Submitted:
25 July 2025
Posted:
29 July 2025
You are already at the latest version
Abstract
The extensive utilization of chemicals in agriculture over the years, particularly those derived from chemical fertilization, has resulted in the proliferation of both natural and human-induced problems. Consequently, there is an urgent need to explore alternative, viable, and environmentally sustainable solutions. In this regard, microalgae and cyanobacteria, as photosynthetic microorganisms, have been postulated as feasible candidates for this purpose. This is primarily due to their ability to capture CO2 from the environment, as well as micro- and macro-elements. Additionally, their extensive metabolic plasticity enables them to produce a multitude of high-added-value compounds, including biostimulant molecules, biocontrol molecules, and biofertilizer capacity due to their nutrient content. Furthermore, it has been demonstrated that certain species possess the capacity to fix atmospheric nitrogen, a property that confers upon them significant agronomic relevance. This review aims to summarize the potential of these microorganisms and their application in this sector, focusing primarily on their biofertilization, biostimulation, and biocontrol capabilities and presents a compilation of the products currently available on the market that are derived from these microorganisms. The present work also identifies the gaps in the use of these microorganisms and provides future prospects for developing a suitable solution for today's agriculture.
Keywords:
microalgae
; cyanobacteria
; biofertilizer
; biostimulant
; biocontrol
; sustainable agriculture
; circular bioeconomy
; plant growth‐promoting microorganisms
1. Introduction
The global population is increasing rapidly worldwide, highlighting an urgent need for sustainable solutions to ensure a suitable food supply [1]. According to the Food and Agriculture Organization (FAO), global food production must increase by 60% by 2050. Central to this strategy is prioritizing the expansion of agriculture as a primary response, aligned with a circular economy plan [2]. However, the overuse of land and the demands to enhance productivity in agricultural fields have resulted in irregular activities, including the misuse of water in irrigation and the excessive application of chemical treatments for fertilization and pesticides [3]. As a consequence of these human activities, environmental conditions have been altered, leading to a decrease in soil health and ultimately impacting crop yield and productivity [4]. Additionally, these practices have contributed to increased greenhouse gas emissions and reduced biodiversity, reinforcing the urgency to adopt more regenerative and low-impact agricultural methods [5].
Soil health is a critical factor in sustainable agriculture, as healthy soil provides the foundation for robust crop growth and long-term productivity. It encompasses the physical, chemical, and biological properties of soil that influence its ability to support plant life. Practices such as crop rotation, reduced tillage, organic fertilization, and cover cropping help maintain and improve soil health by enhancing soil structure, increasing nutrient availability, and fostering beneficial microbial activity [6]. Healthy soil not only boosts crop yields but also contributes to the resilience of agricultural systems against pests, diseases, and climate change impacts [7]. Prioritizing soil health is essential for ensuring food security, conserving biodiversity, and promoting environmentally sustainable farming practices.
Microalgae, cyanobacteria, bacteria and fungi are beneficial microorganisms that form essential microbial communities, contributing to soil stability and resilience. They are gaining attention nowadays as sustainable agriculture inputs due to their rich composition and growth promoting effects. Their presence significantly enhances plant growth across diverse conditions [8]. Within this group, microalgae and cyanobacteria are unicellular photosynthetic organisms that inhabit soil, freshwater and marine ecosystems and represent one of the most ancient and metabolically diverse groups of microorganisms on Earth. With an extraordinary capacity to convert carbon dioxide, sunlight, and inorganic nutrients into a wide array of valuable biomolecules [9]. Thus, microalgae and cyanobacteria have been extensively studied for applications ranging from biofuels to pharmaceuticals [10,11]. Their high photosynthetic efficiency, fast growth rates, and ability to thrive in non-arable land and wastewater environments distinguish them from terrestrial crops and make them ideal candidates for circular bioeconomy models [12].
In recent years, microalgae and cyanobacteria have attracted growing interest in the agricultural sector due to their multifunctional roles in enhancing plant productivity and soil health. The global agricultural biologicals market is projected to reach $14.6 billion by 2025, with algal and microbial biostimulants among the fastest-growing research areas [13]. Additionally, these microorganisms can influence plant development through direct and indirect mechanisms, functioning as biofertilizers, biostimulants or biocontrol, among other roles [9]. These include nutrient enrichment (nitrogen, phosphorus, potassium, and trace elements), production of plant-growth-promoting substances (such as auxins, gibberellins, and cytokinins), improvement of soil microbial communities, and increased tolerance to biotic and abiotic stresses [14]. Their biochemical versatility allows customization depending on the crop needs and environmental conditions. Recent studies further support these effects, showing that the biostimulant activity of microalgae under stress conditions (especially drought and salinity) is linked to improved photosynthetic performance, antioxidant responses, and accumulation of osmo-protectants [15]. Similarly, Brito-Lopez et al., 2025 report the enhanced plant growth and stress resilience across a range of environmental conditions, reinforcing their potential as climate-smart agricultural inputs.
A central challenge in modern agriculture is overreliance on synthetic nitrogen-based fertilizers and chemical NPK inputs. Although these compounds have undeniably contributed to the green revolution and global food security, their continued use has led to serious environmental and health consequences. These include nitrate leaching, eutrophication of aquatic systems, emission of nitrous oxide (a potent greenhouse gas), and deterioration of soil biodiversity [16]. As a result, the search for sustainable, environmentally compatible alternatives has become a global imperative. Photosynthetic organisms such as microalgae and cyanobacteria represent a highly promising, sustainable alternative to conventional fertilizers and agrochemicals [9]. In particular, the exploration of nitrogen-fixing cyanobacteria offers a compelling solution. By mitigating the adverse environmental impacts associated with synthetic fertilizers, the utilization of cyanobacteria for nitrogen fixation holds the potential to address the drawbacks of conventional nitrogen fertilization practices [17]. Integrating these organisms into farming practices can help address critical environmental concerns while enhancing soil fertility, crop resilience, and overall agroecosystem health. As the world confronts the dual challenges of climate change and food security, leveraging the full potential of microalgae in agriculture offers a strategic pathway toward more sustainable and resilient food systems.
Building on this promise, microalgae and cyanobacteria offer specific practical solutions that further underscore their value in sustainable agriculture. Not only can they fix atmospheric carbon and accumulate nitrogen and phosphorus from various sources (including wastewater), but their application to soil can also release nutrients in bioavailable forms, reducing the risk of leaching and volatilization [18]. Furthermore, microalgal biomass can be integrated into regenerative agricultural systems, supporting nutrient recycling, organic matter enrichment, and improved soil structure [19]. Certain species such as Chlorella, Scenedesmus, and Spirulina have already shown considerable promise as biofertilizers in field trials, with positive impacts on yield, root morphology, and chlorophyll content in a variety of crops including wheat, maize, tomatoes, and rice. These benefits are further supported by recent findings showing significant reductions in synthetic fertilizer input without compromising crop yield [20].
From a sustainability perspective, cultivating microalgae for agricultural applications supports circular economy and zero-waste approaches strategies [21]. Algal biomass can be produced using agricultural runoff, animal slurry, or even urban wastewater, thereby reducing waste streams and recovering nutrients that might otherwise cause pollution. Additionally, in comparison to synthetic fertilizers, microalgal products can be produced with lower energy inputs and have significantly reduced carbon footprints when incorporated into biorefinery systems [22]. Despite these advantages, significant challenges remain for the large-scale implementation of microorganisms in agriculture, such as economic feasibility, the scalability of production systems, and the need for standardized protocols and regulatory frameworks. However, ongoing advances in biotechnology, strain selection, photobioreactor design, and downstream processing are steadily improving the viability of microalgae-based agricultural products.
In summary, microalgae and cyanobacteria are emerging as powerful, nature-based solutions for advancing sustainable agriculture. These photosynthetic microorganisms possess the ability to produce a wide range of bioactive compounds that promote plant growth, enhance nutrient use efficiency, and contribute to soil health through nutrient cycling. Their multifunctional roles as biofertilizers, biostimulants, and biocontrol agents make them attractive candidates for reducing dependency on synthetic agrochemicals, offering a path toward reduced environmental impact and improved agricultural resilience. Nevertheless, several challenges remain, particularly regarding cost-effective large-scale production and formulation. This review aims to provide a comprehensive overview of their current applications, market presence, and technological constraints, while also identifying research gaps and future directions necessary to fully realize their potential in environmentally and economically sustainable agriculture. An overview of the present revision article is illustrated in Figure 1.
2. The Use of Microalgae and Cyanobacteria as Biostimulants
Plant biostimulants are defined as substances or microorganisms that, when applied in small amounts, enhance plant nutrition processes, stress tolerance, growth, or quality, independently of direct nutrient supply [23]. Algae are recognized as some of the earliest biostimulants used in agriculture, with historical records dating back to the Roman era, where they were applied as manure to enrich soil fertility and promote plant growth, an application that has continued through the centuries. Thanks to their remarkable diversity and metabolic flexibility, microalgae serve as a rich source of high-value metabolites, including proteins, amino acids, enzymes, pigments, polyunsaturated fatty acids, polysaccharides, vitamins, antioxidants, and phytohormones [19,24]. This biochemical richness explains why microalgae (including cyanobacteria; commonly referred to as “blue-green algae”) are gaining increasing attention as sustainable inputs in agriculture [25]. Their demonstrated growth-promoting properties and environmentally friendly profile have contributed to their rising use, both as biostimulants and biofertilizers, particularly among growers seeking alternatives to synthetic agrochemicals. In the following section key factors in the development of microalgae- and cyanobacteria-based biostimulant products will be explored.
Although microalgae hold great promise for agricultural applications, selecting the most effective and suitable strains remains a complex task. Among the key criteria for strain selection, two are particularly fundamental in determining whether a strain is viable for further development. The first is the strain’s ability to grow rapidly and uniformly while maintaining high productivity (ideally in nutrient rich media or in an ideal case using waste-derived resources) to ensure the production of sufficient biomass. In this way, the strains must be easily cultivated to generate sufficient biomass [26,27]. Species such as Chlorella vulgaris and Scenedesmus spp. are well known for their fast growth rates and short doubling time [28]. The second essential criterion is the strain’s physiological and biochemical potential, particularly its capacity to produce bioactive compounds such as exopolysaccharides, amino acids, proteins, vitamins, and phytohormone-like substances (e.g., auxins and cytokinins). In practice, a decision-making matrix for strain selection should integrate both production performance and biochemical profiles. Based on these parameters, cyanobacteria and green algae currently represent the most widely used groups in biostimulant applications and will be further discussed. In contrast, the application of diatoms in this context is still in its early stages of development.
Cyanobacteria are gaining attention in agriculture thanks to their ability to fix atmospheric nitrogen, solubilize phosphorus, and produce a wide variety of beneficial compounds (like phytohormones) that support plant health. Their unique physiological traits and metabolic flexibility have positioned them as promising biostimulants in crop production systems [29]. Certain genera, including Anabaenopsis, Calothrix, and Anabaena, have demonstrated the capacity to enhance seed germination and promote plant development through the synthesis of phytohormones like cytokinins, gibberellins, and auxins [30,31]. Beyond promoting plant growth, cyanobacteria also help plants cope with stress. For example, they produce extracellular polysaccharides, antioxidants, and signaling molecules that help mitigate the damaging effects of salinity [32]. Under drought conditions, their benefits extend further: cyanobacteria can regulate ion export, modulate the surrounding microbial community, and even improve germination rates under limited water availability. These stress-alleviating traits have been documented across a wide range of crops, as reviewed by Sánchez-Quintero et al., 2023 [33]. Beyond their direct effects on plants, cyanobacteria also play a crucial role in soil health. In addition to enhancing nitrogen fixation and phosphorus availability (discussed in the next section), they foster symbiotic relationships with other soil microorganisms, reinforcing their value in sustainable and regenerative agriculture [2,34]. Unlike seaweed, which are typically collected from the wild, cyanobacteria can be cultivated in controlled environments like open ponds or photobioreactors. This allows for more consistent quality and optimized conditions for producing bioactive compounds [29]. Taken together, the contributions of cyanobacteria to sustainable agriculture are multifaceted. They enhance soil structure by secreting polysaccharides, improve fertility through nitrogen fixation, increasing porosity and water retention, and release growth-promoting substances. Moreover, they aid in reducing salinity, making phosphorus more bioavailable, and facilitating the recycling of agricultural residues. Whether applied as live cell suspensions or processed extracts, cyanobacteria consistently demonstrate significant potential to enhance plant vitality, productivity, and resilience.
Green microalgae have demonstrated a wide range of beneficial effects on crop growth and are increasingly recognized as promising inputs for sustainable agriculture. Microalgae-based products can enhance plant nutrition, improve overall crop performance, support key physiological processes, and increase tolerance to abiotic stresses such as drought and salinity [35]. Among the various genera studied, Chlorella stands out as the most used in agricultural applications [36,37]. Numerous trials using both fresh biomass and extracts from Chlorella strains have reported notable improvements in plant development and yield [38,39]. For instance, treatments with Chlorella fusca have been shown to enhance the growth of Chinese chives under field conditions [40]. In another example, soil-drenching with live Chlorella cells significantly increased biomass production in Medicago truncatula [39]. Complementary findings have been reported for other Chlorella species as well. C. vulgaris and C. pyrenoidosa, for example, have proven effective as biofertilizers in salt-affected soils, supporting the growth of crops such as lettuce, rice, eggplant, and cucumber under saline stress [41]. Likewise, applications of Spirulina platensis have demonstrated growth-promoting effects on leafy vegetables like rocket, red bayam, and pak choi [42], further reinforcing the versatility of microalgae across diverse crop types. A key mechanism by which microalgae enhance plant performance is through the production and secretion of phytohormones, including auxins and cytokinins. In addition, they release exopolysaccharides that contribute to drought and salinity tolerance by improving soil water retention and nutrient availability. These compounds also supply organic carbon to beneficial soil microbes, fostering microbial activity and nutrient cycling. Expanding the range of beneficial species, inoculation with filamentous cyanobacteria such as Calothrix elenkinii has been found to stimulate both the phyllosphere and rhizosphere microbiomes, further supporting plant health and productivity [43]. While research continues to identify and characterize the most effective microalgae strains, current evidence clearly underscores the potential of green microalgae as valuable biostimulants. Their capacity to enhance plant growth, improve resilience to environmental stresses, and promote beneficial soil microbial communities makes them a sustainable and impactful tool for advancing modern agriculture.
Diatoms are an incredibly diverse group of microorganisms, with around 100,000 known species. They are easy and cost-effective to cultivate, which makes them an attractive source of bioactive compounds, many of which are already used in the pharmaceutical industry [44]. Although their application as biostimulants in agriculture is still limited, their unique structural and biochemical characteristics offer great potential. Studies have shown that diatoms, particularly Navicula species, can positively influence plant growth in species such as Salix viminalis, Helianthus tuberosus, and Sida hermaphrodita [45]. One of the most distinctive features of diatoms is their silica-based cell wall and the secretion of mucilage rich in sugars like mannose, fucose, and galactose. These properties can be utilized in the development of diatom-based fertilizers, which may enhance plant biomass and crop yields. Diatoms also possess phytoremediation abilities, as they can absorb heavy metals from the environment (including lead, zinc, nickel, cadmium, and titanium) [46]. While their nitrogen and phosphorus content environment, compared to synthetic fertilizers, the nutrients they provide are often more readily absorbed by plants. As part of the next generation of sustainable fertilizers, diatoms could help improve plant growth and resilience, especially under stress conditions such as extreme climate events [47].
Finally, the use of microalgae–microorganism consortia has emerged as a promising alternative to the traditional application of single microalgal or microbial strains for promoting plant growth. These synergistic partnerships offer significant advantages by providing plants with a broader spectrum of essential nutrients (such as nitrogen, phosphorus, and potassium) through complementary metabolic interactions [48] Thus, when microalgae are combined with nitrogen-fixing bacteria, the consortium can perform complex biological functions that individual strains cannot achieve alone [49]. This cooperative behavior not only enhances nutrient availability but also supports processes with valuable biotechnological applications [50]. Furthermore, the combined activity of microalgae and beneficial microbes can activate plant defense mechanisms, leading to the production of fungal enzymes and antibiotics that protect plants from pests and diseases [51]. Beyond agricultural benefits, microalgae–bacteria consortia, especially those involving nitrogen-fixing species, also hold great promise in the fields of biotechnology [50] .
3. Biofertilizers
A biofertilizer is a substance containing specific living microorganisms, known as plant growth-promoting rhizobacteria (PGPR). These microorganisms improve plant growth by colonizing the rhizosphere or interior of plants. This process enhances the supply or availability of nutrients to the host plant. The product is administered to seeds, plants or soil [9,52,53]. In addition, these eco-friendly approaches have been shown to improve crop productivity, nutrient profile and plant tolerance to abiotic and biotic stress. The use of biofertilizers reduces the problems associated with chemical fertilizers and leads to more sustainable agriculture [54].
The utilization and marketing of biofertilizers commenced in the 18th century with the patenting of 'Nitragin', the first Rhizobium formulation, by Hiltner and Nobbe (1896_Nitragin_Nobbe and Hiltner) classification of biofertilizers is based on their function and mechanism of action. The most widely employed biofertilizers comprise nitrogen-fixers (N-fixers), potassium solubilizers (K solubilizers), phosphorus solubilizers (P solubilizers), zinc solubilizers (Zn solubilizers), iron solubilizers (Fe solubilizers) and phytostimulators [56,57,58].
Beyond the well-known categories of biofertilizers, microalgae and cyanobacteria-based biofertilizers have recently attracted increasing interest. These photosynthetic microorganisms quickly establish themselves as valuable alternatives or supplements to traditional biofertilizers because of their ability to perform multiple functions—such as fixing nitrogen, producing siderophores, synthesizing plant hormones, and solubilizing minerals. Their multifunctional nature, combined with their ability to thrive in a wide range of environmental conditions, makes them a promising tool for promoting sustainable crop production.
3.1. Nitrogen-Fixing Biofertilizers
Nitrogen-fixing biofertilizers are a subject of significant research interest for leguminous plants, with studies focusing on their ability to capture atmospheric nitrogen (N2) from the soil through the action of specific microorganisms. Rhizobial inoculants represent the most prevalent biofertilizers in the global market, with the capacity to curtail annual nitrogen fertilization expenditures [57,59]. Other microorganisms for which the ability to fix nitrogen has been demonstrated comprise the following: Azospirillum spp., Azoarcus spp., Burkholderia spp., Gluconacetobacter diazotrophicus, Herbaspirillum spp., Azotobacter, and Paenibacillus polymyxa [60]. This is evidenced by the successful formulation of efficient N-fixing strains in commercial biofertilizers, such as Rhizobium and Azotobacter spp. [61]. More recently, the use of a biofertilizer derived from chicken feathers and constituted by a consortium of keratinolytic bacteria identified as Bacillus licheniformis MW45 and Bacillus paralicheniformis MW48 has been shown to enhance the growth of spinach for its effectiveness as a nitrogen fertilizer [62]. Furthermore, additional N-fixing microorganisms employed as biofertilizers include different species of microalgae and cyanobacteria [63,64] as will be described in more detail in subsection 3.6.
3.2. Phosphorus and Potassium Solubilizing Biofertilizers
The prevalence of phosphorus (P) deficiency in numerous agricultural soils, in conjunction with the restricted availability of economically viable P deposits, has precipitated an augmented interest in P-solubilizing biofertilizers (PSB) [65]. In addition to enhancing the availability of P in the crop soil, the utilization of PSB has been linked to the biocontrol of crop pests [66]. The utilization of PSB has been extensively employed in various crop cultivation to enhance the availability of phosphorus in the soil and promote increased plant growth. Examples of this include the application of Pseudomonas aeruginosa in rice, Pantoea agglomerans in maize, Pseudomonas sp. in chili pepper, Enterobacter in soybeans, Aspergillus niger in beans, Burkholderia cepacian in peanuts and Azospirillum in wheat [67,68,69,70,71,72,73].
In parallel to P, it has been determined that agricultural soils are also deficient in K, thus giving similarity of relevance to potassium solubilizing biofertilizers (KSB) and PSB. A plethora of research has been carried out which demonstrates that the utilization of KSB in diverse crop cultivation (for instance maize and wheat) leads to a significant improvement in crop growth and productivity through substantial enhancement of germination, nutrient uptake, crop growth and yield [74,75,76,77]. In comparison with chemical fertilizers, the solubilization of potassium by KPS does not fully meet the K requirements of plants. Nevertheless, its utilization is proposed as an environmentally friendly alternative means of reducing the amount of chemical fertilizers [78].
3.3. Zinc Solubilizing Biofertilizers
Intensive agriculture has been demonstrated to reduce the level of soil nutrients, including Zn. The employment of chemical fertilizer is a costly endeavour, and the formulation of such fertilizers is frequently not immediately assimilated by plants [79]. Zinc solubilizing biofertilizers have become increasingly important in the context of crop production. For instance, the process of zinc solubilization by certain bacterial species, including Azospirillum, Azotobacter, Pseudomonas, and Rhizobium, has been demonstrated to enhance zinc assimilation in wheat [80]. Furthermore, three selected different bacteria, Acinetobacter calcoaceticus, Bacillus proteolyticus and Stenotrophomonas pavanii, formulated in both free and encapsulated forms showed improved plant growth parameters and enhanced zinc content in Zea mays and can be applied as biofertilizers to enhance soil fertility [81]. According to [82]. The application of Zn-solubilizing PGP bacteria (Pseudomonas spp.), isolated from cow dung to tomato seedlings leads for improving plant growth and production. Furthermore, this treatment has been found to enhance zinc levels in Solanum lycopersicum. In a recent research study, a Zn-biofertilizer comprising a consortium of Zn-solubilizing strains of Streptomyces sp., Pseudomonas sp., and zinc oxide nanoparticles as a source of Zn was administered to wheat plants. This treatment resulted in a substantial improvement in plant vegetative growth and grain yield. Furthermore, it was observed that this biofertilizer could enhance the Zn status of wheat grains, consequently increasing blood Zn content in rats [80].
3.4. Iron Solubilizing Biofertilizers
Iron is involved in a variety of metabolic pathways within the cell, including photosynthesis, thus rendering it an essential element for plant life [83]. It is noteworthy, however, that siderophores represent a distinct group of low molecular weight compounds (less than 1.5 kDa) that exhibit a high affinity for Fe in environments with low Fe concentrations [34]. The synthesis and secretion of these compounds by different bacterial strains occurs under specific conditions, thereby increasing and regulating the bioavailability of Fe. Consequently, siderophore-producing bacteria have attracted considerable scientific interest because of their potential application as biofertilizers. These bacteria have been demonstrated to enhance soil fertility and increase plant biomass, a finding that is of significant relevance for the development of sustainable agricultural practices [84,85]. Furthermore, the promotional effect of the AS19 strain, a bacterium capable of producing high levels of siderophores and facilitating the absorption of Fe3 by seeds and plants, on the germination of pepper and maize seeds, and the development of shoots and leaves of Gynura divaricata (Linn.) has been demonstrated. AS19 was isolated from the rhizosphere of Paris polyphylla var. yunnanensis, an important traditional Chinese herb [34]. A recent study has demonstrated the significant potential of Fe-solubilizing rhizobacteria (Bacillus spp.) isolated from the maize rhizosphere in calcareous soils as effective biofertilizers. These bacterial strains, namely Bacillus pyramidoids, Bacillus firmicutes, and Bacillus cereus, have been shown to significantly enhance maize growth and physiological attributes under axenic conditions. This enhancement is likely attributable to improved Fe availability and uptake. The findings outlined in this study provide a foundation for the development of biofertilizers that are both eco-friendly and cost-effective. These biofertilizers have the potential to mitigate Fe deficiency in crops, thereby promoting sustainable agriculture practices [86]. Moreover, a further study investigates the capability of siderophore-producing bacterial isolates from the rhizosphere of Pinus pinea, specifically two Pseudomonas strains, Z8.8 and Z10.4, to reverse chlorosis in Fe-starved tomato plants. The delivery of each strain to 4-week-old iron-starved tomatoes resulted in the reversion of chlorosis, which was consistent with enhanced Fe contents up to 40% [87].
Furthermore, microalgae have been demonstrated to play a pivotal role in maintaining ion homeostasis. Dunaliella, a genus of algae, has demonstrated a remarkable capacity for adaptation to environments characterized by low iron levels. Several species of Dunaliella, namely Dunaliella tertiolecta, Dunaliella salina and Dunaliella bardawil, have been identified as originating from radically divergent environments. These species have been found to possess a unique family of siderophore-iron-uptake proteins [88]. In relation to the Cyanobacteria spp, Brick et al., in 2025 highlighted the significant potential of Synechococcus mundulus-derived siderophore in stimulating Zea mays physicochemical growth parameters and iron uptake.
3.5. Phytostimulators
Beyond their direct effects on plant physiology and biostimulation (as have been described in Section 2), phytohormones also play indirect but crucial roles in soil nutrient dynamics. Certain hormones—particularly auxins, cytokinins, and ethylene—modulate root system architecture, increasing root surface area, branching, and exudation. These changes enhance the plant’s capacity to explore the rhizosphere and mobilize nutrients. In parallel, hormone-induced root exudates can stimulate beneficial soil microbiota, including nitrogen-fixing and phosphate-solubilizing microorganisms, creating a feedback loop that improves soil fertility. Additionally, phytohormones can upregulate the expression of nutrient transporter genes in roots, further facilitating the uptake of macro- and micronutrients [90]. Recent studies have shed light on brassinosteroid role in modulating agronomic traits that directly contribute to grain yield in rice (Oryza sativa) [91]. In this context, biofertilizers that can promote plant growth through phytohormone production are known as phytostimulators. Phytostimulators produced by bacteria, microalgae, cyanobacteria and other microorganisms contribute not only to plant growth promotion but also to improved nutrient acquisition through changes in both plant physiology and rhizosphere interactions, positioning them as multifunctional agents in sustainable agriculture. How cyanobacteria and microalgae contribute to this framework will be described in next subsection 3.6.
3.6. Microalgae and Cyanobacteria-Based Biofertilizers
Nostocales cyanobacteria are a large group of photoautotrophic prokaryotes with the ability to fix atmospheric nitrogen to ammonia, a source of nitrogen that can be bio-assimilated by plants [92]. Under nitrogen-starvation conditions, Nostoc cyanobacteria develop heterocysts, specialized cells where nitrogen fixation occurs, which are arranged in an orderly manner along the vegetative cell chain [93].
In addition, these prokaryotic microorganisms improve soil structure by retaining nutrients and water due to their plant growth-promoting traits, such as the ability to form biofilms [94]. Besides, they can increase soil porosity and decrease soil salinity [6]. Nostoc species possess the ability to produce phytohormones (auxins, cytochromes, gibberellins and ethylene), siderophores (iron binders) and mineral solubilizers (e.g., phosphorus, potassium and zinc) [95,96]. These symbiotic cyanobacteria have a wide diversity of associations with plants distributed throughout the plant kingdom such as spore-forming bryophytes, ferns, cycads or rice [97,98,99]. The development of cyanobacteria-based biofertilizer (CBF) has been enabled by the potential characteristics of these prokaryotic photoautotrophic microbes [100,101]. It is worth noting that using cyanobacteria as biofertilizers is not a new concept in agriculture. These organisms have already been studied extensively in various crop systems. For instance, notable advancements have been made in rice cultivation over the past few years, such as in Andalusian paddy fields. Re-inoculation of cyanobacterial isolates from this area as CBF has had a positive, significant effect on plant growth, with a significant increase in plant length of 127% recorded, as well as significant increases in grain weight and number per panicle. Taken together, these results suggest that cyanobacterial biofertilizers could be a promising way to sustain rice production [101]. Additionally, a recent study investigated the potential of two cyanobacteria, Anabaena vaginicola ISB42 and Nostoc spongiaeforme var. tenue ISB65, as promising candidates for producing environmentally friendly biofertilizers for sustainable peppermint cultivation because they improve the quantity and quality of essential oils (EOs) by upregulating the key genes involved in the menthol biosynthetic pathway in Mentha piperita [102]. Furthermore, their potential as biofertilizers for cotton crops has recently been evaluated, demonstrating that reintegrating these beneficial species into agricultural ecosystems can enhance crop growth and maintain a balanced microbial environment. This contributes to the broader goal of achieving sustainable agriculture on a global scale [103].
Special mention should be given to Arthrospira platensis, an edible cyanobacterium known worldwide for its high nutritional value, as well as for the interest in its biological activity and bioactive compounds [104]. Supplementing Arthrospira platensis by drenching the soil resulted in increased growth and productivity of chia plants cultivated under alkaline soil conditions, as well as increased antioxidant levels in the chia seeds. Following microalgae application, the oil content increased, as did the proportion of omega-3 [105]. More recently, according to [106], applying a biofertilizer containing Spirulina maxima, marine Lactobacillus plantarum, molasses, and industrial organic waste (IOW) at a concentration of 0.1% can enhance the growth, development, and nutrient uptake of rosemary plants by generating bioactive compounds, including vitamins, carbohydrates, and phytohormones (auxins, gibberellins, and cytokinins).
Among eukaryotic microalgae, Chlorella vulgaris and Scenedesmus obliquus can stimulate root growth through auxin-like activity while contributing to the mobilization or improved uptake of phosphorus and potassium, leading to better growth under nutrient-limited or stress environments [53,107]. Although Chlamydomonas reinhardtii is not a strong nutrient-mobilizing species on its own, it can participate in associative biofertilization when co-applied with diazotrophic or nutrient-solubilizing bacteria; in such consortia, algal-derived metabolites support microbial activity that increases nitrogen and phosphorus availability to plants [108].
To conclude this section, Table 1 provides a summary of the main microalgae and cyanobacteria species with biofertilizer potential.
4. Biocontrol
Plants are constantly exposed to a wide range of biotic stressors, including fungi, bacteria, nematodes, insects, and viruses. In response, they have evolved complex and sophisticated defense mechanisms. The production of secondary metabolites plays a crucial role due to their effectiveness in pathogen and pest resistance [109]. Compounds such as saponin have been widely recognized for their antifungal activity and are considered key components of plant defense systems. These molecules can be harnessed to develop novel, eco-friendly strategies for disease control, reducing the environmental impact of chemical pesticides [110,111]. While many of these compounds are traditionally derived from plants, recent studies have identified the potential of microalgae as a promising alternative source of secondary metabolites, including alkaloids, flavonoids, terpenes, and essential oils [112]. Microalgae are photosynthetic prokaryotic and eukaryotic microorganisms capable of forming beneficial associations with plants, which can enhance the production of secondary metabolites, especially under abiotic stress conditions. Among these metabolites, allelochemicals produced by microalgae have garnered particular interest in their potential applications in sustainable crop protection and biocontrol strategies (Figure 2).
Recent research has increasingly explored the bioactivity of these allelochemicals, emphasizing their role in agricultural systems [113]. The persistent and widespread use of synthetic pesticides raises serious environmental and health concerns, highlighting the need for alternative pest management tools [114]. Microalgae and specially cyanobacteria represent a prolific source of biologically active compounds involved in allelopathic interactions, many of which could be used for pest control and crop protection (Table 2).
4.1. Phytopathogen Resistance
Allelochemicals from microalgae have demonstrated strong antimicrobial activity against a wide spectrum of phytopathogens. For instance, hapalindole T (an antibacterial alkaloid from Fischerella sp.), nostofungicin (a fungicidal lipopeptide from Nostoc commune), and eicosapentaenoic acid (antimicrobial fatty acid from Phaeodactylum tricornutum) are notable examples [123,124,125,126]. Moreover, cell extracts from Chlorella vulgaris and Tetradesmus obliquus have been successfully applied to protect spinach crops against Fusarium oxysporum infections [127]. Gene editing in Chlamydomonas reinhardtii has also been used to enhance bacterial resistance in tobacco plants [128].
Several studies have evaluated the antifungal activity of microalgal extracts across diverse strains. Anabaena HSSASE11 and Oscillatoria nigroviridis HSSASE15 showed inhibitory effects against Botryodiplodia theobromae and Pythium ultimum, respectively, while Dunaliella HSSASE13 was effective against Fusarium solani. Similar antifungal results were reported for Scenedesmus obliquus extracts against Sclerotium rolfsii [129]. Interestingly, some microalgal extracts also show nematicidal activity. For example, Scenedesmus obliquus, Chlorella vulgaris, and Anabaena oryzae were able to inhibit the banana pathogen Meloidogyne incognita [130]. The antimicrobial activity of these extracts is largely attributed to phenolics, alkaloids, and peptides. However, in many cases, the specific active compounds have not been fully identified or characterized. While the mechanisms remain under investigation, the antifungal effects of phenolic compounds may involve interference with fungal cell wall biosynthesis [131].
4.2. Microalgae as Herbicides
The herbicidal potential of microalgal metabolites is an emerging field with encouraging preliminary findings. Several allelochemicals—mainly from cyanobacteria—have demonstrated phytotoxic activity. For instance, cyanobacterin, a phenolic compound produced by Scytonema hofmanni, inhibits photosynthetic electron transport. Similarly, nostocyclamide (a peptide from Nostoc sp.) and fischerellins (alkaloids from Fischerella sp.) interfere with photosystem II [132,133].
Other compounds like microcystins (peptides that inhibit protein phosphatases) and cryptophycins (polyketides that block microtubule polymerization) also show potential as herbicides [132,134]. Despite these findings, research on herbicidal compounds from eukaryotic microalgae remains limited, presenting an underexplored area with high potential for sustainable weed management.
4.3. Microalgae as Insecticides
Among all properties shown from microalgae extracts along this review, insecticide activity is another one proposed. There are several studies in which microalgae have demonstrated his insecticide properties. In the case of diatoms or chlorophytes, the potential insecticide activity has been investigated. For example, the exploitation of Chlamydomonas reinhardtii extracts in the development of preparations combined with microparticles of zinc oxide was able to improve the larvicidal potential of Tenebrio molitor compared to zinc oxide alone treatment [135]. Another study has demonstrated the extract from Amphora coffeaeformis and Scenedesmus oliquus presented larvicidal activity against Culex pipiens [136].
In another approach, biofilm-forming cyanobacteria were found to enhance plant defense mechanisms against insects. Some studies have demonstrated that biofilm from Fischerella ATCC 43239 increased the mortality of larvae from Chirinimus riparius, demonstrating that the biofilm improves the production of allelochemicals with insecticide activity [137,138,139,140]. Moreover, biomass extracts from Microcystis aeruginosa 205 and Anabaena circinalis 86 showed high toxicity against larvae of Aedes aegypti [141]. Alternatively, some researchers have described both unsaturated and saturated fatty acids are responsible for the insecticide activity of microalgae extracts against larvae from different species. Thus, the possible mechanism of larvicidal activity of fatty acids was investigated. The authors tested the inhibition properties of various fatty acids demonstrating that linolenic and linoleic acids might have a dual mode of action against octopamine signaling pathways [142,143].
5. Commercialized Microalgae- and Cyanobacteria-Based Products
An increasing amount of scientific research demonstrating the effectiveness of microalgae in agriculture has driven the development of an emerging and fast-growing market for microalgae-derived bioproducts. Commercial formulations such as Algafert, PhycoTerra®, and Spiralgrow have already entered the global market, targeting both conventional and organic farming sectors (Table 3). These products are offered as liquid concentrates, dry powders, or granules, and are often marketed for their ability to improve root development, nutrient uptake, and crop resilience under stress. Start-ups and aggrotech companies are also investing in vertically integrated production systems that combine algae cultivation with carbon capture and waste valorization, aligning with global sustainability goals [33]. According to recent market projections, the global microalgae-based biofertilizer segment is expected to grow at a compound annual growth rate (CAGR) exceeding 8% over the next five years, driven by regulatory shifts toward low-input agriculture and increasing demand for eco-certified inputs. This momentum underscores the transition of microalgae from an experimental innovation to a commercially viable and scalable component of sustainable agriculture [107].
6. Bottlenecks and Future Perspectives
Microalgae and cyanobacteria have emerged as highly attractive candidates for sustainable agriculture and bio-based industries due to their distinctive biological and ecological traits. These photosynthetic microorganisms exhibit rapid and continuous growth, short generation times, and remarkable metabolic flexibility. Importantly, their cultivation does not compete with traditional agriculture for critical resources. They can be grown on non-arable land using saline water or even wastewater and rely primarily on sunlight as an energy source. These characteristics underscore their potential as complementary components to existing agri-food systems and as contributors to the circular bioeconomy, particularly in the production of food, feed, and high-value fertilizer products.
However, translating this potential into large-scale, economically viable operations presents significant challenges. Among the foremost constraints are the costs and efficiencies associated with cultivation, harvesting, and biomass processing. On the technical front, optimization of both upstream (e.g., growth conditions, nutrient supply, and biomass accumulation) and downstream (e.g., harvesting, extraction, and formulation) processes is essential to ensure yield consistency, biomass quality, and cost-effectiveness [144]. Infrastructure costs (especially for photobioreactors and open pond systems) are substantial, and operational expenditures such as lighting, temperature regulation, fluid circulation, cleaning, and biomass recovery remain high [145]. Closed systems, while offering advantages in terms of contamination control and process stability, are particularly cost intensive. Inoculum preparation constitutes another critical bottleneck; the reliability of large-scale operations depends on the production of high-quality starter cultures. Any microbial contamination or physiological variability during this stage can lead to serious disruptions, compromising the productivity of the entire system.
Operational stability is further challenged by routine maintenance, system downtime, and unexpected failures. Incorporating automation technologies offers a partial solution, reducing labor costs and human error while enabling real-time monitoring, improved safety, and enhanced process control. Nevertheless, successful deployment of these systems requires a multidisciplinary workforce with expertise in microbiology, bioprocess engineering, chemistry, and systems maintenance. Experienced personnel are indispensable for troubleshooting, process optimization, and quality control.
In parallel, a deeper mechanistic understanding of how microalgae and cyanobacteria interact with plant physiology and soil processes is critical to optimizing their use in agricultural contexts. Elucidating their modes of action (such as modulation of plant hormone levels, nutrient uptake, stress resilience, and microbiome interactions) is key to tailoring applications for specific crops, soil types, and climatic conditions. Such knowledge will improve consistency and efficacy under real-world field conditions. To overcome the current limitations, several strategies are under investigation. Continuous cultivation systems provide a stable operational platform and reduce the need for frequent re-inoculation. Increasing biomass density is another focus area, as it minimizes the volume that must be handled during harvesting. Additionally, phycoprospecting efforts aim to identify naturally robust, high-performing strains, while genetic engineering approaches target enhanced productivity, stress resistance, and metabolite synthesis. Integrated biorefinery models are also gaining prominence, wherein multiple valuable compounds (such as proteins, pigments, fatty acids, and polysaccharides) are co-extracted from a single biomass stream, thereby increasing the overall economic viability of production systems.
In summary, while the upscaling of microalgae and cyanobacteria cultivation is constrained by technical and economic barriers, it also presents a promising frontier for sustainable agriculture and industrial biotechnology. With continued innovation in system design, strain selection, process automation, and bioproduct valorization, these microorganisms have the potential to become foundational components of resilient, resource-efficient, and climate-smart production systems.
9. Conclusion
Microalgae and cyanobacteria have emerged as promising biological resources with multiple applications in agriculture. Their roles as biofertilizers, biostimulants, and biocontrol agents show clear potential to improve nutrient availability, support plant growth, and reduce the impact of biotic and abiotic stress. Beyond their agronomic benefits, they also contribute to more sustainable farming through nutrient recycling, carbon capture, and lower dependence on synthetic fertilizers and pesticides. Their integration into circular economy models further highlights their relevance in the transition toward more environmentally friendly agricultural practices. However, important challenges remain, particularly regarding large-scale production, formulation, and regulatory approval. Continued research and technological development will be essential to address these barriers and bring their practical use closer to reality. Overall, microalgae and cyanobacteria offer a valuable opportunity to improve the sustainability of agriculture.
Author Contributions
Conceptualization, ED-S; Manuscript writing—original draft preparation, AJ-F, LGH-M, GT-C and ED-S; writing—review and editing, GT-C and ED-S; manuscript supervision, GT-C and ED-S. AJ-F and LGH-M contributed equally to this work. The manuscript was corrected, revised and approved by all authors. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Acknowledgments
Figure 1 was created in BioRender. Diaz Santos, E. (2025) https://BioRender.com/9lwoss5.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Ren, C.G.; Kong, C.C.; Li, S.M.; Wang, X.J.; Yu, X.; Wang, Y.C.; Qin, S.; Cui, H.L. Symbiotic Microalgae and Microbes: A New Frontier in Saline Agriculture. Front Microbiol 2025, 16. [Google Scholar] [CrossRef]
- Ren, C.G.; Kong, C.C.; Liu, Z.Y.; Zhong, Z.H.; Yang, J.C.; Wang, X.L.; Qin, S. A Perspective on Developing a Plant ‘Holobiont’ for Future Saline Agriculture. Front Microbiol 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Singh, A. Soil Salinization Management for Sustainable Development: A Review. J Environ Manage 2021, 277. [Google Scholar] [CrossRef]
- Cuevas, J.; Daliakopoulos, I.N.; Del Moral, F.; Hueso, J.J.; Tsanis, I.K. A Review of Soil-Improving Cropping Systems for Soil Salinization. Agronomy 2019, 9. [Google Scholar] [CrossRef]
- Brito-Lopez, C.; Van Der Wielen, N.; Barbosa, M.; Karlova, R. Plant Growth-Promoting Microbes and Microalgae-Based Biostimulants: Sustainable Strategy for Agriculture and Abiotic Stress Resilience. Philosophical Transactions of the Royal Society B: Biological Sciences 2025, 380.
- Crystal-Ornelas, R.; Thapa, R.; Tully, K.L. Soil Organic Carbon Is Affected by Organic Amendments, Conservation Tillage, and Cover Cropping in Organic Farming Systems: A Meta-Analysis. Agric Ecosyst Environ 2021, 312. [Google Scholar] [CrossRef]
- Raihan, A. A Review of Climate Change Mitigation and Agriculture Sustainability through Soil Carbon Sequestration. Journal of Agriculture Sustainability and Environment 2023, 2, 23–56. [Google Scholar] [CrossRef]
- Gonzalez-Gonzalez, L.M.; de-Bashan, L.E. The Potential of Microalgae–Bacteria Consortia to Restore Degraded Soils. Biology (Basel) 2023, 12. [Google Scholar] [CrossRef]
- Alvarez, A.L.; Weyers, S.L.; Goemann, H.M.; Peyton, B.M.; Gardner, R.D. Microalgae, Soil and Plants: A Critical Review of Microalgae as Renewable Resources for Agriculture. Algal Res 2021, 54. [Google Scholar] [CrossRef]
- Spinola, M.V.; Díaz-Santos, E. Microalgae Nutraceuticals: The Role of Lutein in Human Health. In Microalgae Biotechnology for Food, Health and High Value Products; 2020.
- León-Bañares, R.; González-Ballester, D.; Galván, A.; Fernández, E. Transgenic Microalgae as Green Cell-Factories. Trends Biotechnol 2004, 22. [Google Scholar] [CrossRef]
- Maurya, N.; Sharma, A.; Sundaram, S. The Role of PGPB-Microalgae Interaction in Alleviating Salt Stress in Plants. Curr Microbiol 2024, 81. [Google Scholar] [CrossRef] [PubMed]
- Farmonaut® Agriculture Biologicals Market: 2025 Growth & Research Guide. https://farmonaut.com/blogs/agriculture-investments-2025-sustainable-growth-guide.
- Barkia, I.; Saari, N.; Manning, S.R. Microalgae for High-Value Products towards Human Health and Nutrition. Mar Drugs 2019, 17. [Google Scholar] [CrossRef] [PubMed]
- Vangenechten, B.; De Coninck, B.; Ceusters, J. How to Improve the Potential of Microalgal Biostimulants for Abiotic Stress Mitigation in Plants? Front Plant Sci 2025, 16. [Google Scholar] [CrossRef]
- Tyagi, J.; Ahmad, S.; Malik, M. Nitrogenous Fertilizers: Impact on Environment Sustainability, Mitigation Strategies, and Challenges. International Journal of Environmental Science and Technology 2022, 19. [Google Scholar] [CrossRef]
- Prasanna, R.; Sood, A.; Ratha, S.K. ; K.; Singh, P. Cyanobacteria as a “Green” Option for Sustainable Agriculture. In Cyanobacteria: An Economic Perspective; 2013.
- Ferreira, A.; Bastos, C.R.V.; Marques-dos-Santos, C.; Acién-Fernandez, F.G.; Gouveia, L. Algaeculture for Agriculture: From Past to Future. Frontiers in Agronomy 2023, 5. [Google Scholar] [CrossRef]
- Ronga, D.; Biazzi, E.; Parati, K.; Carminati, D.; Carminati, E.; Tava, A. Microalgal Biostimulants and Biofertilisers in Crop Productions. Agronomy 2019, 9, 1–22. [Google Scholar] [CrossRef]
- Yuan, H.; Huang, P.; Yu, J.; Wang, P.; Jiang, H. Bin; Jiang, Y.; Deng, S.; Huang, Z.; Yu, J.; Zhu, W. Efficient Wastewater Treatment and Biomass Co-Production Using Energy Microalgae to Fix C, N, and P. RSC Adv 2025, 15, 14030–14041. [Google Scholar] [CrossRef] [PubMed]
- Parmar, P.; Kumar, R.; Neha, Y.; Srivatsan, V. Microalgae as next Generation Plant Growth Additives: Functions, Applications, Challenges and Circular Bioeconomy Based Solutions. Front Plant Sci 2023, 14. [Google Scholar] [CrossRef]
- Onyeaka, H.; Miri, T.; Obileke, K.C.; Hart, A.; Anumudu, C.; Al-Sharify, Z.T. Minimizing Carbon Footprint via Microalgae as a Biological Capture. Carbon Capture Science and Technology 2021, 1. [Google Scholar] [CrossRef]
- Du Jardin, P. Plant Biostimulants: Definition, Concept, Main Categories and Regulation. Sci Hortic 2015, 196. [Google Scholar] [CrossRef]
- Chiaiese, P.; Corrado, G.; Colla, G.; Kyriacou, M.C.; Rouphael, Y. Renewable Sources of Plant Biostimulation: Microalgae as a Sustainable Means to Improve Crop Performance. Front Plant Sci 2018, 871. [Google Scholar] [CrossRef]
- Chabili, A.; Minaoui, F.; Hakkoum, Z.; Douma, M.; Meddich, A.; Loudiki, M. A Comprehensive Review of Microalgae and Cyanobacteria-Based Biostimulants for Agriculture Uses. Plants 2024, 13. [Google Scholar] [CrossRef]
- León-Vaz, A.; León, R.; Díaz-Santos, E.; Vigara, J.; Raposo, S. Using Agro-Industrial Wastes for Mixotrophic Growth and Lipids Production by the Green Microalga Chlorella sorokiniana. N Biotechnol 2019, 51. [Google Scholar] [CrossRef]
- Rozenberg, J.M.; Sorokin, B.A.; Mukhambetova, A.N.; Emelianova, A.A.; Kuzmin, V.V.; Belogurova-Ovchinnikova, O.Y.; Kuzmin, D.V. Recent Advances and Fundamentals of Microalgae Cultivation Technology. Biotechnol J 2024, 19. [Google Scholar] [CrossRef]
- Janssen, M.; Kuijpers, T.C.; Veldhoen, B.; Ternbach, M.B.; Tramper, J.; Mur, L.R.; Wijffels, R.H. Specific Growth Rate of Chlamydomonas reinhardtii and Chlorella sorokiniana under Medium Duration Light/Dark Cycles: 13-87 s. Prog Ind Microbiol 1999, 35. [Google Scholar] [CrossRef]
- Santini, G.; Biondi, N.; Rodolfi, L.; Tredici, M.R. Plant Biostimulants from Cyanobacteria: An Emerging Strategy to Improve Yields and Sustainability in Agriculture. Plants 2021, 10. [Google Scholar] [CrossRef]
- Jithesh, M.N.; Shukla, P.S.; Kant, P.; Joshi, J.; Critchley, A.T.; Prithiviraj, B. Physiological and Transcriptomics Analyses Reveal That Ascophyllum nodosum Extracts Induce Salinity Tolerance in Arabidopsis by Regulating the Expression of Stress Responsive Genes. J Plant Growth Regul 2019, 38, 463–478. [Google Scholar] [CrossRef]
- Singh, J.S.; Kumar, A.; Rai, A.N.; Singh, D.P. Cyanobacteria: A Precious Bio-Resource in Agriculture, Ecosystem, and Environmental Sustainability. Front Microbiol 2016, 7. [Google Scholar] [CrossRef]
- Nawaz, T.; Saud, S.; Gu, L.; Khan, I.; Fahad, S.; Zhou, R. Cyanobacteria: Harnessing the Power of Microorganisms for Plant Growth Promotion, Stress Alleviation, and Phytoremediation in the Era of Sustainable Agriculture. Plant Stress 2024, 11. [Google Scholar] [CrossRef]
- Sánchez-Quintero, Á.; Fernandes, S.C.M.; Beigbeder, J.B. Overview of Microalgae and Cyanobacteria-Based Biostimulants Produced from Wastewater and CO2 Streams towards Sustainable Agriculture: A Review. Microbiol Res 2023, 277. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, G.; Huang, Y.; Guo, M.; Song, J.; Zhang, T.; Long, Y.; Wang, B.; Liu, H. A Potential Biofertilizer—Siderophilic Bacteria Isolated From the Rhizosphere of Paris polyphylla Var. yunnanensis. Front Microbiol 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Fiorentino, S.; Bellani, L.; Santin, M.; Castagna, A.; Echeverria, M.C.; Giorgetti, L. Effects of Microalgae as Biostimulants on Plant Growth, Content of Antioxidant Molecules and Total Antioxidant Capacity in Chenopodium quinoa Exposed to Salt Stress. Plants 2025, 14. [Google Scholar] [CrossRef]
- Moon, J.; Park, Y.J.; Choi, Y. Bin; Truong, T.Q.; Huynh, P.K.; Kim, Y.B.; Kim, S.M. Physiological Effects and Mechanisms of Chlorella vulgaris as a Biostimulant on the Growth and Drought Tolerance of Arabidopsis thaliana. Plants 2024, 13. [Google Scholar] [CrossRef] [PubMed]
- Minaoui, F.; Hakkoum, Z.; Chabili, A.; Douma, M.; Mouhri, K.; Loudiki, M. Biostimulant Effect of Green Soil Microalgae Chlorella vulgaris Suspensions on Germination and Growth of Wheat (Triticum aestivum Var. achtar) and Soil Fertility. Algal Res 2024, 82, 103655. [Google Scholar] [CrossRef]
- La Bella, E.; Baglieri, A.; Rovetto, E.I.; Stevanato, P.; Puglisi, I. Foliar Spray Application of Chlorella vulgaris Extract: Effect on the Growth of Lettuce Seedlings. Agronomy 2021, 11. [Google Scholar] [CrossRef]
- Gitau, M.M.; Farkas, A.; Balla, B.; Ördög, V.; Futó, Z.; Maróti, G. Strain-Specific Biostimulant Effects of Chlorella and Chlamydomonas Green Microalgae on Medicago truncatula. Plants 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Shim, C.K.; Kim, Y.K.; Ko, B.G.; Park, J.H.; Hwang, S.G.; Kim, B.H. Effect of Biostimulator Chlorella fusca on Improving Growth and Qualities of Chinese Chives and Spinach in Organic Farm. Plant Pathol J (Faisalabad) 2018, 34. [Google Scholar] [CrossRef]
- Abdel-Tawwab, M.; Mousa, M.A.A.; Mamoon, A.; Abdelghany, M.F.; Abdel-Hamid, E.A.A.; Abdel-Razek, N.; Ali, F.S.; Shady, S.H.H.; Gewida, A.G.A. Dietary Chlorella Vulgaris Modulates the Performance, Antioxidant Capacity, Innate Immunity, and Disease Resistance Capability of Nile Tilapia Fingerlings Fed on Plant-Based Diets. Anim Feed Sci Technol 2022, 283. [Google Scholar] [CrossRef]
- Wuang, S.C.; Khin, M.C.; Chua, P.Q.D.; Luo, Y.D. Use of Spirulina Biomass Produced from Treatment of Aquaculture Wastewater as Agricultural Fertilizers. Algal Res 2016, 15, 59–64. [Google Scholar] [CrossRef]
- Priya, H.; Prasanna, R.; Ramakrishnan, B.; Bidyarani, N.; Babu, S.; Thapa, S.; Renuka, N. Influence of Cyanobacterial Inoculation on the Culturable Microbiome and Growth of Rice. Microbiol Res 2015, 171. [Google Scholar] [CrossRef]
- Fimbres-Olivarria, D.; Carvajal-Millan, E.; Lopez-Elias, J.A.; Martinez-Robinson, K.G.; Miranda-Baeza, A.; Martinez-Cordova, L.R.; Enriquez-Ocaña, F.; Valdez-Holguin, J.E. Chemical Characterization and Antioxidant Activity of Sulfated Polysaccharides from Navicula sp. Food Hydrocoll 2018, 75, 229–236. [Google Scholar] [CrossRef]
- Piotrowski, K.; Romanowska-Duda, Z.; Messyasz, B. Cultivation of Energy Crops by Ecological Methods under the Conditions of Global Climate and Environmental Changes with the Use of Diatom Extract as a Natural Source of Chemical Compounds. Acta Physiol Plant 2020, 42. [Google Scholar] [CrossRef]
- Kiran Marella, T.; Saxena, A.; Tiwari, A. Diatom Mediated Heavy Metal Remediation: A Review. Bioresour Technol 2020, 305. [Google Scholar] [CrossRef] [PubMed]
- Elhamji, S.; Haydari, I.; Sbihi, K.; Aziz, K.; Elleuch, J.; Kurniawan, T.A.; Chen, Z.; Yap, P.S.; Aziz, F. Uncovering Applicability of Navicula permitis Algae in Removing Phenolic Compounds: A Promising Solution for Olive Mill Wastewater Treatment. Journal of Water Process Engineering 2023, 56. [Google Scholar] [CrossRef]
- Calatrava, V.; Hom, E.F.Y.; Guan, Q.; Llamas, A.; Fernández, E.; Galván, A. Genetic Evidence for Algal Auxin Production in Chlamydomonas and Its Role in Algal-Bacterial Mutualism. iScience 2024, 27. [Google Scholar] [CrossRef]
- Torres-Tiji, Y.; Fields, F.J.; Yang, Y.; Heredia, V.; Horn, S.J.; Keremane, S.R.; Jin, M.M.; Mayfield, S.P. Optimized Production of a Bioactive Human Recombinant Protein from the Microalgae Chlamydomonas reinhardtii Grown at High Density in a Fed-Batch Bioreactor. Algal Res 2022, 66, 102786. [Google Scholar] [CrossRef]
- Llamas, A.; Leon-Miranda, E.; Tejada-Jimenez, M. Microalgal and Nitrogen-Fixing Bacterial Consortia: From Interaction to Biotechnological Potential. Plants 2023, 12. [Google Scholar] [CrossRef]
- Michalak, I.; Chojnacka, K. Algae as Production Systems of Bioactive Compounds. Eng Life Sci 2015, 15. [Google Scholar] [CrossRef]
- Vessey, J.K. Plant Growth Promoting Rhizobacteria as Biofertilizers; Plant and Soil, 2003, 255, 571-586. [CrossRef]
- Mahanty, T.; Bhattacharjee, S.; Goswami, M.; Bhattacharyya, P.; Das, B.; Ghosh, A.; Tribedi, P. Biofertilizers: A Potential Approach for Sustainable Agriculture Development. Environmental Science and Pollution Research 2017, 24, 3315–3335. [Google Scholar] [CrossRef]
- Bhardwaj, D.; Ansari, M.W.; Sahoo, R.K.; Tuteja, N. Biofertilizers Function as Key Player in Sustainable Agriculture by Improving Soil Fertility, Plant Tolerance and Crop Productivity. Microb Cell Fact 2014, 13. [Google Scholar] [CrossRef]
- Nobbe and Hiltner. Bodenimpfung für anbau von leguminosen. Sächsische. Landwirtschaftliche Zeitschrift, 1896, 44, pp. [Google Scholar]
- Nosheen, S.; Ajmal, I.; Song, Y. Microbes as Biofertilizers, a Potential Approach for Sustainable Crop Production. Sustainability (Switzerland) 2021, 13, 1–20. [Google Scholar] [CrossRef]
- Aloo, B.N.; Tripathi, V.; Makumba, B.A.; Mbega, E.R. Plant Growth-Promoting Rhizobacterial Biofertilizers for Crop Production: The Past, Present, and Future. Front Plant Sci 2022, 13. [Google Scholar] [CrossRef]
- Daniel, A.I.; Fadaka, A.O.; Gokul, A.; Bakare, O.O.; Aina, O.; Fisher, S.; Burt, A.F.; Mavumengwana, V.; Keyster, M.; Klein, A. Biofertilizer: The Future of Food Security and Food Safety. Microorganisms 2022, 10. [Google Scholar] [CrossRef]
- Herridge, D.F.; Peoples, M.B.; Boddey, R.M. Global Inputs of Biological Nitrogen Fixation in Agricultural Systems. Plant Soil 2008, 311, 1–18. [Google Scholar] [CrossRef]
- Elnahal, A.S.M.; El-Saadony, M.T.; Saad, A.M.; Desoky, E.S.M.; El-Tahan, A.M.; Rady, M.M.; AbuQamar, S.F.; El-Tarabily, K.A. The Use of Microbial Inoculants for Biological Control, Plant Growth Promotion, and Sustainable Agriculture: A Review. Eur J Plant Pathol 2022, 162, 759–792. [Google Scholar] [CrossRef]
- Adeleke, R.A.; Raimi, A.R.; Roopnarain, A.; Mokubedi, S.M. Status and Prospects of Bacterial Inoculants for Sustainable Management of Agroecosystems. In; 2019.
- Saleem, I.; Asad, W.; Kiran, T.; Asad, S.B.; Khaliq, S.; Ali Mohammed Al-Harethi, A.; Mallasiy, L.O.; Shah, T.A. Enhancing Spinach Growth with a Biofertilizer Derived from Chicken Feathers Using a Keratinolytic Bacterial Consortium. BMC Microbiol 2025, 25. [Google Scholar] [CrossRef] [PubMed]
- Álvarez, C.; Jimenez-Ríos, L.; Iniesta-Pallares, M.; Jurado-Flores, A.; Molina-Heredia, F.P.; Ng, C.K.Y.; Mariscal, V. Symbiosis between Cyanobacteria and Plants: From Molecular Studies to Agronomic Applications. J Exp Bot 2023, 74, 6145–6157. [Google Scholar] [CrossRef]
- Ma, F.; Li, Y.; Han, X.; Li, K.; Zhao, M.; Guo, L.; Li, S.; Wang, K.; Qin, K.; Duan, J.; et al. Microalgae-Based Biofertilizer Improves Fruit Yield and Controls Greenhouse Gas Emissions in a Hawthorn Orchard. PLoS One 2024, 19. [Google Scholar] [CrossRef]
- Cordell, D.; Drangert, J.O.; White, S. The Story of Phosphorus: Global Food Security and Food for Thought. Global Environmental Change 2009, 19, 292–305. [Google Scholar] [CrossRef]
- Soumare, A.; Boubekri, K.; Lyamlouli, K.; Hafidi, M.; Ouhdouch, Y.; Kouisni, L. From Isolation of Phosphate Solubilizing Microbes to Their Formulation and Use as Biofertilizers: Status and Needs. Front Bioeng Biotechnol 2020, 7. [Google Scholar] [CrossRef]
- Gureev, A.P.; Kryukova, V.A.; Eremina, A.A.; Alimova, A.A.; Kirillova, M.S.; Filatova, O.A.; Moskvitina, M.I.; Kozin, S.V.; Lyasota, O.M.; Gureeva, M.V. Plant-Growth Promoting Rhizobacteria Azospirillum Partially Alleviate Pesticide-Induced Growth Retardation and Oxidative Stress in Wheat (Triticum aestivum L.). Plant Growth Regul 2024, 104, 503–521. [Google Scholar] [CrossRef]
- Gupta, R.; Kumari, A.; Sharma, S.; Alzahrani, O.M.; Noureldeen, A.; Darwish, H. Identification, Characterization and Optimization of Phosphate Solubilizing Rhizobacteria (PSRB) from Rice Rhizosphere. Saudi J Biol Sci 2022, 29, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Sarikhani, M.R.; Khoshru, B.; Greiner, R. Isolation and Identification of Temperature Tolerant Phosphate Solubilizing Bacteria as a Potential Microbial Fertilizer. World J Microbiol Biotechnol 2019, 35. [Google Scholar] [CrossRef]
- Patel, S.K.; Singh, S.; Benjamin, J.C.; Singh, V.R.; Bisht, D.; Lal, R.K. Plant Growth-Promoting Activities of Serratia marcescens and Pseudomonas fluorescens on Capsicum annuum L. Plants. Ecological Frontiers 2024, 44, 654–663. [Google Scholar] [CrossRef]
- Tariq, M.; Tahreem, N.; Zafar, M.; Raza, G.; Shahid, M.; Zunair, M.; Iram, W.; Zahra, S.T. Occurrence of Diverse Plant Growth Promoting Bacteria in Soybean [Glycine Max (L.) Merrill] Root Nodules and Their Prospective Role in Enhancing Crop Yield. Biocatal Agric Biotechnol 2024, 57. [Google Scholar] [CrossRef]
- Ibrahim, E.; Ahmad, A.A.; Abdo, E.S.; Bakr, M.A.; Khalil, M.A.; Abdallah, Y.; Ogunyemi, S.O.; Mohany, M.; Al-Rejaie, S.S.; Shou, L.; et al. Suppression of Root Rot Fungal Diseases in Common Beans (Phaseolus vulgaris L.) through the Application of Biologically Synthesized Silver Nanoparticles. Nanomaterials 2024, 14. [Google Scholar] [CrossRef]
- Han, L.; Zhang, H.; Bai, X.; Jiang, B. The Peanut Root Exudate Increases the Transport and Metabolism of Nutrients and Enhances the Plant Growth-Promoting Effects of Burkholderia pyrrocinia Strain P10. BMC Microbiol 2023, 23. [Google Scholar] [CrossRef] [PubMed]
- Akintokun, A.K.; Ezaka, E.; Akintokun, P.O.; Shittu, O.B.; Taiwo, L.B. Isolation, Screening and Response of Maize to Plant Growth Promoting Rhizobacteria Inoculants. Scientia Agriculturae Bohemica 2019, 50, 181–190. [Google Scholar] [CrossRef]
- Boubekri, K.; Soumare, A.; Mardad, I.; Lyamlouli, K.; Hafidi, M.; Ouhdouch, Y.; Kouisni, L. The Screening of Potassium-and Phosphate-Solubilizing Actinobacteria and the Assessment of Their Ability to Promote Wheat Growth Parameters. Microorganisms 2021, 9, 1–16. [Google Scholar] [CrossRef]
- Wang, J.; Li, R.; Zhang, H.; Wei, G.; Li, Z. Beneficial Bacteria Activate Nutrients and Promote Wheat Growth under Conditions of Reduced Fertilizer Application. BMC Microbiol 2020, 20. [Google Scholar] [CrossRef]
- Devi, R.; Kaur, T.; Negi, R.; Kour, D.; Kumar, S.; Yadav, A.; Singh, S.; Chaubey, K.K.; Rai, A.K.; Shreaz, S.; et al. Bioformulation of Mineral Solubilizing Microbes as Novel Microbial Consortium for the Growth Promotion of Wheat (Triticum aestivum) under the Controlled and Natural Conditions. Heliyon 2024, 10. [Google Scholar] [CrossRef] [PubMed]
- Singh Meena, V.; Ram Maurya, B.; Kumari Meena, S.; Kumar Mishra, P.; Kumar Bisht, J.; Pattanayak, A. Potassium Solubilization: Strategies to Mitigate Potassium Deficiency in Agricultural Soils.
- Cakmak, I.; McLaughlin, M.J.; White, P. Zinc for Better Crop Production and Human Health. Plant Soil 2017, 411, 1–4. [Google Scholar] [CrossRef]
- Hussain Shah, A.; Naz, I.; Ahmad, H.; Nasreen Khokhar, S.; Khan, K. Impact of Zinc Solubilizing Bacteria on Zinc Contents of Wheat. J. Agric. & Environ. Sci 2016, 16, 449–454. [Google Scholar] [CrossRef]
- Sultan, A.A.Y.A.; Gebreel, H.M.; Youssef, H.A.I.A.E. Biofertilizer Effect of Some Zinc Dissolving Bacteria Free and Encapsulated on Zea mays Growth. Arch Microbiol 2023, 205. [Google Scholar] [CrossRef]
- Karnwal, A. Enhancing Zinc Levels in Solanum lycopersicum L. through Biofortification with Plant Growth-Promoting Pseudomonas spp. Isolated from Cow Dung. Biotechnologia 2023, 104, 157–169. [Google Scholar] [CrossRef]
- Briat, J.F.; Dubos, C.; Gaymard, F. Iron Nutrition, Biomass Production, and Plant Product Quality. Trends Plant Sci 2015, 20, 33–40. [Google Scholar] [CrossRef]
- Chaudhary, S.; Sindhu, S.S. Iron Sensing, Signalling and Acquisition by Microbes and Plants under Environmental Stress: Use of Iron-Solubilizing Bacteria in Crop Biofortification for Sustainable Agriculture. Plant Science 2025, 356. [Google Scholar] [CrossRef]
- Timofeeva, A.M.; Galyamova, M.R.; Sedykh, S.E. Bacterial Siderophores: Classification, Biosynthesis, Perspectives of Use in Agriculture. Plants 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Ghazanfar, S.; Hussain, A.; Dar, A.; Ahmad, M.; Anwar, H.; Al Farraj, D.A.; Rizwan, M.; Iqbal, R. Prospects of Iron Solubilizing Bacillus Species for Improving Growth and Iron in Maize (Zea mays L.) under Axenic Conditions. Sci Rep 2024, 14. [Google Scholar] [CrossRef]
- Montero-Palmero, B.; Lucas, J.A.; Montalbán, B.; García-Villaraco, A.; Gutierrez-Mañero, J.; Ramos-Solano, B. Iron Deficiency in Tomatoes Reversed by Pseudomonas Strains: A Synergistic Role of Siderophores and Plant Gene Activation. Plants 2024, 13. [Google Scholar] [CrossRef]
- Davidi, L.; Gallaher, S.D.; Ben-David, E.; Purvine, S.O.; Fillmore, T.L.; Nicora, C.D.; Craig, R.J.; Schmollinger, S.; Roje, S.; Blaby-Haas, C.E.; et al. Pumping Iron: A Multi-Omics Analysis of Two Extremophilic Algae Reveals Iron Economy Management. Proc Natl Acad Sci U S A 2023, 120. [Google Scholar] [CrossRef] [PubMed]
- Brick, M.B.; Hussein, M.H.; Mowafy, A.M.; Hamouda, R.A.; Ayyad, A.M.; Refaay, D.A. Significance of Siderophore-Producing Cyanobacteria on Enhancing Iron Uptake Potentiality of Maize Plants Grown under Iron-Deficiency. Microb Cell Fact 2025, 24. [Google Scholar] [CrossRef] [PubMed]
- Nakano, M.; Omae, N.; Tsuda, K. Inter-Organismal Phytohormone Networks in Plant-Microbe Interactions. Curr Opin Plant Biol 2022, 68. [Google Scholar] [CrossRef] [PubMed]
- Yin, W.; Dong, N.; Li, X.; Yang, Y.; Lu, Z.; Zhou, W.; Qian, Q.; Chu, C.; Tong, H. Understanding Brassinosteroid-Centric Phytohormone Interactions for Crop Improvement. J Integr Plant Biol 2025, 67, 563–581. [Google Scholar] [CrossRef]
- Fujita, Y.; Uesaka, K. Nitrogen Fixation in Cyanobacteria. In Cyanobacterial Physiology: From Fundamentals to Biotechnology; Elsevier, 2022; pp. 29–45 ISBN 9780323961066.
- Neilson, A.; I~ippka, I.; Kv~isawa, R. Heterocyst Formation and Nitrogenase Synthesis in Anabaena sp. A Kinetic Study; Springer-Verlag, 1971; Vol. 76;
- Kollmen, J.; Strieth, D. The Beneficial Effects of Cyanobacterial Co-Culture on Plant Growth. Life 2022, 12. [Google Scholar] [CrossRef]
- Hussain, A.; Shah, S.T.; Rahman, H.; Irshad, M.; Iqbal, A. Effect of IAA on in Vitro Growth and Colonization of Nostoc in Plant Roots. Front Plant Sci 2015, 6. [Google Scholar] [CrossRef]
- Toribio, A.J.; Suárez-Estrella, F.; Jurado, M.M.; López, M.J.; López-González, J.A.; Moreno, J. Prospection of Cyanobacteria Producing Bioactive Substances and Their Application as Potential Phytostimulating Agents. Biotechnology Reports 2020, 26. [Google Scholar] [CrossRef]
- Kimura, J.; Nakano, T. Reconstitution of a Blasia-Nostoc Symbiotic Association under Axenic Conditions. Nova Hedwigia 1990, 50, 191–200. [Google Scholar] [CrossRef]
- Eily, A.N.; Pryer, K.M.; Li, F.W. A First Glimpse at Genes Important to the Azolla–Nostoc Symbiosis. Symbiosis 2019, 78, 149–162. [Google Scholar] [CrossRef]
- Álvarez, C.; Navarro, J.A.; Molina-Heredia, F.P.; Mariscal, V. Endophytic Colonization of Rice (Oryza sativa L.) by the Symbiotic Strain Nostoc punctiforme PCC 73102. Molecular Plant-Microbe Interactions 2020, 33, 1040–1045. [Google Scholar] [CrossRef]
- Bibi, S.; Saadaoui, I.; Bibi, A.; Al-Ghouti, M.; Abu-Dieyeh, M.H. Applications, Advancements, and Challenges of Cyanobacteria-Based Biofertilizers for Sustainable Agro and Ecosystems in Arid Climates. Bioresour Technol Rep 2024, 25. [Google Scholar] [CrossRef]
- Iniesta-Pallarés, M.; Álvarez, C.; Gordillo-Cantón, F.M.; Ramírez-Moncayo, C.; Alves-Martínez, P.; Molina-Heredia, F.P.; Mariscal, V. Sustaining Rice Production through Biofertilization with N2-Fixing Cyanobacteria. Applied Sciences (Switzerland) 2021, 11. [Google Scholar] [CrossRef]
- Ghotbi-Ravandi, A.A.; Shariatmadari, Z.; Riahi, H.; Hassani, S.B.; Heidari, F.; Nohooji, M.G. Enhancement of Essential Oil Production and Expression of Some Menthol Biosynthesis-Related Genes in Mentha piperita Using Cyanobacteria. Iran J Biotechnol 2023, 21, 96–107. [Google Scholar] [CrossRef]
- Jiménez-Ríos, L.; Torrado, A.; González-Pimentel, J.L.; Iniesta-Pallarés, M.; Molina-Heredia, F.P.; Mariscal, V.; Álvarez, C. Emerging Nitrogen-Fixing Cyanobacteria for Sustainable Cotton Cultivation. Science of the Total Environment 2024, 924. [Google Scholar] [CrossRef] [PubMed]
- García-Encinas, J.P.; Ruiz-Cruz, S.; Juárez, J.; Ornelas-Paz, J. de J.; Del Toro-Sánchez, C.L., Ed.; Márquez-Ríos, E. Proteins from Microalgae: Nutritional, Functional and Bioactive Properties. Foods 2025. [Google Scholar]
- Youssef, S.M.; El-Serafy, R.S.; Ghanem, K.Z.; Elhakem, A.; Abdel Aal, A.A. Foliar Spray or Soil Drench: Microalgae Application Impacts on Soil Microbiology, Morpho-Physiological and Biochemical Responses, Oil and Fatty Acid Profiles of Chia Plants under Alkaline Stress. Biology (Basel) 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, H.S.; Hassan, A.; Barakat, K.M.; Ghonam, H.E.B. Improvement of Growth and Biochemical Constituents of Rosmarinus officinalis by Fermented Spirulina maxima Biofertilizer. Plant Physiology and Biochemistry 2024, 208. [Google Scholar] [CrossRef]
- Renuka, N.; Guldhe, A.; Prasanna, R.; Singh, P.; Bux, F. Microalgae as Multi-Functional Options in Modern Agriculture: Current Trends, Prospects and Challenges. Biotechnol Adv 2018, 36, 1255–1273. [Google Scholar] [CrossRef]
- Udaypal, U.; Goswami, R.K.; Verma, P. Strategies for Improvement of Bioactive Compounds Production Using Microalgal Consortia: An Emerging Concept for Current and Future Perspective. Algal Res 2024, 82, 103664. [Google Scholar] [CrossRef]
- Marchiosi, R.; dos Santos, W.D.; Constantin, R.P.; de Lima, R.B.; Soares, A.R.; Finger-Teixeira, A.; Mota, T.R.; de Oliveira, D.M.; Foletto-Felipe, M. de P. ; Abrahão, J.; et al. Biosynthesis and Metabolic Actions of Simple Phenolic Acids in Plants. Phytochemistry Reviews 2020, 19, 865–906. [Google Scholar]
- Xu, H.; Tang, Z.; Yang, D.; Dai, X.; Chen, H. Enhanced Growth and Auto-Flocculation of Scenedesmus quadricauda in Anaerobic Digestate Using High Light Intensity and Nanosilica: A Biomineralization-Inspired Strategy. Water Res 2023, 235. [Google Scholar] [CrossRef]
- Su, Y.; Ren, Y.; Wang, G.; Li, J.; Zhang, H.; Yang, Y.; Pang, X.; Han, J. Microalgae and Microbial Inoculant as Partial Substitutes for Chemical Fertilizer Enhance Polygala Tenuifolia Yield and Quality by Improving Soil Microorganisms. Front Plant Sci 2024, 15. [Google Scholar] [CrossRef]
- Iovinella, M.; Palmieri, M.; Papa, S.; Auciello, C.; Ventura, R.; Lombardo, F.; Race, M.; Lubritto, C.; di Cicco, M.R.; Davis, S.J.; et al. Biosorption of Rare Earth Elements from Luminophores by G. sulphuraria (Cyanidiophytina, Rhodophyta). Environ Res 2023, 239, 117281. [Google Scholar] [CrossRef]
- Shahbaz, A.; Hussain, N.; Saba, S.; Bilal, M. Actinomycetes, Cyanobacteria, and Fungi: A Rich Source of Bioactive Molecules. In Microbial Biomolecules: Emerging Approach in Agriculture, Pharmaceuticals and Environment Management; Elsevier, 2022; pp. 113–133 ISBN 9780323994767.
- Chaïb, S.; Pistevos, J.C.A.; Bertrand, C.; Bonnard, I. Allelopathy and Allelochemicals from Microalgae: An Innovative Source for Bio-Herbicidal Compounds and Biocontrol Research. Algal Res 2021, 54. [Google Scholar] [CrossRef]
- Doan, N.T.; Rickards, R.W.; Rothschild, J.M.; Smith, G.D. Allelopathic Actions of the Alkaloid 12-Epi-Hapalindole E Isonitrile and Calothrixin A from Cyanobacteria of the Genera Fischerella and Calothrix; 2000; Vol. 12;
- Sasso, S.; Pohnert, G.; Lohr, M.; Mittag, M.; Hertweck, C. Microalgae in the Postgenomic Era: A Blooming Reservoir for New Natural Products. FEMS Microbiol Rev 2012, 36, 761–785. [Google Scholar] [CrossRef]
- Baden, D.G.; Bourdelais, A.J.; Jacocks, H.; Michelliza, S.; Naar, J. Natural and Derivative Brevetoxins: Historical Background, Multiplicity, and Effects. Environ Health Perspect 2005, 113, 621–625. [Google Scholar] [CrossRef]
- Yokoyama, A.; Murata, M.; Oshima, Y.; Iwashita, T.; Yasumoto, T. Some Chemical Properties of Maitotoxin, a Putative Calcium Channel Agonist Isolated from a Marine Dinoflagellate1; 1988; Vol. 104;
- DellaGreca, M.; Zarrelli, A.; Fergola, P.; Cerasuolo, M.; Pollio, A.; Pinto, G. Fatty Acids Released by Chlorella Vulgaris and Their Role in Interference with Pseudokirchneriella subcapitata: Experiments and Modelling. J Chem Ecol 2010, 36, 339–349. [Google Scholar] [CrossRef]
- Chiang, I.Z.; Huang, W.Y.; Wu, J.T. Allelochemicals of Botryococcus braunii (Chlorophyceae). J Phycol 2004, 40, 474–480. [Google Scholar] [CrossRef]
- Rastogi, R.P.; Sinha, R.P.; Incharoensakdi, A. The Cyanotoxin-Microcystins: Current Overview. Rev Environ Sci Biotechnol 2014, 13, 215–249. [Google Scholar] [CrossRef]
- Jaki, B.; Orjala, J.; Heilmann, J.; Linden, A.; Vogler, B.; Sticher, O. Novel Extracellular Diterpenoids with Biological Activity from the Cyanobacterium Nostoc commune. J Nat Prod 2000, 63, 339–343. [Google Scholar] [CrossRef] [PubMed]
- Asthana, R.K.; Srivastava, A.; Singh, A.P. ; Deepali; Singh, S. P.; Nath, G.; Srivastava, R.; Srivastava, B.S. Identification of an Antimicrobial Entity from the Cyanobacterium Fischerella sp. Isolated from Bark of Azadirachta Indica (Neem) Tree. J Appl Phycol 2006, 18, 33–39. [Google Scholar] [CrossRef]
- Kajiyama, S.; Kanzakt, H.; Kawazu, K.; Kobayashi, A. Nostofungicidine, an Antifungal Lipopeptide from the Field-Grown Terrestrial Blue-Green Alga Nostoc commune; 1998; Vol. 39;
- Desbois, A.P.; Mearns-Spragg, A.; Smith, V.J. A Fatty Acid from the Diatom Phaeodactylum tricornutum Is Antibacterial against Diverse Bacteria Including Multi-Resistant Staphylococcus aureus (MRSA). Marine Biotechnology 2009, 11, 45–52. [Google Scholar] [CrossRef]
- Ouaddi, O.; Oukarroum, A.; Bouharroud, R.; Alouani, M.; EL blidi, A.; Qessaoui, R.; Radouane, N.; Errafii, K.; Hijri, M.; Hamadi, F.; et al. A Study of Antibacterial Efficacy of Tetraselmis rubens Extracts against Tomato Phytopathogenic Pseudomonas corrugata. Algal Res 2025, 86. [Google Scholar] [CrossRef]
- Viana, C.; Genevace, M.; Gama, F.; Coelho, L.; Pereira, H.; Varela, J.; Reis, M. Chlorella vulgaris and Tetradesmus obliquus Protect Spinach (Spinacia oleracea L.) against Fusarium oxysporum. Plants 2024, 13. [Google Scholar] [CrossRef]
- Sandor, R.; Wagh, S.G.; Kelterborn, S.; Großkinsky, D.K.; Novak, O.; Olsen, N.; Paul, B.; Petřík, I.; Wu, S.; Hegemann, P.; et al. Cytokinin-Deficient Chlamydomonas reinhardtii CRISPR-Cas9 Mutants Show Reduced Ability to Prime Resistance of Tobacco against Bacterial Infection. Physiol Plant 2024, 176. [Google Scholar] [CrossRef] [PubMed]
- Schmid, B.; Coelho, L.; Schulze, P.S.C.; Pereira, H.; Santos, T.; Maia, I.B.; Reis, M.; Varela, J. Antifungal Properties of Aqueous Microalgal Extracts. Bioresour Technol Rep 2022, 18. [Google Scholar] [CrossRef]
- Hamouda, R.; Hamouda, R.A.; El-Ansary, M.S.M. Potential of Plant-Parasitic Nematode Control in Banana Plants by Microalgae as a New Approach Towards Resistance; 2017; Vol. 27;
- Bang, K.H.; Lee, D.W.; Park, H.M.; Rhee, Y.H. Inhibition of Fungal Cell Wall Synthesizing Enzymes by Trans-Cinnamaldehyde. Biosci Biotechnol Biochem 2000, 64, 1061–1063. [Google Scholar] [CrossRef] [PubMed]
- Berry, J. 2011.
- Berry, J.P. Cyanobacterial Toxins as Allelochemicals with Potential Applications as Algaecides, Herbicides and Insecticides. Mar Drugs 2008, 6, 117–146. [Google Scholar] [CrossRef]
- Koodkaew, I.; Sunohara, Y.; Matsuyama, S.; Matsumoto, H. Isolation of Ambiguine D Isonitrile from Hapalosiphon sp. and Characterization of Its Phytotoxic Activity. Plant Growth Regul 2012, 68, 141–150. [Google Scholar] [CrossRef]
- Rankic, I.; Zelinka, R.; Ridoskova, A.; Gagic, M.; Pelcova, P.; Huska, D. Nano/Microparticles in Conjunction with Microalgae Extract as Novel Insecticides against Mealworm beetles, Tenebrio molitor. Sci Rep 2021, 11. [Google Scholar] [CrossRef]
- Hassan, M.E.; Mohafrash, S.M.M.; Fallatah, S.A.; El-Sayed, A.E.K.B.; Mossa, A.T.H. Eco-Friendly Larvicide of Amphora coffeaeformis and Scenedesmus obliquus Microalgae Extracts against Culex pipiens. J Appl Phycol 2021, 33. [Google Scholar] [CrossRef]
- Becher, P.G.; Jüttner, F. Insecticidal Compounds of the Biofilm-Forming Cyanobacterium Fischerella sp. In (ATCC 43239). In Proceedings of the Environmental Toxicology; 2005; Vol. 20; pp. 363–372. [Google Scholar]
- Becher, P.G.; Keller, S.; Jung, G.; Süssmuth, R.D.; Jüttner, F. Insecticidal Activity of 12-Epi-Hapalindole J Isonitrile. Phytochemistry 2007, 68, 2493–2497. [Google Scholar] [CrossRef]
- Becher, P.G.; Jüttner, F. Insecticidal Activity - A New Bioactive Property of the Cyanobacterium Fischerella. Pol J Ecol 2006, 54, 653–662. [Google Scholar]
- Höckelmann, C.; Becher, P.G.; Von Reuß, S.H.; Jüttner, F. Sesquiterpenes of the Geosmin-Producing Cyanobacterium Calothrix PCC 7507 and Their Toxicity to Invertebrates. Zeitschrift fur Naturforschung - Section C Journal of Biosciences 2009, 64, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Delaney, J.M.; Wilkins, R.M. AGAINST VARIOUS INSECT SPECIES; 1995; Vol. 33;
- Aguilar-Marcelino, L.; Pineda-Alegría, J.A.; Salinas-Sánchez, D.O.; Hernández-Velázquez, V.M.; Silva-Aguayo, G.I.; Navarro-Tito, N.; Sotelo-Leyva, C. In Vitro Insecticidal Effect of Commercial Fatty Acids, β-Sitosterol, and Rutin against the Sugarcane aphid, Melanaphis sacchari zehntner (Hemiptera: Aphididae). J Food Prot 2022, 85, 671–675. [Google Scholar] [CrossRef]
- Perumalsamy, H.; Jang, M.J.; Kim, J.R.; Kadarkarai, M.; Ahn, Y.J. Larvicidal Activity and Possible Mode of Action of Four Flavonoids and Two Fatty Acids Identified in Millettia pinnata Seed toward Three Mosquito Species. Parasit Vectors 2015, 8. [Google Scholar] [CrossRef]
- Novoveská, L.; Nielsen, S.L.; Eroldoğan, O.T.; Haznedaroglu, B.Z.; Rinkevich, B.; Fazi, S.; Robbens, J.; Vasquez, M.; Einarsson, H. Overview and Challenges of Large-Scale Cultivation of Photosynthetic Microalgae and Cyanobacteria. Mar Drugs 2023, 21. [Google Scholar] [CrossRef]
- Medeiros, D.L.; Moreira, Í.T.A. Microalgae Biomass Production from Cultivation in Availability and Limitation of Nutrients: The Technical, Environmental and Economic Performance. J Clean Prod 2022, 370, 133538. [Google Scholar] [CrossRef]
Figure 1.
Illustration as an overview of the present revision. (Created using Biorender. Diaz Santos, E. (2025) https://BioRender.com/9lwoss5).
Figure 1.
Illustration as an overview of the present revision. (Created using Biorender. Diaz Santos, E. (2025) https://BioRender.com/9lwoss5).
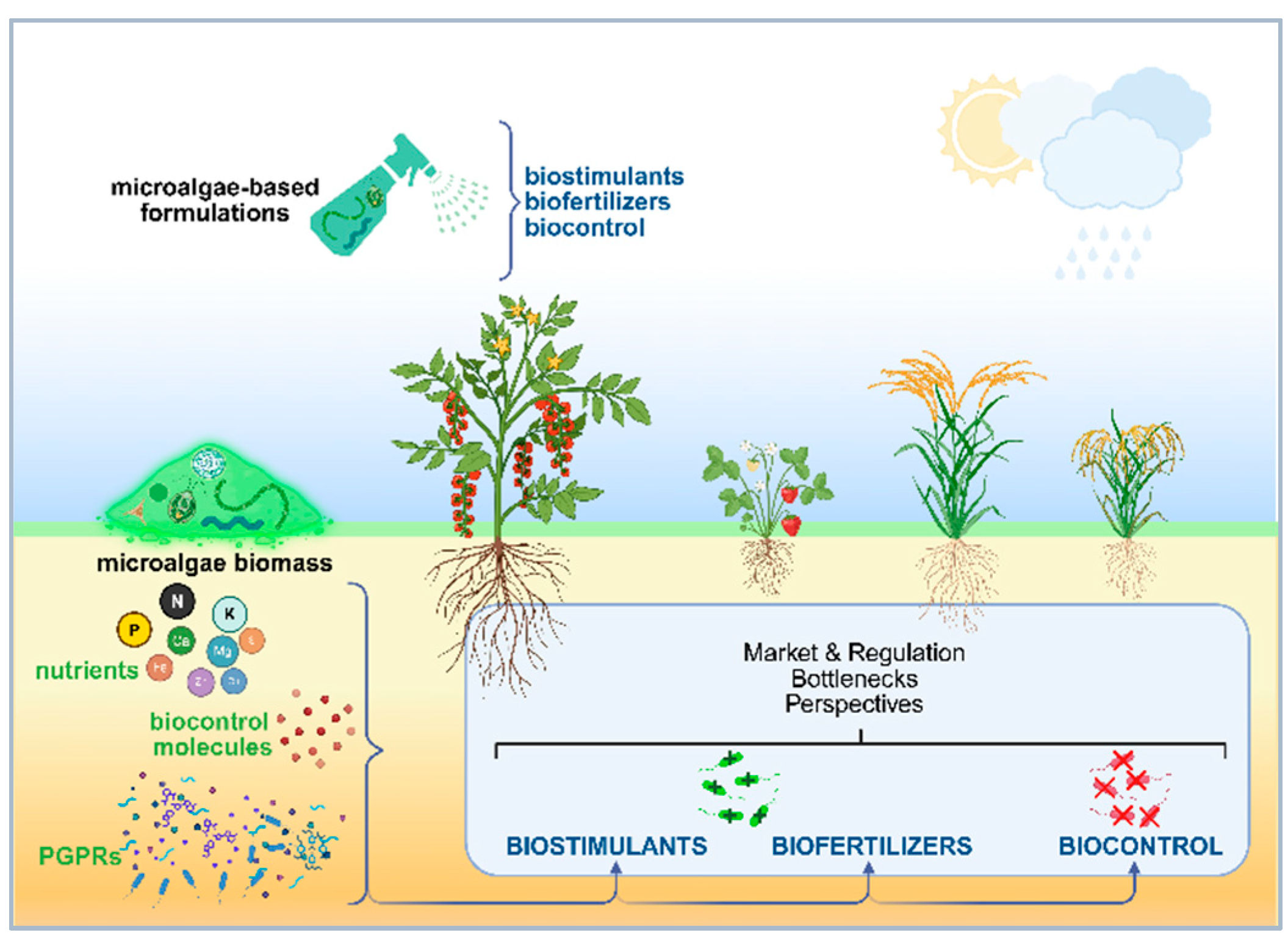
Figure 2.
Schematic representation of biocontrol mediated by extract from microalgae and cyanobacteria, and their activities in plant defense.
Figure 2.
Schematic representation of biocontrol mediated by extract from microalgae and cyanobacteria, and their activities in plant defense.
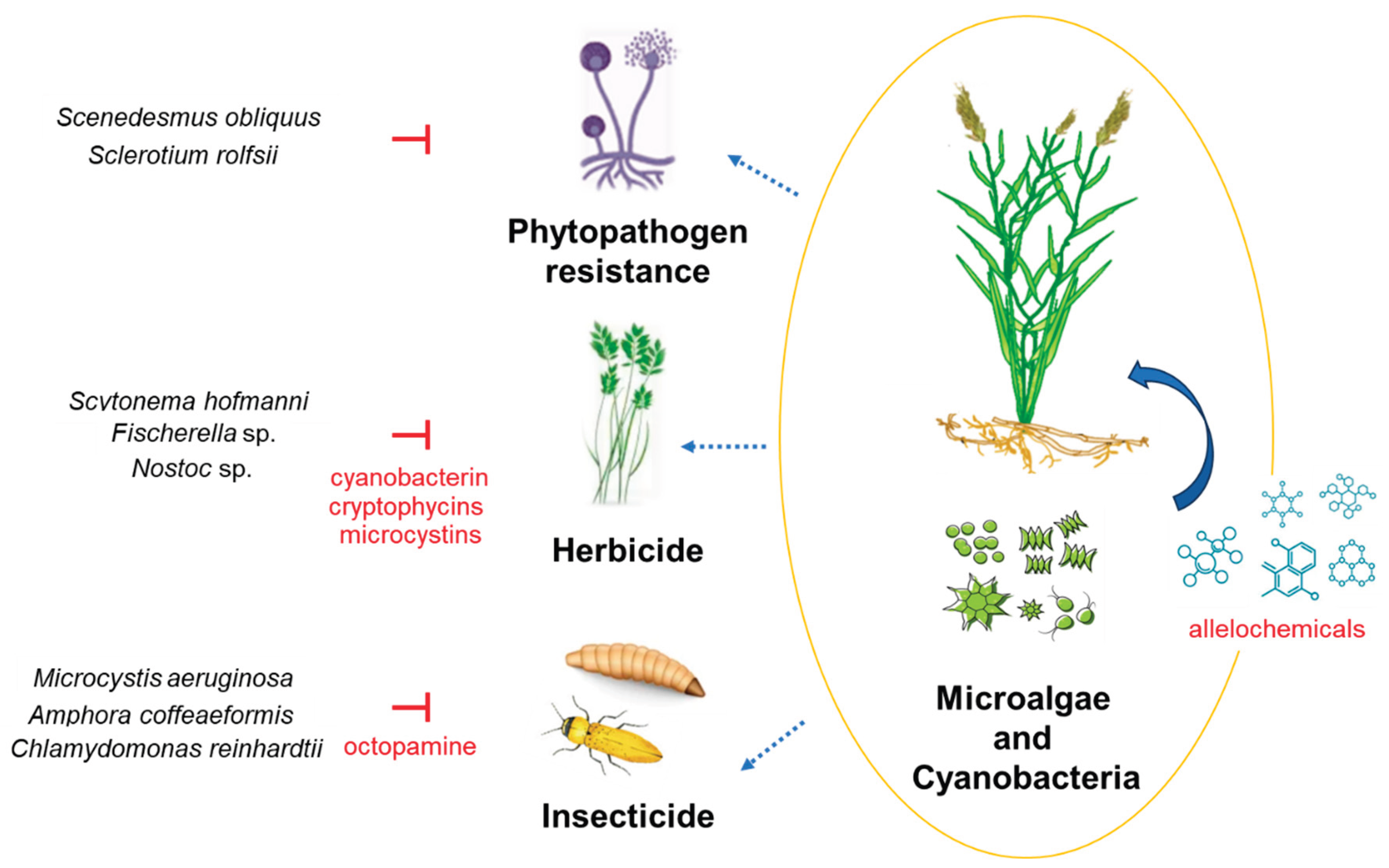

Table 1.
Main microalgae and cyanobacteria species involved in biofertilization.
| Species | Group | Biofertilization mechanism | References |
|---|---|---|---|
| Nostoc sp. | Cyanobacteria | Nitrogen fixation, phytohormone production, solubilization of P/K/Zn | [95,101] |
| Anabaena vaginicola ISB42 | Cyanobacteria | Phytohormone-linked nutrient uptake, peppermint oil enhancement | [102] |
| Nostoc spongiaeforme var. tenue ISB65 | Cyanobacteria | Phytohormone-linked nutrient uptake, peppermint oil enhancement | [102] |
| Synechococcus mundulus | Cyanobacteria | Siderophore production, enhanced Fe uptake in maize | [89] |
| Arthrospira platensis | Cyanobacteria | Phytohormone production, improved nutrient acquisition in chia | [105] |
| Spirulina maxima | Cyanobacteria (marketed as microalgae) |
Bioactive compounds enhancing growth and nutrient uptake in rosemary | [106] |
| Chlorella vulgaris | Microalgae | Phosphorus and potassium solubilization, auxin-like activity | [107] |
| Scenedesmus obliquus | Microalgae | P/K mobilization, root stimulation under stress | [107] |
| Dunaliella salina | Microalgae | Siderophore-mediated Fe uptake under deficiency | [88] |
| Dunaliella bardawil | Microalgae | Siderophore-mediated Fe uptake under deficiency | [88] |
| Dunaliella tertiolecta | Microalgae | Siderophore-mediated Fe uptake | [88] |
| Chlamydomonas reinhardtii | Microalgae | Associative biofertilization via bacterial consortia (N/P uptake) | [108] |
Table 2.
Main secondary metabolites from microalgae and cyanobacteria and their potential applications in agriculture.
Table 2.
Main secondary metabolites from microalgae and cyanobacteria and their potential applications in agriculture.
| Class ofcompound | Characteristics | Producingmicroorganisms | Hypothetical use in agriculture | References |
|---|---|---|---|---|
| Alkaloids | Nitrogen-containing heterocyclic compounds | Fischerella sp., Calothrix sp. | Natural bioinsecticides or antimicrobial agents for biocontrol | [115,116] |
| Polyketides | Structurally diverse metabolites derived from carboxylic acid precursors | Gambierdiscus toxicus, Karenia brevis | Broad-spectrum fungicides or bactericides for crops | [117,118] |
| Fatty acids | Extracellular free fatty acids with allelopathic activity | Chlorella vulgaris, Botryococcus braunii | Natural weed growth inhibitors (bioherbicides) | [119,120] |
| Peptides | Non-ribosomal peptides biosynthesized by multifunctional enzyme complexes |
Anabaena sp. PCC7120, Microcystis sp., Planktothrix sp., Oscillatoria limosa |
Plant defense promoters or biostimulants | [121] |
| Terpenoids | Organic compounds derived from C5 precursors with toxicity to invertebrates | Nostoc commune, Calothrix sp. PCC7507 | Natural insecticides or pest repellents | [122] |
Table 3.
Examples of some commercialized microalgae- and cyanobacteria-based products.
| Product name | Manufacturer | Microalgae used | Formulation | Main effects |
|---|---|---|---|---|
| Algafert | Biorizon Biotech | Spirulina spp. | Dry powder | Provides macro and micronutrients, promotes chlorophyll synthesis. |
| AgriAlgae | AlgaEnergy | Nannochloropsis spp. | Liquid biostimulant |
Enhances photosynthesis, nutrient uptake, and crop vigor. |
| Spiralagrow | GreenCoast | Arhtrospira spp. | Pellet, granulate |
Improves soil fertility and water retention. |
| Ecotop | Herogra | Ascophyllum nodosum and blend of other microalgae | Liquid biostimulant |
Enhances plant vigor, growth and resilience to abiotic/biotic stress. |
| Kelpak | Kelpak | Ecklonia maxima (macroalga, used in synergy with microalgae) | Liquid biostimulant |
Promotes root and shoot development, stress tolerance. |
| AlgaGrow | Agrinos | Proprietary blend including cyanobacteria | Liquid biostimulant |
Increases nutrient uptake and crop yield. |
| Seasol | Seasol International (Australia) | Blend of seaweed and microalgae extracts | Liquid concentrate |
Broad-spectrum plant tonic. |
| Weed-Max | Trade S.A.E. Company (Egypt) | Cyanobacteria extract in powder phase | Dry powder | Suppress soil-borne fungi and enhance the antagonistic abilities of other bioagents. |
| Oligo-X algal | Arabian Group for Agricultural Service | Blue-green algal extracts in liquid phase | Liquid concentrate |
Suppress soil-borne fungi. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



