
Submitted:
31 July 2025
Posted:
01 August 2025
You are already at the latest version
Abstract
Highly Pathogenic Avian Influenza (HPAI) viruses, particularly H5N1, continue to pose a major global threat due to their widespread circulation and high mortality rates in birds. Management of HPAI is complicated by challenges in conserving migratory bird populations, sustaining poultry production, and uncertainties in disease dynamics. Structured decision-making frameworks, such as those based on the PrOACT model, are recommended to improve outbreak response and guide critical actions, especially when HPAI virus (HPAIV) detections occur in sensitive areas like wildlife refuges. Surveillance data from late 2024 to early 2025 show persistent HPAI activity, with 743 detections across 22 European countries and beyond, and notable outbreaks in poultry in nations like Hungary, Iceland, and the UK. The proximity of poultry farms to water sources increases environmental contamination risks. Meanwhile, HPAI A(H5N1) and other H5Nx viruses have been detected in a wide range of mammalian species globally, raising concerns about mammalian adaptation due to mutations like E627K and D701N in the PB2 protein. Human infections with H5N1 have also been reported, with recent cases in North America highlighting zoonotic transmission risks. Molecular studies emphasize the importance of monitoring genetic variations associated with increased virulence and antiviral resistance. Preventive strategies focus on biosafety, personal protective measures, and vaccine development for both avian and human populations. Ongoing genetic characterization and vigilant surveillance remain critical to managing the evolving threat posed by HPAI viruses.
Keywords:
highly pathogenic avian influenza (HPAI)
; H5N1
; H7N9
; zoonotic transmission
; One Health approach
; viral evolution
; global surveillance
1. Introduction
1.1. Background on Avian Influenza Viruses
Avian influenza viruses (AIVs) are members of the Orthomyxoviridae family, classified under the genus Alphainfluenzavirus (formerly known as influenza A virus). These enveloped viruses possess a segmented RNA genome consisting of eight gene segments and are capable of infecting a wide range of vertebrate hosts, primarily avian species but occasionally mammals and humans. AIVs are characterized by combinations of two surface glycoproteins: hemagglutinin (HA) and neuraminidase (NA) [1,27]. In birds, 16 HA and 9 NA antigens have been identified, enabling the classification of numerous subtypes (e.g., H5N1, H7N8). To date, 19 HA (H1–H19) and 11 NA (N1–N11) subtypes have been recognized, with H17N10 and H18N11 isolated exclusively from bats [2].
Based on their virulence in chickens, AIVs are further categorized into two groups: low pathogenic avian influenza viruses (LPAIVs) and highly pathogenic avian influenza viruses (HPAIVs) [1]. LPAIVs typically induce mild or asymptomatic infections, occasionally causing respiratory symptoms, weight loss, or decreased egg production in poultry [1]. In contrast, HPAIVs, historically referred to as "fowl plague," are associated with severe systemic disease and high mortality rates. To date, only viruses of the H5 and H7 subtypes have been shown to evolve into highly pathogenic forms [2].
Among these, HPAI H5N1 has emerged as a particularly concerning zoonotic and pandemic threat, demonstrating expanded host range, enhanced tissue tropism, and increased virulence. The first major zoonotic outbreak of HPAI H5N1 occurred during an epizootic in Hong Kong in 1997 (Table 1), resulting in 18 human infections and 6 fatalities. Although an earlier outbreak of HPAI H5N1 among poultry was recorded in Scotland in 1959, the 1997 Hong Kong event marked the first known human infections [3]. Following a brief period of quiescence, HPAI H5N1 re-emerged in 2003, subsequently establishing a global presence with incursions into East Asia, Southeast Asia, West Asia, and Africa between 2003 and 2008 [4].
More recently, a Eurasian-origin H5N1 virus belonging to clade 2.3.4.4b was introduced into North America via migratory birds in late 2021, resulting in an unprecedented epizootic affecting poultry, wild birds, and numerous mammalian species. By late 2022, the outbreak expanded into Mexico, Central America, and South America. Concurrently, between 2020 and 2021, Europe, Asia, and Africa witnessed a novel wave of clade 2.3.4.4b H5N1 activity, severely impacting wild and domestic avian populations [3].
Separately, a novel reasserting H7N9 virus emerged in March 2013 in China’s Shanghai and Anhui provinces, linked to live poultry markets. Genetic analyses revealed that H7N9 arose through multiple assortment events involving wild bird viruses, with internal genes derived from endemic H9N2 viruses and external HA and NA segments from separate lineages [21]. Domestic chickens subsequently became the primary reservoir for H7N9 [5]. Despite being classified as low pathogenic in birds, H7N9 caused 131 confirmed human infections and 36 deaths within two months of its emergence, raising global concern. The H7N9 epidemic in China evolved through five distinct waves between 2013 and 2017 (Table 1), culminating in the emergence of highly pathogenic variants during the fifth wave. Although large-scale outbreaks have since diminished, H7N9 viruses remain a latent threat to global public health [6].
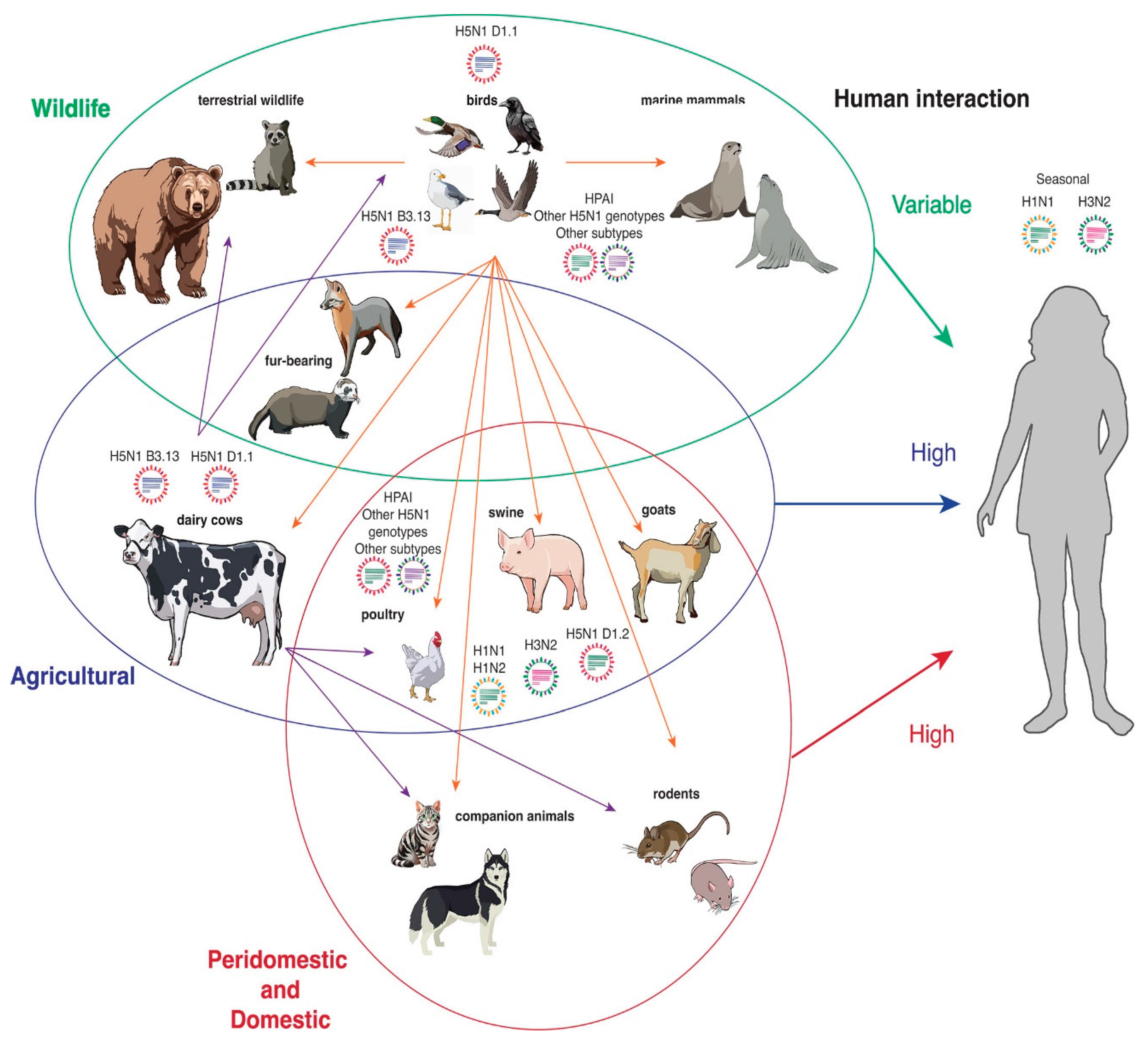
Figure 1.
Animal species infected with H5N1. Wildlife species are encircled in green, agricultural species are encircled in blue, and peridomestic and domestic species are circled in red. Orange arrows depict transmission from an avian host. Purple arrows depict transmission from a bovine host. Viruses indicate influenza. A strains known to circulate in species that present a high risk for re-assortment [18 (Journal of Virology)].
Figure 1.
Animal species infected with H5N1. Wildlife species are encircled in green, agricultural species are encircled in blue, and peridomestic and domestic species are circled in red. Orange arrows depict transmission from an avian host. Purple arrows depict transmission from a bovine host. Viruses indicate influenza. A strains known to circulate in species that present a high risk for re-assortment [18 (Journal of Virology)].
1.2. Significance of the Topic
Avian influenza viruses, particularly HPAI H5N1 and H7N9, continue to pose substantial threats to global health due to their extensive circulation, high mortality rates (Table 2 & Graph 1), and pandemic potential. Persistent viral circulation among avian populations serves as a reservoir, facilitating spillover into mammals, including species of agricultural and economic importance, as well as humans [7].
Recent epizootics, notably the HPAI H5N1 outbreaks involving mammals such as sea lions and dairy cattle, underscore the urgent need for comprehensive Virological surveillance and intersectoral response strategies. The growing number of human infections signals an increasing risk of viral adaptation toward more efficient human-to-human transmission. Although direct avian-to-human transmission remains relatively rare (Table 1), the expanding host range and unprecedented scale of current outbreaks heighten concerns about viral evolution and persistence across species barriers [8].
Of particular alarm is the emergence of HPAI H5N1 infections in dairy cattle. Should cattle become long-term reservoirs, they could facilitate further viral reassortment and adaptation, significantly escalating the risk of zoonotic transmission. Additionally, cross-species transmission among mammals raises the specter of viral mutations favoring sustained mammalian or human transmission [9].
Similarly, H7N9 remains classified as a high-risk virus for pandemic potential. Should either H5 or H7 subtype viruses acquire mutations enabling efficient aerosolized transmission among humans (Table 3), they could precipitate a pandemic with catastrophic public health and socioeconomic consequences [10].
Understanding the epidemiological dynamics, molecular evolution, and zoonotic potential of H5N1 and H7N9 is therefore critical for enhancing pandemic preparedness, informing vaccination strategies, and implementing effective public health interventions. The ongoing global outbreak of avian influenza highlights the fragile balance between human activity and viral ecology, demanding vigilant international cooperation [11].
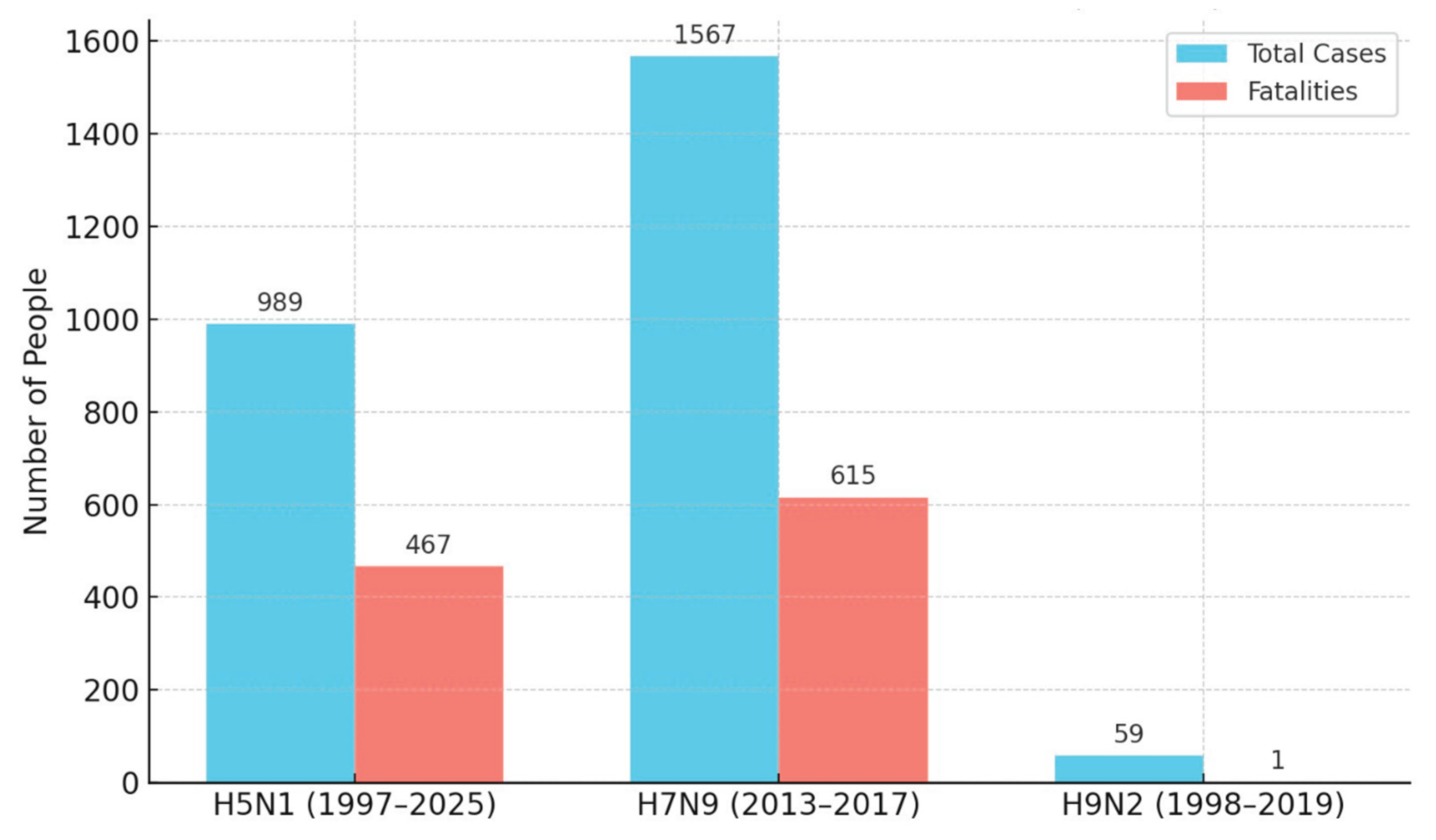
Graph 1.
Proportion of Cumulative Human Cases and deaths for H5N1, H7N9, and H9N2.
1.3. Objectives of the Review
This review seeks to:
- i.
- Examine the epidemiological patterns, virological properties, and clinical outcomes associated with H5N1 and H7N9 infections;
- ii.
- Critically evaluate current global surveillance systems, outbreak response strategies, and mitigation efforts;
- iii.
- Identify critical knowledge gaps and vulnerabilities in pandemic preparedness frameworks;
- iv.
- Propose evidence-based recommendations to enhance early detection, containment, and prevention of future avian influenza outbreaks.
2. Epidemiology of H5n1 and H7n9
2.1. Global Distribution and Spread
2.1.1. H5N1: -
Highly pathogenic avian influenza virus (HPAIV) H5N1 first gained international attention following an epizootic event in Hong Kong in 1997 [12], where it caused 18 human infections and six deaths (Table 2). Although the initial detection of HPAI H5N1 in poultry was documented earlier in Scotland in 1959, the 1997 outbreak marked the virus's first recognized zoonotic transmission to humans [13].
After a temporary hiatus, HPAI H5N1 re-emerged in 2003 and has since become enzootic in several regions, particularly across Europe, Asia, and Africa. Between 2003 and 2008, the virus expanded its geographic footprint to East Asia, Southeast Asia, West Asia, and North Africa. While cases remained relatively low across most of Asia between 2013 and 2015, Egypt experienced a marked surge during 2014–2015 [14].
A major shift occurred between 2020 and 2021, with the emergence of HPAI H5N1 clade 2.3.4.4b across Europe, Asia, and Africa. This lineage caused widespread outbreaks among wild and domestic birds, resulting in the largest recorded HPAI epidemic during the 2021–2022 season. Over 2,000 outbreaks were reported across 37 European countries, necessitating the culling of more than 40 million birds [15].
In late 2021, Eurasian-origin H5N1 clade 2.3.4.4b entered North America via migratory birds, igniting an unprecedented epizootic in poultry, wild birds, and mammals. From 2022 onwards, the virus expanded further into Mexico, Central America, and South America [16]. Between January 2022 and June 2023, H5N1 infections were confirmed in over 7,000 wild birds across all 50 U.S. states and over 800 poultry flocks across 47 states—an unprecedented geographic spread not seen since 2016 [13].
Today, clade 2.3.4.4b H5N1 viruses are endemic in many regions and have been detected on every continent except Australia, extending even into urban areas and Antarctica. The ancestral lineage of these viruses can be traced back to A/goose/Guangdong/1/96, from which multiple genetic clades have evolved [15].
2.1.2. H7N9: -
In March 2013, a novel reassortant avian influenza A (H7N9) virus was first detected in Shanghai and Anhui provinces of China. Genetic analyses revealed that H7N9 originated through multiple reassortment events in wild birds, acquiring internal genes from circulating H9N2 viruses and external HA and NA segments from separate sources. The virus subsequently adapted to domestic poultry, particularly chickens, which became its primary reservoir [17].
Although low pathogenic in birds, H7N9 caused 131 human infections and 36 fatalities within just two months of its emergence (Table 2), raising considerable international concern. Between 2013 and 2017, H7N9 outbreaks occurred in five epidemic waves [5], with each wave beginning in the winter (October–November) and subsiding by early summer (June). The 2016–2017 wave was the most severe, accounting for 759 confirmed human cases due to a broader geographical spread across China.
During the fifth wave, a highly pathogenic variant of H7N9 (HP-H7N9) emerged, increasing the risk to both poultry and humans. Two genetically distinct lineages—the Pearl River Delta and Yangtze River Delta lineages—have since been identified. Although large-scale epidemics have subsided, sporadic poultry outbreaks continue, and H7N9 remains a latent public health threat [19].
2.2. Case Fatality Rates and Clinical Outcomes
2.2.1. H5N1: -
Between 2003 and July 2023, the World Health Organization (WHO) documented 878 confirmed human cases of HPAI H5N1 infection across 23 countries, resulting in 458 fatalities—a case fatality rate (CFR) of approximately 52.2%. The majority of these infections occurred prior to 2016. From 2022 to July 2023 alone, 14 cases and two fatalities were reported (Table 2).
Historically, the CFR for H5N1 infections has been reported at around 60%, substantially higher than that of pandemic influenza viruses. Clinical outcomes are often severe, characterized primarily by viral pneumonia and progression to acute respiratory distress syndrome (ARDS). The pathogenesis of HPAI H5N1 in humans is complex and multifactorial, involving direct viral cytopathic effects and dysregulated host immune responses.
2.2.2. H7N9: -
The initial outbreak of H7N9 in 2013 led to 131 confirmed human cases and 36 deaths within a short timeframe. As of May 2013, 130 confirmed cases (excluding mainland China) had been reported, with at least 27 deaths, indicating a mortality rate of approximately 20% (Table 2).
Across five epidemic waves from 2013 to 2017, H7N9 caused 1,567 confirmed human infections and 615 deaths, corresponding to a cumulative CFR of approximately 39.2%. Severe disease manifestations primarily include viral pneumonia and ARDS. Unlike H5N1, H7N9 infections are often associated with underlying comorbidities.
Low-pathogenic H7N9 viruses were predominant during the first four waves, while highly pathogenic strains emerged in the fifth wave. Although both LP-H7N9 and HP-H7N9 viruses infected humans, current evidence does not suggest a significant difference in virulence or transmissibility between these two variants in human hosts.
2.3. Zoonotic Potential
Role of Poultry Farming, Live Bird Markets, and Migratory Birds
Avian influenza viruses (AIVs), particularly subtypes H1 through H16, are naturally maintained in wild aquatic bird populations, which serve as their primary reservoir. Traditionally, wild waterfowl carried AIVs subclinical. However, since 2005, HPAI H5N1 has increasingly caused morbidity and mortality among wild birds, altering the epidemiological landscape [20].
H5N1 viruses, especially clade 2.3.4.4b, have been frequently detected in wild birds and have caused extensive die-offs, including among endangered species. Transmission from wild birds to domestic poultry occurs primarily via the fecal–oral route, direct contact, and, to a lesser extent, aerosol transmission [3,20].
Migratory birds have played a critical role in disseminating HPAI viruses across continents. Notably, Eurasian-origin clade 2.3.4.4b viruses were introduced into North America in late 2021 via trans-Atlantic migratory pathways, subsequently spreading across the Americas. Wild bird infections with clade 2.3.4.4b viruses have been characterized by unusually high morbidity and mortality [22].
The H7N9 virus, by contrast, is closely associated with live bird markets (LBMs), which act as amplification hubs for viral transmission. Phylogenetic studies suggest that H7N9 emerged through re-assortment in wild birds before establishment in domestic poultry. LBMs have been crucial in maintaining AIV circulation and facilitating zoonotic spillover, particularly given the frequent detection of H9N2 and H7N9 viruses in these environments [23].
Transmission of H5N1 and H7N9 to humans primarily occurs through direct or indirect contact with infected birds or contaminated environments, especially via exposure of mucous membranes (eyes, nose, mouth) to infected secretions [24].
The expanding host range of clade 2.3.4.4b viruses to include domestic and wild mammals significantly raises concerns about further zoonotic transmission events. Although bird-to-human transmission remains infrequent, the unprecedented host range and ecological reach of recent outbreaks substantially increase the likelihood of viral persistence, evolution, and eventual adaptation to humans [25].
3. Evolution of H5N1 and H7N9
The evolutionary dynamics of H5N1 and H7N9 viruses are central to understanding their epidemiology, influencing their geographic distribution, pathogenicity, host range, and potential for zoonotic transmission [26]. Both viruses belong to the Orthomyxoviridae family and the genus Alphainfluenzavirus, characterized by segmented genomes composed of eight single-stranded RNA segments [27].
3.1. Genetic Characteristics
3.1.1. Genomic Differences Between H5N1 and H7N9
H5N1 and H7N9 viruses are classified by their specific hemagglutinin (HA) and neuraminidase (NA) surface glycoproteins—H5 and N1 for H5N1, and H7 and N9 for H7N9.
The H5N1 viruses, particularly those of the A/goose/Guangdong/1/1996 (Gs/Gd) lineage, have diversified extensively into numerous clades and subclades. In contrast, the emergence of H7N9 was a distinct event resulting from multiple reassortment processes. Phylogenetic analyses reveal that H7N9 originated through the acquisition of internal genes from endemic H9N2 viruses, with its HA and NA genes likely derived independently from wild avian species. Notably, the internal gene cassette inherited from H9N2 significantly contributes to the mammalian pathogenicity of H7N9 [28].
After emergence, H7N9 viruses successfully established domestic chickens as the principal reservoir, facilitating their amplification and dissemination.
3.1.2. Mechanisms of Antigenic Drift and Antigenic Shift Between H5N1 and H7N9
Influenza A viruses evolve through two principal mechanisms:
- Antigenic Drift
Antigenic drift involves the gradual accumulation of point mutations—particularly non-synonymous mutations—in the HA and NA genes. These mutations alter critical antigenic sites, allowing the virus to evade host immune responses (Table 3). Antigenic drift is an ongoing process observed across H5N1 clades and H7N9 lineages, driving continuous viral evolution and contributing to epidemic persistence [29].
- Antigenic Shift
Antigenic shift refers to the reassortment of gene segments between two or more influenza viruses co-infecting the same host cell. This process, enabled by the segmented nature of the influenza genome, can give rise to novel viral genotypes with new HA and/or NA combinations. The emergence of H7N9 exemplifies antigenic shift (Table 3), originating from multiple reassortment events involving H7, N9, and H9N2 viruses. Similarly, reassortment has played a pivotal role in the evolution of Gs/Gd-lineage H5N1 viruses. H9N2 viruses, in particular, have been major donors of internal gene segments to several zoonotic influenza strains, including H5N1 and H7N9 [28,30].
3.2. Adaptation to Human Hosts
Avian influenza viruses are naturally adapted to wild aquatic birds, preferentially binding to α2,3-linked sialic acid (SA) receptors in the avian gut. In contrast, human-adapted influenza viruses target α2,6-linked SA receptors found predominantly in the human upper respiratory tract. For successful human infection and transmission, avian viruses must acquire mutations that modify their receptor binding specificity [34].
Key Mutations Enhancing Mammalian Adaptation:
- i.
-
H5N1 AdaptationsAdaptation of H5N1 to human hosts involves several key genetic changes:
- HA mutations, such as Q222L and G224S, promote a shift from α2,3- to α2,6-SA receptor binding [35].
- Additional mutations at HA positions 129 and 134 can further modulate binding affinity.
- Polymerase complex mutations, notably E627K, D701N, and S714R in PB2, enhance viral replication efficiency in mammalian cells [36].
- Mutations in NA can improve viral fitness by enhancing replication in human airway epithelial cells and facilitating immune escape.
- ii.
-
H7N9 AdaptationsThe H7N9 virus has similarly exhibited key adaptations:
- Early isolates demonstrated dual receptor specificity, with mutations such as G186V and Q226L in HA facilitating α2,6-SA binding [37].
- The PB2-E627K substitution has been implicated in increased polymerase activity and replication in mammals.
- Additional mutations in the NP, M, PA, and NA genes contribute to enhanced virulence, replication efficiency, and modulation of the host immune response [38].
- NS1 protein mutations have been associated with antagonism of interferon responses, supporting viral persistence in human hosts.
While these adaptations increase zoonotic potential, neither H5N1 nor H7N9 has yet acquired the ability for efficient or sustained human-to-human transmission [39].
3.3. Antigenic Diversity and Vaccine Challenges
The significant genetic and antigenic evolution observed in H5N1 and H7N9 viruses presents formidable challenges for vaccine development and pandemic preparedness.
Impact of Viral Diversity on Vaccine Development
- H5N1
Gs/Gd-lineage H5N1 viruses have diversified into numerous clades and subclades, each with distinct antigenic properties. The currently dominant clade, 2.3.4.4b, has achieved near-global distribution through wild bird migrations. The continuous emergence of new variants necessitates regular updating of vaccine candidates to ensure antigenic match, complicating mass vaccination efforts.
- H7N9
Over successive epidemic waves, H7N9 viruses have evolved significant genetic and antigenic diversity. For example, vaccines developed based on first-wave strains exhibited poor cross-reactivity against fifth-wave viruses, requiring the development of updated candidate vaccine viruses. Regional diversification and reassortment with H9N2 and other viruses further complicate vaccine design.
Additionally, the high rates of antigenic drift, especially within HA and NA proteins, and reassortment with co-circulating avian viruses such as H9N2, underscore the need for continuous surveillance and the development of broadly protective or "universal" influenza vaccines.
3.4. Current Vaccines and Their Limitations
Vaccination remains a cornerstone of pandemic preparedness against zoonotic influenza viruses like H5N1 and H7N9. Despite substantial advances, several challenges hinder the full effectiveness of current vaccine strategies [40].
3.4.1. H5N1 Vaccine Candidates and Limitations
- i.
-
Approved VaccinesSeveral H5N1 vaccines have been licensed, primarily for pandemic stockpiling or high-risk occupational groups [41]:
- Audenz (FDA, 2020): An adjuvanted, cell-culture-derived H5N1 vaccine for individuals aged six months and older.
- Sanofi Pasteur H5N1 Vaccine (FDA, 2007): Approved for adults aged 18–64 at elevated risk.
- GSK Adjupanrix and Prepandemic Vaccines (EU-approved): Based on A/VietNam/1194/2004 (H5N1) strain.
- Seqirus Vaccines (Celldemic, Incellipan): EMA-approved for avian influenza preparedness.
- CSL Seqirus H5N8 Vaccine: Targets clade 2.3.4.4b HA, with N8 NA, using an MF59 adjuvant.
Research is also advancing alternative platforms, including recombinant HA vaccines, live-attenuated formulations, and mRNA vaccines.
LIMITATIONS
- Antigenic Drift: Rapid viral evolution necessitates frequent updates to vaccine strains.
- Cross-Protection Gaps: Vaccines targeting one clade may not be effective against others.
- Low Immunogenicity: Most H5N1 vaccines require adjuvants and prime-boost regimens.
- Limited Field Data: Real-world effectiveness in preventing disease or transmission remains unproven.
- Stockpile Constraints: Current global vaccine reserves are insufficient for large-scale deployment.
- Low Uptake: Vaccination rates among high-risk occupational groups remain poor.
- Animal Model Limitations: Predictive value of mRNA vaccine studies in ferrets has not translated reliably to humans.
3.4.2. H7N9 Vaccine Candidates and Limitations
Poultry and Human Vaccination Efforts [42].
- Poultry: China introduced a bivalent inactivated H5/H7 vaccine, reducing human cases post-wave 5, though sterilizing immunity remains elusive.
-
Human Vaccines: Multiple candidates have been developed:
- Inactivated Vaccines: Require high doses and adjuvants for robust protection.
- Live-Attenuated Vaccines (LAIVs): Offer stronger T-cell responses and heterosubtypic protection potential.
- Virus-Like Particles (VLPs): Show promise for safety and immunogenicity in early trials.
LIMITATIONS
- Antigenic Drift and Evolution: Ongoing viral changes require frequent updates to vaccine compositions.
- Diverse Viral Lineages: Genetic variability complicates the design of broadly protective vaccines.
- Need for Universal Vaccines: Broader cross-protection strategies remain a critical unmet need.
- Suboptimal Immune Responses: Current vaccines often fail to stimulate sufficient neutralizing antibody and NA-specific responses.
- Persistent Threat: Despite decreased human cases, H7N9 and related viruses continue to circulate in poultry and wild birds, necessitating ongoing vigilance.
4. Clinical Manifestations and Diagnosis
4.1. Symptoms and Disease Progression:
Equally HPAI H5N1 and H7N9 viruses can cause significant illness in humans, often presenting initially with symptoms characteristic of typical influenza. However, they can rapidly progress to severe disease [43].
Common Symptoms:
- i.
- Initial symptoms often include influenza-like illness such as fever (including high fever), cough, muscle or body aches (myalgia), headaches, and fatigue [44].
- ii.
- Other symptoms can include sore throat and shortness of breath or difficulty breathing (dyspnea).
- iii.
- Diarrhea has been reported in both infections, occurring in more than 50% of patients in one H5N1 series from Vietnam but less than 10% in others. It is listed as a less common symptom for H5N1 and among typical flu symptoms for H7N9.
- iv.
- Nausea and vomiting can also occur, sometimes following initial flu-like symptoms in H5N1 [44].
4.2. Differences in Clinical Presentation and Complications:
Severity: Both can cause severe disease. Historically, H5N1 was often associated with severe disease and had a high case fatality rate (CFR), estimated at 48% among reported cases since 2003. H7N9 also has a notable CFR, though it varies depending on the time period and location, with one report noting a nationwide CFR of 39.6% compared to ~13.3% in Shenzhen, China [45], potentially due to intervention measures. Recent human infections with H5N1 clade 2.3.4.4b have frequently been clinically mild and self-resolving, although severe and fatal cases still occur globally. The reasons for the variation in outcome are likely multi-factorial, including virus genotype, duration and route of exposure, viral load, individual health status, personal protective measures, and medical treatment [46].
- i.
- Disease Progression: Both viruses can rapidly attack the lower respiratory tract, leading to severe pneumonia, bilateral pulmonary infiltrates, and acute respiratory distress syndrome (ARDS).
- ii.
- Extrapulmonary Manifestations: H5N1 infection more often leads to extrapulmonary manifestations compared to pandemic influenza viruses. These can include liver impairment with elevated transaminases, particularly in severe cases. Notably, H5N1 infections are characterised by a heightened inclination towards Central Nervous System (CNS) disease manifestation when contrasted with seasonal influenza. In animal models, H5N1 can access the CNS via olfactory and trigeminal nerves, causing severe meningoencephalitis. In animals generally, HPAIV clade 2.3.4.4b spontaneous infections are characterised by remarkable neurotropism and systemic virus spread [46].
- iii.
- Uncommon Features: H7N9 infection presents some uncommon clinical features compared to H5N1 (Table 4), such as conjunctivitis and encephalopathy, while common symptoms like nasal congestion and rhinorrhea may be less apparent. Conversely, recent mild H5N1 cases in the US have predominantly presented with conjunctivitis [47].
- iv.
- Age and Comorbidities: H7N9 infection severity increases with increasing age, with mild diseases observed in young children and more severe disease in adults. Coexisting chronic medical conditions, such as hypertension and diabetes, are commonly observed in H7N9 cases. Historically, H5N1 cases have had a younger mean age and coexisting conditions were uncommon [47].
- v.
- Immune Response: Severe H7N9 and H5N1 infections are associated with high inflammatory cytokine and chemokine levels in the lungs and peripheral blood, referred to as hypercytokinemia or cytokine storm. While potentially correlated with severe disease, hypercytokinemia levels caused by H7N9 may be lower than those caused by H5N6. HP-H7N9 seemed to induce higher cytokine levels than LP-H7N9, but the difference was not significant in one study [48].
- vi.
- Fatal Outcomes: In both infections, fatal outcomes are often associated with the development of complications such as acute respiratory distress syndrome (Table 4), severe pneumonia, and multi-organ failure (including respiratory and renal failure, pulmonary haemorrhage, pneumothorax, and pancytopenia). For H5N1, fatal outcomes have been associated with high viral loads, lymphopenia, and elevated inflammatory cytokines and chemokines. For H7N9, multiple organ failure is a major cause of death. Nosocomial bacterial infections, often with antibiotic-resistant organisms, are also common in severe H7N9 cases [49].
4.3. Diagnostic Tools:
Timely and accurate diagnosis is critical for minimising morbidity and mortality and reducing pandemic potential. Various samples are used, including respiratory, rectal specimens, and serum. For H5N1, throat swabs and lower respiratory specimens are preferred over nasopharyngeal swabs due to the virus's predilection for the lower tract. For recent US H5N1 cases, conjunctival swabs showed high positivity rates, especially in patients with conjunctivitis [51].
4.3.1. RT-PCR and Real-Time PCR:
These molecular techniques are sensitive methods to identify specific gene sequences and are the most sensitive method for H5N1 detection. They provide rapid diagnosis, guiding immediate management. They can also reveal genetic sequences useful for molecular epidemiology [52].
- i.
-
Serological tests: Detection of H5N1-specific antibodies is essential for epidemiological investigations.
- Microneutralisation (MN) assay: More specific than the haemagglutination inhibition (HI) assay for specific H5N1 diagnosis. It is the preferred gold standard for serodiagnosis of H5N1, preferably confirmed by Western blotting. Most individuals develop a positive titre 3 weeks after symptom onset. MN or HI tests can be used to test for seroprevalence in healthy populations, but they need validation. Note that HI has limited value for detecting antibodies against avian viruses in humans/mammals due to low sensitivity [53].
- ii.
- Rapid diagnostic kits / Antigen detection methods: These methods, such as rapid immunochromatographic assays or direct immunofluorescence, can provide rapid diagnosis to guide immediate management. They have a sensitivity of 50–80% and specificity of 90% for virus detection. Reliability depends on factors like specimen type, quality, and timing. Enzyme immunoassays are not widely used for human diagnostics [54].
4.3.2. Virus Isolation: Considered the Gold Standard for Virus Propagation and Detection
- Egg culture: Used for virus isolation.
- Cell culture (e.g., MDCK cells): Provides highly specific laboratory diagnosis. This method is significantly more sensitive than antigen detection but requires a BSL-3 facility, special training, and approval for H5N1 due to it being a select agent in some countries. Sample storage and transportation conditions are important [55].
- iv.
-
Novel Techniques: More recently, user-friendly, cost-effective, specific, and potentially point-of-care methods are being employed, including [56]:
- LAMP (Loop-mediated Isothermal Amplification).
- RT-LAMP (Reverse Transcription Loop-Mediated Isothermal Amplification).
- SAMBA (Simple Amplification-Based Assay).
- NASBA (Nucleic Acid Sequencing-Based Amplification).
- Molecular Integration with Nanotechnology.
- Biosensors Development.
Laboratory findings often include abnormalities such as leucopenia, thrombocytopenia, and elevated liver enzymes (aminotransferase) in both H5N1 and H7N9 cases. Lower white blood cell counts are more likely in fatal H5N1 cases. Severe H7N9 infection is also associated with elevated levels of C-reactive protein (CRP), serum creatinine, myoglobin, and T cell lymphopenia [57].
4.4. Treatment Options:
Prompt initiation of antiviral therapy upon illness onset plays a critical role in reducing the mortality rate.
4.4.1. Neuraminidase Inhibitors (NAIs):
Oseltamivir (Tamiflu®) and zanamivir (Relenza®) are currently the primary treatments for H5N1 virus infections. Peramivir is also mentioned as an NAI. NAIs work by preventing the virus from exiting cells and spreading [58]. Prompt initiation of therapy is associated with reduced symptom duration, decreased reliance on antibiotics, and quicker recovery in seasonal influenza. For H5N1, observational studies support the recommendation for oseltamivir treatment. Most recent US H5N1 cases received oseltamivir, often within 48 hours of symptom onset. Reasonable use of NA inhibitors also contributed to a lower CFR in H7N9 cases in one region [59].
4.4.2. Polymerase Inhibitors:
Baloxavir marboxil (a polymerase acidic endonuclease inhibitor) is an antiviral agent. The CDC has recommended baloxavir as an interim measure in cases of antiviral resistance to oseltamivir or the appearance of symptoms after oseltamivir treatment. Baloxavir has been reported as effective in treating H5N6 infections [60].
4.4.3. Adamantanes:
These agents impede the initial phases of viral replication. They were used in the initial H5N1 outbreak in Hong Kong in 1997, but resistance is now widespread due to M2 protein mutations. Consequently, adamantanes are presently not recommended for treating influenza A due to elevated resistance levels, although susceptible strains are limited to some older clades [61].
4.4.4. Resistance:
While most A(H5N1) viruses from recent US human infections are susceptible to currently available antiviral agents, some viruses have shown mutations conferring minor decreases in susceptibility to neuraminidase inhibitors and baloxavir. The effectiveness of NAIs can be compromised by viral evolution and mutations, such as H274Y in the NA gene, which confers resistance to oseltamivir [62].
4.4.5. Supportive Care:
In severe cases, management includes comprehensive measures such as mechanical ventilation, corticosteroids, antibiotics (to treat secondary bacterial infections), and fluid infusion.
4.4.6. Future Directions:
Further exploration of antiviral therapeutics and treatment strategies is essential. Research should focus on identifying and developing novel antiviral agents with broad-spectrum activity against avian influenza viruses. Investigating host immune responses and developing immunomodulatory therapies could also aid in reducing disease severity and improving patient outcomes [63,64].
5. Global Response and Public Health Measures:
Addressing the threats posed by HPAI H5N1 and H7N9 viruses requires a multi-faceted approach encompassing robust surveillance, proactive prevention strategies, and strong international collaboration. These measures are critical for minimising morbidity and mortality, reducing the potential for pandemics, and managing the impacts on wildlife, agriculture, and human health [65].
5.1. Surveillance Systems:
Strengthening global surveillance systems for avian influenza is crucial for promptly identifying and monitoring emerging strains, particularly those with zoonotic potential. This involves continuous monitoring of wild bird populations, domestic poultry, and high-risk areas. Timely and accurate diagnosis is a critical component of surveillance for minimizing morbidity and mortality in humans [66].
Various Types of Surveillance Are Employed:
- i.
- Monitoring of animal populations: This includes surveillance data and mortality data from poultry and wild birds. For North America, this data is collated from sources like USDA-APHIS, USGS-WHISPers, and CFIA/ACIA. Large-scale animal disease data is managed by various authorities including local, state, indigenous, federal, and transnational agencies, as well as non-governmental research groups. The FAO's EMPRES-i+ database is a primary collation of international regional/country-level data, capturing approximately 30% of H5N1 disease events. WOAH-WAHIS is another reporting system used globally [1,67].
- ii.
- Human case surveillance: Countries should remain vigilant for potential human cases of avian influenza, especially in geographic areas where the virus is highly circulating in poultry, wild birds, or other animals. Healthcare workers in these areas need to be aware of the epidemiological situation and the range of symptoms associated with avian influenza infection in humans. Surveillance systems for seasonal influenza have also highlighted the importance of typing viruses to detect zoonotic avian influenza cases. In the U.S., state and local public health officials have monitored occupationally exposed persons for symptoms after exposure to potentially infected animals and collected specimens from symptomatic persons. Most recent U.S. cases were identified through symptom monitoring. Monitoring of bovine veterinary practitioners for HPAI A(H5) infections has also occurred [68].
- iii.
- Wastewater surveillance: This is another method being used, with the CDC detecting the presence of influenza A virus in wastewater in several U.S. states and cities. Virome sequencing has also identified HPAI H5N1 clade 2.3.4.4b in wastewater. However, identifying specific subtypes at the national level may be challenging with current CDC methods, and the source (avian, bovine, or human) can be uncertain [69].
- iv.
- Genetic surveillance: Ongoing genetic analysis and monitoring are needed due to genetic variations among HPAI H5N1 strains. Studying mutations and reassortment events provides insights into the virus's evolutionary potential, transmission dynamics, and pathogenicity [70].
- v.
- Serological surveillance: Detection of H5N1-specific antibodies is essential for epidemiological investigations. Serological studies, combined with mortality data, can help infer levels of flock immunity in different species. Testing for seroprevalence in healthy populations can be done using MN or HI tests, though HI has limited value for detecting antibodies against avian viruses in humans/mammals due to low sensitivity [71]. Serology testing was conducted in one U.S. state for H5N1.
Challenges exist in understanding changing disease dynamics due to limitations of detection, testing, reporting, and collation across national and international organisations. Not all sick and dead wild birds detected and reported are captured in databases. Surveillance efforts and laboratory characterisation are fundamental to assess the threat posed by emerging H5 and H7 viruses [72].
5.2. Prevention Strategies for HPAI Outbreaks and Transmission:
Range of Strategies to Prevent HPAI Outbreaks: -
A range of strategies are employed to prevent HPAI outbreaks and transmission to humans and other animals [74]:
- i.
- Minimising Exposure: The most effective approach to prevent human H5N1 infection is to minimise exposure to potential sources of the virus. This includes avoiding contact with dead, sick, or abnormal birds and mammals unless properly trained and equipped. Caution with pets is also important [73].
- ii.
- Protection for Exposed Individuals: Public health efforts focus on protecting workers exposed to potentially infected animals. This includes the implementation of prevention measures on farms, including PPE use. Despite the importance, low rates of PPE use have been noted among dairy workers [74].
- iii.
- Biosecurity Measures: Improving biosecurity measures is a priority for the poultry industry to reduce transmission to and within domestic poultry. Good farm biosecurity is paramount. This includes implementing policies for visitors [73].
- iv.
- Live Poultry Market (LPM) Interventions: LPMs have been identified as major sources and hotspots for avian influenza outbreaks and human infection. Closure of live poultry markets has been highly effective in reducing the risk of H7N9 infection in humans. Other effective interventions in the LPM system include rest days and banning live poultry overnight. Enhanced disinfection and regular closure of wet markets also reduced transmission risk [73].
- v.
-
Vaccination:
- Poultry Vaccination: Vaccination in the poultry population is practiced in many countries. A bivalent inactivated H5/H7 vaccine for chickens was introduced in China, and multiple strategies, including this vaccine, seem to have been quite successful against emerging H7N9 viruses. A vaccine against one clade of virus may not protect against other clades or subclades due to antigenic differences. Vaccinating cattle to reduce transmission is also being explored, with research teams in the early stages of developing vaccines for livestock [75].
- Human Vaccination: Development and deployment of effective vaccines against HPAI H5N1 are of paramount importance. Several H5N1 vaccines have received approval, including Audenz (FDA, 2020) for individuals aged six months and older at increased risk, Sanofi Pasteur's vaccine (FDA, 2007) for individuals aged 18-64 at elevated risk [76], GlaxoSmithKline’s prepandemic vaccine (EU, 2008), and Adjupanrix (EU, 2009). The European Medicines Agency (EMA) has recommended vaccines like Celldemic and Incellipan for use in the EU. Research focuses on improving vaccine efficacy, broadening protection, and optimising delivery strategies. mRNA vaccines are being developed for faster production and updating. The U.S. Department of Health and Human Services has plans to produce millions of doses of H5N1 vaccine. Some countries, like Finland and Austria, have made H5 vaccines available to individuals with higher exposure risks like farm workers as a precaution. Strategic deployment of vaccination is needed to minimise human cases. However, existing stockpiles may be insufficient in a pandemic scenario [77].
- vi.
-
Adaptive Management: Effective disease management responses depend on improved systems for decision making, particularly in the face of uncertainty. Formal methods of decision analysis can aid in allocating scarce resources and prioritisation of scientific inquiry to inform management and conservation actions. Management-driven scientific inquiry is urgently needed to establish recommended disease response protocols [78].
- Addressing Underlying Factors: In the longer term, addressing underlying factors such as intensive farming practices and wildlife trade, which create environments conducive to viral mutations, is crucial in preventing pandemics [3].
- Pasteurisation: The U.S. Department of Agriculture (USDA) and the Centers for Disease Control and Prevention (CDC) emphasize the critical need for pasteurisation of dairy products to mitigate the risk of human infection [79].
5.3. International Collaboration:
Addressing the challenges posed by HPAI H5N1 necessitates a proactive approach involving strengthening international collaboration and information sharing among countries [80].
5.3.1. One Health Approach:
A cohesive One Health approach encompassing wildlife, poultry, and humans is crucial for addressing disease dynamics, management response, and reducing future negative impacts. This multisectoral approach recognises the interconnectedness of the health of people, animals, plants, and the environment. Efforts must involve collaboration across human, animal, and environmental health sectors [80].
5.3.2. Information Sharing and Coordination:
Establishing robust communication networks, sharing surveillance data, and coordinating response efforts will facilitate rapid response, early containment, and effective control of outbreaks. The timely sharing of surveillance data and whole genome sequences of isolated viruses has enabled scientists globally to develop virus detection protocols, diagnostic tests, and vaccines. A global network for real-time data sharing and coordination is needed. International cooperation has been proven successful in controlling outbreaks, such as the initial H7N9 outbreak in China [80].
5.3.3. Involvement of International Organisations:
Key international organisations are involved in coordinating global response efforts. These include [79,80]:
- i.
- WHO (World Health Organization).
- ii.
- FAO (Food and Agriculture Organization of the United Nations).
- iii.
- WOAH (World Organisation for Animal Health).
- iv.
- ECDC (European Centre for Disease Prevention and Control).
- v.
- EFSA (European Food Safety Authority).
These organisations contribute through various activities, including joint assessments of public health risk, providing guidance for investigation and management of outbreaks, producing scientific reports and overviews of the situation, and advocating for preparedness measures. The WHO, FAO, and WOAH have conducted updated joint assessments of recent influenza A(H5N1) virus events (Supplementary file no. 1).
5.3.4. Collaboration Between Agencies:
Coordination is needed across multiple scales and between agencies with separate authorities. The Task Force of Joint Prevention and Control System in China, involving different institutes, responded well to the H7N9 outbreak.
Strengthening surveillance systems, increasing testing capacity, investing in research for vaccines and treatments, and fostering international cooperation are critical steps for pandemic preparedness.
6. Lessons Learned and Future Directions
Addressing the complex and evolving challenge of HPAI requires continuous learning from past outbreaks and proactive planning for future threats [81].
6.1. Insights from Past Outbreaks:
Past outbreaks of HPAI H5N1 and H7N9 have provided critical insights into the dynamics of these viruses and effective response strategies. The unprecedented epizootic of Eurasian origin H5N1 in North America since late 2021, affecting poultry, wild birds, and mammals across a wide geographic range, highlights a significant shift in viral dynamics, including widespread symptomatic infections in wild birds and changes in seasonality. This event underscores the potential for HPAI to establish endemism in new regions, as has occurred in Asia, Africa, and Europe. The magnitude and complexity of this spread emphasise the need for effective decision framing and improved systems for disease management, particularly at the interface of wildlife, poultry, and human health, recognising the challenges of complex governance structures and limited resources [81].
The experience with HPAI A(H7N9) in China, which caused five epidemic waves of human infections between 2013 and 2017, demonstrated the effectiveness of coordinated public health intervention strategies. A key lesson learned was the high efficacy of closure of live poultry markets (LPMs) in reducing the risk of H7N9 infection in humans. Other effective interventions within the LPM system included rest days and banning live poultry overnight [82]. The introduction of a bivalent inactivated H5/H7 vaccine for chickens in China also appears to have been quite successful in controlling the H7N9 epidemic, with only sporadic human cases reported since wave 5. Although the H7N9 epidemic appears to have gradually disappeared after these waves, continuous surveillance in LPMs and wild birds still detects the virus, indicating a continuous threat. The response to H7N9 serves as a valuable model for preparedness against other pandemic-related infectious diseases. Both H5 and H7 viruses have characteristics that warrant significant pandemic preparedness efforts, despite factors like HA instability that may limit efficient human-to-human transmission. The Asian lineage of H7N9 is recognised by the CDC as having the highest potential pandemic risk [83].
6.2. Emerging Threats by 2025:
As of 2025, the threat posed by HPAI H5N1 remains significant and dynamic. There is high uncertainty about the future trajectory of HPAIV disease dynamics, but the widespread geographic range and the diversity of affected species in the Americas strongly suggest the likelihood of future endemism. Based on past patterns, a continued increase in transmission and mortality in North American birds is anticipated [84]. The current situation with clade 2.3.4.4b HPAI H5N1 is concerning and presents a significant risk of an influenza pandemic. The overall number of HPAI virus detections in birds in the current 2024–2025 epidemiological year has already surpassed that of the previous year.
A particularly troubling emerging threat is the establishment of HPAI A(H5N1) virus in dairy cattle in the United States. This marks a new chapter, with alarming expansion across multiple states and affected herds as of September 2024. The potential for an pendemic presence in cattle could turn dairy farms into viral reservoirs, facilitating further evolution and increasing the risk of human transmission. Mutations associated with enhanced replication in mammals, such as the PB2 E627K mutation, have already been identified [86]. Detection of the virus in the central nervous system of infected animals, including cats, highlights the virus's broad tissue tropism and severe pathogenic potential. Beyond cattle, poultry outbreaks continue sporadically, and the persistent circulation of HPAI in wild birds in Europe is expected to continue for many years, presenting an ongoing risk to poultry. H5 viruses also remain endemic in poultry in the Middle East, and H7N9 continues to circulate in poultry in China, maintaining a threat [77].
6.3. Recommendations for 2025 and Beyond:
Responding effectively to the ongoing and future threats of HPAI requires a comprehensive, proactive, and coordinated approach. A crucial recommendation is the need for improved decision making, utilising formal analytical methods like Structured Decision Making (SDM). This framework can help management agencies and scientists frame the complex problem, articulate objectives, generate alternatives, predict consequences, and make trade-offs, particularly in the face of high uncertainty and limited resources [81]. Decision analysis can also help prioritise scientific inquiry to address critical knowledge gaps. Coordination across multiple scales, from local properties to global migratory connections, and between agencies with separate authorities is essential for effective response.
Specific Recommended Actions Include:
- i.
- Strengthening Surveillance: Strengthening global surveillance systems for avian influenza is crucial for promptly identifying and monitoring emerging strains, especially those with zoonotic potential (Supplementary file 2). This involves continuous monitoring of wild bird populations, domestic poultry, and high-risk areas. Surveillance should also aim to detect changes in virus pathogenicity and transmissibility and identify potential mammal-to-avian transmission [68,72].
- ii.
- Enhancing Genetic Monitoring: Ongoing genetic analysis and monitoring of viral evolution is needed due to genetic variations, studying mutations and reassortment events to understand evolutionary potential, transmission dynamics, and pathogenicity. Genetic characterisation should be reinforced, especially in areas with mammalian infections. Monitoring viral evolution for changes that stabilise the HA molecule is particularly important [29,35,70].
- iii.
- Advancing Vaccination Strategies: Development and deployment of effective vaccines against HPAI H5N1 are of paramount importance. Research should focus on improving vaccine efficacy, broadening protection across strains, and optimising delivery. Vaccination should be considered in regions where poultry are not vaccinated to protect production and reduce human exposure risk. Controlling the dairy cattle outbreak may require vaccination of cows or better infection control. Vaccination for animals in fur farms and enhanced biosecurity measures are also suggested [11].
- iv.
- v.
- Strengthening Biosecurity: Implementing stringent biosecurity measures and prevention strategies to minimize exposure to potential sources of the virus. This includes avoiding contact with dead, sick, or abnormal birds and mammals unless properly trained and equipped, using appropriate Personal Protective Equipment (PPE) for workers exposed to potentially infected animals, implementing adequate biosafety and biosecurity at occupational settings, and improving biosecurity measures on farms [73].
- vi.
- vii.
- Applying Structured Decision-Making (SDM): Utilize SDM frameworks to guide resource allocation, prioritize research, and develop effective outbreak management protocols under uncertainty [81].
- viii.
- Adopting a One Health Approach: Adopting a cohesive One Health approach encompassing human, animal, and environmental health sectors is crucial for addressing disease dynamics and managing responses effectively. EFSA, ECDC and WHO have provided practical guidance for managing zoonotic outbreaks using this interdisciplinary approach [80].
- ix.
- Addressing Root Causes: Tackle systemic risk factors, including intensive animal farming, wildlife trade, and ecosystem disruption, which drive viral emergence and cross-species transmission [87].
The current outbreak serves as a stark reminder that effective and proactive response is necessary to avert potential global health crises. Surveillance efforts and laboratory characterisation remain fundamental to assessing the threat posed by emerging H5 and H7 viruses [81].
7. Conclusions
Highly pathogenic avian influenza (HPAI), particularly H5N1 and H7N9 viruses, continues to pose a dynamic and significant threat to global health. Insights from past outbreaks and assessment of current and emerging risks reinforce the urgent need for sustained vigilance, coordinated action, and strategic preparedness.
Lessons learned from previous outbreaks emphasize critical strategies and challenges. The unprecedented spread of Eurasian-origin H5N1 in North America since 2021—affecting poultry, wild birds, and mammals across broad geographic regions—demonstrates the virus's ability to establish endemism beyond its traditional range. This event highlighted the complexity of managing disease at the wildlife–poultry–human interface, exacerbated by fragmented governance and limited resources. Similarly, the H7N9 epidemics in China underscored the effectiveness of targeted interventions, notably live poultry market closures and poultry vaccination, in reducing human infections. Despite these successes, the persistence of both H5 and H7 viruses in animal reservoirs continues to demand robust pandemic preparedness efforts.
Looking toward 2025 and beyond, the threats posed by HPAI remain substantial. The widespread geographic distribution, the expanding host range, and the increasing number of detections suggest that clade 2.3.4.4b H5N1 is likely becoming endemic in the Americas. The establishment of H5N1 infections in U.S. dairy cattle—accompanied by mammalian-adaptive mutations such as PB2 E627K and evidence of neuroinvasion—marks a concerning shift that could create new viral reservoirs and facilitate further evolution. Concurrently, sporadic poultry outbreaks, persistent wild bird infections in Europe, and continued H7N9 circulation in China underscore the sustained global threat landscape.
Effectively addressing these challenges requires a comprehensive, proactive strategy centered on:
- i.
- Strengthening surveillance systems across wildlife, poultry, cattle, and high-risk human populations to detect emerging strains and critical evolutionary changes.
- ii.
- Enhancing genetic monitoring to track mutations linked to pathogenicity, host adaptation, and transmission potential.
- iii.
- Accelerating vaccine development and deployment, with a focus on broader protection, rapid scalability, and inclusion of under-vaccinated regions and sectors.
- iv.
- Advancing antiviral research and improving therapeutic strategies.
- v.
- Implementing stringent biosecurity and exposure prevention measures at farms, markets, and occupational settings.
- vi.
- Fostering international collaboration and data sharing to enable rapid response and containment.
- vii.
- Adopting a One Health approach that integrates human, animal, and environmental health responses.
- viii.
- Investing in research and structured decision-making frameworks to prioritize critical knowledge gaps and guide evidence-based policy actions.
- ix.
- Promoting long-term structural changes in farming practices and wildlife trade management to reduce zoonotic spillover risks.
- x.
- Ensuring comprehensive testing and pasteurization of dairy products to minimize risks of foodborne transmission.
The ongoing situation serves as a powerful reminder that early, coordinated, and science-driven interventions are essential to avert future global health crises. Continued surveillance, laboratory characterization, and strategic planning will remain foundational to mitigating the risks posed by emerging H5 and H7 viruses.
Acknowledgments
The authors are grateful to the Deanship of Scientific Research, Prince Sattam bin Abdulaziz University, Al-Kharj, Saudi Arabia for its support and encouragement in conducting the research and publishing this report.
Conflict of Interest
The authors declare no conflicts of interest relevant to this article.
References
- Harvey, J.A.; Mullinax, J.M.; Runge, M.C.; Prosser, D.J. The changing dynamics of highly pathogenic avian influenza H5N1: Next steps for management & science in North America. Biological Conservation 2023, 282, 110041. [Google Scholar] [CrossRef]
- Graziosi, G.; Lupini, C.; Catelli, E.; Carnaccini, S. Highly pathogenic avian influenza (HPAI) H5 clade 2. 3. 4.4 b virus infection in birds and mammals. Animals 2024, 14, 1372. [Google Scholar] [CrossRef]
- Charostad, J.; Rukerd, M.R.Z. , Mahmoudvand, S. ; Bashash, D.; Hashemi, S.M.A., Nakhaie, M.; Zandi, K. A comprehensive review of highly pathogenic avian influenza (HPAI) H5N1: an imminent threat at doorstep. Travel medicine and infectious disease 2023, 55, 102638. [Google Scholar] [CrossRef]
- Yon, L.; Duff, J.P.; Ågren, E.O.; Erdélyi, K.; Ferroglio, E.; Godfroid, J. ; .. & Gavier-Widén, D. Recent changes in infectious diseases in European wildlife. Journal of wildlife diseases 2019, 55, 3–43. [Google Scholar] [CrossRef]
- Poovorawan, Y.; Pyungporn, S.; Prachayangprecha, S.; Makkoch, J. Global alert to avian influenza virus infection: from H5N1 to H7N9. Pathogens and global health 2013, 107, 217–223. [Google Scholar] [CrossRef]
- Nakhaie, M.; Rukerd, M.R.Z. , Shahpar, A. ; Pardeshenas, M.; Khoshnazar, S.M.; Khazaeli, M.;... & Charostad, J. A Closer Look at the Avian Influenza Virus H7N9: A Calm before the Storm?. Journal of Medical Virology 2024, 96, e70090. [Google Scholar] [CrossRef]
- Hosseinian, S.A.; Hajati, M.H. A comprehensive review of the zoonotic potential of avian influenza viruses: a globally circulating threat to pandemic influenza in human. Journal of Zoonotic Diseases 2024. [Google Scholar] [CrossRef]
- Wasik, B.R.; de Wit, E.; Munster, V.; Lloyd-Smith, J.O.; Martinez-Sobrido, L.; Parrish, C.R. Onward transmission of viruses: how do viruses emerge to cause epidemics after spillover? Philosophical Transactions of the Royal Society B 2019, 374, 20190017. [Google Scholar] [CrossRef]
- Mostafa, A.; Naguib, M.M.; Nogales, A.; Barre, R.S.; Stewart, J.P.; García-Sastre, A.; Martinez-Sobrido, L. Avian influenza A (H5N1) virus in dairy cattle: origin, evolution, and cross-species transmission. Mbio 2024, 15, e02542–24. [Google Scholar] [CrossRef]
- Umar, S. (2017). Avian influenza and co-infections: investigation of the interactions in the poultry models (Doctoral dissertation, Institut National Polytechnique de Toulouse-INPT).
- EFSA Panel on Animal Health and Animal Welfare (AHAW), ECDC, Alvarez, J. ; Boklund, A.; Dippel, S.; Dórea, F.;... & Melidou, A. Preparedness, prevention and control related to zoonotic avian influenza. EFSA Journal 2025, 23, e9191. [CrossRef]
- Redrobe, S.P. Avian influenza H5N1: A review of the current situation and relevance to zoos. International Zoo Yearbook 2007, 41, 96–109. [Google Scholar] [CrossRef]
- Charostad, J.; Rukerd, M.R.Z. , Mahmoudvand, S. ; Bashash, D.; Hashemi, S.M.A., Nakhaie, M.; Zandi, K. A comprehensive review of highly pathogenic avian influenza (HPAI) H5N1: an imminent threat at doorstep. Travel medicine and infectious disease 2023, 55, 102638. [Google Scholar] [CrossRef]
- Khan, S.A.; Imtiaz, M.A.; Islam, M.M.; Tanzin, A.Z.; Islam, A.; Hassan, M.M. Major bat-borne zoonotic viral epidemics in Asia and Africa: A systematic review and meta-analysis. Veterinary Medicine and Science 2022, 8, 1787–1801. [Google Scholar] [CrossRef]
- Graziosi, G.; Lupini, C.; Catelli, E.; Carnaccini, S. Highly pathogenic avian influenza (HPAI) H5 clade 2. 3. 4.4 b virus infection in birds and mammals. Animals 2024, 14, 1372. [Google Scholar] [CrossRef]
- Giacinti, J.A.; Signore, A.V.; Jones, M.E.; Bourque, L.; Lair, S.; Jardine, C. ; .. & Soos, C. Avian influenza viruses in wild birds in Canada following incursions of highly pathogenic H5N1 virus from Eurasia in 2021–2022. Mbio 2024, 15, e03203-23. [Google Scholar] [CrossRef]
- Pu, J.; Yin, Y.; Liu, J.; Wang, X.; Zhou, Y.; Wang, Z. ; .. & Liu, J. Reassortment with dominant chicken H9N2 influenza virus contributed to the fifth H7N9 virus human epidemic. Journal of virology 2021, 95, 10–1128. [Google Scholar] [CrossRef]
- Krammer, F.; Hermann, E.; Rasmussen, A.L. Highly pathogenic avian influenza H5N1: history, current situation, and outlook. Journal of Virology 2025, 99, e02209-24. [Google Scholar] [CrossRef] [PubMed]
- Quan, C.; Shi, W.; Yang, Y.; Yang, Y.; Liu, X.; Xu, W. ; .. & Bi, Y. New threats from H7N9 influenza virus: spread and evolution of high-and low-pathogenicity variants with high genomic diversity in wave five. Journal of virology 2018, 92, 10–1128. [Google Scholar] [CrossRef]
- To, K.K.; Ng, K.H.; Que, T.L.; Chan, J.M.; Tsang, K.Y.; Tsang, A.K. ; .. & Yuen, K.Y. Avian influenza A H5N1 virus: a continuous threat to humans. Emerging microbes & infections 2012, 1, 1–12. [Google Scholar] [CrossRef]
- Peacock, T.H.P. , James, J. ; Sealy, J.E.; Iqbal, M. A global perspective on H9N2 avian influenza virus. Viruses 2019, 11, 620. [Google Scholar] [CrossRef]
- Alkie, T.N.; Byrne, A.M.; Jones, M.E.; Mollett, B.C.; Bourque, L.; Lung, O. ; .. & Berhane, Y. Recurring trans-Atlantic incursion of clade 2.3. 4.4 b H5N1 viruses by long distance migratory birds from Northern Europe to Canada in 2022/2023. Viruses 2023, 15, 1836. [Google Scholar] [CrossRef]
- Feoktistova, S.G.; Ivanova, A.O.; Degtyarev, E.P.; Smirnova, D.I.; Volchkov, P.Y.; Deviatkin, A.A. Phylogenetic Insights into H7Nx Influenza Viruses: Uncovering Reassortment Patterns and Geographic Variability. Viruses 2024, 16, 1656. [Google Scholar] [CrossRef]
- Rather, M.A.; Hassan, A.; Aman, M.; Gul, I.; Mir, A.H.; Potdar, V. ; Molecular and ecological determinants of mammalian adaptability in avian influenza virus. Infection 2025, 1–27. [Google Scholar] [CrossRef]
- Zhang, Z.; Lei, Z. The alarming situation of highly pathogenic avian influenza viruses in 2019–2023. Global Medical Genetics 2024, 11, 200–213. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chen, Y.; Shan, N.; Wang, X.; Lin, S.; Ma, K. ; & Qi, W. Genetic diversity, phylogeography, and evolutionary dynamics of highly pathogenic avian influenza A (H5N6) viruses. Virus Evolution 2020, 6, veaa079. [Google Scholar] [CrossRef] [PubMed]
- Ilyicheva, T.N.; Netesov, S.V.; Gureyev, V.N. COVID-19, Influenza, and Other Acute Respiratory Viral Infections: Etiology, Immunopathogenesis, Diagnosis, and Treatment. Part I. COVID-19 and Influenza. Molecular genetics, microbiology and virology 2022, 37, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Aranda, A.J.; Aguilar-Tipacamú, G.; Perez, D.R.; Bañuelos-Hernandez, B.; Girgis, G.; Hernandez-Velasco, X. ; .. & Petrone-Garcia, V.M. Emergence, migration and spreading of the high pathogenicity avian influenza virus H5NX of the Gs/Gd lineage into America. Journal of General Virology 2025, 106, 002081. [Google Scholar] [CrossRef]
- Suntronwong, N. (2020). Identification of genetic and antigenic variation and evolution pattern among influenza a and b viruses in Thailand. [CrossRef]
- Altan, E.; Avelin, V.; Aaltonen, K.; Korhonen, E.; Laine, L.; Lindh, E. ; .. & Österlund, P. Highly Pathogenic Avian Influenza (HPAI) H5N1 virus in Finland in 2021–2023–Genetic diversity of the viruses and infection kinetics in human dendritic cells. Emerging microbes & infections 2025, 14, 2447618. [Google Scholar] [CrossRef]
- Sutton, T. C. The Pandemic Threat of Emerging H5 and H7 Avian Influenza Viruses. Viruses 2018, 10, 461. [Google Scholar] [CrossRef] [PubMed]
- Charostad, J.; Rukerd, M.R.Z. , Mahmoudvand, S. ; Bashash, D.; Hashemi, S.M.A., Nakhaie, M.; Zandi, K. A comprehensive review of highly pathogenic avian influenza (HPAI) H5N1: an imminent threat at doorstep. Travel medicine and infectious disease 2023, 55, 102638. [Google Scholar] [CrossRef]
- Liu, W.J.; Xiao, H.; Dai, L.; Liu, D.; Chen, J.; Qi, X. ; .. & Liu, Y. Avian influenza A (H7N9) virus: from low pathogenic to highly pathogenic. Frontiers of medicine 2021, 15, 507–527. [Google Scholar] [CrossRef]
- Basu Thakur, P.; Bullock, H.A.; Stevens, J.; Kumar, A.; Maines, T.R.; Belser, J.A. Heterogeneity across mammalian-and avian-origin A (H1N1) influenza viruses influences viral infectivity following incubation with host bacteria from the human respiratory tract. bioRxiv 2025, 2025–03. [Google Scholar] [CrossRef]
- Jallow, M.M.; Diagne, M.M.; Ndione, M.H.D. , Barry, M. A.; Ndiaye, N.K.; Kiori, D.E.;... & Dia, N. Genetic and Molecular Characterization of Avian Influenza A (H9N2) Viruses from Live Bird Markets (LBM) in Senegal. Viruses 2025, 17, 73. [Google Scholar] [CrossRef]
- Wang, Y.; Sun, W.; Wang, Z.; Zhao, M.; Zhang, X.; Kong, Y. ; .. & Gao, Y. Amino acid sites related to the PB2 subunits of IDV affect polymerase activity. Virology Journal 2021, 18, 1–8. [Google Scholar] [CrossRef]
- Bisset, A.T.; Hoyne, G.F. Evolution and adaptation of the avian H7N9 virus into the human host. Microorganisms 2020, 8, 778. [Google Scholar] [CrossRef]
- Zhang, H.; Li, X.; Guo, J.; Li, L.; Chang, C.; Li, Y. ; .. & Sun, B. The PB2 E627K mutation contributes to the high polymerase activity and enhanced replication of H7N9 influenza virus. Journal of General Virology 2014, 95, 779–786. [Google Scholar] [CrossRef]
- Joseph, U.; Su, Y.C.; Vijaykrishna, D.; Smith, G.J. The ecology and adaptive evolution of influenza A interspecies transmission. Influenza and other respiratory viruses 2017, 11, 74–84. [Google Scholar] [CrossRef]
- Xu, H.; Zhu, S.; Govinden, R.; Chenia, H.Y. Multiple vaccines and strategies for pandemic preparedness of avian influenza virus. Viruses 2023, 15, 1694. [Google Scholar] [CrossRef] [PubMed]
- Saenz, H.S.C. , & Liliana, C.A. Preventive, safety and control measures against Avian Influenza A (H5N1) in occupationally exposed groups: A scoping review. One Health 2024, 100766. [CrossRef]
- Lopez, C.E.; Legge, K.L. Influenza A virus vaccination: immunity, protection, and recent advances toward a universal vaccine. Vaccines 2020, 8, 434. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.H.; Erazo, E.M.; Ishcol, M.R.C. , Lin, C. Y.; Assavalapsakul, W.; Thitithanyanont, A.; Wang, S.F. Virus-induced pathogenesis, vaccine development, and diagnosis of novel H7N9 avian influenza A virus in humans: a systemic literature review. Journal of International Medical Research 2020, 48, 0300060519845488. [Google Scholar] [CrossRef]
- Wu, Z.Q.; Zhang, Y.; Zhao, N.; Yu, Z.; Pan, H.; Chan, T.C. ; .. & Liu, S.L. Comparative epidemiology of human fatal infections with novel, high (H5N6 and H5N1) and low (H7N9 and H9N2) pathogenicity avian influenza A virus. International Journal of Environmental Research and Public Health 2017, 14, 263. [Google Scholar] [CrossRef]
- Krammer, F.; Hermann, E.; Rasmussen, A.L. Highly pathogenic avian influenza H5N1: history, current situation, and outlook. Journal of Virology 2025, 99, e02209–24. [Google Scholar] [CrossRef]
- Zhou, Y.; Facciuolo, A.; Aubrey, L.; Barron-Castillo, U.; Berube, N.; Norleen, C. ;... & Warner, B. (2024). Highly pathogenic avian influenza virus H5N1 infection in dairy cows confers protective immunity against reinfection. [CrossRef]
- Karimian, P.E.G.A.H. , & Delavar, M. A. Comparative study of clinical symptoms, laboratory results and imaging features of coronavirus and influenza virus, including similarities and differences of their pathogenesis. Pak. J. Med. Health Sci 2020, 14, 1405–1411. [Google Scholar]
- Yang, Y.; Wong, G.; Yang, L.; Tan, S.; Li, J.; Bai, B. ; .. & Gao, G.F. Comparison between human infections caused by highly and low pathogenic H7N9 avian influenza viruses in Wave Five: Clinical and virological findings. Journal of Infection 2019, 78, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Nakhaie, M.; Rukerd, M.R.Z. , Shahpar, A.; Pardeshenas, M., Khoshnazar, S.M., Khazaeli, M., Eds.; ... & Charostad, J. (2024). A Closer Look at the Avian Influenza Virus H7N9: A Calm before the Storm? Journal of Medical Virology, 96, e70090. [Google Scholar] [CrossRef]
- Garg, S.; Reinhart, K.; Couture, A.; Kniss, K.; Davis, C.T.; Kirby, M.K. ; .. & Olsen, S.J. Highly pathogenic avian influenza A (H5N1) virus infections in humans. New England Journal of Medicine 2025, 392, 843–854. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.; Poorvi, Y.R.G. , Pandey, A.; Paul, E.K., Singh, K., Eds.; Paul, R. Avian Influenza: A Comprehensive Systematic Review of Epidemiology, Clinical Manifestations, Diagnostic Approaches, Prevention Strategies, Recent Outbreaks and Global Collaboration. REDVET-Revista electrónica de Veterinaria, 25(1S), 2024. [Google Scholar]
- Raza, M.A.; Ashraf, M.A.; Amjad, M.N.; Din, G.U.; Shen, B.; Hu, Y. The peculiar characteristics and advancement in diagnostic methodologies of influenza A virus. Frontiers in Microbiology 2025, 15, 1435384. [Google Scholar] [CrossRef]
- Waldock, J.; Remarque, E.J.; Zheng, L.; Ho, S.; Hoschler, K.; Neumann, B. ; .. & FLUCOP consortium. Haemagglutination inhibition and virus microneutralisation serology assays: Use of harmonised protocols and biological standards in seasonal influenza serology testing and their impact on inter-laboratory variation and assay correlation: A FLUCOP collaborative study. Frontiers in Immunology 2023, 14, 1155552. [Google Scholar] [CrossRef]
- Madeley, C.R.; Peiris, J.S.M. Methods in virus diagnosis: immunofluorescence revisited. Journal of clinical virology 2002, 25, 121–134. [Google Scholar] [CrossRef]
- Eisfeld, A.J.; Neumann, G.; Kawaoka, Y. Influenza A virus isolation, culture and identification. Nature protocols 2014, 9, 2663–2681. [Google Scholar] [CrossRef]
- Vemula, S.V.; Zhao, J.; Liu, J.; Wang, X.; Biswas, S.; Hewlett, I. Current approaches for diagnosis of influenza virus infections in humans. Viruses 2016, 8, 96. [Google Scholar] [CrossRef]
- Yang, Y.; Wong, G.; Yang, L.; Tan, S.; Li, J.; Bai, B. ; .. & Gao, G.F. Comparison between human infections caused by highly and low pathogenic H7N9 avian influenza viruses in Wave Five: Clinical and virological findings. Journal of Infection 2019, 78, 241–248. [Google Scholar] [CrossRef]
- Bassetti, M.; Sepulcri, C.; Giacobbe, D.R.; Fusco, L. Treating influenza with neuraminidase inhibitors: an update of the literature. Expert Opinion on Pharmacotherapy 2024, 25, 1163–1174. [Google Scholar] [CrossRef]
- Aldhaeefi, M.; Rungkitwattanakul, D.; Saltani, I.; Muirhead, A.; Ruehman, A.J.; Hawkins, W.A.; Daftary, M.N. Update and narrative review of avian influenza (H5N1) infection in adult patients. Pharmacotherapy: The Journal of Human Pharmacology and Drug Therapy 2024, 44, 870–879. [Google Scholar] [CrossRef] [PubMed]
- Smyk, J.M.; Szydłowska, N.; Szulc, W.; Majewska, A. Evolution of influenza viruses—drug resistance, treatment options, and prospects. International Journal of Molecular Sciences 2022, 23, 12244. [Google Scholar] [CrossRef]
- Wang, J.; Sun, Y.; Liu, S. Emerging antiviral therapies and drugs for the treatment of influenza. Expert Opinion on Emerging Drugs 2022, 27, 389–403. [Google Scholar] [CrossRef]
- Bialy, D.; Shelton, H. Functional neuraminidase inhibitor resistance motifs in avian influenza A (H5Nx) viruses. Antiviral Research 2020, 182, 104886. [Google Scholar] [CrossRef]
- Abbas, G.; Yu, J.; Li, G. Novel and alternative therapeutic strategies for controlling avian viral infectious diseases: Focus on infectious bronchitis and avian influenza. Frontiers in Veterinary Science 2022, 9, 933274. [Google Scholar] [CrossRef]
- Meseko, C.; Sanicas, M.; Asha, K.; Sulaiman, L.; Kumar, B. Antiviral options and therapeutics against influenza: history, latest developments and future prospects. Frontiers in Cellular and Infection Microbiology 2023, 13, 1269344. [Google Scholar] [CrossRef]
- Kovács, L.; Farkas, M.; Dobra, P.F.; Lennon, G.; Könyves, L.P.; Rusvai, M. Avian Influenza Clade 2. 3. 4.4 b: Global Impact and Summary Analysis of Vaccine Trials. Vaccines 2025, 13, 453. [Google Scholar] [CrossRef]
- Astill, J.; Dara, R.A.; Fraser, E.D.; Sharif, S. Detecting and predicting emerging disease in poultry with the implementation of new technologies and big data: A focus on avian influenza virus. Frontiers in veterinary science 2018, 5, 263. [Google Scholar] [CrossRef]
- Pardo-Roa, C.; Nelson, M.I.; Ariyama, N.; Aguayo, C.; Almonacid, L.I.; Munoz, G. ;... & Neira, V. (2023). Cross-species transmission and PB2 mammalian adaptations of highly pathogenic avian influenza A/H5N1 viruses in Chile. bioRxiv. [CrossRef]
- Duan, C.; Li, C.; Ren, R.; Bai, W.; Zhou, L. An overview of avian influenza surveillance strategies and modes. Science in One Health 2023, 2, 100043. [Google Scholar] [CrossRef]
- Petrillo, M.; Buttinger, G.; Corbisier, P.; Leoni, G.; Paracchini, V.; Lambrecht, B. ; .. & Marchini, A. In silico design and preliminary evaluation of RT-PCR assays for A (H5N1) bird flu. Nature 2024, 628, 484–485. [Google Scholar] [CrossRef]
- Huang, S.W.; Wang, S.F. The effects of genetic variation on H7N9 avian influenza virus pathogenicity. Viruses 2020, 12, 1220. [Google Scholar] [CrossRef]
- Chen, X.; Wang, W.; Wang, Y.; Lai, S.; Yang, J.; Cowling, B.J. ; .. & Yu, H. Serological evidence of human infections with highly pathogenic avian influenza A (H5N1) virus: a systematic review and meta-analysis. BMC medicine 2020, 18, 1–16. [Google Scholar] [CrossRef]
- Verhagen, J.H.; Fouchier, R.A.; Lewis, N. Highly pathogenic avian influenza viruses at the wild–domestic bird interface in Europe: future directions for research and surveillance. Viruses 2021, 13, 212. [Google Scholar] [CrossRef]
- Abdelwhab, E.M.; Hafez, H.M. Insight into alternative approaches for control of avian influenza in poultry, with emphasis on highly pathogenic H5N1. Viruses 2012, 4, 3179–3208. [Google Scholar] [CrossRef] [PubMed]
- Marshall, K.E. Personal Protective Equipment Use by Dairy Farmworkers Exposed to Cows Infected with Highly Pathogenic Avian Influenza A (H5N1) Viruses—Colorado, 2024. MMWR. Morbidity and Mortality Weekly Report 2024, 73. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Tian, G.; Shi, J.; Deng, G.; Li, C.; Chen, H. Vaccination of poultry successfully eliminated human infection with H7N9 virus in China. Science China Life Sciences 2018, 61, 1465–1473. [Google Scholar] [CrossRef]
- Focosi, D.; Maggi, F. Avian influenza virus A (H5Nx) and prepandemic candidate vaccines: state of the art. International Journal of Molecular Sciences 2024, 25, 8550. [Google Scholar] [CrossRef]
- Perez-Acle, T.; Ravello, C.; Rosemblatt, M. (2024). Are we cultivating the perfect storm for a human avian influenza pandemic? Biological Research, 57, 96. [CrossRef]
- Morgan, O. (2019). How decision makers can use quantitative approaches to guide outbreak responses. Philosophical Transactions of the Royal Society B, 374, 20180365. [CrossRef]
- Owusu, H.; Sanad, Y.M. Comprehensive Insights into Highly Pathogenic Avian Influenza H5N1 in Dairy Cattle: Transmission Dynamics, Milk-Borne Risks, Public Health Implications, Biosecurity Recommendations, and One Health Strategies for Outbreak Control. Pathogens 2025, 14, 278. [Google Scholar] [CrossRef]
- Dhagat, P.; Coan, J.; Ganguly, A.; Puetz, C.; Silvestri, D.; Madad, S. Enhancing healthcare preparedness: Lessons from a tabletop exercise on Highly Pathogenic Avian Influenza (HPAI). Tropical Medicine and Infectious Disease 2025, 10, 47. [Google Scholar] [CrossRef] [PubMed]
- Musa, E.; Nia, Z.M.; Bragazzi, N.L.; Leung, D.; Lee, N.; Kong, J.D. (2024, October). Avian Influenza: Lessons from Past Outbreaks and an Inventory of Data Sources, Mathematical and AI Models, and Early Warning Systems for Forecasting and Hotspot Detection to Tackle Ongoing Outbreaks. In Healthcare (Vol. 12, No. 19, p. 1959). MDPI. [CrossRef]
- Yan, Q.; Tang, S.; Jin, Z.; Xiao, Y. Identifying risk factors of A (H7N9) outbreak by wavelet analysis and generalized estimating equation. International Journal of Environmental Research and Public Health 2019, 16, 1311. [Google Scholar] [CrossRef] [PubMed]
- Cargnin Faccin, F.; Perez, D.R. Pandemic preparedness through vaccine development for avian influenza viruses. Human Vaccines & Immunotherapeutics 2024, 20, 2347019. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Fan, M. Reaction–advection–diffusion model of highly pathogenic avian influenza with behavior of migratory wild birds. Journal of Mathematical Biology 2025, 90, 1–45. [Google Scholar] [CrossRef]
- Mubareka, S.; Amuasi, J.; Banerjee, A.; Carabin, H.; Copper Jack, J.; Jardine, C. ; .. & Jane Parmley, E. Strengthening a One Health approach to emerging zoonoses. Facets 2023, 8, 1–64. [Google Scholar]
- Sreenivasan, C.C.; Li, F.; Wang, D. Emerging Threats of Highly Pathogenic Avian Influenza A (H5N1) in US Dairy Cattle: Understanding Cross-Species Transmission Dynamics in Mammalian Hosts. Viruses 2024, 16, 1703. [Google Scholar] [CrossRef]
- Dhagat, P.; Coan, J.; Ganguly, A.; Puetz, C.; Silvestri, D.; Madad, S. Enhancing healthcare preparedness: Lessons from a tabletop exercise on Highly Pathogenic Avian Influenza (HPAI). Tropical Medicine and Infectious Disease 2025, 10, 47. [Google Scholar] [CrossRef]

| Virus Subtype | Year(s) | Region/Country | Major Events & Notes |
|---|---|---|---|
| H5N1 | 1997 | Hong Kong (SAR) | First human outbreak (Clade 0) |
| 2003 onwards | SE Asia (Vietnam, Laos, Thailand), China | Widespread outbreaks (Clade 1) | |
| 2005–2006 | Asia, Europe, Africa | Intercontinental spread (Clade 2.2) | |
| 2014–2015 | China, Korea, USA, Europe | H5N8 spread (Clade 2.3.4.4a) | |
| 2016–2017 | Global | H5Ny emergence (Clade 2.3.4.4b) | |
| 2020–2021 | Europe, Asia, Africa | Novel H5N1 clade 2.3.4.4b wave | |
| Late 2021–ongoing | North America | Mass poultry/mammal outbreaks | |
| Late 2022–ongoing | Mexico, Central & South America | Spread of clade 2.3.4.4b | |
| 2023 | Europe (Poland, Spain, Finland) | Infections in cats, mammals | |
| 2024–ongoing | USA (Dairy cattle), Global | Dairy cattle infection, continued wild bird circulation | |
| H7N9 | 2013–2017 | China | Five epidemic waves, LPM interventions successful |
| 2003 | Netherlands | H7N7 outbreak (human infections) | |
| 2019 | Cambodia | Emergence of H7N4 | |
| H9N2 | 1998–ongoing | China, Egypt, Pakistan, Oman | Endemic poultry infection; mild human disease |
| Virus Subtype | Year Range | Total Cases | Fatalities | Case Fatality Rate (%) | Notes |
|---|---|---|---|---|---|
| H5N1 | 1997 | 18 | 6 | 33.3% | First human cases (Hong Kong) |
| 2003 | 3 | 2 | 66.7% | Re-emergence in Asia | |
| 2004 | 46 | 32 | 69.6% | Vietnam, Thailand, others | |
| 2003–July 14, 2023 | 878 | 458 | 52.16% | WHO cumulative reports | |
| March–October 2024 (USA) | 46 | Not specified | Not specified | Exposure to cows and poultry | |
| Since 1997–December 11, 2024 | 974 | 464 | 47.6%* | WHO cumulative reports | |
| Since 1997–March 7, 2025 | 989 | 467 | 47.2%* | Updated data | |
| H7N9 | March–May 2013 (China) | 130 | ≥27 | 20% | First wave |
| Since 2013 (China, five waves) | 1567 | 615 | 39.2% | Five epidemic waves | |
| H9N2 | 1998–2019 | 59 | 1 | 1.7% | Mild infections |
*Notes: *Includes USA dairy farm-associated cases. Deaths for H5N1 recorded since 2003. H9N2 infections typically mild.
| Viral Protein/Gene | H5N1 Mutation(s) | H7N9 Mutation(s) | Functional Impact |
| Genome | Segmented, negative single-stranded RNA, 8 segments. Encodes at least 11 proteins | Segmented, negative single-stranded RNA, 8 segments. Encodes at least 11 proteins. | Basic structure of Influenza A viruses. HA and NA determine subtype. High mutation rate in HA and NA. |
| Hemagglutinin (HA) | L129V, S589, S582, S512, S541, L515, L52, L569, L55 (amino acid changes in human isolates, frequencies vary by region). HA gene conservation high (98.3-99.9% similarity in Finnish isolates compared to European). Mutations related to human receptor specificity. HA T108I, S123P, S133A, K218Q, S223R, 327-328del/328del in US/Cambodian 2024/2025 cases [45a, 45b, 45c]. | Acquisition of polybasic amino acids at cleavage site (e.g., PKRKRTA(R/G), PKGKRTA(R/G), PKGKRIA(R/G)). Mutations related to avian-to-human receptor-binding adaptation. | HA binds to host cell receptors. HA cleavage site determines pathogenicity (LP vs HP). Mutations can affect receptor binding and stability. Key determinant of host range. |
| Neuraminidase (NA) | NA H155Y, T365I, N366S found in Finnish isolates. | [Specific mutations listed in sources not available in excerpts]. | NA cleaves sialic acid receptors (facilitates virus release - information not explicitly from sources, but common knowledge about NA function). Amino acid substitutions can affect inhibition by neuraminidase inhibitors. |
| Polymerase basic 2 (PB2) | PB2 K389R, V598T, E627K, G309D found in Finnish isolates. E627K mutation detected in clade 2.3.4.4b H5N1 associated with enhanced mammalian replication (mentioned in conclusion thought process). | Amino acid substitutions can contribute to pathogenicity in mammalian hosts. | Part of the polymerase complex (PA, PB1, PB2) that orchestrates transcription and replication. PB2 E627K is a known adaptation to mammalian hosts. |
| Nucleoprotein (NP) | NP V33I, Y52N, I109V found in Finnish isolates. | [Specific mutations listed in sources not available in excerpts]. | Part of the viral ribonucleoprotein (vRNP), involved in genome encapsidation. |
| Matrix protein 1 (M1) | M1 N30D found in Finnish isolates. | [Specific mutations listed in sources not available in excerpts]. | Found inside the lipid envelope. Involved in viral lifecycle. |
| Nonstructural protein 1 (NS1) | NS1 P3S, R41K, K55E, D70G found in Finnish isolates. | [Specific mutations listed in sources not available in excerpts]. | Non-structural/regulatory protein. Involved in viral lifecycle. |
| Feature | H5N1 | H7N9 | Notes |
|---|---|---|---|
| Severity | Severe; high mortality | Severe; emerged high pathogenicity in wave 5 | Both associated with ARDS, pneumonia |
| Common Symptoms | Respiratory and gastrointestinal symptoms, elevated aminotransferase | Respiratory symptoms; some GI involvement | Symptoms overlap but vary |
| Age Distribution | Median 18 years (USA 2024: adults) | Median 63 years | H9N2 mainly affects young children |
| Sex Ratio (M:F) | 1:1.19 | 1:0.45 (male predominance) | Differences in susceptibility noted |
| Coexisting Conditions | Rare | Common (elderly, hypertension, diabetes) | Underlying conditions worsen outcomes in H7N9 |
| Transmission | Primarily animal-to-human | Primarily animal-to-human (live bird markets) | Human-to-human transmission rare |
| Antiviral Sensitivity | Sensitive to NA inhibitors | Sensitive to NA inhibitors | Treatment feasible with neuraminidase inhibitors |
| Complications | CNS involvement, multi-organ failure | Nosocomial infections, elevated Angiotensin II | Severe systemic complications in both |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



