Submitted:
17 December 2025
Posted:
17 December 2025
You are already at the latest version
Abstract
Ionizing radiation affects enzymes, which are critical for most cellular functions, by inducing chemical alterations in their molecular structures, often resulting in the inhibition of their activities. Understanding the molecular and kinetic mechanisms underlying these effects requires suitable experimental protocols and models tailored to specific enzymes and their substrates. In this study, we present a convenient experimental approach utilizing a medical linear accelerator (LINAC) suite as a precise means for irradiating enzyme solutions. Additionally, we examine the effects of ionizing radiation on the catalytic activities and potential structural changes of two enzymes: invertase (β-fructofuranosidase) and collagenase.
Keywords:
invertase
; collagenase
; enzyme kinetics
; ionizing radiation
; medical linear accelerator (LINAC)
1. Introduction
Ionizing radiation plays a pivotal role in modern medicine, fundamentally reshaping both diagnostics and therapeutics [1,2]. Its utility spans advanced imaging techniques—including roentgenography, computed tomography (CT), and positron emission tomography (PET)—and targeted treatments for malignant and benign pathologies via radiation oncology, radiosurgery, and radionuclide therapy [3,4,5]. Parallel advancements in radioprotection have been essential to these clinical applications. Contemporary research remains focused on optimizing the therapeutic index, striving to enhance clinical outcomes while strictly ensuring safety protocols [6].
The increasing collective radiation dose, exacerbated by technogenic pollutants and pharmaceutical interactions, contributes to synergistic biological toxicity. The specific outcomes of these combined exposures remain obscured by the mechanistic complexity of the factors involved. Consequently, radiobiological research has evolved significantly since the 1950s, with contemporary methodologies now integrating advanced concepts to characterize these multifactorial interactions [3].
Parallel advancements in radiotherapy technology have transitioned from standard high-energy photon sources—such as X-ray tubes and -emitting radioisotopes—to sophisticated particle acceleration systems. The integration of radiofrequency acceleration, microwave generators (magnetrons, klystrons), and waveguides has enabled the development of linear accelerators (LINACs) and microtrons. These devices facilitate the precise delivery of photons and electrons in the MeV energy range.
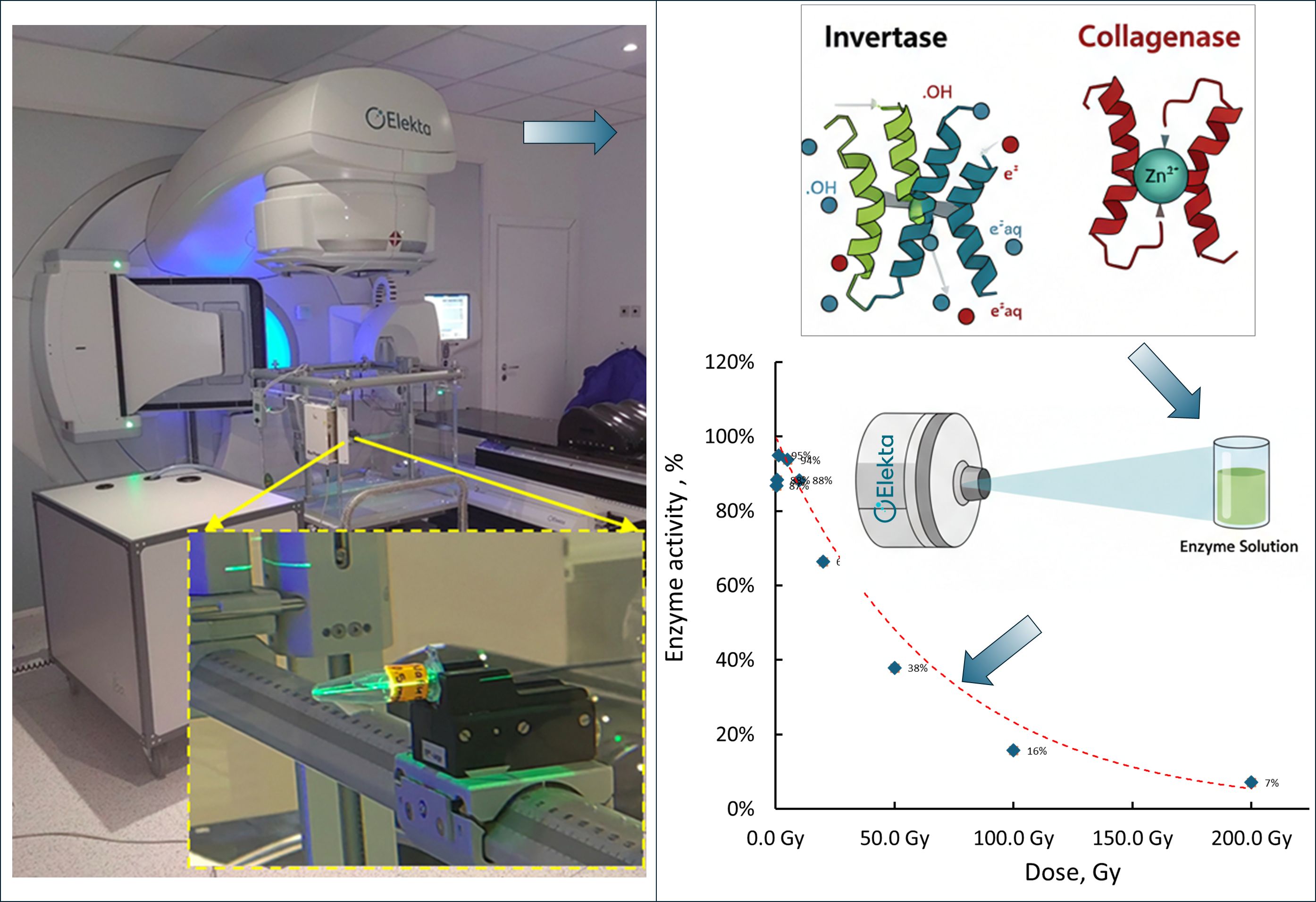
Nowadays LINACs are the most commonly used and most universal generators of ionizing radiation used in external beam radiotherapy. The concept of waveguides and linear accelerators originated in the 1920s, but their medical application became prominent post-World War II, when the Varian brothers advanced microwave technology for particle acceleration using radio frequency [7]. In linear accelerators, electrons are generated by an electron gun, a cathode-anode system that emits electrons from a heated tungsten filament, similar to an X-ray tube. However, instead of being accelerated to an anode target in a single accelerating section by some accelerating (anode) voltage, the electrons are injected in a waveguide and accelerated by intense electromagnetic fields in microwave cavities. The microwave energy is delivered in short-(microseconds) pulses from a klystron or a magnetron. This approach enables acceleration of electrons to relativistic velocities and very high energies with suitable waveguide structures. Medical LINACs produce monoenergetic 4 - 25 MeV electron pencil beams, that can either be used for irradiation with electrons (further applying scattering method, e.g., scattering foils, and collimation for achieving the needed irradiation field dimensions) or, most commonly, for irradiation with high-energy X-rays (directing the electron pencil beam at a high-density (usually tungsten) target. The output photons have a spectrum of energies up to and including the accelerated electrons energy. To form the desired irradiation fields sets of collimators are used, including the now standard multileaf collimators allowing for the creation of very complex radiation fields. The radiation output is monitored with a set of transmission ionization chambers in the radiation beam’s way in the treatment head. LINACs are calibrated in Monitor Units (MUs) in such a way that 1 MU corresponds to certain dose in water at specific reference conditions. Modern LINACs, coupled with state-of-the art dose planning and dosimetric systems allow for precisely controlled and verifiable irradiation and provide significant advantages for accurately irradiating biological samples. (Figure 1). For this study, we utilized a LINAC to expose enzyme solution samples to precisely defined and verified doses of ionizing radiation, ensuring controlled, reproducible conditions, allowing for reliable evaluation of the effects of ionizing radiation on enzyme activity.
Over the past 30 years, the radiation chemistry of proteins, particularly enzymes, has led to numerous applications which include improvements in experimental protocols as well as the development of theoretical and experimental approaches for studying the structure-function relationships in proteins [8]. When exposed to ionizing radiation, the chemical changes in protein molecules arise from both direct and indirect effects. In solutions, indirect effects dominate, while direct effects are negligible [9,10]. Conversely, in lyophilized states or frozen aqueous solutions, proteins are ionized primarily through direct interactions [11,12]. In aqueous solutions, the changes in the structure of protein molecules are due to secondary effects and the fact that proteins react very effectively with primary water free radicals, which are products of radiolysis [13]. The radiolysis of water is well studied, mainly using the pulse radiolysis method, where short electron pulses produce solvated (hydrated) electrons, cations, and excited water molecules (), which are its primary products [14]. The radiolysis products and free radicals, particularly solvated (hydrated) electrons and hydroxyl radicals , react with protein targets, leading to modifications or fragmentation of the polypeptide chain and its amino acid residues [15,16]. Hydrated electrons are known to react with the carbonyl groups of peptide bonds, causing chain cleavage and the release of ammonia through reactions occurring at the free radical level [16]. Ultimately, radiolytic reactions and their effects result in the formation of final products that alter the structures of protein molecules and, consequently, their functions.
The impact of radiation on enzymes, even when the dose is controlled, is not a selective method for the occurrence of chemically active radicals. However, it significantly contributes to the study of the catalytic and kinetic properties of enzymes, which are particularly important in modern pharmaceutical chemistry. The method of radiation inactivation of enzymes is a fundamental approach for investigating the effects of ionizing radiation on enzymes involved in key metabolic pathways, and ultimately for assessing the impact on the human body [15,17].
A key concept in enzymology (bio-catalysis) is the existence of a finite number of permissible conformational equilibrium states for any enzyme, which occur under strictly defined external conditions. Under low-dose irradiation, conformational changes in the enzyme molecule and its active site are thermodynamically reversible [18]. These changes can be controlled by precisely defining the irradiation conditions. Such modifications may occur without loss of catalytic activity or with cyclic variations in activity. However, over time, these changes can ultimately lead to the gradual unfolding of the protein molecule, resulting in an eventual loss of the enzyme's catalytic function [15,18].
The foundation of the present work is the concept that conformational changes induced by ionizing radiation disrupt the balance of thermodynamically and kinetically stable enzyme structures. This imbalance elicits a chemical response from enzymes to adapt to their new environment while attempting to preserve their structure and enzymatic activity. The number of permissible conformations and the degree of protection are determined by the enzyme's specific function. At higher doses of radiation, however, these conformational changes become irreversible, leading to complete denaturation of the molecule, including its active site, and resulting in irreversible inactivation [15].
Numerous enzymes are known to be profoundly affected by radiation-induced damage, yet only a few have been extensively studied under exposure to ionizing radiation [19]. Among those investigated, proteases have received particular attention, primarily due to their critical role in protein processing [15,17]. Collagenases (EC 3.2.24.x) are proteolytic enzymes that break down proteins by hydrolyzing peptide bonds. These enzymes target a major component of the extracellular matrix, the collagen, and degrade it into smaller peptides [20]. Invertase (β-fructofuranosidase, sucrase, EC 3.2.1.26) is an enzyme that belongs to the class of glycosylated enzymes that catalyze the hydrolysis of sucrose into glucose and fructose [21]. Both collagenases and invertase have been previously studied in radiological investigations. The study by Drózdz et al. examined the effects of gamma-ray exposure on collagen fractions and enzyme activity in rats over 30 days, reporting a significant reduction in total collagen content and changes in collagenase and cathepsin activity [22]. Separately, Nagrani and Bisby investigated the radiation inactivation of yeast invertase, finding that in dilute aqueous solutions, the rate of inactivation increased with radiation dose, linked to the reactive radical species formed during water radiolysis [23]. In our recent study [24], we utilized polarimetric methods to investigate the effects of Co-60 ionizing radiation on invertase activity. By determining kinetic parameters, we observed that the values of irradiated enzymes were approximately four times higher than those of unirradiated ones. This significant increase suggests that radiation induces chemical modifications resembling competitive inhibitors, thereby reducing enzyme-substrate binding affinity and catalytic efficiency. Additionally, the ratio further illustrated the diminished overall efficiency of the irradiated invertase.
This article has two distinctive aims. First, it proposes an experimental protocol using a medical linear accelerator (LINAC) suite to achieve homogeneous irradiation of enzyme solutions, enabling precise dose calculation, delivery, and verification. Linear accelerators offer significant advantages over radiation sources like Co-60 for precise dose control in irradiation applications. Unlike the fixed and decaying source of Co-60, LINACs generate X-ray beams through the controlled acceleration of electrons, allowing for dynamic adjustment of beam energy, dose rate, and field shape [25,26]. This is achieved through a series of coordinated subsystems, including a magnetron to precisely control radiofrequency waves and electron injection, as well as quadrupole steering magnets and focusing coils that refine the electron beam to a minimal diameter. Beam shaping is further accomplished by a multi-leaf collimator (MLC), shown schematically in Figure 3, which adaptively modulates the spatial distribution of the X-ray field through the independent motion of multiple tungsten leaves. Crucially, the LINAC incorporates dual independent ionization chambers for continuous, real-time dose monitoring, with an integrated computer system providing synchronized control of all parameters and immediate beam termination upon reaching the prescribed dose. This comprehensive control over beam generation, shaping, and real-time dosimetry ensures superior accuracy, reproducibility, and safety, making LINACs invaluable for applications requiring precise and verifiable radiation delivery, which is often lacking in simpler irradiation setups.
This approach facilitates kinetic studies of enzyme-catalyzed reactions post-irradiation, providing insights into structural and functional changes caused by ionizing radiation. The protocol also allows for the acquisition of kinetic data for reactions catalyzed by enzymes pre-irradiated with varying doses of ionizing radiation, shedding light on potential changes in the active site or molecular structure of the enzymes.
Second, the study investigates how ionizing radiation, delivered in different doses using LINAC, affects the catalytic activities and potentially the molecular structures of two enzymes: invertase and collagenase.
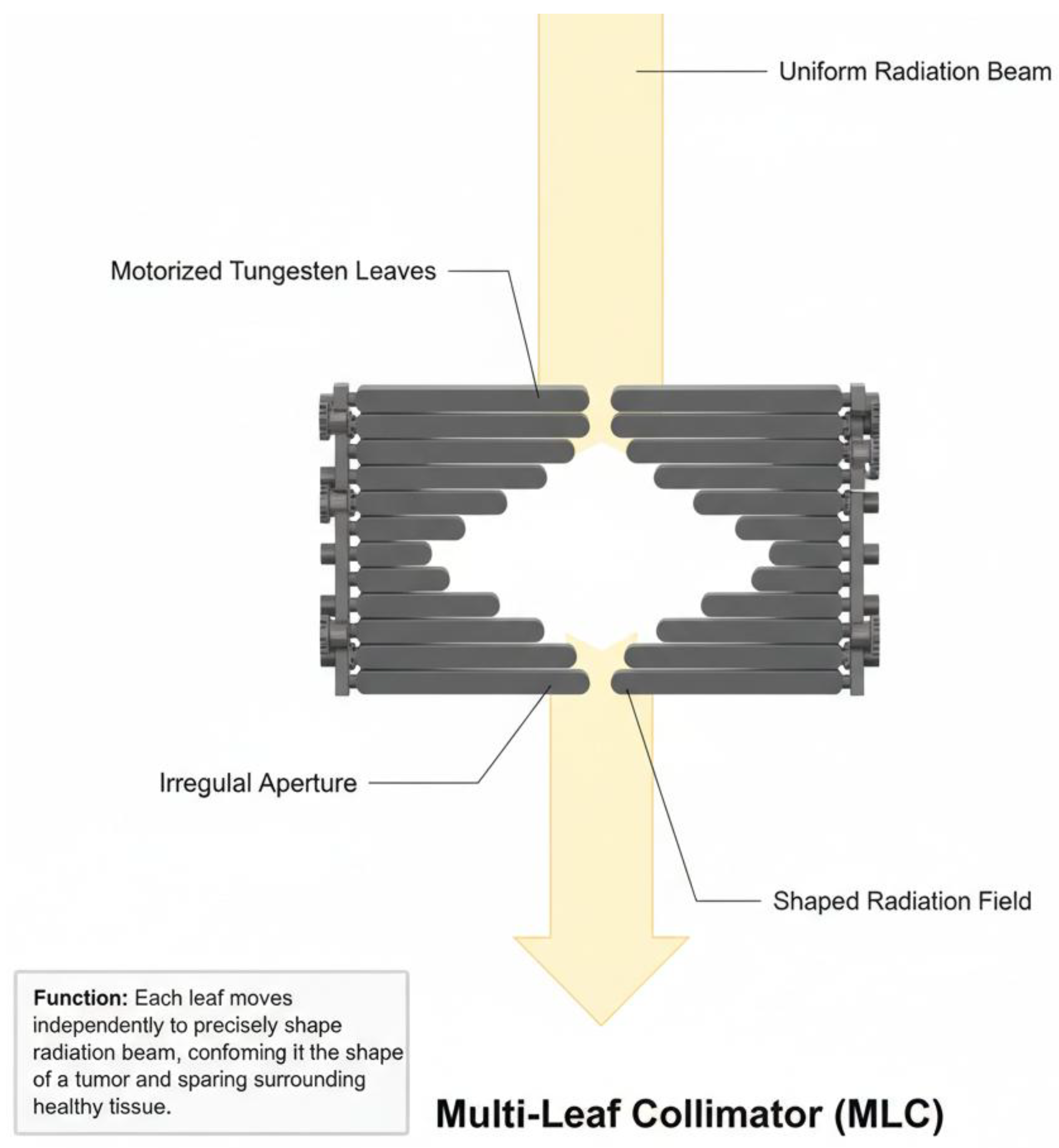
Figure 2.
Schematic illustration of a multi-leaf collimator (MLC), a beam-modulating device incorporated into linear accelerators. Arrays of independently actuated tungsten leaves move laterally to form an irregular aperture, dynamically shaping the radiation beam to match the target geometry. This adaptive field shaping enables precise control of beam distribution and contributes to the superior accuracy, safety, and reproducibility of LINAC-based irradiation in comparison with fixed sources such as Co-60.
Figure 2.
Schematic illustration of a multi-leaf collimator (MLC), a beam-modulating device incorporated into linear accelerators. Arrays of independently actuated tungsten leaves move laterally to form an irregular aperture, dynamically shaping the radiation beam to match the target geometry. This adaptive field shaping enables precise control of beam distribution and contributes to the superior accuracy, safety, and reproducibility of LINAC-based irradiation in comparison with fixed sources such as Co-60.
2. Materials and Methods
A. Materials
Invertase, (I4504, baker's yeast Saccharomyces cerevisiae), was dissolved in acetate buffer (pH 4.7) to create a stock solution at 1.35 mg/ml. A subsequent dilution yielded a 100 ml solution with a concentration of 0.135 mg/ml. Reagents such as DNS (sodium 3,5-dinitrosalicylate) and Copper(II) sulfate were analytical grade from Sigma–Aldrich. Sucrose with 99.5% purity and standard acetate buffer were sourced locally. Deionized water from a Millipore purification system was used for solution preparation where necessary.
Collagenase (SC103) from Sigma-Aldrich (Merck Bulgaria EAD) was prepared in seven concentrations: 1.00, 0.50, 0.25, 0.10, 0.05, 0.025, and 0.01 mg/ml. The solutions were made using 50 mM Tris buffer (pH 7.8) with 1 mM CaCl₂ to maintain enzymatic stability.
B. Irradiation at the LINAC suite
The enzyme solution with appropriate concentrations was placed into 1.5 ml Eppendorf tubes and irradiated with various doses of high-energy photon ionizing radiation using a Versa HD LINAC (Elekta, UK) with 6 MV flattened beams, with static 10 cm by 10 cm field in the isocenter, in service mode (Figure 3). The tubes were submerged (one at a time) at 10 cm depth in a BluePhantom2 water phantom (IBA Dosimetry, Germany) at the LINAC isocenter, with a 90 cm virtual source to water surface distance (Figure 3, the inset). Dose planning was performed with the Monaco 5.51 system, and verification was conducted using an FC65-G Farmer chamber and Dose1 electrometer (IBA Dosimetry).
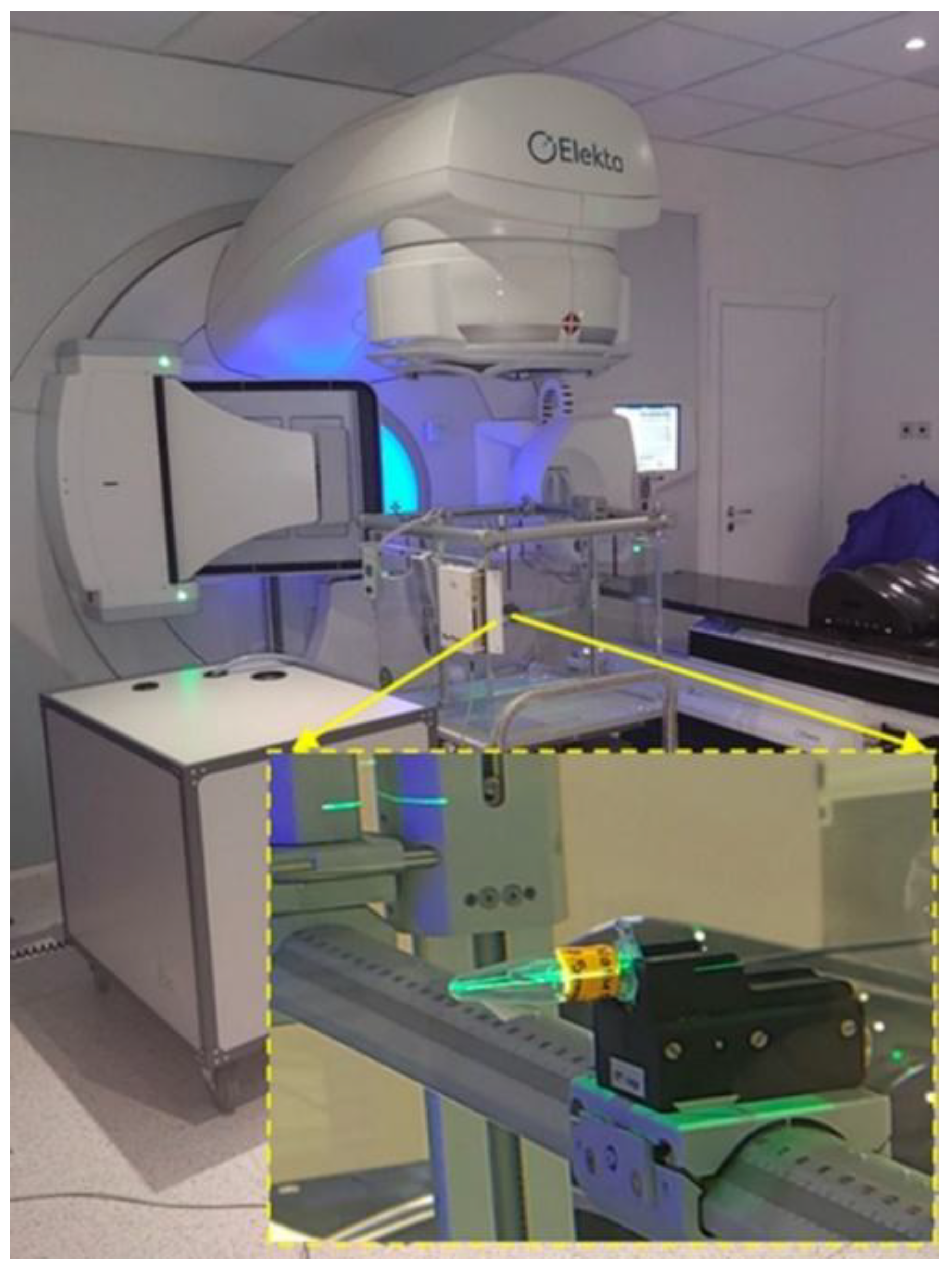
Figure 3.
Image of the LINAC suite used for enzyme solution irradiation. The inset shows a close-up of the sample support positioned within the water phantom.
Figure 3.
Image of the LINAC suite used for enzyme solution irradiation. The inset shows a close-up of the sample support positioned within the water phantom.
C. Enzyme Activity Assays
Invertase catalyzes the hydrolysis of sucrose into an equimolar mixture of glucose and fructose. Unlike sucrose, the products—glucose and fructose—are reducing sugars due to their free glycosidic groups, allowing their concentrations to be quantified calorimetrically. To measure invertase activity, we utilized a spectrophotometric method using DNS (sodium 3,5-dinitrosalicylate) as the analytical reagent, adapting a protocol initially developed for xylanase activity [27]. UV–vis absorption spectra of the samples were recorded using a Jenway 6400 spectrophotometer (Krackeler Scientific Inc., USA).
Collagenase (SC103, Sigma) solutions were prepared at seven concentrations: 1.00, 0.50, 0.25, 0.10, 0.05, 0.025, and 0.01 mg/mL, using 50 mM Tris buffer (pH 7.8) with 1 mM CaCl₂ to maintain enzymatic stability. To investigate the impact of ionizing radiation on collagenase functionality across varying concentrations and doses, two irradiation protocols were employed:
- Fixed Dose Experiment: All collagenase solutions (1.00, 0.50, 0.25, 0.10, 0.05, 0.025, and 0.01 mg/mL) were uniformly irradiated in the LINAC suite with a dose of 50 Gy to assess concentration-specific responses.
- Variable Dose Experiment: The 0.01 mg/mL solution, identified as the most responsive, was aliquoted into 1.5 mL vials and subjected to irradiation at doses of 0.1, 0.5, 1, 5, 10, 20, 50, 100, and 200 Gy to comprehensively evaluate dose-dependent activity changes.
The determination of collagenase activity followed the procedure described elsewhere [28]. Azocoll suspension (10 mg/mL) from Sigma-Aldrich (Merck Bulgaria EAD) was prepared in 50 mM Tris buffer (pH 7.8) containing 1 mM CaCl₂. An equal volume of 0.01 mg/mL collagenase solution was added, and the mixture was vigorously shaken at 21 °C for 90 minutes. After allowing the undigested Azocoll to settle, the clear supernatant was collected from each sample, and its absorbance was measured at 520 nm.
3. Results
3.1. Effect of Ionizing Radiation on Invertase Activity
The rate of enzyme reactions typically decreases over time due to factors such as substrate depletion and product inhibition. However, these factors have minimal impact on the initial reaction rate, which was therefore measured for both unirradiated and irradiated invertases. The specific activities were plotted against the radiation dose. Figure 4 illustrates this relationship (red dots, lower abscissa) along with the dependence of invertase specific activity on copper ion concentration (blue dots, upper abscissa), showing trends under varying conditions.
First, we consider the effect of ionizing radiation on invertase activity. Figure 4 demonstrates a nonlinear decrease in activity with increasing radiation dose. The native (non-irradiated) invertase shows an activity of ~2783 U/mg, aligning with previously reported values (~2700 U/mg) [29]. As radiation doses increase from 0.1 to 10 Gy, invertase activity sharply declines to ~86 U/mg. Beyond 10 Gy, the decrease slows, and at 50 Gy, activity further reduces to ~62 U/mg, reflecting a diminishing effect of higher doses on the enzyme's activity.
To better understand the mechanisms underlying the effects of ionizing radiation, systematic studies involving different enzyme and substrate concentrations (e.g., sucrose) are needed to determine kinetic constants such as the Michaelis–Menten constant. In most experiments, data on enzyme inactivation as a function of increasing radiation dose follow theoretical models that describe an exponential relationship between dose and degree of inactivation [9]. Earlier studies have shown that the inactivation of yeast invertase in dilute aqueous solutions, as well as the increased rate of this process at higher radiation doses, is due to radiolysis products that attack specific amino acid residues in the enzyme molecules, particularly methionine and histidine [23].
In our previous study [24], using same methodology, we investigated the influence of different doses of ionizing radiation on the catalytic activity of invertase and measured the kinetic parameters , and of the enzyme-catalyzed reaction. The data showed that these parameters change significantly upon irradiation—thevalues for irradiated enzymes were approximately four times higher than those for non-irradiated enzymes. This suggests that irradiation likely causes both chemical modifications of the active site and changes in the tertiary structure of the enzyme, resembling the effects of noncompetitive or mixed inhibitors [30].
In the present study, we aim to expand the list of enzymes exposed to ionizing radiation and to compare how their activities change with increasing radiation doses. Therefore, we focus specifically on one enzyme parameter—enzyme activity—and how it is affected by increasing radiation dose.
3.2. Collagenase and Its Treatment with Ionizing Radiation
One early challenge in studying the effects of ionizing radiation on enzymes was the belief that only very high doses could cause inhibition [31]. Later, building on Dale’s idea [9] that lower doses could also inhibit enzymes by reducing their concentration, Barron et al. developed an approach to study the reversible inhibition of sulfhydryl enzymes by X-rays [32] . Their method examined both varying irradiation doses and enzyme concentrations, showing that 50 Gy is a threshold for radiation-induced inhibition, beyond which the mechanisms become more complex and less biologically relevant.
Following a similar experimental logic, we conducted two types of studies. First, we analyzed the relationship between enzyme concentration and the degree of inhibition upon irradiation with a dose of 50 Gy. Solutions of collagenase with concentrations ranging from 1 mg/ml to 0.01 mg/ml were irradiated, and the results are presented in Figure 5. In the same figure, Barron's data for succine oxidase (diamonds) are compared with our results for collagenase (triangles).
The graph shows that at higher concentrations of collagenase (1 and 0.5 mg/ml) its activity decreases by only about 17%, indicating weak inhibition. Conversely, at the lowest concentration tested (0.01 mg/ml) the activity dropped by 79 %, indicating significant inhibition. These results demonstrate that the sensitivity of the enzyme to radiation inactivation is inversely proportional to its concentration, an effect that can be explained by an increased likelihood of direct interaction of radiation-induced reactive species with the enzyme molecules at lower concentrations.
The observed trend is consistent with Barron's findings for succinoxidase and suggests a similar protective mechanism for collagenase. Furthermore, these data provide a rationale for the choice of a concentration of 0.01 mg/ml for subsequent dose-dependent experiments. The pronounced inhibition at this concentration allows higher sensitivity in assessing dose-effect dependence, making it an optimal choice for further kinetic analyses.
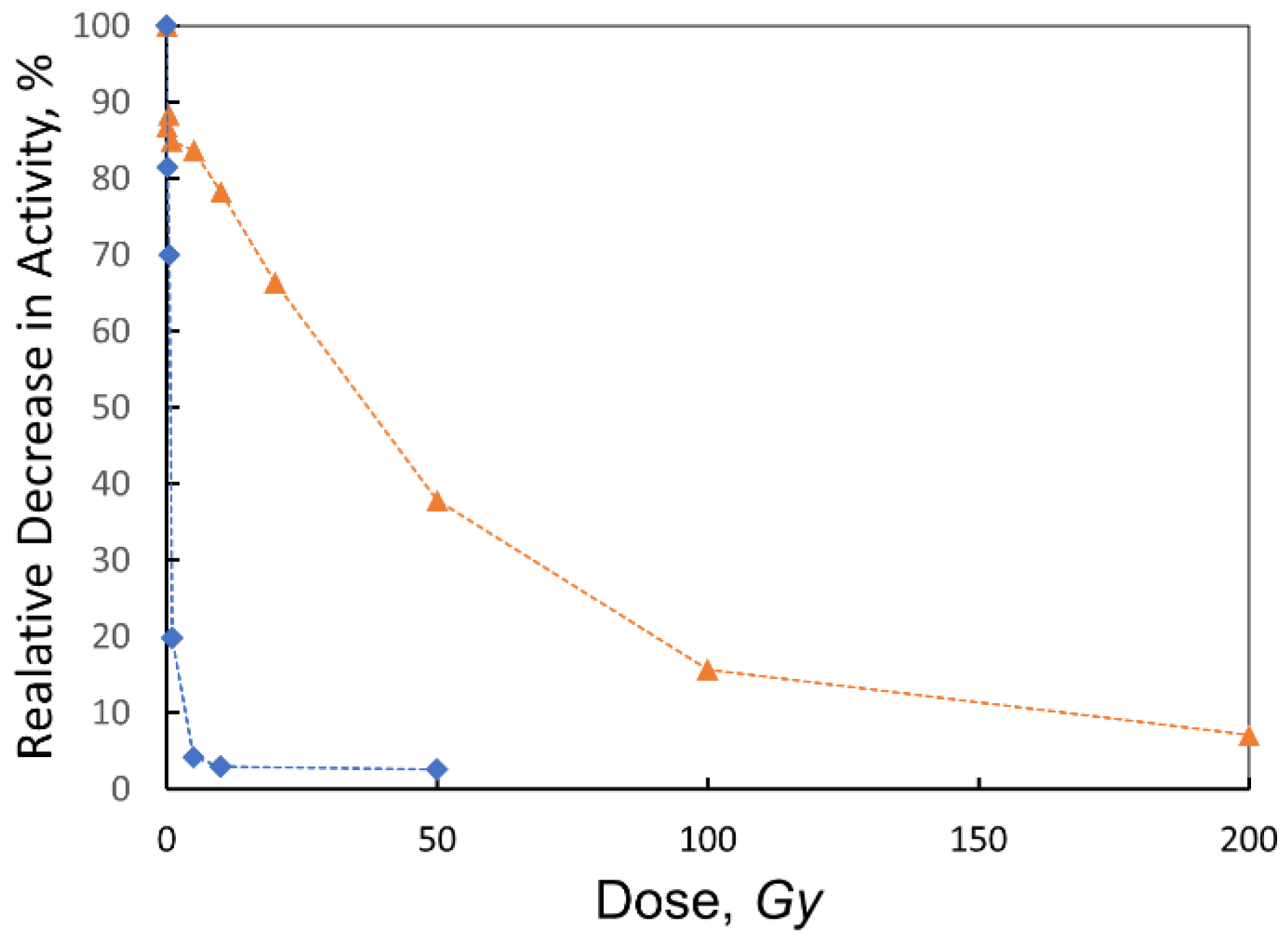
In the next series of experiments, we investigated the effect of ionizing radiation on collagenase activity. A collagenase solution of 0.1 mg/ml was aliquoted into vials and irradiated with doses of 0.1, 0.5, 1, 5, 10, 20, 50, 100 and 200 Gy. The dependence of enzyme activity (expressed as a percentage of initial activity) on irradiation dose is presented in Figure 6. For comparison, the graph also includes the invertase activity data extracted from Figure 4 and presented as percentages, as previously discussed.
Figure 6 shows that within the therapeutic dose range (0.1–10 Gy), collagenase activity decreases only slightly, by about 5–13%, indicating that the enzyme retains 87–95% of its original activity. Beyond 10 Gy, however, collagenase activity declines sharply: dropping to 66% at 20 Gy, 38% at 50 Gy, 16% at 100 Gy, and reaching just 7% at 200 Gy. This trend demonstrates an exponential, dose-dependent inhibition of collagenase activity.
4. Discussion
Over time, various kinetic models have been proposed to explain how enzyme activity changes with radiation dose [10]. These models generally show that enzyme activity decreases as dilute enzyme solutions are irradiated, primarily because "deactivated" enzymes compete with active ones for water radiolysis products. Early models described a linear relationship between enzyme activity and dose, often overlooking experimental deviations. However, it was later recognized that if inactivated and active enzyme molecules interact with aqueous reagents (primarily free radicals) at comparable reaction rates, modified kinetic models predict an exponential relationship between radiation dose and enzyme deactivation [10]. The radiolysis of sulfhydryl proteases (also known as cysteine proteases due to the presence of cysteine in their active sites) has been extensively studied [10,17,33]. A key proposal is that if both active and inactivated enzymes react similarly with free radicals generated from radiolysis, the dose-response relationship follows an exponential model [15]. This relationship is expressed as:
where is the ratio of the amount of irradiated enzyme molecules to native (nonirradiated) enzyme molecules, which is proportional to the relative activity of irradiated enzymes compared to nonirradiated ones, K is the global kinetic constant, and D is the radiation dose. When fitting our collagenase and invertase data (Figure 6) to this model, we obtained and , respectively.
The physical meaning of this empirically determined constant is not straightforward to define, but it allows calculation of the so-called —the dose required to reduce enzyme activity to 50% of its native level. The model yields for collagenase and for invertase. These results indicate that invertase is more than 50 times more susceptible to radiation-induced inactivation than collagenase. This difference is striking, although the applicability of Dale’s model to collagenase should be interpreted with caution. For sulfhydryl proteases, Dale’s exponential dose–response model has been shown to describe enzyme inactivation reliably. However, collagenase is a Zn²⁺-dependent matrix metalloproteinase with a saddle-shaped tertiary structure and a zinc ion at its active site [34]. Therefore, although we applied the same model, the kinetic meaning of the constant K for collagenase likely differs from that for sulfhydryl enzymes. For example, studies on carboxypeptidase A, another Zn-containing enzyme, have shown that inactivation primarily results from conformational changes rather than direct damage to the active site [35]. Interestingly, while other Zn-containing enzymes such as angiotensin-converting enzyme (ACE) also follow an exponential inactivation trend, the reported K value for ACE is about an order of magnitude lower than the K value obtained for collagenase. This suggests that although Zn-containing hydrolases generally share some sensitivity to radiation-induced conformational changes, collagenase shows significantly higher sensitivity—potentially due to differences in structural stability or the dynamics of free radical interactions.
It is also noteworthy that, as reported by Orlova [35], ACE retains about 50% of its activity even after exposure to doses as high as 800 Gy. This indicates that while metalloproteases such as collagenase share some features of their radiation response with serine and sulfhydryl proteases, significant differences remain between these enzyme classes. The mechanisms underlying these differences, particularly their varying radiation sensitivities, require further investigation. A deeper understanding could provide valuable insights into the structural and functional stability of different protease families under ionizing radiation.
5. Conclusions
This study successfully established and applied an experimental protocol leveraging a medical linear accelerator (LINAC) suite to investigate enzyme-catalyzed reactions under precisely controlled irradiation conditions. This methodology offers a significant advantage over traditional radioisotope sources by enabling real-time dose control and accurate dosimetry, crucial for understanding radiation-induced biological effects.
Applying this protocol, we evaluated the catalytic activities of two distinct enzymes: invertase and collagenase. For invertase, our findings revealed a pronounced nonlinear decrease in activity with increasing radiation dose. Native invertase activity experienced a sharp decline between 0.1 and 10 Gy, plummeting to a mere 2.2% of its initial activity at the maximum dose of 50 Gy. This inactivation followed Dale's exponential law, yielding a constant . This observed sensitivity suggests that radical species generated during radiolysis likely induce damage to critical amino acid residues, such as methionine and histidine, within the enzyme's active site or structural integrity.
In the case of collagenase, we thoroughly investigated its response to ionizing radiation across various enzyme concentrations and doses. Through both fixed-dose and variable-dose irradiation protocols, we demonstrated that collagenase activity exhibits an exponential decay with a rate constant of . This finding suggests that collagenase, a metalloprotease, shares certain fundamental radiation-response characteristics with other protease classes, notably serine and sulfhydryl proteases. However, the observed differences in radiation sensitivities among these enzyme classes underscore the necessity for further comprehensive investigation employing a wider array of experimental methods and advanced theoretical approaches to fully elucidate their distinct mechanisms of radiation-induced inactivation.
Our developed LINAC-based experimental framework provides a robust and reproducible platform for future studies exploring the radiation biology of enzymes, offering unparalleled control over irradiation parameters. This precise control is paramount for advancing our understanding of radiation-induced enzyme damage and for developing strategies to mitigate such effects in various applications, including radiotherapy and enzyme biotechnology.
Author Contributions
Conceptualization, K.B. and Ph.M.; methodology, K.B. and S.P.; software, Ph.M.; validation, K.B., S.P. and I.P.; formal analysis, Ph.M.; investigation, Ph.M., K.S., Ts.L. and P.T.; resources, Ph.M. and I.P.; data curation, Ph.M.; writing—original draft preparation, Ph.M.; writing—review and editing, K.B. and I.P.; visualization, Ph.M. and K.B.; supervision, K.B.; project administration, Ph.M.; funding acquisition, Ph.M. and K.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by European Union-NextGenerationEU, through the National Recovery and Resilience Plan of the Republic of Bulgaria, project № BG-RRP-2.004-0003.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
Ph.M. gratefully acknowledges the support of European Union – NextGenerationEU, through the National Recovery and Resilience Plan of the Republic of Bulgaria, project № BG-RRP-2.004-0003. K.B. gratefully acknowledges the support of contract BG-RRP-2.004-0008, funding the project “Sofia University Marking Momentum for Innovation and Technological Transfer” (SUMMIT) under the National Recovery and Resilience Plan, Program for Accelerating Economic Recovery and Transformation through Science and Innovation. The authors acknowledge the use of the AI-assisted tool Gpai to support improvements of Figure 3. In addition, ChatGPT (OpenAI) was used for minor language editing and improvement of text clarity. All conceptual analysis, interpretation of data, and conclusions are solely the responsibility of the authors. All authors have reviewed and approved the final version of the manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| LINAC | Medical linear accelerator |
References
- Arena, V. Ionizing Radiation and Life: An Introduction to Radiation Biology and Biological Radiotracer Methods; Mosby: Saint Louis, 1971; ISBN 978-0-8016-0278-8. [Google Scholar]
- Belli, M.; Indovina, L. The Response of Living Organisms to Low Radiation Environment and Its Implications in Radiation Protection. Front. Public Health 2020, 8, 601711. [Google Scholar] [CrossRef]
- Chang, D.S.; Lasley, F.D.; Das, I.J.; Mendonca, M.S.; Dynlacht, J.R. Basic Radiotherapy Physics and Biology; Springer International Publishing: Cham, 2021; ISBN 978-3-030-61898-8. [Google Scholar]
- Ciernik, I.F.; Dizendorf, E.; Baumert, B.G.; Reiner, B.; Burger, C.; Davis, J.B.; Lütolf, U.M.; Steinert, H.C.; Von Schulthess, G.K. Radiation Treatment Planning with an Integrated Positron Emission and Computer Tomography (PET/CT): A Feasibility Study. International Journal of Radiation Oncology*Biology*Physics 2003, 57, 853–863. [Google Scholar] [CrossRef]
- Namiki, Y.; Fuchigami, T.; Tada, N.; Kawamura, R.; Matsunuma, S.; Kitamoto, Y.; Nakagawa, M. Nanomedicine for Cancer: Lipid-Based Nanostructures for Drug Delivery and Monitoring. Acc. Chem. Res. 2011, 44, 1080–1093. [Google Scholar] [CrossRef]
- Wain, J. Ionising Radiation Protection: A Primer. In Synthesis Lectures on Engineering, Science, and Technology; Springer Nature Switzerland: Cham, 2025; ISBN 978-3-031-65524-1. [Google Scholar]
- Leslie, S.W.; Hevly, B. Steeple Building at Stanford: Electrical Engineering, Physics, and Microwave Research. Proc. IEEE 1985, 73, 1169–1180. [Google Scholar] [CrossRef]
- Houée-Levin, C.; Sicard-Roselli, C. Radiation Chemistry of Proteins. In Studies in Physical and Theoretical Chemistry; Elsevier, 2001; Vol. 87, pp. 553–584. ISBN 978-0-444-82902-3. [Google Scholar]
- Dale, W.M. The Effect of X-Rays on Enzymes. Biochemical Journal 1940, 34, 1367–1373. [Google Scholar] [CrossRef]
- Sanner, T.; Pihl, A. Kinetics of Enzyme Inactivation in Solution by Ionizing Radiation. European Journal of Biochemistry 1967, 1, 33–35. [Google Scholar] [CrossRef]
- Kempner, E.S. Damage to Proteins Due to the Direct Action of Ionizing Radiation. Quart. Rev. Biophys. 1993, 26, 27–48. [Google Scholar] [CrossRef]
- Venkateswara Rao, M.; C. K., S.; Rawson, A.; D. V., C.; N., V. Modifying the Plant Proteins Techno-Functionalities by Novel Physical Processing Technologies: A Review. Critical Reviews in Food Science and Nutrition 2023, 63, 4070–4091. [Google Scholar] [CrossRef]
- Draganić, I.G.; Draganić, Z.D. The Radiation Chemistry of Water; Physical chemistry, a series of monographs; Academic Press: New York, 1971; ISBN 978-0-323-15878-7. [Google Scholar]
- Draganić, I.G. Radiolysis of Water: A Look at Its Origin and Occurrence in the Nature. Radiation Physics and Chemistry 2005, 72, 181–186. [Google Scholar] [CrossRef]
- Orlova, M.A. Radiation-Induced Inactivation of Proteolytic Enzymes. Russ. Chem. Rev. 1993, 62, 493–508. [Google Scholar] [CrossRef]
- Phaniendra, A.; Jestadi, D.B.; Periyasamy, L. Free Radicals: Properties, Sources, Targets, and Their Implication in Various Diseases. Ind J Clin Biochem 2015, 30, 11–26. [Google Scholar] [CrossRef]
- Gribkov, V.A.; Orlova, M.A. On Various Possibilities in Pulsed Radiation Biochemistry and Chemistry. Radiat Environ Biophys 2004, 43, 303–309. [Google Scholar] [CrossRef]
- Orlova, M.A.; Kost, O.A.; Gribkov, V.A.; Gazaryan, I.G.; Dubrovsky, A.V.; Egorov, V.A.; Troshina, N.N. Enzyme Activation and Inactivation Induced by Low Doses of Irradiation. ABAB 2000, 88, 243–256. [Google Scholar] [CrossRef]
- Saha, A. Photo-Induced Inactivation of Dihydroorotate Dehydrogenase in Dilute Aqueous Solution. International Journal of Radiation Biology 1997, 72, 55–61. [Google Scholar] [CrossRef]
- Wu, S.; Zhou, X.; Jin, Z.; Cheng, H. Collagenases and Their Inhibitors: A Review. Collagen & Leather 2023, 5, 19. [Google Scholar] [CrossRef]
- Sainz-Polo, M.A.; Ramírez-Escudero, M.; Lafraya, A.; González, B.; Marín-Navarro, J.; Polaina, J.; Sanz-Aparicio, J. Three-Dimensional Structure of Saccharomyces Invertase. Journal of Biological Chemistry 2013, 288, 9755–9766. [Google Scholar] [CrossRef]
- Drózdz, M.; Piwowarczyk, B.; Olczyk, K. Effects of ionizing radiation on the content of total collagen and its fractions and the activity of collagenolytic enzymes in rat tissues. Med Pr 1981, 32, 317–322. [Google Scholar] [PubMed]
- Nagrani, S.; Bisby, R.H. The Radiation-Induced Inactivation of External Yeast Invertase in Dilute Aqueous Solution. International Journal of Radiation Biology 1989, 55, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Marinov, P.; Petrov, I.; Stoilov, K.; Lazhovski, T.; Temnishki, P.; Petrova, S.; Balashev, K. Effect of Ionizing Radiation on the Activity of the Enzyme Invertase Studied by Polarimetry. C. R. Acad. Bulg. Sci. 2024, 77. [Google Scholar] [CrossRef]
- Ahnesjö, A.; Aspradakis, M.M. Dose Calculations for External Photon Beams in Radiotherapy. Phys. Med. Biol. 1999, 44, R99–R155. [Google Scholar] [CrossRef]
- Bortfeld, T. IMRT: A Review and Preview. Phys. Med. Biol. 2006, 51, R363–R379. [Google Scholar] [CrossRef]
- Sokač, T.; Šalić, A.; Dukarić, A.-M.; Tišma, M.; Planinić, M.; Zelić, B.; Božinović, M. Standardization of 3,5-Dinitrosalicylic Acid (DNS) Assay for Measuring Xylanase Activity: Detecting and Solving Problems. Croat. j. food sci. technol. (Online) 2023, 15, 151–162. [Google Scholar] [CrossRef]
- Chavira, R.; Burnett, T.J.; Hageman, J.H. Assaying Proteinases with Azocoll. Analytical Biochemistry 1984, 136, 446–450. [Google Scholar] [CrossRef]
- Neumann, N.P.; Lampen, J.O. Purification and Properties of Yeast Invertase*. Biochemistry 1967, 6, 468–475. [Google Scholar] [CrossRef] [PubMed]
- Sirisatesuwon, C.; Ninchan, B.; Sriroth, K. Effects of Inhibitors on Kinetic Properties of Invertase from Saccharomyces Cerevisiae. Sugar Tech 2020, 22, 274–283. [Google Scholar] [CrossRef]
- Scott, C.M. Some Quantitative Aspects of the Biological Actions of X and γ-Rays. 1937. [Google Scholar]
- Barron, E.S.G.; Dickman, S.; Muntz, J.A.; Singer, T.P. STUDIES ON THE MECHANISM OF ACTION OF IONIZING RADIATIONS. Journal of General Physiology 1949, 32, 537–552. [Google Scholar] [CrossRef]
- Sanner, T.; Pihl, A. Kinetics of Enzyme Inactivation by Ionizing Radiation. Radiation Research 1963, 19, 12. [Google Scholar] [CrossRef]
- Patel, N.Y.; Baria, D.M.; Pardhi, D.S.; Yagnik, S.M.; Panchal, R.R.; Rajput, K.N.; Raval, V.H. Chapter 16 - Microbial Enzymes in Pharmaceutical Industry. In Biotechnology of Microbial Enzymes (Second Edition); Brahmachari, G., Ed.; Academic Press, 2023; pp. 375–403. ISBN 978-0-443-19059-9. [Google Scholar]
- Orlova, M.A. Radiation-Induced Inactivation of Proteolytic Enzymes. Russ. Chem. Rev. 1993, 62, 493–508. [Google Scholar] [CrossRef]
Figure 1.
Basic structure of a linear accelerator (LINAC). Electrons generated by an electron gun are directed through an accelerator waveguide tube and accelerated. The electron beam is then bent to strike a target, producing X-rays. These X-rays are further shaped by a system of collimators and monitored by an ionization chamber before being delivered to the patient's surface.
Figure 1.
Basic structure of a linear accelerator (LINAC). Electrons generated by an electron gun are directed through an accelerator waveguide tube and accelerated. The electron beam is then bent to strike a target, producing X-rays. These X-rays are further shaped by a system of collimators and monitored by an ionization chamber before being delivered to the patient's surface.
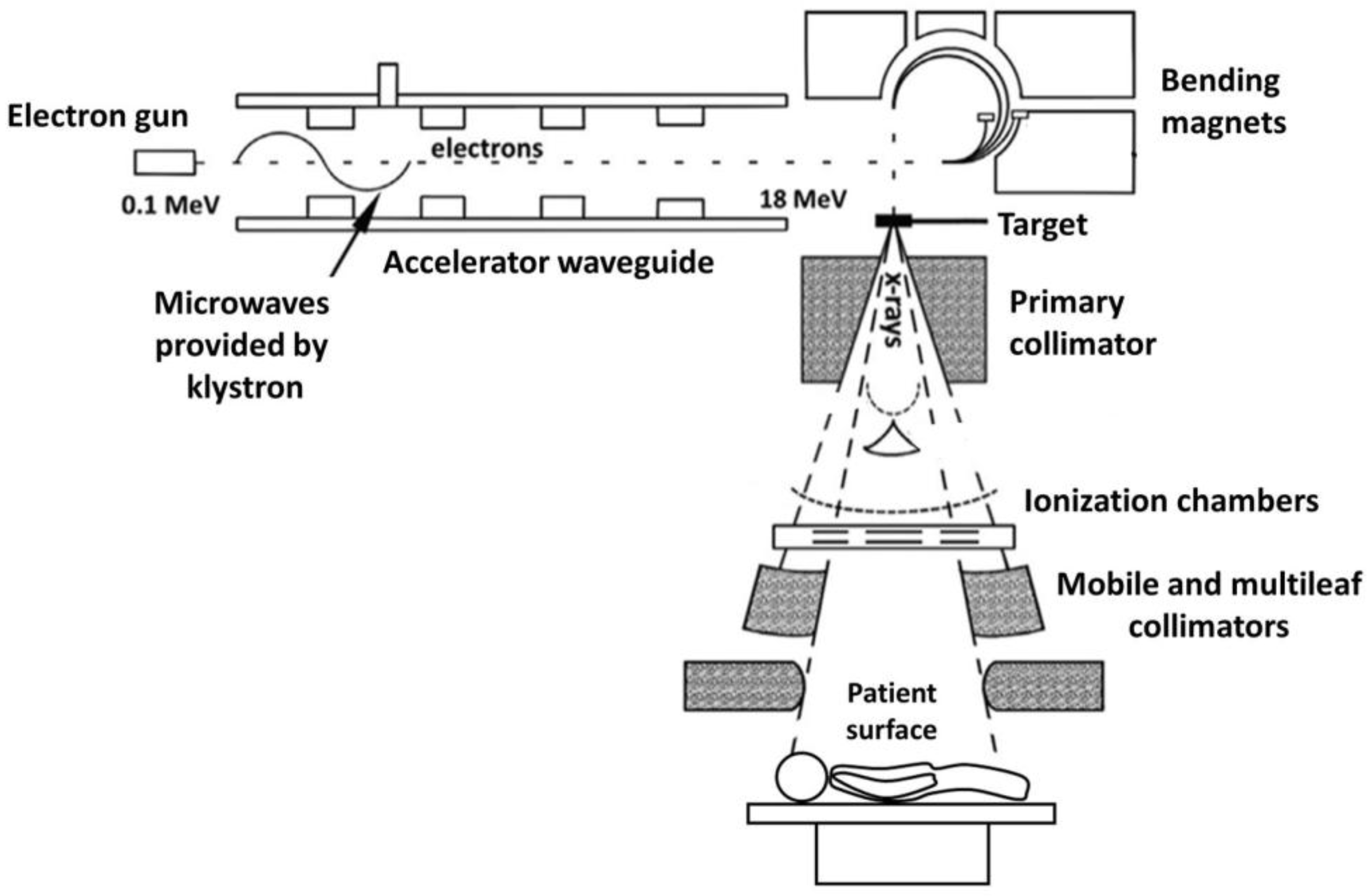
Figure 4.
The specific activity of invertase (SA, μM/min/mg ≡ U/mg) as a function of radiation dose. The dotted line illustrates the trend, connecting the data points as both radiation dose and copper ion concentration increase.
Figure 4.
The specific activity of invertase (SA, μM/min/mg ≡ U/mg) as a function of radiation dose. The dotted line illustrates the trend, connecting the data points as both radiation dose and copper ion concentration increase.
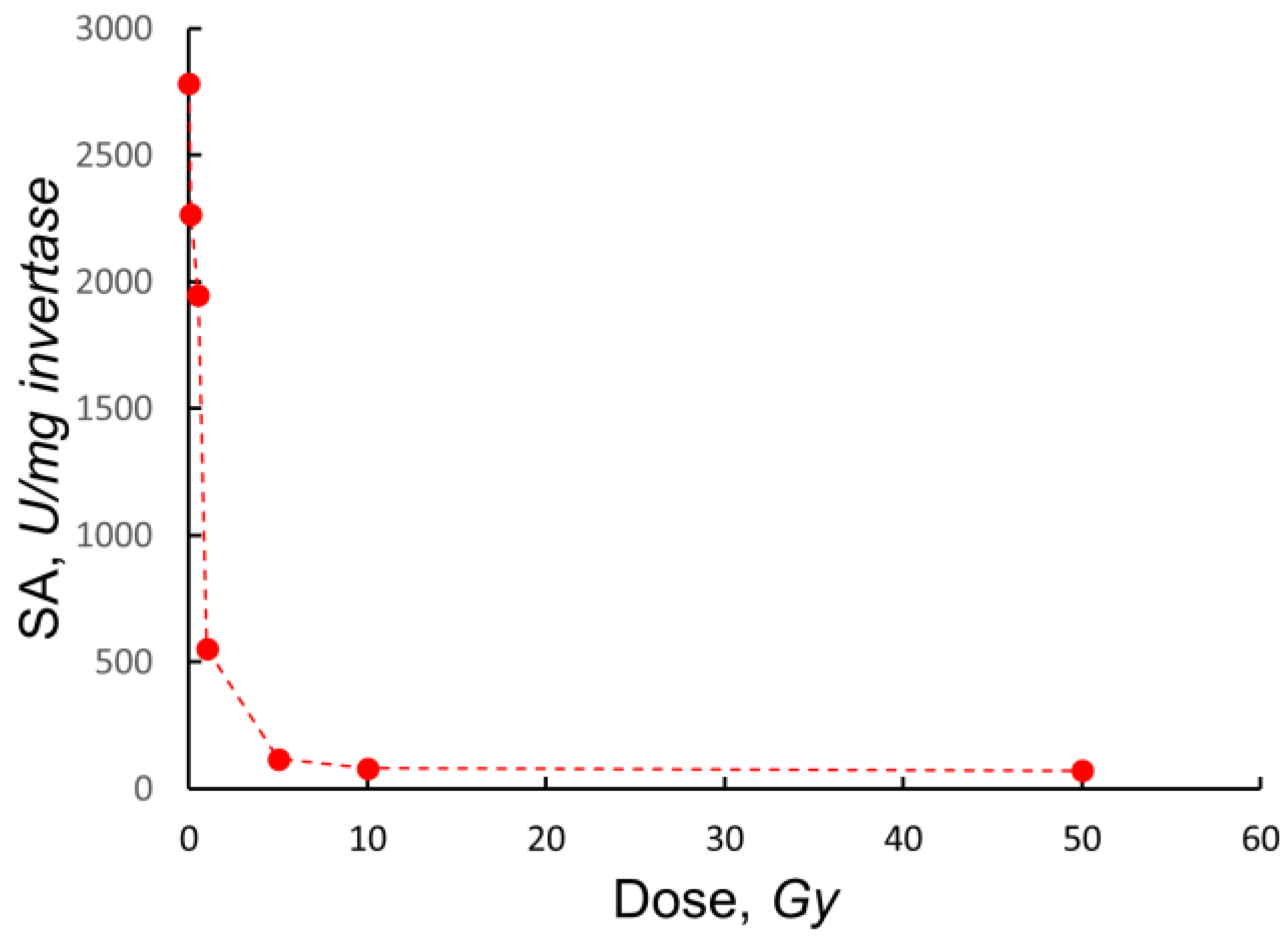
Figure 5.
The graph presents a comparative analysis of the decrease in enzyme activity as a function of enzyme concentration. The first data set, indicated by triangles (▲), shows the experimentally measured decrease in collagenase activity (in percent) at different concentrations expressed in mg/ml (lower abscissa). The second data set, represented by diamonds (◆), illustrates the results of Barron et al. [32] for succine oxidase activity plotted against its concentration in conditional units ("cc"). In both cases, the dotted lines show the trend of change in enzyme activity with increasing concentration. The figure highlights a general trend towards less inhibition at higher concentrations, suggesting a protective effect against radiation inactivation.
Figure 5.
The graph presents a comparative analysis of the decrease in enzyme activity as a function of enzyme concentration. The first data set, indicated by triangles (▲), shows the experimentally measured decrease in collagenase activity (in percent) at different concentrations expressed in mg/ml (lower abscissa). The second data set, represented by diamonds (◆), illustrates the results of Barron et al. [32] for succine oxidase activity plotted against its concentration in conditional units ("cc"). In both cases, the dotted lines show the trend of change in enzyme activity with increasing concentration. The figure highlights a general trend towards less inhibition at higher concentrations, suggesting a protective effect against radiation inactivation.
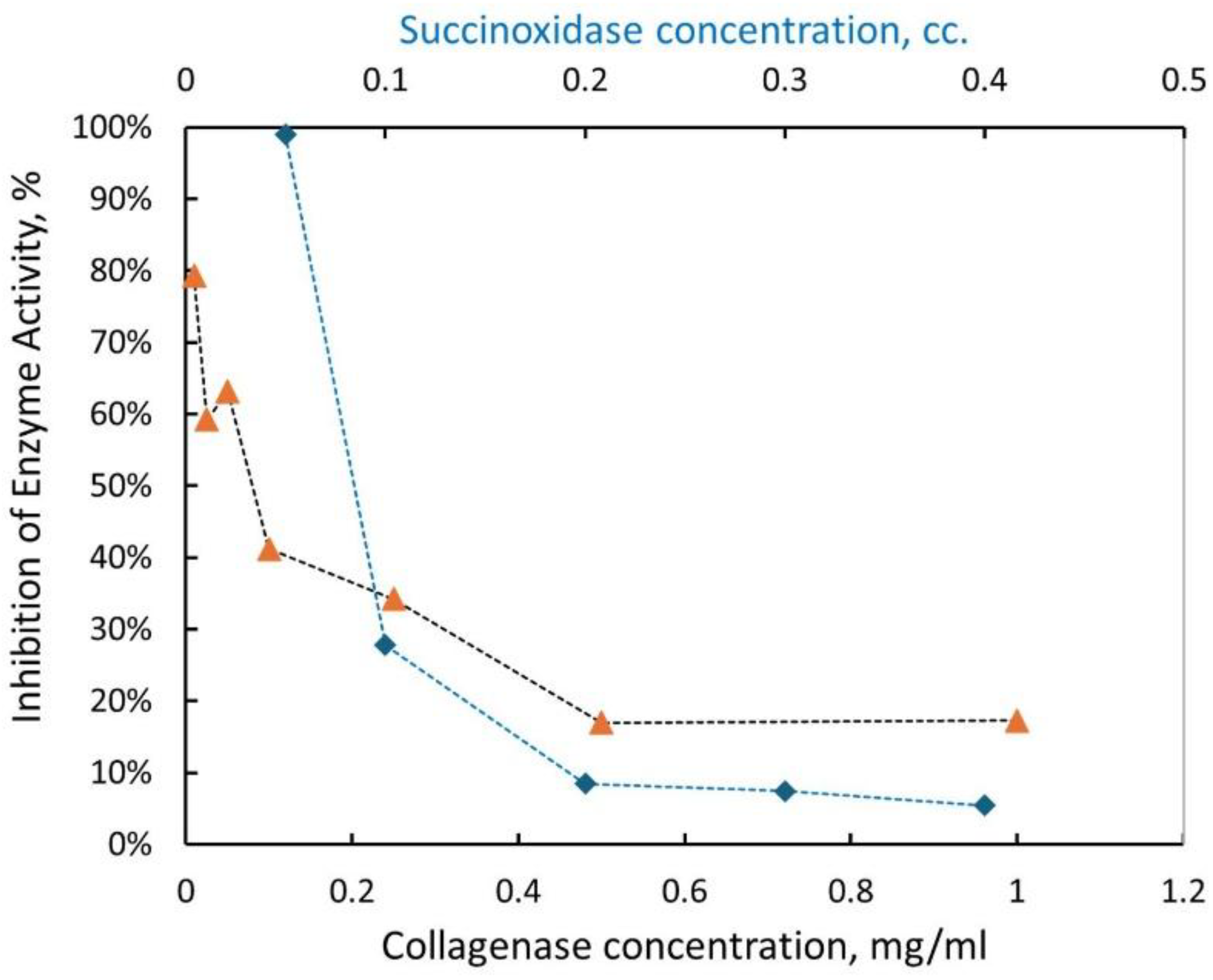
Figure 6.
Decrease in collagenase activity (▲) and invertase activity (◆), shown as a percentage, as a function of radiation dose (Gy). The dotted lines indicate the trend of enzyme activity with increasing dose. The data for both enzymes are fitted to the exponential dose–response model [15], yielding for collagenase and for invertase.
Figure 6.
Decrease in collagenase activity (▲) and invertase activity (◆), shown as a percentage, as a function of radiation dose (Gy). The dotted lines indicate the trend of enzyme activity with increasing dose. The data for both enzymes are fitted to the exponential dose–response model [15], yielding for collagenase and for invertase.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



