Submitted:
05 June 2026
Posted:
08 June 2026
You are already at the latest version
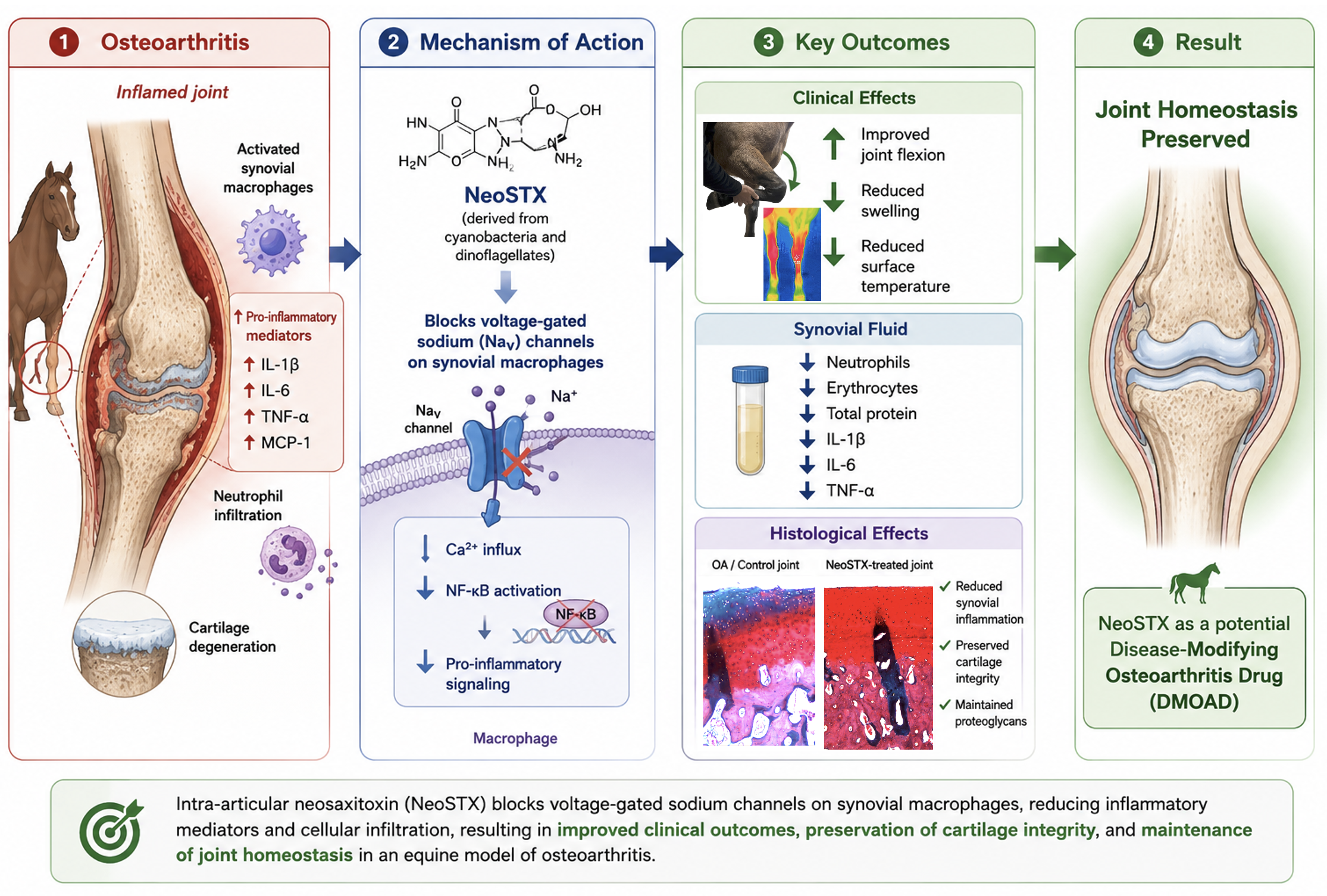
Abstract
Chronic synovial inflammation is a hallmark of osteoarthritis progression and is tightly regulated by synovial macrophages. Voltage-gated sodium channels (NaV) have emerged as modulators of macrophages-driven inflammation, positioning them as novel therapeutic targets. Neosaxitoxin, a highly selective NaV channel blocker, has anesthetic and immunomodulatory properties; however, its intra-articular effects on joint inflammation remain unexplored. Using a bilateral equine carpal osteoarthritis model, this study evaluated the immunomodulatory and tissue-preserving effects of intra-articular neosaxitoxin. Sixteen horses were randomized into two experimental groups (n=8/each): Neosaxitoxin in one joint and triamcinolone (+control) in the contralateral joint; or neosaxitoxin in one joint and saline (-control) in the contralateral joint. Clinical parameters, synovial fluid cytology and cytokine profiles, and histological changes in synovium and cartilage were assessed over 30 days. Neosaxitoxin reduced synovial inflammation, evidenced by decreased synovial effusion and surface temperature, along with improved joint flexion. Further, synovial fluid from neosaxitoxin-treated joints exhibited lower counts of erythrocytes, neutrophils, mononuclear cells, total protein, and key pro-inflammatory mediators (IL-1β, IL-6 and TNF-α). Histologically, neosaxitoxin-treated joints exhibited few synovium inflammatory infiltrate and cartilage integrity preservation. In contrast, control joints exhibited synovial hyperplasia, fibrovascular proliferation, and cartilage degeneration. Intra-articular neosaxitoxin better conserved joint homeostasis by limiting synovial inflammation and preserving cartilage integrity. These results warrant further investigation on NaV channel blockage as a novel disease-modifying strategy for osteoarthritis.
Keywords:
horse
; osteoarthritis
; DMOAD
; neosaxitoxin
; homeostasis
1. Introduction
Osteoarthritis (OA) is a debilitating disease that affects over 30% of the global population [1]. OA remains a major public health concern and represents the second most costly condition in the United States health care system [1,2]. OA is also a highly prevalent condition in domestic animals, and the leading cause of lameness in equine athletes, translating into a substantial economic burden to the equine industry [3,4,5,6,7,8]. Current therapies provide short-term clinical improvement but fail to resolve joint inflammation in the long term and have detrimental side effects [9]. There is a critical need for innovative therapies that can provide marked clinical improvements while promoting the restoration of joint homeostasis.
Synovial inflammation is a central feature of OA progression and is primarily incited by synovial macrophages [10,11,12]. Macrophages are, however, essential for preserving joint homeostasis and resolving joint inflammation [13,14,15]. Upon tissue injuries that exceed their homeostatic functions, synovial macrophages incite an inflammatory response to counteract damage and ultimately resolve inflammation [16]. However, due to a multitude of unknown reasons, this process often results in chronic inflammation [17,18]. This impaired inflammation resolution seen in the development of OA involves persistent signaling of several key molecules. Among them, calcium ions (Ca2+) play a central messenger role and are closely linked to intracellular sodium (Na+) dynamics during cellular activation [19,20,21,22,23,24]. Although ion channels are traditionally known for their critical role in controlling action potential in excitable cells of such as neurons and myocytes [25], there is growing evidence of the role of voltage-gated sodium channels (NaV) in the activation and regulation of immune cells during inflammatory responses [26,27,28,29,30], through regulation of intracellular Na+ and Ca2+. Accordingly, NaV channels have been implicated in different cellular functions such as activation, polarization, migration, adhesion, and phagocytosis [26,30,31,32,33] and are identified in both B- [34] and T-cells [26,30,35]. Recently, NaV channels were identified to regulate signaling pathways in macrophages [36], such as nuclear factor kappa B (NF-kB) signaling, suggesting that NaV channels could be harnessed in the treatment of OA [37,38]. Canonical NF-kB signaling is one the most dominant pro-inflammatory pathways leading to the expression of inflammatory cytokines and metalloproteinases identified in the chronic inflammation in OA [32,39].
Neosaxitoxin (NeoSTX) is a phycotoxin with a high capacity to block NaV channels and has been mainly studied for its local anesthetic properties in various species [40,41,42,43]. Our previous in vitro study on NaV channels in the inflammatory response of macrophages [38] suggest that targeted blockage of these channels with NeoSTX, may reduce joint inflammation and enhance their homeostatic and potentially pro-resolving functions while also reducing synovial membrane nociception [43,44]. Altogether, these observations pose NaV channels as candidate new therapeutics in OA. The aim of this study was to investigate the anti-inflammatory and analgesic effects of intra-articular injection of NeoSTX, in an equine experimental model of OA. We hypothesized that NaV channel blockade with NeoSTX would modulates macrophage-driven inflammation, preserving synovial homeostasis and mitigating osteoarthritis progression.
2. Materials and Methods
2.1. Experimental Design and Animal Selection
To evaluate the role of intraarticular administration of NeoSTX on joint inflammation and OA development, 16 crossbred horses (10 geldings and 6 mares; median age =10.5 years) were subjected to a carpal osteochondral fragment model of OA [45,46]. All procedures were performed under the guidelines and principles of international care and use of animals in medical research, along with the “Institutional Committee of Animal Use and Care” (CICUA, Certificate No: 23716 –VET– UCHe1). The study was conducted at the Faculty of Veterinary and Animal Sciences of the University of Chile in the Departments of Animal Preventive Medicine and Equestria Equine Medical Center, Universidad Andrés Bello.
Horses selected were clinically healthy (with normal CBC and blood chemistry parameters) and lame-free based on a complete musculoskeletal examination before enrollment, including response to joint flexion, gait analysis and diagnostic imaging. Horses showing middle carpal effusion or any clinical or radiographic abnormalities were excluded.
Horses were allocated in two groups (n= 8 horses/each). All horses underwent bilateral middle carpal arthroscopy to induce OA using the osteochondral fragment model [45,46,47,48,49]. In the treatment group, one joint was randomly assigned to receive an intra-articular injection of 20 µg NeoSTX (diluted in 2 mL 0.9% saline), while the contralateral joint received 2 mL (12 mg) triamcinolone (TA) as a positive control [45,49]. In the control group, one joint received 20 µg NeoSTX (0.9% saline), and the contralateral joint received 2 mL of 0.9% saline as a negative control. Overall, horses were stall-confined for 40 days: 10 days of adaptation, OA induction on day 0 (after adaptation), followed by 30 days of clinical evaluation and synovial fluid sampling. The treatment schedule, synovial fluid sampling, and clinical and systemic measurements are detailed in Figure 1.
Outcome measures of joint inflammation included thermographic assessment of joint surface, joint flexion range using a goniometer, and joint circumference. In addition, synovial fluid sampling was performed at baseline (day 0, prior to OA induction) and on days 10, 20, and 30 post-treatments. Samples were collected for cytological analysis (total nucleated cell count and differential cell count) and quantification of total protein, pro-inflammatory and regulatory cytokines, chemokines, and growth factors. At the end of the protocol, horses were sacrificed for gross assessment of joints and synovium and cartilage collection for histology (Figure 1).
2.2. Animal Husbandry
Horses were housed in 3x3 meter stalls with shavings and fed free choice alfalfa-based hay and fresh water provided ad libitum. Throughout the study period, horses were turned out for 4 hours daily in small paddocks (6 × 6 m).
2.3. Neosaxitoxin Purification Procedure
Neosaxitoxin was purified by high-performance liquid chromatography (HPLC). The toxin was extracted from the cyanobacterial cultures (Cyanobacteria aphanizomenon sp.) as previously described by Lagos et al. (1999) [50]. The toxin purity was confirmed by HPLC-MS, and its concentration standardized at doses of 10 µg / mL diluted in 0.9% saline solution.
2.4. Osteochondral Fragment Induction
On day 0, horses underwent bilateral arthroscopy of the middle carpal joints under general anesthesia. Both middle carpal joints were thoroughly examined to rule out pre-existing abnormalities. At this timepoint, if articular damage was detected during arthroscopic evaluation, the procedure was terminated, and the horse was excluded from the study and replaced. An osteochondral fragment was created in both middle-carpal joints in the radial carpal bone (RCB) at the level of the medial synovial plica using an 8-mm curved osteotome directed perpendicular to the articular surface [46]. The fragment was allowed to remain adhered to the joint capsule proximally, and the exposed subchondral bone between the fragment and parent bone was debrided using a motorized arthroburr, creating a 15-mm wide defect in which the bone fragment sat. The skin was closed with 2-0 glyconate monofilament in a simple interrupted pattern. Both operated joints received compressive bandages. Because the evaluation of joint inflammation modulation is a key feature of the study, horses did not receive NSAIDs nor antimicrobials. Bandages were changed every 3 days or when necessary for data collection and maintained until suture removal at 14 days after surgery.
2.5. Joint Treatments Administration
On days 6, 15 and 25 after OA induction, the assigned treatments were delivered (Figure 1). In both groups the treatments were randomly assigned to one experimental joint and maintained throughout the study. In the treatment group (NeoSTX vs. TA) one joint was randomly selected to be injected with 2 mL of NeoSTX (10µg/mL saline) as an experimental treatment and 2 mL of triamcinolone acetonide (6mg/mL) [45,49] in the contralateral joint as positive control. In horses allocated in the control group, one joint was randomly selected to be injected with 2 mL of NeoSTX and 2 mL of saline as negative control, in the contralateral joint. Saline (NaCl 0.9%) was used as control because it was the vehicle for NeoSTX.
The selected dose of 20 µg of NeoSTX was calculated based on doses previously reported humans [43], and what was recently described in horses [51]. Additionally, the treatment protocol (3 injections) was aimed at mimicking the treatment regimens for orthobiological therapies for joint disease [52,53].
2.6. Exercise Program
After 7 days of model induction, horses entered a daily exercise regimen of 7 days/week for 3 more weeks. Each day, horses were actively trotted for 1 hour in an automated equine exerciser (14 meters diameter) [45]. This exercise protocol was adapted to incorporate controlled active trotting (3.2 m/s) while matching the equivalent distance from the original protocol [45]. Nonetheless, as galloping imposes higher impact and loading to the joint, the duration of exercise was increased. This modification was designed to provide consistent and repetitive joint loading sufficient to induce synovial inflammation while ensuring animal safety.
2.7. Clinical Evaluation of Horses
Clinical monitoring included daily assessment of heart rate (HR), respiratory rate (RR), rectal temperature, feed and water intake. Joints were assessed for inflammation by measuring joint circumference and surface temperature using infrared thermography (Flir One® camera), at days 0, 2, 4, 7, 10, 15, 20, 25, and 30. Thermographic measurements were standardized by conducting them in the same enclosed room, maintaining a one-meter distance between the camera and the joint. Musculoskeletal evaluation also included joint palpation and range of motion assessment; joint angle (goniometer) and pain response to flexion were recorded.
2.8. Synovial Fluid (SF) Sampling and Assessment
Synovial fluid (SF) samples (2 mL) were aseptically collected from each treated joint at day 0 (baseline) 10, 20 and 30, and divided into two tubes, one without anticoagulant (Eppendorf® Protein LoBind, Merck, Darmstadt, Germany) and another containing EDTA (Vacutainer™, Becton Dickinson, New Jersey, USA). At day 30, synovial fluid was collected in all horses before slaughter. SF was first examined for physical characteristics including color, turbidity, and viscosity. EDTA-free samples were centrifuged at 2,000 × g for 10 minutes to remove cellular components. A 200 μL aliquot was immediately analyzed using a spectrophotometer (Chemo20®, Shinova, Shanghai, China) to measure total protein [54,55], calcium [56,57], and glucose [58] concentrations. The remaining cell-free supernatants were stored at −20 °C for batch cytokine analyses. EDTA-treated samples were processed within 1 hour of collection using an automated hematology analyzer (Hemo Plus 2900V®, Shinova, Shanghai, China) to determine total nucleated cell counts and differential cell counts for neutrophils, mononuclear, and red blood cells.
2.9. Clinical Evaluation of Horses
Clinical monitoring included daily assessment of heart rate (HR), respiratory rate (RR), rectal temperature, feed and water intake. Joints were assessed for inflammation by measuring joint circumference and surface temperature using infrared thermography (Flir One® camera), at days 0, 2, 4, 7, 10, 15, 20, 25, and 30. Thermographic measurements were standardized by conducting them in the same enclosed room, maintaining a one-meter distance between the camera and the joint. Musculoskeletal evaluation also included joint palpation and range-of-motion assessment; joint angle (goniometer) and pain response to flexion were recorded.
2.9. Cytokine Concentrations
Synovial fluid concentrations of twenty-three analytes including cytokines, chemokines and growth factors were assessed using Luminex® xMAP® technology (Thermo Fischer Scientific, Waltham, MA, USA). Synovial fluid samples were hyaluronidase-digested (100 IU/mL testicular hyaluronidase in 0.05 M acetate buffer pH 4.5, LS005474; Worthington Biochemical, Lakewood, NJ, USA) using 10 µL of hyaluronidase solution per 200 µL of synovial fluid and incubated for 30 min at 37 ◦C. Next, the multiplexing analysis was performed by Eve Technologies Corporation (Calgary, Alberta, Canada) using the Luminex® 200™ system (Luminex Corporation/DiaSorin, Saluggia, Italy) with Bio-Plex Manager™ software (Bio-Rad Laboratories Inc., Hercules, California, USA). Twenty-three markers were measured in the samples using the Eve Technologies’ Equine Cytokine 23-Plex Discovery Assay® Array (EQD23) as per the manufacturer’s instructions for use MILLIPLEX® Equine Cytokine/Chemokine Magnetic Bead Panel Cat. EQCYTMAG-93K, MilliporeSigma (Sigma, Burlington, Massachusetts, USA). The 23-plex consisted of Eotaxin, FGF-2, Fractalkine, G-CSF, GM-CSF, GRO/KC, IFNγ, IL-1α, IL-1β, IL-2, IL-4, IL-5, IL-6, IL-8, IL-10, IL-12p70, IL-13, IL-17A, IL-18, IP-10, MCP-1, RANTES, and TNF-α.
2.10. Synovial Membrane and Cartilage Histology
At day 30, horses were humanely slaughtered and synovial membrane and osteochondral biopsies were harvested using a 6 mm dermal biopsy- and osteochondral punches. For histological assessment, tissue samples were fixed in acetic zinc formalin (AZF Fixative®, Newcomer Supply). Following fixation, tissues were processed and paraffin embedded, sectioned at 5 µm, and stained with haematoxylin and eosin (H&E). Sections were examined for histopathology using the OARSI scoring system [59]. Light microscopy at magnifications ranging from 10× to 1000×. Representative images were captured using an Olympus FSX100 microscope. Samples were evaluated by analyzing the average of five randomly selected fields per section at 200× magnification.
Osteochondral samples were decalcified in neutral ethylenediaminetetracetic acid (EDTA) solution prior to processing. In addition to H&E, osteochondral samples were also stained with Safranin-O to determine proteoglycan content between treatment groups. Samples were blindly analyzed by a board-certified veterinary pathologist using the OARSI scoring guideline [59]. Of the 16 horses included in this study, histopathological samples could be obtained from only 10 animals due to logistical constraints. Among these, five horses belonged to the NeoSTX vs TA group, and five horses were included in the NeoSTX vs saline group.
2.11. Statistical Analysis
The sample size for this study (n = 16; 8 animals per group) was estimated using G*Power software (Düsseldorf, Germany) through an a priori power analysis, considering α, type II error, and effect size (f). Parameters included a 95% confidence level, a significance threshold of 0.05, and 80% statistical power (1–β) with an effect size f of 0.3. For quantitative variables, normality was assessed using the Shapiro–Wilk test. Due to the dataset’s complexity of the, a Principal Component Analysis (PCA) was performed to reduce dimensionality and facilitate data interpretation while preserving key trends. To further account for data’s structure, a multivariate approach was applied to identify relevant patterns. A Generalized Linear Mixed Model (GLMM) was fitted, followed by estimation of marginal means with pairwise post hoc comparisons and 95% confidence intervals, with treatment, time, and their interaction as fixed effects, while horses were included as a random factor. For cytokine values that were below the lower limit of detection (LLOD), LLOD*0.5 imputed values were used in statistical analysis. Data obtained from histopathological samples were analyzed using the non-parametric Kruskal-Wallis test followed by a Pairwise Wilcoxon post hoc test using the Benjamini-Hochberg correction method. All statistical analyses were performed in R software version 4.0.2. Qualitative parameters were evaluated using the Pearson chi-square test. Statistical significance was set at p < 0.05.
3. Results
3.1. Population Demographics
The study included sixteen adult crossbred horses (9 geldings and 7 mares) with a mean age of 9.87 ± 2.68 years (mean ± SD) and a mean body weight of 377 ± 50.87 kg (mean ± SD). The treatment group (NeoSTX vs TA) consisted of 5 geldings and 3 mares, whereas the control group (NeoSTX vs saline) comprised 4 geldings and 4 mares. All subjects recovered from surgery uneventfully. Radiographs obtained on Day 30 clearly identified the experimentally induced fragments, although there was evidence of healing and subchondral sclerosis in the dorso-medial aspect of the radio-carpal bone; no other radiographic lesions, such as osteophytes or subchondral bone lysis, were evidenced (Figure S1).
3.2. Clinical Evaluation of Horses: Joint Flexion Angle, Surface Temperature and Circumference
Induction of the model produced an inflammatory response across all joints, characterized by increased joint flexion angle (indicative of reduced range of motion), increased joint circumference, and elevated surface temperature. Treatment with NeoSTX resulted in a marked improvement in joint flexion over time (Figure 2A). Although early responses at days 4 and 6 showed a similar trend across groups (i.e., increased joint angle), these differences are likely random, as these time points preceded treatment. Additionally, one horse in the TA-treated group appeared unresponsive to the model for this outcome measure, consistently exhibiting lower values compared to the rest of the group. After treatment, NeoSTX-treated joints showed significantly lower angles differences emerging from day 10 throughout the study period (p < 0.018). Although no differences were detected between NeoSTX and TA, the latter exhibited a trend toward greater residual joint angle by the end of the study (p = 0.01) (Figure 2A,B).
Joint circumference followed a similar pattern. NeoSTX-treated joints showed a sustained reduction in perimeter, whereas saline-treated joints exhibited persistently increased joint circumference, reaching significant differences from day 10 onward when compared with NeoSTX-treated joints (p < 0.02). In contrast, NeoSTX and TA demonstrated largely comparable responses, with no statistically significant differences between them (Figure 2C).
Surface temperature changes were consistent with these findings. Following an initial increase associated with model induction, both NeoSTX and TA treatments led to a progressive decrease of joint temperature. In contrast, saline-treated joints maintained elevated temperatures throughout the study, with significant differences compared to NeoSTX at day 30 (Figure 2D).
Collectively, these findings demonstrate that NeoSTX attenuated clinical signs of joint inflammation relative to saline treated controls.
3.3. Principal Component Analysis of the Synovial Fluid
Principal component analysis (PCA) was performed to integrate synovial fluid data, identify variables with the most meaningful differences between groups, and cluster patterns while reducing data dimension. Synovial fluid cytokines/chemokines including eotaxin, fractalkine, G-CSF, GM-CSF, GRO/KC, IL-1α, IL-5, IL-8, IL-12p70, RANTES, as well as calcium and glucose, contributed minimally to variability over time. In contrast, pro-inflammatory cytokines and chemokines (MCP1, IP10, IL-18), clustered primarily along PC1, which accounted for 23.2% of the variance and can be interpreted as an acute synovial inflammatory burden axis, integrating cellularity (neutrophils, RBC), total protein, and cytokine activation (IL-1β, IL-6, TNF-α). NeoSTX-treated joints clustered closer to baseline values, aligning with mononuclear cells and IL-17, consistent with a regulated immune profile. In contrast, saline-treated joints aligned with total proteins, IL-6, neutrophils, RBC, and MCP-1, reflecting a pro-inflammatory profile (Figure 3). TA-treated joints formed a distinct, and more dispersed cluster, characterized by association with pro-inflammatory mediators (IL-1β, TNF-α, IFN-g, IL-2 and IL-18); nonetheless, it was also characterized by a marked upregulation of anti-inflammatory cytokines (IL-4, IL-10) (Figure 3). Together, these findings indicate that NeoSTX maintains a near-baseline, homeostatic biomarker profile, distinct from the pro-inflammatory trajectory observed with saline and the divergent cytokine modulation induced by TA.
3.4. Synovial Fluid Cytology
SF obtained from NeoSTX-treated joints maintained normal color and viscosity, with no signs of hemorrhage. In contrast, saline-treated joints tended to be more reddish characterized by loss of viscosity. SF collected from TA- treated horses showed progressive inflammatory changes, including decreased viscosity and discoloration from translucent yellow to reddish-brown, consistent with indicating hemarthrosis and/or synovial reaction. Moreover, synovial fluid volume was consistently lower.
Red blood cell (RBC) counts were comparable across groups at baseline (pre-treatment). By days 20 and 30, NeoSTX-treated joints, showed significantly lower RBC compared with saline-treated joints (p = 0.0006 and 0.014, respectively) (Figure 4A).
Total nucleated cells (TNCC) counts were also similar at baseline. From day 10 onward, horses treated with TA displayed a pronounced increase in TNCC, which was significant at day 10 (p = 0.0013) and day 30 (p = 0.019), and largely represented by mononuclear cells increase.
Neutrophil proportions increased in all groups at day 10, consistent with acute inflammatory recruitment. Subsequently, both NeoSTX- and TA-treated joints showed a decline toward baseline, whereas saline horses maintained elevated counts at later time points, with significant differences compared with NeoSTX (Figure 4C).
Mononuclear cell dynamics further supported a time-dependent anti-inflammatory effect of NeoSTX. NeoSTX-treated horses exhibited a sustained suppression of mononuclear cells from day 10 through day 30, with relatively low variability over time. Saline-treated horses showed a comparable but less stable pattern. Notably, TA-treated horses demonstrated a progressive increase after day 20, culminating in a pronounced increase at day 30, with higher counts compared to NeoSTX, although, non-significant (Figure 4E). This divergence suggests that, while all groups undergo an early cell type shift phase, NeoSTX is associated with a more controlled and sustained modulation of mononuclear cell populations.
3.5. Synovial Fluid Cytokines and Chemokines
Total protein (TP) concentrations, did not differ between NeoSTX- and TA-treated groups at any time point. A trend toward higher TP in TA-treated horses was observed at day 10. Although all three groups showed a trend toward an increase at day 10, NeoSTX values remained closer to baseline throughout the study. On the other hand, TP decreased significantly in the NeoSTX group at days 20 and 30 compared to saline- treated joints (p = 0.0008 and 0.013, respectively) (Figure 5A).
Several key mediators were selected from the PCA analysis for the statistical analysis of intergroup differences NeoSTX treatment consistently lowered the expression of both pro-inflammatory (IL-1β, IL-6, TNF-α) (Figure 5B, 5C, and 5D) and chemokines (IL-13, IL-18, IP-10, IFN-γ, IL-2, and IL-17) (Figure S2) compared with TA- and saline- treated groups. NeoSTX appears to stabilize IL-1β levels, while saline-treated horses evidenced an upregulation of IL-1β over time. The strongest evidence is on day 20, when IL-1β was significant lower in NeoSTX group compared with saline group (p = 0.018); this trend persisted at day 30 but did not reach significance (Figure 5B). Similar response was observed with IL-6 and TNF-α, inflammatory biomarkers, where NeoSTX maintained a stable cytokine concentration despite the model induction and exercise program whereas saline-treated horses showed a rise in IL-6 on days 20 and 30 (p = 0.031 and 0.017, respectively) (Figure 5D). Although saline-treated horses tended to have higher TNF-α levels than NeoSTX at all post-baseline times, those differences were small and not statistically significant (Figure 5C).
Other pro-inflammatory cytokines, including IFN-γ and IL-2, were also upregulation in the saline- and TA-treated groups when compared with NeoSTX-treated joints, in which cytokine concentrations were down-modulated to near-baseline levels. In contrast, IL-17 did not show statistically significant differences among groups. A similar pattern was observed for other chemokines associated with OA, such as IP-10 and IL-18, which showed significantly higher concentrations on days 20 and 30 post-model induction in the saline-treated joints in comparison with the NeoSTX-treated group. Additionally, IL-18 levels were also increased on day 20 (p= 0.02) in the TA-treated group when compared with the NeoSTX group (Figure S2).
Following model induction, MCP-1 concentrations increased in all three groups. After treatments, MCP-1 levels decreased and subsequently stabilized in the NeoSTX-treated group, similar to the pattern observed in the TA-treated group, although the latter exhibited greater variability. In contrast, the saline group showed a progressive increase in synovial fluid MCP-1 concentrations, when compared with the NeoSTX group at day 30 (Figure 6A).
Regulatory cytokines (IL-4, IL-10) remained at low concentrations in the NeoSTX-treated group compared with the saline- and TA-treated groups. NeoSTX maintained a non-inflammatory, near-baseline cytokine profile without inducing compensatory overactivation of anti-inflammatory pathways. IL-10 levels varied over time in both control groups, peaking around day 10; however, NeoSTX did not significantly alter IL-10 concentrations compared with saline- or TA-treated horses. In contrast, TA treatment was associated with increased IL-10 and IL-4 levels at days 20 and 30. These increases were statistically significant when compared with the NeoSTX group (Figure 6B and 6C). IL-13 concentrations remained stable over time within each group, with no significant differences observed among treatments (Figure S2).
Finally, estimated marginal means analysis revealed time-dependent differences across inflammatory mediators. NeoSTX-treated joints exhibited smaller effect sizes, with confidence intervals largely overlapping zero, indicating a more stable inflammatory profile. In contrast, saline-treated joints showed a progressive increase in pro-inflammatory cytokines and chemokines, with several comparisons versus NeoSTX yielding 95% confidence intervals that excluded zero. TA-treated joints displayed a heterogeneous and more variable response, with some differences relative to NeoSTX emerging at later time points. Overall, these findings suggest that NeoSTX attenuates inflammatory responses compared to saline and, to a lesser extent, triamcinolone (Table S1).
3.6. Hyaline Cartilage and Synovial Membrane Evaluation
3.6.1. Gross Pathology
Gross inspection of NeoSTX- treated joints revealed smooth, glistening articular cartilage surfaces, with uniform coloration and no visible fibrillation or erosions. The synovium appeared pale pink and thin, without villous hypertrophy or hyperemia. No evidence of exudate, hemorrhage, or pannus formation was observed (Figure 7B and 7C).
In contrast, TA-treated joints display the most marked macroscopic changes, including synovial hyperemia, cartilage surface irregularities, and hemorrhagic synovial fluid. TA-treated joints exhibited a more pronounced synovial membrane hyperemia and thickening than saline or NeoSTX. Cartilage focal discoloration was observed in 2 out of 5 joints. Moreover, mild fibrin deposition or pannus-like tissue is visible indicating synovial activation (Figure 7E and 7F).
Finally, saline- treated joints, on gross inspection, consistently evidenced a mildly swollen synovial membrane and hyperemia, compatible with chronic inflammation (Figure 7H and 7I).
3.6.2. Synovial Membrane Histology
Neosaxitoxin- treated synovium displayed a thin intimal lining, typically 1–2 cell layers, with no marked hyperplasia, inflammatory cells, or subintimal edema (Figure 8A). The subintimal layer was loosely organized with occasional small areas of mononuclear cells throughout the sections and slight increase in vessels in focal locations (Figure 8B). Slight subintimal fibrosis was evidenced throughout the sections (Figure 8C). Additionally, OARSI score (cellular infiltration, vascularity, intimal hyperplasia, subintimal edema, and subintimal fibrosis) were all low, near baseline.
TA- treated joints showed a grade 2 intimal hyperplasia observed as synovial villi with 4-5 rows of intimal cells within the sections, with mild increased in number and dilation of vessels in focal locations (Figure 8D). This group showed a moderate cellular infiltration in the subintimal layer, consistent with neutrophils and mononuclear cells infiltrates (Figure 8E and 8F). This group exhibits the highest OARSI scores for cellular infiltration, intimal hyperplasia, and composite score.
Finally, saline group also showed alteration within the synovial membrane characterized by mild intimal hyperplasia (2–4 layers) and slight congestion of synovial villi (Figure 8G). Mild inflammatory infiltrate in subintimal layers was also evidenced, but less intense than TA (Figure 8H), accompanied by occasional inflammatory cells (Figure 8I). The saline group at the end of the study showed mild synovial changes, consistent with an inflammatory process associated with the radio carpal bone osteochondral fragment induction.
3.6.3. Hyaline Cartilage and Subchondral Bone Histology
In the NeoSTX- treated group, showed slight Fibrillation/fissuring of the articular cartilage restricted to surface and superficial zone. Minor disruption of subchondral bone matrix. Lesion occupies <25% of histologic surface and extends no more than 1-2 mm deep to the normal chondrosseous junction. (Figure 9A). There was a homogeneous chondrocyte distribution, with small lacunae and slight chondrocytes clustering (Two chondrocytes (doublets) along superficial aspect). Matrix staining was uniform (Figure 9B). At higher magnification, chondrocytes showed normal morphology, with well-defined lacunae and no evidence of necrosis or pyknosis, and occasionally focal loss (Figure 9C).
TA- treated joints showed surface irregularities and early fissuring that extends into the middle zone in sections. Chondrone formation was more evident, and 2-3 chondrocytes (doublets & triplets) were observed within same lacunae along superficial zone; Subchondral bone displayed prominent vascular channels (Figure 9D), and some fissures or clefts extend into the transitional zone (Figure 9E). Moreover, nuclear pyknosis and cytoplasmic condensation were observed, suggesting focal cell loss (Figure 9F).
Finally, saline-treated horses displayed a hyaline cartilage characterized by a mild surface undulations and occasional fissures (Figure 9G). Chondrone formation was also observed, and 2-3 chondrocytes (doublets & triplets) were seen within same lacunae along superficial aspect of the articular cartilage section. Matrix staining is slightly uneven compared to NeoSTX (Figure 9H). Occasional degenerative chondrocytes were present, with mild focal loss (Figure 9I).
Safranin-O staining of cartilage samples from joints treated with NeoSTX exhibited overall a more preserved or a mild loss of proteoglycan content, with less than 25% loss of staining characteristics (Figure 10A), while those treated with TA showed a mild to significant proteoglycan loss from the superficial layer of the cartilage, with 25-50% loss of staining characteristics (Figure 10B), and saline- treated joints showed a more evident proteoglycan content loss throughout the cartilage layers with 50-75% loss of staining characteristics (Figure 10C). In summary, in our study, NeoSTX- treated joints, showed the lowest OARSI cartilage score assessment between groups.
4. Discussion
Intra-articular (IA) administration of NeoSTX in our model was associated with measurable anti-inflammatory effect and joint homeostasis restoration, accompanied by preservation of hyaline cartilage morphology. To our knowledge, this is the first controlled experimental in-vivo study evaluating NeoSTX, as a joint therapy with emphasis on its immunomodulatory effects. In this study, joint injection with NeoSTX resulted in decreased gross, laboratory, and histological markers of inflammation (cellular and vascular), together with lower concentrations of IL-1β, IL-6, TNF-α in synovial fluid. These findings extend our previous work demonstrating the feasibility of IA NeoSTX administration in healthy joints [51] and highlight its therapeutic potential in inflammatory joint disease.
The modified carpal chip model employed in this study provides a robust and reproducible platform for the induction of osteoarthritis-associated synovial inflammation in horses. Although traditional protocols incorporate high-speed galloping to exacerbate joint loading, accumulating evidence indicates that controlled, repetitive mechanical stimulation is sufficient to trigger the inflammatory cascade and structural changes characteristic of early OA [45,46,49,60]. In the present study, the adapted exercise protocol based on active trotting generated consistent clinical, cytological, and molecular variations, including joint effusion, increased temperature, and inflammatory biomarker profiles, confirming successful model induction. Importantly, the observed responses closely parallel those reported in established equine OA models, supporting the validity of this modified approach as a reliable alternative under controlled experimental conditions. Therefore, this model provides an appropriate framework to investigate targeted immunomodulatory strategies and their effects on joint homeostasis.
TA was used as a positive anti-inflammatory control in this study. IA corticosteroids—particularly triamcinolone acetonide—have historically been among the most used treatments in equine practice due to their ability to provide rapid clinical improvement and suppress local inflammatory mediators, including proteins, prostaglandins, and pro-inflammatory cytokines [45,49,61,62,63]. However, recent reports have raised concerns regarding their long-term effects [64,65]. Chronic or repeated intra-articular corticosteroid administration has been associated with reduced proteoglycan synthesis, synovial membrane fibrosis, increased vascularization, decreased range of motion, and ultimately progression of OA rather than joint protection [9,65,66,67,68]. These effects appear to be dose- and frequency-dependent and are particularly evident in repeated administration protocols [9]. Findings in the present study are consistent with these observations: TA-treated joints exhibited clear changes in synovial fluid cytokine profiles and histopathological alterations of both cartilage and synovial membrane, reflecting loss of joint homeostasis with a biological impact of repeated corticosteroid administration, independent the recommended dose used of 12 mg per joint [69]. Our findings are align with what have been observed in both humans and horses, and studies in where repeated and undiscriminated injection of TA, despite inducing a short term clinical improvement, no long term effectiveness is warranted, and detrimental and toxic effects over the cartilage are prone to be induced [65,70], moreover, it has been reported in patients with active Knee OA, corticosteroids induce cartilage volume lost [68]. While TA remains a cornerstone for short-term inflammation control, its potential deleterious effects on joint tissues over time must be carefully considered when designing therapeutic strategies.
Clinically, our findings, at the short term are consistent with the well-documented anti-inflammatory effects of intra-articular TA, which provides rapid clinical improvement after injection [63,70]. The anti-inflammatory effects of TA are primarily mediated through inhibition of phospholipase A2, thereby preventing the release of arachidonic acid and subsequent activation of the inflammatory cascade. This leads to a reduction in prostaglandin and leukotriene synthesis, resulting in decreased synovial inflammation. Additionally, corticosteroids have been shown to downregulate the expression of pro-inflammatory cytokines such as IL-1β, TNF-α, and IL-6, although inhibition of the NF-κB pathways has been proposed the exact pathways remain incompletely understood [69]. Finally, the analgesic effect of corticosteroids is believed to occur indirectly, through the suppression of local inflammatory mediators and vasoactive amines, which reduces nociceptor sensitization and pain perception [63,70]. On the other hand, the mechanism through which NeoSTX exerts its anti-inflammatory effects remains hypothetical but can be inferred from several lines of evidence reported in the literature and the results obtained in our study. Voltage-gated sodium channels (NaV) have been identified in immune cells, including macrophages, but also in chondrocytes [71]. Altered expression or function of these ion channels has been implicated in the pathophysiology of several inflammatory and degenerative diseases [33,37,72]. This concept has recently gained attention in OA research, where NaV channels have been proposed as potential biomarkers and therapeutic targets due to their involvement in inflammatory signaling and cellular activation [73,74]. The present findings support the notion that NaV channel blockade represents a promising approach to modulating joint inflammation and potentially protecting articular cartilage. Isolated studies have demonstrated that voltage-gated sodium channel (NaV) blockade in macrophages can lead to reduced transcription of mRNA and decreased expression of key pro-inflammatory mediators, including IL-1β, IL-6, TNF-α, and iNOS [38]. In addition, NaV channels have been implicated in the activation of immune cells such as lymphocytes and macrophages, suggesting a broader immunomodulatory role beyond their classical neuronal function [22,25,26,30,31,33,36,75]. Importantly, Na+ and Ca2+ fluxes play a critical role in immune cell activation. Following receptor–ligand engagement—such as Toll-like receptor (TLR) recognition of damage-associated molecular patterns (DAMPs)—an initial influx of sodium occurs through NaV channels expressed on immune cells, particularly macrophages. This event is required to elevate intracellular sodium concentration, which in turn drives a secondary calcium peak via the sodium–calcium exchanger at the plasma membrane. The first, smaller calcium peak originates from endoplasmic reticulum stores immediately after receptor activation, while the second, larger peak—essential for full cellular activation—is sodium-dependent [22].
Accordingly, we proposed that NeoSTX-mediated NaV blockade prevents the initial sodium influx, thereby inhibiting the secondary calcium peak required for downstream signal transduction. Calcium acts as a key second messenger for multiple signaling cascades, including NF-κB and mitogen-activated protein kinase activation. In the absence of this calcium signal, immune cells fail to undergo full activation, resulting in suppression of both signaling. This concept is supported by studies showing that voltage gated sodium channel blockers such as lidocaine can inhibit NF-κB activation and downstream transcription of pro-inflammatory genes [27,76,77,78]. In this context, inhibition of NF-κB leads to the downregulation of transcription factors involved in the expression of pro-inflammatory cytokines and to a reduction in the upregulation of their membrane receptors. This integrated mechanism provides a plausible explanation for the anti-inflammatory and immunomodulatory effects of NeoSTX observed in this study. The proposed mechanism is summarized in Figure 11. NeoSTX may exert its effects also to chondrocytes, as suggested by the histological findings in the present study. In cartilage samples from NeoSTX-treated joints, we observed reduced chondrocyte loss (empty lacunae), less pyknosis, fewer cell clusters, and importantly less extracellular matrix degradation compared with control groups evidence by the Safranin-O staining.
Like TA, the anti-inflammatory action of NeoSTX may also indirectly modulate nociceptor activity within the synovial membrane by decreasing local inflammatory mediators. However, we also propose an additional direct mechanism of action at the synovial membrane, where local blockade of voltage-gated sodium channels inhibits nociception at the joint level, acting similarly to other local anesthetics. What distinguishes NeoSTX from conventional local anesthetics is its long-lasting effect, providing extended analgesia without the need for repeated administrations [40,79,80,81]. This dual mechanism—immunomodulatory at the cellular level and direct local anesthetic at the synovial membrane—may explain the sustained clinical and biological effects observed in NeoSTX-treated joints in this OA model.
Pro-resolving cytokines such as IL-4 and IL-10, well known as anti-inflammatory mediators, are expected to increase its expression during inflammation resolution [10,82], however, such increases are directly proportional to the expression of so-called pro-inflammatory cytokines such as TNF-α [83,84,85]. In NeoSTX-treated joints, their levels returned to baseline after treatment, whereas TA-treated joints exhibited sustained elevation. Although, these cytokines are known as anti-inflammatory they can exhibit dual functions, participating in both resolution and propagation of inflammation [86]. A key determinant of successful inflammation resolution is the acute inflammatory response itself, which serves as a critical trigger for activating pro-resolving pathways [84,85]. We explained our results, firstly for TA-treated joint as a normal physiologic response induced by an acute inflammatory process with sustained increase of TNF-α, IL-6, IL-2 and IFN-γ, to attempt to restore homeostasis. In contrast, the lower cytokine concentration in NeoSTX-treated horses can be explained by a lack of a pathologic inflammatory signaling (early decreased of IL-1β, TNF-α, IL-6, IL-2 and IFN-γ SF concentrations) needed to activate a pro-resolving response, situation also described by other authors when PPARγ agonists were able to decrease the TNF-α concentrations and ultimately associated with decreased concentrations of IL10. These findings were further supported by the proteoglycan preservation in the cartilage of treated joints with NeoSTX.
Another important finding in NeoSTX-treated horses was the maintenance of stable levels of chemokines and growth factors throughout the study, contrasting with the other groups. This regulatory pattern was evident both in synovial fluid cytokine profiling and in the histopathological evaluation of synovial membrane sections, which revealed a significantly lower recruitment of inflammatory cells and red blood cells in NeoSTX-treated joints. These findings strongly support the hypothesis that NeoSTX exerts a broad anti-inflammatory effect, rather than modulating only isolated inflammatory pathways. Going further, our results could also represent that modulation of joint inflammation through blocking NaV channels could upregulate the non-canonical NF-κB pathway and downregulate the canonical pathway, increasing the expression of key gens such as RELB, PPARG [12] nonetheless, this statement needs further investigation.
Despite these promising results, several limitations should be acknowledged. Sample size was a limitation and even more the impossibility of collecting all postmortem tissue samples. Nonetheless, it should be taken in count that our sample size is the often the sample size for equine and translational large animal orthopedic models. Another limitation of this study, despite is just one of the several parameters assessed, is that we were unable to access a controlled temperature room for thermographic data gathering, which could have affected the temperatures obtained during this study, moreover, temperatures could have been bias due to hair growth over the 30 days period of evaluation, nevertheless this factor affected all horses the same; nonetheless, we were aware of this situation, and we took all the measurements at the same time over the days to decrease the measurement bias. We The above limitation was somewhat minimized by measuring other clinical parameters such as joint perimeter and range of joint flexion.
5. Conclusions
In conclusion our findings support targeting NaV channels during early stages of equine OA through repeated IA NeoSTX injections as a promising disease-modifying therapy for OA. Compared with corticosteroids, NeoSTX may offer advantages in long-term joint preservation by combining anti-inflammatory, immunomodulatory, and sustained analgesic effects.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, C.D., N.L., B.M and G. R-T.; methodology, C.D, N.L., G. R-T., B.M.; validation, B.M., G.R-T.; formal analysis, C.D.; investigation, C.D., L.O.; resources, N.L., G.R-T., C.G; data curation, C.D., L.O., C.G.; writing—original draft preparation, C.D.; writing—review and editing, B.M, G.R-T..; visualization, C.D.; supervision, B.M., G.R-T..; project administration, G.R-T.; funding acquisition, C.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research was partially funded by the postgraduate programme Doctorado en Ciencias Silvoagropecuarias y Veterinarias from the Universidad de Chile and ANID programa Beca Doctorado Nacional N 21230257/2023 to C.D.
Institutional Review Board Statement
The protocol was prepared before this study, registered, and approved by the “Comité Institucional de Cuidado y Uso de Animales” of the University of Chile (CICUA, certificate no.: 23716–VET–UCH1e).
Informed Consent Statement
Informed consent was obtained from the horse owners enrolled in this study.
Data Availability Statement
The original contributions presented in this study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Acknowledgments
The authors would like to acknowledge all Equestria Equine Medical Center personnel who helped during all the study development. Slaughterhouse “Don Pedro” (equine primary tissue samples).
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| CBC | Complete blood count |
| DMOAD | Disease modifying osteoarthritic drug |
| GLMM | Generalized linear mixed model |
| IA | Intraarticular |
| LLOD | Lower limit of detection |
| NaV | Voltage-gated sodium channel |
| NeoSTX | Neosaxitoxin |
| OA | Osteoarthritis |
| OARSI | Osteoarthritis Research Society International |
| PCA | Principal component analysis |
| PPARG | Peroxisome proliferator-activated receptor gamma |
| RBC | Red blood cells |
| RCB | Radial carpal bone |
| SF | Synovial fluid |
| TA | Triamcinolone |
| TNCC | Total nucleated cells |
| TP | Total proteins |
References
- Cisternas, M.G.; Murphy, L.; Sacks, J.J.; Solomon, D.H.; Pasta, D.J.; Helmick, C.G. Alternative Methods for Defining Osteoarthritis and the Impact on Estimating Prevalence in a US Population-Based Survey. Arthritis Care Res. 2016, 68, 574–580. [Google Scholar] [CrossRef]
- Boring, M.A.; Hootman, J.M.; Liu, Y.; Theis, K.A.; Murphy, L.B.; Kamil; Barbour, E.; Helmick, C.G.; Brady, T.J.; Croft, J.B. Morbidity and Mortality Weekly Report Prevalence of Arthritis and Arthritis-Attributable Activity Limitation by Urban-Rural County Classification-United States. 2015. [Google Scholar]
- Oke, S.L.; Mcilwraith, C.W.; Acvs, D. Review of the Economic Impact of Osteoarthritis and Oral Joint-Health Supplements in Horses. 2010. [Google Scholar]
- Deloitte, L. The Economic Impact of the Horse Industry on the United States; Consulting | American Horse Council: Washington, DC, 2005. [Google Scholar]
- Egenvall, A.; Bonnett, B.; Wattle, O.; Emanuelson, U. Veterinary-Care Events and Costs over a 5-Year Follow-up Period for Warmblooded Riding Horses with or without Previously Recorded Locomotor Problems in Sweden. Prev. Vet. Med. 2008, 83, 130–143. [Google Scholar] [CrossRef]
- Egenvall, A.; Bonnett, B.N.; Olson, P.; Penell, J.; Emanuelson, U. Association between Costly Veterinary-Care Events and 5-Year Survival of Swedish Insured Warmblooded Riding Horses. Prev. Vet. Med. 2006, 77, 122–136. [Google Scholar] [CrossRef]
- Penell, J.C.; Egenvall, A.; Bonnett, B.N.; Olson, P.; Pringle, J. Specific Causes of Morbidity among Swedish Horses Insured for Veterinary Care between 1997 and 2000. Vet. Rec. 2005, 157, 470–477. [Google Scholar] [CrossRef]
- Dörner, C.A. Locomotor Injuries Morbidity Data Analysis in Chilean Sport Horses: A Retrospective Study (2016-2021). Austral J. Vet. Sci. 2023, 55, 137–141. [Google Scholar] [CrossRef]
- Celeste, C.; Ionescu ’, M.; Poole ’, A.R.; Laverty, S. Repeated Intraarticular Injections of Triamcinolone Acetonide Alter Cartilage Matrix Metabolism Measured by Biomarkers in Synovial Fluid. J. Orthop. Res. 2005, 23, 602–610. [Google Scholar] [CrossRef]
- Menarim, B.C.; Gillis, K.H.; Oliver, A.; Mason, C.; Ngo, Y.; Werre, S.R.; Barrett, S.H.; Luo, X.; Byron, C.R.; Dahlgren, L.A. Autologous Bone Marrow Mononuclear Cells Modulate Joint Homeostasis in an Equine in Vivo Model of Synovitis. FASEB J. 2019, 33, 14337–14353. [Google Scholar] [CrossRef]
- Bondeson, J.; Wainwright, S.D.; Lauder, S.; Amos, N.; Hughes, C.E. The Role of Synovial Macrophages and Macrophage-Produced Cytokines in Driving Aggrecanases, Matrix Metalloproteinases, and Other Destructive and Inflammatory Responses in Osteoarthritis. Arthritis Res. Ther. 2006, 8. [Google Scholar] [CrossRef] [PubMed]
- Chaimbeul, S.F.; Rodrigues, N.N.P.; Thurston, D.D.; Scoggin, K.E.; Janes, J.; Jacobs, C.A.; MacLeod, J.N.; Stone, A. V.; Menarim, B.C. PPARγ Agonism Modulates Synovial Macrophage and Cartilage Responses in an Equine Model of Synovial Inflammation—Implications for Joint Therapy. Biomolecules 2025, 15. [Google Scholar] [CrossRef]
- Estrada Mcdermott, J.; Pezzanite, L.; Goodrich, L.; Santangelo, K.; Chow, L.; Dow, S.; Wheat, W. Role of Innate Immunity in Initiation and Progression of Osteoarthritis, with Emphasis on Horses. Animals 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, A.; Fearon, U.; Veale, D.J.; Godson, C. Macrophages in Synovial Inflammation. Front. Immunol. 2011, 2. [Google Scholar] [CrossRef]
- Fichadiya, A.; Bertram, K.L.; Ren, G.; Yates, R.M.; Krawetz, R.J. Characterizing Heterogeneity in the Response of Synovial Mesenchymal Progenitor Cells to Synovial Macrophages in Normal Individuals and Patients with Osteoarthritis. J. Inflamm. 2016, 13. [Google Scholar] [CrossRef]
- Menarim, B.; MacLeod, J.; Dahlgren, L. Bone Marrow Mononuclear Cells for Joint Therapy: The Role of macrophages in Inflammation Resolution and Tissue Repair. World J. Stem Cells 2021, 7, 825–840. [Google Scholar] [CrossRef]
- Mills, C.D. Anatomy of a Discovery: M1 and M2 Macrophages. Front. Immunol. 2015, 6. [Google Scholar] [CrossRef]
- Olingy, C.E.; San Emeterio, C.L.; Ogle, M.E.; Krieger, J.R.; Bruce, A.C.; Pfau, D.D.; Jordan, B.T.; Peirce, S.M.; Botchwey, E.A. Non-Classical Monocytes Are Biased Progenitors of Wound Healing Macrophages during Soft Tissue Injury. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Desai, B.N.; Leitinger, N. Purinergic and Calcium Signaling in Macrophage Function and Plasticity. Front. Immunol. 2014, 5. [Google Scholar] [CrossRef]
- Zumerle, S.; Calì, B.; Munari, F.; Angioni, R.; Di Virgilio, F.; Molon, B.; Viola, A. Intercellular Calcium Signaling Induced by ATP Potentiates Macrophage Phagocytosis. Cell Rep. 2019, 27, 1–10.e4. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, A.; Sun, Y.; Sukumaran, P.; Quenum Zangbede, F.O.; Jondle, C.N.; Sharma, A.; Evans, D.L.; Chauhan, P.; Szlabick, R.E.; Aaland, M.O.; et al. M1 Macrophage Polarization Is Dependent on TRPC1-Mediated Calcium Entry. iScience 2018, 8, 85–102. [Google Scholar] [CrossRef]
- Roselli, F.; Livrea, P.; Jirillo, E. Voltage-Gated Sodium Channel Blockers as Immunomodulators. Recent Pat. CNS Drug Discov. 2006, 1, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Black, J.A.; Waxman, S.G. Noncanonical Roles of Voltage-Gated Sodium Channels. Neuron 2013, 80, 280–291. [Google Scholar] [CrossRef]
- Oh-hora, M.; Rao, A. Calcium Signaling in Lymphocytes. Curr. Opin. Immunol. 2008, 20, 250–258. [Google Scholar] [CrossRef]
- Stokes, L.; MacKenzie, A.B.; Sluyter, R. Editorial: Roles of Ion Channels in Immune Cells. Front. Immunol. 2016, 7. [Google Scholar] [CrossRef]
- Lo, W.L.; Donermeyer, D.L.; Allen, P.M. A Voltage-Gated Sodium Channel Is Essential for the Positive Selection of CD4 + T Cells. Nat. Immunol. 2012, 13, 880–887. [Google Scholar] [CrossRef]
- Lee, P.Y.; Tsai, P.S.; Huang, Y.H.; Huang, C.J. Inhibition of Toll-like Receptor-4, Nuclear Factor-ΚB and Mitogen-Activated Protein Kinase by Lignocaine May Involve Voltage-Sensitive Sodium Channels. Clin. Exp. Pharmacol. Physiol. 2008, 35, 1052–1058. [Google Scholar] [CrossRef]
- Wilck, N.; Balogh, A.; Markó, L.; Bartolomaeus, H.; Müller, D.N. The Role of Sodium in Modulating Immune Cell Function. Nat. Rev. Nephrol. 2019, 15, 546–558. [Google Scholar] [CrossRef] [PubMed]
- Craner, M.J.; Damarjian, T.G.; Liu, S.; Hains, B.C.; Lo, A.C.; Black, J.A.; Newcombe, J.; Cuzner, M.L.; Waxman, S.G. Sodium Channels Contribute to Microglia/Macrophage Activation and Function in EAE and MS. Glia 2005, 49, 220–229. [Google Scholar] [CrossRef]
- Fraser, S.P.; Diss, J.K.J.; Lloyd, L.J.; Pani, F.; Chioni, A.M.; George, A.J.T.; Djamgoz, M.B.A. T-Lymphocyte Invasiveness: Control by Voltage-Gated Na+ Channel Activity. FEBS Lett. 2004, 569, 191–194. [Google Scholar] [CrossRef] [PubMed]
- Carrithers, M.D.; Dib-Hajj, S.; Carrithers, L.M.; Tokmoulina, G.; Pypaert, M.; Jonas, E.A.; Waxman, S.G. Expression of the Voltage-Gated Sodium Channel NaV1.5 in the Macrophage Late Endosome Regulates Endosomal Acidification. J. Immunol. 2007, 178, 7822–32. [Google Scholar] [CrossRef]
- Dorrington, M.G.; Fraser, I.D.C. NF-ΚB Signaling in Macrophages: Dynamics, Crosstalk, and Signal Integration. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef]
- Besson, P.; Driffort, V.; Bon, É.; Gradek, F.; Chevalier, S.; Roger, S. How Do Voltage-Gated Sodium Channels Enhance Migration and Invasiveness in Cancer Cells? Biochim. Biophys. Acta Biomembr. 2015, 1848, 2493–2501. [Google Scholar] [CrossRef] [PubMed]
- Feher, A.; Pócsi, M.; Papp, F.; Szanto, T.G.; Csoti, A.; Fejes, Z.; Nagy, B.; Nemes, B.; Varga, Z. Functional Voltage-Gated Sodium Channels Are Present in the Human B Cell Membrane. Cells 2022, 11. [Google Scholar] [CrossRef]
- Lahat, A.; Horin, S.B.; Lang, A.; Fudim, E.; Picard, O.; Chowers, Y. Lidocaine Down-Regulates Nuclear Factor-ΚB Signalling and Inhibits Cytokine Production and T Cell Proliferation. Clin. Exp. Immunol. 2008, 152, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Feske, S.; Wulff, H.; Skolnik, E.Y. Ion Channels in Innate and Adaptive Immunity. Annu. Rev. Immunol. 2015, 33, 291–353. [Google Scholar] [CrossRef]
- Sun, H.; Jiang, J.; Gong, L.; Li, X.; Yang, Y.; Luo, Y.; Guo, Z.; Lu, R.; Li, H.; Li, J.; et al. Voltage-Gated Sodium Channel Inhibitor Reduces Atherosclerosis by Modulating Monocyte/Macrophage Subsets and Suppressing Macrophage Proliferation. Biomed. Pharmacother. 2019, 120. [Google Scholar] [CrossRef] [PubMed]
- Montero, M.C.; del Campo, M.; Bono, M.; Simon, M.V.; Guerrero, J.; Lagos, N. Neosaxitoxin Inhibits the Expression of Inflammation Markers of the M1 Phenotype in Macrophages. Mar. Drugs 2020, 18, 1–17. [Google Scholar] [CrossRef]
- Sharif, O.; Bolshakov, V.N.; Raines, S.; Newham, P.; Perkins, N.D. Transcriptional Profiling of the LPS Induced NF-ΚB Response in Macrophages. BMC Immunol. 2007, 8. [Google Scholar] [CrossRef]
- Dörner, C.; Del Campo, M.; Lagos, N. Neosaxitoxin, a Long-Lasting Local Anesthetic and Its Potential Clinical Applications in Horses. Austral J. Vet. Sci. 2023, 55, 147–151. [Google Scholar] [CrossRef]
- Varela, X.; del Campo, M.; Piron, R.; Sepulveda, J.; Bustamante, T.; Hinzpeter, J.; Lagos, N. Effect of Neosaxitoxin on Epidural Anesthesia in Cats: A Promising Alternative to Conventional Anesthetics. Int. Physiol. J. 2019, 2, 4–10. [Google Scholar] [CrossRef]
- Valenzuela, C.; Torres, C.; Muñoz, V.; Simbaina, J.C.; Sánchez, A.; Bustamante, T.; Sepúlveda, J.M.; Piron, R.; Del Campo, M.; Lagos, N. Evaluation of Neosaxitoxin as a Local Anesthetic during Piglet Castration: A Potential Alternative for Lidocaine. Toxicon 2019, 164, 26–30. [Google Scholar] [CrossRef] [PubMed]
- Hinzpeter, J.; Zamorano, A.; Barahona, M.; Möller, G.; Espinoza, J.; del Campo, M.; Piron, R.; Sepúlveda, J.M.; Bustamante, T.; Lagos, N. Management of Arthrofibrosis of the Knee after an Arthroscopic Meniscectomy with Paralytic Shellfish Poisoning Toxin. Case Report. Int. Physiol. J. 2018, 1–6. [Google Scholar] [CrossRef]
- Hinzpeter, J.; Barrientos, C.; Zamorano, Á.; Martinez, Á.; Palet, M.; Wulf, R.; Barahona, M.; Sepúlveda, J.M.; Guerra, M.; Bustamante, T.; et al. Gonyautoxins: First Evidence in Pain Management in Total Knee Arthroplasty. Toxicon 2016, 119, 180–185. [Google Scholar] [CrossRef]
- Frisbie, D.D.; Kawcak, C.E.; McIlwraith, C.W.; Trotter, G.W.; Powers, B.E. Effects of Triamcinolone in an Equine in Vivo Osteochondral Fragment Model. Equine Vet. J. 1996, 29, 270. [Google Scholar]
- McIlwraith, C.W.; Frisbie, D.D.; Kawcak, C.E. The Horse as a Model of Naturally Occurring Osteoarthritis. Bone Jt. Res. 2012, 1, 297–309. [Google Scholar] [CrossRef]
- Foland, J.W.; Mcilwraith, C.W.; Acvs, D.; Trotter, G.W.; Powers, B.E.; Acw, D.; Lamar, C.H. Effect of Betamethasone and Exercise on Equine Carpal Joints With Osteochondral Fragments. Vet. Surg. 1994, 23, 369–76. [Google Scholar] [CrossRef] [PubMed]
- Frisbie, D.; Ghivizzani, S.; Robbins, P.; Evans, C.; McIlwraith, W. Treatment of Experimental Equine Osteoarthritis by in Vivo Delivery of the Equine Interleukin-1 Receptor Antagonist Gene. Gene Ther. 2002, 9, 12–20. [Google Scholar] [CrossRef]
- Kawcak, C.E.; Norrdin, R.W.; Frisbie, D.D.; Trotter, G.W.; Mcilwraith, C.W. Effects of Osteochondral Fragmentation and Intra-Articular Triamcinolone Acetonide Treatment on Subchondral Bone in the Equine Carpus. Equine Vet. J. 1998, 30, 66–71. [Google Scholar] [CrossRef]
- Lagos, N.; Onodera, H.; Zagatto, P.A.; Andrinolo, D.; Azevedo, S.M.F.Q.; Oshima, Y. The First Evidence of Paralytic Shellfish Toxins in the Freshwater Cyanobacterium Cylindrospermopsis Raciborskii, Isolated from Brazil. Toxicon 1999, 37, 1359–1373. [Google Scholar] [CrossRef]
- Dörner, C.; Lagos, N.; Oyaneder, L.; Menarim, B.C.; Ramírez-Toloza, G. Synovial Fluid Biomarker Profile After Intra-Articular Administration of Neosaxitoxin in Horses: A Feasibility Study. Animals 2025, 15, 2453. [Google Scholar] [CrossRef]
- Warner, K.; Lischer, C.J. Komplikationen Nach Intraartikularer Anwendung von ACS (I RAP®) Beim Pferd - Retrospektive Studie. Pferdeheilkunde 2017, 33, 356–362. [Google Scholar] [CrossRef]
- Warner, K.; Schulze, T.; Lischer, C.J. Behandlung von Osteoarthritis Mit ACS (IRAP®) Bei 26 Pferden-Retrospektive Studie. Pferdeheilkunde 2016, 32, 241–248. [Google Scholar] [CrossRef]
- Kawcak, C.E.; McIlwraith, C.W. Comparison of Synovial Fluid in Middle Carpal Joints Undergoing Needle Aspiration, Infusion with Saline, and Infusion with a Combination of N-Acetyl-d-Glucosamine, Hyaluronic Acid, and Sodium Chondroitin Sulfate. J. Equine Vet. Sci. 2011, 31, 155–159. [Google Scholar] [CrossRef]
- Wagner, A.E.; Mcilwraith, C. W.; Martin, G. S. Effect of Intra-Articular Injection of Orgotein and Saline Solution on Equine Synovia The Anti-Inflammatory Drug Orgotein Has Been Used Systemically, Locally, and Intra-Articularly in the Treatment of Inflammatory Conditions in the Horse. Am. J. Vet. Res. 1983, 43, 594–597. [Google Scholar]
- Vinod, E.; Jefferson, T.E.; Amirtham, S.M.; Prince, N.; Geevar, T.; Rebekah, G.; Ramasamy, B.; Kachroo, U. Correlation between Synovial Fluid Calcium Containing Crystal Estimation and Varying Grades of Osteoarthritis Created Using a Rabbit Model: Potential Diagnostic Tool. J. Clin. Orthop. Trauma 2020, 11, S506–S511. [Google Scholar] [CrossRef] [PubMed]
- Yavorskyy, A.; Hernandez-Santana, A.; Shortt, B.; McCarthy, G.; McMahon, G. Determination of Calcium in Synovial Fluid Samples as an Aid to Diagnosing Osteoarthritis. Bioanalysis 2010, 2, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.R.; Phelan, M.M.; Clegg, P.D.; Peffers, M.J.; Rubio-Martinez, L.M. Synovial Fluid Metabolites Differentiate between Septic and Nonseptic Joint Pathologies. J. Proteome Res. 2018, 17, 2735–2743. [Google Scholar] [CrossRef]
- McIlwraith, C.W.; Frisbie, D.D.; Kawcak, C.E.; Fuller, C.J.; Hurtig, M.; Cruz, A. The OARSI Histopathology Initiative - Recommendations for Histological Assessments of Osteoarthritis in the Horse. Osteoarthr. Cartil. 2010, 18, S93–S105. [Google Scholar] [CrossRef]
- Frisbie, D.; Al-Sobayil, F.; Billinghurst, R.; Kawcak, C.; McIlwraith, C.W. Changes in Synovial Fluid and Serum Biomarkers with Exercise and Early Osteoarthritis in Horses. Osteoarthr. Cartil. 2008, 16, 1196–1204. [Google Scholar] [CrossRef]
- Bolt, D.; Ishihara, A.; Weisbrode, S.; Bertone, A. Effects of Triamcinolone Acetonide, Sodium Hyaluronate, Amikacin Sulfate, and Mepivacaine Hydrochloride, Alone and in Combination, on Morphology and Matrix Composition of Lipopolysaccharide-challenged and Unchallenged Equine Articular Cartilage Explants. Am. J. Vet. Res. 2008, 69, 861–867. [Google Scholar] [CrossRef]
- Makrygiannakis, D.; Af Klint, E.; Catrina, S.B.; Botusan, I.R.; Klareskog, E.; Klareskog, L.; Ulfgren, A.K.; Catrina, A.I. Intraarticular Corticosteroids Decrease Synovial RANKL Expression in Inflammatory Arthritis. Arthritis Rheum. 2006, 54, 1463–1472. [Google Scholar] [CrossRef]
- McIlwraith, C.W. The Use of Intra-Articular Corticosteroids in the Horse: What Is Known on a Scientific Basis? Equine Vet. J. 2010, 42, 563–571. [Google Scholar]
- White, G.F.; Gómez Álvarez, C.B.; Lewis, R. Are Biologics More Effective than Corticosteroids for Intra-Articular Treatment of Osteoarthritis? Equine Vet. Educ. 2021, 33, 389–392. [Google Scholar] [CrossRef]
- Stone, S.; Malanga, G.A.; Capella, T. Topical Review Corticosteroids: Review of the History, the Effectiveness, and Adverse Effects in the Treatment of Joint Pain. Pain Physician 2021, S233–S246. [Google Scholar]
- Tokawa, P.K.A.; Baccarin, R.Y.A.; Zanotto, G.M. Intra-Articular Corticosteroids: Systematic Review of Effects of Multiple Injections and Different Doses. Equine Vet. Educ. 2025. [Google Scholar] [CrossRef]
- Pekarek, B.; Osher, L.; Buck, S.; Bowen, M. Intra-Articular Corticosteroid Injections: A Critical Literature Review with up-to-Date Findings. Foot 2011, 21, 66–70. [Google Scholar] [CrossRef] [PubMed]
- McAlindon, T.E.; LaValley, M.P.; Harvey, W.F.; Price, L.L.; Driban, J.B.; Zhang, M.; Ward, R.J. Effect of Intra-Articular Triamcinolone vs Saline on Knee Cartilage Volume and Pain in Patients with Knee Osteoarthritis a Randomized Clinical Trial. J. Am. Med. Assoc. 2017, 317, 1967–1975. [Google Scholar] [CrossRef]
- van Weeren, P.R.; de Grauw, J.C. Pain in Osteoarthritis. Vet. Clin. North Am. 2010, 26, 619–642. [Google Scholar] [CrossRef] [PubMed]
- de Souza, M.V. Osteoarthritis in Horses - Part 2: A Review of the Intraarticular Use of Corticosteroids as a Method of Treatment. Braz. Arch. Biol. Technol. 2016, 59, 1–10. [Google Scholar] [CrossRef]
- Sugimoto, T.; Yoshino, M.; Sato Nagao, M.; Ishii, S.; Yabu’, H. Voltage-Gated Ionic Channels in Cultured Rabbit Articular Chondrocytes. Comp. Biochem. Physiol. C. Pharmacol. Toxicol. Endocrinol. 1996, 115, 223–32. [Google Scholar] [CrossRef]
- Craner, M.J.; Damarjian, T.G.; Liu, S.; Hains, B.C.; Lo, A.C.; Black, J.A.; Newcombe, J.; Cuzner, M.L.; Waxman, S.G. Sodium Channels Contribute to Microglia/Macrophage Activation and Function in EAE and MS. Glia 2005, 49, 220–229. [Google Scholar] [CrossRef]
- Yongming, L.; Yizhe, X.; Zhikai, Q.; Yupeng, W.; Xiang, W.; Mengyuan, Y.; Guoqing, D.; Hongsheng, Z. Identification of Ion Channel-Related Genes as Diagnostic Markers and Potential Therapeutic Targets for Osteoarthritis through Bioinformatics and Machine Learning-Based Approaches. Biomarkers 2024, 29, 285–297. [Google Scholar] [CrossRef]
- Lewis, R.; Barrett-Jolley, R. Changes in Membrane Receptors and Ion Channels as Potential Biomarkers for Osteoarthritis. Front. Physiol. 2015, 6, 357. [Google Scholar] [CrossRef]
- Black, J.A.; Waxman, S.G. Noncanonical Roles of Voltage-Gated Sodium Channels. Neuron 2013, 80, 280–291. [Google Scholar] [CrossRef] [PubMed]
- Karnina, R.; Arif, S.K.; Hatta, M.; Bukhari, A.; Natzir, R.; Hisbullah; Patellongi, I.; Kaelan, C. Systemic Lidocaine Administration Influences NF-Kβ Gene Expression, NF-Kβ and TNF- α Protein Levels on BALB/c Mice with Musculoskeletal Injury. Ann. Med. Surg. 2021, 69. [Google Scholar] [CrossRef]
- Lahat, A.; Horin, S.B.; Lang, A.; Fudim, E.; Picard, O.; Chowers, Y. Lidocaine Down-Regulates Nuclear Factor-ΚB Signalling and Inhibits Cytokine Production and T Cell Proliferation. Clin. Exp. Immunol. 2008, 152, 320–327. [Google Scholar] [CrossRef]
- Feng, G.; Liu, S.; Wang, G.L.; Liu, G.J. Lidocaine Attenuates Lipopolysaccharide-Induced Acute Lung Injury through Inhibiting NF-ΚB Activation. Pharmacology 2007, 81, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Oyaneder, L.; Lagos, N.; Dörner, C. Systemic, Hemodynamic and Neurological Effects of Caudal Epidural Administration of Neosaxitoxin in Horses. Toxicon 2025, 257. [Google Scholar] [CrossRef]
- Rodriguez-Navarro, A.; Lagos, N.; Lagos, M.; Braghetto, I.; Csendes, A.; Hamilton, J.; Figueroa, C.; Truan, D.; Garcia, C.; Rojas, A.; et al. Neosaxitoxin as a Local Anesthetic. Anesthesiology 2007, 106, 339–345. [Google Scholar] [CrossRef]
- Manríquez, V.; Castro Caperan, D.; Guzmán, R.; et al. First Evidence of Neosaxitoxin as a Long-Acting Pain Blocker in Bladder Pain Syndrome. Int. Urogynecol. J. 2015, 26, 853–858. [Google Scholar] [CrossRef]
- Menarim, B.C.; Gillis, K.H.; Oliver, A.; Ngo, Y.; Werre, S.R.; Barrett, S.H.; Rodgerson, D.H.; Dahlgren, L.A. Macrophage Activation in the Synovium of Healthy and Osteoarthritic Equine Joints. Front. Vet. Sci. 2020, 7. [Google Scholar] [CrossRef]
- de Wall Malefyt, R.; Abrams, J.; Bennett, B.; Figdor, C.G.; De Vries, J.E. Interleukin 10(EL,.10) Inhibits Cytokine Synthesis by Human Monocytes: An Autoregulatory Role of IL-10 Produced by Monocytes. J. Exp.Med 1991, 174, 1209–1220. [Google Scholar] [CrossRef]
- Sugimoto, M.A.; Sousa, L.P.; Pinho, V.; Perretti, M.; Teixeira, M.M. Resolution of Inflammation: What Controls Its Onset? Front. Immunol. 2016, 7. [Google Scholar] [CrossRef]
- Leite, C.B.G.; Merkely, G.; Charles, J.F.; Lattermann, C. From Inflammation to Resolution: Specialized Pro-Resolving Mediators in Posttraumatic Osteoarthritis. Curr. Osteoporos. Rep. 2023, 21, 758–770. [Google Scholar] [CrossRef]
- van Helvoort, E.M.; van der Heijden, E.; van Roon, J.A.G.; Eijkelkamp, N.; Mastbergen, S.C. The Role of Interleukin-4 and Interleukin-10 in Osteoarthritic Joint Disease: A Systematic Narrative Review. Cartilage 2022, 13. [Google Scholar] [CrossRef]
Figure 1.
Schematic outlining the study design, treatment assignment and timing for model induction and sampling.
Figure 1.
Schematic outlining the study design, treatment assignment and timing for model induction and sampling.

Figure 2.
Clinical parameters following intra-articular treatments. Representative images of differences in joint flexion range between joints treated with saline and NeoSTX at day 10, respectively (A). Boxplots of the results with boxes representing interquartile range with a median and whiskers representing range, show changes in joint flexion angle (B), joint surface temperature (C), and joint circumference (D), at baseline and during the 30-day follow-up. NeoSTX and TA improved joint mobility, reduced swelling, and decreased surface temperature, whereas saline-treated horses treated with saline showed persistently reduced flexion, a greater joint circumference, and elevated temperature. * Statistical trend between NeoSTX and TA (p = 0.1). p-value = 0.05.
Figure 2.
Clinical parameters following intra-articular treatments. Representative images of differences in joint flexion range between joints treated with saline and NeoSTX at day 10, respectively (A). Boxplots of the results with boxes representing interquartile range with a median and whiskers representing range, show changes in joint flexion angle (B), joint surface temperature (C), and joint circumference (D), at baseline and during the 30-day follow-up. NeoSTX and TA improved joint mobility, reduced swelling, and decreased surface temperature, whereas saline-treated horses treated with saline showed persistently reduced flexion, a greater joint circumference, and elevated temperature. * Statistical trend between NeoSTX and TA (p = 0.1). p-value = 0.05.

Figure 3.
Principal component analysis (PCA) of synovial biomarkers after intra-articular treatments. Cellular, biochemical markers, cytokine and growth factor cluster. Pro-inflammatory variables considered to be important due to it vector length and were contributing to group difference were TNF-α, IL-1β, IL-6, IL-18, IFN-γ, IL-2, IL-4, IL-10, IL-13, FGF-2. Additionally, PMN, Mononuclear cells, red blood cells and Total Proteins were important cellular and metabolic contributors to group differences. NeoSTX-treated horses clustered closer to baseline, maintaining a stable and balanced synovial profile and key markers related with pro-resolving properties (mononuclear cells, MCP-1) were aligned with with this group. Saline controls were associated with pro-inflammatory mediators, chemokines and cells (e.g., neutrophils, red blood cells, IP-10, MCP-1), while triamcinolone separated distinctly, driven by strong upregulation of pro-inflammatory (TNF-α, IL-1β, IL-6, IL-18, IFN-γ, IL-2) and anti-inflammatory (IL-4, IL-10).
Figure 3.
Principal component analysis (PCA) of synovial biomarkers after intra-articular treatments. Cellular, biochemical markers, cytokine and growth factor cluster. Pro-inflammatory variables considered to be important due to it vector length and were contributing to group difference were TNF-α, IL-1β, IL-6, IL-18, IFN-γ, IL-2, IL-4, IL-10, IL-13, FGF-2. Additionally, PMN, Mononuclear cells, red blood cells and Total Proteins were important cellular and metabolic contributors to group differences. NeoSTX-treated horses clustered closer to baseline, maintaining a stable and balanced synovial profile and key markers related with pro-resolving properties (mononuclear cells, MCP-1) were aligned with with this group. Saline controls were associated with pro-inflammatory mediators, chemokines and cells (e.g., neutrophils, red blood cells, IP-10, MCP-1), while triamcinolone separated distinctly, driven by strong upregulation of pro-inflammatory (TNF-α, IL-1β, IL-6, IL-18, IFN-γ, IL-2) and anti-inflammatory (IL-4, IL-10).

Figure 4.
Synovial fluid cellular composition. Boxplots (median, IQR, range) for RBC (A), TNCC (B), neutrophils (%) (C), neutrophils (D), mononuclear cells (%) (E), and mononuclear cells (F) at baseline and days 10, 20, and 30. NeoSTX maintained lower RBC and neutrophil counts compared with saline and triamcinolone. Triamcinolone induced transient TNCC elevations and a mononuclear cell increase at day 30, while saline controls exhibited sustained RBC and neutrophil increases. p-value = 0.05.
Figure 4.
Synovial fluid cellular composition. Boxplots (median, IQR, range) for RBC (A), TNCC (B), neutrophils (%) (C), neutrophils (D), mononuclear cells (%) (E), and mononuclear cells (F) at baseline and days 10, 20, and 30. NeoSTX maintained lower RBC and neutrophil counts compared with saline and triamcinolone. Triamcinolone induced transient TNCC elevations and a mononuclear cell increase at day 30, while saline controls exhibited sustained RBC and neutrophil increases. p-value = 0.05.

Figure 5.
Synovial fluid inflammatory markers concentration following intra-articular treatments. Boxplots of the results with boxes representing interquartile range, with a median and whiskers representing range of changes in in total proteins (A), IL-1β (B), TNF-α (C), and IL-6 (D), concentrations at baseline and days 10, 20, and 30 after OA induction. Horses treated with NeoSTX exhibited significantly lower total protein levels compared with those treated with saline. NeoSTX-treated horses maintained consistently lower levels of pro-inflammatory cytokines (IL-1β, IL-6, TNF-α) compared with saline and triamcinolone groups. NeoSTX and Triamcinolone showed suppression of inflammation nonetheless, NeoSTX sowed lower and stable cytokine concentrations over the study. p-value = 0.05.
Figure 5.
Synovial fluid inflammatory markers concentration following intra-articular treatments. Boxplots of the results with boxes representing interquartile range, with a median and whiskers representing range of changes in in total proteins (A), IL-1β (B), TNF-α (C), and IL-6 (D), concentrations at baseline and days 10, 20, and 30 after OA induction. Horses treated with NeoSTX exhibited significantly lower total protein levels compared with those treated with saline. NeoSTX-treated horses maintained consistently lower levels of pro-inflammatory cytokines (IL-1β, IL-6, TNF-α) compared with saline and triamcinolone groups. NeoSTX and Triamcinolone showed suppression of inflammation nonetheless, NeoSTX sowed lower and stable cytokine concentrations over the study. p-value = 0.05.

Figure 6.
Synovial fluid of key chemokines and anti-inflammatory cytokines concentrations following intra-articular treatments. Boxplots of the results with boxes representing interquartile range with a median and whiskers representing range of changes in MCP-1 (A), IL-4 (B), IL-10 (C) at baseline and days 10, 20, and 30 after OA induction. After reaching a peak at day 10, NeoSTX-treated horses maintained a downregulation of MCP-1 compared with saline- and TA- treated groups. IL-4 and IL-10 showed a downregulation in NeoSTX treated joints being significant than TA group at day 20 and 30. p-value = 0.05.
Figure 6.
Synovial fluid of key chemokines and anti-inflammatory cytokines concentrations following intra-articular treatments. Boxplots of the results with boxes representing interquartile range with a median and whiskers representing range of changes in MCP-1 (A), IL-4 (B), IL-10 (C) at baseline and days 10, 20, and 30 after OA induction. After reaching a peak at day 10, NeoSTX-treated horses maintained a downregulation of MCP-1 compared with saline- and TA- treated groups. IL-4 and IL-10 showed a downregulation in NeoSTX treated joints being significant than TA group at day 20 and 30. p-value = 0.05.

Figure 7.
(A) Synovial fluid collection of NeoSTX- treated joint, showing normal macroscopic characteristics. (B) and (C) Representative (subjects 8 and 12) postmortem middle carpal joint images of NeoSTX- treated joints characterized by less swelling in the synovium and peri-articular and intra-articular hemorrhage. No major changes were evidenced in the cartilage surfaces. (D) Comparison synovial fluid gross appearance between the TA- and NeoSTX- treated joints and (G) between Saline- and NeoSTX- treated joints. There is some blood contamination in the NeoSTX sample, probably due to blood contamination during the procedure. Observe the reddish and scarce amount of synovial fluid in the Triamcinolone group, this outcome was constantly observed in the Triamcinolone group after repeated treatment administration. (E, F, H, and I) Depicting macroscopically inflamed synovium characterized by swelling (black arrowheads) and periarticular and intra-articular hemorrhage in joints treated with TA and Saline respectively (white arrowheads).
Figure 7.
(A) Synovial fluid collection of NeoSTX- treated joint, showing normal macroscopic characteristics. (B) and (C) Representative (subjects 8 and 12) postmortem middle carpal joint images of NeoSTX- treated joints characterized by less swelling in the synovium and peri-articular and intra-articular hemorrhage. No major changes were evidenced in the cartilage surfaces. (D) Comparison synovial fluid gross appearance between the TA- and NeoSTX- treated joints and (G) between Saline- and NeoSTX- treated joints. There is some blood contamination in the NeoSTX sample, probably due to blood contamination during the procedure. Observe the reddish and scarce amount of synovial fluid in the Triamcinolone group, this outcome was constantly observed in the Triamcinolone group after repeated treatment administration. (E, F, H, and I) Depicting macroscopically inflamed synovium characterized by swelling (black arrowheads) and periarticular and intra-articular hemorrhage in joints treated with TA and Saline respectively (white arrowheads).

Figure 8.
Representative hematoxylin and eosin images of the synovial membrane depicting differences in histologic parameters between NeoSTX (A, B, and C), Triamcinolone (D, E, and F), and Saline (G, H, and I) treated joints after 30 days of OA induction and treatment administration. NeoSTX-treated joints exhibited lower scores for all histologic aspects of inflammation, although these differences were not statistically significant. Increase of inflammatory cellular infiltration in Triamcinolone- and Saline-treated synovial membrane (white arrows) in F and I, x400. Hyperplasia of the intimal layer (*) in D and E, x40. Fibrosis of the subintimal collagen fibers (white arrow heads) in G and H, x100. OARSI, Osteoarthritis Research Society International.
Figure 8.
Representative hematoxylin and eosin images of the synovial membrane depicting differences in histologic parameters between NeoSTX (A, B, and C), Triamcinolone (D, E, and F), and Saline (G, H, and I) treated joints after 30 days of OA induction and treatment administration. NeoSTX-treated joints exhibited lower scores for all histologic aspects of inflammation, although these differences were not statistically significant. Increase of inflammatory cellular infiltration in Triamcinolone- and Saline-treated synovial membrane (white arrows) in F and I, x400. Hyperplasia of the intimal layer (*) in D and E, x40. Fibrosis of the subintimal collagen fibers (white arrow heads) in G and H, x100. OARSI, Osteoarthritis Research Society International.

Figure 9.
Representative hematoxylin and eosin images of hyaline cartilage depicting differences in histologic parameters between NeoSTX (A, B, and C), Triamcinolone (D, E, and F), and Saline (G, H, and I) treated joints after 30 days of OA induction (carpal chip model) and treatment administration. NeoSTX-treated joints exhibited lower scores for all histologic aspects of cartilage degradation, although these differences were not statistically significant. Cartilage fissures (white arrowheads) and higher chondrocyte focal loss (*) were observed in triamcinolone- and saline- treated joints, D, G, x40 and E, H, x100. Chondrocyte necrosis near the articular surface is more evident in the Triamcinolone and Saline groups (black arrows), as shown in C, F, and I (x400). Additionally, decreased H&E staining uptake was observed primarily in Saline-treated horses (white arrows), as shown in G and I. OARSI, Osteoarthritis Research Society International.
Figure 9.
Representative hematoxylin and eosin images of hyaline cartilage depicting differences in histologic parameters between NeoSTX (A, B, and C), Triamcinolone (D, E, and F), and Saline (G, H, and I) treated joints after 30 days of OA induction (carpal chip model) and treatment administration. NeoSTX-treated joints exhibited lower scores for all histologic aspects of cartilage degradation, although these differences were not statistically significant. Cartilage fissures (white arrowheads) and higher chondrocyte focal loss (*) were observed in triamcinolone- and saline- treated joints, D, G, x40 and E, H, x100. Chondrocyte necrosis near the articular surface is more evident in the Triamcinolone and Saline groups (black arrows), as shown in C, F, and I (x400). Additionally, decreased H&E staining uptake was observed primarily in Saline-treated horses (white arrows), as shown in G and I. OARSI, Osteoarthritis Research Society International.

Figure 10.
Representative Safranin-O images of hyaline cartilage and subchondral bone depicting differences in histologic parameters between NeoSTX (A), Triamcinolone (B), and Saline (C) treated joints after 30 days of OA induction and treatment administration. Cartilage from joints treated with NeoSTX exhibited preserved proteoglycan content, depicted by red staining, while those treated with Triamcinolone showed proteoglycan loss from the superficial layer of the cartilage, and horses treated with saline showed significantly proteoglycan loss throughout the cartilage.
Figure 10.
Representative Safranin-O images of hyaline cartilage and subchondral bone depicting differences in histologic parameters between NeoSTX (A), Triamcinolone (B), and Saline (C) treated joints after 30 days of OA induction and treatment administration. Cartilage from joints treated with NeoSTX exhibited preserved proteoglycan content, depicted by red staining, while those treated with Triamcinolone showed proteoglycan loss from the superficial layer of the cartilage, and horses treated with saline showed significantly proteoglycan loss throughout the cartilage.

Figure 11.
Proposed mechanisms underlying the effects of intra-articular administration of NeoSTX. (A) NeoSTX blocks voltage-gated sodium channels (NaV), reducing neuronal impulse propagation and joint pain. (B) At the cellular level, blockade of Na+ influx and Na+/Ca2+ transport downregulates intracellular calcium signaling, thereby inhibiting NF-κB activation and reducing the release of pro-inflammatory cytokines (IL-1, IL-6, TNF-α) and chemokine signaling. (C) In cartilage, NeoSTX decreases extracellular matrix degradation (proteoglycans and collagen), limits chondrocyte apoptosis, and reduces fibrillation. (D) In the synovial membrane, NeoSTX attenuates hypertrophy, hyperplasia, angiogenesis, and possibly neuronal proliferation (related to NGF). Collectively, these effects contribute to the recovery of joint homeostasis by modulating cellular responses in cartilage and synovium. Red: Previous research for proposed mechanism [22,26,27,38,40,71,72,80].
Figure 11.
Proposed mechanisms underlying the effects of intra-articular administration of NeoSTX. (A) NeoSTX blocks voltage-gated sodium channels (NaV), reducing neuronal impulse propagation and joint pain. (B) At the cellular level, blockade of Na+ influx and Na+/Ca2+ transport downregulates intracellular calcium signaling, thereby inhibiting NF-κB activation and reducing the release of pro-inflammatory cytokines (IL-1, IL-6, TNF-α) and chemokine signaling. (C) In cartilage, NeoSTX decreases extracellular matrix degradation (proteoglycans and collagen), limits chondrocyte apoptosis, and reduces fibrillation. (D) In the synovial membrane, NeoSTX attenuates hypertrophy, hyperplasia, angiogenesis, and possibly neuronal proliferation (related to NGF). Collectively, these effects contribute to the recovery of joint homeostasis by modulating cellular responses in cartilage and synovium. Red: Previous research for proposed mechanism [22,26,27,38,40,71,72,80].

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



