Submitted:
14 April 2025
Posted:
15 April 2025
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
The Transmembrane Protease Serine S1 subtype 2 (TMPRSS2) and NOD-like receptor protein 3 (NLRP3) inflammasome are the main targets for the prevention and treatment of COVID-19 and influenza. The TMPRSS2 is responsible for the penetration of the SARS-CoV-2 and influenza viruses into the cell, while the hyperactivation of the NLRP3 inflammasome can lead to a cytokine storm, multiorgan damage and death. The correct strategy for preventing illness from COVID-19 and influenza is to preemptively block the TMPRSS2. Preventing the cytokine storm in COVID-19 and influenza is only effective when inhibiting NLRP3 inflammasome. Long-term prophylaxis with the TMPRSS2 inhibitor bromhexine hydrochloride and treatment with high doses of colchicine, able to inhibit the NLRP3 inflammasome, is a very effective, safe and inexpensive method against the spread and complications of COVID-19 and influenza.
Keywords:
COVID-19
; Influenza
; Bromhexine
; TMPRSS2
; NLRP3 inflammasome
; prophylactics
; colchicine
1. Introduction
With the discovery of vaccines and antibiotics, major epidemics of plague, smallpox, cholera, tuberculosis, typhoid fever, poliomyelitis are a thing of the past. However, over the past 100 years the world has been ravaged by deadly influenza and corona virus pandemics.
While the number of deaths in the First World War was around 15 million, the H1N1 pandemic in 1918-1920 caused most likely 50 million deaths (according to some even 100 million) with a world population of 1.86 billion [1].
Now, SARS-CoV-2 continues to claim victims, which are already more than 7.09 million deaths according to WHO [2]. It is more likely, however, that the number of victims may be 2-4 times higher [3]. Seasonal influenza caused by influenza viruses A and B (IAV/B) result in 3–5 million severe cases and 290,000–650,000 deaths annually [4]. A Google scholar search shows 4 980 000 articles with the keyword COVID-19 and 992 000 with the keyword Influenza.
The progress in unraveling the pathogenetic mechanisms of influenza and COVID-19 is impressive. In contrast to the great theoretical progress, the treatment recommended by the WHO is very controversial and ineffective.
2. TMPRSS2 - main target for the prevention of COVID-19 and influenza
2.1. TMPRSS2
Of the 178 serine proteases encoded in the human genome (the total number of proteases in man is 699), 138 belong to the S1 family of trypsin-like proteases [5]. TRansmembrane PRrotease Serine S1 subtype 2 (TMPRSS2) is a type II transmembrane protein (with an intracellular NH2 terminus) which belongs to the human Type II Transmembrane Serine Protease (TTSP) family of trypsin-like membrane-anchored serine proteases. TTSP has 19 members, which are grouped into 4 subfamilies: HAT/DESC, hepsin/TMPRSS (transmembrane protease/serine S1), matriptase, and corin. The TMPRSS2 is a part of the second subfamily, which has seven members - hepsin, TMPRSS2, TMPRSS3, TMPRSS4, TMPRSS5/spinesin, MSPL (mosaic serine protease large-form), and enteropeptidase [6]. As a serine protease, TMPRSS2 is involved in the cleaving peptide bonds of proteins that have serine as the nucleophilic amino acid within the active site [7].
TMPRSS2 is expressed in many epithelial tissues, including prostate (expressed several times higher than in any other tissue), lung (trachea, bronchus), breast, kidney, gastrointestinal tract (small intestine, colon), pancreas, bile duct, thymus, ovary and salivary glands [8-10].
Very little is known about the physiological role of the TMPRSS2. TMPRSS2 regulates the Na+ current by cleaving sodium channels, thereby facilitating ion passage [11-12], but it is not required for normal development, and does not cause pathological changes as demonstrared in TMPRSS2 knockout mice [13].
On the other hand, there is considerable evidence of the involvement of TMPRSS2 in various pathological processes.
v-ets erythroblastosis virus E26 oncogene-homolog (ERG) is an oncogene encoding a family of transcription factors - key regulator of cell proliferation, angiogenesis, differentiation, inflammation and apoptosis [14]. The TMPRSS2:ERG fusion gene is common in prostate cancer, occuring in about 50% of cases and driving early transformation and metastasis [15]. TMPRSS2 has been implicated in other diseases such as hepatitis C [16], chronic hepatitis B and hepatocellular carcinoma [17], ulcerative colitis [18], oncological diseases, including colorectal, gastric, prostate, pulmonary, and hepatic cancers [19].
2.2. Role of TMPRSS2 in Influenza and COVID-19 Infections
2.2.1. Influenza
Influenza viruses (IV) are classified into four categories: IAV, IBV, ICV and IDV. IAV and IBV are the predominant types, causing human seasonal epidemics. IAVs are stratified into subtypes, according to the specific configurations of their surface hemagglutinin. There is a diversity of 18 hemagglutinin (HA) and 11 neuraminidase (NA) different protein subtypes, which allows 144 potential variants [20]. In contrast, IBVs are categorized into distinct lineages: B/Yamagata and B/Victoria [21].
HA and NA are the major IV glycoprotein antigens (Fig. 1). HA is responsible for both attaching the virus to cell surface receptors and the membrane fusion, while NA mediates the release and dissemination of virions from infected cells [22]. HA0 is the inactive precursor of HA. The crucial step for virus infectivity is the cleavage of HA0 by TMPRSS2, resulting in two functional subunits with fusion competence: HA1 and HA2 [23]. TMPRSS2 is the major HA-activating protease of IAV in primary human bronchial cells and of both IAV and IBV in primary human type II pneumocytes [24,25]. HA is cleaved by TMPRSS2 in the Golgi apparatus during assembly [26]. Currently, prevalent IAV subtypes in human circulation include H1N1 and H3N2 [27].
Inhibition of TMPRSS2 would also be useful in battling respiratory parainfluenza viruses as it is an activating protease for them as well [28].
It should be noted that the dependence on TMPRSS2 is different for the different subspecies of IAV. It is bigger for H1N1/1918 and H13, while recombinant IAVs carrying H12 or H17 were not affected by TMPRSS2 knockdown [29].
2.2.2. SARS-CoV-2
SARS-CoV-2 enters into cells via two distinct pathways: one is mediated by TMPRSS2 at the cell surface (Entry via plasma membrane/ Early endocytosis) and the other done by cathepsin L/B in the endosome (Entry via endosomal pathway/Late endocytosis). TMPRSS2 could act in both early and late endosome entry processes [30,31]. TMPRSS2 also plays a critical role in the proteolytic activation of SARS-CoV, MERS-CoV and SARS-CoV-2 [29].
Although TMPRSS2 and cathepsin L/B are the main proteases responsible for cell penetration, the SARS-CoV S protein can be cleaved as well as by HAT, MSPL, DESC1 and Factor Xa [32-34]. However, TMPRSS2 serves as a major cofactor rather than cathepsins for SARS-CoV-2 cell entry [35-37].
TMPRSS2 has been implicated in the regulation of the SARS-CoV assembly in the Golgi apparatus and the release of the mature virus from the host plasma membrane [26,38-40]. TMPRSS2 is also responsible for the viral spread in the infected host [41].
Thus, spike cleavage by TMPRSS2 takes place in the plasma membrane (at cell entry), in Golgi membrane during assembly, and during virus release and spread. All these facts make TMPRSS2 an attractive target for treating both COVID-19, as well as influenza and other infectious diseases.
2.3. Inhibitors of TMPRSS2
2.3.1. Camostat Mesylate
Great hopes were placed on the TMPRSS2 inhibitor Camostat Mesylate/CM (used to treat pancreatitis and reflux esophagitis) for the treatment of COVID-19 [42]. Over 10 prestigious clinical trials have been launched to investigate the effectiveness of CM in COVID-19 and graduated with disappointing results [43-46]. This failure is not difficult to predict, because CM is administered only after the virus has already entered the cell [31].
2.3.2. Bromhexine hydrochloride
Another TMPRSS2 inhibitor is bromhexine hydrochloride (BRH), an over-the-counter, non-invasive, effective, with proven safety, available globally, inexpensive and well-tolerated medicine. Since 1963, the mucolytic cough suppressant BRH has had a long history of use in respiratory tract disorders. In addition, it has been shown to have anti-inflammatory effects, thus reducing swelling and irritation in the respiratory tract [31,47,48].
The conflicting data about the role of BRH in the prevention and treatment of COVID-19 [47-56] can be explained in just a few words - timing and mode of application. With this in mind, we hypothesized that TMPRSS2 inhibition by BRH would be most effective when taken prophylactically or administered immediately after a contact with a sick or infected person (post-exposure prophylaxis).
We recently published our data analyzing the results of 125 people who took BRH prophylactically during the COVID-19 pandemic [31]. The effect of BRH is best when given continuously for prophylaxis during peaks of contagion in the wave of COVID-19. Then the probability of infection drops sharply, and if a disease does occur, it proceeds mildly. BRH is also effective when given by inhalation for post-exposure prophylaxis. When COVID-19 manifests itself clinically, the efficacy of BRH drops sharply because the virus is already in the cell. However, BRH inhalations are useful because they limit the spread of the virus and have an antiinflammatory and possibly antiviral effect [31,57].
3. NLRP3 inflammasome - main target for treatment of COVID-19 and influenza
3.1. Structure and distribution of NLRP3 inflammasome
The NLRP3 inflammasome is a multi-protein complex, the ”cornerstone” of the innate immune system, comprising of the NLRP3 sensor protein, an adaptor protein ASC [apoptosis-associated speck-like protein containing a caspase recruitment domain (CARD)], and an effector protein (pro-caspase-1). The NLRP3 protein, aptly named ”architectural marvel”, consists of three distinct domains: a C-terminal leucine-rich repeat (LRR) sensor domain, a central NACHT domain responsible for oligomerization and an N-terminal pyrin (PYD) domain responsible for protein-protein interactions [58,59]. This intricate structure allows NLRP3 to function as a molecular switch, transitioning from an inactive to an active state in response to cellular danger signals [60].
3.2. Activation of NLRP3 inflammasome
In the absence of activating signals, the inactive oligomeric NLRP3 is in a complex with HSP90 and SGT1 in the cytoplasm and pro-IL-1β is not constitutively expressed in resting macrophages [63].
NF-κB (nuclear factor kappa-light-chain-enhancer of activated B cells) upregulates the expression of NLRP3, which under resting conditions exists at concentrations that are inadequate for initiating inflammasome activation (supramolecular complex) and pro-IL-1β maturation [64,65].
The canonical activation of NLRP3 inflammasomes requires two signals: signal 1 (priming) is provided by TLR (Toll-like receptors), IL-1 and TNF (tumour necrosis factor) receptors activation, which will induce the activation of NF-κB and the up-regulation of inactive pro-IL-1β, pro-IL-18 and inflammasome components [66,67].
According to others, priming signals do not affect the expression levels of pro-IL-18, ASC and pro-caspase-1 [59,68]. Inflammasome activation and inflammatory ‘strength’ is modulated via transcriptional regulation of signal 1 [66].
Signal 2 is provided by various stimuli, including K+ efflux, extracellular ATP activation of the P2X7 receptor, pore-forming toxins, crystalline substances, cellular stress signals as reactive oxygen species (ROS), influenza virus M2/PB1-F2 proteins, ZBP1 (Z-DNA binding protein 1) and RNA viruses. These events lead to the oligomerization of the inflammasome complex through ASC interaction with inactive pro-caspase 1, which is activated. The activated caspase 1 cleaves pro-IL-1β/IL-18 to IL-1β/IL-18 and gasdermin D, which induces pore formation in the plasma membrane and consequently IL-1β/IL-18 release and pyroptotic cell death [66]. IL-1β induces the production of IL-6, another abundantly detected cytokine in severe COVID-19 patients [69,70]. IL-6 stimulates the release of various acute phase proteins, such as the hepatic factors C-reactive protein and ferritin, which are associated with poor prognosis [71,72]. IL-18 has been linked to ferritin production [73] and is another line of evidence for NLRP3 in severe COVID, as its level is significantly higher in symptomatic patients and is increased in accordance with disease severity. IL-1β itself has been associated with severe COVID-19 [74,75]. In macrophages, the priming signal (signal 1) is mandatory for inducing NLRP3 inflammasome activation [68].
SARS-CoV-2 can activate NLRP3 inflammasome by ORF3a (open reading frame 3a) [76,77]; Nucleocapsid [78]; NS6 (non-structural protein 6) [79]; Spike [80,81]; NS5 [82]; viroporin E [77]; viroporin M [83].
Many other viruses can activate the NLRP3 inflammasome such as SARS-CoV, Influenza, Parainfluenza, Adenovirus, Bocavirus, Metapneumovirus, Respiratory syncytial virus, Rhinovirus [84].
The role of activated NLRP3 is to limit the infection by SARS-CoV-2 [85]. When the NLRP3 inflammasome is properly activated, IL-1 β production and the activation of the immune system help to resolve the infection. However, inflammasome inhibition by the virus, leads to easy replication and spread, resulting in worse disease prognosis. The same applies to its hyperactivation, leading to cytokine storm, multiorgan damage, and often death [85]. Thus, the regulation of the NLRP3 complex and pro-IL-1β expression are critical to limit inflammasome function [66].
3.3. Regulation of NLRP3 inflammasome
NLRP3 inflammasome is subjected to strict post-translational regulation including phosphorylation/dephosphorylation, ubiquitination/deubiquitination, SUMOylation, S-nitrosylation, ADP-ribosylation, acetylation, O-GlcNAcylation, nitration of Tyr861, glycosylation, palmitoylation [for. Refs see 67].
For example, negative NLRP3 inflammasome regulators are several ubiquitin ligases, including TRIM31 [86], MARCH7 [87], RNF125 [87], CBL-b [88], FBXL2 [88], ARIH2 [89] and Cullin1 [90].
While most ubiquitin ligases are negative regulators of NLRP3 inflammasome, some as Pellino2 [91] and TRAF6 [92] promote its activation.
In most cases the action of the deubiquitinating enzymes such as BRCC3 [93], USP7 [94], USP47 [94], leads to positive regulation of the NLRP3 inflammasome. However, A20 functions as NLRP3 inflammasome negative regulator [95].
Phosphorylation/dephosphorylation plays a critical role in the NLRP3 inflammasome regulation, with different phosphorylation sites exerting sometimes opposing effects [96]. Positive regulators are protein kinase D [97], protein phosphatase 2A (PP2A) [88], c-Jun N-terminal kinase 1 (JNK1) [98], protein tyrosine phosphatase nonreceptor 22 (PTPN22) [99], whereas negative regulator is PKA [89] while AKT phosphorylation has a dual role [100].
The activated NIMA-related kinase 7 (NEK7) can induce NLRP3 conformational change, leading to disruption of its inactive double-ring structure [90].
Another negative regulator is type I interferon (IFN) expression believed to prevent the induction of a hyperinflammatory NLRP3 inflammasome activation [101,102]. IFN-I is required for the recruitment of pro-inflammatory monocytes and macrophages to the infected lungs [103]. However, the early use of IFNα decreased mortality, whereas late use of IFNα increased mortality [104].
People with severe COVID-19 have symptoms of systemic hyperinflammation, mediated by a rapid release of inflammatory molecules, especially inflammatory cytokines such as interleukin (IL)-1β, IL-18, IL-6 and tumor necrosis factor-α, and the protein Gasdermin D (GSDMD) which is a marker of inflammatory cell death [105].
4. Clinical course of influenza and COVID-19
4.1. FLU
Clinically, the flu progresses as follows: After an incubation period of an average of 2 days, the flu begins acutely and day 1 to day 3 is characterized by worsening of the symptoms. FLU symptoms usually peak between days 2 and 4. A slight decline in induction is noted on day 4, most people feel better after 5–7 days and recover on day 8. In this case, there is a normal response to the NLRP3 inflammasome. However, in some people the desease rebounds on day 7 to levels similar to the onset of infection, suggesting two ‘waves’ of inflammation, leading to a ‘feed-forward’ inflammatory loop [106]. This development of the disease corresponds to hyperactivation of the NLRP3 inflammasome with excessive IL-1β and IL-18 maturation, with subsequent increase in levels of IL-6 and other cytokines (CS).
Thus, the NLRP3 inhibition during the first five days after the onset of IAV infection can be detrimental, while it proves beneficial in the later stages (the period from the seventh day to the ninth day) [106].
4.2. COVID-19
Clinically, COVID-19 progresses as follows: After an incubation period of an average of 2-14 days, day 1 is characterized by mild symptoms, which escalate during day 2 to day 7. Days 8-10 are critical and are followed by either improving or worsening symptoms. Recovery occurs between days 15 and 21. If the NLRP3 inflammasome response is normal, recovery should occur after the critical 8-10 days. However, after hyperactivation of NLRP3 inflammasome, the cytokine storm leads to multiorgan damage, microthrombosis, and death. LDH level on day 8 is the strongest predictor of in-hospital mortality in COVID-19 inpatients and the second one is the decreased number of peripheral blood lymphocytes [107]. Thus, the inhibition of NLRP3 inflammasome is mandatory to prevent cytokine storm.
In short, the incubation period of COVID-19 is longer, and it is more contagious than the flu, the onset of symptoms is milder, the recovery is slower, complications due to NLRP3 inflammasome hyperactivation are more common, and mortality is higher. What is the common clinical feature between COVID-19 and the flu are the critical days during the development of the symptoms (Tabl. 1).
5. Strategies for dealing with COVID-19 and the flu - which one is the winner?
5.1. Antivirals and inhibitors of individual cytokines
The best strategy would be to prevent the virus from entering the host cells.
For a number of reasons, vaccines are not effective enough against either COVID-19 or the flu. COVID-19 vaccines do not appear to be effective against secondary infection. What's more, significantly high number of vaccinated individuals had a secondary encounter and subsequent infection with COVID-19 [108], which is explained by their “high-risk” activities, such as avoiding social distancing, engaging in more-frequent public activities [109].
Drug-based blocking of the virus entry into the cell may be used both as primary and post-exposure prophylaxis. As we emphasized above, the effect of TMPRSS2 inhibitors drops dramatically after the onset of the disease, as the viruses are already in the cells and multiplying and spreading intensively. This also explains the collapse of clinical trials with the TMPRSS2 inhibitor camostat mesylate [43-46].
The WHO and Big Pharma strategy of using antivirals and inhibiting individual cytokines is not productive, because there is no direct correlation between viral load and NLRP3 inflammasome hyperreaction [110]. Furthermore, inhibiting individual cytokines in the presence of ongoing NLRP3 inflammasome hyperactivity is pointless. [48,57,110].
It is clear that the main cause of complications in COVID-19 and influenza is the hyperactivated NLRP3 inflammasome. Its regulation is multifactorial. How all these positive and negative regulatory factors act spatially and temporally, what their cumulative effect is, and why around days 8 and 10 the NLRP3 inflammasome becomes hyperactivated in complicated cases remains a mystery. Moreover, the regulation of the normal NLRP3 inflammasome response, day by day, during the illness is unclear. It is mandatory to inhibit NLRP3 inflammasome when clinical signs of impending complications are present.
5.2. Inhibition of TMPRSS2 and NLRP3 inflammasome
Our strategy for the prevention and treatment of Covid-19 and influenza is based on the following facts:
- 1)
- SARS-CoV-2 and influenza A and B viruses enter the cell mainly through TMPRSS2.
- 2)
- Bromhexine inhibits TMPRSS2.
- 3)
- The effect of BRH is best when taken prophylactically, not after the viruses have already entered the cell.
- 4)
- Post-exposure prophylaxis with BRH is also very effective, especially when done by inhalation.
- 5)
- Complications in both influenza and COVID-19 are due to a NLRP3 inflammasome hyperreaction, which causes a cytokine storm.
- 6)
- Colchicine accumulates in myeloid cells, which explains its NLRP3 inflammasome inhibitory effect at high doses.
- 7)
- Inhibition of the NLRP3 inflammasome prevents cytokine storm and normalizes cytokine levels.
- 8)
- Higher doses of colchicine have been used in the past and are completely safe, provided the rules for administering colchicine are being followed.
- 9)
- With a series of clinical cases, we demonstrated the life-saving effect of high doses of colchicine in critically ill patient [111], those with high levels of obesity [112-114], and the unique case of recovery of a 101-year-old patient infected with COVID-19 in intensive care after major surgery [115]. It is very demonstrative that four patients who mistakenly took more than 12.5 mg of colchicine recovered quickly and completely after discontinuing all therapy [47,116].
- 10)
- Studies on 795 inpatients treated with high doses of colchicine reduced mortality by 2 to 7 times, in direct proportion to increasing doses [117,118]. Outpatients’ high-dose colchicine treatment practically prevents hospitalizations and demonstrates reverse relationship with hospitalization [108]. The maximum loading doses we use of up to 5mg of colchicine (0.045mg/kg) are completely safe [48,57,119].
- 11)
- 12)
- The epidemic situation in Bulgaria for the winter of 2025 is dominated by influenza viruses A(H3N2 and H1N1pdm09) and influenza viruses B/Victoria. We are monitoring several hundred people taking BRH prophylactically. The results will be summarized at the end of the epidemic, but so far none of those taking prophylactic BRH have fallen ill. Our preliminary results were reported at a national conference [121] and published [122].
Conclusion
Prophylaxis with bromhexine and treatment with high doses of colchicine is a very effective and inexpensive method against the spread and complications of COVID-19 and influenza.
Hemagglutinin (HA); Fusion Peptide (FP); Transmembrane domain (TM); Low-Density Lipoprotein Receptor class A (LDLR-A); Scavenger Receptor Cysteine-Rich (SRCR); Serine Protease domain (SP).
Figure 1.
Dependence of Influenza and SARS-CoV-2 viruses on TMPRSS2.
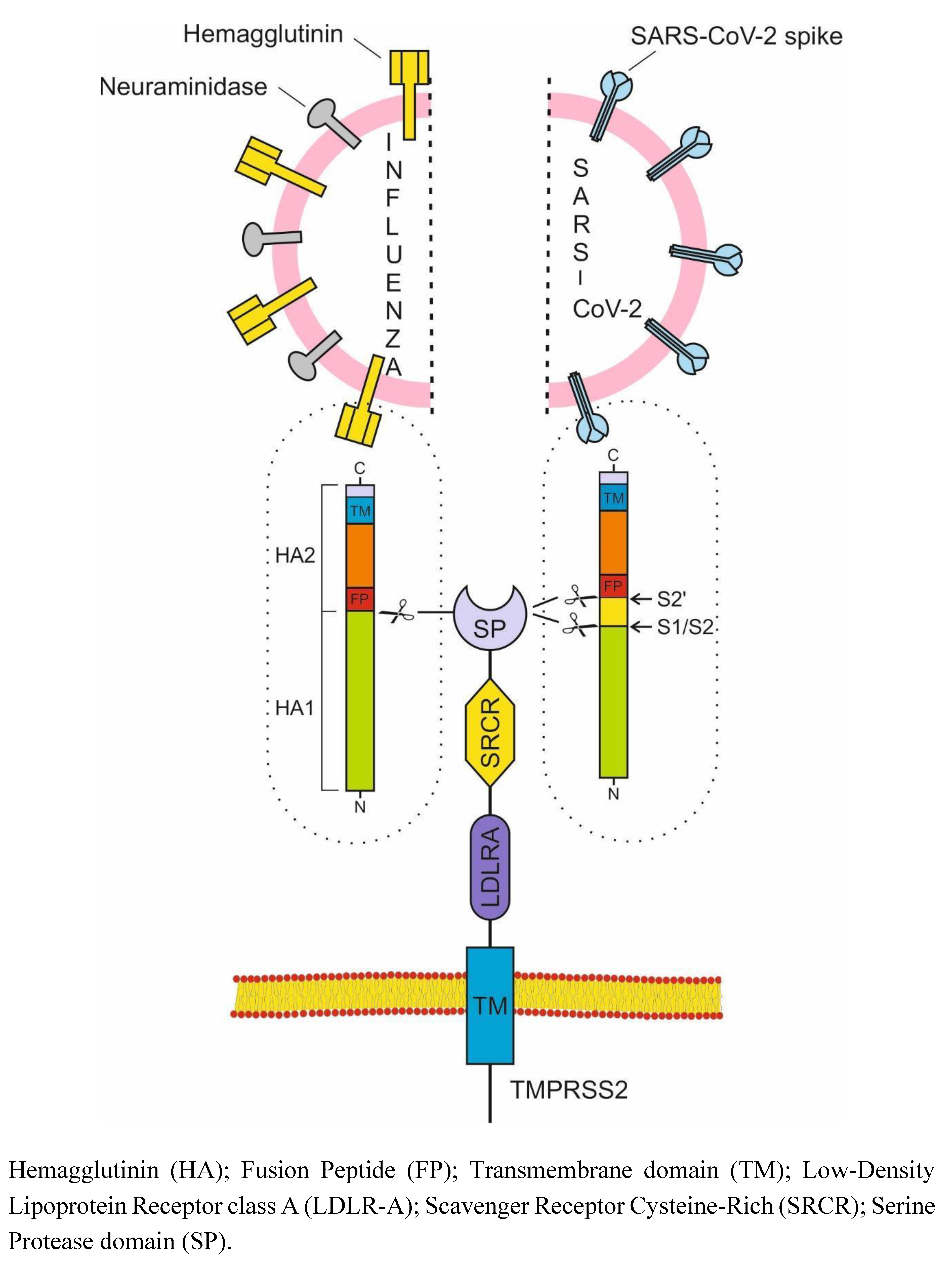

Table 1.
Viral, Clinical and Laboratory Comparison Between Influenza and COVID-19.
| Influenza | COVID-19 | |
|---|---|---|
| Viruses | Spanish influenza in 1918 (H1N1), Asian influenza in 1957 (H2N2), Hong Kong influenza in 1968 (H3N2), Russian influenza in 1977 (H1N1) | SARS-CoV-2, 2019 |
| Virus family | Orthomyxoviridae | Coronaviridae |
| Viral nucleic acid | Negative sense single-stranded RNA | Positive sense single-stranded RNA |
| Vulnerable contingent |
children and adults H1N1 influenza in 1918 - young people |
adults |
| Incubation period | 2 days | 2-14 days |
| R0 | 1.4-2.8 | 5.7 |
| Tropism | Respiratory tract epithelium | Multiple organs |
| Host receptor | α 2,6 sialic acids | ACE2 |
| Viral proteins required for fusion | HA | S |
| Critical protease | TMPRSS2 | TMPRSS2 |
| First symptom | Cough | Fever |
| Contagiousness |
One day before the onset of symptoms and up to five to seven days after | 48 hours before the onset of symptoms and up to 10 days after |
| Most contagious | The first three days after the onset of symptoms | 1-2 days before the onset of symptoms |
| Viral load peak |
2 day | Early at the onset of symptoms, now in a highly immune adult population - 4 day |
| Worst Days Alert | 7-9 | 8-10 |
| Cause of cytokine storm | NLRP3 hyperactivation | NLRP3 hyperactivation |
| IL-6/ IL-1/ IL-18/ D-Dimer/ Hypercoagulative state/ DIC/ Endotheliopathy / Ferritin/ Activated macrophages/ High neutrophil to Lymphocyte ratio/ Immunothrombosis, LDH | ++/++/+/++/++/++/ +++/++/+/++/ ++/+++ |
+++/+++/+++/++/+++/ ++/+++/++/+++/++/++/+++ |
| Radiological findings | Multilobe consolidations | Ground-glass opacities |
| Need for hospitalization | 5.6% | 20% |
| Need for intubation | 4.8% | 10%–15% |
| Mortality | 0.13%–1.36% | 1.40%–3.67% |
| Prophylaxis | BRH | BRH |
| Рost-exposure prophylaxis | Inhaled BRH | Inhaled BRH |
| Cytokine storm treatment | High colchicine doses | High colchicine doses |
ACE2, angiotensin-converting enzyme metallopeptidase 2; COVID-19, coronavirus disease 2019; HA, hemagglutinin; S-spike protein, cytokine storm (CS).
Funding Statement
The work was funded by Project BG-RRP-2.004-0004-C01 financed by Bulgarian National Science Fund. The research is financed by the Bulgarian National Plan for Recovery and Resilience.
References
- Johnson NPAS, Mueller J. Updating the accounts: Global mortality of the 1918-1920 “spanish” influenza pandemic. Bulletin of the History of Medicine [Internet]. 2002;76(1):105–15. Available from: https://pubmed.ncbi.nlm.nih.gov/11875246/.
- World Health Organization. COVID-19 deaths | WHO COVID-19 dashboard [Internet]. World Health Organization Data. 2024. Available from: https://data.who.int/dashboards/covid19/deaths.
- Adam, D. The pandemic’s true death toll: Millions more than official counts. Nature [Internet]. 2022 Jan 18;601(7893):312–5. Available from: https://www.nature.com/articles/d41586-022-00104-8.
- World Health Organization. Influenza (seasonal) [Internet]. Who.int. World Health Organization: WHO; 2023. Available from: https://www.who.int/news-room/fact-sheets/detail/influenza-(seasonal).
- Di Cera, E. Serine Proteases. IUBMB Life. 2009 May;61(5):510–5.
- Bugge TH, Antalis TM, Wu Q. Type II Transmembrane Serine Proteases. Journal of Biological Chemistry. 2009 Aug;284(35):23177–81.
- Fraser BJ, Beldar S, Seitova A, Hutchinson A, Mannar D, Li Y, et al. Structure and activity of human TMPRSS2 protease implicated in SARS-CoV-2 activation. Nature Chemical Biology [Internet]. 2022 Sep 1;18(9):963–71. Available from: https://www.nature.com/articles/s41589-022-01059-7.
- Li X, He L, Luo J, Zheng Y, Zhou Y, Li D, et al. Paeniclostridium sordellii hemorrhagic toxin targets TMPRSS2 to induce colonic epithelial lesions. Nature Communications. 2022 Jul 26;13(1).
- Harbig A, Mernberger M, Bittel L, Pleschka S, Klaus Schughart, Torsten Steinmetzer, et al. Transcriptome profiling and protease inhibition experiments identify proteases that activate H3N2 influenza A and influenza B viruses in murine airways. 2020 Aug 14;295(33):11388–407.
- Sungnak W, Huang N, Bécavin C, Berg M, Queen R, Litvinukova M, et al. SARS-CoV-2 Entry Factors Are Highly Expressed in Nasal Epithelial Cells Together with Innate Immune Genes. Nature Medicine [Internet]. 2020 May 1;26(5):681–7. Available from: https://www.nature.com/articles/s41591-020-0868-6.
- Donaldson SH, Hirsh A, Li DC, Holloway G, Chao J, Boucher RC, et al. Regulation of the Epithelial Sodium Channel by Serine Proteases in Human Airways. Journal of Biological Chemistry. 2002 Mar;277(10):8338–45.
- Florian Sure, Marko Bertog, Afonso S, Alexei Diakov, Rinke R, M. Gregor Madej, et al. Transmembrane serine protease 2 (TMPRSS2) proteolytically activates the epithelial sodium channel (ENaC) by cleaving the channel’s γ-subunit. 2022 Apr 1;298(6):102004–4.
- Kim TS, Heinlein C, Hackman RC, Nelson PS. Phenotypic Analysis of Mice Lacking the Tmprss2-Encoded Protease. Molecular and Cellular Biology. 2006 Feb 1;26(3):965–75.
- Gasi Tandefelt D, Boormans J, Hermans K, Trapman J. ETS Fusion Genes in Prostate Cancer. Endocrine-Related Cancer. 2014 Mar 20;21(3):R143–52.
- Wang Z, Wang Y, Zhang J, Hu Q, Zhi F, Zhang S, et al. Significance of the TMPRSS2:ERG Gene Fusion in Prostate Cancer. Molecular Medicine Reports [Internet]. 2017 Oct 1;16(4):5450–8. Available from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5647090/.
- Esumi M, Ishibashi M, Yamaguchi H, Nakajima S, Tai Y, Kikuta S, et al. Transmembrane Serine Protease TMPRSS2 Activates Hepatitis C Virus Infection. Hepatology. 2015 Jan 20;61(2):437–46.
- Melis M, Diaz G, Kleiner DE, Zamboni F, Kabat J, Lai J, et al. Viral Expression and Molecular Profiling in Liver Tissue versus Microdissected Hepatocytes in Hepatitis B Virus - Associated Hepatocellular Carcinoma. Journal of translational medicine. 2014 Aug 21;12(1).
- Hamamoto Y, Kawamura M, Uchida H, Hiramatsu K, Chiaki Katori, Asai H, et al. Increased ACE2 and TMPRSS2 Expression in Ulcerative Colitis. Pathology - Research and Practice. 2024 Jan 10;254:155108–8.
- Hoang T, Nguyen TQ, Tran TTA. Genetic Susceptibility of ACE2 and TMPRSS2 in Six Common Cancers and Possible Impacts on COVID-19. Cancer Research and Treatment. 2020 Dec 29.
- Barros G, Everton Nencioni, Fábio Thimoteo, Perea C, Fuzaro R, Sasaki SD. TMPRSS2 as a Key Player in Viral Pathogenesis: Influenza and Coronaviruses. Biomolecules. 2025 Jan 7;15(1):75–5.
- Zhang N, Fang S, Wang T, Li J, Cheng X, Zhao C, et al. Applicability of a Sensitive Duplex real-time PCR Assay for Identifying B/Yamagata and B/Victoria Lineages of Influenza Virus from Clinical Specimens. Applied Microbiology and Biotechnology. 2011 Nov 24;93(2):797–805.
- Krammer F, Smith GJD, Fouchier RAM, Peiris M, Kedzierska K, Doherty PC, et al. Influenza. Nature Reviews Disease Primers [Internet]. 2018 Jun 28;4(1). Available from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7097467/.
- Böttcher-Friebertshäuser E, Garten W, Matrosovich M, Klenk HD. The Hemagglutinin: A Determinant of Pathogenicity. Influenza Pathogenesis and Control - Volume I. 2014;3–34.
- BöttcherE, Matrosovich T, Beyerle M, Klenk HD, Garten W, Matrosovich M. Proteolytic Activation of Influenza Viruses by Serine Proteases TMPRSS2 and HAT from Human Airway Epithelium. Journal of Virology. 2006 Oct 1;80(19):9896–8.
- Limburg H, Harbig A, Bestle D, Stein DA, Moulton HM, Jaeger J, et al. TMPRSS2 Is the Major Activating Protease of Influenza a Virus in Primary Human Airway Cells and Influenza B Virus in Human Type II Pneumocytes. Schultz-Cherry S, editor. Journal of Virology. 2019 Nov;93(21).
- Shen LW, Mao HJ, Wu YL, Tanaka Y, Zhang W. TMPRSS2: A potential target for treatment of influenza virus and coronavirus infections. Biochimie [Internet]. 2017 Nov 1;142:1–10. Available from: https://www.sciencedirect.com/science/article/pii/S0300908417301876?
- Bi Y, Yang J, Wang L, Ran L, Gao GF. Ecology and Evolution of Avian Influenza Viruses. Current Biology. 2024 Aug;34(15):R716–21.
- Abe M, Tahara M, Sakai K, Yamaguchi H, Kazuhiko Kanou, Kazuya Shirato, et al. TMPRSS2 Is an Activating Protease for Respiratory Parainfluenza Viruses. Journal of Virology. 2013 Aug 22;87(21):11930–5.
- Schwerdtner M, Schmacke LC, Nave J, Limburg H, Steinmetzer T, Stein DA, et al. Unveiling the Role of TMPRSS2 in the Proteolytic Activation of Pandemic and Zoonotic Influenza Viruses and Coronaviruses in Human Airway Cells. Viruses. 2024 Nov 20;16(11):1798.
- Hoffmann M, Kleine-Weber H, Schroeder S, Krüger N, Herrler T, Erichsen S, et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell. 2020 Mar;181(2):271–80.
- Vanyo Mitev, Tsanko Mondeshki, Ani Miteva, Konstantin Tashkov, Dimitrova V. COVID-19 Prophylactic Effect of Bromhexine Hydrochloride. 2024 Oct 25.
- Simmons G, Gosalia DN, Rennekamp AJ, Reeves JD, Diamond SL, Bates P. Inhibitors of Cathepsin L Prevent Severe Acute Respiratory Syndrome Coronavirus Entry. Proceedings of the National Academy of Sciences [Internet]. 2005 Aug 16;102(33):11876–81. Available from: https://www.pnas.org/content/102/33/11876.
- Simmons G, Zmora P, Gierer S, Heurich A, Pöhlmann S. Proteolytic Activation of the SARS-coronavirus Spike protein: Cutting Enzymes at the Cutting Edge of Antiviral Research. Antiviral Research. 2013 Dec;100(3):605–14.
- Du L, Kao RY, Zhou Y, He Y, Zhao G, Wong C, et al. Cleavage of Spike Protein of SARS Coronavirus by Protease Factor Xa Is Associated with Viral Infectivity. Biochemical and Biophysical Research Communications. 2007 Jul;359(1):174–9.
- Koch J, Uckeley ZM, Doldan P, Stanifer M, Boulant S, Lozach P. TMPRSS2 Expression Dictates the Entry Route Used by SARS-CoV-2 to Infect Host Cells. The EMBO Journal. 2021 Jul 13;40(16).
- Hoffmann M, Kleine-Weber H, Pöhlmann S. A Multibasic Cleavage Site in the Spike Protein of SARS-CoV-2 Is Essential for Infection of Human Lung Cells. Molecular Cell. 2020 May;78(4).
- Mykytyn AZ, Breugem TI, Geurts MH, Beumer J, Schipper D, Romy van Acker, et al. SARS-CoV-2 Omicron Entry Is Type II Transmembrane Serine protease-mediated in Human Airway and Intestinal Organoid Models. Journal of virology. 2023 Aug 31;97(8).
- Ou X, Liu Y, Lei X, Li P, Mi D, Ren L, et al. Characterization of Spike Glycoprotein of SARS-CoV-2 on Virus Entry and Its Immune cross-reactivity with SARS-CoV. Nature Communications. 2020 Mar 27;11(1).
- Maggio R, Corsini GU. Repurposing the Mucolytic Cough Suppressant and TMPRSS2 Protease Inhibitor Bromhexine for the Prevention and Management of SARS-CoV-2 Infection. Pharmacological Research. 2020 Jul;157:104837.
- Depfenhart, M. A SARS-CoV-2 Prophylactic and Treatment; a Counter Argument against the Sole Use of Chloroquine. American Journal of Biomedical Science & Research. 2020 Apr 9;8(4):248–51.
- Iwata-Yoshikawa N, Okamura T, Shimizu Y, Hasegawa H, Takeda M, Nagata N. TMPRSS2 Contributes to Virus Spread and Immunopathology in the Airways of Murine Models after Coronavirus Infection. Gallagher T, editor. Journal of Virology. 2019 Jan 9;93(6).
- Breining P, Frølund AL, Højen JF, Gunst JD, Staerke NB, Saedder E, et al. Camostat Mesylate against SARS-CoV-2 and COVID-19—Rationale, Dosing and Safety. Basic & Clinical Pharmacology & Toxicology. 2020 Nov 22;128(2):204–12.
- Tobback E, Degroote S, Buysse S, Delesie L, Van Dooren L, Vanherrewege S, et al. Efficacy and Safety of Camostat Mesylate in Early COVID-19 Disease in an Ambulatory setting: a Randomized placebo-controlled Phase II Trial. International Journal of Infectious Diseases. 2022 Sep;122:628–35.
- Khan U, Muhammad Mubariz, Yehya Khlidj, Nasir MM, Ramadan S, Saeed F, et al. Safety and Efficacy of Camostat Mesylate for Covid-19: a Systematic Review and Meta-analysis of Randomized Controlled Trials. BMC Infectious Diseases. 2024 Jul 19;24(1).
- Kinoshita T, Shinoda M, Nishizaki Y, Shiraki K, Hirai Y, Kichikawa Y, et al. A multicenter, double-blind, randomized, parallel-group, placebo-controlled Study to Evaluate the Efficacy and Safety of Camostat Mesilate in Patients with COVID-19 (CANDLE study). BMC Medicine. 2022 Sep 27;20(1).
- Chupp G, Spichler-Moffarah A, Søgaard OS, Esserman D, Dziura J, Danzig L, et al. A Phase 2 Randomized, Double-Blind, Placebo-controlled Trial of Oral Camostat Mesylate for Early Treatment of COVID-19 Outpatients Showed Shorter Illness Course and Attenuation of Loss of Smell and Taste. medRxiv: The Preprint Server for Health Sciences [Internet]. 2022 Jan 31 [cited 2022 Oct 17]; 2022.01.28.22270035. Available from: https://pubmed.ncbi.nlm.nih.gov/35132421/.
- Marinov K, Tsanko Mondeshki, Georgiev H, Dimitrova VS, Vanyo Mitev. Effects of long-term Prophylaxis with Bromhexine Hydrochloride and Treatment with High Colchicine Doses of COVID-19. Pharmacia. 2025 Jan 13;72:1–10.
- Vanyo Mitev. Comparison of Treatment of COVID-19 with Inhaled bromhexine, Higher Doses of Colchicine and Hymecromone with WHO-recommended paxlovid, molnupiravir, remdesivir, anti-IL-6 Receptor Antibodies and Baricitinib. Pharmacia/Farmaciâ. 2023 Oct 20;70(4):1177–93.
- Depfenhart M, de Villiers D, Lemperle G, Meyer M, Di Somma S. Potential New Treatment Strategies for COVID-19: Is There a Role for Bromhexine as add-on therapy? Internal and Emergency Medicine. 2020 May 26.
- Ansarin K, Tolouian R, Ardalan M, Taghizadieh A, Varshochi M, Teimouri S, et al. Effect of Bromhexine on Clinical Outcomes and Mortality in COVID-19 patients: a Randomized Clinical Trial. BioImpacts. 2020 Jul 19;10(4):209–15.
- Eslami Ghayour A, Nazari S, Keramat F, Shahbazi F, Eslami-Ghayour A. Evaluation of the Efficacy of N-acetylcysteine and Bromhexine Compared with Standard Care in Preventing Hospitalization of Outpatients with COVID-19: a Double Blind Randomized Clinical Trial. Revista Clínica Española (English Edition) [Internet]. 2024 Feb 1 [cited 2024 Jun 17];224(2):86–95. Available from: https://www.revclinesp.es/en-evaluation-efficacy-n-acetylcysteine-bromhexine-compared-articulo-S225488742400002X?
- Fu Q, Zheng X, Zhou Y, Tang L, Chen Z, Ni S. Re-recognizing Bromhexine hydrochloride: Pharmaceutical Properties and Its Possible Role in Treating Pediatric COVID-19. European Journal of Clinical Pharmacology [Internet]. 2021 [cited 2022 Dec 9];77(2):261–3. Available from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7459257/.
- Ramin Tolouian, Moradi O, Mulla ZD, Shadi Ziaie, Haghighi M, Hadi Esmaily, et al. Bromhexine, for Post Exposure COVID-19 prophylaxis: a Randomized, Double-Blind, Placebo Control Trial. Research Square (Research Square). 2022 May 6.
- Ramin Tolouian, Mulla ZD. Controversy with Bromhexine in COVID-19; Where We Stand. Immunopathologia Persa. 2020 Aug 12;7(2):e12–2.
- Ogbac MK, Tamayo JE. Effect of Bromhexine among COVID-19 Patients - a Meta-anaylsis. 대한결핵및호흡기학회 추계학술발표초록집 [Internet]. 2021 [cited 2025 Apr 7];129(0):412–2. Available from: https://kiss.kstudy.com/Detail/Ar?key=3921594.
- Vila Méndez ML, Antón Sanz C, Cárdenas García A del R, Bravo Malo A, Torres Martínez FJ, Martín Moros JM, et al. Efficacy of Bromhexine versus Standard of Care in Reducing Viral Load in Patients with Mild-to-Moderate COVID-19 Disease Attended in Primary Care: a Randomized Open-Label Trial. Journal of Clinical Medicine [Internet]. 2022 Dec 24 [cited 2024 Mar 20];12(1):142. Available from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC9821213/.
- Vanyo Mitev. Colchicine—The Divine Medicine against COVID-19. Journal of Personalized Medicine. 2024 Jul 16;14(7):756–6.
- Duan Y, Wang J, Cai J, Kelley N, He Y. The leucine-rich Repeat (LRR) Domain of NLRP3 Is Required for NLRP3 Inflammasome Activation in Macrophages. Journal of Biological Chemistry. 2022 Dec 1;298(12):102717–7.
- Kelley N, Jeltema D, Duan Y, He Y. The NLRP3 Inflammasome: an Overview of Mechanisms of Activation and Regulation. International Journal of Molecular Sciences [Internet]. 2019 Jul 6;20(13):3328. Available from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6651423/.
- Anil Akbal, Alesja Dernst, Lovotti M, Mangan M, McManus RM, Latz E. How Location and Cellular Signaling Combine to Activate the NLRP3 Inflammasome. Cellular & Molecular Immunology. 2022 Sep 20;19(11):1201–14.
- Ichinohe T, Lee HK, Ogura Y, Flavell R, Iwasaki A. Inflammasome Recognition of Influenza Virus Is Essential for Adaptive Immune Responses. The Journal of Experimental Medicine [Internet]. 2009 Jan 19;206(1):79–87. Available from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2626661/.
- Allen IC, Scull MA, Moore CB, Holl EK, McElvania-TeKippe E, Taxman DJ, et al. The NLRP3 Inflammasome Mediates in Vivo Innate Immunity to Influenza a Virus through Recognition of Viral RNA. Immunity. 2009 Apr;30(4):556–65.
- Swanson KV, Deng M, Ting JPY. The NLRP3 inflammasome: Molecular Activation and Regulation to Therapeutics. Nature Reviews Immunology [Internet]. 2019 Apr 29;19(8):477–89. Available from:https://www.nature.com/articles/s41577-019-0165-0.
- Dadkhah M, Sharifi M. The NLRP3 inflammasome: Mechanisms of activation, regulation, and Role in Diseases. International Reviews of Immunology. 2024 Oct 14;44(2):98–111.
- Franchi L, Eigenbrod T, Núñez G. Cutting Edge: TNF-α Mediates Sensitization to ATP and Silica via the NLRP3 Inflammasome in the Absence of Microbial Stimulation. The Journal of Immunology. 2009 Jun 19;183(2):792–6.
- Tate MD, Mansell A. An Update on the NLRP3 Inflammasome and influenza: the Road to Redemption or perdition? Current Opinion in Immunology. 2018 Jun 30;54:80–5.
- Beesetti, S. Ubiquitin Ligases in Control: Regulating NLRP3 Inflammasome Activation. Frontiers in Bioscience-Landmark. 2025 Mar 19;30(3).
- Bauernfeind FG, Horvath G, Stutz A, Alnemri ES, MacDonald K, Speert D, et al. Cutting edge: NF-kappaB Activating Pattern Recognition and Cytokine Receptors License NLRP3 Inflammasome Activation by Regulating NLRP3 Expression. Journal of Immunology (Baltimore, Md: 1950) [Internet]. 2009 Jul 15;183(2):787–91. Available from: https://pubmed.ncbi.nlm.nih.gov/19570822/.
- Liu X, Wang H, Shi S, Xiao J. Association between IL-6 and Severe Disease and Mortality in COVID-19 disease: a Systematic Review and meta-analysis. Postgraduate Medical Journal. 2021 Jun 3;postgradmedj-2021-139939.
- Tosato G, Jones K. Interleukin-1 Induces interleukin-6 Production in Peripheral Blood Monocytes. Blood. 1990 Mar 15;75(6):1305–10.
- Chen W, Zheng KI, Liu S, Yan Z, Xu C, Qiao Z. Plasma CRP Level Is Positively Associated with the Severity of COVID-19. Annals of Clinical Microbiology and Antimicrobials. 2020 May 15;19(1).
- Manson JJ, Crooks C, Naja M, Ledlie A, Goulden B, Liddle T, et al. COVID-19-associated Hyperinflammation and Escalation of Patient care: a Retrospective Longitudinal Cohort Study. The Lancet Rheumatology [Internet]. 2020 Oct 1;2(10):e594–602. Available from: https://www.thelancet.com/journals/lanrhe/article/PIIS2665-9913(20)30275-7/fulltext#seccestitle150.
- Slaats J, ten Oever J, van de Veerdonk FL, Netea MG. IL-1β/IL-6/CRP and IL-18/ferritin: Distinct Inflammatory Programs in Infections. Bliska JB, editor. PLOS Pathogens. 2016 Dec 15;12(12):e1005973.
- Potere N, Del Buono MG, Caricchio R, Cremer PC, Vecchié A, Porreca E, et al. Interleukin-1 and the NLRP3 Inflammasome in COVID-19: Pathogenetic and Therapeutic Implications. eBioMedicine. 2022 Nov;85:104299.
- Yin M, Marrone L, Peace CG, O’Neill LAJ. NLRP3, the Inflammasome and COVID-19 Infection. QJM: An International Journal of Medicine [Internet]. 2023 Jan 20 [cited 2025 Feb 11];116(7):502–7. Available from: https://pmc.ncbi.nlm.nih.gov/articles/PMC10382191/.
- Xu H, Akinyemi IA, Chitre SA, Loeb JC, Lednicky JA, McIntosh MT, et al. SARS-CoV-2 Viroporin Encoded by ORF3a Triggers the NLRP3 Inflammatory Pathway. Virology. 2022 Mar;568:13–22.
- Guarnieri JW, Alessia Angelin, Murdock DG, Schaefer P, Prasanth Portluri, Lie T, et al. SARS-COV-2 Viroporins Activate the NLRP3-inflammasome by the Mitochondrial Permeability Transition Pore. Frontiers in Immunology. 2023 Feb 20;14.
- Pan P, Shen M, Yu Z, Ge W, Chen K, Tian M, et al. SARS-CoV-2 N Protein Promotes NLRP3 Inflammasome Activation to Induce Hyperinflammation. Nature Communications. 2021 Aug 2;12(1).
- Sun X, Liu Y, Huang Z, Xu W, Hu W, Yi L, et al. SARS-CoV-2 non-structural Protein 6 Triggers NLRP3-dependent Pyroptosis by Targeting ATP6AP1. Cell Death & Differentiation. 2022 Jan 8;29(6):1240–54.
- Theobald S, Simonis A, Georgomanolis T, Christoph Kreer, Zehner M, Eisfeld HS, et al. Long-lived Macrophage Reprogramming Drives Spike Protein-mediated Inflammasome Activation in COVID-19. 2021 Aug 9;13(8).
- Eisfeld HS, Simonis A, Winter S, Chhen J, Ströh LJ, Krey T, et al. Viral Glycoproteins Induce NLRP3 Inflammasome Activation and Pyroptosis in Macrophages. Viruses. 2021 Oct 15;13(10):2076.
- Planès R, Pinilla M, Santoni K, Hessel A, Passemar C, Lay K, et al. Human NLRP1 Is a Sensor of Pathogenic Coronavirus 3CL Proteases in Lung Epithelial Cells. Molecular Cell [Internet]. 2022 Jul 7;82(13):2385-2400.e9. Available from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC9108100/.
- Ambrożek-Latecka M, Kozlowski P, Hoser G, Bandyszewska M, Karolina Hanusek, Dominika Nowis, et al. SARS-CoV-2 and Its ORF3a, E and M Viroporins Activate Inflammasome in Human Macrophages and Induce of IL-1α in Pulmonary Epithelial and Endothelial Cells. Cell death discovery. 2024 Apr 25;10(1).
- Cerato JA, Silva, Porto BN. Breaking Bad: Inflammasome Activation by Respiratory Viruses. Biology. 2023 Jul 1;12(7):943–3.
- Sefik E, Qu R, Junqueira C, Kaffe E, Mirza H, Zhao J, et al. Inflammasome Activation in Infected Macrophages Drives COVID-19 Pathology. Nature. 2022 Apr 28.
- Song H, Liu B, Huai W, Yu Z, Wang W, Zhao J, et al. The E3 Ubiquitin Ligase TRIM31 Attenuates NLRP3 Inflammasome Activation by Promoting Proteasomal Degradation of NLRP3. Nature Communications. 2016 Dec;7(1).
- Cai B, Zhao J, Zhang Y, Liu Y, Ma C, Yi F, et al. USP5 Attenuates NLRP3 Inflammasome Activation by Promoting Autophagic Degradation of NLRP3. Autophagy [Internet]. 2021 Sep 5 [cited 2024 Nov 26];18(5):990–1004. Available from: https://pmc.ncbi.nlm.nih.gov/articles/PMC9196652/.
- Paik S, Kim JK, Silwal P, Sasakawa C, Jo EK. An Update on the Regulatory Mechanisms of NLRP3 Inflammasome Activation. Cellular & Molecular Immunology. 2021 Apr 13;18(5):1141–60.
- Yang Y, Wang H, Kouadir M, Song H, Shi F. Recent Advances in the Mechanisms of NLRP3 Inflammasome Activation and Its Inhibitors. Cell Death & Disease. 2019 Feb;10(2).
- Ji X, Song Z, He J, Guo S, Chen Y, Wang H, et al. NIMA-related Kinase 7 Amplifies NLRP3 Inflammasome pro-inflammatory Signaling in microglia/macrophages and Mice Models of Spinal Cord Injury. Experimental Cell Research. 2020 Dec 9;398(2):112418–8.
- Zhang E, Li X. The Emerging Roles of Pellino Family in Pattern Recognition Receptor Signaling. Frontiers in Immunology. 2022 Feb 7;13.
- Kim, SK. The Mechanism of the NLRP3 Inflammasome Activation and Pathogenic Implication in the Pathogenesis of Gout. Journal of Rheumatic Diseases. 2022 Jul 1;29(3):140–53.
- Pan X, Wu S, Wei W, Chen Z, Wu Y, Gong K. Structural and Functional Basis of JAMM Deubiquitinating Enzymes in Disease. Biomolecules [Internet]. 2022 Jun 29 [cited 2025 Mar 4];12(7):910. Available from: https://pmc.ncbi.nlm.nih.gov/articles/PMC9313428/.
- Palazón-Riquelme P, Worboys JD, Green J, Valera A, Martín-Sánchez F, Pellegrini C, et al. USP7 and USP47 Deubiquitinases Regulate NLRP3 Inflammasome Activation. EMBO Reports. 2018 Sep 11;19(10).
- Martens A, van Loo G. A20 at the Crossroads of Cell Death, Inflammation, and Autoimmunity. Cold Spring Harbor Perspectives in Biology. 2019 Aug 19;12(1):a036418.
- Song N, Li T. Regulation of NLRP3 Inflammasome by Phosphorylation. Frontiers in Immunology. 2018 Oct 8;9.
- Zhang Z, Meszaros G, He W, Xu Y, de Fatima Magliarelli H, Mailly L, et al. Protein Kinase D at the Golgi Controls NLRP3 Inflammasome Activation. The Journal of Experimental Medicine [Internet]. 2017 Sep 4 [cited 2021 Dec 6];214(9):2671–93. Available from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5584123/.
- Song N, Liu ZS, Xue W, Bai ZF, Wang QY, Dai J, et al. NLRP3 Phosphorylation Is an Essential Priming Event for Inflammasome Activation. Molecular Cell. 2017 Oct;68(1):185-197.e6.
- Spalinger MR, Kasper S, Gottier C, Lang S, Atrott K, Vavricka SR, et al. NLRP3 Tyrosine Phosphorylation Is Controlled by Protein Tyrosine Phosphatase PTPN22. Journal of Clinical Investigation [Internet]. 2016 Apr 4;126(5):1783–800. Available from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4855944/.
- McKee CM, Fischer FA, Bezbradica JS, Coll RC. PHOrming the inflammasome: Phosphorylation Is a Critical Switch in Inflammasome Signalling. Biochemical Society Transactions. 2021 Dec 2;49(6):2495–507.
- Guarda G, Braun M, Staehli F, Tardivel A, Mattmann C, Förster I, et al. Type I Interferon Inhibits Interleukin-1 Production and Inflammasome Activation. Immunity. 2011 Feb;34(2):213–23.
- Masters SL, Mielke LA, Cornish AL, Sutton CE, O’Donnell J, Cengia LH, et al. Regulation of interleukin-1beta by interferon-gamma Is Species specific, Limited by Suppressor of Cytokine Signalling 1 and Influences interleukin-17 Production. EMBO Reports [Internet]. 2010 Aug 1;11(8):640–6. Available from: https://pubmed.ncbi.nlm.nih. 2059.
- Lee JS, Shin EC. The Type I Interferon Response in COVID-19: Implications for Treatment. Nature Reviews Immunology. 2020.
- Wang N, Zhan Y, Zhu L, Hou Z, Liu F, Song P, et al. Retrospective Multicenter Cohort Study Shows Early Interferon Therapy Is Associated with Favorable Clinical Responses in COVID-19 Patients. Cell Host & Microbe. 2020 Sep;28(3):455-464.e2.
- Vora SM, Lieberman J, Wu H. Inflammasome Activation at the Crux of Severe COVID-19. Nature Reviews Immunology. 2021 Aug 9;21(11):694–703.
- Tate MD, Ong JDH, Dowling JK, McAuley JL, Robertson AB, Latz E, et al. Reassessing the Role of the NLRP3 Inflammasome during Pathogenic Influenza a Virus Infection via Temporal Inhibition. Scientific Reports. 2016 Jun 10;6(1).
- Sho Nakakubo, Unoki Y, Kitajima K, Terada M, Hiroyuki Gatanaga, Norio Ohmagari, et al. Serum Lactate Dehydrogenase Level One Week after Admission Is the Strongest Predictor of Prognosis of COVID-19: a Large Observational Study Using the COVID-19 Registry Japan. Viruses. 2023 Mar 2;15(3):671–1.
- Vanyo Mitev, Marinov K, Rumen Tiholov, Konstantin Tashkov, Radoslav Bilyukov, Lilov AI, et al. High Colchicine Doses Are More Effective in COVID-19 Outpatients than Nirmatrelvir/Ritonavir, Remdesivir, and Molnupiravir. 2024.
- Smart SJ, Polachek SW. COVID-19 Vaccine and risk-taking. Journal of risk and uncertainty. 2024 Feb 1;68(1):25–49.
- Kelleni, MT. SARS CoV-2 Viral Load Might Not Be the Right Predictor of COVID-19 Mortality. Journal of Infection. 2020. [Google Scholar]
- Mondeshki T, Bilyukov R, Tomov T, Mihaylov M, Mitev V. Complete, Rapid Resolution of Severe Bilateral Pneumonia and Acute Respiratory Distress Syndrome in a COVID-19 Patient: Role for a Unique Therapeutic Combination of Inhalations with Bromhexine, Higher Doses of Colchicine, and Hymecromone. Cureus [Internet]. 2022 [cited 2024 Mar 21];14(10):e30269. Available from: https://www.ncbi.nlm.nih. 9653.
- Lilov A, Palaveev K, Mitev V. High Doses of Colchicine Act as “Silver Bullets” against Severe COVID-19. Cureus. 2024.
- Tsanko Mondeshki, Vanyo Mitev. High-Dose Colchicine: Key Factor in the Treatment of Morbidly Obese COVID-19 Patients. Cureus. 2024.
- Mitev, V. High colchicine doses are really silver bullets against COVID-19. Acta Medica Bulgarica. 2024, 51(4): 95-96.
- Dimitar Bulanov, Atanas Yonkov, Arabadzhieva E, Vanyo Mitev. Successful Treatment with High-Dose Colchicine of a 101-Year-Old Patient Diagnosed with COVID-19 after an Emergency Cholecystectomy. Cureus. 2024.
- Mondeshki T, Bilyukov R, Mitev V. Effect of an Accidental Colchicine Overdose in a COVID-19 Inpatient with Bilateral Pneumonia and Pericardial Effusion. Cureus. 2023.
- Vanyo Mitev, Mondeshki Tsanko, Marinov K, Radoslav Bilyukov. Colchicine, Bromhexine, and Hymecromone as Part of COVID-19 Treatment-Cold, Warm, Hot. 2023 Jan 30 [cited 2025 Apr 8];106–14. Available from. [CrossRef]
- Rumen Tiholov, Lilov AI, Gergana Georgieva, Palaveev KR, Konstantin Tashkov, Vanyo Mitev. Effect of Increasing Doses of Colchicine on the Treatment of 333 COVID-19 Inpatients. Immunity Inflammation and Disease. 2024 ;12(5). 1 May.
- Vanyo Mitev. What Is the Lowest Lethal Dose of colchicine? Biotechnology & Biotechnological Equipment. 2023 Nov 25;37(1).
- Vanyo Mitev, Tsanko Mondeshki, Ani Miteva, Konstantin Tashkov, Dimitrova V. COVID-19 Prophylactic Effect of Bromhexine Hydrochloride. Preprints 2024. 2024.
- Mitev, V. Prevention and Treatment of COVID-19 and Influenza with Bromhexine and Colchicine. 2025. XXXth Anniversary Medicine and Football International Scientific Conference, 15 april 2025, Sofia, Bulgaria; oral presentation.
- Mitev, V. Prevention and Treatment of COVID-19 and Influenza with Bromhexine and Colchicine. Medizina I Sport. 2025;(1-2):38–9.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



