Submitted:
29 May 2025
Posted:
04 June 2025
You are already at the latest version
Abstract
Environmental DNA and RNA-based virus biosurveillance holds promise as an early warningsystem for detecting pathogen incursions and tracking their evolution across fields. Unliketraditional pathogen monitoring, which typically relies on direct sampling from humans and animalhosts, environmental biosurveillance offers a broader, faster, more accessible, and cost-effectiveapproach. While recent advancements in viral detection methods have expanded their utility, therelative effectiveness and suitability of these methods for point-of-need detection remain unclear.To this end, we evaluate and compare 12 contemporary viral detection methods to assess theirpotential for identifying viruses in environmental samples, with a specific focus on highlypathogenic avian influenza (HPAI) virus. We examine the scalability, practicality, and futurepotential of each method, particularly those that have yet to be applied to environmental samples,while considering the specific requirements for each approach. Our goal is to provide guidance toend-users on selecting the most appropriate methods based on current available approaches anddata. In addition, we outline key requirements for an “optimal” in-field detection method andidentify the steps necessary to enhance environmental virus biosurveillance in the future.
Keywords:
biosecurity
; eDNA
; eRNA
; biomonitoring
; HPAI
; CRISPR
; detection
; environmental surveillance
1. Introduction
Viral disease outbreaks can have devastating consequences for human, animal, and plant health (Jones, 2021; Msemburi et al., 2023; Oldstone, 2020). One prominent example is the H5N1 subtype of highly pathogenic avian influenza (HPAI) virus that is currently spreading worldwide, killing millions of birds and spilling over into a broad range of non-avian hosts, including livestock and humans (Bi et al., 2024; Neumann & Kawaoka, 2024; Plaza et al., 2024). Given the significant health risks associated with such viral outbreaks, there is an increasing need for rapid detection and continuous biosurveillance, ideally using environmental samples. Such efforts would provide a much-needed early warning system for new viral incursions while improving our understanding of pathogen transmission and evolution.
Environmental sampling of DNA and RNA (eDNA/eRNA) has emerged as a powerful tool for viral monitoring (Bass et al., 2023; Farrell et al., 2021), offering a cost-effective, scalable, and less-invasive alternative to traditional host sampling for pathogen detection (Alfano et al., 2021; Farrell et al., 2021; Huver et al., 2015; Kawato et al., 2021). Compared to direct sampling from hosts, environmental surveillance is safer and less labour-intensive (Hood et al., 2021).
The practice of monitoring viruses in environmental samples dates back to the 1940s when cell culture techniques were used to detect human enteric viruses in water sources (Bivins et al., 2022; Metcalf et al., 1995). While slow, cell culture was the gold standard because alternative approaches such as radioimmunoassay, immunofluorescence, and complement fixation lacked the sensitivity needed for detecting low viral counts from environmental samples (Fong & Lipp, 2005; Griffin et al., 2001). Other molecular detection methods including loop-mediated isothermal amplification (LAMP) and enzyme-linked immunosorbent assays (ELISA) proved useful for rapid and sensitive detection of viruses from environmental samples (Kfir & Genthe, 1993; Wong et al., 2018; Zhu et al., 2021), however, it was reverse transcriptase (Temin & Mizutami, 1970) and polymerase chain reaction (PCR) (Mullis & Faloona, 1987) that revolutionised viral detection, enabling rapid, specific and highly sensitive identification of viruses, including strain-specific information, even from low-titre samples (Fu et al., 2023).
Viral environmental detection techniques have developed rapidly in the last five years, largely due to the COVID-19 pandemic. During the COVID-19 pandemic, significant efforts were directed toward optimising environmental sampling, particularly human wastewater surveillance, to detect viral variants (Kumblathan et al., 2021). These methodologies are now being applied to monitor a wide range of viruses, including mpox, dengue, and HPAI, with new assays and technologies continuously emerging (de Jonge et al., 2022; Louis, 2024; Wolfe et al., 2023). In response to the ongoing global spread of HPAI and its potential effect on wildlife, human health, and primary industries (Adlhoch et al., 2022; Plaza et al., 2024), several new molecular detection methods have been deployed. These include CRISPR-based diagnostic platforms (CRISPR-Dx) (J. S. Chen et al., 2018; S.-S. Chen et al., 2024; Gootenberg et al., 2018), digital droplet PCR (ddPCR) (Wolfe et al., 2024), and Oxford Nanopore Technology (ONT)-based sequencing approaches (Croville et al., 2024). Nevertheless, to be effective in real-world settings, viral detection methods must be user-friendly, highly specific, cost-efficient for large-scale use, and capable of detecting viruses from a wide range of diverse, low-concentration environmental samples so that they can guide biosecurity investigations and public health decisions towards tactical deployments (e.g. containment or eradication) (Campbell & Hewitt, 2025). While the development of new environmental nucleic acid detection methods has provided end-users with numerous options, a clear understanding of their advantages and limitations is currently lacking. This knowledge gap presents challenges both in optimising existing methods and in selecting the most suitable technology for new monitoring programs.
To address this knowledge gap, we examine 12 molecular methods to explore their resource requirements and in-field potential for viral detection. We particularly focus on a method’s utility in the current global HPAI outbreak. Herein, we discuss the advantages and disadvantages of these methods, offering end-users a practical guide to selecting the most appropriate approach for viral detection based on several of their requirements. We also discuss key barriers to achieving a cost-effective, low-resource, and field-deployable method for detecting viruses in low-concentration environmental samples, along with potential solutions to overcoming these challenges.
2. An Overview of Current Detection Methods for Monitoring Viruses in the Environment
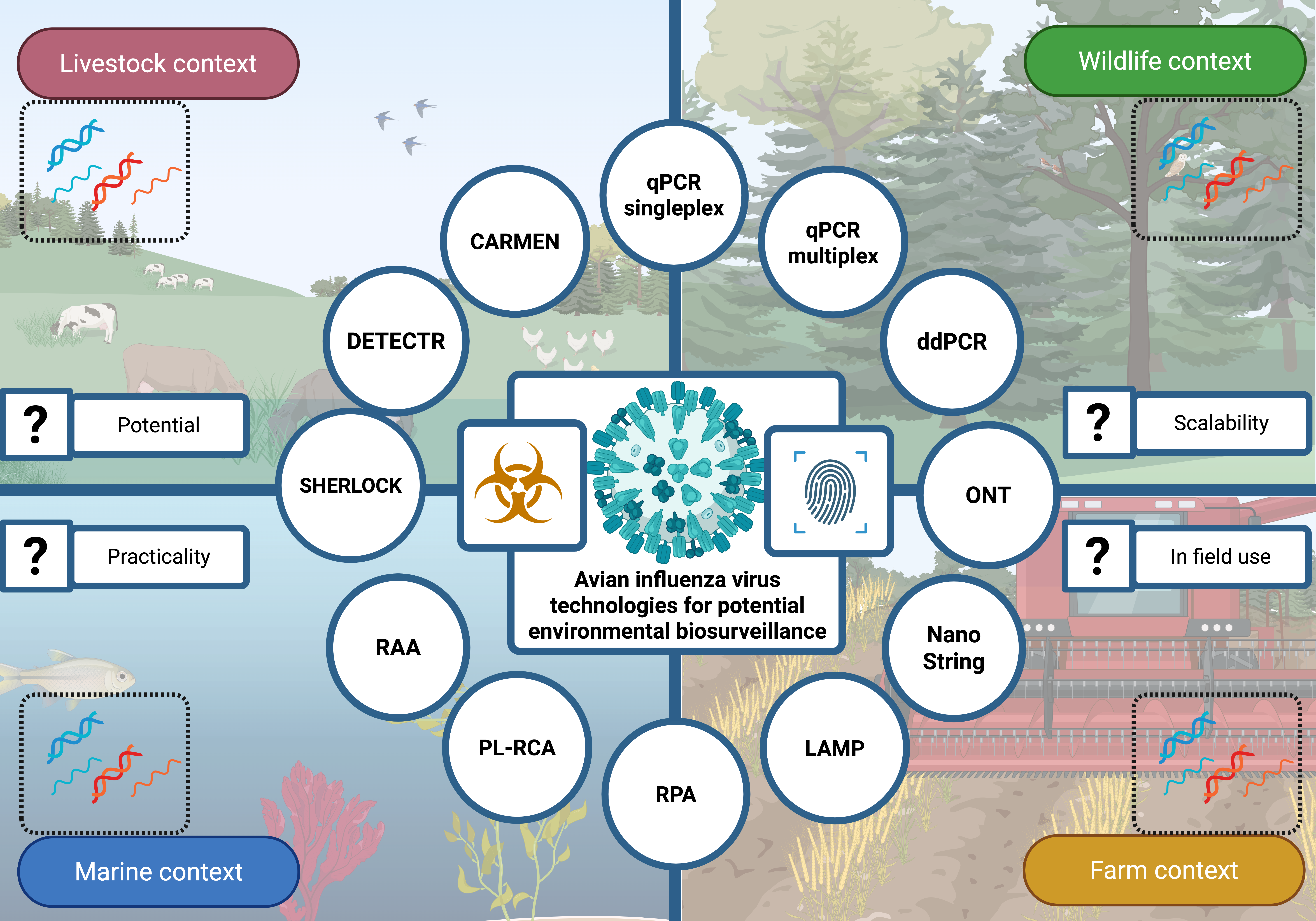
The proliferation of nucleic acid testing (NAT) methods can make it difficult for end-users and researchers to select an approach that best suits their environmental virus monitoring needs. Methods vary in sensitivity, up-front development costs, per-method expenses, and the equipment and expertise required for implementation. To aid in the selection process, we compiled a list of possible methods, some of which are commonly used for detecting viruses in environmental samples, and have summarised their methodological requirements, advantages and limitations (Figure 1; also see Supplementary Table S1 for more detail). While our primary focus is on detection and biosurveillance methods for avian influenza virus (AIV), these methods can, in principle, be applied to any virus (RNA and DNA) or nucleic acid target. We intend this summary to serve as a practical guide for end-users evaluating different detection methods.
We identified 12 distinct viral NAT approaches for AIV detection, including CRISPR-Dx platforms (DETECTR (Chen et al., 2018); SHERLOCK (Gootenberg et al., 2018); and CARMEN (Ackerman et al., 2020)), loop-mediated isothermal amplification (LAMP; Kaneko et al., 2005), NanoString (nCounter; Geiss et al., 2008), ONT (Ji et al., 2020), padlock rolling circle amplification (PL-RCA; Jain et al., 2021), quantitative PCR (qPCR; singleplex (Ding et al., 2024) and multiplex (Vogels et al., 2021)), digital droplet PCR (ddPCR; Ding et al., 2024), recombinase-aided amplification (RAA; Nie et al., 2022), and recombinase polymerase amplification (RPA; Parameswari et al., 2025). Overall, all methods currently rely on a heat source, custom-designed primers, and probes uniquely designed for each method, and, in most cases, nucleic acid extraction. However, there are key notable differences in methodological requirements that present distinct advantages and disadvantages among these approaches. Below, we outline the commonalities and differences in detail.
2.1. Nucleic Acid Extraction
Most methods and samples require nucleic acid extraction (the process of isolating or purifying nucleic acids from a biological sample), with the potential exceptions of some LAMP, CRISPR, and PCR methods, where extraction may not be strictly necessary for target detection (e.g., Barnes et al., 2020; Lalli et al., 2021; Schellenberg et al., 2021; Scriver et al., 2024; Wang et al., 2024). Nucleic acid extraction is needed for RNA targets to prevent degradation, and can help mitigate unwanted complications associated with non-target (e.g. DNA or cellular material) and environmental (e.g. decaying matter and tannins) components. Often this is done through methods including magnetic bead washes and DNase digestions.
2.2. Amplification with Thermocycler
Some methods discussed require resources for nucleic acid amplification. Singleplex and multiplex qPCR and ddPCR, as well as metagenomics approaches, rely on thermocyclers (PCR instruments capable of precisely controlling temperature changes at specific intervals) and sometimes other equipment (e.g. equipment to read fluorescence) for nucleic acid amplification. Some CRISPR-Dx platforms, such as SHERLOCK and DETECTR can simply be run on battery-operated portable heaters (Misra et al., 2022; Zahra et al., 2023), and only require thermocyclers when the reactions are fluorescence-based (Broughton et al., 2020; Myhrvold et al., 2018). Other methods, such as LAMP and RPA, do not necessarily rely on a thermocycler, but still require temperature control (Notomi et al., 2000; Piepenburg et al., 2006).
2.3. Heating and Cooling
All methods require heating steps for strand separation, probe annealing, and enzymatic activity. The complexity of heating and cooling requirements varies, however, ranging from simple heat plates (LAMP methods), to thermocyclers (PCR-based methods), and method-specific heating instruments (e.g., ONT Minion and NanoString Counter Digital Analyzer).
2.4. Additional Equipment Required
At their full optimisation, the equipment requirement needs of the different methods vary dramatically, with some such as NanoString, metagenomics, Carmen Cas13, ONT, and ddPCR often requiring a large amount of equipment compared to others such as CRISPR-Dx platforms and LAMP.
2.5. In-Field Potential
The need for laboratory space depends largely on the method stage. Most methods require a laboratory during early-stage optimisation, but some can be adapted and performed in-field once optimised. Singleplex and multiplex qPCR, ddPCR, SHERLOCK, DETECTR, and ONT sequencing can be adapted for field use with portable equipment (e.g., Boykin et al., 2019). In contrast, methods such as CARMEN, metagenomic sequencing, and NanoString require large, non-portable equipment, making field deployment generally impractical.
2.6. Expertise
Most methods require significant expertise during the development phase, but minimal training once optimised. For example, LAMP requires well-trained staff for primer design and optimisation, but is relatively straightforward to perform afterwards. A benefit of using well-established methods (e.g. qPCR) is that there are often tools available to help with assay design, compared to novel approaches that might require more hands-on time. NanoString, ONT, and metagenomic sequencing methods, however, require specialised expertise at all stages due to their complexity and equipment requirements.
2.7. Sample Concentration
We define sample concentration as low (~1–10 copies/µL of target sequence), medium (~100 copies/µL ), and high (>1000 copies/µL). Most methods can analyse samples across the above mentioned range, with the exception of metagenomic sequencing and ONT (in some cases) which typically recommends higher input concentrations.
2.8. Start-Up and Long-Term Resources
Start-up and long-term resources (that is, the initial resources needed to develop the method and the resources needed to maintain and run the method after the initial start-up) ranges widely and is influenced by expertise, laboratory space, and the desired targets to detect. Methods with established protocols focused on common targets will require some optimisation but have minimal start-up requirements compared to methods that target less-common/custom targets (e.g. specific viral variants). For example, there are multiple well-optimised and highly cited qPCR methods (even commercial kits) for detecting the matrix protein of AIV and thus require little trained technician time to optimise for individual needs. In comparison, methods to detect new emerging variants such as the H5N1 clade 2.3.4.4b often demand more time from trained technicians and laboratory resources to develop and optimise.
Long-term resource needs depend on reagents, technician time, software needs, equipment maintenance, and the variable nature of the target to detect. LAMP, for instance, requires minimal ongoing resources, whereas CARMEN, metagenomic sequencing, NanoString, and ONT require costly reagents, technical expertise, and dedicated laboratory equipment. Technical expertise includes the trained bioinformaticians that are needed to process data (e.g. from metagenomic and ONT sequencing) post sequencing, which can take between days and months depending on the required objective. Lastly, if the viral target is highly-dynamic (i.e. quickly evolving new subtypes/strains) then it will require additional effort to ensure that the method is able to continually detect it.
2.9. Proven Use on Environmental AIV Samples
We found it advantageous if the method had been previously demonstrated to detect AIV from environmental samples. To our knowledge the CRISPR-Dx platforms we analysed (excluding Carmen Cas13), LAMP, and NanoString have no published records of their use on environmental samples for AIV biosurveillance.
Overall advantages and disadvantages. We assessed the methods based on multiple criteria, including cost, speed, portability, sensitivity, and viral subtype detection capability (Figure 1; Supplementary Table S1). Expensive equipment, reagents and/or high technician expertise, and time were generally considered disadvantages. Rapid methods, such as PL-RCA, were considered advantageous, while ONT and metagenomic sequencing were often slower compared to other methods and, while they produce a lot of data, they were considered disadvantageous when time to detection were key criteria. It was advantageous if a method had routinely been used for viral detection, such as singleplex and multiplex PCR, and if the method could be deployed at point-of-need.
Sensitivity, or the ability to detect viral genetic material from low concentration samples, is important, especially for environmental samples. Many methods promised high sensitivity, but were not at the level of singleplex PCR (often considered the gold standard for viral detection). In general, the ability to identify more AIV subtypes was advantageous; however, a high level of detection (e.g. CARMEN can detect up to 169 subtypes in one reaction) may be less advantageous to end-users whose requirement is to detect only a specific subtype in the environment (e.g. H5N1). We found some methods had difficulties assessing samples uniformly, for example LAMP sometimes demonstrated delayed onset of amplicon formation because of inhibitors including tannins, urea, calcium chloride, and humic acid (Nwe et al., 2024), or was affected by pH variability of samples (Brown et al., 2021; Nwe et al., 2024), both often characteristics of environmental samples.
Other disadvantages were method-specific and were associated with the equipment or the nature of the method. For example, ddPCR has droplet variability and ONT/metagenomic sequencing has long-term equipment costs, often with sample quality requirements. By considering these factors, end-users can select the most appropriate technology for their specific application, balancing cost, ease of use, sensitivity, and field deployment potential.
3. Application of These Methods to Avian Influenza Virus Environmental Detection
PCR methods have routinely been the cornerstone for detecting specific AIV genes in environmental samples, including bird guano, wastewater, and sediment (e.g., Hubbard et al., 2023; Jallow et al., 2025; Kuchinski et al., 2024; Tisza et al., 2024). Additionally, ONT has demonstrated utility in identifying influenza viral subtype information directly from bird faeces (de Vries et al., 2022; Nabeshima et al., 2023). These established methodologies provide a strong foundation for AIV biosurveillance; however, emerging NAT approaches can potentially offer additional advantages that may enhance detection capabilities in environmental settings.
While these novel NAT approaches have not yet been shown to detect AIV from environmental samples, we considered their sensitivity, limitations, and start-up/long-term resource requirements based on their use across other sample types to estimate their potential applicability for environmental samples. It is clear that single and multiplex PCR methods have established protocols for environmental samples (e.g., Hubbard et al., 2023), are highly sensitive (Smith & Osborn, 2009), and do not require expensive specialised equipment.
Nevertheless, CRISPR-Dx platforms (e.g. DETECTR and SHERLOCK) are ultrasensitive, specific, scalable, portable, and can be free of extensive equipment when deployed with lateral flow strips (Baerwald et al., 2020; Wei et al., 2022) or even without the need for pre-amplification steps (Bhatt et al., 2022; Huang et al., 2022; Miao et al., 2023; Wang et al., 2024), increasing their point-of-need utility for in-field detection, all of which may outweigh the benefits of other technologies.
Although we focus on one virus type here, the NAT methods we examine can be applied to other viruses besides AIV, including DNA viruses. Viruses naturally vary in their genetic sequences, structures, and hosts; however, their general transmission pathways through environmental mediums such as faeces and water often make them detectable by NAT methods. For example, other avian viruses such as Newcastle disease virus, that are primarily spread through the faecal-oral route (Alexander, 2001), can be present in the environment and thus have the potential to be detected through the NAT methods listed here.
4. Limitations of Environmental Sampling for Viruses
Given recent human and animal health concerns regarding HPAI and other zoonotic pathogenic viruses, there is an urgent need for sensitive, rapid, and cost-effective environmental biosurveillance methods. Unfortunately, methods targeting specific viruses from environmental samples face significant challenges, including heating requirements, low virus concentrations, and nucleic acid isolation limitations. Indeed, a heat source was required for all of the eDNA/eRNA methods that we reviewed. Some methods, such as LAMP and PL-RCA, can function with easily-accessible heat sources (e.g. portable heater, or even body heat). However, PCR, metagenomics, and NanoString require precise temperature control provided by specialised laboratory equipment, making in-field applications difficult or requiring financial investment in portable equipment.
Environmental samples typically contain variable virus concentrations, enzyme inhibitors, and environmental contaminants that hinder detection success (Beng & Corlett, 2020; Bowers et al., 2021). Of the 12 methods reviewed, only five (singleplex qPCR, mulitiplex qPCR, ddPCR, metagenomics, and ONT sequencing) have been applied to environmental AIV detection. These methods have demonstrated high sensitivity, with singleplex RT-PCR capable of detecting AIV RNA at concentrations as low as 1x10-1 EID50 (median egg embryo infectious dose) per mL in wetland water (Hubbard et al., 2024). However, most PCR-based methods require laboratory settings, increasing the delay between sampling and detection. Portable qPCR devices, such as Ubiquitome’s Liberty 16 (https://www.ubiquitomebio.com/) device are becoming more available, but their efficacy for environmental samples remains largely under evaluation (Billington et al., 2021). ONT sequencing has emerged as a promising alternative, demonstrating the ability to recover whole hemagglutinin (HA) genes for AIV subtype and phylogenetic clade identification from environmental samples, including from dust and bird guano (Croville et al., 2024; de Vries et al., 2022). It has also demonstrated a higher sensitivity than conventional culture methods for human pathogen detection (Deng et al., 2022). However, ONT sequencing has several drawbacks, including high cost, an often medium-to-steep learning curve (especially the bioinformatic components), and the general need for RNA extractions prior to sequencing (see Figure 1). There are several extraction-free ONT methods; however, they have not yet been evaluated with environmental samples. The sensitivity of ONT is also often considered a disadvantage of the method. Sensitivity studies are generally lacking for ONT virus detection; however, Croville et al. (2024) concluded that such high throughput sequencing methods, including ONT, were not as sensitive as PCR methods, but that there are demonstrated advantages of combining PCR with high throughput sequencing (e.g., Quick et al., 2017). Lastly, adaptive sequencing is a potential option for increasing ONT detection sensitivity and has been demonstrated to improve the sensitivity of viral clinical detection (Lin et al., 2022); however, more research is needed, especially to determine its use with very low concentration targets, such as those often found in environmental samples.
Nucleic acid extraction was required for all reviewed methods, with the exception of LAMP, RCA, and some Cas12- (Wu et al., 2022) and Cas13-based (Myhrvold et al., 2018) methods when used with specific protocols and sample types. The sample types that were tested so far with extraction-free protocols are often clean of inhibitors and highly-concentrated, such as nasopharyngeal, oropharyngeal, urine, or rash (e.g., mpox) samples (Bhatt et al., 2022; Lalli et al., 2021; Schellenberg et al., 2021; Wang et al., 2024). Kang et al. (2023) demonstrated that RT-RPA with Cas12a and G4 cleavage could detect spiked wastewater samples; however, the samples were processed slightly prior to spiking (centrifuged at 9000 rpm for 10 min). Many extraction-free methods have not been tested for AIV detection in environmental samples, while portability and nucleic acid extraction remains a bottleneck in viral biosurveillance due to its time-consuming nature, laboratory space requirements, and high costs. Several recent advancements aim to facilitate in-field nucleic acid isolation, including S-TECH (https://www.dnaitech.com/), Bento Lab (Pillay et al., 2024), and PDQeX (Stanton et al., 2019). Additionally, novel RT-PCR methods that bypass extraction are under development, such as Cytiva’s RT-PCR anti-inhibitor complex (AIC) and homogenizer-based RNA extraction free methods (Ramírez-Córdova et al., 2023; Struijk et al., 2023). However, these techniques have not yet been validated for environmental samples, which often contain high levels of contaminants, variable pH levels, and fragmented or degraded RNA and DNA (Sahu et al., 2023). In addition, they often have a limit on the number of samples they are able to process at once (often 1—24) making them unsuitable for high throughput monitoring programs when compared to laboratory-based methods that can process up to 16x as many samples in parallel.
Developing viral detection methods is inherently challenging due to the extreme sequence variation and rapid evolution of viruses. Primer and probe design was required for all reviewed methods, except for ONT and metagenomic sequencing. Designing optimised primers and probes requires skilled technicians and can take weeks, sometimes months, to ensure specificity and sensitivity. Primers and probes can be used to amplify a broad range of taxa or very specific taxa of interest. For example, they can be designed to sequence conserved regions of the AIV HA gene to detect general HA subtypes such as H5 and H7, or they can be designed to sequence specific lineages of these subtypes such as H5N1 2.3.4.4b. Tools are available for evaluating and validating targeted eDNA primer and probe methods (e.g., Thalinger et al., 2021) and can be similarly applied to eRNA and viruses).
5. Recommendations for Environmental Biosurveillance of Viruses
Although all of the 12 methods that we reviewed were previously proven to detect AIV, only five have been used with environmental samples to our knowledge. We found several methods to be more user-friendly, less resource-heavy, and/or cost-efficient than others based on their requirements (Figure 2). We found singleplex qPCR still appropriately fits into the “gold standard” rating as it is a well-tested method that has been proven to detect multiple viruses (including AIV) at low concentrations, has equipment and protocols readily available, and has been widely used for environmental samples. There are also methods to specifically enhance the RT-qPCR sensitivity for virus detection from environmental samples such as dust and air samples (Bessière et al., 2023). Multiplex qPCR is another popular option after the protocol is optimised; however, the development and optimisation of the method requires time and needs to work with the requirements of the instrument. Commercial multiplex qPCR kits exist, but are often expensive. Although ddPCR performs absolute quantification and is sometimes more sensitive and more tolerant of sample inhibitors than qPCR (Rački et al., 2014), it requires more time, complex machines, greater expertise, and generally less throughput than qPCR. ddPCR may also be more than what is needed by end-users if absolute quantification is not necessary and sample concentration is within qPCR limits. Other methods such as the CRISPR-Dx platforms appear to hold potential to detect viruses from environmental samples (e.g., Kang et al., 2023), but many of them have not yet been tested on environmental samples. For these methods, we found that the development and optimization effort could be high depending on the computational resources available, but that their thermocycler-free, and potentially nucleic acid extraction-free, requirements allowed them to be highly-portable, low-cost, and field-friendly once optimised (e.g., Barnes et al., 2020) (Figure 2).
6. Conclusion
Ultimately the choice of method for environmental virus detection should align with the specific budget, expertise, and goals of the end-user and their biosurveillance program. We found that, to date, there are no methods that can be easily performed at the point-of-need by non-experts. As an example, ONT sequencing methods have been shown to detect viruses (and sometimes whole viral genomes) in the field (Croville et al., 2024; de Vries et al., 2022); however, these methods require trained technicians, specialised expensive and often inaccessible equipment (e.g. see de Vries et al., 2022). We found that five methods held higher potential than others for detecting AIV in the field requiring lower budgets. Overall, methods free from nucleic acid extraction and PCR steps will ultimately allow for more rapid, sensitive, and cheap ways of detecting viruses in the field.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Allison Miller and Jemma Geoghegan: concept and design. Allison Miller: method development. Allison Miller – original draft. Allison Miller, Eddy Dowle, Benjamin Duran-Vinet, Jo-Ann Stanton, Vinko Besic, Jackson Treece, Stephanie Waller, Gert-Jan Jeunen, David Winter, Neil Gemmell, Jemma Geoghegan: manuscript preparation – review and editing. Jemma Geoghegan and Neil Gemmell: supervision and administration. David Winter and Jemma Geoghegan - Funding.
Ethics Statement
The research meets the ethical guidelines of the study country and the manuscript has not been submitted elsewhere.
Acknowledgments
This work was funded by a project grant (TN/SWC/24/UoOJG) from Te Niwha, New Zealand’s Infectious Disease Research Platform co-hosted by the Institute of Environmental Science and Research and the University of Otago. N.J.G. and B.D.-V. also thank the Ministry of Business, Innovation, and Employment (MBIE) project: A toolbox to underpin and enable tomorrow’s marine biosecurity system (MBIE CAWX1904).
Conflicts of Interest
JS is currently COO of eXymes PLC, a nucleic acid extraction company.
References
- Ackerman, C. M., C. Myhrvold, S. G. Thakku, C. A. Freije, H. C. Metsky, D. K. Yang, and J. Kehe. 2020. Massively multiplexed nucleic acid detection with Cas13. Nature 582, 7811: 277–282. [Google Scholar] [CrossRef] [PubMed]
- Adlhoch, C., F. Baldinelli, A. Fusaro, and C. Terregino. 2022. Avian influenza, a new threat to public health in Europe? Clinical Microbiology and Infection 28, 2: 149–151. [Google Scholar] [CrossRef]
- Alexander, D. J. 2001. Newcastle disease. British poultry science 42, 1: 5–22. [Google Scholar] [CrossRef]
- Alfano, N., A. Dayaram, J. Axtner, K. Tsangaras, M. L. Kampmann, A. Mohamed, and A. D. Greenwood. 2021. Non-invasive surveys of mammalian viruses using environmental DNA. Methods in Ecology and Evolution 12, 10: 1941–1952. [Google Scholar] [CrossRef]
- Baerwald, M. R., A. M. Goodbla, R. P. Nagarajan, J. S. Gootenberg, O. O. Abudayyeh, F. Zhang, and A. D. Schreier. 2020. Rapid and accurate species identification for ecological studies and monitoring using CRISPR-based SHERLOCK. Molecular Ecology Resources 20, 4: 961–970. [Google Scholar] [CrossRef]
- Barnes, K. G., A. E. Lachenauer, A. Nitido, S. Siddiqui, R. Gross, B. Beitzel, and S. B. Mehta. 2020. Deployable CRISPR-Cas13a diagnostic tools to detect and report Ebola and Lassa virus cases in real-time. Nature communications 11, 1: 4131. [Google Scholar] [CrossRef]
- Bass, D., K. W. Christison, G. D. Stentiford, L. S. Cook, and H. Hartikainen. 2023. Environmental DNA/RNA for pathogen and parasite detection, surveillance, and ecology. Trends in Parasitology 39, 4: 285–304. [Google Scholar] [CrossRef] [PubMed]
- Beng, K. C., and R. T. Corlett. 2020. Applications of environmental DNA (eDNA) in ecology and conservation: opportunities, challenges and prospects. Biodiversity and Conservation 29, 7: 2089–2121. [Google Scholar] [CrossRef]
- Bessière, P., B. Hayes, F. Filaire, L. Lèbre, T. Vergne, M. Pinson, and J.-L. Guérin. 2023. Optimizing environmental viral surveillance: bovine serum albumin increases RT-qPCR sensitivity for high pathogenicity avian influenza H5Nx virus detection from dust samples. Microbiology Spectrum 11, 6: e03055-23. [Google Scholar] [CrossRef]
- Bhatt, A., Z. Fatima, M. Ruwali, C. S. Misra, S. S. Rangu, D. Rath, and S. Hameed. 2022. CLEVER assay: A visual and rapid RNA extraction-free detection of SARS-CoV-2 based on CRISPR-Cas integrated RT-LAMP technology. Journal of Applied Microbiology 133, 2: 410–421. [Google Scholar] [CrossRef]
- Bi, Y., J. Yang, L. Wang, L. Ran, and G. F. Gao. 2024. Ecology and evolution of avian influenza viruses. Current biology 34, 15: R716–R721. [Google Scholar] [CrossRef] [PubMed]
- Billington, C., G. Abeysekera, P. Scholes, P. Pickering, and L. Pang. 2021. Utility of a field deployable qPCR instrument for analyzing freshwater quality. Agrosystems, Geosciences & Environment 4, 4: e20223. [Google Scholar]
- Bivins, A., D. Kaya, W. Ahmed, J. Brown, C. Butler, J. Greaves, and S. Sherchan. 2022. Passive sampling to scale wastewater surveillance of infectious disease: Lessons learned from COVID-19. Science of the Total Environment 835: 155347. [Google Scholar] [CrossRef] [PubMed]
- Bowers, H. A., X. Pochon, U. von Ammon, N. Gemmell, J.-A. L. Stanton, G.-J. Jeunen, and A. Zaiko. 2021. Towards the optimization of eDNA/eRNA sampling technologies for marine biosecurity surveillance. Water 13, 8: 1113. [Google Scholar] [CrossRef]
- Boykin, L. M., P. Sseruwagi, T. Alicai, E. Ateka, I. U. Mohammed, J.-A. L. Stanton, and J. Erasto. 2019. Tree lab: Portable genomics for early detection of plant viruses and pests in sub-saharan africa. Genes 10, 9: 632. [Google Scholar] [CrossRef]
- Broughton, J. P., X. Deng, G. Yu, C. L. Fasching, V. Servellita, J. Singh, and A. Sotomayor-Gonzalez. 2020. CRISPR–Cas12-based detection of SARS-CoV-2. Nature biotechnology 38, 7: 870–874. [Google Scholar] [CrossRef]
- Brown, T. A., K. S. Schaefer, A. Tsang, H. A. Yi, J. B. Grimm, A. L. Lemire, and K. Ritola. 2021. Direct detection of SARS-CoV-2 RNA using high-contrast pH-sensitive dyes. Journal of biomolecular techniques: JBT 32, 3: 121. [Google Scholar] [CrossRef]
- Campbell, M. L., and C. L. Hewitt. 2025. A holistic marine biosecurity risk framework that is inclusive of social, cultural, economic and ecological values. Marine Policy 172: 106511. [Google Scholar] [CrossRef]
- Chen, J. S., E. Ma, L. B. Harrington, M. Da Costa, X. Tian, J. M. Palefsky, and J. A. Doudna. 2018. CRISPR-Cas12a target binding unleashes indiscriminate single-stranded DNase activity. Science 360, 6387: 436–439. [Google Scholar] [CrossRef]
- Chen, S.-S., Y.-L. Yang, H.-Y. Wang, T.-K. Guo, R.-M. Azeem, C.-W. Shi, and J.-Z. Wang. 2024. CRISPR/Cas13a-based genome editing for establishing the detection method of H9N2 subtype avian influenza virus. Poultry Science 103, 10: 104068. [Google Scholar] [CrossRef]
- Croville, G., M. Walch, A. Sécula, L. Lèbre, S. Silva, F. Filaire, and J.-L. Guerin. 2024. An amplicon-based nanopore sequencing workflow for rapid tracking of avian influenza outbreaks, France. Frontiers in Cellular and Infection Microbiology 14. [Google Scholar] [CrossRef] [PubMed]
- de Jonge, E. F., C. M. Peterse, J. M. Koelewijn, A.-M. R. van der Drift, R. F. van der Beek, E. Nagelkerke, and W. J. Lodder. 2022. The detection of monkeypox virus DNA in wastewater samples in the Netherlands. Science of the Total Environment 852: 158265. [Google Scholar] [CrossRef] [PubMed]
- de Vries, E. M., N. O. I. Cogan, A. J. Gubala, P. T. Mee, K. J. O’Riley, B. C. Rodoni, and S. E. Lynch. 2022. Rapid, in-field deployable, avian influenza virus haemagglutinin characterisation tool using MinION technology. Scientific reports 12, 1: 11886. [Google Scholar] [CrossRef]
- Deng, Q., Y. Cao, X. Wan, B. Wang, A. Sun, H. Wang, and H. Gu. 2022. Nanopore-based metagenomic sequencing for the rapid and precise detection of pathogens among immunocompromised cancer patients with suspected infections. Frontiers in Cellular and Infection Microbiology 12: 943859. [Google Scholar] [CrossRef]
- Ding, J., X. Xu, Y. Deng, X. Zheng, and T. Zhang. 2024. Comparison of RT-ddPCR and RT-qPCR platforms for SARS-CoV-2 detection: Implications for future outbreaks of infectious diseases. Environment International 183: 108438. [Google Scholar] [CrossRef]
- Farrell, J. A., K. Yetsko, L. Whitmore, J. Whilde, C. B. Eastman, D. R. Ramia, and B. Burkhalter. 2021. Environmental DNA monitoring of oncogenic viral shedding and genomic profiling of sea turtle fibropapillomatosis reveals unusual viral dynamics. Communications Biology 4, 1: 565. [Google Scholar] [CrossRef] [PubMed]
- Fong, T.-T., and E. K. Lipp. 2005. Enteric viruses of humans and animals in aquatic environments: health risks, detection, and potential water quality assessment tools. Microbiology and molecular biology reviews 69, 2: 357–371. [Google Scholar] [CrossRef]
- Fu, X., Q. Wang, B. Ma, B. Zhang, K. Sun, X. Yu, and M. Zhang. 2023. Advances in detection techniques for the H5N1 avian influenza virus. International journal of molecular sciences 24, 24: 17157. [Google Scholar] [CrossRef]
- Geiss, G. K., R. E. Bumgarner, B. Birditt, T. Dahl, N. Dowidar, D. L. Dunaway, and T. Grogan. 2008. Direct multiplexed measurement of gene expression with color-coded probe pairs. Nature biotechnology 26, 3: 317–325. [Google Scholar] [CrossRef]
- Gootenberg, J. S., O. O. Abudayyeh, M. J. Kellner, J. Joung, J. J. Collins, and F. Zhang. 2018. Multiplexed and portable nucleic acid detection platform with Cas13, Cas12a, and Csm6. Science 360, 6387: 439–444. [Google Scholar] [CrossRef]
- Griffin, D. W., E. K. Lipp, M. R. Mclaughlin, and J. B. Rose. 2001. Marine Recreation and Public Health Microbiology: Quest for the Ideal Indicator: This article addresses the historic, recent, and future directions in microbiological water quality indicator research. Bioscience 51, 10: 817–825. [Google Scholar] [CrossRef]
- Hood, G., X. Roche, A. Brioudes, S. von Dobschuetz, F. O. Fasina, W. Kalpravidh, and L. Sims. 2021. A literature review of the use of environmental sampling in the surveillance of avian influenza viruses. Transboundary and emerging diseases 68, 1: 110–126. [Google Scholar] [CrossRef] [PubMed]
- Huang, M., S. Liu, Y. Xu, A. Li, W. Wu, M. Liang, and T. Wang. 2022. CRISPR/Cas12a technology combined with RPA for rapid and portable SFTSV detection. Frontiers in Microbiology 13: 754995. [Google Scholar] [CrossRef] [PubMed]
- Hubbard, L. E., C. E. Givens, E. A. Stelzer, M. L. Killian, D. W. Kolpin, C. M. Szablewski, and R. L. Poulson. 2023. Environmental surveillance and detection of infectious highly pathogenic avian influenza virus in Iowa wetlands. Environmental Science & Technology Letters 10, 12: 1181–1187. [Google Scholar]
- Hubbard, L. E., E. A. Stelzer, R. L. Poulson, D. W. Kolpin, C. M. Szablewski, and C. E. Givens. 2024. Development of a Large-Volume Concentration Method to Recover Infectious Avian Influenza Virus from the Aquatic Environment. Viruses 16, 12: 1898. [Google Scholar] [CrossRef]
- Huver, J., J. Koprivnikar, P. Johnson, and S. Whyard. 2015. Development and application of an eDNA method to detect and quantify a pathogenic parasite in aquatic ecosystems. Ecological applications 25, 4: 991–1002. [Google Scholar] [CrossRef] [PubMed]
- Jain, S., D. S. Dandy, B. J. Geiss, and C. S. Henry. 2021. Padlock probe-based rolling circle amplification lateral flow assay for point-of-need nucleic acid detection. Analyst 146, 13: 4340–4347. [Google Scholar] [CrossRef]
- Jallow, M. M., M. M. Diagne, M. H. D. Ndione, M. A. Barry, N. K. Ndiaye, D. E. Kiori, and M. Fall. 2025. Genetic and Molecular Characterization of Avian Influenza A (H9N2) Viruses from Live Bird Markets (LBM) in Senegal. Viruses 17, 1: 73. [Google Scholar] [CrossRef]
- Ji, P., T. G. Aw, W. Van Bonn, and J. B. Rose. 2020. Evaluation of a portable nanopore-based sequencer for detection of viruses in water. Journal of Virological Methods 278: 113805. [Google Scholar] [CrossRef]
- Jones, R. A. 2021. Global plant virus disease pandemics and epidemics. Plants 10, 2: 233. [Google Scholar] [CrossRef]
- Kaneko, H., T. Iida, K. Aoki, S. Ohno, and T. Suzutani. 2005. Sensitive and rapid detection of herpes simplex virus and varicella-zoster virus DNA by loop-mediated isothermal amplification. Journal of Clinical Microbiology 43, 7: 3290–3296. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y., J. Wang, W. Zhang, Y. Xu, B. Xu, G. Qu, and G. Su. 2023. RNA extraction-free workflow integrated with a single-tube CRISPR-Cas-based colorimetric assay for rapid SARS-CoV-2 detection in different environmental matrices. Journal of Hazardous Materials 454: 131487. [Google Scholar] [CrossRef]
- Kawato, Y., T. Mekata, M. Inada, and T. Ito. 2021. Application of environmental DNA for monitoring Red Sea bream Iridovirus at a fish farm. Microbiology Spectrum 9, 2: e00796-21. [Google Scholar] [CrossRef]
- Kfir, R., and B. Genthe. 1993. Advantages and disadvantages of the use of immunodetection techniques for the enumeration of microorganisms and toxins in water. Water Science and Technology 27, 3–4: 243–252. [Google Scholar] [CrossRef]
- Kuchinski, K. S., M. Coombe, S. C. Mansour, G. A. P. Cortez, M. Kalhor, C. G. Himsworth, and N. A. Prystajecky. 2024. Targeted genomic sequencing of avian influenza viruses in wetland sediment from wild bird habitats. Applied and Environmental Microbiology 90, 2: e00842-23. [Google Scholar] [CrossRef] [PubMed]
- Kumblathan, T., Y. Liu, G. K. Uppal, S. E. Hrudey, and X.-F. Li. 2021. Wastewater-based epidemiology for community monitoring of SARS-CoV-2: progress and challenges. ACS Environmental Au 1, 1: 18–31. [Google Scholar] [CrossRef] [PubMed]
- Lalli, M. A., J. S. Langmade, X. Chen, C. C. Fronick, C. S. Sawyer, L. C. Burcea, and W. J. Buchser. 2021. Rapid and extraction-free detection of SARS-CoV-2 from saliva by colorimetric reverse-transcription loop-mediated isothermal amplification. Clinical Chemistry 67, 2: 415–424. [Google Scholar] [CrossRef]
- Lin, Y., Y. Dai, Y. Liu, Z. Ren, H. Guo, Z. Li, and S. Zhang. 2022. Rapid PCR-based nanopore adaptive sequencing improves sensitivity and timeliness of viral clinical detection and genome surveillance. Frontiers in Microbiology 13: 929241. [Google Scholar] [CrossRef]
- Louis, S. 2024. Wastewater surveillance for influenza A virus and H5 subtype concurrent with the highly pathogenic avian influenza A (H5N1) virus outbreak in cattle and poultry and associated human cases—United States, May 12–July 13, 2024. MMWR. Morbidity and Mortality Weekly Report 73. [Google Scholar] [CrossRef]
- Metcalf, T., J. Melnick, and M. Estes. 1995. Environmental virology: from detection of virus in sewage and water by isolation to identification by molecular biology-a trip of over 50 years. Annual review of microbiology 49: 461–488. [Google Scholar] [CrossRef]
- Miao, J., L. Zuo, D. He, Z. Fang, N. Berthet, C. Yu, and G. Wong. 2023. Rapid detection of Nipah virus using the one-pot RPA-CRISPR/Cas13a assay. Virus research 332: 199130. [Google Scholar] [CrossRef] [PubMed]
- Misra, C. S., S. S. Rangu, R. D. Phulsundar, G. Bindal, M. Singh, R. Shashidhar, and D. Rath. 2022. An improved, simple and field-deployable CRISPR-Cas12a assay for the detection of SARS-CoV-2. Journal of Applied Microbiology 133, 4: 2668–2677. [Google Scholar] [CrossRef] [PubMed]
- Msemburi, W., A. Karlinsky, V. Knutson, S. Aleshin-Guendel, S. Chatterji, and J. Wakefield. 2023. The WHO estimates of excess mortality associated with the COVID-19 pandemic. Nature 613, 7942: 130–137. [Google Scholar] [CrossRef]
- Mullis, K. B., and F. A. Faloona. 1987. [21] Specific synthesis of DNA in vitro via a polymerase-catalyzed chain reaction. In Methods in enzymology. Elsevier: Vol. 155, pp. 335–350. [Google Scholar] [CrossRef]
- Myhrvold, C., C. A. Freije, J. S. Gootenberg, O. O. Abudayyeh, H. C. Metsky, A. F. Durbin, and L. A. Parham. 2018. Field-deployable viral diagnostics using CRISPR-Cas13. Science 360, 6387: 444–448. [Google Scholar] [CrossRef] [PubMed]
- Nabeshima, K., S. Asakura, R. Iwata, H. Honjo, A. Haga, K. Goka, and M. Onuma. 2023. Sequencing methods for HA and NA genes of avian influenza viruses from wild bird feces using Oxford Nanopore sequencing. Comparative Immunology, Microbiology and Infectious Diseases 102: 102076. [Google Scholar] [CrossRef]
- Neumann, G., and Y. Kawaoka. 2024. Highly pathogenic H5N1 avian influenza virus outbreak in cattle: the knowns and unknowns. Nature Reviews Microbiology 22, 9: 525–526. [Google Scholar] [CrossRef]
- Nie, M., Y. Zhou, F. Li, H. Deng, M. Zhao, Y. Huang, and L. Zhu. 2022. Epidemiological investigation of swine Japanese encephalitis virus based on RT-RAA detection method. Scientific reports 12, 1: 9392. [Google Scholar] [CrossRef]
- Notomi, T., H. Okayama, H. Masubuchi, T. Yonekawa, K. Watanabe, N. Amino, and T. Hase. 2000. Loop-mediated isothermal amplification of DNA. Nucleic Acids Research 28, 12: e63. [Google Scholar] [CrossRef]
- Nwe, M. K., N. Jangpromma, and L. Taemaitree. 2024. Evaluation of molecular inhibitors of loop-mediated isothermal amplification (LAMP). Scientific reports 14, 1: 5916. [Google Scholar] [CrossRef]
- Oldstone, M. B. 2020. Viruses, plagues, and history: past, present, and future. Oxford University Press. [Google Scholar]
- Parameswari, B., P. Anbazhagan, A. Rajashree, G. Chaitra, K. Sidharthan, S. Mangrauthia, and B. Bhaskar. 2025. Development of reverse transcription recombinase polymerase amplification assay for rapid diagnostics of Peanut mottle virus. Physiology and Molecular Biology of Plants 31, 1: 131–142. [Google Scholar] [CrossRef]
- Piepenburg, O., C. H. Williams, D. L. Stemple, and N. A. Armes. 2006. DNA detection using recombination proteins. PLoS biology 4, 7: e204. [Google Scholar] [CrossRef] [PubMed]
- Pillay, S., D. Calderón-Franco, and T. Abeel. 2024. Portable In-Field DNA Sequencing for Rapid Detection of Pathogens and Antimicrobial Resistance: A Proof-of-Concept Study. bioRxiv. [Google Scholar]
- Plaza, P. I., V. Gamarra-Toledo, J. R. Euguí, and S. A. Lambertucci. 2024. Recent changes in patterns of mammal infection with highly pathogenic avian influenza A (H5N1) virus worldwide. Emerging Infectious Diseases 30, 3: 444. [Google Scholar] [CrossRef]
- Quick, J., N. D. Grubaugh, S. T. Pullan, I. M. Claro, A. D. Smith, K. Gangavarapu, and N. A. Beutler. 2017. Multiplex PCR method for MinION and Illumina sequencing of Zika and other virus genomes directly from clinical samples. Nature protocols 12, 6: 1261–1276. [Google Scholar] [CrossRef] [PubMed]
- Rački, N., T. Dreo, I. Gutierrez-Aguirre, A. Blejec, and M. Ravnikar. 2014. Reverse transcriptase droplet digital PCR shows high resilience to PCR inhibitors from plant, soil and water samples. Plant methods 10: 1–10. [Google Scholar] [CrossRef]
- Ramírez-Córdova, C., D. Morales-Jadán, S. Alarcón-Salem, A. Sarmiento-Alvarado, M. B. Proaño, I. Camposano, and D. Coello. 2023. Fast, cheap and sensitive: Homogenizer-based RNA extraction free method for SARS-CoV-2 detection by RT-qPCR. Frontiers in Cellular and Infection Microbiology 13: 1074953. [Google Scholar] [CrossRef]
- Sahu, A., N. Kumar, C. P. Singh, and M. Singh. 2023. Environmental DNA (eDNA): Powerful technique for biodiversity conservation. Journal for Nature Conservation 71: 126325. [Google Scholar] [CrossRef]
- Schellenberg, J. J., M. Ormond, and Y. Keynan. 2021. Extraction-free RT-LAMP to detect SARS-CoV-2 is less sensitive but highly specific compared to standard RT-PCR in 101 samples. Journal of Clinical Virology 136: 104764. [Google Scholar] [CrossRef]
- Scriver, M., U. von Ammon, C. Youngbull, X. Pochon, J.-A. L. Stanton, N. J. Gemmell, and A. Zaiko. 2024. Drop it all: extraction-free detection of targeted marine species through optimized direct droplet digital PCR. PeerJ 12: e16969. [Google Scholar] [CrossRef]
- Smith, C. J., and A. M. Osborn. 2009. Advantages and limitations of quantitative PCR (Q-PCR)-based approaches in microbial ecology. FEMS microbiology ecology 67, 1: 6–20. [Google Scholar] [CrossRef]
- Stanton, J.-A. L., A. Muralidhar, C. J. Rand, and D. J. Saul. 2019. Rapid extraction of DNA suitable for NGS workflows from bacterial cultures using the PDQeX. Biotechniques 66, 5: 208–213. [Google Scholar] [CrossRef]
- Struijk, R., A. van den Ouden, J. Louwerse, K. Čurová, R. Burggrave, B. McNally, and G. de Vos. 2023. Ultrafast RNA extraction-free SARS-CoV-2 detection by direct RT-PCR using a rapid thermal cycling approach. Diagnostic Microbiology and Infectious Disease 107, 1: 115975. [Google Scholar] [CrossRef] [PubMed]
- Temin, H. M., and S. Mizutami. 1970. RNA-dependent DNA polymerase in virions of Rous sarcoma virus.
- Thalinger, B., K. Deiner, L. R. Harper, H. C. Rees, R. C. Blackman, D. Sint, and K. Bruce. 2021. A validation scale to determine the readiness of environmental DNA assays for routine species monitoring. Environmental DNA 3, 4: 823–836. [Google Scholar] [CrossRef]
- Tisza, M. J., B. Hanson, J. R. Clark, L. Wang, K. Payne, M. C. Ross, and J. J. Cormier. 2024. Virome Sequencing Identifies H5N1 Avian Influenza in Wastewater from Nine Cities. medRxiv. [Google Scholar]
- Vogels, C. B., M. I. Breban, I. M. Ott, T. Alpert, M. E. Petrone, A. E. Watkins, and J. Goes de Jesus. 2021. Multiplex qPCR discriminates variants of concern to enhance global surveillance of SARS-CoV-2. PLoS biology 19, 5: e3001236. [Google Scholar] [CrossRef]
- Wang, Y., H. Chen, K. Lin, Y. Han, Z. Gu, H. Wei, and R. Jin. 2024. Ultrasensitive single-step CRISPR detection of monkeypox virus in minutes with a vest-pocket diagnostic device. Nature communications 15, 1: 3279. [Google Scholar] [CrossRef]
- Wei, N., B. Zheng, J. Niu, T. Chen, J. Ye, Y. Si, and S. Cao. 2022. Rapid detection of genotype II African swine fever virus using CRISPR Cas13a-based lateral flow strip. Viruses 14, 2: 179. [Google Scholar] [CrossRef]
- Wolfe, M. K., D. Duong, B. Shelden, E. M. Chan, V. Chan-Herur, S. Hilton, and B. J. White. 2024. Detection of hemagglutinin H5 influenza A virus sequence in municipal wastewater solids at wastewater treatment plants with increases in influenza A in spring, 2024. Environmental Science & Technology Letters. [Google Scholar]
- Wolfe, M. K., A. H. Paulos, A. Zulli, D. Duong, B. Shelden, B. J. White, and A. B. Boehm. 2023. Wastewater detection of emerging arbovirus infections: Case study of Dengue in the United States. Environmental Science & Technology Letters 11, 1: 9–15. [Google Scholar]
- Wong, Y. P., S. Othman, Y. L. Lau, S. Radu, and H. Y. Chee. 2018. Loop-mediated isothermal amplification (LAMP): a versatile technique for detection of micro-organisms. Journal of Applied Microbiology 124, 3: 626–643. [Google Scholar] [CrossRef]
- Wu, H., X. Cao, Y. Meng, D. Richards, J. Wu, Z. Ye, and A. J. deMello. 2022. DropCRISPR: A LAMP-Cas12a based digital method for ultrasensitive detection of nucleic acid. Biosensors and Bioelectronics 211: 114377. [Google Scholar] [CrossRef] [PubMed]
- Zahra, A., A. Shahid, A. Shamim, S. H. Khan, and M. I. Arshad. 2023. The SHERLOCK platform: an insight into advances in viral disease diagnosis. Molecular Biotechnology 65, 5: 699–714. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y., X. Wu, A. Gu, L. Dobelle, C. A. Cid, J. Li, and M. R. Hoffmann. 2021. Membrane-based in-gel loop-mediated isothermal amplification (mgLAMP) system for SARS-CoV-2 quantification in environmental waters. Environmental Science & Technology 56, 2: 862–873. [Google Scholar]
Figure 1.
Workflow summarising the 12 methods evaluated. Coloured arrows correspond to 1 of the 12 environmental viral detection methods. The 11 criteria we use to assess the methods are listed in the column on the left and the corresponding responses are listed in the middle. These middle responses range from more advantageous to less advantages going from left to right (e.g., the response of “No” for “Nucleic acid extraction required?” is more advantageous than the “Yes” response). See Supplementary Table S1 for further detail. Figure created with Lucidchart and Biorender.
Figure 1.
Workflow summarising the 12 methods evaluated. Coloured arrows correspond to 1 of the 12 environmental viral detection methods. The 11 criteria we use to assess the methods are listed in the column on the left and the corresponding responses are listed in the middle. These middle responses range from more advantageous to less advantages going from left to right (e.g., the response of “No” for “Nucleic acid extraction required?” is more advantageous than the “Yes” response). See Supplementary Table S1 for further detail. Figure created with Lucidchart and Biorender.
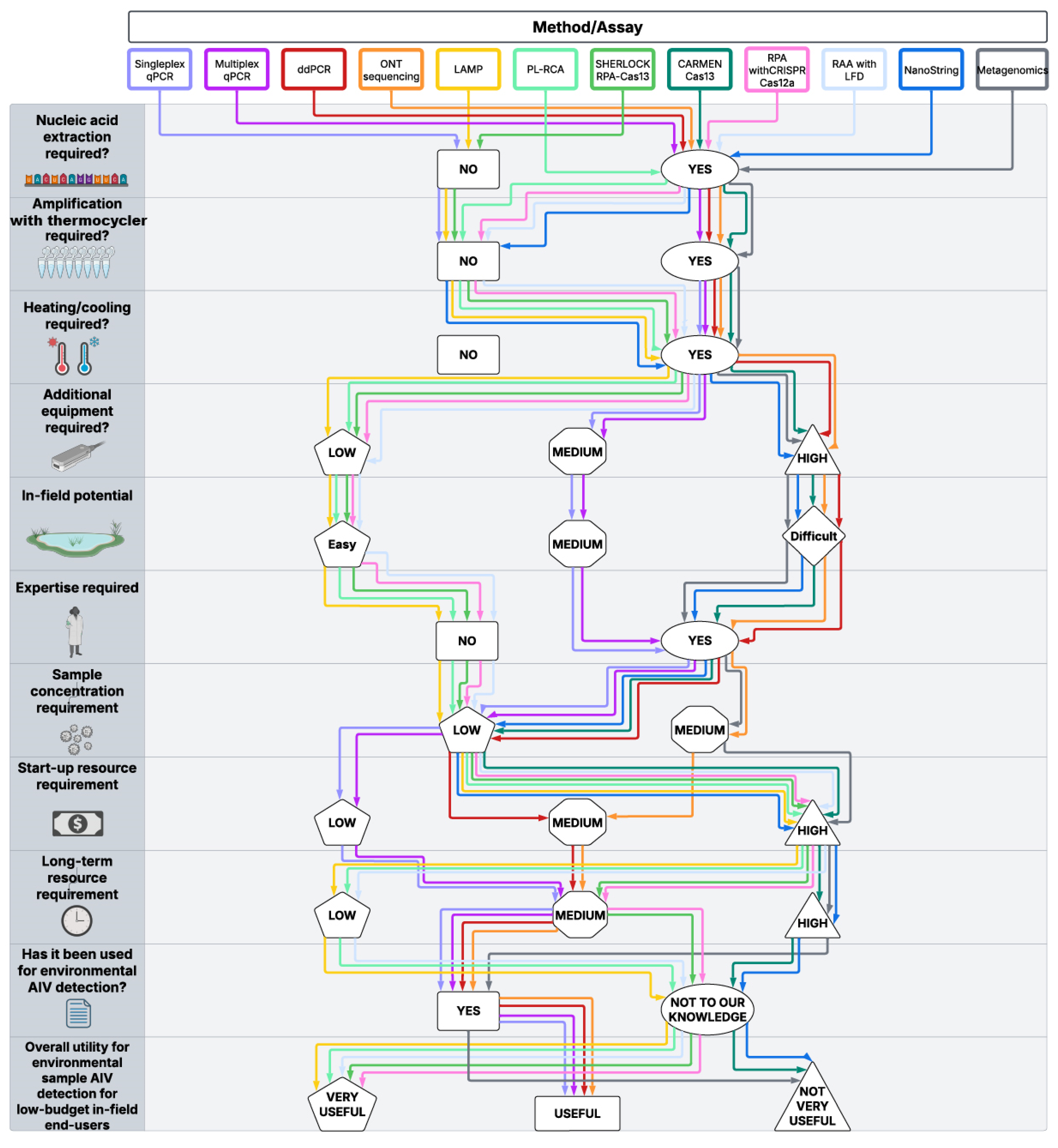
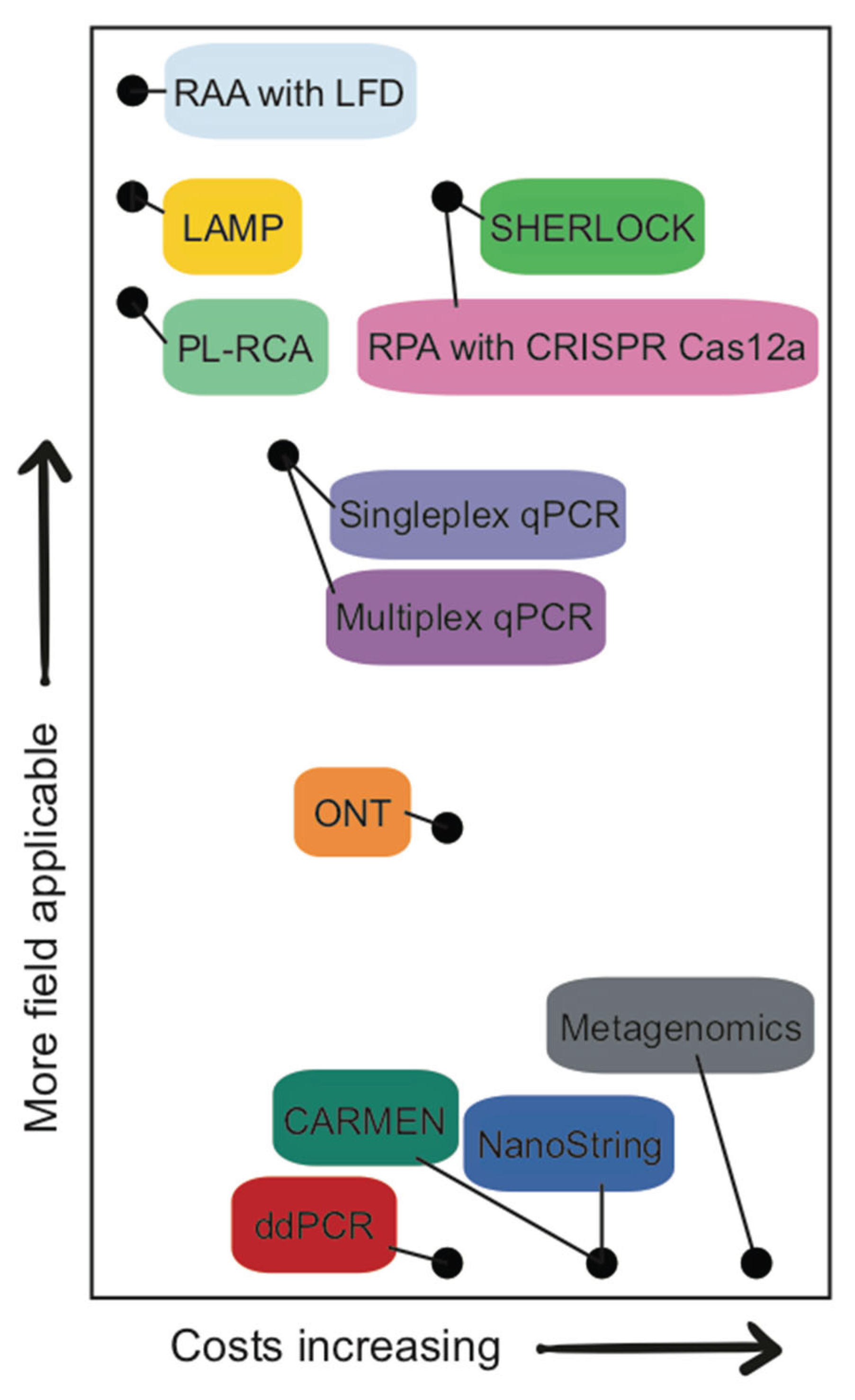
Figure 2.
Cost and in-field potential of environmental viral detection methods. Twelve methods plotted by long-term resource cost and their ability to be deployed in the field using information from Supplementary Table S1. Colours correspond to the methods listed in Figure 1.
Figure 2.
Cost and in-field potential of environmental viral detection methods. Twelve methods plotted by long-term resource cost and their ability to be deployed in the field using information from Supplementary Table S1. Colours correspond to the methods listed in Figure 1.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



