Submitted:
30 May 2025
Posted:
04 June 2025
You are already at the latest version
Abstract
Background: Asthma, a heterogeneous chronic inflammatory airway disorder, imposes a substantial global health burden. Molecular stratification of asthma phenotypes holds promise for elucidating pathobiological pathways and advancing precision therapeutic interventions. Currently, few biomarkers can distinguish between allergic and non-allergic asthma beyond cell concentrations in blood. While inflammatory factors play pivotal roles in asthma's development and progression, the exact molecular interplay driving phenotypic divergence remains incompletely characterized. This study aimed to explore the differences in inflammation-related proteins between allergic and non-allergic asthma. Methods: Utilizing Olink proximity extension assay technology, we conducted comprehensive profiling of 92 inflammation-related proteins in plasma samples from allergic asthma (n=10) and non-allergic asthma (n=15) cohorts. The analysis included the identification of differentially expressed proteins (DEPs), correlation analysis between DEPs and clinical signs, protein-protein interaction (PPI) analysis, area under the receiver operating characteristic curve (AUC) analysis, and function enrichment analysis. Result: Our findings revealed significant upregulation of 17 DEPs in the allergic asthma group compared to the non-allergic asthma group. Proteins such as CD244, CASP-8, TRANCE, SIRT2, and STAMBP exhibited relatively high AUC values (AUC>0.8). Correlation analysis demonstrated that TNFRSF9 was positively associated with eosinophil (%) and VEGFA emerged as a central hub protein in PPI analysis. Furthermore, GO and KEGG enrichment analysis suggested that the DEPs were primarily enriched in biological processes linked to immune response, cytokine-mediated signaling pathways, and cytokine-cytokine receptor interactions. Conclusions: Our results highlight significant differences in the expression of 17 proteins between allergic and non-allergic asthma groups, suggesting differential involvement of inflammation networks and immune pathways. The identified proteins, particularly TRANCE showing diagnostic promise, may represent potential biomarkers for endotype stratification and therapeutic targeting. These findings advance our understanding of heterogeneous immunopathological mechanisms underlying asthma subtypes.
Keywords:
Inflammation
; biomarkers
; allergic asthma
; non-allergic asthma
; Olink proteomics platform
1. Introduction
Asthma is a widespread and heterogeneous chronic inflammatory disease of the airways. Globally, it impacts approximately 339 million individuals, imposing a significant economic and health burden [1]. This disease is primarily categorized by clinical manifestations and distinct pathophysiological mechanisms into two primary subtypes: allergic and non-allergic asthma. The allergic phenotype, accounting for approximately 60% of cases across pediatric and adult populations, demonstrates strong association with T-helper 2 (Th2) lymphocyte-driven immunity [2,3]. In this subtype, environmental triggers including allergens and pollutants activate allergen-specific Th2 cells to secrete type 2 cytokines (IL-4, IL-5, IL-13), initiating a pathological cascade characterized by eosinophilic infiltration, IgE-mediated hypersensitivity, and mucus hypersecretion [4]. In contrast, non-allergic asthma presents greater mechanistic complexity, featuring less well-defined inflammatory pathways. Emerging evidence implicates group 2 innate lymphoid cells (ILC2s) as pivotal contributors to disease pathogenesis in this subtype [5].
Accurate differentiation between these phenotypes carries paramount clinical importance, as phenotype-directed therapeutic strategies informed by molecular mechanisms demonstrate superior clinical efficacy [6]. However, current diagnostic approaches, including combined assessment of allergen sensitivity panels, total IgE levels, fractional exhaled nitric oxide (FeNO), eosinophil counts in peripheral blood and sputum, show limited discriminative capacity for reliable phenotype stratification. This diagnostic challenge stems from substantial symptom overlap between subtypes and the inherent biological variability of asthma manifestations [7]. The identification of biomarkers holds significant potential for distinguishing clinically relevant asthma phenotypes and advancing targeted therapeutic strategies [8]. This underscores the urgency for developing asthma biomarkers rooted in pathophysiological mechanisms.
Asthma pathophysiology involves complex immune system interactions, with inflammatory cytokines serving as pivotal mediators in disease pathogenesis. In allergic asthma, IL-5 orchestrates eosinophil maturation, activation, and survival, while IL-13 drives mucus hypersecretion and airway remodeling. Conversely, non-allergic asthma appears mediated by IL-17 and IL-8 through neutrophil recruitment and inflammatory activation [9]. Nevertheless, the cytokine networks governing these asthma subtypes remain incompletely characterized. Furthermore, broader molecular networks encompassing chemokine signaling, growth factor pathways, and related mediators likely contribute to asthmatic pathogenesis. These knowledge gaps necessitate comprehensive investigations into inflammation-related biomarkers differentiating allergic and non-allergic asthma. The Olink proteomics platform offers a high-throughput analytical approach with exceptional sensitivity and specificity for multiplex protein quantification in biological specimens.
In this study, we employed Olink proteomics platform to profile 92 inflammation-associated proteins in plasma samples from allergic and non-allergic asthma patients. Our analytical framework incorporated differentially expressed proteins (DEPs) analysis, clinical correlation assessments, protein-protein interaction (PPI) network mapping, receiver operating characteristic-area under the curve (ROC-AUC) evaluation, and functional enrichment analysis. This investigation aims to: 1) identify diagnostic biomarkers distinguishing asthma endotypes through high-resolution proteomic profiling, 2) elucidate molecular mechanisms underlying asthma heterogeneity, and 3) provide a foundation for personalized therapeutic strategies.
2. Materials and Methods
2.1. Participants
Asthma patients (n=25) comprising 10 allergic and 15 non-allergic subtypes were recruited from the Peking University Third Hospital (Beijing, China), from August 2021 to May 2024. Demographic and clinical parameters including age, gender, eosinophil (%), neutrophil (%), and allergen-specific IgE levels (kU/L) were systematically documented. The study, conducted in accordance with the Declaration of Helsinki, received ethical approval from the Institutional Review Board of Peking University Third Hospital (IRB No. M2021467), with written informed consent obtained from all participants.
Inclusion Criteria: 1) Age ≥18 years; 2) Meet the asthma diagnostic criteria in the “Global Strategy for Asthma Management and Prevention (2023 update)” report issued by GINA in 2023; 3) Asthma control status assessed by four criteria over the past four weeks: daytime symptoms >2 times/week; nighttime wheezing; rescue medication use >2 times/week; activity limitation due to asthma. The asthma control level is divided into three categories: controlled (0 criteria), partially controlled (1-2 criteria), uncontrolled (3-4 criteria). Exclusion Criteria: 1) Respiratory tract infections within the past month; 2) Concurrent pulmonary pathologies (e.g., pneumonia, tuberculosis, bronchiectasis, cystic fibrosis); 3) Major systemic comorbidities (e.g., metabolic disorders like diabetes, heart failure, acute/chronic infections). Withdrawal Criteria: 1) Voluntary withdrawal by participants; 2) Investigator-determined ineligibility for continued participation.
2.2. Sample Collection
Peripheral venous blood samples (2 mL) were collected from each patient into EDTA tubes, and were immediately centrifuged at 1500 rpm for 15 minutes at 4°C to separate the plasma. Plasma aliquots were cryopreserved at -80°C in single-use vials to maintain biomolecular integrity, with freeze-thaw cycles rigorously avoided.
2.3. Olink Proteomics Analysis
Plasma inflammation-related biomarkers were evaluated using the Olink® target 96 Inflammation panel (Olink Proteomics AB, Uppsala, Sweden) according to the manufacturer's instructions. Detailed information regarding the 92 proteins can be accessed in Table S1. This proximity extension assay utilizes dual antibody probes conjugated with unique DNA oligonucleotides for target-specific binding. Subsequent microfluidic-based proximity-dependent hybridization enables real-time PCR amplification of template DNA, with cycle threshold (Ct) values undergoing rigorous quality control and normalization through adaptive internal/external control algorithms. Normalized Protein Expression (NPX) values, representing relative quantification on a log2 scale, were generated as final outputs, where elevated NPX correlates with increased protein abundance.
2.4. Data Analysis
Differential expression analysis was conducted using the Olink® Analyze R package. Visual analytics including hierarchical clustering heatmaps and comparative volcano plots were implemented using ggplot2 (v3.4.0) and pheatmap (v1.0.12). Pearson’s correlation analysis was used to examine the correlation between DEPs, and the correlation between DEPs and clinical parameters (|r| >0.5 considered clinically relevant). Protein-protein interaction (PPI) networks were constructed using STRINGdb (v11.5) and topologically analyzed in Cytoscape (v3.9.1). Diagnostic potential was evaluated through ROC curve analysis (pROC package v1.18.0), with AUC values ≥0.7 indicating acceptable discriminative capacity. Biological characterization of DEPs was performed through Gene Ontology (GO) term enrichment and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis using the clusterProfiler R package (v4.0.5). The ggplot2 package (v3.4.0) generated graphical representations of significantly enriched terms (adjusted p-value <0.05).
Statistical analysis was conducted following manufacturer-recommended protocols (Olink), and all samples underwent log2 transformation with interplate normalization to standardized NPX values prior to Student's t-test implementation. Proteins exhibiting a p-value less than 0.05 were classified as differentially expressed.
3. Results
3.1. Clinical Characteristics of the Participants
The clinical characteristics of the two groups of patients are shown in Table 1. There were no significant differences in age distribution, eosinophil (%), and neutrophil (%) between the allergic asthma and non-allergic asthma groups. However, marked divergence was noted in immunological profiles, as allergic asthma patients demonstrated significantly elevated serum levels of allergen-specific IgE compared to their non-allergic counterparts (p<0.05). Notably, a distinct gender distribution pattern emerged from the data, with male predominance observed in the allergic asthma group (70% male) contrasting with female predominance in the non-allergic cohort (80% female).
3.2. Identification of Differentially Expressed Inflammation-Related Biomarkers
Using Olink proteomics, we systematically profiled 92 inflammation-related proteins in allergic asthma and non-allergic asthma cohorts, with expression patterns visualized in the heatmap presented in Figure 1A. Differential expression analysis revealed 17 inflammation-related proteins distinguishing the two groups (Figure 1B). Volcano plot visualization further confirmed distinct expression signatures of these DEPs (Figure 1C), all exhibiting significant upregulation in the allergic asthma group. The identified DEPs comprised TNFRSF9, TNFSF14, TRANCE, CXCL1, IL-18, OSM, CD244, CD6, VEGFA, TGF-alpha, HGF, CASP-8, STAMBP, SIRT2, 4E-BP1, ST1A1, and ADA. Comparative expression profiles of these DEPs across groups are detailed in Figure 2.
3.3. Analysis of Differentially Expressed Proteins
The correlations analysis among DEPs were quantitatively assessed using NPX values (Figure 3A). A notable positive correlation was observed between STAM-binding protein (STAMBP) and caspase-8 (CASP-8), as illustrated in Figure 3B. Moreover, the most significant positive correlation was exhibited between STAMBP and SIR2-like protein 2 (SIRT2), shown in Figure 3C.
To further investigate the connections among these proteins, a PPI network for DEPs was constructed (Figure 3D). The PPI analysis revealed that immunomodulatory molecules, including TNFSF14, TRANCE (TNFSF11) and TNFRSF9, were highly activated and strongly interconnected. Additionally, IL-18 was found to interact with cytokines and chemokines, like CXCL1 and OSM. VEGFA, HGF and TGF-alpha also demonstrated strong interactions in the PPI analysis, potentially involving PI3K-Akt signaling pathway in asthma pathology. Notably, VEGFA emerged as a central hub protein, connecting to all the other 11 proteins. These hub proteins are likely to work synergistically, influencing the expression and functionality of other proteins, and play crucial roles in the intricate biological processes associated with asthma.
ROC curve analysis was conducted for several key DEPs, as presented in Figures 3E and 3F. Proteins such as CD244, CASP-8, TRANCE, SIRT2, and STAMBP exhibited relatively high AUC values, suggesting their potential diagnostic utility in distinguishing between allergic and non-allergic asthma. Figure 3G displayed the results of cluster analysis using these five proteins, indicating that their combination possessed a certain degree of discriminatory power between the two asthma subtypes.
3.4. Correlation between Differentially Expressed Inflammation-Related Proteins and Clinical Features
A correlation analysis was performed to explore the interactions between the above-mentioned proteins and clinical characteristics (Figure 4). Only TNFRSF9 was positively associated with eosinophil (%). Five proteins, TNFRSF9, TRANCE, CASP-8, IL-18, and CD6, were positively associated with allergen-specific IgE levels. While, TNFRSF9, ST1A1, and TRANCE were negatively associated with neutrophil (%). Then, some proteins, SIRT2, STAMBP, TRANCE, CASP-8, IL-18, 4E-BP1, ADA, CD6, TNFSF14, OSM, and TGF-alpha were negatively associated with patient age. In addition, CASP-8 and 4E-BP1, were negatively associated with gender.
3.5. Function Enrichment of Differentially Expressed Proteins
GO and KEGG enrichment analysis were conducted to further investigate the potential function of the DEPs. The GO enrichment analysis (Figure 5A,B,C) revealed that the DEPs were mainly enriched in biological processes linked to immune response and cytokine-mediated signaling pathway. In the cellular component category, these proteins were primarily localized to the extracellular space and plasma membrane. In terms of molecular functions, protein binding and growth factor activity emerged as significantly enriched categories.
Furthermore, the KEGG pathway analysis (Figure 5D,E,F) showed that the DEPs were significantly enriched in key pathways, including cytokine-cytokine receptor interaction, EGFR tyrosine kinase inhibitor resistance, and PI3K-Akt signaling pathway. These pathways could potentially be implicated in the regulation of inflammatory processes and immune responses associated with asthma.
4. Discussion
Asthma represents a complex heterogeneous respiratory disorder with two primary phenotypes: allergic and non-allergic asthma. While allergic asthma typically demonstrates T-helper 2 (Th2)-biased immune activation, non-allergic asthma exhibits ambiguous inflammatory characteristics. Current diagnostic biomarkers including peripheral blood/sputum eosinophil counts, fractional exhaled nitric oxide (FeNO), and immunoglobulin E (IgE) levels present limitations: blood eosinophil counts poorly correlate with airway eosinophilic inflammation, and FeNO measurements show susceptibility to various confounders [10]. Crucially, existing biomarkers based on clinical manifestations and conventional molecular profiles lack sufficient discriminative power to differentiate these asthma subtypes, significantly hindering the implementation of phenotype-specific precision therapeutics [11]. Inflammation-related proteins hold particular promise as diagnostic discriminators due to their central role in mediating and sustaining chronic airway inflammation [12]. This highlights the need for developing sensitive, precise, and high-throughput analytical methods to quantify these inflammatory mediators. The emerging Olink proteomic platform offers distinct advantages including superior reproducibility, analytical stability, ultra-low sample requirements (down to 1 μL), and exceptional dynamic range coverage (>10 orders of magnitude). Our investigation employed the Olink Inflammation panel to comprehensively detect 92 inflammation-related proteins in allergic and non-allergic asthma cohorts. The targeted protein array encompasses critical mediators across multiple pathological pathways including inflammatory signaling, immune modulation, angiogenic processes, fibrotic remodeling, and endothelial activation mechanisms.
Inflammation-related proteins in asthma can be assayed from multiple biological sources, including sputum, peripheral blood, and respiratory tract samples. While respiratory tract specimens yield tissue-specific biomarkers, their invasive collection limits clinical utility and repeatability. Sputum analysis offers non-invasive access to airway-specific biomarkers, yet practical challenges hinder routine clinical implementation due to the need for nebulized saline induction and specialized processing for reliable quantification [13]. In contrast, blood-based biomarkers provide systemic insights into physiological and pathological states with minimal collection barriers [14]. Consequently, we prioritized plasma analysis to identify DEPs between allergic and non-allergic asthma cohorts, aiming to discover phenotype-discriminative biomarkers.
Our investigation revealed 17 DEPs in allergic asthma patients, functionally linked to critical biological processes including immune activation and cytokine-mediated signaling pathways (Figure 6). Notably, KEGG pathway analysis revealed marked enrichment of cytokine-cytokine receptor interaction pathways, underscoring their pivotal role in the observed immune dysregulation. The TNF superfamily (TNFSF) and its receptors (TNFRSF)-comprising 19 ligands and 29 receptors-emerged as key regulators, primarily exerting proinflammatory effects via NF-κB pathway activation [15,16]. This molecular family facilitates innate lymphoid cell (ILC2) activation and type 2 cytokine production, mechanisms implicated in allergic asthma pathogenesis [17]. Specifically, allergic asthma patients exhibited elevated levels of TRANCE (TNFSF11), TNFSF14, and TNFRSF9, reflecting heightened inflammatory signaling at the molecular level. PPI network analysis revealed robust connectivity among these mediators, further substantiating the pivotal role of TNFSF/TNFRSF axis dysregulation in perpetuating chronic inflammation characteristic of allergic asthma. These findings align with emerging evidence of TNFSF members influencing tumor microenvironment modulation, underscoring the pleiotropic inflammatory functions of this molecular family.
Beyond cytokine-cytokine receptor interactions, our analysis identified three additional mediators-IL-18, CXCL1, and OSM-as critically associated with allergic asthma pathogenesis. IL-18, an inflammasome-derived cytokine, binds target cell receptors to mediate pleiotropic effects in both T2- and non-T2-driven inflammation [18]. The evidence highlights its dual role as a Th2 differentiation cofactor and a driver of IgE synthesis, chronic airway inflammation, and mucus hypersecretion. Recent clinical studies further corroborate its pathogenic involvement, demonstrating elevated IL-18 levels in the plasma of asthma patients [19,20]. The chemokine CXCL1 emerged as another key player, regulating inflammatory and immune processes by orchestrating the recruitment and migration of immune cells, including neutrophils and T lymphocytes [21]. Then, mechanistic evidence linked CXCL1 to allergic airway hyperreactivity through Th2 response amplification and enhanced trafficking of immune cells to pulmonary tissues and regional lymph nodes [22]. Besides, oncostatin M (OSM)-a member of the gp130 ligand family of cytokines- exhibits strong associations with asthma severity and impaired lung function [23]. Environmental triggers such as pathogens and allergens stimulate OSM production, with its overexpression exacerbating airway inflammation and mucus hypersecretion [24]. Notably, our analysis revealed significantly elevated levels of IL-18, CXCL1, and OSM in allergic asthma patients compared to non-allergic counterparts, highlighting their specific relevance to the allergic phenotype. PPI network analysis further demonstrated robust activation and functional interconnectivity among these mediators, suggesting synergistic roles in driving allergic asthma progression through coordinated inflammatory crosstalk.
Emerging evidence suggests that immune modulatory factors such as CD244 and CD6 may contribute to asthma pathogenesis through immune regulation. CD244, a high-affinity ligand for CD48, functions as an activating receptor on eosinophils. Recent studies highlight the critical role of CD244-CD48 interactions in initiating and perpetuating allergic responses [25,26]. Mechanistically, CD244 crosslinking triggers eosinophil degranulation and subsequent release of cytokines such as IFN-γ and IL-4, potentially explaining its involvement in allergic asthma progression [27,28]. CD6, a transmembrane glycoprotein expressed predominantly on T cells and subsets of B cells, plays incompletely characterized roles in lymphocyte development, activation, and differentiation [29]. Experimental data suggest that allergen exposure induces CD6-mediated T-cell polarization toward Th2 phenotypes via interactions with activated leukocyte cell adhesion molecule (ALCAM). This pathway amplifies Th2 effector responses, driving secretion of pro-inflammatory cytokines including IFN-γ, TNF-α, IL-6, and IL-17, thereby promoting allergic airway inflammation [30]. Our findings demonstrate elevated levels of both CD244 and CD6 in allergic asthma patients compared to non-allergic counterparts, reinforcing their association with the allergic phenotype. Further investigation into the mechanistic roles of these immune regulators could advance our understanding of asthma immunopathology and inform the development of targeted therapeutic strategies.
In addition, our study implicates growth and angiogenic factors-specifically VEGFA, HGF, and TGF-alpha-in allergic asthma pathogenesis, with PPI analysis revealing their strong functional interconnectivity. Vascular endothelial growth factor A (VEGFA), a key isoform of VEGF, emerged as a central hub in the PPI network, interacting with all 11 other identified proteins. Widely recognized as a critical mediator in asthma, VEGFA promotes airway remodeling and immune cell activation within inflammatory cascades [31]. This aligns with clinical observations of elevated VEGF levels in asthmatic biofluids correlating with disease severity [32]. Transforming growth factor-alpha (TGF-alpha), potentially eosinophil-derived, regulates cellular proliferation and differentiation processes. While prior work associates TGF-alpha overexpression in airway basal cells with smoking-related asthma exacerbations [33], our data demonstrate significantly higher plasma TGF-alpha levels in allergic versus non-allergic asthmatics-suggesting an allergy-specific regulatory role beyond smoking-induced effects. Hepatocyte growth factor (HGF) is established as a critical humoral mediator in modulating inflammatory and immune responses. Emerging investigations into its role in asthma pathogenesis have yielded pivotal insights: One study identifies HGF as a central modulator in alleviating allergic asthma by restoring the balance between Th2 and Treg cells [34]. Another independent report demonstrates HGF's inhibitory effects on allergic airway inflammation, revealing its capacity to significantly suppress IL-5-stimulated secretion of TGF-β and VEGF by human eosinophils [35]. Notably, we observed upregulated HGF expression in allergic asthma patients. This discrepancy underscores the need for mechanistic studies to clarify HGF's context-dependent roles in allergic inflammation. Notably, KEGG pathway analysis positioned all three factors within the MAPK signaling cascade, suggesting their coordinated involvement in mitogen-activated inflammatory pathways. These findings collectively highlight the multifactorial nature of growth factor contributions to allergic asthma pathology.
Our investigation further identified altered expression of apoptosis- and signaling-related molecules, notably CASP-8 and STAMBP, potentially implicating these mediators in allergic asthma pathophysiology. Caspase-8 (CASP-8), traditionally recognized for its role in extrinsic apoptosis via death receptor pathways, has recently emerged as a key player in asthma-associated immune dysregulation [36,37]. Environmental allergens may activate type 2 immunity through RIPK1-CASP-8 ripoptosome formation in airway epithelia, triggering downstream caspase-3/7 activation that facilitates IL-33 maturation and release [38]. Concurrently, CASP-8 contributes to allergic inflammation through IL-1β processing and secretion, thereby promoting Th2 polarization that drives pulmonary eosinophilia and airway inflammation [39]. STAM-binding protein (STAMBP), an endosomal deubiquitinating enzyme, regulates receptor homeostasis (e.g., EGFR) and cellular proliferation [40]. Emerging evidence positions STAMBP as a modulator of innate immunity through NLRP3 inflammasome regulation and IL-1β secretion [41]. Notably, both CASP-8 and STAMBP demonstrated significant upregulation in allergic asthma patients compared to non-allergic counterparts. These findings provide preliminary insights into their roles in asthma endotypes, functional studies are required to delineate their precise contributions to allergic asthma mechanisms.
Furthermore, our study uncovered novel molecular candidates potentially linked to asthma pathogenesis. Sirtuin 2 (SIRT2), a class I histone deacetylase, emerged as a critical regulator of cellular homeostasis, oxidative stress responses, and inflammatory modulation [42]. Emerging evidence implicates SIRT2 in inflammatory disorders, with recent work suggesting its involvement in asthma pathophysiology through inflammation regulation [43,44]. Notably, we observed significant SIRT2 upregulation in allergic asthma patients, supported by its robust diagnostic potential (AUC = 0.8133). Eukaryotic translation initiation factor 4E-binding protein 1 (4E-BP1), a downstream mTOR pathway effector, regulates immune responses via IL-10 and COX-2 modulation [45,46]. While its mechanistic involvement in asthma remains uncharacterized, our observation of elevated plasma 4E-BP1 levels in allergic asthmatics suggests potential interactions with key immune regulation pathways. Sulfotransferase 1A1 (ST1A1), pivotal in phase II metabolism, mediates sulfation of diverse endogenous and exogenous substrates [47]. While ST1A1 shows no previously reported association with asthma, our data reveal its elevated plasma level in allergic versus non-allergic asthmatics, highlighting the need for functional studies to delineate its role in disease mechanisms. Intriguingly, adenosine deaminase (ADA)- a hydrolytic enzyme of adenosine metabolism -presented contradictory findings. Although prior studies report reduced ADA activity in asthma patients [48], we detected significantly increased ADA expression in allergic asthma patients. This paradoxical observation underscores the need to clarify ADA's context-dependent roles in allergic inflammation.
In our study, Olink proteomics was effectively used to discover potential biomarkers of asthma. By focusing on inflammation-related proteins, we aimed to establish discriminative signatures between allergic and non-allergic asthma subtypes, which is a critical step toward precision diagnostics and tailored therapeutic approaches. The observed differential protein expression profiles reveal distinct pathophysiological mechanisms underlying these phenotypes, providing valuable insights for mechanistic stratification. Notably, conventional Th2-associated markers (IL-4, IL-5, IL-13) showed no significant elevation in allergic asthma cohorts. This discrepancy may stem from phenotypic heterogeneity within current classification schemes, where overlapping inflammatory pathways potentially exist across subtypes. Our findings underscore the imperative to develop next-generation biomarkers that transcend traditional cytokine paradigms for improved phenotypic discrimination, necessitating refined asthma endotyping frameworks.
Despite the significant findings of this study, three key limitations warrant consideration: First, the modest cohort size necessitates validation in larger, multi-ethnic populations to confirm biomarker generalizability. Second, orthogonal validation using complementary methodologies (e.g., ELISA, Luminex) remains essential to verify proteomic findings. Third, while plasma offers practical advantages, its limited correlation with airway biology emphasizes the need for parallel analyses of localized samples (e.g., sputum) to bridge systemic and mucosal immune profiles. Future studies should prioritize comparative analyses of diverse biofluids-including blood, urine, sputum, and airway lavage samples-to establish standardized protocols that optimize biomarker yield while preserving molecular fidelity across sampling compartments.
5. Conclusions
In summary, this study employed Olink proteomics to profile inflammation-related proteins in plasma samples from allergic and non-allergic asthma patients. We identified 17 DEPs in the allergic asthma cohort, five of which (CD244, CASP-8, TRANCE, SIRT2, and STAMBP) demonstrated superior diagnostic performance (AUC>0.8), positioning them as promising discriminative biomarkers. Then, TNFRSF9 was positively associated with eosinophil (%). These candidates not only enable phenotypic differentiation but also offer novel mechanistic insights into the distinct pathophysiology of asthma subtypes. While the molecular pathways mediating their effects remain uncharacterized, our findings establish a framework for advancing asthma endotyping and precision diagnostics. Future work will focus on expanding cohort sizes to validate biomarker robustness, refining diagnostic algorithms through multi-analyte panels, and translating these discoveries into clinically actionable tools.
Supplementary Materials
The following supporting information can be downloaded at: Preprints.org, Table S1. Inflammation-related proteins panel.
Author Contributions
Wenying Wu: design, experimental studies, data acquisition, data analysis, manuscript preparation, manuscript editing. Yinfeng Wang: literature search, data analysis, manuscript preparation, and manuscript editing. Hao Zhang: definition of intellectual content, clinical studies, manuscript editing. Xiaoyan Sun: literature search and clinical studies. Ran Kong: concept, data analysis, statistical analysis, and manuscript review. Wei Yan: clinical studies, data analysis, statistical analysis, and manuscript editing.
Funding
This work was supported by Beijing Natural Science Foundation, China (7212128).
Institutional Review Board Statement
This study was approved by the Institutional Review Board of Peking University Third Hospital (IRB No. M2021467).
Informed Consent Statement
Informed consent was obtained from all individual participants included in this study.
Acknowledgments
The authors thank LC-Bio Technology CO., Ltd., Hangzhou, China, for technical assistance with the Olink analysis.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Pandey R, Parkash V, Kant S, Verma AK, Sankhwar SN, Agrawal A, et al. An update on the diagnostic biomarkers for asthma. Journal of Family Medicine and Primary Care 2021, 10:1139-1148. [CrossRef]
- Kaur R, Chupp G. Phenotypes and endotypes of adult asthma: Moving toward precision medicine. J Allergy Clin Immunol 2019, 144:1-12. [CrossRef]
- Tiotiu A. Biomarkers in asthma: state of the art. Asthma Research and Practice 2018, 4. [CrossRef]
- Stewart E, Wang X, Chupp GL, Montgomery RR. Profiling cellular heterogeneity in asthma with single cell multiparameter CyTOF. J Leukoc Biol 2020, 108:1555-1564. [CrossRef]
- Barnes PJ. Targeting cytokines to treat asthma and chronic obstructive pulmonary disease. Nature Reviews Immunology 2018, 18:454-466. [CrossRef]
- Anand, M.P. Unveiling Asthma’s Complex Tapestry: Insights from Diverse Perspectives. Diagnostics 2023, 13, 2798. [CrossRef]
- Guida G, Bagnasco D, Carriero V, Bertolini F, Ricciardolo FLM, Nicola S, et al. Critical evaluation of asthma biomarkers in clinical practice. Frontiers in Medicine 2022, 9. [CrossRef]
- Pasha MA, Hopp RJ, Habib N, Tang DD. Biomarkers in asthma, potential for therapeutic intervention. J Asthma 2024, 61:1376-1391. [CrossRef]
- Yamasaki A, Okazaki R, Harada T. Neutrophils and Asthma. Diagnostics 2022, 12, 1175. [CrossRef]
- Wang X, Lin J, Li Z, Wang M. In what area of biology has a “new” type of cell death been discovered? BBA - Reviews on Cancer 2023, 1878. [CrossRef]
- Popović-Grle S, Štajduhar A, Lampalo M, Rnjak D. Biomarkers in Different Asthma Phenotypes. Genes 2021, 12. [CrossRef]
- Januskevicius A, Vasyle E, Rimkunas A, Malakauskas K. Integrative Cross-Talk in Asthma: Unraveling the Complex Interactions Between Eosinophils, Immune, and Structural Cells in the Airway Microenvironment. Diagnostics 2024, 14, 2448. [CrossRef]
- Szefler SJ, Wenzel S, Brown R, Erzurum SC, Fahy JV, Hamilton RG, et al. Asthma outcomes: Biomarkers. J Allergy Clin Immunol 2012;129:S9-S23. [CrossRef]
- Agrawal N, Kraft M. Novel biomarkers in asthma. Curr Opin Pulm Med 2025. [CrossRef]
- Dostert C, Grusdat M, Letellier E, Brenner D. The TNF Family of Ligands and Receptors: Communication Modules in the Immune System and Beyond. Physiol Rev 2019, 99:115-160. [CrossRef]
- de Groot AF, Appelman-Dijkstra NM, van der Burg SH, Kroep JR. The anti-tumor effect of RANKL inhibition in malignant solid tumors – A systematic review. Cancer Treat Rev 2018, 62:18-28. [CrossRef]
- Matsuyama T, Salter BM, Emami Fard N, Machida K, Sehmi R. TNF Superfamily and ILC2 Activation in Asthma. Biomolecules 2024, 14. [CrossRef]
- Kaur D, Chachi L, Gomez E, Sylvius N, Brightling CE. Interleukin-18, IL-18 binding protein and IL-18 receptor expression in asthma: a hypothesis showing IL-18 promotes epithelial cell differentiation. Clin Transl Immunology 2021, 10. [CrossRef]
- Thawanaphong S, Nair A, Volfson E, Nair P, Mukherjee M. IL-18 biology in severe asthma. Front Med 2024, 11. [CrossRef]
- Xu M-H, Yuan F-L, Wang S-J, Xu H-Y, Li C-W, Tong X. Association of interleukin-18 and asthma. Inflammation 2016, 40:324-327. [CrossRef]
- Zhou C, Gao Y, Ding P, Wu T, Ji G. The role of CXCL family members in different diseases. Cell Death Discov 2023, 9. [CrossRef]
- Woo YD, Koh J, Kang H-R, Kim HY, Chung DH. The invariant natural killer T cell–mediated chemokine X-C motif chemokine ligand 1–X-C motif chemokine receptor 1 axis promotes allergic airway hyperresponsiveness by recruiting CD103+ dendritic cells. J Allergy Clin Immunol 2018, 142:1781-1792.e1712. [CrossRef]
- Headland SE, Dengler HS, Xu D, Teng G, Everett C, Ratsimandresy RA, et al. Oncostatin M expression induced by bacterial triggers drives airway inflammatory and mucus secretion in severe asthma. Sci Transl Med 2022, 14:eabf8188. [CrossRef]
- Esnault S, Bernau K, Floerke HL, Dendooven A, Delaunay E, Dill-McFarland KA, et al. Oncostatin-M Is Produced by Human Eosinophils and Expression Is Increased in Uncontrolled Severe Asthma. Allergy 2024, 80:1154-1157. [CrossRef]
- Gangwar RS, Minai-Fleminger Y, Seaf M, Gutgold A, Shikotra A, Barber C, et al. CD48 on blood leukocytes and in serum of asthma patients varies with severity. Allergy 2016, 72:888-895. [CrossRef]
- Branicka O, Jura-Szołtys E, Rogala B, Glück J. sCD48 is elevated in non-allergic but not in allergic persistent rhinitis. Immunopharmacol Immunotoxicol 2021, 43:724-730. [CrossRef]
- McArdel SL, Terhorst C, Sharpe AH. Roles of CD48 in regulating immunity and tolerance. Clin Immunol 2016, 164:10-20. [CrossRef]
- A E El-Shazly, M Henket, P P Lefebvre, R Louis. 2B4 (CD244) is involved in eosinophil adhesion and chemotaxis, and its surface expression is increased in allergic rhinitis after challenge. Int J Immunopathol Pharmacol 2011, 24(4):949-60. [CrossRef]
- Henriques SN, Oliveira L, Santos RF, Carmo AM. CD6-mediated inhibition of T cell activation via modulation of Ras. Cell Commun Signal 2022, 20. [CrossRef]
- Semitekolou M, Xanthou G. Activated Leukocyte Cell Adhesion Molecule: A Novel Regulator of Allergic Inflammation in the Airways. Am J Respir Crit Care Med 2018, 197:973-975. [CrossRef]
- Gomułka K, Liebhart J, Lange A, Mędrala W. Vascular endothelial growth factor-activated basophils in asthmatics. Postepy Dermatol Alergol 2020, 37:584-589. [CrossRef]
- Kim J-H. Serum vascular endothelial growth factor as a marker of asthma exacerbation. Korean J Intern Med 2017, 32:258-260. [CrossRef]
- Sun Y, Miao X, Zhu L, Liu J, lin Y, Xiang G, et al. Autocrine TGF-alpha is associated with Benzo(a)pyrene-induced mucus production and MUC5AC expression during allergic asthma. Ecotoxicol Environ Saf 2022, 241. [CrossRef]
- Fang Q, Wu W, Xiao Z, Zeng D, Liang R, Wang J, et al. Gingival-derived mesenchymal stem cells alleviate allergic asthma inflammation via HGF in animal models. iScience 2024, 27. [CrossRef]
- Yamauchi Y, Ueki S, Konno Y, Ito W, Takeda M, Nakamura Y, et al. The effect of hepatocyte growth factor on secretory functions in human eosinophils. Cytokine 2016, 88:45-50. [CrossRef]
- Zhang W, Zhu C, Liao Y, Zhou M, Xu W, Zou Z. Caspase-8 in inflammatory diseases: a potential therapeutic target. Cell Mol Biol Lett 2024, 29. [CrossRef]
- Pang J, Vince JE. The role of caspase-8 in inflammatory signalling and pyroptotic cell death. Semin Immunol 2023, 70. [CrossRef]
- Brusilovsky M, Rochman M, Rochman Y, Caldwell JM, Mack LE, Felton JM, et al. Environmental allergens trigger type 2 inflammation through ripoptosome activation. Nat Immunol 2021, 22:1316-1326. [CrossRef]
- Qi X, Gurung P, Malireddi RKS, Karmaus PWF, Sharma D, Vogel P, et al. Critical role of caspase-8-mediated IL-1 signaling in promoting Th2 responses during asthma pathogenesis. Mucosal Immunol 2017, 10:128-138. [CrossRef]
- Zhang J, Zhang Y, Liu Y, Zhou T, Pan G, He J, et al. STAMBP is Required for Long-Term Maintenance of Neural Progenitor Cells Derived from hESCs. Stem Cell Rev Rep 2024, 20:1932-1943. [CrossRef]
- Bednash JS, Johns F, Patel N, Smail TR, Londino JD, Mallampalli RK. The deubiquitinase STAMBP modulates cytokine secretion through the NLRP3 inflammasome. Cell Signal 2021, 79. [CrossRef]
- Wang Y, Yang J, Hong T, Chen X, Cui L. SIRT2: Controversy and multiple roles in disease and physiology. Ageing Res Rev 2019, 55. [CrossRef]
- Lee YG, Reader BF, Herman D, Streicher A, Englert JA, Ziegler M, et al. Sirtuin 2 enhances allergic asthmatic inflammation. JCI Insight 2019, 4. [CrossRef]
- Liu Y, Shi G. Roles of sirtuins in asthma. Respir Res 2022;23. [CrossRef]
- Qin X, Jiang B, Zhang Y. 4E-BP1, a multifactor regulated multifunctional protein. Cell Cycle 2016 15:781-786. [CrossRef]
- William M, Leroux L-P, Chaparro V, Lorent J, Graber TE, M’Boutchou M-N, et al. eIF4E-Binding Proteins 1 and 2 Limit Macrophage Anti-Inflammatory Responses through Translational Repression of IL-10 and Cyclooxygenase-2. J Immunol 2018, 200:4102-4116. [CrossRef]
- Isvoran A, Peng Y, Ceauranu S, Schmidt L, Nicot AB, Miteva MA. Pharmacogenetics of human sulfotransferases and impact of amino acid exchange on Phase II drug metabolism. Drug Discov Today 2022, 27. [CrossRef]
- Sharma J, Menon BK, Vijayan VK, Bansal SK. Changes in Adenosine Metabolism in Asthma. A Study on Adenosine, 5’-NT, Adenosine Deaminase and Its Isoenzyme Levels in Serum, Lymphocytes and Erythrocytes. Open J Respir Dis 2015, 05:33-49. [CrossRef]
Figure 1.
Differential expression profiles of inflammation-related biomarkers between allergic asthma and non-allergic asthma group. (A) Unsupervised clustering of 92 differentially expressed proteins (DEPs); (B) Group-specific expression patterns with DEPs; (C) Volcanic visualization of 92 inflammation-related biomarkers, Red, significantly upregulated proteins in asthma group; gray, no differences; blue, significantly downregulated proteins in asthma group.
Figure 1.
Differential expression profiles of inflammation-related biomarkers between allergic asthma and non-allergic asthma group. (A) Unsupervised clustering of 92 differentially expressed proteins (DEPs); (B) Group-specific expression patterns with DEPs; (C) Volcanic visualization of 92 inflammation-related biomarkers, Red, significantly upregulated proteins in asthma group; gray, no differences; blue, significantly downregulated proteins in asthma group.
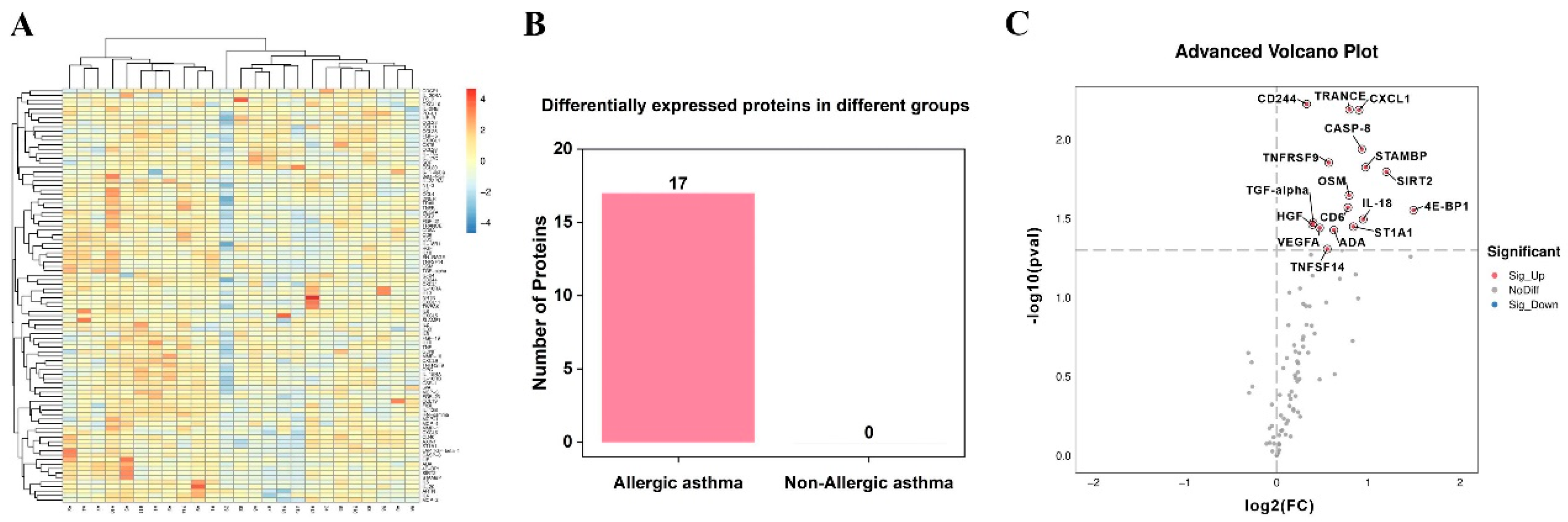
Figure 2.
Comparative analysis of DEPs between allergic asthma and non-allergic asthma group, *p < 0.05, **p < 0.01.
Figure 2.
Comparative analysis of DEPs between allergic asthma and non-allergic asthma group, *p < 0.05, **p < 0.01.
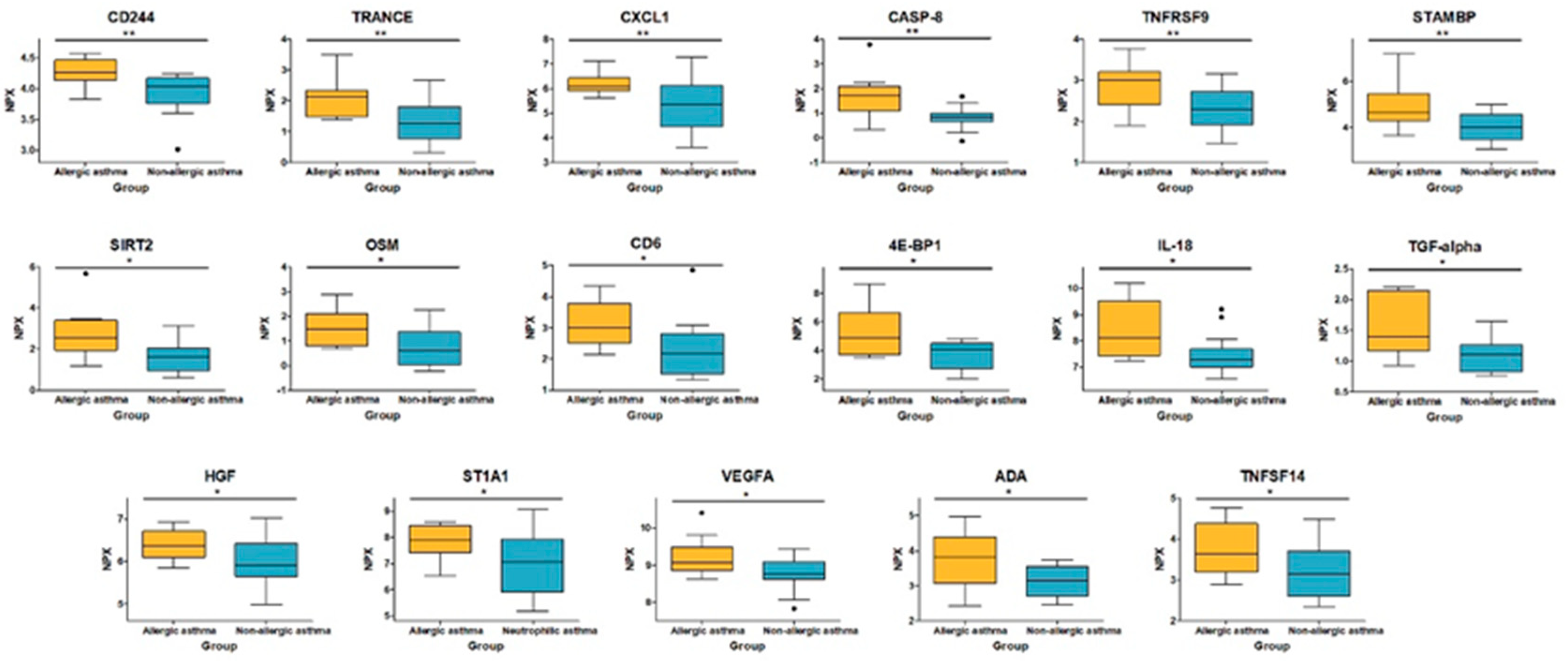
Figure 3.
Differentially expressed proteins profiling. (A) Correlation heatmap between the DEPs. Red, positively related; blue, negatively related; and white, nonrelated; (B) The scatter plots of STAMBP and CASP-8; (C) The scatter plots of STAMBP and SIRT2 (the most significant positive correlation), R: Pearson correlation coefficient; (D) PPI network of DEPs; (E) The ROC curves of individual data set of top 5 proteins; (F) The ROC curve of data combination of top 5 proteins; (G) Clustering heatmap of allergic asthma.
Figure 3.
Differentially expressed proteins profiling. (A) Correlation heatmap between the DEPs. Red, positively related; blue, negatively related; and white, nonrelated; (B) The scatter plots of STAMBP and CASP-8; (C) The scatter plots of STAMBP and SIRT2 (the most significant positive correlation), R: Pearson correlation coefficient; (D) PPI network of DEPs; (E) The ROC curves of individual data set of top 5 proteins; (F) The ROC curve of data combination of top 5 proteins; (G) Clustering heatmap of allergic asthma.
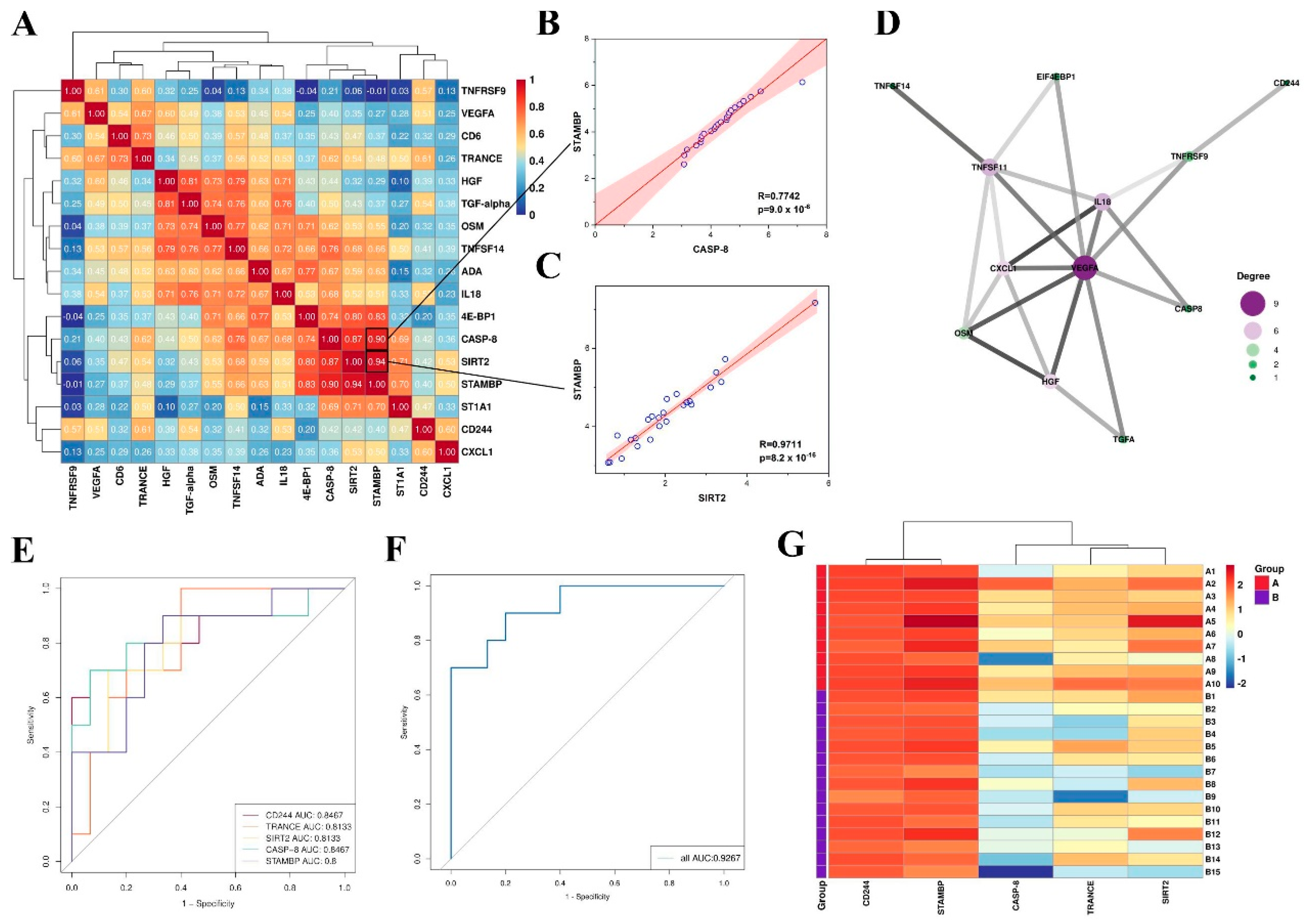
Figure 4.
Correlation heatmap between the DEPs and clinical features, Red, positively related; blue, negatively related; and yellow, nonrelated. *p < 0.05, **p < 0.01.
Figure 4.
Correlation heatmap between the DEPs and clinical features, Red, positively related; blue, negatively related; and yellow, nonrelated. *p < 0.05, **p < 0.01.
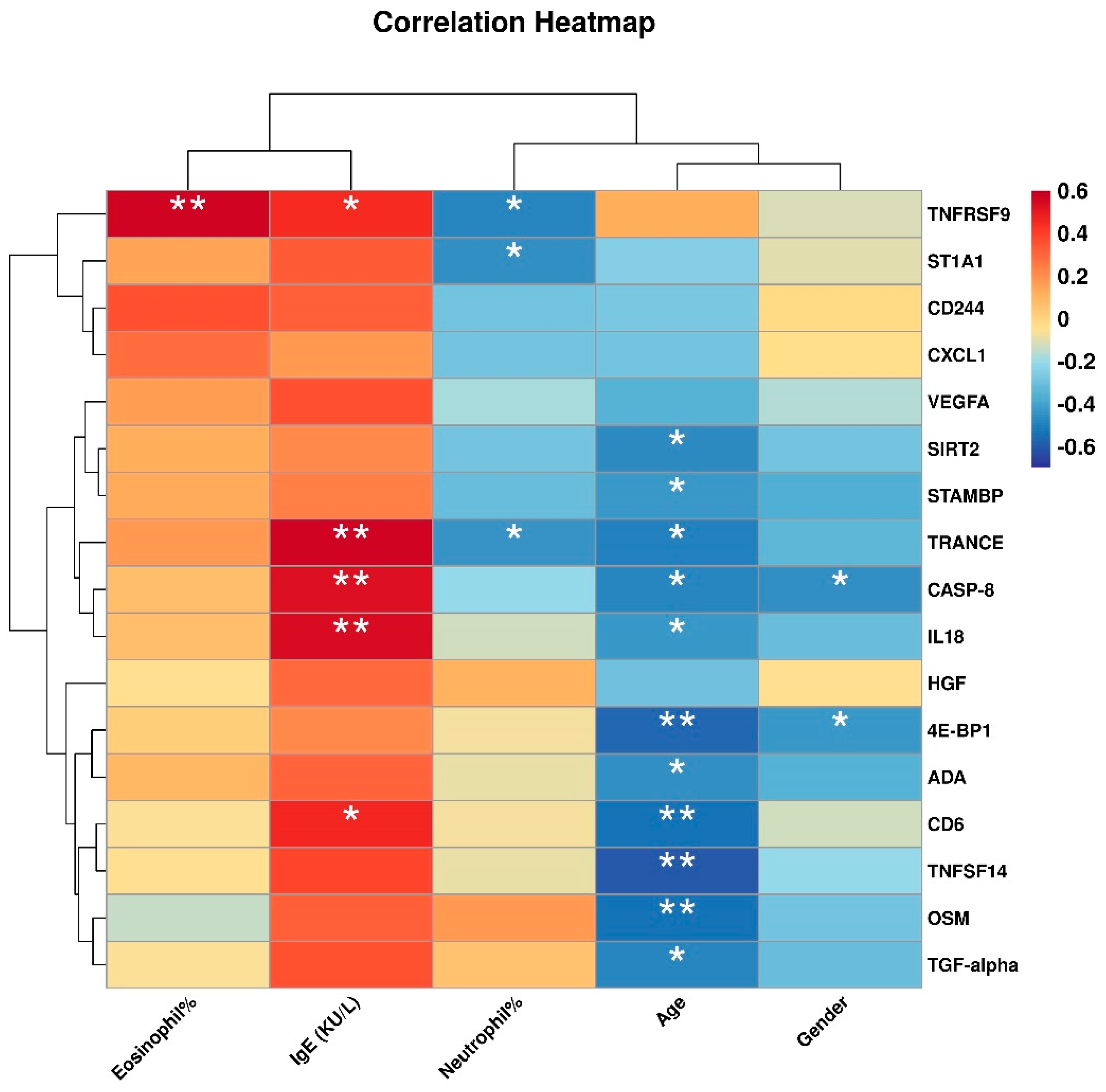
Figure 5.
Functional analysis of DEPs between the allergic asthma and non-allergic asthma groups. GO enrichment analysis of DEPs: (A) scatter plot, (B) Bar plot and (C) circos plot; KEGG enrichment analysis of DEPs: (D) scatter plot, (E) Bar plot and (F) circos plot.
Figure 5.
Functional analysis of DEPs between the allergic asthma and non-allergic asthma groups. GO enrichment analysis of DEPs: (A) scatter plot, (B) Bar plot and (C) circos plot; KEGG enrichment analysis of DEPs: (D) scatter plot, (E) Bar plot and (F) circos plot.
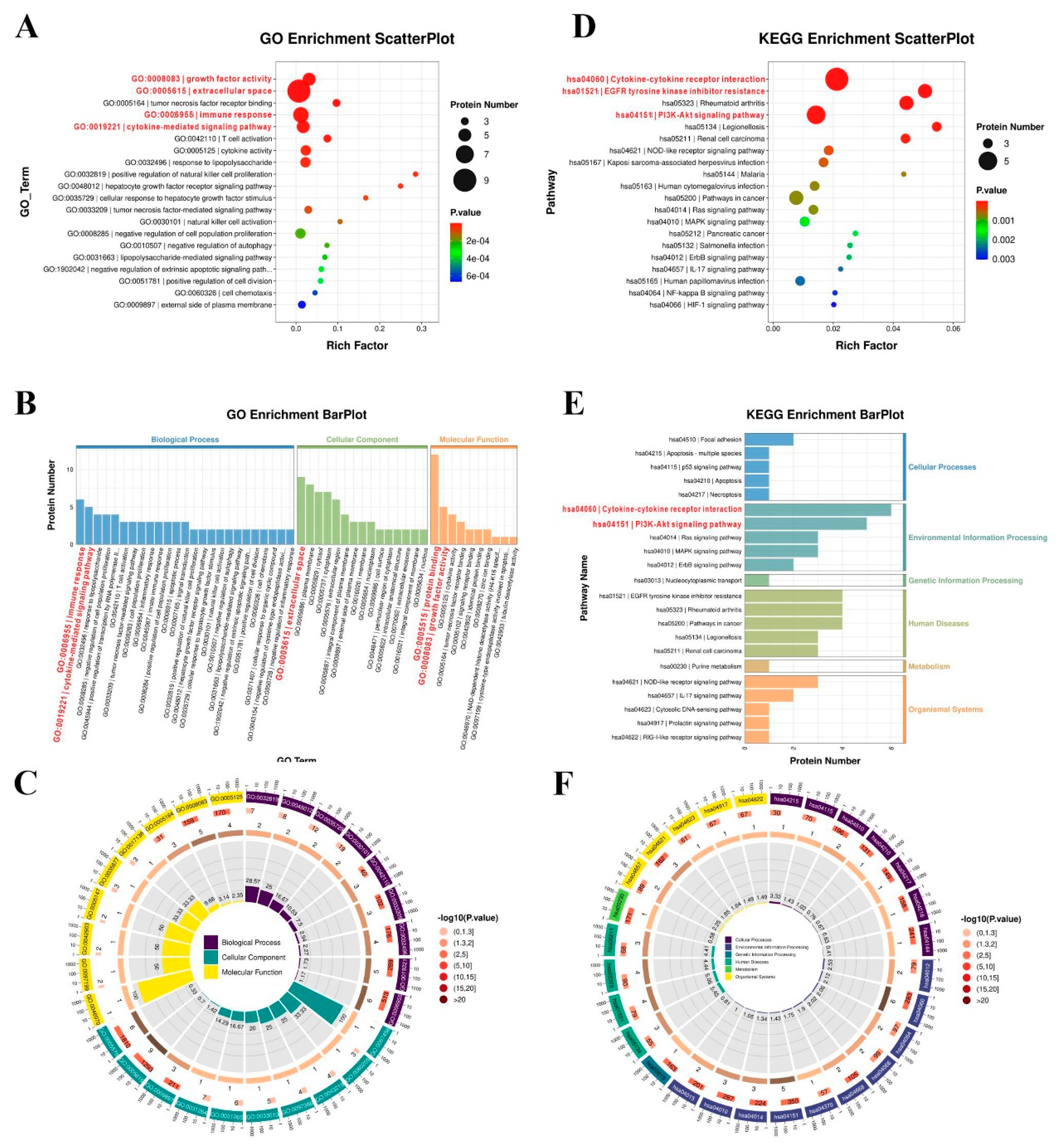
Figure 6.
The potential mechanism linking DEPs in the pathogenesis of allergic asthma.
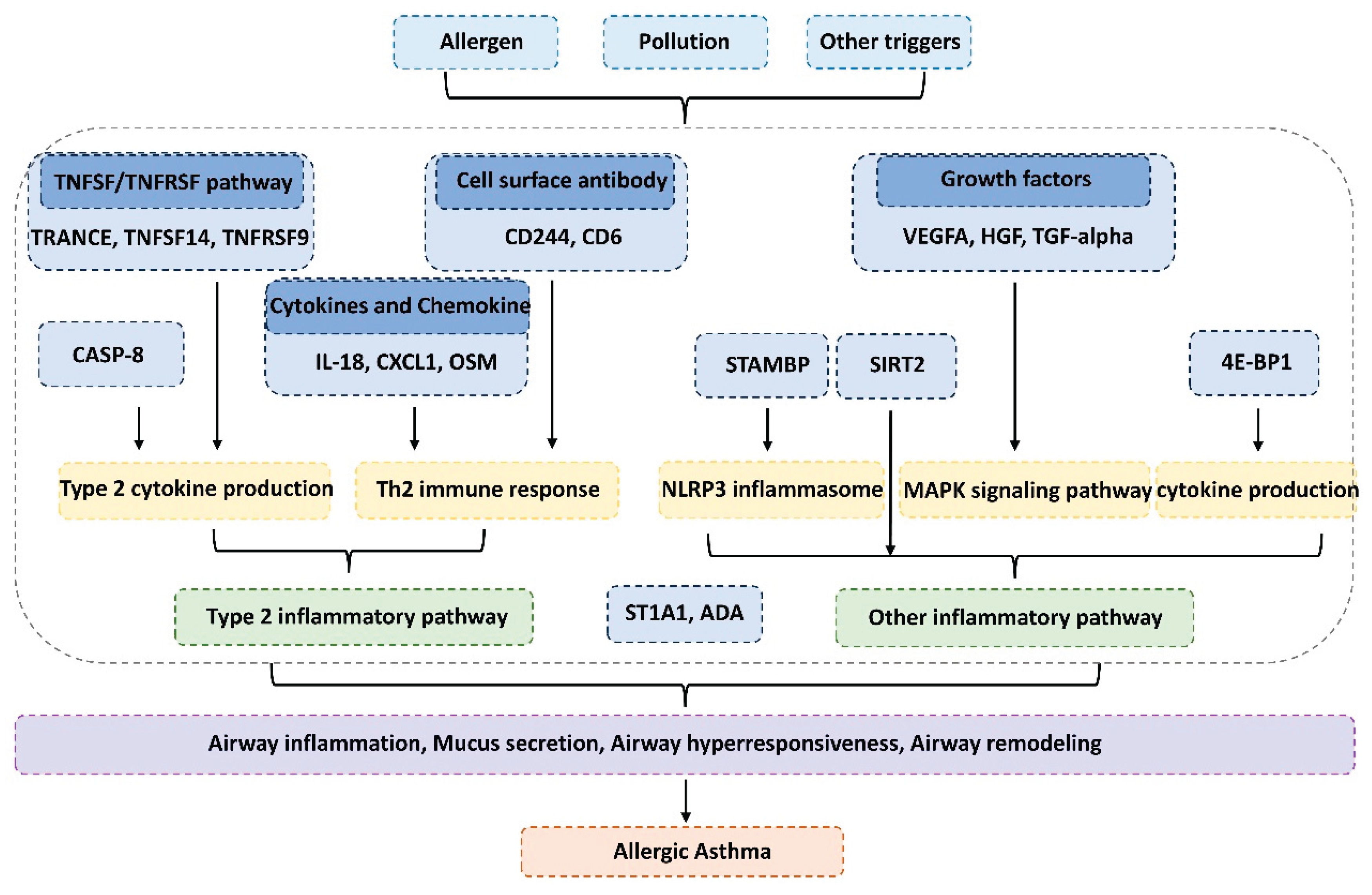

Table 1.
Clinical Characteristics of the Participants.
| Characteristics | Allergic asthma (n=10) | Non-allergic asthma (n=15) |
|---|---|---|
| Gender (Male/Female) | 7/3 | 3/12 |
| Female (%) | 30% | 80% |
| Age (mean ± SD) | 53.0 ± 20.6 | 63.7 ± 13.6 |
| Neutrophil (%) (mean ± SD) | 60.2 ± 10.1 | 74.8 ± 12.5 |
| Eosinophil (%) (mean ± SD) | 7.61 ± 8.87 | 1.52 ± 1.71 |
| Allergen-specific IgE (KU/L) (mean ± SD) | 657.8 ± 474.4 | 191.8 ± 164.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



