Submitted:
04 May 2025
Posted:
05 May 2025
You are already at the latest version
Abstract
Asthma is a chronic inflammatory disease of the respiratory tract with multifactorial origins involving both genetic predisposition and environmental exposures. Recent insights have underscored the pivotal role of epigenetic regulation in the pathogenesis and heterogeneity of asthma. This review focuses on key epigenetically important proteins categorized as writers, erasers, and readers that govern DNA methylation, histone modifications, and RNA modifications. These proteins modulate gene expression without altering the underlying DNA sequence, thereby influencing immune responses, airway remodeling, and disease severity. We highlight the structural and functional dynamics of histone acetyltransferases (e.g., p300/CBP), histone deacetylases (e.g., SIRT family), DNA methyltransferases (DNMT1, DNMT3A), demethylases (TET1), and methyl-CpG-binding proteins (MBD2) in shaping chromatin accessibility and transcriptional activity. Additionally, the m6A RNA modification machinery including METTL3, METTL14, FTO, YTHDF1/2, IGF2BP2, and WTAP is explored for its emerging significance in regulating post-transcriptional gene expression during asthma progression. Structural characterizations of these proteins reveal conserved catalytic domains and interaction motifs, mirroring their respective families such as SIRTs, p300/CBP, DNMT1/3A, and YTHDF1/2 critical to their epigenetic functions, offering mechanistic insight into their roles in airway inflammation and immune modulation. By elucidating these pathways, this review provides a framework for the development of epigenetic biomarkers and targeted therapies. Future directions emphasize phenotype-specific epigenomic profiling and structure-guided drug design to enable precision medicine approaches in asthma management.
Keywords:
p300/CBP
; SIRT family
; DNMT1
; DNMT3A
; YTHDF1/2
; IGF2BP2
1. Introduction
1.1. Background on Asthma
Asthma, a chronic inflammatory respiratory condition, can manifest as symptoms like wheezing, coughing, airflow limitation and chest tightness instigated through chronic airway inflammation, tissue remodeling, mucus hypersecretion and bronchial hyperresponsiveness [1,2,3]. Based on the Global Burden of Disease Study 2019, the asthma affected 10.9% of the population in the United States, encompassing approximately 34 million individuals. This rate significantly surpasses that recorded in China. Despite this, asthma in China often goes undiagnosed and undertreated. The China Pulmonary Health study, a national cross-sectional analysis, indicated that around 45.7 million adults in China were impacted by asthma, yielding a prevalence of 4.2% in 2015 [4]. Over the past two decades, there has been a significant rise in both the incidence and prevalence of asthma worldwide [5]. Asthma impacts more than 300 million people globally and is associated with substantial health risks [6]. Although numerous advancements have been made in treating and diagnosing asthma, a large population still suffers, and it is one of the top infectious diseases around the world and roughly 10% of asthma patients do not sufficiently respond to standard therapy [7,8]. These statistics point to asthma becoming one of the most significant health challenges of our time, emphasizing the urgent need for effective prevention, therapeutics, and disease management. Research suggests that the development of asthma and allergic conditions is multifactorial, with both genetic and environmental factors playing key roles. Environmental factors affect the epigenome, so epigenomic and transcriptomic profiling may be useful in addition to genomic profiling for asthma prediction [9].
1.2. Role of Epigenetics in Asthma
Comparatively to genetics, epigenetics affects gene expression without altering DNA sequence [10]. Epigenetic changes may be triggered by environmental factors, including prenatal smoking and postnatal pollutants, nutrients, and drugs [11]. Asthma is considered a heritable disease according to numerous genetic studies; however, the rising incidence of asthma particularly in the recent years cannot be solely attributed to genetic elements [12]. Epigenetics is regarded as a key contributor to asthma pathogenesis [13]. Numerous studies have highlighted the importance of epigenetic modifications, such as DNA methylation, histone modifications, and RNA modifications , in regulating gene expression patterns relevant to asthma [14]. For instance, a study by Yang et al in 2017 demonstrated substantial differentiations in DNA methylation patterns between asthmatic and non-asthmatic individuals, particularly in genes associated with immune regulation and airway inflammation [15]. This finding underscores the potential role of epigenetic dysregulation in asthma susceptibility and severity. Epigenetic modifications take place in prenatal development, early childhood, and adolescence, as these life stages represent vulnerable periods to various asthma inductions [16]. In addition to the skin, respiratory tract is directly exposed to outer atmosphere, initiating epigenetic changes to nasal, buccal mucosa and bronchial epithelium [16]. In addition, around seventeen studies and meta-analyses have recognized the methylation patterns of immune cells in both whole blood and peripheral blood of individuals with asthma [17].
1.3. Significance of Studying Epigenetically Important Proteins
Epigenetic modifications, which include DNA methylation, histone modifications, chromatin remodeling, and RNA regulation are dynamically regulated by a myriad of epigenetically important proteins, which act as writers, erasers, and readers of epigenetic marks [18,19]. Dysregulation of these proteins has been implicated in various diseases, highlighting the importance of studying their structures, functions, and molecular mechanisms. This review discusses the significance of investigating epigenetically important proteins in asthma in understanding disease pathogenesis, identifying therapeutic targets, and advancing precision medicine strategies.
1.4. Overview of Epigenetic Mechanisms
Epigenetics encompasses molecular processes that induce reversible changes in phenotype by modifying gene expression without modifying the underlying DNA sequence [20]. Within the human genome, approximately 80% of DNA is organized into nucleosomes, with the remaining forming linker regions between these nucleosomes. These nucleosomes are subsequently organized into chromosomes, forming dense three-dimensional structures [21]. Histone proteins constitute the core components of nucleosomes, and they endure numerous posttranslational modifications (PTMs), containing phosphorylation, sumoylation, ubiquitination, methylation and acetylation. These PTMs, particularly present in critical regulatory genomic regions such as promoters or enhancers, can modulate DNA accessibility to the transcriptional machinery, thereby influencing the gene's transcriptional status as active, poised, or silenced. For instance, histone acetylation which is facilitated through histone acetyltransferases (HATs) and reversed via histone deacetylases (HDACs), typically correlates with gene activation [14,22]. DNA methylation encompasses the enzymatic addition of a methyl group to the cytosine ring of DNA, representing another form of epigenetic modification. This DNA methylation is facilitated through DNA methyltransferases, whereas DNA demethylation is carried out by proteins belonging to the ten eleven translocation methylcytosine dioxygenase family. Typically, DNA methylation is allied with gene repression [23]. In addition to traditional epigenetic modifications, post-transcriptional modifications have also been observed in both coding and non-coding RNA molecules, including rRNA, tRNA, mRNA, miRNA, circular RNA (circRNA), and various long non-coding RNAs (lncRNAs), all of which contribute to the epigenetic modulation of gene expression [24]. For instance, miRNAs operate through attaching mature mRNA molecules in the cytoplasm, resulting in mRNA degradation or decreased translational efficiency by ribosomes [25,26]. Epigenetic regulation plays a critical role in asthma pathogenesis, influencing gene expression through DNA methylation, histone modifications, and RNA modification [27,28].
Figure 1.
A basic summary of the primary epigenetic modifications: (A) Chromosomal DNA wraps around histone proteins, forming nucleosomes. The accessibility of nucleosomes to nuclear factors is partially controlled by modifications to the tails of histone proteins. These modifications include acetylation, ubiquitination, phosphorylation and methylation. Acetylation reactions, facilitated by histone acetyltransferase (HAT), alter lysine (K) residues on histone tails. This process neutralizes the positive charges of the lysine side chains, resulting in a less condensed chromatin structure that is conducive to transcription. Conversely, deacetylation reactions, mediated by histone deacetylase (HDAC), remove acetyl groups from histone tails. This increases chromatin packaging, leading to a tighter conformation that inhibits DNA transcription. (B) In DNA modification methylation involves the addition of a methyl group to the carbon-5 position of cytosine within regions known as CG Islands, facilitated by DNA-methyltransferase (DNMT) enzymes. Methylation of CpG islands can lead to gene silencing by either blocking the binding of transcription factors (TFs) to the promoter or by promoting the binding of proteins, such as MeCpG-binding proteins (e.g., MBDs), that specifically recognize methylated CpGs. Furthermore, DNA methylation can reinforce certain histone modifications, such as demethylation and deacetylation, thereby establishing a mechanism for perpetuating these epigenetic marks. (C) RNA modification including mRNA and non-coding RNAs (ncRNAs). Non-coding RNAs are typically categorized as small or long ncRNAs. Functionally, they fall into two main groups: constitutively expressed housekeeping molecules and regulatory molecules, including microRNAs (miRNAs) and long non-coding RNAs (lncRNAs). The modification of m6A in mRNA is regulated by ‘writers’ ‘readers’, and ‘erasers. Writers such as METTL3, METTL14, and WTAP regulate m6A methylation. RNA m6A demethylation is prompted by eraser proteins such as FTO and IGF2BPs have role like other reader proteins.
Figure 1.
A basic summary of the primary epigenetic modifications: (A) Chromosomal DNA wraps around histone proteins, forming nucleosomes. The accessibility of nucleosomes to nuclear factors is partially controlled by modifications to the tails of histone proteins. These modifications include acetylation, ubiquitination, phosphorylation and methylation. Acetylation reactions, facilitated by histone acetyltransferase (HAT), alter lysine (K) residues on histone tails. This process neutralizes the positive charges of the lysine side chains, resulting in a less condensed chromatin structure that is conducive to transcription. Conversely, deacetylation reactions, mediated by histone deacetylase (HDAC), remove acetyl groups from histone tails. This increases chromatin packaging, leading to a tighter conformation that inhibits DNA transcription. (B) In DNA modification methylation involves the addition of a methyl group to the carbon-5 position of cytosine within regions known as CG Islands, facilitated by DNA-methyltransferase (DNMT) enzymes. Methylation of CpG islands can lead to gene silencing by either blocking the binding of transcription factors (TFs) to the promoter or by promoting the binding of proteins, such as MeCpG-binding proteins (e.g., MBDs), that specifically recognize methylated CpGs. Furthermore, DNA methylation can reinforce certain histone modifications, such as demethylation and deacetylation, thereby establishing a mechanism for perpetuating these epigenetic marks. (C) RNA modification including mRNA and non-coding RNAs (ncRNAs). Non-coding RNAs are typically categorized as small or long ncRNAs. Functionally, they fall into two main groups: constitutively expressed housekeeping molecules and regulatory molecules, including microRNAs (miRNAs) and long non-coding RNAs (lncRNAs). The modification of m6A in mRNA is regulated by ‘writers’ ‘readers’, and ‘erasers. Writers such as METTL3, METTL14, and WTAP regulate m6A methylation. RNA m6A demethylation is prompted by eraser proteins such as FTO and IGF2BPs have role like other reader proteins.
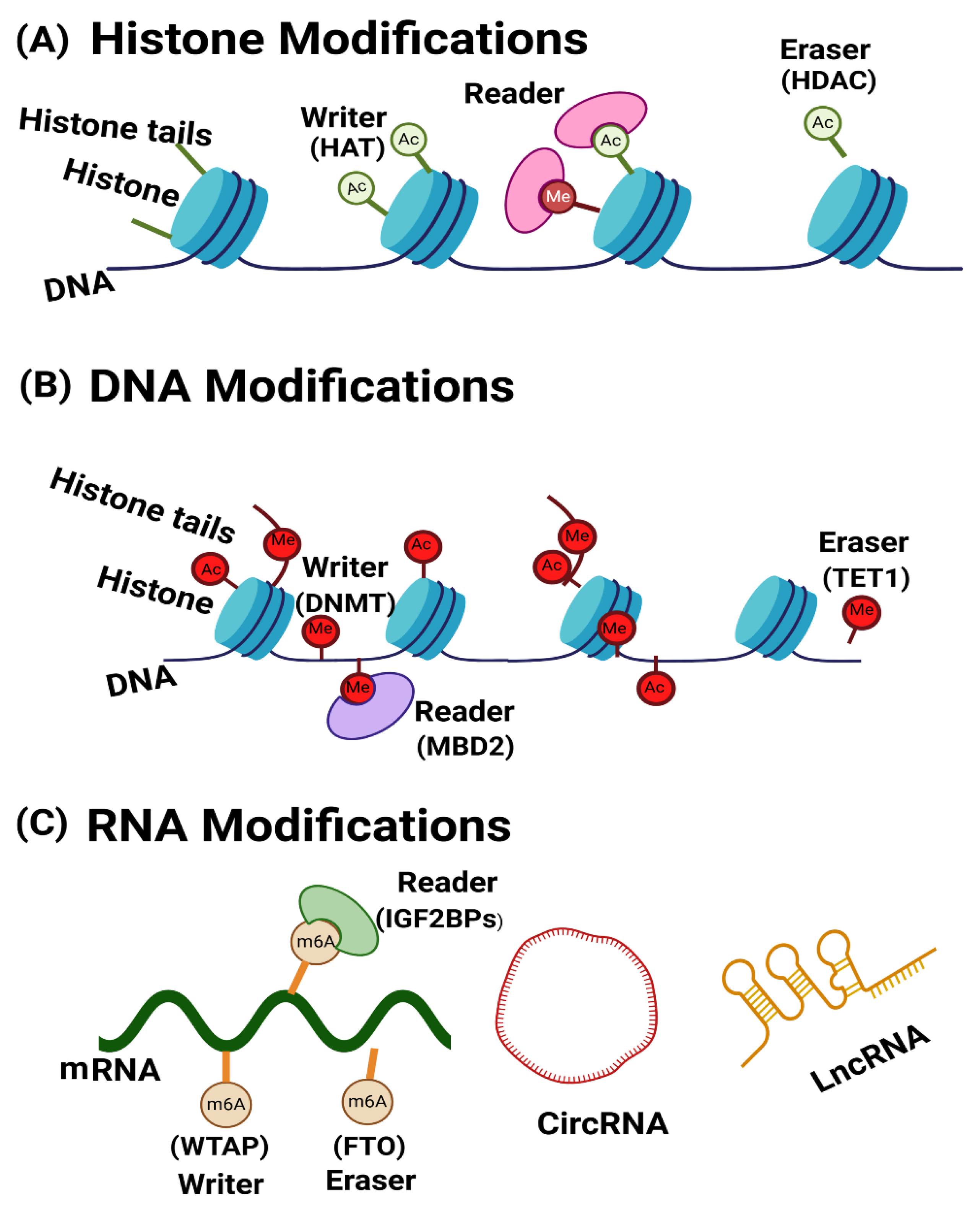
2. Role of Key Epigenetic Proteins in Asthma Pathogenesis
Several key proteins play crucial roles in epigenetic regulation, including DNA modification, histone modification, and RNA modification. These proteins are involved in the addition, removal, or reading of epigenetic marks, which can affect gene expression and contribute to the pathophysiology of asthma. We have categorized them as writers, erasers, and readers based on their respective modifications. Table 1 presents a list of epigenetically significant proteins involved in asthma.
3. Epigenetic Regulators in Asthma: Writers, Readers, and Erasers of Histone Modifications
Histone modifications, including acetylation, methylation, phosphorylation, ubiquitination, and sumoylation, are key regulators of chromatin structure [54]. Histone acetylation takes place at lysine residues positioned on the N-terminal tail of histones, regulated by histone acetyltransferases (HATs) and histone deacetylases (HDACs), which have opposing effects [55,56]. Human HATs can be sorted into 6 main orders based on the structure, features and mode of action of their catalytic domains. Lysine acetyltransferases 2A and 2B are members of the GCN5-related N-acetyltransferase family, which is a class of HATs that acetylate transcription factors and histones. They control cell cycles, centrosome's activity, DNA replication and repair. The MYST family has the following members KAT6A/MYST3/MOZ, KAT6B/MYST4/MORF, KAT7/MYST2/HBO1, KAT8/MYST1/hMOF, and KAT5/Tip60. This HAT group of enzymes is involved in transcription control and DNA repair. The transcriptional factor-related HAT family has the following members KAT4/TAF1/TBP and KAT12/TIFIIIC90, whereas KAT3A and KAT3B enzymes belong to the E1A binding protein p300 (EP300) and CREB binding protein (CREBBP, CBP) family. Lastly, KAT1/HAT1 and HAT4/NAA60 are histone acetyltransferases (HATs) located in the cytoplasmic, whereas KAT13A/SRC1, KAT13B/SCR3/AIB1/ACTR, KAT13C/p600, and KAT13D/CLOCK comprise the steroid receptor coactivators family [55,57,58,59]. Histone deacetylases are classified into 4 categories, that is Class I-IV: class I “(HDAC1, HDAC2, HDAC3, and HDAC8)”, class II “(HDAC4, HDAC5, HDAC6, HDAC7, HDAC9, and HDAC10)”, class III “(SIRT1-SIRT7)”, and class IV “(HDAC11)” [60]. HDACs of Class I, II, and IV rely on Zn²⁺ as a cofactor for their enzymatic activity, whereas the enzymatic function of class III HDACs, also known as Sirtuins, is dependent on NAD⁺ [61]. Histone acetylation "readers" are proteins that recognize acetylation marks, dynamically regulated by HATs ("writers") and HDACs ("erasers"), and influence transcriptional processes. Dysregulation of these readers is often linked to various human diseases [62,63,64]. Ogasawara et al in 2017 highlighted the role of histone acetylation in regulating T cell function in asthma. Bcl6 suppresses IL-4 expression in murine memory Th2 cells by binding to intron 2 of the Il4 locus, an effect modulated by IL-33 and associated with changes in H3K9 and H3K14 acetylation [65]. Table 1 summarizes key histone acetyltransferases (HATs) as writers and histone deacetylases (HDACs) as erasers of histone modifications involved in asthma pathogenesis. These enzymes regulate chromatin accessibility and the expression of inflammatory genes central to asthma. Altered expressions or activity of specific HATs and HDACs are linked to airway inflammation, immune cell activation, and disease severity. Some of these enzymes are detailed in terms of their structure and function, emphasizing their roles in modulating chromatin and regulating asthma-related gene expression.
3.1. The Role of HATs in Asthma
Barens et al has investigated that the levels of histone acetyltransferases (HATs), specifically CBP and p300 have been raised, while the levels of histone deacetylases (HDACs) were notably reduced in the bronchial biopsy samples from asthmatic patients in comparison to those from healthy persons. These changes favor the release of inflammatory factors, which are crucial for the amplification and persistence of inflammatory responses. This mechanism triggers chronic inflammation and the remodeling of the airway [36,66,67]. Research has shown that ORMDL3's promoter region has p300 HAT binding sites. In order to maintain the ORMDL3 gene's basic transcriptional activity, p300 helps transcription factors like ETS-1 and STAT6 via binding to the promotor region of ORMDL3 “[68,69]. P300 is a classic endogenous histone acetyltransferase [70]; it is extensively conserved across species and is integral to cell cycle regulation, cellular transformation, and apoptosis. Currently, three key biological functions of p300 have been identified. (1) Acetylation of histone tails: p300 facilitates transcription via the catalytic function of its KAT domain, which acetylates nucleosomal histones at promoters, leading to chromatin remodeling and relaxation, thereby enhancing DNA accessibility to other critical regulators “[71]. Due to its capacity to alter chromatin structure through histone acetylation, p300 is characterized as a "writer" of the epigenetic code “[72]. (2) As p300 acetylase modifies nonhistone transcription factors via acetylation thus augmenting their activity. (3) P300 acts as a transcriptional activator by recruiting transcription factors to the promoter localities of target genes, thus enhancing transcription [73]. Consequently, it is essential to explore whether histone acetylation mediated via p30 has a key role in the overexpression of ORMDL3 in asthma. Chen et al in 2020 found that increased levels of p300/CBP, a histone acetyltransferase, were associated with elevated inflammatory cytokines and poor prognosis in acute respiratory distress syndrome, highlighting its role in inflammation and disease outcome [74].
3.2. Structure of P300/CBP
Transcription factors (TFs) engage with coactivators such as the Mediator complex and the acetyltransferases CBP (KAT3A, CREBBP gene) and p300 (KAT3B, EP300 gene) via their activation domains; these two paralogous lysine acetyltransferases were discovered in the 1980s and 1990s [75,76,77,78,79,80,81,82,83]. EP300 and CREBBP are comprised of 31 exons and extend around 87 kb and 155 kb, respectively [84,85,86]. They possess a comparable structure with distinct functional domains, encompassing various protein–protein interaction motifs, and overall exhibit 58% of sequence identity (Figure 2). A nuclear receptor interaction domain (NRID) which is capable of binding to PXXP motifs is present on the N terminal region. Three regions rich in cysteine and histidine (C/H1 to C/H3) are implicated in protein–protein interactions. The zinc finger transcriptional adapters like TAZ1 and TAZ2 are present in C/H1 and C/H3 domains, whereas the C/H3 domain further includes a ZZ zinc finger domain and C/H2 have a homeodomain (PHD). The catalytic domain has exhibited significant conservation throughout evolution, with an 86% similarity between p300 and CBP. The structure comprises a KAT domain along with adjacent sections, specifically the Bromodomain, C/H2, and C/H3 regions [87,88,89,90].
The interaction domains are delineated through extensive, inherently disordered regions (IDRs). P300 is activated through autoacetylation [87], a process modulated by cellular signaling via transcription factor activation and dimerization [91]. P300/CBP is a multipurpose protein that serves as both a "writer" and a "reader" of lysine acetylation via its KAT domain and bromodomain (BD), respectively [87,92]. P300/CBP is a versatile lysine acetyltransferase capable of acetylating all four of the canonical histones [93]. There is a significant sequence similarity in other non-catalytic domains. The KIX domain promotes the binding of CREB, especially at the phosphorylated Ser133 residue, to other transcription factors, leading to the formation of a bromodomain (BD) that interacts with acetylated lysines [94]. The C terminal region of p300/CBP features an interferon-binding transactivation domain (IBiD), which encompasses a glutamine-rich domain and nuclear binding coactivator domain, succeeded by a proline-rich PxP motif [95,96].
3.3. The role of Class III histone Deacetylases in Asthma
Sirtuins are NAD-dependent class III histone deacetylases (HDAC), consisting of seven types (Sirt1-7) in human [97]. and can be categorized into four primary groups according to their sequence homology [98]. SIRT1–SIRT3 belong to class I, SIRT4 to class II, SIRT5 to class III and SIRT6 and SIRT7 to class IV [98]. A similar catalytic domain is present between all SIRTs and use NAD+ as a co-substrate. However, they differ in substrate affinities and sub-cellular localization [99,100]. SIRT1, 6 and 7 are principally found into the cellular nucleus, whereas SIRT3, 4 and 5 are mainly mitochondrial enzymes and SIRT2 is exceptional in the group, being a primarily cytoplasmic protein [101]. Asthma impacts individuals across all age groups, with a particular prevalence in children [102]. Current investigations have indicated that targeting SIRT1 could offer a promising new approach for asthma treatment [38,103]. It has been observed that the level of SIRT1 protein was downregulated in the severe asthmatic patients. [104]. SIRT1 has a pivotal role in mitigating inflammation in airway diseases such as asthma. In a study by Tang et al. (2018), SIRT1 was found to modulate inflammation by regulating IL-6 production through the Akt pathway in the context of allergic asthma [105]. Additionally, SIRT1 influences pulmonary function in asthma patients through its effect on IL-6 levels via the Akt signaling pathway [106]. Besides SIRT1, other sirtuins including SIRT2, SIRT3, SIRT6, and SIRT7 have also been linked with asthma. For example, SIRT2 aggravates allergic asthmatic inflammation, where inhibiting SIRT2 pharmacologically alleviates its severity, whereas genetic overexpression of SIRT2 worsens the allergic asthma phenotype [107]. Additionally, SIRT2 augments asthmatic related inflammation via promoting T-helper type 2 responses and macrophage polarization (Kim et al., 2020). In contrast, raising the expression has been shown to reduce apoptosis in the bronchial epithelium and airway inflammation in asthma [40]. Allergic asthma is a persistent inflammatory disorder of the airways, marked by airway remodeling, which severely impairs airflow in the lungs [109]. SIRT6 and SIRT7’s expression levels have been elevated in human bronchial epithelial cells which were obtained from asthma patients [35]. Enhanced SIRT6 alleviates airway remodelling by modulating epithelial-mesenchymal transition in asthma [110]. Conversely, increased SIRT7 facilitates airway remodelling in asthma by modulating TGF-β1-driven proliferation and migration of airway smooth muscle cells [42]; suggesting a distinct role for SIRT6 in airway remodelling.
3.4. Structure of Sirtuin
The sirtuin families demonstrate structural similarity, characterized by a conserved enzymatic core that encompasses a Rossmann fold domain along with a smaller domain composed of zinc-binding and helical domains [111]. Biochemical research on sirtuin activity discloses that deacetylation takes place in a stoichiometric manner, accompanied by the hydrolysis of the bond between nicotinamide and ribose in NAD+. This process results in the creation of a temporary covalent intermediate known as an O-alkylamidate [112]. Avalos et al. (2002) proposed two distinct structure-based mechanisms for the nicotinamide cleavage reaction in sirtuins [113]. Zhao et al in 2004 demonstrated that NAD+ binds to the sirtuin pocket in two different conformations: productive and non-productive [114]. In the productive state, NAD+ positions the positive charge of the nicotinamide ring within a conserved region called the C-site, which contains highly hydrophobic residues that alter the resonance of the delocalized electrons in the nicotinamide ring by distorting the carboxamide group. In the non-productive conformation, the nicotinamide ring does not fit into the C-site [115].
Sirt2, the primary subtype identified via Finnin et al. in 2001 [116], was the subject of the initial characterization of the Sirtuin protein structure, offering valuable insights into the Sirtuin catalytic core (PDB 1J8F). Notably, the structure of Sirt2's catalytic core serves as a nearly representative model for the core region of all Sirtuin isoforms, which contributes to the high degree of conservation observed within the Sirtuin family [117]. SIRT2 comprises a total of 389 residues, with 323 amino acids successfully crystallized at a 1.7 Å resolution, providing a robust foundation for understanding its structural properties [116].
As illustrated in Figure 3B, SIRT2 features an active catalytic core along with helical extension at its N-terminal. The catalytic core comprises a standard NADH-binding site, which is divided into distinct domains: a smaller one and a larger one. The larger domain encompasses an inverted Rossmann fold, a distinctive motif typical of NAD+(H) binding sites, which contains six β-strands (βI-III and βVII-IX) and six α-helices (αI, αVII, αVIII, αX, αXI, and αXII). These structural elements are aligned parallel to the β-sheet (residues “53-89”, “146-186”, and “241-356”) [118]. The smaller domain is further categorized into Zinc binding and helical subdomains. The Zn-binding domain features three antiparallel β-sheets, one α-helix, and a Zn2+ ion coordinated by four conserved cysteine residues (Cys195, Cys200, Cys221, and Cys224) found across all Sir2-like enzymes. The helical domain comprises four helices, two short and two long. These two domains are connected through a short polypeptide chain in the smaller domain and three larger polypeptide chains in the larger domain, forming a prominent groove. NAD+ binding site is present within this groove, and the amino acids involved are highly conserved within the Sirtuin family. The NAD+ cofactor-binding pocket is divided into three regions: the adenine ribose moiety binds to the A-site, the nicotinamide ribose moiety interacts with the B- and C-sites, and the nicotinamide itself resides deep within the catalytic pocket (Figure 3C) [116]. Mutations at these sites lead to a complete loss of deacetylation activity, confirming that this large junction groove forms the core of the catalytic mechanism [116].
4. Epigenetic Regulators in Asthma: Writers, Readers, and Erasers of DNA Modifications
4.1. DNMT1 and DNMT3a Role in Asthma
DNMT1 is the most important enzyme which involved in the epigenetic regulation of gene expression and maintained the methylation pattern during replication. It has been reported by Verma et al in 2013 that overexpression of Socs3 along with low expression of DNMT1 and IL-6 in mouse models of asthma [119]. DNA methyltransferase DNMT3a also regulates Th2, particularly IL-13, expression. Loss of DNMT3a leads to decreased DNA methylation and changes in the H3K27Ac status at the IL-13 locus and is associated with increased lung inflammation in a mouse model of asthma [44].
4.2. Structure of DNMT1 and DNMT3a
Both DNMT1 and DNMT3s are multi-domain proteins, containing a large regulatory region in addition to the C-terminal MTase domain [120,121]. DNMT1 is comprised of ~1600 amino acids, with an N-terminal regulatory region covering two thirds of the sequence, a highly conserved (GK)n repeat and a C-terminal MTase domain [122]. In Ren et al. (2018) paper, the regulatory region is described as containing a 300 amino acid N-terminal domain (NTD) with protein/DNA interaction sites, followed by a replication foci-targeting sequence (RFTS) domain, a CXXC zinc finger domain, and a pair of bromo-adjacent-homology (BAH) domains, as shown in their figures and text [122]. The function of DNMT1 in replication-dependent DNA methylation maintenance is supported by its localization in replication foci during the S phase, and in vitro a 3–40-fold enzymatic preference for hemimethylated CpG sites [120,123], an epigenetic mark enriched at the replication foci [124]. DNMT3A and DNMT3B mediate DNA methylation establishment during gametogenesis and embryogenesis [125,126] and subsequently participate in methylation maintenance. DNMT3A and DNMT3B are highly related in sequence, both containing a largely disordered NTD, followed by a Pro-Trp-Trp-Pro (PWWP) domain, an Atrx-Dnmt3-Dnmt3l (ADD) domain and a highly homologous MTase domain. Ren et al. (2018) explored the structural basis of DNMT1 and DNMT3A-mediated DNA methylation, revealing how their catalytic and regulatory domains collaborate in substrate recognition. These insights are essential for understanding the role of DNMT1 DNMT3A in epigenetic regulation relevant to asthma pathogenesis.
4.3. Methyl-CpG-Binding Domain Protein 2 MBD2 Role in Asthma
Methyl-CpG-binding domain protein 2 (MBD2) is a key epigenetic regulator involved in the interaction between DNA methylation and chromatin remodeling. As a reader of DNA methylation, MBD2 significantly impacts processes such as stem cell differentiation and somatic cell reprogramming [127]. A recent study demonstrated that MBD2 promotes Th17 differentiation in mice through its interaction with MINK1, which regulates IL-17 secretion [128]. Further analysis highlighted MBD2’s potential role in asthma pathogenesis and allergic reactions by influencing CD4+ T cell differentiation. Additionally, MBD2 has been shown to facilitate Th17 cell differentiation by downregulating SOCS3 expression in severe asthmatic mice [129]. Notably, peripheral blood analysis in asthma patients revealed a correlation between serum MBD2 levels and Th17 cell populations, suggesting that MBD2 expression could serve as a biomarker for asthma endotypes [45].
4.4. Structure of Methyl-CpG-Binding Domain Protein 2 (MBD2)
The MBD2 protein consists of four distinct domains, arranged from N-terminal to C-terminal: a glycine-arginine repeat region (GR), a methyl-DNA-binding domain (MBD), an intrinsically disordered region (IDR), and a coiled-coil domain (CC) [130]. The methyl-DNA-binding domain (MBD) of MBD2 is the primary domain responsible for recognizing methylated DNA. It is a highly conserved structure within the protein family in humans and homologous species [131]. This approximately 80-amino acid domain, located between residues 145 and 225 in humans, is present in all MBD2 isoforms [132]. The MBD2MBD domain has been shown to interact with methylated residues across the genome, recognizing motifs such as mCG, mCAG, mCAT, mCC, and mCT di-/tri-nucleotide sequences with varying binding affinities [133]. MBD2MBD conservatively consists of 4 strands of β-sheet and one α-helix loop. β-sheets are centrally located and establish a finger like protrusion for specific pairing with mCpG and for the interaction with major groove of the DNA. Particularly, β2 and β3 are crucial for DNA binding, containing operational amino acids such as Val164, Arg166, Asp176, Tyr178, Arg188, and Ser189 [132,134,135]. The intrinsically disordered region (IDR) in MBD2 has not been fully characterized, as evidenced by AF3 with ID AF-Q9UBB5-F1, which also displays disorder in this region. MBD2 has three different isoforms called MBD2a, MBD2b, and MBD2c(t). There are differences in terms of cell-specific expression and domain architecture between MBD2 isoforms. From the structural perspective, MBD2a is the full-length isoform with 411 amino acids (~43 kDa). On the contrary, MBD2b lacks the N-terminal GR-rich domain because translation initiates from the second start codon in the alternate open reading frame. Thus, it results in a 262- amino acid (~ 29 kDa) peptide. MBD2c(t) is the truncated isoform, which is devoid of the C-terminal completely and includes 301 amino acids (~ 32 kDa). However, all isoforms retain the methyl-DNA-binding domain and DNA-binding properties [136].
4.5. Role of TET1 in Asthma and Its Structural Information
DNA demethylating enzymes (TET1, TET2, and TET3), also known as “erasers”, maintain a complex dynamic equilibrium that regulates gene expression [137]. A study using a mouse model of house dust mite–induced asthma reported an altered methylome, elevated levels of 5-hydroxymethylcytosine (5-hmC), and increased TET1 expression in the lungs, suggesting a potential role for TET1 in the pathogenesis of asthma [138]. Hypomethylation of the TET1 gene (located at 10q21.3) in the nasal epithelium has been associated with the development of asthma in children exposed to traffic-related air pollution. TET1 is responsible for converting 5-methylcytosine into several derivatives, including oxymethylcytosine, 5-formylcytosine, 5-carboxylcytosine, and 5-hydroxymethylcytosine, thereby facilitating DNA demethylation [46]. In mammals, three TET paralogs have been identified: TET1, TET2, and TET3 [139,140,141]. Each has a carboxyl-terminal catalytic core region comprising a Cys-rich domain and a double-stranded β helix (DSBH) domain with a large low-complexity insert [140,142]. The length of this low complexity insert varies greatly among TET paralog members, and the sequence is less conserved [139]. TET1 and TET3 also have an amino-terminal CXXC domain, which was described as a DNA-binding motif [140,141].
5. Epigenetic Regulators in Asthma: Writers, Readers, and Erasers of RNA Modifications
N6-methyladenosine (m6A) methylation is the most abundant post-transcriptional RNA modification in mRNAs, regulating gene expression and influencing RNA fate through the action of m6A “writer” (methyltransferase), “eraser” (demethylase), and “reader” (methylated reading protein) [143,144,145,146]. m6A is involved in inflammatory and lung diseases such as asthma, respiratory distress syndrome, pneumonia, and lung cancer [147,148,149,150]. Presently, m6A methylation gains more and more attention about its function and mechanism. In the asthma pathophysiological process, there is only primary probation and initiatory findings [151,152]. For example, Teng et al in 2021 found that dysregulated or hypermethylated m6A peaks in 329 mRNAs and 150 hypomethylated m6A peaks in 143 mRNAs in asthmatic mice [153]. In addition, Dai et al. 2021 found that 5 candidate m6A regulators (FMR1, KIAA1429, WTAP, YTHDF2, ZC3HAV1) are in close contact with the risk of childhood asthma. Therefore, these literatures inspire us that m6A may participate in the asthma pathology (Dai et al., 2021).
5.1. Role of Reader Protein YTHDF1 and YTHDF2 in Asthma
m6A modification mediates the recruitment of m6A readers that associate m6A-modified RNAs with mRNA processing enzymes to influence RNA export splicing, translation, and degradation [154,155,156,157]. YTHDF1 has been identified as a translation-facilitating m6A reader that recruits translation machinery to its target mRNAs in the cytoplasm (T. Liu et al., 2020; Yan et al., 2023). YTHDF1, a potent m6A reader protein, has been found to influence the macrophage inflammatory pathways, remarkably by modulating the JAK2/STAT3 signaling pathway. Li et al. (2022) indicated that YTHDF1 expression is raised in human macrophages infected with syphilis, where it negatively regulates macrophage-associated inflammation by inhibiting JAK2/STAT3-mRNA translation. This suppression of inflammatory responses also results in the decreased secretion of IL-1β, a critical inflammatory cytokine [160]. This process is functionally essential for asthma, since the inhibition of IL-1β can modulate the Th17/Treg immunological imbalance and reduce neutrophil-driven airway inflammation in an ovalbumin-induced asthma mode [161].
Additionally, YTHDF1's intimate interaction with the JAK2/STAT3 signaling pathway in macrophages has been observed in septicemic rats, enabling the induction of IL-17, which disrupts the Th1/Th17 equilibrium and accelerates inflammatory reactions [162]. This pathway, associated in asthma pathology, contributes to mast cell activation, bronchial smooth muscle thickening, and airway remodeling [163,164,165]. The therapeutic potential of targeting the JAK2/STAT3 pathway has been highlighted, as inhibition of this signaling cascade can reduce neutrophil activation and improve the efficacy of corticosteroids in asthma patients [166]. Recent research demonstrates that the YTHDF1 reader protein is increased in PDGF-BB-induced airway smooth muscle (ASM) cells, promoting cell proliferation and migration, which are critical events in airway remodeling related to asthma. Cyclin D1 (CCND1), a pivotal regulator of the cell cycle, is known as a downstream target of YTHDF1 via an m6A-dependent mechanism. YTHDF1 interacts with CCND1 mRNA, enhancing its stability and expression. The m6A-dependent modulation of CCND1 by YTHDF1 provides a new mechanism in asthma etiology, suggesting possible treatment approaches aimed at the YTHDF1-CCND1 axis [167]. YTHDF1 is significantly elevated in asthmatic patients and supports CLOCK translation in a m6A-dependent way. This drives NLRP3 inflammasome activation and IL-1β release, facilitating inflammatory responses in the airways
[50]. YTHDF2, a m6A reader, regulates AXIN1 mRNA stability in a m6A-dependent manner, governed through a m6A alteration introduced by WTAP. WTAP overexpression reduces AXIN1 stability, promoting excessive airway smooth muscle cell (ASMC) proliferation in asthma. However, the knockdown of YTHDF2 enhances AXIN1 stability, leading to suppression of the Wnt/β-catenin signaling pathway and reduction in ASMC proliferation. This proposes that the WTAP/YTHDF2 axis plays a pivotal role in regulating ASMC growth and offers potential therapeutic targets for asthma treatment [48]. Sun et al in 2021 employed bioinformatics methodologies such as COX regression and logistic regression to evaluate m6A modification patterns in 87 healthy controls and 344 severe asthma patients, identifying YTHDF3 and YTHDC1 as pivotal regulators influencing severe asthma prognosis and immune microenvironment. Both regulators demonstrate substantial adjustments in their methylation peaks in severe asthma, specifically in exons and 3′UTR regions, implying their decisive role in RNA processing. Correlation analysis further highlights their cooperative interaction with other m6A modifiers, indicating their joint contribution to asthma's development. Notably, YTHDF3 has a substantial positive correlation with eosinophil infiltration, highlighting its potential involvement in immune modulation within severe asthma [168].
5.2. Structure of YTHDF Proteins
Most of these diverse functions are realized via the m6A binding proteins containing the YT521-B homology (YTH) domain, so-called m6A “readers”[169,170,171,172]. The experimental structural data for the DF paralogs are limited to their YTH domains [173,174]. The three human DF proteins consist of a YTH domain of 145 residues located in the sequence between unstructured N-terminal and C-terminal segments of about 400 and 35 residues, respectively [175]. The m6A-RNA-binding YTH domains of the DF paralogs span 23–24% of their length and show >90% primary sequence similarity [176]. These domains share a conserved α/β fold with six β-strands and three α-helices [173]. They fold into a β-barrel where the α-helices are packed against it to stabilize the hydrophobic and electronegative m6A-binding cage [173,174,177]. In DF1 and DF3, the adjacent C-terminus and LCD also form α-helices, which further contact the domain [173,174]. Although such folding is predicted for DF2 [178] , a recent set of 19 DF2 crystals spanning its YTH domain reveal variability in the folding and configuration of this adjacent LCD region [179].
Despite their similarity, the available crystal structures reveal differences between the YTH domains of DF paralogs. The conserved α/β folds shape most of the m6A pocket; however, its outer wall forms a considerably flexible region, the m6A recognition loop. This loop contains several of the overall few non-conserved residues across these domains between DF paralogs. This loop has been suggested to easily adopt multiple conformations, to affect m6A binding affinity, and to be more flexible in DF1 via computer simulations [174]. This DF1-restricted property was attributed to the intrinsic flexibility of its non-conserved glycine 459 residue contained in the loop [174]. In addition, the surface electrostatic potential, lipophilicity, and B-factor (an estimate of thermal motion) profiles of the YTH domains demonstrate differences between paralogs [173,174]. Interestingly, the YTH domain of DF1 has a higher overall thermal stability, and this may underlie differences in m6A binding affinities. Importantly, according to glide modeling of the YTH domains of the DF paralogs bound to m6A RNAs, the m6A-binding free energy appear to be lower for DF1 than for DF2 or DF3 [173,174]. This finding suggests less spontaneous binding and thus weaker m6A affinity for the DF1 YTH domain.
Excluding the structurally well-characterized and highly conserved YTH domains, the DF paralog protein sequence identities demonstrate only 42–81% shared identity in their C-terminal regions and 47–64% identity in their N-terminal regions [176]. The N-terminal regions also contain sequences with <30% cross-paralog similarity that span up to 15% (85AA) of the protein [180]. YTH domain containing proteins recognize the m6A-binding sites through a conserved aromatic cage consisting of three tryptophan residues. In this review manuscript, we analyzed the full-length AlphaFold ([178]) predicted structures of YTHDF1(AF-Q9BYJ9-F1) and YTHDF2 (AF-Q9Y5A9-F1). While the N- and C-terminal regions showed low confidence scores, suggesting intrinsic disorder or flexibility, the YTH domains were modeled with high confidence, indicating well-defined and reliable structures (Figure 4). The YTH domain of DF1 consists of β1, α1, β2 from the C termini, α2 from the N-terminus, and the loop between β4 and β5 (Figure 4 (Left)). DF1 specifically recognizes m6A sites using m6A-binding pocket composed of Trp411, Trp465, and Trp470 [173]. DF2 consists of three α helices, eight β strands, and two 310 helices. The hydrophobic core consists of a β-barrel fold (β8–β1–β3–β4–β5–β2) and three α helices (Figure 6 (right)). Trp486 from the β4–β5 loop, Trp432 from the β2 strand and Trp491 from the β4–β5 loop are the basis of aromatic cage and can recognize m6A mononucleotide sites [181].
5.3. WTAP Role in Asthma
According to Deng et al. [182], the upregulation of WTAP protein expression endorsed Wnt signalling activation and triggered the excessive growth of pancreatic ductal adenocarcinoma cells. AXIN1 referred to as an axon, is a gene cloned from a mouse mutant. AXIN1 is known as an inhibitor of the Wnt signalling pathway [183]. Abnormal activation of the Wnt/β-catenin pathway is closely related to the occurrence of asthma [184]. Lin et al evaluated the consequences of RNA modification writers on the immune microenvironment in severe asthma by studying the relationship between immune cell populations and HLA gene expression alongside RNA writer modification patterns. Their findings suggest that WTAP, which was significantly increased in severe asthma, demonstrated a robust positive correlation with multiple immune cell types and HLA genes [185]. Hyperinflammation is frequently linked to elevated WTAP levels in inflammatory diseases. Numerous GEO datasets (GSE13887/137268/69063/97779/166388/208303) were investigated by Ge et al to explore the role of m6A modification in inflammation. The findings indicated that WTAP expression is consistently elevated in patients with systemic lupus erythematosus (SLE), asthma, sepsis, rheumatoid arthritis (RA), psoriasis, and Crohn’s disease, signifying its role in the inflammatory processes of these diseases [186].
5.4. Structure of WTAP Protein
Wilms’ tumor 1-associating protein (WTAP) is encoded on human chromosomal region 6q25.3 [187]. WTAP is a 44 kDa protein which is composed of 396 amino acids (Small et al., 2007). Investigation has shown that WTAP encompasses an elongated N-terminal, coiled-coil domain succeeded by a rather disordered C-terminal region [189]. WTAP localizes to both the nucleus and cytoplasm (Horiuchi et al., 2013; Ping et al., 2014). The entire structural model of the WTAP protein was retrieved from the AlphaFold Protein Structure Database (https://alphafold.ebi.ac.uk/) employing the entry ID AF-Q15007-F1 (Figure 5). The WTAP protein forms a symmetric parallel α-helical coiled-coil homodimer through hydrophobic interactions (X. Yan et al., 2022). WTAP is a key component in m6A modification, forming a complex with VIRMA, RBM15/15B, ZC3H13 (KIAA0853), CBLL1, and METTL3/14 [190]. WTAP encompasses an extended N-terminal, coiled-coil region followed by an unstructured C-terminal part [193]. WTAP regulates the localization of the stable heterodimer core complex of METTL3/14 into nuclear speckles through amino acids 5–13 of the nuclear localization signal (NLS) (-PLPKKVRL- to -PLPGGVGL-) at its N-terminus [189]. Notably, the N-terminal coiled-coil region (1–150 amino acids) that contains the NLS is the binding surface of METTL3, which links to the helical structure at the N-terminus of METTL3, called the leader helix (LH) [189]. Although WT1 was found to interact with WTAP, it was confirmed that WT1 was dispensable for the regulation of m6A modification by WTAP [191]. In the context of the METTL3-METTL14-WTAP-VIRMA complex, WTAP adopts a more compact conformation, interacting with VIRMA through three major interfaces. VIRMA, containing 17 armadillo-like repeats, forms a twisted α-solenoid structure, facilitating preferential m6A modification at 3′ UTRs. The structural integration of WTAP with the multicomponent methyltransferase complex (MTC) core enhances substrate recruitment and catalytic efficiency, underscoring WTAP’s role as a linker between VIRMA and the METTL3-METTL14 core, forming a stable quaternary complex vital for m6A methylation activity [192].
5.5. METTL3 and METTL14 Role in Asthma
The overexpression of METTL3 in asthma models helps to increase the m6A modification of GPX4, a key protein that protects cells from ferroptosis (a type of cell death). By upregulating GPX4, METTL3 inhibits ferroptosis in bronchial epithelial cells, which promotes cell survival, proliferation, and alleviates asthma symptoms. Therefore, METTL3 overexpression plays a protective role in asthma by preventing ferroptosis and supporting cell function [194]. Previous studies have shown that METTL3 reduces and negatively regulates Th2 cell differentiation in allergic asthma [28]. In T2 asthma, a marked decrease in METTL3 expression was observed at clinical, cellular, and animal levels, correlating with increased disease severity and airway inflammation. Low METTL3 levels promote inflammation and exacerbate asthma by activating M2-type macrophages. Additionally, silencing METTL3 in antigen-presenting HBE cells enhanced Th2 cell differentiation, underscoring METTL3's critical role in regulating the immune response and airway inflammation in T2 asthma. These findings highlight METTL3 as a key modulator of asthma pathogenesis [195]. Remarkably, recent research reported that the m6A writer METTL14 is decreased significantly both in mRNA and protein level in lung of mice with allergic asthma compared with control [196]. In another study, METTL14 was suggested to play a crucial role in the development of respiratory allergic diseases and the infiltration of immune cells [197]. Although previous studies have demonstrated the critical role of METTL14 in inflammatory diseases and EMT, the regulatory role of METTL14 in asthma remains obscure. Wang et al. reported that METTL14 expression was significantly downregulated in asthma, as shown by analysis of three GEO datasets (GSE104468, GSE165934, and GSE74986). Consistently, METTL14 levels were reduced in lung tissues of OVA-induced asthmatic mice and in TGF-β1-stimulated human bronchial epithelial cells [195].
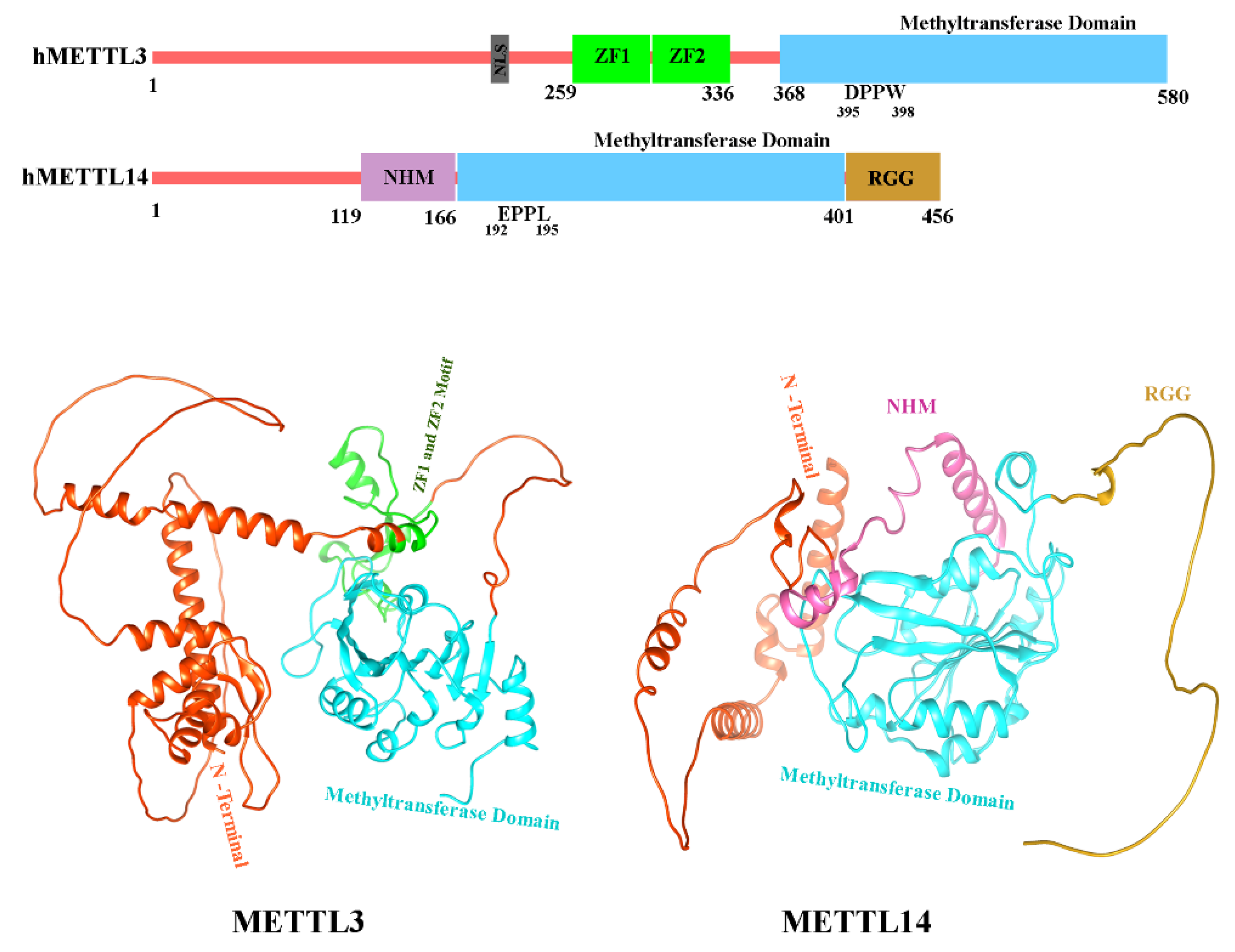
5.6. Structure of METTL3 and METTL14
In humans, two methyltransferases, METTL3 (sometimes stated as MT-A70) and METTL14, function as writer in m6A modification (J. Liu et al., 2014; Y. Wang et al., 2014). Both proteins belong to the class I MTase family [200] and they form a core catalytic complex that is regulated by an additional subunit, Wilms’ tumour 1-associating protein (WTAP) [191,193,201]. Independently, METTL3 and METTL14 demonstrate minimal in vitro methyltransferase activity and need to be in complex with one another for optimal catalytic activity. Interestingly, catalytic activity requires full-length METTL3, while only the central methyltransferase domain of METTL14 is required [202]. METTL3 comprises a low-complexity region at the N-terminus that contains a nuclear localization signal (NLS). This is followed by a zinc finger domain that contains two consecutive CCCH zinc finger motifs (ZF1 and ZF2) responsible for substrate binding, a disordered linker, and a C-terminal methyltransferase domain (Figure 6) with a classic Rossman fold architecture [203,204]. METTL14 contains a non-functional methyltransferase domain which is bordered by an N-terminal helical motif (NHM) and a C-terminal arginine-rich region (RGG; Figure 6) (Yoshida et al., 2022). METTL3’s Rossmann fold holds a catalytic loop with the classical DPPW motif [206], while METTL14 has a nonfunctional EPPL motif (Figure 6). The ZF1 and ZF2 domains of METTL3 act as RNA recognition elements which enable the binding of the mRNA substrate and position the DRACH sequence for methylation. Though, both subunits of the METTL3/14 complex are required for RNA binding (Śledź & Jinek, 2016). The C-terminus RGG of METTL14 exhibits positively charged and additionally participates to the RNA binding of the complex [189]. METTL3 and METTL14 in complex also form a positively charged groove at their protein-protein interface, which is critical for full methyltransferase activity as it also contributes to interactions with the mRNA substrate [204]. More in-depth analysis of the structural aspects of METTL3/14 RNA binding can be found in [202,207,208].
Figure 6.
Domain architecture and 3-dimensional illustration of METTL3 and METTL14. The panel on the top depicts the domain structure of human METTL3 and METTL14. The key domains are marked. The lower panel illustrates the 3D structures of METTL3 (left) and METTL14 (right), with the N-terminal, Methyltransferase Domain, and other significant domains accentuated in different colors for clarity. The protein structures are visualized to demonstrate the spatial arrangement of the domains.
Figure 6.
Domain architecture and 3-dimensional illustration of METTL3 and METTL14. The panel on the top depicts the domain structure of human METTL3 and METTL14. The key domains are marked. The lower panel illustrates the 3D structures of METTL3 (left) and METTL14 (right), with the N-terminal, Methyltransferase Domain, and other significant domains accentuated in different colors for clarity. The protein structures are visualized to demonstrate the spatial arrangement of the domains.
5.7. Role of IGF2BP2 in Asthma
IGF2BPs have recently been noted for their role in modulating biological processes including as development, cancer, and stemness. Recent discoveries reveal that IGF2BP2 is crucial for the recognition of m6A alterations and significantly affects mRNA stability and translation [209]. The significance of IGF2BP2 in macrophage polarization as a m6A reader is still ambiguous. Wang et al. (2021) observed that IGF2BP2 expression raised through macrophage differentiation into M1 and M2 phenotypes, with IL-4-induced overexpression being dependent on STAT6. Their loss-of-function investigations suggested that IGF2BP2 favorably regulates IL-4-driven M2 activation while negatively impacting proinflammatory responses. Furthermore, m6A-RIP-qPCR identified TSC1 as a m6A-modified target recognized by IGF2BP2, pointing to its role in macrophage polarization [52]. This indicates that IGF2BP2 facilitates M2 macrophage polarization, hence intensifying allergic inflammation in asthma.
5.8. Structure of IGF2BP2
The m6A reader protein IGF2BP2 identifies and binds to RNA via its unique structural configuration, which consists of six specific RNA-binding domains. This structure has two RNA recognition motifs (RRM1 and RRM2) and four K Homology (KH) domains (KH1–KH4)
[210]. Structural insights into IGF2BP2 were derived from the protein structure predicted by AlphaFold3, accessible in the AlphaFold Protein Structure Database (ID: AF-F8W930-F1). The KH3-4 didomain is crucial for RNA binding, with the conserved 503GKGG506 motif of the KH4 domain and adjacent hydrophobic residues forming a persistent hydrophobic groove necessary for the selective recognition of the GGm6AC RNA core motif, as evidenced by Fakhar et al in 2024 [211].
5.9. Role of FTO in Asthma
FTO, a m6A demethylase, is pivotal in asthma through modulating inflammatory responses in epithelial cells [212]. Recent evidence demonstrates that FTO, via its action on m6A sites, increases the stability and expression of IKBKB, thereby mitigating epithelial barrier degradation and inflammation in airway epithelial cells exposed to PM2.5 by regulating NF-κB signaling, emphasizing its critical role in asthma cause [53]. Research using mouse models uncovered that FTO knockdown accelerated airway inflammation by increasing IL-13 and decreasing IL-12b expression, potentially through FTO-mediated m6A modification of Foxj1 mRNA, a critical regulator of motor ciliogenesis. These alterations in cytokine profiles, along with impaired Foxj1 expression, contribute to epithelial dysfunction and airway inflammation in asthma [148].
5.10. Structure of FTO
The fat mass and obesity associated gene (FTO) encodes 505 amino acids long protein which belongs to the AlkB family of non-heme iron and 2-oxoglutarate (2-OG)-dependent dioxygenases. This family of enzymes assists in demethylation activities, indicating the valuable function of FTO in oxidative reactions that demethylate RNA [213]. A large number of the human FTO protein X-ray crystallographic structures have been added to the Protein Data Bank (PDB) (https://www.rcsb.org/), revealing thorough structural information about the protein's catalytic processes, substrate selectivity, and interactions against various ligands and inhibitors [214,215,216,217,218,219]. Two separate domains can be observed in the crystal structure of FTOΔ31 (PDB ID:3LFM): the C-terminal domain (CTD; 327–498 AA) and the N-terminal domain (NTD; 32–326 AA). Understanding the structure of FTO's catalytic pocket and enzymatic activity has been made possible by its crystal structure [216]. Due to the lack of the N-terminal 32 residues in the crystallized construct, we utilized the AlphaFold3-predicted structure (Figure 7) (AF-Q9C0B1-F1), which reflects the complete FTO protein, to enable a more thorough structural investigation.
6. Conclusions and Perspective
This study highlights the central role of epigenetically important proteins classified as writers, erasers, and readers of histone, DNA, and RNA modifications in asthma pathogenesis. Dysregulation of histone acetyltransferases (e.g., p300/CBP) and deacetylases (e.g., SIRT1–SIRT7) alters chromatin accessibility, leading to pro-inflammatory gene expression. DNA methylation enzymes such as DNMT1 and DNMT3A, along with demethylases like TET1 and readers like MBD2, modulate immune gene profiles. Additionally, m6A RNA modifiers METTL3/14 (writers), FTO (eraser), YTHDF and IGF2BP2 proteins (readers) emerge as pivotal regulators affecting asthma susceptibility and severity by influencing inflammatory pathways, immune response modulation, and airway remodeling. Structurally, these proteins feature unique and conserved domains crucial for their catalytic and regulatory functions, as highlighted by detailed protein domain analyses, sequence similarities, and structural predictions presented in the manuscript. Understanding these proteins’ molecular mechanisms and their interactions could uncover novel biomarkers for asthma diagnosis and management and open promising avenues for targeted therapies.
Therapeutically, targeting these epigenetic regulators offers promising strategies to reverse aberrant gene expression and improve asthma outcomes. HDAC modulators, DNA methylation inhibitors, and m6A-targeted drugs may complement or enhance existing therapies. Future research should focus on phenotype-specific epigenetic profiling, structure-based drug design, and combinatorial approaches that integrate epigenetic and immunological interventions to achieve precision medicine in asthma care.
Author Contributions
M.F. contributed to the investigation of research and wrote the original draft. M.G. handled data curation and created visualizations. W.L. managed the project, secured funding, provided necessary resources, and reviewed and edited the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Shenzhen Science and Technology Program (Grant No. JCYJ20240813142512017) and the Natural Science Foundation of Guangdong Province, China (Grant No. 2023A1515010471).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
I gratefully acknowledge the unwavering inspiration and joy brought by my daughter, Isbah Fakhar, who was just three months old at the time of this submission. Her presence provided strength and motivation throughout the research journey.
Conflicts of Interest
The authors declare no conflict of interest
Abbreviations
The following abbreviations are used in this manuscript:
| CBP | CREB Binding Protein |
| SIRT | Silent Information Regulator Proteins |
| DNMT1 | DNA (cytosine-5)-Methyltransferase 1 |
| IGF2BP2 | Insulin-like Growth Factor 2 mRNA-Binding Protein 2 |
| MBD2 | Methyl-CpG-binding domain protein 2 |
| TET1 | Ten-Eleven Translocation Methylcytosine Dioxygenase 1 |
| FTO | Fat Mass and Obesity Associated Gene |
| RNA | Ribonucleic Acid |
| DNA | Deoxyribonucleic Acid |
| PTMs | Posttranslational Modifications |
| HATs | Histone Acetylases |
| HDACs | Histone Deacetylases |
| TFs | Transcription Factors |
| METTL3 | Methyltransferase-like 3 |
| WTAP | Wilms’ Tumor 1-Associating Protein |
| IBiD | interferon-binding transactivation domain |
| PWWP | Pro-Trp-Trp-Pro |
| ADD | Atrx-Dnmt3-Dnmt3l |
| DSBH | double-stranded β helix |
| IL-17 | Interleukin-17 |
| ASMC | airway Smooth Muscle Cell |
| ZF1 | zinc finger motifs |
| RRM1 | RNA Recognition Motif 1 |
| RRM2 | RNA Recognition Motif 2 |
| KH | K Homology domain |
References
- Erle, D.J.; Sheppard, D. The Cell Biology of Asthma. Journal of Cell Biology 2014, 205, 621–631. [Google Scholar] [CrossRef] [PubMed]
- Lambrecht, B.N.; Hammad, H. The Immunology of Asthma. Nat Immunol 2015, 16, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Hammad, H.; Lambrecht, B.N. The Basic Immunology of Asthma. Cell 2021, 184, 1469–1485. [Google Scholar] [CrossRef]
- Huang, K.; Yang, T.; Xu, J.; Yang, L.; Zhao, J.; Zhang, X.; Bai, C.; Kang, J.; Ran, P.; Shen, H.; et al. Prevalence, Risk Factors, and Management of Asthma in China: A National Cross-Sectional Study. The Lancet 2019, 394, 407–418. [Google Scholar] [CrossRef]
- Xie, M.; Liu, X.; Cao, X.; Guo, M.; Li, X. Trends in Prevalence and Incidence of Chronic Respiratory Diseases from 1990 to 2017. Respir Res 2020, 21, 1–13. [Google Scholar] [CrossRef]
- Stikker, B.S.; Hendriks, R.W.; Stadhouders, R. Decoding the Genetic and Epigenetic Basis of Asthma. Allergy 2023, 78, 940–956. [Google Scholar] [CrossRef]
- Kapri, A.; Pant, S.; Gupta, N.; Paliwal, S.; Nain, S. Asthma History, Current Situation, an Overview of Its Control History, Challenges, and Ongoing Management Programs: An Updated Review. Proceedings of the National Academy of Sciences, India Section B: Biological Sciences 2023, 93, 539–551. [Google Scholar] [CrossRef]
- Papi, A.; Brightling, C.; Pedersen, S.E.; Reddel, H.K. Seminar Asthma. Lancet 2018, 391, 783–800. [Google Scholar] [CrossRef] [PubMed]
- Edris, A.; den Dekker, H.T.; Melén, E.; Lahousse, L. Epigenome-Wide Association Studies in Asthma: A Systematic Review. Clinical & Experimental Allergy 2019, 49, 953–968. [Google Scholar]
- DeVries, A.; Vercelli, D. Epigenetic Mechanisms in Asthma. Ann Am Thorac Soc 2016, 13, S48–S50. [Google Scholar] [CrossRef]
- Lovinsky-Desir, S.; Miller, R.L. Epigenetics, Asthma, and Allergic Diseases: A Review of the Latest Advancements. Curr Allergy Asthma Rep 2012, 12, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, Z.; Mortaz, E.; Adcock, I.; Moin, M. Role of Epigenetics in the Pathogenesis of Asthma. Iran J Allergy Asthma Immunol 2017, 82–91. [Google Scholar]
- Kabesch, M.; Adcock, I.M. Epigenetics in Asthma and COPD. Biochimie 2012, 94, 2231–2241. [Google Scholar] [CrossRef] [PubMed]
- Alashkar Alhamwe, B.; Miethe, S.; von Strandmann, E.; Potaczek, D.P.; Garn, H. Epigenetic Regulation of Airway Epithelium Immune Functions in Asthma. Front Immunol 2020, 11, 1747. [Google Scholar] [CrossRef]
- Yang, I. V; Pedersen, B.S.; Liu, A.H.; O’Connor, G.T.; Pillai, D.; Kattan, M.; Misiak, R.T.; Gruchalla, R.; Szefler, S.J.; Hershey, G.K.K.; et al. The Nasal Methylome and Childhood Atopic Asthma. Journal of Allergy and Clinical Immunology 2017, 139, 1478–1488. [Google Scholar] [CrossRef]
- Bae, D.-J.; Jun, J.; Chang, H.S.; Park, J.S.; Park, C.-S. Epigenetic Changes in Asthma: Role of DNA CpG Methylation. Tuberc Respir Dis (Seoul) 2020, 83, 1–13. [Google Scholar] [CrossRef]
- Hudon Thibeault, A.-A.; Laprise, C. Cell-Specific DNA Methylation Signatures in Asthma. Genes (Basel) 2019, 10, 932. [Google Scholar] [CrossRef]
- Reinberg, D.; Allis, D.; Jenuwein, T. Epigenetics 2007.
- Arrowsmith, C.H.; Bountra, C.; Fish, P. V; Lee, K.; Schapira, M. Epigenetic Protein Families: A New Frontier for Drug Discovery. Nat Rev Drug Discov 2012, 11, 384–400. [Google Scholar] [CrossRef]
- Lacal, I.; Ventura, R. Epigenetic Inheritance: Concepts, Mechanisms and Perspectives. Front Mol Neurosci 2018, 11, 292. [Google Scholar] [CrossRef]
- Alberts, B.; Johnson, A.; Lewis, J.; Raff, M.; Roberts, K.; Walter, P. Chromosomal DNA and Its Packaging in the Chromatin Fiber. In Molecular Biology of the Cell. 4th edition; Garland science, 2002.
- Harb, H.; Alhamwe, B.A.; Garn, H.; Renz, H.; Potaczek, D.P. Recent Developments in Epigenetics of Pediatric Asthma. Curr Opin Pediatr 2016, 28, 754–763. [Google Scholar] [CrossRef]
- Potaczek, D.P.; Harb, H.; Michel, S.; Alhamwe, B.A.; Renz, H.; Tost, J. Epigenetics and Allergy: From Basic Mechanisms to Clinical Applications. Epigenomics 2017, 9, 539–571. [Google Scholar] [CrossRef]
- Gomes, C.P. da C.; Schroen, B.; Kuster, G.M.; Robinson, E.L.; Ford, K.; Squire, I.B.; Heymans, S.; Martelli, F.; Emanueli, C.; Devaux, Y.; et al. Regulatory RNAs in Heart Failure. Circulation 2020, 141, 313–328. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, O.; Baccarelli, A.A. Environmental Health and Long Non-Coding RNAs. Curr Environ Health Rep 2016, 3, 178–187. [Google Scholar] [CrossRef]
- Hu, G.; Niu, F.; Humburg, B.A.; Liao, K.; Bendi, S.; Callen, S.; Fox, H.S.; Buch, S. Molecular Mechanisms of Long Noncoding RNAs and Their Role in Disease Pathogenesis. Oncotarget 2018, 9, 18648. [Google Scholar] [CrossRef]
- Sharma, S.; Yang, I. V; Schwartz, D.A. Epigenetic Regulation of Immune Function in Asthma. Journal of Allergy and Clinical Immunology 2022, 150, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Liu, L.; Huang, S.; Xiao, W.; Gao, Y.; Zhou, W.; Zhang, C.; Zheng, H.; Yang, L.; Xie, X.; et al. RNA M6A Methylation Modulates Airway Inflammation in Allergic Asthma via PTX3-Dependent Macrophage Homeostasis. Nat Commun 2023, 14, 7328. [Google Scholar] [CrossRef] [PubMed]
- Wawrzyniak, P.; Krawczyk, K.; Swati, A.; Tan, G.; Wawrzyniak, M.; Karouzakis, E.; Dreher, A.; Jakiela, B.; Altunbulakli, C.; Sanak, M.; et al. Inhibition of CpG Methylation Improves the Barrier Integrity of Bronchial Epithelial Cells in Asthma. Authorea Preprints 2020. [Google Scholar] [CrossRef]
- Seumois, G.; Chavez, L.; Gerasimova, A.; Lienhard, M.; Omran, N.; Kalinke, L.; Vedanayagam, M.; Ganesan, A.P. V; Chawla, A.; Djukanović, R.; et al. Epigenomic Analysis of Primary Human T Cells Reveals Enhancers Associated with TH2 Memory Cell Differentiation and Asthma Susceptibility. Nat Immunol 2014, 15, 777–788. [Google Scholar] [CrossRef]
- Gerasimova, A.; Chavez, L.; Li, B.; Seumois, G.; Greenbaum, J.; Rao, A.; Vijayanand, P.; Peters, B. Predicting Cell Types and Genetic Variations Contributing to Disease by Combining GWAS and Epigenetic Data. PLoS One 2013, 8, e54359. [Google Scholar] [CrossRef]
- Stefanowicz, D.; Lee, J.Y.; Lee, K.; Shaheen, F.; Koo, H.-K.; Booth, S.; Knight, D.A.; Hackett, T.-L. Elevated H3K18 Acetylation in Airway Epithelial Cells of Asthmatic Subjects. Respir Res 2015, 16, 1–13. [Google Scholar] [CrossRef]
- Hamamoto, R.; Furukawa, Y.; Morita, M.; Iimura, Y.; Silva, F.P.; Li, M.; Yagyu, R.; Nakamura, Y. SMYD3 Encodes a Histone Methyltransferase Involved in the Proliferation of Cancer Cells. Nat Cell Biol 2004, 6, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Foreman, K.W.; Brown, M.; Park, F.; Emtage, S.; Harriss, J.; Das, C.; Zhu, L.; Crew, A.; Arnold, L.; Shaaban, S.; et al. Structural and Functional Profiling of the Human Histone Methyltransferase SMYD3. PLoS One 2011, 6, e22290. [Google Scholar] [CrossRef]
- Wawrzyniak, P.; Wawrzyniak, M.; Wanke, K.; Sokolowska, M.; Bendelja, K.; Rückert, B.; Globinska, A.; Jakiela, B.; Kast, J.I.; Idzko, M.; et al. Regulation of Bronchial Epithelial Barrier Integrity by Type 2 Cytokines and Histone Deacetylases in Asthmatic Patients. Journal of Allergy and Clinical Immunology 2017, 139, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Bhavsar, P.; Ahmad, T.; Adcock, I.M. The Role of Histone Deacetylases in Asthma and Allergic Diseases. Journal of allergy and clinical immunology 2008, 121, 580–584. [Google Scholar] [CrossRef] [PubMed]
- Leus, N.G.J.; Zwinderman, M.R.H.; Dekker, F.J. Histone Deacetylase 3 (HDAC 3) as Emerging Drug Target in NF-ΚB-Mediated Inflammation. Curr Opin Chem Biol 2016, 33, 160–168. [Google Scholar] [CrossRef]
- Liu, Y.; Shi, G. Roles of Sirtuins in Asthma. Respir Res 2022, 23, 251. [Google Scholar] [CrossRef]
- Kim, Y.-Y.; Hur, G.; Lee, S.W.; Lee, S.-J.; Lee, S.; Kim, S.-H.; Rho, M.-C. AGK2 Ameliorates Mast Cell-Mediated Allergic Airway Inflammation and Fibrosis by Inhibiting FcRI/TGF-β Signaling Pathway. Pharmacol Res 2020, 159, 105027. [Google Scholar] [CrossRef]
- Song, J.; Wang, J. SIRT3 Regulates Bronchial Epithelium Apoptosis and Aggravates Airway Inflammation in Asthma. Mol Med Rep 2022, 25, 144. [Google Scholar] [CrossRef]
- Jang, H.-Y.; Gu, S.; Lee, S.-M.; Park, B.-H. Overexpression of Sirtuin 6 Suppresses Allergic Airway Inflammation through Deacetylation of GATA3. Journal of Allergy and Clinical Immunology 2016, 138, 1452–1455. [Google Scholar] [CrossRef]
- Fang, P.; Xue, Y.; Zhang, Y.; Fan, N.; Ou, L.; Leng, L.; Pan, J.; Wang, X. SIRT7 Regulates the TGF-Β1-Induced Proliferation and Migration of Mouse Airway Smooth Muscle Cells by Modulating the Expression of TGF-β Receptor I. Biomedicine & Pharmacotherapy 2018, 104, 781–787. [Google Scholar]
- Verma, M.; Chattopadhyay, B.D.; Kumar, S.; Kumar, K.; Verma, D. DNA Methyltransferase 1 (DNMT1) Induced the Expression of Suppressors of Cytokine Signaling3 (Socs3) in a Mouse Model of Asthma. Mol Biol Rep 2014, 41, 4413–4424. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Zhou, B.; Zhang, Y.; Nguyen, E.T.; Du, J.; Glosson, N.L.; Kaplan, M.H. DNA Methyltransferase 3a Limits the Expression of Interleukin-13 in T Helper 2 Cells and Allergic Airway Inflammation. Proceedings of the National Academy of Sciences 2012, 109, 541–546. [Google Scholar] [CrossRef]
- Chen, Z.; Yuan, Y.; He, Y.; Wasti, B.; Duan, W.; Jia, J.; Li, D.; Xiao, B.; Zhang, D.; Ma, L.; et al. MBD2 as a Potential Novel Biomarker for Identifying Severe Asthma with Different Endotypes. Front Med (Lausanne) 2021, 8, 693605. [Google Scholar] [CrossRef]
- Somineni, H.K.; Zhang, X.; Myers, J.M.B.; Kovacic, M.B.; Ulm, A.; Jurcak, N.; Ryan, P.H.; Hershey, G.K.K.; Ji, H. Ten-Eleven Translocation 1 (TET1) Methylation Is Associated with Childhood Asthma and Traffic-Related Air Pollution. Journal of Allergy and Clinical Immunology 2016, 137, 797–805. [Google Scholar] [CrossRef]
- Dai, B.; Sun, F.; Cai, X.; Li, C.; Liu, H.; Shang, Y. Significance of RNA N6-Methyladenosine Regulators in the Diagnosis and Subtype Classification of Childhood Asthma Using the Gene Expression Omnibus Database. Front Genet 2021, 12, 634162. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Dai, L. WTAP Promotes the Excessive Proliferation of Airway Smooth Muscle Cells in Asthma by Enhancing AXIN1 Levels Through the Recognition of YTHDF2. Biochem Genet 2024, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Shang, Y.; Zhang, X.; Duan, W.; Li, J.; Zhu, L.; Ma, L.; Xiang, X.; Jia, J.; Ji, X.; et al. METTL3 Mediates SOX5 M6A Methylation in Bronchial Epithelial Cells to Attenuate Th2 Cell Differentiation in T2 Asthma. Heliyon 2024, 10. [Google Scholar] [CrossRef]
- Wang, J.; Zhou, Y.; Zhang, M.; Wu, Y.; Wu, Q.; Su, W.; Xu, M.; Wu, J.; Zhang, M.; Shuai, J.; et al. YTHDF1-CLOCK Axis Contributes to Pathogenesis of Allergic Airway Inflammation through LLPS. Cell Rep 2024, 43. [Google Scholar] [CrossRef]
- Gao, G.; Hao, Y.Q.; Wang, C.; Gao, P. Role and Regulators of N6-Methyladenosine (M6A) RNA Methylation in Inflammatory Subtypes of Asthma: A Comprehensive Review. Front Pharmacol 2024, 15, 1360607. [Google Scholar] [CrossRef]
- Wang, X.; Ji, Y.; Feng, P.; Liu, R.; Li, G.; Zheng, J.; Xue, Y.; Wei, Y.; Ji, C.; Chen, D.; et al. The M6A Reader IGF2BP2 Regulates Macrophage Phenotypic Activation and Inflammatory Diseases by Stabilizing TSC1 and PPARγ. Advanced Science 2021, 8, 2100209. [Google Scholar] [CrossRef]
- Xiong, A.; He, X.; Liu, S.; Ran, Q.; Zhang, L.; Wang, J.; Jiang, M.; Niu, B.; Xiong, Y.; Li, G. Oxidative Stress-Mediated Activation of FTO Exacerbates Impairment of the Epithelial Barrier by up-Regulating IKBKB via N6-Methyladenosine-Dependent MRNA Stability in Asthmatic Mice Exposed to PM2. 5. Ecotoxicol Environ Saf 2024, 272, 116067. [Google Scholar] [CrossRef]
- Potaczek, D.P.; Bazan-Socha, S.; Wypasek, E.; Wygrecka, M.; Garn, H. Recent Developments in the Role of Histone Acetylation in Asthma. Int Arch Allergy Immunol 2024, 185, 641–651. [Google Scholar] [CrossRef] [PubMed]
- Alaskhar Alhamwe, B.; Khalaila, R.; Wolf, J.; von Bülow, V.; Harb, H.; Alhamdan, F.; Hii, C.S.; Prescott, S.L.; Ferrante, A.; Renz, H.; et al. Histone Modifications and Their Role in Epigenetics of Atopy and Allergic Diseases. Allergy, Asthma & Clinical Immunology 2018, 14, 1–16. [Google Scholar]
- Swygert, S.G.; Peterson, C.L. Chromatin Dynamics: Interplay between Remodeling Enzymes and Histone Modifications. Biochimica et Biophysica Acta (BBA)-Gene Regulatory Mechanisms 2014, 1839, 728–736. [Google Scholar] [CrossRef]
- Sun, X.-J.; Man, N.; Tan, Y.; Nimer, S.D.; Wang, L. The Role of Histone Acetyltransferases in Normal and Malignant Hematopoiesis. Front Oncol 2015, 5, 108. [Google Scholar] [CrossRef] [PubMed]
- Wapenaar, H.; Dekker, F.J. Histone Acetyltransferases: Challenges in Targeting Bi-Substrate Enzymes. Clin Epigenetics 2016, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Marmorstein, R.; Zhou, M.-M. Writers and Readers of Histone Acetylation: Structure, Mechanism, and Inhibition. Cold Spring Harb Perspect Biol 2014, 6, a018762. [Google Scholar] [CrossRef]
- Adcock, I.M.; Ford, P.; Ito, K.; Barnes, P.J. Epigenetics and Airways Disease. Respir Res 2006, 7, 1–19. [Google Scholar] [CrossRef]
- Hirschey, M.D. Old Enzymes, New Tricks: Sirtuins Are NAD+-Dependent de-Acylases. Cell Metab 2011, 14, 718–719. [Google Scholar] [CrossRef]
- Potaczek, D.P.; Alashkar Alhamwe, B.; Miethe, S.; Garn, H. Epigenetic Mechanisms in Allergy Development and Prevention. In Allergic diseases–from basic mechanisms to comprehensive management and prevention; Springer, 2021; pp. 331–357.
- Gong, F.; Chiu, L.-Y.; Miller, K.M. Acetylation Reader Proteins: Linking Acetylation Signaling to Genome Maintenance and Cancer. PLoS Genet 2016, 12, e1006272. [Google Scholar] [CrossRef]
- Zhu, H.; Wei, T.; Cai, Y.; Jin, J. Small Molecules Targeting the Specific Domains of Histone-Mark Readers in Cancer Therapy. Molecules 2020, 25, 578. [Google Scholar] [CrossRef]
- Ogasawara, T.; Hatano, M.; Satake, H.; Ikari, J.; Taniguchi, T.; Tsuruoka, N.; Watanabe-Takano, H.; Fujimura, L.; Sakamoto, A.; Hirata, H.; et al. Development of Chronic Allergic Responses by Dampening Bcl6-Mediated Suppressor Activity in Memory T Helper 2 Cells. Proceedings of the National Academy of Sciences 2017, 114, E741–E750. [Google Scholar] [CrossRef] [PubMed]
- Barnes, P.J.; Adcock, I.M.; Ito, K. Histone Acetylation and Deacetylation: Importance in Inflammatory Lung Diseases. European Respiratory Journal 2005, 25, 552–563. [Google Scholar] [CrossRef]
- Gunawardhana, L.P.; Gibson, P.G.; Simpson, J.L.; Powell, H.; Baines, K.J. Activity and Expression of Histone Acetylases and Deacetylases in Inflammatory Phenotypes of Asthma. Clinical & Experimental Allergy 2014, 44, 47–57. [Google Scholar]
- Zhuang, L.-L.; Jin, R.; Zhu, L.-H.; Xu, H.-G.; Li, Y.; Gao, S.; Liu, J.-Y.; Zhou, G.-P. Promoter Characterization and Role of CAMP/PKA/CREB in the Basal Transcription of the Mouse ORMDL3 Gene. PLoS One 2013, 8, e60630. [Google Scholar] [CrossRef] [PubMed]
- Jin, R.; Xu, H.-G.; Yuan, W.-X.; Zhuang, L.-L.; Liu, L.-F.; Jiang, L.; Zhu, L.-H.; Liu, J.-Y.; Zhou, G.-P. Mechanisms Elevating ORMDL3 Expression in Recurrent Wheeze Patients: Role of Ets-1, P300 and CREB. Int J Biochem Cell Biol 2012, 44, 1174–1183. [Google Scholar] [CrossRef] [PubMed]
- Hasan, S.; Hottiger, M.O. Histone Acetyl Transferases: A Role in DNA Repair and DNA Replication. J Mol Med 2002, 80, 463–474. [Google Scholar] [CrossRef]
- Wang, F.; Marshall, C.B.; Ikura, M. Transcriptional/Epigenetic Regulator CBP/P300 in Tumorigenesis: Structural and Functional Versatility in Target Recognition. Cellular and Molecular Life Sciences 2013, 70, 3989–4008. [Google Scholar] [CrossRef]
- Berdasco, M.; Esteller, M. Genetic Syndromes Caused by Mutations in Epigenetic Genes. Hum Genet 2013, 132, 359–383. [Google Scholar] [CrossRef]
- Cheng, Q.; Shang, Y.; Huang, W.; Zhang, Q.; Li, X.; Zhou, Q. P300 Mediates the Histone Acetylation of ORMDL3 to Affect Airway Inflammation and Remodeling in Asthma. Int Immunopharmacol 2019, 76, 105885. [Google Scholar] [CrossRef]
- Chen, Y.; Huang, B.; Zhao, Y.; Qi, D.; Wang, D. Increased P300/CBP Expression in Acute Respiratory Distress Syndrome Is Associated with Interleukin-17 and Prognosis. Clin Respir J 2020, 14, 791–799. [Google Scholar] [CrossRef] [PubMed]
- Yee, S.-P.; Branton, P.E. Detection of Cellular Proteins Associated with Human Adenovirus Type 5 Early Region 1A Polypeptides. Virology 1985, 147, 142–153. [Google Scholar] [CrossRef] [PubMed]
- Whyte, P.; Williamson, N.M.; Harlow, E.D. Cellular Targets for Transformation by the Adenovirus E1A Proteins. Cell 1989, 56, 67–75. [Google Scholar] [CrossRef]
- Chrivia, J.C.; Kwok, R.P.S.; Lamb, N.; Hagiwara, M.; Montminy, M.R.; Goodman, R.H. Phosphorylated CREB Binds Specifically to the Nuclear Protein CBP. Nature 1993, 365, 855–859. [Google Scholar] [CrossRef]
- Avantaggiati, M.L.; Carbone, M.; Graessmann, A.; Nakatani, Y.; Howard, B.; Levine, A.S. The SV40 Large T Antigen and Adenovirus E1a Oncoproteins Interact with Distinct Isoforms of the Transcriptional Co-Activator, P300. EMBO J 1996, 15, 2236–2248. [Google Scholar] [CrossRef]
- Green, M.R. Eukaryotic Transcription Activation: Right on Target. Mol Cell 2005, 18, 399–402. [Google Scholar] [CrossRef] [PubMed]
- Malik, S.; Roeder, R.G. The Metazoan Mediator Co-Activator Complex as an Integrative Hub for Transcriptional Regulation. Nat Rev Genet 2010, 11, 761–772. [Google Scholar] [CrossRef]
- Allen, B.L.; Taatjes, D.J. The Mediator Complex: A Central Integrator of Transcription. Nat Rev Mol Cell Biol 2015, 16, 155–166. [Google Scholar] [CrossRef]
- Merika, M.; Williams, A.J.; Chen, G.; Collins, T.; Thanos, D. Recruitment of CBP/P300 by the IFNβ Enhanceosome Is Required for Synergistic Activation of Transcription. Mol Cell 1998, 1, 277–287. [Google Scholar] [CrossRef]
- Reiter, F.; Wienerroither, S.; Stark, A. Combinatorial Function of Transcription Factors and Cofactors. Curr Opin Genet Dev 2017, 43, 73–81. [Google Scholar] [CrossRef]
- Bartholdi, D.; Roelfsema, J.H.; Papadia, F.; Breuning, M.H.; Niedrist, D.; Hennekam, R.C.; Schinzel, A.; Peters, D.J.M. Genetic Heterogeneity in Rubinstein–Taybi Syndrome: Delineation of the Phenotype of the First Patients Carrying Mutations in EP300. J Med Genet 2007, 44, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, N.; Acosta, A.M.B.F.; Kohlhase, J.; Bartsch, O. Confirmation of EP300 Gene Mutations as a Rare Cause of Rubinstein–Taybi Syndrome. European journal of human genetics 2007, 15, 837–842. [Google Scholar] [CrossRef]
- Giles, R.H.; Dauwerse, H.G.; van Ommen, G.-J.B.; Breuning, M.H. Do Human Chromosomal Bands 16p13 and 22q11-13 Share Ancestral Origins? The American Journal of Human Genetics 1998, 63, 1240–1242. [Google Scholar] [CrossRef] [PubMed]
- Dancy, B.M.; Cole, P.A. Protein Lysine Acetylation by P300/CBP. Chem Rev 2015, 115, 2419–2452. [Google Scholar] [CrossRef]
- Breen, M.E.; Mapp, A.K. Modulating the Masters: Chemical Tools to Dissect CBP and P300 Function. Curr Opin Chem Biol 2018, 45, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Delvecchio, M.; Gaucher, J.; Aguilar-Gurrieri, C.; Ortega, E.; Panne, D. Structure of the P300 Catalytic Core and Implications for Chromatin Targeting and HAT Regulation. Nat Struct Mol Biol 2013, 20, 1040–1046. [Google Scholar] [CrossRef]
- Dyson, H.J.; Wright, P.E. Role of Intrinsic Protein Disorder in the Function and Interactions of the Transcriptional Coactivators CREB-Binding Protein (CBP) and P300. Journal of Biological Chemistry 2016, 291, 6714–6722. [Google Scholar] [CrossRef]
- Ortega, E.; Rengachari, S.; Ibrahim, Z.; Hoghoughi, N.; Gaucher, J.; Holehouse, A.S.; Khochbin, S.; Panne, D. Transcription Factor Dimerization Activates the P300 Acetyltransferase. Nature 2018, 562, 538–544. [Google Scholar] [CrossRef]
- Zeng, L.; Zhang, Q.; Gerona-Navarro, G.; Moshkina, N.; Zhou, M.-M. Structural Basis of Site-Specific Histone Recognition by the Bromodomains of Human Coactivators PCAF and CBP/P300. Structure 2008, 16, 643–652. [Google Scholar] [CrossRef]
- Weinert, B.T.; Narita, T.; Satpathy, S.; Srinivasan, B.; Hansen, B.K.; Schölz, C.; Hamilton, W.B.; Zucconi, B.E.; Wang, W.W.; Liu, W.R.; et al. Time-Resolved Analysis Reveals Rapid Dynamics and Broad Scope of the CBP/P300 Acetylome. Cell 2018, 174, 231–244. [Google Scholar] [CrossRef]
- M Valor, L.; Viosca, J.; P Lopez-Atalaya, J.; Barco, A. Lysine Acetyltransferases CBP and P300 as Therapeutic Targets in Cognitive and Neurodegenerative Disorders. Curr Pharm Des 2013, 19, 5051–5064. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.-J.; Seto, E. Lysine Acetylation: Codified Crosstalk with Other Posttranslational Modifications. Mol Cell 2008, 31, 449–461. [Google Scholar] [CrossRef]
- Bedford, D.C.; Kasper, L.H.; Fukuyama, T.; Brindle, P.K. Target Gene Context Influences the Transcriptional Requirement for the KAT3 Family of CBP and P300 Histone Acetyltransferases. Epigenetics 2010, 5, 9–15. [Google Scholar] [CrossRef]
- Yamagata, K.; Goto, Y.; Nishimasu, H.; Morimoto, J.; Ishitani, R.; Dohmae, N.; Takeda, N.; Nagai, R.; Komuro, I.; Suga, H.; et al. Structural Basis for Potent Inhibition of SIRT2 Deacetylase by a Macrocyclic Peptide Inducing Dynamic Structural Change. Structure 2014, 22, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Penteado, A.B.; Hassanie, H.; Gomes, R.A.; Silva Emery, F. da; Goulart Trossini, G.H. Human Sirtuin 2 Inhibitors, Their Mechanisms and Binding Modes. Future Med Chem 2023, 15, 291–311. [Google Scholar] [CrossRef] [PubMed]
- Schemies, J.; Uciechowska, U.; Sippl, W.; Jung, M. NAD+-Dependent Histone Deacetylases (Sirtuins) as Novel Therapeutic Targets. Med Res Rev 2010, 30, 861–889. [Google Scholar] [CrossRef]
- Vassilopoulos, A.; Fritz, K.S.; Petersen, D.R.; Gius, D. The Human Sirtuin Family: Evolutionary Divergences and Functions. Hum Genomics 2011, 5, 1–12. [Google Scholar] [CrossRef]
- Haigis, M.C.; Sinclair, D.A. Mammalian Sirtuins: Biological Insights and Disease Relevance. Annual Review of Pathology: Mechanisms of Disease 2010, 5, 253–295. [Google Scholar] [CrossRef]
- Luo, Y.; Wang, M.; Tian, Y. Trends and Age-Period-Cohort Effects on Incidence and Mortality of Asthma in Sichuan Province, China, 1990–2019. BMC Pulm Med 2022, 22, 298. [Google Scholar] [CrossRef]
- Fukuda, Y.; Akimoto, K.; Homma, T.; Baker, J.R.; Ito, K.; Barnes, P.J.; Sagara, H. Virus-Induced Asthma Exacerbations: SIRT1 Targeted Approach. J Clin Med 2020, 9, 2623. [Google Scholar] [CrossRef]
- Colley, T.; Mercado, N.; Kunori, Y.; Brightling, C.; Bhavsar, P.K.; Barnes, P.J.; Ito, K. Defective Sirtuin-1 Increases IL-4 Expression through Acetylation of GATA-3 in Patients with Severe Asthma. Journal of Allergy and Clinical Immunology 2016, 137, 1595–1597. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Chen, Q.; Meng, Z.; Sun, L.; Zhu, L.; Liu, J.; Hu, J.; Ni, Z.; Wang, X. Suppression of Sirtuin-1 Increases IL-6 Expression by Activation of the Akt Pathway during Allergic Asthma. Cellular Physiology and Biochemistry 2018, 43, 1950–1960. [Google Scholar] [CrossRef]
- Zhang, Y.Z.; Wu, Q.J.; Yang, X.; Xing, X.X.; Chen, Y.Y.; Wang, H. Effects of SIRT1/Akt Pathway on Chronic Inflammatory Response and Lung Function in Patients with Asthma. Eur Rev Med Pharmacol Sci 2019, 23, 4948–4953. [Google Scholar]
- Lee, Y.G.; Reader, B.F.; Herman, D.; Streicher, A.; Englert, J.A.; Ziegler, M.; Chung, S.; Karpurapu, M.; Park, G.Y.; Christman, J.W.; et al. Sirtuin 2 Enhances Allergic Asthmatic Inflammation. JCI Insight 2019, 4, e124710. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-Y.; Hur, G.; Lee, S.W.; Lee, S.-J.; Lee, S.; Kim, S.-H.; Rho, M.-C. AGK2 Ameliorates Mast Cell-Mediated Allergic Airway Inflammation and Fibrosis by Inhibiting FcRI/TGF-β Signaling Pathway. Pharmacol Res 2020, 159, 105027. [Google Scholar] [CrossRef] [PubMed]
- Elias, J.A. Airway Remodeling in Asthma: Unanswered Questions. Am J Respir Crit Care Med 2000, 161, S168–S171. [Google Scholar] [CrossRef]
- Liu, F.; Shang, Y.-X. Sirtuin 6 Attenuates Epithelial–Mesenchymal Transition by Suppressing the TGF-Β1/Smad3 Pathway and c-Jun in Asthma Models. Int Immunopharmacol 2020, 82, 106333. [Google Scholar] [CrossRef] [PubMed]
- Sanders, B.D.; Jackson, B.; Marmorstein, R. Structural Basis for Sirtuin Function: What We Know and What We Don’t. Biochimica et Biophysica Acta (BBA)-Proteins and Proteomics 2010, 1804, 1604–1616. [Google Scholar] [CrossRef]
- Westphal, C.H.; Dipp, M.A.; Guarente, L. A Therapeutic Role for Sirtuins in Diseases of Aging? Trends Biochem Sci 2007, 32, 555–560. [Google Scholar] [CrossRef]
- Avalos, J.L.; Celic, I.; Muhammad, S.; Cosgrove, M.S.; Boeke, J.D.; Wolberger, C. Structure of a Sir2 Enzyme Bound to an Acetylated P53 Peptide. Mol Cell 2002, 10, 523–535. [Google Scholar] [CrossRef]
- Zhao, K.; Harshaw, R.; Chai, X.; Marmorstein, R. Structural Basis for Nicotinamide Cleavage and ADP-Ribose Transfer by NAD+-Dependent Sir2 Histone/Protein Deacetylases. Proceedings of the National Academy of Sciences 2004, 101, 8563–8568. [Google Scholar] [CrossRef] [PubMed]
- Avalos, J.L.; Boeke, J.D.; Wolberger, C. Structural Basis for the Mechanism and Regulation of Sir2 Enzymes. Mol Cell 2004, 13, 639–648. [Google Scholar] [CrossRef] [PubMed]
- Finnin, M.S.; Donigian, J.R.; Pavletich, N.P. Structure of the Histone Deacetylase SIRT2. Nat Struct Biol 2001, 8, 621–625. [Google Scholar] [CrossRef]
- Wang, Y.; He, J.; Liao, M.; Hu, M.; Li, W.; Ouyang, H.; Wang, X.; Ye, T.; Zhang, Y.; Ouyang, L. An Overview of Sirtuins as Potential Therapeutic Target: Structure, Function and Modulators. Eur J Med Chem 2019, 161, 48–77. [Google Scholar] [CrossRef] [PubMed]
- Bellamacina, C.R. The Nicotinamide Dinucleotide Binding Motif: A Comparison of Nucleotide Binding Proteins. The FASEB Journal 1996, 10, 1257–1269. [Google Scholar] [CrossRef]
- Verma, M.; Chattopadhyay, B.D.; Paul, B.N. Epigenetic Regulation of DNMT1 Gene in Mouse Model of Asthma Disease. Mol Biol Rep 2013, 40, 2357–2368. [Google Scholar] [CrossRef]
- Goll, M.G.; Bestor, T.H. Eukaryotic Cytosine Methyltransferases. Annu. Rev. Biochem. 2005, 74, 481–514. [Google Scholar] [CrossRef]
- Cheng, X.; Blumenthal, R.M. Mammalian DNA Methyltransferases: A Structural Perspective. Structure 2008, 16, 341–350. [Google Scholar] [CrossRef]
- Ren, W.; Gao, L.; Song, J. Structural Basis of DNMT1 and DNMT3A-Mediated DNA Methylation. Genes (Basel) 2018, 9, 620. [Google Scholar] [CrossRef]
- Jeltsch, A. On the Enzymatic Properties of Dnmt1: Specificity, Processivity, Mechanism of Linear Diffusion and Allosteric Regulation of the Enzyme. Epigenetics 2006, 1, 63–66. [Google Scholar] [CrossRef]
- Bird, A.P. Use of Restriction Enzymes to Study Eukaryotic DNA Methylation: II. The Symmetry of Methylated Sites Supports Semi-Conservative Copying of the Methylation Pattern. J Mol Biol 1978, 118, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Okano, M.; Bell, D.W.; Haber, D.A.; Li, E. DNA Methyltransferases Dnmt3a and Dnmt3b Are Essential for de Novo Methylation and Mammalian Development. Cell 1999, 99, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Okano, M.; Xie, S.; Li, E. Cloning and Characterization of a Family of Novel Mammalian DNA (Cytosine-5) Methyltransferases. Nat Genet 1998, 19, 219–220. [Google Scholar] [CrossRef]
- Lekesiz, R.T.; Koca, K.K.; Kugu, G.; Çalşkaner, Z.O. Versatile Functions of Methyl-CpG-Binding Domain 2 (MBD2) in Cellular Characteristics and Differentiation. Mol Biol Rep 2025, 52, 1–12. [Google Scholar] [CrossRef]
- Chen, Z.; Shang, Y.; Yuan, Y.; He, Y.; Wasti, B.; Duan, W.; Ouyang, R.; Jia, J.; Xiao, B.; Zhang, D.; et al. MBD2 Mediates Th17 Cell Differentiation by Regulating MINK1 in Th17-Dominant Asthma. Front Genet 2022, 13, 959059. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Xiao, B.; Jia, A.; Qiu, L.; Zeng, Q.; Liu, D.; Yuan, Y.; Jia, J.; Zhang, X.; Xiang, X. MBD2-Mediated Th17 Differentiation in Severe Asthma Is Associated with Impaired SOCS3 Expression. Exp Cell Res 2018, 371, 196–204. [Google Scholar] [CrossRef]
- Desai, M.A.; Webb, H.D.; Sinanan, L.M.; Scarsdale, J.N.; Walavalkar, N.M.; Ginder, G.D.; Williams Jr, D.C. An Intrinsically Disordered Region of Methyl-CpG Binding Domain Protein 2 (MBD2) Recruits the Histone Deacetylase Core of the NuRD Complex. Nucleic Acids Res 2015, 43, 3100–3113. [Google Scholar] [CrossRef]
- Du, Q.; Luu, P.-L.; Stirzaker, C.; Clark, S.J. Methyl-CpG-Binding Domain Proteins: Readers of the Epigenome. Epigenomics 2015, 7, 1051–1073. [Google Scholar] [CrossRef]
- Çalşkaner, Z.O. Computational Discovery of Novel Inhibitory Candidates Targeting Versatile Transcriptional Repressor MBD2. J Mol Model 2022, 28, 296. [Google Scholar] [CrossRef]
- Liu, K.; Xu, C.; Lei, M.; Yang, A.; Loppnau, P.; Hughes, T.R.; Min, J. Structural Basis for the Ability of MBD Domains to Bind Methyl-CG and TG Sites in DNA. Journal of Biological Chemistry 2018, 293, 7344–7354. [Google Scholar] [CrossRef]
- Du, Q.; Luu, P.-L.; Stirzaker, C.; Clark, S.J. Methyl-CpG-Binding Domain Proteins: Readers of the Epigenome. Epigenomics 2015, 7, 1051–1073. [Google Scholar] [CrossRef]
- Ohki, I.; Shimotake, N.; Fujita, N.; Jee, J.-G.; Ikegami, T.; Nakao, M.; Shirakawa, M. Solution Structure of the Methyl-CpG Binding Domain of Human MBD1 in Complex with Methylated DNA. Cell 2001, 105, 487–497. [Google Scholar] [CrossRef] [PubMed]
- Wood, K.H.; Zhou, Z. Emerging Molecular and Biological Functions of MBD2, a Reader of DNA Methylation. Front Genet 2016, 7, 93. [Google Scholar] [CrossRef]
- Younesian, S.; Yousefi, A.-M.; Momeny, M.; Ghaffari, S.H.; Bashash, D. The DNA Methylation in Neurological Diseases. Cells 2022, 11, 3439. [Google Scholar] [CrossRef] [PubMed]
- Cheng, R.Y.S.; Shang, Y.; Limjunyawong, N.; Dao, T.; Das, S.; Rabold, R.; Sham, J.S.K.; Mitzner, W.; Tang, W.-Y. Alterations of the Lung Methylome in Allergic Airway Hyper-Responsiveness. Environ Mol Mutagen 2014, 55, 244–255. [Google Scholar] [CrossRef]
- Iyer, L.M.; Tahiliani, M.; Rao, A.; Aravind, L. Prediction of Novel Families of Enzymes Involved in Oxidative and Other Complex Modifications of Bases in Nucleic Acids. Cell cycle 2009, 8, 1698–1710. [Google Scholar] [CrossRef] [PubMed]
- Pastor, W.A.; Aravind, L.; Rao, A. TETonic Shift: Biological Roles of TET Proteins in DNA Demethylation and Transcription. Nat Rev Mol Cell Biol 2013, 14, 341–356. [Google Scholar] [CrossRef]
- Williams, K.; Christensen, J.; Helin, K. DNA Methylation: TET Proteins—Guardians of CpG Islands? EMBO Rep 2012, 13, 28–35. [Google Scholar] [CrossRef]
- Hu, L.; Li, Z.; Cheng, J.; Rao, Q.; Gong, W.; Liu, M.; Shi, Y.G.; Zhu, J.; Wang, P.; Xu, Y. Crystal Structure of TET2-DNA Complex: Insight into TET-Mediated 5mC Oxidation. Cell 2013, 155, 1545–1555. [Google Scholar] [CrossRef]
- Liu, K.; Gao, Y.; Gan, K.; Wu, Y.; Xu, B.; Zhang, L.; Chen, M. Prognostic Roles of N6-Methyladenosine METTL3 in Different Cancers: A System Review and Meta-Analysis. Cancer Control 2021, 28, 1073274821997455. [Google Scholar] [CrossRef]
- Sweaad, W.K.; Stefanizzi, F.M.; Chamorro-Jorganes, A.; Devaux, Y.; Emanueli, C. ; others Relevance of N6-Methyladenosine Regulators for Transcriptome: Implications for Development and the Cardiovascular System. J Mol Cell Cardiol 2021, 160, 56–70. [Google Scholar] [CrossRef] [PubMed]
- Tuncel, G.; Kalkan, R. Importance of m N6-Methyladenosine (M6A) RNA Modification in Cancer. Medical Oncology 2019, 36, 36. [Google Scholar] [CrossRef]
- Xu, L.; Zhou, L.; Yan, C.; Li, L. Emerging Role of N6-Methyladenosine RNA Methylation in Lung Diseases. Exp Biol Med 2022, 247, 1862–1872. [Google Scholar] [CrossRef] [PubMed]
- Chao, Y.; Shang, J.; Ji, W. ALKBH5-M6A-FOXM1 Signaling Axis Promotes Proliferation and Invasion of Lung Adenocarcinoma Cells under Intermittent Hypoxia. Biochem Biophys Res Commun 2020, 521, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Lee, Y.; Kim, S.-M.; Jang, S.; Choi, H.; Lee, J.-W.; Kim, T.-D.; Kim, V.N. RNA Demethylation by FTO Stabilizes the FOXJ1 MRNA for Proper Motile Ciliogenesis. Dev Cell 2021, 56, 1118–1130. [Google Scholar] [CrossRef]
- Li, S.-X.; Yan, W.; Liu, J.-P.; Zhao, Y.-J.; Chen, L. Long Noncoding RNA SNHG4 Remits Lipopolysaccharide-Engendered Inflammatory Lung Damage by Inhibiting METTL3–Mediated M6A Level of STAT2 MRNA. Mol Immunol 2021, 139, 10–22. [Google Scholar] [CrossRef]
- Fei, L.; Sun, G.; Sun, J.; Wu, D. The Effect of N6-Methyladenosine (M6A) Factors on the Development of Acute Respiratory Distress Syndrome in the Mouse Model. Bioengineered 2022, 13, 7622–7634. [Google Scholar] [CrossRef]
- Wang, Y.; Su, X.; Zhao, M.; Xu, M.; Chen, Y.; Li, Z.; Zhuang, W. Importance of N6-Methyladenosine RNA Modification in Lung Cancer. Mol Clin Oncol 2021, 14, 128. [Google Scholar] [CrossRef]
- Wu, S.; Li, X.-F.; Wu, Y.-Y.; Yin, S.-Q.; Huang, C.; Li, J. N6-Methyladenosine and Rheumatoid Arthritis: A Comprehensive Review. Front Immunol 2021, 12, 731842. [Google Scholar] [CrossRef]
- Teng, F.; Tang, W.; Wuniqiemu, T.; Qin, J.; Zhou, Y.; Huang, X.; Wang, S.; Zhu, X.; Tang, Z.; Yi, L.; et al. N 6-Methyladenosine Methylomic Landscape of Lung Tissues in Murine Acute Allergic Asthma. Front Immunol 2021, 12, 740571. [Google Scholar] [CrossRef]
- Chen, M.; Wei, L.; Law, C.-T.; Tsang, F.H.-C.; Shen, J.; Cheng, C.L.-H.; Tsang, L.-H.; Ho, D.W.-H.; Chiu, D.K.-C.; Lee, J.M.-F.; et al. RNA N6-Methyladenosine Methyltransferase-like 3 Promotes Liver Cancer Progression through YTHDF2-Dependent Posttranscriptional Silencing of SOCS2. Hepatology 2018, 67, 2254–2270. [Google Scholar] [CrossRef]
- Shi, H.; Wang, X.; Lu, Z.; Zhao, B.S.; Ma, H.; Hsu, P.J.; Liu, C.; He, C. YTHDF3 Facilitates Translation and Decay of N6-Methyladenosine-Modified RNA. Cell Res 2017, 27, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Wang, J.; Huang, H.; Yu, Y.; Ding, J.; Yu, Y.; Li, K.; Wei, D.; Ye, Q.; Wang, F.; et al. YTHDF1 Regulates Pulmonary Hypertension through Translational Control of MAGED1. Am J Respir Crit Care Med 2021, 203, 1158–1172. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Qi, Y.; Yu, J.; Hao, Y.; He, B.; Zhang, M.; Dai, Z.; Jiang, T.; Li, S.; Huang, F.; et al. Nuclear Aurora Kinase A Switches M6A Reader YTHDC1 to Enhance an Oncogenic RNA Splicing of Tumor Suppressor RBM4. Signal Transduct Target Ther 2022, 7, 97. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Wei, Q.; Jin, J.; Luo, Q.; Liu, Y.; Yang, Y.; Cheng, C.; Li, L.; Pi, J.; Si, Y.; et al. The M6A Reader YTHDF1 Promotes Ovarian Cancer Progression via Augmenting EIF3C Translation. Nucleic Acids Res 2020, 48, 3816–3831. [Google Scholar] [CrossRef]
- Yan, S.; Zhou, X.; Wu, C.; Gao, Y.; Qian, Y.; Hou, J.; Xie, R.; Han, B.; Chen, Z.; Wei, S.; et al. Adipocyte YTH N (6)-Methyladenosine RNA-Binding Protein 1 Protects against Obesity by Promoting White Adipose Tissue Beiging in Male Mice. Nat Commun 2023, 14, 1379. [Google Scholar] [CrossRef]
- Li, Z.; Teng, M.; Jiang, Y.; Zhang, L.; Luo, X.; Liao, Y.; Yang, B. YTHDF1 Negatively Regulates Treponema Pallidum-Induced Inflammation in THP-1 Macrophages by Promoting SOCS3 Translation in an M6A-Dependent Manner. Front Immunol 2022, 13, 857727. [Google Scholar] [CrossRef]
- Chen, L.; Hou, W.; Liu, F.; Zhu, R.; Lv, A.; Quan, W.; Mao, S. Blockade of NLRP3/Caspase-1/IL-1β Regulated Th17/Treg Immune Imbalance and Attenuated the Neutrophilic Airway Inflammation in an Ovalbumin-Induced Murine Model of Asthma. J Immunol Res 2022, 2022, 9444227. [Google Scholar] [CrossRef]
- Xing, Y.; Cheng, D.; Shi, C.; Shen, Z. The Protective Role of YTHDF1-Knock down Macrophages on the Immune Paralysis of Severe Sepsis Rats with ECMO. Microvasc Res 2021, 137, 104178. [Google Scholar] [CrossRef]
- Qu, T.; Huang, B.; Lei, L.; Luo, S. Cetirizine Inhibits Activation of JAK2-STAT3 Pathway and Mast Cell Activation in Lung Tissue of Asthmatic Mice. Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi 2022, 38, 193–198. [Google Scholar]
- Wang, R.; Wang, Y.; Liao, G.; Chen, B.; Panettieri, R.A.; Penn, R.B.; Tang, D.D. Abi1 Mediates Airway Smooth Muscle Cell Proliferation and Airway Remodeling via Jak2/STAT3 Signaling. iScience 2022, 25. [Google Scholar] [CrossRef] [PubMed]
- Bai, H.; Xue, Z.; Zhang, W.; Feng, C.; Zhou, Z.; Hu, S.; Zhang, Y.; Qin, Q.; Wu, Y.; Sun, X.; et al. α-Asarone Alleviates Allergic Asthma by Stabilizing Mast Cells through Inhibition of ERK/JAK2-STAT3 Pathway. Biofactors 2023, 49, 140–152. [Google Scholar] [CrossRef]
- Milara, J.; Ballester, B.; de Diego, A.; Calbet, M.; Ramis, I.; Miralpeix, M.; Cortijo, J. The Pan-JAK Inhibitor LAS194046 Reduces Neutrophil Activation from Severe Asthma and COPD Patients in Vitro. Sci Rep 2022, 12, 5132. [Google Scholar] [CrossRef]
- Wang, J.; Wang, L.; Tian, X.; Luo, L. N6-Methyladenosine Reader YTHDF1 Regulates the Proliferation and Migration of Airway Smooth Muscle Cells through M6A/Cyclin D1 in Asthma. PeerJ 2023, 11, e14951. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Yang, H.; Fan, L.; Shen, F.; Wang, Z. M6A Regulator-Mediated RNA Methylation Modification Patterns and Immune Microenvironment Infiltration Characterization in Severe Asthma. J Cell Mol Med 2021, 25, 10236–10247. [Google Scholar] [CrossRef] [PubMed]
- Dominissini, D.; Moshitch-Moshkovitz, S.; Schwartz, S.; Salmon-Divon, M.; Ungar, L.; Osenberg, S.; Cesarkas, K.; Jacob-Hirsch, J.; Amariglio, N.; Kupiec, M.; et al. Topology of the Human and Mouse M6A RNA Methylomes Revealed by M6A-Seq. Nature 2012, 485, 201–206. [Google Scholar] [CrossRef]
- Wang, X.; Lu, Z.; Gomez, A.; Hon, G.C.; Yue, Y.; Han, D.; Fu, Y.; Parisien, M.; Dai, Q.; Jia, G.; et al. N 6-Methyladenosine-Dependent Regulation of Messenger RNA Stability. Nature 2014, 505, 117–120. [Google Scholar] [CrossRef]
- Li, Y.; Bedi, R.K.; Wiedmer, L.; Huang, D.; Śledź, P.; Caflisch, A. Flexible Binding of M6A Reader Protein YTHDC1 to Its Preferred RNA Motif. J Chem Theory Comput 2019, 15, 7004–7014. [Google Scholar] [CrossRef]
- Bedi, R.K.; Huang, D.; Wiedmer, L.; Li, Y.; Dolbois, A.; Wojdyla, J.A.; Sharpe, M.E.; Caflisch, A.; Sledz, P. Selectively Disrupting M6A-Dependent Protein–RNA Interactions with Fragments. ACS Chem Biol 2020, 15, 618–625. [Google Scholar] [CrossRef]
- Xu, C.; Liu, K.; Ahmed, H.; Loppnau, P.; Schapira, M.; Min, J. Structural Basis for the Discriminative Recognition of N6-Methyladenosine RNA by the Human YT521-B Homology Domain Family of Proteins. Journal of Biological Chemistry 2015, 290, 24902–24913. [Google Scholar] [CrossRef]
- Li, Y.; Bedi, R.K.; Moroz-Omori, E. V; Caflisch, A. Structural and Dynamic Insights into Redundant Function of YTHDF Proteins. J Chem Inf Model 2020, 60, 5932–5935. [Google Scholar] [CrossRef] [PubMed]
- Patil, D.P.; Pickering, B.F.; Jaffrey, S.R. Reading M6A in the Transcriptome: M6A-Binding Proteins. Trends Cell Biol 2018, 28, 113–127. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J Mol Biol 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Li, F.; Zhao, D.; Wu, J.; Shi, Y. Structure of the YTH Domain of Human YTHDF2 in Complex with an M6A Mononucleotide Reveals an Aromatic Cage for M6A Recognition. Cell Res 2014, 24, 1490–1492. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Ž\’\idek, A.; Potapenko, A.; et al. Highly Accurate Protein Structure Prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Nai, F.; Nachawati, R.; Zalesak, F.; Wang, X.; Li, Y.; Caflisch, A. Fragment Ligands of the M6A-RNA Reader YTHDF2. ACS Med Chem Lett 2022, 13, 1500–1509. [Google Scholar] [CrossRef] [PubMed]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The Proteomics Server for in-Depth Protein Knowledge and Analysis. Nucleic Acids Res 2003, 31, 3784–3788. [Google Scholar] [CrossRef]
- Zhu, T.; Roundtree, I.A.; Wang, P.; Wang, X.; Wang, L.; Sun, C.; Tian, Y.; Li, J.; He, C.; Xu, Y. Crystal Structure of the YTH Domain of YTHDF2 Reveals Mechanism for Recognition of N6-Methyladenosine. Cell Res 2014, 24, 1493–1496. [Google Scholar] [CrossRef]
- Deng, J.; Zhang, J.; Ye, Y.; Liu, K.; Zeng, L.; Huang, J.; Pan, L.; Li, M.; Bai, R.; Zhuang, L.; et al. N6-Methyladenosine–Mediated Upregulation of WTAPP1 Promotes WTAP Translation and Wnt Signaling to Facilitate Pancreatic Cancer Progression. Cancer Res 2021, 81, 5268–5283. [Google Scholar] [CrossRef]
- Li, Y.; Sheng, H.; Ma, F.; Wu, Q.; Huang, J.; Chen, Q.; Sheng, L.; Zhu, X.; Zhu, X.; Xu, M. RNA M6A Reader YTHDF2 Facilitates Lung Adenocarcinoma Cell Proliferation and Metastasis by Targeting the AXIN1/Wnt/β-Catenin Signaling. Cell Death Dis 2021, 12, 479. [Google Scholar] [CrossRef]
- Hussain, M.; Xu, C.; Lu, M.; Wu, X.; Tang, L.; Wu, X. Wnt/β-Catenin Signaling Links Embryonic Lung Development and Asthmatic Airway Remodeling. Biochimica et Biophysica Acta (BBA)-Molecular Basis of Disease 2017, 1863, 3226–3242. [Google Scholar] [CrossRef]
- Lin, Q.; Ni, H.; Zheng, Z.; Zhong, J.; Nie, H. Cross-Talk of Four Types of RNA Modification Writers Defines the Immune Microenvironment in Severe Asthma. Ann N Y Acad Sci 2022, 1514, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.; Chen, R.; Ling, T.; Liu, B.; Huang, J.; Cheng, Y.; Lin, Y.; Chen, H.; Xie, X.; Xia, G.; et al. Elevated WTAP Promotes Hyperinflammation by Increasing m 6 A Modification in Inflammatory Disease Models. J Clin Invest 2024, 134. [Google Scholar] [CrossRef]
- Little, N.A.; Hastie, N.D.; Davies, R.C. Identification of WTAP, a Novel Wilms’ Tumour 1-Associating Protein. Hum Mol Genet 2000, 9, 2231–2239. [Google Scholar] [CrossRef]
- Small, T.W.; Penalva, L.O.; Pickering, J.G. Vascular Biology and the Sex of Flies: Regulation of Vascular Smooth Muscle Cell Proliferation by Wilms’ Tumor 1–Associating Protein. Trends Cardiovasc Med 2007, 17, 230–234. [Google Scholar] [CrossRef]
- Schöller, E.V.A.; Weichmann, F.; Treiber, T.; Ringle, S.; Treiber, N.; Flatley, A.; Feederle, R.; Bruckmann, A.; Meister, G. Interactions, Localization, and Phosphorylation of the M6A Generating METTL3–METTL14–WTAP Complex. Rna 2018, 24, 499–512. [Google Scholar] [CrossRef]
- Horiuchi, K.; Kawamura, T.; Iwanari, H.; Ohashi, R.; Naito, M.; Kodama, T.; Hamakubo, T. Identification of Wilms’ Tumor 1-Associating Protein Complex and Its Role in Alternative Splicing and the Cell Cycle. Journal of Biological Chemistry 2013, 288, 33292–33302. [Google Scholar] [CrossRef] [PubMed]
- Ping, X.-L.; Sun, B.-F.; Wang, L.U.; Xiao, W.; Yang, X.; Wang, W.-J.; Adhikari, S.; Shi, Y.; Lv, Y.; Chen, Y.-S.; et al. Mammalian WTAP Is a Regulatory Subunit of the RNA N6-Methyladenosine Methyltransferase. Cell Res 2014, 24, 177–189. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Pei, K.; Guan, Z.; Liu, F.; Yan, J.; Jin, X.; Wang, Q.; Hou, M.; Tang, C.; Yin, P. AI-Empowered Integrative Structural Characterization of M6A Methyltransferase Complex. Cell Res 2022, 32, 1124–1127. [Google Scholar] [CrossRef]
- Liu, J.; Yue, Y.; Han, D.; Wang, X.; Fu, Y.; Zhang, L.; Jia, G.; Yu, M.; Lu, Z.; Deng, X.; et al. A METTL3–METTL14 Complex Mediates Mammalian Nuclear RNA N 6-Adenosine Methylation. Nat Chem Biol 2014, 10, 93–95. [Google Scholar] [CrossRef]
- Lin, L.; Hu, X.; Li, Q.; Huang, L. Methyltransferase-like 3 (METTL3) Epigenetically Modulates Glutathione Peroxidase 4 (GPX4) Expression to Affect Asthma. Iran J Allergy Asthma Immunol 2023, 22, 551–560. [Google Scholar] [CrossRef]
- Chen, J.; Li, J.; Yuan, Y.; Tian, Q.; Feng, D.; Zhuang, L.; Cao, Q.; Zhou, G.; Jin, R. ETS1 and RBPJ Transcriptionally Regulate METTL14 to Suppress TGF-Β1-Induced Epithelial-Mesenchymal Transition in Human Bronchial Epithelial Cells. Biochimica et Biophysica Acta (BBA)-Molecular Basis of Disease 2024, 1870, 167349. [Google Scholar] [CrossRef] [PubMed]
- Ni, X.; Li, X.; Hu, B.; Wang, L. Chronic Allergic Asthma Alters M6A Epitranscriptomic Tagging of MRNAs and LncRNAs in the Lung. Biosci Rep 2022, 42, BSR20221395. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, J.; Yan, Z.; Liu, S.; Xu, W. Microenvironment Modulation by Key Regulators of RNA N6-Methyladenosine Modification in Respiratory Allergic Diseases. BMC Pulm Med 2023, 23, 210. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, Y.; Toth, J.I.; Petroski, M.D.; Zhang, Z.; Zhao, J.C. N 6-Methyladenosine Modification Destabilizes Developmental Regulators in Embryonic Stem Cells. Nat Cell Biol 2014, 16, 191–198. [Google Scholar] [CrossRef]
- Liu, J.; Yue, Y.; Han, D.; Wang, X.; Fu, Y.; Zhang, L.; Jia, G.; Yu, M.; Lu, Z.; Deng, X.; et al. A METTL3–METTL14 Complex Mediates Mammalian Nuclear RNA N 6-Adenosine Methylation. Nat Chem Biol 2014, 10, 93–95. [Google Scholar] [CrossRef]
- Iyer, L.M.; Zhang, D.; Aravind, L. Adenine Methylation in Eukaryotes: Apprehending the Complex Evolutionary History and Functional Potential of an Epigenetic Modification. Bioessays 2016, 38, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, S.; Mumbach, M.R.; Jovanovic, M.; Wang, T.; Maciag, K.; Bushkin, G.G.; Mertins, P.; Ter-Ovanesyan, D.; Habib, N.; Cacchiarelli, D.; et al. Perturbation of M6A Writers Reveals Two Distinct Classes of MRNA Methylation at Internal and 5′ Sites. Cell Rep 2014, 8, 284–296. [Google Scholar] [CrossRef]
- Śledź, P.; Jinek, M. Structural Insights into the Molecular Mechanism of the M6A Writer Complex. Elife 2016, 5, e18434. [Google Scholar] [CrossRef]
- Huang, J.; Dong, X.; Gong, Z.; Qin, L.-Y.; Yang, S.; Zhu, Y.-L.; Wang, X.; Zhang, D.; Zou, T.; Yin, P.; et al. Solution Structure of the RNA Recognition Domain of METTL3-METTL14 N6-Methyladenosine Methyltransferase. Protein Cell 2019, 10, 272–284. [Google Scholar] [CrossRef]
- Wang, P.; Doxtader, K.A.; Nam, Y. Structural Basis for Cooperative Function of Mettl3 and Mettl14 Methyltransferases. Mol Cell 2016, 63, 306–317. [Google Scholar] [CrossRef]
- Yoshida, A.; Oyoshi, T.; Suda, A.; Futaki, S.; Imanishi, M. Recognition of G-Quadruplex RNA by a Crucial RNA Methyltransferase Component, METTL14. Nucleic Acids Res 2022, 50, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Corbeski, I.; Vargas-Rosales, P.A.; Bedi, R.K.; Deng, J.; Coelho, D.; Braud, E.; Iannazzo, L.; Li, Y.; Huang, D.; Ethève-Quelquejeu, M.; et al. The Catalytic Mechanism of the RNA Methyltransferase METTL3. Elife 2024, 12, RP92537. [Google Scholar] [CrossRef] [PubMed]
- Oerum, S.; Meynier, V.; Catala, M.; Tisné, C. A Comprehensive Review of M6A/M6Am RNA Methyltransferase Structures. Nucleic Acids Res 2021, 49, 7239–7255. [Google Scholar] [CrossRef]
- Breger, K.; Kunkler, C.N.; O’Leary, N.J.; Hulewicz, J.P.; Brown, J.A. Ghost Authors Revealed: The Structure and Function of Human N 6-Methyladenosine RNA Methyltransferases. Wiley Interdiscip Rev RNA 2024, 15, e1810. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Weng, H.; Sun, W.; Qin, X.; Shi, H.; Wu, H.; Zhao, B.S.; Mesquita, A.; Liu, C.; Yuan, C.L.; et al. Recognition of RNA N 6-Methyladenosine by IGF2BP Proteins Enhances MRNA Stability and Translation. Nat Cell Biol 2018, 20, 285–295. [Google Scholar] [CrossRef]
- Dai, N.; Rapley, J.; Angel, M.; Yanik, M.F.; Blower, M.D.; Avruch, J. MTOR Phosphorylates IMP2 to Promote IGF2 MRNA Translation by Internal Ribosomal Entry. Genes Dev 2011, 25, 1159–1172. [Google Scholar] [CrossRef] [PubMed]
- Fakhar, M.; Gul, M.; Li, W. Interactive Structural Analysis of KH3-4 Didomains of IGF2BPs with Preferred RNA Motif Having m6A through Dynamics Simulation Studies 2024.
- Lian, Z.; Chen, R.; Xian, M.; Huang, P.; Xu, J.; Xiao, X.; Ning, X.; Zhao, J.; Xie, J.; Duan, J.; et al. Targeted Inhibition of M6A Demethylase FTO by FB23 Attenuates Allergic Inflammation in the Airway Epithelium. The FASEB Journal 2024, 38, e23846. [Google Scholar] [CrossRef] [PubMed]
- Jia, G.; Yang, C.-G.; Yang, S.; Jian, X.; Yi, C.; Zhou, Z.; He, C. Oxidative Demethylation of 3-Methylthymine and 3-Methyluracil in Single-Stranded DNA and RNA by Mouse and Human FTO. FEBS Lett 2008, 582, 3313–3319. [Google Scholar] [CrossRef]
- Toh, J.D.W.; Sun, L.; Lau, L.Z.M.; Tan, J.; Low, J.J.A.; Tang, C.W.Q.; Cheong, E.J.Y.; Tan, M.J.H.; Chen, Y.; Hong, W.; et al. A Strategy Based on Nucleotide Specificity Leads to a Subfamily-Selective and Cell-Active Inhibitor of N 6-Methyladenosine Demethylase FTO. Chem Sci 2015, 6, 112–122. [Google Scholar] [CrossRef]
- Peng, S.; Xiao, W.; Ju, D.; Sun, B.; Hou, N.; Liu, Q.; Wang, Y.; Zhao, H.; Gao, C.; Zhang, S.; et al. Identification of Entacapone as a Chemical Inhibitor of FTO Mediating Metabolic Regulation through FOXO1. Sci Transl Med 2019, 11, eaau7116. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Niu, T.; Chang, J.; Lei, X.; Zhao, M.; Wang, Q.; Cheng, W.; Wang, J.; Feng, Y.; Chai, J. Crystal Structure of the FTO Protein Reveals Basis for Its Substrate Specificity. Nature 2010, 464, 1205–1209. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Duan, Z.; Zhang, D.; Xiao, P.; Zhang, T.; Xu, H.; Wang, C.-H.; Rao, G.-W.; Gan, J.; Huang, Y.; et al. Structure–Activity Relationships and Antileukemia Effects of the Tricyclic Benzoic Acid FTO Inhibitors. J Med Chem 2022, 65, 10638–10654. [Google Scholar] [CrossRef] [PubMed]
- Shishodia, S.; Demetriades, M.; Zhang, D.; Tam, N.Y.; Maheswaran, P.; Clunie-O’Connor, C.; Tumber, A.; Leung, I.K.H.; Ng, Y.M.; Leissing, T.M.; et al. Structure-Based Design of Selective Fat Mass and Obesity Associated Protein (FTO) Inhibitors. J Med Chem 2021, 64, 16609–16625. [Google Scholar] [CrossRef]
- Zhang, X.; Wei, L.-H.; Wang, Y.; Xiao, Y.; Liu, J.; Zhang, W.; Yan, N.; Amu, G.; Tang, X.; Zhang, L.; et al. Structural Insights into FTO’s Catalytic Mechanism for the Demethylation of Multiple RNA Substrates. Proceedings of the National Academy of Sciences 2019, 116, 2919–2924. [Google Scholar] [CrossRef]
Figure 2.
Structure of p300 and CBP. The p300 and CBP proteins consist of 2414 and 2442 amino acids respectively and both have a molecular weight of around 265 kilodaltons. The 58% sequence similarity has been observed within their respective domains of both proteins. Both proteins comprise of various domains like on N terminal there is nuclear receptor interaction domain “(NRID)” followed by three regions rich in cysteine and histidine “(C/H1 to C/H3)”, which feature zinc finger transcriptional adapters “TAZ1” and “TAZ2”, and “C/H2”, which contains a “homeodomain (PHD)”, a “KIX domain”, a “bromodomain, a lysine acetyltransferase domain (KAT)”, and an “interferon-binding transactivation domain (IBiD)”. The locations of the various domains are specified in relation to their positions within the amino acid sequence. The catalytic domain has exhibited significant conservation throughout evolution, with 86% similarity between p300 and CBP encompassing the KAT domain and adjacent areas.
Figure 2.
Structure of p300 and CBP. The p300 and CBP proteins consist of 2414 and 2442 amino acids respectively and both have a molecular weight of around 265 kilodaltons. The 58% sequence similarity has been observed within their respective domains of both proteins. Both proteins comprise of various domains like on N terminal there is nuclear receptor interaction domain “(NRID)” followed by three regions rich in cysteine and histidine “(C/H1 to C/H3)”, which feature zinc finger transcriptional adapters “TAZ1” and “TAZ2”, and “C/H2”, which contains a “homeodomain (PHD)”, a “KIX domain”, a “bromodomain, a lysine acetyltransferase domain (KAT)”, and an “interferon-binding transactivation domain (IBiD)”. The locations of the various domains are specified in relation to their positions within the amino acid sequence. The catalytic domain has exhibited significant conservation throughout evolution, with 86% similarity between p300 and CBP encompassing the KAT domain and adjacent areas.
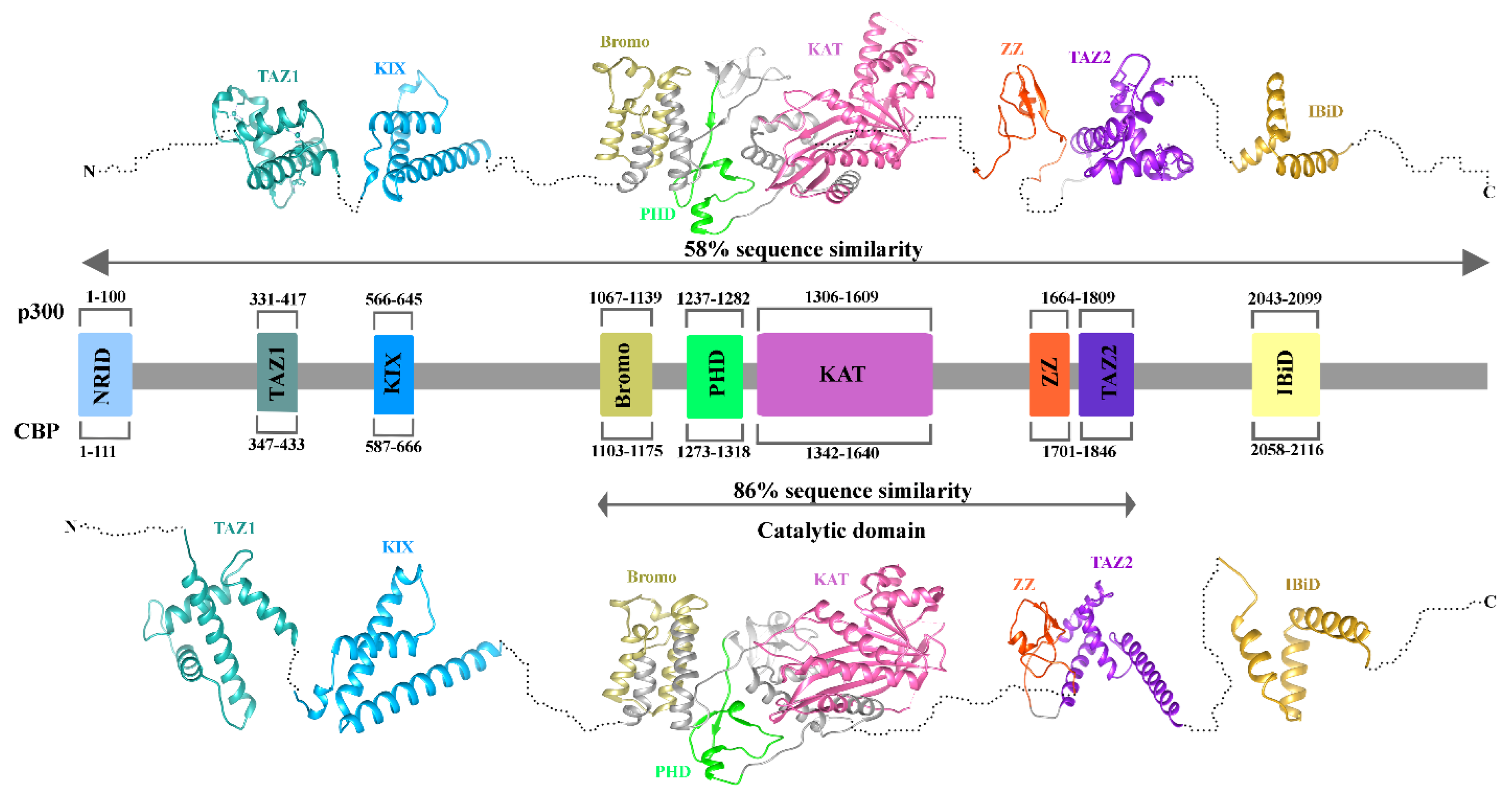
Figure 3.
Conservation Mapping of Sirtuins: (A) The figure depicts the conservation analysis of seven sirtuin proteins (SIRT1–SIRT7), showing their conservation patterns. (B) Ribbon and (C) surface views of SIRT2 3D structure. The top panel displays the conservation profile of the sirtuins, highlighting conserved regions across the family. The bottom panel presents the ribbon and surface views of SIRT2.
Figure 3.
Conservation Mapping of Sirtuins: (A) The figure depicts the conservation analysis of seven sirtuin proteins (SIRT1–SIRT7), showing their conservation patterns. (B) Ribbon and (C) surface views of SIRT2 3D structure. The top panel displays the conservation profile of the sirtuins, highlighting conserved regions across the family. The bottom panel presents the ribbon and surface views of SIRT2.
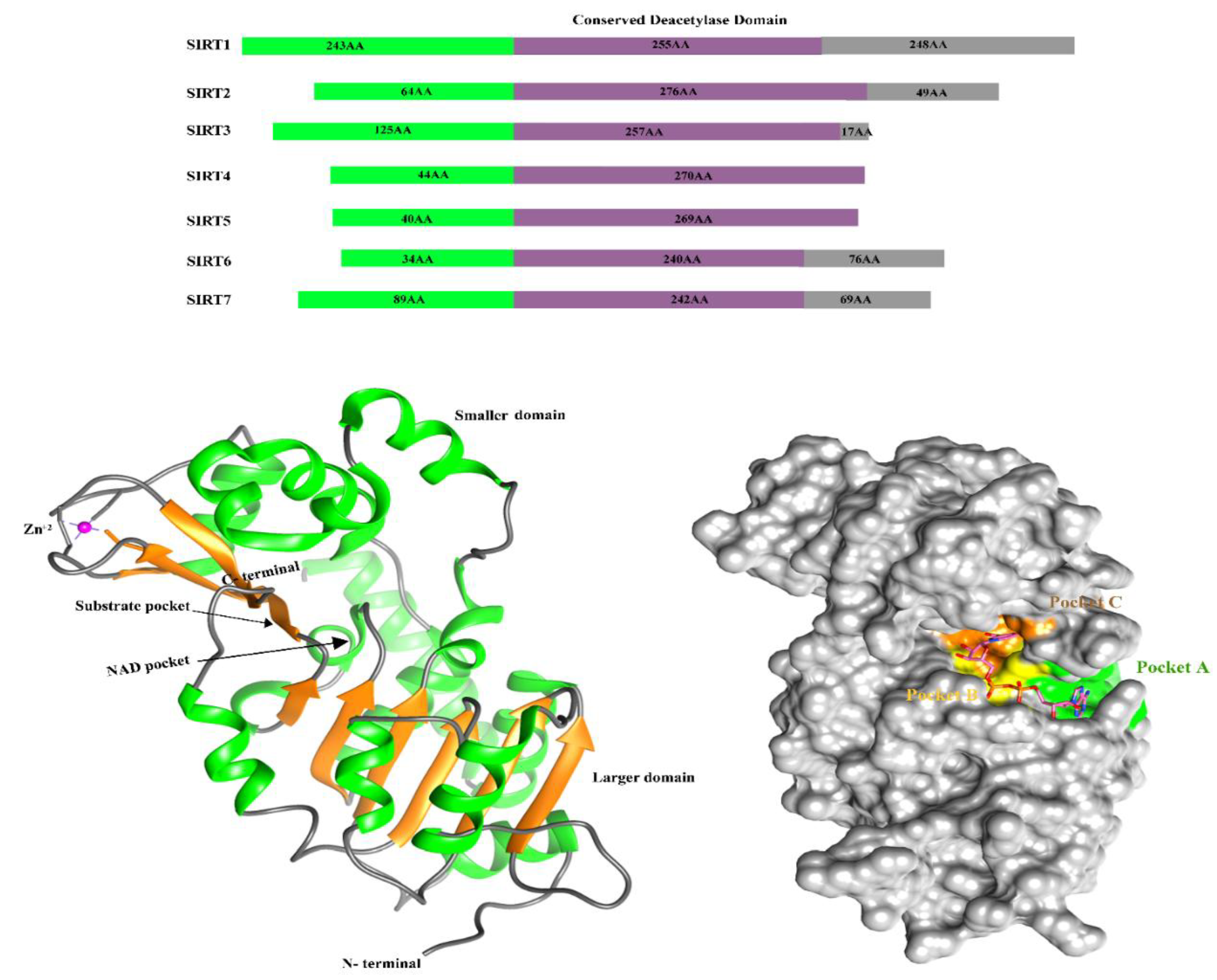
Figure 4.
Structural comparison of YTHDF1 and YTHDF2. The figure depicts the domain organization and 3D structure of YTHDF1 (left) and YTHDF2 (right). The N-terminal is red, the YTH domain is green, and the C-terminal is orange. Protein lengths are indicated in amino acids (AA) at the top.
Figure 4.
Structural comparison of YTHDF1 and YTHDF2. The figure depicts the domain organization and 3D structure of YTHDF1 (left) and YTHDF2 (right). The N-terminal is red, the YTH domain is green, and the C-terminal is orange. Protein lengths are indicated in amino acids (AA) at the top.
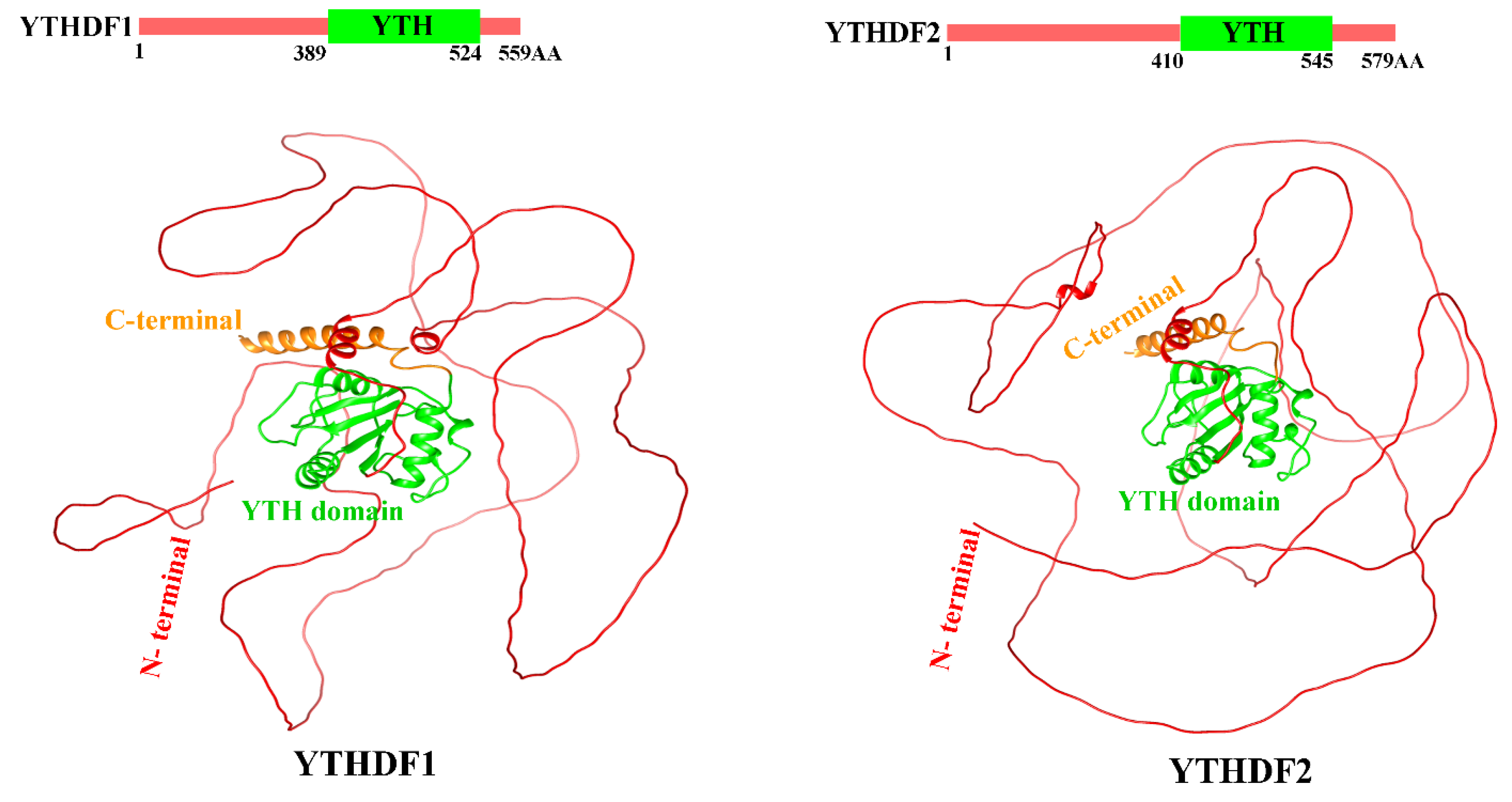
Figure 5.
Structural representation of WTAP: The figure shows the 3D structure of WTAP, with the N-terminal and C-terminal regions colored red, and the coiled-coil region in green. Key structural features are labeled accordingly. The protein's distinct structural domains are visualized for clarity.
Figure 5.
Structural representation of WTAP: The figure shows the 3D structure of WTAP, with the N-terminal and C-terminal regions colored red, and the coiled-coil region in green. Key structural features are labeled accordingly. The protein's distinct structural domains are visualized for clarity.
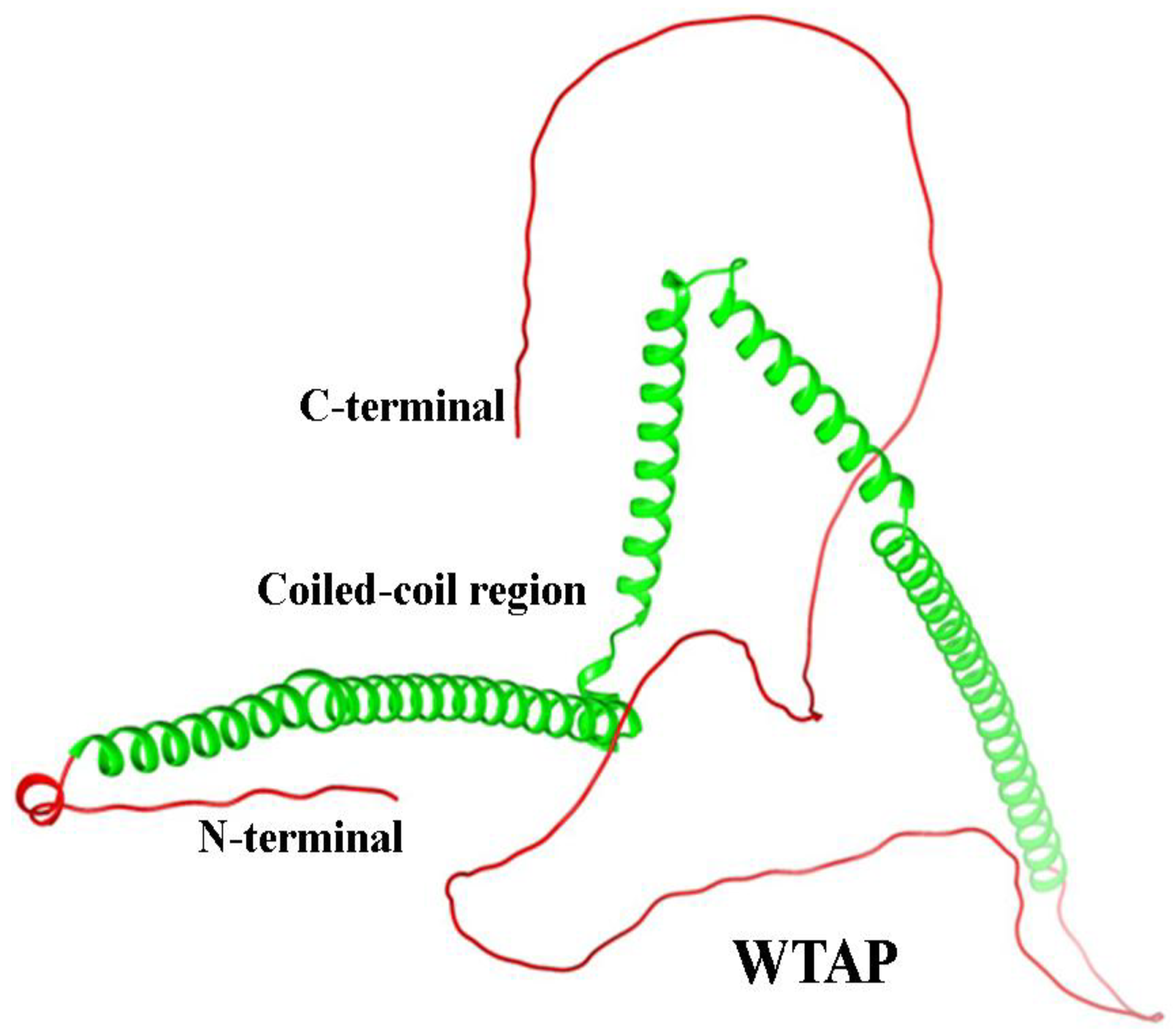
Figure 7.
Structural Domains of the Human FTO Protein. Ribbon representation of the full-length human FTO protein structure, highlighting its two major domains. The N-terminal domain (residues 32–326, shown in grey), while the C-terminal domain (residues 327–498, shown in green). The N- and C-termini are labeled accordingly.
Figure 7.
Structural Domains of the Human FTO Protein. Ribbon representation of the full-length human FTO protein structure, highlighting its two major domains. The N-terminal domain (residues 32–326, shown in grey), while the C-terminal domain (residues 327–498, shown in green). The N- and C-termini are labeled accordingly.
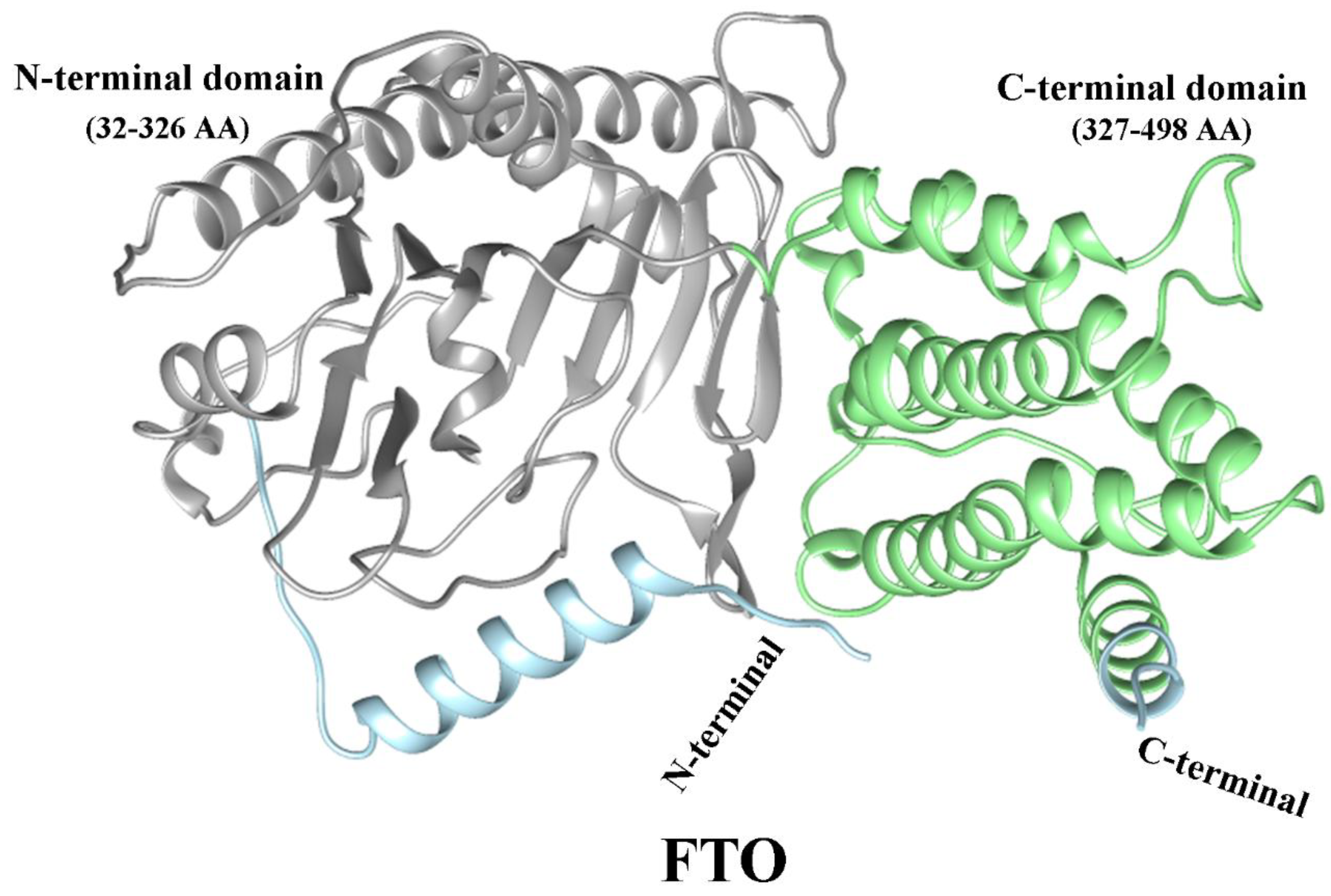

Table 1.
Epigenetic Regulators in Asthma: Writers, Readers and Erasers of DNA, Histone and RNA Modifications.
Table 1.
Epigenetic Regulators in Asthma: Writers, Readers and Erasers of DNA, Histone and RNA Modifications.
| Modification Type | Role | Examples | Function | References |
|---|---|---|---|---|
| Histone Modification | Writer | p300/CBP | p300 and CBP are histone acetyltransferases that, with increased expression in asthma, likely activate pro-inflammatory genes, contributing to chronic airway inflammation. | [29] |
| Gcn5 | H3K27 acetylation via HAT Gcn5 plays a key role in the histone code, with strong associations to asthma through its regulatory impact. | [30,31] | ||
| KAT2A | KAT2A plays a crucial role in acetylating lysine 18 on histone 3, a modification that is found to be elevated in the epithelial cells of individuals with asthma | [32] | ||
| SMYD3 | In asthma, SMYD3 is upregulated in airway fibroblasts, affecting cell cycle regulation through histone methylation. However, its protein-level impact remains unclear, indicating possible additional transcriptional control layers | [33,34] | ||
| Eraser | HDAC1 | HDAC1 was significantly increased in bronchial epithelial cells (HBECs) of asthmatic patients | [35] | |
| HDAC2 | Patients with mild asthma exhibit a slight decrease in HDAC2 activity in bronchial biopsies and alveolar macrophages | [36] | ||
| HDAC3 | HDAC3 regulates NF-κB activity in asthma by deacetylating specific lysine residues, suppressing inflammation. HDAC3 deficiency in macrophages reduces inflammatory gene expression, underscoring its role in controlling asthma-related inflammation. | [37] | ||
| SIRT1 | Both protective and deleterious roles in asthma | [38] | ||
| SIRT2 | SIRT2 exacerbates asthma-associated inflammation by driving Th2 cell responses and macrophage polarization. | [39] | ||
| SIRT3 | Song et al. found that decreased Sirt3 expression in asthmatic mice contributes to increased apoptosis, oxidative stress, and inflammation. | [40] | ||
| SIRT6 | Jiang et al. found that Sirt6 is upregulated in asthmatic mice | [41] | ||
| SIRT7 | Fang et al. found that increased SIRT7 expression in airway smooth muscle cells regulates TGF-β1-induced cell proliferation and migration, highlighting its role in asthmatic airway remodeling. | [42] | ||
| DNA modification | Writer | DNMT1 | DNMT1 maintains DNA methylation patterns, and reduced levels are associated with increased Socs3 expression, promoting inflammation in asthma | [43] |
| DNMT3a | Dnmt3a regulates Th2 responses by modulating IL-13 gene methylation; loss of Dnmt3a decreases methylation, enhancing IL-13 expression and asthma-associated lung inflammation | [44] | ||
| Reader | MBD2 | MBD2 is an epigenetic reader protein recognizing methylated CpG sites, suppressing SOCS3 expression, and promoting Th17 cell differentiation. Elevated MBD2 drives neutrophilic inflammation, contributing to severe asthma. | [45] | |
| Eraser | TET1 | Reduced TET1 promoter methylation (cg23602092) in nasal cells correlates with childhood asthma and traffic-related air pollution, altering TET1 expression and 5hmC.TET1 modulates DNA methylation and epigenetic regulation in asthma. | [46] | |
| RNA modification | Writer | WTAP | WTAP was demonstrated to be abnormally expressed in asthma patients.WTAP knockdown relieves asthma progression by regulating the m6A levels of AXIN1 in a YTHDF2-dependent manner. | [47,48] |
| METTL3 | METTL3 regulates Th2 cell differentiation in T2 asthma by modulating SOX5 m6A methylation in bronchial epithelial cells. This mechanism may offer a potential target for preventing and managing T2 asthma. | [49] | ||
| Reader | YTHDF1 | YTHDF1, highly expressed in airway epithelial cells of allergic and asthmatic individuals, enhances CLOCK translation in an m6A-dependent manner. This triggers NLRP3 inflammasome activation and IL-1β secretion, promoting inflammatory responses in the airways | [50] | |
| YTHDF2 | m6A-YTHDF2 regulates macrophage polarization by inhibiting M1 and promoting M2 phenotypes through NF-κB, MAPK, and STAT pathways, playing a key role in asthma subtypes and targeted therapy. | [51] | ||
| IGF2BP2 | IGF2BP2 promotes asthma by stabilizing Tsc1 mRNA, which helps macrophages adopt the M2 phenotype | [52] | ||
| Eraser | FTO | FTO plays a pivotal role as an eraser of m6A modifications in asthma by regulating the stability of mRNA transcripts such as IKBKB, leading to the activation of the NF-κB pathway and contributing to inflammation and epithelial barrier dysfunction. | [53] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



