Submitted:
27 June 2025
Posted:
27 June 2025
You are already at the latest version
Abstract
Glioblastoma multiforme (GBM) is one of the most aggressive cancers, with limited treatment options due to its immunosuppressive microenvironment. γδ T cells, known for their potent antitumor activity and ability to recognize tumor antigens independently of HLA molecules, have emerged as a promising therapeutic strategy. This review explores the role of γδ T cells in glioblastoma, focusing on their functional plasticity and potential to overcome immune evasion mechanisms. We also discuss potential combinatory approaches to enhance the therapeutic effectiveness of γδ T cell-based strategies in GBM treatment.
Keywords:
Gamma delta T lymphocytes
; glioblastoma
; immunotherapy
Introduction
GBM is the most common and lethal primary brain tumor, accounting for nearly 45% of all primary malignant brain tumors, with a median survival of less than one year, despite aggressive treatments such as surgical resection, radiotherapy, and chemotherapy (also known as Stupp’s treatment). It’s characterized by an incidence of approximately 3 per 100,000 people annually [1,2]. Despite advances in surgical techniques and the use of chemotherapeutic agents like temozolomide (TZM), the recurrence of GBM remains inevitable due to its heterogeneity and the lack of effective therapeutic strategies [3]. A key factor contributing to the poor prognosis of GBM is its immunosuppressive tumor microenvironment (TME), which prevents the immune system from mounting an effective antitumor response. Tumor-associated macrophages (TAMs), myeloid-derived suppressor cells (MDSCs), and regulatory T cells (Tregs) dominate the TME, creating a hostile environment that supports tumor growth while suppressing immune function [4]. The tumor’s highly infiltrative nature and resistance to conventional therapies contribute significantly to the challenges in achieving long-term survival for patients. Additionally, the blood-brain barrier (BBB) restricts the delivery of many therapeutic agents, further complicating treatment options [5]. This makes developing novel and effective immunotherapies a critical area of research in the fight against GBM. One promising avenue of immunotherapy involves immune checkpoint inhibitors (ICIs), which have revolutionized the treatment of several cancers, but their impact on GBM has been limited [6]. Several ICIs have been tested for their efficacy in GBM treatment, among which are those targeting PD-1 (Programmed Death-1) and PD-L1 (Programmed Death Ligand-1), as well as CTLA-4 (Cytotoxic T-Lymphocyte Antigen-4). Pembrolizumab (Keytruda), which targets PD-1, has been explored as a potential treatment for GBM, particularly when combined with other therapeutic approaches. Similarly, Nivolumab (Opdivo), another PD-1 inhibitor, has also been investigated in clinical trials for its efficacy on GBM, although the results have been mixed [7]. Atezolizumab (Tecentriq), which inhibits PD-L1, has been studied in various cancers, including brain tumors, and its effects on GBM have been closely examined [8]. Additionally, Ipilimumab (Yervoy), a CTLA-4 inhibitor primarily used in melanoma, has been tested for GBM treatment, often in combination with other ICIs such as nivolumab [9]. Despite the initial promise surrounding these treatments, the overall effectiveness of ICIs in treating GBM remains limited: although ICIs such as pembrolizumab, nivolumab, and ipilimumab have shown positive results in other types of cancer, their impact on GBM is less pronounced. This is mainly due to GBM's low mutational burden, which results in insufficient immune cell activation within the TME [10]. Furthermore, the presence of an immunosuppressive microenvironment, coupled with the low levels of immune infiltration typically found in GBM, reduces the efficacy of ICI therapies [11]. An emerging approach to GBM immunotherapy involves the use of γδ T cells, a subset of T lymphocytes with innate-like properties that make them particularly effective in recognizing and eliminating tumor cells. γδ T cells do not require the classical antigen presentation through major histocompatibility complex (MHC) molecules, which allows them to target a broader range of tumor antigens, including those expressed on GBM cells [12]. Recent studies suggest that γδ T cells can be activated in vivo or expanded ex vivo to enhance their antitumor activity against GBM [13]. These cells can directly kill tumor cells, modulate the immune response, and produce pro-inflammatory cytokines that can help overcome the immunosuppressive TME of GBM [14]. Furthermore, the ability of γδ T cells to interact with other immune cells, such as dendritic cells and natural killer (NK) cells, provides a unique advantage in promoting a more robust and coordinated immune response against the tumor [14]. γδ T cells can exhibit pro- and anti-tumoral effects depending on their activation state and the surrounding microenvironment and these cells are crucial in counteracting the immunosuppressive milieu created by TAMs, MDSCs, and Tregs, [12]. As a result, researchers are increasingly exploring γδ T cells as a potential therapeutic strategy, either as an adjunct to traditional treatments or in combination with other immunotherapies[14]. This review highlights the role and clinical efficacy using γδ T cells for GBM treatment. We will explore their role in the immune response to GBM, the mechanisms underlying their antitumor activity and the clinical studies investigating their potential in overcoming the challenges posed by the GBM microenvironment.
γδ T Cell Characteristics and Functions
γδ T cells represent a unique subset of CD3+ T lymphocytes, classified as "unconventional" T cells due to their distinct T cell receptor (TCR) structure. Unlike conventional αβ T cells, γδ T cells express a TCR composed of γ and δ chains and account for approximately 1-5% of the total peripheral T lymphocytes in the human blood [15]. γδ T cells bridge innate and adaptive immunity, providing rapid immune responses in collaboration with macrophages and neutrophils against various pathogens. Additionally, they assist in modulating the function of adaptive immune cells, including T cells and B cells [16]. Notably, γδ T cells express receptors typical of NK cells, such as NKG2D, CD94-NKG2A/C, and the natural cytotoxicity receptors (NCRs) NKp30 and NKp44[17], which enable them to target and kill infected, stressed, or neoplastic cells and to activate other immune effectors [16]. Most γδ T cells (around 70%) lack both CD4 and CD8 expression (CD4⁻CD8⁻), while about 30% are CD8⁺CD4⁻, and fewer than 1% are CD4⁺CD8⁻. They are categorized into three main subsets based on their γ and δ chain composition: Vδ1⁺, Vδ2⁺, and Vδ1⁻Vδ2⁻. The Vδ1+ T cells are predominantly localized in mucosal and epithelial tissues, where they exhibit cytotoxic activity through the recognition of stress-induced antigens expressed by transformed cells, such as MICA/MICB and UL16-binding proteins, via NKG2D [14] or NCRs [17]. The Vδ1 chain can associate with various γ chains, including Vγ2, Vγ3, Vγ4, Vγ5, Vγ8, and Vγ10, with Vγ4 showing a notable preferential pairing with Vδ1 in intestinal tissue. The most abundant subset of γδ T cells expresses the Vδ2 chain, commonly paired with Vγ9, forming what are known as Vγ9Vδ2 T cells. These cells account for 50–95% of the total γδ T lymphocyte population and are characterized by their ability to respond to phosphoantigens (PAgs). PAgs are intermediates of the mevalonate metabolic pathway, which is upregulated in tumor cells, or the exogenous 1-deoxy-D-xylulose-5-phosphate (MEP) pathway, which is enhanced during infections [18]. The mevalonate pathway, essential for the biosynthesis of cholesterol, sterols, steroid hormones, and cellular membranes, generates PAgs as intermediates, which serve as signals for γδ T cells to distinguish between normal and abnormal (tumor or infected) cells[19]. High levels PAgs indicate cellular stress or transformation, and promote γδ T cells activation to target and eliminate these altered cells [20]. PAgs-induced γδ T cell activation involves two butyrophilins, BTN2A1 and BTN3A1. The BTN2A1–BTN3A1 complex interacts with the γδ TCR at two distinct sites: BTN2A1 binds to the Vγ9 domain, while BTN3A1 binds to the Vδ2 regions and the γ chains of the TCR [21]. Once activated, γδ T cells target infected, stressed, or tumor cells using mechanisms such as cytotoxicity via perforin and granzyme secretion, antibody-dependent cellular cytotoxicity (ADCC) and the release of pro-inflammatory cytokines like TNF-α and IFN-γ. These mechanisms are also employed by NK and CD8+ T cells to eliminate abnormal cells [22]. Human γδ T cells, particularly the Vγ9Vδ2 subset, exhibit significant phenotypic and functional heterogeneity, allowing them to adopt various differentiation states that influence their responsiveness and antitumor potential and that, in turn, can be influenced by the TME [23]. Understanding this diversity is crucial for the development of γδ T cell-based immunotherapies, as the differentiation state significantly impacts their cytotoxic potential, tissue distribution, and persistence within the TME [24].
Human γδ T Cells in GBM
γδ T cells are involved in the immune response against tumors [25]. These cells are well known for their ability to recognize and target tumor cells in various malignancies, including GBM [14,26]. In GBM, γδ T cells are recruited into the TME, where they can interact with tumor cells and other immune cells, including macrophages and dendritic cells[23]. These cells have been found to infiltrate GBM tissues in mouse models [14,27] and human patients [14,23], with their infiltration correlating with enhanced survival and tumor regression. Moreover, studies have demonstrated that γδ T cells, particularly those expressing the Vγ9Vδ2 TCR, play a key role in immune surveillance in GBM, exhibiting both cytotoxic activity and the capacity to produce pro-inflammatory cytokines that can enhance the immune response to tumor cells [28]. It was observed that Vγ9Vδ2 γδ T cells exhibit potent cytotoxic activity against human glioma cell lines and GBM tumor cells, suggesting their potential for therapeutic applications in GBM [27,29]. However, despite these promising findings, the role of γδ T cells in GBM remains complex. Some studies suggest that these cells may enhance antitumor immunity, while others have indicated that the immunosuppressive TME in GBM can impair their effectiveness [30]. This is partly due to the functional plasticity exhibited by γδ T cells, allowing them to differentiate into different subsets based on the microenvironmental signals present within the tumor such as the cytokine milieu and interactions with other immune and non-immune cells [4,31].
Anti-Tumoral Role of γδ T Cells in GBM
γδ T cells have been shown to play a critical anti-tumoral role in GBM. An analysis of the Cancer Genome Atlas (TCGA) database has shown that γδ T cell activity positively correlates with CD8+ cytotoxic T cell activity and M1 macrophage presence, highlighting their potential to stimulate the immune response against GBM [32]. One of the most well-characterized mechanisms by which γδ T cells mediate anti-tumor responses is their ability to recognize and kill tumor cells directly. γδ T cells express various activating receptors, such as NKG2D, which can bind to stress-induced ligands (e.g., MICA/B, ULBP) expressed on tumor cells. This interaction triggers the release of perforin and granzymes, which mediate the cytotoxicity of γδ T cells against GBM cells. Additionally, γδ T cells produce pro-inflammatory cytokines such as IFN-γ and TNF-α, which enhance their anti-tumor activity by promoting the activation of other immune cells, including CD8+ T cells and NK cells, involved in the anti-tumor immune response [12]. Moreover, γδ T cells can infiltrate GBM tumors and persist in the microenvironment, where they continue to exert their cytotoxic effects. In preclinical models, the adoptive transfer of γδ T cells has been shown to enhance survival and reduce tumor burden in GBM-bearing mice, further supporting their potential as an effective therapeutic option [33]. Importantly, γδ T cells can target tumor cells independently of MHC restriction, giving them an advantage over other immune cells that rely on antigen presentation for activation. This feature is essential in the context of GBM, where tumor cells often downregulate MHC molecules to evade recognition by conventional T cells [34]. Additionally, combining γδ T cell-based therapies with standard treatments such as TMZ has demonstrated synergistic effects, enhancing the therapeutic response and overcoming resistance mechanisms commonly observed in GBM treatment [30]. Recent studies have focused on using γδ T cells in combination with ICIs, such as PD-1/PD-L1 blockers, to further enhance their antitumor effects. These studies have shown that the blockade of immune checkpoint pathways can reactivate γδ T cells within the TME, promoting their anti-tumor activity and improving overall survival rates in animal models of GBM. Furthermore, γδ T cells exhibit a unique capacity for tissue infiltration, including the ability to cross the BBB, which makes them an attractive candidate for targeting CNS tumors like GBM. Taken together, these findings highlight the potential of γδ T cells as both a direct and indirect immune effector against GBM, and underscore the importance of advancing γδ T cell-based immunotherapies for GBM patients [25].
Pro-Tumoral Role of γδ T Cells in GBM
While γδ T cells are generally acknowledged for their role in mediating antitumor immunity, recent studies have shown that they can contribute to tumor progression in specific contexts, particularly in the immunosuppressive microenvironment of GBM. The GBM microenvironment is highly hostile to immune cells, characterized by the presence of various cytokines, including IL-1β, IL-6, TGF-β, and IL-23 [35]. These factors can significantly alter the functional behavior of γδ T cells, driving them to adopt a pro-tumoral, type-3 (Th17-like) phenotype. Although Vγ9Vδ2 T cells are known for their cytotoxic function, they display a dual role. High concentrations of TGF-β divert cells toward pro-tumor subtypes due to the downregulation of NKG2D and the expression of genes encoding granzymes and perforins [14]. Under such conditions, γδ T cells can differentiate into IL-17-producing cells [36] that support tumor progression; the production of IL-17 by these cells has been associated with the recruitment of immunosuppressive cells, including MDSCs and Tregs, which can dampen the efficacy of other immune responses within the tumor [28] . Furthermore, IL-17 can induce angiogenesis by stimulating endothelial cell proliferation and the secretion of pro-angiogenic factors, thereby facilitating the formation of new blood vessels that support tumor growth and metastasis. Another critical aspect of the pro-tumoral role of γδ T cells in GBM is their potential to differentiate into γδ Tregs, a subset of regulatory T cells that contribute to the suppression of immune responses in the TME. γδ Tregs exert their immunosuppressive effects by inhibiting dendritic cell maturation and impairing the activation of cytotoxic T cells and NK cells. This suppressive action further weakens the overall anti-tumor immune response and gives the tumor an advantage in evading immune surveillance. Furthermore, γδ T cells may be recruited to GBM lesions, where they secrete various cytokines that suppress the function of other immune cell populations, potentially aiding in tumor evasion [14] . This dual nature of γδ T cells highlights the complexity of their involvement in GBM and emphasizes the need for further research to clarify their precise role in tumor progression.
γδ T Cells in Cancer Immunotherapy
In recent years, immunotherapy has revolutionized cancer treatment by enhancing the immune system’s ability to eliminate malignant cells. Most immunotherapeutic approaches rely on αβ T lymphocytes, which, although highly specific, are limited by their dependence on MHC restriction, co-stimulatory signals, and recognition of tumor-associated antigens (TAAs) [37]. Many tumors, however, can escape αβ T cell-mediated immune surveillance by downregulating MHC class I molecules or losing antigen expression, thereby limiting the effectiveness of αβ T cell-based therapies [38,39]. γδ T cells provide a promising alternative due to their unique biology. Unlike αβ T cells, γδ T cells recognize stress-induced or transformation-associated antigens in an MHC-independent manner and do not require classical co-stimulation for activation [40]. This allows them to identify and destroy tumor cells that would otherwise evade conventional immune responses. Moreover, γδ T cells exhibit a rapid and potent effector response, including direct cytotoxicity through perforin and granzyme release, production of pro-inflammatory cytokines such as IFN-γ and TNF-α, and modulation of the tumor microenvironment through interactions with dendritic cells, NK cells, and other immune effectors [41,42].The most well-characterized γδ T cell subset in the context of cancer is the Vγ9Vδ2 population, which predominates in peripheral blood. These cells can be activated using PAgs - either naturally derived (e.g., isopentenyl pyrophosphate (IPP)) or synthetic analogs (e.g., BrHPP) - or pharmacologically through nitrogen-containing bisphosphonates (N-BPs), such as zoledronic acid (ZOL) [43]. N-BPs inhibit farnesyl pyrophosphate synthase in the mevalonate pathway, leading to intracellular PAgs accumulation and potent activation of Vγ9Vδ2 T cells [44]. Two main approaches have emerged in the past: in vivo activation using N-BPs and IL-2, and adoptive transfer of ex vivo expanded Vγ9Vδ2 T cells. While in vivo stimulation is less complex from a clinical standpoint, its efficacy may be hindered by transient responses or induction of functional exhaustion following repeated stimulation [45]. In contrast, ex vivo expansion allows more precise control over activation, phenotype and dosing of the infused γδ T cells [46]. Additionally, γδ T cells are genetically engineered to express chimeric antigen receptors (CARs), combining their natural cytotoxic properties with antigen-specific targeting. These CAR-γδ T cells have shown promise in preclinical studies for both hematologic and solid tumors [47,48]. Although Vγ9Vδ2 T cells are widely used in clinical settings due to their abundance and expandability, Vδ1+ γδ T cells are gaining attention. These cells are enriched in epithelial and mucosal tissues, exhibit a strong cytotoxic capacity, and are more resistant to activation-induced cell death, making them ideal candidates for durable antitumor responses [49]. However, the lack of standardized protocols for their isolation and expansion has historically limited their clinical application [50]. Recent developments have enabled the efficient expansion of Vδ1+ cells with potent cytotoxic profiles, making them suitable for adoptive therapy [51].Furthermore, novel strategies are being explored to engage γδ T cells more selectively and effectively. These include bispecific γδ T cell engagers that simultaneously bind to tumor antigens and γδ TCR chains, thereby redirecting γδ T cell cytotoxicity toward tumor cells [52]. For instance, clinical trials investigating bispecific antibodies such as LAVA-051 and LAVA-1207 are currently underway in hematologic and solid tumors (NCT05369000). Adoptive transfer of ex vivo expanded γδ T cells has shown promising efficacy in preclinical models of GBM, melanoma, and other malignancies, with robust tumor infiltration and cytotoxicity (NCT04165941). However, challenges remain, including variability in patient responses and the need for optimal dosing strategies. Other companies are developing allogeneic CAR-γδ T cells, leveraging their lower risk of graft-versus-host disease and reduced incidence of cytokine release syndrome compared to αβ CAR-T cells [48]. While αβ T cell-based therapies, such as CAR-αβ T cells, have achieved remarkable success in hematologic malignancies, their efficacy in solid tumors is limited by antigen heterogeneity and the immunosuppressive TME [39,47]. ICIs targeting PD-1/PD-L1 or CTLA-4 have revolutionized treatment for certain cancers but rely on pre-existing tumor-specific T cell responses, which may be absent in immunologically “cold” tumors [37]. In contrast, γδ T cells’ ability to target stress-induced antigens without MHC restriction makes them an attractive option for overcoming these limitations [40]. Furthermore, the genetic engineering of CAR-γδ T cells combines innate and adaptive immune advantages, potentially addressing antigen escape mechanisms more effectively than conventional CAR-T cells [41].
Despite their promise, γδ T cell-based therapies face several hurdles:
- Exhaustion: Repeated stimulation or chronic activation can lead to functional exhaustion, which reduces efficacy over time [53].
- Off-Target Effects: The MHC-independent recognition of antigens raises concerns about potential off-tumor cytotoxicity [54].
- Tumor Heterogeneity: Variability in the expression of stress-induced antigens across tumors may limit the applicability of γδ T cell therapies [49].
- Expansion Variability: Ex vivo expansion protocols often yield inconsistent results, which can impact the scalability and reproducibility of therapies [50].
Addressing these challenges requires further optimization of expansion protocols, improved understanding of γδ T cell biology, and the development of strategies to mitigate exhaustion and off-target effects. In fact, several biotech companies are at the forefront of advancing γδ T cell therapies. Lava Therapeutics is developing bispecific γδ T cell engagers, with LAVA-051 and LAVA-1207 in clinical trials (NCT05369000). Adicet Bio is exploring allogeneic CAR-γδ T cells to minimize graft-versus-host disease (NCT04735471). IN8bio focuses on utilizing γδ T cells for the treatment of solid tumors, with promising early-phase trial results (NCT04165941). GammaDelta Therapeutics is pioneering methods to enhance the expansion and efficacy of Vδ1+ γδ T cells [51]. These companies’ innovations highlight the growing interest and investment in γδ T cell-based therapies. Several other biotech firms are making significant strides in advancing γδ T cell therapies. TC BioPharm is conducting two investigator-initiated clinical trials for its unmodified γδ T cell product line, including a Phase 2b/3 pivotal trial in the treatment of acute myeloid leukemia using the company's proprietary allogeneic CryoTC technology (NCT04735471). CytoMed Therapeutics has initiated the ANGELICA Trial (NCT05653271), a first-in-human Phase I clinical trial evaluating allogeneic NKG2DL-targeting CAR-gd T cells (CTM-N2D) in patients with advanced solid tumors or hematological malignancies. Acepodia is developing ACE1831, an off-the-shelf γδ T-cell therapy currently undergoing a Phase I clinical trial (NCT05653271) for the treatment of non-Hodgkin's lymphoma. Additionally, Expression Therapeutics, is developing a γδ T cell therapy that, when used in conjunction with standard chemotherapy and immunotherapy, aims to effectively eradicate neuroblastoma tumor cells. These efforts reflect the wide range of strategies aimed at fully leveraging the therapeutic potential of γδ T cells.
γδ T Cells in GBM Immunotherapy
GBM continues to pose a significant therapeutic challenge, with standard treatments such as surgery, radiotherapy, and chemotherapy offering limited survival benefits [55,56]. In this context, γδ T cells have emerged as a promising immunotherapeutic strategy due to their unique properties, including MHC-unrestricted target cell recognition and an immediate response to stress-induced ligands expressed by tumor cells [57,58]. Among the γδ T cell subsets, Vγ9Vδ2 T cells have demonstrated notable antitumor activity against GBM through mechanisms such as perforin/granzyme-mediated lysis and NKG2D-dependent recognition of ligands including ULBP and MIC[13,59]. These ligands are upregulated on tumor cells under stress conditions and are further enhanced by treatments including TMZ and ZOL. ZOL plays a dual role in GBM immunotherapy by inducing the accumulation of isopentenyl-pyrophosphate, a ligand for Vγ9Vδ2 T cells, while also sensitizing glioblastoma cells to immune attack. and by increasing the surface expression of NKG2D ligands, rendering glioblastoma cells more susceptible to γδ T cell-mediated cytotoxicity [60,61]. However, the toxic effects of TMZ on γδ T cells themselves necessitate innovative solutions such as genetic engineering to confer resistance. For instance, γδ T cells modified with lentiviral vectors encoding O6-methylguanine-DNA-methyltransferase (MGMT) retain full functionality in the presence of TMZ, allowing for their concurrent use with chemotherapeutic regimens [62]. Ex vivo expansion of γδ T cells from peripheral blood has proven reliable to generate large numbers of functional cells capable of targeting GBM. These expanded cells have demonstrated significant cytotoxicity against autologous tumor cells while sparing normal tissues, thereby showcasing their safety profile [56,63]. Preclinical models have further validated the efficacy of γδ T cells, with intracranial infusions slowing tumor progression and extending survival in xenograft models [32]. Moreover, combinations of cytokines such as IL-2, IL-12, and IL-15 have synergistically enhanced the proliferation and cytotoxic activity of γδ T cells, offering additional avenues for therapeutic optimization [55,57,58]. Despite their promise, γδ T cells—like other cellular therapies—must overcome the profoundly immunosuppressive TME of GBM, which is characterized by Tregs, MDSCs and TAMs that secrete inhibitory cytokines such as IL-10 and TGF-β [64]. A notable example of the clinical translation of genetically modified γδ T cells is the INB-200 Phase 1 clinical trial, currently underway and sponsored by IN8bio. This study evaluates the safety, tolerability, and potential efficacy of intracranial infusion of autologous, ex vivo expanded γδ T cells in patients with newly diagnosed or recurrent GBM, in combination with TMZ. The cells are administered post-tumor resection alongside standard TMZ therapy, leveraging the drug’s tumor-sensitizing effects without impairing the function of the infused γδ T cells. Unlike conventional CAR-T approaches, this strategy exploits the innate antitumor properties of γδ T cells, which are enhanced through expansion and genetic manipulation, while maintaining a favorable safety profile. Preliminary data released by IN8bio indicate that the infusions are well tolerated, with no treatment-related serious adverse events and early signs of antitumor activity. This trial represents one of the first clinical efforts to validate γδ T cells as a viable form of cellular immunotherapy for GBM, laying the groundwork for future large-scale, controlled studies (NCT04165941, NCT03533816). Additionally, the integration of γδ T cells into combinatorial immunotherapy regimens—including checkpoint inhibitors, NK cell therapies, and dendritic cell vaccines—is under investigation, as synergistic approaches may help reverse TME-mediated immune suppression and improve treatment outcomes [64]. Despite these advances, the immunosuppressive TME of GBM remains a significant barrier. This microenvironment, characterized by inhibitory cytokines and a lack of effective lymphocyte infiltration, limits the full potential of γδ T cell-based therapies [65]. Innovative strategies are being explored to overcome these challenges, including the use of BTN3A1 agonistic monoclonal antibodies and developing CAR γδ T cells targeting GBM-associated antigens. CAR-engineered γδ T cells have shown remarkable preclinical efficacy, highlighting their ability to enhance tumor recognition and destruction [66].
Furthermore, studies have underscored the versatility of γδ T cells in adapting to the complex GBM microenvironment. For instance, γδ T cells co-cultured with GBM cells exhibit a Th1-like profile characterized by increased IFN-γ and TNF-α production, suggesting their ability to reshape the immune landscape within tumors [67]. The infiltration of unique γδ T cell subsets, such as Vγ9Jγ2-Vδ2 T cells, into GBM tissues further emphasizes their tumor-specific adaptations. These cells, distinct from their peripheral counterparts, demonstrate robust antitumor responses, offering insights into their potential for precision immunotherapy [68]. Overall, γδ T cells represent a powerful tool in the fight against GBM, combining intrinsic cytotoxicity with innovative engineering approaches to address the challenges posed by this formidable tumor. As research continues to refine these therapies, γδ T cells are poised to play a central role in developing next-generation immunotherapies for GBM, offering hope for improved outcomes in this devastating disease.
Conclusions
GBM is a highly aggressive and complex brain tumor, characterized by an immunosuppressive microenvironment that limits the efficacy of conventional therapies and contributes to its highly unfavorable prognosis. Immunotherapy represents a promising approach to overcome these limitations, and γδ T cells, with their ability to recognize tumor cells independently of MHC, are emerging as an interesting strategy. However, the application of immunotherapy in GBM presents unique challenges, related to the complexity of the TME and the need to overcome the blood-brain barrier. γδ T cells offer potential to address these challenges, owing to their intrinsic cytotoxicity and the capacity to modulate the immune response. Research focuses on developing strategies to enhance their antitumor activity and the capacity to infiltrate the tumor. Despite promising preclinical results and early-phase clinical trials, the clinical translation of γδ T cell-based immunotherapy for GBM is still in its early stages. Further studies are necessary to optimize therapeutic strategies, fully understand the role of these cells in the context of GBM, and develop novel technologies, such as CAR-γδ T cells and bispecific constructs, to improve their efficacy and specificity [74]. In conclusion, γδ T cell immunotherapy offers a concrete prospect for treating GBM, but its full potential requires further investigation and innovation to overcome the challenges posed by this devastating tumor.
Author Contributions
All authors designed the entire review, wrote the paper, and reviewed and edited the manuscript. All authors contributed to the article and approved the submitted version.
Funding
The research leading to these results has received funding from the European Union - NextGenerationEU through the Italian Ministry of Health under PNRR – M6C2-I2.1, Project Code PNRR-TR1-2023-12377714 “Multidisciplinary and multiomic approach to dissect the cellular network in the glioma microenvironment: translational perspective to improve patient's management” to Francesco Dieli, CUP I73C24000220006. The views and opinions expressed are those of the authors only and do not necessarily reflect those of the European Union or the European Commission. Neither the European Union nor the European Commission can be held responsible for them. .
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest. Francesco Dieli, is an Editor of Cells for section “Cellular Immunology”. This does not alter the authors’ adherence to all the MDPI policies on sharing data and materials. .
Abbreviations
The following abbreviations are used in this manuscript:
| GBM | Glioblastoma Multiforme |
| TMZ | Temozolomide |
| TME | Tumor Microenvironment |
| TAMs | Tumor-associated macrophages |
| MDSCs | Myeloid-derived suppressor cells |
| Tregs | Regulatory T cells |
| BBB | Blood-brain barrier |
| ICIs | Immune checkpoint inhibitors |
| PD-1 | Programmed Death-1 |
| PD-L1 | Programmed Death Ligand-1 |
| CTLA-4 | Cytotoxic T-Lymphocyte Antigen-4 |
| MHC | Major histocompatibility complex |
| NK | Natural killer |
| TCR | T cell receptor |
| NCRs | Natural cytotoxicity receptors |
| PAgs | Phosphoantigens |
| MEP | 1-deoxy-D-xylulose-5-phosphate |
| ADCC | Antibody-dependent cellular cytotoxicity |
| TCGA | The Cancer Genome Atlas |
| TAAs | Tumor-associated antigens |
| N-BPs | Nitrogen-containing bisphosphonates |
| ZOL | Zoledronic acid |
| CARs | Chimeric antigen receptors |
| MGMT | O6-methylguanine-DNA-methyltransferase |
| EGFR | Epidermal Growth Factor |
| N/A | Not Available |
| AE | Adverse Events |
| ROA | Administration Route |
| IC | Intracranial |
| ST-IC | Stereotactic Intracranial |
| IP | Intraperitoneal |
| IT | Intra-tumoral |
| ICV | Intraventricular |
References
- Di Carlo, D.T.; Cagnazzo, F.; Benedetto, N.; Morganti, R.; Perrini, P. Multiple High-Grade Gliomas: Epidemiology, Management, and Outcome. A Systematic Review and Meta-Analysis. Neurosurg Rev 2019, 42, 263–275. [CrossRef]
- Stupp, R.; Mason, W.P.; Van Den Bent, M.J.; Weller, M.; Fisher, B.; Taphoorn, M.J.B.; Belanger, K.; Brandes, A.A.; Marosi, C.; Bogdahn, U.; et al. Radiotherapy plus Concomitant and Adjuvant Temozolomide for Glioblastoma. N Engl J Med 2005, 352, 987–996. [CrossRef]
- Weller, M.; Van Den Bent, M.; Tonn, J.C.; Stupp, R.; Preusser, M.; Cohen-Jonathan-Moyal, E.; Henriksson, R.; Rhun, E.L.; Balana, C.; Chinot, O.; et al. European Association for Neuro-Oncology (EANO) Guideline on the Diagnosis and Treatment of Adult Astrocytic and Oligodendroglial Gliomas. The Lancet Oncology 2017, 18, e315–e329. [CrossRef]
- Lin, H.; Liu, C.; Hu, A.; Zhang, D.; Yang, H.; Mao, Y. Understanding the Immunosuppressive Microenvironment of Glioma: Mechanistic Insights and Clinical Perspectives. J Hematol Oncol 2024, 17, 31. [CrossRef]
- Tang, W.; Fan, W.; Lau, J.; Deng, L.; Shen, Z.; Chen, X. Emerging Blood–Brain-Barrier-Crossing Nanotechnology for Brain Cancer Theranostics. Chem. Soc. Rev. 2019, 48, 2967–3014. [CrossRef]
- Naimi, A.; Mohammed, R.N.; Raji, A.; Chupradit, S.; Yumashev, A.V.; Suksatan, W.; Shalaby, M.N.; Thangavelu, L.; Kamrava, S.; Shomali, N.; et al. Tumor Immunotherapies by Immune Checkpoint Inhibitors (ICIs); the Pros and Cons. Cell Commun Signal 2022, 20, 44. [CrossRef]
- Yang, T.; Kong, Z.; Ma, W. PD-1/PD-L1 Immune Checkpoint Inhibitors in Glioblastoma: Clinical Studies, Challenges and Potential. Hum Vaccin Immunother 2021, 17, 546–553. [CrossRef]
- Lukas, R.V.; Rodon, J.; Becker, K.; Wong, E.T.; Shih, K.; Touat, M.; Fassò, M.; Osborne, S.; Molinero, L.; O’Hear, C.; et al. Clinical Activity and Safety of Atezolizumab in Patients with Recurrent Glioblastoma. J Neurooncol 2018, 140, 317–328. [CrossRef]
- Omuro, A.; Vlahovic, G.; Lim, M.; Sahebjam, S.; Baehring, J.; Cloughesy, T.; Voloschin, A.; Ramkissoon, S.H.; Ligon, K.L.; Latek, R.; et al. Nivolumab with or without Ipilimumab in Patients with Recurrent Glioblastoma: Results from Exploratory Phase I Cohorts of CheckMate 143. Neuro Oncol 2018, 20, 674–686. [CrossRef]
- Yu, M.W.; Quail, D.F. Immunotherapy for Glioblastoma: Current Progress and Challenges. Front. Immunol. 2021, 12, 676301. [CrossRef]
- Wang, X.; Guo, G.; Guan, H.; Yu, Y.; Lu, J.; Yu, J. Challenges and Potential of PD-1/PD-L1 Checkpoint Blockade Immunotherapy for Glioblastoma. J Exp Clin Cancer Res 2019, 38, 87. [CrossRef]
- Ahmedna, T.; Khela, H.; Weber-Levine, C.; Azad, T.D.; Jackson, C.M.; Gabrielson, K.; Bettegowda, C.; Rincon-Torroella, J. The Role of Γδ T-Lymphocytes in Glioblastoma: Current Trends and Future Directions. Cancers 2023, 15, 5784. [CrossRef]
- Bryant, N.L.; Gillespie, G.Y.; Lopez, R.D.; Markert, J.M.; Cloud, G.A.; Langford, C.P.; Arnouk, H.; Su, Y.; Haines, H.L.; Suarez-Cuervo, C.; et al. Preclinical Evaluation of Ex Vivo Expanded/Activated Γδ T Cells for Immunotherapy of Glioblastoma Multiforme. J Neurooncol 2011, 101, 179–188. [CrossRef]
- Kang, I.; Kim, Y.; Lee, H.K. Γδ T Cells as a Potential Therapeutic Agent for Glioblastoma. Front. Immunol. 2023, 14, 1273986. [CrossRef]
- Fonseca, S.; Pereira, V.; Lau, C.; Teixeira, M.D.A.; Bini-Antunes, M.; Lima, M. Human Peripheral Blood Gamma Delta T Cells: Report on a Series of Healthy Caucasian Portuguese Adults and Comprehensive Review of the Literature. Cells 2020, 9, 729. [CrossRef]
- Park, J.H.; Lee, H.K. Function of Γδ T Cells in Tumor Immunology and Their Application to Cancer Therapy. Exp Mol Med 2021, 53, 318–327. [CrossRef]
- Hudspeth, K.; Silva-Santos, B.; Mavilio, D. Natural Cytotoxicity Receptors: Broader Expression Patterns and Functions in Innate and Adaptive Immune Cells. Front. Immunol. 2013, 4. [CrossRef]
- Göbel, A.; Rauner, M.; Hofbauer, L.C.; Rachner, T.D. Cholesterol and beyond - The Role of the Mevalonate Pathway in Cancer Biology. Biochimica et Biophysica Acta (BBA) - Reviews on Cancer 2020, 1873, 188351. [CrossRef]
- Sireci, G.; Espinosa, E.; Sano, C.D.; Dieli, F.; Fournié, J.-J.; Salerno, A. Differential Activation of Human γ δ Cells by Nonpeptide Phosphoantigens. Eur. J. Immunol. 2001, 31, 1628–1635. [CrossRef]
- Cieslak, S.G.; Shahbazi, R. Gamma Delta T Cells and Their Immunotherapeutic Potential in Cancer. Biomark Res 2025, 13, 51. [CrossRef]
- Yuan, L.; Ma, X.; Yang, Y.; Qu, Y.; Li, X.; Zhu, X.; Ma, W.; Duan, J.; Xue, J.; Yang, H.; et al. Phosphoantigens Glue Butyrophilin 3A1 and 2A1 to Activate Vγ9Vδ2 T Cells. Nature 2023, 621, 840–848. [CrossRef]
- Hu, Y.; Hu, Q.; Li, Y.; Lu, L.; Xiang, Z.; Yin, Z.; Kabelitz, D.; Wu, Y. Γδ T Cells: Origin and Fate, Subsets, Diseases and Immunotherapy. Sig Transduct Target Ther 2023, 8, 434. [CrossRef]
- Lo Presti, E.; Pizzolato, G.; Corsale, A.M.; Caccamo, N.; Sireci, G.; Dieli, F.; Meraviglia, S. Γδ T Cells and Tumor Microenvironment: From Immunosurveillance to Tumor Evasion. Front Immunol 2018, 9, 1395. [CrossRef]
- Dieli, F.; Poccia, F.; Lipp, M.; Sireci, G.; Caccamo, N.; Di Sano, C.; Salerno, A. Differentiation of Effector/Memory Vδ2 T Cells and Migratory Routes in Lymph Nodes or Inflammatory Sites. The Journal of Experimental Medicine 2003, 198, 391–397. [CrossRef]
- Zhang, X.; Xu, Y.; Chen, H.; Zhang, J.; He, W. γδT Cells: Alternative Treasure in Antitumor Immunity. Explor Immunol 2022, 32–47. [CrossRef]
- La Manna, M.P.; Di Liberto, D.; Lo Pizzo, M.; Mohammadnezhad, L.; Shekarkar Azgomi, M.; Salamone, V.; Cancila, V.; Vacca, D.; Dieli, C.; Maugeri, R.; et al. The Abundance of Tumor-Infiltrating CD8+ Tissue Resident Memory T Lymphocytes Correlates with Patient Survival in Glioblastoma. Biomedicines 2022, 10, 2454. [CrossRef]
- Meraviglia, S.; Eberl, M.; Vermijlen, D.; Todaro, M.; Buccheri, S.; Cicero, G.; La Mendola, C.; Guggino, G.; D’Asaro, M.; Orlando, V.; et al. In Vivo Manipulation of Vgamma9Vdelta2 T Cells with Zoledronate and Low-Dose Interleukin-2 for Immunotherapy of Advanced Breast Cancer Patients. Clin Exp Immunol 2010, 161, 290–297. [CrossRef]
- Giannotta, C.; Autino, F.; Massaia, M. Vγ9Vδ2 T-Cell Immunotherapy in Blood Cancers: Ready for Prime Time? Front. Immunol. 2023, 14, 1167443. [CrossRef]
- Liu, Y.; Zhang, C. The Role of Human Γδ T Cells in Anti-Tumor Immunity and Their Potential for Cancer Immunotherapy. Cells 2020, 9, 1206. [CrossRef]
- Lafont, V.; Sanchez, F.; Laprevotte, E.; Michaud, H.-A.; Gros, L.; Eliaou, J.-F.; Bonnefoy, N. Plasticity of Î3δ T Cells: Impact on the Anti-Tumor Response. Front. Immunol. 2014, 5. [CrossRef]
- Yuan, M.; Wang, W.; Hawes, I.; Han, J.; Yao, Z.; Bertaina, A. Advancements in γδT Cell Engineering: Paving the Way for Enhanced Cancer Immunotherapy. Front. Immunol. 2024, 15, 1360237. [CrossRef]
- Lee, M.; Park, C.; Woo, J.; Kim, J.; Kho, I.; Nam, D.-H.; Park, W.-Y.; Kim, Y.-S.; Kong, D.-S.; Lee, H.W.; et al. Preferential Infiltration of Unique Vγ9Jγ2-Vδ2 T Cells Into Glioblastoma Multiforme. Front. Immunol. 2019, 10, 555. [CrossRef]
- Lamb, L.S.; Pereboeva, L.; Youngblood, S.; Gillespie, G.Y.; Nabors, L.B.; Markert, J.M.; Dasgupta, A.; Langford, C.; Spencer, H.T. A Combined Treatment Regimen of MGMT-Modified Γδ T Cells and Temozolomide Chemotherapy Is Effective against Primary High Grade Gliomas. Sci Rep 2021, 11, 21133. [CrossRef]
- Zagzag, D.; Salnikow, K.; Chiriboga, L.; Yee, H.; Lan, L.; Ali, M.A.; Garcia, R.; Demaria, S.; Newcomb, E.W. Downregulation of Major Histocompatibility Complex Antigens in Invading Glioma Cells: Stealth Invasion of the Brain. Lab Invest 2005, 85, 328–341. [CrossRef]
- Łaszczych, D.; Czernicka, A.; Gostomczyk, K.; Szylberg, Ł.; Borowczak, J. The Role of IL-17 in the Pathogenesis and Treatment of Glioblastoma—an Update on the State of the Art and Future Perspectives. Med Oncol 2024, 41, 187. [CrossRef]
- Caccamo, N.; La Mendola, C.; Orlando, V.; Meraviglia, S.; Todaro, M.; Stassi, G.; Sireci, G.; Fournié, J.J.; Dieli, F. Differentiation, Phenotype, and Function of Interleukin-17-Producing Human Vγ9Vδ2 T Cells. Blood 2011, 118, 129–138. [CrossRef]
- Schumacher, T.N.; Schreiber, R.D. Neoantigens in Cancer Immunotherapy. Science 2015, 348, 69–74. [CrossRef]
- Chang, C.-H.; Qiu, J.; O’Sullivan, D.; Buck, M.D.; Noguchi, T.; Curtis, J.D.; Chen, Q.; Gindin, M.; Gubin, M.M.; van der Windt, G.J.W.; et al. Metabolic Competition in the Tumor Microenvironment Is a Driver of Cancer Progression. Cell 2015, 162, 1229–1241. [CrossRef]
- Ribas, A.; Wolchok, J.D. Cancer Immunotherapy Using Checkpoint Blockade. Science 2018, 359, 1350–1355. [CrossRef]
- Hayday, A.C. Γδ T Cells and the Lymphoid Stress-Surveillance Response. Immunity 2009, 31, 184–196. [CrossRef]
- Rei, M.; Gonçalves-Sousa, N.; Lança, T.; Thompson, R.G.; Mensurado, S.; Balkwill, F.R.; Kulbe, H.; Pennington, D.J.; Silva-Santos, B. Murine CD27(-) Vγ6(+) Γδ T Cells Producing IL-17A Promote Ovarian Cancer Growth via Mobilization of Protumor Small Peritoneal Macrophages. Proc Natl Acad Sci U S A 2014, 111, E3562-3570. [CrossRef]
- Silva-Santos, B.; Serre, K.; Norell, H. Γδ T Cells in Cancer. Nat Rev Immunol 2015, 15, 683–691. [CrossRef]
- Kunzmann, V.; Bauer, E.; Feurle, J.; Tony, F.W., Hans-Peter; Wilhelm, M. Stimulation of Γδ T Cells by Aminobisphosphonates and Induction of Antiplasma Cell Activity in Multiple Myeloma. Blood 2000, 96, 384–392. [CrossRef]
- Roelofs, A.J.; Thompson, K.; Gordon, S.; Rogers, M.J. Molecular Mechanisms of Action of Bisphosphonates: Current Status. Clinical Cancer Research 2006, 12, 6222s–6230s. [CrossRef]
- Zhang, P.; Zhang, G.; Wan, X. Challenges and New Technologies in Adoptive Cell Therapy. J Hematol Oncol 2023, 16, 97. [CrossRef]
- Boucher, J.C.; Yu, B.; Li, G.; Shrestha, B.; Sallman, D.; Landin, A.M.; Cox, C.; Karyampudi, K.; Anasetti, C.; Davila, M.L.; et al. Large Scale Ex Vivo Expansion of Γδ T Cells Using Artificial Antigen-Presenting Cells. Journal of Immunotherapy 2023, 46, 5–13. [CrossRef]
- Fisher, J.P.; Heuijerjans, J.; Yan, M.; Gustafsson, K.; Anderson, J. Γδ T Cells for Cancer Immunotherapy: A Systematic Review of Clinical Trials. OncoImmunology 2014, 3, e27572. [CrossRef]
- Capsomidis, A.; Benthall, G.; Van Acker, H.H.; Fisher, J.; Kramer, A.M.; Abeln, Z.; Majani, Y.; Gileadi, T.; Wallace, R.; Gustafsson, K.; et al. Chimeric Antigen Receptor-Engineered Human Gamma Delta T Cells: Enhanced Cytotoxicity with Retention of Cross Presentation. Molecular Therapy 2018, 26, 354–365. [CrossRef]
- Siegers, G.M.; Lamb, L.S. Cytotoxic and Regulatory Properties of Circulating Vδ1+ Γδ T Cells: A New Player on the Cell Therapy Field? Molecular Therapy 2014, 22, 1416–1422. [CrossRef]
- Hayday, A.; Dechanet-Merville, J.; Rossjohn, J.; Silva-Santos, B. Cancer Immunotherapy by Γδ T Cells. Science 2024, 386, eabq7248. [CrossRef]
- Correia, D.V.; Lopes, A.C.; Silva-Santos, B. Tumor Cell Recognition by Γδ T Lymphocytes: T-Cell Receptor vs. NK-Cell Receptors. OncoImmunology 2013, 2, e22892. [CrossRef]
- Liguori, L.; Polcaro, G.; Nigro, A.; Conti, V.; Sellitto, C.; Perri, F.; Ottaiano, A.; Cascella, M.; Zeppa, P.; Caputo, A.; et al. Bispecific Antibodies: A Novel Approach for the Treatment of Solid Tumors. Pharmaceutics 2022, 14, 2442. [CrossRef]
- Thommen, D.S.; Schumacher, T.N. T Cell Dysfunction in Cancer. Cancer Cell 2018, 33, 547–562. [CrossRef]
- Chen, D.; Guo, Y.; Jiang, J.; Wu, P.; Zhang, T.; Wei, Q.; Huang, J.; Wu, D. Γδ T Cell Exhaustion: Opportunities for Intervention. Journal of Leukocyte Biology 2022, 112, 1669–1676. [CrossRef]
- Yamaguchi, T.; Suzuki, Y.; Katakura, R.; Ebina, T.; Yokoyama, J.; Fujimiya, Y. Interleukin-15 Effectively Potentiates the in Vitro Tumor-Specific Activity and Proliferation of Peripheral Blood gammadeltaT Cells Isolated from Glioblastoma Patients. Cancer Immunol Immunother 1998, 47, 97–103. [CrossRef]
- Yamaguchi, T.; Fujimiya, Y.; Suzuki, Y.; Katakura, R.; Ebina, T. A Simple Method for the Propagation and Purification of γδT Cells from the Peripheral Blood of Glioblastoma Patients Using Solid-Phase Anti-CD3 Antibody and Soluble IL-2. Journal of Immunological Methods 1997, 205, 19–28. [CrossRef]
- Fujimiya, Y.; Suzuki, Y.; Katakura, R.; Miyagi, T.; Yamaguchi, T.; Yoshimoto, T.; Ebina, T. In Vitro Interleukin 12 Activation of Peripheral Blood CD3(+)CD56(+) and CD3(+)CD56(-) Gammadelta T Cells from Glioblastoma Patients. Clin Cancer Res 1997, 3, 633–643.
- Suzuki, Y.; Fujimiya, Y.; Ohno, T.; Katakura, R.; Yoshimoto, T. Enhancing Effect of Tumor Necrosis Factor (TNF)-Alpha, but Not IFN-Gamma, on the Tumor-Specific Cytotoxicity of gammadeltaT Cells from Glioblastoma Patients. Cancer Lett 1999, 140, 161–167. [CrossRef]
- Cimini, E.; Piacentini, P.; Sacchi, A.; Gioia, C.; Leone, S.; Lauro, G.M.; Martini, F.; Agrati, C. Zoledronic Acid Enhances Vδ2 T-Lymphocyte Antitumor Response to Human Glioma Cell Lines. Int J Immunopathol Pharmacol 2011, 24, 139–148. [CrossRef]
- Nakazawa, T.; Nakamura, M.; Park, Y.S.; Motoyama, Y.; Hironaka, Y.; Nishimura, F.; Nakagawa, I.; Yamada, S.; Matsuda, R.; Tamura, K.; et al. Cytotoxic Human Peripheral Blood-Derived γδT Cells Kill Glioblastoma Cell Lines: Implications for Cell-Based Immunotherapy for Patients with Glioblastoma. J Neurooncol 2014, 116, 31–39. [CrossRef]
- Lamb, L.S.; Bowersock, J.; Dasgupta, A.; Gillespie, G.Y.; Su, Y.; Johnson, A.; Spencer, H.T. Engineered Drug Resistant Γδ T Cells Kill Glioblastoma Cell Lines during a Chemotherapy Challenge: A Strategy for Combining Chemo- and Immunotherapy. PLoS ONE 2013, 8, e51805. [CrossRef]
- Nabors, L.B.; Lamb, L.S.; Goswami, T.; Rochlin, K.; Youngblood, S.L. Adoptive Cell Therapy for High Grade Gliomas Using Simultaneous Temozolomide and Intracranial Mgmt-Modified Γδ t Cells Following Standard Post-Resection Chemotherapy and Radiotherapy: Current Strategy and Future Directions. Front. Immunol. 2024, 15, 1299044. [CrossRef]
- Joalland, N.; Chauvin, C.; Oliver, L.; Vallette, F.M.; Pecqueur, C.; Jarry, U.; Scotet, E. IL-21 Increases the Reactivity of Allogeneic Human Vγ9Vδ2 T Cells Against Primary Glioblastoma Tumors. J Immunother 2018, 41, 224–231. [CrossRef]
- Jiang, D.; Li, Y. Unraveling the Immunosuppressive Microenvironment of Glioblastoma and Advancements in Treatment. Front. Immunol. 2025, 16, 1590781. [CrossRef]
- Chitadze, G.; Kabelitz, D. Immune Surveillance in Glioblastoma: Role of the NKG2D System and Novel Cell-based Therapeutic Approaches. Scand J Immunol 2022, 96, e13201. [CrossRef]
- Wang, Y.; Ji, N.; Zhang, Y.; Chu, J.; Pan, C.; Zhang, P.; Ma, W.; Zhang, X.; Xi, J.J.; Chen, M.; et al. B7H3-Targeting Chimeric Antigen Receptor Modification Enhances Antitumor Effect of Vγ9Vδ2 T Cells in Glioblastoma. J Transl Med 2023, 21, 672. [CrossRef]
- Rosso, D.A.; Rosato, M.; Iturrizaga, J.; González, N.; Shiromizu, C.M.; Keitelman, I.A.; Coronel, J.V.; Gómez, F.D.; Amaral, M.M.; Rabadan, A.T.; et al. Glioblastoma Cells Potentiate the Induction of the Th1-like Profile in Phosphoantigen-Stimulated Γδ T Lymphocytes. J Neurooncol 2021, 153, 403–415. [CrossRef]
- Bryant, N.L.; Suarez-Cuervo, C.; Gillespie, G.Y.; Markert, J.M.; Nabors, L.B.; Meleth, S.; Lopez, R.D.; Lamb, L.S. Characterization and Immunotherapeutic Potential of Gammadelta T-Cells in Patients with Glioblastoma. Neuro Oncol 2009, 11, 357–367. [CrossRef]
- Beck, B.H.; Kim, H.; O’Brien, R.; Jadus, M.R.; Gillespie, G.Y.; Cloud, G.A.; Hoa, N.T.; Langford, C.P.; Lopez, R.D.; Harkins, L.E.; et al. Dynamics of Circulating Γδ T Cell Activity in an Immunocompetent Mouse Model of High-Grade Glioma. PLoS ONE 2015, 10, e0122387. [CrossRef]
- Chitadze, G.; Lettau, M.; Luecke, S.; Wang, T.; Janssen, O.; Fürst, D.; Mytilineos, J.; Wesch, D.; Oberg, H.-H.; Held-Feindt, J.; et al. NKG2D- and T-Cell Receptor-Dependent Lysis of Malignant Glioma Cell Lines by Human Γδ T Cells: Modulation by Temozolomide and A Disintegrin and Metalloproteases 10 and 17 Inhibitors. OncoImmunology 2016, 5, e1093276. [CrossRef]
- Jarry, U.; Chauvin, C.; Joalland, N.; Léger, A.; Minault, S.; Robard, M.; Bonneville, M.; Oliver, L.; Vallette, F.M.; Vié, H.; et al. Stereotaxic Administrations of Allogeneic Human Vγ9Vδ2 T Cells Efficiently Control the Development of Human Glioblastoma Brain Tumors. OncoImmunology 2016, 5, e1168554. [CrossRef]
- Nakazawa, T.; Nakamura, M.; Matsuda, R.; Nishimura, F.; Park, Y.S.; Motoyama, Y.; Hironaka, Y.; Nakagawa, I.; Yokota, H.; Yamada, S.; et al. Antitumor Effects of Minodronate, a Third-Generation Nitrogen-Containing Bisphosphonate, in Synergy with γδT Cells in Human Glioblastoma in Vitro and in Vivo. J Neurooncol 2016, 129, 231–241. [CrossRef]
- Chauvin, C.; Joalland, N.; Perroteau, J.; Jarry, U.; Lafrance, L.; Willem, C.; Retière, C.; Oliver, L.; Gratas, C.; Gautreau-Rolland, L.; et al. NKG2D Controls Natural Reactivity of Vγ9Vδ2 T Lymphocytes against Mesenchymal Glioblastoma Cells. Clinical Cancer Research 2019, 25, 7218–7228. [CrossRef]
- Choi, H.; Kim, T.-G.; Jeun, S.-S.; Ahn, S. Human Gamma-Delta (Γδ) T Cell Therapy for Glioblastoma: A Novel Alternative to Overcome Challenges of Adoptive Immune Cell Therapy. Cancer Letters 2023, 571, 216335. [CrossRef]
- Lobbous, M.; Goswami, T.; Lamb, L.S.; Rochlin, K.; Pillay, T.; Ter Haak, M.; Nabors, L.B. INB-200: Fully Enrolled Phase 1 Study of Gene-Modified Autologous Gamma-Delta (Γδ) T Cells in Patients with Newly Diagnosed Glioblastoma Multiforme (GBM) Receiving Maintenance Temozolomide (TMZ). JCO 2024, 42, 2042–2042. [CrossRef]
- Choi, B.D.; Gerstner, E.R.; Frigault, M.J.; Leick, M.B.; Mount, C.W.; Balaj, L.; Nikiforow, S.; Carter, B.S.; Curry, W.T.; Gallagher, K.; et al. Intraventricular CARv3-TEAM-E T Cells in Recurrent Glioblastoma. N Engl J Med 2024, 390, 1290–1298. [CrossRef]
Figure 1.
Schematic representation of γδ T cells engineered with CARs targeting GBM. The image illustrates γδ T cells expressing CARs specific for the tumor-associated antigens EGFRvIII (Epidermal Growth Factor) and IL-13Rα2, engaging with GBM cells. This strategy highlights the potential of γδ T cells as an alternative platform for CAR-based immunotherapy in the treatment of GBM. Illustration from NIAID NIH BIOART Source.
Figure 1.
Schematic representation of γδ T cells engineered with CARs targeting GBM. The image illustrates γδ T cells expressing CARs specific for the tumor-associated antigens EGFRvIII (Epidermal Growth Factor) and IL-13Rα2, engaging with GBM cells. This strategy highlights the potential of γδ T cells as an alternative platform for CAR-based immunotherapy in the treatment of GBM. Illustration from NIAID NIH BIOART Source.
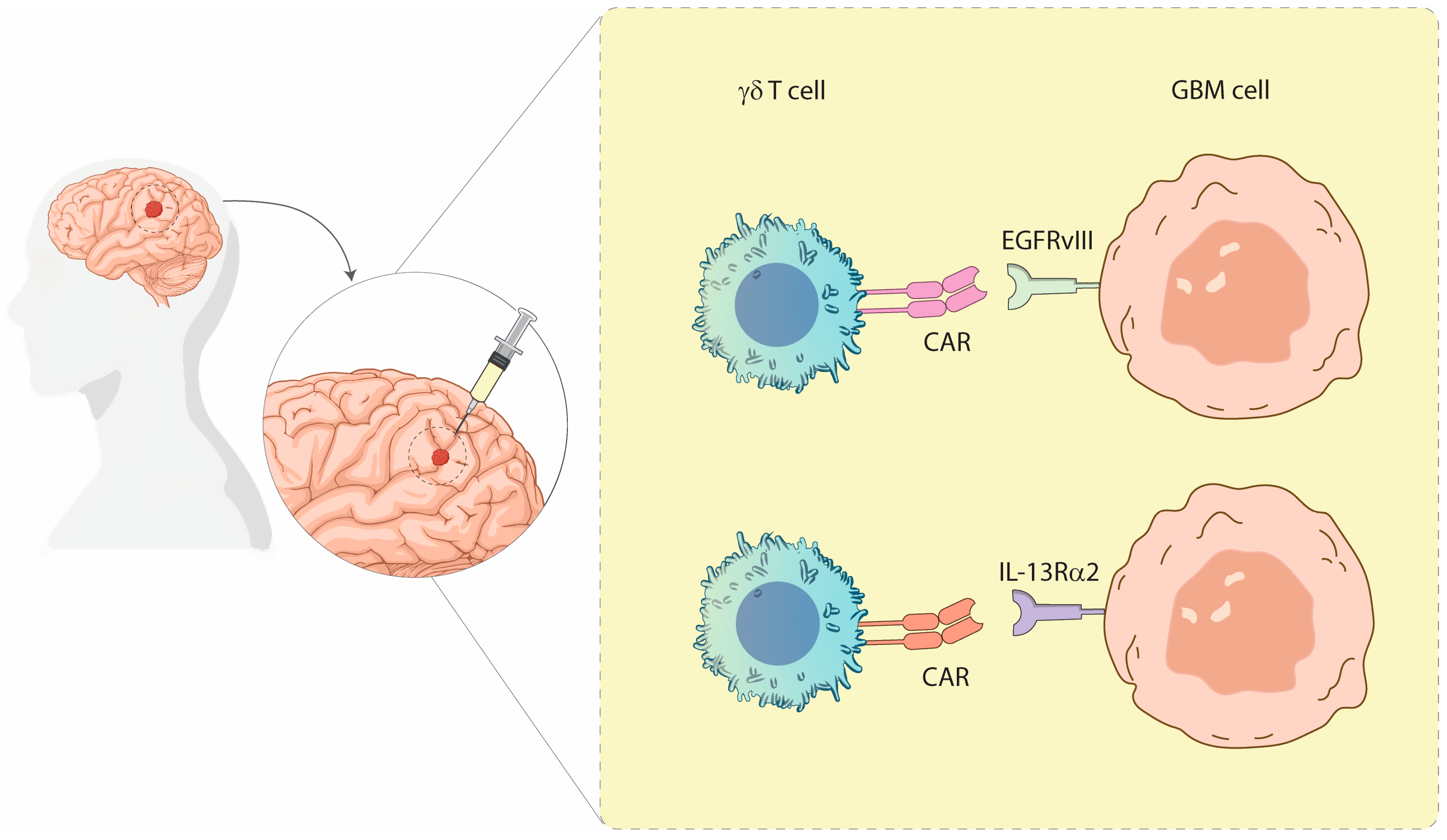

Table 1.
Summary of key studies investigating the therapeutic use of γδ T cells in GBM. The table highlights study characteristics, including type, aim, techniques, main findings, administration route, target, adverse events, and patient population. [Abbreviations: ROA-Administration Route; AE-Adverse Events; N/A-Not Available; IC-Intracranial; ST-IC-Stereotactic Intracranial; IP-Intraperitoneal; IT-Intra-tumoral; ICV-Intraventricular].
Table 1.
Summary of key studies investigating the therapeutic use of γδ T cells in GBM. The table highlights study characteristics, including type, aim, techniques, main findings, administration route, target, adverse events, and patient population. [Abbreviations: ROA-Administration Route; AE-Adverse Events; N/A-Not Available; IC-Intracranial; ST-IC-Stereotactic Intracranial; IP-Intraperitoneal; IT-Intra-tumoral; ICV-Intraventricular].
| Author, Year | Study Type | Aim of the Study | Technique | Findings | ROA (if therapy) |
Target | AE | Patients |
|---|---|---|---|---|---|---|---|---|
| Bryant et al., 2009 [68] | In vitro (preclinical) | Assess the innate immune function of T cells in GBM and their therapeutic potency | T cells from GBM patients and healthy controls were expanded and tested against U251MG, D54MG, U373MG, U87MG and primary GBM explants | GBM patients had fewer and weaker T cells. Activated T cells could kill GBM cells but spared normal astrocytes, indicating selective cytotoxicity. | N/A | GBM cell lines and primary cultures | N/A | GBM patients and healthy controls |
| Bryant et al., 2011 [13] | In vivo (mouse xenograft) | Evaluate T cell migration, infiltration, and antitumor efficacy in GBM | U251MG cells were implanted intracranially in immunodeficient mice, followed by stereotactic injection of expanded T cells | T cells significantly slowed tumor progression and extended survival. Demonstrated in vivo trafficking and tumor infiltration. | IC | U251MG xenografts | N/A | Immunodeficient mouse model |
| Cimini et al., 2011 [59] | In vitro | Investigate whether Zol enhances γδ T cell killing of GBM | Vδ2 T cells activated with phosphoantigens were co-cultured with T70, U251, and U373 GBM cells ± Zol | Zol enhanced perforin-mediated killing and induced apoptosis in a dose-dependent manner, increasing γδ T cell efficacy. | N/A | GBM cell lines | N/A | Cell line model |
|
Lamb et al., 2013 [61] |
In vitro | Determine if MGMT-modified T cells resist TMZ and retain cytotoxicity | T cells were transduced with MGMT and tested on TMZ-resistant U87, U373, SNB-19 GBM lines | MGMT+ T cells resisted TMZ and remained effective against GBM. TMZ increased NKG2D ligands, enhancing T cell sensitivity. | N/A | TMZ-resistant GBM cells | N/A | GBM cell lines |
| Nakazawa et al., 2014[60] | In vitro | Examine the role of Zol in boosting γδ T cell cytotoxicity | U87MG, U138MG, A172 GBM lines were treated with Zol before γδ T cell exposure | Zol sensitized GBM cells, killing from <32% to over 80% in some lines. Blocked by anti-TCR antibody. | N/A | GBM cell lines pretreated with Zol | N/A | GBM cell lines |
| Beck et al., 2015 [69] | In vivo (immunocompetent mouse) | Characterize γδ T cell kinetics and immune suppression in glioma | GL261 murine glioma model used to monitor T cell cytokines and tumor progression | T cells expanded early, then collapsed. Tumors secreted TGF-β, promoting immune evasion. No survival difference in γδ T cell KO mice. | N/A | GL261 gliomas | N/A | Immunocompetent mice |
| Chitadze et al., 2016 [70] | In vitro | Assess impact of TMZ and metalloprotease inhibitors on NKG2DL expression | U87MG, A172, T98G, U251MG treated with TMZ and ADAM10/17 inhibitors; tested with BrHPP-activated γδ T cells | TMZ upregulated ULBP2 on GBM surface. ADAM inhibition prevented shedding, enhancing T cell-mediated lysis. | N/A | NKG2DLs (MICA, MICB, ULBPs) | N/A | Human GBM cell lines |
| Jarry et al., 2016 [71] | In vivo (xenograft) | Test if stereotactic injection of γδ T cells with Zol improves survival | Vγ9Vδ2 T cells were injected into brains of mice bearing U87MG or GBM-10 xenografts ± Zol | Stereotactic delivery led to tumor clearance and improved survival, especially with Zol sensitization. | ST-IC | U87MG, GBM-10 | N/A | Mice with human GBM xenografts |
| Nakazawa et al., 2016 [72] | In vitro & in vivo | Evaluate synergy between minodronate and γδ T cells | Co-culture of GBM cells with γδ T cells ± minodronate; IP injection in immunocompromised mice | Combination induced higher granzyme B, TNF-α release, stronger apoptosis, and tumor inhibition in vivo. | IP (mice) | U87MG, U138MG | N/A | Immunodeficient mouse model |
| Joalland et al., 2018 [63] | In vivo | Investigate effect of IL-21 on γδ T cell cytotoxicity in GBM | Vγ9Vδ2 T cells were pretreated with IL-21 and injected into GBM-bearing mice | IL-21 enhanced cytotoxicity against GBM-1 and U87MG. Survival extended from 41 to 66 days. | ST-IC | GBM-1 (primary), U87MG | N/A | Orthotopic GBM mouse model |
| Chauvin et al., 2019 [73] | In vitro & in vivo | Evaluate immunoreactivity of Vγ9Vδ2 T cells against molecular subtypes of patient-derived GBM cells | Co-culture of Vγ9Vδ2 T cells with mesenchymal (GBM-1) and classical/proneural (GBM-10) cells; tested with/without nitrogen bisphosphonate | Vγ9Vδ2 T cells naturally lysed mesenchymal GBM-1 but not GBM-10. NBPs were required to sensitize GBM-10. Killing depended on TCR and NKG2D pathways. | N/A | Mesenchymal vs classical GBM subtypes | N/A | Patient-derived GBM primary cultures |
| Lee et al., 2019 [32] | In vitro | Characterize tumor-infiltrating γδ T cells in GBM patients | TCR sequencing of tumor and blood-derived γδ T cells from patients | Tumor-infiltrating Vγ9Vδ2 T cells had non-canonical TCR sequences and cytotoxic gene profiles similar to Th1 and M1 macrophages. | N/A | Tumor-infiltrating Vγ9Vδ2 T cells | N/A | GBM patients |
| Choi et al., 2023 [74] | Review | Summarize γδ T cells as a non-MHC-based GBM therapy | Review of existing in vitro/in vivo and early clinical data | Vγ9Vδ2 T cells use NKG2D/DNAM-1 to recognize stress ligands; CAR or checkpoint-enhanced platforms proposed. | IT, ICV (proposed) |
NKG2D, DNAM-1 ligands | N/A | Preclinical evidence |
| Nabors et al., 2024 [62] | Phase 1 clinical trial (ongoing) | Test safety/feasibility of MGMT-modified γδ T cells with TMZ | Drug-resistant immunotherapy: MGMT+ γδ T cells injected during concurrent TMZ | MGMT expression allows T cells to persist during TMZ. Trial ongoing; preliminary data shows feasibility. | IC | NKG2D ligands (e.g., MICA, ULBP2) | Not fully disclosed | Newly diagnosed GBM patients |
| Lobbous et al., 2024 [75] | Phase 1 trial | Determine safety of intracranial MGMT+ γδ T cells during SOC | MGMT+ γδ T cells injected into surgical resection cavity during standard Stupp regimen | No dose-limiting toxicities. Some patients had PFS benefit. Mild hematological AEs only. | IC (resection cavity) |
NKG2D ligands | Mild hematologic; no CRS | Newly diagnosed GBM patients |
| Choi et al., 2024 [76] | Phase 1 trial (dose escalation) | Assess safety of CARv3-TEAM-E T cells targeting EGFR/EGFRvIII | Autologous CAR-T cells delivered intraventricularly; dual-targeting with enhanced cytotoxic modules | Partial tumor regressions observed. No dose-limiting toxicities. Reversible neurotoxicity (grade 3) and fatigue reported. | ICV | EGFRvIII and wild-type EGFR | Grade 3 encephalopathy, fatigue | Recurrent EGFRvIII+ GBM patients |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



