Submitted:
15 August 2025
Posted:
18 August 2025
You are already at the latest version
Abstract
Mercury exposure has been linked to male infertility. Given that mercury chloride (HgCl2) may promote an oxido-inflammatory milieu associated with pathophysiological derangements, it is hypothesised that thymoquinone (TQ), an anti oxido-inflammatory agent, may mitigate the gradual harmful effects of mercury exposure on rat testes, epididymis, and hypothalamus, as these organs are vital to reproductive function. To test this hypothesis, 40 male Wistar rats were randomised into five groups (n=8). After a 7-day acclimation, treatments were dispensed for 28 days as follows: Group I: control (distilled water only); Group II: HgCl2 only (20 µg/mL); Group III: TQ only (2.5 mg/kg); Group IV: HgCl2 + TQ (20 µg/mL + 2.5 mg/kg); and Group V: HgCl2 + TQ (20 µg/mL + 5 mg/kg). Co-treatment with TQ preserved the body and organ weight of the HgCl2-exposed animals. However, TQ did not reduce HgCl2-induced dysfunction in sperm function and morphology. Co-treatment with TQ significantly (p< 0.05) increased the serum levels of follicle-stimulating hormone (FSH), luteinising hormone (LH), and testosterone while decreasing the prolactin level. TQ administration also increased (p< 0.05) the activity of testicular enzymes alkaline phosphatase (ALP), acid phosphatase (ACP), lactate dehydrogenase (LDH), and glucose-6-phosphate dehydrogenase (G6PD), decreased by HgCl2. TQ administration significantly (p< 0.05) restored HgCl2-induced decreases in superoxide dismutase (SOD), catalase (CAT), glutathione peroxidase (GPx), glutathione-s-transferase (GST), glutathione (GSH), and total sulfhydryl groups (TSH) levels in the testes, epididymis, and hypothalamus of experimental rats. Further, TQ reduced HgCl2-mediated increases in reactive oxygen and nitrogen species (RONS), lipid peroxidation (LPO), protein carbonyl (PC), and xanthine oxidase (XO) activity. Furthermore, levels of inflammatory biomarkers including nitric oxide (NO), tumor necrotic factor alpha (TNF-), interleukin-1 beta (IL-1), and myeloperoxidase (MPO) were significantly (p< 0.05) decreased in the co-treated groups, with a higher dose of TQ (5.0 mg/kg) showing a more pronounced protective effect. Additionally, TQ co-administration increased Bax and decreased Bcl-2 and P53 protein levels (p< 0.05), protecting rats' testes, epididymis, and hypothalamus from HgCl2-induced apoptosis. Molecular docking simulation studies showed that TQ may interact with PPAR-α and PPAR- to suppress NF-kB-mediated pro-inflammatory sequela as well as activate Nrf-2-mediated antioxidant defence system. These predicted biological effects of TQ resonate with the findings from the in vivo studies. Therefore, supplementation with TQ may help reduce chemical-induced toxicities, including HgCl2‘s reproductive toxicity.
Keywords:
thymoquinone
; mercury chloride
; reproductive toxicity
; antioxidant
; anti-inflammatory
; apoptosis
1. Introduction
Mercury (Hg) is recognised globally as an environmental toxicant. Humans can be exposed to mercury in various forms: elemental Hg via inhalation, inorganic HgCl2 through occupational or dental exposure, and organic methylmercury from seafood consumption. While HgCl2 does not cross the blood-brain barrier, it accumulates in reproductive organs, causing toxicity; it disrupts testicular architecture and inhibits spermatogenesis in rats (Martinez et al., 2017). Another study confirmed Hg-induced decreases in sperm production, increased DNA damage, and higher levels of reactive oxygen species (ROS) and lipid peroxidation in rat reproductive tissues (Jahan et al., 2019). Similarly, studies revealed that HgCl2 increased malondialdehyde levels (Badary et al., 2021), the expressions of tumour necrosis factor-α and cyclooxygenase-2 (Kandemir et al., 2020), and morphological changes in the reproductive tissues of male rats, including spermatogonium degeneration and tubular wall thinning of the testes (Simsek et al., 2023). Martinez et al. (2017) emphasised that even at low doses, mercury impaired sperm membrane integrity, disrupted germinal epithelium, and increased ROS levels in male rats. Rats exposed to HgCl2 showed significantly reduced testosterone levels and nitric oxide production (Albasher et al., 2020). Additionally, the cellular antioxidant defence, represented by glutathione and enzymes like catalase, superoxide dismutase, and glutathione peroxidase, is significantly compromised upon treatment of experimental animals with HgCl2 (Bhai et al., 2024). In addition, mercury exposure also disturbed cellular apoptotic balance (Almeer et al., 2020). For instance, apoptotic markers such as Bax were upregulated, indicating cell death pathways were activated due to HgCl2 exposure. Given that experimental animals' blood mercury levels were analogous to those seen in humans' post-exposure (Balali-Mood et al., 2021), investigating protective agents against mercury toxicity becomes imperative.
Thymoquinone (TQ) is a bioactive compound from black seed oil (Nigella sativa) (Badary et al., 2021). Studies show TQ can mitigate reproductive toxicities in rats and improve reproductive parameters (Maryamneghari et al., 2021). It decreases sperm DNA fragmentation (p53), modulates apoptosis-related genes, increasing Bax while reducing Bcl-2 (Asaduzzaman Khan et al., 2017). TQ also counteracts lead acetate-induced reductions in sperm quality, testicular antioxidants, and reproductive hormone levels (Al-Sayyed et al., 2014; Hassan et al., 2019), highlighting its protective effects against a heavy metal (Mizuno & Fukuhara, 2022; Owumi et al., 2025). In addition, it modulates pathways crucial for cell survival and inflammation by downregulating TNF-α and NF-κB, and improving testicular histoarchitecture (Algaidi et al., 2022). Given TQ's multi-faceted benefits, this study explored its effects on HgCl2-induced reproductive toxicities in rats, using in vivo experiments and molecular docking studies. Our findings indicate that TQ supplementation reduces HgCl2's adverse effects on sperm function, prevents oxido-inflammatory pathophysiology, and improves cellular histoarchitecture. Molecular docking simulations reveal TQ's interaction with PPAR-α/β activates Nrf-2 and modulates NF-kB, promoting its beneficial antioxidant and anti-inflammatory effects.
2. Methods
2.1. Animals
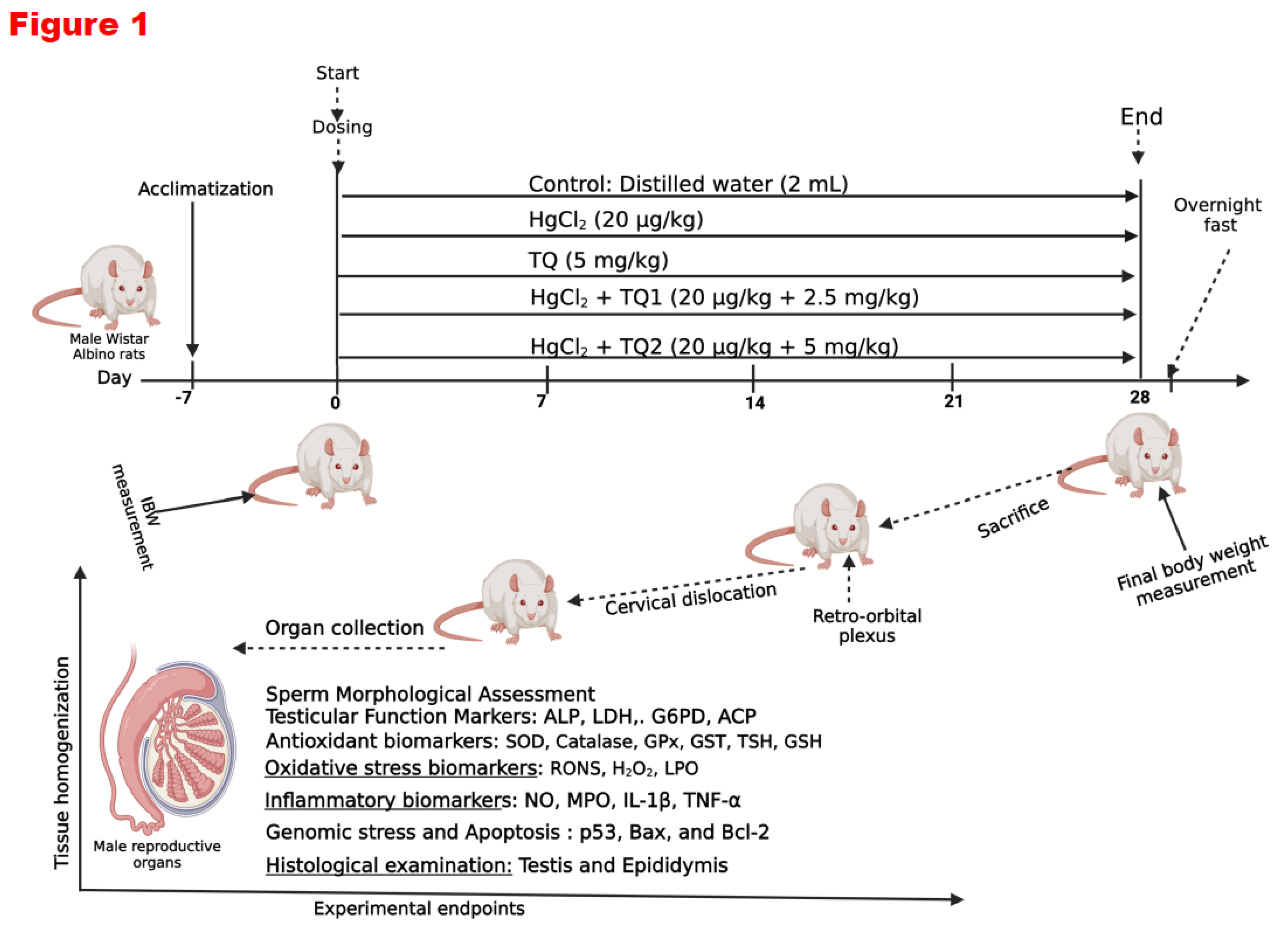
For the investigation, a group of forty male albino Wistar rats, with an average weight of 157 g, was obtained from the University of Ibadan. They were housed in standard cages at the animal facility of the Biochemistry department, University of Ibadan. Before the study, the rats were acclimated to the study environment, provided with a regular rat diet, and given unrestricted access to water. During this adaptation period, the rats were maintained under consistent conditions, including a temperature of 22 ± 2°C, a 12-hour light-dark cycle, and continuous access to food and water. All procedures adhered to the guidelines approved by the University of Ibadan Animal Care and Use Research Ethics Committee (UI-ACUREC), with approval number UI-ACUREC/057-1222/11.
2.2. Chemicals
All chemicals and reagents used in this study were of the highest quality and were obtained commercially from accredited distributors. See Table 1 for a list of chemicals used in this study.
2.3. Experimental Protocol
Forty male Wistar rats were divided into five groups, each consisting of eight animals.
Group 1: Control cohort (allowed free access to distilled water daily).
Group 2: HgCl2 only cohort: (20 µg/L HgCl2 dissolved in the drinking water).
Group 3: TQ only cohort: (TQ 5 mg/kg bw per os /day).
Group 4: HgCl2 + TQ (Low dose TQ) cohort: (20 µg/L HgCl2 dissolved in drinking water + 2.5 mg/kg bw of TQ per os /day).
Group 5: HgCl2 + TQ treated rats (High dose) cohort (20 µg/L of HgCl2 dissolved in drinking water + 5 mg/kg bw of TQ per os /day).
The doses were chosen based on previous studies from the scientific literature (Boujbiha et al., 2009; Moradi Maryamneghari et al., 2021). The substances were administered to non-fasted rats in the morning (between 09:00 and 10:00 h). The first day the animals were treated was considered experimental Day 0. At the end of the 4th week (28 days) of treatment, all animals were sacrificed and dissected. The testis, epididymis and hypothalamus tissues were quickly removed for biochemical and histological examinations.
Figure 1.
Experimental design on the effect of Thymoquinone on mercuric chloride-treated male Wistar Albino rats for 28 days. Created with https://app.biorender.com/ by Uche Arunsi.
Figure 1.
Experimental design on the effect of Thymoquinone on mercuric chloride-treated male Wistar Albino rats for 28 days. Created with https://app.biorender.com/ by Uche Arunsi.
2.4. Study Conclusion and Euthanasia
Twenty-four hours following the final administration, the rats' final body weights were recorded, and blood samples were collected from the retro-orbital venous plexus using plain tubes before euthanasia through cervical dislocation after carbon dioxide asphyxiation. The animals were carefully restrained, and a sterile microcapillary tube was gently inserted into the retro-orbital venous plexus behind the eye. Blood was cautiously aspirated into labelled plain sample bottles. After removing the microcapillary tube, the collected blood was allowed to clot. Serum samples were obtained by centrifuging the clotted blood at 3000 g for 10 minutes. Subsequently, the serum samples were frozen at −20 °C until hormone assays were performed using ELISA diagnostic kits (Amersham, UK). The hypothalamus, testes, and epididymis were promptly excised, weighed, and processed for biochemical and histological analyses. The organo-body weight indices (OBWI) of the hypothalamus, testes, and epididymis were determined using the formula OBWI = 100 × organ weight (g)/body weight (g). The tissues of interest were dissected and washed in sodium phosphate buffer (pH 7.2, 4℃). Post-washing, samples were stored at −80 °C until analysis. The tissues were homogenised using a Teflon homogeniser (Heidolph Silent Crusher M), and the homogenates were subsequently centrifuged. The protein content of the supernatant was determined by the method of Lowry et al. (1951), utilising bovine serum albumin as the standard.
2.5. Assessment of Sperm Progressive Motility
Sperm progressive motility was assessed using the method described by Zemjanis (Zemjanis et al., 1970). The cauda epididymis was cut with surgical blades, releasing sperm onto a sterile glass slide. The sperm was then diluted with a pre-warmed 2.9% sodium citrate dehydrate solution at 37 °C, mixed carefully, and covered with a coverslip (24 × 24 mm). Sperm motility was evaluated by examining at least ten microscopic fields under a phase contrast microscope at 200× magnification. The sperm in the same field were categorised based on their motility as progressive, non-progressive, or immotile. The result was expressed as a percentage of sperm progressive motility.
2.6. Assessment of Epididymal Sperm Count
The epididymal sperm count method follows WHO guidelines (1995). The cauda epididymis is crushed in saline, filtered through nylon mesh, and a 5 μL sample of the suspension is mixed with 95 μL of diluent. After placing 10 μL of the diluted sperm on a hemocytometer and letting it sediment for 5 minutes, sperm cells are counted using an improved Neubauer chamber (Deep 1/10 m; LABART, Munich, Germany) under a light microscope at 400x magnification.
2.7. Assessment of Sperm Morphological Abnormalities and Viability Assay
Sperm viability and morphological abnormalities were evaluated using Wells and Awa's method (1970). Sperm smears were prepared on clean slides. Viability was assessed with 1% eosin and 5% nigrosine in a 3% sodium citrate solution. At least 400 sperm cells per rat were examined for morphological abnormalities by staining with 0.2 g of eosin and 0.6 g fast green, dissolved in distilled water and ethanol (2:1). The percentages of head, mid-piece, and tail abnormalities were recorded for both control and treated rats.
2.8. Assessment of Reproductive Hormone Levels
Serum concentrations of luteinizing hormone (LH), follicle-stimulating Hormone (FSH), prolactin, and testosterone were quantified utilising ELISA plates procured from Elabscience Biotechnology (Wuhan, China), adhering to the manufacturer's protocols. The detection limits of the hormonal assays were established at 0.54 ng/ml for LH, 0.28 ng/ml for FSH, 0.39 ng/ml for prolactin, and 0.58 ng/ml for testosterone. All hormone measurements were performed concurrently to minimise inter-assay variability, with minimal intra-assay coefficients of variation recorded as 2.9%, 3.3%, 2.4%, and 3.8% for FSH, LH, prolactin, and testosterone, respectively.
2.9. Assessment of Testicular Enzyme Function
Acid and alkaline phosphatase (ACP and ALP) activities in the testicular supernatant were assessed using protocols involving p-nitrophenyl-phosphate hydrolysis under acidic (Vanha-Perttula & Nikkanen, 1973) and alkaline (Malymy & Horecker, 1966) conditions. Testicular glucose-6-phosphate dehydrogenase (G6PD) activity was determined using NADP+ and glucose-6-phosphate as substrates, following a modified method based on Wolf et al., 1987. Lactate dehydrogenase-X (LDH-X) activity was evaluated according to Vassault's protocol (Owumi, Otunla, et al., 2022; Vassault, 1983), involving the inter-conversion of pyruvate and lactate.
2.10. Evaluation of Biomarkers of Testes, Epididymis and Hypothalamus Antioxidant Status
The testes, epididymis, and hypothalamus samples from the experimental rats were homogenised in phosphate buffer (0.05 M; pH 7.4). The tissue homogenates were then centrifuged (12,000 × g; 15 min; 4°C) to obtain a clear supernatant, which was collected into appropriately labelled vials for the assessment of oxidative stress, inflammation, and apoptosis. The protein concentrations in the testes, epididymis, and hypothalamus were analysed using Lowry's method (Lowry et al., 1951). The activity of superoxide dismutase (SOD) was measured using the procedure described by Misra and Fridovich (Misra & Fridovich, 1972), while the activity of the catalase (CAT) enzyme was determined using Clairborne's method (Clairborne, 1995). The protocols outlined by Habig (Habig et al., 1974) and Rotruck (Rotruck et al., 1973) were employed to assess the enzyme activities of glutathione-S-transferase (GST) and glutathione peroxidase (GPx). Reduced glutathione (GSH) levels were measured using the methods of Beutler (Beutler et al., 1963), and total sulfhydryl groups (TSH) levels were determined via the Jollow et al. method (Jollow et al., 1974). Additionally, biomarkers of cellular response to oxidative stress in the testes, epididymis, and hypothalamus of experimental animals, specifically NRF-2, HO-1, along with TRX levels and TRX-R activity, were assayed using enzyme-linked immunosorbent assay (ELISA) kits specific for rats, following the manufacturer’s protocol as previously reported (Owumi et al., 2023; Owumi et al., 2022).
2.11. Evaluation of RONS & LPO levels and XO activity in the Testes, Epididymis and Hypothalamus of Rats
An established method based on the RONS-dependent oxidation of 2' 7'-dichlorodihydrofluorescein diacetate (DCFH-DA) to dichlorofluorescein (DCF) was used to assess the RONS production in the testes, epididymis and hypothalamus (Perez-Severiano et al., 2004). Lipid peroxidation was determined by measuring the formation of thiobarbituric acid reactive substances (TBARS) present in the test sample according to the method of Okhawa (Ohkawa, 1979). Malondialdehyde (Badary et al.), formed when fatty acids are peroxidised, combines with the chromogenic agent 2-thiobarbituric acid in an acidic condition to form a pink-coloured complex with a maximum absorbance at 532 nm that may be extracted using an organic solvent like butanol. The result is presented as the amount of free MDA generated, since MDA is frequently used to calibrate this test. A sample was prepared using 40 μL of the testes, epididymis and hypothalamus and was combined with 50 μL of 30% TCA in 160 mL of Tris-KCl buffer. Then, 50 L of thiobarbituric acid (TBA) at 0.75% was added and incubated for 45 min at 80 °C. The mixture was centrifuged at 3000 g for 10 minutes after being cooled to 25 °C. A microplate reader was used to measure the absorbance of 200 μL of the clear supernatant compared to a reference blank of distilled water at 532 nm. The method of Bergmeyer et al. (Bergmeyer et al., 1974) was used to assess the activity of xanthine oxidase (XO) in the testes, epididymis and hypothalamus. As uric acid is created from xanthine by the enzyme XO, the assay depends on the measurement of uric acid absorbance at 290 nm. 8 µL of the sample, 150 µl of phosphate buffer, and 80 µL of xanthine solution were pipetted into a microplate. After mixing, 290 nm absorbance measurements were made every minute for three minutes and read against a blank, which was created by substituting 8 µL of the sample with distilled water.
2.12. Evaluation of Pro-Inflammatory Markers in the Testes, Epididymis and Hypothalamus of Rats
Nitric oxide (NO) levels in the testes, epididymis and hypothalamus samples were assayed following Green’s method (Green et al., 1982). Myeloperoxidase (MPO), a biomarker of polymorphonuclear leukocyte accumulation, was measured using a modified version of the Trush method that was previously published (Trush et al., 1994). MPO oxidises o-dianisidine in the presence of H2O2 to produce a brown-colored molecule with a 470 nm absorbance. Additionally, testicular, epididymal and hypothalamic IL-1β were assessed using SpectraMax Multimodal plate reader (Molecular Devices, CA, USA) and commercially available ELISA Kits, as the manufacturer's guide instructed.
2.13. Evaluation of Apoptosis Biomarkers
Using ELISA Kits from Elabscience (Beijing, China) and following the instructions in the manufacturer's manual, the activity and concentrations of testicular, epididymal and hypothalamic Bcl-2 Associated X Protein (Bax), B-cell Lymphoma 2 (Bcl-2), and protein 53 (P53) were determined. Seven microplate wells were filled with 100 μL of the standard working solution, and the remaining wells were filled with 100 μL of testes, epididymis and hypothalamus homogenates. The plate was sealed with the kit's sealer and left in a water bath at 37 °C for 90 minutes. Following the incubation period, each well's liquid was drained without being washed, and 100μl of biotinylated detection Ab working solution was added immediately. The plate was then gently mixed, covered with a plate sealer, and incubated for a further hour at 37 °C. Then, 350 μL of wash buffer was added, the solution from each well was decanted, and the wells were allowed to soak for 1-2 minutes before being decanted and dried against clean absorbent paper. Following the washing procedure, 90μl of substrate reagent was added to each well in the dark, the plate was sealed, and the mixture was then incubated for 15 min at 37 °C. The M384 SpectraMaxTM Multimodal plate reader (Molecular Devices, San Jose, CA, USA) was then used to measure the OD value at 450 nm immediately after the reaction was stopped by adding 50 µL of stop solution to each well. The amount of Bax, Bcl-2 and P53 in the samples directly relates to how intense the colour developed.
2.14. Histopathological Examination of the Testes, Epididymis and Hypothalamus
Microscopic examination of the assessed testes, epididymis and hypothalamus sections was performed following the method of Bancroft & Gamble (Bancroft & Gamble, 2008). Properly removed and treated with 10% formalin solution were the testes, epididymis and hypothalamus. Following the dehydration processes, the sections were paraffin-embedded. The tissues were divided into sections (4-5 μm slices) using a microtome, then transferred to charged slides and stained using the conventional hematoxylin and eosin stain. A Carl Zeiss Axio light microscope was used to examine the coded tissue slides. A Zeiss Axiocam 512 camera affixed to the microscope was utilised by a pathologist unfamiliar with the various treatment groups from which the slides were taken to record representative images during the assessment.
2.15. Molecular Docking Method
Molecular docking was conducted to assess the interaction between TQ and PPAR-α or PPAR δ/β. The molecular docking scores, reflecting the binding affinity between the ligand and the proteins, were quantified as binding constants (Kd). To achieve this, the 3-D structures of TQ were sourced from PubChem: https://pubchem.ncbi.nlm.nih.gov, while the structures of the PPAR-α (PDB: 1I7G) and PPAR- δ/β (PDB: 3D5F) were acquired from the Protein Database (PDB): https://www.rcsb.org/. Protein preparation was done using UCSF ChimeraX (Meng et al., 2023). Additionally, the ligand (TQ) from the synthesised receptors was loaded into PyRx, and Open Babel was employed for energy minimisation. Following this, molecular docking was conducted using grid boxes designed around the position of the co-crystallised ligand. The docking modes that exhibited the lowest Gibbs free energy were then visualised with PyMol. The binding constant (Kd) was estimated from the general information ΔG = -RTInKd, where R: Gas constant = 0.001987kcal/mol/K, T: Temperature in Kelvin = 298K, G: Gibbs free energy.
2.16. Statistical Analysis
The study's data were analysed using GraphPad Prism version 9.3.1 for Windows (www.graphpad.com; GraphPad, CA, USA). One-way analysis of variance (ANOVA) and a post-hoc test (Bonferroni) were used to compare the means of a group with the mean of another. p<0.05 was selected for statistically significant differences. The data are expressed as the mean ± SD of the replicates.
3. Results
3.1. TQ Conserved the Body Weight of Animals Treated with HgCl2
Table 2 depicts TQ's body weight and organ weight-conserving ability against HgCl2-treated experimental rats. HgCl2 exposure resulted in decreased body weight gain and alterations in the organ weight and relative weights of testes, epididymis, and hypothalamus compared to the control group. However, treatment with low and high doses of TQ (2.5 and 5.0 mg/kg) mitigated these effects induced by HgCl2 exposure, improving body weight gain and restoring the relative organ weights.
3.2. Effect of TQ on Sperm Functional Parameters and Morphological Abnormalities in HgCl2-Treated Rats
Table 3 indicates the effect of TQ on sperm functional parameters and morphological abnormalities in HgCl2-treated rats. Compared to the control, HgCl2 exposure significantly reduced sperm motility. Interestingly, co-treatment with 2.5 and 5 mg/kg TQ failed to reverse these changes. Administration of rats with HgCl2 significantly decreased the sperm count from 132.4 million/mL in the control group to 117 million/mL in animals that received HgCl2 alone. At the same time, there were no changes in sperm volume across all groups. Furthermore, exposure of animals to HgCl2 induced morphological changes to the sperm, which were not reversed upon co-treatment with TQ.
3.3. TQ Improved HgCl2-Induced Alteration in Reproductive Hormones in the Serum of Treated Rats
The impact of TQ co-treatment on the serum level of reproductive hormones in HgCl2-treated experimental rats is shown in Figure 2. Upon exposure to HgCl2 alone, there was a decrease in the serum levels of LH, FSH, and testosterone by 10.60%, 9.91%, and 84.69%, respectively, compared to the control group. Conversely, the serum prolactin level increased by 62.67% with HgCl2 treatment relative to the control. However, co-treatment with TQ (2.5 mg/kg) resulted in 43.81%, 32.11%, and 30.36% increases in LH, FSH, and testosterone serum levels, respectively. Moreover, a decrease in prolactin levels of 29.00% was observed in the TQ co-administered group (2.5 mg/kg). Furthermore, when compared to animals that received HgCl2 alone, 5.0 mg/kg of TQ co-treatment induced even more pronounced changes, as seen in 80.86%, 41.78%, and 515.08% increases in serum levels of LH, FSH, and testosterone, respectively, and a 33.43% decrease in prolactin.
3.4. TQ Co-Administration Increased the Activities of Testicular Enzymes in Experimental Animals
Figure 3 displays the result of TQ co-administration on the activities of testicular enzymes in experimental animals exposed to HgCl2. Exposure of animals to HgCl2 alone significantly decreased the serum activities of ALP, ACP, LDH, and G6PD by 87.53%, 87.53%, 85.85%, and 77.26%, respectively, compared to the control group. However, TQ administration increased the dose-dependent activities of testicular enzymes. Co-treatment with 2.5 mg/kg TQ significantly increased ALP and ACP serum activities by 233.33% each, LDH by 343.27%, and G6PD by 110.47% relative to the HgCl2-alone administered group. Furthermore, a higher dose of TQ co-treatment (5 mg/kg) led to even more significant increases in serum enzyme activities: ALP and ACP each by 1054.96%, LDH by 413.78%, and G6PD by 188.74% when compared to the HgCl2 alone group.
3.5. TQ Co-Treatment Restored the Antioxidant Status of Rats Treated with HgCl2
Figure 4, Figure 5 and Figure 6 depict the effect of TQ co-treatment on the antioxidant status of rats treated with HgCl2. In the testes of rats treated with HgCl2, a marked (p<0.05) decline was observed in the activities of CAT (52.28%), SOD (57.77%), GST (41.67%), GPx (63.39%), along with the levels of GSH (61.68%) and TSH (54.10%). Remarkably, co-treatment with TQ significantly (p<0.05) elevated these parameters, particularly evident at the higher dose (5.0 mg/kg) of TQ, where SOD, GST, and GPx increased by 108.62%, 45.84%, and 112.34% respectively, compared to the HgCl2 group. Furthermore, in the epididymis, HgCl2 treatment substantially reduced the activities of CAT (85.13%), SOD (42.91%), GST (91.50%), GPx (79.56%), along with the levels of GSH (66.43%) and TSH (66.79%). Contrastingly, co-treatment with TQ significantly improved these parameters. Moreover, treatment of experimental animals with HgCl2 significantly downgraded the hypothalamic activities of CAT by 77.45%, SOD by 58.98%, GST by 68.05%, and GPx by 50.72%. The endogenous levels of GSH were equally reduced by 84.13% and TSH by 77.80% upon HgCl2 exposure in the hypothalamus. However, subsequent co-treatment with TQ significantly (p<0.05) ameliorated these disruptions and restored the antioxidant capacity of the hypothalamus.
3.6. TQ Co-Treatment Attenuated HgCl2-Induced Oxidative Stress in the Reproductive Tissues of Rats
The ability of TQ co-treatment to attenuate HgCl2-induced oxidative stress in the reproductive tissues of rats is displayed in Figure 7 and Figure 8. Exposure of rats to HgCl2 resulted in a significant (p<0.05) upsurge in the levels of RONS & LPO and activity of XO by 407.79%, 345.84%, and 225.92% in the testes, compared to the control. However, treatment with TQ alongside HgCl2 (2.5 and 5 mg/kg doses) effectively ameliorated these elevations. Specifically, 2.5 mg/kg co-treatment reduced the levels of RONS & LPO and XO activity by 46.71%, 50.12%, and 48.69%, respectively while treatment with the higher dose of TQ (5 mg/kg) further decreased the levels of RONS & LPO and activity of XO, showing reductions by 60.25%, 61.07%, and 62.21% respectively, in comparison to the HgCl2 treatment alone in the testes. In the epididymis, HgCl2 exposure increased RONS & LPO levels and XO activity by 92.86%, 80.28%, and 198.17% respectively. Remarkably, the 2.5 mg/kg treatment significantly (p<0.05) attenuated these increases, culminating in decreases of 21.48%, 33.29%, and 24.35%, respectively. The group of rats co-treated with the high dose of TQ (5.0 mg/kg) had these levels reduced by 36.26%, 59.16%, and 55.55%, respectively, compared to the HgCl2-alone group. Additionally, the hypothalamus of experimental rats demonstrated a marked increase in RONS & LPO levels and the activity of XO by 622.94%, 47.49%, and 242.38%, respectively, following HgCl2 administration alone. Contrastingly, the co-treatment with 2.5 mg/kg of TQ significantly (p<0.05) reduced the observed increments by 55.63%, 20.08%, and 24.35%, respectively. Animals that received the higher dose (5.0 mg/kg) displayed pronounced protective effects, with significant (p<0.05) decreases in RONS & LPO levels and XO activity by 80.51%, 26.33%, and 53.08%, respectively, compared to the HgCl2-treated group alone. The level of protein carbonyl was significantly raised in the testes (73.29%), epididymis (137.66%), and hypothalamus (307.63%) upon HgCl2 alone administration. However, treatment of rats with TQ (low and high dose) resulted in a significant reduction in the levels of PC in the testes (29.34 & 37.60%), epididymis (22.67 & 37.63%), and hypothalamus (17.38 & 56.68%).
3.7. TQ Co-Administration Reversed Inflammation Caused by HgCl2 Exposure in the Testes, Epididymis and Hypothalamus of Rats
Figure 9 and Figure 10 evidence the anti-inflammatory activity of TQ co-administration in the testes, epididymis and hypothalamus of rats treated with HgCl2. There was a significant (p<0.05) elevation in the levels of NO, TNF-α, IL-1β and the activity of MPO in the testes of animals by 556.03%, 363.41%, 115.90%, and 92.39%, compared to the control group. Contrastingly, animals co-treated with TQ (2.5 mg/kg dose) effectively reduced these elevated levels: NO by 49.03%, TNF-α by 37.46%, MPO by 24.29%, and IL-1β by 25.89%. Rats in the 5.0 mg/kg co-treatment group exhibited more distinct reductions with decreases of 75.89%, 65.78%, 42.24%, and 36.45%, respectively, compared to animals that received HgCl2 alone. The epididymis of rats post-HgCl2 exposure showed significant (p<0.05) increases in NO, TNF-α, MPO, and IL-1β by 173.34%, 135.97%, 185.24%, and 452.82%, compared to control levels. However, 2.5 mg/kg treatment significantly countered these effects, reducing NO levels by 31.42%, TNF-α by 19.52%, MPO activity by 47.26%, and IL-1β by 35.80%. Administration of TQ at the 5.0 mg/kg dose significantly reduced these biomarkers by 53.05%, 34.61%, 56.60%, and 54.70%, respectively, compared to the HgCl2-only group in the epididymis. Post HgCl2 exposure, the hypothalamus was observed to have significant (p<0.05) increments in the levels of NO (226.62%), TNF-α (481.46%), MPO (190.06%), and IL-1β (359.12%) compared to the control. However, TQ co-treatment, 2.5 and 5.0 mg/kg, effectively attenuated these escalations, demonstrated by significant (p<0.05) decreases of 40.99 & 52.56%, 46.54 & 61.35%, 26.43 & 48.75%, 48.25 & 69.60% respectively, in the hypothalamus.
3.8. TQ Co-Treatment Assuaged Apoptosis and Cellular Damage in Experimental Rats Treated with HgCl2
The results of TQ supplementation against HgCl2-induced apoptosis are depicted in Figure 11 and Figure 12. Treatment of animals with HgCl2 alone led to a significant (p<0.05) decline in the expression of Bcl-2 by 94.17%. Conversely, compared to control levels, Bax and P53 levels increased by 100.73% and 228.33%, respectively. However, TQ co-treatment significantly reversed these alterations in Bcl-2 expression. Bax and P53 expressions, on the other hand, decreased by 29.71 & 52.09% and 29.12 & 51.77% in the 2.5 & 5.0 mg/kg treatment, respectively, in the testes, when compared to HgCl2 treatment alone. In the epididymis, HgCl2 treatment markedly decreased Bcl-2 level by 65.87% while raising the expressions of Bax and P53 by 422.09% and 300.29%, respectively, against the control group. However, Bcl-2 levels rose by 568.64 & 317.60%, while Bax expression was reduced by 38.04 & 65.95% and P53 expression by 46.45 & 57.99% for 2.5 and 5.0 mg/kg doses, respectively, compared to HgCl2 only. Furthermore, Bcl-2 expression decreased by 44.54%, while Bax and P53 expressions increased by 153.82% and 333.57%, respectively, compared to control levels in the hypothalamus. However, low and high-dose TQ co-treatment amplified Bcl-2 expression by 455.08 & 233.24% and reduced Bax and P53 expressions by 34.89 & 61.95% and 32.22 & 49.71% respectively.
3.9. TQ Reduced HgCl2-Induced Testicular and Epididymal Histomorphometry Alteration
Corresponding photomicrographs depict the histological findings (Figures 13A and 14A) and histomorphological measurements (Figures 13B and 14B) obtained from the testis and epididymis of experimental rats co-treated with TQ and HgCl2. Figure 13 and Figure 14 show the effect of thymoquinone on the histomorphometry of testicular and epididymal tissues in mercury chloride co-exposure in rats. HgCl2-only exposure rats demonstrated significant atrophy, vacuolation of Sertoli cells, and a notable decrease in the seminiferous tubular distance [F (4, 10) = 11.30, p = 0.001, Figure 13] a decrease in epididymal epithelial thickness [F (4, 10) = 35.61, p < 0.0001, Figure 14] compared with the control. On the other hand, thymoquinone-treated (2.5 and 5 mg/kg) rats partially restored testis and epididymis histoarchitecture with well-organised germinal cells, spermatids, and stored mature sperm cells.
3.10. TQ Mediate Antioxidant and Anti-Inflammatory Effects through PPAR-α or PPAR δ/β Signalling
Following the findings from the in vivo studies, we conducted molecular docking simulations to explore TQ's potential to mediate antioxidant and anti-inflammatory effects. We assessed the binding constant (Kd) from TQ's interactions with PPAR-α or PPAR δ/β. The results showed that TQ had a moderate interaction with PPAR-α or PPAR δ/β, demonstrated by a Kd value of 3.90 × 10⁵ M and 2.32 × 10⁻⁵ M, respectively (Figure 15A). Through this interaction, PPAR δ/β can positively engage Nrf-2 to induce antioxidants, while PPAR-α can modulate the function of NF-kB to induce anti-inflammatory responses (Figure 15 B). Taken together, these results indicate that TQ might possess antioxidant and anti-inflammatory effects by activating PPAR-α and PPAR δ/β.
4. Discussion
Exposure to heavy metals such as mercury has been shown to cause abnormalities in the reproductive system, potentially leading to infertility or subfertility in males (Henriques et al., 2019). This paper aims to investigate the effectiveness of thymoquinone (TQ), an antioxidant with anti-inflammatory and anti-apoptotic properties, in protecting the hypothalamic-gonadal axis of male mice from mercuric chloride (HgCl2)-induced toxicity.
Evaluating body and organ weights is vital for assessing toxicity, as changes in these indices may indicate physiological disturbances in animals (Lazic et al., 2020). In this study, exposure to HgCl2 reduced body weight gain, organ weight, and relative organ weight of the reproductive tissues, as shown in Table 2. A reduction in body weight is symptomatic of mercury toxicity, which is often associated with loss of appetite in experimental animals (Shalan, 2022). This observation is in line with other research that reported insignificant changes in body and organ weights upon HgCl2 administration. However, administration of TQ restored body and organ weights in experimental rats, suggesting its protective effect.
Sperm quality, including parameters such as motility, count, viability, and morphology, is a crucial measure of male fertility in toxicological studies (Tanga et al., 2021). In this study, HgCl2 exposure significantly reduced sperm motility by approximately 27.78% relative to controls, highlighting its adverse effects (Table 3). Sperm motility is essential for fertilisation capabilities, and a decline of 27.78% could jeopardise this function (Table 3). This finding aligns with previous research indicating that HgCl2 impairs sperm membrane integrity and induces morphological defects (Martinez et al., 2017).
Additionally, a significant reduction was observed in sperm count following HgCl2 exposure, with a 25.60% decrease compared to controls, indicating adverse effects on male fertility. Reduced sperm count can lower the chances of successful fertilisation, according to Dabbagh Rezaeiyeh et al. (2022), who found similar results where HgCl2 disrupted germinal epithelium, reducing sperm production (Jahan et al., 2019). Interestingly, semen volume remained consistent across all groups (Table 3), suggesting that while HgCl2 affects spermatogenesis, it does not appear to impact seminal fluid production or ejaculation volume. This observation contrasts with results from Altunkaynak et al. (2015), who noted a reduction in sperm volume when treating rats with elemental mercury (Hg0) as opposed to HgCl2.
While the changes in morphological abnormalities of the sperm between groups post-HgCl2 exposure were insignificant, subtle alterations in sperm morphology can affect their functionality (Moretti et al., 2022), Table 3. The minor increases in abnormalities across the headpiece, midpiece, and tail, albeit statistically insignificant, hint at possible disruptions in spermiogenesis – the final stages of sperm maturation. Previous studies outlined mercury's ability to induce sperm DNA damage (Jahan et al., 2019; Wyatt et al., 2017), which might be corroborated by these observed morphological perturbations. Nevertheless, TQ administration was observed to insignificantly increase sperm morphological and functional parameters in this study Table 3. This result contradicts prior findings, where TQ showcased commendable recuperative actions against various sperm parameter toxicities in rats (Altunkaynak et al., 2015). The subtle increases in morphological abnormalities even post-TQ treatment, especially in the 5.0 mg/kg co-treatment group, might suggest that TQ's protective mechanisms do not entirely circumvent the structural disruptions instigated by mercury.
A complex network of hormones regulates the male reproductive system, including LH, FSH, testosterone, and prolactin (Sokwala, 2021). These hormones are produced in the testes and pituitary glands and are responsible for various functions such as sperm production, sex drive, and secondary sexual characteristics (Sadiq & Tadi, 2024). LH and FSH stimulate the production of testosterone, which is essential for developing and maintaining male reproductive and accessory reproductive organs (Sadiq & Tadi, 2024). Prolactin, on the other hand, plays a role in suppressing the production of LH and FSH, which can lead to a decrease in testosterone levels (Sadiq & Tadi, 2024). The significant diminutions in serum LH, FSH, and testosterone levels upon exposure to HgCl2 corroborate the reports delineated by earlier investigations, where HgCl2's toxicological profile encompassed reproductive harm (Albasher et al., 2020; Sampada & David, 2024), Figure 2. A decline in LH and FSH levels, the primary regulators of spermatogenesis and testosterone synthesis, hints at potential disruptions in the hypothalamic-pituitary-gonadal (HPG) axis Figure 2. A concomitant reduction in testosterone, a pivotal steroid for maintaining male secondary sexual characteristics, sperm production, and libido, provides evidence of this compromised HPG axis, potentially underscoring reduced Leydig cell functionality or disrupted feedback mechanisms. Conversely, the surge in prolactin levels post-HgCl2 exposure presents an intriguing conundrum. In a male physiological context, elevated prolactin can suppress LH and FSH secretion, potentially contributing to the observed declines in these hormones and, by extension, testosterone (Petersenn et al., 2023).
In contrast, co-treatment with TQ (2.5 and 5.0 mg/kg) demonstrated recuperative actions by mitigating the hormonal aberrations induced by HgCl2. This normalisation of hormonal levels, particularly testosterone restoration and prolactin reduction, underscores TQ’s potential in safeguarding male rats’ endocrine system Figure 2. These findings are consistent with previous research where TQ was shown to counteract heavy metal-induced endocrine and reproductive derangements (Hassan et al., 2019). Furthermore, the dose-dependent effectiveness of TQ is evident, as the elevated dose (5.0 mg/kg) results in more pronounced rectifications (515.08%), underscoring its potential therapeutic range. This finding supports the hypothesis that TQ may influence at various levels—shielding testicular Leydig cells, revitalising the hypothalamic-pituitary-gonadal (HPG) axis, and potentially directly modulating steroidogenic enzymes.
Mercury's disruptive influence on the reproductive system, particularly on the testes, has been a focal point of numerous scientific inquiries in recent years (Henriques et al., 2019). Due to its pivotal role in spermatogenesis, the testicular tissue is a highly metabolically active site, relying on specialised enzymes to maintain its structure and function. Alkaline phosphatase (ALP) plays a role in phosphate metabolism and membrane transport, supporting germ cell development and maturation within the seminiferous epithelium (Sekaran et al., 2021). Acid phosphatase (ACP), primarily a lysosomal enzyme, is involved in sperm maturation. At the same time, lactate dehydrogenase (LDH) is essential for energy metabolism in spermatogenic cells, supporting sperm motility and viability through anaerobic glycolysis (Rotimi et al., 2024). Glucose-6-phosphate dehydrogenase (G-6-PD) contributes to redox homeostasis by generating NADPH, which protects testicular cells from oxidative damage and supports steroidogenic activity in Leydig cells (Chen et al., 2022). The current study observed a significant (p<0.05) diminution in the testicular enzymatic activities of ALP, ACP, LDH, and G-6-PD upon exposure to HgCl2 Figure 3. The significant decline in the enzymatic activities of ALP and ACP, which are closely associated with spermatogenic activity and testicular cellular function, indicates the potential compromise of sperm maturation processes upon HgCl2 exposure Figure 3. Similarly, the downturn in LDH, a crucial enzyme for cellular energetics, and G-6-PD, central to redox balance, alludes to a dual blow: an energetic crisis and heightened vulnerability to oxidative perturbations. This complements the findings of Jahan et al. (2019), where mercury exposure translated into compromised sperm production and amplified oxidative damage. Conversely, TQ, especially at the higher dose (5.0 mg/kg), alleviated the HgCl2-induced enzyme activity reductions and augmented them to levels surpassing those seen in controls Figure 3. This resonates with the earlier findings about TQ's protective role against drug and heavy metal-induced reproductive damages (Hassan et al., 2019; Moradi Maryamneghari et al., 2021). Testicular cells, particularly spermatogenic and Leydig cells, are susceptible to oxidative stress, which can impair enzyme function and reduce fertility (Monageng et al., 2023). TQ scavenges reactive oxygen species (ROS) and upregulates endogenous antioxidant defence systems, preserving the structural integrity and function of testicular tissues. By mitigating lipid peroxidation and oxidative damage in the testes, TQ helps restore or enhance the activity of these testicular enzymes.
A consistent theme across studies on mercury toxicity is the perturbation of oxidative balance (Albasher et al., 2020; Boujbiha et al., 2009; Jahan et al., 2019). In this study, a significant (p<0.05) decline in the activities of CAT, SOD, GST, and GPx was observed in Figure 4 and Figure 5, reflecting the adverse impact of HgCl2 on the enzymatic antioxidants of the male rats’ reproductive tissues. The disruption of these enzymes signifies an overwhelmed oxidative defence system, rendering the cells vulnerable to oxidative damage. This aligns with the established understanding that HgCl2 stimulates the production of ROS, which, in turn, can cause cellular damage, inflammation, and apoptosis, central to mercury's reproductive toxicity (Almeer et al., 2020; Kandemir et al., 2020). Furthermore, the reduction in glutathione (GSH) and total sulfhydryl (TSH) levels indicates depleted cellular thiols Figure 6. GSH, a tripeptide, is fundamental in cellular defence against oxidative stress, and its depletion is a well-recognised marker of cellular oxidative damage (Narayanankutty et al., 2019). However, findings from this study demonstrate the protective effects of TQ as co-treatment with TQ, specifically at higher doses (5 mg/kg), significantly elevated the antioxidant enzymes and compounds initially suppressed by HgCl2 Figure 6. This result suggests that TQ might be acting by scavenging the ROS produced, upregulating the antioxidant defence genes, or both. The notable increase in GST activity, especially in the epididymis and hypothalamus following TQ co-treatment, emphasises its role in detoxification. GST is known to conjugate GSH to various electrophilic compounds, aiding their removal and neutralisation (Laborde, 2010). Therefore, the increased activity indicates a mechanism where TQ potentially induces GST expression or provides the necessary cofactors to optimise its activity.
Furthermore, the oxidative stress-inducing ability of HgCl2 is evident from the significant (p<0.05) increases in biomarkers of oxidative damage across the testes, epididymis, and hypothalamus of the exposed rats, specifically, RONS, LPO, and Protein Carbonyl levels and XO activity (Erboga et al., 2016), Figure 7 and Figure 8. These findings are consistent with previously established data, where mercury exposure is associated with an imbalance of the oxidative balance in the reproductive tissues of rats (Albasher et al., 2020; Boujbiha et al., 2009; Jahan et al., 2019). The rise in LPO levels, a marker of cellular lipid membrane damage, underscores the direct damage inflicted on cellular structures by ROS. Elevated levels of LPO are indicative of compromised membrane integrity and function, potentially leading to cellular dysfunction and death (Pardillo-Diaz et al., 2022). Similarly, the surge in XO activity, an enzyme that catalyses the oxidation of hypoxanthine to xanthine, further contributes to ROS generation and consequently intensifies oxidative stress (Battelli et al., 2016).
Additionally, the increase in RONS further corroborates the notion of heightened oxidative and nitrosative stress upon HgCl2 exposure. Contrastingly, TQ significantly (p<0.05) protected the examined tissues from HgCl2-induced oxidative damage Figure 7 and Figure 8. The protective effects of TQ, particularly at higher doses, resonate with its previously documented antioxidant (Algaidi et al., 2022; Mabrouk & Ben Cheikh, 2015). It appears that TQ either directly scavenges the free radicals generated by HgCl2 exposure or upregulates the cellular antioxidant defence system, thereby restoring the oxidative balance. The significant reduction in LPO, XO, and RONS in the testes upon TQ co-treatment suggests an amelioration of HgCl2-induced cellular and enzymatic damages. This has profound implications for male fertility, as the testes are pivotal for sperm production, the epididymis for sperm maturation and storage, and the hypothalamus for regulating numerous physiological functions, including reproductive hormone synthesis. Oxidative stress in these organs can compromise spermatogenesis, reducing sperm count, motility and increasing sperm abnormalities (Algaidi et al., 2022; Mabrouk & Ben Cheikh, 2015). However, the attenuation of oxidative markers in the epididymis suggests that TQ might preserve the structural and functional integrity of maturing sperm, ensuring their viability and motility and preserving endocrine homeostasis under oxidative insults.
It is evident from previous work that damaged cellular components, as a result of oxidative stress, can act as danger signals or "alarmins" which are recognised by specific receptors on immune cells, leading to the activation of various inflammatory pathways (Vona et al., 2021). Also, lipid peroxidation products can activate the nucleotide-binding oligomerisation domain-like receptors (NOD), leucine-rich repeats (LRR), and pyrin domain-containing protein 3 (NLRP3) inflammasome, resulting in the production and release of pro-inflammatory cytokines like TNF-α and IL-1β (Gora et al., 2021). Moreover, ROS can directly activate transcription factors such as NF-κB, which plays a central role in the expression of genes responsible for inflammatory responses (Khan et al., 2021). Therefore, the sustained oxidative stress, as is potentially caused by HgCl2 exposure, can perpetuate a chronic inflammatory state in the testes, epididymis and hypothalamus. Inflammation, depending on the duration, is a two-edged sword. While acute inflammation is protective and aids tissue repair, chronic or persistent inflammation is detrimental and has been implicated in many pathological conditions, including reproductive dysfunctions (Soliman & Barreda, 2023). In this study, a pronounced inflammatory response was observed in the testes, epididymis, and hypothalamus upon HgCl2 administration, as evidenced by the elevation in inflammatory markers, NO, MPO, IL-1β, and TNF-α (Figure 9 and Figure 10). Although a crucial signalling molecule in numerous physiological processes, NO can react with superoxide radicals at elevated levels to form peroxynitrite, a potent oxidant that can induce cellular damage (Sharifi-Rad et al., 2020). The significant upsurge in NO post-HgCl2 exposure indicates the oxidative and nitrative stress inflicted on the examined tissues. The concomitant rise in pro-inflammatory cytokines observed in the current study, such as TNF-α and IL-1β, and the myeloperoxidase marker of neutrophil infiltration, suggests an active inflammatory process (Figure 9 and Figure 10).
The increase in protein carbonyl levels further highlights oxidative protein damage, potentially compromising proteins' structural and functional integrity within the testes. Additionally, the significant increase in IL-1β levels, a potent pro-inflammatory cytokine, coupled with the elevation in other inflammatory markers (NO and MPO), suggests an epididymal environment that may not be conducive to sperm maturation and might compromise sperm motility and function. Moreover, chronic inflammation in the hypothalamus can potentially disrupt hormonal homeostasis, affecting downstream reproductive processes like hormone production and other vital functions regulated by this brain region (Sheng et al., 2020). This inflammatory and oxidative milieu can impair spermatogenesis, reduce testosterone synthesis, and affect overall testicular health. On the flip side, co-administration of TQ effectively counteracted the HgCl2-induced inflammation in rats' testes, epididymis, and hypothalamus. The higher dose (5.0 mg/kg) manifested more pronounced effects, suggesting a dose-dependent protective role of TQ. TQ has been shown to exert an anti-inflammatory effect via its inhibition of inflammatory cytokines and processes, such as TNF-α, inducible NOS, cyclooxygenase-2 (COX-2), 5-lipoxygenase, and cyclin D1 (Fouad & Jresat, 2015). TQ also modulates the nuclear factor-κΒ and mitogen-activated protein kinase signalling pathways, which regulate inflammation and cell proliferation (Leong et al., 2021).
Apoptosis is an essential physiological process that ensures cellular homeostasis by removing damaged or unnecessary cells; the balance between pro-apoptotic and anti-apoptotic molecules determines cellular fate (Singh et al., 2022). Bax (Bcl-2-associated X protein) induces the permeabilisation of the mitochondrial outer membrane, releasing cytochrome c into the cytosol (Singh et al., 2019). Once released, cytochrome c triggers a cascade of events culminating in cell death, making Bax a pro-apoptotic protein. Bax forms pores or channels in the mitochondrial membrane, facilitating the release of cytochrome c and other pro-apoptotic factors (Singh et al., 2019). On the other hand, B-cell lymphoma 2 or Bcl-2 prevents the permeabilisation of the mitochondrial membrane and subsequent release of cytochrome c by forming heterodimers with Bax, thereby inhibiting cell death (Edlich, 2018). P53 is a tumour suppressor protein that regulates cellular responses to stress and DNA damage (Vaddavalli & Schumacher, 2022). It is a transcription factor that activates genes involved in cell cycle arrest, DNA repair, senescence, and apoptosis. This transcription factor promotes cell death by upregulating the expression of pro-apoptotic genes, including Bax, while simultaneously repressing anti-apoptotic genes, such as Bcl-2 (Shen et al., 2023). By modulating the balance between pro- and anti-apoptotic factors, p53 helps to eliminate cells with irreparable DNA damage or those at risk of becoming cancerous, thus earning it the title "guardian of the genome (Vaddavalli & Schumacher, 2022)."
Additionally, p53 can directly induce apoptosis through transcription-independent mechanisms, such as interacting with pro-apoptotic proteins or promoting mitochondrial outer membrane permeabilisation (Shen et al., 2023). The significant decrease in Bcl-2 expression observed in the HgCl2-alone group across the testes, epididymis, and hypothalamus signifies the potent ability to induce apoptosis Figure 11 and Figure 12. Since Bcl-2 is vital in inhibiting apoptosis, its reduction would predispose cells to death, possibly accounting for the observed pathological changes and functional impairments in the reproductive tissues and the regulatory centre (hypothalamus). Concomitantly, the substantial upregulation of Bax and P53 upon HgCl2 exposure underscores the activation of apoptotic pathways. Bax initiates the intrinsic apoptotic cascade by promoting the release of cytochrome c from the mitochondria, while P53 facilitates apoptosis in response to DNA damage (Vaddavalli & Schumacher, 2022). The protective role of TQ was evident, as shown in the significant increase in Bcl-2 expression across all tissues upon TQ co-administration, implying that TQ can bolster cellular defences against apoptosis Figure 11 and Figure 12, thereby promoting cell survival. This is particularly crucial in the testes, where preserving the population of various cells, including the sperm-producing germ cells and testosterone-producing Leydig cells, is imperative for maintaining reproductive health. In the epididymis, safeguarding cells ensure optimal conditions for sperm maturation, and in the hypothalamus, neuronal survival is essential for endocrine and physiological regulation (Erboga et al., 2016; Sheikhbahaei et al., 2016).
Additionally, the concurrent reduction in Bax and P53 expressions upon TQ treatment in all tissues suggests that TQ can block the apoptotic signals induced by HgCl2. This observation concurs with earlier studies (Adinew et al., 2022; Homayoonfal et al., 2022). The significant downregulation of these pro-apoptotic molecules indicates that TQ potentially prevents the cellular cascades that culminate in apoptosis, safeguarding cellular integrity and function.
Our histomorphometry findings are consistent with our result on sperm morphology, which showed atrophy of the testicular tubules with a disarranged germ cell layer and vacuolation of Sertoli cells, which confirm impairment in spermatogenesis and degenerated germinal epithelium of the epididymis confirming impaired deficit in sperm maturation and count, viability and motility (Table 3) following oral exposure to HgCl2. This finding corroborated earlier studies that indicated ultrastructural impairment of the testis and epididymis concerning mercury chloride exposure (El-Desoky et al., 2013; Kalender et al., 2013). TQ-treated rat cohorts showed protection against testicular ultrastructural damage caused by HgCl2.
Peroxisome proliferator-activated receptors (PPARs), specifically PPAR-α, PPAR-γ, and PPAR δ/β, are an excellent therapeutic target. They have been known to induce antioxidant and anti-inflammatory functions (Owumi et al., 2024). They promote optimal health by conferring antioxidant and anti-inflammatory responses (Billin, 2008; Lee et al., 2017; Okada-Iwabu et al., 2013; Savkur & Miller, 2006). Molecular docking simulations demonstrated that PPAR δ/β positively engage Nrf-2 and modulate NF-kB function, promoting antioxidants and anti-inflammatory responses. Thus, TQ anti-oxidative and anti-inflammatory effects are by agonistic activation of PPAR-α and PPAR δ/β. Our in-silico studies showed that TQ could act as a PPAR-α and PPAR δ/β agonist, thereby increasing their potential to activate Nrf-2 and inhibit NF-kB. The activation of Nrf-2 upregulates the expression of cytoprotective genes, while inhibition of Nf-kB represses the expression of pro-inflammatory mediators (Owumi et al., 2024). Upon confirming our current observations in conjunction with previous studies, we propose that Thymoquinone (TQ) mitigates the reproductive toxicities induced by mercuric chloride (HgCl2) by enhancing the levels of endogenous antioxidants, which can counteract the reactive oxygen species (ROS) generation instigated by HgCl2. This mechanism diminishes pro-inflammatory mediators, apoptosis, and the histological aberrations orchestrated by the toxicant to the reproductive system. Specifically, superoxide dismutase (SOD) catalyses the conversion of superoxide anion radicals resulting from HgCl2-induced oxidative DNA damage—arising from the excessive release of purines, hypoxanthine, and xanthine—into hydrogen peroxide. In the absence of transformation into water by catalase (CAT) or glutathione peroxidase (GPx), the resultant hydrogen peroxide has the potential to react with labile iron species (Fe2+/Fe3+) in the Fenton reaction, generating hydroxyl radicals, which represent a more toxic form of ROS. These hydroxyl radicals may interact with membrane lipids, culminating in lipid peroxidation, as evidenced by elevated malondialdehyde levels (MDA or LPO).
Furthermore, should hydrogen peroxide accumulate without effective scavenging, it may react with tissue chlorides in the presence of myeloperoxidase (MPO) to produce hypochlorous acid—another ROS that incites pro-inflammatory responses within tissues. In addition to alleviating oxidative stress, TQ displays an anti-inflammatory effect against HgCl2 by curtailing the levels of nitric oxide (NO) that HgCl2 prompts via increased activity of inducible nitric oxide synthase, which elevates tissue NO concentrations. Moreover, TQ demonstrates anti-apoptotic characteristics within the reproductive system by modulating the levels of p53, Bax (a pro-apoptotic protein) and augmenting the activity of Bcl-2 (an anti-apoptotic protein), thereby attenuating the downstream repercussions on caspases-9 and -3. Through these mechanisms, TQ effectively modulates HgCl2-induced reproductive toxicities in male Wistar albino rats (Figure 16).
This study has some limitations. We did not measure daily food intake, which limits our understanding of how thymoquinone (TQ) mitigates the effects of mercuric chloride on body weight gain and organ weight. Additionally, the bioavailability of TQ was not assessed, as it was beyond the scope of this study, leaving uncertainty about its systemic absorption and distribution. While molecular docking simulations were employed to identify potential therapeutic targets underlying TQ's antioxidant and anti-inflammatory effects, these findings are preliminary and require further experimental validation.
5. Conclusions
Collectively, this study found that TQ supplementation reduced the adverse effects of HgCl2 on sperm parameters such as motility, count, and morphology, indicating potential benefits for male fertility. TQ treatment also restored testicular enzymatic activities, hormonal levels, and oxidative balance, while reducing inflammation, apoptosis, and histomorphometric changes in reproductive tissues. The effects of TQ were dose-dependent, with higher doses leading to more significant outcomes. These results demonstrate various mechanisms by which TQ may counteract HgCl2-induced hepatorenal and reproductive toxicity (Owumi et al., 2025), through anti-oxidative, anti-inflammatory, and anti-apoptotic pathways. TQ may be considered a candidate for addressing the effects of heavy metal exposure on male reproductive health.
Authors Contribution
All authors partook in the design, interpretation, and analysis of the data generated from the study. SO conceptualised the study; MO, PA, BO, JOB, JC, CI, AA, UA, and OO conducted the research and preliminary data analysis. SO and AKO supervised the investigation, and SO, MO, PA, BO, JOB, JC, UA, CI, AA, OO and AKO proofread data for errors. The manuscript was written and revised by SSO, MO, PA, BO, JOB, JC, CI, AA, UA, OO and AKO.
Funding
The authors privately funded this research through their contribution and received no external grant from funding agencies in the commercial, not-for-profit, or public sectors.
Data Availability Statement
The datasets used and analysed during the current study are available from the corresponding author upon reasonable request
Conflicts of Interest
The authors reported no potential conflict of interest
References
- Adinew, G. M., Messeha, S. S., Taka, E., Badisa, R. B., Antonie, L. M., & Soliman, K. F. A. (2022). Thymoquinone Alterations of the Apoptotic Gene Expressions and Cell Cycle Arrest in Genetically Distinct Triple-Negative Breast Cancer Cells. Nutrients, 14(10). [CrossRef]
- Albasher, G., Alkahtani, S., & Alarifi, S. (2020). Berberine mitigates oxidative damage associated with testicular impairment following mercury chloride intoxication. J Food Biochem, 44(9), e13385. [CrossRef]
- Algaidi, S. A., Faddladdeen, K. A., Alrefaei, G. I., Qahl, S. H., Albadawi, E. A., HM, A. L., & Ayuob, N. N. (2022). Thymoquinone protects the testes of hypothyroid rats by suppressing pro-inflammatory cytokines and oxidative stress and promoting SIRT1 testicular expression. Front Pharmacol, 13, 1040857o-inflammatory cytokines and oxidative stress and promoting SIRT1 testicular expression. [CrossRef]
- Almeer, R. S., Albasher, G., Kassab, R. B., Ibrahim, S. R., Alotibi, F., Alarifi, S., Ali, D., Alkahtani, S., & Abdel Moneim, A. E. (2020). Ziziphus spina-christi leaf extract attenuates mercury chloride-induced testicular dysfunction in rats. Environ Sci Pollut Res Int, 27(3), 3401–3412. [CrossRef]
- Altunkaynak, M. E., Akgul, N., Yahyazadeh, A., Altunkaynak, B. Z., Turkmen, A. P., Akgul, H. M., Aksak, S., & Unal, B. (2015). A stereological and histopathological study of the effects of exposure of male rat testes to mercury vapor. Biotech Histochem, 90(7), 529–534. [CrossRef]
- Badary, O. A., Hamza, M. S., & Tikamdas, R. (2021). Thymoquinone: A Promising Natural Compound with Potential Benefits for COVID-19 Prevention and Cure. Drug Des Devel Ther, 15, 1819–1833. [CrossRef]
- Bancroft, J. D., & Gamble, M. (2008). Theory and practise of histological techniques (6th ed.). Churchill Livingstone Elsevier.
- Battelli, M. G., Polito, L., Bortolotti, M., & Bolognesi, A. (2016). Xanthine Oxidoreductase-Derived Reactive Species: Physiological and Pathological Effects. Oxid Med Cell Longev, 2016, 3527579. [CrossRef]
- Bergmeyer, H. I., Gawehn, K., & Grassl, M. (1974). Methods of Enzymatic analysis (H. U. Bergmeyer, Ed. 2 ed., Vol. 1). Academic Press Inc.
- Billin, A. N. (2008). PPAR-beta/delta agonists for Type 2 diabetes and dyslipidemia: an adopted orphan still looking for a home. Expert Opin Investig Drugs, 17(10), 1465–1471. [CrossRef]
- Boujbiha, M. A., Hamden, K., Guermazi, F., Bouslama, A., Omezzine, A., Kammoun, A., & El Feki, A. (2009). Testicular toxicity in mercuric chloride treated rats: association with oxidative stress. Reprod Toxicol, 28(1), 81–89. [CrossRef]
- Chen, P. H., Tjong, W. Y., Yang, H. C., Liu, H. Y., Stern, A., & Chiu, D. T. (2022). Glucose-6-Phosphate Dehydrogenase, Redox Homeostasis and Embryogenesis. Int J Mol Sci, 23(4). [CrossRef]
- Edlich, F. (2018). BCL-2 proteins and apoptosis: Recent insights and unknowns. Biochem Biophys Res Commun, 500(1), 26–34. [CrossRef]
- El-Desoky, G. E., Bashandy, S. A., Alhazza, I. M., Al-Othman, Z. A., Aboul-Soud, M. A., & Yusuf, K. (2013). Improvement of mercuric chloride-induced testis injuries and sperm quality deteriorations by Spirulina platensis in rats. PLoS One, 8(3), e59177. [CrossRef]
- Erboga, M., Aktas, C., Kurt, O., Uygur, R., Caglar, V., Turan, B. C., Topcu, B., Fidanol Erboga, Z., Gurel, A., & Ozen, O. A. (2016). Protective effects of thymoquinone on experimental testicular ischaemia-reperfusion injury: an apoptotic, proliferative and biochemical study. Andrologia, 48(2), 222–230. [CrossRef]
- Fouad, A. A., & Jresat, I. (2015). Thymoquinone therapy abrogates toxic effect of cadmium on rat testes. Andrologia, 47(4), 417–426. [CrossRef]
- Gora, I. M., Ciechanowska, A., & Ladyzynski, P. (2021). NLRP3 Inflammasome at the Interface of Inflammation, Endothelial Dysfunction, and Type 2 Diabetes. Cells, 10(2). [CrossRef]
- Green, L. C., Wagner, D. A., Glogowski, J., Skipper, P. L., Wishnok, J. S., & Tannenbaum, S. R. (1982). Analysis of nitrate, nitrite, and [15N]nitrate in biological fluids. Anal Biochem, 126(1), 131–138. [CrossRef]
- Hassan, E., El-Neweshy, M., Hassan, M., & Noreldin, A. (2019). Thymoquinone attenuates testicular and spermotoxicity following subchronic lead exposure in male rats: Possible mechanisms are involved. Life Sci, 230, 132–140. [CrossRef]
- Henriques, M. C., Loureiro, S., Fardilha, M., & Herdeiro, M. T. (2019). Exposure to mercury and human reproductive health: A systematic review. Reprod Toxicol, 85, 93–103. [CrossRef]
- Homayoonfal, M., Asemi, Z., & Yousefi, B. (2022). Potential anticancer properties and mechanisms of thymoquinone in osteosarcoma and bone metastasis. Cellular & Molecular Biology Letters, 27(1), 21. [CrossRef]
- Jahan, S., Azad, T., Ayub, A., Ullah, A., Afsar, T., Almajwal, A., & Razak, S. (2019). Ameliorating potency of Chenopodium album Linn. and vitamin C against mercuric chloride-induced oxidative stress in testes of Sprague Dawley rats. Environ Health Prev Med, 24(1), 62. [CrossRef]
- Kalender, S., Uzun, F. G., Demir, F., Uzunhisarcikli, M., & Aslanturk, A. (2013). Mercuric chloride-induced testicular toxicity in rats and the protective role of sodium selenite and vitamin E. Food Chem Toxicol, 55, 456–462. [CrossRef]
- Kandemir, F. M., Caglayan, C., Aksu, E. H., Yildirim, S., Kucukler, S., Gur, C., & Eser, G. (2020). Protective effect of rutin on mercuric chloride-induced reproductive damage in male rats. Andrologia, 52(3), e13524. [CrossRef]
- Khan, A. Q., Rashid, K., AlAmodi, A. A., Agha, M. V., Akhtar, S., Hakeem, I., Raza, S. S., & Uddin, S. (2021). Reactive oxygen species (ROS) in cancer pathogenesis and therapy: An update on the role of ROS in anticancer action of benzophenanthridine alkaloids. Biomed Pharmacother, 143, 112142. [CrossRef]
- Laborde, E. (2010). Glutathione transferases as mediators of signaling pathways involved in cell proliferation and cell death. Cell Death & Differentiation, 17(9), 1373–1380. [CrossRef]
- Lee, T. W., Bai, K. J., Lee, T. I., Chao, T. F., Kao, Y. H., & Chen, Y. J. (2017). PPARs modulate cardiac metabolism and mitochondrial function in diabetes. J Biomed Sci, 24(1), 5. [CrossRef]
- Leong, X. F., Choy, K. W., & Alias, A. (2021). Anti-Inflammatory Effects of Thymoquinone in Atherosclerosis: A Mini Review [Mini Review]. Front Pharmacol, 12, 758929. [CrossRef]
- Mabrouk, A., & Ben Cheikh, H. (2015). Thymoquinone supplementation reverses lead-induced oxidative stress in adult rat testes. Gen Physiol Biophys, 34(1), 65–72. [CrossRef]
- Meng, E. C., Goddard, T. D., Pettersen, E. F., Couch, G. S., Pearson, Z. J., Morris, J. H., & Ferrin, T. E. (2023). UCSF ChimeraX: Tools for structure building and analysis. Protein science : a publication of the Protein Society, 32(11), e4792. [CrossRef]
- Mizuno, M., & Fukuhara, K. (2022). Antioxidant and Prooxidant Effects of Thymoquinone and Its Hydroquinone Metabolite. Biological & pharmaceutical bulletin, 45(9), 1389–1393. [CrossRef]
- Monageng, E., Offor, U., Takalani, N. B., Mohlala, K., & Opuwari, C. S. (2023). A Review on the Impact of Oxidative Stress and Medicinal Plants on Leydig Cells. Antioxidants (Basel), 12(8). [CrossRef]
- Moradi Maryamneghari, S., Shokri-Asl, V., Abdolmaleki, A., & Jalili, C. (2021). Genetic, biochemical and histopathological evaluations of thymoquinone on male reproduction system damaged by paclitaxel in Wistar rats. Andrologia, 53(10), e14192. [CrossRef]
- Moretti, E., Signorini, C., Noto, D., Corsaro, R., & Collodel, G. (2022). The relevance of sperm morphology in male infertility. Front Reprod Health, 4, 945351. [CrossRef]
- Narayanankutty, A., Job, J. T., & Narayanankutty, V. (2019). Glutathione, an Antioxidant Tripeptide: Dual Roles in Carcinogenesis and Chemoprevention. Curr Protein Pept Sci, 20(9), 907–917. [CrossRef]
- Ohkawa, H. O., N.; Yagi, K.. (1979). Assay for lipid peroxidation in animal tissues by Thiobarbituric acid reaction Anal Biochem 95, 351–358.
- Okada-Iwabu, M., Yamauchi, T., Iwabu, M., Honma, T., Hamagami, K., Matsuda, K., Yamaguchi, M., Tanabe, H., Kimura-Someya, T., Shirouzu, M., Ogata, H., Tokuyama, K., Ueki, K., Nagano, T., Tanaka, A., Yokoyama, S., & Kadowaki, T. (2013). A small-molecule AdipoR agonist for type 2 diabetes and short life in obesity. Nature, 503(7477), 493–499. [CrossRef]
- Owumi, S., Chimezie, J., Otunla, M., Oluwawibe, B., Agbarogi, H., Anifowose, M., Arunsi, U., & Owoeye, O. (2024). Prepubertal Repeated Berberine Supplementation Enhances Cerebrocerebellar Functions by Modulating Neurochemical and Behavioural Changes in Wistar Rats. Journal of Molecular Neuroscience, 74(3), 72. [CrossRef]
- Owumi, S., Otunla, M., Akindipe, P., Oluwawibe, B., Babalola, J. O., Chimezie, J., Arunsi, U., Owoeye, O., & Oyelere, A. K. (2025). Thymoquinone modulates oxidative stress and inflammation, correcting mercury-induced HO-1/NRF/Trx pathway disruption in experimental rat hepatorenal system: an in vivo and in silico study. Biometals, 38(4), 1179–1202. [CrossRef]
- Pardillo-Diaz, R., Perez-Garcia, P., Castro, C., Nunez-Abades, P., & Carrascal, L. (2022). Oxidative Stress as a Potential Mechanism Underlying Membrane Hyperexcitability in Neurodegenerative Diseases. Antioxidants (Basel), 11(8). [CrossRef]
- Perez-Severiano, F., Santamaria, A., Pedraza-Chaverri, J., Medina-Campos, O. N., Rios, C., & Segovia, J. (2004). Increased formation of reactive oxygen species, but no changes in glutathione peroxidase activity, in striata of mice transgenic for the Huntington's disease mutation. Neurochem Res, 29(4), 729–733. [CrossRef]
- Petersenn, S., Fleseriu, M., Casanueva, F. F., Giustina, A., Biermasz, N., Biller, B. M. K., Bronstein, M., Chanson, P., Fukuoka, H., Gadelha, M., Greenman, Y., Gurnell, M., Ho, K. K. Y., Honegger, J., Ioachimescu, A. G., Kaiser, U. B., Karavitaki, N., Katznelson, L., Lodish, M.,…Melmed, S. (2023). Diagnosis and management of prolactin-secreting pituitary adenomas: a Pituitary Society international Consensus Statement. Nature Reviews Endocrinology, 19(12), 722–740. [CrossRef]
- Rotimi, D. E., Iyobhebhe, M., Oluwayemi, E. T., Olajide, O. P., Akinsanola, B. A., Evbuomwan, I. O., Asaleye, R. M., & Ojo, O. A. (2024). Energy metabolism and spermatogenesis. Heliyon, 10(19), e38591. [CrossRef]
- Sadiq, N. M., & Tadi, P. (2024). Physiology, Pituitary Hormones. In StatPearls. StatPearls Publishing.
- Copyright © 2024, StatPearls Publishing LLC.
- Sampada, M., & David, M. (2024). Mercuric chloride induced reproductive toxicity associated with oxidative damage in male Wistar albino rat, Rattus norvegicus. Naunyn-Schmiedeberg's Archives of Pharmacology, 1–27. [CrossRef]
- Savkur, R. S., & Miller, A. R. (2006). Investigational PPAR-gamma agonists for the treatment of Type 2 diabetes. Expert Opin Investig Drugs, 15(7), 763–778. [CrossRef]
- Sekaran, S., Vimalraj, S., & Thangavelu, L. (2021). The Physiological and Pathological Role of Tissue Nonspecific Alkaline Phosphatase beyond Mineralization. Biomolecules, 11(11), 1564.
- Sharifi-Rad, M., Anil Kumar, N. V., Zucca, P., Varoni, E. M., Dini, L., Panzarini, E., Rajkovic, J., Tsouh Fokou, P. V., Azzini, E., Peluso, I., Prakash Mishra, A., Nigam, M., El Rayess, Y., Beyrouthy, M. E., Polito, L., Iriti, M., Martins, N., Martorell, M., Docea, A. O.,…Sharifi-Rad, J. (2020). Lifestyle, Oxidative Stress, and Antioxidants: Back and Forth in the Pathophysiology of Chronic Diseases [Review]. Front Physiol, 11, 694. [CrossRef]
- Sheikhbahaei, F., Khazaei, M., Rabzia, A., Mansouri, K., & Ghanbari, A. (2016). Protective Effects of Thymoquinone against Methotrexate-Induced Germ Cell Apoptosis in Male Mice. Int J Fertil Steril, 9(4), 541–547. [CrossRef]
- Shen, J., Wang, Q., Mao, Y., Gao, W., & Duan, S. (2023). Targeting the p53 signaling pathway in cancers: Molecular mechanisms and clinical studies. MedComm (2020), 4(3), e288. [CrossRef]
- Sheng, J. A., Bales, N. J., Myers, S. A., Bautista, A. I., Roueinfar, M., Hale, T. M., & Handa, R. J. (2020). The Hypothalamic-Pituitary-Adrenal Axis: Development, Programming Actions of Hormones, and Maternal-Fetal Interactions [Review]. Front Behav Neurosci, 14, 601939. [CrossRef]
- Singh, R., Letai, A., & Sarosiek, K. (2019). Regulation of apoptosis in health and disease: the balancing act of BCL-2 family proteins. Nature Reviews Molecular Cell Biology, 20(3), 175–193. [CrossRef]
- Singh, V., Khurana, A., Navik, U., Allawadhi, P., Bharani, K. K., & Weiskirchen, R. (2022). Apoptosis and Pharmacological Therapies for Targeting Thereof for Cancer Therapeutics. Sci, 4(2), 15.
- Sokwala, S. (2021). Chapter 2 - Reproductive endocrine physiology. In R. Rehman & A. Sheikh (Eds.), Subfertility (pp. 39–64). Elsevier. [CrossRef]
- Soliman, A. M., & Barreda, D. R. (2023). Acute Inflammation in Tissue Healing. International Journal of Molecular Sciences, 24(1), 641.
- Trush, M. A., Egner, P. A., & Kensler, T. W. (1994). Myeloperoxidase as a biomarker of skin irritation and inflammation. Food Chem Toxicol, 32(2), 143–147. [CrossRef]
- Vaddavalli, P. L., & Schumacher, B. (2022). The p53 network: cellular and systemic DNA damage responses in cancer and aging. Trends Genet, 38(6), 598–612. [CrossRef]
- Vona, R., Pallotta, L., Cappelletti, M., Severi, C., & Matarrese, P. (2021). The Impact of Oxidative Stress in Human Pathology: Focus on Gastrointestinal Disorders. Antioxidants (Basel), 10(2), 201. [CrossRef]
- Wyatt, L. H., Luz, A. L., Cao, X., Maurer, L. L., Blawas, A. M., Aballay, A., Pan, W. K., & Meyer, J. N. (2017). Effects of methyl and inorganic mercury exposure on genome homeostasis and mitochondrial function in Caenorhabditis elegans. DNA Repair (Amst), 52, 31–48. [CrossRef]
Figure 2.
The effect of Thymoquinone on reproductive hormones in rats treated with mercuric chloride for 28 days. Results are shown as mean ± SD (n=8), p < 0.05, 0.01, 0.001, 0.0001 (*, **, ***, ****). FSH: follicle-stimulating hormone, and LH: luteinising hormone.
Figure 2.
The effect of Thymoquinone on reproductive hormones in rats treated with mercuric chloride for 28 days. Results are shown as mean ± SD (n=8), p < 0.05, 0.01, 0.001, 0.0001 (*, **, ***, ****). FSH: follicle-stimulating hormone, and LH: luteinising hormone.
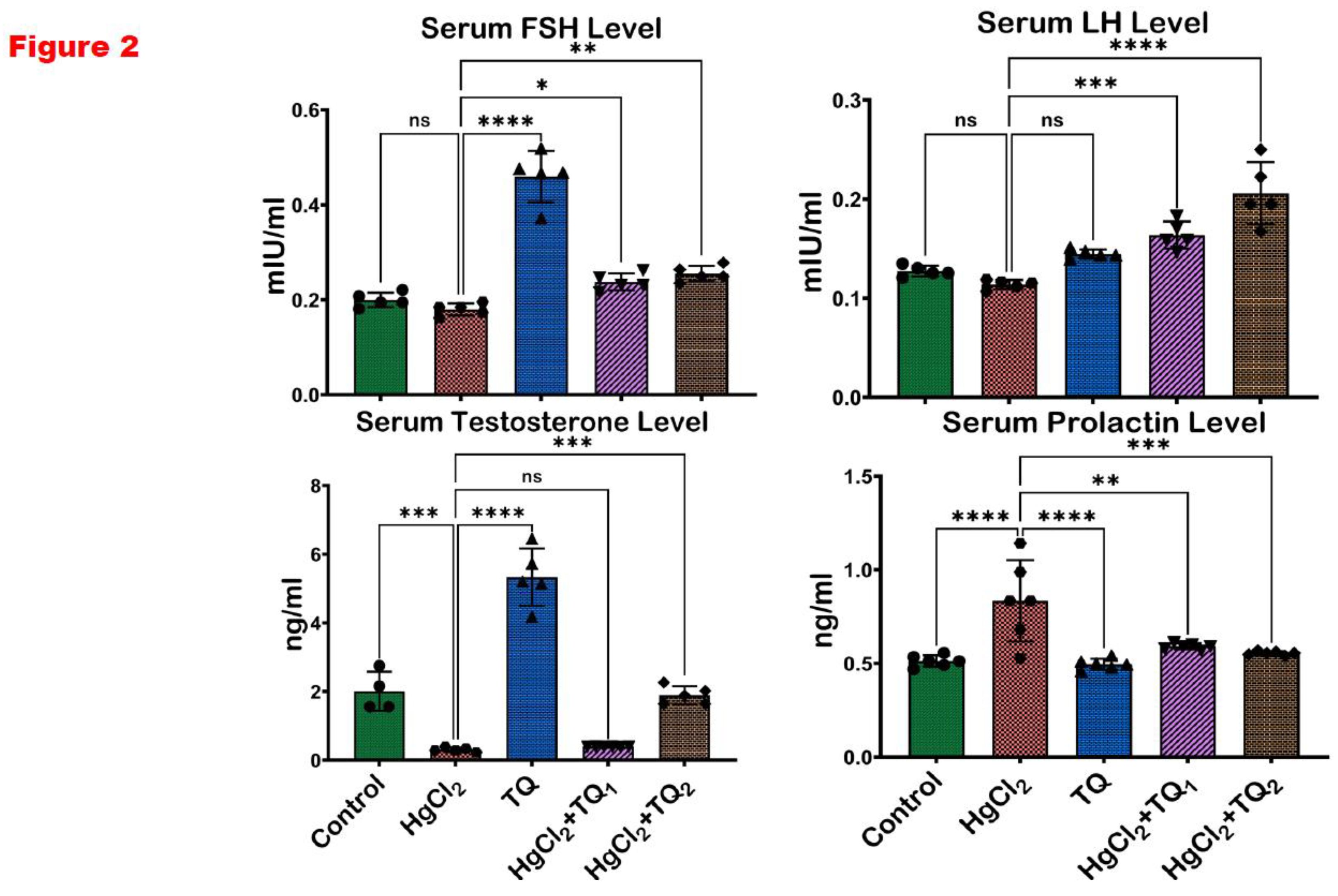
Figure 3.
The effect of Thymoquinone on testicular enzyme activities in rats treated with mercuric chloride for 28 days. Results are shown as mean ± SD (n=8), p < 0.05, 0.01, 0.001, 0.0001 (*, **, ***, ****). ALP: Alkaline phosphatase; ACP: Acid phosphatase; LDH: Lactate Dehydrogenase; G6PD: Glucose-6-phosphate dehydrogenase.
Figure 3.
The effect of Thymoquinone on testicular enzyme activities in rats treated with mercuric chloride for 28 days. Results are shown as mean ± SD (n=8), p < 0.05, 0.01, 0.001, 0.0001 (*, **, ***, ****). ALP: Alkaline phosphatase; ACP: Acid phosphatase; LDH: Lactate Dehydrogenase; G6PD: Glucose-6-phosphate dehydrogenase.
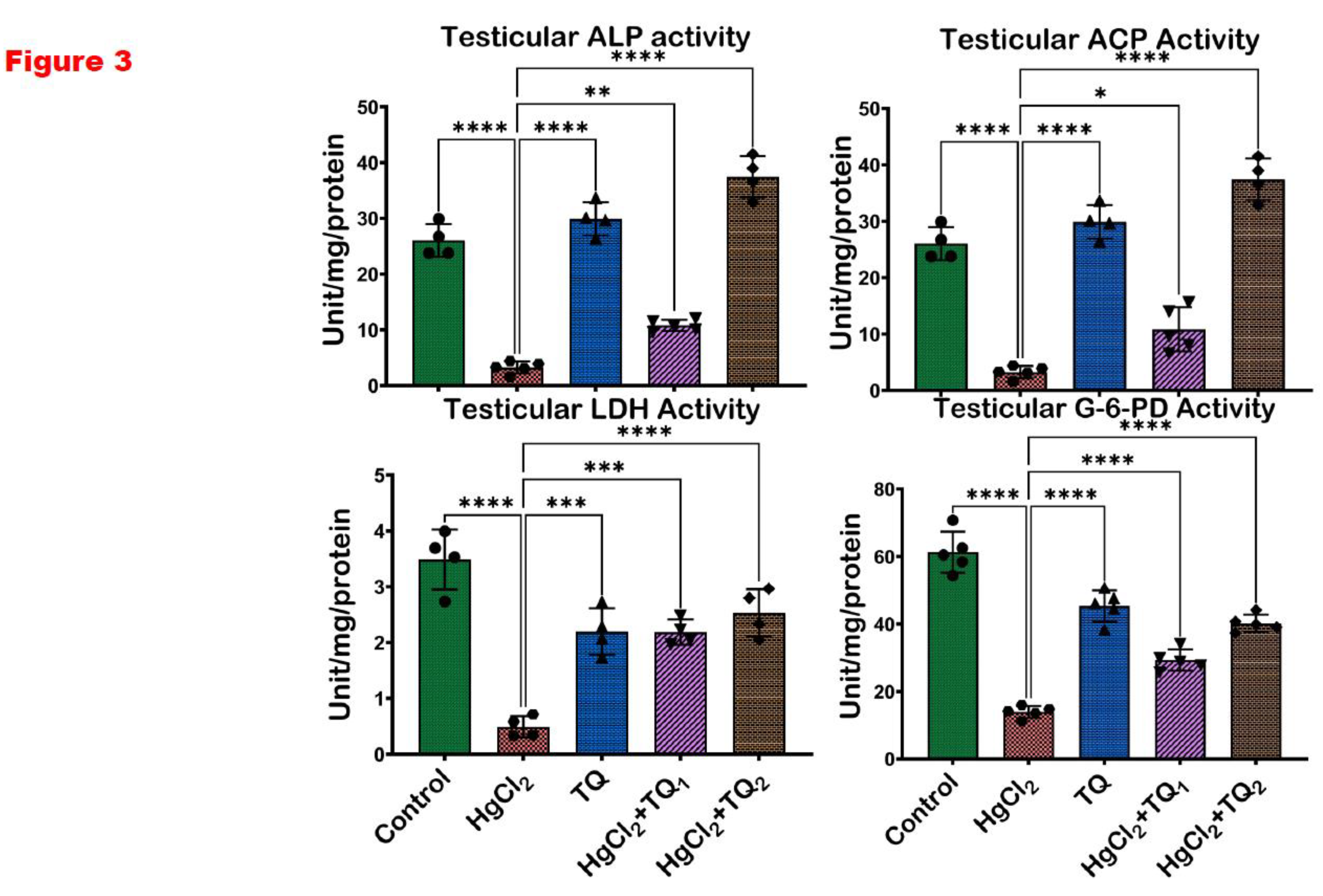
Figure 4.
The impact of Thymoquinone (TQ) on catalase (CAT) and superoxide dismutase activities in the testes, epididymis and hypothalamus of male Wistar rats treated with mercuric chloride for a period of 28 days. Results are shown as mean ± SD (n=8).
Figure 4.
The impact of Thymoquinone (TQ) on catalase (CAT) and superoxide dismutase activities in the testes, epididymis and hypothalamus of male Wistar rats treated with mercuric chloride for a period of 28 days. Results are shown as mean ± SD (n=8).
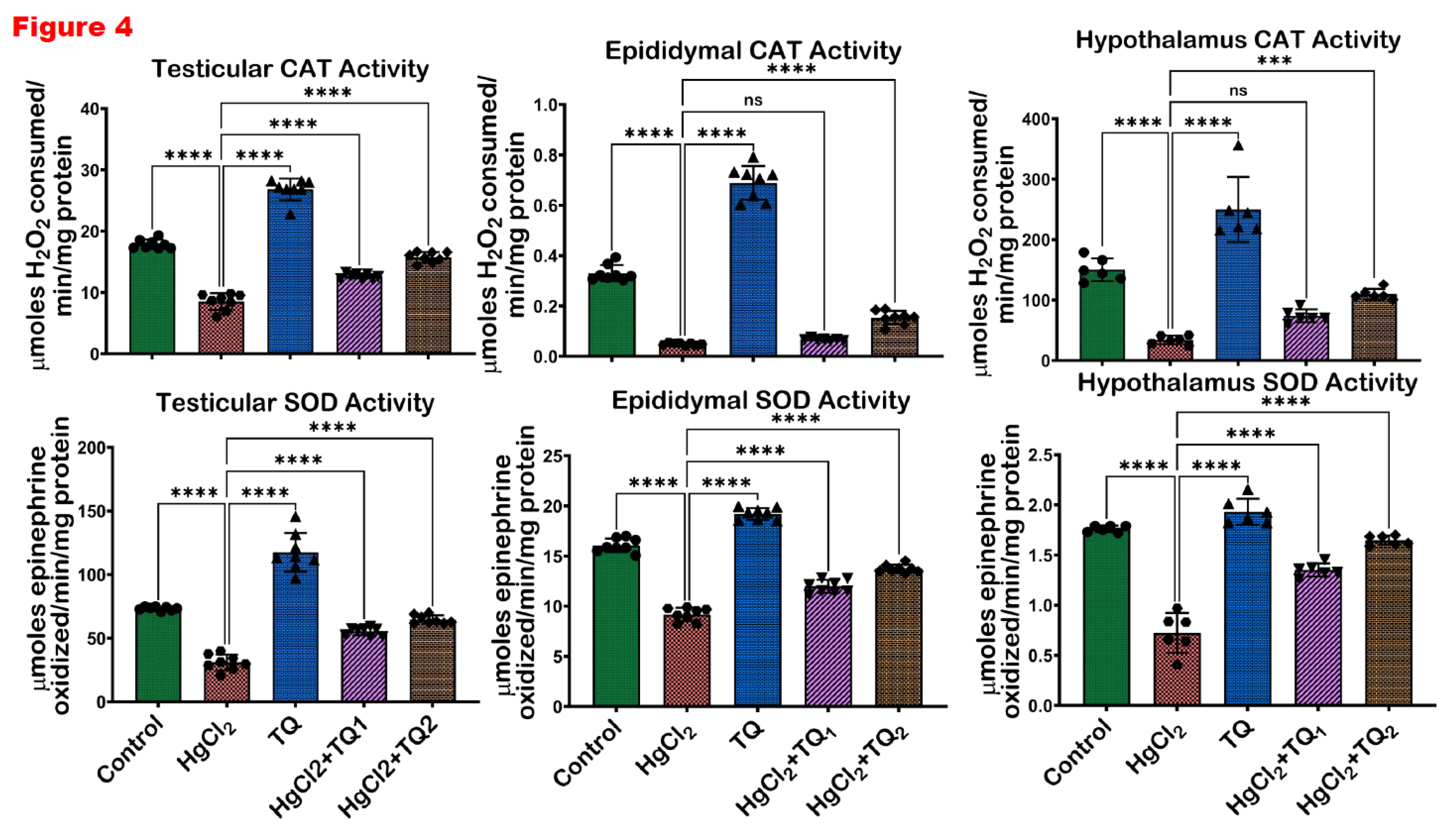
Figure 5.
The outcome of Thymoquinone (TQ) administration on Glutathione peroxidase (GPx) and Glutathione-S-Transferase (GST) activities in the testes, epididymis and hypothalamus of experimental rats treated with mercuric chloride for a period of 28 days. Results are shown as mean ± SD (n=8), p < 0.05, 0.01, 0.001, 0.0001 (*, **, ***, ****). .
Figure 5.
The outcome of Thymoquinone (TQ) administration on Glutathione peroxidase (GPx) and Glutathione-S-Transferase (GST) activities in the testes, epididymis and hypothalamus of experimental rats treated with mercuric chloride for a period of 28 days. Results are shown as mean ± SD (n=8), p < 0.05, 0.01, 0.001, 0.0001 (*, **, ***, ****). .
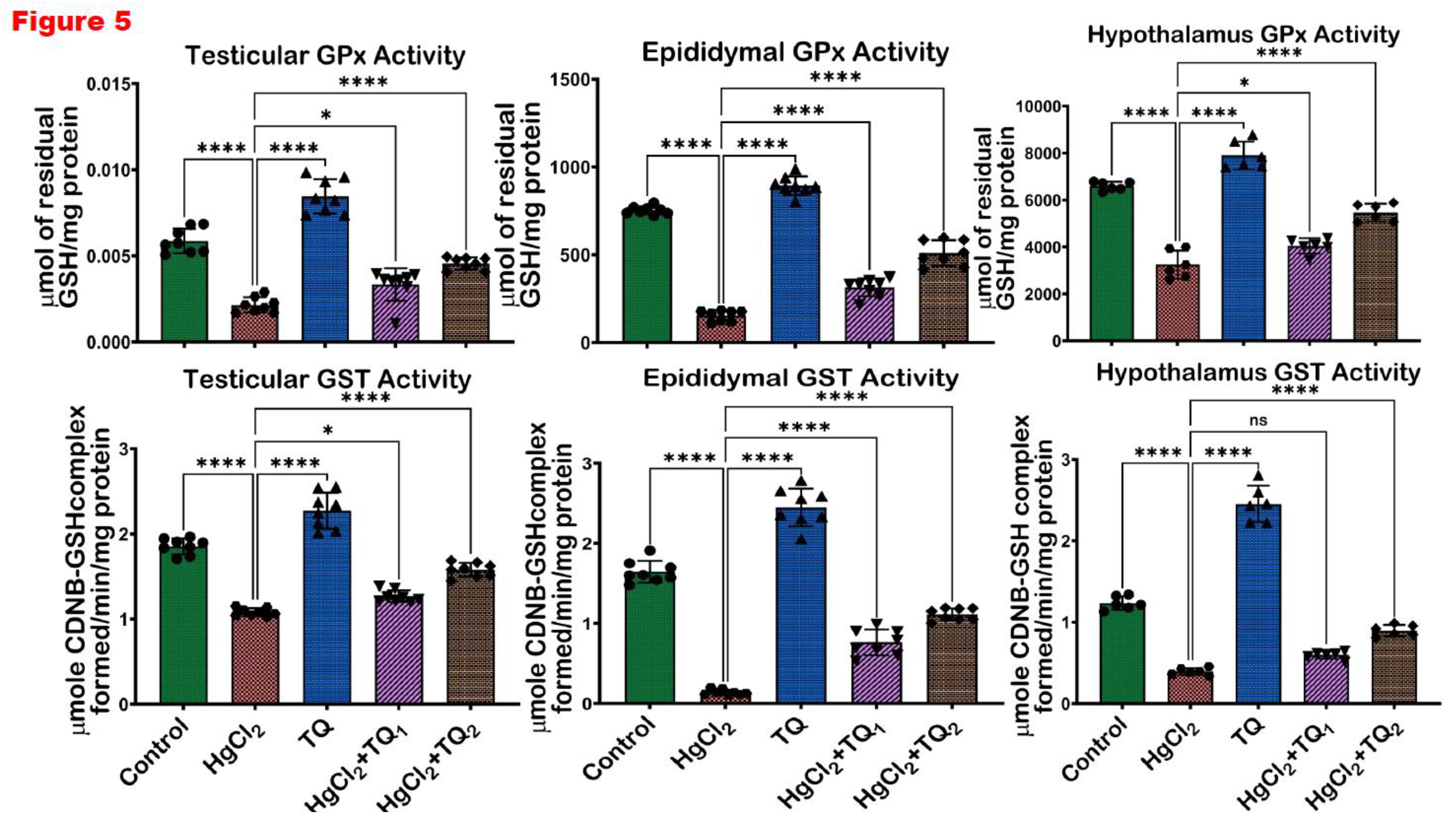
Figure 6.
The impact of Thymoquinone (TQ) treatment on the levels of glutathione (GSH) and total sulfhydryl groups (TSH) in the testes, epididymis and hypothalamus of male Wistar rats administered with mercuric chloride for a period of 28 days. Results are shown as mean ± SD (n=8), p < 0.05, 0.01, 0.001, 0.0001 (*, **, ***, ****).
Figure 6.
The impact of Thymoquinone (TQ) treatment on the levels of glutathione (GSH) and total sulfhydryl groups (TSH) in the testes, epididymis and hypothalamus of male Wistar rats administered with mercuric chloride for a period of 28 days. Results are shown as mean ± SD (n=8), p < 0.05, 0.01, 0.001, 0.0001 (*, **, ***, ****).
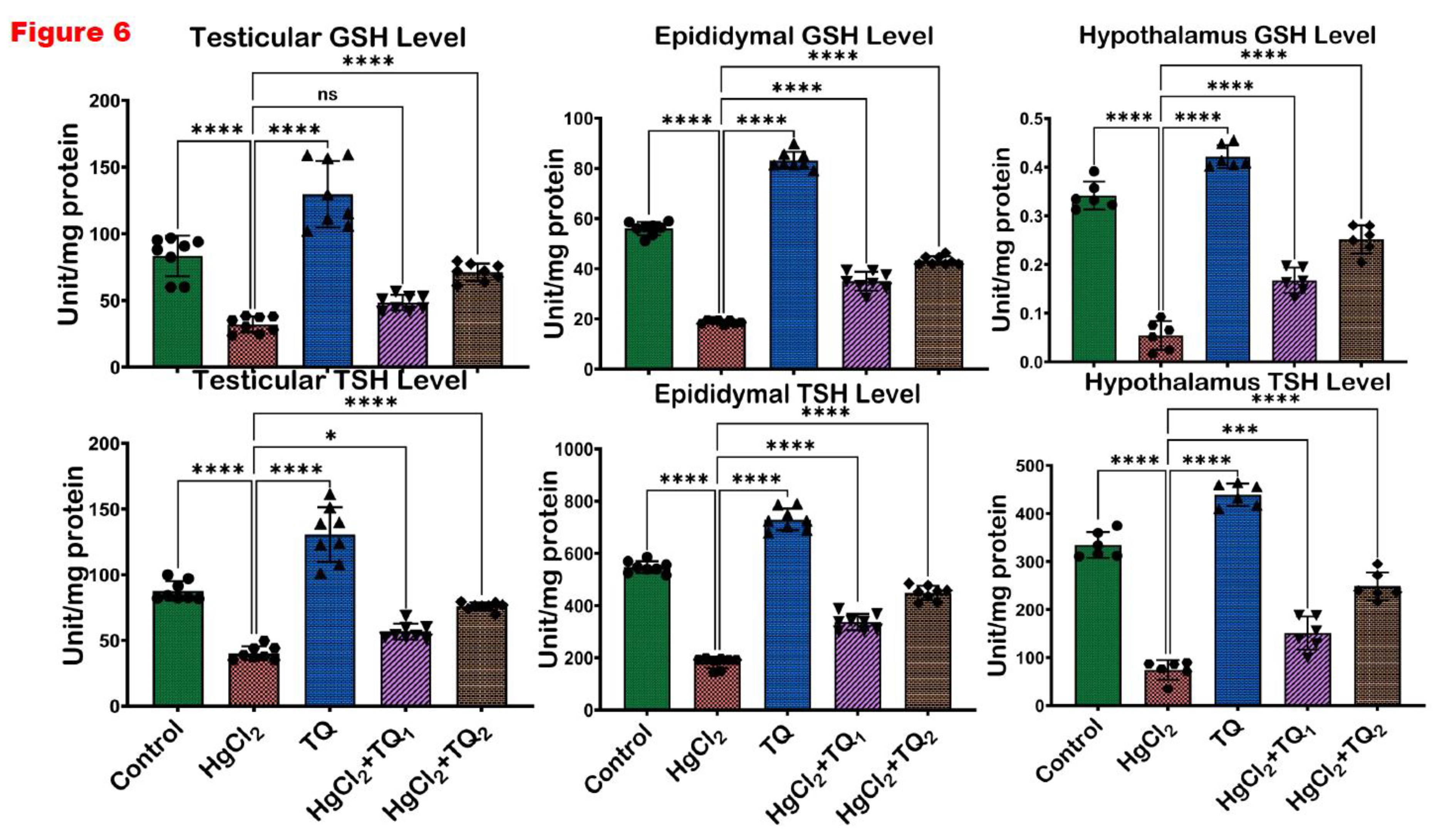
Figure 7.
The effect of Thymoquinone (TQ) on the levels of reactive oxygen and nitrogen species and lipid peroxidation (LPO) in the testes, epididymis and hypothalamus of mercuric chloride-treated male Wistar rats for 28 days. Results are shown as mean ± SD (n=8), p < 0.05, 0.01, 0.001, 0.0001 (*, **, ***, ****). .
Figure 7.
The effect of Thymoquinone (TQ) on the levels of reactive oxygen and nitrogen species and lipid peroxidation (LPO) in the testes, epididymis and hypothalamus of mercuric chloride-treated male Wistar rats for 28 days. Results are shown as mean ± SD (n=8), p < 0.05, 0.01, 0.001, 0.0001 (*, **, ***, ****). .
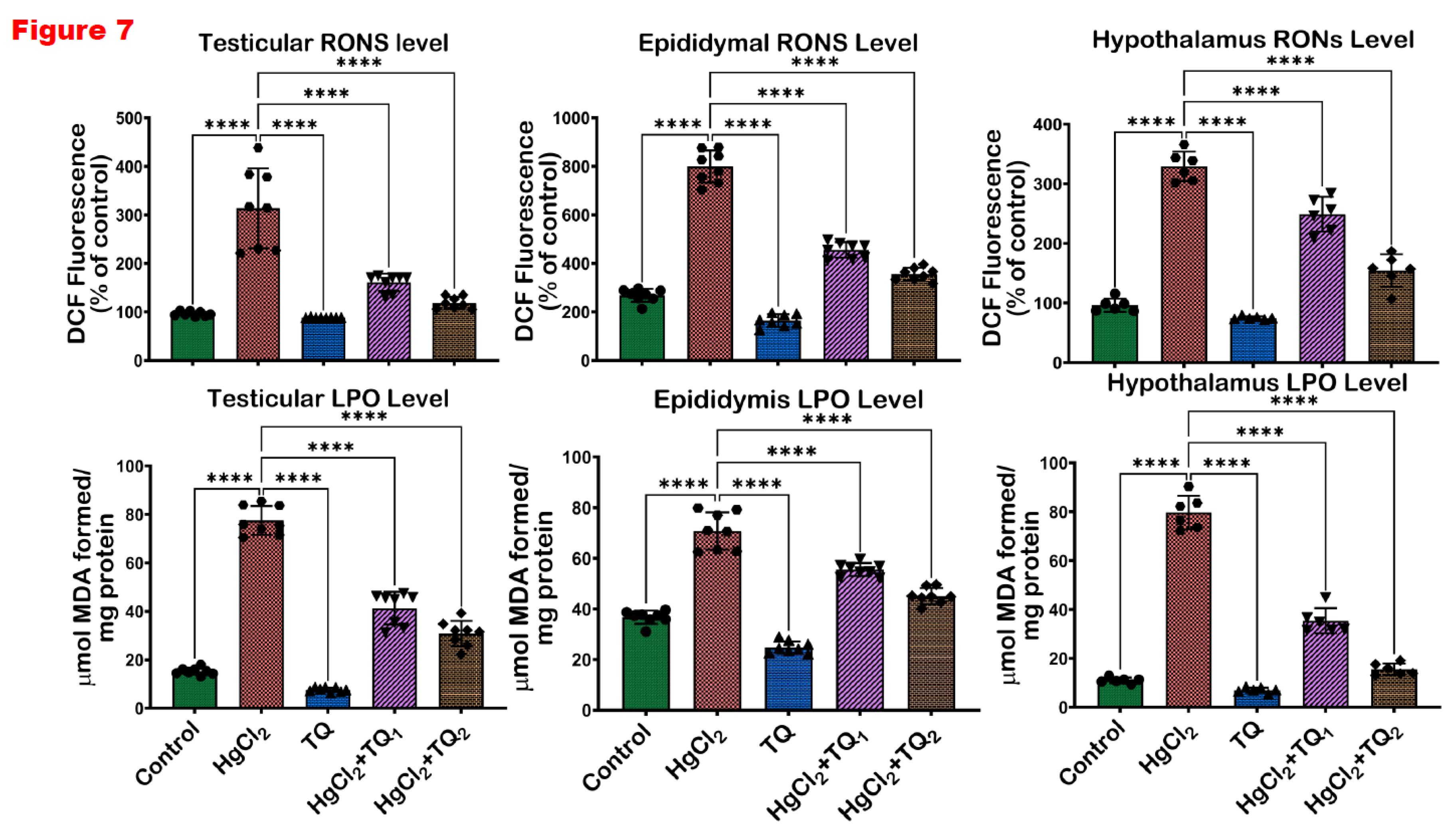
Figure 8.
The effect of Thymoquinone on the activities of xanthine oxidase (XO) and protein carbonyl in the testes, epididymis and hypothalamus of mercuric chloride-treated male Wistar rats for 28 days. Results are shown as mean ± SD (n=8), p < 0.05, 0.01, 0.001, 0.0001 (*, **, ***, ****). .
Figure 8.
The effect of Thymoquinone on the activities of xanthine oxidase (XO) and protein carbonyl in the testes, epididymis and hypothalamus of mercuric chloride-treated male Wistar rats for 28 days. Results are shown as mean ± SD (n=8), p < 0.05, 0.01, 0.001, 0.0001 (*, **, ***, ****). .
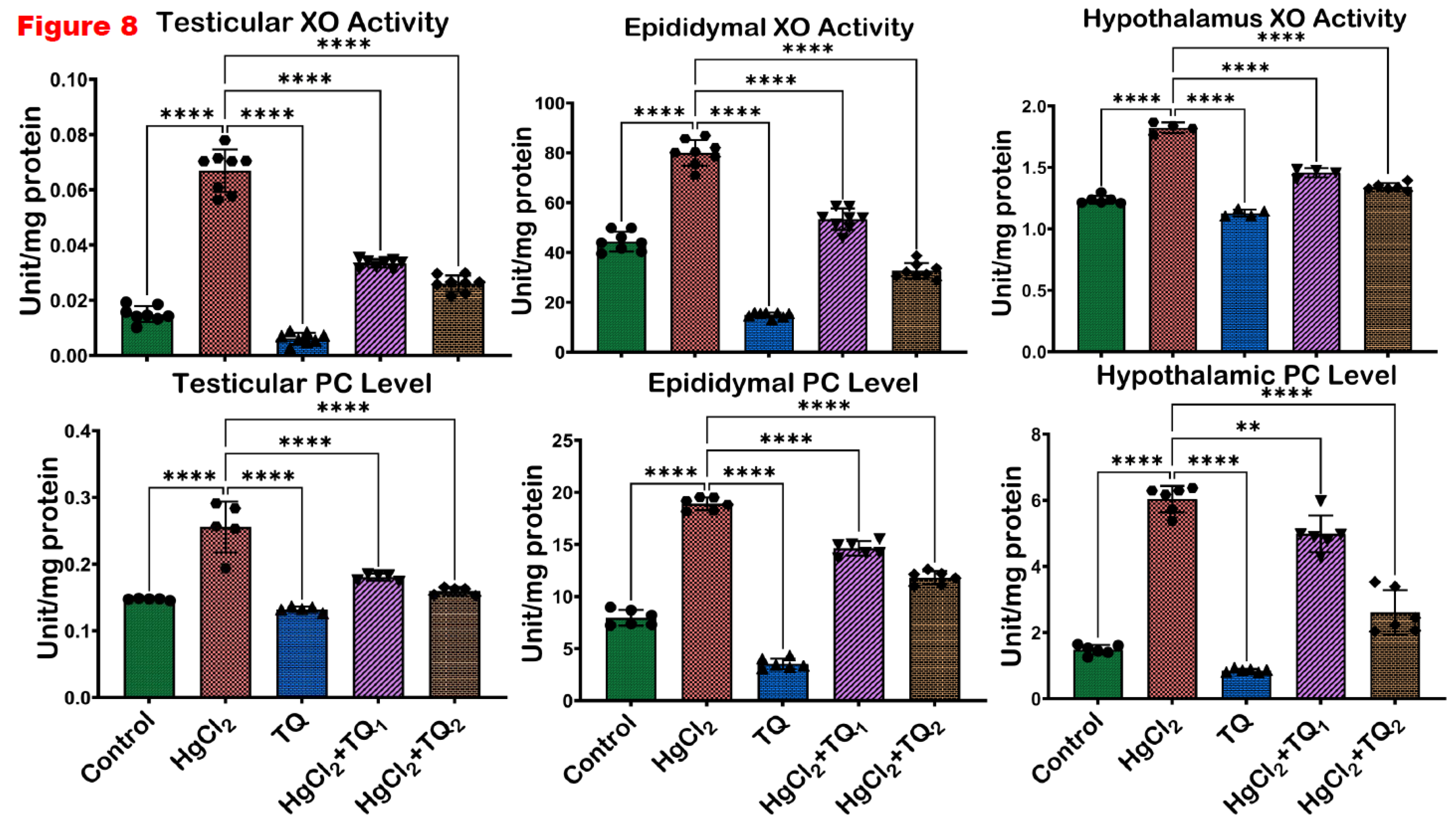
Figure 9.
The result of Thymoquinone (TQ) treatment on biomarkers of inflammation- level of Nitric oxide (NO) and the activities of myeloperoxidase in the testes, epididymis and hypothalamus of mercuric chloride-treated male Wistar rats for a period of 28 days. Results are shown as mean ± SD (n=8), p < 0.05, 0.01, 0.001, 0.0001 (*, **, ***, ****). .
Figure 9.
The result of Thymoquinone (TQ) treatment on biomarkers of inflammation- level of Nitric oxide (NO) and the activities of myeloperoxidase in the testes, epididymis and hypothalamus of mercuric chloride-treated male Wistar rats for a period of 28 days. Results are shown as mean ± SD (n=8), p < 0.05, 0.01, 0.001, 0.0001 (*, **, ***, ****). .
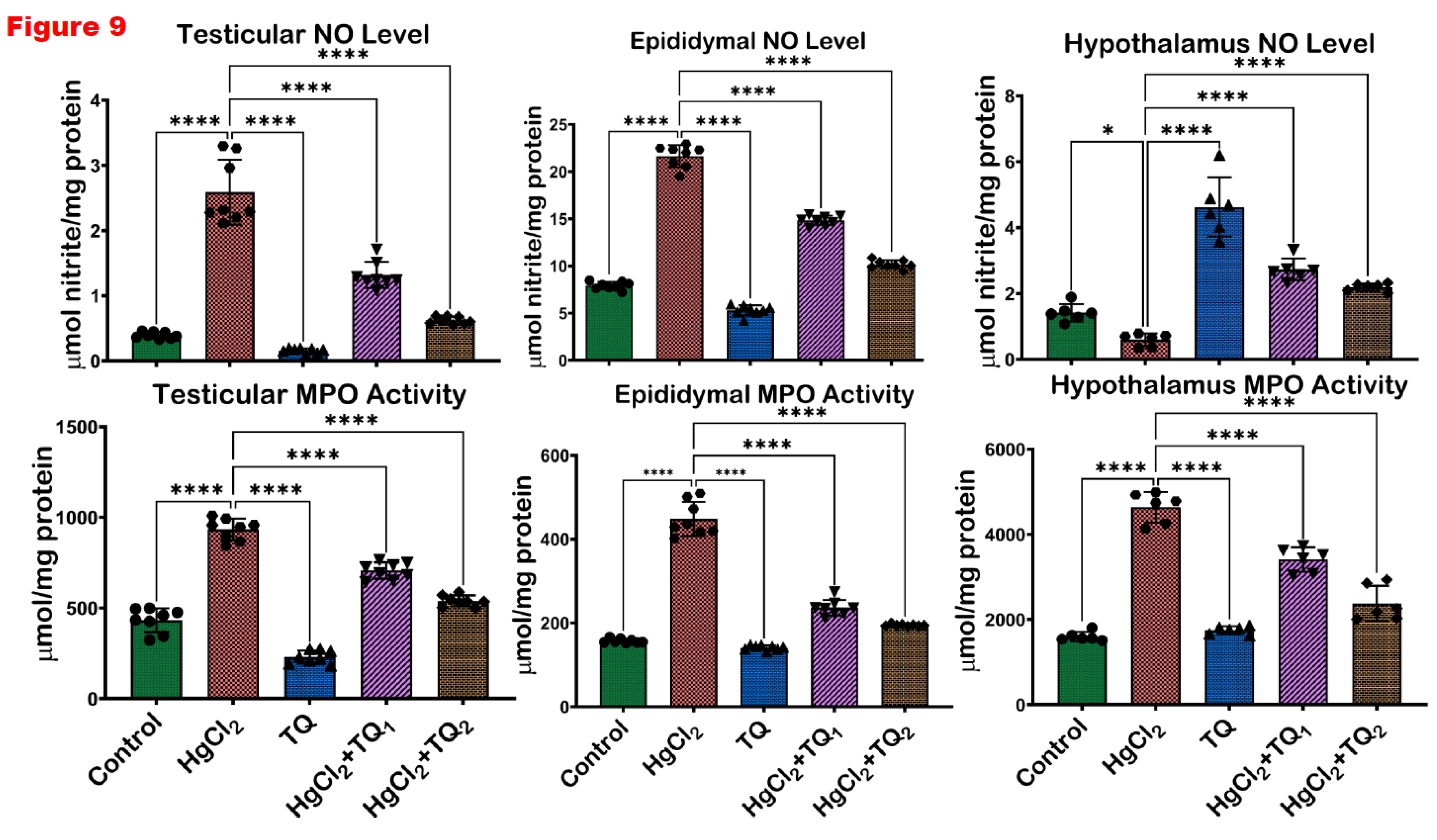
Figure 10.
The influence of Thymoquinone (TQ) administration on biomarkers of inflammation- level of Interleukin-1 beta (IL-1β) and Tumour Necrotic Factor alpha (TNF-α) in the testes, epididymis and hypothalamus of mercuric chloride-treated male Wistar rats for 28 consecutive days. Results are shown as mean ± SD (n=8), p < 0.05, 0.01, 0.001, 0.0001 (*, **, ***, ****). .
Figure 10.
The influence of Thymoquinone (TQ) administration on biomarkers of inflammation- level of Interleukin-1 beta (IL-1β) and Tumour Necrotic Factor alpha (TNF-α) in the testes, epididymis and hypothalamus of mercuric chloride-treated male Wistar rats for 28 consecutive days. Results are shown as mean ± SD (n=8), p < 0.05, 0.01, 0.001, 0.0001 (*, **, ***, ****). .
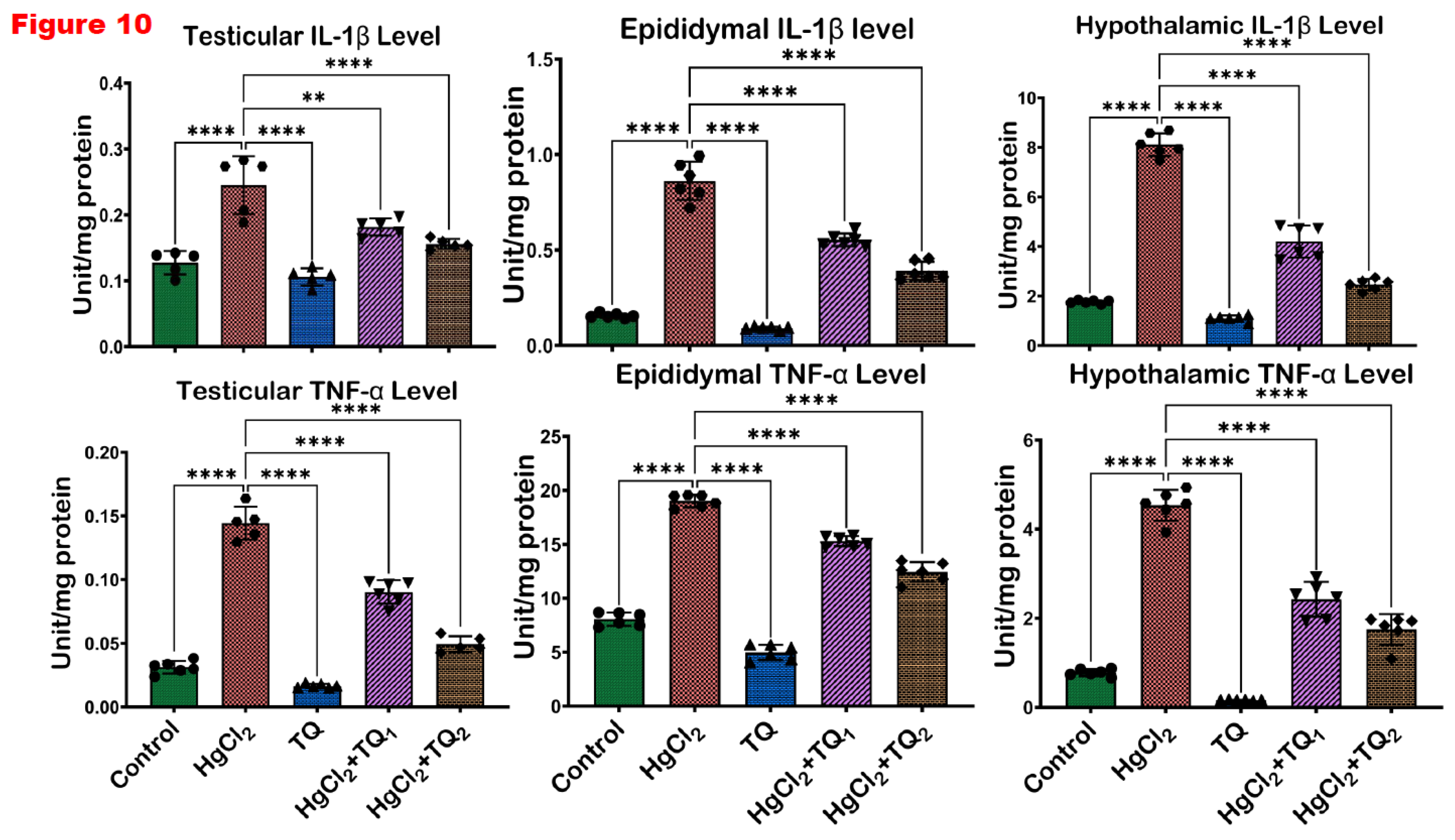
Figure 11.
The influence of Thymoquinone (TQ) administration on the levels of apoptotic markers- Bax and Bcl-2 in the testes, epididymis and hypothalamus of male Wistar rats treated with mercuric chloride for a period of 28 days. Results are shown as mean ± SD (n=8), p < 0.05, 0.01, 0.001, 0.0001 (*, **, ***, ****). .
Figure 11.
The influence of Thymoquinone (TQ) administration on the levels of apoptotic markers- Bax and Bcl-2 in the testes, epididymis and hypothalamus of male Wistar rats treated with mercuric chloride for a period of 28 days. Results are shown as mean ± SD (n=8), p < 0.05, 0.01, 0.001, 0.0001 (*, **, ***, ****). .
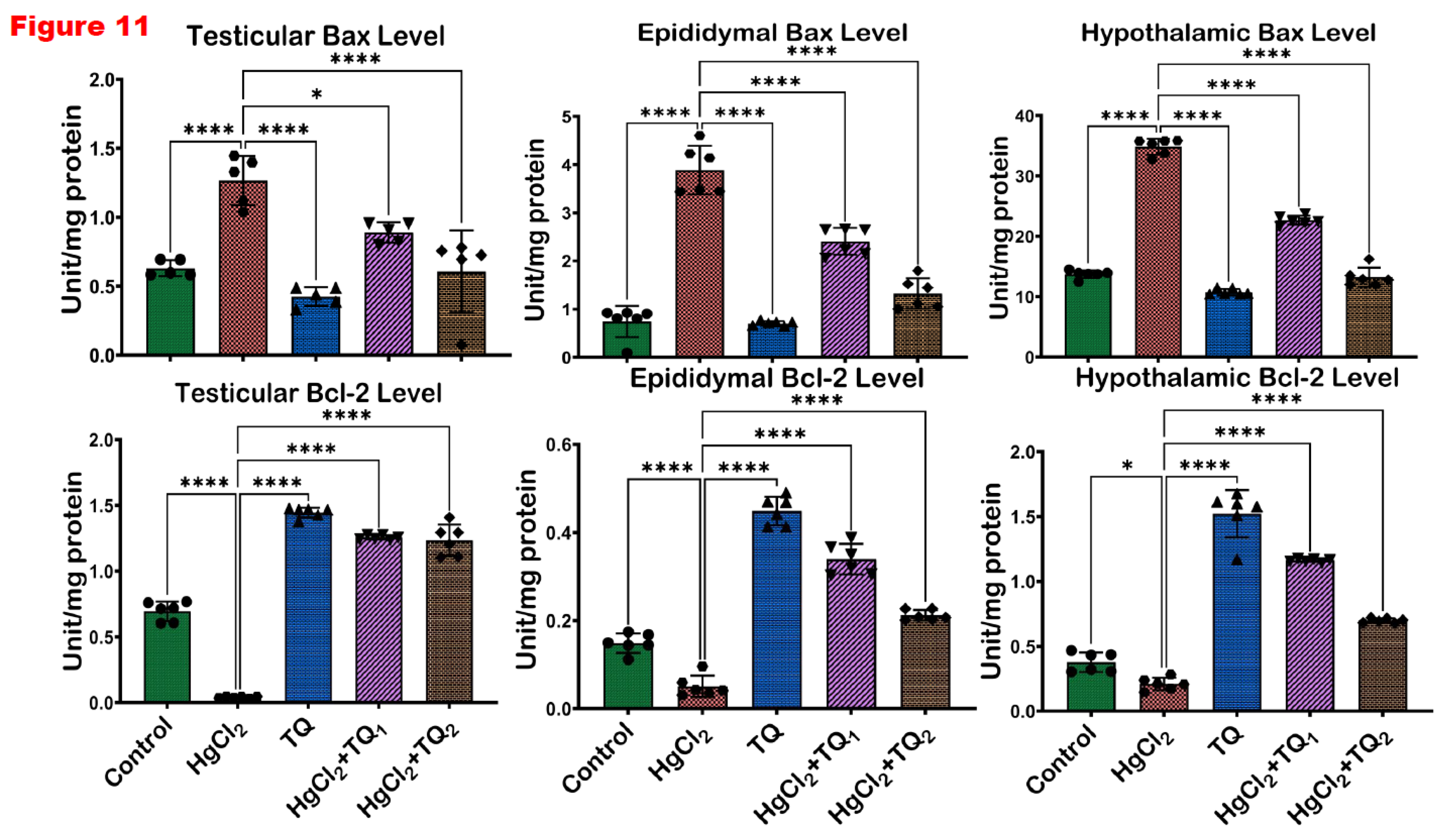
Figure 12.
The effect of Thymoquinone (TQ) on the level of Protein 53 (P53), a marker of DNA damage in the testes, epididymis and hypothalamus of mercuric chloride-treated male Wistar rats for 28 days. Results are shown as mean ± SD (n=8), p < 0.05, 0.01, 0.001, 0.0001 (*, **, ***, ****). .
Figure 12.
The effect of Thymoquinone (TQ) on the level of Protein 53 (P53), a marker of DNA damage in the testes, epididymis and hypothalamus of mercuric chloride-treated male Wistar rats for 28 days. Results are shown as mean ± SD (n=8), p < 0.05, 0.01, 0.001, 0.0001 (*, **, ***, ****). .
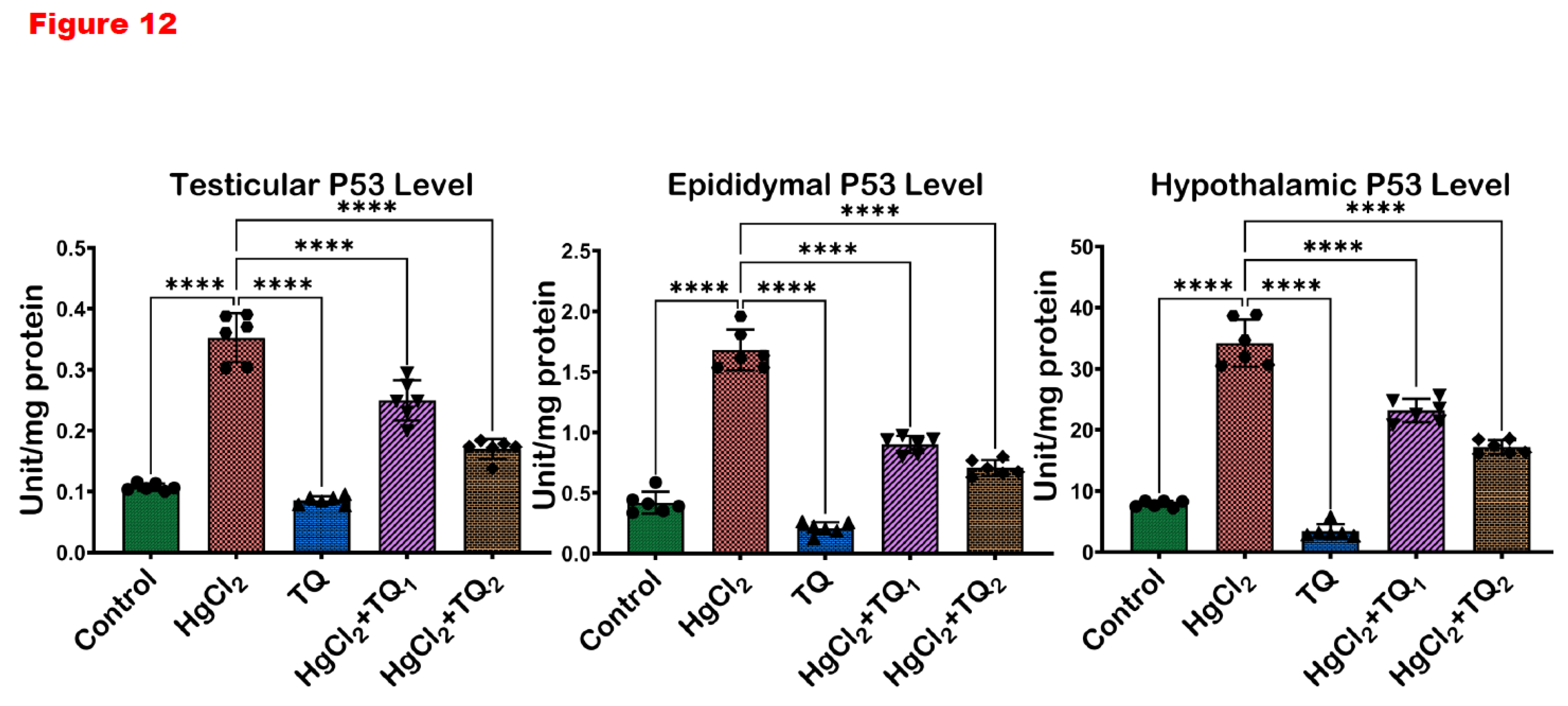
Figure 13.
Histological representation of the effect of thymoquinone on testicular damage caused by mercury chloride. The controls and TQ-treated groups show typical testicular structure with well-organised testicular tubules and germinal epithelium. HgCl2-only exposed rat shows testicular degeneration with vacuolation of the seminiferous tubule and severe loss of germinal epithelium. Each bar represents mean values with asterisks indicating significant differences at *p<0.05, **p<0.01, and ***p<0.001 versus control and HgCl2 from histomorphometric data. Thymoquinone (TQ), mercury chloride (HgCl2). H & E section of the testis, scale bar=50µm, magnification: x100 (top row) and x400 (bottom row). .
Figure 13.
Histological representation of the effect of thymoquinone on testicular damage caused by mercury chloride. The controls and TQ-treated groups show typical testicular structure with well-organised testicular tubules and germinal epithelium. HgCl2-only exposed rat shows testicular degeneration with vacuolation of the seminiferous tubule and severe loss of germinal epithelium. Each bar represents mean values with asterisks indicating significant differences at *p<0.05, **p<0.01, and ***p<0.001 versus control and HgCl2 from histomorphometric data. Thymoquinone (TQ), mercury chloride (HgCl2). H & E section of the testis, scale bar=50µm, magnification: x100 (top row) and x400 (bottom row). .
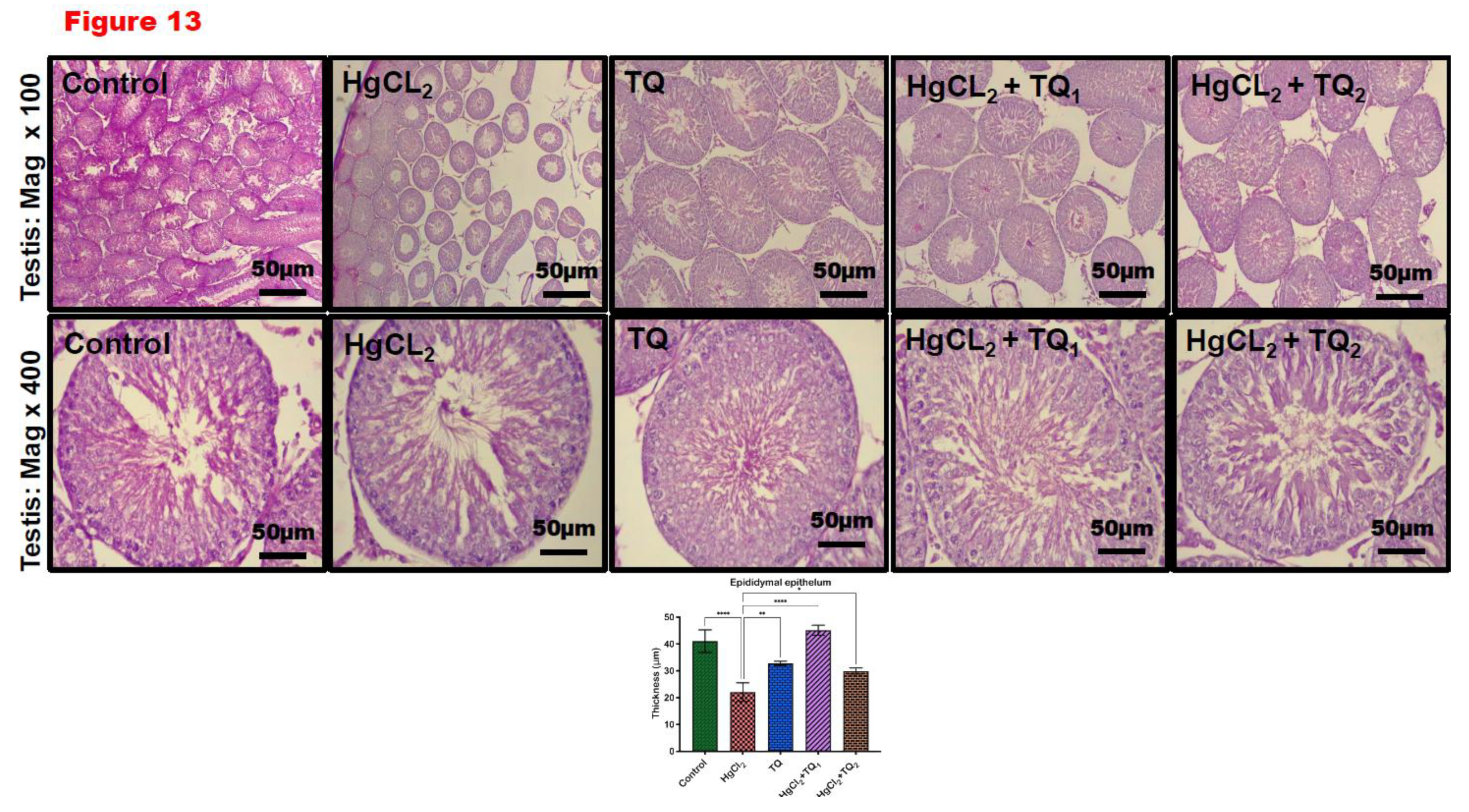
Figure 14.
Epididymal section of thymoquinone and mercury chloride. The control and TQ-treated groups show typical testicular structures with well-organised epididymal epithelium and mature stored spermatozoa. HgCl2-only exposed rat shows epididymal degeneration with fewer stored mature spermatozoa. Each bar represents mean values with asterisks indicating significant differences at *p<0.05, **p<0.01, ***p<0.001 and ****p<0.0001 versus control and HgCl2 from histomorphometric data. Thymoquinone (TQ), mercury chloride (HgCl2). H & E section of the testis, scale bar=50µm, magnification: x100 (top row) and x400 (bottom row). .
Figure 14.
Epididymal section of thymoquinone and mercury chloride. The control and TQ-treated groups show typical testicular structures with well-organised epididymal epithelium and mature stored spermatozoa. HgCl2-only exposed rat shows epididymal degeneration with fewer stored mature spermatozoa. Each bar represents mean values with asterisks indicating significant differences at *p<0.05, **p<0.01, ***p<0.001 and ****p<0.0001 versus control and HgCl2 from histomorphometric data. Thymoquinone (TQ), mercury chloride (HgCl2). H & E section of the testis, scale bar=50µm, magnification: x100 (top row) and x400 (bottom row). .
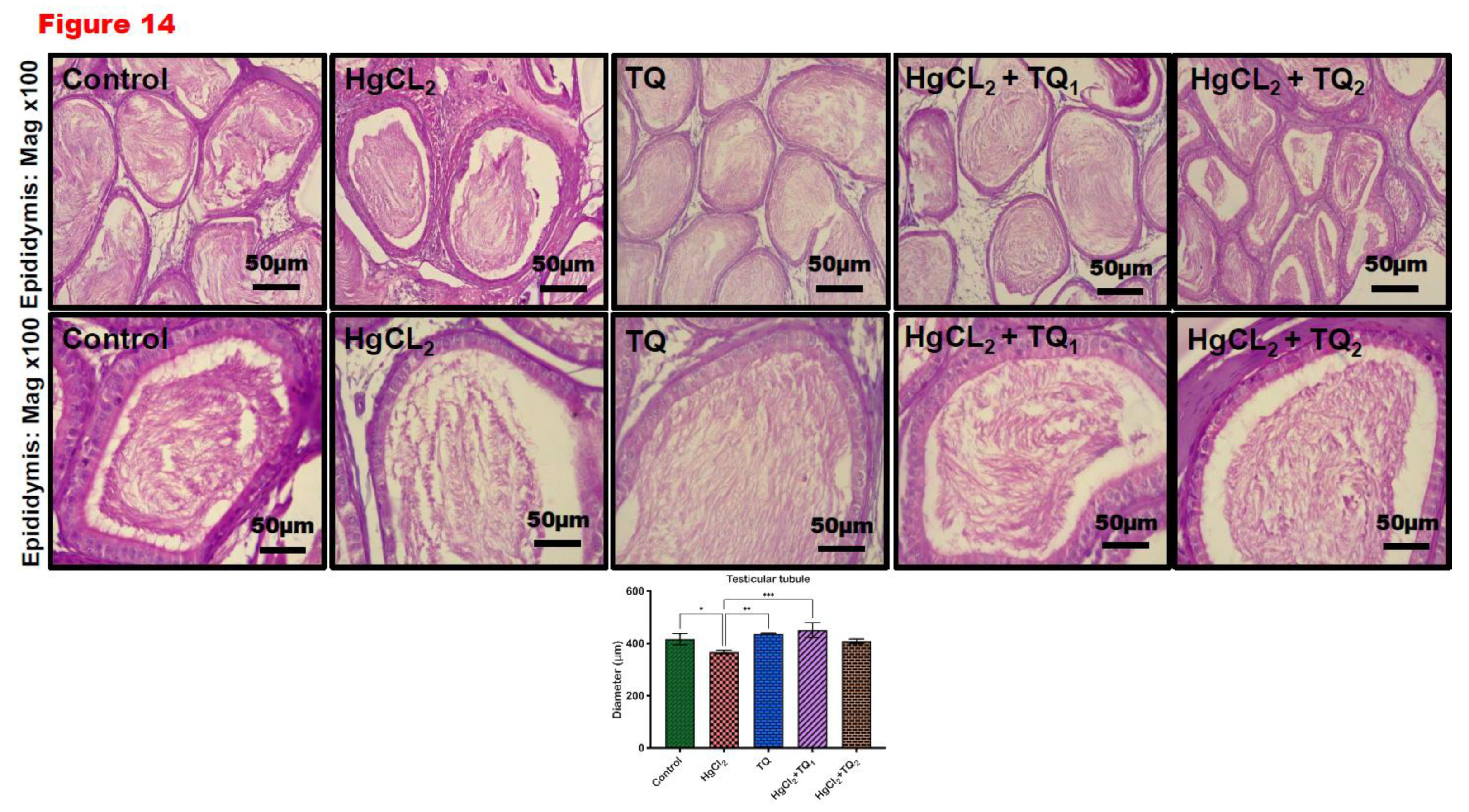
Figure 15.
A: Molecular docking illustration showing the binding of thymoquinone with PPAR-α (PDB: 1I7G) and PPAR- δ/β (PDB: 3D5F) with ΔG values of –5.9 Kcal/mol and –6.2 Kcal/mol. B: TQ can activate PPAR-α or PPAR-δ/β to mediate anti-oxidant effect or resolve inflammation. Specifically, PPAR-α blocks the activation of NF-kB, thereby preventing the expression of pro-inflammatory cytokines. Additionally, PPAR-δ/β induces an antioxidant effect by activating Nrf-2, which upregulates the expression of antioxidant genes.
Figure 15.
A: Molecular docking illustration showing the binding of thymoquinone with PPAR-α (PDB: 1I7G) and PPAR- δ/β (PDB: 3D5F) with ΔG values of –5.9 Kcal/mol and –6.2 Kcal/mol. B: TQ can activate PPAR-α or PPAR-δ/β to mediate anti-oxidant effect or resolve inflammation. Specifically, PPAR-α blocks the activation of NF-kB, thereby preventing the expression of pro-inflammatory cytokines. Additionally, PPAR-δ/β induces an antioxidant effect by activating Nrf-2, which upregulates the expression of antioxidant genes.
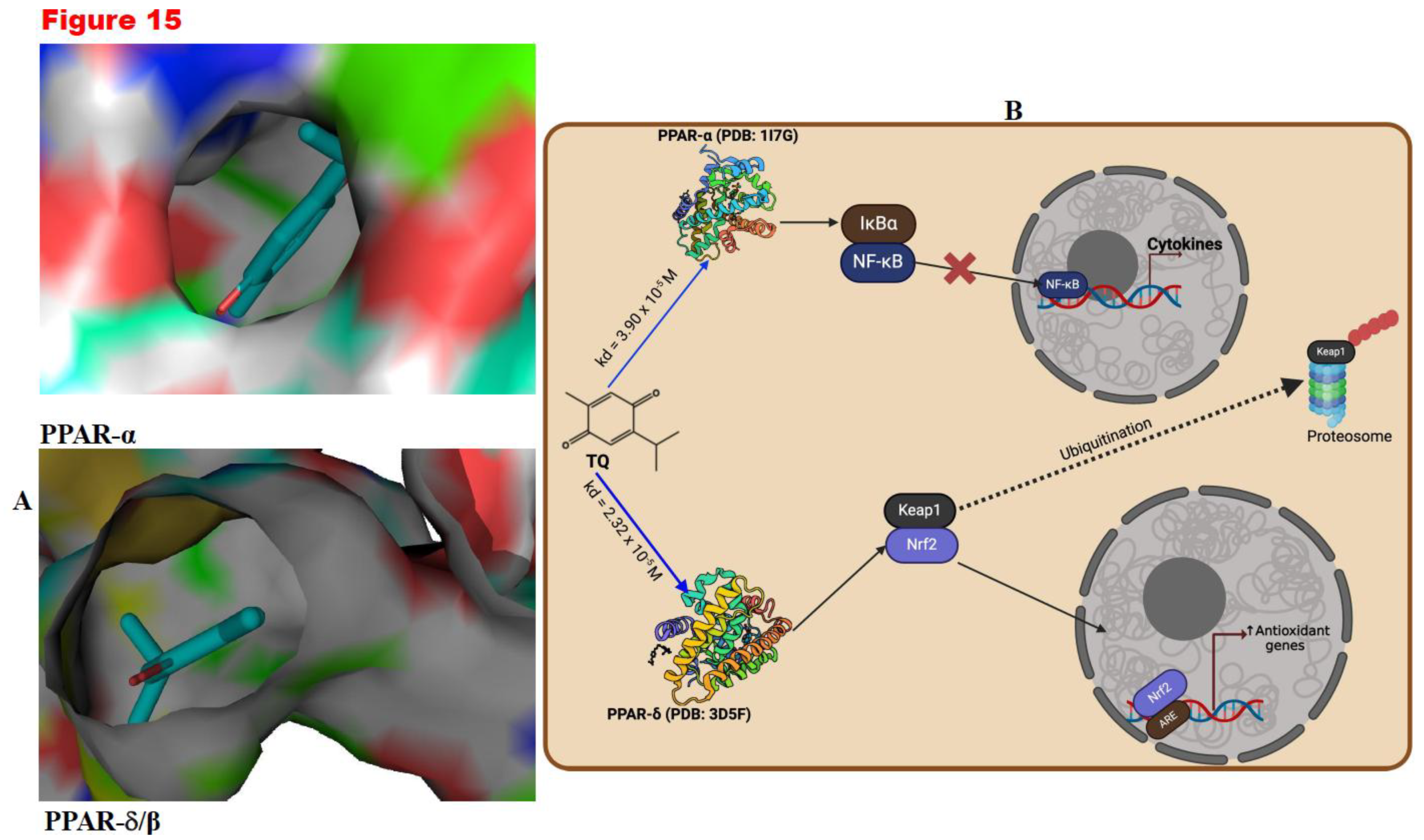
Figure 16.
Proposed mechanism of action indicating how Thymoquinone (TQ) suppresses mercuric chloride-alteration of male Wistar rats' testes, epididymis and hypothalamus after a 28-day study. Mercuric chloride is obtained by the action of chlorine on mercury or mercury(I)chloride. It can also be produced by the addition of hydrochloric acid to a hot, concentrated solution of mercury(I) compounds such as the nitrate. The activation of mitogen-activated protein kinase (MAPK), nuclear factor erythroid 2-related factor 2 (Nrf2), and the formation of DNA adducts are part of the cellular responses to stress and damage caused by mercury exposure. While activated, NF-kB mediates pro-inflammatory gene transcription, leading to inflammation. DNA adducts upregulate the expression of p53, which upregulates the expression of PUMA. PUMA then activates Bax. Bax then enters the mitochondria and induces MPTP, releasing cytochrome C. Cytochrome C, combined with Apaf1 and procaspase-9, forms the apoptosome, converting procaspase-9 into active Casp-9. Caspase-9 then activates procaspase-3 to active caspase-3, eventually executing programmed cell death. However, the action of thymoquinone inhibits inflammation and the formation of DNA adducts. It promotes the expression of detoxification enzymes, such as heme oxygenase-1 (HO-1), glutathione S-transferase (GST) and other antioxidant enzymes from the activation of Nrf2. Created with https://app.biorender.com/ by Uche Arunsi.
Figure 16.
Proposed mechanism of action indicating how Thymoquinone (TQ) suppresses mercuric chloride-alteration of male Wistar rats' testes, epididymis and hypothalamus after a 28-day study. Mercuric chloride is obtained by the action of chlorine on mercury or mercury(I)chloride. It can also be produced by the addition of hydrochloric acid to a hot, concentrated solution of mercury(I) compounds such as the nitrate. The activation of mitogen-activated protein kinase (MAPK), nuclear factor erythroid 2-related factor 2 (Nrf2), and the formation of DNA adducts are part of the cellular responses to stress and damage caused by mercury exposure. While activated, NF-kB mediates pro-inflammatory gene transcription, leading to inflammation. DNA adducts upregulate the expression of p53, which upregulates the expression of PUMA. PUMA then activates Bax. Bax then enters the mitochondria and induces MPTP, releasing cytochrome C. Cytochrome C, combined with Apaf1 and procaspase-9, forms the apoptosome, converting procaspase-9 into active Casp-9. Caspase-9 then activates procaspase-3 to active caspase-3, eventually executing programmed cell death. However, the action of thymoquinone inhibits inflammation and the formation of DNA adducts. It promotes the expression of detoxification enzymes, such as heme oxygenase-1 (HO-1), glutathione S-transferase (GST) and other antioxidant enzymes from the activation of Nrf2. Created with https://app.biorender.com/ by Uche Arunsi.
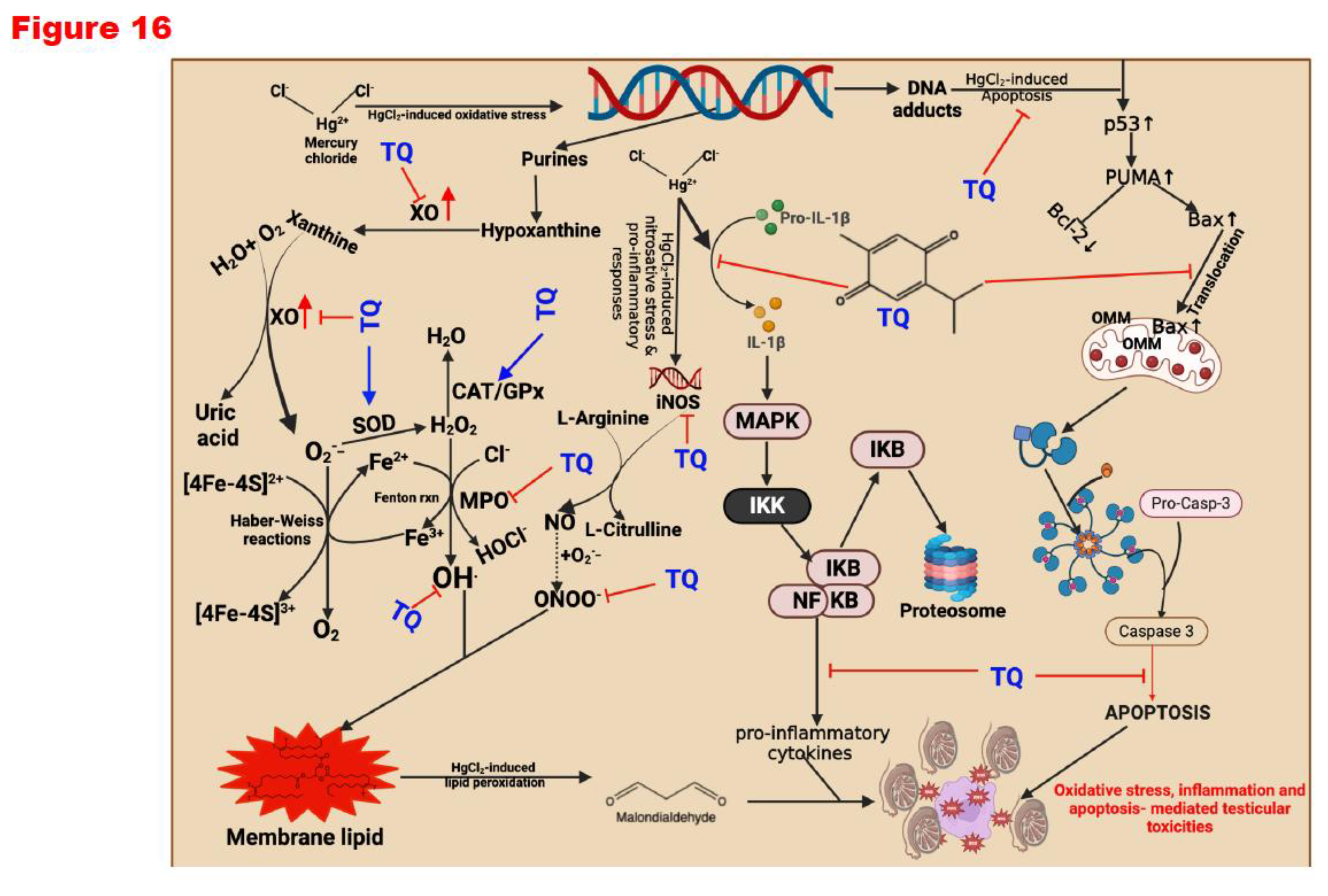

Table 1.
Materials Used for the Study.
| Chemical name | Catalogue No. | Company |
| 2’,7’-dichlorodihydrofluorescin diacetate | 4091-99-0 | Sigma-Aldrich Inc. (St Louis, MO, USA) |
| 5,5’-dithiobis-(2-nitrobenzoic acid) | 69-78-3 | Sigma-Aldrich Inc. (St Louis, MO, USA) |
| Copper sulfate pentahydrate | 7758-99-8 | Sigma-Aldrich Inc. (St Louis, MO, USA) |
| Dipotassium hydrogen phosphate trihydrate | 7758-11-4 | AK Scientific, Union City, USA |
| Epinephrine | 51-43-4 | Sigma-Aldrich Inc. (St Louis, MO, USA) |
| Folin-Ciocalteu reagent | 125629 | J.T Baker (Phillisburg, PH, USA) |
| Glucose 6-phosphate dehydrogenase (G6PD) | 9001-40-5 | Elabscience Biotechnology Company (Wuhan, China) |
| Hydrogen peroxide (H2O2) | 7722-84-1 | Sigma-Aldrich Inc. (St Louis, MO, USA) |
| Hydrochloric acid | 7647-01-0 | Sigma-Aldrich Inc. (St Louis, MO, USA) |
| Mercury chloride | AK Scientific, Union City, USA | |
| O-Dianisidine | 119-90-4 | Sigma-Aldrich Inc. (St Louis, MO, USA) |
| Potassium Chloride | 7447-40-7 | AK Scientific, Union City, USA |
| Potassium dihydrogen phosphate | 7778-77-0 | AK Scientific, Union City, USA |
| Reduced glutathione (GSH) | 70-18-8 | Sigma-Aldrich Inc. (St Louis, MO, USA) |
| Sodium azide | 26628-22-8 | Sigma-Aldrich Inc. (St Louis, MO, USA) |
| Sodium hydroxide | 1310-73-2 | Sigma-Aldrich Inc. (St Louis, MO, USA) |
| Sodium-Potassium tartrate | 6381-59-5 | Sigma-Aldrich Inc. (St Louis, MO, USA) |
| Sulphosalicylic acid | 5965-83-3 | Sigma-Aldrich Inc. (St Louis, MO, USA) |
| Trichloroacetic acid | 76-03-9 | Sigma-Aldrich Inc. (St Louis, MO, USA) |
| Thiobarbituric acid (TBA) | 504-17-6 | Sigma-Aldrich Inc. (St Louis, MO, USA) |
| Thymoquinone | 490-91-5 | Merck KGaA, Darmstadt, Germany. |
| Trichloroacetic acid | 76-03-9 | Molychem, Mumbai India |
| Xanthine | 69-89-6 | Sigma-Aldrich Inc. (St Louis, MO, USA) |
Table 2.
Body weight gain and relative Testis, Epididymis, and Hypothalamus weight of rats following exposure to HgCl2 for 28 days.
Table 2.
Body weight gain and relative Testis, Epididymis, and Hypothalamus weight of rats following exposure to HgCl2 for 28 days.
|
*Total rats per grouping |
Control (8) |
HgCl2 (8) |
TQ (8) |
HgCl2+ TQ1 (8) |
HgCl2+ TQ2 (8) |
| Body weight gain (g) | 60.86 ± 8.07 | 57.00 ± 12.45 | 61.29 ± 14.20 | 65.25 ± 10.26 | 72.63 ± 16.18 |
| Testes weight (g) | 2.50 ± 0.12 | 2.32 ± 0.07 | 2.52 ± 0.13 | 2.63 ± 0.17 | 2.73 ± 0.22 |
| Relative Testes weight (%) | 1.18 ± 0.18 | 1.09 ± 0.08 | 1.14 ± 0.13 | 1.18 ± 0.09 | 1.21 ± 0.10 |
| Epididymis weight (g) | 0.29 ± 0.01 | 0.29 ± 0.01 | 0.35 ± 0.04 | 0.30 ± 0.03 | 0.34 ± 0.05 |
| Relative Epididymis weight (%) | 0.14 ± 0.01 | 0.14 ± 0.01 | 0.16 ± 0.02 | 0.14 ± 0.01 | 0.14 ± 0.01 |
| Hypothalamus Weight (g) | 0.09 ± 0.01 | 0.07 ± 0.03 | 0.05 ± 0.01 | 0.08 ± 0.02 | 0.07 ± 0.02 |
| Relative Hypothalamus Weight (%) | 0.04 ± 0.01 | 0.02 ± 0.01 | 0.03 ± 0.01 | 0.03 ± 0.01 | 0.04 ± 0.01 |
HgCl2 (20µg/L dissolved in drinking water); TQ1 and TQ2 (2.5 and 5.0 mg/Kg respectively) body weight; n=8. Data are expressed as mean ± SD. *: p<0.05 versus Control; #: p<0.05 versus HgCl2 alone. HgCl2: Mercuric chloride; TQ: Thymoquinone.
Table 3.
Sperm analysis and sperm abnormalities of rats following exposure to HgCl2 for 28 days.
|
*Total rats per grouping |
Control (8) |
HgCl2 (8) |
TQ (8) |
HgCl2+ TQ1 (8) |
HgCl2+ TQ2 (8) |
| Sperm Functional Analysis | |||||
| Motility | 90.00±4.62 | 75.00±5.34**** | 72.50±4.62 | 67.50±4.62* | 65.00±5.34** |
| Viability | 96.50±1.60 | 96.13±1.55 | 96.50±1.60 | 96.50±1.64 | 94.88±4.25 |
| Sperm Volume | 5.16±0.05 | 5.17±0.04 | 5.17±0.04 | 5.18±0.03 | 5.18±0.05 |
| Epididymal Sperm Count | 132.40±9.89 | 117.00±11.10* | 113.00±8.55 | 101.90±8.25* | 98.50±10.61** |
| Sperm Abnormalities | |||||
| Abnormality of the Head (%) | 2.08±0.27 | 2.14±0.13 | 2.15±0.30 | 2.23±0.31 | 2.17±0.53 |
| Abnormality of the Mid-piece (%) | 4.21±0.38 | 4.65±0.25 | 4.67±0.35 | 4.86±0.38 | 5.14±0.25 |
| Abnormality of the Tail (%) | 5.23±0.45 | 5.61±0.53 | 5.56±0.56 | 5.88±0.93 | 6.25±0.48 |
| Total Abnormality (%) | 11.52±0.44 | 12.41±0.63 | 12.38±0.63 | 12.98±1.28 | 13.56±1.10 |
HgCl2 (20µg/L dissolved in drinking water); TQ1 and TQ2 (2.5 and 5.0 mg/Kg respectively) body weight; n=8. Data are expressed as mean ± SD. *: p<0.05, **: p<0.01 and ****: p<0.0001. HgCl2: Mercuric chloride; TQ: Thymoquinone.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



