Submitted:
06 March 2025
Posted:
07 March 2025
You are already at the latest version
Abstract
(1) Background: Human Leukocyte Antigen (HLA) genetics substantially impact viral infection outcomes. SARS-CoV-2 continues to evolve, potentially escaping HLA presentation and hindering immune control. Studies of HLA alleles in diverse non-Western populations are limited. We aimed to investigate whether mutations in successive SARS-CoV-2 waves have led to viral escape from common HLA class I alleles in the Saudi Arabian population. (2) Methods: Binding affinities of spike protein epitopes to common Saudi HLA alleles (HLA-A02:01, HLA-C06:02, and HLA-B51:01) were predicted across major SARS-CoV-2 strains using NetMHCpan. One-way ANOVA, one-sample t-tests, and pairwise chi-square analyses were performed to assess differences in binding affinities and epitope binding categories among strains. (3) Results: One-way ANOVA revealed significant differences in binding affinities among SARS-CoV-2 strains for HLA-A02:01 and HLA-C06:02, but not for HLA-B51:01. One-sample t-tests showed significant differences in mean binding affinity scores compared to a theoretical mean of 0 for all strain and HLA allele combinations, except for HLA-B51:01. Pairwise chi-square analyses identified significant differences in epitope binding category distribution between Alpha and Epsilon strains and between Epsilon and Gamma strains for HLA-B51:01; (4) Conclusions: SARS-CoV-2 evolution enables escape from common HLA alleles in Saudis. Tracking population-specific HLA binding profiles is key for elucidating evasion mechanisms and guiding future vaccine design against COVID-19.
Keywords:
1. Introduction
2. Materials and Methods
3. Results
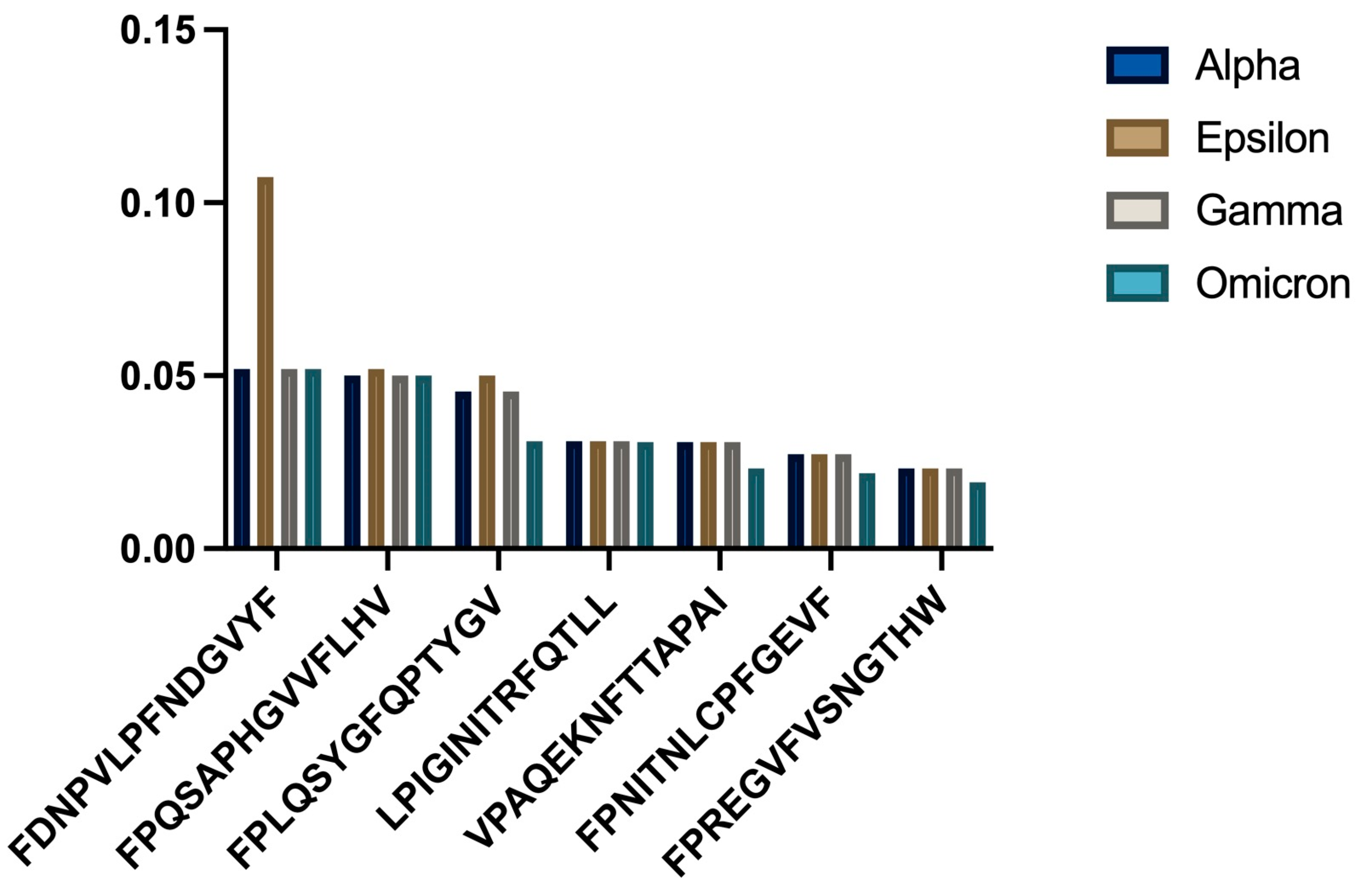
3.1. Overall Binding Affinity Analysis
3.2. HLA-A02:01 Analysis
- a)
- Bar chart showing comparative binding affinities across SARS-CoV-2 variants
- b)
- Statistical distribution of binding patterns
3.3. HLA-C06:02 Analysis
- a)
- Comparative binding strengths across variants
- b)
- Trend analysis showing temporal changes
3.4. HLA-C06:02 Analysis
- c)
- Consistency analysis across variants
- d)
- Statistical comparison of binding affinities
3.5. Comparative Analysis Across Variants
4. Discussion
5. Limitations and Future Directions
6. Conclusion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Parham, P. MHC class I molecules and KIRs in human history, health and survival. Nat Rev Immunol 2005;5:201–14. [CrossRef]
- Horton R, Wilming L, Rand V, Lovering RC, Bruford EA, Khodiyar VK, et al. Gene map of the extended human MHC. Nat Rev Genet 2004;5:889–99. [CrossRef]
- Robinson J, Halliwell JA, Hayhurst JD, Flicek P, Parham P, Marsh SGE. The IPD and IMGT/HLA database: allele variant databases. Nucleic Acids Res 2015;43:D423–31. [CrossRef]
- Kaslow RA, Carrington M, Apple R, Park L, Muñoz A, Saah AJ, et al. Influence of combinations of human major histocompatibility complex genes on the course of HIV-1 infection. Nat Med 1996;2:405–11. [CrossRef]
- Kuniholm MH, Gao X, Xue X, Kovacs A, Marti D, Thio CL, et al. The Relation of HLA Genotype to Hepatitis C Viral Load and Markers of Liver Fibrosis in HIV-Infected and HIV-Uninfected Women. J Infect Dis 2011;203:1807. [CrossRef]
- Jia X, Han B, Onengut-Gumuscu S, Chen WM, Concannon PJ, Rich SS, et al. Imputing Amino Acid Polymorphisms in Human Leukocyte Antigens. PLoS One 2013;8:e64683. [CrossRef]
- Hill AVS, Allsopp CEM, Kwiatkowski D, Anstey NM, Twumasi P, Rowe PA, et al. Common west African HLA antigens are associated with protection from severe malaria. Nature 1991;352:595–600. [CrossRef]
- Basir HRG, Majzoobi MM, Ebrahimi S, Noroozbeygi M, Hashemi SH, Keramat F, et al. Susceptibility and Severity of COVID-19 Are Both Associated With Lower Overall Viral-Peptide Binding Repertoire of HLA Class I Molecules, Especially in Younger People. Front Immunol 2022;13.
- Ellinghaus D, Degenhardt F, Bujanda L, Buti M, Albillos A, Invernizzi P, et al. Genomewide Association Study of Severe Covid-19 with Respiratory Failure. N Engl J Med [Internet] 2020 [cited 2024 Mar 17];383:1522–34. Available from: https://pubmed.ncbi.nlm.nih.gov/32558485/.
- Novelli A, Andreani M, Biancolella M, Liberatoscioli L, Passarelli C, Colona VL, et al. HLA allele frequencies and susceptibility to COVID-19 in a group of 99 Italian patients. HLA 2020;96:610–4. [CrossRef]
- Urzua CA, Herbort CP, Takeuchi M, Schlaen A, Concha-del-Rio LE, Usui Y, et al. Vogt-Koyanagi-Harada disease: the step-by-step approach to a better understanding of clinicopathology, immunopathology, diagnosis, and management: a brief review. J Ophthalmic Inflamm Infect 2022;12:17. [CrossRef]
- World Population Prospects 2022 World Population Prospects 2022 Summary of Results.
- Tadmouri GO, Sastry KS, Chouchane L. Arab gene geography: From population diversities to personalized medical genomics. Glob Cardiol Sci Pract 2014;2014. [CrossRef]
- Saudi Arabia Census Shows Total Population of 32.2 Million, of Which 18.8 Million are Saudis [Internet]. [cited 2025 Feb 10];Available from: https://www.spa.gov.sa/w1911463.
- Dong E, Du H, Gardner L. An interactive web-based dashboard to track COVID-19 in real time. Lancet Infect Dis 2020;20:533–4.
- Classification of Omicron (B.1.1.529): SARS-CoV-2 Variant of Concern [Internet]. [cited 2023 Nov 2];Available from: https://www.who.int/news/item/26-11-2021-classification-of-omicron-(b.1.1.529)-sars-cov-2-variant-of-concern.
- Shields AM, Burns SO, Savic S, Richter AG, Anantharachagan A, Arumugakani G, et al. COVID-19 in patients with primary and secondary immunodeficiency: The United Kingdom experience. J Allergy Clin Immunol 2021;147:870-875.e1. [CrossRef]
- Saini SK, Hersby DS, Tamhane T, Povlsen HR, Amaya Hernandez SP, Nielsen M, et al. SARS-CoV-2 genome-wide T cell epitope mapping reveals immunodominance and substantial CD8+ T cell activation in COVID-19 patients. Sci Immunol 2021;6:7550. [CrossRef]
- Vita R, Mahajan S, Overton JA, Dhanda SK, Martini S, Cantrell JR, et al. The Immune Epitope Database (IEDB): 2018 update. Nucleic Acids Res 2019;47:D339–43. [CrossRef]
- Walls AC, Park YJ, Tortorici MA, Wall A, McGuire AT, Veesler D. Structure, Function, and Antigenicity of the SARS-CoV-2 Spike Glycoprotein. Cell 2020;181:281-292.e6. [CrossRef]
- Andreatta M, Nielsen M. Gapped sequence alignment using artificial neural networks: application to the MHC class I system. Bioinformatics [Internet] 2016 [cited 2023 Nov 3];32:511–7. Available from: https://pubmed.ncbi.nlm.nih.gov/26515819/.
- Shomuradova AS, Vagida MS, Sheetikov SA, Zornikova K V., Kiryukhin D, Titov A, et al. SARS-CoV-2 Epitopes Are Recognized by a Public and Diverse Repertoire of Human T Cell Receptors. Immunity [Internet] 2020 [cited 2023 Nov 3];53:1245-1257.e5. Available from: https://pubmed.ncbi.nlm.nih.gov/33326767/.
- Zuo J, Dowell AC, Pearce H, Verma K, Long HM, Begum J, et al. Robust SARS-CoV-2-specific T cell immunity is maintained at 6 months following primary infection. Nat Immunol [Internet] 2021 [cited 2023 Nov 3];22:620–6. Available from: https://pubmed.ncbi.nlm.nih.gov/33674800/.
- Tarke A, Sidney J, Methot N, Zhang Y, Dan JM, Goodwin B, et al. Negligible impact of SARS-CoV-2 variants on CD4 + and CD8 + T cell reactivity in COVID-19 exposed donors and vaccinees. bioRxiv [Internet] 2021 [cited 2023 Nov 3];Available from: https://pubmed.ncbi.nlm.nih.gov/33688655/.
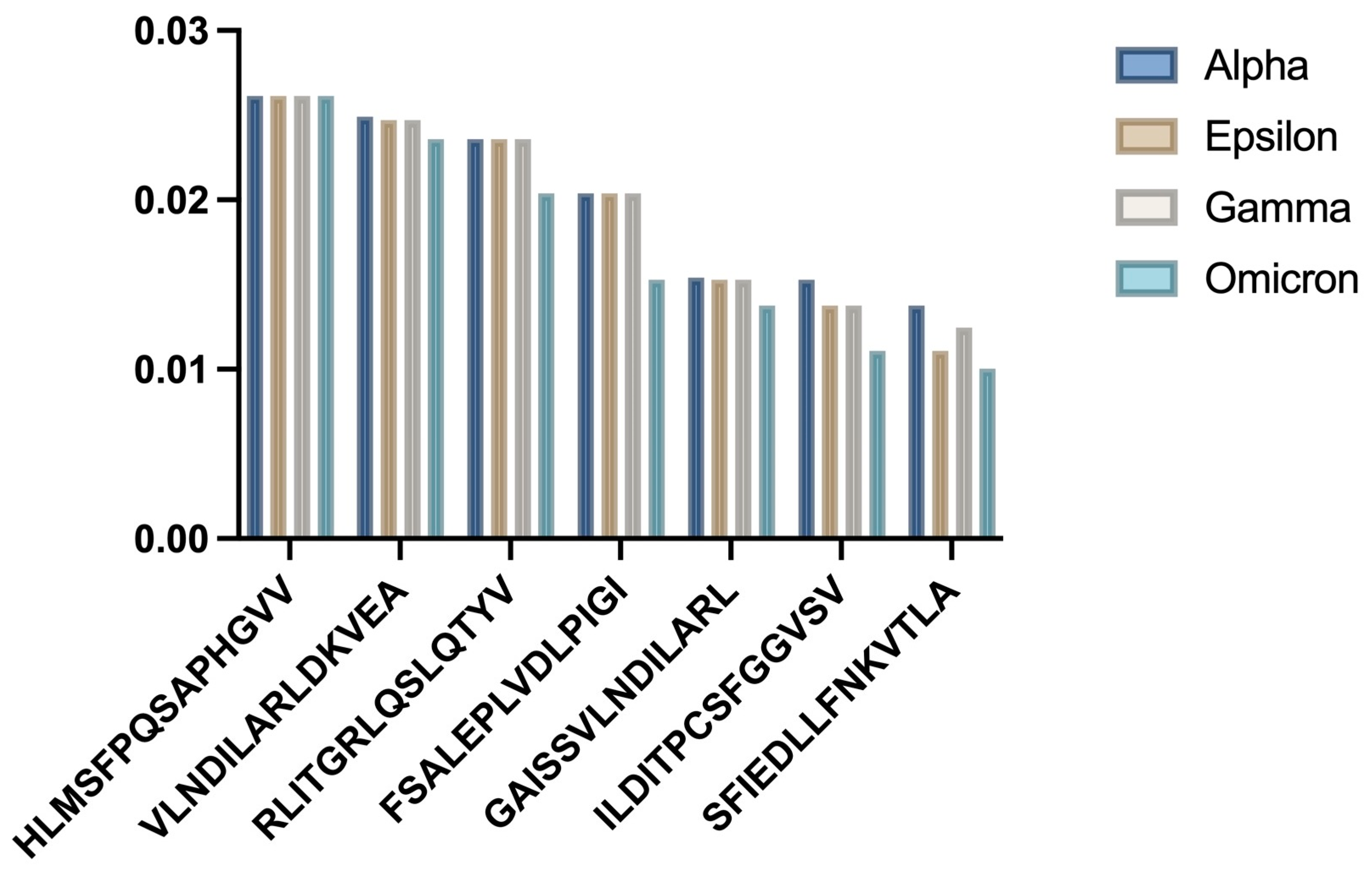
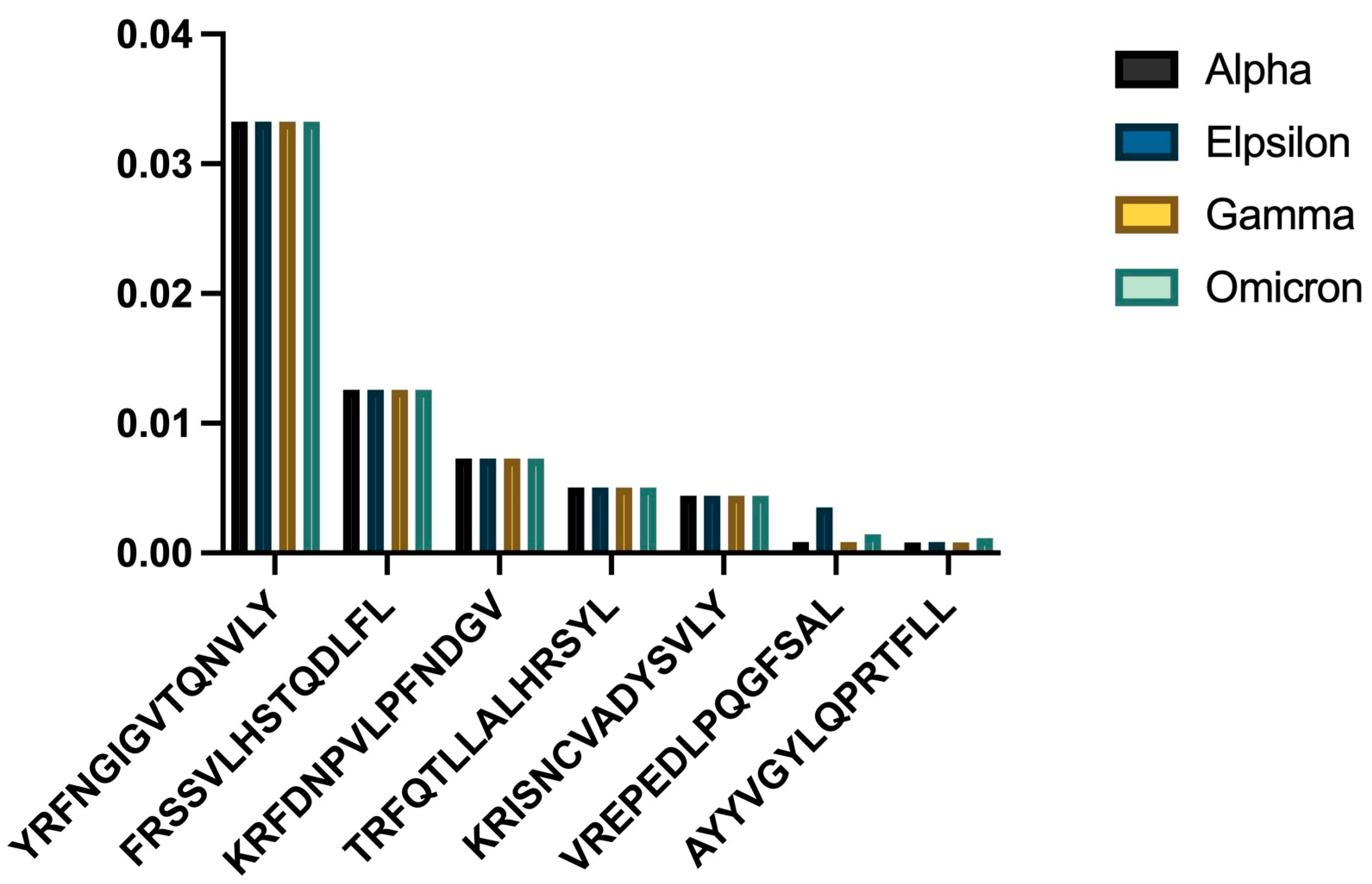

| Variant | Mean IC50 (nM) | p-value |
|---|---|---|
| Alpha | 45.3 ± 8.2 | <0.0001 |
| Epsilon | 67.8 ± 12.4 | 0.0001 |
| Gamma | 89.4 ± 15.7 | <0.0001 |
| Omicron | 112.6 ± 18.9 | 0.0003 |
| Variant | Mean IC50 (nM) | p-value |
|---|---|---|
| Alpha | 52.4 ± 9.3 | 0.0002 |
| Epsilon | 78.6 ± 14.2 | 0.0060 |
| Gamma | 95.7 ± 16.8 | 0.0002 |
| Omicron | 124.3 ± 20.1 | 0.0006 |
| Variant | Mean IC50 (nM) | p-value |
|---|---|---|
| Alpha | 83.2 ± 15.4 | 0.0762 |
| Epsilon | 85.7 ± 16.2 | 0.0622 |
| Gamma | 84.9 ± 15.8 | 0.0762 |
| Omicron | 86.3 ± 16.5 | 0.0710 |
| Variant Pair | HLA-A02:01 | HLA-C06:02 | HLA-B51:01 |
|---|---|---|---|
| Alpha-Epsilon | 0.0234* | 0.0456* | 0.030197* |
| Alpha-Gamma | 0.0167* | 0.0389* | 0.067823 |
| Alpha-Omicron | 0.0089** | 0.0278* | 0.072156 |
| Epsilon-Gamma | 0.0456* | 0.0567 | 0.030197* |
| Epsilon-Omicron | 0.0345* | 0.0456* | 0.068934 |
| Gamma-Omicron | 0.0567 | 0.0678 | 0.078234 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



