Submitted:
24 June 2025
Posted:
24 June 2025
You are already at the latest version
Abstract
Background: The therapeutic potential of sulfur-containing mineral water (SMW) has been recognised since ancient times. Given the substantial amount of data on the bene-ficial effects of sulfur-containing active ingredients in foods, interest in SMW has been increasing in recent years, particularly regarding its contribution to human health as part of the daily diet. The anti-inflammatory potential of sulforaphane (SFN), allicin (ALL), and taurine (TAU) is well documented. This study aimed to analyse and com-pare the anti-inflammatory potential of SMW, SFN, ALL, and TAU on human intesti-nal epithelial cells (HIEC-6). Methods: A cell viability assay was conducted, selecting three concentrations from each sulfur-containing compound for analysis. In a model of lipopolysaccharide-induced inflammation, total RNA was extracted, and the gene ex-pressions of cyclooxygenase-2 (COX-2), tumour necrosis factor-alpha (TNFα), inter-leukin-1ß (IL-1ß), and IL-1ß receptor antagonist (IL1RN) were analysed. Results: With the exception of ALL, all studied compounds significantly decreased the expression of COX-2 in a dose-dependent manner. No change in IL-1ß expression was observed in SMW- and SFN-treated cells. All sulfur compounds, including SMW, significantly de-creased TNF-α expression, and this effect was particularly pronounced in TAU-treated cells. SMW, SFN, and ALL, but not TAU, increased the mRNA levels of IL-1RN in un-stimulated cells. Conclusions: SMW significantly modulated the expression of the studied markers, and these effects were comparable to those of SFN, TAU, and ALL. These results demonstrate the anti-inflammatory potential of SMW and elucidate the underlying mechanisms behind its therapeutic effects.
Keywords:
sulfur-containing mineral water
; sulforaphane
; taurine
; allicin
; COX-2
; TNF-
; IL-1
; IL-1RN
; HIEC-6
; LPS-induced inflammation
1. Introduction
The healing properties of mineral waters have been recognised since ancient times. Due to the extensive use of mineral waters in balneology, dermatology, and rheumatology, their anti-inflammatory properties are the focus of scientific interest [1,2,3,4]. Sulphur-containing mineral waters possess a wide range of therapeutic and balneological applications. The concentration of sulphur-containing compounds in mineral waters is considered a determinant of their anti-inflammatory or pro-inflammatory effects on human metabolism [5]. In a human intervention study, the daily intake of SMW demonstrated its antioxidant and anti-inflammatory potential by improving total thiol levels in plasma and reducing plasma C-reactive protein (CRP) in healthy volunteers [6,7]. Furthermore, SMW, prescribed as a complementary therapy for patients with specific pathological conditions, was reported to improve blood glucose concentration and to reduce plasma concentration of reactive oxygen metabolites [8].
The health benefits of SMW are attributed to its sulfur content, primarily in the form of hydrogen sulfide (H2S) and soluble sulfides, which are thought to modulate various biological processes [9,10].
Hydrogen sulfide is a gaseous transmitter that has various biological effects, including immunomodulatory effects by inhibiting the production of interleukin 6 (IL-6) and TNF-α in macrophages via the suppression of histone H3 acetylation at their promoters. This chromatin remodelling results in decreased gene transcription of these pro-inflammatory cytokines, highlighting an epigenetic mechanism through which H2S exerts anti-inflammatory effects [11] (Rios et al., 2015). Another study suggests that H₂S-mediated sulfhydration of nuclear factor kappa B (NF-κB) p65 plays a crucial role in its transcriptional activity and thus regulates immune responses [12]. A comprehensive review highlighted the dual role of H2S in inflammation. Under certain conditions, H2S can have pro-inflammatory effects or be anti-inflammatory by modulating mechanisms that underpin the relationship between oxidative stress and inflammation [13]. However, we should not overlook that SMW also contains other components, such as soluble sulfides and micro and macro elements, which may significantly influence metabolism and contribute to the biological effects.
Recently, the number of in vitro studies involving SMW has risen. Essentially, in vitro studies provide a foundation for exploring the mechanisms of action of various biologically active compounds. They enable rapid and detailed monitoring of changes in target markers following specific treatments. In the field of balneotherapy, primary cells or immortalised cell lines are employed to study the biological effects of a single inorganic molecule [5,14,15], an organic component extracted from mineral water [16], or the whole mineral water [5,17,18]. Moreover, it has been suggested that the sulfur-based compounds may exhibit a mild pro-inflammatory hormesis effect, which primes the body for protection against inflammation. However, despite the rising scientific interest in the beneficial effects of mineral waters and the mechanisms underlying these effects, SMW remains underestimated as a part of the daily diet and the molecular mechanisms of its anti-inflammatory effects are not fully elucidated.
Conversely, plant-derived organosulphur compounds and sulphur-containing amino acids attract significant scientific interest, with their biological effects being the focus of numerous in vitro, in vivo, and human interventional studies [19,20,21]. A wealth of scientific data accumulated in recent decades reveals the potential of organosulfides found in broccoli and garlic to contribute to the prevention of various pathological processes, including chronic inflammation, by suppressing the expression and activity of several inflammatory mediators such as COX-2, nitric oxide (NO), IL-1β, IL-6, TNF-α, among others [22,23,24]. Furthermore, these compounds have been shown to activate the antioxidant response by stimulating the Nrf2 signalling pathway [25] and display remarkable cytoprotective effects [26].
Sulforaphane (1-isothiocyanato-4-(methylsulfinyl)butane) is one of the most studied isothiocyanates, primarily due to its antioxidant, anticancer, and anti-inflammatory properties [27]. It is a dietary organosulfur compound found in plants from the Brassica genus, such as cauliflower, broccoli, kale, cabbage, collards, and others [28]. Consuming cruciferous vegetables has been linked to a reduced risk of certain metabolic disorders closely associated with low-grade inflammation, such as diabetes [29]. The anti-inflammatory potential of SFN has been well demonstrated in various types of in vitro models of inflammation, including lipopolysaccharide (LPS)-stimulated expression of COX-2 and TNF-α [30,31]. In addition to this data, the anti-inflammatory effects of SFN were demonstrated in in vivo models of lung injury, muscle inflammation, hypoxic pulmonary hypertension, and diabetes [31,32,33]. Furthermore, several clinical studies have demonstrated the potential of SFN as an adjuvant to anti-inflammatory therapies in humans. Unlike many other plant-derived biologically active compounds, such as polyphenols, SFN has good bioavailability and relatively low toxicity [34,35].
Allicin [S-(2-propenyl)-2-propene-1-sulfinothioate] is the most bioactive organosulfur compound found in garlic, which is released after crushing, cutting, or chewing the garlic cells. [21,36,37]. It is a thioester of sulfenic acid and is also known as allyl thiosulfinate, synthesized from alliin (S-allylcysteine sulfoxide) during the mechanical breakdown of garlic tissue [36].
Garlic organosulfur compounds, including allicin, have attenuated the release of proinflammatory mediators from various types of inflammatory cells, thereby reducing chronic inflammatory processes [38,39]. They can also inhibit the transcription factor NF-kB, which plays a key role in inflammation, and the phosphorylation of p38 mitogen-activated protein kinases. Lang et al. demonstrated that allicin has the potential to reduce intestinal inflammation by exerting an inhibitory effect on spontaneous and TNF-α-induced secretion of proinflammatory chemokines in HT-29 and Caco-2 epithelial cell lines in a dose-dependent manner [38].
Taurine is a conditionally essential sulfonated beta-amino acid synthesised in the human body from cysteine, and is often included in food supplements. Despite being one of the few non-protein amino acids, it is among the most abundant amino acids in the human body. It is found in high concentrations in various tissues, particularly in the brain, retina, muscle tissue, and phagocytic cells. Taurine has been reported to act as an antioxidant during oxidative stress-induced inflammation, exhibit cytoprotective effects, and potentially maintain homeostasis during both acute and chronic inflammation [40].
Scientific data accumulated in recent decades reveal the potential of active substances in foods, primarily of plant origin, to suppress inflammatory responses by inhibiting the activity of numerous pro-inflammatory markers in various in vitro models [30,31,41,42]. However, data comparing the biological effects of sulfur-containing active substances of natural origin on intestinal epithelial cell models are scarce. The gastrointestinal tract is likely the first site where bioactive compounds from foods and water exert their biological effects. Therefore, intestinal cell lines may serve as a valuable model for studying the mechanisms of these effects in both normal and pathological conditions, including inflammation models.
The present study aimed to perform a comparative analysis of the biological effects of sulfur-containing biologically active compounds (SBAC) from foods and waters on human intestinal epithelial cells. We investigated and compared the biological impact of SMW from the Varna basin with SBAC entering the body with foods, such as SFN from broccoli, ALL and TAU.
2. Materials and Methods
2.1. Sampling of Mineral Water
Twenty-four hours before the analyses the SMW samples were collected from a public fountain in Varna (Bulgaria) in clean plastic bottles and were stored in the dark at 4 °C in a refrigerator. Two hours before the experiment, the water samples were left at room temperature. Immediately before each experiment, a sample of 50 ml SMW was filtered through a 0.2 μm pore size filter and used to prepare working concentrations with nutritional medium.
2.2. Studied Compounds, Materials and Reagents
L-Sulforaphane (Cat. No. 14797) and Alliin (Cat. No. 14001) were purchased from Cayman Chemical Company, Ann Arbor, MI, USA. Taurine was obtained from Thermo Fisher Scientific, Inc., Waltham, MA, USA (Cat.No. 166540250). Reduced-Serum Medium Opti-MEM™ I with Phenol red (Cat. No. 31985-070), Phenol red-free Reduced-Serum Medium Opti-MEM™ I (Cat. No. 11058-021), Еpidermal growth factor (Cat. No. PHG 0311), and GlutaMAX™ Supplement (Cat. No. 35050061) were sourced from Thermo Fisher Scientific Inc. (Gibco, Waltham, MA, USA). Amplifyme SG Universal Mix (Cat. No. AM 02-200) was purchased from Blirt S.A. (Gdansk, Poland). PCR Red Mastermix (Cat. No. M3029.0500) was acquired from Genaxxon Bioscience GmbH (Ulm, Germany). Fetal Bovine Serum (FBS) (Cat. No. F7524), Phosphate Buffered Saline (PBS) (pH 7.4) (Cat. No. 806552), Dulbecco’s Phosphate Buffered Saline (DPBS) (Cat. No. 56064C), HEPES (pH 5-6.5) (Cat. No. H3375), and antibiotics were supplied by Sigma-Aldrich (St. Louis, MO, USA). Primezol TM reagent (Cat. No. AN1100) was obtained from Canvax Biotech, S.L. (Cordoba, Spain). 3-(4,5-Dimethylthiazol-2-yl)-2,5-Diphenyltetrazolium Bromide (MTT) (Cat. No. A2231.0005) was sourced from ITW Reagents, S.R.L. (AppliChem, Monza (MB), Italy). Dimethyl sulfoxide (DМSO) (Cat. No. BP2311), RevertAid™ First Strand cDNA Synthesis Kit (Cat. No. K1622), and RevertAid™ First Strand cDNA Synthesis Kit (Cat. No. K1622) were purchased from Thermo Fisher Scientific, (Waltham, MA, USA). Trypsin/EDTA 10x (Cat. No. 392-0459) was procured from VWR (Radnor, PA, USA). Salicylic acid, 99.5% (Cat. No. С351) was supplied by Chimtex LTD (Dimitrovgrad, Bulgaria). Lipopolysaccharides (LPS) from Escherichia coli (serotype O26:B6, Cat. No. L8274) were provided by Merck (Sigma Aldrich), Taufkirchen, Germany.
2.2. Cell Line and Subcultivation
The human intestinal epithelial cells HIEC-6 (ATCC® CRL3266™) were obtained from the American Type Culture Collection (ATCC) and cultured in OptiMEM reduced serum medium, containing 20 mM HEPES, 10 mM GlutaMAX, 10 ng/mL epidermal growth factor (EGF), 4% fetal bovine serum (FBS), and a penicillin/streptomycin mixture to final concentrations of 1% for each component. Cells were cultured in 75 cm² flasks and incubated at 37°C in a humidified atmosphere of 95% air and 5% CO2.
2.3. Cell Viability Assay
The 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay was employed to determine cell viability. The cells were collected and seeded in 12-well flasks using phenol red-free Opti-MEM™ I medium at a density of 6.5 x 10⁴ cells per well. Following 24 hours of incubation, the culture medium was removed and the cells were incubated in fresh medium containing increasing percentages of mineral water (v/v): 1%, 2%, 4%, 8%, 16%, and 20%, or sulfur-containing bioactive compounds, as follows: SFN (g/ml) 0.05, 0.1, 0.2, 0.25, 0.5, 1, 2, 5, 10; ALL (g/ml) 2.5, 5, 10, 25, 30, 50, 75, 100; TAU (mM/L) 0.1, 0.25, 0.5, 1, 2, 4, 5. After 20 hours of incubation, 100 µL of MTT working solution (2 mg/mL in phosphate buffer saline) was added to each well, and incubation continued for 4 hours. Following the incubation period, the MTT solution was removed, and 1 ml of dimethyl sulfoxide (DMSO) was added to each well to lyse the cells and dissolve the formazan crystals. Finally, absorbance was determined using a Synergy 2 plate reader (BioTek) at a wavelength of 550 nm. The viability of the treated cells was expressed as a percentage of the viability of the non-treated cells (controls), which was considered 100%. All treatments were performed in triplicate, and data were presented as mean ± SEM. Based on the MTT assay, three values of SMW percentage content in the culture medium, along with concentrations of SFN, ALL, and TAU, were selected for further analyses.
2.4. Induction of Inflammation
HIEC cells were seeded in Opti-MEM™ I medium using 12-well flasks at a density of 6.5 x 10^4 cells per well. After 24 hours, the cells were pre-treated and incubated for another 24 hours with selected concentrations of SMW (4, 6, and %) and sulfur-containing compounds: SFN [μg/ml] 0.5, 1, and 2; ALL [μg/ml] 10, 25, and 30; TAU [mM/ml] 1, 2, and 5. The cells were treated with 1 μg/ml LPS to induce inflammation. Additionally, SA was used as a reference compound with an anti-inflammatory effect (100 μM per well). Each treatment was conducted in triplicate.
2.5. Gene Expression Analysis
Two-step quantitative Real-Time PCR was employed to determine the gene expression levels of selected genes in the cell culture. The nucleotide sequences of the primers used for analysing the selected genes are presented in Table 1. The primers were designed using the Real-Time PCR gene expression Design Tool and were synthesised by Sigma–Aldrich and Integrated DNA Technologies.
2.5.1. Total RNA Extraction
After the experiment, the cells were treated with Primezol reagent to isolate total RNA. The protocol adhered fully to the manufacturer’s guidelines. Before proceeding to cDNA synthesis, the concentration of isolated RNA from each sample was measured using a multifunctional reader (Synergy 2, BioTek) and was utilised for cDNA synthesis.
2.5.2. Reverse Transcription
Extracted RNA (200 ng) was reverse transcribed and employed for cDNA synthesis using the First Strand cDNA Synthesis Kit, which contained oligo (dT)18 primer, reaction buffer, nuclease-free water, RNase inhibitor, nucleotide mix (dNTP), and reverse transcriptase enzyme. The reaction was performed according to the manufacturer’s instructions on the GeneAmp PCR System 9700 (Applied Biosystems), with a final reaction volume of 20 μl. Following synthesis, each cDNA sample was diluted with 60 μL of molecular biology-grade water and stored at -20°C.
2.5.3. Quantitative Real-Time Polymerase Chain Reaction (RT PCR)
As a template for quantitative real-time PCR, a preamplifier of 0.50 µL cDNA was used in a final reaction volume of 5 µl. The final concentration of the primers in the reaction mixture was 300 nm. Two-step RT PCR analysis was performed with the AMPLIFYME SG Universal Mix kit (Blirt S.A.) using Rox Low fluorescent binding dye. Reactions were conducted in 96-well plates with the following reaction parameters: enzyme activation and denaturation at 95°C for 3 minutes; denaturation at 95 °c for 5 seconds for 40 cycles; annealing at 60°C for 10 seconds for 40 cycles; extension and fluorescence detection at 72 °c for 18 seconds for 40 cycles, followed by melting curve analyses. The analyses were performed on the QuantStudio 5 (Applied Biosystems). For the calculation of gene expression levels, the 2-ΔΔCt method was applied [43] and mRNA levels were presented as relative units (RU). The expression of genes of interest was assessed by comparison with untreated cells (controls), where the mRNA levels were considered equal to 1. Actin beta was used as an endogenous control gene.
2.6. Statistical Analysis
Statistical analyses were conducted using Microsoft Excel and GraphPad Prism 10.0. An unpaired two-tailed Student’s t-test was employed to compare the differences between treated cells and the controls. All samples were measured in triplicate, and the mean values were utilised for analysis. The data are presented as mean ± standard error of the mean (SEM). A p value of less than 0.05 indicated a significant difference between the groups.
3. Results
3.1. Cell Viability
The effect of SMW and sulfur-containing bioactive compounds on cell viability was assessed using the MTT assay. Cells were treated with increasing concentrations of the compounds under investigation. Only concentrations that resulted in a survival rate exceeding 80% were deemed nontoxic. Based on the MTT results (Figure 1A–D), three concentrations of SMW and each sulfur-containing compound were chosen for further analyses.
The effect of SMW on HIEC-6 viability is illustrated in Figure 1A. It was observed that cell viability decreased sharply compared to the controls (p = 0.0003) at the highest percentage of SMW content in the culture medium (20%). However, even at this high concentration of SMW, cell viability remained above 80%. Values between 1% and 16% SMW content did not significantly alter cell viability. Based on these results, all applied concentrations of SMW were considered non-toxic for HIEC-6. Three concentrations of SMW in the culture medium were selected for further analyses: 4%, 6%, and 8%.
3.2. Expression of Target Genes in a Model of LPS-Induced Inflammation
To explore the anti-inflammatory potential of SMW and SBAC on HIEC-6, we employed a model of LPS-induced inflammation. As detailed in the previous chapter, cells were pre-treated for 24 hours with the substances under investigation, after which they were exposed to 1 μg/ml LPS. Some of the cells were also exposed to the same concentration of LPS alone, without any pretreatment. Furthermore, we utilised 3 mM salicylic acid as a reference substance known for its anti-inflammatory effects. As previously described, cells were treated with three selected concentrations of each studied compound, in accordance with the MTT assay. The mRNA levels of COX-2, TNF-α, IL-1β, and IL1RN were compared with those of untreated cells.
Interestingly, SFN significantly stimulated cell proliferation at 0.25, 0.5, and 1 µg/ml (Figure 1B). However, cell viability noticeably decreased at 5 µg/ml SFN. Furthermore, the highest concentration of SFN markedly inhibited cell viability (74%) and may be deemed cytotoxic (p<0.001). Concentrations of 0.5, 1, and 2 µg/ml SFN were employed in the subsequent analyses.
As demonstrated in Figure 1C, the TAU concentrations of 0.2-5 mM did not affect cell viability. The slight decrease in viability of cells treated with 10 mM TAU was not statistically significant (95%, p=0.2). A statistically significant reduction in cell viability was observed in cells treated with 20 mM TAU (p=0.03). Nevertheless, even at this concentration, cell viability remained sufficiently high (89%) to conclude that all TAU concentrations applied were non-toxic to HIEC-6. The TAU concentrations used in the subsequent analyses were 1, 2, and 5 mM.
Cell viability in ALL-treated cells (Figure 1D) significantly decreased at the three highest concentrations. Notably, the viability of cells treated with the highest concentration of 100 μg/ml approached the 80% threshold. Based on these results, the chosen concentrations of allicin for the subsequent analyses were 10, 25, and 30 µg/ml.
3.2.1. Effect of SMW and SBAC on COX-2 Expression
We explored the potential of SMW and SBAC to influence COX-2 expression. Data are presented in Figure 2A–D. As observed, in cells treated with LPS, the gene expression was 1.8-fold higher (p=0.0003). Conversely, SA significantly inhibited the expression of COX-2.
Figure 2A illustrates the mRNA levels of the COX-2 gene in SMW-treated cells. We observed that pretreatment with SMW significantly reduced COX-2 transcription levels in LPS-stimulated HIEC-6. The lowest concentration of SMW significantly decreased mRNA levels in unstimulated cells (p=0.05). Conversely, the highest concentration of 8% SMW significantly suppressed the expression of the enzyme, both in unstimulated cells and under conditions of induced inflammation, suggesting a concentration-dependent effect of SMW on COX-2 expression.
The effect of SFN on COX-2 expression was comparable (Figure 2B). Notably, the highest concentration of 2 μg/ml inhibited gene expression in both LPS-stimulated and unstimulated cells (p=0.037 and p=0.04, respectively).
Taurine pretreatment significantly reduced COX-2 transcription levels in LPS-stimulated HIEC-6 across all three concentrations tested, with this effect being markedly more pronounced in cells treated with the highest concentration (Figure 2C).
None of the applied concentrations of ALL changed the expression levels of the COX-2 gene (Figure 2D).
3.2.2. Effect of SMW and SBAC on TNF-α Expression
Another target pro-inflammatory gene in our study was TNF-α. The results illustrating the effect of SMW and SBACon on TNF-α expression under conditions of LPS-induced inflammation are summarised in Figure 3A–D. Regarding COX-2, in cells treated with LPS alone, the TNF-α gene was significantly overexpressed compared to the control cells (p=0.02). Salicylic acid inhibited the expression of the gene (p=0.0047).
The inhibitory effect of SMW on TNFα expression was significant and dependent on concentration (Figure 3A). Notably, while unstimulated cells show that only the highest concentration exhibits an inhibitory effect on gene expression, all three SMW concentrations demonstrate such an effect under induced inflammation.
TNF-α expression was significantly inhibited in stimulated cells that were pretreated with 1 and 2 μg/ml SFN (Figure 3B). A concentration of 0.5 μg/ml SFN did not influence the expression of this gene, and its mRNA levels remained comparable to those measured in untreated controls.
Taurine demonstrated significant potential to inhibit TNF-α expression, both in conditions of induced inflammation and in unstimulated cells, and this effect was concentration-dependent (Figure 3C)
The effect of ALL on TNF-α expression was also investigated. In the absence of inflammation, we observed a statistically significant decrease in cytokine expression in cells treated with 10 µg/ml allicin compared to the control. Under conditions of induced inflammation, a concentration of 25 µg/ml significantly reduced mRNA levels (Figure 3D).
3.2.3. Effect of SMW and SBACon IL-1β Expression
The effect of SMW and SBACon IL-1β expression is illustrated in Figure 4A–D. Similarly to the previous target genes, LPS significantly stimulated IL-1β gene expression, whereas in cells treated with SA, mRNA levels were notably lower than those in the controls. No change in IL-1β gene expression was observed in SMW and SFN-treated cells at any of the applied concentrations (Figure 4A and B, respectively). In cells with LPS-induced inflammation, taurine demonstrated a statistically significant inhibitory effect on mRNA levels for the IL-1β gene (Figure 4C). At concentrations of 25 and 30 µg/ml, ALL exhibited an inhibitory effect on IL-1β expression, but only in cells with induced inflammation. In other instances, no effect of allicin on gene expression was found (Figure 4D).
3.2.4. Effect of SMW and SBACon IL-1RN Expression
We investigated the effect of SMW and SBAC on the gene expression for the anti-inflammatory marker IL-1RN (Figure 5A–D). In cells treated with LPS alone, significantly lower mRNA levels for the gene were observed compared to untreated cells. Salicylic acid had no effect on the gene expression of this marker.
The results presented in Figure 5 indicate that SMW, SFN, and ALL increased the expression of IL-1RN, but only in unstimulated cells. It is worth noting that for allicin, this effect is pronounced; the IL-1RN gene was significantly overexpressed at all applied concentrations (Figure 5A,B,D). No changes in mRNA levels were observed in cells treated with taurine (Figure 5C).
4. Discussion
For many years, scientists have concentrated their efforts on functional foods and nutrition, primarily from plant sources, as providers of active substances that contribute to human health. In this respect, it can be said that mineral waters have long been underestimated as a source of biologically active substances within the human body, with knowledge of their beneficial effects relying chiefly on empirical data.
Since H2S has been identified as a signalling gas molecule with numerous biological effects, interest in SMW has grown significantly. A 2013 study demonstrated the immunomodulatory and antioxidant properties of SMW and compared them to other exogenous donors of H2S in patients with respiratory diseases [44]. A physico-chemical analysis reported that the dominant microelement in Varna mineral waters is sulphur in the form of soluble sulfides and H2S [45].
In the present study, we compared the SMW and SBAC in terms of their potential to modulate pro- and anti-inflammatory markers from foods in human intestinal epithelial cells. To our knowledge, this is the first study to explore the molecular mechanisms behind the anti-inflammatory potential of SMW in comparison with well-studied sulfur-containing compounds.
4.1. Effects of SMW and SBAC on Cell Viability
The first part of our study involved determining the range of non-toxic concentrations of all tested substances (Figure 1A–D). As shown in Figure 1A, the highest concentration of SMW significantly decreased cell viability. However, even at this concentration of SMW in the culture medium, cell viability did not fall below the threshold of 80%. For the subsequent experiments, we selected three medium concentrations, aiming to ensure 100% cell viability while maintaining a sufficient presence of active ingredients in the water [45].
An intriguing result emerged from the SFN cytotoxicity test, revealing that three of the concentrations had a proliferative effect, increasing cell viability to approximately 109% (Figure 1B). SFN has been extensively researched for its anticancer potential, demonstrated as cytotoxic activity on cancer cell lines [27]. Low doses of SFN may enhance cell survival, while high doses of the compound may inhibit cell viability. In this study, the intermediate concentrations stimulated cell proliferation. However, the cell line used in the present study is derived from normal intestinal epithelial cells with intact homeostasis, which likely influences the effects of SFN on cell viability, suggesting that the biological effects of SFN depend on its concentration, cell type, and conditions.
Regarding taurine, cell viability remained unchanged across all applied concentrations. Even at the highest concentration, where a statistically significant decrease was noted, approximately 90% cell survival was observed (Figure 1C). Based on these results, we can infer that all taurine concentrations are non-toxic. The last three concentrations of allicin, 50, 75, and 100 g/ml, significantly reduced cell viability to 96% (p=0.002), 93% (p=0.002), and 87% (p=0.006), respectively (Figure 1D). Allicin has been documented to exhibit high cytotoxic activity in vitro across various cancer cell lines, similar to sulforaphane. [21,46]. However, in our study, even these high concentrations of allicin did not reduce cell viability below 80%.
In summary, all applied concentrations of SMW, TAU, and Allicin can be considered non-toxic to the tested cells. Conversely, SFN exhibits a complex effect on cell viability, stimulating proliferation at the medium concentrations used while displaying cytotoxicity at the highest concentration.
4.2. Effects of SMW and SBAC on COX-2 Gene Expression
We investigated the potential of SMW to influence the expression of COX-2 in a model of LPS-induced inflammation. The results indicated that a 4% concentration of SMW in the nutrient medium inhibited COX-2 expression in unstimulated cells only. However, at the highest concentration, SMW significantly suppressed the enzyme’s expression in both unstimulated and LPS-stimulated cells. This suggests a concentration-dependent effect of SMW on COX-2 gene expression. Interestingly, the effect of SMW closely resembled that of SFN on COX-2 expression (Figure 2B). This similarity is not surprising, as SFN’s anti-inflammatory properties are well established and considered fundamental to its anticancer potential as an organosulfur compound [30,47].
Dietary TAU is believed to significantly prevent cardiovascular disease and type 2 diabetes due to its beneficial biological effects, including reducing pro-inflammatory cytokines [40,48]. In our study, TAU suppressed the expression of COX-2 solely in LPS-stimulated cells, and this effect was concentration-dependent (Figure 2C). Studies suggest that one potential mechanism underpinning the anti-inflammatory effects of TAU relates to its ability to decrease mRNA expression of Toll-Like Receptor-2 (TLR-2), thereby inhibiting the expression of pro-inflammatory cytokines [48,49]. Therefore, TAU could be regarded as a therapeutic agent with the potential to mitigate inflammatory signals.
Cyclooxygenase-2 is the inducible isoform of the enzyme involved in the synthesis of prostanoids (prostaglandins, prostacyclins, and thromboxanes) from arachidonic acid [50]. Its expression is activated in response to various pro-inflammatory stimuli. Unlike the constitutive isoform COX-1, the inducible isoform is not subject to biochemical regulation, causing the prostaglandins produced under the action of COX-2 to lose their beneficial physiological effects and instead act as mediators of acute and chronic inflammation and pain perception [41]. For these reasons, COX-2 is a primary target in the development of anti-inflammatory drugs—the therapeutic effect of non-steroidal anti-inflammatory drugs (NSAIDs) stems from their inhibitory effect on cyclooxygenase. Most synthetic NSAIDs lack selectivity for the inducible isoform; hence, a significant undesirable effect of their use is ulcer formation. Scientists are focused on creating NSAIDs with high selectivity for COX-2 and a safer profile to address this issue [51]. In this context, scientific data on the anti-inflammatory effects of active substances from natural sources, which can potentially suppress the activity or expression of the enzyme, could form the basis for developing new generations of selective NSAIDs without undesirable side effects.
Surprisingly, ALL pretreatment did not affect COX-2 expression in HECs at any of the concentrations used. Alongside SFN, allicin has been extensively studied for its anti-inflammatory properties, both in vitro and in vivo. The effects are likely to vary depending on the cell type and the stimulus applied.
4.3. Effects of SMW and SBAC on TNF-α Gene Expression
Tumour necrosis factor alpha is a cytokine recognised as a pivotal regulator of inflammatory processes and is known to be involved in the pathogenesis of several inflammatory and autoimmune diseases [52]. TNF-α can mediate various aspects of inflammation, such as enhancing endothelial cell permeability, thereby facilitating leukocyte migration and releasing other cytokines [53].
In this study, SMW and all SBAC demonstrated a potential to suppress TNF-α expression; for SMW, SFN, and TAU, this effect was concentration-dependent and pronounced under induced inflammation conditions. The highest concentrations of SMW and TAU suppressed the expression of the marker in both unstimulated cells and LPS-stimulated inflammation conditions (Figure 3A and C, respectively). Compared to the control cells, ALL significantly decreased TNF-α expression in unstimulated cells treated with 10 µg/ml and in LPS-stimulated cells treated with 25 µg/ml (Figure 3D).
There is a substantial body of data in the scientific literature supporting the potential of SFN, TAU, and ALL to suppress TNF-α expression. Organosulfur compounds from garlic and cruciferous vegetables exhibit significant anti-inflammatory properties through multiple mechanisms. Sulforaphane, for example, was shown to alleviate LPS-induced cell damage in Caco-2 cells by activating the AMPK/SIRT1/PGC-1α cascade [54]. Furthermore, SFN and ALL have been reported to attenuate the expression of pro-inflammatory cytokines by downregulating transcription factors such as NF-kB and Nrf-2 [55,56].
The anti-inflammatory potential of TAU in inhibiting TNF-α is also well documented. Taurine supplementation has been shown to reduce cytokine levels in various contexts, including sepsis models and in response to LPS stimulation [57]. It has been reported that in stimulated pulmonary macrophages, the TAU metabolite taurine chloride inhibits NF-κB migration to the nucleus by stabilising IκB in the cytoplasm [58].
As was mentioned before, the main active component in the SMW we studied is H2S. There is sufficient scientific evidence for its anti-inflammatory effects. A recently published review summarises the data revealing the mechanisms by which this transmitter gas regulates inflammation. In specific inflammatory conditions, H2S alleviates inflammation by decreasing the expression of pro-inflammatory mediators, including TNF-α, and by inhibiting important signalling pathways, including phosphatidylinositol 3-kinase (PI3K), mitogen-activated protein kinase (MAPK), and NF-κB [59].
This data suggests that SMW and SBACs may affect specific targets in multiple signalling pathways while also modulating common markers of pro-inflammatory signals.
4.4. Effects of SMW and SBAC on IL-1β Gene Expression
Interleukin-1 beta (IL-1β) is a multifunctional pro-inflammatory cytokine that plays a crucial role in both acute and chronic inflammation [60]. It exhibits a pyrogenic effect and has the potential to induce pain signals [61]. IL-1β is expressed in a wide range of tissues and various cell types, particularly in macrophages found in lymphoid organs such as the thymus, spleen, lymph nodes, and bone marrow. In non-lymphoid organs, IL-1β is present in tissue macrophages of the lung, digestive tract, and liver and in various specific cell types, including neutrophils, keratinocytes, epithelial and endothelial cells, lymphocytes, smooth muscle cells, and fibroblasts [60]. The binding of IL-1β to its receptor activates several intracellular signalling pathways in target cells. One of the primary effects of transducing these signals is the creation of a pro-inflammatory environment within the cell through the release of a broad network of pro-inflammatory mediators, including cytokines, chemokines, metalloproteinases, acute phase proteins, prostaglandins, and nitric oxide [62].
In our study, TAU and ALL significantly inhibited gene expression of IL-1β, but only under conditions of LPS-induced inflammation (Figure 4B,D). This result supports the data reported by other studies on the therapeutic potential of both SBACs. On the other hand, neither SMW nor SFN demonstrated such potential. The anti-inflammatory potential of the studied biologically active substances in vitro is likely dependent on the target signalling pathways, the model, and the cell type. A class of receptors, known as NLR family pyrin domain containing 3 (NLRP3), expressed in macrophages, is activated by various harmful stimuli, such as reactive oxygen species, crystalline uric acid, and LPS-induced inflammation, subsequently leading to the release of IL-1β. The HIEC-6 cell line used in our study is fetal, and as a result, specific proteins are expressed very weakly or not at all in these cells, including NLRP3. A recent study reported that in LPS-stimulated HIEC-6, activation of IL-1β is possible through an NLRP3-independent pathway involving caspase-4 [63]. It can be expected that caspase-4 could also be a target for the anti-inflammatory effects of TAU and ALL, which remains to be confirmed in future studies.
4.5. Effects of SMW and SBAC on IL1RN Gene Expression
The IL-1β receptor antagonist (IL1RN; IL-1Ra) is a natural inhibitor of the cytokine’s binding to its receptor and, therefore, plays an important role as an anti-inflammatory mediator [64,65]. The balance between IL-1β and IL1RN is a crucial regulatory factor in determining the outcome of the inflammatory process activated by IL-1β [66,67].
The antagonist IL1RN is synthesised in response to various proinflammatory stimuli, acting as an IL-1β inhibitor by competitively binding to its receptor [60]. Once bound to the receptor, IL1RN prevents the binding of IL-1β, thereby inhibiting the signalling pathway and reducing the release of several proinflammatory factors, including IL-6, IL-8, and TNF-α [62].
Research has demonstrated that the inhibition of IL1RN results in an increase in proinflammatory signals and tissue damage. Thus, the balance between IL-1β and IL1RN is crucial for maintaining a robust anti-inflammatory status [68]. Given the scientific evidence supporting a physiological relationship between these two markers and the regulatory role of IL1RN on IL-1β activity, it can be inferred that IL1RN itself could serve as a potential target for the development of new anti-inflammatory drugs [60].
In our study, overexpression of IL1RN was detected in cells pretreated with SMW, SFN, and ALL, but only in unstimulated cells. This result highlights the potential of these natural compounds to enhance immune defence in uncompromised cells, which is likely to increase their resistance and survival when proinflammatory stimuli are present.
Studies examining the factors influencing transcriptional regulation and the relationship between IL-1β and IL1RN are relatively scarce. In the present study, we analysed the effects of the investigated SMW and SBAC on the expression of the genes for IL-1β and IL1RN. We believe that the results obtained provide a valuable contribution to clarifying this relationship and enhancing our understanding of the anti-inflammatory potential of biologically active substances of natural origin.
In summary, this study examined the potential of SMW from the Varna basin to influence the expression of markers related to the inflammatory response. The results discussed above indicated that the anti-inflammatory effects of SMW are comparable to those of organosulfur compounds known for their well-established anti-inflammatory properties.
5. Conclusions
The results obtained confirm the data on the anti-inflammatory potential of sulforaphane, allicin, and taurine, revealing for the first time the ability of sulfur-containing mineral water to inhibit the expression of genes related to the inflammatory response, such as COX-2, TNF-α, and IL-1β, both under LPS-induced inflammation and in unstimulated cells. In some cases, the effect of the SMW is even more pronounced than that of SFN and ALL. For SMW, these represent the first scientific data of their kind supporting our understanding of its anti-inflammatory effect and its potential to modulate gene expression related to the inflammatory response in intestinal cells.
The results obtained in this study contribute to enhancing our understanding of the effects of active substances from food and mineral waters on the markers associated with the inflammatory response. These findings may also assist in developing new strategies for personalised nutrition, with implications for human health.
Funding
This study was supported by European Union-NextGenerationEU, through the National Recovery and Resilience Plan of the Republic of Bulgaria, Project No BG-RRP-2.004-0009-C02 MUVE-TEAM); Medical University of Varna, Science Fund, Project No 22003.
Author Contributions Conceptualization
M.N.; methodology: M.N. and D.V.; validation: M.N., D.V. and T.S.; formal analysis: M.N.; investigation: M.N. and S.S.; writing—original draft preparation: M.N.; writing—review and editing: M.N., D.V. and T.S.; visualization: M.N.; supervision: M.N.; project administration: M.N. and S.S.; funding acquisition: M.N.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in this study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| A | Alliin |
| COX-2 | Inducible cyclooxygenase |
| CRP | C-reactive protein |
| H2S | Hydrogen sulfide |
| IL-1RN | IL-1β receptor antagonist |
| IL-1β | Interleukin -1β |
| IL-6 | Interleukin 6 |
| NF-κB | Nuclear factor kappa B |
| NO | Nitric oxide |
| PGE2 | Prostaglandin E2 |
| SBAC | Sulfur-containing biologically active compounds |
| SFN | Sulforaphane |
| SMW | Sulphur-containing mineral waters |
| TAU | Taurine |
| TNF-α | Tumour necrosis factor alpha |
References
- Bálint, G.P.; Buchanan, W.W.; Adám, A.; Ratkó, I.; Poór, L.; Bálint, P.V.; Somos, E.; Tefner, I.; Bender, T. The effect of the thermal mineral water of Nagybaracska on patients with knee joint osteoarthritis—a double blind study. Clin. Rheumatol. 2007, 26(6), 890–894. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.P.; Choi, Y.J.; Cho, K.A.; Woo, S.Y.; Yun, S.T.; Lee, J.T.; Kim, H.J.; Lee, K.H.; Kim, J.W. Effect of spa spring water on cytokine expression in human keratinocyte HaCaT cells and on differentiation of CD4(+) T cells. Ann. Dermatol. 2012, 24(3), 324–336. [Google Scholar] [CrossRef]
- Seite, S. Thermal waters as cosmeceuticals: La Roche-Posay thermal spring water example. Clin. Cosmet. Investig. Dermatol. 2013, 6, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Branco, M.; Rêgo, N.N.; Silva, P.H.; Archanjo, I.E.; Ribeiro, M.C.; Trevisani, V.F. Bath thermal waters in the treatment of knee osteoarthritis: a randomized controlled clinical trial. Eur. J. Phys. Rehabil. Med. 2016, 52(4), 422–430. [Google Scholar] [PubMed]
- Carbajo, J.M.; Maraver, F. Sulphurous mineral waters: New applications for health. Evid.-Based Complement. Alternat. Med. 2017, 2017, 8034084. [Google Scholar] [CrossRef]
- Roussev, B.; Sokrateva, T.; Nashar, M.; Radanova, M.; Komosinska-Vassev, K.; Olczyk, P.; Potoroko, I.; Ivanova, D. Effect of sulfur-containing mineral water on the renal function: a human interventional study. Compt. Rend. Acad. Bulg. Sci. 2019, 72(11), 1577–1584. [Google Scholar]
- Sokrateva, T.; Roussev, B.; Nashar, M.; Kiselova-Kaneva, Y.; Mihaylova, G.; Todorova, M.; Pasheva, M.; Tasinov, O.; Nazifova-Tasinova, N.; Vankova, D.; Ivanova, D.P.; Radanova, M.; Galunska, B.; Vlaykova, T.; Ivanova, D.G. Effects of sulfur-containing mineral water intake on oxidative status and markers for inflammation in healthy subjects. Arch. Physiol. Biochem. 2021, 127(4), 327–336. [Google Scholar] [CrossRef]
- Costantino, M.; Conti, V.; Corbi, G.; Filippelli, A. Hydropinotherapy with sulphurous mineral water as complementary treatment to improve glucose metabolism, oxidative status, and quality of life. Antioxidants 2021, 10, 1773. [Google Scholar] [CrossRef]
- Karagülle, M.Z; Karagülle, M.; Kılıç, S.; et al. . In vitro evaluation of natural thermal mineral waters in human keratinocyte cells: a preliminary study. Int J Biometeorol, 1661. [Google Scholar] [CrossRef]
- Cheleschi, S.; Gallo, I.; Tenti, S. A comprehensive analysis to understand the mechanism of action of balneotherapy: why, how, and where they can be used? Evidence from in vitro studies performed on human and animal samples. Int. J. Biometeorol. 2020, 64(7), 1247–1261. [Google Scholar] [CrossRef]
- Rios, E.C.; Szczesny, B.; Soriano, F.G.; Olah, G.; Szabo, C. Hydrogen sulfide attenuates cytokine production through the modulation of chromatin remodeling. Int. J. Mol. Med. 2015, 35(6), 1741–1746.Sen, N.; Paul, B.D.; Gadalla, M.M.; Mustafa, A.K.; Sen, T.; Xu, R.; Kim, S.; Snyder, S.H. Hydrogen sulfide-linked sulfhydration of NF-κB mediates its antiapoptotic actions. Mol. Cell 2012, 45, 13–24. [Google Scholar]
- Sen, N.; Paul, B.D.; Gadalla, M.M.; Mustafa, A.K.; Sen, T.; Xu, R.; Kim, S.; Snyder, S.H. Hydrogen sulfide-linked sulfhydration of NF-κB mediates its antiapoptotic actions. Mol. Cell 2012, 45, 13–24. [Google Scholar] [CrossRef]
- Bhatia, M.; Gaddam, R.R. Hydrogen Sulfide in Inflammation: A Novel Mediator and Therapeutic Target. Antioxid. Redox Signal. 2021, 34, 1368–1377. [Google Scholar] [CrossRef] [PubMed]
- Wallace, J.L.; Wang, R. Hydrogen sulfide-based therapeutics: exploiting a unique but ubiquitous gasotransmitter. Nat. Rev. Drug Discov. 2015, 14, 329–345. [Google Scholar] [CrossRef]
- Viegas, J.; Esteves, A.F.; Cardoso, E.M.; Arosa, F.A.; Vitale, M.; Taborda-Barata, L. Biological Effects of Thermal Water-Associated Hydrogen Sulfide on Human Airways and Associated Immune Cells: Implications for Respiratory Diseases. Front. Public Health 2019, 7, 128. [Google Scholar] [CrossRef] [PubMed]
- Gerencsér, G.; Szabó, I.; Szendi, K.; Hanzel, A.; Raposa, B.; Gyöngyi, Z.; Varga, C. Effects of medicinal waters on the UV-sensitivity of human keratinocytes - a comparative pilot study. Int. J. Biometeorol. 2019, 63, 1417–1423. [Google Scholar] [CrossRef] [PubMed]
- Fioravanti, A.; Lamboglia, A.; Pascarelli, N.A.; Cheleschi, S.; Manica, P.; Galeazzi, M.; Collodel, G. Thermal water of Vetriolo, Trentino, inhibits the negative effect of interleukin 1β on nitric oxide production and apoptosis in human osteoarthritic chondrocyte. J. Biol. Regul. Homeost. Agents 2013, 27, 891–902. [Google Scholar]
- Silva, A.; Oliveira, A.S.; Vaz, C.V.; Correia, S.; Ferreira, R.; Breitenfeld, L.; Martinez-de-Oliveira, J.; Palmeira-de-Oliveira, R.; Pereira, C.M.F.; Palmeira-de-Oliveira, A.; Cruz, M.T. Anti-inflammatory potential of Portuguese thermal waters. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef]
- Petropoulos, S.; Di Gioia, F.; Ntatsi, G. Vegetable Organosulfur Compounds and their Health Promoting Effects. Curr. Pharm. Des. 2017, 23, 19. [Google Scholar] [CrossRef]
- Chen, C.-J.; Cheng, M.-C.; Hsu, C.-N.; Tain, Y.-L. Sulfur-Containing Amino Acids, Hydrogen Sulfide, and Sulfur Compounds on Kidney Health and Disease. Metabolites 2023, 13, 688. [Google Scholar] [CrossRef]
- Stoeva, S.; Nashar, N.; Sokratreva, T.; Vankova, D.; Zamtikova, M.; Zaykov, H.; Varbanov, P. Anti-inflammatory Properties of Plant-Derived Organosulfur Compounds: Insights from Sulforaphane and Allicin. Scr. Sci. Pharm.
- Esteve, M. Mechanisms Underlying Biological Effects of Cruciferous Glucosinolate-Derived Isothiocyanates/Indoles: A Focus on Metabolic Syndrome. Front. Nutr. 2020, 7, 111. [Google Scholar] [CrossRef]
- Lu, P.D.; Zhao, Y.H. Targeting NF-κB Pathway for Treating Ulcerative Colitis: Comprehensive Regulatory Characteristics of Chinese Medicines. Chin. Med. 2020, 15. [Google Scholar] [CrossRef]
- Kamal, R.M.; Abdull Razis, A.F.; Mohd Sukri, N.S.; Perimal, E.K.; Ahmad, H.; Patrick, R.; et al. Beneficial Health Effects of Glucosinolates-Derived Isothiocyanates on Cardiovascular and Neurodegenerative Diseases. Molecules 2022, 27, 624. [Google Scholar] [CrossRef] [PubMed]
- Caglayan, B.; Kilic, E.; Dalay, A.; Altunay, S.; Tuzcu, M.; Erten, F.; et al. Allyl Isothiocyanate Attenuates Oxidative Stress and Inflammation by Modulating Nrf2/HO-1 and NF-kB Pathways in Traumatic Brain Injury in Mice. Mol. Biol. Rep. 2019, 46, 241–250. [Google Scholar] [CrossRef]
- Egbujor, M.C.; Petrosino, M.; Zuhra, K.; Saso, L. The Role of Organosulfur Compounds as Nrf2 Activators and Their Antioxidant Effects. Antioxidants 2022, 11, 1255. [Google Scholar] [CrossRef] [PubMed]
- Baralić, K.; Živanović, J.; Marić, Đ.; Bozic, D.; Grahovac, L.; Antonijević Miljaković, E.; Ćurčić, M.; Buha Djordjevic, A.; Bulat, Z.; Antonijević, B.; Đukić-Ćosić, D. Sulforaphane—A Compound with Potential Health Benefits for Disease Prevention and Treatment: Insights from Pharmacological and Toxicological Experimental Studies. Antioxidants 2024, 13, 147. [Google Scholar] [CrossRef]
- Fahey, J.; Zalcmann, A.; Talalay, P. The Chemical Diversity and Distribution of Glucosinolates and Isothiocyanates among Plants. Phytochemistry 2001, 56, 5–51. [Google Scholar] [CrossRef]
- Latté, K.P.; Appel, K.E.; Lampen, A. Health benefits and possible risks of broccoli – An overview. Food Chem. Toxicol. 2011, 49, 3287–3309. [Google Scholar] [CrossRef] [PubMed]
- Shan, Y.; Wu, K.; Wang, W.; et al. Sulforaphane down-regulates COX-2 expression by activating p38 and inhibiting NF-kappaB-DNA-binding activity in human bladder T24 cells. Int. J. Oncol. 2009, 34, 1129–1134. [Google Scholar] [CrossRef]
- Ruhee, R.T.; Roberts, L.A.; Ma, S.; Suzuki, K. Organosulfur Compounds: A Review of Their Anti-inflammatory Effects in Human Health. Front. Nutr. 2020, 7, 64. [Google Scholar] [CrossRef]
- Gu, J.; Cheng, Y.; Wu, H.; Kong, L.; Wang, S.; Xu, Z.; et al. Metallothionein is downstream of Nrf2 and partially mediates sulforaphane prevention of diabetic cardiomyopathy. Diabetes 2017, 66, 529–542. [Google Scholar] [CrossRef]
- Pan, J.; Wang, R.; Pei, Y.; Wang, D.; Wu, N.; Ji, Y.; et al. Sulforaphane alleviated vascular remodeling in hypoxic pulmonary hypertension via inhibiting inflammation and oxidative stress. J. Nutr. Biochem. 2023, 111, 109182. [Google Scholar] [CrossRef] [PubMed]
- Houghton, C.A. Sulforaphane: Its “Coming of Age” as a Clinically Relevant Nutraceutical in the Prevention and Treatment of Chronic Disease. Oxid. Med. Cell Longev. 2019, 2716870. [Google Scholar] [CrossRef]
- Zuo, M.; Chen, H.; Liao, Y.; He, P.; Xu, T.; Tang, J.; Zhang, N. Sulforaphane and bladder cancer: a potential novel antitumor compound. In Frontiers in Pharmacology, Vol. 14; Frontiers Media SA: 2023. [CrossRef]
- Ilic, D.; Nikolic, V.; Nikolic, L.; Stankovic, M.; Stanojevic, L.; Cakic, M. Allicin and related compounds: Biosynthesis, synthesis and pharmacological activity. Facta Univ. Ser. Phys. Chem. Technol. 2011, 9, 9–20. [Google Scholar] [CrossRef]
- El-Saber Batiha, G.; Magdy Beshbishy, A.; Wasef, L.G.; Elewa, Y.H.A.; Al-Sagan, A.A.; Abd El-Hack, M.E.; Taha, A.E.; Abd-Elhakim, Y.M.; Devkota, H.P. Chemical Constituents and Pharmacological Activities of Garlic (Allium sativum L.): A Review. Nutrients 2020, 12, 872. [Google Scholar] [CrossRef]
- Lang, A.; Lahav, M.; Sakhnini, E.; et al. Allicin inhibits spontaneous and TNF-alpha induced secretion of proinflammatory cytokines and chemokines from intestinal epithelial cells. Clin. Nutr. 2004, 23, 1199–1208. [Google Scholar] [CrossRef]
- Xu, C.; Mathews, A.E.; Rodrigues, C.; Eudy, B.J.; Rowe, C.A.; O’Donoughue, A.; Percival, S.S. Aged garlic extract supplementation modifies inflammation and immunity of adults with obesity: A randomized, double-blind, placebo-controlled clinical trial. Clin. Nutr. ESPEN 2018, 24, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Qaradakhi, T.; Gadanec, L.K.; McSweeney, K.R.; Abraham, J.R.; Apostolopoulos, V.; Zulli, A. The Anti-Inflammatory Effect of Taurine on Cardiovascular Disease. Nutrients 2020, 12, 2847. [Google Scholar] [CrossRef]
- Attiq, A.; Jalil, J.; Husain, K.; Ahmad, W. Raging the War Against Inflammation With Natural Products. Front. Pharmacol. 2018, 9, 976. [Google Scholar] [CrossRef]
- Lee, D.Y.; Li, H.; Lim, H.J.; Lee, H.J.; Jeon, R.; Ryu, J.H. Anti-inflammatory activity of sulfur-containing compounds from garlic. J. Med. Food 2012, 15, 992–999. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Prandelli, C.; Parola, C.; Buizza, L.; Delbarba, A.; Marziano, M.; Salvi, V.; Zacchi, V.; Memo, M.; Sozzani, S.; Calza, S.; Uberti, D.; Bosisio, D. Sulphurous thermal water increases the release of the anti-inflammatory cytokine IL-10 and modulates antioxidant enzyme activity. Int. J. Immunopathol. Pharmacol. 2013, 26, 633–646. [Google Scholar] [CrossRef]
- Sokrateva, T.; Ivanova, D.; Galunska, B.; Todorova, M.; Ivanov, D. Physicochemical analysis of Varna Basin mineral water. Surveying Geology & Mining Ecology Management (SGEM). [CrossRef]
- Zhang, Y.; Liu, X.; Ruan, J.; Zhuang, X.; Zhang, X.; Li, Z. Phytochemicals of garlic: Promising candidates for cancer therapy. Biomed. Pharmacother. 2020, 123, 109730. [Google Scholar] [CrossRef] [PubMed]
- Habtemariam, S. Anti-Inflammatory Therapeutic Mechanisms of Isothiocyanates: Insights from Sulforaphane. Biomedicines 2024, 12, 1169. [Google Scholar] [CrossRef] [PubMed]
- Swiderski, J.; Sakkal, S.; Apostolopoulos, V.; Zulli, A.; Gadanec, L.K. Combination of Taurine and Black Pepper Extract as a Treatment for Cardiovascular and Coronary Artery Diseases. Nutrients 2023, 15, 2562. [Google Scholar] [CrossRef] [PubMed]
- Miao, J.; Zheng, L.; Zhang, J.; Ma, Z.; Zhu, W.; Zou, S. The effect of taurine on the toll-like receptors/nuclear factor kappa B (TLRs/NF-κB) signaling pathway in Streptococcus uberis-induced mastitis in rats. Int. Immunopharmacol. 2011, 11, 1740–1746. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Y.; Sun, J.; Zhang, W.; Guo, Z.; Ma, Q. Arachidonic acid metabolism in health and disease. MedComm 2023, 4, e363. [Google Scholar] [CrossRef]
- Ju, Z.; Li, M.; Xu, J.; Howell, D.C.; Li, Z.; Chen, F.E. Recent development on COX-2 inhibitors as promising anti-inflammatory agents: The past 10 years. Acta Pharm. Sin. B 2022, 12, 2790–2807. [Google Scholar] [CrossRef]
- Jang, D.I.; Lee, A.H.; Shin, H.Y.; Song, H.R.; Park, J.H.; Kang, T.B.; Lee, S.R.; Yang, S.H. The Role of Tumor Necrosis Factor Alpha (TNF-α) in Autoimmune Disease and Current TNF-α Inhibitors in Therapeutics. Int. J. Mol. Sci. 2021, 22, 2719. [Google Scholar] [CrossRef]
- You, K.; Gu, H.; Yuan, Z.; Xu, X. Tumor Necrosis Factor Alpha Signaling and Organogenesis. Front. Cell Dev. Biol. 2021, 9, 727075. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Wu, Q. Sulforaphane protects intestinal epithelial cells against lipopolysaccharide-induced injury by activating the AMPK/SIRT1/PGC-1ɑ pathway. Bioengineered 2021, 12, 4349–4360. [Google Scholar] [CrossRef]
- Ruhee, R.T.; Ma, S.; Suzuki, K. Sulforaphane Protects Cells against Lipopolysaccharide-Stimulated Inflammation in Murine Macrophages. Antioxidants 2019, 8, 577. [Google Scholar] [CrossRef] [PubMed]
- Lv, C.; Wang, C.; Li, P.; Huang, Y.; Lu, X.; Shi, M.; Zeng, C.; Qin, S. Effect of Garlic Organic Sulfides on Gene Expression Profiling in HepG2 Cells and Its Biological Function Analysis by Ingenuity Pathway Analysis System and Bio-Plex-Based Assays. Mediators Inflamm. 2021, 7681252. [Google Scholar] [CrossRef]
- Ma, Y.; Zhang, Y.; Li, R.; Deng, S.; Qin, Q.; Ran, C.; Hao, Y.; Zhang, J.; Zhu, L. Mechanism of taurine reducing inflammation and organ injury in sepsis mice. Cell. Immunol. 2022, 375, 104503. [Google Scholar] [CrossRef] [PubMed]
- Barua, M.; Liu, Y.; Quinn, M.R. Taurine chloramine inhibits inducible nitric oxide synthase and TNF-alpha gene expression in activated alveolar macrophages: decreased NF-kappaB activation and IkappaB kinase activity. J. Immunol. 2001, 167, 2275–2281. [Google Scholar] [CrossRef]
- Shahid, A.; Bhatia, M. Hydrogen Sulfide: A Versatile Molecule and Therapeutic Target in Health and Diseases. Biomolecules 2024, 14, 1145. [Google Scholar] [CrossRef]
- Kaneko, N.; Kurata, M.; Yamamoto, T.; et al. The role of interleukin-1 in general pathology. Inflamm. Regener. 2019, 39, 12. [Google Scholar] [CrossRef] [PubMed]
- Ren, K.; Torres, R. Role of interleukin-1beta during pain and inflammation. Brain Res. Rev. 2009, 60, 57–64. [Google Scholar] [CrossRef]
- Wang, P.; Qian, H.; Xiao, M.; Lv, J. Role of signal transduction pathways in IL-1β-induced apoptosis: pathological and therapeutic aspects. Immun. Inflamm. Dis. 2023, 11, e762. [Google Scholar] [CrossRef]
- Chan, A.; Burgener, S.; Vezyrgiannis, K.; Wang, X.; Acklam, J.; Von Pein, J.; Pizzuto, M.; Labzin, L.; Boucher, D.; Schroder, K. Caspase-4 dimerisation and D289 auto-processing elicit an interleukin-1β-converting enzyme. Life Sci. Alliance 2023, 6, e202301908. [Google Scholar] [CrossRef]
- Luotola, K. IL-1 Receptor Antagonist (IL-1Ra) Levels and Management of Metabolic Disorders. Nutrients 2022, 14, 3422. [Google Scholar] [CrossRef]
- Villatoro, A.; Cuminetti, V.; Bernal, A.; et al. Endogenous IL-1 receptor antagonist restricts healthy and malignant myeloproliferation. Nat. Commun. 2023, 14, 12. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, J.M.; Duarte, S.; Ribeiro, A.C.; Mascarenhas, P.; Noronha, S.; Alves, R.C. Association between IL-1A, IL-1B and IL-1RN Polymorphisms and Peri-Implantitis: A Systematic Review and Meta-Analysis. Appl. Sci. 2022, 12, 6958. [Google Scholar] [CrossRef]
- Maculewicz, E.; Antkowiak, B.; Antkowiak, O.; et al. The interactions between interleukin-1 family genes: IL1A, IL1B, IL1RN, and obesity parameters. BMC Genomics 2022, 23, 112. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, L.A.; Dutreil, M.; Fattman, C.; Pandey, A.C.; Torres, G.; Go, K.; Phinney, D.G. Interleukin 1 receptor antagonist mediates the antiinflammatory and antifibrotic effect of mesenchymal stem cells during lung injury. Proc. Natl. Acad. Sci. USA 2007, 104, 11002–11007. [Google Scholar] [CrossRef]
Figure 1.
Effect of SMW (A), SFN (B), TAU (C) and A (D) on HIEC-6 cells viability. Data are presented as mean ± SD of three independent experiments; *p<0.05, **p<0.01 and ***p≤0.001, versus control (untreated cells).
Figure 1.
Effect of SMW (A), SFN (B), TAU (C) and A (D) on HIEC-6 cells viability. Data are presented as mean ± SD of three independent experiments; *p<0.05, **p<0.01 and ***p≤0.001, versus control (untreated cells).
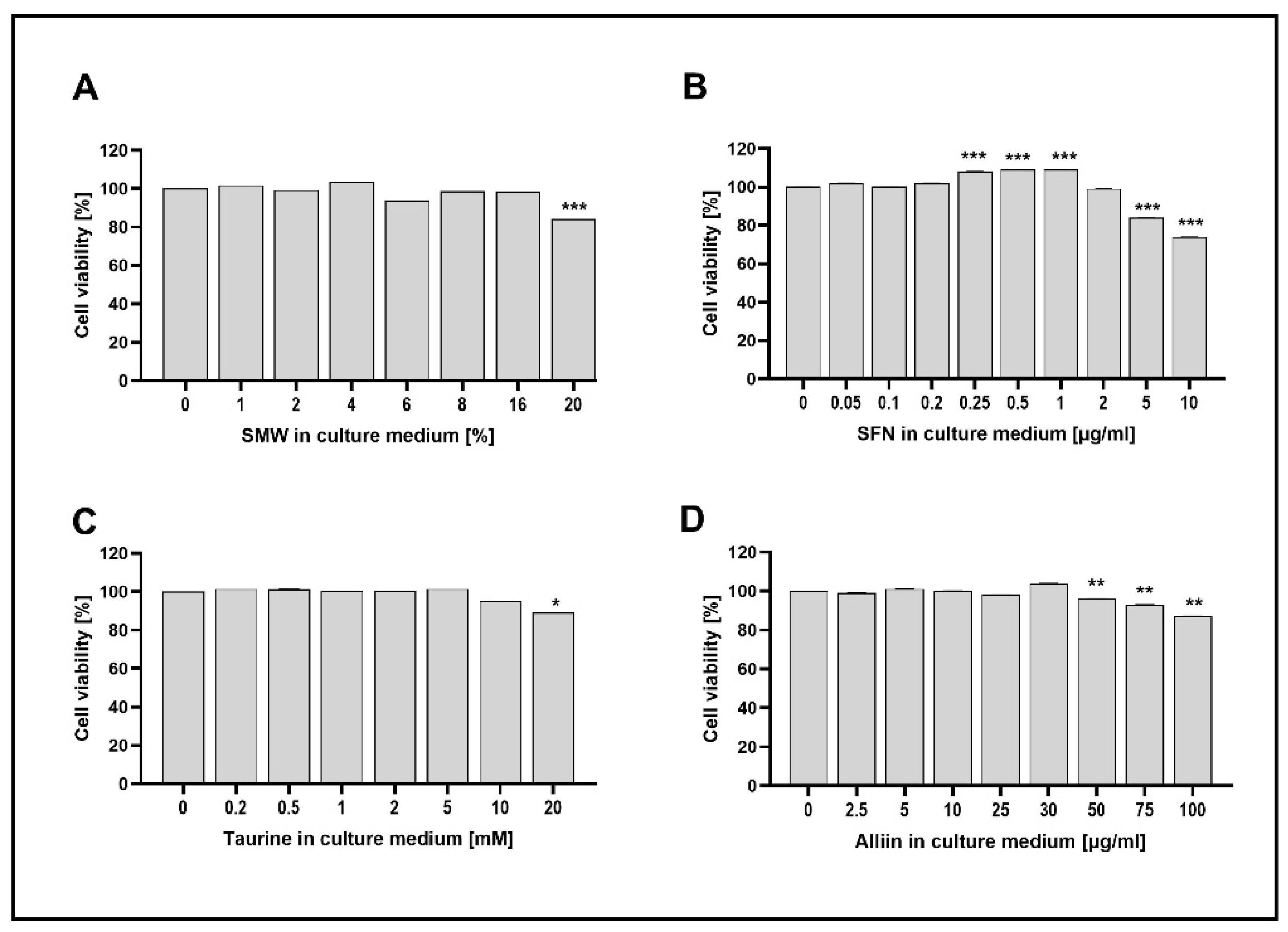
Figure 2.
Effect of SMW and SBAC on COX-2 expression in HIEC-6, under conditions of LPS-induced inflammation. Levels mRNA are presented as relative units (RU), with untreated cells – controls taken as 1; C – control (untreated cells); LPS – lipopolysaccharides; SA – salicylic acid; SMW4, SMW6, SMW8 – cells pretreated with 4%, 6% or 8% sulfur-containing mineral water, respectively (A); SFN0.5, SFN 1, SFN 2 – cells pretreated with sulforaphane 0.5, 1 or 2 µg/ml, respectively (B); T1, T2, T5 – cells pretreated with 1, 2 or 5 mM taurine, respectively (C); A10, A25, A30 – cells pretreated with 10, 25 or 30 µg/ml alliin (D); LPS ⁻/+ cells without/with LPS-induced inflammation (*p ≤ 0.05; **p ≤ 0.01; ***p ≤ 0.001 versus C).
Figure 2.
Effect of SMW and SBAC on COX-2 expression in HIEC-6, under conditions of LPS-induced inflammation. Levels mRNA are presented as relative units (RU), with untreated cells – controls taken as 1; C – control (untreated cells); LPS – lipopolysaccharides; SA – salicylic acid; SMW4, SMW6, SMW8 – cells pretreated with 4%, 6% or 8% sulfur-containing mineral water, respectively (A); SFN0.5, SFN 1, SFN 2 – cells pretreated with sulforaphane 0.5, 1 or 2 µg/ml, respectively (B); T1, T2, T5 – cells pretreated with 1, 2 or 5 mM taurine, respectively (C); A10, A25, A30 – cells pretreated with 10, 25 or 30 µg/ml alliin (D); LPS ⁻/+ cells without/with LPS-induced inflammation (*p ≤ 0.05; **p ≤ 0.01; ***p ≤ 0.001 versus C).
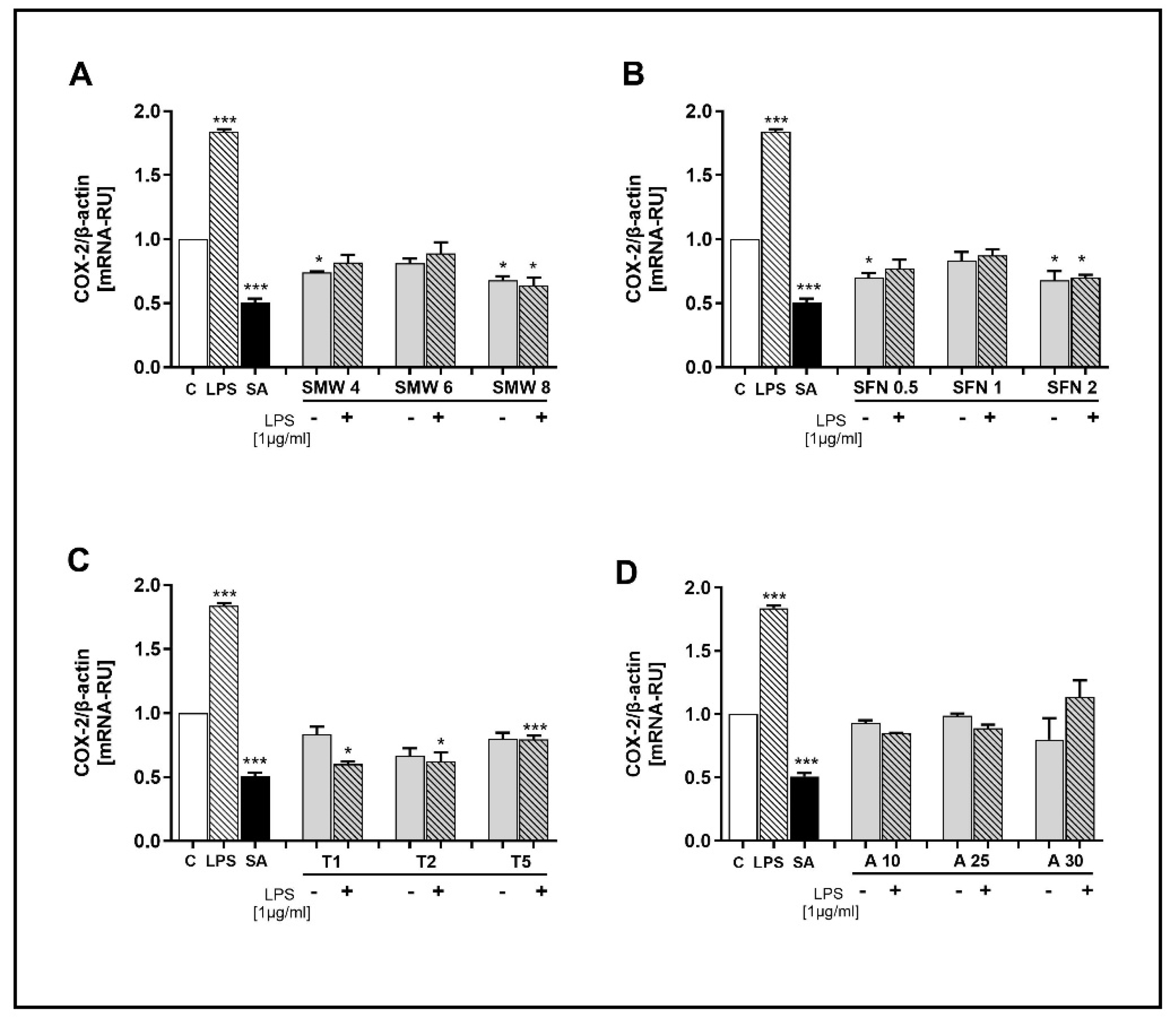
Figure 3.
Effect of SMW and SBAC on TNF-α expression in HIEC-6, under conditions of LPS-induced inflammation. Levels mRNA are presented as relative units (RU), with untreated cells – controls taken as 1; C – control (untreated cells); LPS – lipopolysaccharides; SA – salicylic acid; SMW4, SMW6, SMW8 – cells pretreated with 4%, 6% or 8% sulfur-containing mineral water, respectively (A); SFN0.5, SFN 1, SFN 2 – cells pretreated with sulforaphane 0.5, 1 or 2 µg/ml, respectively (B); T1, T2, T5 – cells pretreated with 1, 2 or 5 mM taurine, respectively (C); A10, A25, A30 – cells pretreated with 10, 25 or 30 µg/ml alliin (D); LPS ⁻/+ cells without/with LPS-induced inflammation (*p ≤ 0.05; **p ≤ 0.01; ***p ≤ 0.001 versus C).
Figure 3.
Effect of SMW and SBAC on TNF-α expression in HIEC-6, under conditions of LPS-induced inflammation. Levels mRNA are presented as relative units (RU), with untreated cells – controls taken as 1; C – control (untreated cells); LPS – lipopolysaccharides; SA – salicylic acid; SMW4, SMW6, SMW8 – cells pretreated with 4%, 6% or 8% sulfur-containing mineral water, respectively (A); SFN0.5, SFN 1, SFN 2 – cells pretreated with sulforaphane 0.5, 1 or 2 µg/ml, respectively (B); T1, T2, T5 – cells pretreated with 1, 2 or 5 mM taurine, respectively (C); A10, A25, A30 – cells pretreated with 10, 25 or 30 µg/ml alliin (D); LPS ⁻/+ cells without/with LPS-induced inflammation (*p ≤ 0.05; **p ≤ 0.01; ***p ≤ 0.001 versus C).
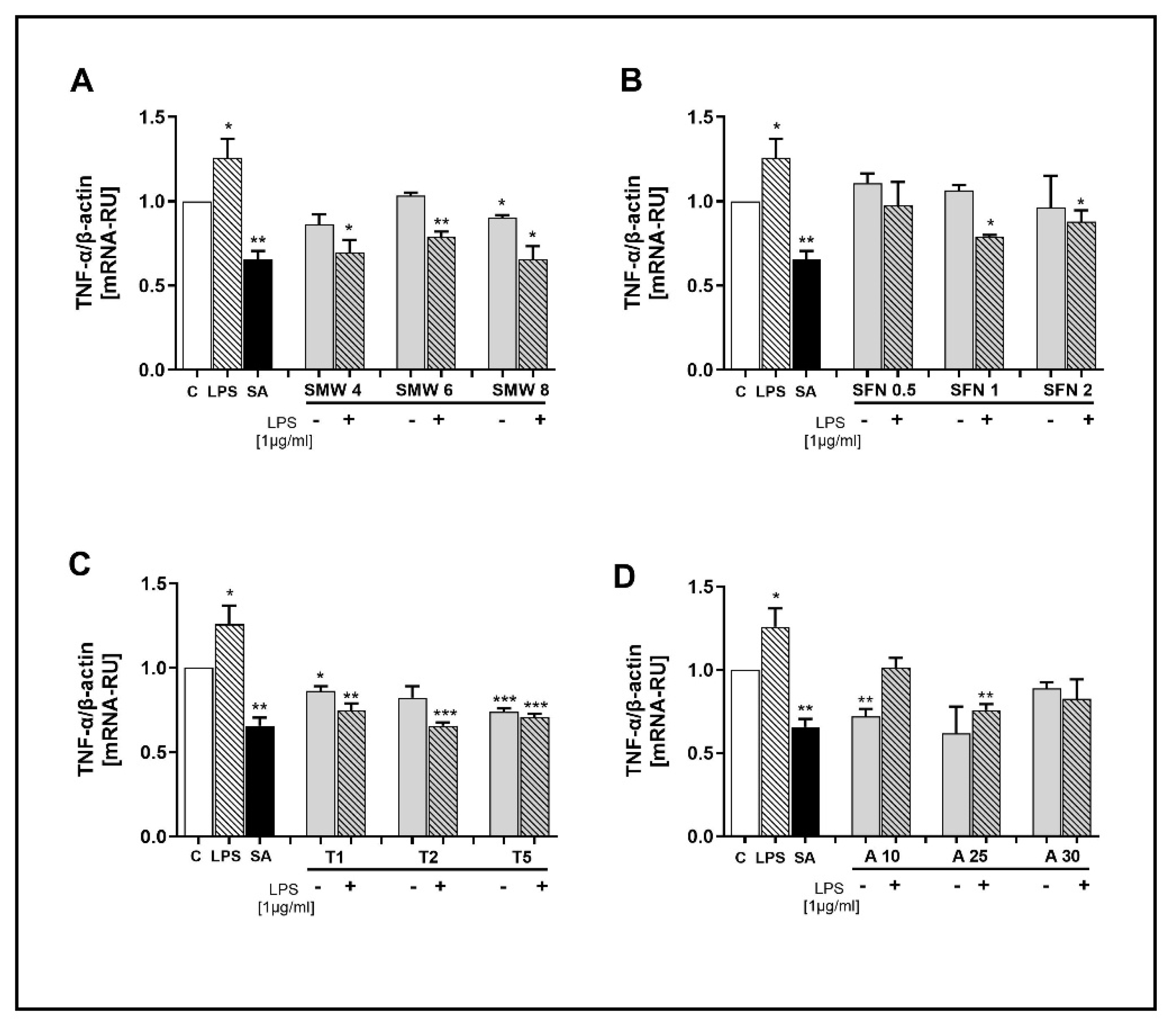
Figure 4.
Effect of SMW and SBAC on IL-1β expression in HIEC-6, under conditions of LPS-induced inflammation. Levels mRNA are presented as relative units (RU), with untreated cells – controls taken as 1; C – control (untreated cells); LPS – lipopolysaccharides; SA – salicylic acid; SMW4, SMW6, SMW8 – cells pretreated with 4%, 6% or 8% sulfur-containing mineral water, respectively (A); SFN0.5, SFN 1, SFN 2 – cells pretreated with sulforaphane 0.5, 1 or 2 µg/ml, respectively (B); T1, T2, T5 – cells pretreated with 1, 2 or 5 mM taurine, respectively (C); A10, A25, A30 – cells pretreated with 10, 25 or 30 µg/ml alliin (D); LPS ⁻/+ cells without/with LPS-induced inflammation (*p ≤ 0.05; **p ≤ 0.01; ***p ≤ 0.001 versus C).
Figure 4.
Effect of SMW and SBAC on IL-1β expression in HIEC-6, under conditions of LPS-induced inflammation. Levels mRNA are presented as relative units (RU), with untreated cells – controls taken as 1; C – control (untreated cells); LPS – lipopolysaccharides; SA – salicylic acid; SMW4, SMW6, SMW8 – cells pretreated with 4%, 6% or 8% sulfur-containing mineral water, respectively (A); SFN0.5, SFN 1, SFN 2 – cells pretreated with sulforaphane 0.5, 1 or 2 µg/ml, respectively (B); T1, T2, T5 – cells pretreated with 1, 2 or 5 mM taurine, respectively (C); A10, A25, A30 – cells pretreated with 10, 25 or 30 µg/ml alliin (D); LPS ⁻/+ cells without/with LPS-induced inflammation (*p ≤ 0.05; **p ≤ 0.01; ***p ≤ 0.001 versus C).
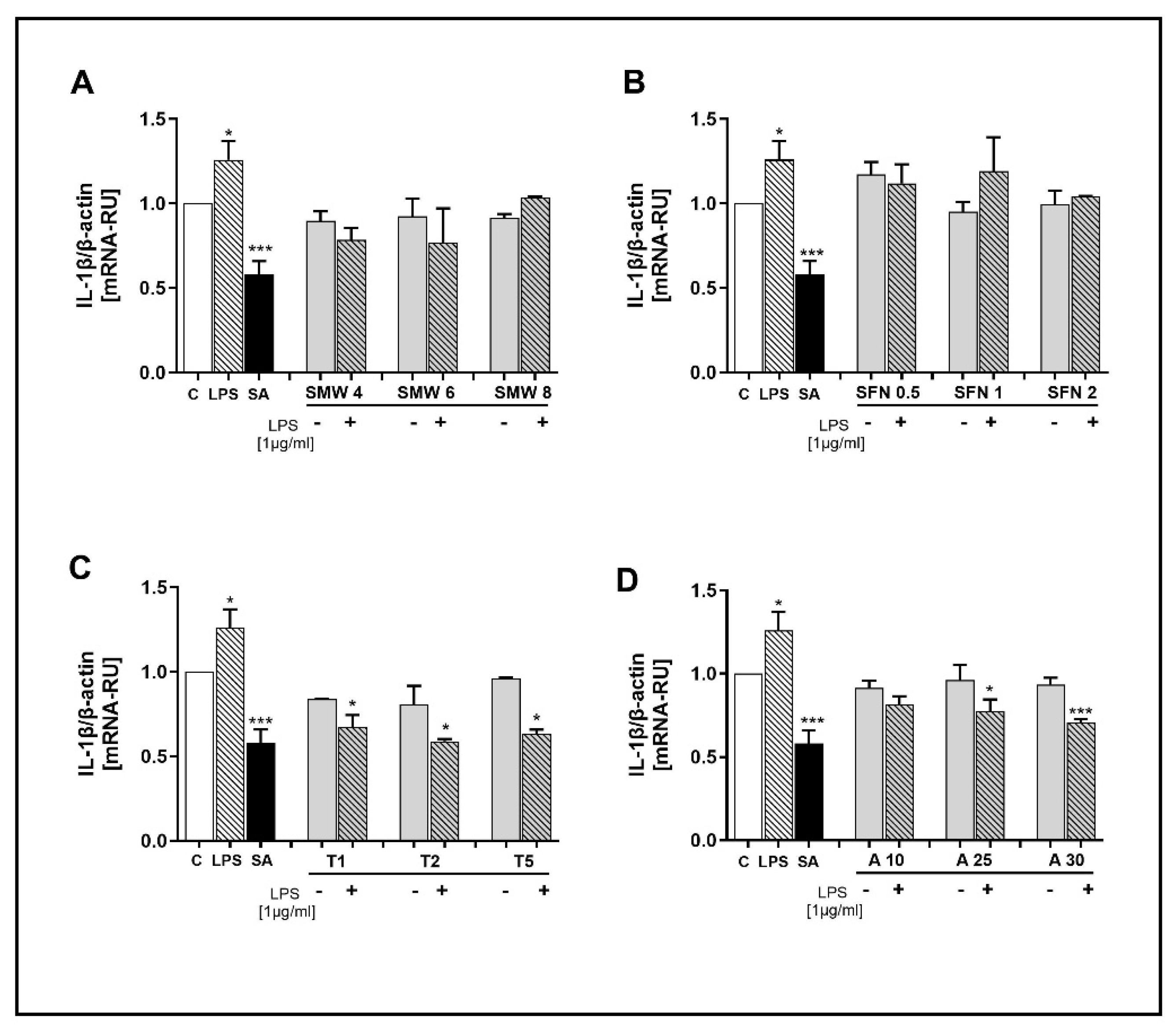
Figure 5.
Effect of SMW and SBAC on IL-1RN expression in HIEC-6, under conditions of LPS-induced inflammation. Levels mRNA are presented as relative units (RU), with untreated cells – controls taken as 1; C – control (untreated cells); LPS – lipopolysaccharides; SA – salicylic acid; SMW4, SMW6, SMW8 – cells pretreated with 4%, 6% or 8% sulfur-containing mineral water, respectively (A); SFN0.5, SFN 1, SFN 2 – cells pretreated with sulforaphane 0.5, 1 or 2 µg/ml, respectively (B); T1, T2, T5 – cells pretreated with 1, 2 or 5 mM taurine, respectively (C); A10, A25, A30 – cells pretreated with 10, 25 or 30 µg/ml alliin (D); LPS ⁻/+ cells without/with LPS-induced inflammation (*p ≤ 0.05; versus C).
Figure 5.
Effect of SMW and SBAC on IL-1RN expression in HIEC-6, under conditions of LPS-induced inflammation. Levels mRNA are presented as relative units (RU), with untreated cells – controls taken as 1; C – control (untreated cells); LPS – lipopolysaccharides; SA – salicylic acid; SMW4, SMW6, SMW8 – cells pretreated with 4%, 6% or 8% sulfur-containing mineral water, respectively (A); SFN0.5, SFN 1, SFN 2 – cells pretreated with sulforaphane 0.5, 1 or 2 µg/ml, respectively (B); T1, T2, T5 – cells pretreated with 1, 2 or 5 mM taurine, respectively (C); A10, A25, A30 – cells pretreated with 10, 25 or 30 µg/ml alliin (D); LPS ⁻/+ cells without/with LPS-induced inflammation (*p ≤ 0.05; versus C).
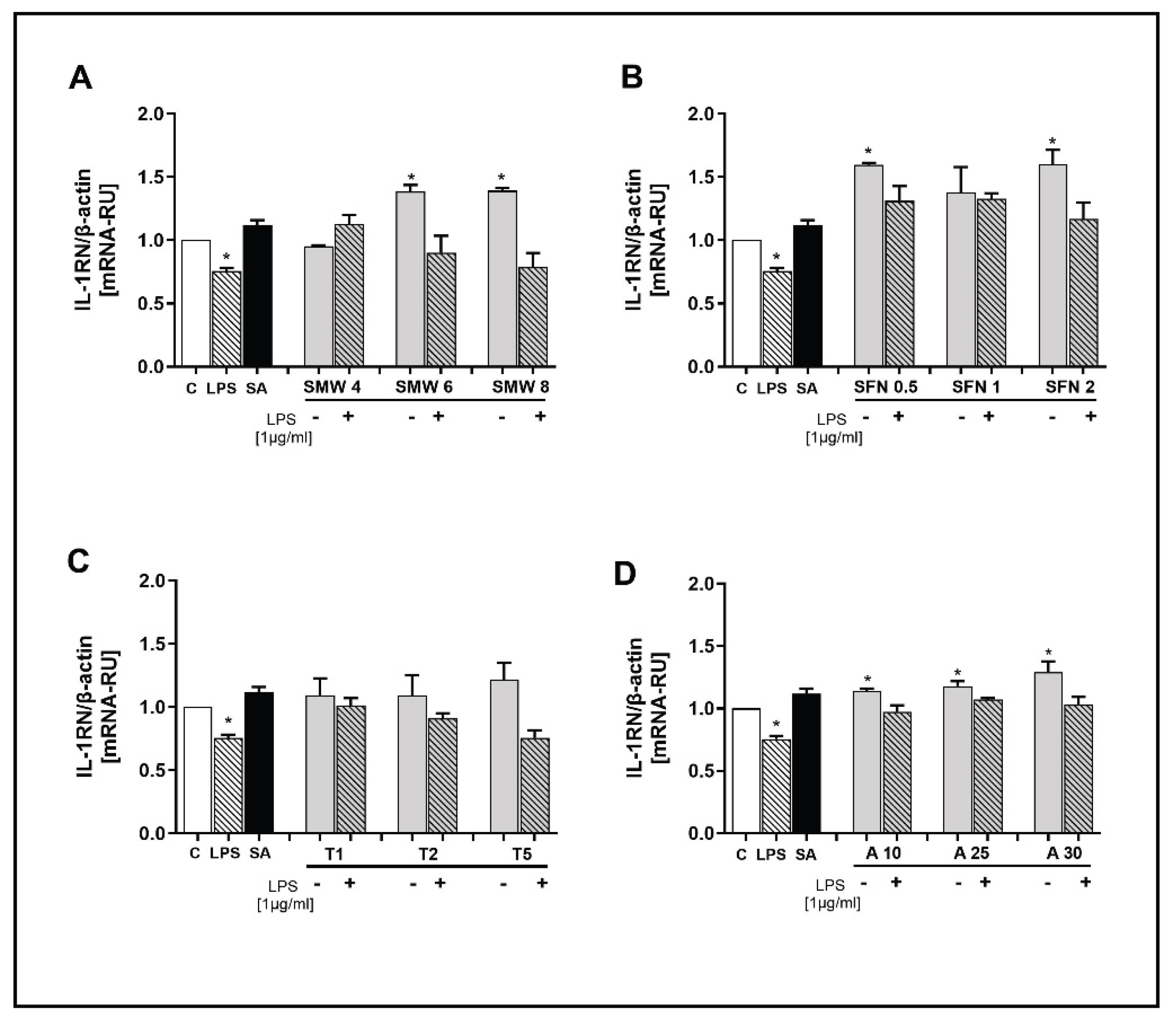

Table 1.
Primers nucleotide sequences.
| Target gene (Human) |
Nucleotide sequence (5`-3`) |
|---|---|
| Actin beta | F GTG GCC GAG GAC TTT GAT TGR CCT GTA ACA ACG CAT CTC ATA |
| COX-2 | F GAA ACA GAG AAG TTG GCA GCAR GGC AGG ATA CAG CTC CAC AG |
| TNF-α | F CTC TTC TGC CTG CTG CAC TTTR ATG GGC TAC AGG CTT GTC ACT |
| IL-1β | F CCA CCT CCA GGG ACA GGA TAR AGA ATT AGC AAG CTG CCA GGA |
| IL1RN | F GCC CAT CCT CAG GAC CTT TCR ATG TCC TAG CCA TCC CCA CT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



