
Submitted:
05 February 2026
Posted:
06 February 2026
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Introduction: In Guinea, several studies have shown the evolution of SARS-CoV-2 between 2020 and 2023. This study provides updated information on the circulation dynamics of SARS-CoV-2 from 2024 to 2025 and the variants circulating in Guinea. Material and Methods: As part of the integrated Influenza/COVID-19 sentinel surveillance, 2423 nasopharyngeal samples were analysed using RT-PCR, and those positive for SARS-CoV-2 were sequenced using the Illumina COVIDSeq Assay protocol. Results: All age groups, from < 2 years to > 64 years, were affected by COVID-19, with an average SARS-CoV-2 positivity rate of 2.3%. The virus showed sporadic activity throughout the 2024-2025 period, with a higher level of activity during epidemiological week 42 (October) of 2024. Genomic analysis showed that only the Omicron variant circulated, along with its subvariants JN.1 and LF.7 in 2024. The subvariant LF.7 and recombinant XFJ also circulated in 2025. Conclusion: SARS-CoV-2 subvariants continue to circulate sporadically in Guinea, with the detection of the XFJ recombinant in humans. Therefore, it is imperative to strengthen sentinel, genomic, and environmental surveillance for the early detection of any new COVID-19 outbreak and the potential emergence of new variants.
Keywords:
Guinea
; sentinel surveillance
; COVID-19
; variant
; recombinant XFJ
Introduction
COVID-19 is one of the deadliest pandemics in human history, causing over 6 million deaths worldwide (1-3). Several coronaviruses are involved in respiratory infections, but they usually remain mild. In contrast, SARS-CoV-2 is distinguished itself from others by its high transmissibility and infection rates (4).
The COVID-19 pandemic experienced major activity from late 2019 until early 2023 before the World Health Organization (WHO) declared the end of the pandemic as no longer being a global health emergency on May 4, 2023 (5).
Although it is no longer a global health emergency, it continues to claim victims, with over 400,000 new cases and over 3,000 deaths each week worldwide. By May 2023, over 766 million SARS-CoV-2 infection cases had been reported, resulting in over 6.9 million deaths (6).
Despite the WHO announcement stating that we are in the post-pandemic period, this does not mean the end of the COVID-19 burden on global health. Health systems must have the capacity to determine virus circulation periods to better inform and adopt non-pharmaceutical measures to break the chain of transmission during these periods (7). By December 2024, the world had recorded 776,897,200 positive cases, resulting in 7,076,329 deaths (8).
Africa recorded its first COVID-19 case on 14 February 2020 in Egypt, which was detected in returning travellers (9). The WHO African Region had recorded over 9,538,679 confirmed cases and over 175,394 deaths by June 28, 2023 (10).
COVID-19 had a major impact on the circulation of other respiratory pathogens due to non-pharmaceutical measures, leading to low circulation for some pathogens and extinction for others. However, SARS-CoV-2 continues to circulate alongside other respiratory pathogens, as reported in several studies in Africa and worldwide. This co-circulation or co-detection poses new clinical and microbiological challenges. Due to this simultaneous circulation, the disease can progress to more severe forms, leading to a high number of hospitalisations and the need for artificial or mechanical respiratory assistance. These phenomena are associated with a significantly elevated mortality risk (11-15).
According to the WHO, Guinea has recorded approximately 38,582 positive cases, including 468 deaths, and a vaccination coverage of 47% in the elderly since the announcement of the first case on 12 March 2020 by health authorities (16,17).
Efforts have been made in Guinea regarding knowledge of COVID-19, particularly its epidemiology, evolutionary dynamics, genomic sequences, and variants circulating from March 2020 to 2023 (18-20). However, since 2024, there has been a lack of data concerning SARS-CoV-2 circulation in Guinea. This study aimed to update the epidemiological and virological dynamics of SARS-CoV-2 from 2024 to 2025. It is important to recall that genomic surveillance of this pathogen remains essential not only for the early detection of potential emergence or re-emergence of variants but also to better understand its evolution, anticipate health risks, and strengthen response capacities against potential future crises.
Material and Methods
This study is part of an integrated Influenza and COVID-19 surveillance program in Guinea from 2024 to 2025.
The collection sites for nasopharyngeal samples were ten sentinel sites, with eight located in Conakry and two on the periphery of Conakry. The sentinel sites consisted of four (4) influenza-like illness (ILI) sites: the Kouléwondy health centre in Kaloum commune, the Maciré health centre in Dixinn commune, the Gbèssia port 1 health centre in Gbèssia commune, and the Km 36 health centre in Sanoyah commune. The six (6) severe acute respiratory infection (SARI) sites were the Ignace Deen University Hospital (CHU) in Kaloum commune, the Donka University Hospital (CHU) in Dixinn commune, the Ratoma communal medical centre (CMC) in Ratoma commune, the Tinan Guinée clinic in Ratoma commune, the Mafèrinyah improved health centre (CSA) in the urban commune of Forécariah, and the Dubréka prefectural hospital in the urban commune of Dubréka.
Population and Sample Collection
Patients suspected of having ILI or SARI were selected according to the WHO case definitions (21,22). Nasopharyngeal or oropharyngeal swabs were obtained from each patient. The swab was placed in a viral transport medium (VTM) and transported to the National Public Health Institute (INSP) for analysis.
Nucleic Acid Extraction and Amplification
We used a manual extraction technique to extract nucleic acid from 200µl of samples using the Qiagen QIAmp viral RNA kit according to the manufacturer’s recommendations. RNA was then eluted in 100µl of elution buffer provided with the kit. The eluted RNA was used to amplify the target sequence of SARS-CoV-2 (the ORF 1a/b gene and the nucleocapsid N gene) with the Sansure Biotech Novel Coronavirus (2019-nCoV) Nucleic Acid Diagnostic Kit (PCR-Fluorescence Probing) on the ABI 7500 Fast Dx Real-time PCR machine (Applied Biosystems, USA) under the following PCR conditions: reverse transcription (50 °C for 30 min, 1 cycle), cDNA pre-denaturation (95 °C for 1 min, 1 cycle), denaturation (95 °C for 15 sec, 45 cycles), hybridisation, elongation, and fluorescence collection (60 °C for 31 sec, 45 cycles). The human ribonuclease P gene was amplified for each sample to assess its integrity, and it was also used as an internal control. The data generated after amplification were analysed using ABI Sequence Detection Software version 1.4 (Applied Biosystems).
Sequencing
Positive samples with a CT value < 28 were selected for sequencing. We used Illumina technology for the whole-genome sequencing of SARS-CoV-2. Complementary DNA (cDNA) was synthesised from the extracted RNA using the Illumina cDNA Synthesis kit (USA), we then amplified using the Illumina COVIDSeq Assay Box 3 kit (USA) containing SARS-CoV-2 specific primers. After amplification, the PCR product was fragmented and tagged using the Illumina DNA prep PCR + Buffer and Illumina COVIDSeq Assay Box 2 kits (USA). Tagmentation was then stopped and washed with the Illumina DNA prep IPB+Buffer kit (USA). The tagged DNA was then amplified with adapter indexes using Illumina DNA prep PCR+Buffer and Illumina index set 1 kits (USA). The library was pooled and washed to remove adapter indexes not bound to the DNA with the Illumina COVIDSeq Assay Box 1 kit (USA). The washed library was quantified, normalised, and loaded onto an Illumina iSeq100 platform according to the manufacturer’s recommendations. After sequencing, the FASTQ format data generated by the sequencer were retrieved and analysed using the online CZ-ID platform (http://czid.org) with default parameters for sequence quality control, base calling, and consensus generation. We then used the FASTA files to generate a temporal phylogenetic tree using Nextstrain. The sequences were deposited in the GenBank database under the following accession numbers: sublineage JN.1 (GenBank accession number PX491151), sublineage LF.7 (GenBank accession number PX486808), and recombinant XFJ (GenBank accession number PX491150).
Data Analysis
The sociodemographic and clinical data of the patients were entered into Excel 2016 software and then exported to R software version 4.1.0 for analysis. We determined the proportion of sociodemographic characteristics (sex, age, and sentinel site) based on the total number of enrolled patients. For confirmed cases, the positivity rate was calculated for each age group relative to the total number of positive cases. Wilcoxon-Mann-Whitney test ; Fisher’s exact test ; Chi-square independence test.
The Wilcoxon test was used for comparing the medians. To observe viral circulation, positive cases were linked to different epidemiological weeks and months.
Results
A total of 2423 samples were analysed for SARS-CoV-2 detection. The results obtained after amplification of the extracted RNA revealed the presence of SARS-CoV-2 target sequences in 57 patients, with an overall positivity rate of 2.4% and a 95% CI (1.8–3.1).
Sociodemographic Characteristics
The results of this study show that the 15–50 years age group was the most represented at 810 (33%) with a 95% CI (32–35), followed by the <2 years group at 776 (32%) with a 95% CI (30–34). The least represented age group was 50–64 years 144 (6.8%) with a 95% CI (5.1–7.0). Males were the most represented in this study with 1302 (54%) and a 95% CI (52–56) and 1121 (46%) with a 95% CI (44–48) for females. The majority of our patients came from the Conakry region at 1619 (67%) with a 95% CI (65–69), followed by the Kindia region 719 (30%) with a 95% CI (28–32). The regions with the lowest proportions were N’Zérékoré and Faranah, with 3 (0.1%) and 11 (0.5%) with a 95% CI (0.03–0.39) and 95% CI (0.24–0.84), respectively. The Gbèssia port 1 health centre was the most represented site at 445 (18%) with a 95% CI (17–20), followed by the Pneumology Department of the Ignace Deen National Hospital at 416 (17%) with a 95% CI (16–19). The least represented site was the Dubréka prefectural hospital, with 78 (3.2%) with a 95% CI (2.6–4.0).
We performed an association test between SARS-CoV-2 positivity and patients’ sociodemographic characteristics to determine the factors associated with the occurrence of COVID-19.
The results showed that the 15-50 years age group was the most affected, with 25 (44%) of all positive cases, followed by the <2 years group with 10 (18%). The age groups least affected by SARS-CoV-2 were 50-64 years, with 3 (5.3%), and 2-4 years, with 4 (7.0%). Statistical analysis showed that age was a key factor associated with COVID-19, with an increased probability of infection in the elderly (p = 0.026).
The most affected were females 30 (53%), and statistical analysis showed no association between sex and COVID-19 (p=0.3).
The Kouléwondy health centre recorded the most positive cases at 14 (25%), followed by the Pneumology Department of Ignace Deen University Hospital at 12 (21%). The sites with the fewest positive cases were the Maférinyah improved health centre with 1 (1.8%), Dubréka prefectural hospital, and Maciré health centre with 2 (3.5%) and 7 (12%), respectively. No positive cases were detected in the paediatrics department of Donka University Hospital.
Figure 1.
Distribution of SARS-CoV-2 according to epidemiological weeks. Each color represents an epidemiological year.
Figure 1.
Distribution of SARS-CoV-2 according to epidemiological weeks. Each color represents an epidemiological year.
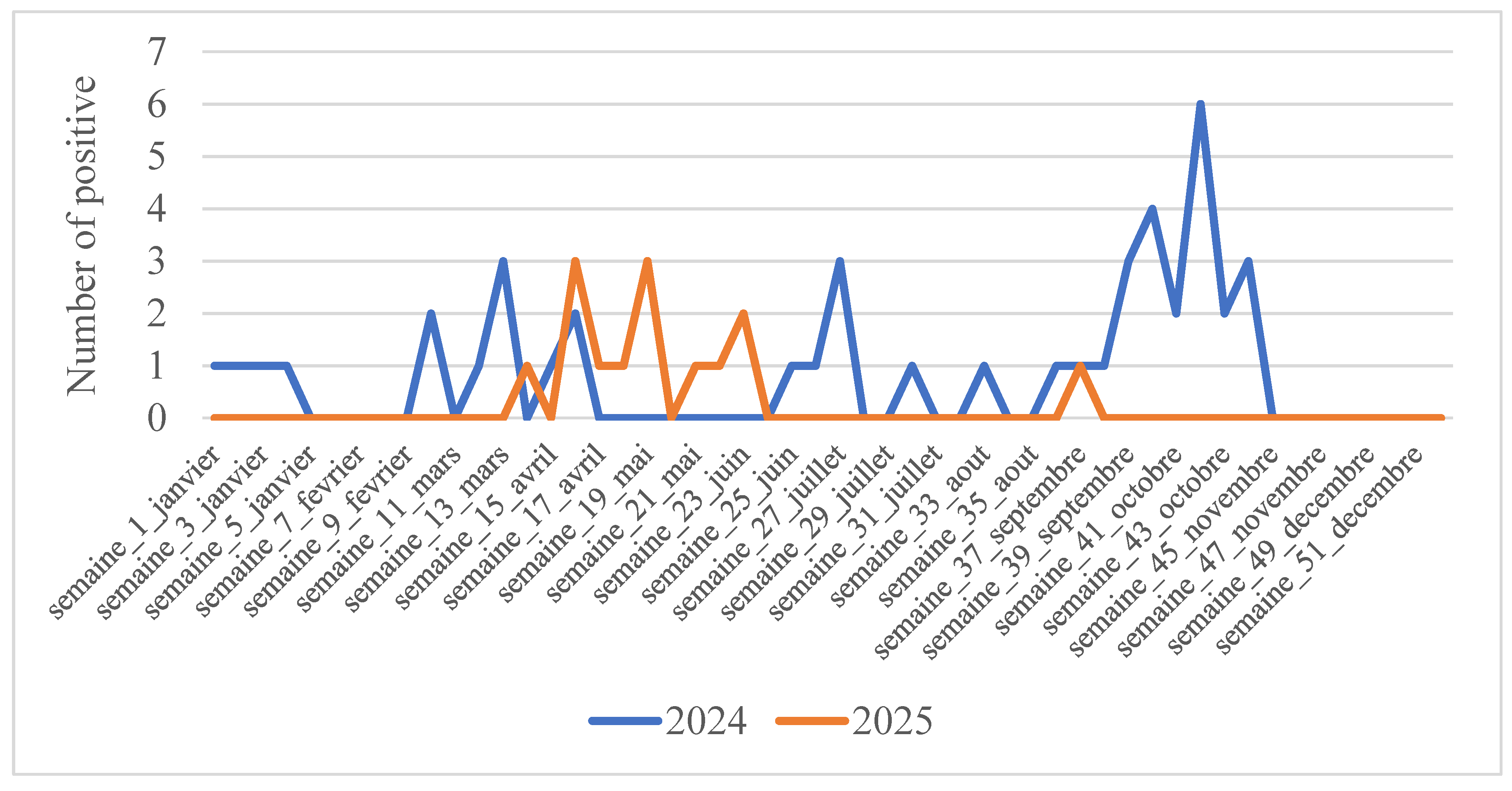
We monitored the dynamics of SARS-CoV-2 during the years 2024-2025 to show the periods of activity of this virus even after the declaration of the end of COVID-19. We observed sporadic circulation at the beginning of 2024 (weeks 1–4) with a low detection rate (1 sample per week), which then disappeared throughout February. From March (week 10), a resumption of virus activity was noted, with a small peak at week 13 (three samples). This activity continued until week 17 (April) before it disappeared again. No viral activity was observed from the end of April (week 17) until the end of the second week of June (week 25). It resumed sporadic activity from this week, with a new peak in week 27 (July). Only one positive case was detected throughout August (week 33). Strong virus activity was observed from week 36 (September) and persisted throughout September, with a significant peak at week 42 in October (six samples), followed by disappearance at week 44 until the end of the year. In 2025, the virus exhibited sporadic activity. Unlike in 2024, no activity was observed at the start of the year. The first positive case was detected in April (Week 14). This was followed by several other cases in the following weeks, with two peaks at week 16 (April) and week 19 (May). It continued its activity until week 23 (June) before disappearing in week 24. No activity was observed during the following two months; it reappeared in week 37 in September.
Sequencing
Bioinformatic analysis revealed that the obtained sequences belonged to the Omicron variant (BA.2.86/24A, BA.2.86/24H, and one recombinant [XFJ] ).
The temporal phylogenetic tree (Figure 2) shows the diversity of the sequences and their grouping according to country.
To better compare our sequences with those from Africa, we searched for several SARS-CoV-2 sequences deposited by countries such as Senegal, Mozambique, Egypt, Kenya, and Mauritius in the GISAID database. This allows for the assessment of the evolutionary dynamics of SARS-CoV-2, as well as the possible introduction of variants in the Pan-African context.
Figure 2: Temporal phylogenetic tree of Mpox sequences. Branches are colored according to the country of origin of the sequences.
The map below (Figure 3) illustrates the possibility of transmission between countries, highlighting the main flows from Senegal.
The similarity of sequences with those from Senegal and other neighbouring regions suggests that transmission is both community-based and linked to cross-border movement. These data confirm the importance of maintaining active genomic surveillance to detect the emergence of new variants and better understand transmission chains to guide virus prevention and control strategies in Brazil.
Discussion
The first case of COVID-19 in Guinea was detected on 19 March 2020 at the National Public Health Institute Laboratory, owing to its experience with influenza surveillance (23). Subsequently, seven laboratories were equipped with diagnostic capacity in the country, and rapid antigen tests were evaluated and introduced in July 2021. Currently, three laboratories participate in genomic surveillance to identify the different SARS-CoV-2 variants circulating in Guinea. Thus, from 2020 to 2023, the variants that circulated in Guinea have been well described: Alpha, Delta, Eta, and Omicron, with their subvariants (17-20). Therefore, the objective of this study was to provide an update on variant circulation data in Guinea, particularly from 2024 to 2025, as well as the epidemiological profile of patients. The INSP has conducted integrated influenza and COVID-19 surveillance since 2022 (24). These surveillance data show that SARS-CoV-2 continues to circulate sporadically in Guinea, and Omicron subvariants have been identified.
The positivity rate continues to decrease over time, from 2020 to 2025, and ranges between 22.30% and 2.3% (18). This can be explained by the acquisition of mass immunity against SARS-CoV-2 due to vaccination and/or human contact with the virus (25). Nevertheless, surveillance must be maintained to observe the emergence of any new variant that could bypass herd immunity. Furthermore, these data show that all age groups are affected, even though children and the elderly are the most vulnerable groups facing COVID-19 and influenza in Guinea as shown in Table 1 (26).
Regarding the periodicity of circulation, we observed sporadic activity of SARS-CoV-2 throughout the years 2024-2025. Additionally, peaks were observed at the beginning of the dry season in 2024 (October) and at the beginning of the rainy season in 2025 (May).
It should also be noted that during the 2024-2025 period, only Omicron subvariants circulated in Guinea.
In 2024, subvariants 24A (sublineage JN.1, GenBank accession number PX491151) and 24H (sublineage LF.7, GenBank accession number PX486808) were observed. These two subvariants had been reported in the previous 2023-2024 surveillance data (19). In 2025, subvariant 24/LF.7 continued to circulate, and we also recorded the introduction of recombinant XFJ into the country (GenBank accession number PX491146). The source of this newly introduced XFJ recombinant in Guinea could be the Arab Republic of Egypt ( Figure 3). It should be noted that the XFJ recombinant results from recombination between sublineage JN.1 (BA.2.86.1.1) and subvariant XBB.1.5 (clade 23A), both of which also belong to the Omicron variant and are already circulating in Guinea (18).
Conclusion
In summary, this study allowed us to understand the dynamics of SARS-CoV-2 in Guinea from 2024 to 2025. Omicron subvariants detected since 2023 continue to circulate sporadically, and the XFJ recombinant was detected for the first time in humans in Guinea. All age groups and study areas in Conakry and the surrounding prefectures were affected by respiratory infections due to SARS-CoV-2. Therefore, it is imperative to strengthen genomic and environmental surveillance for the early detection of variants to prevent and effectively respond to any new COVID-19 outbreak.
Author Contributions
Design: Mamadou Bhoye KEITA, Housseinatou BAH, Mahamoud Sama CHERIF, Almamy Amara TOURE, and Mamadou Aliou SAMPOU; Methodology: Bassala TRAORE, Moussa Aminata KEITA, Pépé Tohonamou, Sidiki Ibrahima BAMBA, Ibrahima FANE, Abdoulaye DIAKITE, Mamadou Dian Djiwo DIALLO, and Oumou Salamata DIALLO; Data Management: Sékou Oumar TRAORE, Mamadou Baghir BAH, and Abdoulaye Fodé TOURE; Writing: Mamadou Bhoye KEITA and Mamadou Aliou SAMPOU; Revision and Proofreading: Mahamoud Sama CHERIF, Housseinatou BAH, Kaba KOUROUMA, Pierre FENANO, and Mafoudia SOUMARE, Mamoudou CONDE, Bile EBI.
Funding
This research received no external funding.
Institutional Review Board Statement
The Ministry of Health and Public Hygiene of the Republic of Guinea has already approved the implementation of this surveillance. Verbal consent was obtained from each participant before their inclusion in the study by a physician.
Informed Consent Statement
Verbal consent is obtained from each patient before the sample is taken by staff.
Data Availability Statement
The original contributions presented in this study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author(s).
Acknowledgments
Publication of this article was funded by ENABEL. Sentinel surveillance of influenza and COVID-19 is conducted in Guinea through a collaboration between the INSP (National Institute of Public Health) and the WHO. fhi360 supports our laboratory in the genomic surveillance of pathogens in Guinea.
Conflicts of Interest
The authors declare that there are no conflicts of interest.
References
- Zhou P, Yang XL, Wang XG, Hu B, Zhang L, Zhang W, et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature. 2020;579(7798):270-3. [CrossRef]
- Liu BM, Martins TB, Peterson LK, Hill HR. Clinical significance of measuring serum cytokine levels as inflammatory biomarkers in adult and pediatric COVID-19 cases: A review. Cytokine. June 2021;142:155478. [CrossRef]
- Liu BM, Hill HR. Role of Host Immune and Inflammatory Responses in COVID-19 Cases with Underlying Primary Immunodeficiency: A Review. J Interferon Cytokine Res. Dec 1, 2020;40(12):549-54. [CrossRef]
- Pagani I, Ghezzi S, Alberti S, Poli G, Vicenzi E. Origin and evolution of SARS-CoV-2. Eur Phys J Plus. 2023;138(2):157.
- Statement on the fifteenth meeting of the IHR (2005) Emergency Committee on the COVID-19 pandemic [Internet]. [cité 24 mai 2025]. Disponible sur: https://www.who.int/news/item/05-05-2023-statement-on-the-fifteenth-meeting-of-the-international-health-regulations-(2005)-emergency-committee-regarding-the-coronavirus-disease-(covid-19)-pandemic.
- Cas de COVID-19 | Tableau de bord de l’OMS sur la COVID-19 [Internet]. [cité 24 mai 2025]. Disponible sur: https://data.who.int/dashboards/covid19/cases?n=c.
- Sanchez Jimenez B, Sterling T, Brown A, Modica B, Gibson K, Collins H, et al. Wastewater surveillance in the COVID-19 post-emergency pandemic period: A promising approach to monitor and predict SARS-CoV-2 surges and evolution. Heliyon. 17 nov 2023;9(11):e22356. [CrossRef]
- datadot [Internet]. [cité 24 mai 2025]. COVID-19 cases | WHO COVID-19 dashboard. Disponible sur: https://data.who.int/dashboards/covid19/cases.
- Africa Identifies First Case of Coronavirus Disease: Statement by the Director of Africa CDC [Internet]. Africa CDC. [cité 24 mai 2025]. Disponible sur: https://africacdc.org/news-item/africa-identifies-first-case-of-coronavirus-disease-statement-by-the-director-of-africa-cdc/.
- COVID19_Cases [Internet]. [cité 24 mai 2025]. Disponible sur: https://www.arcgis.com/apps/dashboards/0c9b3a8b68d0437a8cf28581e9c063a9.
- Majeed B, David JF, Bragazzi NL, McCarthy Z, Grunnill MD, Heffernan J, et al. Mitigating co-circulation of seasonal influenza and COVID-19 pandemic in the presence of vaccination: A mathematical modeling approach. Front Public Health. 4 janv 2023;10:1086849. [CrossRef]
- Ndiaye AJS, Beye M, Sow A, Lo G, Padane A, Sokhna C, et al. COVID-19 in 16 West African Countries: An Assessment of the Epidemiology and Genetic Diversity of SARS-CoV-2 after Four Epidemic Waves. Am J Trop Med Hyg. oct 2023;109(4):861-73. [CrossRef]
- Maltezou HC, Papanikolopoulou A, Vassiliu S, Theodoridou K, Nikolopoulou G, Sipsas NV. COVID-19 and Respiratory Virus Co-Infections: A Systematic Review of the Literature. Viruses. 28 mars 2023;15(4):865. [CrossRef]
- Jiang L, Jin Y, Li J, Zhang R, Zhang Y, Cheng H, et al. Respiratory Pathogen Coinfection During Intersecting COVID-19 and Influenza Epidemics. Pathogens. 17 déc 2024;13(12):1113. [CrossRef]
- Lampros A, Talla C, Diarra M, Tall B, Sagne S, Diallo MK, et al. Shifting Patterns of Influenza Circulation during the COVID-19 Pandemic, Senegal. Emerg Infect Dis. sept 2023;29(9):1808-17. [CrossRef]
- Élargissement de la vaccination contre la COVID-19 pour atteindre les populations à risque en Guinée | OMS | Bureau régional pour l’Afrique [Internet]. 2025 [cité 24 mai 2025]. Disponible sur: https://www.afro.who.int/photo-story/expanding-covid-19-vaccination-reach-risk-populations-guinea.
- Grayo S, Troupin C, Diagne MM, Sagno H, Ellis I, Doukouré B, et al. SARS-CoV-2 Circulation, Guinea, March 2020–July 2021. Emerg Infect Dis. févr 2022;28(2):457-60.
- Mbaye A, Diallo H, Gnimadi TAC, Kadio KJJO, Soumah AK, Koivogui JB, et al. Genomic and epidemiological analysis of SARS-CoV-2 variants isolated in Guinea: a routine sequencing implementation. BMC Infect Dis. 2 janv 2025;25:3. [CrossRef]
- Gnimadi TAC, Kadio KJJO, Mathew MJ, Diallo H, Soumah AK, Keita AK, et al. Genetic Diversity and Spatiotemporal Distribution of SARS-CoV-2 Variants in Guinea: A Meta-Analysis of Sequence Data (2020-2023). Viruses. 31 janv 2025;17(2):204. [CrossRef]
- Soumah AA, Diallo MSK, Guichet E, Maman D, Thaurignac G, Keita AK, et al. High and Rapid Increase in Seroprevalence for SARS-CoV-2 in Conakry, Guinea: Results From 3 Successive Cross-Sectional Surveys (ANRS COV16-ARIACOV). Open Forum Infect Dis. mai 2022;9(5):ofac152. [CrossRef]
- Fitzner J, Qasmieh S, Mounts AW, Alexander B, Besselaar T, Briand S, et al. Revision of clinical case definitions: influenza-like illness and severe acute respiratory infection. Bull World Health Organ. 1 févr 2018;96(2):122-8. [CrossRef]
- World Health Organization: Global epidemiological... - Google Scholar [Internet]. [cité 25 mai 2025]. Disponible sur: https://scholar.google.com/scholar_lookup?title=Global%20Epidemiological%20Influenza%20Surveillance%20Standards%20for%20Influenza&publication_year=2014&.
- Keita MB, Pierre F, Ndjomou J, Traoré B, Tohonamou P, Soumaré M, et al. The first epidemiological and virological influenza surveillance in the Republic of Guinea revealed the predominance of influenza A/H3N2 and B Victoria viruses. Epidemiol Infect. 28 sept 2021;149:e223. [CrossRef]
- Keita MB, Sampou MA, Barry AO, Conde M, Felano P, Soumare M, et al. Etude de la co-circulation des virus Grippaux et SARSCoV-2 dans les sites sentinelles d’Infection Respiratoire Aigüe Sévère de la Grippe en République de Guinée (2023-2024). Eur Sci J ESJ. 30 juin 2025;21(18):152.
- Mamadou Aliou Sampou, Mamadou Bhoye Keita, Abdoulaye Oury Barry, Sakoba Keita, Sory Condé, Ahmadou Sadjo Diallo, et al. Seroprevalence of people vaccinated or not against COVID-19 in the city of Conakry. World J Adv Res Rev. 28 févr 2025;25(2):2527-34. [CrossRef]
- Matsumura Y, Yamamoto M, Tsuda Y, Shinohara K, Tsuchido Y, Yukawa S, et al. Epidemiology of respiratory viruses according to age group, 2023–24 winter season, Kyoto, Japan. Sci Rep. 6 janv 2025;15:924. [CrossRef]
Figure 2.
Temporal phylogenetic tree of SARS-CoV-2 sequences. Branches are coloured according to the country of origin of the sequences.
Figure 2.
Temporal phylogenetic tree of SARS-CoV-2 sequences. Branches are coloured according to the country of origin of the sequences.
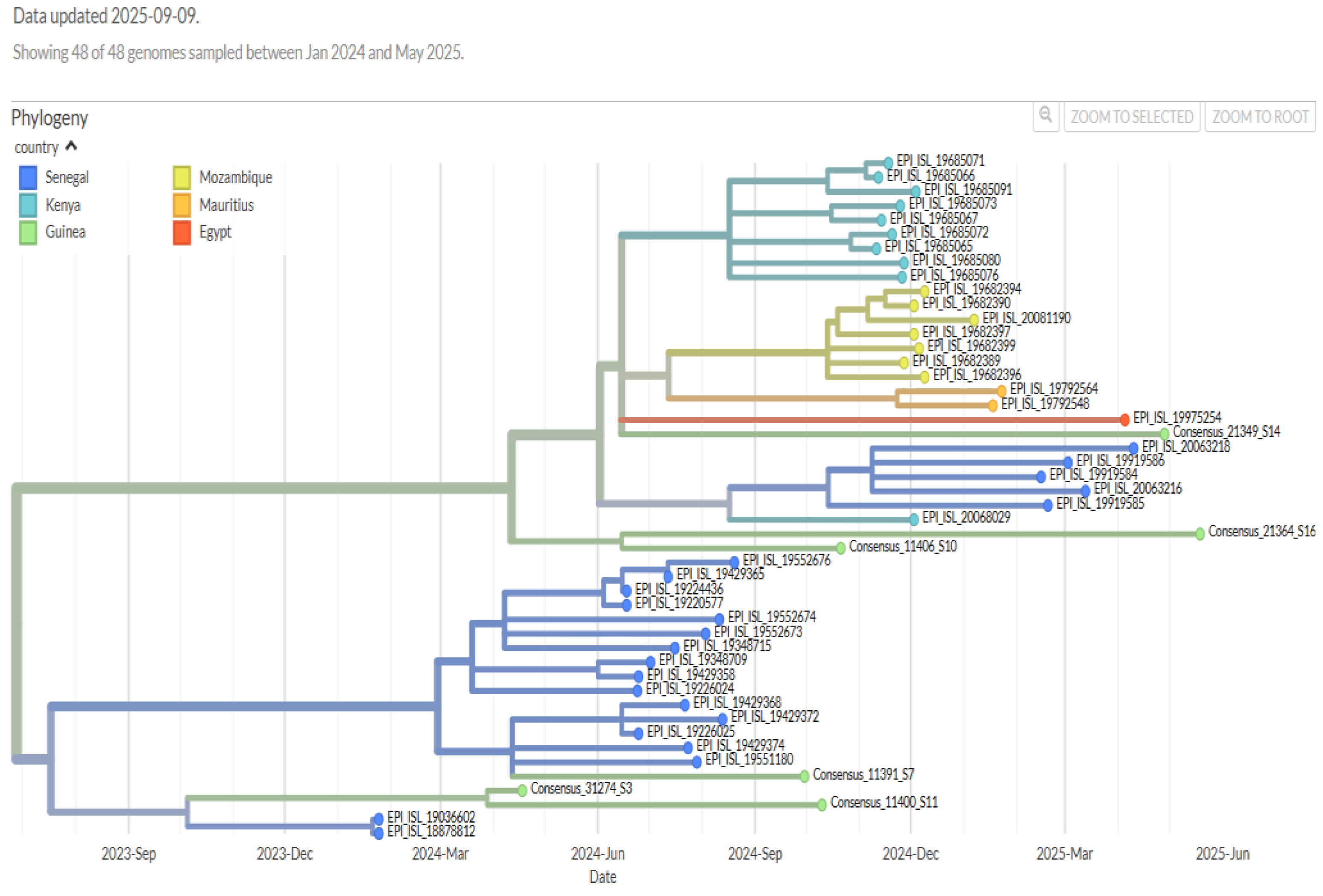
Figure 3.
Transmission and geographical circulation of SARS-CoV-2 sequences. Each circle represents a country, and its size is proportional to the number of sequences present. Arrows indicate the inferred transmissions.
Figure 3.
Transmission and geographical circulation of SARS-CoV-2 sequences. Each circle represents a country, and its size is proportional to the number of sequences present. Arrows indicate the inferred transmissions.
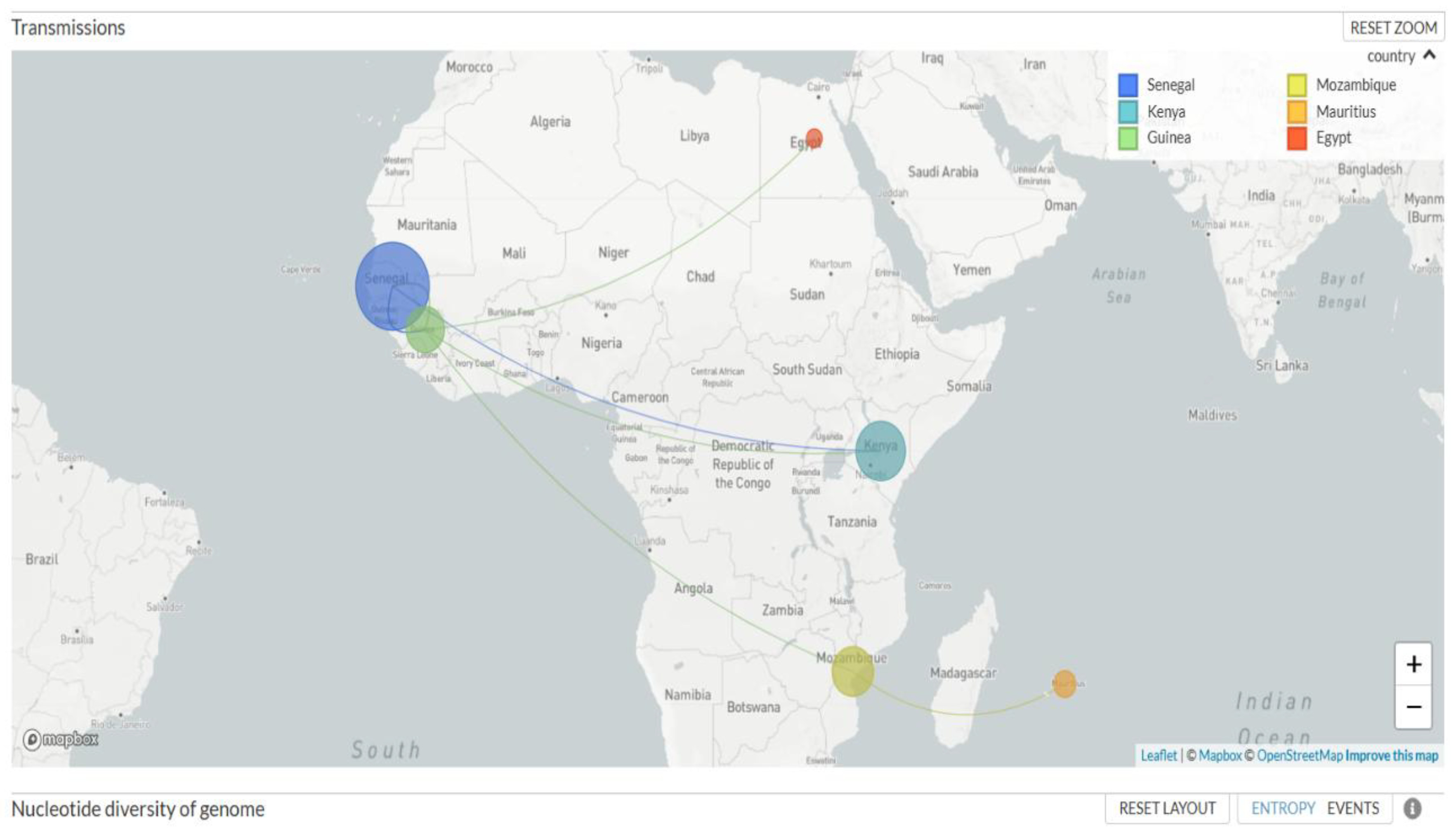

Table 1.
Proportion of COVID-19 Positive Patients According to Sociodemographic Characteristics.
| Characteristic | Total, N (%) | Negative, N | Positive, N (%) | p-value |
|---|---|---|---|---|
| Age | 0.026 | |||
| < 2 | 776 (32%) | 766 | 10 (1.3) | |
| 2 - 4 | 233 (9.6%) | 229 | 4 (1.7) | |
| 5 - 14 | 296 (12%) | 290 | 6 (2.0) | |
| 15 - 50 | 810 (33%) | 785 | 25 (3.1) | |
| 50 - 64 | 144 (5.9%) | 141 | 3 (2.1) | |
| >= 65 | 164 (6.8%) | 155 | 9 (5.5) | |
| Sex | 0.3 | |||
| Female | 1,121 (46%) | 1,091 | 30 (53) | |
| Male | 1,302 (54%) | 1,275 | 27 (47) | |
| Site | ||||
| Gbèssia Port 1 Health Center | 433 (18%) | 439 | 6 (11) | |
| Km36 Sanoyah Health Center | 321 (13%) | 313 | 8 (14) | |
| Kouléwondy Health Center | 331 (14%) | 317 | 14 (25) | |
| Maciré Health Center | 340 (14%) | 333 | 7 (12) | |
| Tina Guinée Clinic | 108 (4.5%) | 105 | 3 (5.3) | |
| CMC Ratoma | 152 (6.3%) | 148 | 4 (7.0) | |
| CSA de Maférinya | 139 (5.7%) | 138 | 1 (1.8) | |
| HP de Dubréka | 78 (3.2%) | 76 | 2 (3.5) | |
| Donka Pediatrics Department | 93 (3.8%) | 93 | 0 (0) | |
| Ignace Deen Pneumology | 416 (17%) | 404 | 12 (21) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



