Submitted:
02 September 2025
Posted:
05 September 2025
You are already at the latest version
Abstract
Motor control arises from dynamic interactions among distributed brain networks that integrate cognitive, sensory, and autonomic processes. Among these, the Action-Mode Network (AMN; formerly the cingulo-opercular network) and the Somato-Cognitive Action Network (SCAN) are central for initiating and sustaining goal-directed behavior. The AMN orchestrates global action readiness by coordinating arousal, attention, and motor planning, whereas the SCAN executes whole-body motor output in concert with physiological adjustments. In Functional Movement Disorders (FMD), impaired AMN–SCAN coupling may disrupt the automatic integration of intention and movement, resulting in abnormal motor output and loss of agency. To address this, we propose a neurorehabilitation framework that combines action observation, motor imagery, and motor execution in conjunction with ultrasound neuromodulation of the AMN–SCAN interface at the dorsal anterior cingulate cortex (dACC)–supplementary motor complex (SMC). Neuroimaging-guided neuromodulation of the dACC–SMC integrated with biofeedback training has the potential to restore flexible network transitions between default-mode and action-mode states, normalize efferent signaling, and reduce oscillatory motor errors. This network-level approach reframes FMD as a disorder of brain–body state transitions and identifies a testable target for neuromodulation.
Keywords:
action-mode network
; somato-cognitive action network
; functional movement disorders
; neuromodulation
1. Introduction
The ability to initiate and control movement is fundamental to human behaviour, arising from a hierarchy of brain processes that span intention, planning, execution, and sensory feedback. Classic models of motor control have long emphasized segregated pathways, e.g., cortical motor areas driving spinal effectors, or cerebellar loops adjusting motor commands. However, accumulating evidence suggests that motor control is orchestrated by distributed brain networks that integrate cognitive, sensory, and autonomic components of action [1]. In particular, this new research highlights the importance of brain states and arousal in modulating these networks [2]. Here, it is postulated that the brain transitions between internal, self-referential modes and external, action-oriented modes depending on context. During active, goal-directed behaviour, the brain shifts into an “action-mode” characterized by heightened arousal and outward-focused attention, in contrast to the inward-focused, lower-arousal default-mode during rest. This shift in state, much like establishing a new initial condition in a dynamical system [3], is observed across species and is thought to reflect an evolutionary trade-off between exploiting the environment (action mode) and engaging in experience replay (default mode) [2]. An extreme shift in state to hyperarousal can transition the organism into a freeze state [4] when overwhelming, inescapable threat triggers a parasympathetic shutdown via the dorsal vagal complex as hypothesized in the polyvagal theory [5]. Neuroimaging shows that freezing engages amygdala–periaqueductal gray (PAG) circuits, with ventral PAG mediating heart-rate deceleration and shifts to active defence involving the perigenual anterior cingulate and frontal control regions [4].
In functional neurological disorder, reduced amygdala habituation, heightened sensitivity to negative emotions, and weakened prefrontal regulation of amygdala–PAG activity may underlie increased emotional reactivity and protective responses [6]. Recent advances in human neuroimaging have delineated altered brain networks [7]. The default-mode network (DMN) is active during rest and self-referential thought, and shows deactivation during many goal-oriented tasks. Opposite to the DMN is an Action-Mode Network (AMN), a large-scale network that becomes engaged during active behaviour and maintains the brain in an action-ready state [2]. The AMN, anatomically aligned with the former cingulo-opercular network, overlaps extensively with the “salience” network that comprises the dorsal anterior cingulate cortex (dACC), anterior insula, dorsal amygdala, PAG, and hypothalamus. The AMN comprises a set of interconnected cortical and subcortical nodes, including the dACC and adjacent cingulate motor zones, the anterior insula (frontal operculum), anterior middle frontal gyri near the frontal eye fields, and the supplementary motor area (SMA) along the dorsal midline. Subcortical components include the anterior putamen (receiving inputs from premotor cortex), portions of the central thalamus such as the intralaminar centromedian nucleus, and the cerebellar vermis and lateral cerebellar cortex across anterior and posterior lobes. This distributed anatomy positions the AMN to influence both cortical and subcortical motor systems. Notably, these AMN regions have been found to carry a remarkably diverse array of signals ranging from cognitive control and action initiation to autonomic arousal, error feedback and pain reflecting their broad role in enabling goal-directed behaviour. Another network, the somato-cognitive action network (SCAN), has been identified within the motor cortical system [1] that consists of nodes interleaved with the classical motor homunculus in primary motor cortex. In between the discrete areas for foot, hand, and mouth movements, there are inter-effector regions that are strongly interconnected with each other and with higher motor control areas. These interdigitated SCAN nodes lack specificity for a single effector i.e., they are co-activated by movements of different body parts and instead appear to coordinate whole-body actions and physiological adjustments. The SCAN’s cortical nodes are functionally coupled with midline structures like the SMA and caudal dACC, with the centromedian (CM) intralaminar thalamus, dorsal striatum (posterior putamen), and specific cerebellar lobules (paravermal Crus VI/ VIIIa). These subcortical connections place the SCAN at the interface of traditional motor control loops of the cortico–basal ganglia–thalamo circuit and the cortico–cerebello–thalamo–cortical circuit. Indeed, deep brain stimulation targets for movement disorders, such as the subthalamic nucleus (STN), globus pallidus internus (GPi), and ventral intermediate nucleus of the thalamus (VIM), have been found to lie within the SCAN connectivity map. Parkinson’s disease, for example, involves abnormalities in these nodes and their cortical connections, suggesting that the SCAN underpins a broad, multi-effector motor circuit implicated in movement disorders [8].
In this opinion piece, we synthesize all current knowledge and describe how brain networks interact to support motor execution, motor imagery, and action observation functions that can be harnessed for portable brain imaging–guided neuromodulation during action-simulation. We highlight the pivotal role of arousal systems that can be neuromodulated [9] to facilitate initial switching from a high-salience, high-motor readiness resting-state and enabling transition to a stable motor task-set. Our rehabilitation approach is framed within the principles of predictive coding hypothesis whereby the brain continually generates and updates predictions about the sensory consequences of actions, and we consider how these brain networks might implement or modulate such predictions [3]. We propose that targeting the action-mode network during action simulation under reward gameplay (operant conditioning [10]) may restore patients’ capacity to initiate and control movement in a volitional, adaptive manner.
2. Distinctive Connectivity Patterns Underlying Functional Movement Disorders
Functional movement disorders (FMD), a subtype of functional neurological disorders, exemplify a condition where patients experience involuntary or exaggerated movements without overt structural lesions [12,13]. Waugh and colleagues [11] applied a support vector machine classifier to resting-state fMRI data from 61 FMD patients and 59 controls, identifying a restricted set of 29 region-to-region connectivity features that classified patients with 80% accuracy. Seed regions were derived from prior FMD/FND studies (66 ROIs across motor, limbic, and executive networks – see Supplementary Table S1), and recursive feature elimination highlighted connections particularly involving the right sensorimotor cortex, left dorsolateral prefrontal cortex, cerebellum, and posterior insula as shown in Figure 1. Supplementary analyses (see Supplementary Table S2) detailed the specific ROI pairs most predictive of FMD status, including strong connectivity between the right precentral gyrus and left posterior insula, right SMA and left cerebellum, and SMA–amygdala links. The selected features revealed three functional clusters, motor, limbic, and executive where limbic regions often mediated interactions between motor and executive systems – see the thick grey edges showing strong connectivity in Figure 1. Together, these findings support the hypothesis that FMD arises from disrupted integration of emotion, interoception, and motor control, producing a brain connectivity signature. Indeed, abnormal communication among motor planning areas, limbic regions, and agency-related regions (e.g., the right temporoparietal junction) has been reported in FMD [7]. Therefore, the pathophysiology of FMD is increasingly viewed as a network-level disturbance [14,15]. For instance, patients with motor FMD show altered resting-state dynamics where Weber and colleagues found that patients less frequently enter a brain state characterized by co-activation of the salience network (SN) with the DMN (with deactivation of executive and motor networks), and more frequently dwell in a state of simultaneous somatomotor and salience network hyperactivation with DMN suppression [7]. These findings suggest FMD patients may be “stuck” in a high-salience, high-motor readiness resting-state without the flexibility to switch to the executive and motor networks [3]. Dosenbach’s latest hypothesis portrays the SN, AMN, and SCAN as distinct yet functionally interconnected components where the salience network handles reward and motivational processing which transitions into goal-directed and action-control functions via the AMN and culminates in coordinated whole-body movement through the SCAN [16]. Here, the SN detects and initiates shifts in arousal, allowing response to salient internal and external demands; the AMN then works to stabilize arousal in support of sustained, goal-directed action, while the SCAN translates these regulated states into coordinated bodily movement [3,7] – see Supplementary Table S1 and S2 for ROIs overlap with SN, AMN, and SCAN. Therefore, effective motor task maintenance depends on ongoing interactions between the AMN and SCAN i.e., if the AMN fails to appropriately stabilize arousal and guide transitions between action states, or if the SCAN’s integration of intention and motor execution is disrupted, the result can be a breakdown in the sense of agency and the emergence of uncontrolled movements, as seen in FMD. Understanding how the SN, AMN, and SCAN interact is crucial for neuroengineering solutions to movement dysfunction in FMD [3,14].
3. Meta-Analysis By Linking Neurosynth Studies To Subnetworks To Guide Ultrasond Neuromodulation
The ROIs in the functional clusters (see Supplementary Table S2) were used for meta-analysis by linking Neurosynth studies and then testing which cognitive terms are differentially associated with those subnetworks [17]. The Matlab (Mathworks Inc., USA) script (see Supplementary Script S3) loads term weights per paper from Neurosynth studies, each paper’s MNI peak coordinates, and ROI node coordinates for each subnetwork. A paper is considered to “hit” a subnetwork if at least a set fraction of its ROI nodes (coverage_thresh, e.g., 75% used) lie within a distance threshold (distance_thresh, e.g., 5 mm used) of the paper’s peaks. Using only the papers that match each subnetwork, it performs a one-way ANOVA for every term to compare term weights across subnetworks, then applies Benjamini–Hochberg FDR (q=0.05) to identify significant terms. Topic terms derived using Latent Dirichlet Allocation [18] applied to the Neurosynth text corpus of neuroimaging abstracts [17], where each topic_xxx reflects a distribution of semantically related words capturing broader cognitive or functional domains – see Figure 2.
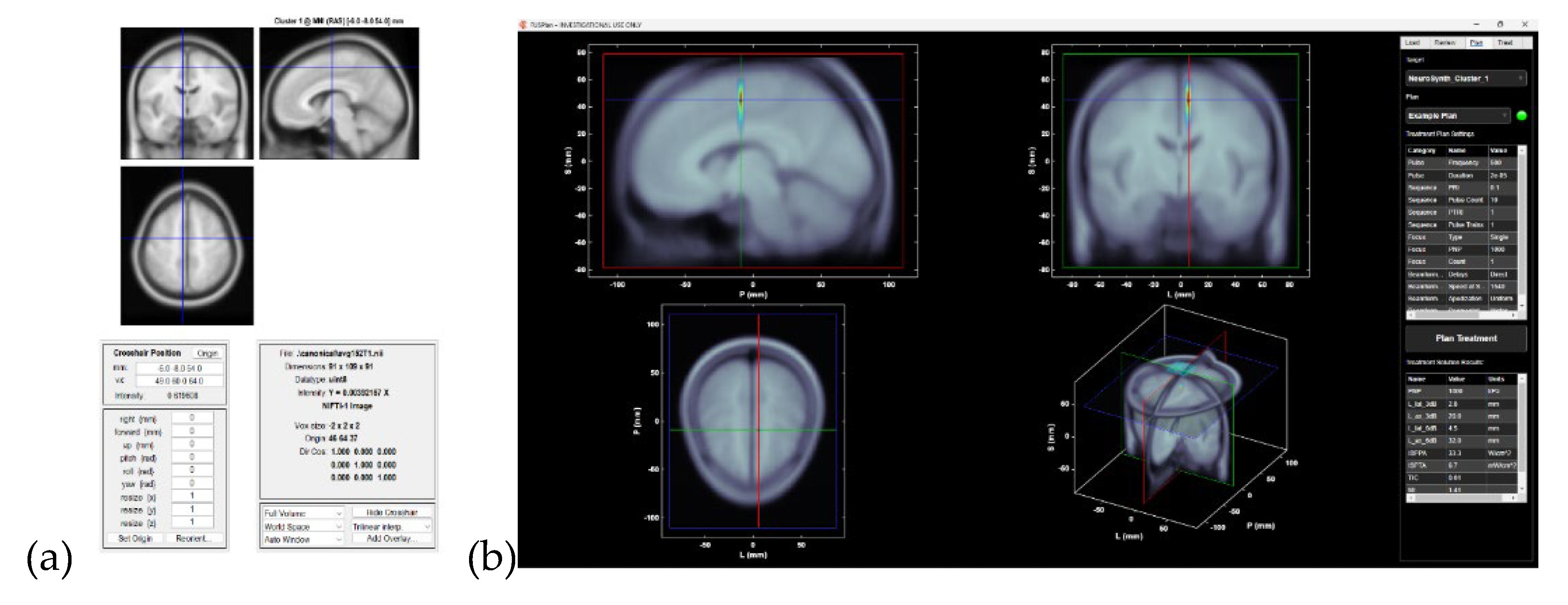
We identified consensus activation regions across several thresholded Neurosynth maps [17] from Latent Dirichlet Allocation [18]. The Matlab (Mathworks Inc., USA) script (see Supplementary Script S4) first loaded multiple Z maps on a common grid, binarized them at a chosen threshold (thr=7 used to get a single consensus activation Cluster 1). Then, it created logical-AND mask of voxels significant in all maps which it clustered and reported with MNI peak coordinates and atlas labels. Then, it computed a voxel wise overlap count and performed a binomial k-of-N test (probability of observing ≥K hits given per-map alpha=0.01 used) and applied Benjamini–Hochberg FDR across voxels to obtain a significant binomial mask and a q-value map. The binomial-FDR masks were clusterised and cluster mask results were written out as NIfTI images (see Supplementary Image S5) with a cluster table (see Supplementary Table S6) listing cluster size, MNI peak, and atlas ID/name (from AAL3v1 https://www.oxcns.org/aal3.html). Then, a separate Matlab (Mathworks Inc., USA) script (see Supplementary Script S7) converted the cluster peak from the cluster table into usable coordinates to be used with Open-LIFU 2.0 (OpenWater, USA) wearable, steerable, transcranial low intensity focused ultrasound system [19]. The Matlab (Mathworks Inc., USA) script loaded the cluster table, extracted the voxel indices of a chosen cluster (Cluster 1), and converted them into MNI (RAS) millimeter coordinates using a 1 mm MNI152 template header. These RAS coordinates are then transformed into LPS millimeters, which are the convention used by focused ultrasound (FUS) targeting software. We visualized the location in SPM’s orthviews [20], writing out a small spherical ROI mask around the peak as a NIfTI, and creating a fus.Point target object for use in the openLIFU workflow (https://github.com/OpenwaterHealth/OpenLIFU-app) [19]. In short, the Matlab script bridged Neurosynth cluster output with SPM visualization [20], ROI generation, and FUS targeting – see Figure 3.
4. Action-Mode Network In Relation To SCAN For Neuromodulation
The SCAN is tightly interconnected with the AMN to the point that one might view the SCAN as an effector of the AMN within the sensorimotor cortex. The SCAN’s cortical nodes are three bilateral regions embedded in M1 that “alternate” with the classic motor homunculus regions for foot, hand, and mouth [21]. These nodes are strongly interconnected with each other across hemispheres (forming a bilateral integrative network distinct from unilateral effector areas). They also show strong connectivity with nodes of the AMN especially along the dorsal midline [2]. In fact, the strongest SCAN–AMN connections link the SCAN’s M1 nodes with the SMA and caudal dACC which themselves are part of the AMN [2]. This effectively connects the SCAN and AMN into a contiguous system spanning from higher-order frontal control regions to primary motor areas. Additionally, the SCAN connects to subcortical hubs that encompasses the centromedian (CM) thalamus and ventral intermediate (VIM) thalamus, dorsolateral striatum (putamen), and cerebellar regions (paravermal lobules) that are adjacent to (but distinct from) the cerebellar representations of specific effectors [21]. These connections endow the SCAN with the ability to influence spinal and brainstem motor outputs both via cortico-spinal pathways (through M1 and SMA outputs) and subcortical loops (basal ganglia and cerebellum). Notably, animal studies show that stimulating cortex in SCAN regions can evoke complex, multi-joint movements and even affect autonomic functions [21]. For example, stimulation in anterior M1 (likely a SCAN zone) in monkeys can produce coordinated postural or whole-body adjustments rather than isolated finger twitches, and this same region has connectivity to autonomic targets like the adrenal medulla, stomach, kidneys, and heart [22]. These findings subserve the SCAN’s role in integrating visceromotor (internal organ control) with somatomotor (skeletal muscle control) as part of allostasis during action [21]. For example, consider the act of standing up quickly where the SCAN/AMN system would anticipate the need for blood pressure regulation (via sympathetic activation) i.e., a whole-body adjustment concurrent with the movement. Here, a pure motor circuit might execute the muscle activations, but without anticipatory adjustments leading to orthostatic dizziness [23]. Given their structural connections, the AMN and SCAN can be thought of as two levels of an integrated action control hierarchy. The AMN sets the goals, context, and necessary initial conditions (in terms of attention, arousal, and readiness), and the SCAN translates those into concrete motor outputs with coordinated muscle group activations, postural sets, and even hormone releases (like adrenaline surge) to support the action. Therefore, the AMN operates at the level of global brain state and high-level action control (“initiation transient”), whereas the SCAN implements the detailed execution of actions in concert with bodily adjustments (stable task-set maintenance) – “the SCAN functions as the AMN’s actuator, implementing goal-directed actions via coordinated skeletal and smooth muscle movement and hormone release” [24]. If the AMN determines that the appropriate response is to “flee from danger,” the SCAN would orchestrate coordinated whole-body movements for escape while simultaneously activating sympathetic responses such as increased heart rate and enhanced blood flow to the limbs. Conversely, if the AMN decides to “freeze in danger,” the SCAN would halt whole-body movement and, in parallel, trigger parasympathetic shutdown via the dorsal vagal complex. In this integrative framework, the caudal dACC–to–SMA complex part of the AMN likely relay efferent signalling [25] and is proposed target for neuromodulation. Dysfunctional efferent signalling could explain many functional symptoms [23] viz. normal voluntary movement might be impeded by an inability to generate the required preparatory state such that the body is, in a sense, unprepared or in the wrong state for the action leading to a disconnect between intention and execution [3].
A key function emerging for the AMN–SCAN ensemble is in initiating self-generated actions. Evidence from neurology and neurostimulation studies indicates that damage to or disruption of the AMN/SCAN network impairs internally driven, voluntary behaviour while sparing externally triggered movements [21]. Patients with lesions in dACC, anterior insula, or medial frontal areas (all part of AMN/SCAN connectivity) can exhibit abulia or akinetic mutism where they do not initiate actions or speech on their own yet may still react normally to external commands or sensory cues. Similarly, in monkeys, lesions of anterior (higher-order) motor areas result in loss of spontaneous behavioural initiation but preserved execution of movements in response to stimuli, whereas lesions of posterior M1 produce the opposite (no challenge with initiation, but problems executing precise movements). These double dissociations confirm that the brain has a dedicated system for volitional drive located on the medial and anterior network (AMN/SCAN) that is separate from the implementation system for precise motor output.
5. Discussion
In the context of predictive coding and agency [3], the AMN/SCAN can be seen as generating the “prior” or intention for movement which is then delivered to the effector apparatus. If that prior is not properly generated or conveyed, the sense of agency (the feeling “I willed this”) can be disrupted. It is notable that electrical stimulation of the SCAN or connected regions can produce unusual experiential effects with patients having reported an urge to move without actual movement [26], a sense of having moved when they did not, or conversely performing a movement without sensing that they did it. Phenomena elicited by stimulating dACC or anterior insula regions are essentially mismatches between intention and action, or between action and awareness, and mirror the disturbances in sense of agency seen in FMD [27]. If caudal dACC–to–SMA complex part of the AMN is perturbed, one can expect a predictive coding error either an volitional action is executed but not registered as self-initiated (viz. functional seizure) or a prediction of movement occurs without volitional execution (viz. functional stroke). Here, we approach rehabilitation as a network coordination dysfunction where the solution then is to reteach the networks to communicate and transition properly. The AMN must be re-enabled to properly turn on the action-mode (neither under- nor over-shooting arousal [28]), the SCAN must be recalibrated to link intention with execution (without aberrant efferent signalling [25]), and the cerebellar loops must be harnessed for autonomous error-correcting function (rather than contributing to oscillatory errors [29]). Combining physical therapy with targeted neuromodulation and biofeedback training could synergistically achieve this in a way not possible with isolated approaches. Moreover, Dosenbach’s reframing the cingulo-opercular network as the AMN reflects a shift from anatomy-based labels to function-based understanding [2]. This perspective unifies its diverse roles from conflict monitoring, pain, arousal under the theme of action control. Here, the AMN coordinates cognition and attention for action while the SCAN integrates motor plans with whole-body adaptation. In FMD neurorehabilitation, neuromodulation approach would involve modulating the salience network via vagus nerve stimulation [9,30] while using ultrasound neuromodulation to engage the AMN/SCAN interface at the caudal dACC–SMA complex during biofeedback training with suggestion and imagery [10]. Clinically, recognizing the action-mode vs default-mode distinction can guide practice such as keeping patients goal-focused during movement training or down-regulating the AMN when unwanted movements emerge. Integrating the AMN into motor control frameworks suggests that movement disorders without structural lesions often reflect network dysregulation that may be amenable to suggestion and imagery [10]. Future closed-loop stimulation guided by real-time wearable neuroimaging could restore the balance between the internal self (default-mode) and acting self (action-mode), reclaiming the sense of agency and improving interaction with the external world.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Pushpinder Walia: Methodology, Investigation. Abhijit Das: Validation, Writing – Review & Editing. Anirban Dutta: Conceptualization, Methodology, Investigation, Visualization, Resources, Writing – Original Draft, Writing – Review & Editing.
Competing interests: The author declares no competing interests.
References
- E. M. Gordon et al., ‘A somato-cognitive action network alternates with effector regions in motor cortex’, Nature, vol. 617, no. 7960, pp. 351–359, May 2023. [CrossRef]
- N. U. F. Dosenbach, M. E. N. U. F. Dosenbach, M. E. Raichle, and E. M. Gordon, ‘The brain’s action-mode network’, Nat. Rev. Neurosci., vol. 26, no. 3, pp. 158–168, Mar. 2025. [CrossRef]
- Dutta, ‘Neurocomputational Mechanisms of Sense of Agency: Literature Review for Integrating Predictive Coding and Adaptive Control in Human–Machine Interfaces’, Brain Sciences, vol. 15, no. 4, p. 396, Apr. 2025. [CrossRef]
- K. Roelofs, ‘Freeze for action: neurobiological mechanisms in animal and human freezing’, Philosophical Transactions of the Royal Society B: Biological Sciences, vol. 372, no. 1718, p. 20160206, Feb. 2017. [CrossRef]
- S. W. Porges, ‘Orienting in a defensive world: mammalian modifications of our evolutionary heritage. A Polyvagal Theory’, Psychophysiology, vol. 32, no. 4, pp. 301–318, Jul. 1995. [CrossRef]
- A. Milano, M. A. Milano, M. Moutoussis, and L. Convertino, ‘The neurobiology of functional neurological disorders characterised by impaired awareness’, Front. Psychiatry, vol. 14, Mar. 2023. [CrossRef]
- S. Weber, J. Bühler, T. A. W. Bolton, and S. Aybek, ‘Altered brain network dynamics in motor functional neurological disorders: the role of the right temporo-parietal junction’, Transl Psychiatry, vol. 15, no. 1, p. 167, May 2025. [CrossRef]
- J. Ren et al., ‘The somato-cognitive action network is the core neuromodulation target in Parkinson’s disease’, Brain Stimulation: Basic, Translational, and Clinical Research in Neuromodulation, vol. 18, no. 1, pp. 523–524, Jan. 2025. [CrossRef]
- Das and, A. Dutta, ‘Functional seizure therapy via transauricular vagus nerve stimulation’, Medical Hypotheses, vol. 191, p. 111462, Oct. 2024. [CrossRef]
- Dutta, *!!! REPLACE !!!*; et al. , ‘Extended Reality Biofeedback for Functional Upper Limb Weakness: Mixed Methods Usability Evaluation’, JMIR XR and Spatial Computing (JMXR), vol. 2, no. 1, p. e68580, Jun. 2025. [CrossRef]
- R. E. Waugh, J. A. R. E. Waugh, J. A. Parker, M. Hallett, and S. G. Horovitz, ‘Classification of Functional Movement Disorders with Resting-State Functional Magnetic Resonance Imaging’, Brain Connect, vol. 13, no. 1, pp. 4–14, Feb. 2023. [CrossRef]
- J. Stone, A. J. Stone, A. Zeman, and M. Sharpe, ‘Functional weakness and sensory disturbance’, Journal of Neurology, Neurosurgery & Psychiatry, vol. 73, no. 3, pp. 241–245, Sep. 2002. [CrossRef]
- V. Voon et al., ‘Emotional stimuli and motor conversion disorder’, Brain, vol. 133, no. 5, pp. 1526–1536, May 2010. [CrossRef]
- T. Grippe, N. T. Grippe, N. Desai, T. Arora, and R. Chen, ‘Use of non-invasive neurostimulation for rehabilitation in functional movement disorders’, Front Rehabil Sci, vol. 3, p. 1031272, Nov. 2022. [CrossRef]
- W. Maurer, K. W. Maurer, K. LaFaver, R. Ameli, S. A. Epstein, M. Hallett, and S. G. Horovitz, ‘Impaired self-agency in functional movement disorders’, Neurology, vol. 87, no. 6, pp. 564–570, Aug. 2016. [CrossRef]
- Badke, D’Andrea; et al. , ‘Action-mode subnetworks for decision-making, action control, and feedback’, Proceedings of the National Academy of Sciences, vol. 122, no. 27, p. e2502021122, Jul. 2025. [CrossRef]
- T. Yarkoni, R. A. T. Yarkoni, R. A. Poldrack, T. E. Nichols, D. C. Van Essen, and T. D. Wager, ‘Large-scale automated synthesis of human functional neuroimaging data’, Nat Methods, vol. 8, no. 8, pp. 665–670, Jun. 2011. [CrossRef]
- R. A. Poldrack, J. A. R. A. Poldrack, J. A. Mumford, T. Schonberg, D. Kalar, B. Barman, and T. Yarkoni, ‘Discovering Relations Between Mind, Brain, and Mental Disorders Using Topic Mapping’, PLOS Computational Biology, vol. 8, no. 10, p. e1002707, Oct. 2012. [CrossRef]
- R. Bawiec et al., ‘A Wearable, Steerable, Transcranial Low-Intensity Focused Ultrasound System’, J Ultrasound Med, vol. 44, no. 2, pp. 239–261, Feb. 2025. [CrossRef]
- ‘SPM - Statistical Parametric Mapping’. Accessed: Jun. 17, 2018. [Online]. Available: http://www.fil.ion.ucl.ac.
- M. Gordon et al., ‘A somato-cognitive action network alternates with effector regions in motor cortex’, Nature, vol. 617, no. 7960, pp. 351–359, May 2023. [CrossRef]
- R. P. Dum, D. J. R. P. Dum, D. J. Levinthal, and P. L. Strick, ‘Motor, cognitive, and affective areas of the cerebral cortex influence the adrenal medulla’, Proceedings of the National Academy of Sciences, vol. 113, no. 35, pp. 9922–9927, Aug. 2016. [CrossRef]
- S. Gilmour et al., ‘Management of functional neurological disorder’, J Neurol, vol. 267, no. 7, pp. 2164–2172, Jul. 2020. [CrossRef]
- N. Dosenbach, M. N. Dosenbach, M. Raichle, and E. Gordon, ‘The brain’s cingulo-opercular action-mode network’, Jan. 27, 2024, OSF. [CrossRef]
- Dutta, ‘”Hyperbinding” in functional movement disorders: role of supplementary motor area efferent signalling’, Brain Communications, vol. 7, no. 1, p. fcae464, Feb. 2025. [CrossRef]
- M. Desmurget and A. Sirigu, ‘Revealing humans’ sensorimotor functions with electrical cortical stimulation’, Philosophical Transactions of the Royal Society B: Biological Sciences, vol. 370, no. 1677, p. 20140207, Sep. 2015. [CrossRef]
- J. P. Ospina, R. J. P. Ospina, R. Jalilianhasanpour, and D. L. Perez, ‘The Role of the Anterior and Middle Cingulate Cortices in the Neurobiology of Functional Neurological Disorder’, Handb Clin Neurol, vol. 166, pp. 267–279, 2019. [CrossRef]
- V. Breton-Provencher, G. T. V. Breton-Provencher, G. T. Drummond, and M. Sur, ‘Locus Coeruleus Norepinephrine in Learned Behavior: Anatomical Modularity and Spatiotemporal Integration in Targets’, Front. Neural Circuits, vol. 15, Jun. 2021. [CrossRef]
- R. C. Miall, D. J. R. C. Miall, D. J. Weir, D. M. Wolpert, and J. F. Stein, ‘Is the cerebellum a smith predictor?’, J Mot Behav, vol. 25, no. 3, pp. 203–216, Sep. 1993. [CrossRef]
- A. Monti, N. A. Monti, N. Wintering, F. Vedaei, A. Steinmetz, F. B. Mohamed, and A. B. Newberg, ‘Changes in brain functional connectivity associated with transcutaneous auricular vagus nerve stimulation in healthy controls’, Front Hum Neurosci, vol. 19, p. 1531123, Mar. 2025. [CrossRef]
Figure 1.
Brain network of the 29 predictive rs-fMRI connections in Functional Movement Disorder. Nodes are plotted at their MNI coordinates (seed list from Supplementary Table S1); node size encodes weighted degree within the 29 SVM-selected connections (Supplementary Table S2). Node color denotes functional modules (Motor=Red, Limbic=Yellow, Executive=Green) as used in the Waugh and colleagues [11] clustering; edges depict the 29 predictive region-to-region links, weighted by the model’s feature scores. Prominent hubs (larger nodes) include right sensorimotor cortex (pre/postcentral and SMA), left posterior insula, left cerebellum, and dorsolateral prefrontal cortex, illustrating the limbic system’s mediation between motor and executive circuits.
Figure 1.
Brain network of the 29 predictive rs-fMRI connections in Functional Movement Disorder. Nodes are plotted at their MNI coordinates (seed list from Supplementary Table S1); node size encodes weighted degree within the 29 SVM-selected connections (Supplementary Table S2). Node color denotes functional modules (Motor=Red, Limbic=Yellow, Executive=Green) as used in the Waugh and colleagues [11] clustering; edges depict the 29 predictive region-to-region links, weighted by the model’s feature scores. Prominent hubs (larger nodes) include right sensorimotor cortex (pre/postcentral and SMA), left posterior insula, left cerebellum, and dorsolateral prefrontal cortex, illustrating the limbic system’s mediation between motor and executive circuits.
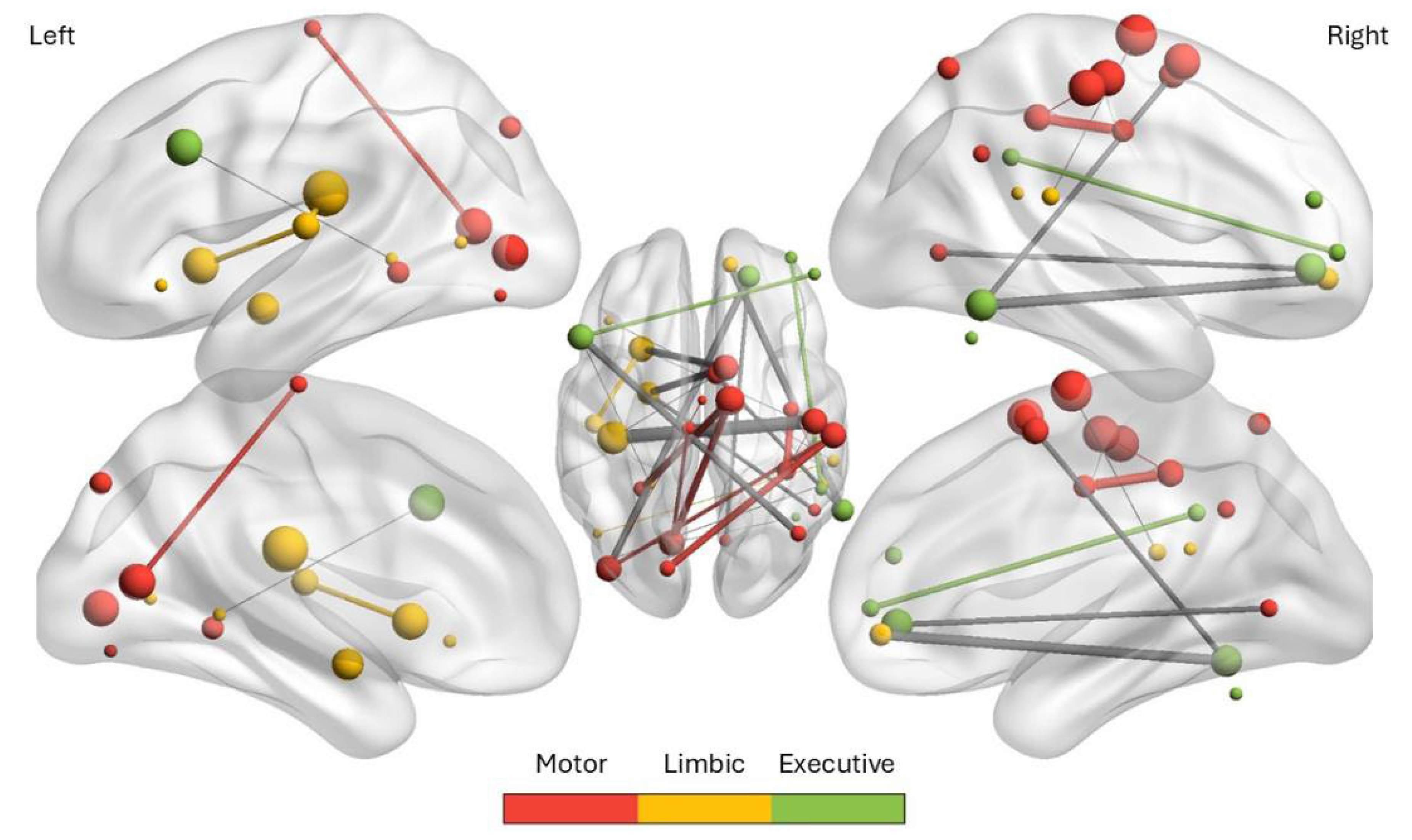
Figure 2.
Word cloud of significant Neurosynth topics identified by MANA analysis that shows keywords from topic terms that were significantly associated with one or more subnetworks after ANOVA and false discovery rate (FDR) correction (q < 0.05). Word size reflects the relative frequency of keywords across significant topics, highlighting supplementary, motor, control most consistently linked to the matched subnetworks.
Figure 2.
Word cloud of significant Neurosynth topics identified by MANA analysis that shows keywords from topic terms that were significantly associated with one or more subnetworks after ANOVA and false discovery rate (FDR) correction (q < 0.05). Word size reflects the relative frequency of keywords across significant topics, highlighting supplementary, motor, control most consistently linked to the matched subnetworks.
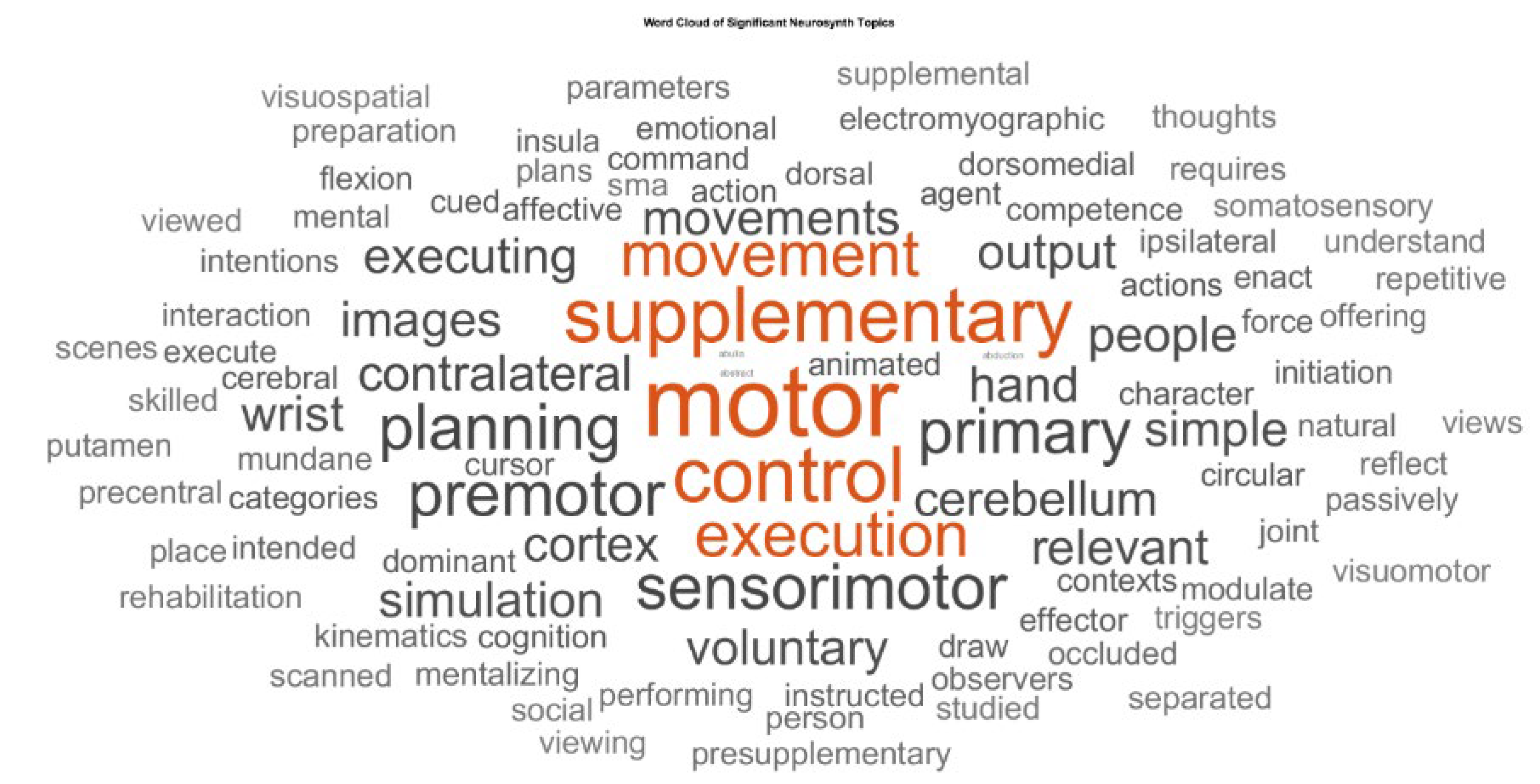
Figure 3.
(a) SPM Orthoview display showing cluster peak (left SMA) overlaid on the MNI template in coronal, sagittal, and axial slices. (b) Screenshot from the Openwater planning software. The left panel displays the anatomical MRI with the transducer, treatment target (cluster peak), and simulated acoustic pressure field overlaid. The right panel shows the Plan interface which provides treatment protocol details and analysis of the planned sonication solution.
Figure 3.
(a) SPM Orthoview display showing cluster peak (left SMA) overlaid on the MNI template in coronal, sagittal, and axial slices. (b) Screenshot from the Openwater planning software. The left panel displays the anatomical MRI with the transducer, treatment target (cluster peak), and simulated acoustic pressure field overlaid. The right panel shows the Plan interface which provides treatment protocol details and analysis of the planned sonication solution.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



