonal Carcinoma Cells in Differentiating Aggregates. Dev. Biol. 1981, 85, 190–194. [CrossRef]
- Selfa Aspiroz, L.; Mennecozzi, M.; Batlle, L.; Corneo, B.; Healy, L.; Kotter, M.; Kurtz, A.; Ludwig, T.E.; Mummery, C.; Pera, M.; et al. Promoting the Adoption of Best Practices and Standards to Enhance Quality and Reproducibility of Stem Cell Research. Stem Cell Rep. 2025, 20, 102531. [CrossRef]
- Rizalar, F.S.; Haucke, V. Generation of Glutamatergic Human Neurons from Induced Pluripotent Stem Cells. Methods Mol. Biol. 2025, 2910, 27–36. [CrossRef]
- Wang, Y.; Wang, Z.; Wang, L.; Sun, Y.; Song, H.; Cheng, X.; He, X.; Gao, Z.; Sun, Y. Human Induced Pluripotent Stem Cells: Directed Differentiation Methods and Applications in Brain Diseases. J. Neurosci. Res. 2025, 103, e70027. [CrossRef]
- Lin, H.-C.; He, Z.; Ebert, S.; Schörnig, M.; Santel, M.; Nikolova, M.T.; Weigert, A.; Hevers, W.; Kasri, N.N.; Taverna, E.; et al. NGN2 Induces Diverse Neuron Types from Human Pluripotency. Stem Cell Rep. 2021, 16, 2118–2127. [CrossRef]
- Shan, X.; Zhang, A.; Rezzonico, M.G.; Tsai, M.-C.; Sanchez-Priego, C.; Zhang, Y.; Chen, M.B.; Choi, M.; Andrade López, J.M.; Phu, L.; et al. Fully Defined NGN2 Neuron Protocol Reveals Diverse Signatures of Neuronal Maturation. Cell Rep. Methods 2024, 4, 100858. [CrossRef]
- Yang, Z.; Teaney, N.A.; Buttermore, E.D.; Sahin, M.; Afshar-Saber, W. Harnessing the Potential of Human Induced Pluripotent Stem Cells, Functional Assays and Machine Learning for Neurodevelopmental Disorders. Front. Neurosci. 2024, 18, 1524577. [CrossRef]
- Thaploo, S.; Lin, D.; Wang, P.; Appajodu, A.; Thurgam, P.; Baig, A.; Vargas, L.; Li, Y.; Brewer, G.; Nenadić, Z.; et al. Long-Term Maintenance of Human Stem Cell-Derived Neural Networks for Electrophysiology Studies 2025.
- Andersen, J.; Revah, O.; Miura, Y.; Thom, N.; Amin, N.D.; Kelley, K.W.; Singh, M.; Chen, X.; Thete, M.V.; Walczak, E.M.; et al. Generation of Functional Human 3D Cortico-Motor Assembloids. Cell 2020, 183, 1913-1929.e26. [CrossRef]
- Chacón, P.E.A.; Irineo-Moreno, V.; Loera-Valencia, R. Three-Dimensional Cell Culture Technologies in the Study of Alzheimer’s Disease: A Narrative Review. Adv. Technol. Neurosci. 2025, 2, 85. [CrossRef]
- Gordon, A.; Yoon, S.-J.; Tran, S.S.; Makinson, C.D.; Park, J.Y.; Andersen, J.; Valencia, A.M.; Horvath, S.; Xiao, X.; Huguenard, J.R.; et al. Long-Term Maturation of Human Cortical Organoids Matches Key Early Postnatal Transitions. Nat. Neurosci. 2021, 24, 331–342. [CrossRef]
- Onesto, M.M.; Amin, N.D.; Pan, C.; Chen, X.; Kim, J.-I.; Reis, N.; Kanton, S.; Valencia, A.M.; Hudacova, Z.; McQueen, J.P.; et al. Midline Assembloids Reveal Regulators of Human Axon Guidance. Science 2025, 389, 282–289. [CrossRef]
- Ajongbolo, A.O.; Langhans, S.A. Brain Organoids and Assembloids—From Disease Modeling to Drug Discovery. Cells 2025, 14, 842. [CrossRef]
- Chen, X.; Sun, G.; Tian, E.; Zhang, M.; Davtyan, H.; Beach, T.G.; Reiman, E.M.; Blurton-Jones, M.; Holtzman, D.M.; Shi, Y. Modeling Sporadic Alzheimer’s Disease in Human Brain Organoids under Serum Exposure. Adv. Sci. 2021, 8, e2101462. [CrossRef]
- Raja, W.K.; Mungenast, A.E.; Lin, Y.-T.; Ko, T.; Abdurrob, F.; Seo, J.; Tsai, L.-H. Self-Organizing 3D Human Neural Tissue Derived from Induced Pluripotent Stem Cells Recapitulate Alzheimer’s Disease Phenotypes. PLOS ONE 2016, 11, e0161969. [CrossRef]
- Zhang, Y.; Tang, C.; He, Y.; Zhang, Y.; Li, Q.; Zhang, T.; Zhao, B.; Tong, A.; Zhong, Q.; Zhong, Z. Semaglutide Ameliorates Alzheimer’s Disease and Restores Oxytocin in APP/PS1 Mice and Human Brain Organoid Models. Biomed. Pharmacother. Biomedecine Pharmacother. 2024, 180, 117540. [CrossRef]
- Zhao, J.; Fu, Y.; Yamazaki, Y.; Ren, Y.; Davis, M.D.; Liu, C.-C.; Lu, W.; Wang, X.; Chen, K.; Cherukuri, Y.; et al. APOE4 Exacerbates Synapse Loss and Neurodegeneration in Alzheimer’s Disease Patient iPSC-Derived Cerebral Organoids. Nat. Commun. 2020, 11, 5540. [CrossRef]
- Kim, J.; Imaizumi, K.; Jurjuț, O.; Kelley, K.W.; Wang, D.; Thete, M.V.; Hudacova, Z.; Amin, N.D.; Levy, R.J.; Scherrer, G.; et al. Human Assembloid Model of the Ascending Neural Sensory Pathway. Nature 2025, 642, 143–153. [CrossRef]
- Edler, M.C.; Faber, K.; Lacy, K.; Jackson, J.; Foroud, T. NCRAD: Advancing Alzheimer’s Research through High-Quality Biospecimens and Data. Alzheimers Dement. J. Alzheimers Assoc. 2025, 21, e70682. [CrossRef]
- O’Shea, O.; Steeg, R.; Chapman, C.; Mackintosh, P.; Stacey, G.N. Development and Implementation of Large-Scale Quality Control for the European Bank for Induced Pluripotent Stem Cells. Stem Cell Res. 2020, 45, 101773. [CrossRef]
- Maina, M.B.; Isah, M.B.; Marsh, J.A.; Muhammad, Z.; Babazau, L.; Idris, A.A.; Aladyeva, E.; Miller, N.; Starr, E.; Miller, K.J.; et al. Somatic and Stem Cell Bank to Study the Contribution of African Ancestry to Dementia: African iPSC Initiative. Alzheimers Dement. J. Alzheimers Assoc. 2025, 21, e70145. [CrossRef]
- Ramirez, A.M.; Nasciben, L.B.; Moura, S.; Coombs, L.; Rajabli, F.; DeRosa, B.A.; Whitehead, P.L.; Adams, L.D.; Starks, T.D.; Mena, P.R.; et al. Ancestral Genomic Functional Differences in Oligodendroglia: Implications for Alzheimer’s Disease. Alzheimers Dement. 2025, 21, e70593. [CrossRef]
- Maguire, E.; Winston, J.; Ellwood, S.H.; O’Donoghue, R.; Shaw, B.; Morales, A.C.; Keat, S.; Evans, A.; Marshall, R.; Luckcuck, L.; et al. Modeling Common Alzheimer’s Disease with High and Low Polygenic Risk in Human iPSC: A Large-Scale Research Resource. Stem Cell Rep. 2025, 20, 102570. [CrossRef]
- Park, J.-C.; Jang, S.-Y.; Lee, D.; Lee, J.; Kang, U.; Chang, H.; Kim, H.J.; Han, S.-H.; Seo, J.; Choi, M.; et al. A Logical Network-Based Drug-Screening Platform for Alzheimer’s Disease Representing Pathological Features of Human Brain Organoids. Nat. Commun. 2021, 12, 280. [CrossRef]
- Majerníková, N.; Marmolejo-Garza, A.; Salinas, C.S.; Luu, M.D.A.; Zhang, Y.; Trombetta-Lima, M.; Tomin, T.; Birner-Gruenberger, R.; Lehtonen, Š.; Koistinaho, J.; et al. The Link between Amyloid β and Ferroptosis Pathway in Alzheimer’s Disease Progression. Cell Death Dis. 2024, 15, 782. [CrossRef]
- Iyer, A.K.; Vermunt, L.; Mirfakhar, F.S.; Minaya, M.; Acquarone, M.; Koppisetti, R.K.; Renganathan, A.; You, S.-F.; Danhash, E.P.; Schache, K.J.; et al. Cell Autonomous Microglia Defects in a Stem Cell Model of Frontotemporal Dementia Tau. Mol. Psychiatry 2025, 30, 4776–4791. [CrossRef]
- Zhang, J.; Lim, Y.W.; Medina, A.; Carrasco Carvajal, M.; Lee, E.; Ferrer, M. Human iPSC-Derived Microglia-Containing 3D Brain Region-Specific Neural Spheroids for Disease Modeling and Therapeutic Development 2118. J. Immunol. 2025, 214, vkaf283.082. [CrossRef]
- Sarhan, M.; Wohlfeld, C.; Perry-Mills, A.; Meyers, J.; Fadel, J.; Murphy, E.A.; Bonilha, L.; Fan, D. The Pathophysiology of Mixed Alzheimer’s Disease and Vascular Dementia. Theranostics 2025, 15, 9793–9818. [CrossRef]
- Khan, M.S.; Qureshi, N.; Khan, R.; Son, Y.-O.; Maqbool, T. CRISPR/Cas9-Based Therapeutics as a Promising Strategy for Management of Alzheimer’s Disease: Progress and Prospects. Front. Cell. Neurosci. 2025, 19. [CrossRef]
- Pantazis, C.B.; Yang, A.; Lara, E.; McDonough, J.A.; Blauwendraat, C.; Peng, L.; Oguro, H.; Kanaujiya, J.; Zou, J.; Sebesta, D.; et al. A Reference Human Induced Pluripotent Stem Cell Line for Large-Scale Collaborative Studies. Cell Stem Cell 2022, 29, 1685-1702.e22. [CrossRef]
- Rehbach, K.; Fernando, M.B.; Brennand, K.J. Integrating CRISPR Engineering and hiPSC-Derived 2D Disease Modeling Systems. J. Neurosci. Off. J. Soc. Neurosci. 2020, 40, 1176–1185. [CrossRef]
- He, C.; Chen, B.; Yan, C.; Zhou, X. Stem Cell and CRISPR/Cas9 Gene Editing Technology in Alzheimer’s Disease Therapy: From Basic Research to Clinical Innovation. Front. Genome Ed. 2025, 7. [CrossRef]
- Maksour, S.; Finol-Urdaneta, R.K.; Hulme, A.J.; Cabral-da-Silva, M.E.C.; Targa Dias Anastacio, H.; Balez, R.; Berg, T.; Turner, C.; Sanz Muñoz, S.; Engel, M.; et al. Alzheimer’s Disease Induced Neurons Bearing PSEN1 Mutations Exhibit Reduced Excitability. Front. Cell. Neurosci. 2024, 18, 1406970. [CrossRef]
- Rybarikova, M.; Rey, M.; Hasanovic, E.; Sipion, M.; Rambousek, L.; Déglon, N. Gene Editing for Spinocerebellar Ataxia Type 3 Taking Advantage of the Human ATXN3L Paralog as Replacement Gene. Gene Ther. 2025, 32, 462–474. [CrossRef]
- Busquets, O.; Li, H.; Syed, K.M.; Jerez, P.A.; Dunnack, J.; Bu, R.L.; Verma, Y.; Pangilinan, G.R.; Martin, A.; Straub, J.; et al. iSCORE-PD: An Isogenic Stem Cell Collection to Research Parkinson’s Disease. BioRxiv Prepr. Serv. Biol. 2025, 2024.02.12.579917. [CrossRef]
- Monzón-Sandoval, J.; Burlacu, E.; Agarwal, D.; Handel, A.E.; Wei, L.; Davis, J.; Cowley, S.A.; Cader, M.Z.; Webber, C. Lipopolysaccharide Distinctively Alters Human Microglia Transcriptomes to Resemble Microglia from Alzheimer’s Disease Mouse Models. Dis. Model. Mech. 2022, 15, dmm049349. [CrossRef]
- Hasselmann, J.; Blurton-Jones, M. Human iPSC-Derived Microglia: A Growing Toolset to Study the Brain’s Innate Immune Cells. Glia 2020, 68, 721–739. [CrossRef]
- Roy, N.; Haq, I.; Ngo, J.C.; Bennett, D.A.; Teich, A.F.; De Jager, P.L.; Olah, M.; Sher, F. Elevated Expression of the Retrotransposon LINE-1 Drives Alzheimer’s Disease-Associated Microglial Dysfunction. Acta Neuropathol. (Berl.) 2024, 148, 75. [CrossRef]
- Washer, S.J.; Perez-Alcantara, M.; Chen, Y.; Steer, J.; James, W.S.; Trynka, G.; Bassett, A.R.; Cowley, S.A. Single-Cell Transcriptomics Defines an Improved, Validated Monoculture Protocol for Differentiation of Human iPSC to Microglia. Sci. Rep. 2022, 12, 19454. [CrossRef]
- Daskoulidou, N.; Shaw, B.; Zelek, W.M.; Morgan, B.P. The Alzheimer’s Disease-Associated Complement Receptor 1 Variant Confers Risk by Impacting Glial Phagocytosis. Alzheimers Dement. J. Alzheimers Assoc. 2025, 21, e70458. [CrossRef]
- Bedford, L.M.; Tutrow, K.D.; Hooper, K.; Messenger, E.J.; Hernandez, M.; Lamb, B.T.; Meyer, J.S.; Richardson, T.I.; Bissel, S.J. Alzheimer’s Disease–Associated PLCG2 Variants Alter Microglial State and Function in Human Induced Pluripotent Stem Cell–Derived Microglia-like Cells. Alzheimers Dement. 2025, 21, e70772. [CrossRef]
- McQuade, C.A.; Pathology, R.G. of D.S.J.-I.M.-O. Exploring the Involvement of Oligodendrocytes in Alzheimer’s Disease Using Human iPSCs. Alzheimers Dement. 2023, 19, e079773. [CrossRef]
- Palm, T.; Bolognin, S.; Meiser, J.; Nickels, S.; Träger, C.; Meilenbrock, R.-L.; Brockhaus, J.; Schreitmüller, M.; Missler, M.; Schwamborn, J.C. Rapid and Robust Generation of Long-Term Self-Renewing Human Neural Stem Cells with the Ability to Generate Mature Astroglia. Sci. Rep. 2015, 5, 16321. [CrossRef]
- Tcw, J.; Wang, M.; Pimenova, A.A.; Bowles, K.R.; Hartley, B.J.; Lacin, E.; Machlovi, S.I.; Abdelaal, R.; Karch, C.M.; Phatnani, H.; et al. An Efficient Platform for Astrocyte Differentiation from Human Induced Pluripotent Stem Cells. Stem Cell Rep. 2017, 9, 600–614. [CrossRef]
- Lish, A.M.; Galle, P.C.; Orme, G.A.; Ashour, N.; Heuer, S.E.; Curle, A.J.; Muratore, C.R.; Young-Pearse, T.L. Protocol for Generating a Human iPSC-Derived Tri-Culture Model to Study Interactions between Neurons, Astrocytes, and Microglia. STAR Protoc. 2025, 6, 104152. [CrossRef]
- Neyrinck, K.; Van Den Daele, J.; Vervliet, T.; De Smedt, J.; Wierda, K.; Nijs, M.; Vanbokhoven, T.; D’hondt, A.; Planque, M.; Fendt, S.-M.; et al. SOX9-Induced Generation of Functional Astrocytes Supporting Neuronal Maturation in an All-Human System. Stem Cell Rev. Rep. 2021, 17, 1855–1873. [CrossRef]
- Maioli, S.; Nalvarte, I.; Ankarcrona, M.; Schultzberg, M.; Zuloaga, K.L.; Goikolea, J.; Visser, P.J.; De Strooper, B.; Winblad, B.; Pizzo, P.; et al. Bioenergetics and Lipid Metabolism in Alzheimer’s Disease: From Cell Biology to Systemic Health. J. Intern. Med. 2026, 299, 20–43. [CrossRef]
- Cheng, Y.-J.; Wang, F.; Feng, J.; Yu, B.; Wang, B.; Gao, Q.; Wang, T.-Y.; Hu, B.; Gao, X.; Chen, J.-F.; et al. Prolonged Myelin Deficits Contribute to Neuron Loss and Functional Impairments after Ischaemic Stroke. Brain 2024, 147, 1294–1311. [CrossRef]
- Ehrlich, M.; Mozafari, S.; Glatza, M.; Starost, L.; Velychko, S.; Hallmann, A.-L.; Cui, Q.-L.; Schambach, A.; Kim, K.-P.; Bachelin, C.; et al. Rapid and Efficient Generation of Oligodendrocytes from Human Induced Pluripotent Stem Cells Using Transcription Factors. Proc. Natl. Acad. Sci. U. S. A. 2017, 114, E2243–E2252. [CrossRef]
- Xu, J.; Zhao, J.; Wang, R.; Zhang, Y.; Shen, L.; Xiao, Q.; Xie, Y.; Jiang, J.; Nie, Y.; Deng, W. Shh and Olig2 Sequentially Regulate Oligodendrocyte Differentiation from hiPSCs for the Treatment of Ischemic Stroke. Theranostics 2022, 12, 3131–3149. [CrossRef]
- Livesey, M.R.; Magnani, D.; Cleary, E.M.; Vasistha, N.A.; James, O.T.; Selvaraj, B.T.; Burr, K.; Story, D.; Shaw, C.E.; Kind, P.C.; et al. Maturation and Electrophysiological Properties of Human Pluripotent Stem Cell-Derived Oligodendrocytes. Stem Cells 2016, 34, 1040–1053. [CrossRef]
- Dolciotti, C.; Righi, M.; Grecu, E.; Trucas, M.; Maxia, C.; Murtas, D.; Diana, A. The Translational Power of Alzheimer’s-Based Organoid Models in Personalized Medicine: An Integrated Biological and Digital Approach Embodying Patient Clinical History. Front. Cell. Neurosci. 2025, 19, 1553642. [CrossRef]
- Yagi, T.; Ito, D.; Okada, Y.; Akamatsu, W.; Nihei, Y.; Yoshizaki, T.; Yamanaka, S.; Okano, H.; Suzuki, N. Modeling Familial Alzheimer’s Disease with Induced Pluripotent Stem Cells. Hum. Mol. Genet. 2011, 20, 4530–4539. [CrossRef]
- Krattli, R.P.; Markarian, M.; Madan, S.; Swami, D.; McQuade, A.; Baulch, J.E.; Blurton-Jones, M.; Acharya, M.M. Comparing Functional Consequences of Human iPSC-Microglia and Neural Stem Cell-Derived Extracellular Vesicles in Mitigating Cognitive Decline in Alzheimer’s Disease. Aging Cell 2026, 25, e70341. [CrossRef]
- Ransom, L.S.; Liu, C.S.; Dunsmore, E.; Palmer, C.R.; Nicodemus, J.; Ziomek, D.; Williams, N.; Chun, J. Human Brain Small Extracellular Vesicles Contain Selectively Packaged, Full-Length mRNA. Cell Rep. 2024, 43, 114061. [CrossRef]
- Malaguarnera, M.; Cabrera-Pastor, A. Emerging Role of Extracellular Vesicles as Biomarkers in Neurodegenerative Diseases and Their Clinical and Therapeutic Potential in Central Nervous System Pathologies. Int. J. Mol. Sci. 2024, 25, 10068. [CrossRef]
- Ramos-Zaldívar, H.M.; Polakovicova, I.; Salas-Huenuleo, E.; Corvalán, A.H.; Kogan, M.J.; Yefi, C.P.; Andia, M.E. Extracellular Vesicles through the Blood-Brain Barrier: A Review. Fluids Barriers CNS 2022, 19, 60. [CrossRef]
- Li, R.-L.; Zou, Y.-Z.; Kang, S. Decoding Aging through iPSC Reprogramming: Advances and Challenges. Aging Dis. 2025. [CrossRef]
- Reed, X.; Weller, C.A.; Saez-Atienzar, S.; Beilina, A.; Solaiman, S.; Portley, M.; Kaileh, M.; Roy, R.; Ding, J.; Zenobia Moore, A.; et al. Characterization of DNA Methylation in PBMCs and Donor-Matched iPSCs Shows Methylation Is Reset during Stem Cell Reprogramming. BioRxiv Prepr. Serv. Biol. 2024, 2024.12.13.627515. [CrossRef]
- Matthew, R.; Jia, W.; Sharma, A.; Zhao, Y.; Clarke, L.E.; Cheng, X.; Wang, H.; Salli, U.; Vrana, K.E.; Robertson, G.P.; et al. Robust Activation of the Human but Not Mouse Telomerase Gene during the Induction of Pluripotency. FASEB J. 2010, 24, 2702–2715. [CrossRef]
- Lapasset, L.; Milhavet, O.; Prieur, A.; Besnard, E.; Babled, A.; Aït-Hamou, N.; Leschik, J.; Pellestor, F.; Ramirez, J.-M.; De Vos, J.; et al. Rejuvenating Senescent and Centenarian Human Cells by Reprogramming through the Pluripotent State. Genes Dev. 2011, 25, 2248–2253. [CrossRef]
- Ocampo, A.; Reddy, P.; Martinez-Redondo, P.; Platero-Luengo, A.; Hatanaka, F.; Hishida, T.; Li, M.; Lam, D.; Kurita, M.; Beyret, E.; et al. In Vivo Amelioration of Age-Associated Hallmarks by Partial Reprogramming. Cell 2016, 167, 1719-1733.e12. [CrossRef]
- Scesa, G.; Adami, R.; Bottai, D. iPSC Preparation and Epigenetic Memory: Does the Tissue Origin Matter? Cells 2021, 10, 1470. [CrossRef]
- Efrat, S. Epigenetic Memory: Lessons From iPS Cells Derived From Human β Cells. Front. Endocrinol. 2021, 11. [CrossRef]
- Haubenreich, C.; Lenz, M.; Schuppert, A.; Peitz, M.; Koch, P.; Zenke, M.; Brüstle, O. Epigenetic and Transcriptional Shifts in Human Neural Stem Cells after Reprogramming into Induced Pluripotent Stem Cells and Subsequent Redifferentiation. Int. J. Mol. Sci. 2024, 25, 3214. [CrossRef]
- Kim, K.; Doi, A.; Wen, B.; Ng, K.; Zhao, R.; Cahan, P.; Kim, J.; Aryee, M.J.; Ji, H.; Ehrlich, L.I.R.; et al. Epigenetic Memory in Induced Pluripotent Stem Cells. Nature 2010, 467, 285–290. [CrossRef]
- Aversano, S.; Caiazza, C.; Caiazzo, M. Induced Pluripotent Stem Cell-Derived and Directly Reprogrammed Neurons to Study Neurodegenerative Diseases: The Impact of Aging Signatures. Front. Aging Neurosci. 2022, 14, 1069482. [CrossRef]
- Herdy, J.; Schafer, S.; Kim, Y.; Ansari, Z.; Zangwill, D.; Ku, M.; Paquola, A.; Lee, H.; Mertens, J.; Gage, F.H. Chemical Modulation of Transcriptionally Enriched Signaling Pathways to Optimize the Conversion of Fibroblasts into Neurons. eLife 2019, 8, e41356. [CrossRef]
- Hu, W.; Qiu, B.; Guan, W.; Wang, Q.; Wang, M.; Li, W.; Gao, L.; Shen, L.; Huang, Y.; Xie, G.; et al. Direct Conversion of Normal and Alzheimer’s Disease Human Fibroblasts into Neuronal Cells by Small Molecules. Cell Stem Cell 2015, 17, 204–212. [CrossRef]
- Tang, Y.; Liu, M.-L.; Zang, T.; Zhang, C.-L. Direct Reprogramming Rather than iPSC-Based Reprogramming Maintains Aging Hallmarks in Human Motor Neurons. Front. Mol. Neurosci. 2017, 10, 359. [CrossRef]
- Yang, Y.; Chen, R.; Wu, X.; Zhao, Y.; Fan, Y.; Xiao, Z.; Han, J.; Sun, L.; Wang, X.; Dai, J. Rapid and Efficient Conversion of Human Fibroblasts into Functional Neurons by Small Molecules. Stem Cell Rep. 2019, 13, 862–876. [CrossRef]
- Puri, D.; Wagner, W. Epigenetic Rejuvenation by Partial Reprogramming. BioEssays News Rev. Mol. Cell. Dev. Biol. 2023, 45, e2200208. [CrossRef]
- Gill, D.; Parry, A.; Santos, F.; Okkenhaug, H.; Todd, C.D.; Hernando-Herraez, I.; Stubbs, T.M.; Milagre, I.; Reik, W. Multi-Omic Rejuvenation of Human Cells by Maturation Phase Transient Reprogramming. eLife 2022, 11, e71624. [CrossRef]
- Haase, F.D.; Coorey, B.; Riley, L.; Cantrill, L.C.; Tam, P.P.L.; Gold, W.A. Pre-Clinical Investigation of Rett Syndrome Using Human Stem Cell-Based Disease Models. Front. Neurosci. 2021, 15, 698812. [CrossRef]
- Sun, X.-Y.; Ju, X.-C.; Li, Y.; Zeng, P.-M.; Wu, J.; Zhou, Y.-Y.; Shen, L.-B.; Dong, J.; Chen, Y.-J.; Luo, Z.-G. Generation of Vascularized Brain Organoids to Study Neurovascular Interactions. eLife 2022, 11, e76707. [CrossRef]
- Dinasarapu, A.R.; Sutcliffe, D.J.; Ozel, E.; Thite, A.; Grychowski, L.; Visser, J.E.; Hess, E.J.; Kolk, S.M.; Jinnah, H.A. Modeling Rare Genetic Disease with Gene-Edited Induced Pluripotent Stem Cells: Relevance of the Starting Stock Line. Stem Cells Transl. Med. 2025, 14, szaf065. [CrossRef]
- Dhaiban, S.; Chandran, S.; Noshi, M.; Sajini, A.A. Clinical Translation of Human iPSC Technologies: Advances, Safety Concerns, and Future Directions. Front. Cell Dev. Biol. 2025, 13. [CrossRef]
- Castillo Ransanz, L.; Van Altena, P.F.J.; Heine, V.M.; Accardo, A. Engineered Cell Culture Microenvironments for Mechanobiology Studies of Brain Neural Cells. Front. Bioeng. Biotechnol. 2022, 10, 1096054. [CrossRef]
- Li, T.; Yin, J.; Hao, Y.; Gao, W.; Li, Q.; Feng, Q.; Tao, B.; Hao, M.; Liu, Y.; Lin, C.; et al. Single-Cell Sequencing and Organoids: Applications in Organ Development and Disease. Mol. Biomed. 2025, 6, 134. [CrossRef]
- Awada, B.; Williams, I.; Moreau, T.; Charrier-Savournin, F.; Roux, T.; Trask, O.J.; Blackmore, M.; Bixby, J.L.; Lemmon, V.P.; Al-Ali, H. Phenotypic Screening with Primary and Human iPSC-Derived Neurons. In Assay Guidance Manual; Markossian, S., Grossman, A., Baskir, H., Arkin, M., Auld, D., Austin, C., Baell, J., Brimacombe, K., Chung, T.D.Y., Coussens, N.P., Dahlin, J.L., Devanarayan, V., Foley, T.L., Glicksman, M., Gorshkov, K., Grotegut, S., Hall, M.D., Hoare, S., Inglese, J., Iversen, P.W., Lal-Nag, M., Li, Z., Manro, J.R., McGee, J., Norvil, A., Pearson, M., Riss, T., Saradjian, P., Sittampalam, G.S., Tarselli, M.A., Trask, O.J., Weidner, J.R., Wildey, M.J., Wilson, K., Xia, M., Xu, X., Eds.; Eli Lilly & Company and the National Center for Advancing Translational Sciences: Bethesda (MD), 2004.
- Gameiro-Ros, I.; Tengolics, A.J.; Prytkova, I.; Kamarajan, C.; Pang, Z.P.; Goate, A.M.; Hart, R.P.; Slesinger, P.A. High-Throughput Measurements of Neuronal Activity in Single Human iPSC-Derived Glutamate Neurons 2025, 2025.04.07.646449.
- Farasati Far, B.; Omidvar, K.; Heidari, E.; Ebrahimi, M.; Mohammadi, Y.; Ertas, Y.N. Neuronal Organoid Engineering and Disease-Focused High-Throughput Neuropharmacology: Advances, Limitations, and Translational Strategies. ACS Pharmacol. Transl. Sci. 2026, 9, 1–19. [CrossRef]
- Bye, C.R.; Qian, E.; Lim, K.; Daniszewski, M.; Garton, F.C.; Trần-Lê, B.C.; Liang, H.H.; Lin, T.; Lock, J.G.; Crombie, D.E.; et al. Large-Scale Drug Screening in iPSC-Derived Motor Neurons from Sporadic ALS Patients Identifies a Potential Combinatorial Therapy. Nat. Neurosci. 2026, 29, 40–52. [CrossRef]
- Maxwell, H.; Smith, J.; Campbell, T.; Clark, J.; La Rocca, R.; Paonessa, F.; Sitnikov, S.; Evans, M.; Maycox, P.; Livesey, F.J. A Phenotypic Screen for Novel Small Molecules That Correct Tau-Mediated Pathologies in Human Frontotemporal Dementia Neurons. Alzheimers Dement. J. Alzheimers Assoc. 2025, 21, e70620. [CrossRef]
- Strong, C.E.; Zhang, J.; Carrasco, M.; Kundu, S.; Boutin, M.; Vishwasrao, H.D.; Liu, J.; Medina, A.; Chen, Y.-C.; Wilson, K.; et al. Functional Brain Region-Specific Neural Spheroids for Modeling Neurological Diseases and Therapeutics Screening. Commun. Biol. 2023, 6, 1211. [CrossRef]
- Shin, H.; An, Y.; Lee, J.-H.; Kim, J.H.; Prasad, R.; Kim, K.-T.; Kang, H.-C.; Sun, W.; Cho, S.-W.; Cho, I.-J. One-Step Drug Screening System Utilizing Electrophysiological Activity in Multiple Brain Organoids. Adv. Sci. 2025, 12, e04913. [CrossRef]
- Tsefou, E.; Bez, S.; Birkle, T.J.Y.; Foiani, M.; Watamura, N.; Bourdenx, M.; Gavriouchkina, D.; Turkes, E.; Crawford, S.; Coneys, R.; et al. Scalable Human Neuronal Models of Tauopathy Producing Endogenous Seed-Competent 4R Tau 2025, 2025.07.11.664346.
- Nguyen, L.D.; Wei, Z.; Silva, M.C.; Barberán-Soler, S.; Zhang, J.; Rabinovsky, R.; Muratore, C.R.; Stricker, J.M.S.; Hortman, C.; Young-Pearse, T.L.; et al. Small Molecule Regulators of microRNAs Identified by High-Throughput Screen Coupled with High-Throughput Sequencing. Nat. Commun. 2023, 14, 7575. [CrossRef]
- Francis, N.; Aho, J.; Ben-Nun, I.F.; Bharti, K.; Dianat, N.; Makovoz, B.; Nouri, P.; Rothberg, J.; Song, H.; Zamilpa, R.; et al. Scaling up Pluripotent Stem Cell-Based Therapies - Considerations, Current Challenges and Emerging Technologies: Perspectives from the ISCT Emerging Regenerative Medicine Working Group. Cytotherapy 2025, 27, 1031–1042. [CrossRef]
- Marei, H.E.; Khan, M.U.A.; Hasan, A. Potential Use of iPSCs for Disease Modeling, Drug Screening, and Cell-Based Therapy for Alzheimer’s Disease. Cell. Mol. Biol. Lett. 2023, 28, 98. [CrossRef]
Figure 1.
Types and Generation of Pluripotent Stem Cell culture. Created in BioRender. Sirvinskas, D. (2026) pq7mv90.
Figure 1.
Types and Generation of Pluripotent Stem Cell culture. Created in BioRender. Sirvinskas, D. (2026) pq7mv90.

Figure 2.
iPSC Quality Control and Characterization. Created in BioRender. Sirvinskas, D. (2026) https://BioRender.com/t0s6bnj.
Figure 2.
iPSC Quality Control and Characterization. Created in BioRender. Sirvinskas, D. (2026) https://BioRender.com/t0s6bnj.

Figure 3.
Applications in Neuroscience: Monolayer vs 3D Organoid. Created in BioRender. Sirvinskas, D. (2026) https://BioRender.com/w4tte6i.
Figure 3.
Applications in Neuroscience: Monolayer vs 3D Organoid. Created in BioRender. Sirvinskas, D. (2026) https://BioRender.com/w4tte6i.

Figure 4.
Epigenetic Memory in iPSC Reprogramming. Created in BioRender. Sirvinskas, D. (2026) https://BioRender.com/07s6gz5.
Figure 4.
Epigenetic Memory in iPSC Reprogramming. Created in BioRender. Sirvinskas, D. (2026) https://BioRender.com/07s6gz5.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



