Submitted:
03 September 2025
Posted:
04 September 2025
You are already at the latest version
Abstract
Acacetin, a naturally occurring flavonoid, has attracted increasing attention due to its broad anticancer potential. In vitro and in vivo studies using diverse tumor models have demonstrated that acacetin modulates oncogenic signaling, suppresses angiogenesis, and induces apoptosis and other regulated cell death pathways. With the rising demand for multi-target therapeutics, network pharmacology and molecular docking have emerged as powerful tools to unravel the complex molecular mechanisms of phytochemicals. This review integrates computational network analysis with molecular docking and experimental validation, revealing that acacetin directly targets critical oncogenic proteins such as the epidermal growth factor receptor (EGFR), signal transducer and activator of transcription 3 (STAT3), and the serine/threonine kinase AKT, also known as protein kinase B (PKB). These findings indicate a multi-target mode of action across key oncogenic pathways, underscoring the role of acacetin as a multi-target agent capable of interfering with several cancer hallmarks. By combining systems biology approaches with experimental evidence, this review provides a comprehensive perspective on the pharmacological profile of acacetin and highlights its potential as a promising candidate for the future development of cancer therapy.
Keywords:
acacetin
; cancer
; network pharmacology
; molecular docking
; EGFR
; STAT3
; AKT
1. Introduction
Cancer is one of the most formidable global public health challenges, consistently ranking as the second leading cause of death worldwide and affecting millions of individuals and families across diverse populations [1,2,3,4,5]. Cancer is characterized by uncontrolled cellular proliferation, invasion of surrounding tissues, and the potential for distant metastasis, all of which together contribute to its high morbidity and mortality [6].
Despite significant advances, traditional cancer therapies, including surgery, chemotherapy, radiation therapy, and hormonal treatments, continue to grapple with intrinsic limitations such as incomplete tumor eradication due to heterogeneity, systemic toxicity, development of drug resistance, and adverse effects on patients’ quality of life [1,7]. Although these modalities remain the mainstay of oncology, therapeutic barriers underscore the urgent need for novel and selective interventions [8]. Moreover, innovative strategies such as targeted therapies, immunotherapies, gene therapy, and nanomedicine have emerged, offering improved specificity and promise for precision medicine. However, obstacles such as antigenic variation, manufacturing complexity, limited tissue penetration, and high costs are still encountered [9]. In this context, bioactive natural compounds, particularly flavonoids, are attracting research interest due to their multi-target actions, favorable safety profiles, and ability to regulate complex oncogenic signaling networks [10].
Acacetin, a naturally occurring flavonoid that belongs to the flavone subclass present in various plant species, has demonstrated diverse pharmacological actions, including anticancer, anti-inflammatory, antioxidant, and hepatoprotective effects through modulation of key intracellular pathways such as mitogen-activated protein kinase (MAPK)/c-Jun N-terminal kinase (JNK)/extracellular signal-regulated kinase (ERK), nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB), nuclear factor erythroid 2–related factor 2 (Nrf2), phosphoinositide 3-kinase (PI3K)/serine/threonine kinase (AKT, also known as protein kinase B, PKB)/mechanistic target of rapamycin (mTOR), and cyclooxygenase (COX)-2 [11]. Nonetheless, a detailed understanding of the molecular mechanisms by which acacetin exerts its anticancer activity remains limited owing to the inherently networked and redundant signaling architecture of cancer cells [12].
In recent years, network pharmacology, defined as a systems-level approach that integrates bioinformatics, cheminformatics, and systems biology to map interactions between drugs and multiple molecular targets, has emerged as a powerful system-level framework for predicting interactions between bioactive compounds and disease-associated targets, facilitating the identification of hub proteins and enriched signaling pathways through integrated protein–protein interaction (PPI) networks and pathway analysis [13]. Molecular docking offers a computational method for estimating binding affinities and interactions between compounds and target proteins, providing structural insights to support experimental validation [14,15,16,17,18]. These computational strategies have been successfully applied to phytochemicals such as curcumin in osteosarcoma and herbal extracts in prostate and cervical cancers, highlighting their value in uncovering multi-target anticancer mechanisms [19,20,21].
Therefore, in this review, we aimed to synthesize the pharmacological evidence of acacetin’s anticancer activities in vitro and in vivo and complement this with a combined network pharmacology and molecular docking approach to identify key targets and binding interactions. Such efforts may illuminate the potential of acacetin as a multi-target therapeutic agent across diverse cancer types and may inform future strategies for translational and combinatory treatment development.
In preparing this review, we systematically searched the literature using the keyword “acacetin” in the PubMed and Google Scholar databases. The search covered publications available until August 2025 and was restricted to full-text articles written in English. Emphasis is placed on preclinical investigations, particularly those providing mechanistic insights, reporting in vitro and in vivo pharmacological activities, and presenting experimental evidence supporting the predicted molecular targets.
2. Overview of Acacetin
2.1. Chemical Structure and Sources
Acacetin (5,7-dihydroxy-4′-methoxyflavone) is an O-methylated flavone (C16H12O5) belonging to the flavonoid family (Figure 1). Its A-ring bears hydroxyl groups at C-5 and C-7, whereas the B-ring carries a methoxy group at C-4′, and these are features that shape its physicochemical behavior and target engagement in biological systems [11]. In nature, acacetin acts as an aglycone and glycoside in multiple edible or medicinal taxa. It has been isolated from black locust (Robinia pseudoacacia), bee propolis, Dracocephalum moldavica, Turnera diffusa, and Betula pendula, as evidenced by chemical profiling in peer-reviewed studies. These sources include widely used foods or ethnomedicinal plants from which acacetin and its conjugates have been chemically characterized [22,23,24].
2.2. Pharmacological Activities of Acacetin
Acacetin exerts diverse pharmacological effects. In oncology, it modulates key oncogenic pathways, including PI3K/AKT, JAK/signal transducer and activator of transcription 3 (STAT3), NF-κB, and MAPK, thereby inducing apoptosis, cell cycle arrest, and inhibition of metastasis [25,26,27,28]. Its anti-inflammatory action has been demonstrated in RAW 264.7 macrophages, where acacetin suppresses lipopolysaccharide (LPS)-induced expression of inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2) by inhibiting the NF-κB pathway through reduced IκBα phosphorylation and blockade of NF-κB nuclear translocation [29]. In cardiovascular research, acacetin selectively inhibits atrial potassium currents, including ultra-rapid delayed rectifier K⁺ current and transient outward, prolonging atrial action potential duration and preventing atrial fibrillation in canine models without affecting ventricular repolarization [30,31]. Furthermore, in apolipoprotein E-deficient mice, acacetin attenuated atherosclerosis by activating the Nrf2 signaling pathway and upregulating the antioxidant enzyme methionine sulfoxide reductase A [32].
Beyond these effects, acacetin exhibits antiviral activity against multiple viruses, including herpes simplex virus type 1, dengue virus type 2, influenza virus, and human immunodeficiency virus, underscoring its broad-spectrum potential [33,34,35,36]. It displays antimicrobial efficacy, particularly against methicillin-resistant Staphylococcus aureus and various Gram-positive and Gram-negative bacteria, partly through synergy with antibiotics and inhibition of virulence factors such as sortase A [37,38,39]. Moreover, acacetin exerts anti-obesity effects by suppressing adipogenesis, enhancing lipolysis, and activating AMP-activated protein kinase (AMPK)/sirtuin 1 (SIRT1) signaling, consistent with reduced body weight observed in high-fat diet–induced obese mice [40]. These multifaceted activities highlight acacetin’s potential as a therapeutic candidate for a wide range of chronic diseases.
3. Pharmacological Evidence of Acacetin’s Anticancer Activity
Several mechanistic and pharmacological studies have evaluated the anticancer activity of acacetin in different tumor types. As outlined in Table 1, these findings indicate that acacetin exerts multi-faceted effects both in vitro and in vivo, including apoptosis induction, proliferation inhibition, cell-cycle regulation, suppression of metastasis, and inhibition of angiogenesis.

Table 1.
Summary of acacetin’s anticancer effects by cancer type.
| Interventions Cancer Type |
Cell Line(s) / Model | Up-Regulation | Down-Regulation | Mechanisms | Refs. |
|---|---|---|---|---|---|
| Breast | T-47D, MDA-MB-231, MCF-7, MDA-MB-468, |
DNA fragmentation, sub-G1 cells, Cyt c (cytosolic) AIF (cytosolic), p-SAPK/ JNK1/2, p-c-Jun, JNK1/2, ROS, cleaved caspase-7,-8 and -9, cleaved PARP, p-ERK, Chk1, Bax, Mitochondrial superoxide generation, RIP1, RIP3 p21, p27 |
Bcl-2, MMP, Cyt c (mitochondrial), AIF (mitochondrial) CDK1, CDK2, Cdc25C, Cyclin B1, Cyclin E, AKT, FADD, p-AKT, AKT, p38, ERK |
Apoptosis, Necroptosis, G2/M arrest, ERK activation Migration and EMT inhibition |
[41,42,43] |
| Colorectal |
HT-29, HCT 116, SW480 | ROS, MMP (mitochondrial), AIF | β-catenin, c-Myc, | Apoptosis, S and G2/M arrest |
[44,45] |
| Esophageal | TE-1, TE-10 | Bax | Ki-67, MMP-2 and -9, Bcl-2, p-JAK2, p-STAT3 | Apoptosis, JAK2/STAT3 inhibition, Migration suppression |
[26] |
| Gastric | MKN45, MKN45 xenograft, MGC803, AGS | E-cadherin, Bax, cleaved PARP, cleaved caspase-3, DNA fragmentation, Sub-G1 cells, ROS, Cyt c (cytosolic) caspase-3,-8, and -9 activity, Fas, FasL, cleaved Bid, p53 |
N-cadherin, MMP-2 and -9, Snail, p-PI3K, p-AKT, p-EGFR, Bcl-xL, p-STAT3, p-ERK, p-EGFR, PCNA, DFF-45, MMP | Apoptosis, EMT suppression, Anti-metastasis, PI3K/AKT/Snail inhibition |
[12,46,47] |
| Liver | HepG2, HepG2/RARγ xenografts | DNA fragmentation, cleaved PARP, p21, p53, FasL, mFasL, sFasL, caspase-8 activity, Bax, cleaved caspase-3, | p-STAT3, p-JAK1,p-JAK2, p-AKT, p-Src, Cyclin D1, Bcl-2, Bcl-xL, Mcl-1, Survivin, Mcl-2, VEGF, p-IκBα (cytosolic), p-p65 (cytosolic), pro-casapase-3, RARγ, p-GSK-3β, Ki67 | Apoptosis, STAT3 inhibition, Invasion inhibition, Angiogenesis inhibition, G1 arrest, Apoptosis via non-genomic RARγ-AKT-p53 pathway |
[48,49,50] |
| Lung | A549, H460, A549 xenografts | Bak, p53, miR-34a, IκBα(cytosolic), G1 phase, p21, Fas, FasL | Cyclin B1, , Cyclin D, Bcl-2, PD-L1, MMP-2 and -9, u-PA, p-p38α, p-MKK3, 6, p-MLK3, NF-κB, NF-κB p50 (nuclear), NF-κB p65 (nuclear) AP-1, c-Fos, c-Jun, p-IκBα (cytosolic) | Proliferation, invasion, and migration inhibition, G2/M arrest, Apoptosis |
[27,28,51,52] |
| Ovarian | SKOV3, OVCAR-3, A2780, A2780 (CAM assay in vivo model) | AKT | PCNA, MMP-2 and -9, IL-6 and -8, RAGE, p-PI3K, p-AKT, VEGF, HIF-1α | Cell proliferation and invasion inhibition, Apoptosis, RAGE-PI3K/AKT inhibition, Angiogenesis inhibition, AKT/HIF-1α inhibition |
[53,54] |
| Osteosarcoma | HOS | Bax, cleaved PARP, cleaved caspase-3, -8, and -9, Cyt c , ROS, p-c-Jun, c-Jun, p-JNK, | Colony-formation, Bcl-2, Survivin, MMP | Apoptosis, ROS/JNK activation | [55] |
| Prostate | DU145, DU145 xenograft, LNCaP | p53, IκBα, Bax, cleaved PARP, sub-G1 cells, ROS, p-JAK2, p-p38, p21 |
p-AKT, p-GSK-3β, p-NF-κB p65, p-IκBα, Bcl-2, XIAP, COX-2, p-STAT3 (Y705), Cyclin D1, Bcl-2, Bcl-xL, Mcl-1, Survivin, STAT3 activity, CDK2, CDK4, CDK6, Cdc25C, Cdc2, Cyclin B1 |
Apoptosis, STAT3 inhibition |
[25,56,57] |
| Skin (Melanoma) | SK-MEL-28, SK-MEL-28 xenograft | G1 arrest | p-AKT (Thr308), p-AKT (Ser473), p-GSK3β, Cyclin D1, Tumor volume | PI3K p110 binding, PI3K/AKT/p70S6K inhibition | [58] |
AIF, apoptosis-inducing factor; AP-1, activator protein 1; AKT, serine/threonine kinase (also known as protein kinase B, PKB); Bax, Bcl-2-associated X protein; Bak, Bcl-2 homologous antagonist/killer; Bcl-2, B-cell lymphoma 2; Bcl-xL, B-cell lymphoma-extra large; CAM assay, chorioallantoic membrane assay; CDK, cyclin-dependent kinase; COX-2, cyclooxygenase-2; Caspase, cysteine-aspartic protease; Chk1, checkpoint kinase 1; Cyt c, cytochrome c; DFF-45, DNA fragmentation factor 45; E-cadherin, epithelial cadherin; EGFR, epidermal growth factor receptor; EMT, epithelial-mesenchymal transition; ERK, extracellular signal-regulated kinase; FADD, Fas-associated death domain protein; Fas, first apoptosis signal receptor (CD95/APO-1); FasL, Fas ligand; GSK-3β, glycogen synthase kinase-3 beta; HIF-1α, hypoxia-inducible factor 1-alpha; IL-6, interleukin-6; IkBα, inhibitor of nuclear factor kappa-B alpha; JAK2, Janus kinase 2; JNK, c-Jun N-terminal kinase; MKK, mitogen-activated protein kinase kinase; MMP, mitochondrial membrane potential; MMP-2, matrix metalloproteinase-2; Mcl-1, myeloid cell leukemia-1; NF-κB, nuclear factor kappa-light-chain-enhancer of activated B cells; p-, phosphorylated; PARP, Poly (ADP-ribose) polymerase; PCNA, proliferating cell nuclear antigen; PD-L1, programmed death-ligand 1; PI3K, phosphoinositide 3-kinase; RAGE, receptor for advanced glycation end-products; RARγ, retinoic acid receptor gamma; RIP, receptor-interacting protein kinase; ROS, reactive oxygen species; SAPK, stress-activated protein kinase; STAT3, signal transducer and activator of transcription 3; VEGF, vascular endothelial growth factor; XIAP, X-linked inhibitor of apoptosis protein; u-PA, urokinase-type plasminogen activator.
3.1. In Vitro Effects: Apoptosis, Proliferation, and Cell Cycle Arrest
Acacetin exerts potent proapoptotic activity in diverse cancer cell lines by simultaneously activating the intrinsic (mitochondrial) and extrinsic (death receptor) apoptotic pathways. The intrinsic pathway involves disruption of mitochondrial homeostasis with cytochrome c release and activation of initiator caspase-9 followed by effector caspase-3/-7, accompanied by downregulation of anti-apoptotic proteins such as B-cell lymphoma 2 (Bcl-2) and B-cell lymphoma-extra large (Bcl-xL) and, in certain contexts, upregulation of pro-apoptotic mediators including Bcl-2-associated X protein (Bax) and Bcl-2 homologous antagonist/killer (Bak), thereby shifting the balance toward apoptosis [41,47,48,49,59]. The extrinsic pathway is promoted by the upregulation of the first apoptosis signal receptor (Fas, also known as CD95 or APO-1) and Fas ligand (FasL) signaling and the activation of caspase-8, which also engages the mitochondrial pathway via BH3-interacting domain death agonist (Bid) cleavage [47,60].
In breast cancer models, acacetin not only induces classical apoptosis but also triggers receptor-interacting protein kinase 1 (RIP1)-dependent necroptosis through sustained ERK1/2 phosphorylation, suggesting its potential to bypass apoptosis resistance mechanisms [42]. In colorectal carcinoma, suppression of the Wingless-related integration site (Wnt)/β-catenin/cellular myelocytomatosis oncogene (c-Myc) axis and nuclear β-catenin accumulation is coupled with apoptosis-inducing factor (AIF) translocation from mitochondria to the nucleus, leading to caspase-independent cell death [45].
Acacetin disrupts cell cycle progression in several cancer types, inducing G1 arrest in hepatocellular carcinoma via p53 and p21 upregulation [49,50] and S-phase arrest in colorectal carcinoma through the downregulation of cyclin A and cyclin-dependent kinase 2 (CDK2) [45]. These cell cycle regulatory effects are often accompanied by the inhibition of key pro-survival signaling pathways, including PI3K/AKT, MAPK/ERK, and signal transducer and activator of transcription 3 (STAT3) that are commonly dysregulated in malignant cells [25,42,46,56,61].
3.2. Anti-Metastatic and Anti-Angiogenic Activities
Metastasis suppression by acacetin has been demonstrated most clearly in gastric cancer, where the inhibition of epithelial–mesenchymal transition (EMT) is mediated through the downregulation of PI3K/AKT/Snail signaling. This results in the restoration of the epithelial marker E-cadherin and the reduction of mesenchymal markers such as N-cadherin, along with decreased expression and activity of matrix metalloproteinase (MMP)-2 and MMP-9, thereby reversing the EMT phenotype [46].
In non-small cell lung cancer (NSCLC) models, acacetin also attenuates tumor cell invasion by downregulating MMP-2 and urokinase-type plasminogen activator (u-PA) through inactivation of JNK and reduced NF-κB/activator protein-1 (AP-1) DNA-binding activity, ultimately suppressing extracellular matrix degradation [27].
Regarding angiogenesis, acacetin decreases vascular endothelial growth factor (VEGF) and hypoxia-inducible factor-1α (HIF-1α) by inhibiting the AKT/HIF-1α pathway, and this translates into significant suppression of ovarian cancer cell-induced angiogenesis and tumor growth in vivo (as demonstrated in animal models), thereby linking its molecular effects to anti-angiogenic efficacy in tumors [54].
3.3. In Vivo Efficacy and Safety Profiles
Multiple animal models and human tumor xenografts have been used to confirm the anticancer efficacy of acacetin in vitro [46,51,56,58]. In gastric cancer xenograft models, acacetin treatment significantly reduced tumor volume and weight, consistent with the suppression of the phosphoinositide 3-kinase/protein kinase B/Snail (PI3K/AKT/Snail) signaling pathway [46]. In a separate gastric cancer xenograft study, acacetin administration was associated with a lower Ki-67 proliferation index and increased levels of apoptotic markers, including cleaved caspase-3 and poly(ADP-ribose) polymerase (PARP), in tumor tissues, in parallel with the inhibition of STAT3 signaling. In lung cancer xenograft models, acacetin treatment led to a marked reduction in metastatic nodules in the lungs accompanied by decreased programmed death-ligand 1 (PD-L1) expression, suggesting both antiproliferative and immunomodulatory effects [51]. In prostate cancer xenografts, tumor regression is associated with the strong inhibition of STAT3 Tyr705 phosphorylation, achieved through the direct binding of acacetin to STAT3, which contributes to reduced tumor cell proliferation and increased apoptosis in tumor tissues [56]. In a skin cancer xenograft model, acacetin directly targets the p110 catalytic subunit of PI3K, resulting in the inhibition of AKT phosphorylation and significant suppression of tumor growth [58]. Collectively, these in vivo findings underscore the ability of acacetin to inhibit tumor progression across diverse cancer types by modulating multiple oncogenic pathways. Although detailed toxicological evaluations remain limited, no severe adverse effects or significant body weight loss has been reported in the available studies, suggesting a favorable preliminary safety profile that warrants further investigation in preclinical and clinical settings [23,46,51,56,58].
4. Network Pharmacology-Based Target Prediction of Acacetin
The predicted and validated targets of acacetin identified in previous network pharmacology and molecular docking studies are summarized in Table 2. This table lists each target protein, associated cancer types, implicated pathways, type of evidence (computational prediction, docking simulation, or experimental validation), functional outcomes, and corresponding references. Notably, several targets such as the epidermal growth factor receptor (EGFR), STAT3, and AKT1 appeared as high-ranking hub genes in network analyses and as experimentally confirmed molecular targets, underscoring the concordance between in silico prediction and empirical data.
4.1. Methodological Pipeline and Identified Targets
Previous network-pharmacology studies investigating acacetin have reported its potential multi-target spectrum in oncology through integrated computational workflows that combine ligand-based target prediction [68,69,70], disease-gene mapping [71,72,73,74], PPI analysis [75], enrichment analysis [76], and, in some cases, molecular docking [71,77]. As summarized in Table 2, these studies identified targets across multiple functional classes, including EGFR [12,62,63], AKT [62,64,67], proto-oncogene tyrosine-protein kinase Src (SRC) [62,67], transcription factors (STAT3 [56,67]), and molecular chaperones or inflammatory mediators such as (heat shock protein 90 alpha family class A member 1(HSP90AA1) [62,67] and tumor necrosis factor (TNF) [62].
In the reported workflows, the compound–target prediction stage is generally conducted using ligand-based virtual screening platforms such as SwissTargetPrediction[68], Search Tool for Interacting Chemicals (STITCH) [69], or the Similarity Ensemble Approach (SEA) [70], that generates candidate protein interactions based on chemical similarity metrics and known bioactivity profiles. Molecular docking simulations were performed using AutoDock Vina [71] to evaluate the binding affinities between acacetin and its predicted targets. To ensure relevance to oncology, predicted protein targets were cross-referenced with cancer-associated genes obtained from databases, including GeneCards (human gene compendium) [72], Online Mendelian Inheritance in Man (OMIM) [73], and the disease–gene association network database (DisGeNET) [74] (e.g., EGFR, AKT1 [62]).
The intersection sets of predicted and disease-related genes were subsequently analyzed through PPI network construction using the Search Tool for the Retrieval of Interacting Genes/Proteins (STRING) database [75], followed by network visualization and topological analysis in Cytoscape [76].
The resulting PPI networks enabled the visualization of interactions among acacetin-associated targets and identification of nodes with higher connectivity that were considered more likely to play central roles in the pharmacological activity of the compound. Several of these computationally prioritized targets such as EGFR [12,62,63], STAT3 [56,67], and AKT [62,64,67] are also supported by experimental studies as direct or indirect mediators of the anticancer effects of acacetin (see Section 5), thereby indicating concordance between network-based predictions and empirical evidence.
5. Molecular Docking and Experimental Validation of Key Targets
As outlined in Section 4 and summarized in Table 2, network pharmacology analyses highlighted several acacetin-associated targets of potential oncological relevance. In this section, we focus on the targets for which molecular docking simulations and experimental studies provide complementary evidence.
5.1. Overview of Key Network-Derived Targets
Acacetin was further examined against several network pharmacology–highlighted targets, most notably EGFR [12,62,63], STAT3 [56,67], and AKT [62,64,67], by using molecular docking simulations and experimental validation (Table 2). These proteins consistently emerged as central nodes in the predicted protein–protein interaction networks and were mechanistically linked to critical oncogenic signaling pathways, including PI3K/AKT, JAK/STAT, and MAPK/ERK [12,56,62,63,64,67].
5.2. EGFR
EGFR molecular docking using the crystal structure of the EGFR kinase domain predicted a stable binding conformation for acacetin within the kinase active site that was characterized by a favorable docking score. Target engagement was further supported by experimental validation using drug affinity responsive target stability (DARTS) and cellular thermal shift assays (CETSA), both of which demonstrated a direct interaction between acacetin and EGFR in gastric cancer cells [78,79]. Consistent with these in silico and biophysical findings, in vitro assays demonstrated that acacetin treatment reduced EGFR phosphorylation and suppressed activation of downstream STAT3 and ERK pathways, leading to decreased proliferation. In vivo, acacetin administration in gastric cancer xenografts significantly inhibited tumor growth in vivo without marked systemic toxicity [12]. In addition to these gastric cancer findings, docking studies in colorectal cancer models predicted a strong binding affinity of -8.3 kcal/mol [62], while bladder cancer models exhibited a binding affinity of -4.0 kcal/mol [63], further supporting the potential of acacetin as a broadly relevant EGFR-targeting compound across multiple tumor types.
5.3. STAT3
For STAT3, docking simulations demonstrated strong binding of acacetin to the SH2 domain of STAT3, stabilized by three hydrogen bonds and a cation–π interaction. Biophysical assays, including DARTS, CETSA, and pull-down experiments, confirmed a direct interaction between acacetin and STAT3 in prostate cancer DU145 cells. In vitro, acacetin treatment decreased p-STAT3 (Tyr705) levels, downregulated downstream STAT3-regulated anti-apoptotic and cell cycle–related proteins such as cyclin D1, Bcl-2, Bcl-xL, Mcl-1, and Survivin, and increased pro-apoptotic Bax expression. These effects were accompanied by PARP and caspase-3 cleavage, Annexin V positivity, and elevated reactive oxygen species (ROS) generation. In vivo administration of acacetin to DU145 xenograft models significantly suppressed tumor growth and reduced STAT3 Tyr705 phosphorylation. Collectively, these findings indicate that acacetin functions as a STAT3 inhibitor and is a potential drug candidate for targeting STAT3 in cancer therapy [56]. Furthermore, network pharmacology and molecular docking in gastric cancer models predicted a strong binding affinity of acacetin for STAT3 (-9.0 kcal/mol), reinforcing computational evidence for STAT3 as a potential target across multiple tumor types [67].
5.4. AKT
For AKT1, docking simulations predicted that acacetin occupies the ATP-binding pocket of the kinase domain, forming a hydrogen bond with Thr211 and exhibiting a strong binding affinity (-9.2 kcal/mol) that is indicative of potential competitive inhibition. Experimental validation in colorectal cancer HT-29 cells confirmed these predictions, indicating that acacetin treatment reduced p-AKT and p-PI3K levels, leading to the downregulation of Survivin and decreased cell migration and proliferation. Acacetin increases p53 expression and induces caspase-3 cleavage, resulting in enhanced apoptosis. These molecular effects are consistent with the inhibition of the PI3K/AKT/p53 signaling axis, supporting AKT1 as a direct target of acacetin [62]. Additionally, in lung cancer models acacetin inhibited HSP90AB1, a molecular chaperone that stabilizes AKT, leading to reduced p-AKT and p-mTOR levels, suppression of EMT markers, decreased migration and invasion, and restoration of E-cadherin expression. These effects were reversed by the HSP90 agonist terazosin [64]. Additionally, gastric cancer–based network pharmacology and molecular docking analyses reported a notable binding affinity of acacetin for AKT1 (-8.3 kcal/mol) [67], providing complementary computational support for its potential role as an AKT1-targeting compound across diverse malignancies.
5.5. Additional Network-Derived Targets
In addition to EGFR, STAT3, and AKT1, acacetin was associated with several other targets, and this was supported primarily by docking predictions, with a subset corroborated by experimental validation. In colorectal cancer–focused analyses, SRC exhibited a docking affinity of -6.9 kcal/mol, the docking affinity of HSP90AA1 was -7.1 kcal/mol, and that of TNF was -6.4 kcal/mol [62]. In bladder cancer models, IL-6 and MYC exhibited relatively weaker predicted affinities of -4.12 kcal/mol and -4.37 kcal/mol, respectively [63]. Among the targets with functional corroboration, prostaglandin-endoperoxide synthase 2 (PTGS2) in nasopharyngeal carcinoma demonstrated < -7 kcal/mol docking with a key Tyr385 contact and facilitated in vitro suppression of proliferation and migration [66]. In skin cancer models, acacetin directly targeted PI3K (p110), as validated by pull-down/kinase assays and xenografts, and docked to the ATP-binding site (H-bonds: Val828, Glu826, and Asp911; hydrophobic: Trp760, Ile777, and Tyr813) [58]. In breast cancer systems, PI3Kγ inhibition by acacetin was supported by kinase and Western blot assays, while docking analyses indicated binding at the ATP site through hydrogen bonds (Ser806, Ala885, and Val882) and hydrophobic interactions (Lys833 and Asp964) [65]. Additionally, a gastric cancer–based network pharmacology study reported “high” docking affinities (mostly < -7.3 kcal/mol) for a broader panel that included STAT3, AKT1, MAPK1, HSP90AA1, HRAS, SRC, PIK3CA, PIK3R1, FYN, and RHOA [67]. Although many of these associations remain supported primarily by computational predictions, they provide testable hypotheses where systematic investigation may reveal whether acacetin exerts pleiotropic anticancer effects via the concurrent modulation of multiple oncogenic signaling pathways.
6. Challenges and Future Prospects
Despite a growing body of preclinical evidence supporting the anticancer potential of acacetin, several challenges remain to be addressed before its effective clinical translation.
6.1. Bioavailability and Pharmacokinetics
Acacetin exhibits very low aqueous solubility and poor oral absorption, with instability across physiological pH levels and gastrointestinal fluids, resulting in limited intestinal uptake. Consequently, its oral bioavailability in rats is extremely low (~2%), while intravenous studies have demonstrated rapid plasma clearance and a short half-life (~1.5 h) [80,81]. These findings highlight the urgent need for the development of advanced formulation strategies.
6.2. Nanoparticle-Based Delivery and Structural Optimization
The pharmacokinetic limitations of acacetin have spurred the exploration of nanotechnology-based delivery approaches, including polymeric nanoparticles, liposomes, and solid lipid nanoparticles that can enhance bioavailability, prolong circulation time, and increase tumor selectivity via enhanced permeability and retention effect [80,82,83,84]. Surface modification with tumor-targeting ligands (e.g., folate, RGD [Arg-Glu-ASP] peptides, and antibodies) has demonstrated reduced off-target toxicity in preclinical studies, suggesting the potential of acacetin formulations [85,86,87]. Parallel to delivery strategies, structural optimization through medicinal chemistry, such as prodrug derivatization, may improve the stability and absorption, distribution, metabolism, and excretion (ADME) properties [88,89,90,91]. Collectively, nanoparticle-mediated delivery and structural modification represent synergistic strategies to overcome the pharmacokinetic barriers of acacetin and enable its development into clinically viable formulations [92,93].
6.3. Synthetic Derivatives and Structural Analogs of Acacetin
Synthetic derivatives and structural analogs of acacetin have been developed to overcome its pharmacokinetic limitations and enhance its anticancer efficacy. Aminoalkyl and Mannich base derivatives of acacetin-7-O-methyl ether demonstrated enzyme inhibitory activity, supporting the feasibility of structural diversification [94]. Simple glycosylated analogs such as linarin and linarin acetate exhibited much weaker activity than that of acacetin, indicating that certain glycosylation patterns can reduce anticancer potency [57]. In contrast, semisynthetic 7-O-β-D glycosides of acacetin prepared from naringin exhibited measurable cytotoxicity across multiple cancer cell lines (HL-60, SMMC-7721, A549, MCF-7, and SW480), suggesting that specific glycosylation patterns may preserve or even restore biological activity [95]. Methylated flavone derivatives exhibit improved hepatic metabolic stability and intestinal absorption compared to that of their unmethylated counterparts, supporting methylation as a viable strategy for orally available analogs [88,96]. In parallel, biotechnological methods enable scalable production of methylated flavonoids with favorable pharmacological profiles [90]. Collectively, these findings highlight the ability of methylation to consistently improve pharmacological stability, whereas the effects of glycosylation are context-dependent. In some cases, glycosylation reduces activity, whereas in others, it can yield analogs with favorable pharmacological profiles. These structural modifications complement the formulation strategies for enhancing the translational potential of acacetin [57,88,90,95].
6.4. Synergistic Drug Combinations
The most direct evidence of synergy comes from non-small-cell lung carcinoma, where acacetin enhances the therapeutic efficacy of doxorubicin by reducing clonogenic survival, inducing G2/M arrest, and suppressing multidrug resistance protein 1 (MDR1)-mediated efflux, thereby increasing the intracellular retention of doxorubicin [97]. Beyond this, although acacetin’s inhibition of EGFR [12,62,63], STAT3 [56,67], and PI3K/AKT [62,64,65,67] signaling, as well as its suppression of PD-L1 expression [51] and VEGF/HIF-1α–mediated angiogenesis [54], are mechanistically compatible with combination strategies, formal synergy studies remain lacking. Thus, a systematic evaluation of well-designed preclinical models is required to establish the role of acacetin as an adjuvant in combination therapies, both in vitro and in vivo.
6.5. Need for Clinical Translation
Despite accumulating preclinical evidence demonstrating the anticancer efficacy of acacetin in vitro and in various animal models, no clinical trials have evaluated its safety, tolerability, or therapeutic effectiveness in humans, and this represents a critical translational void. Most research remains confined to cell culture and xenograft models that report pro-apoptotic, anti-proliferative, and anti-metastatic effects; however, these effects remain clinically unverified [23,98]. Human pharmacokinetic and toxicity data are lacking, precluding the determination of dosing regimens, dose-limiting toxicities, or metabolic pathways [23,99]. Therefore, early phase safety studies are essential for clinical advancements [100].
7. Conclusions
Acacetin has exhibited promising preclinical anticancer activity, as supported by extensive in vitro and in vivo studies and has been systematically characterized using an integrated approach combining network pharmacology and molecular docking. Mechanistic investigations have demonstrated that acacetin suppresses tumor progression by inhibiting oncogenic signaling and angiogenesis while inducing apoptosis, cell cycle arrest, anti-metastatic activity, and other regulated cell death modes. These effects are mediated primarily through EGFR, STAT3, and AKT, as well as additional critical pathways. Molecular docking and experimental validation further confirmed the ability of acacetin to directly target key oncogenic proteins such as EGFR, STAT3, and AKT, underscoring its multi-target potential (Figure 2).
Despite this progress, significant challenges remain in clinical translation. Poor aqueous solubility, rapid systemic clearance, and metabolic instability hinder effective bioavailability, highlighting the need for innovative delivery systems, such as nanoparticles, and rational structural optimization. Additionally, synthetic derivatives and structural analogs of acacetin, particularly glycosylated and methylated forms, have been investigated to improve metabolic stability and absorption, and in certain cases cytotoxic potency, underscoring structural modification as a complementary strategy for future drug development. Moreover, while preliminary studies suggest synergy with standard chemotherapeutics such as doxorubicin, systematic evaluation of acacetin in well-designed preclinical combination models and early phase clinical trials is still required.
Taken together, the current evidence suggests that acacetin holds promise as a multi-target anticancer agent with potential applications in personalized medicine and combination therapies. Future research should prioritize translational studies integrating advanced formulation strategies, structural modifications of synthetic derivatives, rational drug combinations, and biomarker-driven patient stratification to establish acacetin as a clinically viable anticancer therapeutic agent.
Author Contributions
Conceptualization, J.Y.J., D.K., E.I., N.K.L., and N.D.K.; investigation, J.Y.J.; writing—original draft preparation, J.Y.J.; writing—review and editing, D.K., E.I., N.K.L., and N.D.K.; supervision, N.K.L. and N.D.K.; project administration, N.K.L.; funding acquisition, N.D.K. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by a two-year research grant by Pusan National University (N.K.L.)
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data presented in this study are available upon request from the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| ADME | Absorption, distribution, metabolism, and excretion |
| AIF | Apoptosis-inducing factor |
| AKT | Serine/threonine kinase (also known as protein kinase B, PKB) |
| AMPK | AMP-activated protein kinase |
| Bak | Bcl-2 homologous antagonist/killer |
| Bax | Bcl-2–associated X protein |
| Bcl-xL | B-cell lymphoma-extra large |
| Bcl-2 | B-cell lymphoma 2 |
| Bid | BH3-interacting domain death agonist |
| CDK2 | Cyclin-dependent kinase 2 |
| CETSA | Cellular thermal shift assay |
| c-Myc | Cellular myelocytomatosis oncogene |
| DARTS | Drug affinity responsive target stability |
| EGFR | Epidermal growth factor receptor |
| EMT | Epithelial–mesenchymal transition |
| ERK | Extracellular signal-regulated kinase |
| FYN | Proto-oncogene tyrosine-protein kinase Fyn |
| HIF-1α | Hypoxia-inducible factor-1 alpha |
| HRAS | Harvey rat sarcoma viral oncogene homolog |
| HSP90AA1 | Heat shock protein 90 alpha family class A member 1 |
| IL-6 | Interleukin-6 |
| JAK | Janus kinase |
| JNK | c-Jun N-terminal kinase |
| MDR1 | Multidrug resistance protein 1 |
| MAPK | Mitogen-activated protein kinase |
| mTOR | Mechanistic target of rapamycin |
| MYC | Myelocytomatosis oncogene |
| NF-κB | Nuclear factor kappa-light-chain-enhancer of activated B cells |
| Nrf2 | Nuclear factor erythroid 2–related factor 2 |
| PAMPA | Parallel artificial membrane permeability assay |
| PD-L1 | Programmed death-ligand 1 |
| PIK3CA | Phosphatidylinositol-4,5-bisphosphate 3-kinase catalytic subunit alpha |
| PIK3R1 | Phosphoinositide-3-kinase regulatory subunit 1 |
| PI3K | Phosphoinositide 3-kinase |
| PPI | Protein–protein interaction |
| PTGS2 (COX-2) | Prostaglandin-endoperoxide synthase 2 (cyclooxygenase-2) |
| RHOA | Ras homolog family member A |
| RIP1 | Receptor-interacting protein kinase 1 |
| SIRT1 | Sirtuin 1 |
| STAT3 | Signal transducer and activator of transcription 3 |
| TNF | Tumor necrosis factor |
| VEGF | Vascular endothelial growth factor |
| Wnt | Wingless-related integration site |
References
- Zafar, A.; Khatoon, S.; Khan, M.J.; Abu, J.; Naeem, A. Advancements and limitations in traditional anti-cancer therapies: A comprehensive review of surgery, chemotherapy, radiation therapy, and hormonal therapy. Discov. Oncol. 2025, 16, 607. [Google Scholar] [CrossRef]
- Bray, F.; Laversanne, M.; Sung, H.; Ferlay, J.; Siegel, R.L.; Soerjomataram, I.; Jemal, A. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA: Cancer J. Clin. 2024, 74, 229–263. [Google Scholar] [CrossRef]
- Bray, F.; Laversanne, M.; Weiderpass, E.; Soerjomataram, I. The ever-increasing importance of cancer as a leading cause of premature death worldwide. Cancer 2021, 127, 3029–3030. [Google Scholar] [CrossRef]
- Jang, J.Y.; Kim, D.; Kim, N.D. Recent developments in combination chemotherapy for colorectal and breast cancers with topoisomerase inhibitors. Int. J. Mol. Sci. 2023, 24, 8457. [Google Scholar] [CrossRef]
- Jang, J.Y.; Kim, D.; Im, E.; Kim, N.D. Etoposide as a key therapeutic agent in lung cancer: Mechanisms, efficacy, and emerging strategies. Int. J. Mol. Sci. 2025, 26, 796. [Google Scholar] [CrossRef]
- Li, Y.; Liu, F.; Cai, Q.; Deng, L.; Ouyang, Q.; Zhang, X.H.F.; Zheng, J. Invasion and metastasis in cancer: Molecular insights and therapeutic targets. Signal Transduct. Target. Ther. 2025, 10, 57. [Google Scholar] [CrossRef]
- Jang, J.Y.; Im, E.; Kim, N.D. Mechanism of resveratrol-induced programmed cell death and new drug discovery against cancer: A review. Int. J. Mol. Sci. 2022, 23, 13689. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Zhou, H.; Tan, L.; Siu, K.T.H.; Guan, X.-Y. Exploring treatment options in cancer: Tumor treatment strategies. Signal Transduct. Target. Ther. 2024, 9, 175. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Kantoff, P.W.; Wooster, R.; Farokhzad, O.C. Cancer nanomedicine: Progress, challenges and opportunities. Nat. Rev. Cancer 2017, 17, 20–37. [Google Scholar] [CrossRef] [PubMed]
- Rudzińska, A.; Juchaniuk, P.; Oberda, J.; Wiśniewska, J.; Wojdan, W.; Szklener, K.; Mańdziuk, S. Phytochemicals in cancer treatment and cancer prevention—review on epidemiological data and clinical trials. Nutrients 2023, 15, 1896. [Google Scholar] [CrossRef]
- Zhou, Z.; Li, T.; Qin, H.; Wang, X.; He, S.; Fan, Z.; Ye, Q.; Du, Y. Acacetin as a natural cardiovascular therapeutic: Mechanisms and preclinical evidence. Front. Pharmacol. 2025, 16, 1493981. [Google Scholar] [CrossRef]
- Zhang, G.; Dong, J.; Lu, L.; Liu, Y.; Hu, D.; Wu, Y.; Zhao, A.; Xu, H. Acacetin exerts antitumor effects on gastric cancer by targeting EGFR. Front. Pharmacol. 2023, 14, 1121643. [Google Scholar] [CrossRef]
- Noor, F.; Tahir Ul Qamar, M.; Ashfaq, U.A.; Albutti, A.; Alwashmi, A.S.S.; Aljasir, M.A. Network pharmacology approach for medicinal plants: Review and assessment. Pharmaceuticals 2022, 15, 572. [Google Scholar] [CrossRef]
- Pinzi, L.; Rastelli, G. Molecular docking: Shifting paradigms in drug discovery. Int. J. Mol. Sci. 2019, 20, 4331. [Google Scholar] [CrossRef] [PubMed]
- Ahn, Y.R.; Jang, J.Y.; Kang, Y.J.; Oh, H.J.; Kang, M.K.; Yoon, D.; Kim, H.S.; Moon, H.R.; Chung, H.Y.; Kim, N.D. MHY446 induces apoptosis via reactive oxygen species-mediated endoplasmic reticulum stress in HCT116 human colorectal cancer cells. J. Chemother. 2024, 36, 483–500. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.J.; Jang, J.Y.; Kwon, Y.H.; Lee, J.H.; Lee, S.; Park, Y.; Jung, Y.-S.; Im, E.; Moon, H.R.; Chung, H.Y. MHY2245, a sirtuin inhibitor, induces cell cycle arrest and apoptosis in HCT116 human colorectal cancer cells. Int. J. Mol. Sci. 2022, 23, 1590. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.Y.; Kang, Y.J.; Sung, B.; Kim, M.J.; Park, C.; Kang, D.; Moon, H.R.; Chung, H.Y.; Kim, N.D. MHY440, a novel topoisomerase Ι inhibitor, induces cell cycle arrest and apoptosis via a ROS-dependent DNA damage signaling pathway in AGS human gastric cancer cells. Molecules 2018, 24, 96. [Google Scholar] [CrossRef]
- Eom, S.H.; Liu, S.; Su, M.; Noh, T.H.; Hong, J.; Kim, N.D.; Chung, H.Y.; Yang, M.H.; Jung, J.H. Synthesis of phthalimide derivatives as potential PPAR-γ ligands. Mar. Drugs 2016, 14, 112. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Yan, H.; Li, H.; Feng, Y.; Sun, W.; Ren, Y.; Ma, L.; Zeng, W.; Huang, F.; Jiang, Z.; et al. Use of network pharmacology and molecular docking to explore the mechanism of action of curcuma in the treatment of osteosarcoma. Sci. Rep. 2023, 13, 9569. [Google Scholar] [CrossRef]
- Lin, Z.; Zhang, Z.; Ye, X.; Zhu, M.; Li, Z.; Chen, Y.; Huang, S. Based on network pharmacology and molecular docking to predict the mechanism of Huangqi in the treatment of castration-resistant prostate cancer. PloS one 2022, 17, e0263291. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, L.; Li, L. The feasibility of using the compound kushen injection to treat cervical cancer based on network pharmacology and transcriptomics. Medicine 2023, 102, e35135. [Google Scholar] [CrossRef] [PubMed]
- Veitch, N.C.; Elliott, P.C.; Kite, G.C.; Lewis, G.P. Flavonoid glycosides of the black locust tree, Robinia pseudoacacia (Leguminosae). Phytochemistry 2010, 71, 479–486. [Google Scholar] [CrossRef]
- Singh, S.; Gupta, P.; Meena, A.; Luqman, S. Acacetin, a flavone with diverse therapeutic potential in cancer, inflammation, infections and other metabolic disorders. Food Chem. Toxicol. 2020, 145, 111708. [Google Scholar] [CrossRef]
- Alibi, S.; Crespo, D.; Navas, J. Plant-derivatives small molecules with antibacterial activity. Antibiotics 2021, 10, 231. [Google Scholar] [CrossRef]
- Kim, H.R.; Park, C.G.; Jung, J.Y. Acacetin (5, 7-dihydroxy-4-methoxyflavone) exhibits in vitro and in vivo anticancer activity through the suppression of NF-κB/Akt signaling in prostate cancer cells. Int. J. Mol. Med. 2014, 33, 317–324. [Google Scholar] [CrossRef]
- Wang, W.; Renquan, Z. Acacetin restrains the malignancy of esophageal squamous carcinoma cells via regulating JAK2/STAT3 pathway. Chem. Biol. Drug Des. 2023, 102, 564–573. [Google Scholar] [CrossRef] [PubMed]
- Fong, Y.; Shen, K.H.; Chiang, T.A.; Shih, Y.W. Acacetin inhibits TPA-nduced MMP-2 and u-PA expressions of human lung cancer cells through inactivating JNK signaling pathway and reducing binding activities of NF-kappaB and AP-1. J. Food Sci. 2010, 75, H30–H38. [Google Scholar] [CrossRef] [PubMed]
- Chien, S.-T.; Lin, S.-S.; Wang, C.-K.; Lee, Y.-B.; Chen, K.-S.; Fong, Y.; Shih, Y.-W. Acacetin inhibits the invasion and migration of human non-small cell lung cancer A549 cells by suppressing the p38α MAPK signaling pathway. Mol. Cell. Biochem. 2011, 350, 135–148. [Google Scholar] [CrossRef]
- Pan, M.-H.; Lai, C.-S.; Wang, Y.-J.; Ho, C.-T. Acacetin suppressed LPS-induced up-expression of iNOS and COX-2 in murine macrophages and TPA-induced tumor promotion in mice. Biochem. Pharmacol. 2006, 72, 1293–1303. [Google Scholar] [CrossRef]
- Wu, H.-J.; Sun, H.-Y.; Wu, W.; Zhang, Y.-H.; Qin, G.-W.; Li, G.-R. Properties and molecular determinants of the natural flavone acacetin for blocking hKv4. 3 channels. PloS one 2013, 8, e57864. [Google Scholar] [CrossRef]
- Li, G.-R.; Wang, H.-B.; Qin, G.-W.; Jin, M.-W.; Tang, Q.; Sun, H.-Y.; Du, X.-L.; Deng, X.-L.; Zhang, X.-H.; Chen, J.-B.; et al. Acacetin, a natural flavone, selectively inhibits human atrial repolarization potassium currents and prevents atrial fibrillation in dogs. Circulation 2008, 117, 2449–2457. [Google Scholar] [CrossRef]
- Wu, Y.; Song, F.; Li, Y.; Li, J.; Cui, Y.; Hong, Y.; Han, W.; Wu, W.; Lakhani, I.; Li, G. Acacetin exerts antioxidant potential against atherosclerosis through Nrf2 pathway in apoE−/− Mice. J. Cell. Mol. Med. 2021, 25, 521–534. [Google Scholar] [CrossRef]
- Simões, L.; Maciel, G.; Brandão, G.; Kroon, E.; Castilho, R.; Oliveira, A. Antiviral activity of Distictella elongata (Vahl) Urb.(Bignoniaceae), a potentially useful source of anti-dengue drugs from the state of Minas Gerais, Brazil. Lett. Appl. Microbiol. 2011, 53, 602–607. [Google Scholar] [CrossRef]
- Critchfield, J.W.; Butera, S.T.; Folks, T.M. Inhibition of HIV activation in latently infected cells by flavonoid compounds. AIDS Res. Hum. Retroviruses 1996, 12, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.-Q.; Chen, K.; Shi, Q.; Kilkuskie, R.E.; Cheng, Y.-C.; Lee, K.-H. Anti-AIDS agents, 10. acacetin-7-O-β-D-galactopyranoside, an anti-HIV principle from chrysanthemum morifolium and a structure-activity correlation with some related flavonoids. J. Nat. Prod. 1994, 57, 42–51. [Google Scholar] [CrossRef]
- Wu, Q.; Yu, C.; Yan, Y.; Chen, J.; Zhang, C.; Wen, X. Antiviral flavonoids from Mosla scabra. Fitoterapia 2010, 81, 429–433. [Google Scholar] [CrossRef]
- Cha, J.-D.; Choi, S.-M.; Park, J.H. Combination of acacetin with antibiotics against methicillin resistant Staphylococcus aureus isolated from clinical specimens. Adv. Biosci. Biotechnol. 2014, 5, 398–408. [Google Scholar] [CrossRef]
- Komape, N.P.M.; Aderogba, M.; Bagla, V.P.; Masoko, P.; Eloff, J.N. Anti-bacterial and anti-oxidant activities of leaf extracts of Combretum vendee (combretecacea) and the isolation of an anti-bacterial compound. Afr. J. Tradit. Complement. Altern. Med. 2014, 11, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Bi, C.; Dong, X.; Zhong, X.; Cai, H.; Wang, D.; Wang, L. Acacetin protects mice from Staphylococcus aureus bloodstream infection by inhibiting the activity of sortase A. Molecules 2016, 21, 1285. [Google Scholar] [CrossRef]
- Liou, C.-J.; Wu, S.-J.; Chen, L.-C.; Yeh, K.-W.; Chen, C.-Y.; Huang, W.-C. Acacetin from traditionally used Saussurea involucrata Kar. et Kir. suppressed adipogenesis in 3T3-L1 adipocytes and attenuated lipid accumulation in obese mice. Front. Pharmacol. 2017, 8, 589. [Google Scholar] [CrossRef] [PubMed]
- Shim, H.-Y.; Park, J.-H.; Paik, H.-D.; Nah, S.-Y.; Kim, D.S.; Han, Y.S.J.M.; cells. Acacetin-induced apoptosis of human breast cancer MCF-7 cells involves caspase cascade, mitochondria-mediated death signaling and SAPK/JNK1/2-c-Jun activation. 2007, 24, 95-104.
- Kandhari, K.; Mishra, J.P.N.; Agarwal, R.; Singh, R.P. Acacetin induces sustained ERK1/2 activation and RIP1-dependent necroptotic death in breast cancer cells. Toxicol. Appl. Pharmacol. 2023, 462, 116409. [Google Scholar] [CrossRef] [PubMed]
- Kandhari, K.; Mishra, J.; Singh, R. Acacetin inhibits cell proliferation, survival, and migration in human breast cancer cells. Int. J. Pharm. Biol. Sci. 2019, 9, 443–452. [Google Scholar] [CrossRef]
- Aslan, B.T.; Ertugrul, B.; Iplik, E.S.; Cakmakoglu, B. Apoptotic effects of acacetin in human colon cancer HT-29 and HCT 116 cells. J. Cancer Res. Ther. 2021, 17, 1479–1482. [Google Scholar] [CrossRef]
- Prasad, N.; Sharma, J.R.; Yadav, U.C.S. Induction of growth cessation by acacetin via β-catenin pathway and apoptosis by apoptosis inducing factor activation in colorectal carcinoma cells. Mol. Biol. Rep. 2020, 47, 987–1001. [Google Scholar] [CrossRef]
- Zhang, G.; Li, Z.; Dong, J.; Zhou, W.; Zhang, Z.; Que, Z.; Zhu, X.; Xu, Y.; Cao, N.; Zhao, A. Acacetin inhibits invasion, migration and TGF-β1-induced EMT of gastric cancer cells through the PI3K/Akt/Snail pathway. BMC Complement. Med. Ther. 2022, 22, 10. [Google Scholar] [CrossRef]
- Pan, M.-H.; Lai, C.-S.; Hsu, P.-C.; Wang, Y.-J. Acacetin induces apoptosis in human gastric carcinoma cells accompanied by activation of caspase cascades and production of reactive oxygen species. J. Agric. Food Chem. 2005, 53, 620–630. [Google Scholar] [CrossRef]
- Alfwuaires, M.; Elsawy, H.; Sedky, A. Acacetin inhibits cell proliferation and induces apoptosis in human hepatocellular carcinoma cell lines. Molecules 2022, 27, 5361. [Google Scholar] [CrossRef]
- Hsu, Y.-L.; Kuo, P.-L.; Lin, C.-C. Acacetin inhibits the proliferation of Hep G2 by blocking cell cycle progression and inducing apoptosis. Biochem. Pharmacol. 2004, 67, 823–829. [Google Scholar] [CrossRef] [PubMed]
- Zeng, W.; Zhang, C.; Cheng, H.; Wu, Y.-L.; Liu, J.; Chen, Z.; Huang, J.-g.; Ericksen, R.E.; Chen, L.; Zhang, H.; et al. Targeting to the non-genomic activity of retinoic acid receptor-gamma by acacetin in hepatocellular carcinoma. Sci. Rep. 2017, 7, 348. [Google Scholar] [CrossRef]
- Li, J.; Zhong, X.; Zhao, Y.; Shen, J.; Xiao, Z.; Pilapong, C. Acacetin inhibited non-small-cell lung cancer (NSCLC) cell growth via upregulating miR-34a in vitro and in vivo. Sci. Rep. 2024, 14, 2348. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.-L.; Kuo, P.-L.; Liu, C.-F.; Lin, C.-C. Acacetin-induced cell cycle arrest and apoptosis in human non-small cell lung cancer A549 cells. Cancer letters 2004, 212, 53–60. [Google Scholar] [CrossRef]
- Tian, M.; Tang, Y.; Huang, T.; Liu, Y.; Pan, Y. Amelioration of human peritoneal mesothelial cell co-culture-evoked malignant potential of ovarian cancer cells by acacetin involves LPA release-activated RAGE-PI3K/AKT signaling. Cell. Mol. Biol. lett. 2021, 26, 51. [Google Scholar] [CrossRef]
- Liu, L.Z.; Jing, Y.; Jiang, L.L.; Jiang, X.E.; Jiang, Y.; Rojanasakul, Y.; Jiang, B.H. Acacetin inhibits VEGF expression, tumor angiogenesis and growth through AKT/HIF-1α pathway. Biochem. Biophys. Res. Commun. 2011, 413, 299–305. [Google Scholar] [CrossRef]
- Wang, S.; Lin, B.; Liu, W.; Wei, G.; Li, Z.; Yu, N.; Xue, X.; Ji, G. Acacetin induces apoptosis in human osteosarcoma cells by modulation of ROS/JNK activation. Drug Des. Devel. Ther. 2020, 5077–5085. [Google Scholar] [CrossRef] [PubMed]
- Yun, S.; Lee, Y.-J.; Choi, J.; Kim, N.D.; Han, D.C.; Kwon, B.-M. Acacetin inhibits the growth of STAT3-activated DU145 prostate cancer cells by directly binding to signal transducer and activator of transcription 3 (STAT3). Molecules 2021, 26, 6204. [Google Scholar] [CrossRef]
- Singh, R.P.; Agrawal, P.; Yim, D.; Agarwal, C.; Agarwal, R. Acacetin inhibits cell growth and cell cycle progression, and induces apoptosis in human prostate cancer cells: Structure–activity relationship with linarin and linarin acetate. Carcinogenesis 2005, 26, 845–854. [Google Scholar] [CrossRef]
- Jung, S.K.; Kim, J.E.; Lee, S.Y.; Lee, M.H.; Byun, S.; Kim, Y.A.; Lim, T.G.; Reddy, K.; Huang, Z.; Bode, A.M.; et al. The P110 subunit of PI3-K is a therapeutic target of acacetin in skin cancer. Carcinogenesis 2014, 35, 123–130. [Google Scholar] [CrossRef]
- Jang, J.Y.; Sung, B.; Kim, N.D. Role of induced programmed cell death in the chemopreventive potential of apigenin. Int. J. Mol. Sci. 2022, 23, 3757. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.Y.; Im, E.; Choi, Y.H.; Kim, N.D. Mechanism of bile acid-induced programmed cell death and drug discovery against cancer: A review. Int. J. Mol. Sci. 2022, 23, 7184. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.Y.; Im, E.; Kim, N.D. Therapeutic potential of bioactive components from Scutellaria baicalensis Georgi in inflammatory bowel disease and colorectal cancer: A review. Int. J. Mol. Sci. 2023, 24, 1954. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.-L.; Yang, B.-W.; Wang, X.-Y.; Chen, X.; Li, W.-D.; Zhai, H.-Y.; Wu, Y.; Cui, M.-Y.; Wu, J.-H.; Meng, Q.-H.J.W.J.o.G.O. Targeting colorectal cancer with Herba Patriniae and Coix seed: Network pharmacology, molecular docking, and in vitro validation. 2024, 16, 3539. [CrossRef]
- Yu, H.; Dong, Y.; Zou, G.-c.; Yang, Y.-j.; Liu, M.-z.; Han, C.-h. Exploring the molecular mechanism of Xiao Ji (Cirsium setosum) in treating bladder cancer using network pharmacology and molecular docking. Asian Biomed.(Res. Rev. News) 2025, 19, 94. [Google Scholar] [CrossRef]
- Guo, Z.; Yin, H.; Wu, T.; Wu, S.; Liu, L.; Zhang, L.; He, Y.; Zhang, R.; Liu, N. Study on the mechanism of Cortex Lycii on lung cancer based on network pharmacology combined with experimental validation. J. Ethnopharmacol. 2022, 293, 115280. [Google Scholar] [CrossRef]
- Zhang, H.-W.; Hu, J.-J.; Fu, R.-Q.; Liu, X.; Zhang, Y.-H.; Li, J.; Liu, L.; Li, Y.-N.; Deng, Q.; Luo, Q.-S. Flavonoids inhibit cell proliferation and induce apoptosis and autophagy through downregulation of PI3Kγ mediated PI3K/AKT/mTOR/p70S6K/ULK signaling pathway in human breast cancer cells. Sci. Rep. 2018, 8, 11255. [Google Scholar] [CrossRef]
- Huang, T.; Han, B.; Tao, Z.; Chen, X. Network pharmacology and experimental validation-based investigation of the underlying mechanism of Yi-Yi-Fu-Zi-Bai-Jiang-San of nasopharyngeal carcinoma. J. Cancer 2025, 16, 2212. [Google Scholar] [CrossRef]
- Lu, X.; Zheng, Y.; Wen, F.; Huang, W.; Chen, X.; Ruan, S.; Gu, S.; Hu, Y.; Teng, Y.; Shu, P. Study of the active ingredients and mechanism of Sparganii rhizoma in gastric cancer based on HPLC-Q-TOF–MS/MS and network pharmacology. Sci. Rep. 2021, 11, 1905. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissTargetPrediction: Updated data and new features for efficient prediction of protein targets of small molecules. Nucleic acids Res. 2019, 47, W357–W364. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Santos, A.; Von Mering, C.; Jensen, L.J.; Bork, P.; Kuhn, M. STITCH 5: Augmenting protein–chemical interaction networks with tissue and affinity data. Nucleic Acids Res. 2016, 44, D380–D384. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Liang, L.; Yin, Z.; Lin, J. Improving chemical similarity ensemble approach in target prediction. J. Cheminform. 2016, 8, 20. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- Stelzer, G.; Rosen, N.; Plaschkes, I.; Zimmerman, S.; Twik, M.; Fishilevich, S.; Stein, T.I.; Nudel, R.; Lieder, I.; Mazor, Y. The GeneCards suite: From gene data mining to disease genome sequence analyses. Curr. Protoc. Bioinformatics 2016, 54, 1.30–1. [Google Scholar] [CrossRef] [PubMed]
- Amberger, J.S.; Bocchini, C.A.; Schiettecatte, F.; Scott, A.F.; Hamosh, A. OMIM.org: Online Mendelian Inheritance in Man (OMIM®), an online catalog of human genes and genetic disorders. Nucleic Acids Res. 2014, 43, D789–D798. [Google Scholar] [CrossRef] [PubMed]
- Piñero, J.; Bravo, À.; Queralt-Rosinach, N.; Gutiérrez-Sacristán, A.; Deu-Pons, J.; Centeno, E.; García-García, J.; Sanz, F.; Furlong, L.I. DisGeNET: A comprehensive platform integrating information on human disease-associated genes and variants. Nucleic Acids Res. 2016, 45, D833–D839. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P.; et al. The STRING database in 2021: Customizable protein–protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2020, 49, D605–D612. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Friesner, R.A.; Banks, J.L.; Murphy, R.B.; Halgren, T.A.; Klicic, J.J.; Mainz, D.T.; Repasky, M.P.; Knoll, E.H.; Shelley, M.; Perry, J.K. Glide: A new approach for rapid, accurate docking and scoring. 1. method and assessment of docking accuracy. J. Med. Chem. 2004, 47, 1739–1749. [Google Scholar] [CrossRef] [PubMed]
- Lomenick, B.; Hao, R.; Jonai, N.; Chin, R.M.; Aghajan, M.; Warburton, S.; Wang, J.; Wu, R.P.; Gomez, F.; Loo, J.A. Target identification using drug affinity responsive target stability (DARTS). Proc. Natl. Acad. Sci. USA 2009, 106, 21984–21989. [Google Scholar] [CrossRef]
- Molina, D.M.; Jafari, R.; Ignatushchenko, M.; Seki, T.; Larsson, E.A.; Dan, C.; Sreekumar, L.; Cao, Y.; Nordlund, P. Monitoring drug target engagement in cells and tissues using the cellular thermal shift assay. Science 2013, 341, 84–87. [Google Scholar] [CrossRef]
- Han, D.G.; Cha, E.; Joo, J.; Hwang, J.S.; Kim, S.; Park, T.; Jeong, Y.S.; Maeng, H.J.; Kim, S.B.; Yoon, I.S. Investigation of the factors responsible for the poor oral bioavailability of acacetin in rats: Physicochemical and biopharmaceutical aspects. Pharmaceutics 2021, 13. [Google Scholar] [CrossRef]
- Fan, L.-h.; Li, X.; Chen, D.-y.; Zhang, N.; Wang, Y.; Shan, Y.; Hu, Y.; Xu, R.-a.; Jin, J.; Ge, R.-S. Determination of acacetin in rat plasma by UPLC-MS/MS and its application to a pharmacokinetic study. J. Chromatogr. B 2015, 986, 18–22. [Google Scholar] [CrossRef]
- Subhan, M.A.; Yalamarty, S.S.K.; Filipczak, N.; Parveen, F.; Torchilin, V.P. Recent advances in tumor targeting via EPR effect for cancer treatment. J. Pers. Med. 2021, 11, 571. [Google Scholar] [CrossRef] [PubMed]
- Sun, R.; Xiang, J.; Zhou, Q.; Piao, Y.; Tang, J.; Shao, S.; Zhou, Z.; Bae, Y.H.; Shen, Y. The tumor EPR effect for cancer drug delivery: Current status, limitations, and alternatives. Adv. Drug Deliv. Rev. 2022, 191, 114614. [Google Scholar] [CrossRef]
- Ranjbar, S.; Emamjomeh, A.; Sharifi, F.; Zarepour, A.; Aghaabbasi, K.; Dehshahri, A.; Sepahvand, A.M.; Zarrabi, A.; Beyzaei, H.; Zahedi, M.M. Lipid-based delivery systems for flavonoids and flavonolignans: Liposomes, nanoemulsions, and solid lipid nanoparticles. Pharmaceutics 2023, 15, 1944. [Google Scholar] [CrossRef]
- Yan, S.; Na, J.; Liu, X.; Wu, P. Different targeting ligands-mediated drug delivery systems for tumor therapy. Pharmaceutics 2024, 16, 248. [Google Scholar] [CrossRef] [PubMed]
- Jurczyk, M.; Jelonek, K.; Musiał-Kulik, M.; Beberok, A.; Wrześniok, D.; Kasperczyk, J. Single-versus dual-targeted nanoparticles with folic acid and biotin for anticancer drug delivery. Pharmaceutics 2021, 13, 326. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Tang, D.; Cui, J.; Jiang, H.; Yu, J.; Guo, Z. RGD-based self-assembling nanodrugs for improved tumor therapy. Front. Pharmacol. 2024, 15, 1477409. [Google Scholar] [CrossRef] [PubMed]
- Walle, T. Methylation of dietary flavones greatly improves their hepatic metabolic stability and intestinal absorption. Mol. Pharm. 2007, 4, 826–832. [Google Scholar] [CrossRef]
- Tang, S.; Wang, B.; Liu, X.; Xi, W.; Yue, Y.; Tan, X.; Bai, J.; Huang, L. Structural insights and biological activities of flavonoids: Implications for novel applications. Food Front. 2025, 6, 218–247. [Google Scholar] [CrossRef]
- Koirala, N.; Thuan, N.H.; Ghimire, G.P.; Thang, D.V.; Sohng, J.K. Methylation of flavonoids: Chemical structures, bioactivities, progress and perspectives for biotechnological production. Enzyme Microb. Technol. 2016, 86, 103–116. [Google Scholar] [CrossRef]
- Gogineni, V.; Nael, M.A.; Chaurasiya, N.D.; Elokely, K.M.; McCurdy, C.R.; Rimoldi, J.M.; Cutler, S.J.; Tekwani, B.L.; León, F. Computationally assisted lead optimization of novel potent and selective MAO-B inhibitors. Biomedicines 2021, 9, 1304. [Google Scholar] [CrossRef]
- Zverev, Y.F.; Rykunova, A.Y. Modern nanocarriers as a factor in increasing the bioavailability and pharmacological activity of flavonoids. Appl. Biochem. Microbiol. 2022, 58, 1002–1020. [Google Scholar] [CrossRef] [PubMed]
- Stevens Barrón, J.C.; Chapa González, C.; Álvarez Parrilla, E.; De la Rosa, L.A. Nanoparticle-mediated delivery of flavonoids: Impact on proinflammatory cytokine production: A systematic review. Biomolecules 2023, 13. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.R.; Men, X.; Gao, X.H.; Liu, L.B.; Fan, H.Q.; Xia, X.H.; Wang, Q.A. Discovery of potent and selective acetylcholinesterase (AChE) inhibitors: acacetin 7-O-methyl ether Mannich base derivatives synthesised from easy access natural product naringin. Nat. Prod. Res. 2018, 32, 743–747. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Chen, L.; Cai, S.; Wang, Q. Semisynthesis of apigenin and acacetin-7-O-β-D-glycosides from naringin and their cytotoxic activities. Carbohydr. Res. 2012, 357, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Walle, T. Methylation of dietary flavones increases their metabolic stability and chemopreventive effects. Int. J. Mol. Sci. 2009, 10, 5002–5019. [Google Scholar] [CrossRef]
- Punia, R.; Raina, K.; Agarwal, R.; Singh, R.P. Acacetin enhances the therapeutic efficacy of doxorubicin in non-small-cell lung carcinoma cells. PLoS One 2017, 12, e0182870. [Google Scholar] [CrossRef]
- Chen, K.; Gao, Z. Acacetin, a natural flavone with potential in improving liver disease based on its anti-inflammation, anti-cancer, anti-infection and other effects. Molecules 2024, 29, 4872. [Google Scholar] [CrossRef]
- Fasolo, A.; Sessa, C. Translational research in phase I trials. Clini. Transl. Oncol. 2009, 11, 580–588. [Google Scholar] [CrossRef]
- Goldblatt, E.M.; Lee, W.H. From bench to bedside: the growing use of translational research in cancer medicine. Am. J. Transl. Res. 2010, 2, 1–18. [Google Scholar]
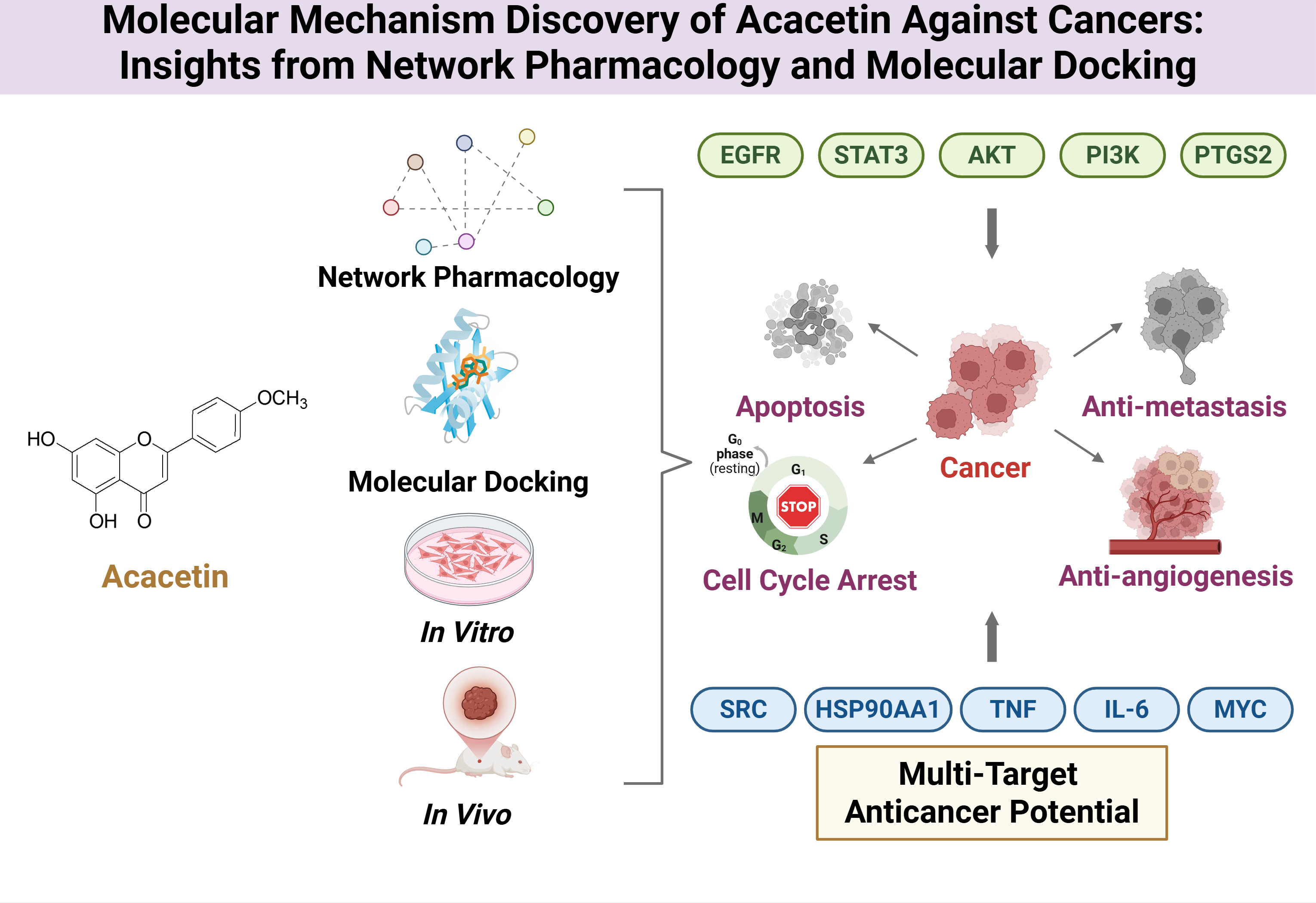
Figure 1.
Molecular structure of acacetin.
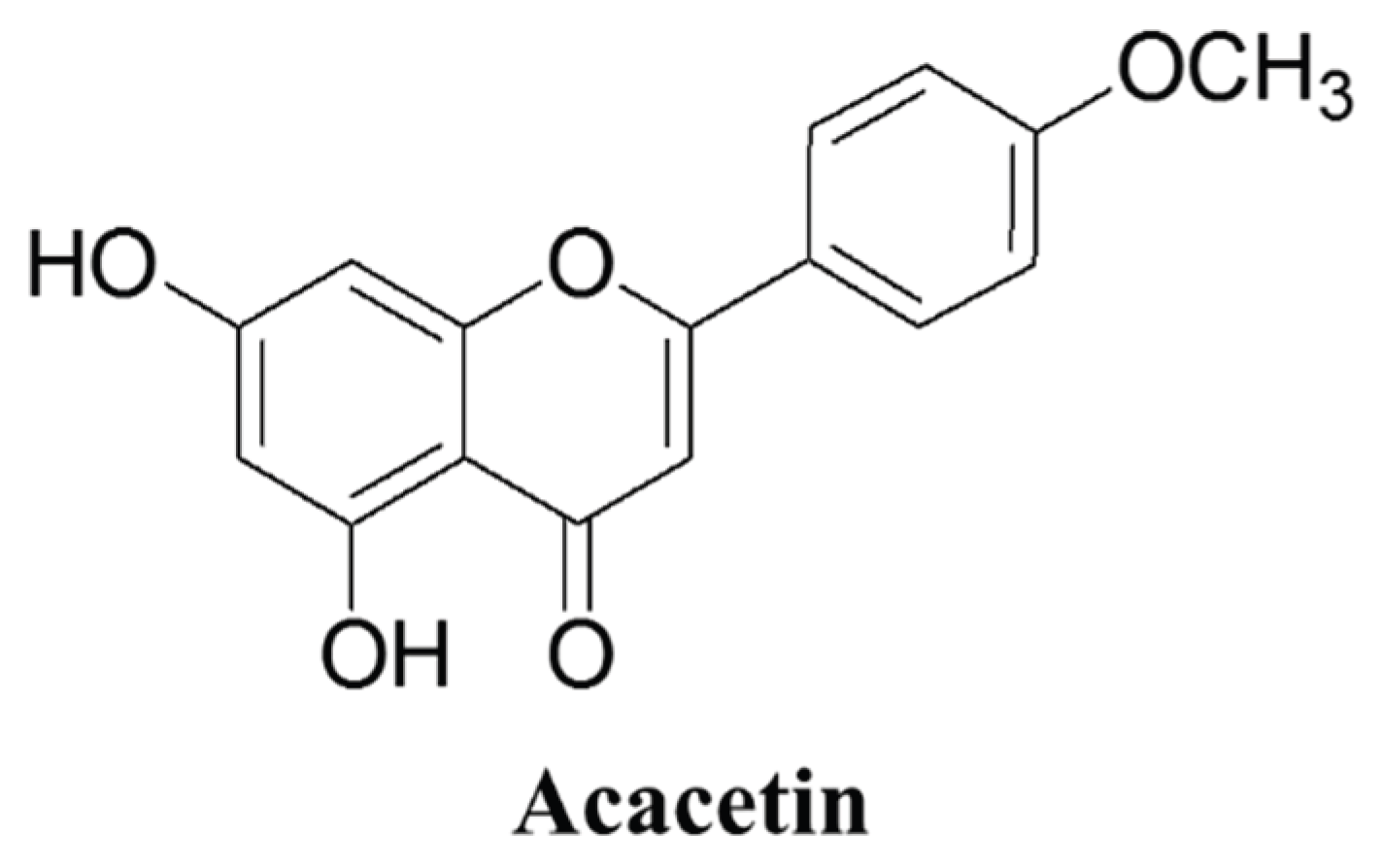
Figure 2.
Schematic overview of the multi-target anticancer mechanisms of acacetin based on network pharmacology, molecular docking, and experimental validation. Acacetin exerts anticancer activity through an integrated approach combining network pharmacology and molecular docking (in silico) with in vitro and in vivo validation. These analyses identified multiple oncogenic protein targets, including EGFR, STAT3, AKT, PI3K, and PTGS2 (validated by docking, in vitro, and in vivo studies), as well as SRC, HSP90AA1, TNF, IL-6, and MYC (supported by docking only), which collectively regulate critical signaling pathways. Functionally, acacetin induces apoptosis, promotes cell cycle arrest, and exerts anti-metastatic and anti-angiogenic effects, thereby underscoring its potential as a multi-target anticancer therapeutic. Figure created with BioRender.com. AKT, serine/threonine kinase (also known as protein kinase B, PKB); EGFR, epidermal growth factor receptor; HSP90AA1, heat shock protein 90 alpha family class A member 1; IL-6, interleukin-6; MYC, myelocytomatosis oncogene; PI3K, phosphoinositide 3-kinase; PTGS2, prostaglandin-endoperoxide synthase 2 (COX-2); SRC, proto-oncogene tyrosine-protein kinase Src; STAT3, signal transducer and activator of transcription 3; TNF, tumor necrosis factor.
Figure 2.
Schematic overview of the multi-target anticancer mechanisms of acacetin based on network pharmacology, molecular docking, and experimental validation. Acacetin exerts anticancer activity through an integrated approach combining network pharmacology and molecular docking (in silico) with in vitro and in vivo validation. These analyses identified multiple oncogenic protein targets, including EGFR, STAT3, AKT, PI3K, and PTGS2 (validated by docking, in vitro, and in vivo studies), as well as SRC, HSP90AA1, TNF, IL-6, and MYC (supported by docking only), which collectively regulate critical signaling pathways. Functionally, acacetin induces apoptosis, promotes cell cycle arrest, and exerts anti-metastatic and anti-angiogenic effects, thereby underscoring its potential as a multi-target anticancer therapeutic. Figure created with BioRender.com. AKT, serine/threonine kinase (also known as protein kinase B, PKB); EGFR, epidermal growth factor receptor; HSP90AA1, heat shock protein 90 alpha family class A member 1; IL-6, interleukin-6; MYC, myelocytomatosis oncogene; PI3K, phosphoinositide 3-kinase; PTGS2, prostaglandin-endoperoxide synthase 2 (COX-2); SRC, proto-oncogene tyrosine-protein kinase Src; STAT3, signal transducer and activator of transcription 3; TNF, tumor necrosis factor.
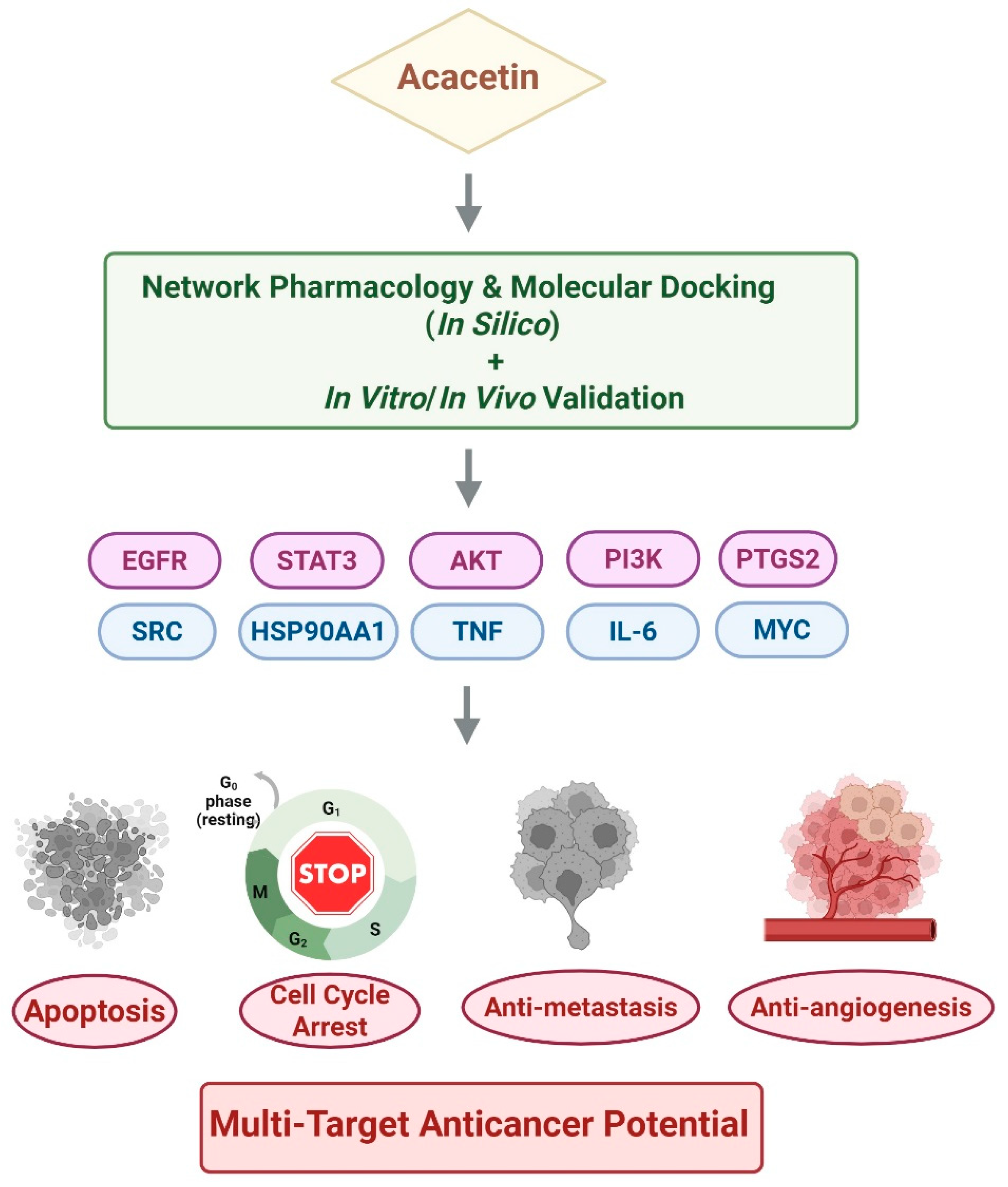
Table 2.
Confirmed and predicted molecular targets of acacetin based on docking and network pharmacology.
Table 2.
Confirmed and predicted molecular targets of acacetin based on docking and network pharmacology.
| Target | Cancer Type(s) | Function | Docking / Prediction / Validation Method | Validation Level | Functional Outcome | Refs. |
|---|---|---|---|---|---|---|
| EGFR | Gastric | EGFR-mediated signaling (STAT3, ERK) | Target prediction (SwissTargetPrediction, GeneCards, RNA-seq); network analysis (STRING, Cytoscape); docking (Schrödinger Maestro); validation (DARTS, CETSA, WB, TUNEL, colony formation, xenograft) | Confirmed (docking + in vitro + in vivo) | ↓: p-EGFR, p-STAT3, p-ERK (transient), Ki-67, PCNA, Bcl-xL, Colony formation, Tumor growth | [12] |
| ↑: Bax, cleaved caspase-3, cleaved PARP, Apoptosis, | ||||||
| Strong EGFR binding (docking) | ||||||
| Colorectal | - | Target prediction (SwissTargetPrediction, TCMSP, GeneCards, OMIM, DisGeNET); network analysis (STRING, Cytoscape); docking (AutoDock Vina) | Docking only | Binding affinity -8.3 kcal/mol | [62] | |
| Bladder | EGFR-mediated signaling | Target prediction (TCMSP, GeneCards, OMIM); network analysis (STRING, Cytoscape); docking (AutoDock Vina) | Docking only | Binding affinity -4.0 kcal/mol | [63] | |
| STAT3 | Prostate | STAT3-mediated signaling (proliferation, survival) | Docking (Glide, Schrödinger Maestro); binding validation (DARTS, CETSA, pull-down); functional assays (WB, Annexin V/PI, xenograft) | Confirmed (docking + in vitro + in vivo) | ↓: p-STAT3 (Tyr705), Cyclin D1, Bcl-2, Bcl-xL, Mcl-1, Survivin, Tumor volume, | [56] |
| ↑: Bax, cleaved PARP, cleaved caspase-3, Annexin V, p-JAK2, p-p38, ROS, Apoptosis | ||||||
| Strong STAT3 SH2 binding (3 H-bonds + cation–π; Glide) | ||||||
| AKT1 |
Colorectal | PI3K/AKT/p53 signaling | Target prediction (SwissTargetPrediction, TCMSP, GeneCards, OMIM, DisGeNET); network analysis (STRING, Cytoscape); docking (AutoDock Vina); validation (WB, CCK-8, Annexin V-FITC, scratch assay) | Confirmed (docking + multiple in vitro assays) | ↓: p-AKT, p-PI3K, Survivin, Migration, Proliferation | [62] |
| ↑: p53, cleaved caspase-3, Apoptosis, | ||||||
| Strong AKT1 binding (-9.2 kcal/mol; 1 H-bond, Thr211) | ||||||
| HSP90AB1 → AKT | Lung | Chaperone-mediated PI3K/AKT/mTOR activation | Network pharmacology; docking (AutoDock Tools); validation (WB, EMT, invasion assay ± terazosin) | Confirmed (docking + in vitro + in vivo) | ↓: p-AKT, p-mTOR, Migration, Invasion, EMT markers | [64] |
| ↑: E-Cadherin; effect reversed by HSP90 agonist terazosin | ||||||
| PI3Kγ | Breast | PI3K/AKT/mTOR/p70S6K/ULK signaling | Docking (SYBYL2.0); PI3Kγ kinase assay; validation (WB [PI3Kγ, p-AKT, p-mTOR, p-p70S6K, p-ULK1], Apoptosis assay, G2/M arrest, LC3 puncta) | Confirmed (docking + in vitro) | ↓: PI3Kγ, p-AKT, p-mTOR, p-p70S6K, p-ULK1 | [65] |
| ↑: Apoptosis, Autophagy, G2/M arrest | ||||||
| PI3Kγ ATP-site binding (H-bonds: Ser806, Ala885, Val882; hydrophobic: Lys833, Asp964) | ||||||
| SRC | Colorectal | Predicted PI3K/AKT signaling; proliferation/migration in CRC | Target prediction (SwissTargetPrediction, TCMSP, GeneCards, OMIM, DisGeNET); network analysis (STRING, Cytoscape); docking (AutoDock Vina) | Docking only | Binding affinity -6.9 kcal/mol | [62] |
| HSP90AA1 | Colorectal | Predicted chaperone-mediated PI3K/AKT; cell survival in CRC | Target prediction (SwissTargetPrediction, TCMSP, GeneCards, OMIM, DisGeNET); network analysis (STRING, Cytoscape); docking (AutoDock Vina) | Docking only | Binding affinity -7.1 kcal/mol | [62] |
| TNF | Colorectal | Predicted inflammatory/apoptotic signaling; CRC progression | Target prediction (SwissTargetPrediction, TCMSP, GeneCards, OMIM, DisGeNET); network analysis (STRING, Cytoscape); docking (AutoDock Vina) | Docking only | Binding affinity -6.4 kcal/mol | [62] |
| IL-6 | Bladder | Inflammatory cytokine signaling | Target prediction (TCMSP, GeneCards, OMIM); network analysis (STRING, Cytoscape); docking (AutoDock Vina) | Docking only | Binding affinity -4.12 kcal/mol | [63] |
| MYC | Bladder | Cell proliferation and transcription | Target prediction (TCMSP, GeneCards, OMIM); network analysis (STRING, Cytoscape); docking (AutoDock Vina) | Docking only | Binding affinity -4.37 kcal/mol | [63] |
| PTGS2 | Nasopharyngeal carcinoma | Inflammation, Tumor proliferation and migration | Target prediction (TCMSP, SwissTargetPrediction, SEA, GeneCards, OMIM); network analysis (STRING, Cytoscape); docking (AutoDock Vina), MD simulation; validation (CCK-8, colony formation, migration, WB [PTGS2]) | Confirmed (docking + in vitro) | ↓: PTGS2 (docking: < -7 kcal/mol; Tyr385 contact), Proliferation, Migration | [66] |
| PI3K (p110 subunit) | Skin | PI3K-AKT-p70S6K signaling | Binding assays (pull-down, kinase); cell transformation (in vitro); docking (Glide/induced-fit, Maestro); xenograft | Confirmed (docking + in vitro + in vivo) | ↓: PI3K activity, p-AKT, p-p70S6K, Tumor growth, Colony formation, | [58] |
| ↑: G1 arrest; docking to PI3K-p110 ATP-binding site (H-bonds: Val828, Glu826, Asp911; hydrophobic: Trp760, Ile777, Tyr813) | ||||||
| STAT3, AKT1, MAPK1, HSP90AA1, HRAS, SRC, PIK3CA, PIK3R1, FYN, RHOA, |
Gastric | PI3K-AKT, MAPK, JAK/STAT3, and cytoskeleton regulation pathways | HPLC-Q-TOF–MS/MS identification; network pharmacology (STRING, Cytoscape); docking (AutoDock Vina) | Docking only | High binding affinity (mostly < -7.3 kcal/mol) |
[67] |
AKT1, RAC-alpha serine/threonine-protein kinase (protein kinase B); Bax, Bcl-2-associated X protein; Bcl-2, B-cell lymphoma 2; Bcl-xL, B-cell lymphoma-extra large; CCK-8, cell counting kit-8; CETSA, cellular thermal shift assay; CRC, colorectal cancer; Cytoscape, Cytoscape network visualization software; DARTS, drug affinity responsive target stability; EGFR, epidermal growth factor receptor; EMT, epithelial–mesenchymal transition; HSP90AA1, heat shock protein 90 alpha family class A member 1; JAK2, Janus kinase 2; MAPK, mitogen-activated protein kinase; Mcl-1, myeloid cell leukemia-1; MD, molecular dynamics; OMIM, Online Mendelian Inheritance in Man; PARP, poly (ADP-ribose) polymerase; PCNA, proliferating cell nuclear antigen; PI3K, phosphoinositide 3-kinase; PTGS2, prostaglandin-endoperoxide synthase 2 (cyclooxygenase-2); Pull-down assay, biotinylated compound-based target validation assay; RNA-seq, RNA sequencing; ROS, reactive oxygen species; SEA, similarity ensemble approach; Schrödinger Maestro, Schrödinger molecular modeling suite; SRC, proto-oncogene, non-receptor tyrosine kinase Src; STAT3, signal transducer and activator of transcription 3; STRING, Search Tool for the Retrieval of Interacting Genes/Proteins; SwissTargetPrediction, SwissTargetPrediction in silico target prediction tool; TCMSP, Traditional Chinese Medicine Systems Pharmacology database; TNF, tumor necrosis factor; TUNEL, terminal deoxynucleotidyl transferase dUTP nick end labeling; WB, western blot. ↑, increase; ↓, decrease.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



