
Submitted:
22 May 2025
Posted:
22 May 2025
You are already at the latest version
Abstract
Immunoglobulin Y (IgY), the major antibody class in birds, has gained increasing attention in recent years as a versatile and ethically sustainable alternative to mammalian immunoglobulins. IgY has demonstrated strong potential in diagnostics, prophylaxis, and therapy across a wide range of fields, including infectious diseases, allergy management, oral health, and food safety. Its applications in animal health—particularly in poultry, livestock, and companion animals—further underscore its relevance within the One Health framework. This review provides a comprehensive synthesis of IgY technology, starting with its physiological role in maternal immunity and the structural characteristics that distinguish it from mammalian immunoglobulin G (IgG). Current production and purification strategies are outlined, followed by an overview of its biomedical and veterinary applications mainly based on studies published over the past five years. The final section addresses the current limitations of IgY technology, such as variability in protocols, stability challenges, and the need for safety assessment, while highlighting the importance of harmonized guidelines to support broader implementation. With growing scientific interest, expanding clinical research, and increasing availability of commercial products, IgY is well-positioned to become a valuable immunobiological tool for both human and veterinary applications.
Keywords:
immunoglobulin Y
; hyperimmune egg
; One Health
; antimicrobial resistance
1. Introduction
Maternal immunity refers to the transfer of protective antibodies from the mother to her offspring, providing temporary immune defense during the early stages of life—when the immune system of the neonate is not yet fully developed [1]. In mammals, this transfer occurs either transplacentally before birth or through the intestinal absorption of immunoglobulin G from colostrum within the first 24 hours after birth [2]. In birds, however, the process involves immunoglobulin Y (IgY), the predominant antibody class, which also plays a central role in the passive transfer of immunity. This transfer occurs via the egg yolk through a specialized, two-step mechanism: first, IgY is transported from the maternal bloodstream into the yolk; then, during embryonic development, it is absorbed from the yolk into the embryo’s circulation via the yolk sac membrane [3].
In recent years, IgY has emerged as a promising tool in biomedical research and applications, due to its phylogenetic distance from mammals, which enhances the immune response to conserved mammalian proteins. Moreover, hens can be immunized with specific antigens to generate large quantities of high-affinity antibodies, which are then non-invasively harvested from egg yolks. This production system supports the ethical principles of the 3Rs (Replacement, Reduction, Refinement), while also offering cost-efficiency and scalability. The resulting antibodies demonstrate remarkable thermal and pH stability, which enhances their usability in diverse formulations for oral, topical, or intranasal delivery.
In addition to its diagnostic potential, IgY has shown efficacy in treating or preventing infectious diseases in both human and veterinary contexts. It has been successfully applied against pathogens such as Helicobacter pylori, Pseudomonas aeruginosa, Clostridioides difficile, and SARS-CoV-2, with minimal side effects and no reported risk of inducing resistance. The versatility of IgY positions it as an attractive alternative to antibiotics, particularly in the context of growing antimicrobial resistance.
This article presents a detailed analysis of IgY, beginning with its physiological and structural characteristics, followed by a brief overview of current production technologies. It then reviews the latest biomedical applications of IgY, focusing on findings from recent studies, including clinical trials. The synthesis concludes by discussing the current advantages, limitations, and practical considerations of IgY technology, emphasizing its potential for broader implementation in both research and applied immunotherapy.
2. Structure and Function of IgY
Immunoglobulin Y is a class of antibodies found in birds, reptiles, amphibians and lungfish [4]. IgY is similar to IgG found in mammals and it was called IgG until it was discovered that distinct differences exist between the two, prompting to switch to the term ‘IgY’ [5,6].
The structure of immunoglobulin Y is comparable to mammalian IgG, with their basic unit structure consisting of two light and two heavy polypeptide chains that form a monomeric unit (Figure 1). The two heavy chains are connected to each other via disulfide bonds, while each heavy chain is attached to a light chain through a disulfide bond as well. In any immunoglobulin molecule, the two heavy chains and two light chains are identical, resulting in an antibody with two identical antigen-binding sites. This symmetry enables the antibody to bind simultaneously to two identical structures. [7]. However, while IgG has three constant domains per each heavy chain, IgY is comprised of four constant domains per heavy chain, an aspect that accounts for the higher molecular weight of IgY (180kDa for IgY compared to 150kDa for IgG) [8,9]. The extra domain of the heavy chain of IgY can also play a significant role in providing more molecular stability as compared to IgG [10].
Another important structural difference of IgY compared to IgG is the absence of a genetically encoded hinge, which results in limited flexibility. This has implications in its biological properties, such as its inability to precipitate antigens at physiological salt concentrations [11], a phenomenon observed in both chickens [12] and ducks [13]. One notable advantage of the higher chain stiffness of IgY is its potential association with increased resistance to proteolytic degradation and fragmentation, as reported in previous studies [14].
The Y-shaped structure comprises two main regions; the fragment-crystallizable (Fc) and the fragment-antigen-binding (Fab) region [15]. Similar to IgG, the Fc region of IgY mediates most of its biological effector functions, including complement fixation and opsonization [16]. Cell surface Fc receptors interact with the constant (Fc) region of the immunoglobulin, contributing significantly to the regulation and modulation of the immune response [17]. The Fab (fragment antigen-binding) region of an immunoglobulin is primarily responsible for recognizing and binding to specific antigens [18].
Fc portion is unable to activate human complement [19] which is crucial for ensuring analytical accuracy in immunological assays. This property prevents interference caused by varying complement activity in patient samples, reducing analytical errors and enhancing assay reliability [16]. Immunoglobulin Y does not bind to the rheumatoid factor, nor does it activate human anti-mouse antibodies [20], furthermore proving its potential to reduce interference problems in immunological assays [16]. In addition, chickens generate a stronger immune response against conserved mammalian proteins due to their significant phylogenetic distance from mammals [21]. This makes chicken antibodies particularly effective for targeting highly conserved epitopes.
In humans and mice, immunoglobulin (Ig) genes are formed through a random assortment of variable (V), diversity (D), and joining (J) segments, generating a diverse repertoire of antibodies [22]. After antigen stimulation, this diversity is further expanded by somatic hypermutation, and the Ig isotype can be altered through class switch recombination. In chickens, however, the contribution of V(D)J recombination to immunogenetic diversity is minimal, as only a single locus undergoes rearrangement to produce a functional gene [4]. Instead, chickens rely on gene conversion to achieve antibody diversity. During this process, short sequences in the V-region gene are replaced with homologous sequences donated by pseudogenes, significantly enhancing the diversity of IgY and compensating for the limited role of V(D)J rearrangement [17].
IgY exhibits remarkable stability across a range of conditions: it remains stable at temperatures between 4°C and 60°C and within a pH range of 4.0 to 10.0. Additionally, IgY maintained its stability for 35 days at 4°C, 25°C, and 37°C [23]. Also, eggs stored for up to one month at room temperature, or for up to 6 months at +4°C showed no significant reduction of antibody titers [24]. The stability of IgY under heat stress (75–80°C), acidic conditions (Ph 3) or high pressure (5000 kg/cm2) was found to be increased by the addition of high concentrations of sucrose [25].
3. Immunoglobulin Y Production and Extraction
Understanding the production and extraction process of IgY is essential for exploiting its full potential in biomedical applications. The technology bridges immunology, biotechnology, and translational research, and involves a series of coordinated steps, from antigen selection and hen immunization to antibody extraction and purification. Each stage can influence the quality, specificity, and usability of the final commercial product. As IgY continues to gain recognition as a noteworthy tool for prophylactic, therapeutic, and diagnostic purposes, it becomes increasingly important to ensure not only efficient production but also rigorous testing. Evaluating its interactions with target antigens and the host is critical to confirming safety and efficacy for downstream applications (Figure 2).
3.1. Immunization Protocol
Optimizing the immunization process remains a critical challenge regarding IgY technology. Key challenges include selecting the appropriate antigen type, choosing the best adjuvant, and designing an effective immunization protocol to maximize IgY yield and specificity. These aspects came into increased attention during 1990, leading to the course of workshops organized by the European Centre for the Validation of Alternative Methods (ECVAM) and the subsequent report [6] (Table 1). Since then, various protocols have been published and utilized in IgY studies [9,26], emphasizing the need for close monitoring of antibody titers throughout the immunization period to enable adjustments to the protocol based on the observed immune response [17].
The first step in the obtaining of IgY is the choice of an antigen to be used for immunizing hens. Selecting an antigen of the right type and of high quality is paramount for inducing a strong humoral response and, subsequently, the production of IgY antibodies. Immunogenicity, which refers to the ability to stimulate cellular and humoral immune responses, and antigenicity, the capacity to be specifically recognized by the antibodies produced during the immune response, are two essential antigen characteristics for generating high-performance antibodies [17,27].
Various antigen types can be used to produce specific IgY in birds: complex antigens from whole viruses, bacteria, parasites or fungi [28,29,30], recombinant proteins [31,32,33,34] or other antigens with immunogenic properties, such as toxoids, venom, allergenic antigens and even other antibodies [35,36,37,38]. More recent technologies include the use of haptens, small molecules that are converted to antigens by linking to a carrier protein, virus-like particles, bacterial ghosts, virosomes or nucleic acid vaccines [17,26].
Particularly when working with pathogenic or toxic antigens, a crucial step is antigen inactivation. Various methods can be employed, with the overall aim of preserving immunogenicity and antigenicity while at the same time protecting animal welfare and ensuring the safety of the laboratory team [17]. Various methods can be employed to inactivate antigens, such as chemical inactivation using formaldehyde [39,40,41], physical inactivation through heat [42] or ultrasonication [43] or by combining different types of methods [44].
The antigen dose is another key aspect for IgY production, as insufficient doses may fail to elicit an adequate immune response, while excessively high doses can overstimulate or overwhelm the immune system, potentially leading to immune tolerance or reduced antibody production. Early studies showed that 0.1 or 0.5 mg of mouse-IgG elicit a maximum response regarding the antibody titre, while increasing to 1 mg can lead to a reduction in the immune response [26]. However, the majority of studies use between 50 and 200 µg of antigen per inoculation [31,32,33,45]. A more recent study further supports the importance of antigen dose in IgY production, demonstrating that while doses as low as 2 µg can elicit significant antibody levels after multiple immunizations, the highest antibody titers were observed with doses of 20 and 200 µg [46].
Other important factors regarding the production of antibodies include the genetic background [47], nutrition [48], age [49], keeping conditions [50] or biorhythm [51].
Choosing the suitable adjuvant also plays a significant role regarding the outcome of the immunization, since the inactivated antigens used in the inoculum are usually less immunogenic compared to the modified live microbes [52]. Adjuvants enhance IgY production by improving antigen retention, stimulating immune responses, and increasing specific antibody titers. To produce IgY antibodies in birds, the most frequently used adjuvants are complete and incomplete Freund's adjuvants (FCA and FIA) [53]. FCA, which is a mixture of a mineral oil and heat-killed and dried mycobacteria, has been the gold standard for generating high levels of antibodies in animals [54], however its main disadvantage is the potential to induce local and systemic side-effects [26]. FIA does not contain mycobacteria, which usually is responsible for the adverse reactions seen with FCA [54], therefore numerous studies now use FCA for the first immunization and FIA for the boosters injections [40,55,56]. Other commercial adjuvants can be used, such as Montanide or Polygen or aluminium hydroxide [33,46,57,58], while there is still research being done in order to obtain a better, safer adjuvant. [59,60].
Usually, antigen administration for IgY production in chicken is done through the intramuscular route, most common into the breast muscle, as administration into the thigh muscle may lead to lameness [4]. The second most common route is the subcutaneous route [32,57,61], which may be associated with higher yields of antibodies [26] but may also be less suited for young animals [9]. The intravenous route should only be used without adjuvants and performed very slowly in order to avoid an anaphylactic reaction [17], while the oral route has been successfully used [62] and may be useful for large scale immunizations.
Immunization should be performed when the animals are of egg-laying age, with the overall aim being to make the peak of laying and the peak of antibody production to occur at the same time [63]. Although the total IgY content of an egg has been reported to increase with age from 32 to 52 week-old breeders [64], a recent study showed that the content of IgY from spent laying hens (age 83 weeks) was lower than that isolated from new laying hens (age 27 weeks) [49]. The interval between multiple immunization varies greatly between different studies, with early studies considering a primary vaccination and a booster given before the laying period, at 4-6 weeks interval [6], or every 3-5 weeks [65]. The usual interval is now at least 4 weeks, to allow for the generating of immune memory [17], while a general rule is to administer a booster immunization whenever the titer of IgY reaches a plateau or begins to decrease [63]. Other studies reported administering the booster injection at 7 days [61], 8 days [56], 10 days [41] or 14 days [39,40,66]. After the first immunization, IgY begins to appear in the serum in approximately 4 days, while a delay of 3-4 days was observed between the occurrence of IgY in the serum and its occurrence into the egg yolk. [17]. High titers of IgY can be kept for more than 150 days through booster immunizations [63].
3.2. Extraction and Purification of IgY
To get a better understanding of the technology of IgY production, it is important to discuss the structure of egg yolk. The egg yolk contains numerous nutrients and preservative substances, highlighting its role as an embryonic chamber [67] The composition of yolk is approximately 48% water, 32% lipid and 16% proteins [68] and its main components include phosvitin, low-density lipoprotein (LDL), high-density lipoprotein (HDL), and lecithin [69].
From a structural point of view, the egg yolk has two parts: an aqueous plasma consisting of 86% low density lipoproteins and the remainder α-livetin (serum albumin), β-livetin (α2glycoprotein), and γ-livetin (IgY), and granules of different sizes composed of 70% high-density lipoproteins (α- and β-lipovitellins), 16% phosvitin (glycophospoprotein), and 12% low-density lipoproteins (LDL) [17]. Since IgY is part of the aqueous plasma, the first step of IgY extraction consists of removing lipids to obtain the water-soluble components (water-soluble filtrate – WSF), while the second step involves the precipitation of antibodies present in the WSF by different techniques [9].
Regardless of the method employed, it is important that the egg is washed with clean water and cleaned with 75% ethanol to ensure that contaminants don’t affect the IgY extract. After cracking the eggshell, the removal of egg white is important, which can be easily done by using an egg separator or by rinsing the yolk in flowing water or rolling it on filter paper [17].
By de-lipidation, the egg yolk granules, containing lipids, precipitate and separate from the WSF that contains IgY together with other proteins. The usual de-lipidation technique is the water dilution method, which results in the aggregation of yolk lipids at low ionic strength [70]. The degree of dilution varies among different authors and ranges between 6 and 10-fold dilution [9], while variations of the technique include ph adjustment of water around 5-5.2 [71] or adding a freeze-thaw cycle for the diluted egg yolk [72]. At the end of the process, the mixture is centrifugated and the WSF is collected in the supernatant. The water dilution method has the advantage that no toxic products are used, making it suitable for the purification of IgY for oral administration [17] Other methods of de-lipidation include the use of Polyethylene Glycol [73], Anionic Polysaccharides such as pectin or λ-carrageenan [74,75,76], organic solvents such as chloroform [77] or specific chemicals such as caprylic acid [78].
After de-lipidation, the concentrated fraction obtained also contains other water-soluble proteins as well as some minor lipids or lipoproteins [70]. Four main types of methods can be used to purify the contaminants and concentrate the IgY from the delipidated plasma: precipitation, chromatography, filtration, and aqueous biphasic systems [17].
Salt precipitation is commonly employed in protein purification, the main mechanism being that most proteins require at least a small concentration of salt to remain stable. At very low salt concentrations, proteins with charged regions tend to aggregate. Salt ions help neutralize surface charges, preventing aggregation, while at high concentrations, excessive charge at the surface of proteins can induce aggregation again. [79]. Different saturated salt solutions can be used, such as ammonium sulphate [30], sodium sulphate [80], polyethylene glycol [81] or sodium chloride [33].
Chromatographic methods are generally employed in industrial settings to obtain samples of IgY with the highest purity, due to their high resolution, selectivity and efficiency [17]. Different methods can be used, such as affinity chromatography [36], ion exchange chromatography [82] or hydrophobic interaction chromatography [83,84]. Regarding affinity chromatography, the conventional method using protein A or protein G is ineffective for IgY purification, as IgY, unlike IgG, does not bind to these proteins [85], therefore other ligands have been developed, such as an elastin-like polypeptide-tagged immunoglobulin-binding domain of streptococcal protein G [86]. Although chromatographic methods have proven advantages as mentioned beforehand, they are generally expensive and therefore impractical for large-scale use [9].
Filtration methods, such as funnel filtration, column-filtration or ultrafiltration, offer another possibility for IgY purification, with the main principle being that the solution is passed through a barrier that retains some species based on their size and shape while allowing other species to pass through [17]. Column and ultrafiltration are feasible for large scale use and industrial applications [70].
Aqueous biphasic systems represent a more recent alternative to conventional IgY purification methods, research showing that these systems maintain the structural integrity of IgY, offer satisfactory levels of purity and represent a cost-effective, simple and scalable technique for IgY purification [87].
At the same time, numerous studies employ a combination of different techniques in their IgY extraction protocol, usually salt precipitation by polyethylene glycol or ammonium sulphate followed by chromatographical methods [31,36,88] or ultrafiltration [41], or a combination of all three types of methods [56].
3.3. Evaluation of IgY
Antibodies can be characterized by various methods to determine their purity, quantity, and biological activity. These parameters are evaluated independently, and confirmation through a single analytical method is usually sufficient for polyclonal antibodies. There is a close relationship between purity and quantity, as each additional step in the extraction methodology leads to a loss of protein content [17].
The determination of protein concentration in IgY extracts is typically performed prior to detailed antibody evaluation. The total protein concentration in IgY extracts is primarily determined using the Bradford method [36,45,89], the Lowry method [81] or the bicinchoninic acid (BCA) assay [66]. In addition, protein concentration can be assessed by ultraviolet absorption at 280 nm, according to the Lambert-Beer law [49,90], although accurate determination requires establishing the correct extinction coefficient (molar absorptivity), which may be 1.33 [91,92], 1.25 [93] or 1.094 [94], depending on the solvent used for the measurement [17].
Since IgY has a known molecular weight of 180 kDa, its purity can be evaluated by SDS-PAGE (sodium dodecyl sulfate polyacrylamide gel electrophoresis), considered the gold standard technique and widely used to assess the purity of IgY isolated from egg yolk [9]. Separation by SDS-PAGE under standard conditions will reveal a single protein band around the 180 kDa marker, while the addition of a reducing agent will result in two protein bands corresponding to the heavy and light chains of IgY [17]. Western blotting is a method commonly used in conjunction with SDS-PAGE to confirm the presence and evaluate the purity and specificity of IgY isolated from immunized egg yolks [41,61,95], while high-performance liquid chromatography (HPLC) can be employed as a complementary technique to further assess purity and identify protein composition with high resolution [59].
3.4. Stability and Routes of Administration for IgY Products
The route of administration is a critical factor in determining the efficacy, safety, and practicality of immunoglobulin Y (IgY)–based interventions, both for therapeutic and for prophylactic reasons. Due to their structural and functional differences from mammalian antibodies, IgYs exhibit unique pharmacokinetic and pharmacodynamic profiles. Understanding how these antibodies behave in vivo—particularly at mucosal surfaces—is essential for the development of targeted, non-invasive therapies that minimize systemic side effects.
IgY antibodies do not bind to human Fc receptors and do not activate the mammalian complement system [96]. Additionally, due to their large molecular size, systemic penetration is limited [97]. A human study involving intranasal administration of IgY demonstrated no evidence of a systemic inflammatory immune response following topical treatment [98], supporting the notion that IgY is among the safest therapeutic agents available [97].
Another factor limiting the intravenous use of IgY is the lack of interaction with Fc receptors, which are typically required for active immunoglobulin transport across biological membranes. This absence of Fc binding helps explain the low alveolar concentrations observed after IV administration of IgY in a porcine pneumonia model [99].
IgY exhibits hydrophobicity and notable physicochemical stability. It maintains biological activity for up to six months at ambient temperature and can be stored for over five years at 4 °C without significant degradation. Functionally, IgY remains active within a broad pH range (pH 3.5–11) and demonstrates thermal stability up to 70 °C for 15 minutes. IgY is quite resistant against trypsin and chymotrypsin inactivation, but degraded by pepsin, the stability against the latest being highly dependent on pH and the enzyme/substrate ratio [100]. However, limited data are available regarding its in vivo half-life. In human studies, active IgY has been detected in saliva up to 12 hours following oral administration via gargling[101], in neonatal piglets, the reported serum half-life of IgY is approximately 1.85 days, while in the gastrointestinal tract it is significantly shorter, averaging 1.73 hours [102].
Despite these limitations, IgY antibodies can be effectively utilized for mucosal passive immunization, offering local protection at respiratory, oropharyngeal, and gastrointestinal surfaces [97]. In a clinical trial, oral administration of anti-Helicobacter pylori IgY three times daily for two weeks led to significant improvement in clinical symptoms and quality of life in H. pylori-infected patients, while also increasing the bacterial eradication rate by 30.6% [103]. Similarly, intranasal administration of anti-SARS-CoV-2 IgY three times daily for 14 days demonstrated an excellent safety profile [98]. Oral delivery via IgY-based sprays has also been proposed to carry minimal systemic risk and a lower likelihood of resistance development [104].
Recent in vivo studies have further explored intranasal administration of IgY in various animal models, including mice [105,106,107] and hamsters [108,109]. Oral administration has also been studied in veterinary settings, such as in calves [29,110], broiler chickens [111,112,113], and domestic cats. In the latter, a six-month safety evaluation of cat food supplemented with anti-Fel d 1 IgY reported no increase in mutagenicity or chromosomal aberrations [114].
4. Biomedical Applications of IgY
The unique immunological properties of immunoglobulin Y (IgY)—including high antigen specificity, lack of interaction with mammalian Fc receptors, and minimal background reactivity—make it a valuable tool for diagnostic applications. As outlined in a recent review, IgY can be developed and evaluated through established in vitro methods for the detection of a wide range of biomolecules [9]. Recent studies have demonstrated its effectiveness across various diagnostic platforms, addressing both infectious and non-infectious disease targets.
4.1. Avian Antibodies in Diagnostic
Although the primary focus of the reviewed studies is therapeutic and prophylactic, diagnostic applications of IgY are emerging, particularly in research and tool development. Anti-SpCas9 IgY antibodies have been developed for the immunodetection of CRISPR-Cas components in biological samples, demonstrating high sensitivity and specificity in Leishmania braziliensis promastigotes expressing SpCas9 [45]. Such tools are essential in molecular biology for tracking gene editing processes and validating CRISPR efficacy.
Immunoglobulin Y has also been employed to target bacterial pathogens. Anti-SpA IgY antibodies against Staphylococcus aureus demonstrated high specificity without cross-reactivity, reliable performance across various sample types, and advantages over IgG-based assays in terms of reduced false positives. Moreover, the same antibodies exhibited bacteriostatic activity and reduced biofilm formation, highlighting both diagnostic and neutralizing functions [115]. Similarly, IgY antibodies targeting the LTB subunit of the heat-labile enterotoxin produced by enterotoxigenic Escherichia coli (ETEC) were used to develop a sandwich ELISA, achieving an analytical sensitivity of 39 ng, consistent with previously reported detection ranges [116]. IgY-based assays have also been developed for the rapid detection and discrimination of toxigenic Vibrio cholerae, targeting outer membrane protein W (OmpW) and cytotoxin B (CtxB) antigens [117].
In oncology, anti-human B7-H4 IgY antibodies were successfully developed and used to establish a double-antibody sandwich ELISA capable of detecting soluble B7-H4 in human serum with higher sensitivity than a commercial kit [95]. In the context of viral infections, multiple studies have confirmed the potential of IgY-based platforms for the detection of SARS-CoV-2. These include an electrochemical immunosensor utilizing anti-spike IgY with a detection limit of 5.65 pg/mL [88], and an IgY-scFv construct with high affinity for the S1 subunit of the spike protein, capable of recognizing multiple epitopes and showing promise for both diagnostic and therapeutic applications [118]. Additionally, neutralizing IgY antibodies against human papillomavirus (HPV) have been characterized using pseudovirus assays, suggesting their potential utility in neutralization-based diagnostic platforms to assess viral infectivity and vaccine responses [119].
For parasitic infections, IgY antibodies have shown strong diagnostic potential. Anti-Ascaris suum IgY achieved high avidity and recognized multiple antigenic bands, with immunofluorescence and immune complex detection methods yielding sensitivity and specificity rates of 80% and 90%, respectively [120]. Similarly, IgY antibodies against Toxoplasma gondii were successfully conjugated with latex particles for the development of a particle-based detection platform, with potential applications across health, food, and biotechnology sectors [121,122]. A coproantigen sandwich ELISA test was also developed using recombinant Opisthorchis viverrini cathepsin F (rOv-CF) for the diagnostic of O. viverrini infections, the conventional stool examination making the diagnostic difficult given the low intensity of the infection and intestinal coinfections, in the context of fatal cholangiocarcinoma caused by chronic and repeated infections with liver flukes [123].
Beyond human medicine, IgY-based diagnostic tools have shown promising results in veterinary and animal health applications. In canine diagnostics, anti-DEA1.1 IgY antibodies were successfully produced and purified from egg yolk for use in blood typing. The resulting test is simpler to standardize than traditional canine blood group assays, and may facilitate routine screening in clinical practice [124]. In poultry, a lateral flow assay employing anti-IBDV IgY enabled rapid detection of infectious bursal disease virus (IBDV), demonstrating strong specificity by avoiding cross-reactivity with avian influenza and Newcastle disease viruses [125].
In aquaculture, IgY has been utilized for the diagnosis of hemorrhagic septicemia caused by Aeromonas hydrophila in Piaractus mesopotamicus. Immunohistochemical analysis confirmed the presence of A. hydrophila within host phagocytes and surrounding tissues, supporting the applicability of IgY in aquatic diagnostics [126]. Additionally, anti-vitellogenin IgY was developed for detecting reproductive biomarkers in Cichlasoma festae, with ELISA assays successfully quantifying vitellogenin concentrations between 10–1280 ng/mL [56].
In the area of food safety, IgY antibodies have been applied for the detection of contaminants and allergens. An indirect competitive ELISA (ic-ELISA) using anti-zearalenone (ZEN) IgY was developed to detect ZEN in post-fermented tea, showing strong correlation with HPLC results and demonstrating potential for rapid screening of mycotoxins in the beverage industry [30]. Similarly, an IgY-based immunosensor targeting dust mite protein was successfully used for the detection of allergens in flour [127].
In research settings, IgY has been used to detect and isolate minor bioactive components. For example, affinity measurements between lactoferrin and anti-lactoferrin IgY revealed that IgY is well suited for immunochemical assays and chromatographic applications targeting low-abundance molecules [128]. Furthermore, conjugated anti-mouse IgG IgY has been proven as a reliable secondary antibody in ELISA, Western blot, and immunofluorescence assays. These conjugates displayed high thermal and pH stability, and performed comparably to conventional goat anti-mouse IgG, supporting their use in broad immunological workflows [129].
All these findings underscore the versatility of IgY in biomedical research and suggest a promising future for its diagnostic deployment, especially where mammalian antibodies might cause cross-reactivity or ethical constraints.
4.2. The Role of IgY in Prevention and Therapy
The use of IgY antibodies as a passive immunization strategy has shown promising results in combating viral respiratory infections, particularly due to SARS-CoV-2 and MERS-CoV. In vivo studies have demonstrated that intranasal administration of anti-SARS-CoV-2 IgY significantly reduces viral load, lung inflammation, and histopathological damage in hamster models [108,109], while also inhibiting spike-mediated cell–cell fusion and persisting in the nasal and oral cavities for several hours after administration [106]. Radiolabeled IgY exhibited preferential accumulation in the trachea and no adverse effects in mice and rats, supporting the safety of mucosal delivery [98,107]. Additionally, IgY antibodies produced against inactivated SARS-CoV-2 demonstrated potent neutralizing activity and suitability for large-scale production [106]. In the case of MERS-CoV, anti-S1 IgY significantly reduced viral antigen expression and lung inflammation in infected mice, further confirming its therapeutic potential against emerging coronaviruses [130].
IgY antibodies have also demonstrated prophylactic and therapeutic potential against a variety of other viral pathogens. In a murine model, intranasal administration of IgY specific to H5N1 influenza virus protected mice from weight loss, lung pathology, and clinical symptoms following infection, without impairing the development of adaptive immunity or immunological memory [105]. Against norovirus, anti-HuNoV IgY effectively disrupted virus binding and replication in a human intestinal enteroid model, showing promise for oral prophylaxis, particularly in populations lacking access to effective vaccines [66]. In the case of dengue virus, specific IgY antibodies neutralized viral infection in vitro, while one variant also conferred significant protection in mice against lethal challenge [83]. Rotavirus-specific IgY conferred partial protection in cynomolgus monkeys and effectively neutralized viral activity in cell cultures, reducing viral loads in vitro [131,132]. IgY antibodies raised against multiple strains of human papillomavirus (HPV) achieved neutralizing titers of up to 1:2000, suggesting their applicability as therapeutic agents, particularly post-infection [133]. Additionally, in a murine pseudovirus challenge model, high-dose IgY antibodies against Ebola virus provided post-exposure protection and displayed excellent thermostability—an advantage for deployment in resource-limited settings [134].
Regarding antibacterial activity, IgY antibodies targeting Escherichia coli and Helicobacter pylori have shown strong potential as alternative or adjunctive agents in both food safety and therapeutic applications. In the context of functional food development, anti-E. coli O157:H7 and anti-H. pylori IgY were successfully incorporated into fortified ice cream, with 0.6 mg/mL being sufficient for antimicrobial effectiveness, suggesting a novel delivery vehicle for gastrointestinal prophylaxis [89]. In a murine model, oral administration of anti-urease IgY significantly reduced gastric colonization, indicating its preventive efficacy (dose-dependent, up to 500 mg/kg) [135], while clinically, a two-week dietary intake of multivalent anti-H. pylori IgY improved symptoms, enhanced quality of life, and increased eradication rates by 30.6% [103]. In a murine model of acute graft-versus-host disease, dietary anti-E. coli IgY administration reduced intestinal inflammation and improved outcomes by lowering the expression of inflammatory markers and pathogen-associated receptors [42]. IgY targeting enterotoxigenic E. coli (ETEC) reduced bacterial adherence to epithelial cells by 74%, neutralized heat-labile toxin effects, and significantly reduced fluid accumulation in an ileal loop model, confirming both antitoxin and anti-adhesion properties [34]. Oral administration of anti-ETEC IgY in mice further ameliorated clinical signs, restored intestinal integrity, and suppressed proinflammatory cytokine production during enterotoxigenic E. coli infection [136]. Similar efficacy was observed for IgY targeting diarrheagenic E. coli, which inhibited bacterial growth and adhesion and significantly reduced colonization in an in vivo infection model [82]. In a murine model of urinary tract infection, IgY-rich egg-based formulations significantly reduced uropathogenic E. coli (UPEC) burden over a 15-day treatment period, supporting their potential use as adjuvant therapies [137].
The application of IgY antibodies in the treatment of Pseudomonas aeruginosa (PA) infections has yielded mixed results across various experimental models. Several studies have reported encouraging outcomes in murine models of pneumonia and burn wound infections, where passive immunization with polyclonal anti-PA IgY conferred broad-spectrum protection, likely due to its multivalency and strain-independent targeting capability [28]. Anti-flagellin IgY antibodies also showed dose-dependent, non-type-specific protection, achieving full protection in acute pneumonia models and partial efficacy in burn wound infections depending on the flagellin subtype [138]. Immunotherapy with anti-OprF IgY significantly improved survival in PA-infected burned mice and inhibited bacterial invasion in epithelial cell lines, supporting its role in passive protection [139]. Synergistic effects between IgY and antibiotics such as azithromycin, ceftazidime, imipenem, and meropenem were observed both in vitro and in vivo, suggesting that IgY can potentiate conventional antimicrobial therapies and help suppress pulmonary colonization, including in models of cystic fibrosis [140,141]. Furthermore, inhaled co-administration of anti-PA IgY with the macrocyclic peptide POPSICAL led to a >99% reduction in biofilm and bacterial burden in mouse lungs, demonstrating promising antibiofilm activity [142]. However, several studies have reported limited or no protective efficacy under certain conditions. In a rabbit sepsis model, anti-chimeric PilQ/PilA IgY failed to reduce mortality or bacterial burden [143]. Similarly, in a porcine model of ventilator-associated pneumonia, intravenous IgY administration did not significantly reduce pulmonary PA loads, likely reflecting the limited systemic bioavailability of IgY when delivered intravenously [99].
IgY antibodies have also demonstrated antimicrobial potential against a variety of other bacterial pathogens. In a comparative study, anti-Staphylococcus aureus IgY produced using Freund’s incomplete adjuvant significantly inhibited bacterial growth, with higher antigen-binding activity compared to other adjuvants [75]. In a murine model of Salmonella typhimurium infection, specific IgY antibodies, when delivered via alginate nanoparticles, provided full protection, whereas non-encapsulated IgY was only partially protective, highlighting the value of nanoformulations for enhancing efficacy [144]. Studies on Vibrio cholerae showed that anti-V. cholerae O1 IgY could significantly reduce mortality in infected mice, with survival rates improving with higher IgY doses [40], while antibodies targeting a chimeric outer membrane protein similarly demonstrated high immunoreactivity and reduced colonization [33]. In the case of Enterococcus faecalis, IgY antibodies against cytolysin protected microbiota-humanized mice from ethanol-induced liver damage by neutralizing cytotoxic effects in primary hepatocytes [145].
The use of IgY antibodies in dentistry has emerged as a promising non-invasive strategy for the prevention and management of oral infectious diseases, particularly dental caries, periodontitis, and dentin hypersensitivity. Anti-Streptococcus mutans IgY significantly reduced bacterial adhesion and growth in caries-active children following administration via mouth spray or lozenges, indicating its potential as a passive immunotherapeutic in high-risk pediatric populations [104,146]. Similarly, anti-Porphyromonas gingivalis IgY inhibited bacterial adhesion and invasion in human periodontal ligament fibroblasts, supporting its use in the prevention and treatment of periodontal disease [41]. Broader-spectrum formulations targeting multiple periodontitis-associated pathogens, such as Aggregatibacter actinomycetemcomitans and Porphyromonas gingivalis, demonstrated the ability to bind bacterial cell walls, promote agglutination, and disrupt biofilm formation, while maintaining stability under varied pH, temperature, and storage conditions [147]. In clinical evaluations, the application of IgY as a local drug delivery agent or mouthwash adjunct to scaling and root planing significantly reduced colony-forming units in both gingivitis and periodontitis patients [148]. Additionally, an IgY–amorphous calcium phosphate (ACP) complex demonstrated dual functionality by occluding dentinal tubules under acid challenge and inhibiting Streptococcus mutans biofilms, highlighting its promise in dentin hypersensitivity (DH) management [149].
In the context of fungal infections, IgY antibodies have demonstrated promising antifungal activity against Candida albicans, a common opportunistic pathogen. IgY raised against the iron permease Ftr1 from C. albicans exhibited high-affinity binding and fungicidal activity, leading to increased survival in Galleria mellonella larvae infected with C. albicans [31]. In a murine model of vaginal candidiasis, hyperimmune egg-based IgY formulations significantly reduced fungal burden—achieving a 99.6% reduction in colony-forming units (CFUs) by day 15—and diminished inflammation and filamentous yeast forms in vaginal lavage samples, supporting its utility as an adjuvant therapy [150].
The use of IgY has extended beyond human medicine into a wide variety of veterinary applications. In broiler chickens, feed supplementation with microencapsulated IgY significantly improved average daily gain and reduced feed conversion ratio, outperforming conventional antibiotics and supporting more sustainable practices [151]. Anti-Clostridium perfringens IgY alleviated clinical signs and normalized hematological and biochemical parameters in experimentally infected birds [113], while maternally or orally delivered IgY targeting Escherichia coli, Salmonella enteritidis, and Clostridium perfringens reduced pathogen colonization, improved immune responses, and was more effective when used prophylactically [111]. Against viral infections, intramuscular administration of anti-Newcastle disease virus IgY improved hemagglutination inhibition titers and survival in broilers, with better performance observed for intramuscular compared to subcutaneous routes [152]. In ducks, specific IgY provided 100% protection against Riemerella anatipestifer when administered shortly after infection [153], and markedly reduced mortality from avian influenza H5 virus in Muscovy ducks [81] .
IgY-based immunotherapies have also shown promising results in companion animals and livestock. In dogs, chicken-derived IgY-scFv antibodies targeting the capsid protein of canine parvovirus were used for diagnostic and neutralization purposes [154]. In cats, dietary anti-Fel d 1 IgY reduced exposure to the major allergen without affecting feline health or growth [35], and a human proof-of-concept study demonstrated that feeding cats anti-Fel d 1 IgY led to a notable reduction in allergy symptoms among sensitized individuals [155]. In calves, E. coli K99-specific IgY, combined with probiotics, improved growth, reduced diarrhea incidence, and supported immune function during the pre-weaning period [110], while IgY targeting bovine coronavirus significantly delayed the onset and reduced the severity of clinical episodes and viral shedding in neonatal diarrhea models, with successful scale-up for industrial production [29]. In equine applications, IgY against Salmonella enterica serovar Newport inhibited bacterial motility and growth in vitro in a dose-dependent manner [156]. More broadly, IgY has demonstrated cross-species growth-inhibitory effects against key zoonotic pathogens including Campylobacter jejuni, Salmonella spp., and Escherichia coli, supporting its utility as a sustainable alternative to antibiotics in livestock management [157].
The frequent use of antibiotics in aquaculture to control bacterial diseases has led to the rise of the antimicrobial resistance of the pathogenic bacteria in this sector of activity. As an alternative, IgY antibodies can successfully be used for their protective effects against viruses like nervous necrosis virus -NNV [158,159], tilapia lake virus – TiLV [160] and infections produced by Vibrio parahaemolyticus [161], Aeromonas veronii [39], Aeromonas hydrophila [162], Edwardsiella tarda [163], or Yersinia ruckeri [164].
IgY-based antivenoms have also been developed, demonstrating the ability to neutralize cobra venom, (Naja Naja)[165,166], krait (Bungarus caerulens)[165], white-lipped pit viper (Trimeresurus albolabris)[167], crossed pit viper (Bothrops alternatus)[168], scorpion (Tityus caripitensis)[169], bee venom (Apis mellifera) [170].
Further applications in aquaculture, poultry, and human dental care reveal the broad spectrum of IgY utility, highlighting its role as an alternative to antibiotics. Collectively, these findings support IgY’s incorporation in One Health strategies to combat antimicrobial resistance.
4.3. Clinical Trials Using IgY
Clinical trials investigating IgY have shown promising results across diverse medical conditions, including infectious, gastrointestinal, respiratory, and inflammatory diseases. As summarized in Table 2, studies registered on ClinicalTrials.gov highlight both prophylactic and therapeutic applications of IgY. For instance, a completed trial (NCT02705885) evaluated IgY antibodies against Porphyromonas gingivalis as a dietary supplement in patients with chronic periodontitis, supporting its role in oral health. Notably, several trials (NCT00633191, NCT01455675) explored the efficacy of anti-Pseudomonas IgY in cystic fibrosis patients, demonstrating its potential in preventing recurrent pulmonary infections. Furthermore, Phase I studies (NCT04567810, NCT06702280) assessed the safety of IgY against SARS-CoV-2 and its impact on gut microbiota in healthy individuals. Another investigation (NCT02721355) tested anti-Helicobacter pylori IgY for chronic gastritis, while NCT03058224 targeted fibromyalgia with IgY formulations against E. coli and Salmonella. These findings underscore IgY’s expanding clinical relevance, with a favourable safety profile and broad-spectrum applicability in human health.
5. Advantages and Limitations of Using IgY Compared to IgG
Although chicken egg yolk immunoglobulins are not yet fully utilized, they offer several advantages over their mammalian counterparts. First, IgY technology enhances animal welfare and helps better meet the requirements of the 3R principle: reduction, because laying hens produce a higher yield of immunoglobulins per month (1000-2800 mg/chicken/month) compared to rabbits (200 mg/rabbit/month), thus reducing the number of animals required [17]; replacement and refinement, as antibodies are secreted directly into the egg, eliminating the need for invasive collection procedures, such as blood sampling or sacrifice [63].
IgY technology also possesses economical advantages over conventional antibody production, as the cost of maintaining a laying hen is less expensive that maintaining mice or rabbits, while the lower number of animals needed further contributing to the cost-effectiveness of this method [4]. A chicken usually lays around 280 eggs in a year, and with an average egg yolk containing 100-150mg of IgY, this can result in a yield of 28-42 g of IgY per year per chicken [16]. Although the laying capacity decreases in the second year of egg laying, this is compensated by a greater yield of IgY per egg [175]. Other avian species can be used for IgY production, such as ducks, geese, ostriches, quails or emu. [17,176].
The structural and phylogenetic differences between IgY and IgG result in distinct molecular and biochemical interactions. The Fc region of IgY does not activate the human complement system, bind to rheumatoid factor, or interact with protein G [4]. Avian immunoglobulin does not cross-react with human anti-mouse antibodies (HAMA), and due to their phylogenetic divergence, avian species elicit a stronger immune response against conserved mammalian proteins [17]. Compared to IgG antibodies produced by mammals, IgY antibodies have similar affinity and avidity [53]. Hens can also generate a strong immune response with lower antigen quantities compared to mammals, thereby reducing the amount of antigen needed for antibody production [16].
In regards to oral IgY therapy, inactivation by digestive processes can be an important challenge, as IgY is sensitive to gastric acidity and protease enzymes such as pepsin [177]. In order to overcome this challenge, the use of microencapsulation using liposomes, chitosan-alginate or whey protein-based microcapsules, macroencapsulation in enteric coated gelatin capsules or even the concurrent use of cytoprotective agent sucralfate have been proposed, with various degrees of success [175].
Another concern relates to the triggering of allergic, systemic and local responses, as IgY may function as an antigen itself. The repeated long-term administration of three IgY formulations in non-microbially-challenged mice showed that in the absence of significant amounts of their target antigens, IgYs can influence various immunological processes in ways that are not yet completely understood or easy to predict [178].
The lack of a standardized IgY production protocol may be another factor that hampers the widespread use of this technology. Although attempts have been made to standardize different steps of production [6,26], variability exists in immunization schedules, adjuvant selection, IgY extraction and purification methods, as well as storage conditions, all of which can influence yield and antibody quality. The absence of standardized guidelines makes it challenging to compare results across studies and implement large-scale production efficiently. Further research and collaboration are needed to establish optimized, reproducible protocols that ensure consistency and facilitate the broader adoption of IgY technology.
6. Conclusions
Immunoglobulin Y represents a promising immunotechnological platform, offering several advantages over conventional mammalian IgG-based approaches. In the context of rising antimicrobial resistance and the integrative framework of the One Health concept, IgY is increasingly recognized as a valuable tool in diagnostics, prophylaxis, and therapeutics—at the intersection of human health, animal health, and food safety. The use of IgY is steadily expanding, supported by a growing body of published research and the emergence of commercial products. With continued efforts to address current limitations, standardize production protocols, and further evaluate its safety profile, IgY technology is well-positioned to become an accessible and versatile solution for a broad range of applications in the near future.
Author Contributions
Conceptualization, R.C. and M.M.; methodology, R.C., D.C.-S., V.N., M.M.; software, R.C., D.C-S; resources, R.C., D.C.-S., A.-C.B.-I.; writing—original draft preparation, R.C., D.C.-S., A.-C.B.-I., V.N., M.M.; writing—review and editing, R.C., D.C.-S., M.M.; visualization, R.C., D.C.-S., A.-C.B.-I., V.N., M.M.; supervision, M.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
All figures created with BioRender.com.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| IgY | Immunoglobulin Y |
| IgG | Immunoglobulin G |
| Fc | Fragment crystallizable |
| Fab | Fragment antigen-binding |
| Fv | Variable fragment |
| VH | Variable region of the heavy chain |
| VL | Variable region of the light chain |
| CL | Constant region of the light chain |
| CH | Constant region of the heavy chain |
| FcRY | Fc receptor for IgY (avian neonatal Fc receptor) |
| ECVAM | European Centre for the Validation of Alternative Methods |
| WSF | Water-soluble filtrate |
| SDS-PAGE | Sodium dodecyl sulfate polyacrylamide gel electrophoresis |
| HPLC | High-performance liquid chromatography |
| BCA | Bicinchoninic acid assay |
| ELISA | Enzyme-linked immunosorbent assay |
| ETEC | Enterotoxigenic Escherichia coli |
| LTB | Heat-labile toxin subunit B |
| OmpW | Outer membrane protein W |
| CtxB | Cholera toxin subunit B |
| HPDLF | Human periodontal ligament fibroblast |
| DH | Dentin hypersensitivity |
| CFU | Colony-forming unit |
| VN | Virus neutralization |
| HAMA | Human anti-mouse antibodies |
| CRISPR | Clustered Regularly Interspaced Short Palindromic Repeats |
| SpCas9 | Streptococcus pyogenes Cas9 |
| SARS-CoV-2 | Severe Acute Respiratory Syndrome Coronavirus 2 |
| COVID-19 | Coronavirus Disease 2019 |
| NCT | ClinicalTrials.gov identifier number |
| PA | Pseudomonas aeruginosa |
| AI | Avian influenza |
| IBDV | Infectious Bursal Disease Virus |
| ZEN | Zearalenone |
| BCoV | Bovine Coronavirus |
| NCD | Neonatal Calf Diarrhea |
References
- Langel, S.N.; Blasi, M.; Permar, S.R. Maternal Immune Protection against Infectious Diseases. Cell Host Microbe 2022, 30, 660–674. [Google Scholar] [CrossRef]
- Niewiesk, S. Maternal Antibodies: Clinical Significance, Mechanism of Interference with Immune Responses, and Possible Vaccination Strategies. Front. Immunol. 2014, 5, 446. [Google Scholar] [CrossRef]
- Okamoto, M.; Sasaki, R.; Ikeda, K.; Doi, K.; Tatsumi, F.; Oshima, K.; Kojima, T.; Mizushima, S.; Ikegami, K.; Yoshimura, T.; et al. FcRY Is a Key Molecule Controlling Maternal Blood IgY Transfer to Yolks during Egg Development in Avian Species. Front. Immunol. 2024, 15. [Google Scholar] [CrossRef]
- Pereira, E.P.V.; van Tilburg, M.F.; Florean, E.O.P.T.; Guedes, M.I.F. Egg Yolk Antibodies (IgY) and Their Applications in Human and Veterinary Health: A Review. Int. Immunopharmacol. 2019, 73, 293–303. [Google Scholar] [CrossRef]
- Leslie, G.A.; Clem, L.W. Phylogen of Immunoglobulin Structure and Function. 3. Immunoglobulins of the Chicken. J. Exp. Med. 1969, 130, 1337–1352. [Google Scholar] [CrossRef]
- Schade, R.; Staak, C.; Hendriksen, C.; Erhard, M.; Hugl, H.; Koch, G.; Larsson, A.; Pollmann, W.; van Regenmortel, M.; Rijke, E.; et al. The Production of Avian (Egg Yolk) Antibodies: IgY: The Report and Recommendations of ECVAM Workshop 211,2. Altern. Lab. Anim. 1996, 24, 925–934. [Google Scholar] [CrossRef]
- Charles A Janeway, J.; Travers, P.; Walport, M.; Shlomchik, M.J. The Structure of a Typical Antibody Molecule. In Immunobiology: The Immune System in Health and Disease. 5th edition; Garland Science, 2001.
- Hersh, R.T.; Kubo, R.T.; Leslie, G.A.; Benedict, A.A. Molecular Weights of Chicken, Pheasant, and Quail IgG Immunoglobulins. Immunochemistry 1969, 6, 762–765. [Google Scholar] [CrossRef]
- Karachaliou, C.-E.; Vassilakopoulou, V.; Livaniou, E. IgY Technology: Methods for Developing and Evaluating Avian Immunoglobulins for the in Vitro Detection of Biomolecules. World J. Methodol. 2021, 11, 243–262. [Google Scholar] [CrossRef]
- Gandhi, S.; Alshehri, S.M. Molecular Stability of the Rabbit and Chicken Egg Yolk Immunoglobulins. Front. Biosci. Elite Ed. 2021, 13, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Härtle, S.; Magor, K.E.; Göbel, T.W.; Davison, F.; Kaspers, B. Chapter 5 - Structure and Evolution of Avian Immunoglobulins. In Avian Immunology (Third Edition); Kaspers, B., Schat, K.A., Göbel, T.W., Vervelde, L., Eds.; Academic Press: Boston, 2022; pp. 101–119 ISBN 978-0-12-818708-1. [Google Scholar]
- Gallagher, J.S.; Voss, E.W. Immune Precipitation of Purified Chicken Antibody at Low pH. Immunochemistry 1970, 7, 771–785. [Google Scholar] [CrossRef]
- Higgins, D.A. Precipitating Antibodies of the Duck (Anas Platyrhynchos). Comp. Biochem. Physiol. Part B Comp. Biochem. 1989, 93, 135–144. [Google Scholar] [CrossRef]
- Spillner, E.; Braren, I.; Greunke, K.; Seismann, H.; Blank, S.; du Plessis, D. Avian IgY Antibodies and Their Recombinant Equivalents in Research, Diagnostics and Therapy. Biologicals 2012, 40, 313–322. [Google Scholar] [CrossRef]
- Lee, W.; Syed Atif, A.; Tan, S.C.; Leow, C.H. Insights into the Chicken IgY with Emphasis on the Generation and Applications of Chicken Recombinant Monoclonal Antibodies. J. Immunol. Methods 2017, 447, 71–85. [Google Scholar] [CrossRef]
- Karlsson, M.; Kollberg, H.; Larsson, A. Chicken IgY: Utilizing the Evolutionary Advantage. Worlds Poult. Sci. J. 2004, 60, 341–348. [Google Scholar] [CrossRef]
- IgY-Technology: Production and Application of Egg Yolk Antibodies: Basic Knowledge for a Successful Practice; Zhang, X.-Y., Vieira-Pires, R.S., Morgan, P.M., Schade, R., Eds.; Springer International Publishing: Cham, 2021; ISBN 978-3-030-72686-7. [CrossRef]
- Chiu, M.L.; Goulet, D.R.; Teplyakov, A.; Gilliland, G.L. Antibody Structure and Function: The Basis for Engineering Therapeutics. Antibodies 2019, 8, 55. [Google Scholar] [CrossRef]
- Carlander, D.; Larsson, A. Avian Antibodies Can Eliminate Interference Due To Complement Activation In ELISA. Ups. J. Med. Sci. 2001, 106, 189–195. [Google Scholar] [CrossRef]
- Leiva, C.L.; Gallardo, M.J.; Casanova, N.; Terzolo, H.; Chacana, P. IgY-Technology (Egg Yolk Antibodies) in Human Medicine: A Review of Patents and Clinical Trials. Int. Immunopharmacol. 2020, 81, 106269. [Google Scholar] [CrossRef]
- Gassmann, M.; Thömmes, P.; Weiser, T.; Hübscher, U. Efficient Production of Chicken Egg Yolk Antibodies against a Conserved Mammalian Protein. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 1990, 4, 2528–2532. [Google Scholar] [CrossRef]
- Wang, X.; Li, S.; Zhao, R.; Li, H.; Gao, P.; Jin, C.; Fang, C.; Liu, Y.; Hei, A.; Zhou, J.; et al. Development of a Novel Recombinant Full-Length IgY Monoclonal Antibody against Human Thymidine Kinase 1 for Automatic Chemiluminescence Analysis on a Sandwich Biotin-Streptavidin Platform for Early Tumour Discovery. J. Immunol. Res. 2023, 2023, 7612566. [Google Scholar] [CrossRef]
- Li, X.; He, P.; Yu, L.; He, Q.; Jia, C.; Yang, H.; Lu, M.; Wei, X.; Zhao, S. Production and Characteristics of a Novel Chicken Egg Yolk Antibody (IgY) against Periodontitis-Associated Pathogens. J. Oral Microbiol. 12, 1831374. [CrossRef]
- Nilsson, E.; Stålberg, J.; Larsson, A. IgY Stability in Eggs Stored at Room Temperature or at +4°C. Br. Poult. Sci. 2012, 53, 42–46. [Google Scholar] [CrossRef]
- Shimizu, M.; Nagashima, H.; Hashimoto, K.; Suzuki, T. Egg Yolk Antibody (Ig Y) Stability in Aqueous Solution with High Sugar Concentrations. J. Food Sci. 1994, 59, 763–765. [Google Scholar] [CrossRef]
- Schade, R.; Gutierrez Calzado, E.J.; Sarmiento, R.; Chacana, P.; Porankiewicz-Asplund, J.; Horacio Raúl, T. Chicken Egg Yolk Antibodies (IgY-Technology): A Review of Progress in Production and Use in Research and Human and Veterinary Medicine. Altern. Lab. Anim. ATLA 2005, 33, 129–154. [Google Scholar] [CrossRef]
- Bachtiar, E.W.; Bachtiar, B.M.; Soejoedono, R.D.; Wibawan, I.W.; Afdhal, A. Biological and Immunogenicity Property of IgY Anti S. Mutans ComD. Open Dent. J. 2016, 10, 308–314. [Google Scholar] [CrossRef]
- Ahmadi, T.S.; Behrouz, B.; Mousavi Gargari, S.L. Polyclonal Anti-Whole Cell IgY Passive Immunotherapy Shields against P. Aeruginosa-Induced Acute Pneumonia and Burn Wound Infections in Murine Models. Sci. Rep. 2024, 14, 405. [Google Scholar] [CrossRef]
- Bok, M.; Vega, C.G.; Castells, M.; Colina, R.; Wigdorovitz, A.; Parreño, V. Development of an IgY-Based Treatment to Control Bovine Coronavirus Diarrhea in Dairy Calves. Viruses 2023, 15, 708. [Google Scholar] [CrossRef]
- Qiu, T.; Zhang, H.; Lei, H.; Zhang, L.; Zhang, Y.; Shen, X.; Xu, B.; Zhu, J.; Xiao, W.; Zheng, J.; et al. Preparation of Anti-Zearalenone IgY and Development of an Indirect Competitive ELISA Method for the Measurement of Zearalenone in Post-Fermented Tea. Foods 2023, 12, 4478. [Google Scholar] [CrossRef]
- de Souza, P.C.; Corrêa, A.E. do N.; Gameiro, J.G.; de Oliveira Júnior, A.G.; Panagio, L.A.; Venancio, E.J.; Almeida, R.S. Production of IgY against Iron Permease Ftr1 from Candida Albicans and Evaluation of Its Antifungal Activity Using Galleria Mellonella as a Model of Systemic Infection. Microb. Pathog. 2023, 181, 106166. [Google Scholar] [CrossRef] [PubMed]
- Hadi, N.; Nazarian, S.; Rouhi, S.; Hosseini, S.E.; Fathi, J. Production of Egg Yolk Antibody (IgY) against a Chimeric Protein Containing IpaD, StxB, and TolC Antigens from Shigella: An Investigation of Its Prophylactic Effects against Shiga Toxin (Stx) and Shigella Dysenteriae in Vitro and in Vivo. Heliyon 2024, 10, e26361. [Google Scholar] [CrossRef]
- Hoseini, H.S.N.; Ahmadi, T.S.; Gargari, S.L.M.; Nazarian, S. IgY-Mediated Protection against Vibrio Cholerae Infection: Efficacy of Avian Antibodies Targeting a Chimeric Recombinant Protein. BioImpacts 2024. [Google Scholar] [CrossRef]
- Mohammadkhani, F.; Mousavi Gargari, S.L.; Nazarian, S.; Mafi, M. Protective Effects of Anti-CfaB-EtpA-LTB IgY Antibody against Adherence and Toxicity of Enterotoxigenic Escherichia Coli (ETEC). J. Appl. Microbiol. 2023, 134, lxad013. [Google Scholar] [CrossRef]
- Hedrick, E.D.; Matulka, R.A.; Conboy-Schmidt, L.; May, K.A. Evaluation of Anti-Fel d 1 IgY Ingredient for Pet Food on Growth Performance in Kittens. Front. Vet. Sci. 2024, 11. [Google Scholar] [CrossRef] [PubMed]
- Kpordze, S.W.; Mobegi, V.A.; Kikuvi, G.M.; Gikunju, J.K.; Setsoafia Saba, C.K.; Moshe, J.; Kimotho, J.H. Generation of Chicken-Based IgY Polyclonal Antibodies against Dendroaspis Polylepis and Preclinical Evaluation of Envenomation-Neutralizing Efficacy Vis-à-Vis Selected Commercial Antivenoms. Toxicon X 2024, 23, 100201. [Google Scholar] [CrossRef] [PubMed]
- Mubarak, A.; Alturaiki, W.; Dawoud, T.; El-Tayeb, M.; Elbadawi, Y.; Moussa, I. Production of Anti-Camel IgY for Diagnosis of Infectious Diseases Affecting Camels Located in Kingdom of Saudi Arabia. J. King Saud Univ. - Sci. 2023, 35, 102421. [Google Scholar] [CrossRef]
- TETİK, K.; SAREYYÜPOĞLU, B. Production, Purification, and Specificity Serologically Determination of Immunoglobulin-Y (IgY) from Chicken Eggs against Clostridium Tetani Toxoid. Turk. J. Vet. Anim. Sci. 2024, 48, 165–173. [Google Scholar] [CrossRef]
- Li, Y.; Huang, B.; Sun, S.; Liu, N.; Li, Y.; Lan, M.; Wang, X.; Zhang, Y.; Wu, A.; Yang, S.; et al. Immunoprotection Effects of Chicken Egg Yolk Immunoglobulins (IgY) against Aeromonas Veronii Infection in Sinocyclocheilus Grahami. Aquaculture 2023, 563, 738935. [Google Scholar] [CrossRef]
- Shoushtari, M.; Shooshtari, A.B.; Asadi, S.; Karami, Y.; Honari, M.; Alizadeh, G.A.; Zeinoddini, M.; Fathi, J. Production of Egg Yolk Antibody (IgY) Against Vibrio Cholerae O1: Protective Effect in Mice. Avicenna J. Med. Biotechnol. 2023, 15, 239–244. [Google Scholar] [CrossRef]
- Su, H.; Wei, K.; Zhao, M.; Li, X.; Zhang, Y. Research Note: A Novel Method for Preparation of Egg Yolk Immunoglobulin Y against Porphyromonas Gingivalis. Poult. Sci. 2023, 102, 102863. [Google Scholar] [CrossRef]
- Bouazzaoui, A.; Bogari, N.M.; Al-Allaf, F.A.; Ekram, S.N.; Athar, M.; Dannoun, A.; Schubert, T.; Syed, S.N.; Youssef, A.-R.; Alqahtani, M.; et al. Anti-E. Coli Immunoglobulin Yolk (IgY): Reduction of Pathogen Receptors and Inflammation Factors Could Be Caused by Decrease in E. Coli Load. Heliyon 2023, 9, e13876. [Google Scholar] [CrossRef]
- Guo, Y.; Ma, Y.; Endri, H.; Huang, Y.; Pan, X.; Qiu, S.; Cao, Y.; Gu, W.; Du, J.; Wang, L.; et al. Protective Effect of Chicken Egg Yolk Immunoglobulin (IgY) against Spiroplasma Eriocheiris Infection in Chinese Mitten Crab. Aquaculture 2023, 572, 739488. [Google Scholar] [CrossRef]
- Liu, X.; Xiao, H.; Cui, P.; Chen, J.; Chao, J.; Wu, X.; Lu, J.; Zhang, X.; Xu, G.; Liu, Y. Differential Polyvalent Passive Immune Protection of Egg Yolk Antibodies (IgY) against Live and Inactivated Vibrio Fluvialis in Fish. Fish Shellfish Immunol. 2024, 151, 109751. [Google Scholar] [CrossRef]
- León, E.; Ortiz, V.; Pérez, A.; Téllez, J.; Díaz, G.J.; Ramírez H, M.H.; Contreras R, L.E. Anti-SpCas9 IgY Polyclonal Antibodies Production for CRISPR Research Use. ACS Omega 2023, 8, 33809–33818. [Google Scholar] [CrossRef] [PubMed]
- Montini, M.P.O.; Fernandes, E.V.; Ferraro, A.C.N. dos S.; Almeida, M.A.; da Silva, F.C.; Venancio, E.J. Effects of Inoculation Route and Dose on Production and Avidity of IgY Antibodies. Food Agric. Immunol. 2018, 29, 306–315. [Google Scholar] [CrossRef]
- Emam, M.; Mehrabani-Yeganeh, H.; Barjesteh, N.; Nikbakht, G.; Thompson-Crispi, K.; Charkhkar, S.; Mallard, B. The Influence of Genetic Background versus Commercial Breeding Programs on Chicken Immunocompetence. Poult. Sci. 2014, 93, 77–84. [Google Scholar] [CrossRef]
- Forte, C.; Moscati, L.; Acuti, G.; Mugnai, C.; Franciosini, M.P.; Costarelli, S.; Cobellis, G.; Trabalza-Marinucci, M. Effects of Dietary Lactobacillus Acidophilus and Bacillus Subtilis on Laying Performance, Egg Quality, Blood Biochemistry and Immune Response of Organic Laying Hens. J. Anim. Physiol. Anim. Nutr. 2016, 100, 977–987. [Google Scholar] [CrossRef]
- Meethenkul (PDF) Comparison of Immunoglobulin Y Antibody Production in New and Spent Laying Hens. ResearchGate 2024. [CrossRef]
- Erhard, M.H.; Özpinar, H.; Bilal, T.; Abbas, Y.; Kutay, C.; Eseceli, H.; Stangassinger, M. The Humoral Immune Response and the Productivity of Laying Hens Kept on the Ground or in Cages. Altern. Lab. Anim. 2000, 28, 699–705. [Google Scholar] [CrossRef]
- He, J.-X.; Thirumalai, D.; Schade, R.; Zhang, X.-Y. Chronobiological Studies of Chicken IgY: Monitoring of Infradian, Circadian and Ultradian Rhythms of IgY in Blood and Yolk of Chickens. Vet. Immunol. Immunopathol. 2014, 160, 266–272. [Google Scholar] [CrossRef]
- Burakova, Y.; Madera, R.; McVey, S.; Schlup, J.R.; Shi, J. Adjuvants for Animal Vaccines. Viral Immunol. 2018, 31, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, B.L.B.; Nogueira, C.P.; Venancio, E.J. IgY Antibodies from Birds: A Review on Affinity and Avidity. Animals 2023, 13, 3130. [Google Scholar] [CrossRef] [PubMed]
- Marcq, C.; Marlier, D.; Beckers, Y. Improving Adjuvant Systems for Polyclonal Egg Yolk Antibody (IgY) Production in Laying Hens in Terms of Productivity and Animal Welfare. Vet. Immunol. Immunopathol. 2015, 165, 54–63. [Google Scholar] [CrossRef]
- Hu, B.; Yang, X.; Guo, E.; Zhou, P.; Xu, D.; Qi, Z.; Deng, L. The Preparation and Antibacterial Effect of Egg Yolk Immunoglobulin (IgY) against the Outer Membrane Proteins of Vibrio Parahaemolyticus. J. Sci. Food Agric. 2019, 99, 2565–2571. [Google Scholar] [CrossRef]
- Tirado, J.; Caza, J.; Tufiño, A.; Pardo, H.; Muñoz, D.; Manjunatha, B. DEVELOPMENT, PURIFICATION, AND CHARACTERIZATION OF IGY ANTIBODIES AGAINST VITELLOGENIN OF THE "VIEJA COLORADA” FISH (CICHLASOMA FESTAE), A SPECIES NATIVE TO THE GUAYAS RIVER, ECUADOR. J. Microbiol. Biotechnol. Food Sci. 2022, 12, e3159–e3159. [Google Scholar] [CrossRef]
- Felegary, A.; Nazarian, S.; Zafarmand-Samarin, M.; Sadeghi, D.; Fathi, J.; Samiei-Abianeh, H. Evaluation of the Prophylactic Effect of Egg Yolk Antibody (IgY) Produced against the Recombinant Protein Containing IpaD, IpaB, StxB, and VirG Proteins from Shigella. Mol. Immunol. 2024, 173, 53–60. [Google Scholar] [CrossRef]
- Schön, J.; Aebischer, A.; Halwe, N.J.; Ulrich, L.; Hoffmann, D.; Reiche, S.; Beer, M.; Grund, C. Evaluation of SARS-CoV-2-Specific IgY Antibodies: Production, Reactivity, and Neutralizing Capability against Virus Variants. Int. J. Mol. Sci. 2024, 25, 7976. [Google Scholar] [CrossRef]
- Kubo, N.; Nishii, M.; Osada-Oka, M.; Hatta, H. A Comparative Study on Egg Yolk IgY Production with Different Adjuvants and Their Inhibitory Effects on Staphylococcus Aureus. J. Poult. Sci. 2021, 58, 192–199. [Google Scholar] [CrossRef]
- Rizkiantino, R.; Wibawan, I.W.T.; Pasaribu, F.H.; Soejoedono, R.D.; Poetri, O.N.; Arnafia, W.; Sasi, K.D.; Reisinta, D. THE POTENTIAL OF ADJUVANT AGAINST PRODUCTION OF ANTISTREPTOCOCCAL IMMUNOGLOBULIN Y (IGY) IN AQUACULTURE. J. Kedokt. Hewan - Indones. J. Vet. Sci. 2020, 14. [Google Scholar] [CrossRef]
- Sun, B.-Y.; Kou, H.-Y.; Jian, P.-Y.; Kong, L.-J.; Fang, J.; Meng, P.-K.; Wu, K.; Yang, C.-G.; Yang, G.; Song, X.-H. Protective Effects of Egg Yolk Immunoglobulins (IgY) against CyHV-2 Infection in Gibel Carp (Carassius Gibelio). Aquaculture 2023, 569, 739371. [Google Scholar] [CrossRef]
- Amro, W.A.; Al-Qaisi, W.; Al-Razem, F. Production and Purification of IgY Antibodies from Chicken Egg Yolk. J. Genet. Eng. Biotechnol. 2018, 16, 99–103. [Google Scholar] [CrossRef]
- Marcq, C.; Théwis, A.; Portetelle, D.; Beckers, Y. Refinement of the Production of Antigen-Specific Hen Egg Yolk Antibodies (IgY) Intended for Passive Dietary Immunization in Animals. A Review. BASE 2013. [Google Scholar]
- Santos, J.S.; Araújo, I.C.S.; Lacerda, M.J.R.; Andrade, M.A.; Café, M.B.; Leandro, N.S.M.; Stringhini, J.H. Effects of Broiler Breeder Age on Immune System Development of Progeny. R Bras Zootec 2022, 51. [Google Scholar] [CrossRef]
- Leenaars, M.; Hendriksen, C.F.M. Critical Steps in the Production of Polyclonal and Monoclonal Antibodies: Evaluation and Recommendations. ILAR J. 2005, 46, 269–279. [Google Scholar] [CrossRef]
- Artman, C.; Idegwu, N.; Brumfield, K.D.; Lai, K.; Hauta, S.; Falzarano, D.; Parreño, V.; Yuan, L.; Geyer, J.D.; Goepp, J.G. Feasibility of Polyclonal Avian Immunoglobulins (IgY) as Prophylaxis against Human Norovirus Infection. Viruses 2022, 14, 2371. [Google Scholar] [CrossRef]
- Zambrowicz, A.; Dąbrowska, A.; Bobak, Ł.; Szołtysik, M. Egg Yolk Proteins and Peptides with Biological Activity. Postępy Hig. Med. Dośw. 2014, 68, 1524–1529. [Google Scholar] [CrossRef]
- Ahn, D.; Kim, S.; Shu, H. Effect of Egg Size and Strain and Age of Hens on the Solids Content of Chicken Eggs. Poult. Sci. 1997, 76, 914–919. [Google Scholar] [CrossRef]
- Zhao, Y.; Feng, F.; Yang, Y.; Xiong, C.; Xu, M.; Tu, Y. Gelation Behavior of Egg Yolk under Physical and Chemical Induction: A Review. Food Chem. 2021, 355, 129569. [Google Scholar] [CrossRef]
- De Meulenaer, B.; Huyghebaert, A. Isolation and Purification of Chicken Egg Yolk Immunoglobulins: A Review. Food Agric. Immunol. 2001, 13, 275–288. [Google Scholar] [CrossRef]
- Akita, E.M.; Nakai, S. Immunoglobulins from Egg Yolk: Isolation and Purification. J. Food Sci. 1992, 57, 629–634. [Google Scholar] [CrossRef]
- Hodek, P.; Trefil, P.; Šimůnek, J.; Hudecek, J.; Stiborova, M. Optimized Protocol of Chicken Antibody (IgY) Purification Providing Electrophoretically Homogenous Preparations. Int. J. Electrochem. Sci. 2013, 8, 113–124. [Google Scholar] [CrossRef]
- Polson, A.; von Wechmar, M.B.; van Regenmortel, M.H. Isolation of Viral IgY Antibodies from Yolks of Immunized Hens. Immunol. Commun. 1980, 9, 475–493. [Google Scholar] [CrossRef]
- Ko, K.Y.; Ahn, D.U. Preparation of Immunoglobulin Y from Egg Yolk Using Ammonium Sulfate Precipitation and Ion Exchange Chromatography. Poult. Sci. 2007, 86, 400–407. [Google Scholar] [CrossRef]
- Kubo, N.; Nishii, M.; Osada-Oka, M.; Hatta, H. A Comparative Study on Egg Yolk IgY Production with Different Adjuvants and Their Inhibitory Effects on Staphylococcus Aureus. J. Poult. Sci. 2021, 58, 192–199. [Google Scholar] [CrossRef]
- Tan, S.H.; Mohamedali, A.; Kapur, A.; Lukjanenko, L.; Baker, M.S. A Novel, Cost-Effective and Efficient Chicken Egg IgY Purification Procedure. J. Immunol. Methods 2012, 380, 73–76. [Google Scholar] [CrossRef]
- Asemota, H.; Curtello, S.; Vaillant, A.; Mohammed, W.; Vuma, S.; AV, C.; Kurhade, A.; Kissoon, S.; Smikle, M.; Wisdom, B.; et al. Purification of Avian IgY with Trichloroacetic Acid (TCA). J Chromatogr. Separat Tech. 2013, 4. [Google Scholar] [CrossRef]
- McLaren, R.D.; Prosser, C.G.; Grieve, R.C.J.; Borissenko, M. The Use of Caprylic Acid for the Extraction of the Immunoglobulin Fraction from Egg Yolk of Chickens Immunised with Ovine α-Lactalbumin. J. Immunol. Methods 1994, 177, 175–184. [Google Scholar] [CrossRef]
- Duong-Ly, K.C.; Gabelli, S.B. Chapter Seven - Salting out of Proteins Using Ammonium Sulfate Precipitation. In Methods in Enzymology; Lorsch, J., Ed.; Laboratory Methods in Enzymology: Protein Part C; Academic Press, 2014; Vol. 541, pp. 85–94. [CrossRef]
- Deignan, T.; Kelly, J.; Alwan, A.; O’Farrelly, C. Comparative Analysis of Methods of Purification of Egg Yolk Immunoglobulin. Food Agric. Immunol. 2000, 12, 77–85. [Google Scholar] [CrossRef]
- Radwan, F.; Elshemy, A.; Toaleb, N.; Bastamy, M.; Amer, M. SEPARATION AND PURIFICATION OF CHICKEN IgY WITH ITS FIELD EFFICACY IN CONTROLLING AVIAN INFLUENZA IN MUSCOVY DUCKS. 2023. [CrossRef]
- Indhuprakash, S.T.; S, P.; C, D.R.; Thirumalai, D. Efficacy Evaluation of Anti-DEC-IgY against Antibiotic-Resistant Diarrhoeagenic Escherichia Coli. J. Med. Microbiol. 2024, 73, 001801. [Google Scholar] [CrossRef]
- O’Donnell, K.L.; Espinosa, D.A.; Puerta-Guardo, H.; Biering, S.B.; Warnes, C.M.; Schiltz, J.; Nilles, M.L.; Li, J.; Harris, E.; Bradley, D.S. Avian Anti-NS1 IgY Antibodies Neutralize Dengue Virus Infection and Protect against Lethal Dengue Virus Challenge. Antiviral Res. 2020, 183, 104923. [Google Scholar] [CrossRef]
- Tong, H.-F.; Lin, D.-Q.; Pan, Y.; Yao, S.-J. A New Purification Process for Goose Immunoglobulin IgY(ΔFc) with Hydrophobic Charge-Induction Chromatography. Biochem. Eng. J. 2011, 56, 205–211. [Google Scholar] [CrossRef]
- Łupicka-Słowik, A.; Walczak, M.; Grzywa, R.; Bobrek, K.; Łęcka, M.; Boivin, S.; Gaweł, A.; Stefaniak, T.; Oleksyszyn, J.; Sieńczyk, M. Generation and Application of Polyclonal Igy Antibodies Specific for Full-Length and Nicked Prostate-Specific Antigen. Bioanalysis 2014, 6, 3197–3213. [Google Scholar] [CrossRef]
- Xia, W.; Lu, H.; Li, Y.; Cao, J.; Zhou, X.; Zhang, X.; Xia, X.; Sun, H. Purification of Chicken IgY by Binding Capture Using Elastin-like Polypeptide-Tagged Immunoglobulin-Binding Domain of Streptococcal Protein G. Vet. Immunol. Immunopathol. 2017, 192, 13–19. [Google Scholar] [CrossRef]
- Vicente, F.A.; Castro, L.S.; Mondal, D.; Coutinho, J.A.P.; Tavares, A.P.M.; Ventura, S.P.M.; Freire, M.G. Purification of Immunoglobulin Y from Egg Yolk Using Thermoresponsive Aqueous Micellar Two-Phase Systems Comprising Ionic Liquids. Sep. Purif. Technol. 2022, 288, 120589. [Google Scholar] [CrossRef]
- Al-Qaoud, K.M.; Obeidat, Y.M.; Al-Omari, T.; Okour, M.; Al-Omari, M.M.; Ahmad, M.I.; Alshadfan, R.; Rawashdeh, A.M. The Development of an Electrochemical Immunosensor Utilizing Chicken IgY Anti-Spike Antibody for the Detection of SARS-CoV-2. Sci. Rep. 2024, 14, 748. [Google Scholar] [CrossRef]
- Sabahi, S.; Mortazavi, S.A.; Nassiri, M.; Ghazvini, K.; Shahidi, F.; Abbasi, A. Production of Functional Ice Cream Fortified by Immunoglobulin Y against Escherichia Coli O157:H7 and Helicobacter Pylori. BIOINTERFACE Res. Appl. Chem. 2023, 13, 188. [Google Scholar] [CrossRef]
- Eriksson, M.; Nylén, S.; Grönvik, K.-O. Passive Immunization of Mice with IgY Anti-H5N1 Protects against Experimental Influenza Virus Infection and Allows Development of Protective Immunity. Vaccine 2024. [Google Scholar] [CrossRef]
- Pauly, D.; Chacana, P.A.; Calzado, E.G.; Brembs, B.; Schade, R. IgY Technology: Extraction of Chicken Antibodies from Egg Yolk by Polyethylene Glycol (PEG) Precipitation. J. Vis. Exp. JoVE 2011, 3084. [Google Scholar] [CrossRef]
- Shimelis, E. Extraction, Purification and Invitro Challenging of IgY of Commercial Chicken Eggs with the Newcastle Disease. Biomed. J. Sci. Tech. Res. 2022, 45. [Google Scholar] [CrossRef]
- Goldring, J.P.D.; Coetzer, T.H.T. Isolation of Chicken Immunoglobulins (IgY) from Egg Yolk. BAMBEd 2003, 31, 185–187. [Google Scholar] [CrossRef]
- Hodek, P.; Trefil, P.; Simunek, J.; Hudecek, J.; Stiborova, M. Optimized Protocol of Chicken Antibody (IgY) Purification Providing Electrophoretically Homogenous Preparations. Int. J. Electrochem. Sci. 2013, 8, 113–124. [Google Scholar] [CrossRef]
- Zhang, L.; Fan, Z.; Zhang, W.; Huang, H.; Wang, J.; Hei, A.; Cong, Y.; Xu, J. Preparation and characterization of chicken anti-human B7-H4 IgY polyclonal antibody. Chin. J. Cell. Mol. Immunol. 2024, 40, 257–266. [Google Scholar]
- El-Kafrawy, S.A.; Abbas, A.T.; Oelkrug, C.; Tahoon, M.; Ezzat, S.; Zumla, A.; Azhar, E.I. IgY Antibodies: The Promising Potential to Overcome Antibiotic Resistance. Front. Immunol. 2023, 14. [Google Scholar] [CrossRef]
- Hakalehto, E. Chicken IgY Antibodies Provide Mucosal Barrier against SARS-CoV-2 Virus and Other Pathogens. Isr. Med. Assoc. J. 2021, 23, 208–211. [Google Scholar]
- Frumkin, L.R.; Lucas, M.; Scribner, C.L.; Ortega-Heinly, N.; Rogers, J.; Yin, G.; Hallam, T.J.; Yam, A.; Bedard, K.; Begley, R.; et al. Egg-Derived Anti-SARS-CoV-2 Immunoglobulin Y (IgY) With Broad Variant Activity as Intranasal Prophylaxis Against COVID-19. Front. Immunol. 2022, 13, 899617. [Google Scholar] [CrossRef]
- Otterbeck, A.; Skorup, P.; Hanslin, K.; Larsson, A.; Stålberg, J.; Hjelmqvist, H.; Lipcsey, M. Intravenous Anti-P. Aeruginosa IgY-Antibodies Do Not Decrease Pulmonary Bacterial Concentrations in a Porcine Model of Ventilator-Associated Pneumonia. Innate Immun. 2022, 28, 224–234. [Google Scholar] [CrossRef]
- Rahman, S.; Nguyen, S.V.; Jr, F.C.I.; Umeda, K.; Kodama, Y. Oral Passive IgY-Based Immunotherapeutics. Hum. Vaccines Immunother. 2013, 9, 1039–1048. [Google Scholar] [CrossRef]
- Kollberg, H.; Carlander, D.; Olesen, H.; Wejåker, P.-E.; Johannesson, M.; Larsson, A. Oral Administration of Specific Yolk Antibodies (IgY) May Prevent Pseudomonas Aeruginosa Infections in Patients with Cystic Fibrosis: A Phase I Feasibility Study. Pediatr. Pulmonol. 2003, 35, 433–440. [Google Scholar] [CrossRef]
- Yokoyama, H.; Peralta, R.C.; Sendo, S.; Ikemori, Y.; Kodama, Y. Detection of Passage and Absorption of Chicken Egg Yolk Immunoglobulins in the Gastrointestinal Tract of Pigs by Use of Enzyme-Linked Immunosorbent Assay and Fluorescent Antibody Testing. Am. J. Vet. Res. 1993, 54, 867–872. [Google Scholar] [CrossRef]
- Guo, R.; Wu, S.; Guan, L.; Xie, Y.; Yang, X.; Li, Z.; Zhang, Z. Dietary Multivalent Anti-Helicobacter Pylori Immunoglobulin Y Significantly Increase the H. Pylori Eradication and Improve the Clinical Symptoms in Patients. Helicobacter 2021, 26, e12843. [Google Scholar] [CrossRef]
- Panigrahi, A.; Pattanaik, S.; Mohammad, N.; Sahoo, S.N.; Patnaik, S.; Sarangi, D. Effect of Passive Immunotherapeutics (IgY) on S. Mutans in Caries Active Children 8-11 Years of Age. J. Pharm. Bioallied Sci. 2024, 16, S4448. [Google Scholar] [CrossRef]
- Eriksson, M.; Nylén, S.; Grönvik, K.-O. Passive Immunization of Mice with IgY Anti-H5N1 Protects against Experimental Influenza Virus Infection and Allows Development of Protective Immunity. Vaccine 2024, S0264-410X(24)00796-5. [CrossRef]
- Shen, H.; Cai, Y.; Zhang, H.; Wu, J.; Ye, L.; Yang, P.; Lin, X.; Jiang, S.; Liao, M. Anti-SARS-CoV-2 IgY Isolated from Egg Yolks of Hens Immunized with Inactivated SARS-CoV-2 for Immunoprophylaxis of COVID-19. Virol. Sin. 2021, 36, 1080–1082. [Google Scholar] [CrossRef]
- Wongso, H.; Mahendra, I.; Arnafia, W.; Idar, I.; Yusuf, M.; Achmad, A.; Holik, H.A.; Kurniawan, A.; Halimah, I.; Sriyani, M.E.; et al. Preclinical Evaluation of Chicken Egg Yolk Antibody (IgY) Anti-RBD Spike SARS-CoV-2—A Candidate for Passive Immunization against COVID-19. Vaccines 2022, 10, 128. [Google Scholar] [CrossRef]
- Agurto-Arteaga, A.; Poma-Acevedo, A.; Rios-Matos, D.; Choque-Guevara, R.; Montesinos-Millán, R.; Montalván, Á.; Isasi-Rivas, G.; Cauna-Orocollo, Y.; Cauti-Mendoza, M. de G.; Pérez-Martínez, N.; et al. Preclinical Assessment of IgY Antibodies Against Recombinant SARS-CoV-2 RBD Protein for Prophylaxis and Post-Infection Treatment of COVID-19. Front. Immunol. 2022, 13, 881604. [Google Scholar] [CrossRef] [PubMed]
- Madai, M.; Hanna, D.; Hetényi, R.; Földes, F.; Lanszki, Z.; Zana, B.; Somogyi, B.; Papp, H.; Kuczmog, A.; Faragó-Sipos, O.; et al. Evaluating the Protective Role of Intranasally Administered Avian-Derived IgY Against SARS-CoV-2 in Syrian Hamster Models. Vaccines 2024, 12, 1422. [Google Scholar] [CrossRef] [PubMed]
- Karamzadeh-Dehaghani, A.; Towhidi, A.; Zhandi, M.; Mojgani, N.; Fouladi-Nashta, A. Combined Effect of Probiotics and Specific Immunoglobulin Y Directed against Escherichia Coli on Growth Performance, Diarrhea Incidence, and Immune System in Calves. Animal 2021, 15, 100124. [Google Scholar] [CrossRef]
- Qandoos P27 | The Efficacy of Maternally Delivered Immunoglobulin (Y) against Some Gut Pathogens in Chickens. J. Vet. Pharmacol. Ther. 2023, 46, 98–98. [CrossRef]
- Soltani, N.; Rahimi, S.; Khaki, P.; Karimi Torshizi, M.A.; Eskandari, B.; Grimes, J. Efficacy of Hyperimmunized Egg Yolk Antibodies (IgY) against Campylobacter Jejuni: In Vitro and In Vivo Evaluations. Poult. Sci. 2025, 104, 104718. [Google Scholar] [CrossRef]
- Ul Abadeen, Z.; Javed, M.T.; Rizvi, F.; Rahman, S.U. Salutary Effects of Anti-Clostridium Perfringens Type A Egg Yolk Antibodies (IgY) on Growth Performance and Hemato-Biochemical Parameters in Experimentally Infected Broiler Chicken. Pak. Vet. J. 2021, 41, 562–566. [Google Scholar] [CrossRef]
- Matulka, R.A.; Thompson, L.; Corley, D. Multi-Level Safety Studies of Anti Fel d 1 IgY Ingredient in Cat Food. Front. Vet. Sci. 2020, 6, 477. [Google Scholar] [CrossRef]
- Kota, R.K.; Reddy, P.N.; Sreerama, K. Application of IgY Antibodies against Staphylococcal Protein A (SpA) of Staphylococcus Aureus for Detection and Prophylactic Functions. Appl. Microbiol. Biotechnol. 2020, 104, 9387–9398. [Google Scholar] [CrossRef]
- Vansofla, A.N.; Nazarian, S.; Kordbache, E.; Fathi, J. An IgG/IgY Sandwich-ELISA for the Detection of Heat-Labile Enterotoxin B Subunit of Enterotoxigenic Escherichia Coli. Gene Rep. 2021, 23, 101099. [Google Scholar] [CrossRef]
- Bayat, M.; Khabiri, A.; Hemati, B. Development of IgY-Based Sandwich ELISA as a Robust Tool for Rapid Detection and Discrimination of Toxigenic Vibrio Cholerae. Can. J. Infect. Dis. Med. Microbiol. 2018, 2018, 4032531. [Google Scholar] [CrossRef]
- Ge, S.; Wu, R.; Zhou, T.; Liu, X.; Zhu, J.; Zhang, X. Specific Anti-SARS-CoV-2 S1 IgY-scFv Is a Promising Tool for Recognition of the Virus. AMB Express 2022, 12, 18. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Xiao, H.; Lin, M.; Zhou, J.; Xuan, Q.; Cui, X.; Zhao, S. Preparation and Evaluation of IgY against Human Papillomavirus. J. Virol. Methods 2025, 334, 115115. [Google Scholar] [CrossRef] [PubMed]
- Lopes, C.A.; Faria, L.S. de; Sousa, J.E.N. de; Borges, I.P.; Ribeiro, R.P.; Bueno, L.L.; Ávila, V.M.R.; Júnior, Á.F.; Costa-Cruz, J.M. Anti-Ascaris Suum Immunoglobulin Y as a Novel Biotechnological Tool for the Diagnosis of Human Ascariasis. J. Helminthol. 2020, 94, e71. [Google Scholar] [CrossRef]
- Cakir-Koc, R.; Budama-Kilinc, Y.; Ustun, E.; Babur, C. Conjugation and Characterization of Latex Particles with Toxoplasma Gondii-Specific Immunoglobulin Y Antibodies for Diagnostic Aim and Evaluation Efficiency in In Vitro Culture. J. Equine Vet. Sci. 2020, 92, 103145. [Google Scholar] [CrossRef]
- Cakir-Koc, R. Production of Anti-SAG1 IgY Antibody against Toxoplasma Gondii Parasites and Evaluation of Antibody Activity by ELISA Method. Parasitol. Res. 2016, 115, 2947–2952. [Google Scholar] [CrossRef]
- Teimoori, S.; Arimatsu, Y.; Laha, T.; Kaewkes, S.; Sereerak, P.; Sripa, M.; Tangkawattana, S.; Brindley, P.J.; Sripa, B. Chicken IgY-Based Coproantigen Capture ELISA for Diagnosis of Human Opisthorchiasis. Parasitol. Int. 2017, 66, 443–447. [Google Scholar] [CrossRef]
- Prasertsincharoen, N.; Metheenukul, P. Production of Immunoglobulin Y against Dog Erythrocyte Antigen (DEA) 1.1 from Egg Yolk. Thai J. Vet. Med. 2020, 50, 381–387. [Google Scholar] [CrossRef]
- Syahruni, S.; Hartati, Y.W.; Yusuf, M.; Kusumawardani, S.; Wibawan, I.W.T.; Arnafia, W.; Sibit, G.; Subroto, T. Development of Lateral Flow Assay Based on Anti-IBDV IgY for the Rapid Detection of Gumboro Disease in Poultry. J. Virol. Methods 2021, 291, 114065. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, D.C.; Eto, S.F.; Claudiano, G. da S.; Marcusso, P.F.; Ramos-Espinoza, F.C.; Engracia de Moraes, J.R. Production and Use of Immunoglobulin Y in the Diagnosis of Haemorrhagic Septicemia Caused by Aeromonas Hydrophila in Piaractus Mesopotamicus. Aquac. Res. 2022, 53, 2930–2936. [Google Scholar] [CrossRef]
- Alves, C.F.; Notário, A.F.O.; Correia, L.I.V.; Ferreira, K.N.; Araujo, K.C.L.; Taketomi, E.A.; Souza, G.R.L.; Medeiros, E.S.; Cunha-Júnior, J.P.; Alves, R.P.; et al. A Novel Electrochemical Immunosensor Based on Anti-IgY and PLA/PEG Nanofibrous Mats Used for Allergens Detection in Farinaceous Products. J. Electrochem. Soc. 2023, 170, 037509. [Google Scholar] [CrossRef]
- Tu, Y.-Y.; Ma, C.-Y.; Ho, S.-B.; Chen, C.-C.; Chang, H.-M. Afffinity Measurement of Lactoferrin (LF)-Anti-LF Immunoglobulin in Yolk (IgY) Complexes by Competitive Indirect Enzyme-Linked Immunosorbent Assay (CI-ELISA). J. Food Drug Anal. 2020, 14. [Google Scholar] [CrossRef]
- Zhang, Q.; He, D.; Xu, L.; Ge, S.; Wang, J.; Zhang, X. Generation and Evaluation of Anti-Mouse IgG IgY as Secondary Antibody. Prep. Biochem. Biotechnol. 2020, 50, 788–793. [Google Scholar] [CrossRef]
- Abbas, A.T.; El-Kafrawy, S.A.; Sohrab, S.S.; Tabll, A.A.; Hassan, A.M.; Iwata-Yoshikawa, N.; Nagata, N.; Azhar, E.I. Anti-S1 MERS-COV IgY Specific Antibodies Decreases Lung Inflammation and Viral Antigen Positive Cells in the Human Transgenic Mouse Model. Vaccines 2020, 8, 634. [Google Scholar] [CrossRef]
- Bentes, G.A.; Lanzarini, N.M.; Guimarães, J.R.; Heinemann, M.B.; Volotão, E. de M.; da Silva, A. dos S.; Heneine, L.G.D.; de Oliveira, J.M.; Pinto, M.A. Production and Evaluation of Chicken Egg Yolk Immunoglobulin (IgY) against Human and Simian Rotaviruses. Viruses 2022, 14, 1995. [Google Scholar] [CrossRef] [PubMed]
- Bentes, G.A.; Guimarães, J.R.; Volotão, E. de M.; Lanzarini, N.M.; da Silva, A. dos S.; Gardinali, N.R.; Marchevsky, R.S.; Leite, J.P.G.; de Oliveira, J.M.; Pinto, M.A. Passive Immunotherapy of Cynomolgus Monkeys with Anti-Rotavirus IgY. Pharmaceutics 2024, 16, 1149. [Google Scholar] [CrossRef]
- Chen, W.; Xiao, H.; Lin, M.; Zhou, J.; Xuan, Q.; Cui, X.; Zhao, S. Preparation and Evaluation of IgY against Human Papillomavirus. J. Virol. Methods 2025, 334, 115115. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wei, Y.; Li, Y.; Wang, X.; Liu, Y.; Tian, D.; Jia, X.; Gong, R.; Liu, W.; Yang, L. IgY Antibodies against Ebola Virus Possess Post-Exposure Protection in a Murine Pseudovirus Challenge Model and Excellent Thermostability. PLoS Negl. Trop. Dis. 2021, 15, e0008403. [Google Scholar] [CrossRef]
- Hao, N.; Liu, B.; Zhao, M.; Lu, M.; Chen, F.; Kang, J.; Tang, X.; Zhang, Y.; Dang, C. Real-World Evidence of a Novel Tetravalent Immunoglobulin Y Effectiveness and Safety in Patients with the Refractory Helicobacter Pylori Infection. BMC Infect. Dis. 2024, 24, 647. [Google Scholar] [CrossRef]
- Han, S.; Wen, Y.; Yang, F.; He, P. Chicken Egg Yolk Antibody (IgY) Protects Mice Against Enterotoxigenic Escherichia Coli Infection Through Improving Intestinal Health and Immune Response. Front. Cell. Infect. Microbiol. 2021, 11, 662710. [Google Scholar] [CrossRef]
- Capotă, R.; Iliescu, A.C.B.; Năstasă, V.; Minea, B.; Sliwa, D.C.; Mareș, M. Hyperimmune Egg-Based IgY-Rich Formulations as Adjuvant Therapy in a Murine Model of Urinary Tract Infection. J. Glob. Antimicrob. Resist. 2024, 39, 48. [Google Scholar] [CrossRef]
- Ahmadi, T.S.; Mousavi Gargari, S.L.; Talei, D. Anti-Flagellin IgY Antibodies Protect against Pseudomonas Aeruginosa Infection in Both Acute Pneumonia and Burn Wound Murine Models in a Non-Type-Specific Mode. Mol. Immunol. 2021, 136, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Norouzi, F.; Behrouz, B.; Ranjbar, M.; Mousavi Gargari, S.L. Immunotherapy with IgY Antibodies toward Outer Membrane Protein F Protects Burned Mice against Pseudomonas Aeruginosa Infection. J. Immunol. Res. 2020, 2020, 7840631. [Google Scholar] [CrossRef]
- Sanches, R.F.; dos Santos Ferraro, A.C.N.; Marroni, F.E.C.; Venancio, E.J. Synergistic Activity between Beta-Lactams and Igy Antibodies against Pseudomonas Aeruginosa in Vitro. Mol. Immunol. 2022, 148, 1–5. [Google Scholar] [CrossRef]
- Thomsen, K.; Christophersen, L.; Lerche, C.J.; Holmgaard, D.B.; Calum, H.; Høiby, N.; Moser, C. Azithromycin Potentiates Avian IgY Effect against Pseudomonas Aeruginosa in a Murine Pulmonary Infection Model. Int. J. Antimicrob. Agents 2021, 57, 106213. [Google Scholar] [CrossRef] [PubMed]
- Wannigama, D.L.; Hurst, C.; Monk, P.N.; Abe, S.; Chatsuwan, T.; Stick, S.M.; Kicic, A. Novel Inhale Treatment of IgY Immunoglobulin Combination with Macrocyclic Peptide Eliminate Multidrug-Resistant Pseudomonas Aeruginosa Pulmonary Infections. RESPIROLOGY 2023, 28, 6–6. [Google Scholar]
- Zamani, K.; Irajian, G.; Zahedi Bialvaei, A.; Zahraei Salehi, T.; Khormali, M.; Vosough, A.; Masjedian Jazi, F. Passive Immunization with Anti- Chimeric Protein PilQ/PilA –DSL Region IgY Does Not Protect against Mortality Associated with Pseudomonas Aeruginosa Sepsis in a Rabbit Model. Mol. Immunol. 2022, 141, 258–264. [Google Scholar] [CrossRef]
- Zafarmand-Samarin, M.; Nazarian, S.; Aghaie, S.M.; Sadeghi, D.; Samiei-Abianeh, H.; Felegary, A. Therapeutic Efficacy of SipD/LptD-Specific IgY Entrapped in Alginate Nanoparticles against Salmonella Typhimurium Infection. Heliyon 2024, 10, e39650. [Google Scholar] [CrossRef]
- Cabré, N.; Hartmann, P.; Llorente, C.; Kouno, T.; Wang, Y.; Zeng, S.; Kim, H.Y.; Zhang, X.; Kisseleva, T.; Iyer, S.; et al. Immunoglobulin Y Antibodies against Cytolysin Reduce Ethanol-Induced Liver Disease in Mice. Hepatol. Baltim. Md 2023, 78, 295–306. [Google Scholar] [CrossRef]
- Mary, D.J.; Mathew, M.G. Effect of Streptococcal IgY on Quantity of Streptococcus Mutans in High Caries Risk Children. J. Pharm. Res. Int. 2020, 6–11. [Google Scholar] [CrossRef]
- Li, X.; He, P.; Yu, L.; He, Q.; Jia, C.; Yang, H.; Lu, M.; Wei, X.; Zhao, S. Production and Characteristics of a Novel Chicken Egg Yolk Antibody (IgY) against Periodontitis-Associated Pathogens. J. Oral Microbiol. 2020, 12, 1831374. [Google Scholar] [CrossRef]
- Sindhu, B.U.; Praveen, S.; Sandeep, J.N.; Avinash, J.L.; Rajiv, N.P.; Rudraswamy, S. Evaluation of Antimicrobial Efficacy of Immunoglobulin Y (IgY) against Periodontal Biofilm. J. Oral Maxillofac. Pathol. JOMFP 2023, 27, 499–506. [Google Scholar] [CrossRef]
- Yan, Y.; Guan, Y.; Luo, L.; Lu, B.; Chen, F.; Jiang, B. Effects of Immunoglobulin Y-Loaded Amorphous Calcium Phosphate on Dentinal Tubules Occlusion and Antibacterial Activity. Front. Bioeng. Biotechnol. 2022, 10, 921336. [Google Scholar] [CrossRef] [PubMed]
- Mares, M.; Iliescu, A.C.B.; Minea, B.; Capota, R.; Sliwa, D.C.; Nastasa, V. Hyperimmune Egg-Based IgY-Rich Formulations as Adjuvant Therapy in a Murine Model of Vaginal Candidiasis. J. Glob. Antimicrob. Resist. 2024, 39, 63. [Google Scholar] [CrossRef]
- Jin, Y.; Lv, H.; Wang, M.; Cho, C.-S.; Shin, J.; Cui, L.; Yan, C. Effect of Microencapsulation of Egg Yolk Immunoglobulin Y by Sodium Alginate/Chitosan/Sodium Alginate on the Growth Performance, Serum Parameters, and Intestinal Health of Broiler Chickens. Anim. Biosci. 2023, 36, 1241–1251. [Google Scholar] [CrossRef]
- Radwan, F.M.; EL-Shemy, A.; Bosila, M.M.; Bastamy, M.A.; Amer, M.M. Evaluation of The Efficacy of Purified Egg Yolk Immunoglobulin (IgY) in Preventing and Controlling Newcastle Disease Virus Infection in Broiler Chickens. Egypt. J. Vet. Sci. 2024, 55, 931–943. [Google Scholar] [CrossRef]
- Yang, D.; Mai, K.; Zhou, Q.; Zhu, Y.; Xing, J.; Luo, C.; Liu, S.; Zhou, Q.; Huang, W.; Luo, J.; et al. The Protective Efficacy of Specific Egg Yolk Immunoglobulin Y(IgY) against Riemerella Anatipestifer Infections. Vet. Microbiol. 2020, 243, 108642. [Google Scholar] [CrossRef] [PubMed]
- Ge, S.; Xu, L.; Li, B.; Zhong, F.; Liu, X.; Zhang, X. Canine Parvovirus Is Diagnosed and Neutralized by Chicken IgY-scFv Generated against the Virus Capsid Protein. Vet. Res. 2020, 51, 110. [Google Scholar] [CrossRef]
- Bousquet, J.; Gherasim, A.; de Blay, F.; Mathieu-Dupas, E.; Batot, G.; Laune, D.; Sousa-Pinto, B.; Zuberbier, T.; Pham-Thi, N.; Group, M. study Proof-of-Concept Study of Anti-Fel d 1 IgY Antibodies in Cat Food Using the MASK-Air® App. Clin. Transl. Allergy 2024, 14, e12353. [Google Scholar] [CrossRef]
- Bustos, C.P.; Leiva, C.L.; Gambarotta, M.; Guida, N.; Chacana, P.A. In Vitro Inhibitory Activity of IgY Antibodies Against Salmonella Ser. Newport Isolated from Horses. J. Equine Vet. Sci. 2021, 103, 103657. [Google Scholar] [CrossRef]
- Czoska, P.; Tarsalewska, K.; Ponichtera, M.; Rybicka, M.; Sowa-Rogozinska, N.; Sominka-Pierzchlewicz, H.; Stodolna, A.; Ogonowska, P.; Kosciuk, A.; Glosnicka, R.; et al. Growth-Inhibitory Effect of Chicken Egg Yolk Polyclonal Antibodies (IgY) on Zoonotic Pathogens Campylobacter Jejuni, Salmonella Spp. and Escherichia Coli, In Vitro. Int. J. Mol. Sci. 2025, 26, 1040. [Google Scholar] [CrossRef]
- Liu, J.; Qin, Y.; Yan, L.; Liu, W.; Shi, H.; Lu, Y.; Liu, X. Protective Effects of Egg Yolk Immunoglobulins (IgY) on Juvenile Groupers (Epinephelus Fuscoguttatus × Epinephelus Lanceolatus) with Red-Spotted Grouper Nervous Necrosis Virus Infection. Aquaculture 2021, 545, 737218. [Google Scholar] [CrossRef]
- Liang, Z.; Ning, Y.; Cao, J.; Liu, S.; Liang, X.; Peng, X.; Huang, Y.; Wei, J.; Xiao, S.; Qin, Q.; et al. The Protective Effect of Specific Yolk Antibody against Nervous Necrosis Virus Infection in Mandarin Fish(Siniperca Chuatsi). Fish Shellfish Immunol. 2024, 155, 109996. [Google Scholar] [CrossRef] [PubMed]
- Setthawong, P.; Yamkasem, J.; Khemthong, M.; Tattiyapong, P.; Metheenukul, P.; Prasertsincharoen, N.; Lertwanakarn, T.; Thengchaisri, N.; Surachetpong, W. Development of IgY-Based Passive Immunization Against Tilapia Lake Virus: Development and In Vitro Neutralization Assays. Viruses 2025, 17, 448. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, R.; Pedrosa-Gerasmio, I.R.; Alenton, R.R.R.; Nozaki, R.; Kondo, H.; Hirono, I. Anti-PirA-like Toxin Immunoglobulin (IgY) in Feeds Passively Immunizes Shrimp against Acute Hepatopancreatic Necrosis Disease. J. Fish Dis. 2019, 42, 1125–1132. [Google Scholar] [CrossRef]
- James, T.; Rahiman KM ,Mujeeb; Sebastian ,Denoj; S ,Suja Rani; and MD, D. The Combinational Effect of Avian IgY Antibodies and Broad-Spectrum Antibiotics (Ciprofloxacin and Chloramphenicol) in Vitro Reduces the Incidence of Aeromonas Hydrophila. J. Appl. Aquac. 2025, 1–16. [CrossRef]
- Xu, L.; Che, J.; Xu, Y.; Chen, Y.; Li, Y.; Murtaza, B.; Wang, L.; Zhang, M.; Li, X. Oral Administration of Microencapsulated Egg Yolk Immunoglobulin (IgY) in Turbot (Scophthalmus Maximus) to Combat against Edwardsiella Tarda 2CDM001 Infections. Fish Shellfish Immunol. 2020, 106, 609–620. [Google Scholar] [CrossRef]
- Soto-Dávila, M.; Rodríguez-Cornejo, T.; Benito, V.W.; Rodríguez-Ramos, T.; Mahoney, G.; Supinski, R.; Heath, G.; Dang, X.; Valle, F.M.; Hurtado, C.; et al. Innate and Adaptive Immune Response of Rainbow Trout (Oncorhynchus Mykiss) Naturally Infected with Yersinia Ruckeri. Fish Shellfish Immunol. 2024, 151, 109742. [Google Scholar] [CrossRef] [PubMed]
- Gopal, G.; Antonysamy ,Michael; Visaga Ambi ,Senthil; and Thirumalai, D. Development of Chicken Egg Yolk Antibodies (IgY) against Venom of Cobra (Naja Naja) and Krait (Bungarus Caeruleus) and a Study on Its Neutralization Potential. Toxin Rev. 2024, 43, 484–504. [CrossRef]
- Nguyen, V.T.; Vu, T.T.H.; Hoang, V.T.; Nguyen, T.V.; Lam, T.D.; Nguyen, K.C.; Than, V.T.; Le, V.P. Immunological Properties of the Chicken Egg Yolk (IgY) Antibodies against Vietnamese Cobra Naja Naja Venom. IOP Conf. Ser. Mater. Sci. Eng. 2020, 991, 012012. [Google Scholar] [CrossRef]
- Duan, H.; He, Q.; Zhou, B.; Wang, W.; Li, B.; Zhang, Y.; Deng, Q.; Zhang, Y.; Yu, X. Anti-Trimeresurus Albolabris Venom IgY Antibodies: Preparation, Purification and Neutralization Efficacy. J. Venom. Anim. Toxins Trop. Dis. 2016, 22, 23. [Google Scholar] [CrossRef]
- Leiva, C.L.; Cangelosi, A.; Mariconda, V.; Farace, M.; Geoghegan, P.; Brero, L.; Fernández-Miyakawa, M.; Chacana, P. IgY-Based Antivenom against Bothrops Alternatus: Production and Neutralization Efficacy. Toxicon 2019, 163, 84–92. [Google Scholar] [CrossRef]
- Alvarez, A.; Montero, Y.; Jiménez, E.; Zerpa, N.; Parrilla, P.; Malavé, C. IgY Antibodies Anti-Tityus Caripitensis Venom: Purification and Neutralization Efficacy. Toxicon Off. J. Int. Soc. Toxinology 2013, 74. [Google Scholar] [CrossRef] [PubMed]
- Leiva, C.L.; Geoghegan, P.; Lammer, M.; Cangelosi, A.; Mariconda, V.; Celi, A.B.; Brero, M.L.; Chacana, P. In Vivo Neutralization of Bee Venom Lethality by IgY Antibodies. Mol. Immunol. 2021, 135, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, S.V.; Nguyen, M.T.H.; Tran, B.C.; Ho, M.T.Q.; Umeda, K.; Rahman, S. Evaluation of Lozenges Containing Egg Yolk Antibody against Porphyromonas Gingivalis Gingipains as an Adjunct to Conventional Non-Surgical Therapy in Periodontitis Patients: A Randomized Controlled Clinical Trial. J. Periodontol. 2018, 89, 1334–1339. [Google Scholar] [CrossRef]
- Nilsson, E.; Larsson, A.; Olesen, H.V.; Wejåker, P.-E.; Kollberg, H. Good Effect of IgY against Pseudomonas Aeruginosa Infections in Cystic Fibrosis Patients. Pediatr. Pulmonol. 2008, 43, 892–899. [Google Scholar] [CrossRef]
- Immunoglobulin IgY Pseudomonas A Clinical Trial for Cystic Fibrosis Treatment | FP7. Available online: https://cordis.europa.eu/project/id/261095/reporting (accessed on 24 April 2025).
- Liu, S.T.H.; Mirceta, M.; Lin, G.; Anderson, D.M.; Broomes, T.; Jen, A.; Abid, A.; Reich, D.; Hall, C.; Aberg, J.A. Safety, Tolerability, and Pharmacokinetics of Anti-SARS-CoV-2 Immunoglobulin Intravenous (Human) Investigational Product (COVID-HIGIV) in Healthy Adults: A Randomized, Controlled, Double-Blinded, Phase 1 Study. Antimicrob. Agents Chemother. 2023, 67, e01514-22. [Google Scholar] [CrossRef]
- Kovacs-Nolan, J.; Mine, Y. Egg Yolk Antibodies for Passive Immunity. Annu. Rev. Food Sci. Technol. 2012, 3, 163–182. [Google Scholar] [CrossRef]
- Yamaki, K.; Ohta, K.; Kobayashi, N.; Morita, I.; Kiguchi, Y.; Oyama, H.; Ito, K.; Nanbo, A.; Oh-oka, H.; Koyama, Y.; et al. Purification of Emu IgY for Therapeutic and Diagnostic Use Based on Monoclonal Secondary Antibodies Specific to Emu IgY. Biol. Pharm. Bull. 2022, 45, 1022–1026. [Google Scholar] [CrossRef] [PubMed]
- Esmaeili, Z.; Shahsavar, S.K.; Keikha, M.; Ghazvini, K. Chicken Immunoglobulin (IgY) as an Alternative Treatment for Bacterial Infections, Emphasizing Advantages, Disadvantages and Mechanisms. J. Microbiol. Infect. Dis. 2024, 14, 95–95. [Google Scholar] [CrossRef]
- Nastasa, V.; Minea, B.; Pasca, A.-S.; Bostanaru-Iliescu, A.-C.; Stefan, A.-E.; Gologan, D.; Capota, R.; Foia, L.-G.; Mares, M. Long-Term Oral Administration of Hyperimmune Egg-Based IgY-Rich Formulations Induces Mucosal Immune Response and Systemic Increases of Cytokines Involved in Th2- and Th17-Type Immune Responses in C57BL/6 Mice. Int. J. Mol. Sci. 2024, 25, 8701. [Google Scholar] [CrossRef]
Figure 1.
Structural comparison between immunoglobulin G (IgG) and immunoglobulin Y (IgY). Fab – fragment antigen-binding; Fc – fragment crystallizable; Fv – variable fragment; VH – variable region of the heavy chain; VL – variable region of the light chain; CL – constant region of the light chain; CH – constant region of the heavy chain.
Figure 1.
Structural comparison between immunoglobulin G (IgG) and immunoglobulin Y (IgY). Fab – fragment antigen-binding; Fc – fragment crystallizable; Fv – variable fragment; VH – variable region of the heavy chain; VL – variable region of the light chain; CL – constant region of the light chain; CH – constant region of the heavy chain.
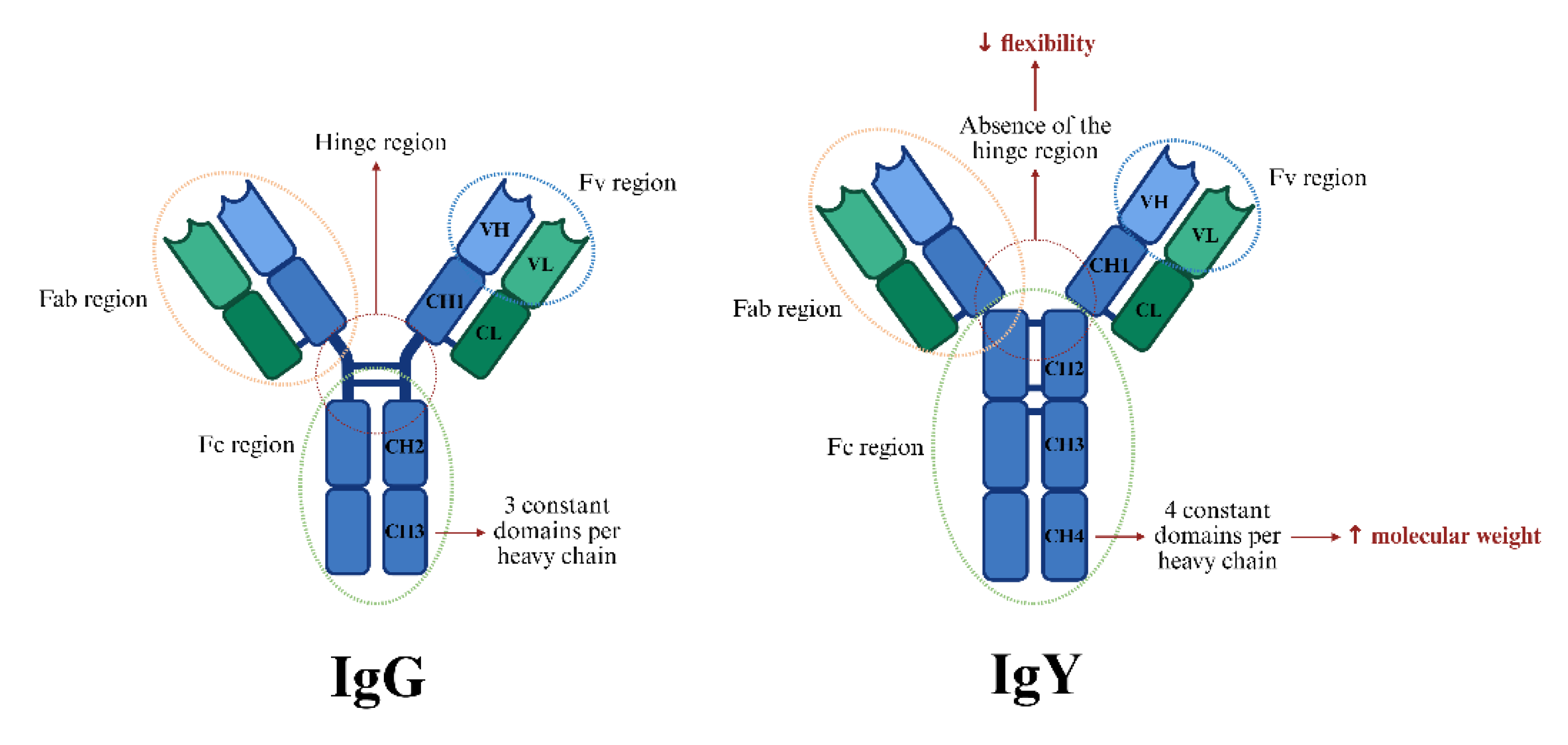
Figure 2.
Schematic representation of immunoglobulin Y production technology.
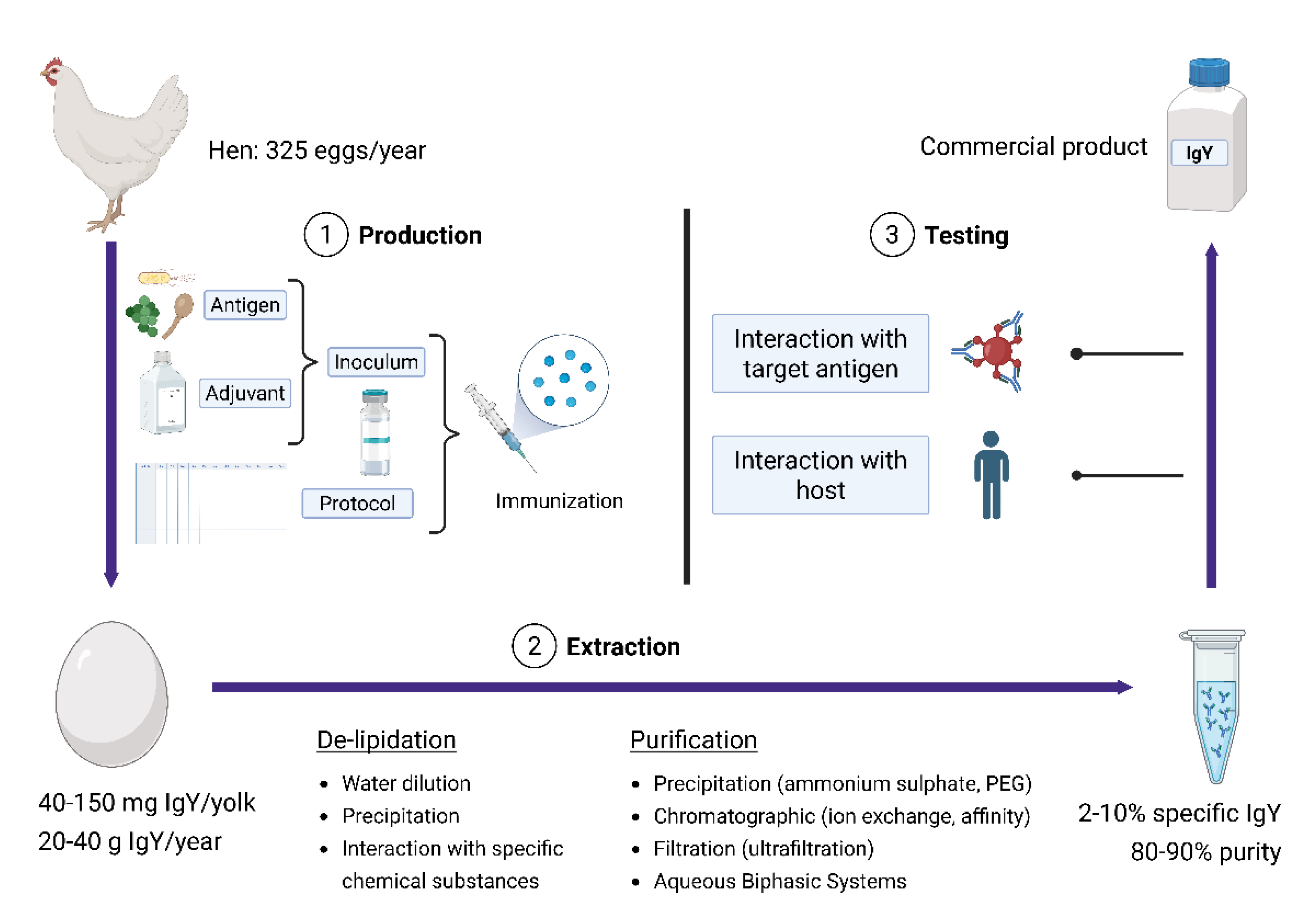

Table 1.
ECVAM Recommendation relating to chicken immunisation protocols [6].
Table 1.
ECVAM Recommendation relating to chicken immunisation protocols [6].
| Factor | Recommendation |
|---|---|
| Adjuvant | Freund’s incomplete adjuvant, Specol, lipopeptide (Pam₃-Cys-Ser-[Lys]₄; 250 µg) |
| Antigen dose | 10 ng–1 mg (preferably 10–100 µg) |
| Injection site | Intramuscular (field studies; young laboratory chickens) Subcutaneous (older laboratory chickens) |
| Injection volume | < 1 ml |
| Injection frequency | 2–3 times; boosters during laying period |
| Vaccination interval | 4–8 weeks |
| Use of chickens | Entire laying period (about 1 year) |
Table 2.
Clinical trials (ongoing and complete) investigating the use of IgY in human health.
| Study overview | Condition/Goal | Intervention/ Treatment | Study completion |
|---|---|---|---|
| Evaluation of IgY Antibody Effectiveness in Supportive Therapy of Periodontitis Patients [171] Study Id Number: NCT02705885 |
Chronic Periodontitis | IgY antibodies against Porphyromonas gingivalis gingipains as dietary supplement | 2015-03 |
| Phase I and II: Post Marketing Study of Anti-pseudomonas IgY in Prevention of Recurrence of Pseudomonas Aeruginosa Infections Infections in Cystic Fibrosis (CF) Patients [172] Study Id Number: NCT00633191 |
Cystic fibrosis. Infection with Pseudomonas aeruginosa |
Anti-pseudomonas IgY gargle | 2012-12 |
| Phase III Study to Evaluate Clinical Efficacy and Safety of Avian Polyclonal Anti-Pseudomonas Antibodies (IgY) in Prevention of Recurrence of Pseudomonas Aeruginosa Infection in Cystic Fibrosis Patients [173] Study Id Number: NCT01455675 |
Cystic fibrosis | Avian polyclonal anti-pseudomonas antibodies (IgY) | 2017-06 |
| Phase I: Assessing the Safety of an IGY Supplement on the Gut Microbiome Study Id Number: NCT06702280 |
Healthy | Dietary Supplement: IgY supplement | Estimated 2025-10 |
| Phase I and II: A Randomized, Double-blind, Placebo-controlled, Parallel Dose Ranging Study on the Influence of IgY Max on Inflammatory Markers and the Gut Microbiome Study Id Number: NCT02972463 |
Healthy | Immunoglobulin Y | 2018-01 |
| A Phase 1 Study in Healthy Participants to Evaluate the Safety, Tolerability, and Pharmacokinetics of Single -Ascending and Multiple Doses of an Anti-Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Chicken Egg Antibody (IgY) [174] Study Id Number: NCT04567810 |
COVID19 | anti-SARS-CoV-2 IgY | 2020-12 |
| Evaluation of a Health Food Supplement Containing Anti-Helicobacter Pylori Urease IgY Antibody on Patients with Chronic Gastritis in Hanoi, Vietnam Study Id Number: NCT02721355 |
Chronic Gastritis Caused by Helicobacter Pylori | GastimunHP | 2016-06 |
| Randomized, Double-blind, Placebo-controlled Exploratory Trial to Investigate Efficacy and Safety of IGN-ES001 in Patients with Chronic Widespread Pain with or Without Fibromyalgia Study Id Number: NCT03058224 |
Chronic Widespread Pain Fibromyalgia |
Polyclonal avian immunoglobulin IgY containing specific IgY against E. coli F18ab and S. typhimurium in partially delipidated avian egg yolk powder | 2017-12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



