
Submitted:
14 June 2025
Posted:
16 June 2025
You are already at the latest version
Abstract
Autism Spectrum Disorder (ASD) is a complex neurodevelopmental condition characterized by difficulty with social communication, behavior, and sensory integration. With its prevalence rising worldwide in recent decades, understanding and mitigating the origins of ASD has become a priority. Though its etiology is multifactorial, current research highlights two major contributors: genetic susceptibilities and environmental inflammatory exposures leading to oxidative stress during critical developmental periods. We explore how genetic variations, particularly those affecting cerebral folate metabolism, and various inflammatory triggers intersect to influence the development of ASD, each giving rise to specific symptoms seen in autism.
Keywords:
Autism
; ASD
; Leucovorin
; Folate
; Cerebral Folate Deficiency
; Inflammation
; Oxidative Stress
; Maternal Immune Activation
1. Introduction
Autism diagnosis has seen a dramatic rise in the 21st century, increasing approximately five-fold, and becoming a public health priority [1]. There are striking parallels between ASD and other multifactorial diseases such as cancer. In both conditions, risk is influenced by an interplay of genetic predispositions and environmental factors, particularly triggers of inflammation, which lead to oxidative stress. Just as cancer risk can be increased by inherited mutations and external exposures like toxins or infections, ASD may also arise from a combination of genetic vulnerability and environmental stressors. In both diseases, these factors modulate susceptibility rather than determining certainty of onset, effectively raising the probability of development becoming neurodivergent.
Furthermore, recent findings in both fields suggest an important role for the gut microbiome in modulating immune and neurological outcomes. Nutritional and dietary choices appear to influence disease trajectories in both ASD and cancer. For example, diets that reduce inflammation or support detoxification pathways are known to reduce cancer risk and may contribute to better outcomes in ASD as well.
Another similarity with cancer is that ASD is presumed to have multiple variants, with differing etiologies. To date, over 70 genes have been correlated with ASD, with hundreds more believed to be related to ASD susceptibility [2,3,4]. the wide array of genes may help explain the diversity of expression of ASD, as well as point to a rationale for some of the comorbidities commonly diagnosed with ASD [5].
These insights invite a new perspective on ASD, not just as a static diagnosis, but as a dynamic condition potentially influenced by modifiable risk factors. This opens the door to discussing strategies for reducing risk, and even the possibility of partial or complete remission under certain therapeutic regimens. Understanding the contributing causes of ASD could lead to more personalized, preventive, and responsive treatments.
2. Genetic Factors in ASD
Many genes have been implicated as having a role in autism, with multiple categories involved [6,7,8]. There is evidence from brain organoids of a form of idiopathic autism that some of the genes involved lead to an imbalance in excitatory cortical neurons [9]. Given that there are presumed to be several autism variants, the associated genes are ones that are correlated with a greater risk of developing ASD. Some have specific pathways that are involved and others are correlated with ASD but in a non-defined manner. Section 2.3 and 2.4 provide a brief look at some of the developmental genes that have traditionally been considered for their role in ASD.
Genes associated with folate transport (Section 2.1) are related to folate delivery to the brain and a biomarker for cerebral folate deficiency (CFD). Therapeutic intervention can overcome CFD, as described in this section. A potentially related genetic risk is for mitochondrial dysfunction (Section 2.2), which may play a role in the autism regression seen in some children as they grow.
2.1. Cerebral Folate Deficiency
2.1.1. Folate Metabolism and Brain Development
Folate is essential for DNA synthesis, repair, and methylation, all of which are essential processes during early brain development. The transport of folate into the central nervous system (CNS) is tightly regulated by specific transport mechanisms, primarily the folate receptor alpha (FRα) and the reduced folate carrier (RFC).
2.1.2. Genetic Variants Impairing Folate Transport
Mutations in the FOLR1 gene can impair FRα function, leading to CFD. Similarly, polymorphisms such as the SLC19A1 variant rs1051266 can reduce RFC efficiency. Both impair the brain’s ability to maintain adequate folate levels, resulting in developmental impairments and ASD features, such as decreased social communication and decreased emotional recognition [10,11].
2.1.3. Folate Receptor Alpha Autoantibodies (FRAAs)
FRAAs, which block or bind to FRα, are much more prevalent in ASD children (about 70% of ASD children, compared with 5-10% in the general population). These autoantibodies suppress folate transport across the blood-brain barrier, contributing to CFD. FRAAs are believed to result from environmental exposure,[12] further linking genetic and inflammatory pathways.
2.1.4. Therapeutic Interventions
Folinic acid supplementation has shown promise in reversing CFD symptoms in children with ASD in multiple clinical trials (based in USA, France, or India) published between 2018-2025 [13,14,15,16]. Dietary modifications, such as avoiding cow’s milk products (which may trigger FRAAs),[12] have also demonstrated potential benefits. These findings support the clinical importance of identifying and treating CFD in ASD patients.
2.2. Mitochondrial Dysfunction
There is growing evidence that mitochondrial dysfunction may contribute to neurodevelopmental regression in children [17,18,19,20]. This condition is seen when an ASD child shows strong learning, language and memory initially, only to regress later in childhood [21]. Of note, while non-regressed children showed inflammation and immune dysregulation occurs first, while in regressed children mitochondrial dysfunction is first observed, with an impact on immune activity possibly occurring later [19].
2.3. Synapse and Signaling Genes
2.4. Regulation, Cell Growth and Plasticity Genes
3. Inflammation During Critical Developmental Periods and ASD Risk
3.1. The Developing Brain and Vulnerability to Inflammation
Neurodevelopment is highly sensitive to environmental insults during prenatal and early postnatal periods. Inflammation during these times can disrupt neuronal migration, synaptogenesis, synaptic pruning, and brain connectivity, all of which are critical to neurotypical brain function [32]. Many causes of inflammation have been documented, some of which are acute, such as viral infection and fever, and others are chronic, such as microplastics and malnutrition. When inflammation occurs during critical periods in brain development, then neurotypical development can be delayed or arrested, resulting in a child reaching milestones later, or aberrantly, or possibly not at all if the inflammation exceeds the duration of the critical period [32,33,34,35]. Summary schematics of the categories of inflammation and their presumed action are outlined in Figure 1 and Figure 2.
3.2. Categories of Inflammation
3.2.1. Viral Infections
Maternal infections such as influenza, rubella, or COVID-19 have been linked to increased ASD risk due to maternal immune activation (MIA). Postnatal viral infections can also contribute to neuroinflammatory states [33,35,36,37,38].
Evaluation of the studies in a meta-analysis shows there is an association between maternal infection and subsequent autism diagnosis in the child, with little variation based on when the infection occurred during pregnancy or what type of infection it was [39,40]. A genetic analysis showed the increased risk for ASD with maternal infections may be correlated with a genetically distinct subtype of autism, a subtype that come from the interaction between genetic susceptibility and exposure to infections in pregnancy [41]. Thus, since the maternal infections may not be causal in the development of ASD, it is unclear if prevention of infection may reduce autism incidence [42].
3.2.2. Air Pollution & Maternal Asthma
Environmental pollutants are associated with ASD incidence [43]. Exposure to particulate matter (e.g., PM2.5) during pregnancy has been associated with neuroinflammation[44] and developmental disorders, with an increased risk of development of ASD [32,44,45,46,47,48,49]. Airborne toxins can activate microglia and oxidative stress pathways in the fetal brain [50].
Additional studies have examined air pollution from roadways as being particularly problematic, pointing to a potential link to plasticizers used in the manufacture of tires and tailpipe exhaust, as risk was related to distance from roadways [51,52]. Given that immune dysregulation is a common comorbid feature with ASD, the link to maternal exposure to air pollution may be indicating a mechanism of action in the development of autism. In this regard, multiple studies have found elevated risk of autism in infants born to asthmatic mothers, and that this risk is related to an increase in inflammatory biomarkers [53,54,55].
Of concern, in a study of air pollution pre-conception, women exposed to particulate matter and nitric oxide in the three months prior to conception had children with greater growth in body weight in their first two years of life [56]. The authors report that the critical period of exposure is the three months prior to conception.
3.2.3. Maternal Immune Activation
Maternal inflammation during pregnancy is associated with an increased risk of ASD, with this maternal immune activation (MIA) being evaluated in animal models, where it leads to behavioral changes resembling ASD [57]. The immune activation raises levels of cytokines, especially interleukin-6 (IL-6) and interleukin 17a (IL-17a), which are implicated in disrupting fetal brain development, in what may lead to some ASD behaviors. The effect of the MIA on the developing fetus may be impacted by the fetus’ genetic makeup, with certain genetic profiles making the fetal brain more impacted by the maternal inflammation, implicating a gene-environment interaction in the etiology of ASD [57,58,59,60].
3.2.4. Microplastics
Emerging studies in animal models suggest that prenatal and early postnatal exposure to microplastics, especially bisphenol A and phthalates, may lead to inflammation-driven behavioral changes reminiscent of ASD [61,62,63,64,65]. The action may be through immune and epigenetic mechanisms, or as endocrine disrupters. In this regard, plastic food containers and films were tested and found to contain thousands of chemicals, with many being endocrine disruptors (including an estrogen receptor alpha activator, and an androgen receptor inhibitor) and metabolism disruptors [66]. Samples tested that were made with fewer chemicals also had fewer toxic chemicals that entered the food. Additionally, exposure to polystyrene nanoplastics causes depression and anxiety in a mouse model,[67] and it decreased oligodendrocyte activity and enhanced hyperactivity and aggression in zebrafish,[68] while exposure to polyethelene led to repetitive behaviors and diminished social interaction [69].
3.2.5. Malnutrition
Deficiencies in nutrients with anti-inflammatory properties, such as omega-3 fatty acids, zinc, and vitamins B12, B9, A, D, and K may increase susceptibility to inflammation and neurodevelopmental disorders [70,71,72,73,74,75]. The vitamin A deficiency may have its effect by impact on the gut microbiome [76,77]. Other micronutrients appear to play important roles in neurotypical development [78]. There is also evidence that autistic children, who frequently eat the same food each day, are lacking in certain nutrients,[79,80] or are receiving an abundance of one macronutrient to the exclusion of others, which may contribute to symptoms [81].
3.2.6. Emotional Stress
Maternal stress during pregnancy can result in elevated cortisol and pro-inflammatory cytokines, potentially disrupting fetal brain development. Chronic stress in the early life of the child may also affect immune and neurological outcomes [39,82,83,84,85,86]. One measure of such stress on the child can be quantified by evaluating adverse childhood experiences (ACEs), as ASD children who are at increased risk of bullying, parental divorce, income and food insecurity, have an increased risk of comorbid health disorders [87].
3.2.7. Pharmaceuticals
Certain medications, particularly selective serotonin reuptake inhibitors (SSRIs), have been implicated in altering fetal brain development when used during pregnancy [88]. SSRIs cross the placental barrier and can influence serotonin signaling pathways critical to neurodevelopment. Research suggests that prenatal SSRI exposure may disrupt synaptogenesis and increase neuroinflammation, particularly in genetically susceptible offspring [89]. These effects may contribute to ASD-like phenotypes in animal models and warrant careful consideration of risk-benefit ratios in clinical practice [88,90,91,92].
3.2.8. Maternal Disorders, Diabetes
A recent study in California found higher ASD incidence for children born to mothers with asthma or obesity, and a much higher incidence for mothers with both conditions [93]. The authors speculate that the incidence increase may be due to the increased inflammatory conditions during pregnancy, and that earlier screening of children born to mothers with asthma and obesity during pregnancy may be warranted.
Meta-analysis of over 200 studies, and including over 50 million pregnancies, has revealed that children born to mothers with diabetes have an increased risk of neurodevelopmental disorders. This was especially true for mothers with pre-gestational diabetes, and less so for gestational diabetes. There was a 25-30% increased risk for ASD, ADHD and intellectual disability, with a 15-20% increased risk for communication learning disorders [94].
3.2.9. Synthetic Vitamins
Excessive or poorly metabolized synthetic folic acid may provoke immune responses, especially in individuals with impaired folate pathways. This imbalance can exacerbate inflammation and contribute to FRAA development, which leads to CFD. Additionally, intake of folic acid in excess of a few hundred micrograms per day (the typical level of folic acid that can be converted to usable folate in the gut) has been shown to result in unmetabolized folic acid in the blood, which interferes with absorption of folate across the blood brain barrier or across the placenta [95,96,97,98].
3.2.10. Vaccines
While the scientific consensus supports vaccine safety, there is speculation that vaccines trigger inflammatory responses in genetically susceptible individuals, as detailed in a recent comprehensive report [99]. If such inflammation occurs during critical periods, it can elevate risk of altering brain development. This was a larger concern in previous decades, for example when the earlier pertussis vaccine (in the 1980s) triggered fever in some children, or when the thiomersal (thimerosal) preservative in the measles vaccine was used (until 2001) that could be inflammatory in some children. The fact that diagnosed autism cases has increased five-fold from 2000-2025[1,100] while inflammation from vaccines has diminished indicates this is likely not a significant factor in this neurodevelopmental disorder.
3.2.11. Microbiome and Metabolic Disorders
This topic may merit a separate category, as processing by the microbiome seems to be essential for absorption of key nutrients as well as inactivation of inflammatory agents, and digestive disorders are frequently comorbid with ASD [100,101,102,103]. In this regard, restoring microbiome function with probiotics may alleviate some ASD symptoms,[104] and use of fecal transfer has shown some effectiveness in treating autism symptoms [105].
Several studies have shown a link between the gut microbiome and neurological disorders [106]. Propionic acid, a microbial byproduct, is known to cause neuroinflammation and over-proliferation of glia in mice offspring born to mothers exposed to propionic acid through pregnancy and weaning [107]. This provides direction as to a mechanism of action by which inflammation may be caused and neural correlates of its action.
Screening of the gut microbiome of autistic children and non-autistic siblings revealed categories of bacteria and fungi that were at different levels in the ASD individuals [108]. When these were fed to mice, there was a concomitant change in behavioral patterns, indicating the use of certain probiotics may provide some benefit in autism. A similar study found differences in ASD and non-ASD microbiomes, and observed that ASD regression children had higher levels of Proteobacteria [109]. This link to ASD regression may provide another avenue to explore.
Additional evaluation of mitochondrial metabolism finds changes in energy pathways in ASD children [110]. when metabolic analysis is made of cord blood, to look at such parameters at birth, and in maternal mid-gestational blood, a machine-learning analysis found that cord blood was more predictive of ASD and that this was true for girls and boys [111]. A later study showed that elevation of a specific cord blood component, acylcarnitine, was predictive of both ASD and ADHD outcomes [112].
3.3. Mechanistic Insights
Inflammation can alter neural circuitry by affecting synaptic pruning, neuronal connectivity, and neurotransmitter systems, and this may occur due to alterations in the neural immune system. Genetic predispositions, such as those affecting folate metabolism, may intensify the brain’s vulnerability to these inflammatory insults. Microglial priming and sustained cytokine exposure during development may create a neuroimmune environment conducive to ASD [37,113,114,115,116,117].
3.4. Therapeutic Interventions
Therapeutic intervention to reduce neuroinflammation typically involves reducing exposure to the causative agents. Given the great diversity of such potential sources, and the rampant exposure in daily living to most of these agents, such action is likely to be futile. Some things clearly can be done, such as reducing exposure to those agents that are one has control over, and seeking medical attention reduce risk from illnesses during pregnancy.
Additionally, where mechanisms are posited for inflammatory action, and when the impact of such action can be ameliorated, then use of specific products may provide benefit. In this regard, with maternal infection there is a concomitant reduction in N-acetyl cysteine and taurine associated with neuroinflammation due to a decrease in microglial cells. Each of these compounds can be supplemented into diet to reduce impact on the brain’s immune function to deter fetal injury.
There is a recent report in a mouse model of autism that an extract of basil, Ocimum basilicum, reduced damage in the pups that was induced by maternal separation, and reduced autism behaviors via antioxidant and anti-inflammatory properties of the extract [118]. This opens an avenue to considering other proven antioxidants in an ASD model. In this regard, the comparison with cancer may be apt, as there are decades of research identifying mechanisms of action of antioxidant nutritive extracts on cancer cell cultures [119,120]. Some of these may also prove to have an impact in deferring ASD development.
4. Integrative Perspective: Gene-Environment Interactions in ASD
4.1. Synergistic Effects
Genetic susceptibilities, such as impaired folate transport (described above), may amplify the effects of environmental insults. For example, exposure to synthetic folic acid or infections in a child with FOLR1 or RFC polymorphisms may result in a greater risk of ASD due to insufficient levels of natural folate reaching the brain or the uterus [74,98,117].
4.2. Epigenetic Modifications
Disruptions in folate metabolism may lead to altered DNA methylation and gene expression, affecting neurodevelopmental outcomes. These epigenetic changes may bridge the gap between genetic and environmental risk factors.
4.3. Implications for Prevention and Treatment
Screening for CFD in the fetus by testing for parental FRAAs, nutritional support (including prenatal vitamins with natural folate instead of folic acid), and inflammation-reducing strategies during pregnancy could reduce ASD risk [35].
The use of machine learning systems to successfully identify ASD in children could provide much earlier diagnosis [121,122]. Such machine learning provides hope that a similar system may be devised that can identify future ASD risk during pregnancy.
Personalized approaches considering both genetic background and environmental exposures hold promise for future interventions. Follow on care in early childhood may successfully identify developing ASD risk, and reduce severity autism severity by treating CFD and nutritional deficits at earlier ages, and by reducing inflammatory stimuli or reducing widespread neural inflammation.
We project that ASD treatment may parallel other multifactorial disorders, such as cancer, with a targeted approach to lessen risk and improve neurotypical outcomes, with a similar goal of extended remission to provide an outcome of raised qualify of life. Given that such treatment has a lower societal and health maintenance cost, and has been demonstrated to have no deleterious effects, we propose engaging such a model of treatment now.
5. Conclusions
ASD arises from a complex interplay between genetic and environmental factors. Genetic variants impairing folate metabolism and inflammatory exposures during critical periods each play significant roles in disrupting brain development. Understanding and addressing these risk factors through early detection, targeted interventions, and public health strategies may help reduce the incidence and severity of ASD in future generations.
Identifying the genetic causes and treating their ramifications, such as treating CFD with levo-leucovorin, has been shown repeatedly to make a substantial difference in resolving the communication difficulties of a young autistic child. It is time to have this become the standard of practice for all ASD children at the earliest ages.
Addressing inflammation causes can also be accomplished now, focusing on the most readily addressable ones first. In this way, reducing infection in pregnancy, eliminating malnutrition of mothers and children, and find ways to alleviate stress may have a lasting impact in reducing the severity of autism. And providing support during inflammation to help retain immune functioning, such as has been reported by using N-acetyl cysteine and taurine to replenish their levels when depleted by inflammation, may be an appropriate next step. It is most important to address the readily resolvable inflammation causes, specifically stress, malnutrition and health protection from disease now, and to extend efforts to work as a society to reduce the impact of inflammatory agents such as microplastics and pollution in order to provide the healthiest environment for neurotypical brain development in all children, and thus build a better and more productive future for all.
Funding
This research received no external funding
Institutional Review Board Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Conflicts of Interest
The author declares no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| ASD | Autism Spectrum Disorder |
| CNS | Central Nervous System |
| FRα | Folate Receptor alpha |
| RFC | Reduced Folate Carrier |
| FRAA | Folate Receptor Auto Antibody |
| CFD | Cerebral Folate Deficiency |
| MIA | Maternal Immune Activation |
| SSRI | Selective Serotonin Reuptake Inhibitor |
References
- Shaw KA. Prevalence and Early Identification of Autism Spectrum Disorder Among Children Aged 4 and 8 Years — Autism and Developmental Disabilities Monitoring Network, 16 Sites, United States, 2022. MMWR Surveill Summ. 2025;74. [CrossRef]
- Gogate A, Kaur K, Khalil R, et al. The genetic landscape of autism spectrum disorder in an ancestrally diverse cohort. Npj Genomic Med. 2024;9(1):1-22. [CrossRef]
- Havdahl A, Niarchou M, Starnawska A, Uddin M, van der Merwe C, Warrier V. Genetic contributions to autism spectrum disorder. Psychol Med. 51(13):2260-2273. [CrossRef]
- Fu JM, Satterstrom FK, Peng M, et al. Rare coding variation provides insight into the genetic architecture and phenotypic context of autism. Nat Genet. 2022;54(9):1320-1331. [CrossRef]
- Khachadourian V, Mahjani B, Sandin S, et al. Comorbidities in autism spectrum disorder and their etiologies. Transl Psychiatry. 2023;13(1):71. [CrossRef]
- Rolland T, Cliquet F, Anney RJL, et al. Phenotypic effects of genetic variants associated with autism. Nat Med. 2023;29(7):1671-1680. [CrossRef]
- Manoli DS, State MW. Autism Spectrum Disorder Genetics and the Search for Pathological Mechanisms. Am J Psychiatry. 2021;178(1):30-38. [CrossRef]
- Wan L, Yang G, Yan Z. Identification of a molecular network regulated by multiple ASD high risk genes. Hum Mol Genet. 2024;33(13):1176-1185. [CrossRef]
- Jourdon A, Wu F, Mariani J, et al. Modeling idiopathic autism in forebrain organoids reveals an imbalance of excitatory cortical neuron subtypes during early neurogenesis. Nat Neurosci. 2023;26(9):1505-1515. [CrossRef]
- Ayoub G. Neurodevelopment of Autism: Critical Periods, Stress and Nutrition. Cells. 2024;13(23):1968. [CrossRef]
- Qiu S, Qiu Y, Li Y, Cong X. Genetics of autism spectrum disorder: an umbrella review of systematic reviews and meta-analyses. Transl Psychiatry. 2022;12(1):249. [CrossRef]
- Ramaekers VT, Sequeira JM, Blau N, Quadros EV. A milk-free diet downregulates folate receptor autoimmunity in cerebral folate deficiency syndrome. Dev Med Child Neurol. 2008;50(5):346-352. [CrossRef]
- Frye RE, Slattery J, Delhey L, et al. Folinic acid improves verbal communication in children with autism and language impairment: a randomized double-blind placebo-controlled trial. Mol Psychiatry. 2018;23(2):247-256. [CrossRef]
- Renard E, Leheup B, Guéant-Rodriguez RM, Oussalah A, Quadros EV, Guéant JL. Folinic acid improves the score of Autism in the EFFET placebo-controlled randomized trial. Biochimie. 2020;173:57-61. [CrossRef]
- Panda PK, Sharawat IK, Saha S, Gupta D, Palayullakandi A, Meena K. Efficacy of oral folinic acid supplementation in children with autism spectrum disorder: a randomized double-blind, placebo-controlled trial. Eur J Pediatr. 2024;183(11):4827-4835. [CrossRef]
- Soetedjo F, Kristijanto JA, Durry F. Folinic acid and autism spectrum disorder in children: A systematic review and meta-analysis of two double-blind randomized placebo-controlled trials. AcTion Aceh Nutr J. 2025;10:194. [CrossRef]
- Frye RE, McCarty PJ, Werner BA, Rose S, Scheck AC. Bioenergetic signatures of neurodevelopmental regression. Front Physiol. 2024;15:1306038. [CrossRef]
- Frye RE. Mitochondrial Dysfunction in Autism Spectrum Disorder: Unique Abnormalities and Targeted Treatments. Semin Pediatr Neurol. 2020;35:100829. [CrossRef]
- Gevezova M, Ivanov Z, Pacheva I, et al. Bioenergetic and Inflammatory Alterations in Regressed and Non-Regressed Patients with Autism Spectrum Disorder. Int J Mol Sci. 2024;25(15):8211. [CrossRef]
- Scott O, Shi D, Andriashek D, Clark B, Goez HR. Clinical clues for autoimmunity and neuroinflammation in patients with autistic regression. Dev Med Child Neurol. 2017;59(9):947-951. [CrossRef]
- The Mystery of Regressive Autism—Mental Health. Accessed May 21, 2025. https://www.enotalone.com/article/mental-health/the-mystery-of-regressive-autism-r20878/.
- Betancur C, Buxbaum JD. SHANK3 haploinsufficiency: a “common” but underdiagnosed highly penetrant monogenic cause of autism spectrum disorders. Mol Autism. 2013;4(1):17. [CrossRef]
- Glessner JT, Wang K, Cai G, et al. Autism genome-wide copy number variation reveals ubiquitin and neuronal genes. Nature. 2009;459(7246):569-573. [CrossRef]
- Jamain S, Quach H, Betancur C, et al. Mutations of the X-linked genes encoding neuroligins NLGN3 and NLGN4 are associated with autism. Nat Genet. 2003;34(1):27-29. [CrossRef]
- Sanders SJ, Campbell AJ, Cottrell JR, et al. Progress in Understanding and Treating SCN2A-Mediated Disorders. Trends Neurosci. 2018;41(7):442-456. [CrossRef]
- Trifonova EA, Mustafin ZS, Lashin SA, Kochetov AV. Abnormal mTOR Activity in Pediatric Autoimmune Neuropsychiatric and MIA-Associated Autism Spectrum Disorders. Int J Mol Sci. 2022;23(2):967. [CrossRef]
- Curatolo P, Moavero R, Vries PJ de. Neurological and neuropsychiatric aspects of tuberous sclerosis complex. Lancet Neurol. 2015;14(7):733-745. [CrossRef]
- Panwar V, Singh A, Bhatt M, et al. Multifaceted role of mTOR (mammalian target of rapamycin) signaling pathway in human health and disease. Signal Transduct Target Ther. 2023;8(1):1-25. [CrossRef]
- Bernier R, Golzio C, Xiong B, et al. Disruptive CHD8 Mutations Define a Subtype of Autism Early in Development. Cell. 2014;158(2):263-276. [CrossRef]
- Amir RE, Van den Veyver IB, Wan M, Tran CQ, Francke U, Zoghbi HY. Rett syndrome is caused by mutations in X-linked MECP2, encoding methyl-CpG-binding protein 2. Nat Genet. 1999;23(2):185-188. [CrossRef]
- Bassell GJ, Warren ST. Fragile X Syndrome: Loss of Local mRNA Regulation Alters Synaptic Development and Function. Neuron. 2008;60(2):201-214. [CrossRef]
- Yenkoyan K, Mkhitaryan M, Bjørklund G. Environmental Risk Factors in Autism Spectrum Disorder: A Narrative Review. Curr Med Chem. 2024;31(17):2345-2360. [CrossRef]
- Usui N, Kobayashi H, Shimada S. Neuroinflammation and Oxidative Stress in the Pathogenesis of Autism Spectrum Disorder. Int J Mol Sci. 2023;24(6):5487. [CrossRef]
- Chen Y, Du X, Zhang X, et al. Research trends of inflammation in autism spectrum disorders: a bibliometric analysis. Front Immunol. 2025;16:1534660. [CrossRef]
- Ayoub G. Neurodevelopment of Autism: Critical Periods, Stress and Nutrition. Cells. 2024;13(23):1968. [CrossRef]
- Ellul P, Maruani A, Vantalon V, et al. Maternal immune activation during pregnancy is associated with more difficulties in socio-adaptive behaviors in autism spectrum disorder. Sci Rep. 2023;13(1):17687. [CrossRef]
- Carter M, Casey S, O’Keeffe GW, Gibson L, Gallagher L, Murray DM. Maternal Immune Activation and Interleukin 17A in the Pathogenesis of Autistic Spectrum Disorder and Why It Matters in the COVID-19 Era. Front Psychiatry. 2022;13:823096. [CrossRef]
- Yin H, Wang Z, Liu J, et al. Dysregulation of immune and metabolism pathways in maternal immune activation induces an increased risk of autism spectrum disorders. Life Sci. 2023;324:121734. [CrossRef]
- Alamoudi RA, Al-Jabri BA, Alsulami MA, Sabbagh HJ. Prenatal maternal stress and the severity of autism spectrum disorder: A cross-sectional study. Dev Psychobiol. 2023;65(2):e22369. [CrossRef]
- Tioleco N, Silberman AE, Stratigos K, et al. Prenatal maternal infection and risk for autism in offspring: A meta-analysis. Autism Res. 2021;14(6):1296-1316. [CrossRef]
- Nudel R, Thompson WK, Børglum AD, et al. Maternal pregnancy-related infections and autism spectrum disorder—the genetic perspective. Transl Psychiatry. 2022;12(1):1-9. [CrossRef]
- Brynge M, Sjöqvist H, Gardner RM, Lee BK, Dalman C, Karlsson H. Maternal infection during pregnancy and likelihood of autism and intellectual disability in children in Sweden: a negative control and sibling comparison cohort study. Lancet Psychiatry. 2022;9(10):782-791. [CrossRef]
- Duque-Cartagena T, Dalla MDB, Mundstock E, et al. Environmental pollutants as risk factors for autism spectrum disorders: a systematic review and meta-analysis of cohort studies. BMC Public Health. 2024;24(1):2388. [CrossRef]
- Phiri YVA, Canty T, Nobles C, Ring AM, Nie J, Mendola P. Neonatal intensive care admissions and exposure to satellite-derived air pollutants in the United States, 2018. Sci Rep. 2025;15(1):420. [CrossRef]
- Bragg MG, Gorski-Steiner I, Song A, et al. Prenatal air pollution and children’s autism traits score: Examination of joint associations with maternal intake of vitamin D, methyl donors, and polyunsaturated fatty acids using mixture methods. Environ Epidemiol Phila Pa. 2024;8(4):e316. [CrossRef]
- Kang N, Sargsyan S, Chough I, et al. Dysregulated metabolic pathways associated with air pollution exposure and the risk of autism: Evidence from epidemiological studies. Environ Pollut Barking Essex 1987. 2024;361:124729. [CrossRef]
- Amnuaylojaroen T, Parasin N, Saokaew S. Exploring the association between early-life air pollution exposure and autism spectrum disorders in children: A systematic review and meta-analysis. Reprod Toxicol. 2024;125:108582. [CrossRef]
- Ritz B, Liew Z, Yan Q, et al. Air pollution and autism in Denmark. Environ Epidemiol. 2018;2(4):e028. [CrossRef]
- Ojha SK, Amal H. Air pollution: an emerging risk factor for autism spectrum disorder. Brain Med. 2024;1(1):31-34. [CrossRef]
- Dutheil F, Comptour A, Morlon R, et al. Autism spectrum disorder and air pollution: A systematic review and meta-analysis. Environ Pollut Barking Essex 1987. 2021;278:116856. [CrossRef]
- Carter SA, Rahman MM, Lin JC, et al. In utero exposure to near-roadway air pollution and autism spectrum disorder in children. Environ Int. 2022;158:106898. [CrossRef]
- Flanagan E, Malmqvist E, Rittner R, Gustafsson P, Källén K, Oudin A. Exposure to local, source-specific ambient air pollution during pregnancy and autism in children: a cohort study from southern Sweden. Sci Rep. 2023;13(1):3848. [CrossRef]
- Autism likelihood in infants born to mothers with asthma is associated with blood inflammatory gene biomarkers in pregnancy—ScienceDirect. Accessed May 20, 2025. https://www.sciencedirect.com/science/article/pii/S2666354624001236.
- Gong T, Lundholm C, Rejnö G, et al. Parental asthma and risk of autism spectrum disorder in offspring: A population and family-based case-control study. Clin Exp Allergy. 2019;49(6):883-891. [CrossRef]
- Neural Regeneration Research. Accessed May 20, 2025. https://journals.lww.com/nrronline/fulltext/2025/04000/evidence_supporting_the_relationship_between.26.aspx.
- Liao J, Yan W, Zhang Y, et al. Associations of preconception air pollution exposure with growth trajectory in young children: A prospective cohort study. Environ Res. 2025;267:120665. [CrossRef]
- Gardner RM, Brynge M, Sjöqvist H, Dalman C, Karlsson H. Maternal Immune Activation and Autism in Offspring: What Is the Evidence for Causation? Biol Psychiatry. Published online November 2024:S0006322324017608. [CrossRef]
- Brynge M. Immune Dysregulation in Early Life and Risk of Autism. thesis. Karolinska Institutet; 2022. Accessed May 20, 2025. https://openarchive.ki.se/articles/thesis/Immune_dysregulation_in_early_life_and_risk_of_autism/26904367/1.
- McLellan J, Kim DHJ, Bruce M, Ramirez-Celis A, Van de Water J. Maternal Immune Dysregulation and Autism–Understanding the Role of Cytokines, Chemokines and Autoantibodies. Front Psychiatry. 2022;13. [CrossRef]
- Robinson-Agramonte M de los A, Noris García E, Fraga Guerra J, et al. Immune Dysregulation in Autism Spectrum Disorder: What Do We Know about It? Int J Mol Sci. 2022;23(6):3033. [CrossRef]
- Kumar P, Kumar A, Kumar D, et al. Microplastics influencing aquatic environment and human health: A review of source, determination, distribution, removal, degradation, management strategy and future perspective. J Environ Manage. 2025;375:124249. [CrossRef]
- Thongkorn S, Kanlayaprasit S, Kasitipradit K, et al. Investigation of autism-related transcription factors underlying sex differences in the effects of bisphenol A on transcriptome profiles and synaptogenesis in the offspring hippocampus. Biol Sex Differ. 2023;14(1):8. [CrossRef]
- Parenti M, Schmidt RJ, Ozonoff S, et al. Maternal Serum and Placental Metabolomes in Association with Prenatal Phthalate Exposure and Neurodevelopmental Outcomes in the MARBLES Cohort. Metabolites. 2022;12(9):829. [CrossRef]
- Parenti M, Slupsky CM. Disrupted Prenatal Metabolism May Explain the Etiology of Suboptimal Neurodevelopment: A Focus on Phthalates and Micronutrients and their Relationship to Autism Spectrum Disorder. Adv Nutr Bethesda Md. 2024;15(9):100279. [CrossRef]
- Symeonides C, Vacy K, Thomson S, et al. Male autism spectrum disorder is linked to brain aromatase disruption by prenatal BPA in multimodal investigations and 10HDA ameliorates the related mouse phenotype. Nat Commun. 2024;15(1):6367. [CrossRef]
- Stevens S, McPartland M, Bartosova Z, Skåland HS, Völker J, Wagner M. Plastic Food Packaging from Five Countries Contains Endocrine- and Metabolism-Disrupting Chemicals. Environ Sci Technol. 2024;58(11):4859-4871. [CrossRef]
- Su Z, Kong R, Huang C, et al. Exposure to polystyrene nanoplastics causes anxiety and depressive-like behavior and down-regulates EAAT2 expression in mice. Arch Toxicol. Published online February 28, 2025. [CrossRef]
- Lin J, Pan D, Zhu Y, et al. Polystyrene nanoplastics chronic exposure cause zebrafish visual neurobehavior toxicity through TGFβ-crystallin axis. J Hazard Mater. 2025;492:138255. [CrossRef]
- Zaheer J, Kim H, Ko IO, et al. Pre/post-natal exposure to microplastic as a potential risk factor for autism spectrum disorder. Environ Int. 2022;161:107121. [CrossRef]
- Cortés-Albornoz MC, García-Guáqueta DP, Velez-van-Meerbeke A, Talero-Gutiérrez C. Maternal Nutrition and Neurodevelopment: A Scoping Review. Nutrients. 2021;13(10):3530. [CrossRef]
- Siracusano M, Riccioni A, Abate R, Benvenuto A, Curatolo P, Mazzone L. Vitamin D Deficiency and Autism Spectrum Disorder. Curr Pharm Des. 2020;26(21):2460-2474. [CrossRef]
- Zwierz M, Suprunowicz M, Mrozek K, et al. Vitamin B12 and Autism Spectrum Disorder: A Review of Current Evidence. Nutrients. 2025;17(7):1220. [CrossRef]
- Pancheva R, Toneva A, Bocheva Y, Georgieva M, Koleva K, Yankov I. Prevalence of vitamin D deficiency in children with cerebral palsy and autism spectrum disorder: a comparative pilot study. Folia Med (Plovdiv). 2024;66(6):787-794. [CrossRef]
- Stefanyshyn V, Stetsyuk R, Hrebeniuk O, et al. Analysis of the Association Between the SLC19A1 Genetic Variant (rs1051266) and Autism Spectrum Disorders, Cerebral Folate Deficiency, and Clinical and Laboratory Parameters. J Mol Neurosci MN. 2025;75(2):42. [CrossRef]
- Gusso D, Prauchner GRK, Rieder AS, Wyse ATS. Biological Pathways Associated with Vitamins in Autism Spectrum Disorder. Neurotox Res. 2023;41(6):730-740. [CrossRef]
- Kacimi FE, Ed-Day S, Didou L, et al. Narrative Review: The Effect of Vitamin A Deficiency on Gut Microbiota and Their Link with Autism Spectrum Disorder. J Diet Suppl. 2024;21(1):116-134. [CrossRef]
- Kacimi FE, Didou L, Ed Day S, et al. Gut microbiota, vitamin A deficiency and autism spectrum disorder: an interconnected trio—a systematic review. Nutr Neurosci. 2025;28(4):492-502. [CrossRef]
- Savino R, Medoro A, Ali S, Scapagnini G, Maes M, Davinelli S. The Emerging Role of Flavonoids in Autism Spectrum Disorder: A Systematic Review. J Clin Med. 2023;12(10):3520. [CrossRef]
- Schimansky S, Jasim H, Pope L, et al. Nutritional blindness from avoidant-restrictive food intake disorder—recommendations for the early diagnosis and multidisciplinary management of children at risk from restrictive eating. Arch Dis Child. 2024;109(3):181-187. [CrossRef]
- Panchawagh SJ, Kumar P, Srikumar S, et al. Role of Micronutrients in the Management of Autism Spectrum Disorders: A Systematic Review and Meta-Analysis. Indian J Med Spec. 2023;14(4):187-196. [CrossRef]
- Sumathi T, Manivasagam T, Thenmozhi AJ. The Role of Gluten in Autism. Adv Neurobiol. 2020;24:469-479. [CrossRef]
- Usui N, Kobayashi H, Shimada S. Neuroinflammation and Oxidative Stress in the Pathogenesis of Autism Spectrum Disorder. Int J Mol Sci. 2023;24(6):5487. [CrossRef]
- Woo T, King C, Ahmed NI, et al. microRNA as a Maternal Marker for Prenatal Stress-Associated ASD, Evidence from a Murine Model. J Pers Med. 2023;13(9):1412. [CrossRef]
- Love C, Sominsky L, O’Hely M, Berk M, Vuillermin P, Dawson SL. Prenatal environmental risk factors for autism spectrum disorder and their potential mechanisms. BMC Med. 2024;22(1):393. [CrossRef]
- Subashi E, Lemaire V, Petroni V, Pietropaolo S. The Impact of Mild Chronic Stress and Maternal Experience in the Fmr1 Mouse Model of Fragile X Syndrome. Int J Mol Sci. 2023;24(14):11398. [CrossRef]
- Ahmavaara K, Ayoub G. Stress and Folate Impact Neurodevelopmental Disorders. J Health Care Res. 2024;5(1):1-6. [CrossRef]
- Hoover DW, Kaufman J. Adverse childhood experiences in children with autism spectrum disorder. Curr Opin Psychiatry. 2018;31(2):128-132. [CrossRef]
- Liu K, Garcia A, Park JJ, Toliver AA, Ramos L, Aizenman CD. Early Developmental Exposure to Fluoxetine and Citalopram Results in Different Neurodevelopmental Outcomes. Neuroscience. 2021;467:110-121. [CrossRef]
- Bravo K, González-Ortiz M, Beltrán-Castillo S, Cáceres D, Eugenín J. Development of the Placenta and Brain Are Affected by Selective Serotonin Reuptake Inhibitor Exposure During Critical Periods. In: Gonzalez-Ortiz M, ed. Advances in Maternal-Fetal Biomedicine: Cellular and Molecular Mechanisms of Pregnancy Pathologies. Springer International Publishing; 2023:179-198. [CrossRef]
- Arzuaga AL, Teneqexhi P, Amodeo K, Larson JR, Ragozzino ME. Prenatal stress and fluoxetine exposure in BTBR and B6 mice differentially affects autism-like behaviors in adult male and female offspring. Physiol Behav. 2025;295:114891. [CrossRef]
- Arzuaga AL, Edmison DD, Mroczek J, Larson J, Ragozzino ME. Prenatal stress and fluoxetine exposure in mice differentially affect repetitive behaviors and synaptic plasticity in adult male and female offspring. Behav Brain Res. 2023;436:114114. [CrossRef]
- Lan Z, Tachibana RO, Kanno K. Chronic exposure of female mice to selective serotonin reuptake inhibitors during lactation induces vocal behavior deficits in pre-weaned offspring. Pharmacol Biochem Behav. 2023;230:173606. [CrossRef]
- Croen LA, Ames JL, Qian Y, et al. Inflammatory Conditions During Pregnancy and Risk of Autism and Other Neurodevelopmental Disorders. Biol Psychiatry Glob Open Sci. 2024;4(1):39-50. [CrossRef]
- Ye W, Luo C, Zhou J, et al. Association between maternal diabetes and neurodevelopmental outcomes in children: a systematic review and meta-analysis of 202 observational studies comprising 56·1 million pregnancies. Lancet Diabetes Endocrinol. 2025;0(0). [CrossRef]
- Fardous AM, Heydari AR. Uncovering the Hidden Dangers and Molecular Mechanisms of Excess Folate: A Narrative Review. Nutrients. 2023;15(21):4699. [CrossRef]
- Miraglia N, Dehay E. Folate Supplementation in Fertility and Pregnancy: The Advantages of (6S)5-Methyltetrahydrofolate. Altern Ther Health Med. 2022;28(4):12-17.
- Xu X, Zhang Z, Lin Y, Xie H. Risk of Excess Maternal Folic Acid Supplementation in Offspring. Nutrients. 2024;16(5):755. [CrossRef]
- Ledowsky CJ, Schloss J, Steel A. Variations in folate prescriptions for patients with the MTHFR genetic polymorphisms: A case series study. Explor Res Clin Soc Pharm. 2023;10:100277. [CrossRef]
- Maguire G. Vaccine Induced Autoimmunity May Cause Autism and Neurological Disorders. Arch Microbiol Immunol. 2025;9(1):103-132.
- Sotelo-Orozco J, Schmidt RJ, Slupsky CM, Hertz-Picciotto I. Investigating the Urinary Metabolome in the First Year of Life and Its Association with Later Diagnosis of Autism Spectrum Disorder or Non-Typical Neurodevelopment in the MARBLES Study. Int J Mol Sci. 2023;24(11):9454. [CrossRef]
- Parenti M, Shoff S, Sotelo-Orozco J, Hertz-Picciotto I, Slupsky CM. Metabolomics of mothers of children with autism, idiopathic developmental delay, and Down syndrome. Sci Rep. 2024;14(1):31981. [CrossRef]
- Parenti M, Schmidt RJ, Tancredi DJ, Hertz-Picciotto I, Walker CK, Slupsky CM. Neurodevelopment and Metabolism in the Maternal-Placental-Fetal Unit. JAMA Netw Open. 2024;7(5):e2413399. [CrossRef]
- Merchak AR, Bolen ML, Tansey MG, Menees KB. Thinking outside the brain: Gut microbiome influence on innate immunity within neurodegenerative disease. Neurotherapeutics. 2024;21(6). [CrossRef]
- Sanctuary MR, Kain JN, Chen SY, et al. Pilot study of probiotic/colostrum supplementation on gut function in children with autism and gastrointestinal symptoms. PloS One. 2019;14(1):e0210064. [CrossRef]
- Dossaji Z, Khattak A, Tun KM, Hsu M, Batra K, Hong AS. Efficacy of Fecal Microbiota Transplant on Behavioral and Gastrointestinal Symptoms in Pediatric Autism: A Systematic Review. Microorganisms. 2023;11(3):806. [CrossRef]
- Zang Y, Lai X, Li C, Ding D, Wang Y, Zhu Y. The Role of Gut Microbiota in Various Neurological and Psychiatric Disorders-An Evidence Mapping Based on Quantified Evidence. Mediators Inflamm. 2023;2023:5127157. [CrossRef]
- Lagod PP, Abdelli LS, Naser SA. An In Vivo Model of Propionic Acid-Rich Diet-Induced Gliosis and Neuro-Inflammation in Mice (FVB/N-Tg(GFAPGFP)14Mes/J): A Potential Link to Autism Spectrum Disorder. Int J Mol Sci. 2024;25(15):8093. [CrossRef]
- Retuerto M, Al-Shakhshir H, Herrada J, McCormick TS, Ghannoum MA. Analysis of Gut Bacterial and Fungal Microbiota in Children with Autism Spectrum Disorder and Their Non-Autistic Siblings. Nutrients. 2024;16(17):3004. [CrossRef]
- Plaza-Díaz J, Gómez-Fernández A, Chueca N, et al. Autism Spectrum Disorder (ASD) with and without Mental Regression is Associated with Changes in the Fecal Microbiota. Nutrients. 2019;11(2):337. [CrossRef]
- Brister D, Rose S, Delhey L, et al. Metabolomic Signatures of Autism Spectrum Disorder. J Pers Med. 2022;12(10):1727. [CrossRef]
- Che X, Roy A, Bresnahan M, et al. Metabolomic analysis of maternal mid-gestation plasma and cord blood in autism spectrum disorders. Mol Psychiatry. 2023;28(6):2355-2369. [CrossRef]
- Vacy K, Thomson S, Moore A, et al. Cord blood lipid correlation network profiles are associated with subsequent attention-deficit/hyperactivity disorder and autism spectrum disorder symptoms at 2 years: a prospective birth cohort study. EBioMedicine. 2024;100:104949. [CrossRef]
- Nabetani M, Mukai T, Taguchi A. Cell Therapies for Autism Spectrum Disorder Based on New Pathophysiology: A Review. Cell Transplant. 2023;32:9636897231163217. [CrossRef]
- Ahrens AP, Hyötyläinen T, Petrone JR, et al. Infant microbes and metabolites point to childhood neurodevelopmental disorders. Cell. 2024;187(8):1853-1873.e15. [CrossRef]
- Freitas BC, Beltrão-Braga PCB, Marchetto MC. Modeling Inflammation on Neurodevelopmental Disorders Using Pluripotent Stem Cells. Adv Neurobiol. 2020;25:207-218. [CrossRef]
- Tsukada T, Shimada H, Sakata-Haga H, Iizuka H, Hatta T. Molecular mechanisms underlying the models of neurodevelopmental disorders in maternal immune activation relevant to the placenta. Congenit Anom. 2019;59(3):81-87. [CrossRef]
- Frye RE, Rincon N, McCarty PJ, Brister D, Scheck AC, Rossignol DA. Biomarkers of mitochondrial dysfunction in autism spectrum disorder: A systematic review and meta-analysis. Neurobiol Dis. 2024;197:106520. [CrossRef]
- Amini-Khoei H, Taei N, Dehkordi HT, et al. Therapeutic Potential of Ocimum basilicum L. Extract in Alleviating Autistic-Like Behaviors Induced by Maternal Separation Stress in Mice: Role of Neuroinflammation and Oxidative Stress. Phytother Res. 2025;39(1):64-76. [CrossRef]
- Nguyen V, Tang J, Oroudjev E, et al. Cytotoxic Effects of Bilberry Extract on MCF7-GFP-Tubulin Breast Cancer Cells. J Med Food. 2010;13(2):278-285. [CrossRef]
- Jessica Tang, Emin Oroudjev, Leslie Wilson, George Ayoub. Delphinidin and cyanidin exhibit antiproliferative and apoptotic effects in MCF7 human breast cancer cells. Integr Cancer Sci Ther. 2015;2(1). [CrossRef]
- Lai M, Lee J, Chiu S, et al. A machine learning approach for retinal images analysis as an objective screening method for children with autism spectrum disorder. EClinicalMedicine. 2020;28:100588. [CrossRef]
- Farooq MS, Tehseen R, Sabir M, Atal Z. Detection of autism spectrum disorder (ASD) in children and adults using machine learning. Sci Rep. 2023;13(1):9605. [CrossRef]
Figure 1.
Schematic of maternal inflammatory categories and their impact on fetal development that can lead to conditions conducive for development of autism spectrum disorder. A genetic propensity and agents causing oxidative stress are presumed to be the preconditions for neurodivergent fetal development. A similar schematic is seen in Figure 2 for the child.
Figure 1.
Schematic of maternal inflammatory categories and their impact on fetal development that can lead to conditions conducive for development of autism spectrum disorder. A genetic propensity and agents causing oxidative stress are presumed to be the preconditions for neurodivergent fetal development. A similar schematic is seen in Figure 2 for the child.
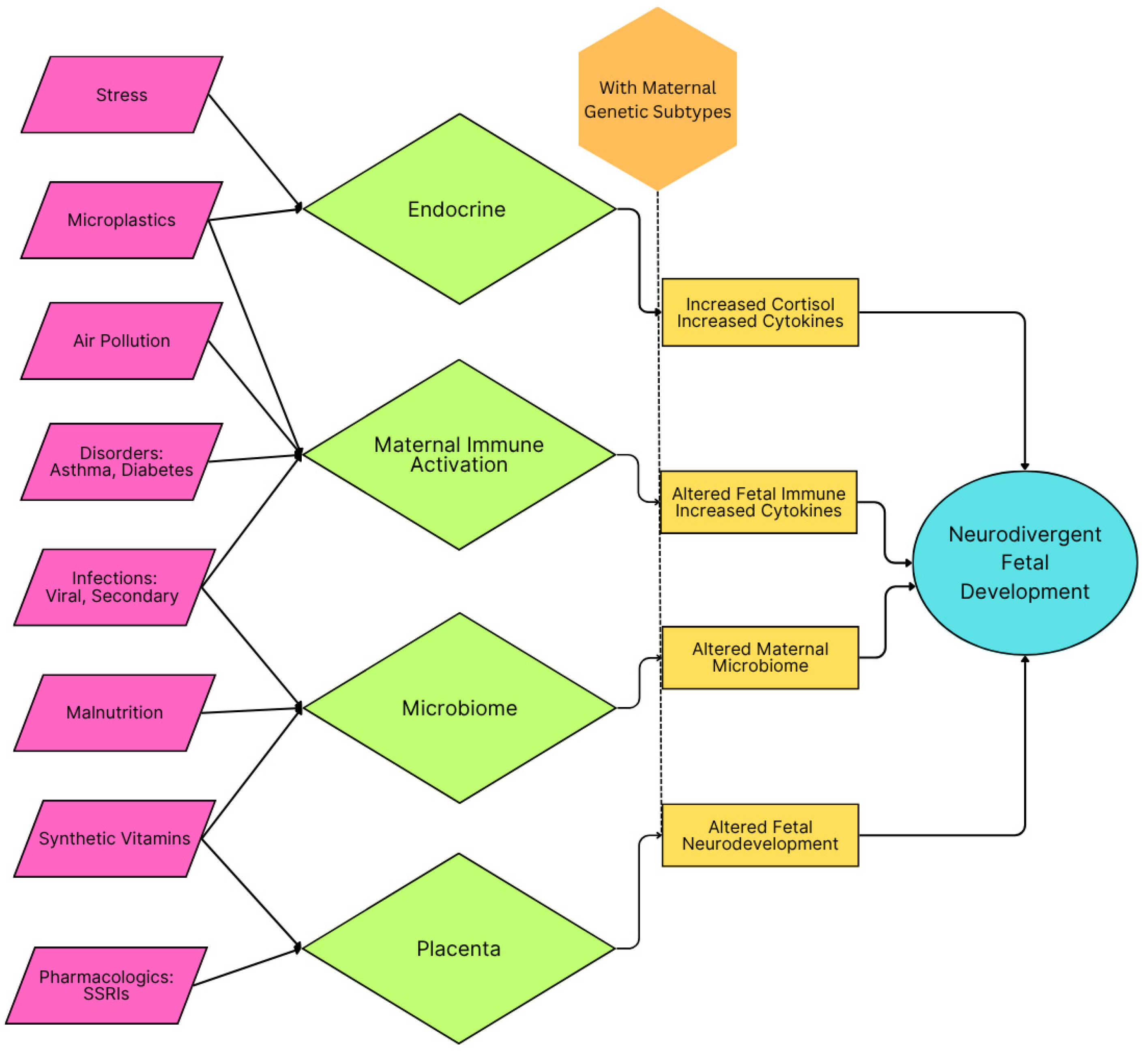
Figure 2.
Schematic of inflammatory categories in childhood and their impact on early childhood development that can lead to autism spectrum disorder symptoms. A genetic propensity and agents causing oxidative stress are presumed to be the preconditions for neurodivergent development.
Figure 2.
Schematic of inflammatory categories in childhood and their impact on early childhood development that can lead to autism spectrum disorder symptoms. A genetic propensity and agents causing oxidative stress are presumed to be the preconditions for neurodivergent development.
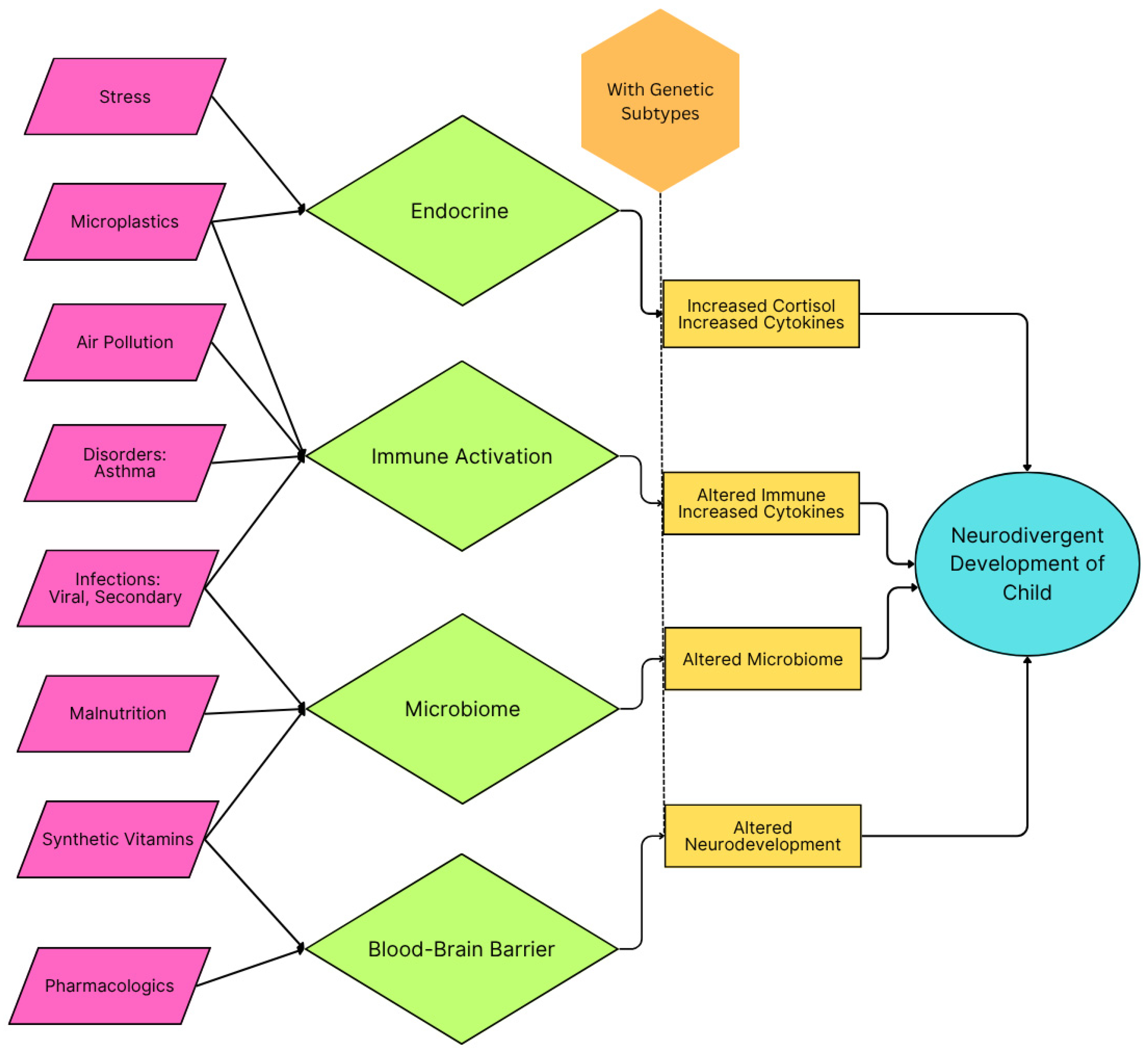
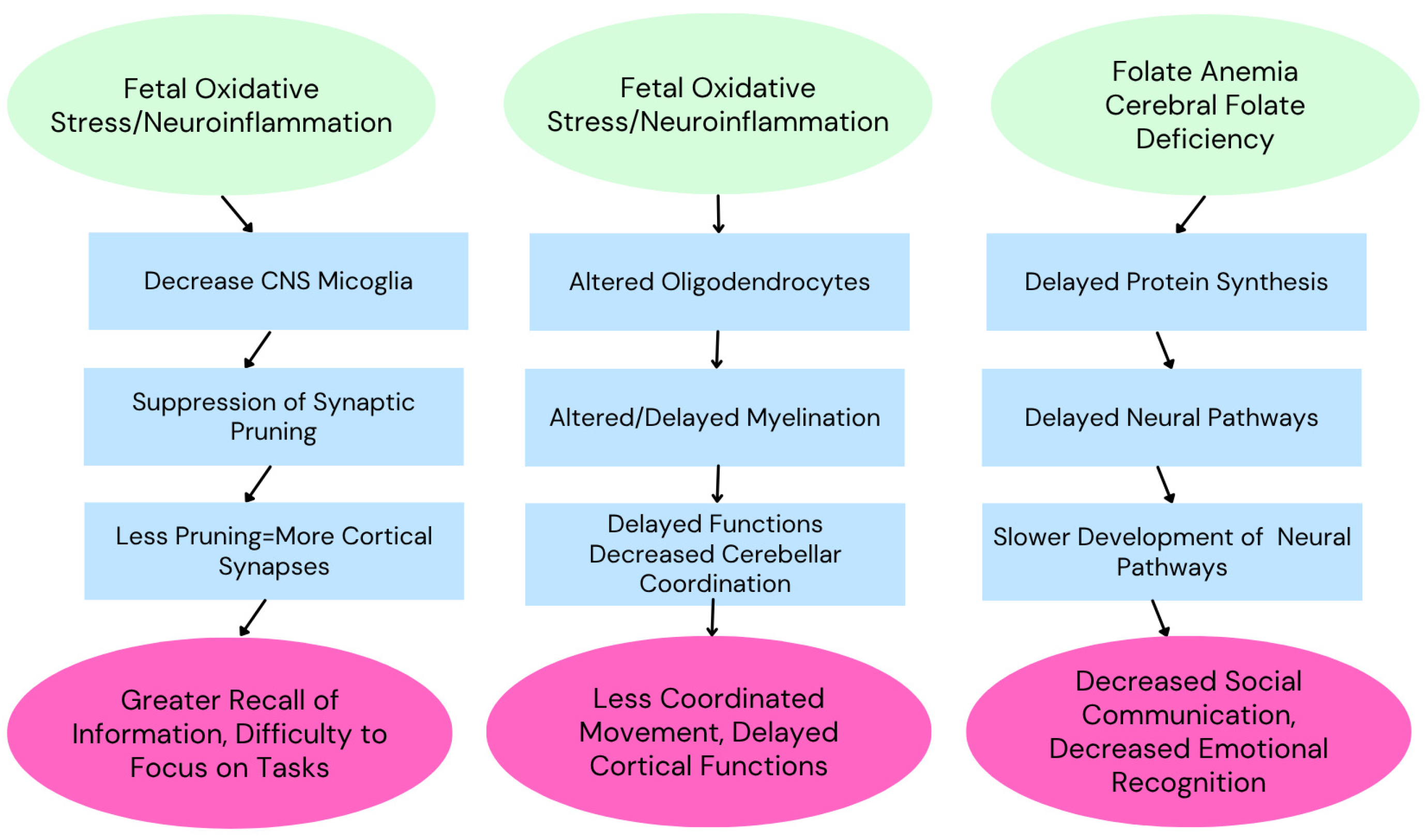
Figure 3.
Development of autism spectrum disorder is multifactorial, with various factors presumably responsible for the various symptoms observed. Adapted from Ayoub (2024)[35].
Figure 3.
Development of autism spectrum disorder is multifactorial, with various factors presumably responsible for the various symptoms observed. Adapted from Ayoub (2024)[35].
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



