Submitted:
08 May 2025
Posted:
09 May 2025
You are already at the latest version
Abstract
This study evaluated the histopathological impact of three commonly used pesticides—pirimiphos-methyl, propamocarb hydrochloride, and 2,4-dichlorophenoxyacetic acid (2,4-D) on the kidneys of common carp (Cyprinus carpio Linnaeus, 1758) after 96-hour acute exposure. The histopathological analysis demonstrated that all three tested pesticides induced structural changes. The histopathological changes were assessed using a semi-quantitative scoring system and categorised into circulatory, degenerative, proliferative, and inflammatory alterations. While circulatory alterations were absent in all treatments, clear and statistically significant degenerative, proliferative, and inflammatory responses were recorded, which escalated with increasing the pesticide concentrations. In addition, different statistical analyses were performed to interpret the lesions in the kidney structure and function. Prior to the statistical analysis, normality and variance homogeneity were assessed using the Shapiro–Wilk and Levene’s tests, respectively. Due to non-normal data distribution, non-parametric methods were applied. Hence, the non-parametric statistical methods showed distinct group-level differences in the kidney damage indices. The Kruskal–Wallis test revealed significant differences across treatments (p < 0.001), and Mann–Whitney U tests identified specific pairwise differences. The degenerative and proliferative lesions were most prominent in fish exposed to 2,4-D at 100 µg/L (IK = 34), followed by pirimiphos-methyl and propamocarb hydrochloride. Inflammatory changes were mainly observed in the pirimiphos-methyl groups. The histopathological lesions were concentration-dependent, with 2,4-D causing irreversible renal damage at higher concentrations. These findings highlight the nephrotoxic risks posed by common pesticides and validate that the use of histopathological indices, combined with robust non-parametric testing, provides a reliable approach to evaluating organ-specific pesticide toxicity. These biomarkers offer sensitive early warning indicators of environmental risk and reinforce the suitability of common carp as a model species for ecotoxicological assessment.
Keywords:
common carp
; kidney
; histological biomarkers
; pesticides
; pollution
1. Introduction
Pesticides are chemicals that prevent, destroy, repel, or mitigate pests. They can also be used as plant regulators, defoliants, or desiccants [1]. The global use of pesticides, associated with a risk of exposure to all organisms and the environment, has increased manifold in recent years [2,3]. Therefore, the application of pesticides is recognised as the largest intentional input of biologically active substances into terrestrial ecosystems [4]. Over the past few decades, the intensification of international trade has corresponded with a notable 50% increase in pesticide application from 1990 to 2022. The increase in pesticide use has reflected the environmental and health implications, especially in low-income regions where pesticide application has soared due to agricultural growth. [5]. Thus, today the modern agriculture is highly dependent on the use of pesticides, primarily for crop protection and yield enhancement [6,7,8,9]. The benefit of their application is undeniable, but on the other hand, there is clear evidence that the widespread importation of pesticide substances causes irreversible damage to ecosystems and their inhabitants, including humans [7,10,11].
Fish are important bioindicators for pollution in aquatic ecosystems, as they allow the use of biological approaches in environmental biomonitoring. The changes that occur in their bodies as a result of pollution can be used as reliable biomarkers in determining the degree and type of pollution in aquatic ecosystems. In ecotoxicological studies, biomarkers are useful tools in monitoring the health of aquatic ecosystems. Incorporating complex biomarkers at different levels of biological organisation is a suitable approach to detect pollutant-induced responses [12]. Biomarkers are also important assessment tools because they provide specific information about the biological effects of a particular pollutant. They can be used for monitoring purposes and to clarify the causal relationship between the risk to aquatic health and the toxicant concentration. In this regard, morphological methods are applied in ecotoxicological research because they provide the opportunity to assess the pollutant effects on specific target organs and tissues. In addition, morphological changes in cellular and tissue structure are important parameters in determining the potential toxicity of pollutants. By studying the extent of each morphological change, the sensitivity of an organism to the level of toxicity to which it is exposed can be also determined.
The kidneys of fish play a key role in maintaining body fluid homeostasis, as well as in the excretion of toxic xenobiotic residues. Studies related to pathological disorders in kidney tissues can also be considered an important parameter that indirectly reveals the intensity of environmental pollution [13]. Therefore, fish kidneys can be successfully applied in ecotoxicological studies, however, they are less commonly used compared to fish gills and liver. Their function and the established changes in the histological structure under the influence of various toxicants are associated with disruptions in homeostasis [14]. On this basis, conducting research related to morphological changes can provide information about the state of the studied organ as a result of a certain environmental pollutant’s action, which can affect the health of the whole organism [15].
The present laboratory study aims to assess for the first time the histopathological changes in the kidney of common carp (Cyprinus carpio Linnaeus, 1758) after 96-hour exposure to three pesticides (pirimiphos-methyl, propamocarb hydrochloride, and 2,4-dichlorophenoxyacetic acid) and their experimental concentrations. To our knowledge, this is the very first study on the toxicological effects of pirimiphos-methyl and propamocarb hydrochloride on fish renal health.
2. Materials and Methods
2.1. Experimental fish
Common carp is one of the most preferred species for industrial breeding, compared to other freshwater fish species. It is widely distributed in almost all countries of the world and it is an important fish for human consumption [16,17]. Common carp has a high commercial value worldwide, but it is also used as a bioindicator for water pollution [18] because it is a demersal fish, with feeding habits that expose it to many different types of environmental pollutants. In addition, it is easily caught and maintained under laboratory conditions [19].
2.2. Experimental pesticides
Pirimiphos-methyl is an organophosphorus insecticide that is often used for the prevention and control of insects during the storage of agricultural production [20]. However, residues in the crop resulting from its excessive application pose a health hazard to humans and animals [21]. It is widely used in major grain-producing countries to protect against insect attack, as this organophosphate insecticide has a long-lasting activity [22]. Pirimiphos-methyl is also widely used against populations of the malaria mosquito Anopheles spp. (Meigen 1818), especially in Africa [23,24]. The mechanisms of resistance to pirimiphos-methyl are poorly understood, but organophosphates, similarly to carbamates, block the action of the acetylcholinesterase (AChE) enzyme by competitively binding to its active site [25]. Therefore, pirimiphos-methyl is responsible for the phosphorylation of AChE, which regulates the hydrolysis of acetylcholine in the synaptic cleft of the insect nervous system [26,27,28]. Just like AChE, the circulating enzyme butyrylcholinesterase (BchE), also known as pseudocholinesterase, is also a target for phosphorylation and inhibition by this insecticide [29].
Propamocarb hydrochloride was first introduced into the European markets to control pathogens in ornamental crops and some vegetables in 1978 [30]. The fungicide has good protective and curative activity against downy mildew without phytotoxic effects in fruits and vegetables, such as tomatoes, potatoes, and cucumbers [31,32,33,34,35]. It belongs to the carbamate pesticide family and limits fungal growth by inhibiting the phosphoric acid and fatty acid synthesis pathways [36,37]. This pesticide is classified as slightly toxic by oral, dermal, and eye acute contact and practically nontoxic by inhalation [38]. Propamocarb hydrochloride has systemic activity after absorption through the leaves, stems, and roots and by transport through the vascular system in treated plants [39].
2,4-Dichlorophenoxyacetic acid (2,4-D) is a synthetic auxin herbicide that, due to its effectiveness, selectivity, low cost, and broad spectrum in pest control, has become one of the most frequently used herbicides in agricultural and urban areas worldwide [40]. It is commonly used to control dicotyledonous weeds in cereal crops, such as maize (Zea mays L.) and wheat (Triticum aestivum L.). Despite its agricultural use for over eight decades, the development of resistance to 2,4-D has been slow, with 25 weeds globally having developed resistance to 2,4-D as of 2021 [41]. In 2012, 2,4-D was the most widely used herbicide in home and garden areas, roughly equal to glyphosate used in combined non-agricultural areas[42]. Furthermore, 2,4-D is registered as an ingredient in approximately 1500 agricultural and domestic pesticides, either as the sole active ingredient or in combination with others [43,44]. However, even at low concentrations, 2,4-D has herbicidal effects on dicotyledons due to its ability to induce uncoordinated cell growth [45], damage to vascular tissues and roots, and malformation of leaves and stems [46]. Studies have shown the ability of this herbicide to bioaccumulate in non-target organisms exposed for a short period of time [47,48]. Due to its high water solubility, acidic characteristics and mobility, 2,4-D has become a serious problem for soil and groundwater contamination [49,50], which means that it also has a serious environmental and health risk on non-target organisms [51,52].
2.3. Experimental set-up
Juvenile fish (n=105, 6 months old) were used in the present experiment. They were purchased from the Institute of Fisheries and Aquaculture in Plovdiv, Bulgaria, where they are bred and raised under strictly controlled conditions. The experimental individuals were of the same size and age group (Table 1), without external pathological changes. For the experiment, 15 individuals were used for each experimental concentration, as well as in the control group.
Acclimatisation was carried out in static conditions for a week. A total of 7 aquariums with a capacity of 100 L were used in the experiment; six of them were filled with dechlorinated water and treated with different concentrations of the pesticides and the seventh - was used for control with no added chemicals. Two concentrations of each pesticide were prepared (pirimiphos-methyl – 10 μg/L and 60 μg/L; propamocarb hydrochloride – 40 μg/L and 80 μg/L; 2,4-D – 50 μg/L and 100 μg/L, Table 2). All aquariums were equipped with aerator pumps to supply the necessary oxygen levels. The fish were fed daily with pelleted dry food (Aller Aqua, Poland) at 3–5% body weight. Water parameters, such as t (oC), pH, dissolved oxygen (mg/L), and electrical conductivity (μS/cm) in all aquaria were measured 3 times per day (Table 3) until the experiment stopped after 96 hours [53]. The experiments were repeated twice.
2.4. Histopathological assessment
The fish were dissected according to the procedure given by Rosseland et al. (2003) [54], complying with the requirements for the human treatment of experimental animals according to Directive 2010/63/EU [55]. The histpathological processing of the materials was carried out according to the standard methodology of Romeis (1989) [56], which includes fixation of the sample in 10% neutral buffered formalin, tissue processing (alcohol dehydration, xylene clearing, paraffin wax infiltration), embedding, microtome sectioning and hematoxylin & eosin (H&E) staining. The histopathological changes in the kidney were assessed according to the Bernet et al. (1999) [57] scale. The alterations were classified into four groups – circulatory, proliferative, degenerative, and inflammation-related changes, with each group including specific changes affecting functional units of the organ or the entire organ. In addition, a 5-point scale was used to determine the severity of each change according to Saraiva et al. (2015) [58], as follows: (0) – no changes in the kidney structure (up to 10% of the organ structure); (1) – very mild changes in the kidney structure (from 10% to 20% of the organ structure); (2) – mild degree of changes in the kidney structure (from 20% to 30% of the organ structure); (3) – moderate degree of changes in kidney structure (from 30% to 50% of the organ structure); (4) – severe degree of changes in the kidney structure (from 50% to 80% of organ structure); (5) – very severe degree of changes in the kidney structure (over 80% of the organ structure). The pathological degree of each change was determined using the significance factor (W) according to Bernet et al. (1999) [57] - W is a constant value and is categorised according to the author as follows: (1) – minimal pathological significance, reversible change after cessation of the toxicant; (2) – low pathological significance, the lesion is reversible in most cases if the stressor is neutralized; (3) – high pathological significance, irreversible changes, leading to partial or complete loss of organ function. The indices of histopathological changes are the result of the product of the degree of each specific change in the group multiplied by W. The summation of these final values for the organ determines the index (I) for the respective group of changes (circulatory IC, degenerative IR, proliferative IP, and inflammatory II). The sum of all indices also determines the organ index (IO) for pathological change - IO refers to a class, according to Zimmerli et al. (2007) [59], as follows: class I (Index ≤ 10) – normal histological structure with mild pathological changes (reversible); class II (Index 11–20) – normal histological structure with moderate pathological changes (reversible); class III (Index 21–30) – moderate degree of change in the histological structure (reversible); class IV (Index 31–40) – severe degree of change in the histological structure (irreversible); class V (Index > 40) – very severe degree of change in the histological structure (irreversible).
2.5. Statistical analysis
Before statistical testing, the data were assessed for normality using the Shapiro–Wilk test and for homogeneity of variances using Levene’s test. As assumptions for parametric tests were not met, non-parametric methods were applied. The Kruskal–Wallis H test was used to evaluate the overall differences in the histopathological index values (Ik) among the treatment groups [60]. When significant differences were detected, pairwise comparisons were conducted using the Mann–Whitney U test [61]. A significance threshold of p < 0.05 was used, and the pairwise differences were adjusted for multiple testing where appropriate. A bar plot was generated displaying the final index for organ (Ik) values per treatment to visualise the group differences. Different lowercase letters were used to denote the statistically distinct groups based on the post hoc tests. Furthermore, a Principal Component Analysis (PCA) was performed on the kidney histopathological scores to evaluate the separation of treatment groups based on the histopathological alterations. The dataset comprised 22 histopathological parameters assessed across six experimental groups (n = 10 per group), including six pesticide-treated groups. The control group was excluded from the PCA, as all histopathological scores for the control specimens were zero, resulting in no variance and thus no contribution to the analysis. The experimental groups were as follows: (Pir-10), exposed to 10 µg/L pirimiphos-methyl; (Pir-60), exposed to 60 µg/L pirimiphos-methyl; (Pro-40), exposed to 40 µg/L propamocarb hydrochloride; (Pro-80), exposed to 80 µg/L propamocarb hydrochloride; and (2,4-D 50) and (2,4-D 100), exposed to 50 and 100 µg/L 2,4-dichlorophenoxyacetic acid, respectively. The following histopathological variables were used in the PCA: Kidney-level changes included haemorrhage (KHem), hyperaemia (KHyp), and aneurysms (KAne); Tubular degenerative changes consisted of vacuolar degeneration (TVacDeg), hyaline degeneration (THyalDeg), necrobiosis (TNecBio), and necrosis (TNec); Glomerular lesions included dilatation of Bowman’s capsule (GDilBow), contraction (GCont), necrobiosis (GNecBio), and necrosis (GNec); Interstitial tissue changes involved necrosis (INec); Proliferative responses were assessed through hypertrophy and hyperplasia in tubules (TProlHyp and TProlHpl), glomeruli (GProlHyp and GProlHpl), and thickening of Bowman’s capsular membrane (GBowThick); Interstitial tissue proliferation was evaluated through hypertrophy (IProlHyp) and edema (IEdema); Inflammatory reactions in the kidney were described by infiltration (KInfl) and activation of melano-macrophages (KActMM). All variables were standardised before analysis using z-score transformation to ensure comparability. PCA was carried out using the scikit-learn library [62], and the first two principal components (PCA1 and PCA2) were visualized in a two-dimensional scatter plot. Each treatment group was represented with a unique color and symbol. Confidence ellipses (95%) were added to highlight the distribution of individuals within each treatment group. Vectors representing histopathological parameters were overlaid to illustrate their contribution to the separation of treatments, except for variables located at or near the origin, which were excluded from the labeling to avoid visual clutter. The final figure was generated in Python using matplotlib [63] and seaborn for visual aesthetics. All statistical analyses were conducted using Python 3.10 with the scipy.stats and seaborn libraries.
3. Results and Discussion
The results showed normal morphology of the histological structure of the kidney in the control fish group. According to the five-point (0–5) scale for the severity of changes, the histopathological sections of the kidney were assigned a grade of 0, although in some individuals, the histopathological changes were found to occupy less than 10% of the section surface. The normal histological structure of a common carp kidney is shown in Table 4 and Figure 1.
The teleostean kidney consists of the anterior kidney (also known as head kidney) and posterior kidney (also known as body or trunk kidney). Embryologically, the anterior kidney derives from the pronephros, and the posterior kidney from mesonephros [64]. The anterior kidney is integrated into the endocrine system of the fish and is very important for the stress response, mediated by the hypothalamus–pituitary–interrenal cell (HPI) axis and by the hypothalamus–sympathetic nervous system–chromaffin tissue (HSC) axis [65,66]. In addition, the head of the kidney contains endocrine elements - the chromaffin cells and interrenal tissue, which are located around the blood vessels. The posterior kidney contains the nephrons with variable quantities of hemopoietic and lymphoid tissue in the interstitium. It is mainly composed of renal corpuscles that are made up of the Bowman’s capsules, glomeruli, renal tubules and collecting ducts. In vertebrates, three types of kidney are found: the pronephros, mesonephros and metanephros [67,68]. Therefore, the healthy kidney is involved in the maintenance of body fluid homeostasis, it produces urine, which acts as an excretory route for the metabolites of a variety of xenobiotics to, which the fish may be exposed, it also excretes other nitrogen-containing waste products from metabolism, such as ammonia and creatinine and last but not least, the kidney performs an important function related to electrolyte and water balance and the maintenance of a stable internal environment [69].
The histopathological sections of the pirimiphos-methyl treated fish kidney showed no significant changes in the circulatory system, associated with haemorrhage, hyperemia and aneurysms. Therefore, the index of these changes (IKC) at both tested concentrations was 0, respectively (Table 4).
Degenerative changes were observed in the renal parenchyma, which were localised in the renal tubules, glomeruli and interstitial hematopoietic tissue. Considering the degree of involvement of the disorders, a general trend of increasing severity with the increasing concentration of pirimiphos-methyl applied was observed. In the cytoplasm of single epithelial cells of the covering epithelium, which structures the wall of the renal tubules, vacuolar degeneration with a mild degree of manifestation was detected, similarly at both tested concentrations (Figure 2 A, B). Along with this, a more severe degenerative change was also observed in the epithelial cells, namely hyaline-droplet degeneration. In single cells, processes of necrobiosis are also observed, associated with the presence of changes in the nucleus - karyopyknosis, karyorrhexis and karyolysis. Necrosis was also reported in the epithelial cells of the renal tubules. The hyaline-droplet degeneration, necrobiosis and necrosis were detected, but only at the higher applied concentration, and the degree of manifestation was determined as mild.
In the area of the Bowman's capsule in the glomerulus, a well-pronounced dilatation was observed, which, with the involvement of the organ, was determined to a mild degree at a concentration of 10 μg/L and a moderate degree at a concentration of 60 μg/L pirimiphos-methyl (Table 4, Figure 2 C, D). Shrinkage of the renal corpuscles was observed to a very mild degree, similar at both concentrations. Necrobiosis and necrosis of the glomerulus, as well as necrosis of the interstitial tissue, were not observed at any of the concentrations used. The degenerative change index (IKR) at a concentration of 10 μg/L was 5, and at a concentration of 60 μg/L it was 12.
Proliferative changes were present in the renal tubules, glomeruli and interstitial tissue. Mild renal tubule hypertrophy was found at both tested concentrations, while tubular hyperplasia was not observed. Hypertrophy and hyperplasia of the glomerulus were also not detected. A thickening of the membrane of the Bowman's capsule in the glomerulus was observed only at the higher concentration to a very mild degree. Among the changes in the interstitial tissue, moderate hypertrophy and very mild edema were observed at both concentrations tested (Figure 2 E). The proliferative change index (IKP) at a concentration of 10 μg/L is 7, and at a concentration of 60 μg/L it is 9.
Regarding the changes related to inflammation, lymphocyte infiltration was not observed. Melanomacrophage activation was found to be moderate at both tested concentrations (Figure 2 A). The index of these changes (IKI) was 6 for both concentrations.
After calculating the indices of all changes, it was found that the kidney index (IK) of fish treated with pirimiphos-methyl at a concentration of 10 μg/L was 18, and in those treated with a concentration of 60 μg/L, it was 27 (Table 6). Based on the results obtained and the scale proposed by Zimmerli et al. (2007) [59], the IK at a concentration of 10 μg/L pirimiphos-methyl falls into class II (index 11-20), which means that the organ has a normal histological structure with moderate pathological changes that are reversible. Moreover, the IK at a concentration of 60 μg/L pirimiphos-methyl falls into class III (index 21-30), which indicates that the organ has a moderate degree of change in the histological structure that is still reversible.
The histopathological examination of kidney sections from common carp exposed to pirimiphos-methyl for 96 hours revealed no significant circulatory system alterations, such as haemorrhage, hyperemia, or aneurysms, at either of the tested concentrations (10 μg/L and 60 μg/L). Accordingly, the Index for Circulatory Changes (IKC) remained 0 across all groups. This was confirmed by statistical analysis, where the Kruskal–Wallis test showed no significant differences between treatment groups (Kruskal–Wallis test, Hc = 0, p = 1.000) (Table 4).
In contrast, degenerative changes were observed in the renal parenchyma, affecting the renal tubules, glomeruli, and interstitial hematopoietic tissue. The index for degenerative changes (IKR) showed a concentration-dependent increase (control: 0, 10 μg/L: 5, 60 μg/L: 12). Alterations included vacuolar degeneration (mild, at both concentrations), hyaline-droplet degeneration, necrobiosis (including karyopyknosis, karyorrhexis, and karyolysis), and necrosis, all observed at the higher concentration. These changes were visible in the cytoplasm and nuclei of single epithelial cells lining the renal tubules (Figure 2A, B). The statistical analysis showed significant differences between groups (Kruskal–Wallis test, Hc = 73.80, p < 0.001), and post hoc comparisons confirmed significant pairwise differences across all groups (Mann–Whitney U test, p < 0.001) (Table 4).
Proliferative changes were also detected, including tubular and interstitial hypertrophy and mild edema. The index for proliferative changes (IKP) increased from 0 (control) to 7 (10 μg/L) and 9 (60 μg/L). These changes were statistically significant (Kruskal–Wallis test, Hc = 47.69, p < 0.001), with significant differences between the control group and both exposure groups (Mann–Whitney U test, p < 0.001), while the difference between the two exposed groups was not statistically significant (Mann–Whitney U test, p = 0.1335) (Table 4).
Regarding inflammatory processes, the primary finding was the activation of melanomacrophages in both exposed groups. The index for inflammatory changes (IKI) increased from 0 in the control group to 3 in the exposed groups. The Kruskal–Wallis test indicated a significant difference among groups (Kruskal–Wallis test, Hc = 12.53, p < 0.01), with post hoc tests showing significant differences between the control and both treatment groups (Mann–Whitney U test, p < 0.01), but no significant difference between the two concentrations (Mann–Whitney U test, p = 0.5776) (Table 4).
The total organ index (IK) increased progressively with concentration: 0 (control), 18 (10 μg/L), and 27 (60 μg/L). Statistical analysis confirmed this trend with a highly significant difference between groups (Kruskal–Wallis test, Hc = 125.4, p < 0.001), and all pairwise comparisons were statistically significant (Mann–Whitney U test, p < 0.001) (Table 4).
The kidney of fish treated with propamocarb hydrochloride revealed changes in the circulatory system of the organ in single areas in only one of the individuals. Since the changes do not cover the necessary commitment of the authority to be included in the scale of degree of changes (0-5), this change was assigned a degree of 0 - no changes. Therefore, the index of these changes (IKC) was also 0 (Table 5).
Vacuolar degeneration in the cytoplasm of the renal tubular epithelial cells was detected to a very mild extent at the lower concentration, while at the higher concentration it was determined to be moderately pronounced (Figure 3 A, B, D, E, F). Hyaline-droplet degeneration in the epithelial cells of the renal tubules was observed to a very mild extent at a concentration of 40 μg/L and to a mild extent at a concentration of 80 μg/L. Necrobiosis and necrosis of the renal tubules were not detected.
Dilatation of the Bowman's capsule in the glomerulus was found and was expressed to a mild extent at both tested concentrations (Figure 3 D, F). Shrinkage of the renal corpuscles, necrobiosis and necrosis in the glomerulus, as well as necrosis in the interstitial hematopoietic tissue, were not observed at both tested concentrations of the pesticide. The index of degenerative changes (IKR) in the kidney of treated fish after exposure to propamocarb hydrochloride at the lower concentration was 4, while at the higher tested concentration, the value was 7.
Regarding the proliferative changes in the renal tubules, hypertrophy was observed (Figure 3 C), involving epithelial cells to a very mild degree, similar at both concentrations, while hyperplasia was not detected at any of the concentrations tested. Of the proliferative changes in the glomerulus, hypertrophy was reported to be to a very mild degree, similar at both concentrations. Also, at both experimental concentrations, hypertrophy was observed in the glomerulus with a very mild degree of manifestation, while hyperplasia was absent in the analysed histological sections. Thickening of the membrane of the Bowman's capsule in the glomerulus was recorded only at the higher concentration of the applied pesticide, with a very mild degree of manifestation. Proliferative changes in the interstitial tissue were not detected at any of the fungicide concentrations. The calculated index of proliferative changes (IKP) in the kidney of fish treated with a concentration of 40 μg/L was 2, while in those treated with 80 μg/L fungicide, the value was 4.
Of the inflammatory changes in the kidney, lymphocyte infiltration was not observed at either fungicide concentrations tested. Melanomacrophage activation was detected to a very slight extent at a concentration of 40 μg/L, and a slight extent at a concentration of 80 μg/L (Figure 3 E). The calculated inflammatory index (IKI) for the lower concentration was 2, while for the higher concentration, the value was 4. The total kidney index (IK) of fish treated with propamocarb hydrochloride at a concentration of 40 μg/L was 8, and at a concentration of 80 μg/L it was 15 (Table 7).
From the results obtained and according to the scale proposed by Zimmerli et al. (2007) [59], it was calculated that IK at the lower concentration falls into class I (index ≤10), which means that the organ has a normal histological structure with mild pathological changes that are reversible. At the higher applied concentration, IK falls into class II (index 11-20), which indicates that the kidney of the treated fish has a normal histological structure and the presence of moderate pathological changes, which are also reversible. The results indicate that the changes in the kidney increase their degree of manifestation in direct proportion to the increase in the applied concentrations of propamocarb hydrochloride.
The kidney of common carp exposed to propamocarb hydrochloride for 96 hours showed no significant changes in the circulatory system (Kruskal–Wallis test, Hc = 0, p = 1.000). Therefore, all samples received a score of 0 for the circulatory system alterations, and the index for circulatory changes (IKC) was 0 in all groups (Table 5).
In contrast, degenerative changes were observed in the renal parenchyma, particularly within the tubular epithelium. Vacuolar degeneration was noted to a very mild extent (score 1) at 40 μg/L, and increased to a moderate degree (score 3) at 80 μg/L. Similarly, hyaline-droplet degeneration was scored as very mild (1) and mild (2) at the lower and higher concentrations, respectively. Necrobiosis and necrosis of the renal tubules were not observed at either concentration. In the glomeruli, dilatation of Bowman’s capsule was found at a mild level (score 2) in both exposed groups (Figure 3A, B, D, E, F). The index for degenerative changes (IKR) increased accordingly: 0 (control), 4 (40 μg/L), and 7 (80 μg/L). The statistical analysis showed a significant difference between the treatment groups (Kruskal–Wallis test, Hc = 34.04, p < 0.001). The pairwise comparisons confirmed significant differences between all group pairs (Mann–Whitney U test, p < 0.05) (Table 5).
Proliferative changes in the kidney were also evident. Tubular hypertrophy was observed at both concentrations with a very mild degree of expression (score 1), while hyperplasia was absent. In the glomeruli, hypertrophy was also scored as very mild at both exposure levels. Thickening of Bowman’s capsule membrane was noted only at the higher concentration, with a very mild degree of change. No proliferative changes were found in the interstitial tissue (Figure 3C). The index for proliferative changes (IKP) increased from 0 (control) to 2 (40 μg/L) and 4 (80 μg/L). These differences were statistically significant (Kruskal–Wallis test, Hc = 29.83, p < 0.001), with pairwise comparisons confirming significant differences between all groups (Mann–Whitney U test, p < 0.05) (Table 5).
Regarding inflammatory processes, lymphocyte infiltration was not observed. However, activation of melanomacrophages was recorded at both concentrations - very mild (score 1) at 40 μg/L and mild (score 2) at 80 μg/L (Figure 3E). The index for inflammatory changes (IKI) was 0 (control), 2 (40 μg/L), and 4 (80 μg/L). The Kruskal–Wallis test revealed a significant difference between groups (Kruskal–Wallis test, Hc = 10.81, p < 0.01), with post hoc comparisons showing that the treated groups were significantly different from the control (Mann–Whitney U test, p < 0.01), but not from each other (p = 0.3783) (Table 5).
The overall organ index (IK), reflecting total kidney damage, increased progressively with concentration: 0 (control), 8 (40 μg/L), and 15 (80 μg/L). This increase was statistically significant (Kruskal–Wallis test, Hc = 74.5, p < 0.001), and all pairwise comparisons revealed significant differences (Mann–Whitney U test, p < 0.05) (Table 5).
Based on the classification of Zimmerli et al. (2007) [59], the total organ index (IK) of 8 in fish exposed to 40 μg/L of propamocarb hydrochloride places them in Class I (IK ≤ 10), indicating a normal histological structure with mild, reversible pathological changes. At 80 μg/L, an IK of 15 corresponds to Class II (IK 11–20), suggesting a normal histological structure with moderate pathological changes that remain reversible (Table 5).
After a short-term treatment with the herbicide 2,4-D, changes in the circulatory system of the kidney, similarly to the other two pesticides, were not detected. Therefore, the index of these changes (IKC) at both applied concentrations was 0 (Table 8).

Table 6.
Histopathological changes in the kidney of common carp after acute (96-hour) exposure to 2,4-D.
Table 6.
Histopathological changes in the kidney of common carp after acute (96-hour) exposure to 2,4-D.
| Reaction pattern | Орган | Alteration |
Importance factor |
Score value | ||
|---|---|---|---|---|---|---|
|
Control |
2, 4 - D | |||||
| 50 μg/L | 100 μg/L | |||||
| Changes in the circulatory system | Kidney | Haemorrhage | WKC1 = 1 | 0A | 0A | 0A |
| Hyperaemia | WKC2 = 1 | 0A | 0A | 0A | ||
| Aneurysms | WKC3 = 1 | 0A | 0A | 0A | ||
| Index for changes in the circulatory system | IKC =0A | IKC =0A | IKC =0A | |||
| Degenerative changes | Tubule | Vacuolar degeneration | WKR1 = 1 | 0A | 2B | 4C |
| Hyaline degeneration | WKR2 = 1 | 0A | 0A | 0A | ||
| Necrobiosis | WKR3 = 2 | 0A | 0A | 0A | ||
| Necrosis | WKR4 = 3 | 0A | 0A | 0A | ||
| Glomerulus | Dilatation of the Bowman's capsule | WKR5 = 1 | 0A | 1B | 2C | |
| Contraction | WKR6 = 1 | 0A | 0A | 1B | ||
| Necrobiosis* | WKR7 = 2 | 0A | 1B | 1B | ||
| Necrosis | WKR8 = 3 | 0A | 1B | 1B | ||
|
Interstitial tissue |
Necrosis | WKR9 = 3 | 0A | 0A | 1B | |
| Index forthedegenerative changes | IKR = 0A | IKR=5B | IKR=15C | |||
| Proliferative changes | Tubule | Hypertrophy | WKP1 = 1 | 0A | 0A | 1B |
| Hyperplasia | WKP2 = 2 | 0A | 0A | 0A | ||
| Glomerulus | Hypertrophy | WKP3 = 1 | 0A | 1B | 3C | |
| Hyperplasia | WKP4 = 2 | 0A | 0A | 1B | ||
| Thickening of Bowman's capsular membrane | WKP5 = 2 | 0A | 0A | 0A | ||
|
Interstitial tissue |
Hypertrophy | WKP6 = 1 | 0A | 1B | 1B | |
| Edema | WKP7 = 2 | 0A | 2B | 4C | ||
| Index fortheproliferative changes | IKP = 0A | IKP =6B | IKP=15C | |||
| Inflammation | Kidney | Infiltration | WKI1 = 2 | 0A | 0A | 0A |
| Activation of melano-macrophages | WKI2 = 2 | 0A | 2B | 2B | ||
| Index fortheinflammatory processes | IKI = 0A | IKI =4B | IKI =4B | |||
| Index for organ IK | IK = 0A | IK=15B | IK=34C | |||
In addition, a 5-point scale was used to determine the severity of each change according to Saraiva et al. (2015) [58], as follows: (0) – no changes in the kidney structure (up to 10% of the organ structure); (1) – very mild changes (10–20% of the organ structure); (2) – mild changes (20–30%); (3) – moderate changes (30–50%); (4) – severe changes (50–80%); (5) – very severe changes (over 80%). Different superscript letters (A, B, C) indicate statistically significant differences between groups according to the Mann–Whitney U test (p < 0.05). * Necrobiosis includes karyopyknosis, karyorrhexis, and karyolysis.
Degenerative changes were reported in the epithelial cells of the renal tubules, expressed in vacuolar degeneration localised in the cytoplasm of the cells, which was determined to be mild at a concentration of 50 μg/L and severe at a concentration of 100 μg/L of 2,4-D (Figure 4 A, D, E, F). Hyaline-droplet degeneration, necrobiosis and necrosis were not observed at any of the applied concentrations. Of the degenerative changes in the glomerulus, dilation of the Bowman's capsule was found to a very slight extent at the lower concentration and to a slight extent at the higher concentration of the applied pesticide (Figure 4 C). Renal corpuscle shrinkage was observed to a very mild extent only at the higher pesticide concentration used. Necrobiosis, including karyorrhexis, karyopyknosis and karyolysis, as well as glomerular necrosis, were observed to a very mild extent, similarly at both experimental concentrations (Figure 4 B, C). The index of degenerative changes (IKR) in the kidney for the lower applied fungicide concentration is 5, while at the higher concentration it has a value of 15.
From the proliferative changes in the renal tubule, hypertrophy was found at a concentration of 100 μg/L to a very mild degree, while hypertrophy at a concentration of 50 μg/L, as well as hyperplasia, at both concentrations, were absent. Glomerular hypertrophy was described to be very mild at the lower concentration and moderate at the higher concentration. Glomerular hyperplasia was very mild, only at a concentration of 100 μg/L. Thickening of the membrane of the Bowman's capsule in the glomerulus was not observed at any of the concentrations. Hypertrophy of the interstitial tissue was present to a very mild extent at both fungicide concentrations used. Edema in the interstitial hematopoietic tissue was found to be mild at a concentration of 50 μg/L, and severe at the higher concentration. The organ proliferative change index (IKP) at the lower concentration was 6, and at the higher concentration, 15.
Among the changes associated with inflammatory processes, activation of melanomacrophages was found, which was determined to be mild in both concentrations (Figure 4 F). At the same time, lymphocytic infiltration was not detected in any of the used concentrations of 2,4-D. When calculating the inflammatory process index (IKI), a value of 4 was found for both fungicide concentrations.
The kidney index (IK) of fish treated with 2,4-D, at a concentration of 50 μg/L was 15, while the index of those treated with a concentration of 100 μg/L was 34 (Table 8). Based on the results obtained and according to the scale proposed by Zimmerli et al. (2007) [59], it was calculated that the kidney index (IK) at the lower concentration falls into class II (index 11-20). This indicates that the organ has a normal histological structure with moderate pathological changes that are reversible. However, at the higher concentration, IK falls into class IV (index 31-40). This indicates a severe degree of change in the histological structure of the organ, and the processes are considered irreversible. The results indicate that the higher concentration of the applied fungicide 2,4-D causes significantly more serious pathological lesions in the histological structure of the fish kidney treated with this herbicide.
Following the 96-hour exposure to the herbicide 2,4-D, no histopathological changes were observed in the circulatory system of the kidney, consistent with previous results obtained for pirimiphos-methyl and propamocarb hydrochloride. Consequently, the index for circulatory changes (IKC) was 0 in all treatment groups, and no statistically significant differences were found (Kruskal–Wallis test, Hc = 0, p = 1.000) (Table 6).
In contrast, degenerative changes were detected, primarily affecting the tubular and glomerular compartments. Vacuolar degeneration in renal tubular epithelial cells was determined to be mild at 50 μg/L and severe at 100 μg/L (Figure 4A, D, E, F). Hyaline-droplet degeneration, necrobiosis, and necrosis of the tubules were not observed at either concentration. In the glomeruli, dilatation of Bowman’s capsule was found at very mild (score 1) and mild (score 2) levels for the 50 and 100 μg/L groups, respectively (Figure 4C). Additional changes, such as glomerular contraction, necrobiosis (including karyopyknosis, karyorrhexis, and karyolysis), and necrosis, were observed to a very mild extent at both concentrations (Figure 4B, C). The index for degenerative changes (IKR) increased significantly with concentration: 0 (control), 5 (50 μg/L), and 15 (100 μg/L). These differences were statistically significant (Kruskal–Wallis test, Hc = 79.8, p < 0.001), with pairwise comparisons confirming significant differences between all groups (Mann–Whitney U test, p < 0.001) (Table 6).
Proliferative changes were also observed. Tubular hypertrophy was absent at 50 μg/L but present at 100 μg/L with a very mild degree (score 1). Glomerular hypertrophy was noted as very mild at 50 μg/L and moderate at 100 μg/L, while glomerular hyperplasia was observed to a very mild extent only at the higher concentration. Thickening of Bowman’s capsule was not present. In the interstitial tissue, hypertrophy was observed at both concentrations, and edema was detected as mild at 50 μg/L and severe at 100 μg/L. The Index for proliferative changes (IKP) rose from 0 (control) to 6 (50 μg/L) and 15 (100 μg/L). These increases were statistically significant (Kruskal–Wallis test, Hc = 70.49, p < 0.001), with all pairwise comparisons between groups showing significant differences (Mann–Whitney U test, p < 0.001) (Table 6).
Regarding inflammatory processes, lymphocytic infiltration was not observed at any concentration. However, activation of melanomacrophages was found to a mild extent in both exposed groups (Figure 4F). The index for inflammatory changes (IKI) was 0 (control) and 4 (for both 50 and 100 μg/L groups). These differences were statistically significant (Kruskal–Wallis test, Hc = 12.04, p < 0.01), though no significant difference was detected between the two exposure groups (Mann–Whitney U test, p > 0.05) (Table 6).
The total organ index (IK) showed a strong dose-dependent increase: 0 (control), 15 (50 μg/L), and 34 (100 μg/L). These differences were highly significant (Kruskal–Wallis test, Hc = 152.7, p < 0.001), and all pairwise comparisons between groups were statistically significant (Mann–Whitney U test, p < 0.01) (Table 6).
Based on the classification system of Zimmerli et al. (2007) [59], the IK score of 15 at 50 μg/L places the kidneys into Class II (index 11–20), indicating moderate pathological changes that are generally reversible. In contrast, the IK score of 34 at 100 μg/L corresponds to Class IV (index 31–40), representing a severe and likely irreversible degree of histological alteration. These results clearly demonstrate that the herbicide 2,4-D induces significantly more serious renal damage at higher concentrations in common carp (Table 6, Figure 4).
To assess the cumulative impact of each tested pesticide on kidney histopathology, the index for organ (IK) was calculated as the sum of severity scores across all observed alterations in each individual. This composite index allowed directly comparing the overall kidney damage caused by different treatments.
The control group exhibited no histopathological changes, resulting in an IK value 0. Among the exposed groups, the lowest IK value was recorded in fish treated with propamocarb hydrochloride at a concentration of 40 μg/L (IK = 8), indicating a relatively low level of pathological damage. Pirimiphos-methyl at 10 μg/L and 2,4-D at 50 μg/L both resulted in moderate damage, with IK values of 18 and 15, respectively. Slightly higher but still moderate alterations were observed in the kidneys of fish exposed to propamocarb hydrochloride at 80 μg/L (IK = 15). Pirimiphos-methyl at 60 μg/L caused a more pronounced level of damage (IK = 27), while the most severe histopathological response was observed following exposure to 2,4-D at 100 μg/L, with an IK value of 34, representing substantial tissue disruption.
The statistical analysis using the Kruskal–Wallis test revealed a highly significant difference in IK values among the treatment groups (Hc = 58.70, p < 0.001). The pairwise post hoc analysis using the Mann–Whitney U test confirmed significant differences between most treatment pairs. Specifically, the control group differed significantly from all treated groups. However, no significant differences were found among the groups treated with pirimiphos-methyl at 10 μg/L, propamocarb hydrochloride at 80 μg/L, and 2,4-D at 50 μg/L (Mann–Whitney U test, p > 0.05). These treatments were therefore considered statistically similar in terms of their overall impact on kidney histopathology. In contrast, 2,4-D at 100 μg/L induced significantly more severe lesions than all other treatments, highlighting its strong nephrotoxic potential under the conditions tested (Figure 5.).
Figure 6 presents the PCA based on the histopathological scores from carp kidneys, showing a clear separation between treatment groups along the first two principal components (PCA1 = 35.4%, PCA2 = 22.5% of total variance). A clear separation was observed among the treatment groups along the first two principal components. Notably, the group exposed to 100 µg/L 2,4-dichlorophenoxyacetic acid (2,4-D 100) formed a distinct cluster, largely influenced by severe vacuolar degeneration of tubular cells, interstitial edema, glomerular hyperplasia, interstitial necrosis, glomerular necrobiosis, glomerular necrosis, and glomerular hypertrophy. In contrast, the groups treated with pirimiphos-methyl at both 10 µg/L (Pir-10) and 60 µg/L (Pir-60) were separated from others primarily due to thickening of Bowman’s capsular membrane, tubular necrosis, tubular hypertrophy, tubular necrobiosis, activation of melano-macrophages, interstitial hypertrophy, dilatation of Bowman’s capsule, and glomerular contraction. These patterns indicate that different pesticide compounds induced distinct histopathological responses, with glomerular and interstitial lesions being particularly influential in group differentiation. Vectors near the origin, representing negligible influence on separation, were intentionally left unlabeled for clarity. This multivariate approach reinforces the distinct pathological profiles induced by specific pesticide exposures (Fig 6).
Our findings are consistent with previously documented pesticide-induced histopathological effects in fish kidneys, including glomerular atrophy, vacuolization, hypertrophy, and necrosis [70,71,72]. However, our study is among the first to assess and statistically confirm the specific toxic profiles of pirimiphos-methyl and propamocarb hydrochloride under controlled laboratory conditions.
The results of the histopathological analysis of the kidney, after treatment with pirimiphos-methyl, propamocarb hydrochloride and 2,4-D, showed the presence of morphological changes in the structure of the organ. The observed proliferative changes increased in direct proportion to the applied concentrations for all three experimental pesticides. On the other hand, the degenerative changes, which also increased with the increasing pesticide concentration, were relatively less pronounced compared to those found in the liver. Inflammatory changes in the organ were more pronounced compared to those in the liver, despite the lower degrees of manifestation of degenerative disorders. This was probably because the kidneys have a weaker regenerative potential than the liver. Circulatory changes were not found in the kidney after the pesticide exposure.
Similarly to our results on the toxic effects of pesticides on the normal histostructure of the fish kidney, many previous studies have reported that pesticides cause multiple histological changes in the kidney tissues of fish. Al-Otaibi et al. (2019) [70] demonstrated that diazinon exposure caused glomerular hypertrophy and haemorrhages in the kidney tissues of African sharptooth catfish (Clarias gariepinus Burchell, 1822). Inflammation and necrosis were observed in the kidney tissues of rainbow trout (Oreochromis mykiss Walbaum, 1792) exposed to the fungicide captan [73]. Also, Nile tilapia (Oreochromis niloticus Linnaeus, 1758) exposed to sumithione showed deformed and pyknotic nuclei and vacuolization in kidney tissues [74]. Cypermethrin caused necrosis, karyolysis and ruptured renal tubules in Tengara catfish (Mystus tengara F. Hamilton, 1822) [75]. Cengiz et al. (2006) [76] and Vinodhini & Narayanan (2009) [77] also revealed similar changes in carp exposed to deltamethrin and toxic metals. Java barb (Barbonymus gonionotus Bleeker, 1849) exposed to chronic sublethal concentrations of quinalphos showed renal changes, such as degeneration of renal corpuscles, vacuolization, severely degenerated and dilated renal tubules and hematopoietic tissue, changes in nuclear structure, mild to severe necrosis, and haemorrhage [78]. After glyphosate treatment, renal tubules become severely dilated, and some of them are characterized by loss of cellular integrity. Enlargement, edema, and hypertrophied nuclei are also observed. Glomeruli show vacuolization and disorganized blood capillaries. Necrosis and pyknotic nuclei are observed in the mesenchymal tissue [79]. Ayoola & Ajani (2008) [80] reported that exposure to cypermethrin caused tubular fusion and condensation of the glomerular contents. Ortiz et al. (2003) [81] reported epithelial lifting and necrosis in the tubule walls due to the toxic effects of lindane. Edema, atrophy of the tubule lumen, necrosis, pyknosis, congestion, and glomerular degeneration were found in the kidney of fish treated with fipronil [71]. In fish exposed to deltamethrin for 24 hours, the kidney showed degeneration in renal tubular epithelial cells, pyknotic nuclei in hematopoietic tissue, dilation of glomerular capillaries, narrowing of the tubular lumen, intracytoplasmic vacuoles in renal tubular epithelial cells with hypertrophied cells, and glomerular degeneration [69]. Dilatation of the tubular lumen diameter, pyknotic nuclei of some epithelial cells, hyaline degeneration, hydropic edema and vacuolization were also observed in the renal tubules of Cyprinus carpio after exposure to dimethoate. Das & Mukherjee (2000) [82] reported dilatation of the renal tubules and necrotic changes characterized by karyorrhexis and karyolysis in rohu (Labeo rohita F. Hamilton, 1822) exposed to hexachlorocyclohexane. Tilak et al. (2001) [83] observed severe necrosis, edema in the renal tubules, cellular hypertrophy, granular cytoplasm, and vacuolization in the renal tissues of grass carp (Ctenopharyngodon idella Cuvier & Valenciennes, 1844) after exposure to fenvalerate. Degeneration in renal tubular epithelial cells, pyknotic nuclei in hematopoietic tissue, dilation of glomerular capillaries, glomerular degeneration, intracytoplasmic vacuoles in epithelial cells with hypertrophy and narrowing of the tubular lumen were observed in renal tissues of fish exposed to deltamethrin [69]. Velmurugan et al. (2007) [84] reported pyknotic nuclei in the renal tubular epithelium, hypertrophied tubular epithelial cells, glomerular shrinkage and expansion of the space within the Bowman's capsule in the kidney of mrigal (Cirrhinus mrigala Hamilton, 1822) exposed to monocrotophos. The result of histopathological examination revealed remarkable damages such as vacuolization of renal parenchyma and dilation of renal tubules in fish exposed to glyphosate [72]. Histopathological examination of kidney cells from African sharptooth catfish exposed to 2,4-D showed glomerular loss, vascular congestion, and intrarenal haemorrhage [85]. In contrast to these authors, at the same exposure but with a lower concentration of the herbicide in the glomerulus, a low degree of dilation of the Bowman's capsule and very mild necrobiotic changes in the epithelial cells of the renal tubules and necrosis were found in our study.
Overall, the observed histopathological alterations were grouped into four categories - changes in the circulatory system, proliferative, degenerative changes and changes related to inflammatory processes. Post hoc Mann–Whitney U tests revealed specific pairwise distinctions, notably highlighting the severity of 2,4-D at 100 µg/L as the most toxic treatment, inducing irreversible alterations. The degenerative changes were more pronounced following exposure to 2,4-D and pirimiphos-methyl, with 2,4-D at 100 µg/L producing the highest index for degenerative changes (IKR = 15). Proliferative responses also followed a concentration-dependent trend, with glomerular and interstitial hypertrophy being the most common. The inflammatory responses - particularly melanomacrophage activation were more intense in fish treated with pirimiphos-methyl, indicating potential immunological involvement. Comparatively, the severity of kidney damage followed a consistent rank order across all indices: 2,4-D showed the highest nephrotoxic potential at higher doses, often causing irreversible tissue changes. Pirimiphos-methyl induced moderate to severe alterations depending on concentration, including significant proliferative and inflammatory lesions. Propamocarb hydrochloride, though toxic, triggered only mild to moderate and largely reversible changes, as confirmed statistically. Regarding the comparative analysis of the toxic pesticide impact on the kidney structure of common carp, the degree of toxicity of the tested pesticides is presented in Figure 7 and can be summarised in the following descending order: for proliferative changes: 2,4-D > pirimiphos-methyl > propamocarb hydrochloride; for degenerative changes: 2,4-D > pirimiphos-methyl > propamocarb hydrochloride; for inflammation: pirimiphos-methyl > 2,4-D > propamocarb hydrochloride.
Since no changes in the renal circulatory system were found, a comparison between the three pesticides on this criterion could not be made. In the case of proliferative and degenerative changes, the most severe degree of manifestation was observed after exposure to 2,4-D, followed by pirimiphos-methyl, and the least pronounced changes were found after exposure to propamocarb hydrochloride. Regarding the changes related to inflammatory processes, the most severe degree of manifestation was caused by pirimiphos-methyl, and the mildest by propamocarb hydrochloride (Figure 7).
4. Conclusions
In summary, we can conclude that all three applied pesticides negatively affect the histological structure of the kidney of common carp. Along with this, compensatory-adaptive mechanisms were activated in the fish organ, which affect the kidney functionality and inevitably lead to a deterioration in the overall fish health. There was a tendency towards an increase in morphological changes, and the degree of their expression is directly proportional to the increasing concentrations of the applied pesticides. The results of the present study on the identification of histopathological biomarkers in the bioindicator species common carp could be used to determine maximum permissible concentrations of organic pollutants in biota, as well as for ecological biomonitoring, applying an assessment model based on correlation dependencies between the identified biomarkers.
This study confirms that pirimiphos-methyl, propamocarb hydrochloride, and 2,4-D induce dose-dependent histopathological alterations in the kidney of common carp, even during short-term (96-hour) exposure. Among the three tested pesticides, 2,4-D exhibited the most pronounced and irreversible damage, particularly at 100 µg/L, as demonstrated by high kidney index scores and strong statistical separation in group comparisons. Pirimiphos-methyl caused moderate to severe lesions, whereas propamocarb hydrochloride showed comparatively mild effects.
The applied histopathological scoring system, reinforced by Kruskal–Wallis and Mann–Whitney U tests, effectively distinguished treatment effects and revealed significant concentration-dependent differences. These findings highlight the value of integrating morphological and statistical assessment to quantify sublethal organ damage in ecotoxicology. The study underscores the importance of regulating pesticide exposure levels and validates kidney biomarkers as effective tools for ecological risk assessment and environmental monitoring.
Funding
This work was supported by the European Union-NextGenerationEU, through the National Recovery and Resilience Plan of the Republic of Bulgaria, project № BG-RRP-2.004-0001-C01.
Acknowledgements
This research was funded by the European Union - NextGenerationEU, through the National Recovery and Resilience Plan of the Republic of Bulgaria, grant № BG-RRP-2.004-0001-C01, DUEcoS.
Conflicts of Interest
The authors declare no conflict of interest.
References
- US EPA. What is a Pesticide? Chapter 6—Insecticides and environmental pesticide control subchapter II—Environmental pesticide control. Se. 136—Definitions (2013).
- Tang, F.H.M. , et al., Risk of pesticide pollution at the global scale. Nature Geoscience, 2021. 14(4): p. 206-210. [CrossRef]
- Zaller, J.G., Daily Poison: Pesticides - an Underestimated Danger. Springer Nature 2020.
- Hvezdova, M. , et al., Currently and recently used pesticides in Central European arable soils. Sci Total Environ, 2018. 613-614: p. 361-370. [CrossRef]
- Hamed, A.S.A. and N. Hidayah, Global Trade and Pesticide Use, in The Interplay of Pesticides and Climate Change: Environmental Dynamics and Challenges, B.R. Babaniyi and E.E. Babaniyi, Editors. 2025, Springer Nature Switzerland: Cham. p. 111-126.
- Sharma, A. , et al., Worldwide pesticide usage and its impacts on ecosystem. SN Applied Sciences, 2019. 1(11): p. 1446. [CrossRef]
- Sjerps, R.M.A. , et al., Occurrence of pesticides in Dutch drinking water sources. Chemosphere, 2019. 235: p. 510-518. [CrossRef]
- Rizzo, G., J.P. Monzon, and O. Ernst, Cropping system-imposed yield gap: Proof of concept on soybean cropping systems in Uruguay. Field Crops Research, 2021. 260: p. 107944. [CrossRef]
- Syafrudin, M. , et al., Pesticides in Drinking Water-A Review. Int J Environ Res Public Health, 2021. 18(2). [CrossRef]
- Pérez-Parada, A. , et al., Recent advances and open questions around pesticide dynamics and effects on freshwater fishes. Current Opinion in Environmental Science & Health, 2018. 4: p. 38-44. [CrossRef]
- de Souza, R.M. , et al., Occurrence, impacts and general aspects of pesticides in surface water: A review. Process Safety and Environmental Protection, 2020. 135: p. 22-37.
- Souza-Bastos, L.R. , et al., Evaluation of the water quality of the upper reaches of the main Southern Brazil river (Iguacu river) through in situ exposure of the native siluriform Rhamdia quelen in cages. Environ Pollut, 2017. 231(Pt 2): p. 1245-1255.
- Ortiz, J.B., M.L.G. De Canales, and C. Sarasquete, Histopathological changes induced by lindane (γ-HCH) in various organs of fishes. SCIENTIA MARINA, 2003. 67(1): p. 53-61. [CrossRef]
- Iqbal, F., I. Qureshi, and M. Ali, Histopathological changes in the kidney of common carp, Cyprinus carpio following nitrate exposure. J. Res. Sci, 2004. 15.
- Sachi, I.T.C. , et al., Biochemical and morphological biomarker responses in the gills of a Neotropical fish exposed to a new flavonoid metal-insecticide. Ecotoxicol Environ Saf, 2021. 208: p. 111459. [CrossRef]
- Weber, M.J. and M.L. Brown, Relationships among invasive common carp, native fishes and physicochemical characteristics in upper Midwest (USA) lakes. Ecology of Freshwater Fish, 2011. 20(2): p. 270-278. [CrossRef]
- Kloskowski, J. , Impact of common carp Cyprinus carpio on aquatic communities: Direct trophic effects versus habitat deterioration. Fundamental and Applied Limnology / Archiv für Hydrobiologie, 2011. 178: p. 245-255. [CrossRef]
- Ahmad, H. , et al., Pollution Problem in River Kabul: Accumulation Estimates of Heavy Metals in Native Fish Species. Biomed Res Int, 2015. 2015: p. 537368. [CrossRef]
- Xing, H. , et al., Oxidative stress response and histopathological changes due to atrazine and chlorpyrifos exposure in common carp. Pesticide Biochemistry and Physiology, 2012. 103(1): p. 74-80. [CrossRef]
- Mhadhbi, L. and R. Beiras, Acute Toxicity of Seven Selected Pesticides (Alachlor, Atrazine, Dieldrin, Diuron, Pirimiphos-Methyl, Chlorpyrifos, Diazinon) to the Marine Fish (Turbot, Psetta maxima). Water, Air, & Soil Pollution, 2012. 223(9): p. 5917-5930. [CrossRef]
- Stepic, S. , et al., Effects of individual and binary-combined commercial insecticides endosulfan, temephos, malathion and pirimiphos-methyl on biomarker responses in earthworm Eisenia andrei. Environ Toxicol Pharmacol, 2013. 36(2): p. 715-723. [CrossRef]
- Fleurat-Lessard, F. , et al., Effects of processing on the distribution of pirimiphos-methyl residues in milling fractions of durum wheat. Journal of Stored Products Research, 2007. 43(4): p. 384-395. [CrossRef]
- Oxborough, R.M. , Trends in US President's Malaria Initiative-funded indoor residual spray coverage and insecticide choice in sub-Saharan Africa (2008-2015): urgent need for affordable, long-lasting insecticides. Malar J, 2016. 15: p. 146.
- Dengela, D. , et al., Multi-country assessment of residual bio-efficacy of insecticides used for indoor residual spraying in malaria control on different surface types: results from program monitoring in 17 PMI/USAID-supported IRS countries. Parasit Vectors, 2018. 11(1): p. 71. [CrossRef]
- Oakeshott, J.G. , et al., Comparing the organophosphorus and carbamate insecticide resistance mutations in cholin- and carboxyl-esterases. Chem Biol Interact, 2005. 157-158: p. 269-75. [CrossRef]
- Donarski, W.J. , et al., Structure-activity relationships in the hydrolysis of substrates by the phosphotriesterase from Pseudomonas diminuta. Biochemistry, 1989. 28(11): p. 4650-5. [CrossRef]
- Elersek, T. and M. Filipic, Organophosphorous Pesticides - Mechanisms of Their Toxicity. 2011.
- Khan, H.A.A. , Variation in susceptibility to insecticides and synergistic effect of enzyme inhibitors in Pakistani strains of Trogoderma granarium. Journal of Stored Products Research, 2021. 91: p. 101775. [CrossRef]
- John, H. , et al., Chapter 56 - Toxicokinetic Aspects of Nerve Agents and Vesicants, in Handbook of Toxicology of Chemical Warfare Agents (Second Edition), R.C. Gupta, Editor. 2015, Academic Press: Boston. p. 817-856.
- Pieroh, E.A. , Krass, W., Hemmen, C., 1978. Propamocarb, einneues Fungizid zur Abwehr von Oomyceten im Zierpflanzen- und Gemüsebau. Meded. Fac. Landbouw. Rijksuniv. Gent, 43, 933-942. 1978.
- Taylor, J.C. , et al., Determination of residues of propamocarb in wine by liquid chromatography-electrospray mass spectrometry with direct injection. Food Addit Contam, 2004. 21(6): p. 572-7. [CrossRef]
- Hiemstra, M. and A. de Kok, Determination of propamocarb in vegetables using polymer-based high-performance liquid chromatography coupled with electrospray mass spectrometry. J Chromatogr A, 2002. 972(2): p. 231-9.
- Sahoo, S., R.S. Battu, and B. Singh, Development and Validation of QuEChERS Method for Estimation of Propamocarb Residues in Tomato (Lycopersicon esculentum Mill) and Soil. American Journal of Analytical Chemistry, 2011. 02. [CrossRef]
- Abd-Alrahman, S.H. and M.M. Almaz, Degradation of propamocarb-hydrochloride in tomatoes, potatoes and cucumber using HPLC-DAD and QuEChERS methodology. Bull Environ Contam Toxicol, 2012. 89(2): p. 302-5.
- Mohamed, H. Mpina, F.H., Fenamidone + Propamocarb Hydrochloride: A Promising Package for the Control of Early and Late Blights of Tomatoes in Northern Tanzania. Int. J. Agr. Forestry, 3(3), 1-7, 2016.
- Wang, Z. , et al., Exogenous 24-epibrassinolide regulates antioxidant and pesticide detoxification systems in grapevine after chlorothalonil treatment. Plant Growth Regulation, 2017. 81(3): p. 455-466. [CrossRef]
- Li, C. , et al., Predicting the Relative Bioavailability of DDT and Its Metabolites in Historically Contaminated Soils Using a Tenax-Improved Physiologically Based Extraction Test (TI-PBET). Environ Sci Technol, 2016. 50(3): p. 1118-25. [CrossRef]
- USEPA. 1995. U.S. Environmental Protection Agency. Propamocarb Hydrochloride R.E.D. FACTS - Prevention, P.а.T.S.W.
- Elliott, M., S.F. Shamoun, and G. Sumampong, Effects of systemic and contact fungicides on life stages and symptom expression of Phytophthora ramorum in vitro and in planta. Crop Protection, 2015. 67: p. 136-144. [CrossRef]
- Dehnert, G.K. , et al., Effects of low, subchronic exposure of 2,4-Dichlorophenoxyacetic acid (2,4-D) and commercial 2,4-D formulations on early life stages of fathead minnows (Pimephales promelas). Environ Toxicol Chem, 2018. 37(10): p. 2550-2559.
- Heap, I. 2022. The International Herbicide-Resistant Weed Database. https://www.weedscience.org.
- USEPA. 2015. U.S. Environmental Protection Agency. Pesticides Industry Sales and Usage 2008–2012 Market Estimates. https://www.epa.gov/pesticides/pesticides-industrysales-and-usage-2008-2012-market-estimates.
- Islam, F. , et al., 2,4-D attenuates salinity-induced toxicity by mediating anatomical changes, antioxidant capacity and cation transporters in the roots of rice cultivars. Sci Rep, 2017. 7(1): p. 10443. [CrossRef]
- Song, Y. , Insight into the mode of action of 2,4-dichlorophenoxyacetic acid (2,4-D) as an herbicide. J Integr Plant Biol, 2014. 56(2): p. 106-13.
- von Stackelberg, K. , A Systematic Review of Carcinogenic Outcomes and Potential Mechanisms from Exposure to 2,4-D and MCPA in the Environment. J Toxicol, 2013. 2013: p. 371610.
- RED. 2006. Reregistration eligibility decision (RED) 2, -.D.U.s.e.P.A.
- da Fonseca, M.B. , et al., The 2,4-D herbicide effects on acetylcholinesterase activity and metabolic parameters of piava freshwater fish (Leporinus obtusidens). Ecotoxicol Environ Saf, 2008. 69(3): p. 416-20.
- Kiljanek, T. , et al., Multi-residue method for the determination of pesticides and pesticide metabolites in honeybees by liquid and gas chromatography coupled with tandem mass spectrometry--Honeybee poisoning incidents. J Chromatogr A, 2016. 1435: p. 100-14. [CrossRef]
- Souza, F.L. , et al., Solar-powered electrokinetic remediation for the treatment of soil polluted with the herbicide 2,4-D. Electrochimica Acta, 2016. 190: p. 371-377. [CrossRef]
- Gul, U. and H. Silah, Monitoring 2,4-D removal by filamentous fungi using electrochemical methods. Biotech Studies, 2025. 34: p. 52-65. [CrossRef]
- Islam, F. , et al., Potential impact of the herbicide 2,4-dichlorophenoxyacetic acid on human and ecosystems. Environ Int, 2018. 111: p. 332-351. [CrossRef]
- Bartczak, P. , et al., Saw-sedge Cladium mariscus as a functional low-cost adsorbent for effective removal of 2,4-dichlorophenoxyacetic acid from aqueous systems. Adsorption, 2016. 22(4): p. 517-529. [CrossRef]
- APHA. 2005. Standard methods for examination of water and wastewater, s.E.W., American Public Health Association.
- Rosseland, B.O. , et al., Brown Trout in Lochnagar: Population and Contamination by Metals and Organic Micropollutants, in Lochnagar: The Natural History of a Mountain Lake, N.L. Rose, Editor. 2007, Springer Netherlands: Dordrecht. p. 253-285.
- relevance, D.E.o.t.E.P.a.o.t.C.o.S.o.t.p.o.a.u.f.s.p.T.w.E.
- Romeis, B. , Mikroskopische technik. München: Urban und Schwarzenberg, p. 697. Radic, Z., Taylor, P. 2006. Structure and function of cholinesterases. Toxicol. Organo-phosphat. Carbamate Comp., 1, 161-186. 1989.
- Bernet, D. , et al., Histopathology in fish: proposal for a protocol to assess aquatic pollution. Journal of Fish Diseases, 1999. 22(1): p. 25-34.
- Saraiva, A. , et al., A histology-based fish health assessment of farmed seabass (Dicentrarchus labrax L.). Aquaculture, 2015. 448: p. 375-381. [CrossRef]
- Zimmerli, S. , et al., Assessment of fish health status in four Swiss rivers showing a decline of brown trout catches. Aquatic Sciences, 2007. 69(1): p. 11-25. [CrossRef]
- Kruskal, W.H. and W.A. and Wallis, Use of Ranks in One-Criterion Variance Analysis. Journal of the American Statistical Association, 1952. 47(260): p. 583-621.
- Mann, H.B. and D.R. Whitney, On a test of whether one of two random variables is stochastically larger than the other. Annals of Mathematical Statistics, 1947. 18: p. 50-60.
- Pedregosa, F. , et al., Scikit-learn: Machine Learning in Python. Journal of Machine Learning Research, 2012. 12.
- Hunter, J.D. , Matplotlib: A 2D Graphics Environment. Computing in Science & Engineering, 2007. 9(3): p. 90-95.
- Takashima, F. and T. Hibiya, An Atlas of Fish Histology: Normal and Pathological Features. 1995: Kodansha Limited.
- Donaldson, E.M. pituitary-interrenal axis as an indicator of stress in fish. 1981.
- Wendelaar Bonga, S.E. , The stress response in fish. Physiol Rev, 1997. 77(3): p. 591-625. [CrossRef]
- Fedorova, S. , et al., Renal glomerulogenesis in medaka fish, Oryzias latipes. Dev Dyn, 2008. 237(9): p. 2342-52. [CrossRef]
- Mobjerg, N., A. Jespersen, and M. Wilkinson, Morphology of the kidney in the West African caecilian, Geotrypetes seraphini (Amphibia, Gymnophiona, Caeciliidae). J Morphol, 2004. 262(2): p. 583-607. [CrossRef]
- Cengiz, E.I. , Gill and kidney histopathology in the freshwater fish Cyprinus carpio after acute exposure to deltamethrin. Environ Toxicol Pharmacol, 2006. 22(2): p. 200-4. [CrossRef]
- Al-Otaibi, A.M. , et al., Toxicity bioassay and sub-lethal effects of diazinon on blood profile and histology of liver, gills and kidney of catfish, Clarias gariepinus. Braz J Biol, 2019. 79(2): p. 326-336. [CrossRef]
- Ghaffar, A. , et al., Fipronil (Phenylpyrazole) induces hemato-biochemical, histological and genetic damage at low doses in common carp, Cyprinus carpio (Linnaeus, 1758). Ecotoxicology, 2018. 27(9): p. 1261-1271. [CrossRef]
- Ma, J., Y. Bu, and X. Li, Immunological and histopathological responses of the kidney of common carp (Cyprinus carpio L.) sublethally exposed to glyphosate. Environ Toxicol Pharmacol, 2015. 39(1): p. 1-8. [CrossRef]
- Boran, H. , et al., Assessment of acute toxicity and histopathology of the fungicide captan in rainbow trout. Exp Toxicol Pathol, 2012. 64(3): p. 175-9. [CrossRef]
- Sharmin, S. , et al., Sumithion induced structural erythrocyte alteration and damage to the liver and kidney of Nile tilapia. Environ Sci Pollut Res Int, 2021. 28(27): p. 36695-36706. [CrossRef]
- Haque, S. , et al., Toxic effects of agro-pesticide cypermethrin on histological changes of kidney in Tengra, Mystus tengara. Asian Journal of Medical and Biological Research, 2018. 3: p. 494. [CrossRef]
- Cengiz, E.I. and E. Unlu, Sublethal effects of commercial deltamethrin on the structure of the gill, liver and gut tissues of mosquitofish, Gambusia affinis: A microscopic study. Environ Toxicol Pharmacol, 2006. 21(3): p. 246-53. [CrossRef]
- Rajamanickam, V. and M. Narayanan, Heavy Metal Induced Histopathological Alterations in Selected Organs of the Cyprinus carpio L.(Common Carp). International Journal of Environmental Research (ISSN: 1735-6865) Vol 3 Num 1, 2009. 3.
- Mohammod Mostakim, G. , et al., Alteration of Blood Parameters and Histoarchitecture of Liver and Kidney of Silver Barb after Chronic Exposure to Quinalphos. J Toxicol, 2015. 2015: p. 415984. [CrossRef]
- Sigamani, D. , Effect of herbicides on fish and histological evaluation. 2015: p. 437-440.
- Ayoola, S.O.a.A., E.K. , Histopathological Effect of Cypermethrin on Juvenile African Catfish (Clarias gariepinus). World Journal of Biological Research. Vol.1(2): 1-14. 2008.
- Ortiz-Delgado, J., M. Canales, and C. Sarasquete, Cambios histopatológicos inducidos por lindano (?HCH) en varios órganos de peces. Scientia Marina, 2003. 67.
- Das, B. and S. Mukherjee, A histopathological study of carp (Labeo rohita) exposed to hexachlorocyclohexane. Veterinarski Arhiv, 2000. 70.
- Tilak, K., K. Veeraiah, and K. Yacobu, Studies on histopathological changes in the Gill, liver and kidney of Ctenopharyngodon idellus (Valenciennes) exposed totechnical fenvalerate and EC 20%. Pollution Research, 2001. 20: p. 387-393.
- Velmurugan, B. , et al., The effects of monocrotophos to different tissues of freshwater fish Cirrhinus mrigala. Bull Environ Contam Toxicol, 2007. 78(6): p. 450-4. [CrossRef]
- Olatubi, O.K. and A. Ojokoh, Effects of Fermentation and Extrusion on the Proximate Composition of Corn-Groundnut Flour Blends. Nigerian Journal of Biotechnology, 2016. 30: p. 59. [CrossRef]
Figure 1.
A normal histological structure of the kidney from the common carp control group (A, B), x400, H&E.
Figure 1.
A normal histological structure of the kidney from the common carp control group (A, B), x400, H&E.
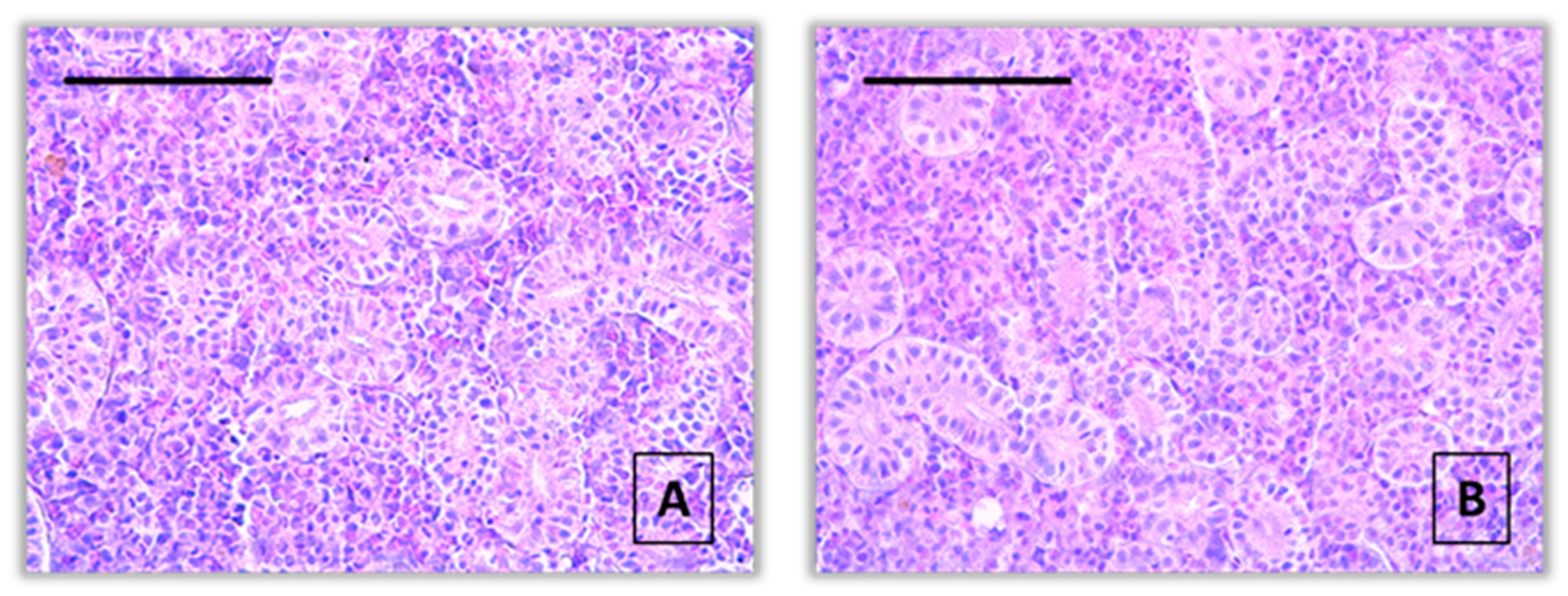
Figure 2.
Histopathological alterations in kidney of common carp after acute (96-hour) exposure with pirimiphos-methyl (H&E): A – activation of melanomacrophages (red arrow) and vacuolar degeneration (thin black arrow) (10 μg/L), x400; B – vacuolar degeneration (thin black arrow) (10 μg/L), x400; C – dilatation of Bowman’s capsule (thick black arrow) (10 μg/L), x400; D - dilatation of Bowman’s capsule (thick black arrow) (60 μg/L), x400; E – vacuolar degeneration (thin black arrow) (60 μg/L), x400; F – hypertrophy of interstitial tissue (star) (60 μg/L), x400.
Figure 2.
Histopathological alterations in kidney of common carp after acute (96-hour) exposure with pirimiphos-methyl (H&E): A – activation of melanomacrophages (red arrow) and vacuolar degeneration (thin black arrow) (10 μg/L), x400; B – vacuolar degeneration (thin black arrow) (10 μg/L), x400; C – dilatation of Bowman’s capsule (thick black arrow) (10 μg/L), x400; D - dilatation of Bowman’s capsule (thick black arrow) (60 μg/L), x400; E – vacuolar degeneration (thin black arrow) (60 μg/L), x400; F – hypertrophy of interstitial tissue (star) (60 μg/L), x400.
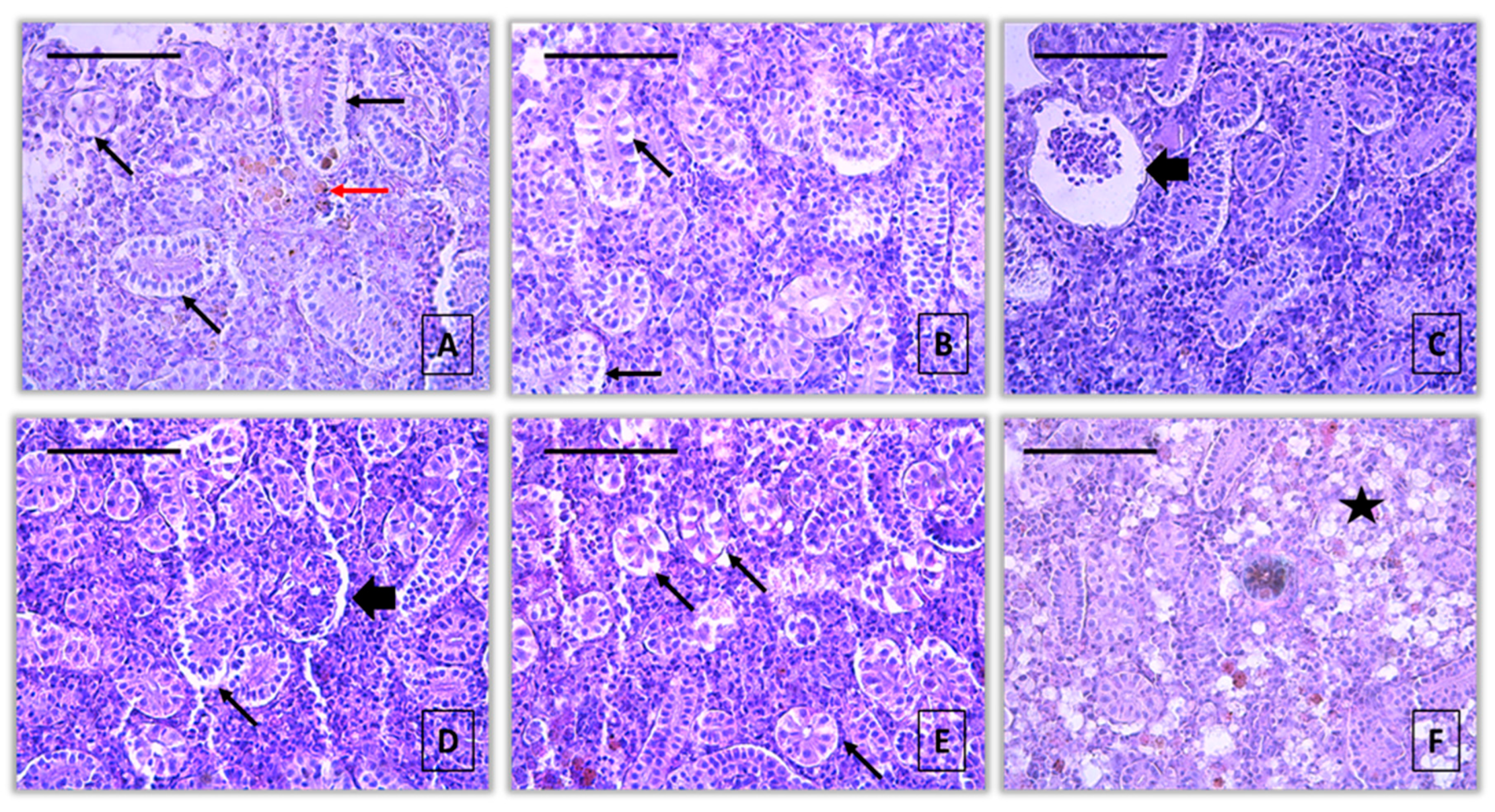
Figure 3.
Histopathological alterations in kidney of common carp after acute (96-hour) exposure with propamocarb hydrochloride (H&E): A, B – vacuolar degeneration (thin black arrow) (40 μg/L), x400; C – hypertrophy of the tubule (thick red arrow) (40 μg/L), x400; D – dilatation of Bowman’s capsule (thick black arrow) and vacuolar degeneration (thin black arrow) (80 μg/L), x400; E - vacuolar degeneration (thin black arrow) (80 μg/L), x400; F – vacuolar degeneration (thin black arrow), dilatation of Bowman’s capsule (thick black arrow) and activation of melanomacrophages (thin red arrow) (80 μg/L), x400.
Figure 3.
Histopathological alterations in kidney of common carp after acute (96-hour) exposure with propamocarb hydrochloride (H&E): A, B – vacuolar degeneration (thin black arrow) (40 μg/L), x400; C – hypertrophy of the tubule (thick red arrow) (40 μg/L), x400; D – dilatation of Bowman’s capsule (thick black arrow) and vacuolar degeneration (thin black arrow) (80 μg/L), x400; E - vacuolar degeneration (thin black arrow) (80 μg/L), x400; F – vacuolar degeneration (thin black arrow), dilatation of Bowman’s capsule (thick black arrow) and activation of melanomacrophages (thin red arrow) (80 μg/L), x400.
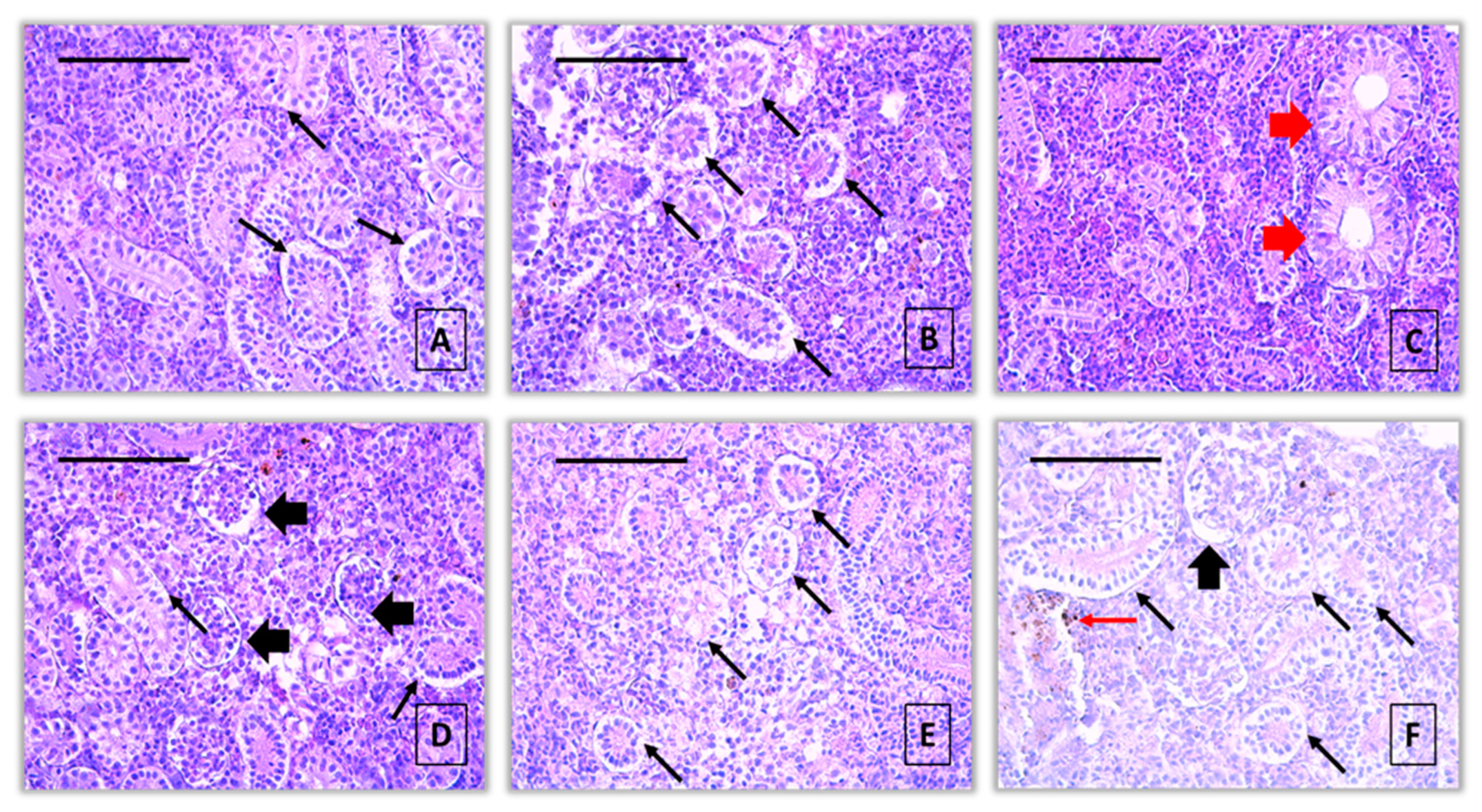
Figure 4.
Histopathological alterations in kidney of common carp after acute (96-hour) exposure with 2, 4 - D (H&E): A – vacuolar degeneration (thin black arrow) (50 μg/L), x400; B – necrosis of glomerulus (rhombus) (50 μg/L), x400; C – necrosis of glomerulus (rhombus) and dilatation of Bowman’s capsule (thick black arrow) (50 μg/L), x400; D, E – vacuolar degeneration (thin black arrow) (100 μg/L), x400; F - vacuolar degeneration (thin black arrow) (80 μg/L), x400; F – vacuolar degeneration (thin black arrow) and activation of melanomacrophages (thin red arrow) (100 μg/L), x400.
Figure 4.
Histopathological alterations in kidney of common carp after acute (96-hour) exposure with 2, 4 - D (H&E): A – vacuolar degeneration (thin black arrow) (50 μg/L), x400; B – necrosis of glomerulus (rhombus) (50 μg/L), x400; C – necrosis of glomerulus (rhombus) and dilatation of Bowman’s capsule (thick black arrow) (50 μg/L), x400; D, E – vacuolar degeneration (thin black arrow) (100 μg/L), x400; F - vacuolar degeneration (thin black arrow) (80 μg/L), x400; F – vacuolar degeneration (thin black arrow) and activation of melanomacrophages (thin red arrow) (100 μg/L), x400.
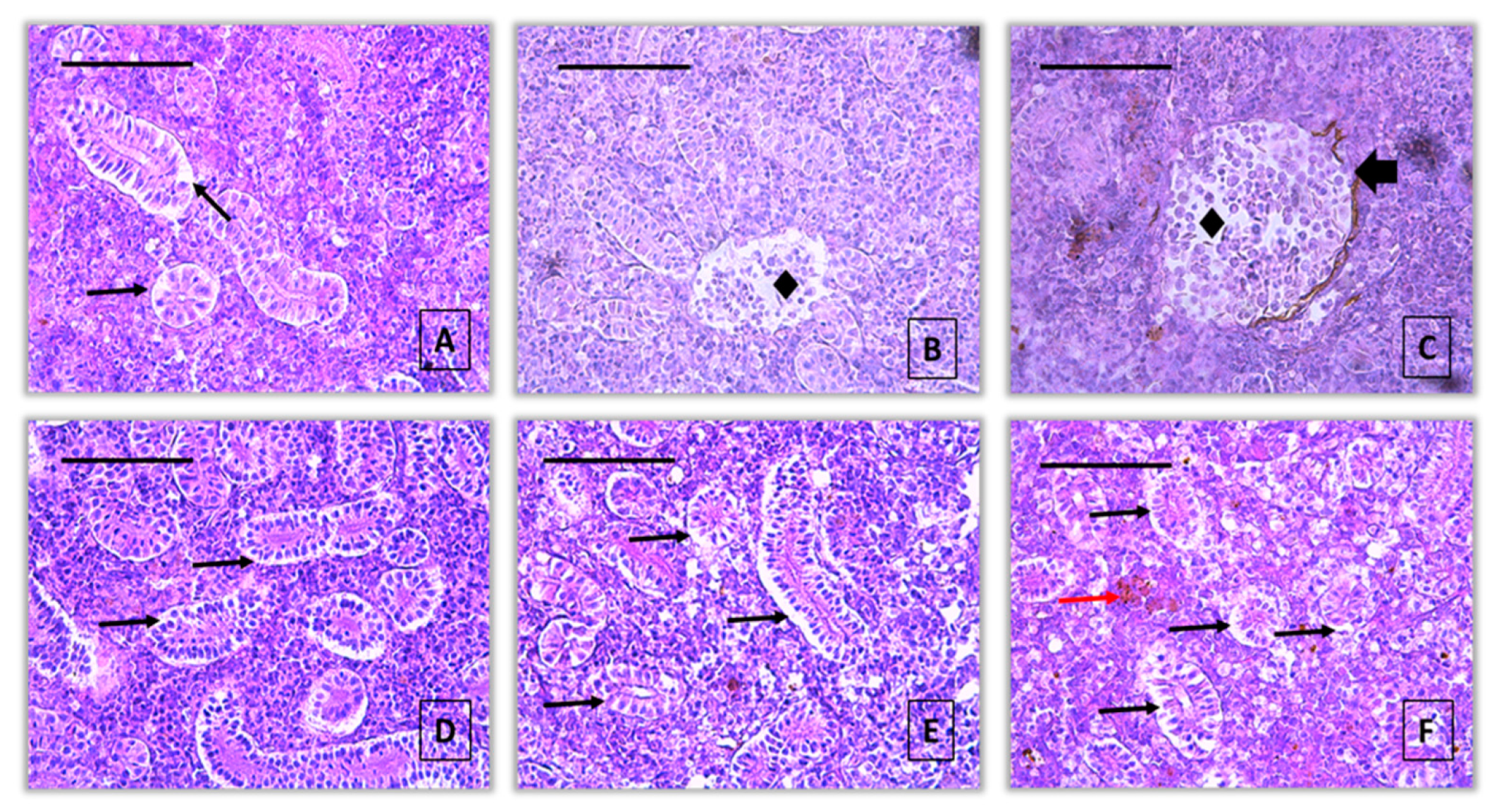
Figure 5.
Index for organ (IK) in the kidney of common carp after acute (96-hour) exposure to the tested concentrations of pirimiphos-methyl, propamocarb hydrochloride, and 2,4-D. Different letters indicate statistically significant differences between treatment groups (Mann–Whitney U test, p < 0.05).
Figure 5.
Index for organ (IK) in the kidney of common carp after acute (96-hour) exposure to the tested concentrations of pirimiphos-methyl, propamocarb hydrochloride, and 2,4-D. Different letters indicate statistically significant differences between treatment groups (Mann–Whitney U test, p < 0.05).
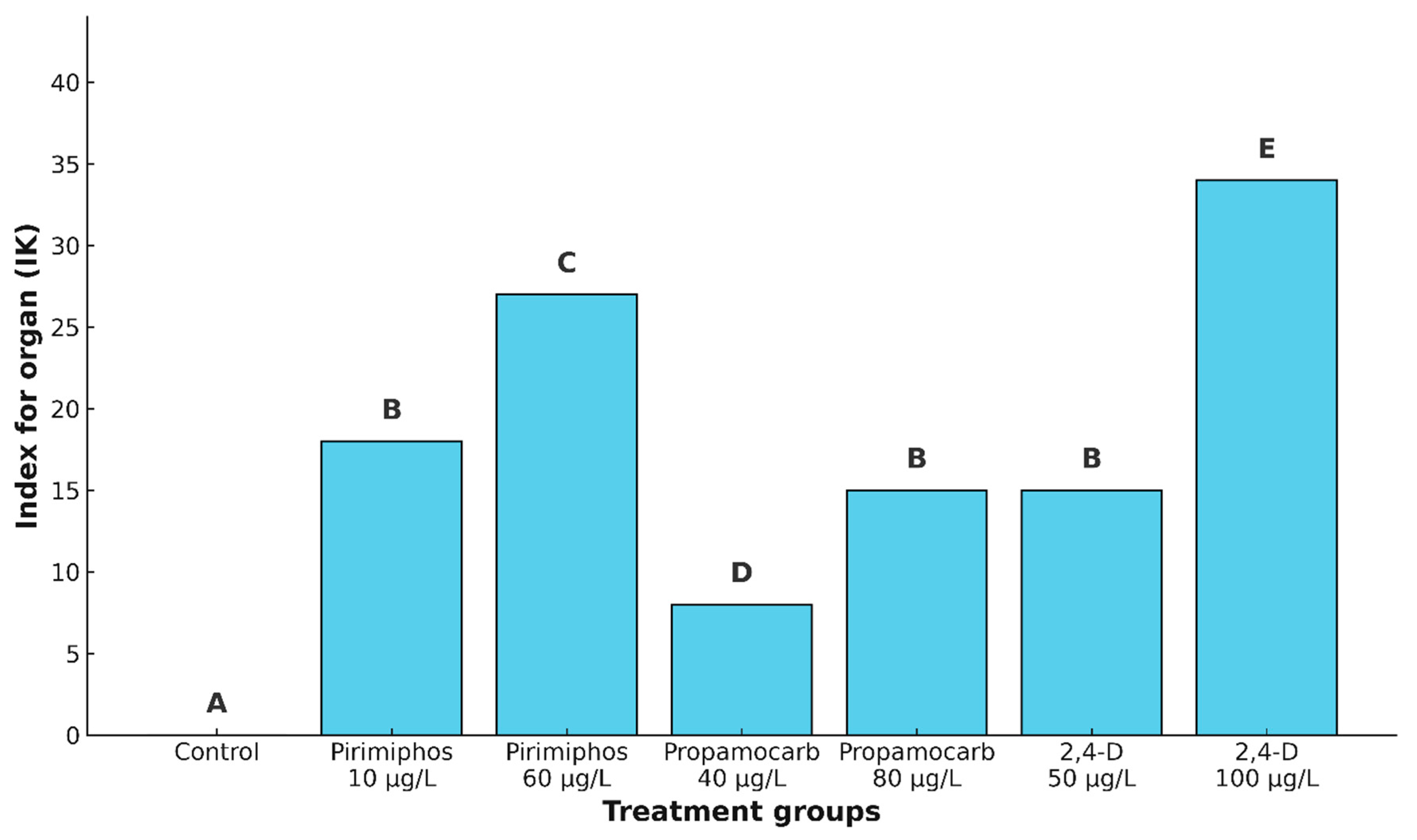
Figure 6.
Principal component analysis (PCA) of kidney histopathological alterations in common carp following acute (96-hour) pesticide exposure. PCA biplot showing the distribution of common carp (n = 10 per group) based on kidney histopathological alterations after 96-hour exposure to different pesticide treatments. Each point represents an individual specimen, grouped by treatment and enclosed by 95% confidence ellipses. Vectors represent the histopathological variables contributing to separation along PCA1 and PCA2 axes (explaining 35.4% and 25.5% of total variance, respectively); only vectors with meaningful influence (i.e., not located at the origin) are labeled. Treatment groups:(Pir-10): 10 µg/L pirimiphos-methyl; (Pir-60): 60 µg/L pirimiphos-methyl; (Pro-40): 40 µg/L propamocarb hydrochloride; (Pro-80): 80 µg/L propamocarb hydrochloride; (2,4-D 50): 50 µg/L 2,4-dichlorophenoxyacetic acid; (2,4-D 100): 100 µg/L 2,4-dichlorophenoxyacetic acid. Abbreviations of histopathological variables (vectors): KHem – haemorrhage; KHyp – hyperaemia; KAne – aneurysms; TVacDeg – vacuolar degeneration; THyalDeg – hyaline degeneration; TNecBio – tubular necrobiosis; TNec – tubular necrosis; GDilBow – dilatation of Bowman’s capsule; GCont – glomerular contraction; GNecBio – glomerular necrobiosis; GNec – glomerular necrosis; INec – interstitial necrosis; TProlHyp – tubular hypertrophy; TProlHpl – tubular hyperplasia; GProlHyp – glomerular hypertrophy; GProlHpl – glomerular hyperplasia; GBowThick – thickening of Bowman’s capsular membrane; IProlHyp – interstitial hypertrophy; IEdema – interstitial edema; KInfl – infiltration; KActMM – activation of melano-macrophages.
Figure 6.
Principal component analysis (PCA) of kidney histopathological alterations in common carp following acute (96-hour) pesticide exposure. PCA biplot showing the distribution of common carp (n = 10 per group) based on kidney histopathological alterations after 96-hour exposure to different pesticide treatments. Each point represents an individual specimen, grouped by treatment and enclosed by 95% confidence ellipses. Vectors represent the histopathological variables contributing to separation along PCA1 and PCA2 axes (explaining 35.4% and 25.5% of total variance, respectively); only vectors with meaningful influence (i.e., not located at the origin) are labeled. Treatment groups:(Pir-10): 10 µg/L pirimiphos-methyl; (Pir-60): 60 µg/L pirimiphos-methyl; (Pro-40): 40 µg/L propamocarb hydrochloride; (Pro-80): 80 µg/L propamocarb hydrochloride; (2,4-D 50): 50 µg/L 2,4-dichlorophenoxyacetic acid; (2,4-D 100): 100 µg/L 2,4-dichlorophenoxyacetic acid. Abbreviations of histopathological variables (vectors): KHem – haemorrhage; KHyp – hyperaemia; KAne – aneurysms; TVacDeg – vacuolar degeneration; THyalDeg – hyaline degeneration; TNecBio – tubular necrobiosis; TNec – tubular necrosis; GDilBow – dilatation of Bowman’s capsule; GCont – glomerular contraction; GNecBio – glomerular necrobiosis; GNec – glomerular necrosis; INec – interstitial necrosis; TProlHyp – tubular hypertrophy; TProlHpl – tubular hyperplasia; GProlHyp – glomerular hypertrophy; GProlHpl – glomerular hyperplasia; GBowThick – thickening of Bowman’s capsular membrane; IProlHyp – interstitial hypertrophy; IEdema – interstitial edema; KInfl – infiltration; KActMM – activation of melano-macrophages.
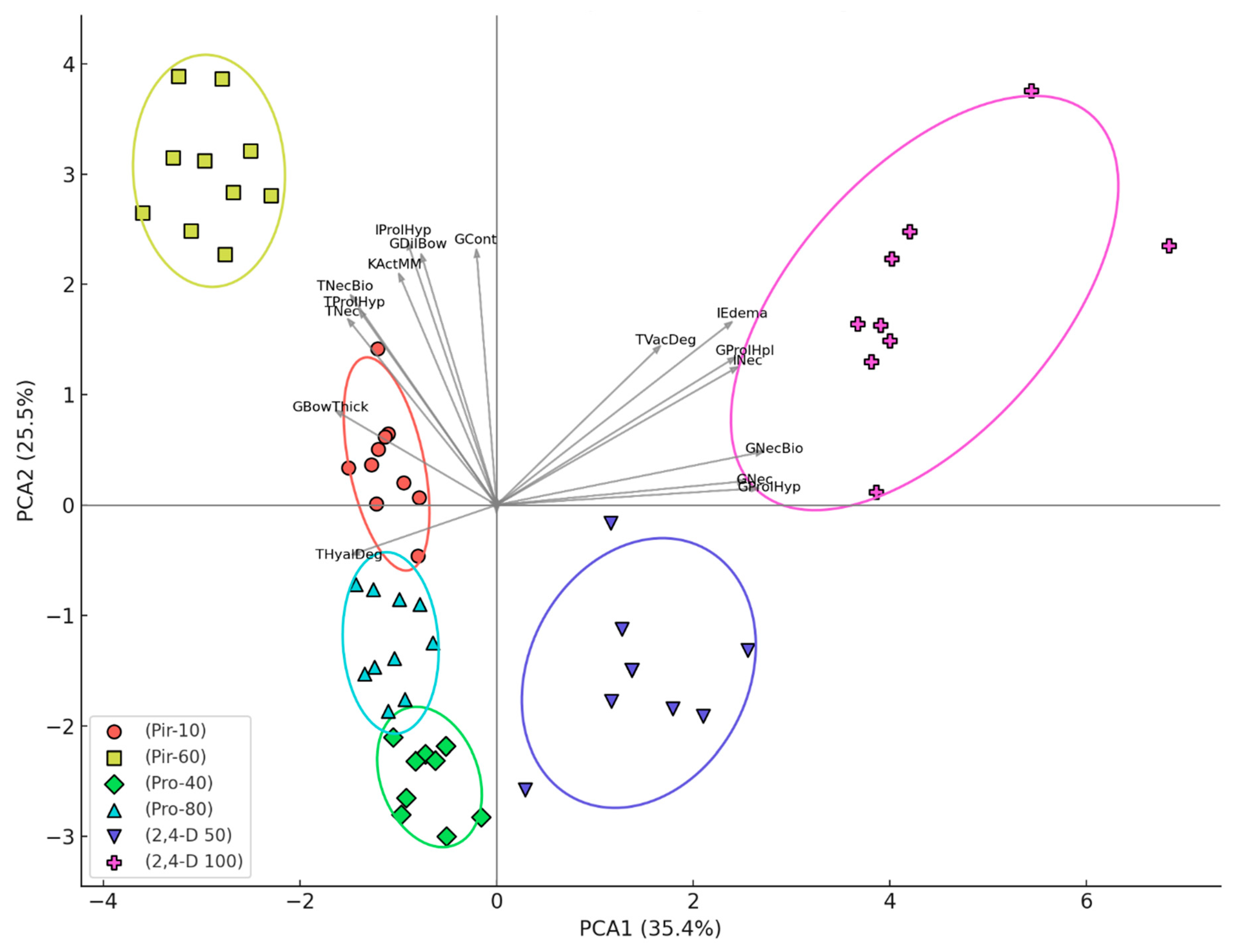
Figure 7.
Comparative analysis of the acute (96-hour) pesticide impact on the kidney structure of common carp.
Figure 7.
Comparative analysis of the acute (96-hour) pesticide impact on the kidney structure of common carp.
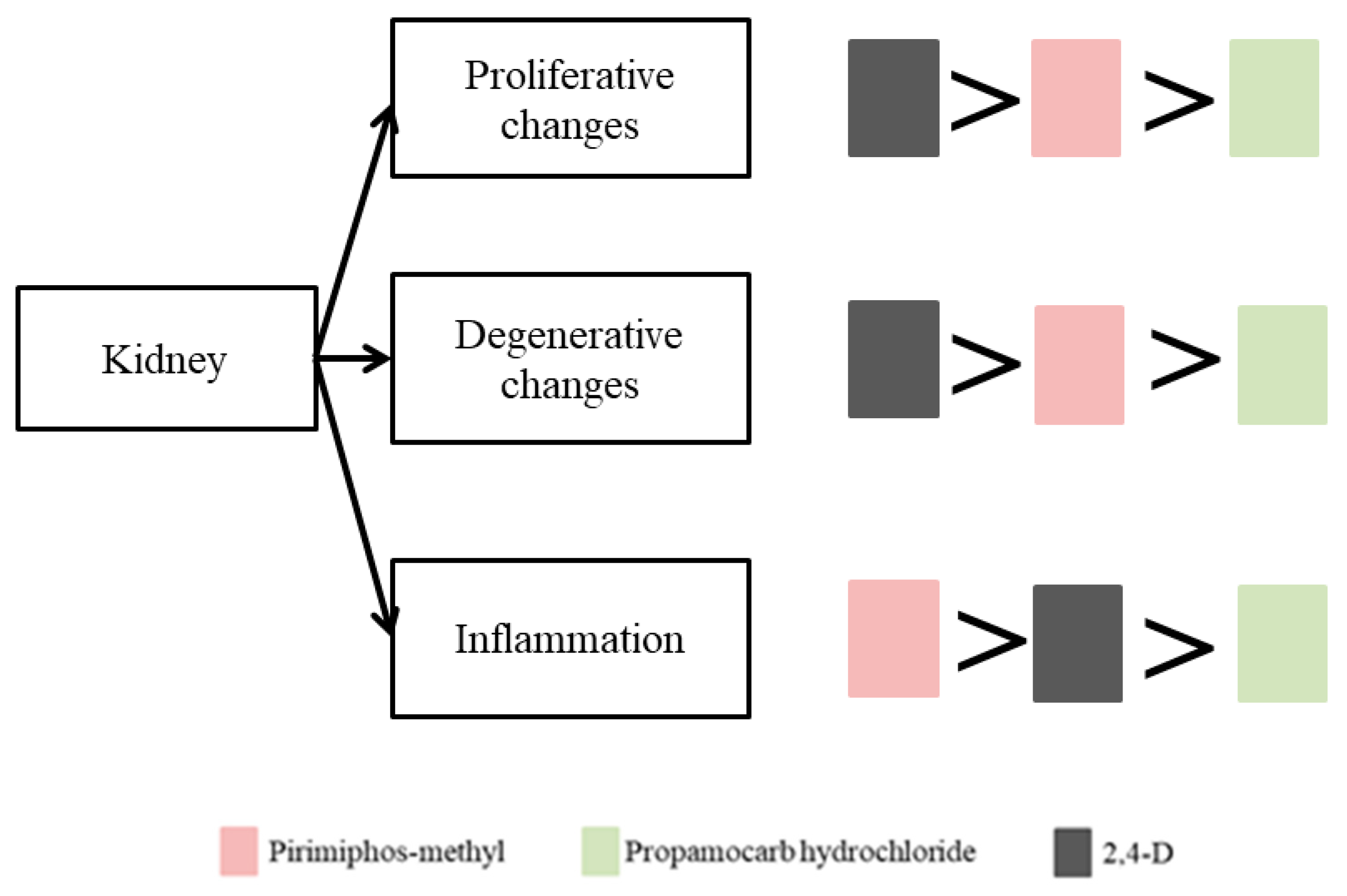
Table 1.
Length and weight of the experimental specimens.
| Indicator | Control | Pirimiphos-methyl | Propamocarb hydrochloride | 2,4-D | |||
|---|---|---|---|---|---|---|---|
| 10 μg/L | 60 μg/L | 40 μg/L | 80 μg/L | 50 μg/L | 100 μg/L | ||
| Total length(cm) | 9,16±0,4 | 10,44±0,8 | 11,49±0,5 | 9,64±0,63 | 8,67±2,9 | 9,31±0,2 | 11,13±0,3 |
| Weight(g) | 18,83±5,7 | 19,25±2,8 | 18,89±2.4 | 19,11±3,1 | 19,57±2,7 | 16,28±2,5 | 15,87±3,5 |
Table 2.
Experimental concentrations of the applied pesticides.
| Pesticide | Recommended dose | Dilution relative to LC50 | Experimental concentration |
|---|---|---|---|
| Pirimiphos-methyl | 0.00015 mL/m3 - 1 mL/dka | x 60 000 | 10 μg/L |
| x 10 000 | 60 μg/L | ||
| Propamocarb hydrochloride | 0.01-0.3 mL/dka | x 2 000 | 40 μg/L |
| x 1 000 | 80 μg/L | ||
| 2,4-D | 120-200 mL/dka | x 2 000 | 50 μg/L |
| x 1 000 | μg/L |
Table 3.
Water parameters during the pesticide exposure, 96 hours.
| Pirimiphos-methyl | pH | To | Dissolved oxygen mg/L |
Electrical conductivity μS/cm |
|---|---|---|---|---|
| Control | 8,3±,.05 | 17,5±0,05 | 9±0,05 | 365±0.05 |
| 10 μg/L | 8,2±0,05 | 17±0,05 | 8,5±0,05 | 426±0.05 |
| 60 μg/L | 8,1±0,05 | 17,5±0,05 | 7,3±0,05 | 473±0.05 |
| Propamocarb hydrochloride | pH | To |
Dissolved oxygen mg/L |
Electrical conductivity μS/cm |
| Control | 8,5±0,05 | 17,5±0,05 | 9±0,05 | 365±0,05 |
| 40 μg/L | 8,1±0,05 | 18±0,05 | 8,1±0,05 | 433±0,05 |
| 80 μg/L | 7,9±0,05 | 18,5±0,05 | 7,5±0,05 | 485±0,05 |
| 2,4-D | pH | To |
Dissolved oxygen mg/L |
Electrical conductivity μS/cm |
| Control | 8,5±0,05 | 17,5±0,05 | 9±0,05 | 365±0,05 |
| 50 μg/L | 7,5±0,05 | 17,5±0,05 | 8,2±0,05 | 451±0,05 |
| 100 μg/L | 6,9±0,05 | 18,5±0,05 | 7,7±0,05 | 469±0,05 |
Table 4.
Histopathological changes in the kidney of common carp after acute (96-hour) exposure to pirimiphos-methyl.
Table 4.
Histopathological changes in the kidney of common carp after acute (96-hour) exposure to pirimiphos-methyl.
| Reaction pattern |
Organ | Alteration | Importance factor | Score value | ||
|---|---|---|---|---|---|---|
| Control | Pirimiphos-methyl | |||||
| 10 μg/L | 60 μg/L | |||||
| Changes in the circulatory system | Kidney | Haemorrhage | WKC1 = 1 | 0A | 0A | 0A |
| Hyperaemia | WKC2 = 1 | 0A | 0A | 0A | ||
| Aneurysms | WKC3 = 1 | 0A | 0A | 0A | ||
| Index for the circulatory system | IKC =0A | IKC =0A | IKC =0A | |||
| Degenerative changes | Tubule | Vacuolar degeneration | WKR1 = 1 | 0A | 2B | 2B |
| Hyaline degeneration | WKR2 = 1 | 0A | 0A | 1B | ||
| Necrobiosis | WKR3 = 2 | 0A | 0A | 1B | ||
| Necrosis | WKR4 = 3 | 0A | 0A | 1B | ||
| Glomerulus | Dilatation of the Bowman's capsule | WKR5 = 1 | 0A | 2B | 3C | |
| Contraction | WKR6 = 1 | 0A | 1B | 1B | ||
| Necrobiosis* | WKR7 = 2 | 0A | 0A | 0A | ||
| Necrosis | WKR8 = 3 | 0A | 0A | 0A | ||
| Interstitial tissue | Necrosis | WKR9 = 3 | 0A | 0A | 0A | |
| Index forthedegenerative changes | IKR = 0A | IKR=5B | IKR=12C | |||
| Proliferative changes | Tubule | Hypertrophy | WKP1 = 1 | 0A | 2B | 2B |
| Hyperplasia | WKP2 = 2 | 0A | 0A | 0A | ||
| Glomerulus | Hypertrophy | WKP3 = 1 | 0A | 0A | 0A | |
| Hyperplasia | WKP4 = 2 | 0A | 0A | 0A | ||
| Thickening of Bowman's capsular membrane | WKP5 = 2 | 0A | 0A | 1B | ||
| Interstitial tissue | Hypertrophy | WKP6 = 1 | 0A | 3B | 3B | |
| Edema | WKP7 = 2 | 0A | 1B | 1B | ||
| Index fortheproliferative changes | IKP = 0A | IKP = 7B | IKP = 9B | |||
| Inflammation | Kidney | Infiltration | WKI1 = 2 | 0A | 0A | 0A |
| Activation of melanomacrophages | WKI2 = 2 | 0A | 3B | 3B | ||
| Index fortheinflammatory processes | IKI = 0A | IKI =6B | IKI =6B | |||
| Index fortheorgan | IK = 0A | IK=18B | IK=27C | |||
In addition, a 5-point scale was used to determine the severity of each change according to Saraiva et al. (2015) [58], as follows: (0) – no changes in the kidney structure (up to 10% of the organ structure); (1) – very mild changes (10–20% of the organ structure); (2) – mild changes (20–30%); (3) – moderate changes (30–50%); (4) – severe changes (50–80%); (5) – very severe changes (over 80%). Different superscript letters (A, B, C) indicate statistically significant differences between groups according to the Mann–Whitney U test (p < 0.05). * Necrobiosis includes karyopyknosis, karyorrhexis, and karyolysis.
Table 5.
Histopathological changes in the kidney of common carp after acute (96-hour) exposure to propamocarb hydrochloride.
Table 5.
Histopathological changes in the kidney of common carp after acute (96-hour) exposure to propamocarb hydrochloride.
| Reaction pattern |
Organ | Alteration | Importance factor | Score value | |||
|---|---|---|---|---|---|---|---|
| Control | Propamocarb hydrochloride | ||||||
| 40 μg/L | 80 μg/L | ||||||
| Changes in the circulatory system | Kidney | Haemorrhage | WKC1 = 1 | 0A | 0A | 0A | |
| Hyperaemia | WKC2 = 1 | 0A | 0A | 0A | |||
| Aneurysms | WKC3 = 1 | 0A | 0A | 0A | |||
| Index for changes in the circulatory system | IKC =0A | IKC =0A | IKC =0A | ||||
| Degenerative changes | Tubule | Vacuolar degeneration | WKR1 = 1 | 0A | 1B | 3C | |
| Hyaline degeneration | WKR2 = 1 | 0A | 1B | 2C | |||
| Necrobiosis | WKR3 = 2 | 0A | 0A | 0A | |||
| Necrosis | WKR4 = 3 | 0A | 0A | 0A | |||
| Glomerulus | Dilatation of the Bowman's capsule | WKR5 = 1 | 0A | 2B | 2B | ||
| Contraction | WKR6 = 1 | 0A | 0A | 0A | |||
| Necrobiosis* | WKR7 = 2 | 0A | 0A | 0A | |||
| Necrosis | WKR8 = 3 | 0A | 0A | 0A | |||
|
Interstitial tissue |
Necrosis | WKR9 = 3 | 0A | 0A | 0A | ||
| Index forthedegenerative changes | IKR = 0A | IKR=4B | IKR=7C | ||||
| Proliferative changes | Tubule | Hypertrophy | WKP1 = 1 | 0A | 1B | 1B | |
| Hyperplasia | WKP2 = 2 | 0A | 0A | 0A | |||
| Glomerulus | Hypertrophy | WKP3 = 1 | 0A | 1B | 1B | ||
| Hyperplasia | WKP4 = 2 | 0A | 0A | 0A | |||
| Thickening of Bowman's capsular membrane | WKP5 = 2 | 0A | 0B | 1B | |||
| Interstitial tissue | Hypertrophy | WKP6 = 1 | 0A | 0A | 0A | ||
| Edema | WKP7 = 2 | 0A | 0A | 0A | |||
| Index fortheproliferative changes | IKP = 0A | IKP =2B | IKP =4C | ||||
| Inflammation | Kidney | Infiltration | WKI1 = 2 | 0A | 0A | 0A | |
| Activation of melano-macrophages | WKI2 = 2 | 0A | 1B | 2C | |||
| Index for inflammatory processes | IKI = 0A | IKI =2B | IKI =4B | ||||
| Index for organ IK | IK = 0A | IK=8B | IK=15C | ||||
In addition, a 5-point scale was used to determine the severity of each change according to Saraiva et al. (2015) [58], as follows: (0) – no changes in the kidney structure (up to 10% of the organ structure); (1) – very mild changes (10–20% of the organ structure); (2) – mild changes (20–30%); (3) – moderate changes (30–50%); (4) – severe changes (50–80%); (5) – very severe changes (over 80%). Different superscript letters (A, B, C) indicate statistically significant differences between groups according to the Mann–Whitney U test (p < 0.05). * Necrobiosis includes karyopyknosis, karyorrhexis, and karyolysis.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



