Submitted:
04 August 2025
Posted:
05 August 2025
You are already at the latest version
Abstract
Tuberculosis (TB), a disease caused by Mycobacterium tuberculosis, affects nearly one third of the world population. It is estimated that TB infects around ten million people worldwide with not less than two million fatalities annually. It is one of the treatable diseases thanks to the development of diagnostic tools and therapeutic agents. However, the disease remains a threat to humankind due to the emergence of multi-and-extensive drug resistance M. tuberculosis. This has driven many researchers to look for new antitubercular medications with better efficacy, safety and affordability. As has always been the case, natural products have provided huge potential as a source of remedies for various infectious and non-infectious diseases. The aim of this review is to report new discoveries and updates pertaining to antitubercular natural products with minimum inhibitory concentration (MIC) values of less than or 10µg/mL or 50 µM and selectivity indices of greater than 10. This review discusses 49 naturally occurring compounds from various classes, isolated from both terrestrial and aquatic organisms, including higher plants and microorganisms. Perusal of the literature reveals that most of these promising compounds are alkaloids, terpenoids, steroids and peptides. The most effective compound against the M. tuberculosis H37Rv sensitive strain was found to be rufomycin I (MIC= 0.02 µM), which outperformed isoniazid (MIC=0.23 µM). It was proposed that this cyclic peptide interferes with protein hemostasis through the transcription of ClpC1, a protein that is crucial to M. tuberculosis hemostasis. Current TB drugs have paradoxically turned out to become less effective; so due to its potent activity, selectivity, and novelty, different natures of products, including hapalindole A and rufomycin I, are now centered as the best leads against TB.
Keywords:
tuberculosis
; mechanisms of action
; minimum inhibitory concentration
; selectivity index
; natural products
1. Introduction
Tuberculosis (TB), one of the leading causes of death, is caused by Mycobacterium tuberculosis [1,2]. Around 2 billion people are latently infected with tuberculosis, yet only 10% of them develop active disease during their lifetime [3]. TB is treatable thanks to the development of diagnostic instruments, procedures, and medications in the 20th century [4,5]. Nevertheless, the emergence of HIV infection and the spread of multidrug-resistant TB (MDR-TB) in the 1980s and 1990s dealt TB management a serious blow [4]. According to WHO [2] TB annual report, an estimated 10.6 million people acquired TB, of which 7.5 million were newly diagnosed with huge burden (87%) shared by South-East Asian, African, and Western Pacific regions with the death of 1.3 million people [2]. Quarters of these deaths are because of coinfection with HIV/AIDS [2].
The current pharmacotherapy of TB is challenged by emergence of resistant bacterial strains, side effects and log duration of treatment. Hence, there is a need for search of new alternative treatment with new mode of action, good oral bioavailability, short duration of treatment, treating MDR- and extensively drug-resistant (XDR) TB, availability and affordability, decreasing pill burden, and low dosage frequency, and minimum drug-drug interactions [6]. This could be achieved through looking into natural products or synthetic molecules. Natural products provide a large pool of molecules that could have antimycobacterial effect or scaffold in anti-TB drug discovery [7].
Natural products could be screened for their antitubercular activity based on phenotypic (drug to target) or the target to drug screening [8,9]. Most anti-TB medications advanced to clinics through whole-cell screening, followed by analysis of the mechanism of action and identification of potential targets [10]. But even for early screening, antibacterial new drug discovery will be efficient and effective if it begins with a target that has been identified [8].
2. Targets of Mycobacteria for Antitubercular Drugs Discovery
2.1. Cell Wall of M. tuberculosis
The cell wall of M. tuberculosis is extremely intricately designed and contains elements that are involved in bacterial pathogenicity, host cell communication, and treatment resistance [11]. So, several medications have been produced that target the pathogen cell wall. Mycolic acid production pathway is one of the potential targets in M. tuberculosis cell wall [12]. The production of mycolic acid is carried out by two enzymes, enoyl acyl carrier protein reductase (InhA) and -ketoacyl synthase, both of which are members of the fatty acid synthase (FAS) family [13]. Lysis of the bacterial cell results from the inhibition of these enzymes, which prevent the bacteria’s ability to synthesize cell walls [4].
Since InhA is only present in bacteria, it makes it a good target for antibacterial treatments [14,15]. It is one of the enzymes responsible for producing mycolic acid, a component of the cell wall of M. tuberculosis [12]. There are three known binding sites on this NADH-dependent enzyme. Site I of NAD contains the tyrosine and ribose residues [16], Site II contains hydrophobic pockets that allow the binding of alky groups [17], and Site III has not been thoroughly explored but is thought to provide hydrophilic binding in which the phosphate group of NAD binds [18]. Isoniazid (INH) and ethionamide (ETH), two medications that are currently on the market, target this enzyme. Isoniazid is a prodrug that must be activated by catalase peroxidase in order for it to become an unstable free radical that binds to NADH of InhA and forms covalent bonds with it, rendering it inactive [15]. The bacteria will be killed off by this. To prevent the activation of INH, where the bacteria would otherwise develop drug resistance, the bacteria undergo KatG mutation. Therefore, research on compounds that will prevent InhA from activating through KatG as antimycobacterial drugs will be ideal [19].
In addition to InhA, β-hydroxyacyl- acyl carrier protein (ACP) dehydrase (HadABC), β-ketoacyl ACP reductase (MabA), and -ketoacyl-acyl carrier protein (ACP) synthases (KasA and KasB, mtFabH, mtFab) are enzymes involved in the production of mycolic acid [20]. As a result, these enzymes could serve as a target for antimycobacterial drugs.
2.2. Nucleic Acids
Nucleic acids play important role in the survival of mycobacteria. Notwithstanding, there are few drugs approved for treatment of TB on these targets [21].
2.2.1. Purine and Pyrimidine Ribonucleotide Synthesis
In purine biosynthesis, an aminoimidazole moiety is produced and connected to a ribose by a sequence of reactions that are metabolized by the enzyme phosphoribosylpyrophosphate (PRPP) synthetase, which is dependent on the metabolite inorganic phosphate, which is essential for the bacteria’s existence [22,23]. The C4 and C5 of imidazole then undergo cyclization, resulting in the production of inosine-5’-monophosphate (IMP), which is then transformed into guanosine 5’-monophosphate (GMP) by IMP dehydrogenase (IMPDH, GuaB) [21,24]. GuaB, one of the three IMPDH homologues that are currently known, is thought to be a viable therapeutic target because of its critical involvement in the metabolism of guanine nucleotides and as the cascade’s rate-limiting enzyme [21,25].
Uridine monophosphate, a precursor for pyrimidine nucleotides, is produced during the multi-enzymatic cascade reaction of pyrimidine biosynthesis [24]. Promising therapeutic targets have been investigated, including the cascade that orotate phosphoribosyltransfrase (OPRT) catalyzes to transform orotate into orotidine 5’-monophosphate (OMP) [22,23]. Additionally, PRPP synthase, an enzyme that synthesize PRPP, which in turn is necessary to produce both pyrimidine and purine nucleotides. Therefore, blocking this enzyme will also likely be a target for drugs [21].
2.2.2. Deoxyribonucleic Acid (DNA) Replication
A multistep, multiprotein replisome in bacteria does DNA replication in a highly controlled and coordinated manner [26]. These replisome proteins catalyze the production of RNA primers, clamp loading, DNA polymerization, and DNA unwinding [21]. Nearly 3,950 genes make up the genome of M. tuberculosis, and 10% of them are necessary for the bacteria to survive [27,28]. The DnaA replication initiator, the primosomal (P) helicase, the DnaB helicase, the DnaG primase, the single-stranded DNA binding proteins (SSB), the clamp loader subunits, the DNA polymerases I and III, the DnaN-clamp, the DNA ligase I, and type I and II topoisomerases are among the 15 genes that are necessary for DNA replication [26]. The single topoisomerase II class member, DNA gyrase (mostly DNA gyrA), is the only target for fluoroquinolones in the treatment of MDR-TB [29]. Therefore, there is a great potential to search for targets as DNA replication inhibitors and anti-mycobacterial drugs.
2.2.3. DNA Repair
Due to the release of reactive oxygen species (ROS) and reactive nitrogen species (RNS), M. tuberculosis continuously sustains DNA damage in its host cell [30]. One of the RNS, nitrogen monoxide, interacts with oxygen to form nitrous anhydride, which then nitrosates amines and amides to produce potent DNA alkylating agents that endanger the survival of the bacteria because of genome instability [21]. Multi-enzymatic systems including Nucleotide Excision Repair (NER) and Base Excision Repair (BER), recombination repair systems, and single proteins that directly reverse DNA damage are all part of M. tuberculosis’ DNA repair mechanisms [31]. As a result, antimycobacterial agents that target this bacterial pathway will have potential. However, they possess a challenge of selectivity [21].
2.3. Protein Synthesis (RNA Translation)
The ribosome of M. tuberculosis is the key component in the translation of RNA into proteins [12]. The smaller subunit of ribosomes, known as 30S, contains 21 ribosomal proteins (RPs) and 16S ribosomal RNA (rRNA), which are used to decode messenger RNA (mRNA) sequences, and the larger subunit, known as 50S, contains 37 RPs and 20S and 5S rRNA and is used to form peptide bonds via the peptidyl transferase center (PTC) [12,32]. The 70S functional ribosome is made up of the two subunits [33]. The two subunits came together to create the 70S functioning ribosome [32]. Since these subunits’ interface is where the incoming transfer RNA (tRNA) passes via the aminoacyl (A-site), peptidyl (+P-site), and exit (E-site) sites, it is a crucial translational step [12]. The decoding center in the 30S and PTC and polypeptide exit site at 50S subunit on the ribosome are prospective targets on the basis of the already available antibiotics (aminoglycosides) [34,35].
2.4. Energy Metabolism
Mycobacterium tuberculosis produces ATP through oxidative phosphorylation, which is essential for growth and survival [36]. M. tuberculosis can produce ATP by substrate level phosphorylation, however it is insufficient [37]. Therefore, the primary source of energy for the bacteria is oxidative phosphorylation. NADH dehydrogenases and succinate dehydrogenases aid in the entry of the electron transport chain that transfers electrons to menaquinone, generating a protein complex and proton motive force (PMF), in the process of oxidative phosphorylation [37,38]. Adenosine triphosphatase (ATP synthase) uses the energy of this enzyme to produce ATP [36]. Since oxidative phosphorylation differs significantly between prokaryotes and eukaryotes, there are questions about the selectivity of antitubercular medications that target this route [37]. Despite this, there are already available clinical and experimental medications that more specifically block the routes used by mycobacteria to synthesize ATP, revealing alternate targets for treating MDR and XDR infections.
3. Natural Products with Potent Antimycobacterial Activities
Numerous natural compounds derived from both aquatic and terrestrial sources, such as microbes and higher plants, have demonstrated potent anti-mycobacterial properties against various strains of mycobacteria. In vitro assays are used to determine the inhibitory concentration (IC) or minimum inhibitory concentration (MIC) of natural compounds [39]. MIC offers numerical outcomes [40]. For further study as potential anti-TB drug candidates, natural compounds with in vitro MIC values of less than or equal to 10 µg/mL or 50 µM are regarded as having a considerable antimycobacterial action and therefore considered for further investigations provided that their selectivity indices are greater than 10 [41,42]. This review focuses on natural products discovered between 2000 and 2024, with particular emphasis on compounds identified from 2020 to 2024 that exhibit potent in vitro activity against Mycobacterium, as demonstrated by minimum inhibitory concentration (MIC) values of ≤10 µg/mL or 50 µM, along with selectivity indices (SI) greater than 10. These bioactive compounds primarily belong to the terpene, alkaloid, peptide, and steroid classes.
3.1. Alkaloids
Alkaloids, which are a diverse group of nitrogen-containing natural products, have demonstrated promising potential against tuberculosis (TB), especially drug-resistant M. tuberculosis. Many plants, cyanobacteria, and marine-derived alkaloids have demonstrated potent antimycobacterial activity, with high selectivity indices. The bioactive compounds target M. tuberculosis in different ways and provide scaffolds for new anti-TB treatments.
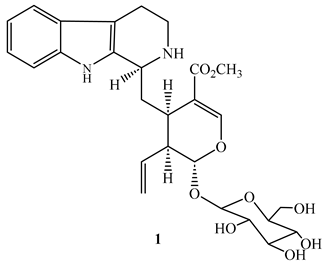
Strictosidine (1), a monoterpene indole obtained from the methanol extract of the leaves and twigs of Psychotria nuda (Cham. & Schltdl.) Wawra (Rubiaceae), showed an MIC value of 7.1 µg/mL against M. tuberculosis H37Rv with cytotoxic effect of 90 µg/mL against RAW264.7 cells in the MTT assay [43].
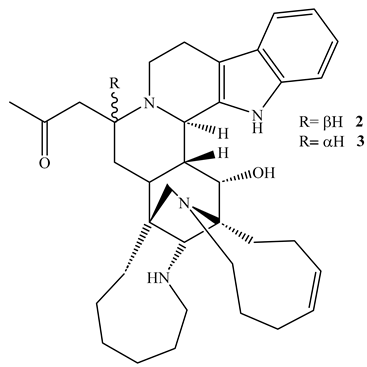
Two indole alkaloids namely, manadomanzamine A (2) and manadomanzamine B (3), isolated from the acetone extract of the sponge Acanthostrongylophora sp., (Petrosiidae) demonstrated potent activity against M. tuberculosis H37Rv with MIC values of 1.9 µg/mL and 1.5 µg/mL, respectively, in the Microplate Alamar Blue Assay (MABA) with no noticeable toxicity against normal Vero cell line at 5 µg/mL [44]. These alkaloids also possess anti-HIV effect implying they could be alternative treatments for TB-HIV infections [44].
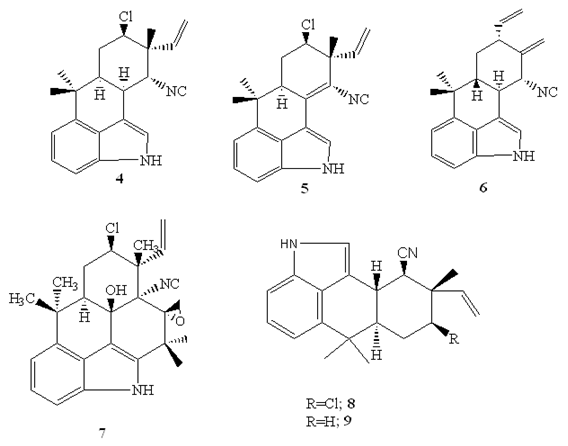
The hapalindole-type alkaloids, hapalindoleisonitrile A (4), I (5), X (6), isolated from the cultured cyanobacteria otherwise called bule-green algae (Westiellopsis sp. and Fischerella muscicola) exhibited substantial activities against M. tuberculosis H37Rv, though only hapalindole A, and hapalindole X exhibited potent activity against M. smegmatis in the MABA method with SI> 10 (Table 1) [45].
Fischambiguine B (7) (MIC=22 µM) and hapalindole G (8) (MIC=6.8 µM), hapalindole alkaloids produced by the filamentous, branched, sheathed blue green algae Fischerella ambigua (Fischerellaceae) showed activity against M. tuberculosis [46,47]. In addition, hapalindole H (9) (MIC= 39.6 µM), fischambiguine B (MIC=23.4 µM), and hapalindole G (MIC=34 µM) were reported to be effective against M. smegmatis [46,47]. According to Mo et al. [46], only fischambiguine B demonstrated superior specificity (SI>60) toward M. tuberculosis H37Rv and M. smegmatis, while hapalindole G demonstrated specificity (SI>18) toward M. tuberculosis H37Rv when tested against Vero cell [48]. SAR study of fischambiguine B revealed that the epoxide at C25-26 improve activity [47].
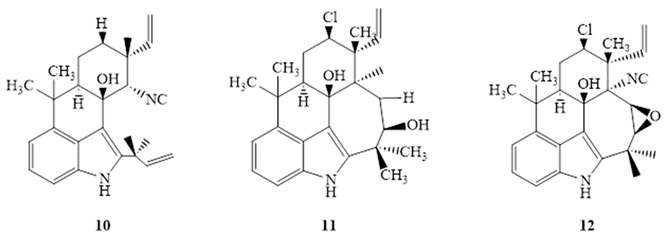
Mo et al. [46] isolated ambiguine isonitriles from the CH2Cl2: MeOH (1:1) extract of Fischerella ambigua. potent activity was reported for ambiguine C (10) isonitrle, and ambiguine M isonitrle (11) against M. tuberculosis H37Rv (Table 2) [46]. On the other hand, ambiguine E (12) isonitrle was found to be highly active against M. smegmatis in the MABA method [46]. Despite their strong antimycobacterial activity, only ambiguine E (12) isonitrile (SI=10.67) and ambiguine C isonitrile (SI=11.2) showed specificity toward M. tuberculosis H37Rv, and ambiguine E isonitrile (SI=30.4) toward M. smegmatis [45,46]. SAR studies indicated that the chlorine substituent at C-13, did not make any significant difference on activity toward the two strains [46].
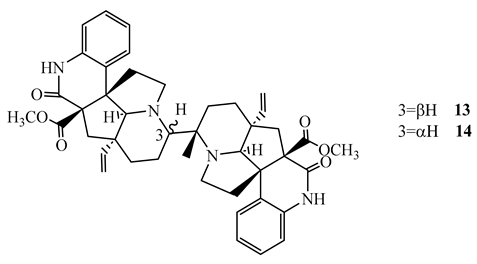
Suadamin A (13) and suadamin B (14), dimeric monoterpenoid quinoline alkaloids isolated from the twigs and leaves of Melodinus suaveolens, showed activity against M. tuberculosis H37Rv with an MIC value of 6.76 μM and 33.47 μM, respectively, in the MABA method. Suadamine A (SI=21.2) showed a better selectivity index against Vero cell [49]. SAR study revealed that the difference in activity between the two isomers was caused by their difference in stereochemistry at C-3 [49].
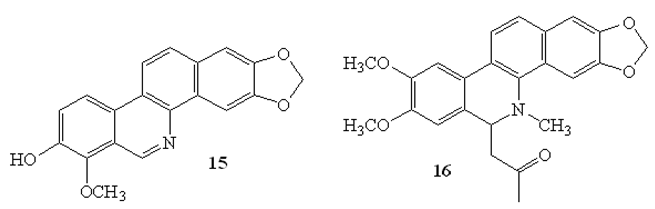
Other isoquinoline-based alkaloids namely, decarine (15), and 6-acetonyldihydronitidine (16) isolated from the roots of Zanthoxylum capense were shown to exhibit substantial activities against M. tuberculosis H37Ra with MIC values of 3.1 μg/mL and 6.2 μg/mL, respectively, by microbroth dilution assay. In addition, decarine was found to be active against M. tuberculosis H37Rv strain (MIC=1.6 μg/mL with SI>10 [50].
3.2. Simple Amides and Peptides
Cyclic peptides and usual amides have become potential antimycobacterial drugs, showing potent activity against both drug-sensitive and drug-resistant M. tuberculosis strains. The cyclic peptides and amides contained in this review are mostly derived from Streptomyces and Zanthoxylum capense and have been shown to exhibit bactericidal activity at low MIC values with low cytotoxicity. The mechanisms of action of the cyclic peptides and amides and particularly their activity in inhibition of ClpC1 ATPase provide a new direction for the discovery of new therapeutics against tuberculosis.
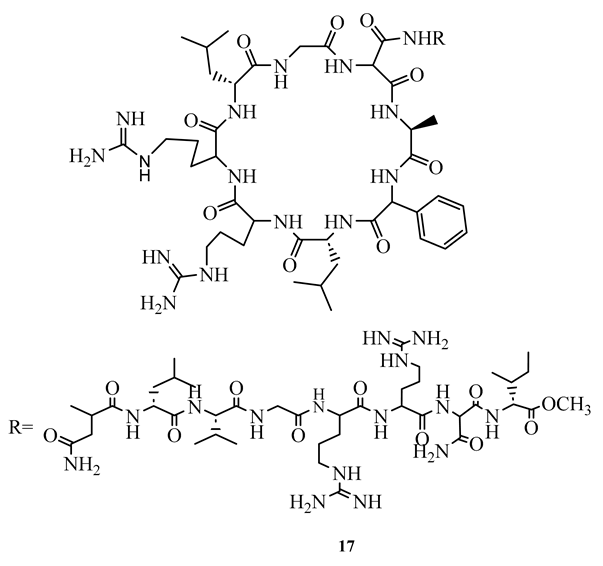
Lassomycin (17), a cyclic peptide isolated from Lentzea kentuckyensis, has demonstrated an excellent bactericidal action against M. smegmatis, M. tuberculosis H37Rv, M. avium subsp. paratuberculosis, MDR and XDR strains, with MIC ranging between 0.125 µg/mL and 3.1 µg/mL with less toxicity (IC50 =350 µg/mL) against human NIH 3T3 and HepG2 cells [9]. Molecular docking study indicated that its mechanism of action is binding to the acidic region of the caseinolytic protein, ClpC1 of ATPase where it interferes with proteolysis activity of ATPase [9].
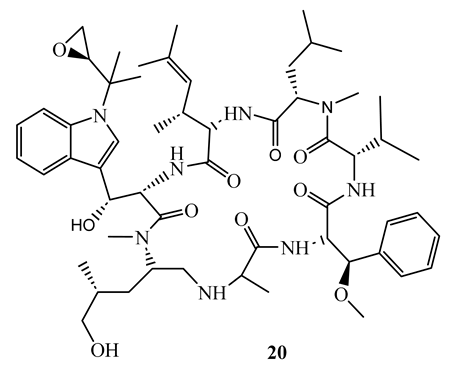
Rufomycin I (18), ecumicin (19), and cyclomarin A (20), cyclic peptides isolated from Streptomyces sp. demonstrated potent activity (MIC ranging between 0.02 µM and 4.85 µM) against M. tuberculosis H37Rv (both sensitive and resistant strains), M. smegmatis, and M. avium. The most potent was rufomycin I (MIC= 0.02 µM) which showed better activity than isoniazid (MIC=0.23 µM) against M. tuberculosis H37Rv sensitive strain [51]. In another study, ecumicin was shown to have MIC value of 0.26 µg/mL against M. tuberculosis H37Rv when tested using the MABA method [52]. It was demonstrated that the selective indices of the above compounds towards mycobacteria were more than 10 [51,52]. A hypothesis was put forward that the mechanism of action of these cyclic peptides is through interference with protein hemostasis by targeting ClpC1, which plays an important role in the hemostasis of protein in M. tuberculosis [51]. Considering the less cytotoxic nature of the above peptides, they were considered to be potential candidates for the discovery of new canon in the fight against both sensitive and resistant mycobacteria infections.
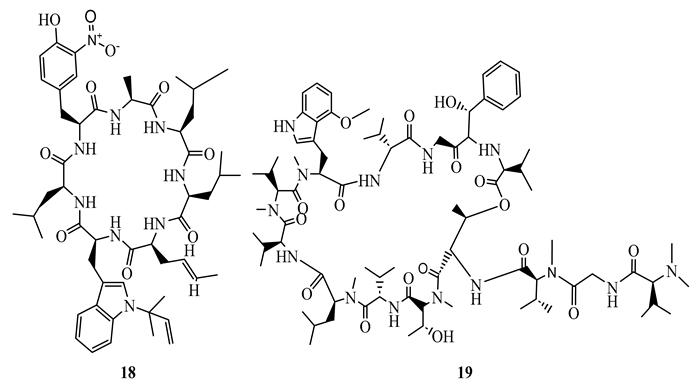
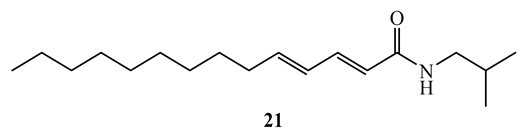
N-isobutyl-(2E,4E)-2,4-tetradecadienamide (21), a simple amide isolated from the roots of Zanthoxylum capense demonstrated antimycobacterial activity against M. tuberculosis H37Rv and H37Ra with MIC values of 1.6 µg/mL (SI=38.2) and 6.2 µg/mL (SI=9.9), respectively, in broth micro-dilution method [50].
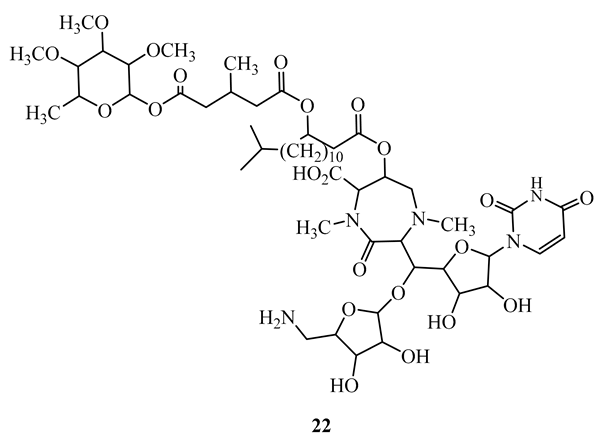
Caprazamycin B (22), a novel peptide produced by a different species of Streptomyces, displayed potent activity (MIC=3.13 µg/mL) against the M. tuberculosis H37Rv Kurono, and M. bovis Ravenel strains. In addition, it showed activity against drug susceptible and multidrug-resistant M. tuberculosis with MIC ranging between 6.25 and 12.5 µg/mL. It did not exhibit any significant cytotoxicity at 5000 µg/mL in mice [53].
3.3. Terpenoids
Naturally occurring products have shown significant antimycobacterial activity against M. tuberculosis and have different scaffolds such as triterpenoids, diterpenes, isonitrile compounds. Those include eurylene, 12-deacetoxyscalarin derivatives, and amphilectene-based compounds, showing remarkable low MICs, moderate to high selectivity indices, and potential for further development as therapeutics for tuberculosis. Their proposed mechanisms of action might be membrane disruption through inhibition of enzymes including farnesyl transferase has raised the possibility for these compounds to be valuable lead structures in the development of anti-TB drugs.
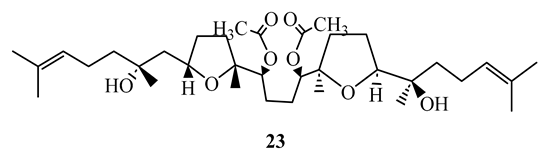
Strong activity against M. tuberculosis H37Rv and M. tuberculosis M299 were demonstrated by the squalene type triterpenoid eurylene (23), isolated from the roots of Homalolepis suffruticosa, with MIC= 1.4 µg/mL and 2.0 µg/mL, respectively. These compounds had better selectivity (SI>20) when tested by the MTT method [54].
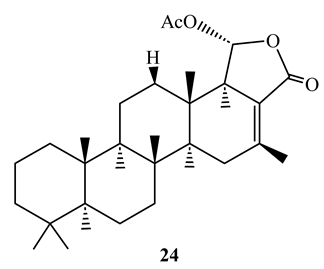
12-Deacetoxyscalarin 19-acetate (24), displayed antimycobacterial activity with MIC value of 4 µM against M. tuberculosis H37Ra when tested by the MABA method [55]. Its mechanism of action was shown to be inhibition of farnesyl transferase which interferes with membrane of the bacteria [56].
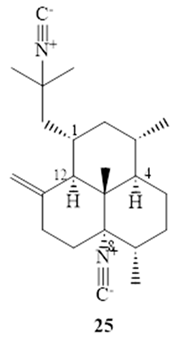
Nieves et al. [57] reported that (−)-8,15-diisocyano-11(20)-amphilectene (25), exhibits strong activity (MIC=8 µM) against M. tuberculosis H37Ra by using MABA with marginal selectivity (SI=10.2) [57].
3.4. Steroids
Steroidal compounds have displayed potent inhibitory activity against M. tuberculosis, including the drug-sensitive strains, particularly the triterpene and phytosterol derivatives. The steroids, including saringosterol and stigmastene, were made from plant and marine algae, and had relatively low MIC values, some equaling the potency of standard drugs such as rifamycin. The selectivity indices and structure–activity relationships were promising, particularly in terms of the role of unsaturated ketone moieties, indicating the steroids could have utility as scaffolds for anti-TB drug development.
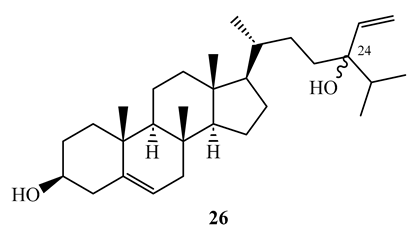
Saringosterol (26) is a triterpene steroid isolated from CH2Cl2-CH3OH (1:1) extract of Lessonia nigrescens (brown alga). It was obtained as a 1:1 mixture of 24S and 24R epimers. Saringosterol was found to have strong activity (MIC=0.25 µg/mL) against M. tuberculosis H37Rv. Its activity was comparable with that of rifamycin used as a positive control [58]. In the same study, it was shown that the pure 24S and 24R epimers possess MIC values of 1 µg/mL, and 0.125 µg/mL, respectively, against same strain in the BACTEC method [58]. The compound has shown a selectivity index greater than 10 against Vero cell [58]. The difference in activity between the two isomers should be studied further to understand the SAR.
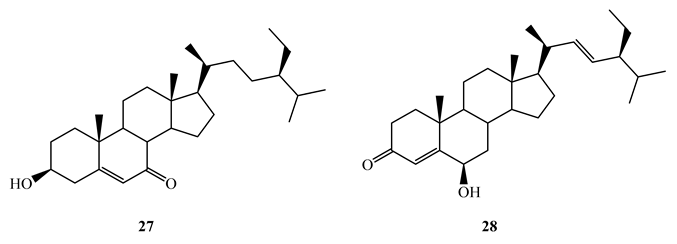
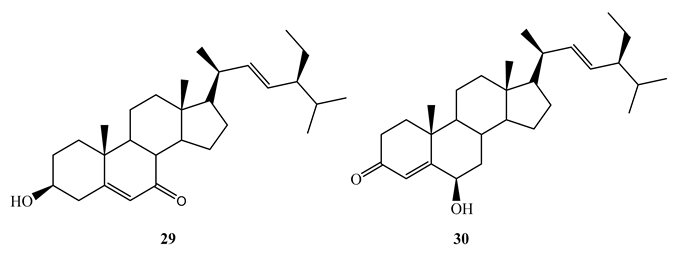
Bioassay guided fractionation of the aerial parts of Thalia multiflora Horkel ex Koernicke (Marantaceae) with dichloromethane-methanol (1:1) led to the isolation of stigmast-5-ene-3β-ol-7-one (MIC=1.98 µg/mL) (27), stigmast-4-ene-6β-ol-3-one (MIC=4.298 µg/mL) (28), stigmast-5,22-diene-3β-ol-7-one (29) (MIC=1.0 µg/mL) and stigmast-4,22-diene-6β-ol-3-one (30) (MIC=1.0 µg/mL) against M. tuberculosis H37Rv [59]. It was revealed that that the presence of α, β unsaturated ketone either on ring A or B has significant effect on antitubercular activity. The above compounds showed higher in vitro selectivity (SI>10) against Vero cells making them potential candidates for further studies as antitubercular agents.
3.5. Miscellaneous Compounds
There exists a wide variety of structurally diverse natural products with antimycobacterial activities, including fatty acid derivatives, polyketides, lactones, organic acids, and others. Certain compounds (e.g., falcarindiol, micromolide, plumericin) exhibited low values of MIC with selectivity indices and good mechanisms of action (e.g., inhibition of the biosynthesis of para-aminobenzoate (PABA) and succinate dehydrogenase). The potent activity of these compounds against drug-sensitive and resistant strains of M. tuberculosis supports and may indicate their potential for use in future explorations for anti-TB drugs.
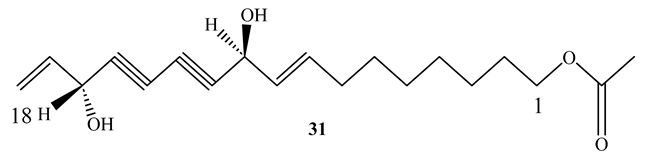
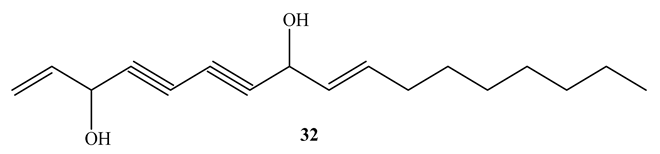
11(S),16(R)-Dihydroxy-octadeca-9Z,17-dien-12,14-diyn-1-yl acetate (31) (MIC=1.4 µg/mL) and falcarindiol (32) (MIC=6 µg/mL), obtained from the methanol extract of Angelica sinensis roots, showed potent activity against M. tuberculosis Erdman. Both compounds showed selectivity against M. tuberculosis Erdman with SI>117 and SI>19, respectively [60]. SAR investigations confirmed that an OH group at C16 and unsaturation at C9-C10 are necessary for their antimycobacterial activity [60]. This implies that the compounds are good candidates for more research in the evaluation of antimycobacterial properties [41].
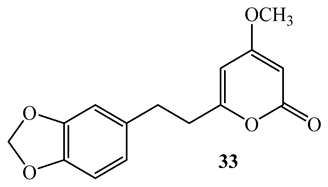
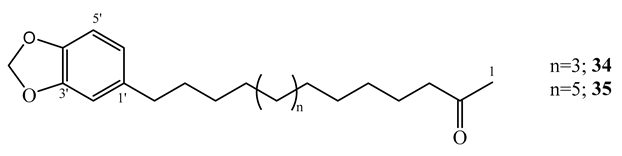
By using the MABA assay, the kawalactone, 5,6-dehydro-7,8-dihydromethysticin (33), 2-oxo-14-(3′,4′-methylenedioxyphenyl) tetradecane (34), and 2-oxo-16-(3′,4′-methylenedioxyphenyl) hexadecane (35) isolated from the dichloromethane-methanol (1:1) extract of the leaves and stem of Piper sanctum, demonstrated activity against M. tuberculosis H37Rv (MIC=4-6.25 µg/mL) with selectivity indices of greater than 10 against Vero cell [61].
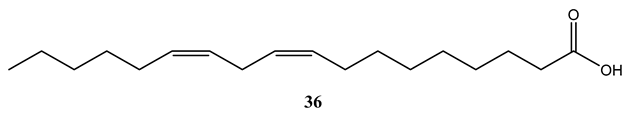
Linoleic acid (36), isolated from the CH2Cl2 extract of the stem bark of Warburgia ugandensis showed potent activity against M. aurum and M. phlei with MIC values of 4 µg/mL and individually and M. fortuitum (MIC=8 µg/mL) [62]. Mycobacterium tuberculosis has evolved to utilize long-chain fatty acids (LCFAs) as a preferred carbon source. However, LCFAs have long been known to exhibit bactericidal effects against M. tuberculosis in vitro by disrupting the bacterial membrane, leading to hyperpolarization and subsequent cell death. A deeper understanding of how M. tuberculosis regulates the uptake and metabolism of LCFAs, and how these pathways can be therapeutically targeted, is essential for the development of novel anti-tuberculosis agents.
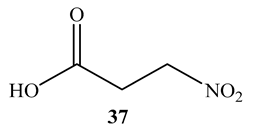
3-Nitropropionic acid (37) obtained from the endophytic fungal strain, Phomopsis sp. strain usia5, displayed strong activity (MIC = 3.3 μM) against M. tuberculosis H37Ra and undetected cytotoxicity against Vero cell [63]. Inhibition of succinate dehydrogenase has been proposed to be the mechanism of action of 3-nitropropionic [63].
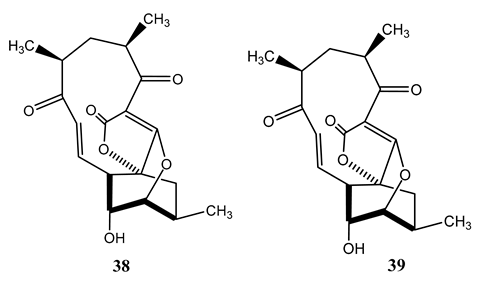
(_)-Abyssomicin C (38) and (_)-atrop-abyssomicin C (39) are polyketide-like compounds produced by Verrucosispora maris, a Gram-positive bacterium isolated from deep-sea sediments from the Sea of Japan [64]. Both compounds showed strong antimycobacterial activity (MIC=1.2-10 µg/mL) against Mycobacterium species in the MABA method. The most potent activity (MIC=1.2 µg/mL) was reported against M. tuberculosis H37Rv [65]. Their mechanism of action is associated with inhibition of the para-amino benzoate (PABA) biosynthetic pathway [66]. Since there is no such biosynthetic pathway in human [65], these class of drugs have potential for further development.
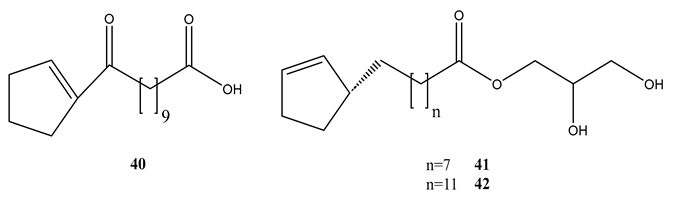
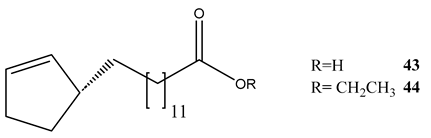
Phytochemical evaluation of the seeds of Hydnocarpus anthelminthica led to the isolation of five organic acids or their derivatives. All the compounds showed variable activities against M. bovis BCG and M. tuberculosis H37Rv (Table 3) [67]. Antihemithicin C was found to inhibit the metabolic pathway between chorismate and PABA [67].
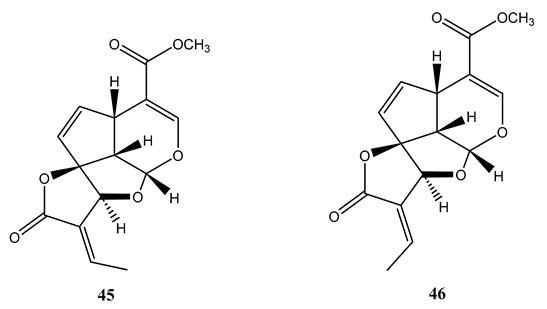
Plumeria bicolo bark extract yielded plumericin (45) and isoplumericin (46), which showed antimycobacterial activity against M. tuberculosis H37Rv and MDR strains with MIC values ranging between 1.3 µg/mL and 2.6 µg/mL. Maximum activity (MIC= 1.3 µg/mL) was observed for plumercin against MDR strain [68]. The activities of the compounds were better than the positive controls isoniazid and rifampicin against MDR strains. The compounds were found to be non-toxic to 74G8 murine macrophage cell lines at the tested concentration [68].
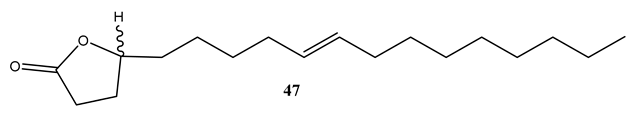
Micromolide (47), a lactone derivative of oleic acid, isolated from the CH2Cl2 extract of the stem bark of Micromelum hirsutum was found to possess potent activity (MIC=1.5 µg/mL) against M. tuberculosis H37Rv with selectivity index (SI) of 63 by using MABA method [69]. Furthermore, micromolide exhibited strong activity (MIC=5.6 µg/mL) against M. tuberculosis Erdman on J774 mouse macrophage cell line [69]. This indicates that the compound will be a good candidate for in vivo studies as antimycobacterial agent.
Maritinone (48) and 3,3ʹ-biplumbagin (49), naphthoquinones isolated from the n-hexane extract of the stem bark of Diospyros anisandra, demonstrated strong activity against sensitive M. tuberculosis H37Rv with an MIC value of 3.13 µg/mL each. Both compounds showed a high selectivity (SI>10) against Vero cell [70]. Sarkar et al. [71] reported that such anthraquinones target thymidylate synthase (ThyX) of the Mycobacterium [71].

4. Conclusion
Current control strategies have had little impact on TB control and new therapies are urgently needed. Some strains of M. tuberculosis are extensively resistant to all existing therapeutic agents for TB treatment, indicating the requirement for new drugs with novel modes of action and targets. A number of natural products from different sources showed particular promise as anti-mycobacterial agents with clinical potential. However, most of antitubercular drugs are discovered through phenotypic screening rather than on the basis of target-based drug discovery. This will make antitubercular drug discovery challenging. Despite these issues, natural products such as hapalindole A, lassomycin, rufomycin I, ecumicin, and cyclomarin A were found to be potent antimycobacterial agents in in vitro models with higher selectivity index. This indicates that natural products remain one of the most productive sources of drug leads to this day, and no doubt will continue to benefit antimycobacterial drug discovery. However, structure activity relationships should be performed on those promising compounds so that safe and cost-effective anti-tuberculosis agents that can be useful in the fight against tuberculosis are developed.
Author Contributions
Conceptualization PB, AK, DB, YM; software and data curation, PB, AK, MY writing-original draft preparation, BP, AK, BD; writing-review and editing, BP, YM, AK, DB, have likewise contributed to this work. All authors have read and agreed to the published version of the manuscript.
Funding
This study was partially funded by the International Science Program (ISP), Uppsala University, through the ETH:02 project.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
There is no additional data associated with this article.
Acknowledgments
One of the authors (B.P.) would like to express his gratitude for financial support to International Science Program (ISP), Uppsala University, through the ETH:02 project.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Abrahams, K.A.; Besra, G.S. Mycobacterial Drug Discovery. RSC Med. Chem. 2020, 11, 1354–1365. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. The WHO Global Tuberculosis Report 2023.
- Bueno-Sánchez, J.G.; Martínez-Morales, J.R.; Stashenko, E.E.; Ribón, W. Anti-Tubercular Activity of Eleven Aromatic and Medicinal Plants Occurring in Colombia. Biomed. Rev. Inst. Nac. Salud 2009, 29, 51–60. [Google Scholar]
- Zhang, Y.; Amzel, L.M. Tuberculosis Drug Targets. Curr. Drug Targets 2002, 3, 131–154. [Google Scholar] [CrossRef] [PubMed]
- Lawn, S.D.; Zumla, A.I. Tuberculosis. Lancet Lond. Engl. 2011, 378, 57–72. [Google Scholar] [CrossRef]
- Shehzad, A.; Rehman, G.; Ul-Islam, M.; Khattak, W.A.; Lee, Y.S. Challenges in the Development of Drugs for the Treatment of Tuberculosis. Braz. J. Infect. Dis. 2013, 17, 74–81. [Google Scholar] [CrossRef]
- Verpoorte, R. Exploration of Nature’s Chemodiversity: The Role of Secondary Metabolites as Leads in Drug Development. Drug Discov. Today 1998, 3, 232–238. [Google Scholar] [CrossRef]
- Kana, B.D.; Karakousis, P.C.; Parish, T.; Dick, T. Future Target-Based Drug Discovery for Tuberculosis? Tuberculosis 2014, 94, 551–556. [Google Scholar] [CrossRef]
- Gavrish, E.; Sit, C.S.; Cao, S.; Kandror, O.; Spoering, A.; Peoples, A.; Ling, L.; Fetterman, A.; Hughes, D.; Bissell, A.; et al. Lassomycin, a Ribosomally Synthesized Cyclic Peptide, Kills Mycobacterium tuberculosis by Targeting the ATP-Dependent Protease ClpC1P1P2. Chem. Biol. 2014, 21, 509–518. [Google Scholar] [CrossRef]
- Oh, S.; Trifonov, L.; Yadav, V.D.; Barry, C.E.; Boshoff, H.I. Tuberculosis Drug Discovery: A Decade of Hit Assessment for Defined Targets. Front. Cell. Infect. Microbiol. 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Barry, C.E. Interpreting Cell Wall “virulence Factors” of Mycobacterium tuberculosis. Trends Microbiol. 2001, 9, 237–241. [Google Scholar] [CrossRef]
- Shetye, G.S.; Franzblau, S.G.; Cho, S. New Tuberculosis Drug Targets, Their Inhibitors, and Potential Therapeutic Impact. Transl. Res. 2020, 220, 68–97. [Google Scholar] [CrossRef]
- Martínez-Hoyos, M.; Perez-Herran, E.; Gulten, G.; Encinas, L.; Álvarez-Gómez, D.; Alvarez, E.; Ferrer-Bazaga, S.; García-Pérez, A.; Ortega, F.; Angulo-Barturen, I.; et al. Antitubercular Drugs for an Old Target: GSK693 as a Promising InhA Direct Inhibitor. EBioMedicine 2016, 8, 291–301. [Google Scholar] [CrossRef]
- Campaniço, A.; Moreira, R.; Lopes, F. Drug Discovery in Tuberculosis. New Drug Targets and Antimycobacterial Agents. Eur. J. Med. Chem. 2018, 150, 525–545. [Google Scholar] [CrossRef]
- Purohit, H.J.; Cheema, S.; Lal, S.; Raut, C.P.; Kalia, V.C. In Search of Drug Targets for Mycobacterium tuberculosis. Infect. Disord. Drug Targets 2007, 7, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Kuo, M.R.; Morbidoni, H.R.; Alland, D.; Sneddon, S.F.; Gourlie, B.B.; Staveski, M.M.; Leonard, M.; Gregory, J.S.; Janjigian, A.D.; Yee, C.; et al. Targeting Tuberculosis and Malaria through Inhibition of Enoyl Reductase: Compound Activity and Structural Data. J. Biol. Chem. 2003, 278, 20851–20859. [Google Scholar] [CrossRef] [PubMed]
- Rozwarski, D.A.; Vilchèze, C.; Sugantino, M.; Bittman, R.; Sacchettini, J.C. Crystal Structure of the Mycobacterium tuberculosis Enoyl-ACP Reductase, InhA, in Complex with NAD+ and a C16 Fatty Acyl Substrate. J. Biol. Chem. 1999, 274, 15582–15589. [Google Scholar] [CrossRef]
- Shirude, P.S.; Madhavapeddi, P.; Naik, M.; Murugan, K.; Shinde, V.; Nandishaiah, R.; Bhat, J.; Kumar, A.; Hameed, S.; Holdgate, G.; et al. Methyl-Thiazoles: A Novel Mode of Inhibition with the Potential to Develop Novel Inhibitors Targeting InhA in Mycobacterium tuberculosis. J. Med. Chem. 2013, 56, 8533–8542. [Google Scholar] [CrossRef] [PubMed]
- Almeida Da Silva, P.E.A.; Palomino, J.C. Molecular Basis and Mechanisms of Drug Resistance in Mycobacterium tuberculosis: Classical and New Drugs. J. Antimicrob. Chemother. 2011, 66, 1417–1430. [Google Scholar] [CrossRef]
- Duan, X.; Xiang, X.; Xie, J. Crucial Components of Mycobacterium Type II Fatty Acid Biosynthesis (Fas-II) and Their Inhibitors. FEMS Microbiol. Lett. 2014, 360, 87–99. [Google Scholar] [CrossRef]
- Miggiano, R.; Morrone, C.; Rossi, F.; Rizzi, M. Targeting Genome Integrity in Mycobacterium tuberculosis: From Nucleotide Synthesis to DNA Replication and Repair. Molecules 2020, 25, 1205. [Google Scholar] [CrossRef]
- Biochemical and Structural Investigations on Phosphoribosylpyrophosphate Synthetase from Mycobacterium smegmatis. PLoS ONE. Available online: https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0175815 (accessed on 11 December 2022). [CrossRef]
- Breda, A.; Martinelli, L.K.B.; Bizarro, C.V.; Rosado, L.A.; Borges, C.B.; Santos, D.S.; Basso, L.A. Wild-Type Phosphoribosylpyrophosphate Synthase (PRS) from Mycobacterium tuberculosis: A Bacterial Class II PRS? PLoS ONE 2012, 7, e39245. [Google Scholar] [CrossRef]
- Warner, D.F.; Evans, J.C.; Mizrahi, V. Nucleotide Metabolism and DNA Replication. Microbiol. Spectr. 2014, 2. [Google Scholar] [CrossRef]
- Hedstrom, L. IMP Dehydrogenase: Structure, Mechanism, and Inhibition. Chem. Rev. 2009, 109, 2903–2928. [Google Scholar] [CrossRef]
- Ditse, Z.; Lamers, M.H.; Warner, D.F. DNA Replication in Mycobacterium tuberculosis. Microbiol. Spectr. 2017, 5. [Google Scholar] [CrossRef]
- Wang, T.-C.; Chen, F.-C. The Evolutionary Landscape of the Mycobacterium tuberculosis Genome. Gene 2013, 518, 187–193. [Google Scholar] [CrossRef]
- DeJesus, M.A.; Gerrick, E.R.; Xu, W.; Park, S.W.; Long, J.E.; Boutte, C.C.; Rubin, E.J.; Schnappinger, D.; Ehrt, S.; Fortune, S.M.; et al. Comprehensive Essentiality Analysis of the Mycobacterium tuberculosis Genome via Saturating Transposon Mutagenesis. mBio 2017, 8, e02133-16. [Google Scholar] [CrossRef] [PubMed]
- Reiche, M.A.; Warner, D.F.; Mizrahi, V. Targeting DNA Replication and Repair for the Development of Novel Therapeutics against Tuberculosis. Front. Mol. Biosci. 2017, 4, 75. [Google Scholar] [CrossRef] [PubMed]
- Durbach, S.I.; Springer, B.; Machowski, E.E.; North, R.J.; Papavinasasundaram, K.G.; Colston, M.J.; Böttger, E.C.; Mizrahi, V. DNA Alkylation Damage as a Sensor of Nitrosative Stress in Mycobacterium tuberculosis. Infect. Immun. 2003, 71, 997–1000. [Google Scholar] [CrossRef] [PubMed]
- Mizrahi, V.; Andersen, S.J. DNA Repair in Mycobacterium tuberculosis. What Have We Learnt from the Genome Sequence? Mol. Microbiol. 1998, 29, 1331–1339. [Google Scholar] [CrossRef]
- Kumar, N.; Sharma, S.; Kaushal, P.S. Protein Synthesis in Mycobacterium tuberculosis as a Potential Target for Therapeutic Interventions. Mol. Aspects Med. 2021, 81, 101002. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Chang, J.; Cui, Z.; Li, X.; Meng, R.; Duan, L.; Thongchol, J.; Jakana, J.; Huwe, C.M.; Sacchettini, J.C.; Zhang, J. Structural Insights into Species-Specific Features of the Ribosome from the Human Pathogen Mycobacterium tuberculosis. Nucleic Acids Res 2017, 45, 10884–10894. [Google Scholar] [CrossRef]
- Wilson, D.N. Ribosome-Targeting Antibiotics and Mechanisms of Bacterial Resistance. Nat. Rev. Microbiol. 2014, 12, 35–48. [Google Scholar] [CrossRef]
- Poehlsgaard, J.; Douthwaite, S. The Bacterial Ribosome as a Target for Antibiotics. Nat. Rev. Microbiol. 2005, 3, 870–881. [Google Scholar] [CrossRef]
- Bald, D.; Koul, A. Respiratory ATP Synthesis: The New Generation of Mycobacterial Drug Targets? FEMS Microbiol. Lett. 2010, 308, 1–7. [Google Scholar] [CrossRef]
- Bald, D.; Villellas, C.; Lu, P.; Koul, A. Targeting Energy Metabolism in Mycobacterium tuberculosis, a New Paradigm in Antimycobacterial Drug Discovery. mBio 2017, 8, e00272-17. [Google Scholar] [CrossRef]
- Wellington, S.; Hung, D.T. The Expanding Diversity of Mycobacterium tuberculosis Drug Targets. ACS Infect. Dis. 2018, 4, 696–714. [Google Scholar] [CrossRef]
- Bueno, J.; Kouznetsov, V. Antimycobacterial Susceptibility Testing Methods for Natural Products Research. Braz. J. Microbiol. Publ. Braz. Soc. Microbiol. 2010, 41, 270–277. [Google Scholar] [CrossRef]
- Jorgensen, J.H.; Ferraro, M.J. Antimicrobial Susceptibility Testing: A Review of General Principles and Contemporary Practices. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2009, 49, 1749–1755. [Google Scholar] [CrossRef] [PubMed]
- Orme, I.; Program, T.D.S. Search for New Drugs for Treatment of Tuberculosis. Antimicrob. Agents Chemother. 2001, 45, 1943–1946. [Google Scholar] [CrossRef] [PubMed]
- Phillips, O.A.; Udo, E.E.; Varghese, R. Antimycobacterial Activities of Novel 5-(1H-1,2,3-Triazolyl)Methyl Oxazolidinones. Tuberc. Res. Treat. 2012, 2012, 289136. [Google Scholar] [CrossRef]
- de Carvalho Junior, A.R.; Oliveira Ferreira, R.; de Souza Passos, M.; da Silva Boeno, S.I.; Glória das Virgens, L.d.L.; Ventura, T.L.B.; Calixto, S.D.; Lassounskaia, E.; de Carvalho, M.G.; Braz-Filho, R.; et al. Antimycobacterial and Nitric Oxide Production Inhibitory Activities of Triterpenes and Alkaloids from Psychotria nuda (Cham. & Schltdl.) Wawra. Mol. 2019, 24, 1026. [Google Scholar] [CrossRef]
- Peng, J.; Hu, J.-F.; Kazi, A.B.; Li, Z.; Avery, M.; Peraud, O.; Hill, R.T.; Franzblau, S.G.; Zhang, F.; Schinazi, R.F.; et al. Manadomanzamines A and B: A Novel Alkaloid Ring System with Potent Activity against Mycobacteria and HIV-1. J. Am. Chem. Soc. 2003, 125, 13382–13386. [Google Scholar] [CrossRef]
- Kim, H.; Lantvit, D.; Hwang, C.H.; Kroll, D.J.; Swanson, S.M.; Franzblau, S.G.; Orjala, J. Indole Alkaloids from Two Cultured Cyanobacteria, Westiellopsis Sp. and Fischerella muscicola. Bioorg. Med. Chem. 2012, 20, 5290–5295. [Google Scholar] [CrossRef] [PubMed]
- Mo, S.; Krunic, A.; Chlipala, G.; Orjala, J. Antimicrobial Ambiguine Isonitriles from the Cyanobacterium Fischerella ambigua. J. Nat. Prod. 2009, 72, 894–899. [Google Scholar] [CrossRef] [PubMed]
- Mo, S.; Krunic, A.; Santarsiero, B.D.; Franzblau, S.G.; Orjala, J. Hapalindole-Related Alkaloids from the Cultured Cyanobacterium Fischerella ambigua. Phytochemistry 2010, 71, 2116–2123. [Google Scholar] [CrossRef]
- Raveh, A.; Carmeli, S. Antimicrobial Ambiguines from the Cyanobacterium Fischerella Sp. Collected in Israel. J. Nat. Prod. 2007, 70, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.-H.; Fan, Y.-Y.; Liu, Q.-F.; Cho, S.-H.; Pauli, G.F.; Chen, S.-N.; Yue, J.-M. Suadimins A-C, Unprecedented Dimeric Quinoline Alkaloids with Antimycobacterial Activity from Melodinus suaveolens. Org. Lett. 2019, 21, 7065–7068. [Google Scholar] [CrossRef]
- Luo, X.; Pires, D.; Aínsa, J.A.; Gracia, B.; Duarte, N.; Mulhovo, S.; Anes, E.; Ferreira, M.-J.U. Zanthoxylum capense Constituents with Antimycobacterial Activity against Mycobacterium tuberculosis in vitro and ex vivo within Human Macrophages. J. Ethnopharmacol. 2013, 146, 417–422. [Google Scholar] [CrossRef]
- Choules, M.P.; Wolf, N.M.; Lee, H.; Anderson, J.R.; Grzelak, E.M.; Wang, Y.; Ma, R.; Gao, W.; McAlpine, J.B.; Jin, Y.-Y.; et al. Rufomycin Targets ClpC1 Proteolysis in Mycobacterium tuberculosis and M. abscessus. Antimicrob. Agents Chemother. 2019, 63, e02204-18. [Google Scholar] [CrossRef]
- Gao, W.; Kim, J.-Y.; Chen, S.-N.; Cho, S.-H.; Choi, J.; Jaki, B.U.; Jin, Y.-Y.; Lankin, D.C.; Lee, J.-E.; Lee, S.-Y.; et al. Discovery and Characterization of the Tuberculosis Drug Lead Ecumicin. Org. Lett. 2014, 16, 6044–6047. [Google Scholar] [CrossRef]
- Igarashi, M.; Nakagawa, N.; Doi, N.; Hattori, S.; Naganawa, H.; Hamada, M. Caprazamycin B, a Novel Anti-Tuberculosis Antibiotic, from Streptomyces Sp. J. Antibiot. 2003, 56, 580–583. [Google Scholar] [CrossRef] [PubMed]
- Boeno, S.I.S.; Passos, M.d.S.; Félix, M.; Calixto, S.D.; Júnior, A.R.C.; Barbosa Siqueira, L.F.; Muzitano, M.F.; Braz-Filho, R.; Vieira, I.J.C. Antimycobacterial Activity of Milemaronol, a New Squalene-Type Triterpene, and Other Isolate? Nat. Prod. Commun. 2020, 15, 1934578X20925589. [Google Scholar] [CrossRef]
- Wonganuchitmeta, S.; Yuenyongsawad, S.; Keawpradub, N.; Plubrukarn, A. Antitubercular Sesterterpenes from the Thai Sponge Brachiaster Sp. J. Nat. Prod. 2004, 67, 1767–1770. [Google Scholar] [CrossRef]
- Ledroit, V.; Debitus, C.; Ausseil, F.; Raux, R.; Menou, J.-L.; Hill, B. Heteronemin as a Protein Farnesyl Transferase Inhibitor. Pharm. Biol. 2004, 42, 454–456. [Google Scholar] [CrossRef]
- Nieves, K.; Prudhomme, J.; Le Roch, K.G.; Franzblau, S.G.; Rodríguez, A.D. Natural Product-Based Synthesis of Novel Anti-Infective Isothiocyanate- and Isoselenocyanate-Functionalized Amphilectane Diterpenes. Bioorg. Med. Chem. Lett. 2016, 26, 854–857. [Google Scholar] [CrossRef]
- Wächter, G.A.; Franzblau, S.G.; Montenegro, G.; Hoffmann, J.J.; Maiese, W.M.; Timmermann, B.N. Inhibition of Mycobacterium tuberculosis Growth by Saringosterol from Lessonia nigrescens. J. Nat. Prod. 2001, 64, 1463–1464. [Google Scholar] [CrossRef]
- Gutierrez-Lugo, M.-T.; Wang, Y.; Franzblau, S.G.; Suarez, E.; Timmermann, B.N. Antitubercular Sterols from Thalia multiflora Horkel Ex Koernicke. Phytother. Res. PTR 2005, 19, 876–880. [Google Scholar] [CrossRef]
- Deng, S.; Wang, Y.; Inui, T.; Chen, S.-N.; Farnsworth, N.R.; Cho, S.; Franzblau, S.G.; Pauli, G.F. Anti-TB Polyynes from the Roots of Angelica sinensis. Phytother. Res. PTR 2008, 22, 878–882. [Google Scholar] [CrossRef]
- Mata, R.; Morales, I.; Pérez, O.; Rivero-Cruz, I.; Acevedo, L.; Enriquez-Mendoza, I.; Bye, R.; Franzblau, S.; Timmermann, B. Antimycobacterial Compounds from Piper sanctum. J. Nat. Prod. 2004, 67, 1961–1968. [Google Scholar] [CrossRef]
- Wube, A.A.; Bucar, F.; Gibbons, S.; Asres, K. Sesquiterpenes from Warburgia ugandensis and Their Antimycobacterial Activity. Phytochemistry 2005, 66, 2309–2315. [Google Scholar] [CrossRef]
- Chomcheon, P.; Wiyakrutta, S.; Sriubolmas, N.; Ngamrojanavanich, N.; Isarangkul, D.; Kittakoop, P. 3-Nitropropionic Acid (3-NPA), a Potent Antimycobacterial Agent from Endophytic Fungi: Is 3-NPA in Some Plants Produced by Endophytes? J. Nat. Prod. 2005, 68, 1103–1105. [Google Scholar] [CrossRef]
- Goodfellow, M.; Stach, J.E.M.; Brown, R.; Bonda, A.N.V.; Jones, A.L.; Mexson, J.; Fiedler, H.-P.; Zucchi, T.D.; Bull, A.T. Verrucosispora Maris Sp. Nov., a Novel Deep-Sea Actinomycete Isolated from a Marine Sediment Which Produces Abyssomicins. Antonie Van Leeuwenhoek 2012, 101, 185–193. [Google Scholar] [CrossRef]
- Freundlich, J.S.; Lalgondar, M.; Wei, J.-R.; Swanson, S.; Sorensen, E.J.; Rubin, E.J.; Sacchettini, J.C. The Abyssomicin C Family as in vitro Inhibitors of Mycobacterium tuberculosis. Tuberc. Edinb. Scotl. 2010, 90, 298–300. [Google Scholar] [CrossRef] [PubMed]
- Bister, B.; Bischoff, D.; Ströbele, M.; Riedlinger, J.; Reicke, A.; Wolter, F.; Bull, A.T.; Zähner, H.; Fiedler, H.-P.; Süssmuth, R.D. Abyssomicin C-A Polycyclic Antibiotic from a Marine Verrucosispora Strain as an Inhibitor of the p-Aminobenzoic Acid/Tetrahydrofolate Biosynthesis Pathway. Angew. Chem. Int. Ed Engl. 2004, 43, 2574–2576. [Google Scholar] [CrossRef]
- Wang, J.-F.; Dai, H.-Q.; Wei, Y.-L.; Zhu, H.-J.; Yan, Y.-M.; Wang, Y.-H.; Long, C.-L.; Zhong, H.-M.; Zhang, L.-X.; Cheng, Y.-X. Antituberculosis Agents and an Inhibitor of the Para-Aminobenzoic Acid Biosynthetic Pathway from Hydnocarpus anthelminthica Seeds. Chem. Biodivers. 2010, 7, 2046–2053. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Singh, A.; Sharma, U.; Singh, D.; Dobhal, M.P.; Singh, S. Anti-Mycobacterial Activity of Plumericin and Isoplumericin against MDR Mycobacterium tuberculosis. Pulm. Pharmacol. Ther. 2013, 26, 332–335. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Case, R.J.; Wang, Y.; Zhang, H.-J.; Tan, G.T.; Van Hung, N.; Cuong, N.M.; Franzblau, S.G.; Soejarto, D.D.; Fong, H.H.; et al. Anti-Tuberculosis Constituents from the Stem Bark of Micromelum hirsutum. Planta Med. 2005, 71, 261–267. [Google Scholar] [CrossRef]
- Uc-Cachón, A.H.; Borges-Argáez, R.; Said-Fernández, S.; Vargas-Villarreal, J.; González-Salazar, F.; Méndez-González, M.; Cáceres-Farfán, M.; Molina-Salinas, G.M. Naphthoquinones Isolated from Diospyros anisandra Exhibit Potent Activity against Pan-Resistant First-Line Drugs Mycobacterium tuberculosis Strains. Pulm. Pharmacol. Ther. 2014, 27, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, A.; Ghosh, S.; Shaw, R.; Patra, M.M.; Calcuttawala, F.; Mukherjee, N.; Das Gupta, S.K. Mycobacterium tuberculosis Thymidylate Synthase (ThyX) Is a Target for Plumbagin, a Natural Product with Antimycobacterial Activity. PLoS ONE 2020, 15, e0228657. [Google Scholar] [CrossRef]

Table 1.
MIC and IC50 values for hapalindoles and their derivatives isolated from cyanobacteria Westiellopsis sp. and Fischerella muscicola against Mycobacterium tuberculosis H37Rv and Mycobacterium smegmatis.
Table 1.
MIC and IC50 values for hapalindoles and their derivatives isolated from cyanobacteria Westiellopsis sp. and Fischerella muscicola against Mycobacterium tuberculosis H37Rv and Mycobacterium smegmatis.
| Compound | MIC (µM) | Cytotoxicity (IC50 (µM, Vero) | |
| M. tuberculosis H37Rv | M. smegmatis | ||
| Hapalindole A (4) | <0.6 | 18.2 | 31.9 |
| Hapalindole I (5) | 2 | >100 | >100 |
| Hapalindole X (6) | 2.5 | 78.8 | 35.2 |
Table 2.
MIC and IC50 values for ambiguine isonitriles isolated from Fischerella ambigua against Mycobacterium tuberculosis H37Rv and Mycobacterium smegmatis.
Table 2.
MIC and IC50 values for ambiguine isonitriles isolated from Fischerella ambigua against Mycobacterium tuberculosis H37Rv and Mycobacterium smegmatis.
| Compound | MIC (µM) | Cytotoxicity (IC50, µM, Vero) | |
| M. tuberculosis H37Rv | M. smegmatis | ||
| Ambiguine C isonitrile (10) | 7.0 | 59.6 | 78.3 |
| Ambiguine M isonitrile (11) | 7.5 | 25.8 | 79.8 |
| Ambiguine E isonitrile (12) | 21 | 1.4 | 42.6 |
Table 3.
MIC values of organic acids or their derivatives isolated from Hydnocarpus anthelminthica against Mycobacterium tuberculosis H37Rv and Mycobacterium bovis BCG.
Table 3.
MIC values of organic acids or their derivatives isolated from Hydnocarpus anthelminthica against Mycobacterium tuberculosis H37Rv and Mycobacterium bovis BCG.
| Compound | MIC (µM) | |
| M. tuberculosis H37Rv | M. bovis BCG | |
| Anthelminthicin A (40) | 5.54 | 13.8 |
| Anthelminthicin B (41) | 16.7 | 0.52 |
| Anthelminthicins C (42) | 4.38 | 11.2 |
| Chaulmoogric acid (43) | 9.82 | 2.36 |
| Ethyl chaulmoograte (44) | 16.8 | 0.26 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



