
Submitted:
05 May 2025
Posted:
09 May 2025
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Background/Objectives: Since their approval in early 2020, mRNA vaccine has gained significant attention since COVID-19 pandemic as a potential therapeutic approach to tackle several infectious diseases. The aim of this article is to review the current state of mRNA vaccine technology and its use against other diseases. Methods: To get accurate and reliable data, we carefully search the clinicaltrial.gov, and individual company’s website for current ongoing clinical trials reports. Also, we accessed different NCBI databases for recent articles or reports of clinical trials, innovative design of mRNA vaccine and reviews. Results: Significant progress have been made in the design and improvement of mRNA vaccine technology. Currently, there are hundreds of ongoing clinical trials on mRNA vaccine against different cancer types, infectious diseases, and genetic and rare diseases which showcase the advancement in this technology and their potential therapeutic advantages over traditional vaccine platforms. Finally, we project what could be potential future direction in designing more effective mRNA vaccine, particularly against cancer. Conclusions: Results of many of the ongoing clinical trials have shown significant positive outcomes with many of the trials already at phase III. Despite this outlook however, some have been terminated or withdrawn for several reasons, some of which are not made available. This means that despite the advancement, there is need for more research and critical evaluation of each innovation to better understand their immunological benefits and long-term effects.
Keywords:
mRNA vaccine
; infectious diseases
; cancer
; personalized medicine
; Rare and genetic diseases
; Dendritic cells
; Lipid nanoparticles
; Exosomes
1. Introduction
The first mRNA therapeutic attempt was designed in 1995 to target cancer, loading the protein on dendritic cells (DCs) [1,2]. After this breakthrough research by Hsu and his colleagues, the technology of mRNA vaccine seemed to be in the shadow until the emergence of COVID-19 pandemic when mRNA vaccines were rolled out to tackle the diseases, which became the fastest vaccines to be approved during any pandemic in history. Despite this great success, the liquid nanoparticles (LNPs)-based mRNA vaccines were not without some drawbacks. Beside the need for storage and transportation under very low temperature, the associated cytokine storm that leads to many adverse immunologic effect have been reasons for serious concern [3,4]. Given their potentials however, many companies and institutions have been working to advance this technology and targeting other infectious and genetic diseases.
Some of the strategies to improve mRNA vaccines beside untranslated (UTR) and codon optimizations and nucleotide modification which were earlier used by many companies including Moderna and Pfizer in their vaccine designs, other recent approaches include, alternative delivery system such as polymer-based delivery system, exosome/peptide, ionizable lipid [5], and the use of next-generation lipid nanoparticles. Other advancements are the use of self-amplifying mRNA (sa-mRNA), thermostability, adjuvant integration, the design of personalized and targeted vaccine, and the use of dendritic cell-based mRNA vaccines which seems to have gained more attention among other strategies [6,7,8,9].
Currently, numerous clinical trials are ongoing with the use of these innovative mRNA vaccines as therapies for infectious diseases such as influenza, rabies, Zika virus, Epstein-Barr Virus (EBV), Human Metapneumovirus (hMPV), Respiratory Syncytial Virus (RSV), Herpes Simplex Virus (HSV), and HIV (see Table 1). Available clinical trial data shows significant numbers of genetic and rare diseases are being investigated for potential cure using mRNA therapeutics, and different cancer types have also been targeted with mRNA vaccine formulations (see Table 2 and 3).
2.1. The Structure and Mechanism of Action of mRNA Vaccines
mRNA vaccines contain synthetic mRNA molecules that direct the production of antigens to elicit immune responses. Structurally, the Invitro transcribed mRNA mimics the endogenous mRNA. The two main mRNA types under investigation as vaccine antigens are self-amplifying RNA and non-replicating mRNA [10]. The non-replicating (conventional) mRNA contains five components: 5ʹ cap, 5ʹ untranslated regions (UTR), an open reading frame (ORF), 3ʹ UTR, and 3ʹ end poly(A) tail. The self-amplifying mRNA contains all these components with an extra coding region in their ORF that codes for the viral replication machinery, allowing for continuous intracellular RNA amplification and subsequent increased antigen production [11]. The 5ʹcap contains 7-methylguanosine nucleoside (m7G) linked to the 5′-triphosphate bridge. The cap increases the stability of mRNA against 5ʹ exonucleolytic degradation. Next to the 5ʹcap, the 5ʹ UTR contains important regulatory areas such as the internal ribosome entry site (IRES) and ribosome binding site (RBS). These components recruit the ribosome and other translation factors to initiate and regulate translation. The open reading frame is the most important part of the vaccine as it codes for the area that is translated into the protein. The overall translation efficiency of mRNA can be maximized by selecting the right codons in this region. This can be done by replacing rarely used codons with more frequently occurring codons that encode the same amino acid residue [12,13]. The polyadenylated tail at the 3ʹ end regulates the stability and translation efficiency of the mRNA by interacting with proteins that control its degradation and translation initiation (see Figure 1). Modified nucleosides are added to enhance translation. This modification prevents recognition by the antigen-presenting cells, leading to a high level of translation and producing a significant number of needed proteins/antigens (Jain et al., 2021). The mRNA is encapsulated with lipid nanoparticles for efficient delivery into the cells.
Upon injection of the vaccine, the muscular cells take up the LNP-mRNA. It escapes the endosome, releasing the mRNA into the cytoplasm [14]. Following its release, the host ribosome recognizes it and is translated into protein/antigen. The protein can be cleaved by the proteasome into peptides or transported outside the cell by the Golgi apparatus [15]. The danger-associated signals produced by the LNP recruit antigen-presenting cells like dendritic cells and neutrophils, which present the peptides to cytotoxic T lymphocytes on the cell surface by the major histocompatibility complex (MHC) class I. MHC class II proteins can also convey released antigens on the cell surface to helper T lymphocytes after they have been absorbed by cells and broken down inside endosomes [16,17]. As a result, CD8T cells generate humoral immunity that can neutralize subsequent infections from the same pathogen and establish a rapid immune response. Antigen-specific antibodies are produced by B cells and are triggered by either direct antigen recognition at the B cell receptor (BCR) or CD4T cell assistance [13,18]. Later, these antibodies can identify the same antigen in subsequent exposures, quickly triggering an immune response that prevents infection and growth. It is this humoral defense that brings long-term vaccine-mediated immunity.
2.3. Key Advantages over Traditional Vaccine Platforms
Unlike conventional vaccines (inactivated, live-attenuated, subunit, recombinant, and other conjugate types), mRNA vaccines offer several distinct advantages. First, the mRNA vaccine does not contain the pathogen itself. Hence, there is no chance of infection after vaccination. The cell-free In vitro production also reduces safety issues such as viral contamination and cell-derived impurities commonly present in other platforms [19]. Second, since mRNA expression does not require nuclear entrance, it is safer than DNA-based vaccines and has a nearly minimal chance of random genome integration [20]. Third, its ability to encode several distinct antigens in a single formulation enables accurate and strong immune responses. It promotes humoral and cellular immune responses and induces the innate immune system. Fourth, the mRNA vaccine platform's adaptability is particularly beneficial for manufacturing since it allows for standardization of production, as changes to the encoded antigen do not affect the physical-chemical properties of the mRNA backbone (Armbruster et al., 2019). Fifth, it has a wide range of applications. mRNA technology treats infectious diseases, genetic diseases, cancer, and diabetes. Lastly, mRNA is transiently active; hence, it is readily degraded by the metabolic pathways [21].
3. Applications Beyond COVID-19
3.1. Infectious Diseases
Infectious diseases remain a significant global threat to human health, particularly in low-income countries where emerging pathogens exist without approved vaccines or widely available treatments, resulting in devastating consequences [32,33]. The ongoing burden of infectious diseases on both health and the economy has been a persistent concern, driving extensive research in vaccine development [34,35]. Given that vaccines are a powerful means of enhancing global health security, several have been created to combat the threat of infectious diseases, significantly transforming disease prevention strategies [35,36,37]. With reduced concerns regarding insertional mutagenesis [11,19], remarkable progress has been made in applying mRNA technology for both infectious and non-infectious diseases. This technology has proven effective in treating cancer and infectious diseases [38]. The therapeutic and preventive roles of mRNA technology have been explored to support the treatment of infectious diseases. Recent advancements in mRNA design, delivery systems, and production scalability have expanded its applications to oncology, autoimmune disorders, and genetic diseases. The emergence of the COVID-19 pandemic has accelerated research into mRNA vaccines for other infectious diseases, highlighting their potential as a groundbreaking platform for vaccine development. This progress is particularly evident in the enhancement of delivery systems for these vaccines [33,36,39,40]. mRNA vaccines hold promising applications for various human infectious diseases caused by pathogens such as filoviruses (including Ebola), arboviruses (like Zika, Dengue, and Chikungunya), influenza, HIV, and rabies. Recent studies have underscored the potential of mRNA vaccines against the Zika virus (ZIKV). Starting with the first-generation mRNA-1325 Zika vaccine developed by Moderna, which is based on structural proteins from a 2007 Micronesia Zika virus isolate to the triple repeat domain III mRNA vaccine, 3xEIII, encapsulated in lipid nanoparticles (LNPs), both elicited neutralizing antibodies and long-term immunity in mice [27]. Similarly, other Zika virus vaccines, including the mRNA vaccine encoding ZIKV prM-E proteins in LNPs, provided protection in AG129 mice against lethal ZIKV challenges [41]. A self-replicating mRNA vaccine encoding ZIKV prM-E proteins demonstrated efficacy in IFNAR1-/- C57BL/6 mice. However, its effectiveness was inversely related to the dose in wild-type mice due to type I interferon responses [42]. An optimized mRNA vaccine, mRNA-1893, produced comparable neutralizing antibody titers to its predecessor at a lower dose, ensuring complete protection in non-human primates [43]. The HIV vaccine development has faced extreme difficulties consequent to the challenge of the high variability of the viral envelope antigens of HIV, driving the huge genetic diversity of HIV, and the frequent escape mutations consistent with the virus [44,45]. The fact that most people infected with HIV are infected with about 20% of HIV variants further compounds the problem of successfully developing a vaccine with broad neutralising antibodies (bNAb) [46,47,48]. COVID-19 mRNA vaccine success has propelled numerous research and approaches in HIV mRNA development. Presently, the HIV-1 trimeric envelope (Env) glycoprotein (gp160/gp140) has been targeted by mRNA-1574 Moderna as a vaccine candidate, while the engineered gp120 (outer domain of Env) and stabilised HIV-1 Env gp140 trimer developed by the eOD-GT8 60mer mRNA (IAVI/Moderna) and BG505 MD39.3 mRNA (NIH/NIAID) respectively [49,50,51]. Currently, the HIV mRNA vaccines are being tested in healthy individuals and HIV-infected patients with the aim of passive immunisation (HIV mRNA Vaccine | List Results | ClinicalTrials.Gov, n.d.).
The human avian influenza (H5N1) virus has been reported in more than 23 countries since its first report and has remained a significant threat to global health, resulting in continuous seasonal economic loss. Global yearly seasonal influenza epidemics record about 3 million people developing illnesses, of which up to 600,000 died (majorly children) from influenza cases worldwide [52,53]. For a long time, embryonated chicken eggs have been the platform for the generation of influenza vaccines, although this approach presented greater scalability, effectiveness was about 60% due to the constantly changing antigenic drift and shift, limited duration of immunity, and antigenic mismatch between the selected vaccine strains and circulating influenza strains [54] since influenza mutation is seasonally influenced [55], posing a challenge for the development of a global influenza vaccine. Although the mRNA influenza vaccine targeting Hemagglutinin (HA) has long been reported [56], the most significant mRNA vaccines for influenza came after the success of the mRNA vaccine for the COVID-19 pandemic. The trivalent mRNA vaccine candidate with cross-specific humoral immune response in mice was reported by Elena et al [57]. Others in clinical trials include the ARCT-2304 encoding viral glycoproteins (HA and neuraminidase) and the mRNA-1018 mRNA vaccine developed by Arcturus Therapeutics and Moderna, respectively [58]. The recent outbreak of influenza in the US has prompted the US Department of Health and Human Services (HHS) to announce £590 million in support of Moderna to expedite the development of mRNA vaccines against potential pandemic flu viruses [58,59].
3.2. Cancer Immunology
A recent publication by Chen et al (2023) estimated the global economic cost of cancers to be at $25.2 trillion from 2020 to 2050, imposing a marked toll on the economy through reduced productivity, unemployment, labour losses, and capital investment reductions. Cancer treatments, including surgery, chemotherapy, radiation, and targeted therapies, have significantly improved patient outcomes in recent decades, but all these methods have come with limited outcomes [27,60], requiring a dare need for the alternative best option. Cancer immunotherapy’s desired outcome is to achieve long-lasting positive responses in patients, therefore, the quest to develop effective vaccines and therapy for cancer treatment has advanced lately with the introduction of several technologies and approaches in therapeutics [27,60,61]. Cancer vaccines target treatment rather than prevention, unlike vaccines for infectious diseases. Their goal is to stimulate the patient’s immune system using tumour antigens, triggering an antitumor response to help eliminate the tumour [62]. The successes of mRNA technology during COVID-19 revolutionised vaccinology and paved the way for numerous breakthroughs in disease treatment, particularly in cancer. The uniqueness of mRNA technology for cancer therapy includes the versatility and flexibility of mRNA to encode multiple antigens against diverse tumour-associated antigens (TAA), the ability to induce both humoral and cellular immune responses, leading to broader and more robust protection against cancer in terms of vaccine, the use of mRNA to encode tumour-specific antigens (TSA) allows the immune system to recognise and eliminate cancer cells displaying these antigens [27]. mRNA therapy offers numerous advantages over traditional cancer immunotherapy, primarily its ability to be used to achieve both monotherapy and synergistic effect for cancer immunotherapy, addressing the known features of tumour immune evasion and resistance, limited response rate and specificity, immune-related adverse effects [61,63]. Issues of safety, delivery methods and personalised treatments are addressed in most mRNA candidates for cancer immunotherapy [60]. Despite its advantage in inducing cross-presentation of multiple epitopes in antigen-presenting cells (APCs) with patient-specific human leukocyte antigens (HLA), reducing HLA type restrictions and boosting T cell responses, a lot of research is still required in personalised mRNA cancer vaccines. Numerous mRNA cancer vaccines are currently in clinical trials, ranging from ex vivo mRNA-based DC vaccines to in vivo mRNA cancer vaccines. Promising results from clinical trials, such as the Lipo-MERIT trial, have demonstrated the effects of mRNA vaccines targeting melanoma-associated antigens, resulting in long-lasting immune responses in patients with advanced melanoma [64]. The Phase 3 clinical study of Adjuvant V940 (mRNA-4157) with Pembrolizumab shows promising results as a personalized cancer vaccine targeting tumour neoantigens in high-risk stage II-IV melanoma patients. This treatment significantly prolongs recurrence-free survival (RFS) and reduces the risk of recurrence or death by 44% [60,65].
Prostate cancer is the leading cancer among men in sub-Saharan Africa, with increasing incidence rates and high mortality rates [66]. Recent immunotherapies target either key tumour-associated antigens (TAA) or tumour-specific antigens (TSA). Tumour-associated antigens (TAA) are the most common prostate cancer antigens. They are characterized by weak tumour immunogenicity and specificity. To address this issue, mixtures of TAAs are being utilized to enhance the efficacy of cancer vaccines [67]. TAA serves as the target for the multivalent mRNA vaccine, CV9103 (CureVac), which encodes the PSA, PSCA, PSMA, and STEAP1 antigens for the treatment of patients with advanced prostate cancer [68]. In Phase I/IIa clinical study involving 44 patients with advanced castration-resistant prostate cancer (CRPC), CV9103 was well tolerated and immunogenic, leading to an enhanced version of the vaccine, CV9104, which includes prostatic acid phosphatase (PAP) and mucin 1 (MUC1). The RNActive® prostate cancer vaccine CV9104 was evaluated in a randomised, placebo-controlled Phase II trial to determine its clinical efficacy in prolonging survival for patients with asymptomatic or minimally symptomatic castrate-resistant metastatic prostate cancer [69] However, the trial was terminated because the results showed no recent updates indicating a significant difference in overall survival (OS) between the treatment and control groups [70].
3.3. Personalised Medicine
- -
- Potential in creating individualised vaccines for patients based on genetic profiles
The Human Genome Project has set the foundation for personalised medicine by providing a comprehensive mapping of the human genome. This mapping illustrates that an individual possesses a unique genetic makeup, which is valuable information for disease prevention, prognosis, diagnosis, and treatment [71]. Personalised medicine, also known as "precision medicine," has revolutionised patient treatment by overcoming the limitations of conventional pharmacotherapies. [72]. In personalised medicine, individualised treatment can be designed based on the specific characteristics of the individual identified through genomic investigations, specifically pharmacogenetic therapies. Lately, cancer immunotherapy has harnessed the specific characteristics of each tumour, developing vaccines and treatments with personalised approaches. mRNA vaccines have become a groundbreaking technology in oncology and personalised medicine, showing great potential for treating cancer. Their quick production and adaptability significantly advance immunotherapy [73]. mRNA vaccines targeting either key tumour-associated antigens (TAA) or tumour-specific antigens (TSA) have been developed [70,74]. and have shown promising results. Tumour-specific antigens (TSA) or neoantigens provide a robust vaccination, providing better tumour specificity, and are not known to generate autoimmunity [75]. Because of their uniqueness and specificity to cancer cells, rationally, ongoing clinical trial outcomes have favoured more TSA vaccine development [60,76]. Most cancer mrna personalised vaccines are TSA-based. TSA-based vaccines combine large-scale sequencing, bioinformatics algorithms, and patient immunology. Recently, Merck and Moderna developed an individualised neoantigen therapy (INT) approach for treating patients with resected high-risk melanoma with mRNA-4157 (V940) in combination with pembrolizumab [77]. This approach demonstrated clinically meaningful improvements in both recurrence-free survival and distant metastasis-free survival (62% reduction in the risks of distant metastasis and death) compared to pembrolizumab monotherapy, a contrast to the traditional checkpoint inhibitors with evidence of melanoma recurrence [77,78]. The mRNA-4157 (V940) is in a phase III clinical trial stage. BioNTech recently published three-year follow-up data from a Phase 1 trial, which showed that 8 out of 16 patients exhibited sustained polyspecific T cell responses in patients with resected pancreatic ductal adenocarcinoma. This was associated with a delayed recurrence of tumours over three years following treatment with their mRNA (iNeST) or BNT122 vaccine, developed in collaboration with Genentech [79], encouraging the initiation of a phase II trial [74]. Furthermore, Gritstone Bio’s neoantigen cancer vaccine GRANITE (GRT-C901/GRT-R902) phase 2/3 study indicates a potential for clinical benefits for colorectal cancers in combination with immune checkpoint inhibitors [80]. Other personalised mrna vaccines, such as Frame Therapeutic FRAME-001, in a phase II trial for the treatment of III-IV non-small cell lung cancer [81]. Indeed, the emergence of mRNA vaccine technology represents a significant shift in personalised cancer immunotherapy. Continual advances in high-throughput sequencing, neoantigen prediction, and nanocarrier systems sustain this strategy. Also, combining these vaccines with immune checkpoint inhibitors enhances their efficacy against tumour immune evasion. As these technologies improve, pharmacogenomics is expected to become cost-effective and more practical, promoting wider adoption of precision immuno-oncology in the future.
4. Innovations in mRNA Vaccine Designs
The technology of mRNA therapeutics has been in existence over the past few decades, but not until during the COVID19 pandemic that its fame spread across after Pfizer and Moderna announced their mRNA vaccine for COVID-19. Stemirna Therapeutics is a mRNA-based technology company, established in 2016 to maximize the potential of RNA science. In spite of the successes recorded with the use of LNP for mRNA vaccine delivery during COVID19, the technology faces many limitations. They are sensitive to temperature fluctuations and require ultra-cold storage (between -20 to -80oC). Some LNP components can trigger innate immune responses leading to inflammation and adverse reactions. For a detailed review on current strategies to reduce the risks of mRNA vaccine toxicity, we recommend [82]. Following the development and successful delivery of mRNA COVID-19 vaccine using lipid nanoparticles (LNP), there have been several attempts to improve and to design new platforms that offers efficient and more targeted delivery of vaccines with reduced immune toxicity.
4.1. Advances in Lipid Nanoparticle (LNP) Delivery Systems
Lipid nanoparticles (LNP) are non-viral vectors that have gained wide application as efficient delivery tools. They have low cytotoxicity and immunogenicity, high tissue penetration, high nucleic acid encapsulation, and efficient transfection with good biocompatibility [22]. Structurally, LNP consists of phospholipids, cholesterol, and polyethylene glycol (PEG) containing lipids. Phospholipids improve stability, enhance ease of mRNA encapsulation, and promote cellular absorption. Ionizable lipids have unique pH-responsive qualities that allow them to become positively charged in acidic environments, like the endosomes, and stay neutral during systemic circulation [23]. They are neutral at physiological pH, encouraging stable LNP production, prolonging bloodstream circulation, and inhibiting immune cell clearance. In the acidic environment, they become protonated (positively charged) and directly interact with the negatively charged phospholipids, promoting the release of cargo into the cytoplasm and aiding in the rupture of the endosomal membrane. Cholesterol stabilizes LNP structure, regulates the fluidity and permeability of membranes, and improves particle stability [24,25].
Lipid nanoparticles are classified into four groups: liposomes, solid lipid nanoparticles (SLNs), nanostructured lipid carriers (NLCs), and hybrid lipid-polymeric nanoparticles. Liposomes are the early version of LNPs. They contain a phospholipid bilayer, which is typically made of phosphatidylcholine (PC) or other phospholipids, which self-assemble into bilayers due to their amphiphilic nature, the aqueous core and hydrocarbon chain [2,26]. The aqueous interior of liposomes contains hydrophilic drugs, as the hydrocarbon chain portion of the lipid bilayer entraps hydrophobic drugs. Its size ranges between 20-1000nm. Liposomes can be synthesized as unilamellar or multilamellar. Small Unilamellar Vesicles (SUVs): 20–100 nm consists of a single bilayer. Large Unilamellar Vesicles (LUVs): 100–500 nm, have a single bilayer but larger encapsulation volume. Multilamellar Vesicles (MLVs): 0.1–10 µm, composed of multiple concentric bilayers. The size determines the half-life and drug encapsulation of the liposome. The small unilamellar liposomes exhibit higher encapsulation efficiency, Increased drug half-life, and the ability to evade the immune system [27]. Liposomes are challenging to produce on a large scale, they require complex production using organic solvents. Solid and nanostructured lipid nanoparticles are produced to overcome these challenges.
SLNs are solid colloidal particles that range in size from 4 to 1000 nm. They contain physiological, biodegradable, and biocompatible lipids and surfactants in their formulation, which can incorporate both lipophilic and hydrophilic drugs inside the lipid matrix [24,28]. The solid lipid serves as the dispersed phase, while the surfactant acts as an emulsifier. The solid lipid is typically made from triglycerides, glyceride mixtures, or even waxes, and it stays solid at room and body temperature; the surfactant concentration range is usually 0.5 to 5% (w/v) to improve formulation stability [2,29]. The selection of lipids and surfactants significantly impacts the nanoparticle formulation's physicochemical properties. The structure of SLN is determined by the production method and the solubility of the component and component formulation. Three structural types have been described: the homogenous matrix model, the drug-enriched shell, and the drug-enriched core. A homogenous matrix model is used for highly lipophilic drugs, the particles are created using a cold or hot homogenization process. The drug is either dissolved in a lipid matrix, a high-pressure homogenization process, and mechanical breakings leading to nanoparticle formation, or the lipid is dissolved in a lipid matrix with increased temperatures, leading to nanoparticle formation. It offers slow and controlled drug release. The drug-enriched shell is produced by hot homogenization as the system cools down the lipid precipitate, forming the lipid core. The concentration of the drug increases until its solubility limit is reached. When this limit is reached, the mix of drug and melted lipid crystalizes to form an outer shell. It results in a faster release of drugs. In the drug-enriched core, the drug crystallizes before the lipid and gets trapped in the core, with a pure lipid shell forming around it. Occurs when the drug has higher solubility in the molten lipid but separates during solidification [25,29]. Unlike liposomes, SLNs have prolonged release and better stability. However, the solid lipid's crystalline structure causes its poor integration rate, and it tends to gelate. NLCs are developed to overcome the disadvantages of SLNs.
NLCs are modified forms of SLNs that contain a mixture of solid and liquid phases (oil), forming a formless matrix, improving stability and capacity loading [22,27,28]. The lipids used here are biologically compatible, hence reducing toxicity. Depending on their structure, they can be classified into three: The imperfect crystal type comprises lipids with different chain lengths forming a matrix with several voids and imperfections that provide a more suitable environment for drug incorporation. The amorphous type is obtained using medium chain-length triglycerides and solid lipids. The solid lipids do not undergo recrystallization, thus resulting in solid particles with an amorphous structure. The drug's unwanted release is reduced, extending its shelf life. The multiple LNC has nanocompartments of oil containing dissolved drugs [27,30,31].
Methods like high-pressure homogenization, ultrasonication, spraying dying, solvent injection, double emulsion, and microfluidics are used to prepare lipid nanoparticles. Microfluidic is the recent form that leads to the production of nanoparticles with uniform particles; it is a promising approach for the large-scale production of SLNs and NLCs [24,25].
4.2. Improvement in Delivery Systems
The FDA-approved COVID-19 vaccines (Pfizer-BioNTech and Moderna) utilized LNPs with proprietary lipids like ALC-0315 (Pfizer) and SM-102 (Moderna) [23]. These LNPs used ionizable lipids that self-assemble with mRNA at acidic pH which promotes endosomal escape. Beside LNP, platforms such as improved ionized lipids, polymer-based delivery systems, peptide-based delivery, and exosome and extracellular vesicle (EV) delivery are being investigated to bypass LNPs limitations. A 2022 study conducted by Kamerker and her colleagues demonstrated engineered exosomes delivered mRNA to pancreatic cancer cells with higher efficiency [83].
While mRNA vaccine for COVID-19 remains one of the greatest achievements in modern medicine, the idea of using this concept for cancer started over 20 years ago when Hsu and his colleagues demonstrated for the first time, the potential of using autologous tumor-specific antigen loaded on dendritic cells to induce antitumor immune response in cancer patients [1]. This groundbreaking clinical trial of using DCs for vaccine in cancer, laid a crucial foundation for what has now become a golden approach, and many researchers now focused on the use of monocyte-derived DCs (moDC) generated in vitro for mRNA vaccine production. The efficacy of this type of DCs have led to the approval of Sipuleucel-T (a DC-based vaccine) for prostate cancer by the FDA. Recently, however, due to extensive culture periods and supplements required to produce mature moDCs, many researchers have sought for alternative ways of generating DCs. Myeloid DC (mDC) and plasmacytoid DC (pDC) are two alternatives that have shown promising immunologic and clinical outcomes in metastatic melanoma patients [7,9], and lung cancer [6]. pDCs have also been shown as promising tools for multispecific viral and tumor antigen-specific T cells [8]. For a comprehensive review on the use of dendritic-based vaccine in immunotherapy, we refer readers to a recent review on this subject [84]. Furthermore, to enhance targeted delivery, LNPs are now being conjugated with antibodies or peptides to enable cell-specific delivery. For example, targeting dendritic cells via CD40 ligands enhances antigen presentation, as shown in cancer vaccine trials [85]. Pulsing DCs with human cytomegalovirus (CMV) pp65-LAMP (lysosomal-associated membrane protein) mRNA to target gliomas is currently being investigated in the clinical trial (NCT03688178), and it will be interesting to know the outcome of the trial. This cytomegalovirus unique protein has been reported in significant amount of people suffering from malignant gliomas (MGs), and targeting the proteins within cancer microenvironment provide alternative therapeutic potential to gliomas and this approach might be relevant to other cancer types.
4.3. Strategies for Enhancing Stability and Immunogenicity
As already mentioned, one of the challenges of LNPs is the need for extremely low temperature for their storage and transportation. Hence, there is need for improved and higher storage condition.
The technology involved in the development of ARCT-154 (Kostaive) vaccine by CSL and Arcturus Therapeutics that was given approval in Japan includes addition of an active substance, zapomeran which is a self-amplifying messenger RNA (mRNA). This self-amplifying mRNA (also called sa-mRNA) has an instructional mechanism for making an enzyme replicase, which in turn helps to make additional copies of mRNA upon delivery. Unlike other approved COVID-19 vaccines, Kostaive can be kept at higher temperature of about 2oC. Furthermore, development of this sa-mRNA vaccine is reported to significantly reduce immunogenicity as low doses are required to stimulate potent cellular immune responses. This will effectively reduce immune toxicity and other adverse effect associated with other COVID-19 vaccine. Also, mRNA1283, a next-generation COVID vaccine developed by Moderna which is currently in phase 3 trial, and it can be kept at a temperature of about 2-5oC [86]. Also, addition of adjuvants to the mRNA vaccine is another notable innovation that would enhance vaccine’s protection. Co-delivering immune stimulants (for example, TLR agonists) enhances will improve responses. BioNTech’s FixVac platform incorporates uridine-rich mRNA to activate TLR7/8, boosting T-cell immunity in melanoma trials [64].
4.4. Multivalent Vaccine Designs for Multiple Pathogens or Cancer Antigens
Viruses are known for their unique ability to mutate rapidly. This variability within the same species of viruses reduces the ability of monovalent vaccines to protect against different serotypes of some viruses of global significance. The emergence of variants of concerns (VOCs) necessitates the proposal for booster doses for COVID-19 pandemic. Another example is influenza virus. One of the major challenges for the development of vaccine for influenza virus is the multiple subtypes with varying degree of mismatches. Following recent advances in nucleic acid-based vaccine however, Arevalo and colleagues have developed a multivalent vaccine that encodes all the 20 known subtypes [87]. There are applications of this method also in cancer immunotherapy.
Currently, there are numerous ongoing clinical trials that are investigating the potential and immunogenic properties of several mRNA vaccines encoding tumor-specific neoantigen (see Table 2) to specific cancer. Personalized neoantigen vaccines (for example, BioNTech’s BNT111) target patient-specific tumor mutations. A 2021 trial by Sahin et al. reported prolonged survival in melanoma patients [88].
5. Current Challenges
5.1. Manufacturing and Scale-Up
- -
- Challenges in production and supply chain scalability.
Over the last decade and a half period, mRNA vaccine development has undergone significant progress. The emergence of the Coronavirus disease 2019 (COVID-19) pandemic put the mRNA technology for vaccines to a test and this was a success story with the first production of mRNA vaccines in less than a year of the pandemic [89]. This has resulted in a significant reduction in the time of production, cost, and ease of vaccine production leading to a more rapid response to disease outbreaks as demonstrated during the COVID-19 pandemic. There have been efforts to improve the technology further as demand for mRNA vaccines continues to increase. Wei et al worked on the development of a universal integrated platform aimed at streamlining and enhancing on-demand preparation [90]. More advances in artificial intelligence have been propelling mRNA technology further, however, some challenges may persist especially with differential global capacity.
- -
- Addressing regional manufacturing gaps.
Vaccine production capacity is limited in most LMICs, for example, there are 11 vaccine manufacturing facilities in Africa but their production capacities could hardly meet up to 1% vaccine for the over 50 countries of the continent. Significant deficiencies in infrastructure, technology, and human capital [91]. Conscious efforts and deliberate investment will be required to reverse this and the African Union (AU) should lead the continental efforts.
5.2. Cost and Accessibility
Generally, the production of mRNA vaccines has been documented to be cheaper when compared with traditional vaccine production. mRNA vaccines possess the advantage of fast manufacturing as short as half a year in contrast to the traditional method which may take 10-15 years. Notwithstanding these forgoing merits, the mRNA vaccine requires higher capital and starting materials. This becomes more significant in the low-and-middle-income countries that do not have adequate resources (material and human) to compete favorably with countries that are well-developed [92]. This challenge may further worsen the vaccine inequality that characterized COVID-19 vaccination at the peak of the pandemic between the global north and south. For example, Africa only supplies 1% of global vaccines [93]. The implication of this is that Africa which constitutes almost 20% of the world will have to continue to depend on other nations to meet its vaccine needs.
- -
- High costs of mRNA technology compared to traditional vaccines.
There should be concerted efforts to ensure global vaccine equity. This could be led by the World Health Organization and other vaccine multinational groups like GAVI, the vaccine alliance. This may include but is not limited to technology transfer and license sharing. Health system strengthening and improvement is another important factor in vaccine equity especially in many low-and-middle-income countries [94].
- -
- Strategies for reducing costs and ensuring global equity.
Global equity for vaccines is not just right but is in the best interest of global health. It is important to learn from the lesson of COVID-19 vaccine nationalism and other barriers to the equitable distribution of mRNA vaccines especially in LMIC. For example, local manufacturing of vaccines in Africa requires urgent attention and investment. Africa consumes one-quarter of the global vaccines but 99% of these vaccines are produced outside of the continent. Scaling up local capacities for vaccine manufacturing will reverse this. The steps to take will include technology transfer, regulatory system strengthening, training, and workforce development [91]. In the long term, these efforts will drastically reduce the cost of vaccines and ensure global equity.
5.3. Safety Concerns and Public Acceptance
- -
- Addressing adverse effects (e.g., myocarditis).
The novelty of SARS-CoV-2 at the start of COVID-19 led to many uncertainties not just about the virus but also about the introduction of the COVID-19 vaccine which was developed at an unprecedented speed of less than a year. The implementation and emergency authorization of the new technology of utilization of mRNA vaccine in humans for the first time raised many concerns. One such concern was the adverse events that could be associated with the new vaccines. While overall the mRNA vaccines for COVID-19 have been shown to be safe, there was a report of unusual adverse events. In a multinational study of over 99 million participants with combined doses of approximately 220 million of two COVID-19 mRNA vaccines (BNT162b2 and mRNA-1273), there was confirmation of pre-established safety signals relating to the cardiovascular system (myocarditis and pericarditis), neurological system (acute disseminated encephalomyelitis) and the identification of more safety signals including facial Bell`s palsy, febrile and generalized seizure, idiopathic thrombocytopenia, cerebral venous sinus thrombosis, splanchnic vein thrombosis, and pulmonary embolism [95]. It is important to further assess these findings in association studies rather than observational cohorts and to determine the clinical significance and implications. In another trial setback, the FDA has put on hold all RSV vaccine trials in small children citing an increase in severe illness following the negative outcome of clinical trial conducted by Moderna among children aged 2-5 years old [96]. Furthermore, two clinical trials on DC-based mRNA vaccine for cancers were terminated by Oslo University Hospital. However, no information has been provided for these terminations.
- -
- Public skepticism in the wake of misinformation.
The global mobilization of human and material resources to tackle the COVID-19 pandemic faced the challenge of misinformation and disinformation resulting in public skepticism, vaccine hesitancy, and anti-vaccine ideologies. Some of the related skepticism include mutation or genetic changes posed by the vaccines, induction of autism, the vaccines were chips, and that they could cause injury and death [97]. Social media were instrumental to the spread of misinformation and disinformation on the vaccines. The WHO and other global and local health authorities however rose to the occasion to address this challenge.
6. Regulatory and Ethical Considerations
- -
- Regulatory landscape for mRNA vaccines beyond COVID.
The application of mRNA vaccines has expanded beyond their use in infectious diseases, there are various mRNA technology uses in immunotherapy and cancer. Regulatory oversight will be very critical to ensure consistent ethical application of the rapidly evolving technology. There are no formal regulatory guidelines that are specific to mRNA vaccines according to the World Health Organization although key regulations for manufacturing and quality control are in place. Similarly, no regulatory guideline exists in the European Union while the United Kingdom and United States regulation is based on the Human Medicines Regulation of 2012 and the Food and Drug Administration (FDA) respectively [98]. These are the same with regulatory bodies in other part of the world. This is an important gap that needs to be addressed and tackled on time.
- -
- Ethical concerns related to personalized mRNA vaccines.
The application of personalized mRNA cancer vaccine is becoming a reality and the issue of associated issues such as cost and equitable access, data privacy, and potential unintended side effects especially in the long-term. The concern of vaccine nationalism as it was with the COVID-19 vaccine may also resurface here [99]. The issue of technology transfer to LMIC given the enormous resources that are associated with the development of these vaccines. All these and other related matters will require attention.
7.0. Future Direction
The current trend of designing multivalent mRNA vaccines for infectious diseases is a laudable approach that will help to protect millions of people across the world. Considering the number of ongoing clinical trials involving the use of dendritic cell-based mRNA vaccine, especially against cancers, one other approach that will be worth exploring is to load tumor-specific neoantigen mRNA on multiple combined dendritic cells (for example pDC and mDC). This will most likely increase immunological response by activating more MHC molecules, and invariably, activation of cytotoxic T cells to target cancer cells (see Figure 3). Beside activating MHC I and II, DCs can provide signals for co-stimulatory molecules and cytokine secretion ensuring specific T cell activation [100]. Moreover, these DCs are considered to be relatively easy to propagate ex vivo. One important tool that will be valuable in the design of more targeted and personalized mRNA vaccine is the use of AI.
8.0. Conclusion
It is encouraging to see the level of progress that have been made over the past few years since the emergence of mRNA vaccine. Despite public skepticism, the available clinical trials data reveal a profound justification for continuous research and collaboration to addressing global health challenges. With increasing number of clinical trials in the use of mRNA vaccines, particularly, not just against infectious diseases but also cancer, this technology will usher in a new era in vaccinology. The collaboration between companies to optimize different structural components of the vaccine should be replicated among universities and institutions’ mRNA experts in order to overcome current challenges facing this area of research. Finally, it would be desirable to see more funding support from government and all stakeholders to ensure equity and global distribution of this technology to people who need them the most.
References
- Hsu FJ, Benike C, Fagnoni F, Liles TM, Czerwinski D, Taidi B, et al. Vaccination of patients with B–cell lymphoma using autologous antigen–pulsed dendritic cells. Nat Med 1996;2:52–8. [CrossRef]
- Hou X, Zaks T, Langer R, Dong Y. Lipid nanoparticles for mRNA delivery. Nat Rev Mater 2021;6:1078–94. [CrossRef]
- Igyártó BZ, Qin Z. The mRNA-LNP vaccines – the good, the bad and the ugly? Front Immunol 2024;15:1336906. [CrossRef]
- Iqbal Z, Rehman K, Mahmood A, Shabbir M, Liang Y, Duan L, et al. Exosome for mRNA delivery: strategies and therapeutic applications. J Nanobiotechnology 2024;22:395. [CrossRef]
- Han X, Zhang H, Butowska K, Swingle KL, Alameh M-G, Weissman D, et al. An ionizable lipid toolbox for RNA delivery. Nat Commun 2021;12:7233. [CrossRef]
- Hannani D, Leplus E, Laurin D, Caulier B, Aspord C, Madelon N, et al. A New Plasmacytoid Dendritic Cell-Based Vaccine in Combination with Anti-PD-1 Expands the Tumor-Specific CD8+ T Cells of Lung Cancer Patients. Int J Mol Sci 2023;24:1897. [CrossRef]
- Charles J, Chaperot L, Hannani D, Bruder Costa J, Templier I, Trabelsi S, et al. An innovative plasmacytoid dendritic cell line-based cancer vaccine primes and expands antitumor T-cells in melanoma patients in a first-in-human trial. OncoImmunology 2020;9:1738812. [CrossRef]
- Lenogue K, Walencik A, Laulagnier K, Molens J-P, Benlalam H, Dreno B, et al. Engineering a Human Plasmacytoid Dendritic Cell-Based Vaccine to Prime and Expand Multispecific Viral and Tumor Antigen-Specific T-Cells. Vaccines 2021;9:141. [CrossRef]
- Perez CR, De Palma M. Engineering dendritic cell vaccines to improve cancer immunotherapy. Nat Commun 2019;10:5408. [CrossRef]
- Schlake T, Thess A, Fotin-Mleczek M, Kallen K-J. Developing mRNA-vaccine technologies. RNA Biol 2012;9:1319–30. [CrossRef]
- Pardi N, Hogan MJ, Weissman D. Recent advances in mRNA vaccine technology. Curr Opin Immunol 2020;65:14–20. [CrossRef]
- Chaudhary N, Weissman D, Whitehead KA. mRNA vaccines for infectious diseases: principles, delivery and clinical translation. Nat Rev Drug Discov 2021;20:817–38. [CrossRef]
- Jin L, Zhou Y, Zhang S, Chen S-J. mRNA vaccine sequence and structure design and optimization: Advances and challenges. J Biol Chem 2025;301:108015. [CrossRef]
- Iavarone C, O’hagan DT, Yu D, Delahaye NF, Ulmer JB. Mechanism of action of mRNA-based vaccines. Expert Rev Vaccines 2017;16:871–81. [CrossRef]
- Bettini E, Locci M. SARS-CoV-2 mRNA Vaccines: Immunological Mechanism and Beyond. Vaccines 2021;9:147. [CrossRef]
- Cagigi A, Loré K. Immune Responses Induced by mRNA Vaccination in Mice, Monkeys and Humans. Vaccines 2021;9:61. [CrossRef]
- Gergen J, Petsch B. mRNA-Based Vaccines and Mode of Action. In: Yu D, Petsch B, editors. MRNA Vaccines, vol. 440, Cham: Springer International Publishing; 2020, p. 1–30. [CrossRef]
- Ramachandran S, Satapathy SR, Dutta T. Delivery Strategies for mRNA Vaccines. Pharm Med 2022;36:11–20. [CrossRef]
- Pardi N, Hogan MJ, Porter FW, Weissman D. mRNA vaccines — a new era in vaccinology. Nat Rev Drug Discov 2018;17:261–79. [CrossRef]
- Rosa SS, Prazeres DMF, Azevedo AM, Marques MPC. mRNA vaccines manufacturing: Challenges and bottlenecks. Vaccine 2021;39:2190–200. [CrossRef]
- Sahin U, Karikó K, Türeci Ö. mRNA-based therapeutics — developing a new class of drugs. Nat Rev Drug Discov 2014;13:759–80. [CrossRef]
- Swetha K, Kotla NG, Tunki L, Jayaraj A, Bhargava SK, Hu H, et al. Recent Advances in the Lipid Nanoparticle-Mediated Delivery of mRNA Vaccines. Vaccines 2023;11:658. [CrossRef]
- Schoenmaker L, Witzigmann D, Kulkarni JA, Verbeke R, Kersten G, Jiskoot W, et al. mRNA-lipid nanoparticle COVID-19 vaccines: Structure and stability. Int J Pharm 2021;601:120586. [CrossRef]
- Parhi R, Suresh P. Preparation and Characterization of Solid Lipid Nanoparticles-A Review. Curr Drug Discov Technol 2012;9:2–16. [CrossRef]
- Viegas C, Patrício AB, Prata JM, Nadhman A, Chintamaneni PK, Fonte P. Solid Lipid Nanoparticles vs. Nanostructured Lipid Carriers: A Comparative Review. Pharmaceutics 2023;15:1593. [CrossRef]
- Yang L, Gong L, Wang P, Zhao X, Zhao F, Zhang Z, et al. Recent Advances in Lipid Nanoparticles for Delivery of mRNA. Pharmaceutics 2022;14:2682. [CrossRef]
- Wang J, Ding Y, Chong K, Cui M, Cao Z, Tang C, et al. Recent Advances in Lipid Nanoparticles and Their Safety Concerns for mRNA Delivery. Vaccines 2024;12:1148. [CrossRef]
- Naseri N, Valizadeh H, Zakeri-Milani P. Solid Lipid Nanoparticles and Nanostructured Lipid Carriers: Structure, Preparation and Application. Adv Pharm Bull 2015;5:305–13. [CrossRef]
- Xu L, Wang X, Liu Y, Yang G, Falconer RJ, Zhao C-X. Lipid Nanoparticles for Drug Delivery. Adv NanoBiomed Res 2022;2:2100109. [CrossRef]
- Akbari J, Saeedi M, Ahmadi F, Hashemi SMH, Babaei A, Yaddollahi S, et al. Solid lipid nanoparticles and nanostructured lipid carriers: a review of the methods of manufacture and routes of administration. Pharm Dev Technol 2022;27:525–44. [CrossRef]
- Yoon G, Park JW, Yoon I-S. Solid lipid nanoparticles (SLNs) and nanostructured lipid carriers (NLCs): recent advances in drug delivery. J Pharm Investig 2013;43:353–62. [CrossRef]
- Klepac P, Funk S, Hollingsworth TD, Metcalf CJE, Hampson K. Six challenges in the eradication of infectious diseases. Epidemics 2015;10:97–101. [CrossRef]
- Parhiz H, Atochina-Vasserman EN, Weissman D. mRNA-based therapeutics: looking beyond COVID-19 vaccines. The Lancet 2024;403:1192–204. [CrossRef]
- Keddy KH, Gobena T. The continuing challenge of infectious diseases. Lancet Infect Dis 2024;24:800–1. [CrossRef]
- Rodrigues CMC, Plotkin SA. Impact of Vaccines; Health, Economic and Social Perspectives. Front Microbiol 2020;11:1526. [CrossRef]
- Khormi AHI, Qohal RMM, Masrai AYA, Hakami KHH, Ogdy JA, Almarshad AA, et al. Emerging Trends in mRNA Vaccine Technology: Beyond Infectious Diseases. Egypt J Chem 2024;67:1567–74. [CrossRef]
- Tripathi T. Advances in vaccines: revolutionizing disease prevention. Sci Rep 2023;13:11748, s41598-023-38798-z. [CrossRef]
- Chaudhary N, Weissman D, Whitehead KA. mRNA vaccines for infectious diseases: principles, delivery and clinical translation. Nat Rev Drug Discov 2021;20:817–38. [CrossRef]
- Overmars I, Au-Yeung G, Nolan TM, Steer AC. mRNA vaccines: a transformative technology with applications beyond COVID -19. Med J Aust 2022;217:71–5. [CrossRef]
- Szabó GT, Mahiny AJ, Vlatkovic I. COVID-19 mRNA vaccines: Platforms and current developments. Mol Ther 2022;30:1850–68. [CrossRef]
- Medina-Magües LG, Gergen J, Jasny E, Petsch B, Lopera-Madrid J, Medina-Magües ES, et al. mRNA Vaccine Protects against Zika Virus. Vaccines 2021;9:1464. [CrossRef]
- Yu X, Wei L, Chen H, Niu X, Dou Y, Yang J, et al. Development of Colloidal Gold-Based Immunochromatographic Assay for Rapid Detection of Goose Parvovirus. Front Microbiol 2018;9:953. [CrossRef]
- Bollman B, Nunna N, Bahl K, Hsiao CJ, Bennett H, Butler S, et al. An optimized messenger RNA vaccine candidate protects non-human primates from Zika virus infection. Npj Vaccines 2023;8:58. [CrossRef]
- Berkley SF, Koff WC. Scientific and policy challenges to development of an AIDS vaccine. The Lancet 2007;370:94–101. [CrossRef]
- Morris L. mRNA vaccines offer hope for HIV. Nat Med 2021;27:2082–4. [CrossRef]
- Ahmed Y, Tian M, Gao Y. Development of an anti-HIV vaccine eliciting broadly neutralizing antibodies. AIDS Res Ther 2017;14:50. [CrossRef]
- Barouch DH. Challenges in the development of an HIV-1 vaccine. Nature 2008;455:613–9. [CrossRef]
- Boomgarden AC, Upadhyay C. Progress and Challenges in HIV-1 Vaccine Research: A Comprehensive Overview. Vaccines 2025;13:148. [CrossRef]
- Ahmed S, Herschhorn A. mRNA-based HIV-1 vaccines. Clin Microbiol Rev 2024;37:e00041-24. [CrossRef]
- Khalid K, Padda J, Khedr A, Ismail D, Zubair U, Al-Ewaidat OA, et al. HIV and Messenger RNA Vaccine. Cureus 2021. [CrossRef]
- Moderna. Moderna Announces First Participant Dosed in Phase 1 Study of its HIV Trimer mRNA Vaccine. n.d.
- Iuliano AD, Roguski KM, Chang HH, Muscatello DJ, Palekar R, Tempia S, et al. Estimates of global seasonal influenza-associated respiratory mortality: a modelling study. The Lancet 2018;391:1285–300. [CrossRef]
- Paules C, Subbarao K. Influenza. The Lancet 2017;390:697–708. [CrossRef]
- Gouma S, Anderson EM, Hensley SE. Challenges of Making Effective Influenza Vaccines. Annu Rev Virol 2020;7:495–512. [CrossRef]
- Becker T, Elbahesh H, Reperant LA, Rimmelzwaan GF, Osterhaus ADME. Influenza Vaccines: Successes and Continuing Challenges. J Infect Dis 2021;224:S405–19. [CrossRef]
- Fleeton MN, Chen M, Berglund P, Rhodes G, Parker SE, Murphy M, et al. Self-Replicative RNA Vaccines Elicit Protection against Influenza A Virus, Respiratory Syncytial Virus, and a Tickborne Encephalitis Virus. J Infect Dis 2001;183:1395–8. [CrossRef]
- Mazunina EP, Gushchin VA, Kleymenov DA, Siniavin AE, Burtseva EI, Shmarov MM, et al. Trivalent mRNA vaccine-candidate against seasonal flu with cross-specific humoral immune response. Front Immunol 2024;15:1381508. [CrossRef]
- Leonard A, Weiss MJ. Hematopoietic stem cell collection for sickle cell disease gene therapy. Curr Opin Hematol 2024;31:104–14. [CrossRef]
- Hatta M, Hatta Y, Choi A, Hossain J, Feng C, Keller MW, et al. An influenza mRNA vaccine protects ferrets from lethal infection with highly pathogenic avian influenza A(H5N1) virus. Sci Transl Med 2024;16:eads1273. [CrossRef]
- Yaremenko AV, Khan MM, Zhen X, Tang Y, Tao W. Clinical advances of mRNA vaccines for cancer immunotherapy. Med 2025;6:100562. [CrossRef]
- Zhang G, Tang T, Chen Y, Huang X, Liang T. mRNA vaccines in disease prevention and treatment. Signal Transduct Target Ther 2023;8:365. [CrossRef]
- Bidram M, Zhao Y, Shebardina NG, Baldin AV, Bazhin AV, Ganjalikhany MR, et al. mRNA-Based Cancer Vaccines: A Therapeutic Strategy for the Treatment of Melanoma Patients. Vaccines 2021;9:1060. [CrossRef]
- Wang B, Pei J, Xu S, Liu J, Yu J. Recent advances in mRNA cancer vaccines: meeting challenges and embracing opportunities. Front Immunol 2023;14:1246682. [CrossRef]
- Sahin U, Oehm P, Derhovanessian E, Jabulowsky RA, Vormehr M, Gold M, et al. An RNA vaccine drives immunity in checkpoint-inhibitor-treated melanoma. Nature 2020;585:107–12. [CrossRef]
- Merck Sharp & Dohme LLC. A Clinical Study of V940 Plus Pembrolizumab in People With High-Risk Melanoma (V940-001). n.d.
- Seraphin TP, Joko-Fru WY, Kamaté B, Chokunonga E, Wabinga H, Somdyala NIM, et al. Rising Prostate Cancer Incidence in Sub-Saharan Africa: A Trend Analysis of Data from the African Cancer Registry Network. Cancer Epidemiol Biomarkers Prev 2021;30:158–65. [CrossRef]
- Zhang G, Tang T, Chen Y, Huang X, Liang T. mRNA vaccines in disease prevention and treatment. Signal Transduct Target Ther 2023;8:365. [CrossRef]
- Kübler H, Scheel B, Gnad-Vogt U, Miller K, Schultze-Seemann W, Vom Dorp F, et al. Self-adjuvanted mRNA vaccination in advanced prostate cancer patients: a first-in-man phase I/IIa study. J Immunother Cancer 2015;3:26. [CrossRef]
- Varaprasad GL, Gupta VK, Prasad K, Kim E, Tej MB, Mohanty P, et al. Recent advances and future perspectives in the therapeutics of prostate cancer. Exp Hematol Oncol 2023;12:80. [CrossRef]
- Lin G, Elkashif A, Saha C, Coulter JA, Dunne NJ, McCarthy HO. Key considerations for a prostate cancer mRNA vaccine. Crit Rev Oncol Hematol 2025;208:104643. [CrossRef]
- Carrasco-Ramiro F, Peiró-Pastor R, Aguado B. Human genomics projects and precision medicine. Gene Ther 2017;24:551–61. [CrossRef]
- Goetz LH, Schork NJ. Personalized medicine: motivation, challenges, and progress. Fertil Steril 2018;109:952–63. [CrossRef]
- Lin F, Lin EZ, Anekoji M, Ichim TE, Hu J, Marincola FM, et al. Advancing personalized medicine in brain cancer: exploring the role of mRNA vaccines. J Transl Med 2023;21:830. [CrossRef]
- Fu Q, Zhao X, Hu J, Jiao Y, Yan Y, Pan X, et al. mRNA vaccines in the context of cancer treatment: from concept to application. J Transl Med 2025;23:12. [CrossRef]
- Liu Y, Yan Q, Zeng Z, Fan C, Xiong W. Advances and prospects of mRNA vaccines in cancer immunotherapy. Biochim Biophys Acta BBA - Rev Cancer 2024;1879:189068. [CrossRef]
- Hu C, Bai Y, Liu J, Wang Y, He Q, Zhang X, et al. Research progress on the quality control of mRNA vaccines. Expert Rev Vaccines 2024;23:570–83. [CrossRef]
- Weber JS, Carlino MS, Khattak A, Meniawy T, Ansstas G, Taylor MH, et al. Individualised neoantigen therapy mRNA-4157 (V940) plus pembrolizumab versus pembrolizumab monotherapy in resected melanoma (KEYNOTE-942): a randomised, phase 2b study. The Lancet 2024;403:632–44. [CrossRef]
- Weichenthal M, Svane IM, Mangana J, Leiter U, Meier F, Ruhlmann C, et al. Real-World efficiency of pembrolizumab in metastatic melanoma patients following adjuvant anti-PD1 treatment. EJC Skin Cancer 2024;2:100271. [CrossRef]
- Jim Stallard. In Early-Phase Pancreatic Cancer Clinical Trial, Investigational mRNA Vaccine Induces Sustained Immune Activity in Small Patient Group 2025. https://www.mskcc.org/news/can-mrna-vaccines-fight-pancreatic-cancer-msk-clinical-researchers-are-trying-find-out.
- Chitwood, H. & Myers, A. Use of Circulating Tumor DNA to Monitor Minimal Residual Disease Among Patients With Colorectal Cancer. Clin J Oncol Nurs 2023. [CrossRef]
- Oosting LT, Franke K, Martin MV, Kloosterman WP, Jamieson JA, Glenn LA, et al. Development of a Personalized Tumor Neoantigen Based Vaccine Formulation (FRAME-001) for Use in a Phase II Trial for the Treatment of Advanced Non-Small Cell Lung Cancer. Pharmaceutics 2022;14:1515. [CrossRef]
- Bitounis D, Jacquinet E, Rogers MA, Amiji MM. Strategies to reduce the risks of mRNA drug and vaccine toxicity. Nat Rev Drug Discov 2024;23:281–300. [CrossRef]
- Kremsner PG, Ahuad Guerrero RA, Arana-Arri E, Aroca Martinez GJ, Bonten M, Chandler R, et al. Efficacy and safety of the CVnCoV SARS-CoV-2 mRNA vaccine candidate in ten countries in Europe and Latin America (HERALD): a randomised, observer-blinded, placebo-controlled, phase 2b/3 trial. Lancet Infect Dis 2022;22:329–40. [CrossRef]
- Sheykhhasan M, Ahmadieh-Yazdi A, Heidari R, Chamanara M, Akbari M, Poondla N, et al. Revolutionizing cancer treatment: The power of dendritic cell-based vaccines in immunotherapy. Biomed Pharmacother 2025;184:117858. [CrossRef]
- Kranz LM, Diken M, Haas H, Kreiter S, Loquai C, Reuter KC, et al. Systemic RNA delivery to dendritic cells exploits antiviral defence for cancer immunotherapy. Nature 2016;534:396–401. [CrossRef]
- Stewart-Jones GBE, Elbashir SM, Wu K, Lee D, Renzi I, Ying B, et al. Development of SARS-CoV-2 mRNA vaccines encoding spike N-terminal and receptor binding domains 2022. [CrossRef]
- Arevalo CP, Bolton MJ, Le Sage V, Ye N, Furey C, Muramatsu H, et al. A multivalent nucleoside-modified mRNA vaccine against all known influenza virus subtypes. Science 2022;378:899–904. [CrossRef]
- Sahin U, Muik A, Vogler I, Derhovanessian E, Kranz LM, Vormehr M, et al. BNT162b2 vaccine induces neutralizing antibodies and poly-specific T cells in humans. Nature 2021;595:572–7. [CrossRef]
- Xu S, Yang K, Li R, Zhang L. mRNA Vaccine Era—Mechanisms, Drug Platform and Clinical Prospection. Int J Mol Sci 2020;21:6582. [CrossRef]
- Wei H, Rong Z, Liu L, Sang Y, Yang J, Wang S. Streamlined and on-demand preparation of mRNA products on a universal integrated platform. Microsyst Nanoeng 2023;9:97. [CrossRef]
- Doua J, Ndembi N, Auerbach J, Kaseya J, Zumla A. Advancing local manufacturing capacities for vaccines within Africa - Opportunities, priorities and challenges. Vaccine 2025;50:126829. [CrossRef]
- Davidopoulou C, Kouvelas D, Ouranidis A. COMPARING vaccine manufacturing technologies recombinant DNA vs in vitro transcribed (IVT) mRNA. Sci Rep 2024;14:21742. [CrossRef]
- Seth Berkley. Ramping up Africa’s vaccine manufacturing capability is good for everyone. Here’s why 2022.
- Van De Pas R, Widdowson M-A, Ravinetto R, N Srinivas P, Ochoa TJ, Fofana TO, et al. COVID-19 vaccine equity: a health systems and policy perspective. Expert Rev Vaccines 2022;21:25–36. [CrossRef]
- Faksova K, Walsh D, Jiang Y, Griffin J, Phillips A, Gentile A, et al. COVID-19 vaccines and adverse events of special interest: A multinational Global Vaccine Data Network (GVDN) cohort study of 99 million vaccinated individuals. Vaccine 2024;42:2200–11. [CrossRef]
- Mahase E. FDA pauses all infant RSV vaccine trials after rise in severe illnesses. BMJ 2024:q2852. [CrossRef]
- Meghana GVR, Chavali DP. Examining the Dynamics of COVID-19 Misinformation: Social Media Trends, Vaccine Discourse, and Public Sentiment. Cureus 2023;15:e48239. [CrossRef]
- Bouderhem R. Challenges Faced by States and the WHO in Efficiently Regulating the Use of mRNA Vaccines. IECV 2023, MDPI; 2023, p. 1. [CrossRef]
- Kumar A, Dixit S, Srinivasan K, M D, Vincent PMDR. Personalized cancer vaccine design using AI-powered technologies. Front Immunol 2024;15:1357217. [CrossRef]
- Clemente B, Denis M, Silveira CP, Schiavetti F, Brazzoli M, Stranges D. Straight to the point: targeted mRNA-delivery to immune cells for improved vaccine design. Front Immunol 2023;14:1294929. [CrossRef]
Figure 1.
(A), an illustration of mRNA structure. B: the mechanism of mRNA vaccine in vivo. 1. After injection, the LNP encapsulated vaccine is internalized by APCs like dendritic cells and macrophages through endocytosis. 2, in the cytoplasm, the mRNA is released to be translated into tumor-specific antigen in the ribosome. 3, the synthesized antigens are processed into peptide fragment by proteosome. 4, the peptide fragments are loaded on major histocompatibility complex (MHC) molecules. MCH class I molecules present the peptides to CD8+ cytotoxic T cell, while MHC class II molecules present them to CD4+ helper T cells. 5, the interactions between MHC-antigen complexes and T cell receptors (TCRs) activates T cell which target and destroy tumor cells expressing the same antigen. The MHC II-antigen complex activates helper T cells which in turn activates B cell. 6, the interaction between helper T cell activates B cell for antibodies production and/or the B cell recognizes tumor-specific antigen on cell surface and form a complex via B cell receptors (BCRs) to activate B cells and lead to production of antibodies.
Figure 1.
(A), an illustration of mRNA structure. B: the mechanism of mRNA vaccine in vivo. 1. After injection, the LNP encapsulated vaccine is internalized by APCs like dendritic cells and macrophages through endocytosis. 2, in the cytoplasm, the mRNA is released to be translated into tumor-specific antigen in the ribosome. 3, the synthesized antigens are processed into peptide fragment by proteosome. 4, the peptide fragments are loaded on major histocompatibility complex (MHC) molecules. MCH class I molecules present the peptides to CD8+ cytotoxic T cell, while MHC class II molecules present them to CD4+ helper T cells. 5, the interactions between MHC-antigen complexes and T cell receptors (TCRs) activates T cell which target and destroy tumor cells expressing the same antigen. The MHC II-antigen complex activates helper T cells which in turn activates B cell. 6, the interaction between helper T cell activates B cell for antibodies production and/or the B cell recognizes tumor-specific antigen on cell surface and form a complex via B cell receptors (BCRs) to activate B cells and lead to production of antibodies.
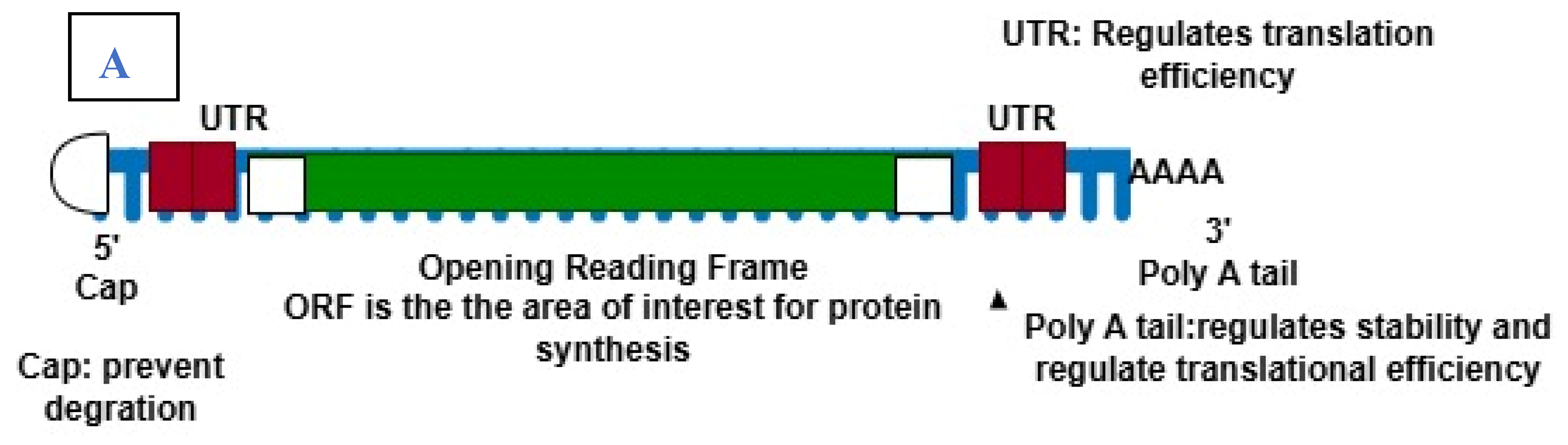
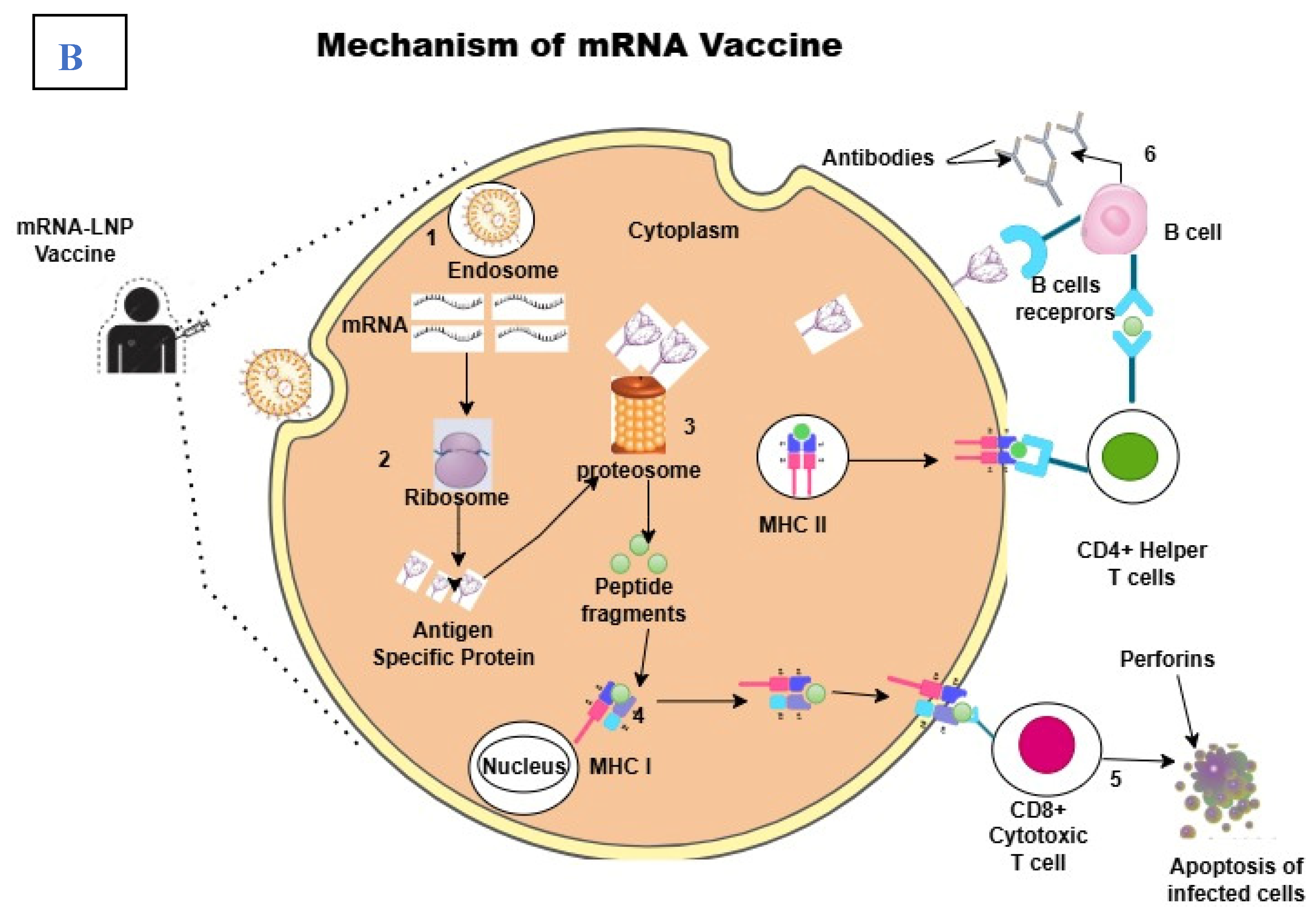
Figure 2.
Summary of current innovations in mRNA vaccine design. 1. Structural optimization of mRNA to enhance packaging and delivery. 2. New methods aimed at improving delivery systems over the traditional lipid nanoparticles such as the use of exosome/peptide, ionizable lipids for easy processing and the release of mRNA molecules upon injection. 3. Self-amplifying mRNA (sa-mRNA) for lower dose requirement of mRNA vaccine as the incorporated molecules like zapomeran helps to produce more copies of mRNAs. 4. Thermostability is another strategy to overcome the need for low temperature for storage and transportation of mRNA-LNPs vaccines. These vaccines are stable at temperature of about 2-80C. 5. Personalized and targeted vaccines based on patients’ genetic profile. 6. Adjuvant integration of additional molecules like cytokines or combination of vaccine with traditional immune blockade therapies.
Figure 2.
Summary of current innovations in mRNA vaccine design. 1. Structural optimization of mRNA to enhance packaging and delivery. 2. New methods aimed at improving delivery systems over the traditional lipid nanoparticles such as the use of exosome/peptide, ionizable lipids for easy processing and the release of mRNA molecules upon injection. 3. Self-amplifying mRNA (sa-mRNA) for lower dose requirement of mRNA vaccine as the incorporated molecules like zapomeran helps to produce more copies of mRNAs. 4. Thermostability is another strategy to overcome the need for low temperature for storage and transportation of mRNA-LNPs vaccines. These vaccines are stable at temperature of about 2-80C. 5. Personalized and targeted vaccines based on patients’ genetic profile. 6. Adjuvant integration of additional molecules like cytokines or combination of vaccine with traditional immune blockade therapies.
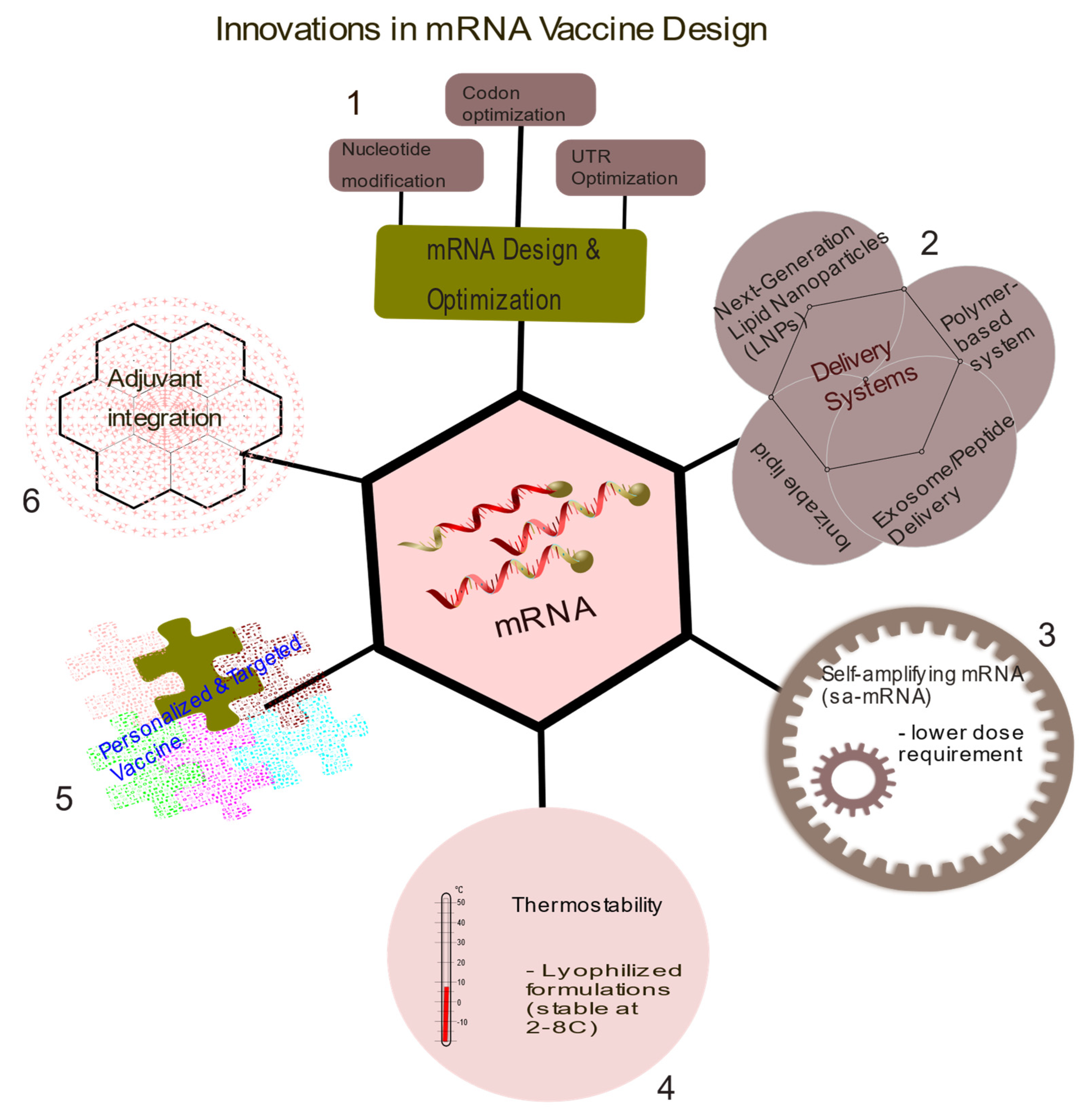
Figure 3.
scheme for multiple dendritic cell-based mRNA vaccine against cancers. The combination of myeloid DC (mDC) and plasmacytoid DC (pDC) could significantly improve immune activation and antibodies production against cancer.
Figure 3.
scheme for multiple dendritic cell-based mRNA vaccine against cancers. The combination of myeloid DC (mDC) and plasmacytoid DC (pDC) could significantly improve immune activation and antibodies production against cancer.
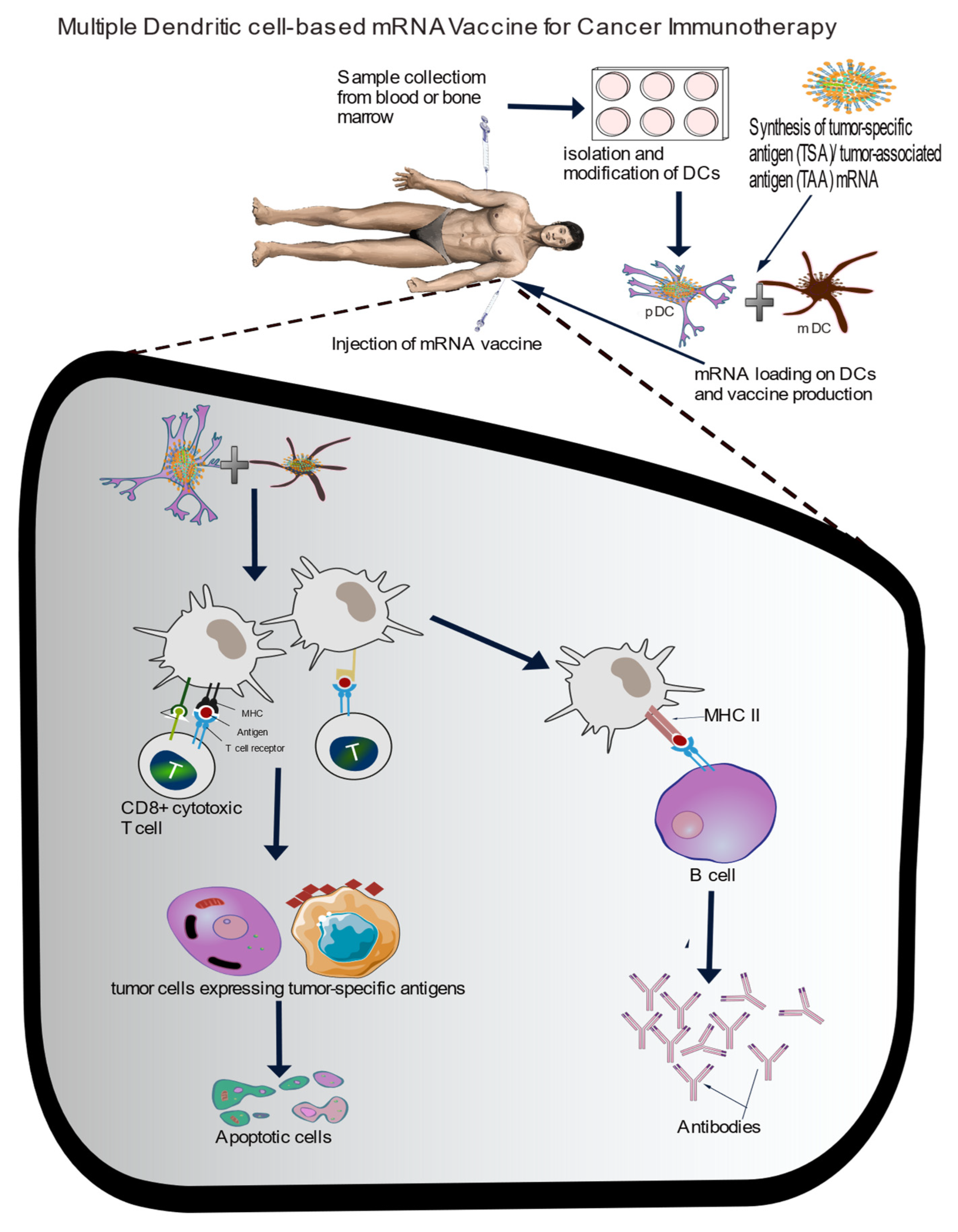

Table 1.
Ongoing clinical trials on infectious diseases.
| Company | Candidate | Product Type | Disease | Status | CT Ref |
|---|---|---|---|---|---|
| Sanofi | VAV00027 | mRNA/LNP | RSV/hMPV | Phase I | NCT06237296 |
| VBS00001 | mRNA vaccine | Pandemic flu H5 | Phase I/II | NCT06727058 | |
| MRT5421, MRT5424, and MRT5429 | mRNA vaccine | Influenza | Phase I/II | NCT06361875 | |
| mRNA vaccine | Influenza | Phase I/II | NCT06744205 | ||
| mRNA-NA | mRNA vaccine | Influenza | Phase I | NCT05426174 | |
| mRNA vaccine | Influenza | Phase I | NCT05829356 | ||
| mRNA vaccine | Acne | Phase I/II | NCT06316297 | ||
| Rabipur® CV7202 | mRNA vaccine | Rabies | Phase I | NCT03713086 | |
| RNActive® CV7201 | mRNA vaccine | Rabies | Phase I | NCT02241135 | |
| MRT5413 | mRNA vaccine | Influenza | Phase I/II | NCT05650554 | |
| mRNA vaccine | Influenza | Phase I | NCT06118151 | ||
| Moderna TX, Inc. | mRNA-1189 | mRNA vaccine | Epstein-Barr Virus (EBV) | Phase I/II | NCT05164094 |
| mRNA-1644 mRNA-1574 |
HIV | Phase I/II | NCT03547245 | ||
| mRNA-1215 | Nipah Virus (NiV) | Phase I | NCT05398796 | ||
| mRNA-1769 | Mpox | Phase I/II | NCT05995275 | ||
| mRNA-1403 | Novovirus/(Acute Gastroenteritis) | NCT06592794 | |||
| mRNA-1647 | Cytomegalovirus (CMV) | Phase III | NCT05085366 | ||
| mRNA-1325, mRNA-1893 |
mRNA vaccine | Zika virus | Phase I Phase II |
NCT03014089 NCT04064905 |
|
| mRNA-1010 | Seasonal Flu | Phase III | NCT06602024 | ||
| mRNA-1011.1, mRNA-1011.2, mRNA-1012.1 | Next-Gen mRNA vaccines | Influenza Virua | Phase I/II | NCT05827068 | |
| mRNA-1345 | Respiratory Syncytial Virus (RSV) | Phase III | NCT06067230 | ||
| mRNA-1365 | Human Metapneumovirus (hMPV) | Phase I | NCT05743881 | ||
| mRNA-1608 | Herpes Simplex Virus | Phase I/II | NCT06033261 | ||
| mRNA-1468 | Herpes Zoster (HZ) | Phase I/II | NCT05701800 | ||
| VAL-339851 | Seasonal influenza | Phase I | NCT03345043 | ||
| mRNA-1944 | Chikungunya Virus | Phase I | NCT03829384 | ||
| CanSino Biologics | mRNA vaccine | COVID-19 | Phase I/II | NCT05373485 NCT05373472 |
|
| National Institute of Allergy and Infectious Diseases (NIAID) | HVTN 302 | BG505 MD39.3 mRNA/BG505 MD39.3 gp151 mRNA/BG505 MD39.3 gp151 CD4KO mRNA | HIV | Phase I | NCT05217641 |
| mRNA-1273 | mRNA-LNP | COVID-19 | Phase I | NCT04283461 | |
| mRNA-1215 | mRNA vaccine | Nipah Virus (NiV) | Phase I | NCT05398796 | |
| Massachusetts General Hospital | mRNA-transfected autologous dendritic cells | HIV-1 | Phase I/II | NCT00833781 | |
| AstraZeneca | AZD9838/ AZD6563 | mRNA-VLP | SARS-CoV-2 | Phase I | NCT06147063 |
| CSPC ZhongQi Pharmaceutical Technology Co., Ltd. | SYS6006 | mRNA vaccine | SARS-CoV-2 | Phase II | NCT05439824 |
| Immorna Biotherapeutics, Inc. | JCXH-221 | mRNA vaccine | COVID-19 | Phase II | NCT05743335 |
| JCXH-108 | mRNA vaccine | RSV | Phase I | NCT06564194 | |
| CNBG-Virogin Biotech (Shanghai) Ltd. | ZSVG-02-O | mRNA vaccine | COVID-19 | Phase II | NCT06113731 |
| Vaxart | VXA-CoV2-3.1 | Oral SARS-CoV-2 Vaccine Tablet | COVID-19 | Phase II | NCT06672055 |
| Shenzhen Shenxin Biotechnology Co., Ltd | IN001 | mRNA vaccine | Herpes Zoster | Phase I | NCT06375512 |
| *** Lemonex | LEM-mR203 | mRNA-DegradaBALL vaccine | COVID-19 | Phase I | NCT06032000 |
| Barinthus Biotherapeutics | ChAdOx1-HBV | mRNA HBV vaccine | Chronic HBV infection | Phase I | NCT04297917 |
| RinuaGene Biotechnology Co., Ltd. | RG002 | mRNA vaccine | HPV16/18 associated Cervical Intraepithelial Neoplasia Grade 2 or 3(CIN2/3). | Phase I/II | NCT06273553 |
| Speransa Therapeutics | PRIME-2-CoV_Beta | COVID-19 | Phase I | NCT05367843 | |
| Albert B. Sabin Vaccine Institute | Sinovac | mRNA vaccine | COVID-19 | Phase IV | NCT05343871 |
| Sinocelltech Ltd. | SCTV01E-2 | mRNA vaccine | COVID-19 | Phase II | NCT05933512 |
| SK Bioscience Co., Ltd. | GBP560-A GBP560-B |
mRNA vaccine | SK Japanese Encephalitis virus disease | Phase I/II | NCT06680128 |
| Gritstone bio, Inc. Seqirus Stemirna Therapeutics |
GRT-R912, GRT-R914, and GRT-R918 | samRNA Vaccine | COVID-19/HIV | Phase I | NCT05435027 |
| V202_01 | Sa-mRNA vaccine | Influenza | Phase I | NCT06028347 | |
| SWC002 | mRNA vaccine | COVID-19 | Phase I/II | NCT05144139 | |
| SW-BIC-213 | mRNA vaccine | COVID-19 | Phase III | NCT05580159 | |
| SWIM816 | mRNA vaccine | COVID-19 | Phase II/III | NCT05911087 |
Table 2.
List of ongoing clinical trials involving mRNA therapies for cancers.
| Company | Candidate | Product Type | Cancer Type | Status | CT Ref |
|---|---|---|---|---|---|
| BioNTech | BNT111 TYR, vaccine, CTAG1B | mRNA vaccine | Malignant melanoma | Phase II | NCT04526899 |
| BNT113 vaccine | mRNA vaccine | Head and neck cancer | Phase II | NCT04534205 | |
| BioNTech/Regeneron Pharmaceuticals | BNT116 vaccine, TAA | mRNA vaccine | Non-small cell lung cancer (NSCLC) | Phase II | NCT05557591 |
| BioNTech/Roche | BNT112 (autogene cevumeran) vaccine, neoantigen | mRNA vaccine | Malignant melanoma | Phase II | NCT03815058 |
| Colorectal cancer | Phase II | NCT04486378 | |||
| Other metastatic tumors | Phase II | NCT03289962 | |||
| Pancreatic ductal adenocarcinoma (PDAC) | Phase II | NCT05968326 | |||
| Muscle invasive urothelial carcinoma (MIUC) | Phase II | NCT06534983 | |||
| **** | Metastatic castration resistance prostate cancer (mCRPC) | Phase I/II | NCT04382898 | ||
| Moderna TX, Inc. | mRNA-4359 + pembrolizumab | Advanced solid tumors | Phase I/II | NCT05533697 | |
| mRNA-4106 + Nivolumab/Relatlimab | Solid tumors | Phase I | NCT06880549 | ||
| mRNA-2752 | Refractory solid tumors/lymphoma | Phase I | NCT03739931 | ||
| mRNA-2736 | Relapsed/refractory multiple myeolema | Phase I | NCT05918250 | ||
| Moderna/ Merck & Co | mRNA-4157 vaccine/ neoantigen | mRNA vaccine | Bladder cancer | Phase I/II | NCT06305767 |
| non-small cell lung cancer (NSCLC) | Phase III | NCT06077760 | |||
| Renal cell carcinoma | Phase II | NCT06307431 | |||
| Malignant melanoma | Phase III | NCT05933577 | |||
| Others | Phase II/III | NCT06295809 | |||
| Stemirna | mRNA vaccine | Advanced malignant solid tumors | Preclinical | NCT05949775 | |
| Personalized tumor vaccine for esophageal and non-small cells lung cancers | Preclinical | NCT03908671 | |||
| CureVac | CV09050101 | mRNA vaccine (CVGBM) | Glioblastoma (GBM) | Phase I | NCT05938387 |
| Fudan University | PANC-IIT-RGL-mRNA vaccine | mRNA vaccine | Pancreatic adenocarcinoma | Preclinical/phase I | NCT06156267 |
| HRXG-K-1939 | mRNA vaccine + adebrelimab | Advanced solid tumors | Phase I | NCT05942378 | |
| Guangzhou Medical University | ZZVACCINE-mRNA-020 | Neoantigen mRNA vaccine | Advanced Solid Tumors | Phase I | NCT06195384 |
| National cancer institute *** | NCI-4650 | mRNA-based personalized vaccine | Metastatic epithelial cancer | Phase I/II | NCT03480152 |
| Ruijin Hospital | Xp-004 | mRNA tumor vaccine | Recurrent pancreatic cancer | Phase I | NCT06496373 |
| WGc-043 | EBV mRNA vaccine | Refractory Lymphoma | Phase I | NCT06788600 | |
| Guangdong Provincial People's Hospital | Neoantigen mRNA vaccine | non-small cell lung cancer (NSCLC). | Phase I | NCT06735508 | |
| University Hospital Tuebingen | RNA-Mel-03 | mRNA vaccine coding for Melan-A, Mage-A1, Mage-A3, Survivin, GP100 and Tyrosinase | Malignant melanoma | Phase I/II | NCT00204516 |
| Shanghai Zhongshan Hospital | Personalized mRNA vaccine | Posto perative hepatocellular carcinoma | Preclinical | NCT05761717 | |
| Steinar Aamdal*** | DC-004 | mRNA vaccine with dendritic cells | Metastatic malignant melanoma | Phase I/II | NCT00961844 |
| Radboud University Medical left | TLR-DC and Trimix DC loaded with mRNA encoding melanoma-associated tumor antigens (gp100 and tyrosinase) | Metastatic melanoma | Phase I/II | NCT01530698 | |
| *** Memorial Sloan Kettering Cancer left | Vaccination With CT7, MAGE-A3, and WT1 mRNA-electroporated Autologous Langerhans-type Dendritic Cells as Consolidation | Multiple myeloma | Phase I | NCT01995708 | |
| Autologous Langerhans-type Dendritic Cells with mRNA encoding tumor antigen | Melanoma | Phase I | NCT01456104 | ||
| Shanghai Jiao Tong University School of Medicine | mRNA-0523-L001 | Individualized mRNA neoantigen vaccine | Advanced endocrine tumor | Phase I | NCT06141369 |
| John Sampson, Duke University | BTSC mRNA-loaded DCs | Personalized cancer vaccine | glioblastoma multiforme (GBM) | Phase I | NCT00890032 |
| Sir Run Run Shaw Hospital | iNeo-Vac-R01 | Personalized mRNA vaccine | Advanced Digestive System Neoplasms | Phase I | NCT06019702 |
| iNeo-Vac-R01 + standard adjuvant therapy | NCT06026774 | ||||
| Inge Marie Svane, Herlev Hospital | mRNA transfected dendritic cell | Metastatic prostate cancer | Phase II | NCT01446731 | |
| Steinar Aamdal, Oslo University Hospital *** | DC-004 ** | mRNA vaccine therapy with dendritic cells | Metastatic malignant melanoma | Phase I/II | NCT00961844 |
| DC-006 ** | Dendric cells with amplified ovarian cancer stem cell vaccine | Recurrent Platinum Sensitive Epithelial Ovarian cancer | Phase I/II | NCT01334047 | |
| DC-005 | Dendritic cells with cancer mRNA vaccine | Prostate cancer | Phase I/II | NCT01197625 | |
| DC-CAST-GMB | Dendritic cells with cancer mRNA vaccine | gliobalstoma | Phase I/II | NCT00846456 | |
| Nanjing Tianyinshan Hospital | RGL-270 | MRNA Vaccine + Adebrelimab | Non-small cells lung cancer | Phase I | NCT06685653 |
| Radboud University Medical left | TLR-DC and Trimix DC | Autologous dendritic cell vaccine | Melanoma | Phase I/II | NCT01530698 |
| mRNA vaccine | Melanoma stage III/IV | Phase I/II | NCT00243529 | ||
| MiHA-loaded PD-L-silenced DC vaccine | Hematological malignancies | Phase I/II | NCT02528682 | ||
| CEA-loaded dendritic cell vaccine | Colorectal cancer | Phase I/II | NCT00228189 | ||
| ***Ludwig Institute for Cancer Research | BI 1361849 | mRNA vaccine + durvalumab + tremelimumab | Metastatic non-small cells lung cancer | Phase I/II | NCT03164772 |
| YueJuan Cheng, Peking Union Medical College Hospital | KY- 1007 | Personalized mRNA neoantigen vaccine | Advanced solid tumors | Early phase I | NCT05359354 |
| Guangdong 999 Brain Hospital | PERCELLVAC3 | Personalized cellular vaccine | Glioblastoma | Phase I | NCT02808416 |
| PerCellVac2 | Personalized cellular vaccine | Glioblastoma | Phase I | NCT02808364 | |
| Jinling Hospital, China | SJ-Neo006 | Camrelizumab + personalized neoantigen vaccines | Pancreatic cancer | Early phase I | NCT06326736 |
| Wu Wenming, Peking Union Medical College Hospital | XH001 | Neoantigen cancer vaccine + Ipilimumab | Pancreatic cancer | Early phase I | NCT06353646 |
| University of Florida *** | RNA PRIME | pp65 RNA LP (DP1 & DP2) vaccines | Pediatric Recurrent Intracranial malignancies | Phase I/II | NCT05660408 |
| pp65 RNA-LP | mRNA vaccine | Recurrent glioblastoma | Phase I | NCT06389591 | |
| PNOC020 | RNA-loaded lipid particle (RNA-LP) vaccine | Pediatric High-Grade Gliomas (pHGG), and Adult Glioblastoma | Phase I/II | NCT04573140 | |
| pp65-shLAMP *** | pp65-shLAMP mRNA DCs with GM-CSF | Glioblastoma Multiforme | Phase II | NCT02465268 | |
| jianming xu, The Affiliated Hospital of the Chinese Academy of Military Medical Sciences | mRNA neoantigen tumor vaccine | Advanced gastric, esophageal, and liver cancers | Early phase I | NCT05192460 | |
| XH001 | Neoantigen tumor vaccine, XH001 + sintilimab | Advanced solid tumors | Preclinical | NCT05940181 | |
| RinuaGene Biotechnology Co., Ltd. | RG002 | mRNA therapeutic vaccine | HPV16/18 associated Cervical Intraepithelial neoplasia | Phase I/II | NCT06273553 |
| Gary Archer Ph.D., Duke University | Dendric cell-based mRNA vaccine | Brain tumor | Phase I | NCT00639639 | |
| DC-based mRNA vaccine + Nivolumab | Brain tumors | Phase I | NCT02529072 | ||
| University Medical left Groningen | W_ova1 vaccine | Liposome formulated mRNA vaccine | Ovarian cancer | Phase I | NCT04163094 |
| Zwi Berneman, University Hospital, Antwerp | CCRG12-001 | DC vaccine | Acute myeloid leukemia (AML) | Phase II | NCT01686334 |
| Svein Dueland, Oslo University Hospital | mRNA vaccine | Malignant melanoma | Phase I/II | NCT01278940 | |
| mRNA vaccine | Androgen Resistant Metastatic Prostate cancer | Phase I/II | NCT01278914 | ||
| Inge Marie Svane, Herlev Hospital | DC vaccine | Breast cancer | Phase I | NCT00978913 | |
| Peking Union Medical College Hospital | ABOR2014(IPM511) | Neoantigen mRNA vaccine | Advanced hepatocellular carcinoma | Preclinical | NCT05981066 |
| Diakonos Oncology Cooperation | DOC1021 | Dendric cell-based mRNA vaccine loaded with tumor lysate | Adult Glioblastoma | Phase II | NCT06805305 |
| Peking University Hospital & Institute | JCXH-212 | Tumor neoantigen mRNA vaccine | Malignant solid tumors | Early phase I | NCT05579275 |
| *** Merck Sharp & Dohme LLC | mRNA 5679/V941 | mRNA vaccine | Metastatic Non-Small Cell Lung Cancer, Colorectal or Pancreatic Adenocarcinoma | Phase I | NCT03948763 |
| University Hospital, Antwerp | CCRG 05-001 | Dendritic-based mRNA vaccine | Acute Myeloid Leukemia | Phase I | NCT00834002 |
| DC vaccine | Pediatric High Grade Gliomas, and Diffuse Intrinsic Pontine Gliomas | Phase I/II | NCT04911621 | ||
| The Affiliated Hospital Of Guizhou Medical University | InnoPCV | mRNA vaccine | Advanced solid tumors | Early phase I | NCT06497010 |
Table 3.
multivalent vaccines in trials.
| Company/sponsor | Candidate | Product Type | Disease(s) | Status | Reference |
| Duke University | |||||
| BioNTech/Roche | BNT112 (autogene cevumeran) vaccine, neoantigen | mRNA vaccine | Malignant melanoma | Phase II | NCT03815058 |
| Colorectal cancer | Phase II | NCT04486378 | |||
| Other metastatic tumors | Phase II | NCT03289962 | |||
| Pancreatic ductal adenocarcinoma (PDAC) | Phase II | NCT05968326 | |||
| Muscle invasive urothelial carcinoma (MIUC) | Phase II | NCT06534983 | |||
| Moderna/ Merck & Co | mRNA-4157 vaccine/ neoantigen | mRNA vaccine | Bladder cancer | Phase I/II | NCT06305767 |
| non-small cell lung cancer (NSCLC) | Phase III | NCT06077760 | |||
| Renal cell carcinoma | Phase II | NCT06307431 | |||
| Malignant melanoma | Phase III | NCT05933577 | |||
| Others | Phase II/III | NCT06295809 | |||
| mRNA-1195 | mRNA vaccine |
Multiple Sclerosis | Phase II | NCT06735248 | |
| mRNA-3927 | Propionic Acidemia | Phase I/II | NCT04159103 | ||
| mRNA-3745 | Glycogen Storage Disease Type 1a (GSD1a) | Phase I/II | NCT05095727 | ||
| mRNA-3705 | methylmalonic acidemia (MMA) | Phase I/II | NCT05295433 | ||
| mRNA-1083 | COVID-19 + Influenza | Phase III | NCT06694389 | ||
| mRNA-1975, mRNA-1982 | Lyme Disease | Phase I/II | NCT05975099 | ||
| mRNA-1653 | hMPV + Parainfluenza Virus Type 3 (PIV3) | Phase I | NCT04144348 | ||
| mRNA-1045 | Influenza + RSV | Phase I | NCT05585632 | ||
| mRNA-1230 | Influenza + RSV + SARS-CoV-2 | Phase I | NCT05585632 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



