Submitted:
01 September 2025
Posted:
02 September 2025
You are already at the latest version
Abstract
Phosphorus is a key macronutrient central to the processes of energy and information storage and exchange in the cell. Single-celled photosynthetic organisms including microalgae accumulate intracellular reserves of phosphorus (mostly in form of polyphosphate) essential for maintenance of cell homeostasis during fluctuations of external phosphorus availability. The polyphosphate reserves in microalgal cells are formed by polyphosphate polymerases—a ubiquitous enzyme family represented mainly by prokaryotic (PPK-type, typical of prokaryotes e.g. cyanobacteria) and VTC-type polyphosphate polymerases harbored by eukaryotic microalgae, although certain species possess both PPK- and VTC-types of the enzyme. This enzyme is important for environmental fitness of microalgae dwelling in diverse habitats as well as for efficiency of microalgae-based systems for biocapture of phosphate from waste streams and for upcycling this valuable nutrient to agricultural ecosystems via biofertilizer from microalgal biomass. This review summarizes the recent progress in the field of structure, regulation, and functioning of VTC in microalgae. In conclusion, biotechnological implications, and perspectives of VTC as a target of microalgal cell engineering and bioprocess design for improved phosphate bioremoval efficiency and culture robustness are considered.
Keywords:
microalgae
; polyphosphate
; luxury uptake
; phosphorus bioremoval
1. Introduction
Phosphorus (P) is a key macronutrient central to the processes of energy and information storage and exchange in the cell [1,2,3]. The most bioavailable form of P is inorganic phosphate, Pi, which is directly involved in many metabolic reactions [4,5,6]. Both shortage and excess of Pi are deteriorative for normal functioning of the cell. Therefore, maintenance of Pi homeostasis is essential for the normal operation of biological systems including photosynthetic organisms [7,8,9]. This goal is accomplished through the concerted operation of Pi uptake via Pi transporter system and turnover of P reserves in the cells to match its metabolic demand vs. availability of P [10]. As balancing of supply and demand, stockpiles, and border controls constitute core concepts of economics, P uptake, storage, and turnover of its reserves in the cell can be collectively termed “P economy of the cell” which is implemented via sophisticated network of precisely orchestrated mechanisms [8,11]. The bulk of P reserves in single-celled photosynthetic organisms including eukaryotic microalgae and cyanobacteria are represented by inorganic polyphosphate (polyP)—a ubiquitously present, ancient, multifunctional molecule [1,2,12,13,14]. To date, the capability of synthesizing and accumulating polyP was confirmed in more than 100 microalgal species [15]. These species dwell in diverse habitats including terrestrial (soil), marine, and freshwater environments featuring a broad range of environmental conditions from arctic to hot acidic springs [15]. Such a broad representation indicates that this ability is not of limited significance e.g., for acclimation to a specific niche, but of fundamental importance.
Experimental evidence accumulated to the date supports the importance of the enzymatic machinery implementing turnover of polyP for the maintenance of cell P homeostasis (or, in frame of the analogy drawn above, for the maintenance of viable P economy of the cell) [1,16]. In addition to this, polyP fulfills a plethora of other functions in different organisms from bacteria to human. Impaired polyP biosynthesis is associated with impaired mitochondrial metabolism, failure of cell apoptosis, bone mineralization, it also promotes procoagulant and proinflammatory responses, and disturbs mTOR signaling [2,17,18,19,20,21,22]. As efficient ion chelator, polyP modulates intracellular monovalent and divalent cation availability, serves the function of molecular chaperone preventing protein aggregation under stress [22], binds to proteins and regulates their functions, activates ribosomal protein degradation in response to nitrogen starvation [8,23,24,25]. The latter mechanism is particularly important for the induction of valuable compound biosynthesis in microalgae by nitrogen starvation [26].
In microalgae, polyP is synthesized by polyphosphate polymerase. This enzyme family includes eukaryote-type polyP polymerase (commonly termed VTC since it was first described as Vacuolar Transport Chaperone proteins [27]) and prokaryote-type enzymes, PPK [28]. Certain eukaryotic microalgae harbor both types of enzymes (see below and [29]). Polyphosphate polymerases are conserved throughout microalgae and other major groups of living beings [27]. Although polyP were found in all cells studied so far [1,12], eukaryotic polyP polymerases, VTC, remained poorly studied due to their complexity, but recent breakthroughs deepened considerably our understanding of their structure and function [30,31].
The VTC enzyme and VTC-dependent biosynthesis of polyP gained increasing attention of the researchers during last decade (see [15,30,32,33] and references therein). It became clear that the functioning of VTC and its regulation is at the core of P flows and partitioning in individual cells, microbial consortia, and larger ecosystems [16]. The VTC-mediated biosynthesis of polyP is of primary importance for the efficiency of microalgae-based biotechnologies suggested as a promising solution for sustainable usage of P. Illustrious examples of those include biocapture of Pi from waste streams and wastewater treatment facilities with microalgal cultures [33,34,35,36] and re-routing it to agroecosystems via “green” biofertilizer made from the microalgal biomass [37].
Experimental evidence on functional significance and biotechnological importance of polyP polymerases in microalgae has been rapidly accumulated during last decade while the generalizations on these topics mostly lagged. In view of this, recently obtained experimental facts about polyP polymerases of microalgae are summarized below. Towards this end, handpicked reports were carefully selected, both experimental papers and recent reviews reflecting, in our view, most significant breakthroughs in the field of polyP in microalgae. Certain seminal works were included to help in understand the developments in the field. The review starts from the general outline of polyP polymerase, its subcellular localization, structure, and function (mostly on the example of VTC), continues to the regulation of this enzyme, and concludes by its implications for biotechnology.
2. Localization in the Cell
Orthophosphate, if taken up in a large amount e.g., during luxury uptake and/or overcompensation response [5,38,39,40,41], can potentially displace the equilibria of phosphorylation-dephosphorylation reactions in the cell. Hence important roles of polyP polymerase in the cell includes the conversion of the surplus Pi into polyP and compartmentalization of the latter in a less metabolically compartments e.g., vacuole (Figure 1). This function is particularly important since short-chain polyP synthesized outside the ‘isolating’ vesicle or vacuolar compartment are believed to be toxic for the cell [42,43].
In the eukaryotic cell, polyP polymerase is typically associated with membranes of specialized vacuoles harboring polyP and called acidocalcisomes [44] or acidocalcisome-like vacuoles [45,46,47]. These vacuoles are hallmarked by the presence of polyP in form of electron-opaque granules, initially described in yeast as “volutin” [48] and colocalized with abundant mono- and divalent cations (e.g., K+, Ca2+) equilibrating the strong negative charge of polyP [8,11]. At the same time, ultrastructural studies of microalgae frequently reveal the presence of polyP-like granules in numerous other compartments and subcompartments [40,49,50] suggesting the existence of other sites of polyP biosynthesis in the cell. Thus, alternative sites for the initiation of polyP synthesis have been suggested for certain microalgae e.g., for Cyanidioschyzon merolae where small polyP granules are synthesized in cytosolic vesicles. Later, the small vacuoles merge with larger vacuoles where big polyP granules are eventually assembled [50]. These peculiarities in polyP biosynthesis were associated with the differences in the genetic determinants of the polyP biosynthesis in these organisms (see below).
Recently, highly ordered ultrastructure of polyP granules in microalgal vacuoles has been documented [51]. The authors suggested that such structure can result from concerted operation of several VTC complexes grouped in the vacuolar membrane into raft-like units. Such VTC arrays simultaneously synthesize several polyP chains wrapped in a sheath of organic substance resembling poly-(R)-3-hydroxybutyrate (PHB) so the whole assembly appeared on TEM images as a “multi-wire electric cable” (Figure 2, see also [38]). The whole picture partially resembled that of the VTC-mediated accumulation of polyP in yeast acidocalcisomes [31] of yeast [43]. As summarized in [38], the sophisticated structure of polyP chains interleaved with PHB was suggested earlier [52] and indirectly confirmed later in Parachlorella kessleri NIES-2152 [53].
3. Polyphosphate Polymerases in Microalgae
The homologs of the key components of the VTC enzyme complex are conserved in many unicellular eukaryotes [16,29,31] but animals and certain plant lineages (mostly Streptophyta and red algae) lack genes encoding this type of polyP polymerase [54]. The phylogenetic distribution of these genes suggests that the ancestral eukaryote possessed a homolog of the catalytic unit of VTC, VTC4 (see below) but multiple losses have occurred during eukaryotic evolution [16,55].
In vascular plants, the absence of a VTC4 homolog seemingly correlates with the loss of the polyP storage capacity: most of their P reserves are stored in form of Pi in the central vacuole of the cells or as phytic acid [54,55,56,57]. However, this loss does not seem to be complete: presence of polyP granules in higher plant cells have been reconfirmed recently along with the genes involved in their biosynthesis [55]. Overall, the presence of genes coding for polyP polymerases is considered to be an indication of the capability of the polyP biosynthesis. At the moment of this writing, the genes coding for VTC-type polyP kinase were detected in genomes of more than 50 microalgal species [15] but many more such discoveries are expected as a result of ongoing genome mining effort.
Current phylogenetic evidence on the putative VTC4-encoding sequences predicted by the homology search with BLAST and its 3D structure inferred with AlphaFold suggests that the capability of polyP biosynthesis represents a rather ancient ability. Likely, it has evolved with the species of the microalgal lineage. Still the results of the in silico genome mining and other findings made by bioinformatic tools need proper experimental validation.
The importance of this trait for the environmental fitness explains why the functionality of the VTC-type polyP polymerase persisted throughout evolution [15]. Phylogenetic analysis of the predicted VTC4-encoding genomic sequences of microalgae revealed that their phylogenetic relationships largely align with the taxonomic relationships between the species harboring corresponding VTC4 sequences [15]. These similarities and evolutionary relationships suggest that polyP synthesis represents an ancient ability that has evolved as the microalgal lineage has spread out over time. Thus, Cliff et al. [15] showed that VTC4 sequences from Auxenochlorella protothecoides, Micractinium conductrix, Volvox reticuliferus, and Haematococcus lacustris appear to be isoforms originating from a single gene for each species. The analysis carried out in the same work demonstrated that species of Chlorellales form two clades, one of which is more closely related to the remaining Chlorophyta sequences than the other clade and differing primarily by the sequences encoding the transmembrane domain. Recent genomic and transcriptomic analyses of the oleaginous microalga Nannochloropsis oceanica revealed four genes encoding proteins harboring SPX domain (named after the first proteins in which it was discovered—SYG1, Pho81, and XPR1) including SPX-VTC (NoSPX2) encoding a protein associated with vacuolar or acidocalcisomal membrane homologous to the VTC4 [58].
A consensus regarding the representation and functionality of polyP polymerases in eukaryotic microalgae is yet to be developed: while certain workers argue that the VTC is the only known polyP eukaryotic polymerase, there is a ground to believe that, at least in some species, VTC is complemented by the homologs of the bacterial type polyP polymerase [29]. Interestingly, the phylogenetic relationships and representation of PPK1, a highly conserved, bacterial-type polyP polymerase also typical for cyanobacteria [2], scarcely overlaps with the representation of VTC4 homologs. This observation suggests the possibility of differential selection between genes of distinct polyP polymerase types in each lineage [15] (although some chlorophytes possess both homologs [16]).
The results of genomic studies presented in [59] suggest that polyP kinases of some simple eukaryotic species like microalgae Ostreococcus tauri, O. lucimarinus, Porphyra yezoensis, and Cyanidioschyzon merolae and the moss Physcomitrium patens derived from prokaryotic enzymes e.g., PPK1 and PPK2 [60]. Most of the PPK homologs harbored by eukaryotic genomes are fused with the genes encoding other enzyme types known as nucleoside diphosphate-linked moiety X (Nudix) hydrolases belonging to the diphosphoinositol polyphosphate phosphohydrolase (DIPP) family specific to eukaryotes [61]. The genes coding for these enzymes were likely acquired as a result of several individual horizontal transfer events (for more detail on phylogenetic analysis of representation and distribution of PPK homologs in eukaryotes including microalgae, see [59]). The authors of this work argue that simpler “prokaryotic version” of polyP polymerase might give an advantage such as increased tolerance to stresses including desiccation or nutrient shortage for the simple eukaryotes that lack more sophisticated adaptations. Acquisition of the “bacterial type” polyP kinases they hypothetically associate with the boost of marine phytoplankton abilities to sequester Pi, hence accelerating global carbon fixation and transformation of ancient biosphere.
During recent decades, functional significance of many proteins harboring SPX domains and involved in metabolism of P has been revealed. Thus, SPX1-type polypeptides were found to be nuclear repressors of constitutively expressed phosphate starvation response 1 (PHR1); in plants, paralogs of PHR1 (PHL, PHR-like), are the transcription factors of the plant-specific MYB-CC class. Genome-wide analysis of differential gene expression, mostly in model microalga Chlamydomonas reinhardtii [62], revealed many genes and proteins responsible for acclimation of unicellular green algae to fluctuating Pi availability collectively referred to as “Pi starvation inducible” genes or PSI genes. These genes were also found in genomes of marine microalgae where they encode homologs of the yeast Pho84 and Pho89 high-affinity Pi transporters, VTC complex, and a broad diversity of enzymes (mostly phosphatases and nucleases) implementing Pi scavenging from surplus RNA and phospholipids [63,64,65].
The most studied VTC4 gene of microalgae is the one in the genome of Ch. reinhardtii [13,15,32,63,66,67]. Studies of VTC4 in other species are complicated by incompleteness and/or incorrect truncating of the sequences that were predicted to encode VTC4 protein and deposited into public databases: they appear to be missing either N-terminal SPX domain or transmembrane domain [15]. Still, homologs of VTC4 were found within > 50 species of microalgae inhabiting marine, freshwater, and terrestrial biotopes. Analysis of protein models generated by Cliff at al. for the predicted VTC4 proteins from four eukaryotic species (Chlorella vulgaris, Desmodesmus cf. armatus, Gonium pectorale, and Microcystis aeruginosa) demonstrated that the microalgae share the conserved VTC catalytic core and SPX phosphate-sensing domains found in the yeast VTC4 proteins. These microalgae also demonstrated the capability of enhanced polyP accumulation upon replenishment of Pi to P-starving cultures.
4. Structure and Function
The polyP polymerase catalyzes the elongation of polyP chain with Pi units from ATP hence adequate supply of this substrate is crucial for biosynthesis of polyP. In eukaryotic cells, the elongation of PolyP chain takes place along with the translocation of the nascent polyP chain through the vacuole membrane into the vacuole lumen [43]. These mechanistic findings were based on the biochemical and genetic evidence and recently confirmed by structure-guided functional analyses in yeast Saccharomyces cerevisiae [30]. The microalgal VTC protein sequences and corresponding yeast protein sequences displayed a high structural homology (38–44 % sequence identity with ca. 60 % sequence coverage including the VTC catalytic core) so their predicted structures turned to be very close except their unstructured outer loops [15]. These findings form a ground to believe in close similarity of structure and function of these enzymes in yeast and eukaryotic microalgae.
The VTC complex includes the catalytic subunit with PolyP polymerase activity (VTC4) and a set of accessory subunits (Table 1) whose composition depends on the subcellular localization of the complex and the organism [68]. Thus, VTC2 and VTC3 are homologous to VTC4 but are catalytically inactive; VTC1 is homologous to the transmembrane domains of the other subunits [69]. According to the current knowledge, microalgae only possess homologues of the VTC1 and VTC4 subunits of S. cerevisae, so the structure of the microalgal VTC complex is expected to differ from that of the yeast VTC. The 3D models of the predicted VTC4 proteins generated by Cliff et al. from the four eukaryotic species and showed that at least the studied microalgal species share the conserved VTC catalytic core and SPX phosphate-sensing domains initially mapped in the VTC4 proteins of yeast [15]. These findings further confirm the polyP polymerase function of the VTC4 protein in microalgae and supports the similarity of VTC4 regulation in microalgae and yeast. Still, the precise details of the assembly of the microalgal VTC complex and the mechanisms in control of this process are not known.
The role of the membrane-integrated vacuolar transporter chaperone (VTC) complex in the biosynthesis of polyP has been initially demonstrated in S. cerevisiae. Later it has been proved by reconstitution of an active Vtc1/Vtc3/Vtc4 (a pentamer with the stoichiometric ratio of 3:1:1) complex exhibiting polyP polymerase activity and capable of polyP synthesis activated by inositol (poly)phosphate in vivo [30,43]. In microalgae, the function of VTC4 was initially demonstrated in Ch. reinhardtii albeit no information on its structure was available at that time. Later, the VTC catalytic core and the SPX regulatory domain were shown as conserved among microalgae and yeast suggesting that VTC4 is functioning as a polyP polymerase in other microalgae as well. In addition, the affinity of the SPX domain to inositol phosphates was shown via molecular docking calculations [15].
Forward genetics indicated that the components if polyP polymerase complex, namely VTC1 are involved in acclimation to fluctuations [7] of the availability of other nutrients, such as sulfur (S) or nitrogen (N). Thus, the ars76 mutant deficient in arylsulfatase is also impaired in acidocalcisome formation and more prone to death under S- or N-deprived conditions compared with wild-type [66].
Recent breakthrough in the structure of the VTC complex was achieved by Guan et al. [30] employing cryo-electron microscopy to resolve the structure of yeast VTC complex at 3.0 Å resolution (Figure 3). Although VTC complex was initially predicted to have nine transmembrane helices, subsequent cryo-EM structural study revealed that the complexes actually possess fifteen trans-membrane helices, three helices per subunit. In yeast, 15 trans-membrane helices form a novel membrane channel (Figure 3) enabling the transport of newly synthesized polyP chain into the vacuolar lumen. Notably, the SPX and TTM domains of VTC4 are tightly interacting (Figure 3a). The exit tunnel of the catalytic TTM domain is positioned at the entrance of the trans-membrane channel (Figure 3), the latter is connected to TTM by a well-defined loop interacting with the N-terminus of Vtc1A protomer at the membrane-proximal region (Figure 3b).
Negatively charged ligands e.g., Pi, pyrophosphate (PPi), sulfate, and polyP) can also bind to the TTM domain enabling the initial binding of PPias a primer for the synthesis of polyP [31]. Two Pi ions and two lipid molecules bind to the cytoplasmic and luminal side of the VTC transmembrane channel, respectively (Figure 3b, c).
Bacterial-type polyP polymerase, PPK1 is a dimer forming a catalytic tunnel, in which polyP is elongated by the transfer of Pi from ATP and the elongating polyP chain exits from the other side [70]. The amino acid sequence of PPK1 consists of highly conserved amino- and carboxy-terminal domains (N and C domains) and a less conserved middle ‘head’ (H) domain; the N and C domains provide the binding site for ATP, and the C domain is responsible for the catalytic function [70]. In addition to the forward reaction of polyP chain elongation, PPK1 catalyzes the reverse reaction (cleavage of Pi from the PolyP chain) as well as nucleoside diphosphate kinase reactions with Pi transferred from PolyP, and 5′-tetraphosphate by transferring of PPi to guanosine diphosphate [71].
5. Regulation of Polyphosphate Polymerase Function
Crucial insights into the regulation of VTC functioning were achieved only recently (see e.g., [30]). In yeast and eukaryote microalgae, polyP synthesis is regulated by inositol pyrophosphate (PP-InsP) nutrient messengers directly sensed by the VTC complex. In eukaryotic microalgae, as in yeast [16] or animals (and humans) [72], cell Pi level and also its energy status is manifested by signal molecules normally present in small (<5 µM) concentrations. Inositol pyrophosphates, PP-InsP like 1,5-IP8 can serve as an example [73,74]. Thus, the SPX domains of VTC3 and/or VTC4 subunits of the VTC complex can bind PP-InsP molecules to the surface collectively formed by basic amino acid residues called phosphate binding cluster (PBC) and lysine surface cluster (KSC) together provide a binding surface for PP-InsPs (for more detail, see [30,31]). In addition to that, InsP6-activated VTC complex binds molecules of InsP6.
Overall, the VTC complex harbors three InsP6 molecules: one bound to the basic surface patch of the PP-InsP sensing SPX domain (Figure 3a), second bound simultaneously to the tunnel domains of VTC3 and VTC4, likely promoting their association (Figure 3c), and the third InsP6 was found in the positively charged tunnel domain of VTC4, in a binding site that is normally occupied by the ATP substrate. Binding PP-InsP results in the orientation of the catalytic polymerase domain at the entrance of the trans-membrane channel activating the enzyme and coupling PolyP synthesis with its translocation through the vacuolar membrane (see Figure 3 in [30]).
Coupling of PP-InsP with P- level is implemented via two-stage phosphorylation: first of IP6 to 5-IP7 (by inositol phosphate kinase 1, ITPK1), then 5-IP7 to 1,5-IP8 (by VIH1/VIH2 or Vip1 homolog kinases, see Figure 4). Replete Pi and ample ATP in the cell boost VIH kinase activity while Pi shortage reciprocally fosters phosphatase activity of the VIH kinase [75,76]. The net results of this, as summarized in [16], is oscillation of the energy metabolites in synchrony with derivative signal molecules to adjust and orchestrate metabolism according to Pi supply. The term “INPHORS pathway” was coined for collective designation of these processes [73].
Interestingly, inositol pyrophosphates assumed the role of central regulators of Pi homeostasis in animal and plant like Arabidopsis thaliana and Oryza sativa [73] cells as well. In microalgae, PP-InsPs are directly involved in control of polyP biosynthesis and important P reserves stored in vacuole [77,78]. The SPX-type receptors were also encountered in plant and human Pi transporters indicating potential similarities in the mechanisms of their regulation. The SPX sensor domains responsive to Pi in the cell interior were discovered in four plant protein families. They harbor a single SPX domain constituting the entire protein (these proteins were named ‘stand-alone’ SPX polypeptides exemplified by SPX1–SPX4 proteins of A. thaliana) or occupying the N-terminal region followed by additional domains. The latter protein type includes SPX–MFS protein (PHT5/VPT) family, SPX-EXS proteins (PHO1 family) and SPX– RING proteins (NLA family) [79,80].
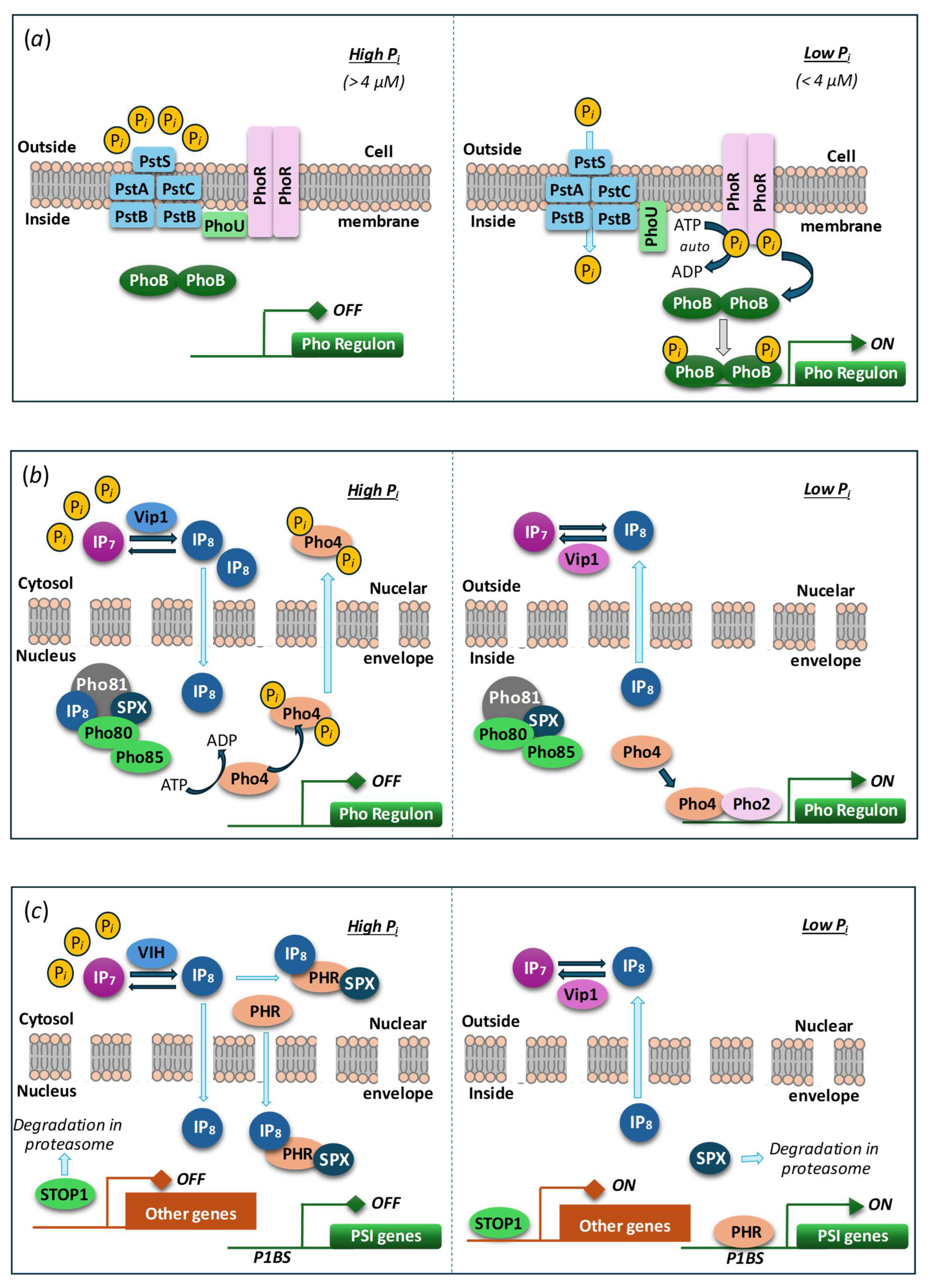
The mechanisms of Pi sensing and response to its level in microalgae and yeast are also distinct: the transcription factor Phosphate Starvation Response 1 (PSR1) is a key mediator of P starvation responses in algae while PhoR/PhoB regulon is operating in bacteria (like Escherichia coli) and Pho80/Pho85/Pho4 regulon—in yeast (S. cerevisiae), cf. panels (a)-(c) in Figure 4.
PSR1 and other orthologs of PHR1/PHL being master transcriptional regulators of the systemic Pi starvation response regulates the expression of many PSI genes in algae [81]. PSR1 belongs to a large Myb (Myeloblastosis) family of nuclear transcription factors common for eukaryotic organisms [65,82]. Overall, the discovery of SPX proteins and VTC homologs regulated via SPX domains across three phyla of microalgae might be indicative of a conserved mechanisms of intracellular Pi sensing via PP-InsP molecules [15,54,65].
Figure 4.
Minimal models of Pi sensing and core transcriptional response circuits. (a) Pho regulation in bacteria (presumably, including cyanobacteria). The Pi-specific transporter (Pst), composed of four different Pst subunits (blue), functions as a sensor of extracellular Pi. In Pi sufficiency, Pi binds to PstS and inactivates PhoR, a histidine sensor kinase, via PhoU-mediated recruitment to PstB. In Pi limitation, Pi is taken up by Pst, which activates autophosphorylation of PhoR, subsequent phosphorylation and activation of PhoB, a transcriptional response regulator of diverse Pho box-containing promoters (Pho regulon). (b) Pho regulation in yeast (S. cerevisiae). In Pi sufficiency, Vip1, a bifunctional IP7 kinase/IP8 phosphatase signals intracellular Pi status via the dynamics of IP8 level to the cell nucleus. IP8 binds to the SPX domain of Pho81, an inhibitor of the cyclin-dependent protein kinase complex, Pho85 (kinase)–Pho80 (cyclin), which causes partial Pho81 dissociation and thus Pho85–Pho80 kinase activation. Phosphorylation of Pho4, a basic helix-loop-helix transcription factor facilitates nuclear Pho4 export and Pho gene repression. In Pi limitation, IP8 level drops (activation of Vip1 IP8 phosphatase), which represses Pho85–Pho80 kinase activity via tighter binding to Pho81. Dephosphorylated, nuclear Pho4 interacts with Pho2, a homeodomain transcription factor, to activate Pho gene expression. (c) Regulation of PSI genes in land plants (A. thaliana). Like in yeast, the dynamics of IP7/IP8 levels, responding to intracellular Pi and energy status via the bifunctional kinase/ phosphatase VIH, control the activation of Pi-responsive genes. In high Pi, IP8 mediates the recruitment of SPX proteins to PHR transcription factors of the plant-specific MYB-CC class to repress PSI gene activation. When IP8 levels drop in low Pi conditions, PHR–SPX complexes dissociate, leading to PSI gene activation and SPX protein degradation. Unlike the systemic Pi response, local Pi sensing of extracellular Pi availability is controlled by yet unknown processes (involving proteolysis), STOP1, a zinc finger transcription factor, which activates some of its target genes in low Pi, such as ALMT1. For details, see the recent reviews (e.g., [73,83]). Reprinted from [16].
Figure 4.
Minimal models of Pi sensing and core transcriptional response circuits. (a) Pho regulation in bacteria (presumably, including cyanobacteria). The Pi-specific transporter (Pst), composed of four different Pst subunits (blue), functions as a sensor of extracellular Pi. In Pi sufficiency, Pi binds to PstS and inactivates PhoR, a histidine sensor kinase, via PhoU-mediated recruitment to PstB. In Pi limitation, Pi is taken up by Pst, which activates autophosphorylation of PhoR, subsequent phosphorylation and activation of PhoB, a transcriptional response regulator of diverse Pho box-containing promoters (Pho regulon). (b) Pho regulation in yeast (S. cerevisiae). In Pi sufficiency, Vip1, a bifunctional IP7 kinase/IP8 phosphatase signals intracellular Pi status via the dynamics of IP8 level to the cell nucleus. IP8 binds to the SPX domain of Pho81, an inhibitor of the cyclin-dependent protein kinase complex, Pho85 (kinase)–Pho80 (cyclin), which causes partial Pho81 dissociation and thus Pho85–Pho80 kinase activation. Phosphorylation of Pho4, a basic helix-loop-helix transcription factor facilitates nuclear Pho4 export and Pho gene repression. In Pi limitation, IP8 level drops (activation of Vip1 IP8 phosphatase), which represses Pho85–Pho80 kinase activity via tighter binding to Pho81. Dephosphorylated, nuclear Pho4 interacts with Pho2, a homeodomain transcription factor, to activate Pho gene expression. (c) Regulation of PSI genes in land plants (A. thaliana). Like in yeast, the dynamics of IP7/IP8 levels, responding to intracellular Pi and energy status via the bifunctional kinase/ phosphatase VIH, control the activation of Pi-responsive genes. In high Pi, IP8 mediates the recruitment of SPX proteins to PHR transcription factors of the plant-specific MYB-CC class to repress PSI gene activation. When IP8 levels drop in low Pi conditions, PHR–SPX complexes dissociate, leading to PSI gene activation and SPX protein degradation. Unlike the systemic Pi response, local Pi sensing of extracellular Pi availability is controlled by yet unknown processes (involving proteolysis), STOP1, a zinc finger transcription factor, which activates some of its target genes in low Pi, such as ALMT1. For details, see the recent reviews (e.g., [73,83]). Reprinted from [16].
Despite recent discoveries, the reports on the differential expression of VTC components and/or other polyP polymerases in green microalgae are still scarce in the literature. Current evidence shows that massive uptake of Pi is followed by a transient spike of polyP biosynthesis which is preceded by a peak of the expression of the gene encoding VTC in Ch. reinhardtii [32] and Chlorella vulgaris [39]. Similar indications of the relationships between VTC expression, polyP, and P availability were found in the coccolithophore Emiliania huxleyi [84,85] and the diatom Thalassiosira pseudonana [86,87].
Plausible mechanisms in control of the genes responsible to Piavailability in bacteria (including cyanobacteria), yeast, and vascular plants are summarized in [16], see also Figure 4. The latter is thought to be the closest to the one operation in eukaryotic microalgae but actual mechanism of Pi sensing and gene expression control in these organisms can include a mix of components homologous to those in yeast and plants. It is also conceivable that a similar mechanism can be involved in the regulation of VTC-encoding genes. Briefly, under ample Pi conditions, proteins with SPX domains bind 1,5-IP8 with high affinity (7–50 µM) [78]. The [SPX : 1,5-IP8] complex then docks to the PHR1/PHL protein. As a result, its dimer dissociates and the monomers interact with the MYB-like DNA-binding domain blocking target gene activation [79]. Under low-Pi conditions, a decrease in 1,5-IP8 level declines the [SPX:1,5-IP8] complex formation and its interaction with other proteins promoting PHR1/PHL (hetero)oligomerization and activation of PSI genes via recognition of PHR1-binding sequences (P1BS) promoter elements. Positive and negative feedback loops for the expression of the genes harboring SPX domains are provided by the degradation of ligand-free SPX and P1BS-mediated proteins activation provide, respectively. Obviously, in this regulatory framework the expression of the VTC-encoding genes will be activated under ample Pi conditions and repressed under shortage of this nutrient.
6. Eco-Physiological Significance of Polyphosphate Polymerases
As outlined above, polyP polymerase is involved in modulation of cellular Pi storage by synthesizing polyP when Pi is ample in response to stimulation by high PP-InsPs levels. Its important role is participation in the responses of microalgae to fluctuations in bioavailability of P [88]. Accordingly, the function of polyP polymerase is at the foundation of microalgal resilience to environmental stresses, especially nutrient shortage, and their ability to survive or even thrive in niches with high level of competition e.g., phytoplankton [89,90]. This might be an explanation of the famous “paradox of phytoplankton” (which is characterized by a large diversity of species although a dominance of the most competitive is expected). This hypothesis also connects within the “starve out the competitor” strategy of competition in microbial communities [91]. Overall, microalgal species capable of rapid uptake and intracellular storing of P during short periods of its high availability will have the chance to outcompete other species less capable of doing so. All this highlights the importance of the intracellular P storing mechanisms and VTC as their important component for environmental fitness and competitiveness of microorganisms, including photosynthetic ones. In a similar way, cyanobacterial PPK1 is essential for the dynamic regulation of P metabolism, including hyperaccumulation of polyP and to maintain physiological fitness under other nutrient (e.g., sulfur) starvation conditions [28]. Thus, knocking out of NoSPX2 gene, a VTC4 homolog from Nannochloropsis oceanica, diminished polyP levels and hindered the uptake of Pi, confirming its role in polyP biosynthesis and Pi homeostasis [58].
Another role of VTC, the long-term storage of energy, is debated until now. On one hand, the Pi units of polyP chains are connected by high-energy phosphor anhydride bonds, so the energy content of polyP is deemed to be significant [92,93]. Furthermore, knock-out strains lacking polyP polymerase display higher productivity [94]. On the other hand, the contribution of polyP synthesized by VTC to the total energy budget of the cell is questioned on the grounds of little or no effect of active polyP biosynthesis on carbon assimilation and biomass accumulation [15]. At the same time, active biosynthesis of polyP normally takes place when the cell division rate is slow (during lag-phase after Pi replenishment following P starvation or at the stationary phase) i.e., in situations when the energy demand is not so high as in the periods of active cell division. Therefore, the assessment of real impact of polyP biosynthesis on assimilatory processes in the cell is complicated.
Although polyP are thought to function as chaperone protecting proteins against stress-induced misfolding [95], short-chain polyP seemingly can interfere with folding of proteins in certain situations e.g., when the Pi is ample, but the energy is in short supply and cannot support the simultaneous elongation of many polyP chains [96,97] initiated outside the vesicles or vacuoles [42]. The symptoms of Pi toxicity, namely cell division slowdown and cell structure disorganization, recently described in microalgae like Chlorella [96] and Micractinium [97] are hypothesized to be mediated by the ‘ectopic’ initiation of short-chain polyP [42] observed in P-starved microalgae upon abrupt replenishment of Pi. This hypothesis explains the phenomenon of failed Pi tolerance (which is normally quite high [49]) by analogy with the toxic effect of short-chain PolyP initially described in the yeast [43] and, recently, implied in Chlorella regularis [96,98]. The P-toxicity syndrome links the failure of Pi resilience of microalgae initiation of polyP upon an abrupt rise in cytoplasmic Pi at the VTC complexes with subsequent shortage in ATP, the essential substrate for the polyP chain elongation, likely due to metabolic shutdown. It was shown (so far in yeast) that the cell can cope with the toxicity of non-vacuolar polyP by expressing polyphosphatases degrading these polyP [42]. Currently accumulated evidence suggests that the presence, activity of, and adequate energy supply to VTC are crucial determinants of the tolerance to the exogenic Pi spikes. Namely, VTC mediates compartmentalization of the Pi taken up in excess and translocation of the nascent polyP chain into the vacuole preventing thereby toxicity of short-chained polyP [49,97,99].
7. Implications for Biotechnology and Sustainability of P Usage
Currently, humanity relies very much on mined P minerals which are finite, non-renewable, and non-uniformly distributed [100,101,102,103]. This problem is exacerbated by extremely inefficient use of mined P and the products derived from it, especially mineral P fertilizers [101,104,105,106]. The usage of microalgal biotechnologies was proposed as a promising vehicle for making use of P resources more sustainable by means of capturing P from waste streams and re-routing it to agricultural ecosystems (see [107,108,109,110] and references therein).
As outlined above, the capability for biosynthesis of polyP is important for environmental fitness of microalgae. Therefore, it is no surprise that microalgae isolated from different habitats display the ability to synthesize polyP, and that VTC4 is broadly conserved. Continuously facing fluctuating P availability, microalgae evolved to be very efficient at P acquisition and as intracellular storing as polyP. This capability could be harnessed for sustainable usage of P, namely, to recycle P from waste streams and side streams and to mitigate water pollution. Indeed, the efficiency and competitiveness of the biotechnological solutions for sustainable use of P is determined by the rate and completeness of P bioremoval as well as by the size of cell P quota (influenced profoundly by polyP accumulation capacity) of microalgal strains used in these solutions.
Environmental parameters such as N:P ratio, light intensity, or temperature influencing P accumulation in microalgal cells during wastewater treatment, are sensed and relayed in the cells to modulate eventually the expression of specific P-related genes and proteins (including VTC), but the underlying mechanisms of these processes are currently not known. The findings of [32,64] indicated that the differences in the expression of the genes participating in the biosynthesis of polyP cannot completely explain the variation of polyP yield and kinetic, even in the model microalga Ch. reinhardtii. Accordingly, the synthesis of polyP is believed to be regulated at the post-transcriptional level in microalgae, like in other organisms such as yeast, by interaction with PP-InP [8].
The considerations presented above make obvious the strong dependence the capability of microalgal cultures to uptake and store Pi in their cells and the performance of polyP polymerase [36,111]. Therefore, microalgal strains capable of rapid mobilization and uptake of P from diverse waste streams with subsequent conversion of the Pi taken up into polyP are in high demand. These goals can be achieved by bioprospecting new strains with desired traits and/or by gaining a better control on P metabolism. Another plausible way is cell selection, especially in frame of approach called “adaptive laboratory evolution” [112,113,114].
As the efficiency of microalgae in bioremoval of P is directly related with VTC which is a very efficient enzyme [31], deeper insights into the structure, functioning and its regulation of VTC are needed to gain more control over P-metabolism in microalgae. For example, an ambitious goal is constituted by engineering of VTC for triggering polyP synthesis on-demand which can be indispensable for biotechnological P-upcycling. Site-directed mutagenesis can be an approach to boosting the activity of VTC: for example, mutation of K24 and R31 amino acids to alanine in yeast VTC1 increased activity of the whole VTC complex [30]. This might be of direct consequences for the efficiency of microalgal technologies for bioremoval of P from waste streams. It is also conceivable that microalgal strains with boosted VTC activity will be not only more efficient at Pi scavenging but also less prone to short-chain polyP-mediated Pi toxicity (see above) and hence more tolerant to Pi spikes in the wastewater. Recently, polyP came to spotlight as a valuable commodity [36,115], potential antiviral [116] and anti-inflammatory agent [117], so boosting polyP accumulation in microalgae can be another driver for engineering of VTC for productivity.
On the other hand, knocking down the genes responsible for the conversion of Pi into polyP might be considered for enhancing the P deprivation effects, such as stimulation of accumulation reserve lipids and other target compounds in microalgal cultures. Thus, disruption of Nospx2 gene encoding a putative polyP polymerase in N. oceanica considerably increased triacylglycerol and biomass accumulation under Pi deficiency conditions [58]. A knock-out strain of the cyanobacterium Synechocystis sp. PCC 6803 lacking the polyphosphate kinase (ppk) and harboring a foreign ethylene-forming enzyme was obtained [94]. The engineered strain displayed higher ATP levels, faster growth, and better ethylene productivity than the wild type under optimal conditions but lagged under stress [94]. So, the gain in productivity achieved by knocking out polyP-mediated biosynthesis of polyP comes at a cost of lower stress resilience requiring more effort to maintain optimal cultivation conditions. Finally, a reservation should be made regarding the current legislation which frequently prohibits the release of engineered strains into environment making their cultivation possible only in closed photobioreactors [118].
Adequate supply of energy is important for uninterrupted flow of substrate (ATP) to VTC and hence for unhindered accumulation of polyP. The preferred source of ATP for the biosynthesis of polyP is photophosphorylation whereas dark respiration is less efficient in this regard; the formation of polyP is possible but is impaired profoundly under anaerobic conditions [119], likely due to the shortage of ATP. Therefore, the cultivation systems for microalgal bioremoval of Pi and generation polyP-rich biomass should be designed with the requirement of sufficient light energy supply in mind.
Finally, the representation and abundance of VTC in metagenomes can be a useful criterion in the selection of efficient algal-bacterial consortia for environmental biotechnology via functional profiling of the consortia metagenomes. Although many biotechnological applications of microalgae such as production of high-value metabolites are based on axenic cultures, the use of the mutualistic consortia comprised by microalgae and associated heterotrophic bacteria quickly comes to foreground, especially in nutrient bioremoval, bioremediation, and wastewater treatment [120]. This approach using mining of metagenomes to reveal the genetic determinants of the robustness and productivity of the consortia quickly becomes a hot research direction and a basis for the designing of artificial consortia tailored to the application. Thus, consortia of “high-performance P absorbers” for application e.g., in bioscrubbers used in wastewater polishing for can be engineered from individual strains featuring active VTC and hence a high capacity for taking up exogenic Pi and storing it in form of polyP. Although the mainstream method for metagenomic studies was high-throughput sequencing (HTS), the advent and rapid development of the 3rd generation nanopore sequencing made recent breakthrough in this field. Development of novel bioinformatic algorithms for processing the metagenomic datasets opened new opportunities for the analysis of structure and physiology of microalgal-bacterial communities. From the practical perspective, the new HTS techniques became a time- and labor-savers in discovery of new microalgae with a high potential for the accumulation of valuable metabolites, biodegradation of hazardous micropollutants, and biosequestration of nutrients from waste streams. The insights from the both short-read and long-read metagenomics will form a solid foundation for the rational design of microalgal-bacterial consortia for biotechnology.
8. Conclusions and Outlook
Recent years were hallmarked by important breakthroughs in our understanding of the genetic determinants, structure, and function of polyP polymerases in microalgae. Fundamental discoveries in the relevant model organism, S. cerevisiae paved the way for these breakthroughs which shed light on numerous aspects of P metabolism and storage in microalgal cells. These findings are important not only for basic science but for diverse applications in microalgal biotechnology as well. At the same time, many questions related with P metabolism in microalgae remain unanswered. Thus, the exact mechanism of polyP catabolism, especially of polyP compartmentalized in vacuoles and packed into large PolyP granules, are not known. In bacteria, polyP is degraded by exopolyphosphatase cleaving terminal phosphate off the polyP chains by hydrolysis or by endophosphatases removing larger fragments of the polyP chain, a similar mechanism operates in yeast. PolyP can also be catabolized via the reactions of phosphorylation of glucose, adenosine monophosphate, or NAD involving polyP as Pi donors [121]. However, as was inferred from the studies of Ch. reinhardtii, microalgae differ from yeast regarding polyP turnover since the microalga lacks homologs of the yeast exopolyphosphatase PPX1. Although Plouviez et al. [32] suggested that a NUDIX-box containing protein (see also [59,61]) may be responsible for polyP degradation in C. reinhardtii, microalgal pathways for degradation of polyP remain vaguely known. Still, this question is of considerable importance given the existence of different polyP fractions in microalgal cells was discovered more than 60 years back [122,123,124,125] but have not yet fully explained. Whether different polyP polymerases are involved in the formation of low-mobility and high-mobility polyP fractions, or the main difference between these fractions is in their compartmentalization in the cell, remains unknown, too. At the same time, assessing the contributions of different polyP fractions to cell P quota and finding the conditions for mobilization of low-mobility polyP fractions would be valuable insights for biotechnology. In this context, polyP toxicity and the role of polyP polymerases in its induction and mitigation also comes into foreground, but now our understanding of this problem is limited.
Of special importance are further structural studies of microalgal VTC-type polyP polymerases. One might expect that these studies will help to explain, or even predict the species-specific differences in the capacity for PolyP accumulation and yield further mechanistic insights about post-translational regulation of polyP synthesis. Progress in this field requires establishing the protocols for production of microalgal polyP polymerases by heterologous expression and subsequent in vitro characterization: Cliff et al. [15] rightly noted that current hypotheses about the regulation of PolyP synthesis in microalgae (e.g., by PP-InP binding to VTC) require experimental confirmation. At the same time, new and improved algorithms for modelling and prediction of VTC characteristics in silico (including the new machine-learning algorithms like AlphaFold 3 [126]) could be used as a screening tool for the assessment of potential efficiency of PolyP synthesizing capacity in microalgae. This method will facilitate the selection of “top performing” microalgae in terms of P bioremoval from waste streams and accumulation of valuable PolyP. It is also an indispensable tool for functional profiling of algal-bacterial consortia and assessment of their potential for the same purpose. Both approaches will be useful for the development of microalgal biotechnologies for sustainable usage of P.
Finally, development of adaptive technologies for up-cycling of P from waste streams suitable for integration into conventional processes of waste treatment will be a major challenge in the nearest future. There is a ground to believe that deeper insights into the function and regulation of microalgal polyP polymerase function would help immensely to cope with this challenge.
Funding
The study was carried out under the state assignment of Lomonosov Moscow State University.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Abbreviations
The following abbreviations are used in this manuscript:
| P | Phosphorus |
| PHB | Poly-(R)-3-hydroxybutyrate |
| polyP | Polyphosphate |
| PP-InsP | Inositol pyrophosphate |
| PPK | Polyphosphate kinase |
| VTC | Vacuolar Transport Chaperone |
References
- Brown, M.R.; Kornberg, A. Inorganic polyphosphate in the origin and survival of species. Proceedings of the National Academy of Sciences of the United States of America 2004, 101, 16085–16087. [CrossRef]
- Rao, N.N.; Gómez-García, M.R.; Kornberg, A. Inorganic polyphosphate: essential for growth and survival. Annual Review of Biochemistry 2009, 78, 605–647. [CrossRef]
- Wimmer, J.L.E.; Kleinermanns, K.; Martin, W.F. Pyrophosphate and irreversibility in evolution, or why PPi is not an energy currency and why nature chose triphosphates. Frontiers in Microbiology 2021, 12. [CrossRef]
- Dyhrman, S.T. Nutrients and their acquisition: phosphorus physiology in microalgae. In The Physiology of Microalgae, Borowitzka, M.A., Beardall, J., Raven, J.A., Eds. Springer International Publishing: Cham, 2016; 10.1007/978-3-319-24945-2_8pp. 155–183.
- Cembella, A.D.; Antia, N.J.; Harrison, P.J. The utilization of inorganic and organic phosphorous compounds as nutrients by eukaryotic microalgae: A multidisciplinary perspective: Part I. Critical reviews in microbiology 1982, 10, 317–391. [CrossRef]
- Blank, L.M. The cell and P: from cellular function to biotechnological application. Current Opinion in Biotechnology 2012, 23, 846–851. [CrossRef]
- Thomas, M.R.; O’Shea, E.K. An intracellular phosphate buffer filters transient fluctuations in extracellular phosphate levels. Proceedings of the National Academy of Sciences 2005, 102, 9565–9570. [CrossRef]
- Azevedo, C.; Saiardi, A. Eukaryotic phosphate homeostasis: the inositol pyrophosphate perspective. Trends in biochemical sciences 2017, 42, 219–231.
- Tiwari, B. Chapter 7 - Phosphate metabolism in cyanobacteria: fundamental prospective and applications. In Cyanobacteria, Mishra, A.K., Singh, S.S., Eds. Academic Press: 2024; 10.1016/B978-0-443-13231-5.00002-7pp. 159–175.
- Mason-Jones, K.; Robinson, S.L.; Veen, G.F.C.; Manzoni, S.; van der Putten, W.H. Microbial storage and its implications for soil ecology. ISME Journal 2021, 10.1038/s41396-021-01110-w. [CrossRef]
- Blaby-Haas, C.E.; Merchant, S.S. Regulating cellular trace metal economy in algae. Curr Opin Plant Biol 2017, 39, 88–96. [CrossRef]
- Achbergerová, L.; Nahálka, J. Polyphosphate-an ancient energy source and active metabolic regulator. Microb Cell Fact 2011, 10, 14170–14175. [CrossRef]
- Sanz-Luque, E.; Bhaya, D.; Grossman, A.R. Polyphosphate: A Multifunctional Metabolite in Cyanobacteria and Algae. Frontiers in Plant Science 2020, 11. [CrossRef]
- Denoncourt, A.; Downey, M. Model systems for studying polyphosphate biology: a focus on microorganisms. Curr Genet 2021, 10.1007/s00294-020-01148-x. [CrossRef]
- Cliff, A.; Guieysse, B.; Brown, N.; Lockhart, P.; Dubreucq, E.; Plouviez, M. Polyphosphate synthesis is an evolutionarily ancient phosphorus storage strategy in microalgae. Algal Research 2023, 7. [CrossRef]
- Abel, S.; Naumann, C. Evolution of phosphate scouting in the terrestrial biosphere. Philosophical Transactions B 2024, 379, 20230355. [CrossRef]
- Wang, L.; Fraley, C.D.; Faridi, J.; Kornberg, A.; Roth, R.A. Inorganic polyphosphate stimulates mammalian TOR, a kinase involved in the proliferation of mammary cancer cells. Proceedings of the National Academy of Sciences 2003, 100, 11249–11254. [CrossRef]
- Müller, F.; Mutch, N.J.; Schenk, W.A.; Smith, S.A.; Esterl, L.; Spronk, H.M.; Schmidbauer, S.; Gahl, W.A.; Morrissey, J.H.; Renné, T. Platelet polyphosphates are proinflammatory and procoagulant mediators in vivo. Cell 2009, 139, 1143–1156. [CrossRef]
- Omelon, S.J.; Grynpas, M.D. Relationships between polyphosphate chemistry, biochemistry and apatite biomineralization. Chemical reviews 2008, 108, 4694–4715. [CrossRef]
- Hernandez-Ruiz, L.; González-García, I.; Castro, C.; Brieva, J.A.; Ruiz, F.A. Inorganic polyphosphate and specific induction of apoptosis in human plasma cells. Haematologica 2006, 91, 1180–1186.
- Abramov, A.Y.; Fraley, C.; Diao, C.T.; Winkfein, R.; Colicos, M.A.; Duchen, M.R.; French, R.J.; Pavlov, E. Targeted polyphosphatase expression alters mitochondrial metabolism and inhibits calcium-dependent cell death. Proceedings of the National Academy of Sciences 2007, 104, 18091–18096. [CrossRef]
- Gray, M.J.; Wholey, W.-Y.; Wagner, N.O.; Cremers, C.M.; Mueller-Schickert, A.; Hock, N.T.; Krieger, A.G.; Smith, E.M.; Bender, R.A.; Bardwell, J.C. Polyphosphate is a primordial chaperone. Molecular cell 2014, 53, 689–699. [CrossRef]
- Kuroda, A.; Nomura, K.; Ohtomo, R.; Kato, J.; Ikeda, T.; Takiguchi, N.; Ohtake, H.; Kornberg, A. Role of inorganic polyphosphate in promoting ribosomal protein degradation by the Lon protease in E. coli. Science 2001, 293, 705–708. [CrossRef]
- Cremers, C.M.; Knoefler, D.; Gates, S.; Martin, N.; Dahl, J.-U.; Lempart, J.; Xie, L.; Chapman, M.R.; Galvan, V.; Southworth, D.R. Polyphosphate: a conserved modifier of amyloidogenic processes. Molecular cell 2016, 63, 768–780. [CrossRef]
- Xie, L.; Jakob, U. Inorganic polyphosphate, a multifunctional polyanionic protein scaffold. Journal of Biological Chemistry 2019, 294, 2180–2190. [CrossRef]
- Abdelkarim, O.H.; Verhagen, R.A.; Wijffels, R.H.; Barbosa, M.J. Physiological, biochemical, and morphological responses to nitrogen starvation and biomass-specific photon supply rates of Nannochloropsis oceanica and Microchloropsis gaditana. Journal of Applied Phycology 2025, 10.1007/s10811-025-03569-y. [CrossRef]
- Cohen, A.; Perzov, N.; Nelson, H.; Nelson, N. A novel family of yeast chaperons involved in the distribution of V-ATPase and other membrane proteins. Journal of Biological Chemistry 1999, 274, 26885–26893.
- Sato, N.; Endo, M.; Nishi, H.; Fujiwara, S.; Tsuzuki, M. Polyphosphate-kinase-1 dependent polyphosphate hyperaccumulation for acclimation to nutrient loss in the cyanobacterium, Synechocystis sp. PCC 6803. Frontiers in Plant Science 2024, 15. [CrossRef]
- Yagisawa, F.; Fujiwara, T.; Yamashita, S.; Hirooka, S.; Tamashiro, K.; Izumi, J.; Kanesaki, Y.; Onuma, R.; Misumi, O.; Nakamura, S. A fusion protein of polyphosphate kinase 1 (PPK1) and a Nudix hydrolase is involved in inorganic polyphosphate accumulation in the unicellular red alga Cyanidioschyzon merolae. Plant Molecular Biology 2025, 115, 1–21. [CrossRef]
- Guan, Z.; Chen, J.; Liu, R.; Chen, Y.; Xing, Q.; Du, Z.; Cheng, M.; Hu, J.; Zhang, W.; Mei, W., et al. The cytoplasmic synthesis and coupled membrane translocation of eukaryotic polyphosphate by signal-activated VTC complex. Nat Commun 2023, 14, 718. [CrossRef]
- Hothorn, M.; Neumann, H.; Lenherr, E.D.; Wehner, M.; Rybin, V.; Hassa, P.O.; Uttenweiler, A.; Reinhardt, M.; Schmidt, A.; Seiler, J.J.S. Catalytic core of a membrane-associated eukaryotic polyphosphate polymerase. Science 2009, 324, 513–516. [CrossRef]
- Plouviez, M.; Fernandez, E.; Grossman, A.R.; Sanz-Luque, E.; Sells, M.; Wheeler, D.; Guieysse, B. Responses of Chlamydomonas reinhardtii during the transition from P-deficient to P-sufficient growth (the P-overplus response): The roles of the vacuolar transport chaperones and polyphosphate synthesis. J Phycol 2021, 10.1111/jpy.13145. [CrossRef]
- Plouviez, M.; Brown, N. Polyphosphate accumulation in microalgae and cyanobacteria: recent advances and opportunities for phosphorus upcycling. Curr Opin Biotechnol 2024, 90, 103207. [CrossRef]
- Yu, D.; Yan, L.; Shi, J.; Liu, Y.; Zhang, A.; Wang, Y.; Zhang, Y.; Xie, T. Phosphorus removal and recovery during microalgae-based wastewater treatment: a mini-review. International Journal of Environmental Research 2024, 18, 34. [CrossRef]
- Demling, P.; Baier, M.; Deitert, A.; Fees, J.; Blank, L.M. Biotechnological polyphosphate as an opportunity to contribute to the circularization of the phosphate economy. Current Opinion in Biotechnology 2024, 87, 103107. [CrossRef]
- Blank, L.M. (Poly)phosphate biotechnology: Envisaged contributions to a sustainable P future. Microb Biotechnol 2023, 16, 1616–1622. [CrossRef]
- Solovchenko, A.; Plouviez, M.; Khozin-Goldberg, I. Getting grip on phosphorus: potential of microalgae as a vehicle for sustainable usage of this macronutrient. Plants 2024, 13, 1834. [CrossRef]
- Solovchenko, A.E.; Ismagulova, T.T.; Lukyanov, A.A.; Vasilieva, S.G.; Konyukhov, I.V.; Pogosyan, S.I.; Lobakova, E.S.; Gorelova, O.A. Luxury phosphorus uptake in microalgae. Journal of Applied Phycology 2019, 31, 2755–2770. [CrossRef]
- Solovchenko, A.; Khozin-Goldberg, I.; Selyakh, I.; Semenova, L.; Ismagulova, T.; Lukyanov, A.; Mamedov, I.; Vinogradova, E.; Karpova, O.; Konyukhov, I., et al. Phosphorus starvation and luxury uptake in green microalgae revisited. Algal Research 2019, 43, 101651. [CrossRef]
- Solovchenko, A.; Gorelova, O.; Karpova, O.; Selyakh, I.; Semenova, L.; Chivkunova, O.; Baulina, O.; Vinogradova, E.; Pugacheva, T.; Scherbakov, P. Phosphorus feast and famine in cyanobacteria: is luxury uptake of the nutrient just a consequence of acclimation to its shortage? Cells 2020, 9, 1933. [CrossRef]
- Voronkov, A.; Sinetova, M. Polyphosphate accumulation dynamics in a population of Synechocystis sp. PCC 6803 cells under phosphate overplus. Protoplasma 2019, 10.1007/s00709-019-01374-2. [CrossRef]
- McCarthy, L.; Abramchuk, I.; Wafy, G.; Denoncourt, A.; Lavallée-Adam, M.; Downey, M. Ddp1 cooperates with Ppx1 to counter a stress response initiated by nonvacuolar polyphosphate. mBio 2022, 10.1128/mbio.00390-22. [CrossRef]
- Gerasimaite, R.; Sharma, S.; Desfougeres, Y.; Schmidt, A.; Mayer, A. Coupled synthesis and translocation restrains polyphosphate to acidocalcisome-like vacuoles and prevents its toxicity. J Cell Sci 2014, 127, 5093–5104. [CrossRef]
- Goodenough, U.; Heiss, A.A.; Roth, R.; Rusch, J.; Lee, J.-H. Acidocalcisomes: ultrastructure, biogenesis, and distribution in microbial eukaryotes. Protist 2019, 170, 287–313. [CrossRef]
- Docampo, R.; de Souza, W.; Miranda, K.; Rohloff, P.; Moreno, S.N. Acidocalcisomes? Conserved from bacteria to man. Nature Reviews Microbiology 2005, 3, 251–261. [CrossRef]
- Docampo, R.; Huang, G. Acidocalcisomes of eukaryotes. Current opinion in cell biology 2016, 41, 66–72. [CrossRef]
- Ruiz, F.A.; Marchesini, N.; Seufferheld, M.; Govindjee; Docampo, R. The polyphosphate bodies of Chlamydomonas reinhardtii possess a proton-pumping pyrophosphatase and are similar to acidocalcisomes. Journal of Biological Chemistry 2001, 276, 46196–46203. [CrossRef]
- Nagel, L. Volutin. The Botanical Review 1948, 14, 174–184. [CrossRef]
- Lobakova, E.S.; Selyakh, I.O.; Semenova, L.R.; Scherbakov, P.N.; Fedorenko, T.A.; Chekanov, K.A.; Chivkunova, O.B.; Baulina, O.I.; Vasilieva, S.G.; Solovchenko, A.E. Hints for understanding microalgal phosphate-resilience from Micractinium simplicissimum IPPAS C-2056 (Trebouxiophyceae) isolated from a phosphorus-polluted site. Journal of Applied Phycology 2022, 34, 2409–2422. [CrossRef]
- Yagisawa, F.; Kuroiwa, H.; Fujiwara, T.; Kuroiwa, T. Intracellular structure of the unicellular red alga Cyanidioschyzon merolae in response to phosphate depletion and resupplementation. Cytologia 2016, 81, 341–347. [CrossRef]
- Shebanova, A.; Ismagulova, T.; Solovchenko, A.; Baulina, O.; Lobakova, E.; Ivanova, A.; Moiseenko, A.; Shaitan, K.; Polshakov, V.; Nedbal, L., et al. Versatility of the green microalga cell vacuole function as revealed by analytical transmission electron microscopy. Protoplasma 2017, 254, 1323–1340. [CrossRef]
- Reusch, R. Transmembrane ion transport by polyphosphate/poly-(R)-3-hydroxybutyrate complexes. Biochemistry Moscow 65, 280–295.
- Ota, S.; Yoshihara, M.; Yamazaki, T.; Takeshita, T.; Hirata, A.; Konomi, M.; Oshima, K.; Hattori, M.; Bisova, K.; Zachleder, V., et al. Deciphering the relationship among phosphate dynamics, electron-dense body and lipid accumulation in the green alga Parachlorella kessleri. Scienific Reports 2016, 6, 25731. [CrossRef]
- Wang, L.; Jia, X.; Zhang, Y.; Xu, L.; Menand, B.; Zhao, H.; Zeng, H.; Dolan, L.; Zhu, Y.; Yi, K. Loss of two families of SPX domain-containing proteins required for vacuolar polyphosphate accumulation coincides with the transition to phosphate storage in green plants. Molecular Plant 2021, 14, 838–846. [CrossRef]
- Zhu, J.; Loubéry, S.; Broger, L.; Zhang, Y.; Lorenzo-Orts, L.; Utz-Pugin, A.; Fernie, A.R.; Young-Tae, C.; Hothorn, M. A genetically validated approach for detecting inorganic polyphosphates in plants. The Plant Journal 2020, 102, 507–516. [CrossRef]
- Silva, V.M.; Putti, F.F.; White, P.J.; Dos Reis, A.R. Phytic acid accumulation in plants: Biosynthesis pathway regulation and role in human diet. Plant physiology and biochemistry 2021, 164, 132–146. [CrossRef]
- Lorenzo-Orts, L.; Couto, D.; Hothorn, M. Identity and functions of inorganic and inositol polyphosphates in plants. New Phytologist 2020, 225, 637–652. [CrossRef]
- Okazaki, K.; Hori, K.; Iwai, M.; Kurita, T.; Shimizu, S.; Nomura, S.; Saito, F.; Maeda, S.; Takami, A.; Yamamoto, T., et al. Knockout of an SPX-related gene for polyphosphate synthetase accelerates phosphate starvation responses in the oleaginous microalga Nannochloropsis oceanica. Journal of Experimental Botany 2025, 10.1093/jxb/eraf171, eraf171. [CrossRef]
- Whitehead, M.P.; Hooley, P.; Brown, M.R. Horizontal transfer of bacterial polyphosphate kinases to eukaryotes: implications for the ice age and land colonisation. BMC research notes 2013, 6, 221. [CrossRef]
- Sun, Q.-W.; Gao, Y.; Wang, J.; Fu, F.-x.; Yong, C.-W.; Li, S.-Q.; Huang, H.-L.; Chen, W.-Z.; Wang, X.-W.; Jiang, H.-B. Molecular mechanism of a coastal cyanobacterium Synechococcus sp. PCC 7002 adapting to changing phosphate concentrations. Marine Life Science & Technology 2024, 6, 562–575. [CrossRef]
- Srouji, J.R.; Xu, A.; Park, A.; Kirsch, J.F.; Brenner, S.E. The evolution of function within the Nudix homology clan. Proteins: Structure, Function, and Bioinformatics 2017, 85, 775–811. [CrossRef]
- Blaby, I.K.; Blaby-Haas, C.E.; Tourasse, N.; Hom, E.F.Y.; Lopez, D.; Aksoy, M.; Grossman, A.; Umen, J.; Dutcher, S.; Porter, M., et al. The Chlamydomonas genome project: a decade on. Trends in Plant Science 2014, 19, 672–680. [CrossRef]
- Grossman, A.R.; Aksoy, M. Algae in a phosphorus-limited landscape. In Annual Plant Reviews, Phosphorus Metabolism in Plants, Plaxton, W., Lambers, H., Eds. Wiley-Blackwell: 2015; Vol. 48, pp. 337–374.
- Plouviez, M.; Abyadeh, M.; Hasan, M.; Mirzaei, M.; Paulo, J.A.; Guieysse, B. The proteome of Chlamydomonas reinhardtii during phosphorus depletion and repletion. Algal Research 2023, 71. [CrossRef]
- Kumar Sharma, A.; Mühlroth, A.; Jouhet, J.; Maréchal, E.; Alipanah, L.; Kissen, R.; Brembu, T.; Bones, A.M.; Winge, P. The Myb-like transcription factor phosphorus starvation response (PtPSR) controls conditional P acquisition and remodelling in marine microalgae. New Phytologist 2020, 225, 2380–2395. [CrossRef]
- Aksoy, M.; Pootakham, W.; Grossman, A.R. Critical function of a Chlamydomonas reinhardtii putative polyphosphate polymerase subunit during nutrient deprivation. Plant Cell 2014, 26, 4214–4229. [CrossRef]
- Sanz-Luque, E.; Grossman, A.R. Chapter 4 - Phosphorus and sulfur uptake, assimilation, and deprivation responses. In The Chlamydomonas Sourcebook (Third Edition), Grossman, A.R., Wollman, F.-A., Eds. Academic Press: London, 2023; 10.1016/B978-0-12-821430-5.00006-7pp. 129–165.
- Desfougeres, Y.; Gerasimaite, R.U.; Jessen, H.J.; Mayer, A. Vtc5, a novel subunit of the vacuolar transporter chaperone complex, regulates polyphosphate synthesis and phosphate homeostasis in yeast. J Biol Chem 2016, 291, 22262–22275. [CrossRef]
- Ogawa, N.; DeRisi, J.; Brown, P.O. New components of a system for phosphate accumulation and polyphosphate metabolism in Saccharomyces cerevisiae revealed by genomic expression analysis. Molecular biology of the cell 2000, 11, 4309–4321. [CrossRef]
- Zhu, Y.; Huang, W.; Lee, S.S.K.; Xu, W. Crystal structure of a polyphosphate kinase and its implications for polyphosphate synthesis. EMBO reports 2005, 6, 681–687. [CrossRef]
- Tzeng, C.-M.; Kornberg, A. The multiple activities of polyphosphate kinase of Escherichia coli and their subunit structure determined by radiation target analysis. Journal of Biological Chemistry 2000, 275, 3977–3983. [CrossRef]
- Qi, J.; Shi, L.; Zhu, L.; Chen, Y.; Zhu, H.; Cheng, W.; Chen, A.F.; Fu, C. Functions, mechanisms, and therapeutic applications of the inositol pyrophosphates PP-InsP5 and InsP8 in mammalian cells. Journal of cardiovascular translational research 2024, 17, 197–215.
- Austin, S.; Mayer, A. Phosphate homeostasis − a vital metabolic equilibrium maintained through the INPHORS signaling pathway. Frontiers in Microbiology 2020, Volume 11 - 2020. [CrossRef]
- Shears, S.B. Inositol pyrophosphates: Why so many phosphates? Advances in Biological Regulation 2015, 57, 203–216. [CrossRef]
- Gu, C.; Nguyen, H.-N.; Hofer, A.; Jessen, H.J.; Dai, X.; Wang, H.; Shears, S.B. The significance of the bifunctional kinase/phosphatase activities of diphosphoinositol pentakisphosphate kinases (PPIP5Ks) for coupling inositol pyrophosphate cell signaling to cellular phosphate homeostasis. Journal of Biological Chemistry 2017, 292, 4544–4555. [CrossRef]
- Zhu, J.; Lau, K.; Puschmann, R.; Harmel, R.K.; Zhang, Y.; Pries, V.; Gaugler, P.; Broger, L.; Dutta, A.K.; Jessen, H.J., et al. Two bifunctional inositol pyrophosphate kinases/phosphatases control plant phosphate homeostasis. eLife 2019, 8, e43582. [CrossRef]
- Szijgyarto, Z.; Garedew, A.; Azevedo, C.; Saiardi, A. Influence of inositol pyrophosphates on cellular energy dynamics. Science 2011, 334, 802–805. [CrossRef]
- Wild, R.; Gerasimaite, R.; Jung, J.-Y.; Truffault, V.; Pavlovic, I.; Schmidt, A.; Saiardi, A.; Jessen, H.J.; Poirier, Y.; Hothorn, M., et al. Control of eukaryotic phosphate homeostasis by inositol polyphosphate sensor domains. Science 2016, 352, 986–990. [CrossRef]
- Zhou, J.; Hu, Q.; Xiao, X.; Yao, D.; Ge, S.; Ye, J.; Li, H.; Cai, R.; Liu, R.; Meng, F., et al. Mechanism of phosphate sensing and signaling revealed by rice SPX1-PHR2 complex structure. Nature Communications 2021, 12, 7040. [CrossRef]
- Li, X.; Gu, C.; Hostachy, S.; Sahu, S.; Wittwer, C.; Jessen, H.J.; Fiedler, D.; Wang, H.; Shears, S.B. Control of XPR1-dependent cellular phosphate efflux by InsP8 is an exemplar for functionally-exclusive inositol pyrophosphate signaling. Proceedings of the National Academy of Sciences 2020, 117, 3568–3574. [CrossRef]
- Rubio, V.; Linhares, F.; Solano, R.; Martín, A.C.; Iglesias, J.; Leyva, A.; Paz-Ares, J. A conserved MYB transcription factor involved in phosphate starvation signaling both in vascular plants and in unicellular algae. Genes & development 2001, 15, 2122–2133.
- Fiore, C.L.; Alexander, H.; Soule, M.C.K.; Kujawinski, E.B. A phosphate starvation response gene (psr1-like) is present and expressed in Micromonas pusilla and other marine algae. Aquatic Microbial Ecology 2021, 86, 29–46. [CrossRef]
- Paz-Ares, J.; Puga, M.I.; Rojas-Triana, M.; Martinez-Hevia, I.; Diaz, S.; Poza-Carrión, C.; Miñambres, M.; Leyva, A. Plant adaptation to low phosphorus availability: Core signaling, crosstalks, and applied implications. Molecular Plant 2022, 15, 104–124. [CrossRef]
- Dyhrman, S.; Chappell, P.; Haley, S.; Moffett, J.; Orchard, E.; Waterbury, J.; Webb, E. Phosphonate utilization by the globally important marine diazotroph Trichodesmium. Nature 2006, 439, 68. [CrossRef]
- Dyhrman, S.T.; Haley, S.T.; Birkeland, S.R.; Wurch, L.L.; Cipriano, M.J.; McArthur, A.G.J.A.; Microbiology, E. Long serial analysis of gene expression for gene discovery and transcriptome profiling in the widespread marine coccolithophore Emiliania huxleyi. 2006, 72, 252–260. [CrossRef]
- Alexander, H.; Jenkins, B.D.; Rynearson, T.A.; Saito, M.A.; Mercier, M.L.; Dyhrman, S.T.J.F.i.m. Identifying reference genes with stable expression from high throughput sequence data. Front Microbiol 2012, 3, 385. [CrossRef]
- Dyhrman, S.T.; Jenkins, B.D.; Rynearson, T.A.; Saito, M.A.; Mercier, M.L.; Alexander, H.; Whitney, L.P.; Drzewianowski, A.; Bulygin, V.V.; Bertrand, E.M., et al. The transcriptome and proteome of the diatom Thalassiosira pseudonana reveal a diverse phosphorus stress response. PLOS ONE 2012, 7, e33768. [CrossRef]
- Xiao, M.; Burford, M.A.; Wood, S.A.; Aubriot, L.; Ibelings, B.W.; Prentice, M.J.; Galvanese, E.F.; Harris, T.D.; Hamilton, D.P. Schindler’s legacy: from eutrophic lakes to the phosphorus utilization strategies of cyanobacteria. FEMS Microbiol Rev 2022, 46. [CrossRef]
- Hutchinson, G.E. The paradox of the plankton. The American Naturalist 1961, 95, 137–145. [CrossRef]
- Sanz-Luque, E.; Saroussi, S.; Huang, W.; Akkawi, N.; Grossman, A. Metabolic control of acclimation to nutrient deprivation dependent on polyphosphate synthesis. Science Advances 2020, 6, eabb5351. [CrossRef]
- de Mazancourt, C.; Schwartz, M.W. Starve a competitor: evolution of luxury consumption as a competitive strategy. Theoretical Ecology 2010, 5, 37–49. [CrossRef]
- Müller, W.E.; Schröder, H.C.; Wang, X. Inorganic polyphosphates as storage for and generator of metabolic energy in the extracellular matrix. Chemical reviews 2019, 119, 12337–12374. [CrossRef]
- Pavlov, E.; Aschar-Sobbi, R.; Campanella, M.; Turner, R.J.; Gómez-García, M.R.; Abramov, A.Y. Inorganic polyphosphate and energy metabolism in mammalian cells. Journal of Biological Chemistry 2010, 285, 9420–9428. [CrossRef]
- Sebesta, J.; Cantrell, M.; Schaedig, E.; Hou, H.J.M.; Pastore, C.; Chou, K.J.; Xiong, W.; Guarnieri, M.T.; Yu, J. Polyphosphate kinase deletion increases laboratory productivity in cyanobacteria. Front Plant Sci 2024, 15, 1342496. [CrossRef]
- Kampinga, H.H. Chaperoned by prebiotic inorganic polyphosphate molecules: an ancient transcription-independent mechanism to restore protein homeostasis. Molecular cell 2014, 53, 685–687. [CrossRef]
- Li, Q.; Fu, L.; Wang, Y.; Zhou, D.; Rittmann, B.E. Excessive phosphorus caused inhibition and cell damage during heterotrophic growth of Chlorella regularis. Bioresource Technology 2018, 268, 266–270. [CrossRef]
- Lobakova, E.; Gorelova, O.; Selyakh, I.; Semenova, L.; Scherbakov, P.; Vasilieva, S.; Zaytsev, P.; Shibzukhova, K.; Chivkunova, O.; Baulina, O. Failure of Micractinium simplicissimum phosphate resilience upon abrupt re-feeding of its phosphorus-starved cultures. International Journal of Molecular Sciences 2023, 24, 8484.
- Fu, L.; Li, Q.; Yan, G.; Zhou, D.; Crittenden, J.C. Hormesis effects of phosphorus on the viability of Chlorella regularis cells under nitrogen limitation. Biotechnol Biofuels 2019, 12, 121. [CrossRef]
- Vasilieva, S.; Lobakova, E.; Gorelova, O.; Baulina, O.; Scherbakov, P.; Chivkunova, O.; Semenova, L.; Selyakh, I.; Lukyanov, A.; Solovchenko, A. Photosynthetic and ultrastructural responses of the chlorophyte Lobosphaera to the stress caused by a high exogenic phosphate concentration. Photochemical & Photobiological Sciences 2022, 21, 2035–2051. [CrossRef]
- Haneklaus, S.; Bloem, H.; Schnug, E. Hungry plants—a short treatise on how to feed crops under stress. Agriculture 2018, 8. [CrossRef]
- Cordell, D.; White, S. Life’s bottleneck: implications of global phosphorus scarcity and pathways for a sustainable food system. Annual Review of Environment and Resources 2014, 39. [CrossRef]
- Daneshgar, S.; Callegari, A.; Capodaglio, A.; Vaccari, D. The potential phosphorus crisis: resource conservation and possible escape technologies: a review. Resources 2018, 7. [CrossRef]
- Bennett, E.; Elser, J. A broken biogeochemical cycle. Nature 2011, 478, 29–31. [CrossRef]
- Elser, J.J.; Call, D.F.; Deaver, J.A.; Duckworth, O.W.; Mayer, B.K.; McLamore, E.; Rittmann, B.; Mahmood, M.; Westerhoff, P. The phosphorus challenge: biotechnology approaches for a sustainable phosphorus system. Curr Opin Biotechnol 2024, 90, 103197. [CrossRef]
- Chen, M.; Graedel, T. A half-century of global phosphorus flows, stocks, production, consumption, recycling, and environmental impacts. Global Environmental Change 2016, 36, 139–152. [CrossRef]
- Cordell, D.; Rosemarin, A.; Schroder, J.J.; Smit, A.L. Towards global phosphorus security: a systems framework for phosphorus recovery and reuse options. Chemosphere 2011, 84, 747–758. [CrossRef]
- Solovchenko, A.; Verschoor, A.M.; Jablonowski, N.D.; Nedbal, L. Phosphorus from wastewater to crops: An alternative path involving microalgae. Biotechnology advances 2016, 34, 550–564. [CrossRef]
- Cakmak, E.K.; Hartl, M.; Kisser, J.; Cetecioglu, Z. Phosphorus mining from eutrophic marine environment towards a blue economy: the role of bio-based applications. Water Research 2022, 10.1016/j.watres.2022.118505. [CrossRef]
- de-Bashan, L.E.; Bashan, Y. Recent advances in removing phosphorus from wastewater and its future use as fertilizer (1997–2003). Water Research 2004, 38, 4222–4246. [CrossRef]
- Akuku, V.; Satognon, F. Sustainable phosphorus recovery from wastewater using microalgae: economic, environmental, and qgronomic implications for future phosphorus fertilizer solutions. Cleaner Waste Systems 2025, 10.1016/j.clwas.2025.100377. [CrossRef]
- Brown, N.; Shilton, A. Luxury uptake of phosphorus by microalgae in waste stabilisation ponds: current understanding and future direction. Reviews in Environmental Science and Bio/Technology 2014, 13, 321–328. [CrossRef]
- Wu, F.; Zhou, Y.; Beardall, J.; Raven, J.A.; Peng, B.; Xu, L.; Zhang, H.; Li, J.; Xia, J.; Jin, P. The dynamics of adaptive evolution in microalgae in a high-CO2 ocean. New Phytol 2024, 10.1111/nph.20323. [CrossRef]
- Amin, M.; Tahir, F.; Akbar, I.; Alessa, A.H.; Alsaigh, A.A.; Liu, C.-G.; Xin, F.; Chi, Z.; Syafiuddin, A.; Mehmood, M.A., et al. Towards environmental sustainability: employing adaptive laboratory evolution to develop elite algae strains for industrial and environmental applications. Current Pollution Reports 2025, 11. [CrossRef]
- Marchetto, F.; Conde, T.; Sliwinska, M.A.; Rewerski, B.; Lebiedzinska-Arciszewska, M.; Szymanski, J.; Wieckowski, M.R.; Matlakowska, R.; Domingues, M.R.; Kargul, J. Adaptive laboratory evolution of extremophilic red microalga Cyanidioschyzon merolae under high nickel stress enhances lipid production and alleviates oxidative damage. Bioresour Technol 2025, 434, 132826. [CrossRef]
- Kulakovskaya, T.V.; Vagabov, V.M.; Kulaev, I.S. Inorganic polyphosphate in industry, agriculture and medicine: Modern state and outlook. Process Biochemistry 2012, 47, 1–10. [CrossRef]
- Lorenz, B.; Leuck, J.; Köhl, D.; Müller, W.E.; Schröder, H.C. Anti-HIV-1 activity of inorganic polyphosphates. JAIDS Journal of Acquired Immune Deficiency Syndromes 1997, 14, 110–118.
- Feng, G.; Dong, S.; Huang, M.; Zeng, M.; Liu, Z.; Zhao, Y.; Wu, H. Biogenic polyphosphate nanoparticles from a marine cyanobacterium Synechococcus sp. PCC 7002: production, characterization, and anti-inflammatory properties in vitro. Marine Drugs 2018, 16. [CrossRef]
- Glass, D.J. Government regulation of the uses of genetically modified algae and other microorganisms in biofuel and bio-based chemical production. In Algal Biorefineries: Volume 2: Products and Refinery Design, Prokop, A., Bajpai, R.K., Zappi, M.E., Eds. Springer International Publishing: Cham, 2015; 10.1007/978-3-319-20200-6_2pp. 23–60.
- Vasilieva, S.; Petrova, E.; Lobakova, E.; Solovchenko, A.; Antal, T.; Gorelova, O. Effect of hydrogenase deficiency on accumulation of phosphorus-rich inclusions in Chlamydomonas reinhardtii. Russian Journal of Plant Physiology 2024, 71, 48. [CrossRef]
- Trebuch, L.M.; Sohier, J.; Altenburg, S.; Oyserman, B.O.; Pronk, M.; Janssen, M.; Vet, L.E.M.; Wijffels, R.H.; Fernandes, T.V. Enhancing phosphorus removal of photogranules by incorporating polyphosphate accumulating organisms. Water Research 2023, 10.1016/j.watres.2023.119748. [CrossRef]
- Kulaev, I.; Vagabov, I.; Kulakovskaya, T. The Biochemistry of Inorganic Polyphosphates, 2 ed.; John Wiley & Sons, Ltd.: Chichester, England, 2004.
- Miyachi, S.; Kanai, R.; Mihara, S.; Miyachi, S.; Aoki, S. Metabolic roles of inorganic polyphosphates in Chlorella cells. Biochimica et Biophysica Acta 1964, 93, 625–634. [CrossRef]
- Miyachi, S.; Miyachi, S. Modes of formation of phosphate compounds and their turnover in Chlorella cells during the process of life cycle as studied by the technique of synchronous culture. Plant and Cell Physiology 1961, 2, 415–424. [CrossRef]
- Miyachi, S.; Tamiya, H. Some observations on the phosphorus metabolism in growing Chlorella cells. Biochimica et Biophysica Acta 1961, 46, 200–202. [CrossRef]
- Miyachi, S.; Tamiya, H. Distribution and turnover of phosphate compounds in growing Chlorella cells. Plant and Cell Physiology 1961, 2, 405–414. [CrossRef]
- Abramson, J.; Adler, J.; Dunger, J.; Evans, R.; Green, T.; Pritzel, A.; Ronneberger, O.; Willmore, L.; Ballard, A.J.; Bambrick, J. Accurate structure prediction of biomolecular interactions with AlphaFold 3. Nature 2024, 630, 493–500. [CrossRef]
Figure 1.
The Pi taken up by the cell in excess of current metabolic demand is converted into polyphosphate with participation of VTC enzyme complex. VTC, vacuolar transport chaperone; DOP, dissolved organic phosphorus. Designations: PTA, PTB, PTC,—Pi transporters. Adapted from [37].
Figure 1.
The Pi taken up by the cell in excess of current metabolic demand is converted into polyphosphate with participation of VTC enzyme complex. VTC, vacuolar transport chaperone; DOP, dissolved organic phosphorus. Designations: PTA, PTB, PTC,—Pi transporters. Adapted from [37].
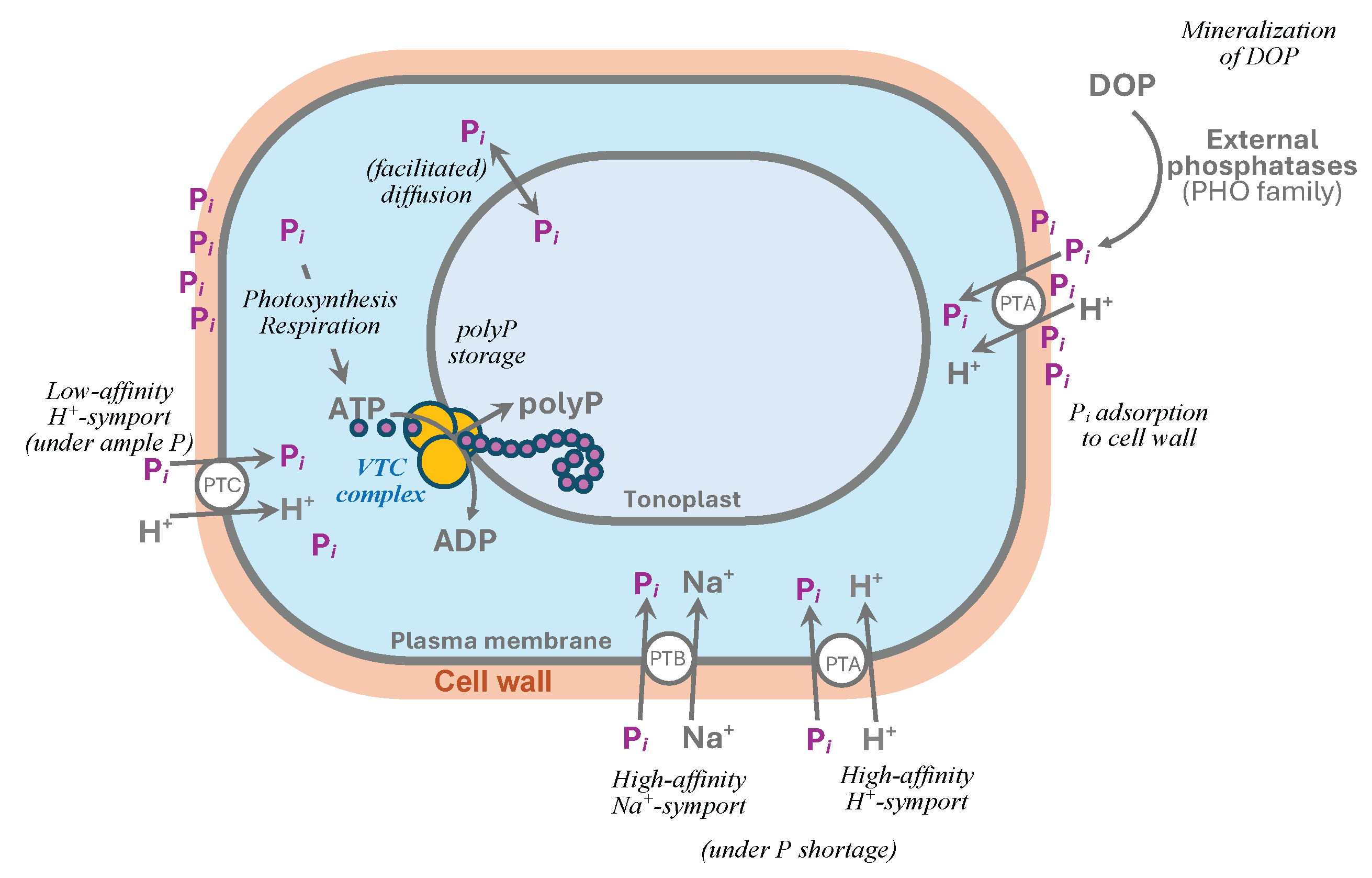
Figure 2.
The ultrastructure of Chlorella vulgaris CCALA 256 cells with the vacuolar polyphosphate granules from the stationary phase (9 days from re-supplementation of Pi to P-starving cells) culture: (a) survey micrograph view of the cells and (b) TEM image of the polyP granule in the vacuole and (c) its enlarged fragment together with (d) a hypothetical schematic representation of the coupled biosynthesis of the polyP chains and their translocation into the vacuole by the tonoplast-bound VTC-like enzyme complexes leading to the formation of the P-rich inclusions. The polyP particle chains are likely enclosed within poly-(R)-3-hydroxybutirate (PHB) and interlinked with bridge-like structures. The “multiwire cable” ultrastructural pattern likely arises from the simultaneous operation of many VTC-like complexes organized as raft(s) floating in the tonoplast. Ch, chloroplast; M, mitochondrion; P, polyP granule; Py, pyrenoid; S, starch grain; V, vacuoles with granules. In c, the longitudinally cut (asterisk) and crosscut (#) regions of the “multiwire cable” chains of the electron-dense particles are shown (see (d)). Scale bars, 1 μm (a) and 0.1 μm (b, c). (b, c). Reprinted from [38] with kind permission from Springer Nature.
Figure 2.
The ultrastructure of Chlorella vulgaris CCALA 256 cells with the vacuolar polyphosphate granules from the stationary phase (9 days from re-supplementation of Pi to P-starving cells) culture: (a) survey micrograph view of the cells and (b) TEM image of the polyP granule in the vacuole and (c) its enlarged fragment together with (d) a hypothetical schematic representation of the coupled biosynthesis of the polyP chains and their translocation into the vacuole by the tonoplast-bound VTC-like enzyme complexes leading to the formation of the P-rich inclusions. The polyP particle chains are likely enclosed within poly-(R)-3-hydroxybutirate (PHB) and interlinked with bridge-like structures. The “multiwire cable” ultrastructural pattern likely arises from the simultaneous operation of many VTC-like complexes organized as raft(s) floating in the tonoplast. Ch, chloroplast; M, mitochondrion; P, polyP granule; Py, pyrenoid; S, starch grain; V, vacuoles with granules. In c, the longitudinally cut (asterisk) and crosscut (#) regions of the “multiwire cable” chains of the electron-dense particles are shown (see (d)). Scale bars, 1 μm (a) and 0.1 μm (b, c). (b, c). Reprinted from [38] with kind permission from Springer Nature.
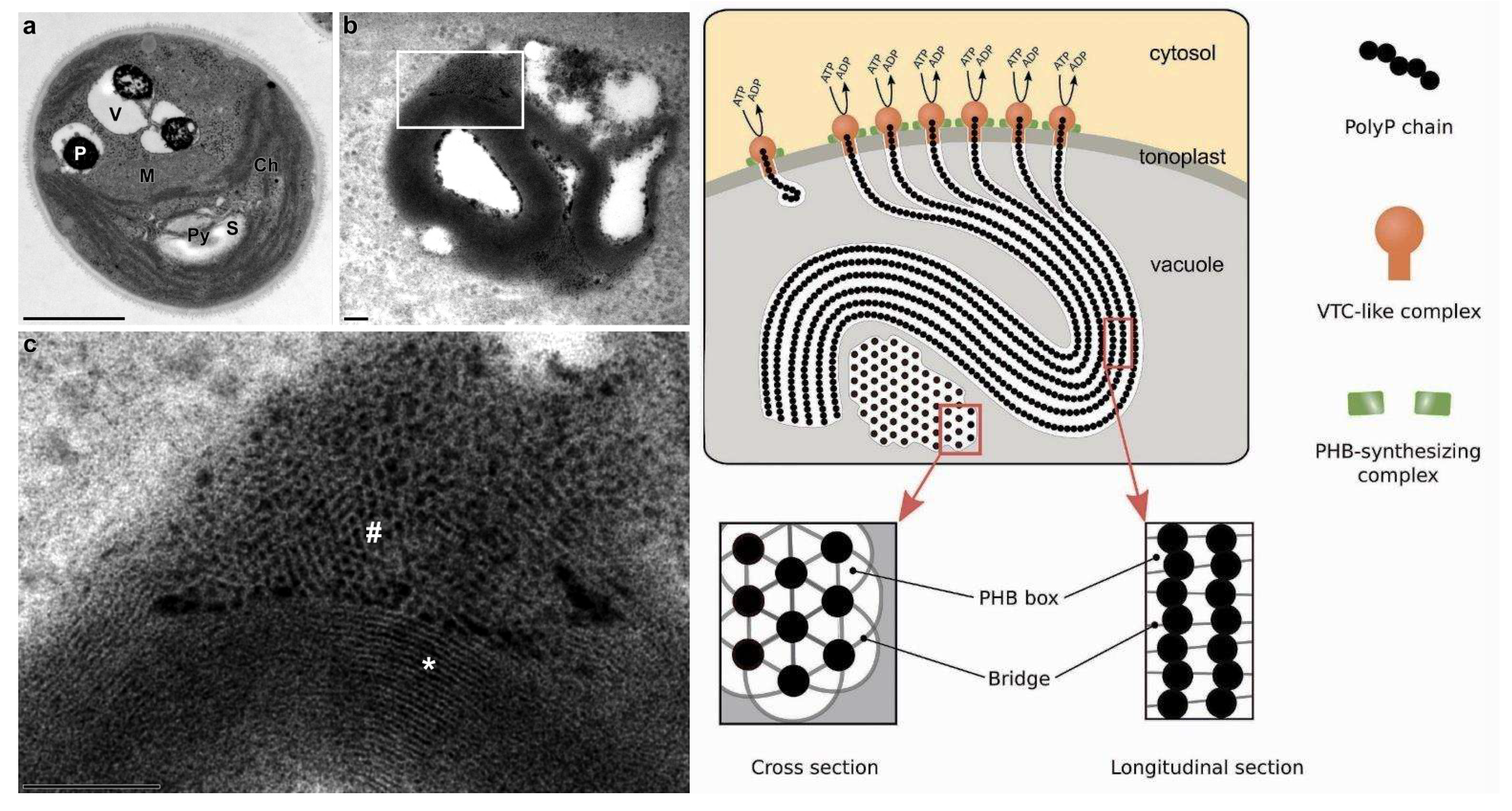
Figure 3.
Structure of the InsP6-activated VTC complex. (a–c) Overall structure of Vtc1/Vtc3/Vtc4R264A/R266A/E426N in complex with inositol phosphate, InsP6. Surface (Top) and cartoon (Bottom) representations are displayed in the same perspective. Vtc3 and Vtc4 are colored in blue and pink, respectively. The protomer A, B, and C of Vtc1 are colored separately in gradient green. Molecules of InsP6, phosphate and lipid are shown in stick representations. The exit tunnel of catalytic TTMVtc4 and the entrance channel of the trans-membrane region are marked with white arrows. Reprinted from [30] under Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/).
Figure 3.
Structure of the InsP6-activated VTC complex. (a–c) Overall structure of Vtc1/Vtc3/Vtc4R264A/R266A/E426N in complex with inositol phosphate, InsP6. Surface (Top) and cartoon (Bottom) representations are displayed in the same perspective. Vtc3 and Vtc4 are colored in blue and pink, respectively. The protomer A, B, and C of Vtc1 are colored separately in gradient green. Molecules of InsP6, phosphate and lipid are shown in stick representations. The exit tunnel of catalytic TTMVtc4 and the entrance channel of the trans-membrane region are marked with white arrows. Reprinted from [30] under Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/).
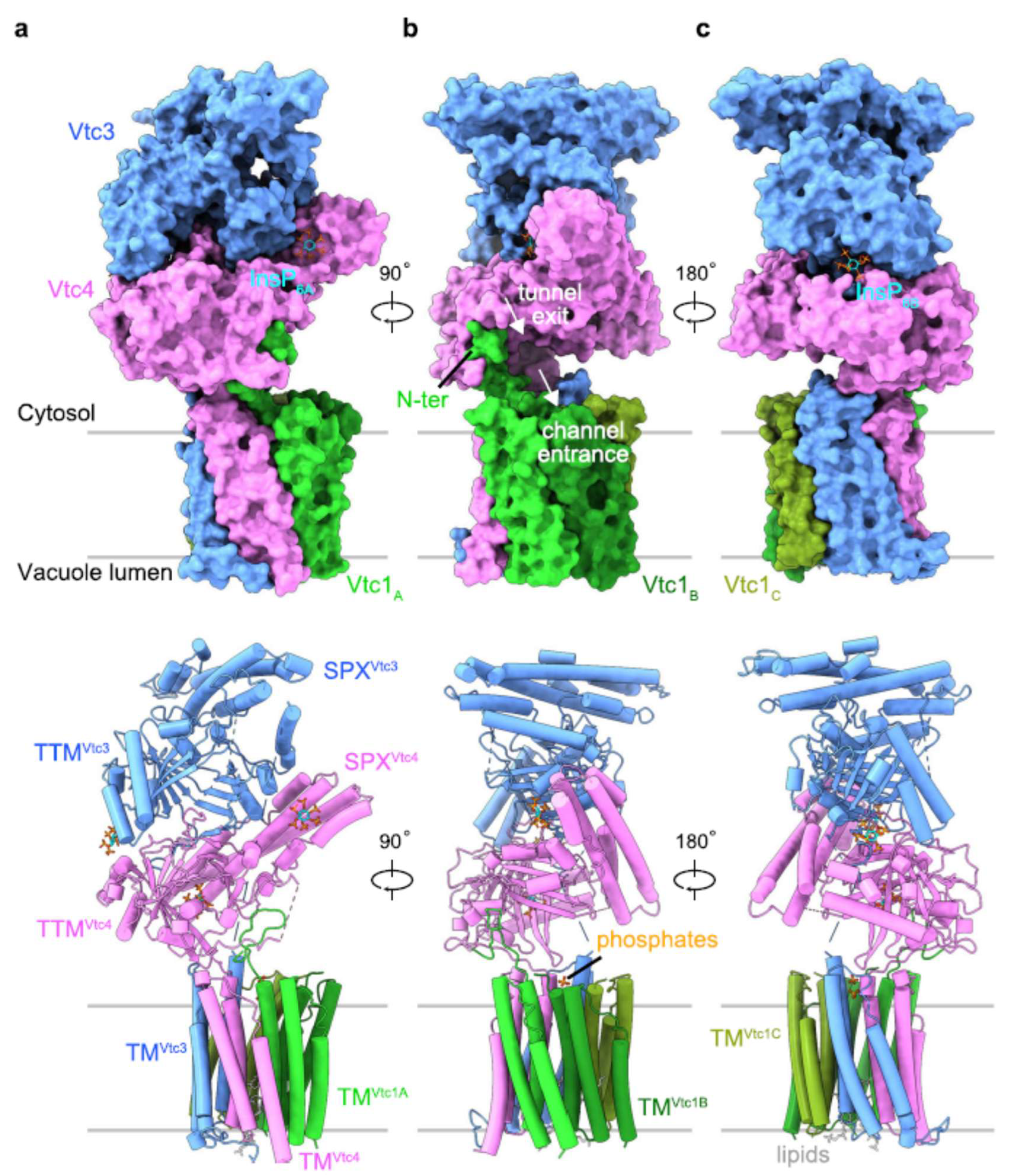

Table 1.
The subunit composition of VTC complex (according to [30]).
Table 1.
The subunit composition of VTC complex (according to [30]).
| Subunit | Domains harbored [PFAM accession] | Function |
|---|---|---|
| Vtc1 | Transmembrane domain, composed by three protomers (VTC1A, VTC1B, VTC1C) | Formation of the transmembrane channel guiding the nascent PolyP chain into the vacuole lumen |
| Vtc2 | Auxiliary subunit; catalytically inactive1 | |
| Vtc3 | N-terminal SPX domain [PF03105] | Auxiliary subunit; not essential for VTC regulation1 |
| TM1 domain | Discharging of polyP chain | |
| Vtc4 | Catalytic tunnel domain (triphosphate tunnel metalloenzyme, TTM domain) [PF09359] | Polymerization of polyP |
| N-terminal SPX domain [PF03105] | Regulation (Pi level sensing via PP-InsP) | |
| Transmembrane domain [PF02656] | Anchoring the whole complex to the vacuolar membrane and enabling translocation of the formed polyP chain into the vacuole | |
| Vtc5 | SPX domain [PF03105] | Accessory subunit for activation of the VTC complex. The only protein acting directly on the VTC complex to stimulate polyP production2 |
1 Homologous to VTC4. 2 Presents only in yeast.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



