Submitted:
03 February 2025
Posted:
04 February 2025
You are already at the latest version
Abstract
Glioblastoma multiforme (GBM) is the most common brain tumour and one of the most aggressive, with a median overall survival (OS) of only 15-18 months. These characteristics make it necessary to identify new targets for the improvement of prognosis and better prediction of response to therapies currently available for GBM patients. One possible candidate target could be the evaluation of miRNAs. miRNAs are small non-coding RNAs that play important roles in post-transcriptional gene regulation. Due to their functions, miRNAs also control biological processes underlying the development of GBM and may be considered as possible targets with a clinical role. This narrative review aims to introduce the concept of miRNAs in GBM from a clinical and a molecular perspective, and then addresses the specific miRNAs that are most described in the literature as relevant for the development, the prognosis and the response to therapies for patients affected by GBM.
Keywords:
GBM
; miRNAs
; prognosis
; survival
; treatment
; TMZ
‡ These authors share co-last authorship.
1. Introduction
Each year, approximately 85,000 people in the United States are diagnosed with a primary brain tumor, with about 29% of these being malignant [1]. In adults, 80% to 85% of these are gliomas. The fifth edition of the WHO Classification of Central Nervous System (CNS) Tumors identifies these entities as “adult-type diffuse gliomas” [2]. Among this group, glioblastoma (GBM) stands out as the most common malignant primary brain tumor in adults and it is the leading cause of death in patients with primary brain tumors. GBM typically appears after the age of 40, peaking in incidence among those aged 75 to 84 years [3]. Survival rates for GBM decrease with age: only 5% of all diagnosed individuals survive for five years, and this figure falls to 2% for patients aged 65 years and older [4]. Despite advancements in understanding the biology of these tumors, significant improvements in treatment and patient outcomes are still needed.
2. General Molecular Features
Fewer than 5% of adults diagnosed with malignant brain tumors report a family history of brain tumors or a predisposition syndrome for malignancies. Around 5% of all gliomas are considered to be familial, with several rare Mendelian inherited syndromes associated with GBM and other gliomas [5]. Interestingly, the presence of germline variants appears to be higher than anticipated, with up to 13% of glioma patients carrying at least one harmful or potentially harmful genetic mutation in their germline [6].
The causes of most glioblastomas remain largely unidentified. While gliomas have been experimentally induced in rats using chemical carcinogens like ethylnitrosourea and methylnitrosourea, there is no strong evidence that these chemicals contribute to glioma development in humans. Similarly, although genome sequences and proteins from polyomaviruses (e.g., SV40, BK virus, and JC virus) have been detected in gliomas, their presence was infrequent in a more recent large series. A small percentage of glioblastomas occur in multiple members of the same family or are linked to inherited genetic tumor syndromes, such as Lynch syndrome, constitutional mismatch repair deficiency syndrome, Li–Fraumeni syndrome, and neurofibromatosis type 1. Genome-wide association studies have uncovered genetic variants in genes like TERT, EGFR, CCDC26, CDKN2B, PHLDB1, TP53, and RTEL1 that are associated with an elevated risk of gliomas. Additionally, specific single nucleotide polymorphisms (SNPs) have been identified as increasing glioma risk, distinct from those linked to other types of brain tumors.
The rising incidence of glioblastoma suggests a potential role for environmental factors, but despite extensive research, most environmental exposures, including non-ionizing radiation (e.g., from mobile phones) and occupational hazards, have not been conclusively linked to the disease. The only confirmed environmental risk factor is exposure to ionizing radiation targeting the head and neck. For instance, patients treated for acute lymphoblastic leukemia or atomic bomb survivors are at greater risk of glioblastoma, though no association has been found with diagnostic radiation. Interestingly, individuals with a history of allergies or atopic conditions appear to have a reduced risk of developing the disease [2].
3. Glioblastoma Management
The current management of glioblastoma multiforme (GBM), one of the most aggressive and lethal brain tumors, necessarily involves a comprehensive, multidisciplinary approach. Despite 20 years have been passed from its first publications, Stupp Protocol, which foresees a combined chemo-radiation therapy, remains the gold-standard treatment [7]. Initially, the treatment strategy centers on maximal safe surgical resection, aiming to remove as much of the tumor as possible while preserving neurological function. This surgery helps to reduce the tumor burden and alleviate symptoms.
Following surgery, patients typically undergo radiotherapy combined with chemotherapy using temozolomide. Radiotherapy targets residual cancer cells, while temozolomide, an oral alkylating agent, enhances the cytotoxic effects of radiation and has shown to improve overall survival rates [8,9].
Despite aggressive treatment, glioblastomas are notorious for their high recurrence rates and, consequently, for the severe and short course. As a consequence, research is increasingly focused on novel therapies and combination treatments. Tumor-treating fields (TTF), a non-invasive treatment that uses alternating electric fields to disrupt cancer cell division, has emerged as a promising adjunct to standard therapy, extending survival in some patients [10].
Moreover, advancements in molecular and genetic profiling of glioblastomas are paving the way for personalized medicine [11]. Identifying specific genetic mutations and molecular markers allows for tailored treatment strategies, potentially improving efficacy and reducing side effects. Immunotherapies, including immune checkpoint inhibitors and CAR-T cell therapy, are under investigation, aiming to harness the body's immune system to recognize and destroy cancer cells [12].
Clinical trials continue to explore the potential of targeted therapies, such as inhibitors of specific signaling pathways involved in glioblastoma progression. Bevacizumab, an anti-angiogenic agent, has been used to manage symptoms and prolong progression-free survival, although its impact on overall survival remains unclear [13].
Despite these advancements, glioblastoma management remains challenging, with a median survival time of approximately 15-18 months post-diagnosis. The aggressive nature of the tumor, its location within the brain, and its genetic heterogeneity contribute to its poor prognosis. Continuous research and clinical trials are crucial in the ongoing effort to discover more effective treatments and improve patient outcomes.
4. Clinical Relevance of the Molecular Profile
The rise in molecular profiling and the use of machine learning techniques have led to more accurate prognostic assessments and personalized treatment strategies. Discovering new alterations within GBM presents opportunities for developing targeted drugs, while correlating these genic dysregulations with specific clinical courses improves our ability to diagnose and predict disease severity.
In 2017, the establishment of the Consortium to Inform Molecular and Practical Approaches to Central Nervous System Tumor Taxonomy (cIMPACT-NOW) was announced. This consortium aims to evaluate and suggest updates to the WHO classification of brain tumors, incorporating the latest molecular insights and their practical implications for clinical practice [14].
The updated WHO criteria and nomenclature introduced in 2021 have placed a stronger emphasis on the role of molecular genetics in diagnosing GBM:
- Astrocytoma, IDH-mutant: previously, IDH-mutant tumors were classified as diffuse astrocytoma, anaplastic astrocytoma, or GBM. The latest classification now consolidates these into a single type of IDH-mutant astrocytoma, graded as 2, 3, or 4.
- Grading criteria: the grading of IDH-mutant diffuse astrocytic tumors is no longer based solely on histology. It also considers the presence of the CDKN2A/B homozygous deletion mutation, which results in a CNS WHO grade of 4, even if microvascular proliferation or necrosis is absent.
- GBM, IDH-wildtype: this classification identifies specific molecular markers for this tumor, such as the presence of a TERT promoter mutation (associated with increased telomerase activity, crucial for tumor cell immortalization) or EGFR gene amplification, leading to overexpression of the receptor, and the combined gain of chromosome 7 and loss of chromosome 10 (+7/−10). If any of these markers are found in an IDH-wildtype diffuse astrocytic glioma in adults, the diagnosis should be GBM, IDH-wildtype.
- Pediatric patients: the diagnostic criteria for IDH-wildtype diffuse astrocytomas differ in pediatric patients, who are diagnosed using different categories of pediatric-type gliomas [2].
IDH-wildtype GBMs are defined by the absence of mutations in IDH1 codon 132 and IDH2 codon 172, as well as the absence of mutations in H3 p.K28X (formerly p.K27X) or H3 p.G35X (formerly p.G34X) [15]. For patients aged 55 years or older at diagnosis, with a histologically classic GBM that is not located in midline structures and has no history of pre-existing lower-grade glioma, the lack of immunoreactivity for IDH1 p.R132H is sufficient to diagnose IDH-wildtype GBM [16]. In such cases, further DNA sequencing is unnecessary, as the probability of non-canonical IDH mutations is very low (less than 1%) in GBMs of patients aged 55 years or older [17].
However, for patients younger than 55 years, or those with a history of lower-grade glioma and/or tumors showing immunohistochemical loss of nuclear ATRX expression, negative IDH1 p.R132H immunostaining should be followed by DNA sequencing to identify less common IDH1 or IDH2 mutations. If no IDH mutations are found through sequencing, these tumors are classified as GBM, IDH-wildtype [18].
Tumors located in midline structures should be evaluated for the H3 p.K28M mutation (formerly known as p.K27M) to exclude diffuse midline glioma, H3 p.K28X–altered. In hemispheric tumors, especially in younger patients, it is important to rule out H3 p.G34X–mutant diffuse hemispheric gliomas by using immunohistochemistry for H3.3 p.G35R (formerly p.G34R) or H3.3 p.G35V (formerly p.G34V) mutations or through sequencing of H3-3A (H3F3A) [15]. This comprehensive approach ensures accurate diagnosis and proper classification of GBMs based on the presence or absence of specific genetic mutations.
The presence of at least one of these molecular aberrations in an IDH- and H3-wildtype diffuse glioma is sufficient for the diagnosis of IDH-wildtype GBM, even in the absence of microvascular proliferation and/or necrosis [19].
In addition to these genetic alterations, DNA methylation profiling has emerged as a valuable tool for diagnosing and stratifying GBMs. A significant calibrated score for the DNA methylation profile of IDH-wildtype GBM is sufficient for diagnosis. DNA methylation profiles can further classify GBMs into molecular subgroups, with the RTK1, RTK2/classic, and mesenchymal subgroups being the most common in adult patients [20]. While the clinical relevance of methylation-based subgroups in adult patients is somewhat limited, high-grade gliomas in children and adolescents may exhibit less common DNA methylation profiles that are associated with significantly longer survival [21].
Overall, DNA methylation profiling can assist in diagnosing challenging cases and differentiating GBM from histologically similar entities, enhancing our understanding of the molecular diversity within this aggressive brain tumor [20].
Molecular profiling has enabled researchers to identify common genetic mutations and core pathways shared among sporadic glioblastomas, leading to the identification of three primary subgroups:
- Proneural Group: characterized by proneural gene expression patterns and RTK I/LGm6 DNA methylation profiles. This subgroup often shows amplifications of genes such as cyclin-dependent kinase 4 (CDK4) and platelet-derived growth factor alpha (PDGFRA). It is more prevalent among younger adults.
- Classical Group: exhibits classical gene expression patterns and classic-like RTK II DNA methylation profiles. It is marked by frequent EGFR amplifications and the loss of CDKN2A/B genes.
- Mesenchymal Group: enriched for tumors with neurofibromatosis type 1 (NF1) loss and increased infiltration by macrophages [22]. This subgroup is associated with a mesenchymal or mesenchymal-like subtype.
These subgroups are defined by specific genetic alterations and provide insights into the molecular diversity of GBMs.
These three distinct subgroups, along with mixed entities that exhibit characteristics from multiple subgroups, encompass the majority of GBMs. While molecular classification has provided valuable insights and a framework for research, its clinical utility in GBM treatment remains uncertain. None of the GBM subtypes can reliably predict responses to current therapies. Moreover, assigning a specific subtype can be challenging as tumors may display mixed subtypes simultaneously, and subtype characteristics may evolve throughout the disease course [20].
Currently, the sole predictive biomarker for treatment response to temozolomide (TMZ) is the status of MGMT-mediated DNA repair silencing. This silencing typically results from MGMT promoter methylation and loss of the second allele of chromosome 10 [23,24].
MGMT promoter methylation status is routinely assessed in IDH-wildtype GBMs because it provides crucial clinical information regarding chemotherapy response and patient survival [25]. Specifically, it helps predict how patients will respond to treatment with drugs like TMZ or TMZ plus lomustine (CCNU) [26,27]. In elderly patients, MGMT promoter methylation status can guide treatment decisions, helping determine whether chemotherapy or radiotherapy is more appropriate [28,29].
Furthermore, the presence of IDH1/2 mutations in adult diffuse gliomas is associated with prolonged patient survival. A rapid and cost-effective initial screening method for IDH mutation is mutation-specific immunohistochemistry, particularly for the common variant IDH1 p.R132H, which accounts for the vast majority of IDH mutations in GBM. A positive result in this test confirms the presence of an IDH mutation [2].
For cases where initial immunohistochemistry is negative (i.e., "antibody-negative" GBM), further testing such as targeted sequencing may be considered. However, the decision for additional testing depends on various factors, including the patient’s age. Non-canonical IDH mutations are extremely rare in older patients (over 55 years). Additionally, GBMs with microthrombi and/or clear pseudopalisading necrosis at initial diagnosis are highly unlikely to harbor an IDH mutation [30].
Genomic profiling has significantly advanced our understanding of the molecular mechanisms underlying GBM and identified potential avenues for developing targeted therapies tailored to specific patient groups or for identifying specific morhphologic patterns. . Particularly, BRAF p.V600E mutation, relatively rare in IDH-wildtype GBMs, is detectable in up to 50% of GBMs with epithelioid histology. It is highly prevalent (79%) in pleomorphic xanthoastrocytoma–like tumors, 35% of adult-type IDH-wildtype GBMs, but absent in pediatric RTK1 tumors [31].
Despite providing valuable insights into tumor biology and heterogeneity, however, gene expression profiles have not yet gained clear significance in routine clinical diagnostics for GBM. [14].
It is important to note that none of the genetic alterations mentioned are specific or sufficient for defining the respective morphological subtypes or patterns of GBMs. The molecular characterization of these tumors is complex, and individual GBMs can exhibit multiple genetic changes, especially after chemo-radiation therapies. Following initial treatment, indeed, which typically involves surgical resection, radiation therapy, and chemotherapy, distinct subgroups of tumor cells may emerge with unique characteristics. For example, approximately 10% of recurrent GBMs, occurring after treatment with TMZ, exhibit significantly higher mutation rates [32]. This phenomenon of DNA "hypermutation" is often associated with underlying genetic deficiencies in DNA mismatch repair (MMR) genes. Additionally, hypermutation can develop due to exposure to DNA alkylating agents, particularly in gliomas with MGMT methylation, including those with IDH mutations [33].
Moreover, comparisons between tumor samples obtained at diagnosis and those from recurrence show that approximately 80% of mutations and copy-number variations remain consistent between primary and recurrent tumors. Genetic events like mutations in PIK3CA, TERT alterations, and EGFR amplifications found in the primary tumor typically persist in the recurrent tumor. Conversely, events such as PDGFRA amplifications, EGFR mutations, and the presence of EGFR variant III (EGFRvIII) rearrangement are more likely to be lost. Commonly acquired genetic changes in recurrent tumors include mutations in TP53, EGFR, and phosphatase and tensin homolog (PTEN) [34].
Emerging sequencing technologies provide deeper insights into intratumoral heterogeneity and the evolution of GBMs. Single-cell transcriptomics have revealed that GBMs contain cells representing each of the three gene expression subtypes, rather than fitting neatly into a single category. This supports earlier findings from bulk gene expression profiling across multiple tumor sectors.
Moreover, sequencing circulating tumor DNA (ctDNA) present in cerebrospinal fluid (CSF) can provide a genetically accurate snapshot of the glioma genome in up to 50% of patients. This advancement may potentially obviate the need for repeat tumor biopsies in certain cases. Ongoing technological advancements may also make it feasible to evaluate plasma ctDNA in the future.
Further research is exploring novel predictive biomarkers for molecularly targeted therapies in subsets of GBM patients. Promising biomarkers include high tumor mutation burden, BRAF p.V600E mutation, NTRK or FGFR gene family fusions, and MET amplification or fusions. These markers hold potential for tailoring therapeutic approaches in specific GBM subgroups and improving treatment outcomes [35].
5. microRNAs (miRNAs) and GBM
5.1. miRNAs Function and Biogenesis
miRNAs are small non-coding molecules of RNAs of the length of 19-23 nucleotides that exerts post-transcriptional regulatory effects on many genes. Their action is based on a process of silencing of the mRNAs defined as RNA interference (RNAi). RNAi can bring to mRNA degradation or sequence-specific mRNA inhibition [35]. It has been estimated that nearly 30% of genes in the human genome are regulated by miRNAs [36,37].
miRNAs derive from primary transcripts (pri-miRNAs) which are transcribed from the genome by the RNA polymerase II. In the nucleus the pri-miRNAs are then modified, with the 5`cap and the 3` polyadenilation tail, in 60-70 nucleotides pre-miRNA by the RNAse III-Drosha complex. Subsequently, pre-miRNAs are exported in the cytoplasm through the nuclear transporter exportin 5. To conclude the process of miRNA biogenesis, in the cytoplasm pre-miRNAs are cutted and separated into two strands of 19-23 nucleotides by the Dicer protein, an endoribonuclease. These strands form a duplex where the antisense strand is incorporated in the RISC complex (developing the miRNA-induced silencing complex, miRISC) while the sense strand is degraded. The complementary attachment of miRISC complex to the specific mRNA in the 3` untranslated region of mRNA can bring to mRNA degradation, if there is a high degree of complementarity, or to inhibition of mRNA, if the complementarity is partial [38,39] (Figure 1).
5.2. miRNAs in Cancer
Since miRNAs are involved in the regulation of a variety of genes, they are also implicated in several pathological conditions, especially cancer. In the development of tumours they can act like oncogenes (oncomiR) or as tumor suppressor genes [40]. OncomiR are upregulated and promote cancer through inhibition of the genes involved in the control of cell differentiation, apoptosis and/or through the negative regulation of tumor suppressor genes. Instead the miRNAs acting like tumor suppressor genes are downregulated and they can act through regulation of the genes for cell differentiation or apoptosis or through regulation of oncogenes [40]. The evaluation of the expression of miRNAs signature can be a diagnostic tool for the definition of cancer disease stage and survival. In addition, in some cases, can help for the decision of a tailored treatment based on the specific characterization of miRNAs expression status [41]. In particular, miRNAs expression disorders in cancer affect cell proliferation, invasiveness, angiogenesis and recurrence [42,43]. Their role in tumorigenis is further demonstrated by the fact that their genes are located in general near chromosomes fragile sites which are known to be more susceptible to amplifications, to deletions, to point mutations and even to DNA methylation disorders [44,45].
5.3. miRNAs in GBM
Recent years have seen an increase in publications focusing on the function of miRNAs in GBM, demonstrating how these molecules can influence tumour development, prognosis, response to therapies and, in some cases, even the classification of this cancer [35,46,47]. Indeed, miRNAs in GBM can influence the promotion of sustained signalling of proliferation or evasion from tissues [47]. In a tumour the expression levels of miRNAs dynamically change at the various stages [47].
In this chapter we report a list and the description of some of the most relevant miRNAs involved in GBM, a summary is reported in Table 1 and Table 2. In this list, the miRNAs are subdivided between the ones that act like oncomiR (Table 1) (Figure 2) or like tumor suppressor miRNAs (Table 2) (Figure 3) in GBM development.
Abbreviations: EMT, Epithelial–mesenchymal transition; GBM, Glioblastoma; GSC, GBM stem cells; OS, Overall survival; TMZ, Temozolomide.
5.3.1. OncomiR-Upregulated miRNAs
5.3.1.1. OncomiR Involved in GBM Biogenesis
- miR-17-92 cluster
This miRNA cluster regulates, through inhibition, the tumour suppressor genes that act as cell cycle inhibitors (e.g. PTEN and CDKN1A), thus promoting GBM Stem Cells (GSCs) differentiation and blocking apoptosis (Figure 2).
The overexpression of this miRNA cluster is higher in GBM tumour samples and cell lines than in normal brain tissues. High levels of miR-17-92 cluster are associated with high aggressiveness of the tumour because the expression of this cluster associates to higher invasion and replication capability [48].
5.3.1.2. OncomiR Involved in GBM prognosis
- miR-9
MiR-9 promotes GBM tumour cell proliferation and inflammation targeting markers of the extracellular matrix and proteins involved in cell duplication and transformation including RAS and MYC [49,50] (Figure 2). MiR-9 activates oncogenic RAS which induces VEGF expression and which is able to potentiate MYC activity to promote tumour angiogenesis [49,50].
In addition, when upregulated, miR-9 downregulates the PTCH1 tumor suppressor gene resulting in the activation of Sonic HedgeHog (SHH), a protein with neuronal morphogenic activity that, when activated, increases the cell proliferation signal [49] (Figure 2). In GBM cell lines models the expression of this oncomiR brought to resistance to the standard therapy based on TMZ, this could be associated to the PTCH1-SHH pathway. Indeed, in TMZ resistant cells, PTCH1 mRNA was significantly (p < 0.05) increased but the protein was decreased, suggesting that TMZ could induce post-transcriptional regulation of PTCH1 [51]. Due to TMZ resistance in vitro, miR-9 expression has been evaluated in patients comparing short term and long term survivors and it was found that it is associated to poor survival [49,51,52].
- miR-10a and miR-10b
MiR-10a and miR-10b block cell cycle inhibitors activity (such as CDKN1A, BIM, BCL2, TEAP2C and PTEN) and HOXD10, a protein involved in differentiation [53] (Figure 2). As a consequence, these OncomiRNAs promote cell proliferation, invasion, migration, epithelial-mesenchymal transition (EMT) in addition to an effect on GSCs [53,54,55]. The expression of miR-10b has been associated to the degree of the brain tumour being higher in GBM than in other gliomas [56,57,58].
For what concerns the treatments based on this marker hypothetically the development of a drug able to block up-regulation of miR10 should restore the correct target genes expression and could reduce glioblastoma cell growth [47]. Recently also the relevance of the synergistic action between miR-10b and miR-222 has been described. These two miRNAs promote cell proliferation blocking PTEN, which activates p53. Additionally, this pair of miRNAs regulates apoptosis modulating BIM, an apoptotic factor. As a matter of fact, they have been associated with poor survival and, due to their joint effect on GBM development, in the future both these miRNAs will be considered as putative new target for the treatment of GBM [59].
- miR-148a
The main target of miR-148a is the cell adhesion molecule 1 (CADM1). The inhibition of CADM1 brings to increase of the STAT3 pathway [53,60] (Figure 2). Another target of miR-148a is the factor inhibiting HIF-1 (FIH1) that through inhibition of HIF-1, a regulator of homeostasis, is able to influence angiogenesis [61,62].
This miRNA has been included by the The Cancer Genome Atlas (TCGA) in the list of the miRNAs involved in GBM development. The first fact justifying this classification is that miR-148a has been found overexpressed in the plasma from the serum of patients with GBM if compared to healthy cases [60]. The second one is the role in GBM. In a 2011 study, Srinivasan and colleagues identified a signature of ten miRNAs whose expression is associated with GBM patient survival [61]. One of these miRNA is the miR-148a which has been found to be expressed in the high risk group (i.e. patients characterized by low survival). The relevant role of this miRNA in high risk GBMs is justified by its aforementioned biological activity of angiogenesis regulation bringing to stimulation of the invasion of the tumour [53,60,61].
- miR-182
The main targets of miR-182 are: BCL2L12, HIF2A, MET, CYLD, LRRC4 leading to uncontrolled proliferation, and stemness but, in contrast, also to apoptosis [47,63] (Figure 2).
The expression of this miRNA is associated with the molecular status of brain tumour malignancy. In particular, the cluster miR-183/96/182 including miR-182 has been reported to be upregulated in tumors with EGFR amplification [64,65,66]. To date, the influence of this miRNA on GBM survival is still debated. In many publications the expression of miR-182 correlates with better response to TMZ based chemotherapy and, as a consequence, with better survival. In particular Zhao et al found that three serum miRNAs (miR-182, 15b-5p and 145-5p) including miR-182 are significantly associated (p=0.04) with 2-years overall survival and Kouri et al reported how this miRNA is detected at high levels in patients with better survival (p=0.01) [67,68]. This could be due to the pro-apoptotic effect and to the downregulation of cancer cells migration [67,68]. On the other side, some research groups have described the effect of miR-182 on growth promotion and on migration and, consequently, have associated its expression with worse prognosis [69,70]. In particular, it has been found that the cumulative 5-year overall survival rate is lower when miR-182 expression is high than in the low miR-182 group for what concerns overall survival (OS) (p=0.003) and disease-free survival (DFS) (p=0.006) [69].
- miR-196a and miR-196b
These miRNAs target different markers associated to proliferation and to the regulation of apoptosis such as: HOXB8, HOXC8, HOXD8, HOXA7, HOXB7, ERG, HMGA2, ANXA1 [71], as a consequence, when miR-196 is overexpressed in glioma cells proliferation is induced [72] (Figure 2). A study published by Lakomy et al in 2011 demonstrated how this miRNA can influence sensitivity to TMZ [73]. Indeed, miR-196b correlated in a positive way with OS (p= 0.0492) [73]. In addition to the results of this group, the only ones published so far, our group, in 2023, obtained the same results but in a larger and consecutively enrolled cohort [74]. In particular, we found a significant correlation between GBM OS and miR-196b expression (p=0.008) where a better OS associated to miR-196 expression. In our study the same data were not statistically significant for what concerns progression-free survival (PFS) [74].
5.3.1.3. OncomiR with Effect on Therapy Efficacy
- miR-26a
MiR-26a binds to PTEN and ATM causing their inhibition and the promotion of tumour formation. In vitro (testing U87 cells, a cell line from malignant glioma) it has been reported that the overexpression of this miRNA, reducing DNA repair ability, enhances radio sensitivity to radiotherapy [75,76,77] (Figure 2). In addition, the knock down of this miRNA act in the opposite way, reducing this sensitivity [76]. From the molecular point of view the response to radiotherapy is modulated by this miRNA through the targeting of the 3`UTR region of ATM gene which leads to reduced ATM levels and to the consequent inhibition of homologous recombination repair system. In the future, these in vitro studies could be expanded on patient tissues to clarify whether miR-26 could play a role as a radiosensitiser in GBM. However, to date, in vivo results on GBM patients are not yet available.
- miR-648
miR-648 is involved in the regulation of Myelin-associated oligodendrocyte basic protein isoform 1 (MOBP) [78] (Figure 2)and has been associated with the low expression of miR-181c, miR-181d, and miR-195 [79]. miR-648 is a member of a group of miRNAs that have been found, by a bioinformatically guided experimental approach by Kreth et al, as capable to downregulate MGMT expression independently of promoter methylation by elongation of the 3’-UTR end of the MGMT mRNA [80]. In particular, this research group described how this miRNA is able to regulate MGMT at post-transcriptional level [80] and, only in cell lines, they observed that the expression by transfection of miR-648 enhanced responsivity of TMZ in MGMT expressing T98G glioma cells [80]. To the best of our knowledge, until now, few data have been obtained concerning association of miR-648 and TMZ response in samples from patients. A study published by our group in 2023 reported correlation between OS and miR-648 expression (p=0.02), the high expression of this miRNA correlates to better survival but not PFS in GBM treated with TMZ according with the Stupp scheme (60 Gray radiotherapy and concomitant chemotherapy with TMZ, followed by six cycles of maintenance with TMZ) [74].
5.3.1.4. OncomiR Involved in GBM Biogenesis that in the Future Will Have a Clinical Role Through Their Inhibition
- miR-21
MiR-21 is overexpressed in many tumours, including GBM, in which its role is to regulate the inhibition of PTEN, p53 and the activation of EGFR, Cyclin D1 and AKT2 (Figure 2). Other targets of miR-21 are SPOCK1, a proteoglycan, and transcription regulators such as RB1CC1 [35]. In addition, miR-21 enhances not only pro-proliferative action but also tumour invasion and migration targeting the factors that regulate matrix metalloproteinase (e.g. RECK and TIMP3). Historically, this miRNA has been the first to be described as an oncogenic miRNA able to give a contribute to the progression of GBM. Based on the overexpression of these miRNAs, it has been suggested the idea that, in the future, different silencing mechanisms could be implemented in order to improve the chance of treatment of GBM [48,53,55]. These different mechanisms are generally based on RNA interference and could be of two types: genome derived miRNAs and exogenous miRNAs transferred inside exosomes. However, the mechanisms of inhibition are still at the drawing stage and data have not been published so far.
5.3.2. Tumor Suppressor miRNAs-Downregulated miRNAs
5.3.2.1. Tumor Suppressor miRNAs Involved in GBM Biogenesis
- miR-1
This miRNA binds connexin-43 and can target glucose-6-phosphate dehydrogenase (G6PD) causing the inhibition of tumour cell proliferation and of tumour cells migration [53,81,82] (Figure 3). Some in vitro experiments demonstrated that miR-1 inhibition can enhance the sensitivity of GBM cells towards TMZ and, when downregulated, it increases tumorigenesis leading to cells proliferation [53,81,82]. In particular it has been described how the expression of miR-1 in GBM cell lines targets fibronectin. Subsequently, it has been observed that, when fibronectin is higly expressed in GBM, there is a poor patient survival [53]. To date, to the best of our knowledge, no additional data have been obtained in tumour tissues.
5.3.2.2. Tumor Suppressor miRNAs Involved in GBM Prognosis
- miR-128
This miRNA has several targets in the apoptosis and cell growth pathways: WNT, ERK, EGFR, IGF1R, Bcl2, PDGFRA and caspase [53,83,84] (Figure 3). When miR-128 is down-regulated, cell duplication and reduced apoptosis are favoured. MiR-128, beside the role in carcinogenesis, permits to distinguish between high and low-grade gliomas; in particular it has been reported how its low expression can be associated to high-grade glioma cell lines and, consequently, to a worst prognosis [81].
- miR-137
MiR-137 exerts its role as a tumour suppressor by inhibiting angiogenesis and through inhibition of the EZH2 protein, a proliferation factor [53,85]. When it is downregulated, glioma development is favoured because EZH2 is consequently over-expressed, and angiogenesis and proliferation are stimulated [85] (Figure 3). Sun and colleagues found out that the expression level of miR-137 was down-regulated in GBM cells, and they described how the low level of this miRNA was related to poor prognosis in GBM patients [53,85,86]. MiR-137 expression could be a marker for poor prognosis in GBM patients, but in the future, might also be a new treatment approach for GBM treatment [86]. Indeed, Sun and colleagues demonstrated how the expression of miR-137 inhibited tumor growth and angiogenesis in mouse models [86].
- miR-181 family
MiR-181 family is a group of tumor suppressor miRNAs (miR-181a, miR-181b, miR-181c, miR-181d). MiR-181a targets CD133 and BMI1, markers that are related to stemness and cell proliferation [81] (Figure 3). MiR-181b causes growth inhibition, apoptosis and invasion inhibition in glioma cells by directly targeting CCN1, an inducible growth factor that promotes the adhesion of endothelial cells [87]. The most relevant for GBM development are miR-181a and miR-181b. Indeed, they could be analyzed, in the future, in diagnostic routine for a more precise classification of high-grade gliomas [81]. The miR-181 low expression can be associated to high-grade glioma cell lines and, consequently, to a worst prognosis [81]. For what concerns miR-181c and miR-181d, which regulate in cluster the WNT pathway, a paper published by our group demonstrated how low expression of miR-181c or low expression of miR-181d in combination with expression of miR-648, predict the worst prognosis [79]. Lakomy et al observed higher levels of expression of this miRNA in the group of patients with time to progression shorter than 6 months (p=0.0010) [73].
5.3.2.3. Tumor Suppressor miRNAs with Effect on Therapy Efficacy
- miR-370-3p
miR-370-3p suppresses cell migration and proliferation. This function is based on regulation of WNT signalling pathway through the stabilization of β-catenin and on regulation of FOXO1, FOXM1 and TGFβ proteins [88] (Figure 3). In cell lines, it has been described that, when miR-370-3p is upregulated, GBM growth is inhibited and a longer upregulation is associated with longer survival [89]. This miRNA is not strictly adequate for the classification as high or low grade glioma because it can be found as down-regulated in both these situations [89].
5.3.2.4. Tumor Suppressor miRNAs Involved in GBM Biogenesis that in the Future Will Have a Clinical Role Through Their Inhibition
- miR-7
This miRNA regulates the expression of EGFR mRNA and of different mRNAs involved in the AKT/PI3K pathway, and, as a consequence, has an impact on cell division (Figure 3). In addition, it regulates also PKM2, which is responsible for net ATP production within the glycolytic sequence. The increase of miR-7 inhibits the glucose metabolic capacity acting on the knockdown of IGF-R, which is upstream of the AKT pathway [90].
Beside the influence on cell division, miR-7 regulates also the growth and the differentiation of GSCs [91].
The transfection of this miRNA in U373-MG GBM cell line resulted in significant suppression of EGFR mRNA and protein bringing to the inhibition of cells duplication. This permit to affirm that, in the future, after tests on tumour tissues and trials including GBM patients, the treatment with this miRNA, if found as overexpressed, will inhibit the extension of the GBM tumour [35]. Until now, no data have been reported concerning this.
- miR-34
MiR-34 regulates many genes, including Bcl2, NOTCH, NUMB (involved respectively in apoptosis and in the development of the nervous system), CDK6, EGFR and c-Met (the last one is involved in cell proliferation and invasion) [92,93,94] (Figure 3). The inhibition of these genes by miR-34 brings to cell proliferation and to a block of apoptosis causing the development of glioma.
The expression of miR-34 can be used to treat GBM: indeed, it has been demonstrated a possibility of transmission through viral vectors and extracellular vesicles [95]. In this field, good results have been obtained by Francipane and colleagues. This research group focused their attention on the use of Zika virus (ZIKV) in GBM clinic. In GSC cultures the infection by ZIKV induced miR-34 expression inhibiting the anti-apoptotic protein Bcl-2 and Numb, the antagonist of Notch, both involved in GSC invasiveness. Similar data have been obtained in mouse models where ZIKV reduced brain tumor size [96,97] and metastasis [98]. The infection by ZIKV has also the advantage to persist for months in blood after the infection lowering the risk of tumour recurrence and the need for repeated viral infusions. Another fact associated to ZIKV is the enhanced effector/memory CD4+ T cell response after the transfection suggesting that in the future this virus could be a potential adjuvant to vaccine based immunotherapies against GBM [99].
6. Conclusions
The knowledge about miRNA involvement in tumor development, including GBM, has increased dramatically over the last decade. The miRNAs described in this review are the most important examples of how these non-coding RNAs can influence the development, prognosis and efficacy of TMZ-based treatments in GBM. The information here reported shows that the incorporation of miRNA, if not all at least those for which the clinical role is quite universally accepted, into the clinical setting may provide GBM patients with a more accurate description of the treatment or prognosis of their tumour. As a consequence, the evaluation of these miRNAs could improve treatment and the definition of prognosis.
Further studies will be needed in the future to better define how these markers can be used for tumour treatment by amplification of their expression in the case of tumour suppressor miRNAs or by their inhibition in the case of OncomiR (Table 1 and Table 2). The treatment focused on miRNAs could be a new starting point to improve the survival and quality of life of a very severe neoplastic disease, GBM, possibly by designing specific miRNAs-based therapies.
Author Contributions
Conceptualization, S.E., F.M., M.F.; formal analysis S.E., P.S., I.Z., F.M, M.F.; investigation, S.E., P.S., I.Z., F.M., M.F, A.C.; data curation, S.E., P.S., I.Z., F.M., M.F, A.C.; writing—original draft preparation, S.E., P.S., I.Z., F.M., M.F., A.C.; writing—review and editing, S.E., P.S., I.Z., F.M., M.F, A.C.; supervision, F.M., M.F, A.C. All authors have read and agreed to the published version of the manuscript.”.
Funding
This research received no external funding. Check carefully that the details given are accurate and use the standard spelling of funding agency names at https://search.crossref.org/funding. Any errors may affect your future funding.
Institutional Review Board Statement
The study did not require ethical approval.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Schaff, L.R. , Mellinghoff, I.K. Glioblastoma and Other Primary Brain Malignancies in Adults: A Review. JAMA 2023, 329(7), 574-587. [CrossRef]
- Louis, D.N. , Perry, A., Wesseling, P., Brat, D.J., Cree, I.A., Figarella-Branger, D., Hawkins, C., Ng, H.K., Pfister, S.M., Reifenberger, G., Soffietti, R., et al. The 2021 WHO Classification of Tumors of the Central Nervous System: a summary. Neuro Oncol, 1: 23(8), 1231-1251. [Google Scholar] [CrossRef]
- Wen, P.Y. , Weller, M., Lee, E.Q., Alexander, B.M., Barnholtz-Sloan, J.S., Barthel, F.P., Batchelor, T.T., Bindra, R.S., Chang, S.M., Chiocca, E.A., et al. Glioblastoma in adults: a Society for Neuro-Oncology (SNO) and European Society of Neuro-Oncology (EANO) consensus review on current management and future directions. Neuro Oncol, 2020, 22(8), 1073-1113. [Google Scholar] [CrossRef]
- Ostrom, Q.T. , Cioffi, G., Waite, K., Kruchko, C., Barnholtz-Sloan, J.S. CBTRUS Statistical Report: Primary Brain and Other Central Nervous System Tumors Diagnosed in the United States in 2014-2018. Neuro Oncol, 2021: 23 (12 Suppl 2):iii1-iii105. [CrossRef]
- Vijapura, C. , Saad, Aldin. E., Capizzano, A.A., Policeni, B., Sato, Y., Moritani, T. Genetic Syndromes Associated with Central Nervous System Tumors. Radiographics. 2017, 37(1), 258–280. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, P. , Lin, A.L., Young, R.J., DiStefano, N.M., Hyman, D.M., Li, B.T., Berger, M.F., Zehir, A., Ladanyi, M., Solit, D.B. et al. Genomic Correlates of Disease Progression and Treatment Response in Prospectively Characterized Gliomas. Clin Cancer Res, 5: 25(18), 5537-5547. [Google Scholar] [CrossRef]
- Stupp, R. , Mason, W.P., van den Bent, M.J., Weller, M., Fisher, B., Taphoorn, M.J., Belanger, K., Brandes, A.A., Marosi, C., Bogdahn, U. et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N Engl J Med, 2005: 352(10):987-96. [CrossRef]
- Hanna, C. , Kurian, K.M., Williams, K., Watts, C., Jackson, A., Carruthers, R., Strathdee, K., Cruickshank, G., Dunn, L,, Erridge, S. et al. Pharmacokinetics, safety, and tolerability of olaparib and temozolomide for recurrent glioblastoma: results of the phase I OPARATIC trial. Neuro Oncol, 2020, 22(12): 1840-1850. [Google Scholar] [CrossRef]
- Hau, E. , Shen, H., Clark, C., Graham, P.H., Koh, E.S., L, McDonald, K. The evolving roles and controversies of radiotherapy in the treatment of glioblastoma. J Med Radiat Sci. 2016, 63(2), 114–23. [Google Scholar] [CrossRef] [PubMed]
- Stupp, R. , Taillibert, S., Kanner, A., Read, W., Steinberg, D., Lhermitte, B., Toms, S., Idbaih, A., Ahluwalia, M.S., Fink, K., et al. Effect of Tumor-Treating Fields Plus Maintenance Temozolomide vs Maintenance Temozolomide Alone on Survival in Patients With Glioblastoma: A Randomized Clinical Trial. JAMA. 2017, 318(23), 2306-2316; Erratum in: JAMA. 2018 May 1;319(17):1824. 10.1001/jama.2018.3431. [Google Scholar] [CrossRef]
- Brennan, C.W. , Verhaak, R.G., McKenna, A., Campos, B., Noushmehr, H., Salama, S.R., Zheng, S., Chakravarty, D., Sanborn, J.Z., Berman, S.H., et al. The somatic genomic landscape of glioblastoma. Cell. 2013, 155(2), 462–77. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.J. , Mashouf, L.A., Lim, M. CAR T Cell Therapy in Primary Brain Tumors: Current Investigations and the Future. Front Immunol, 2022, 13, 817296. [Google Scholar] [CrossRef]
- Wick, W. , Gorlia, T., Bendszus, M., Taphoorn, M., Sahm, F., Harting, I., Brandes, A.A., Taal, W,, Domont, J., Idbaih, A., et al. Lomustine and Bevacizumab in Progressive Glioblastoma. N Engl J Med. 2017, 377(20), 1954–1963. [Google Scholar] [CrossRef]
- Louis, D.N. , Aldape, K., Brat, D.J., Capper, D., Ellison, D.W., Hawkins, C., Paulus, W., Perry, A., Reifenberger, G, et al. Announcing cIMPACT-NOW: the Consortium to Inform Molecular and Practical Approaches to CNS Tumor Taxonomy. Acta Neuropathol 2017, 133(1), 1-3. [CrossRef]
- Leske, H. , Dalgleish, R., Lazar, A.J., Reifenberger, G., Cree, I.A. A common classification framework for histone sequence alterations in tumours: an expert consensus proposal. J Pathol 2021, 254(2), 109-120. [CrossRef]
- Louis, D.N. , Perry, A., Reifenberger, G., von Deimling, A., Figarella-Branger, D., Cavenee, W.K., Ohgaki, H., Wiestler, O.D., Kleihues, P., Ellison, D.W. The 2016 World Health Organization Classification of Tumors of the Central Nervous System: a summary. Acta Neuropathol 2016, 131(6), 803-20. [CrossRef]
- Chen, L. , Voronovich, Z., Clark, K., Hands, I., Mannas, J., Walsh, M., Nikiforova, M.N., Durbin, E.B., Weiss, H., Horbinski, C. Predicting the likelihood of an isocitrate dehydrogenase 1 or 2 mutation in diagnoses of infiltrative glioma. Neuro Oncol. 2014, 16(11), 1478–83. [Google Scholar] [CrossRef]
- Capper, D. , Jones, D.T.W., Sill, M., Hovestadt, V., Schrimpf, D., Sturm, D., Koelsche, C., Sahm, F., Chavez, L., Reuss, D.E., et al. DNA methylation-based classification of central nervous system tumours. Nature, 2018: 22, 555(7697): 469-474. [CrossRef]
- Louis, D.N. , Wesseling, P., Aldape, K., Brat, D.J., Capper, D., Cree, I.A., Eberhart, C., Figarella-Branger, D., Fouladi, M., Fuller, G,N., et al. cIMPACT-NOW update 6: new entity and diagnostic principle recommendations of the cIMPACT-Utrecht meeting on future CNS tumor classification and grading. Brain Pathol 2020, 30(4), 844-856. [CrossRef]
- Capper, D. , Stichel, D., Sahm, F., Jones, D.T.W., Schrimpf, D., Sill, M., Schmid, S., Hovestadt, V., Reuss, D.E., Koelsche, C., et al. Practical implementation of DNA methylation and copy-number-based CNS tumor diagnostics: the Heidelberg experience. Acta Neuropathol 2018, 136(2), 181-210. [CrossRef]
- Korshunov, A. , Schrimpf, D., Ryzhova, M., Sturm, D., Chavez, L., Hovestadt, V., Sharma, T., Habel, A., Burford, A., Jones, C., et al. H3-/IDH-wild type pediatric glioblastoma is comprised of molecularly and prognostically distinct subtypes with associated oncogenic drivers. Acta Neuropathol 2017, 134(3): 507-516. [CrossRef]
- Wang, Q. , Hu, B., Hu, X., Kim, H., Squatrito, M., Scarpace, L., deCarvalho, A.C., Lyu, S., Li, P., Li, Y., et al. Tumor Evolution of Glioma-Intrinsic Gene Expression Subtypes Associates with Immunological Changes in the Microenvironment. Cancer Cell 2017, 32(1), 42-56.e6, Erratum in: Cancer Cell. 2018 Jan 8;33(1):152. [CrossRef]
- Stupp, R. , Hegi, M.E., Mason, W.P., van den Bent, M.J., Taphoorn, M.J., Janzer, R.C., Ludwin, S.K., Allgeier, A., Fisher, B., Belanger, K., et al. Effects of radiotherapy with concomitant and adjuvant temozolomide versus radiotherapy alone on survival in glioblastoma in a randomised phase III study: 5-year analysis of the EORTC-NCIC trial. Lancet Oncol 2009, 10(5), 459-66. [CrossRef]
- Reifenberger, G. , Weber, R.G., Riehmer, V., Kaulich, K., Willscher, E., Wirth, H., Gietzelt, J., Hentschel, B., Westphal, M., Simon, M., Schackert, G., et al. Molecular characterization of long-term survivors of glioblastoma using genome- and transcriptome-wide profiling. Int J Cancer, 2014, 135(8), 1822-31. [Google Scholar] [CrossRef]
- Weller, M. , Felsberg, J., Hartmann, C., Berger, H., Steinbach, J.P., Schramm, J., Westphal, M., Schackert, G., Simon, M., Tonn, J.C., et al. Molecular predictors of progression-free and overall survival in patients with newly diagnosed glioblastoma: a prospective translational study of the German Glioma Network. J Clin Oncol. 2009, 27(34), 5743-50. [Google Scholar] [CrossRef]
- Hegi, M.E. , Diserens, A.C., Gorlia, T., Hamou, M.F., de Tribolet, N., Weller, M., Kros, J.M., Hainfellner, J.A., Mason, W., Mariani, L., et al. MGMT gene silencing and benefit from temozolomide in glioblastoma. N Engl J Med, 2005, 352(10), 997-1003. [Google Scholar] [CrossRef]
- Weller, M. , Tabatabai, G., Kästner, B., Felsberg, J., Steinbach, J.P., Wick, A., Schnell, O., Hau, P., Herrlinger, U., Sabel, M.C., et al. MGMT Promoter Methylation Is a Strong Prognostic Biomarker for Benefit from Dose-Intensified Temozolomide Rechallenge in Progressive Glioblastoma: The DIRECTOR Trial. Clin Cancer Res, 2015, 21(9), 2057-64. [Google Scholar] [CrossRef]
- Malmström, A. , Grønberg, B.H., Marosi, C., Stupp, R., Frappaz, D., Schultz, H., Abacioglu, U., Tavelin, B., Lhermitte, B., Hegi, M.E., et al. Temozolomide versus standard 6-week radiotherapy versus hypofractionated radiotherapy in patients older than 60 years with glioblastoma: the Nordic randomised, phase 3 trial. Lancet Oncol 2012, 13(9), 916-26. [CrossRef]
- Tandel, S. G,, Biswas, M., Kakde, G.O., Tiwari, A., Suri, S.H., Turk, M., Laird, J.R., Asare, C.K., Ankrah, A.A., Khanna, N.N. A Review on a Deep Learning Perspective in Brain Cancer Classification. Cancers (Basel). 2019, 11(1), 111. [Google Scholar] [CrossRef]
- DeWitt, J.C. , Jordan, J.T., Frosch, M.P., Samore, W.R., Iafrate, A.J., Louis, D.N., Lennerz, J.K. Cost-effectiveness of IDH testing in diffuse gliomas according to the 2016 WHO classification of tumors of the central nervous system recommendations. Neuro Oncol. 2017, 19(12), 1640–1650. [Google Scholar] [CrossRef]
- Korshunov, A. , Chavez, L., Sharma, T., Ryzhova, M., Schrimpf, D., Stichel, D., Capper, D., Sturm, D., Kool, M., Habel, A., et al. Epithelioid glioblastomas stratify into established diagnostic subsets upon integrated molecular analysis. Brain Pathol. 2018, 28(5), 656–662. [Google Scholar] [CrossRef]
- Körber, V. , Yang, J., Barah, P., Wu, Y., Stichel, D., Gu, Z., Fletcher, M.N.C., Jones, D., Hentschel, B., Lamszus, K., et al. Evolutionary Trajectories of IDHWT Glioblastomas Reveal a Common Path of Early Tumorigenesis Instigated Years ahead of Initial Diagnosis. Cancer Cell 2019, 35(4), 692-704.e12. [CrossRef]
- Touat, M. , Li, Y.Y., Boynton, A.N., Spurr, L.F., Iorgulescu, J.B., Bohrson, C.L., Cortes-Ciriano, I., Birzu, C., Geduldig, J.E., Pelton, K., Lim-Fat, M.J., et al. Mechanisms and therapeutic implications of hypermutation in gliomas. Nature, . 2020, 580(7804), 517-523. [Google Scholar] [CrossRef]
- Barthel FP, Johnson KC, Varn FS, Moskalik AD, Tanner G, Kocakavuk E, Anderson KJ, Abiola O, Aldape K, Alfaro KD, et al. Longitudinal molecular trajectories of diffuse glioma in adults. Nature. 2019, 576(7785), 112–120. [Google Scholar] [CrossRef]
- Hasan, H.; Afzal, M.; Castresanam, J.S.; Shahi, M.H. A Comprehensive Review of miRNAs and Their Epigenetic Effects in Glioblastoma. Cells. 2023, 12(12), 1578. [Google Scholar] [CrossRef]
- Lewis, B.P. , Burge, C.B., Bartel, D.P. Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets. Cell 2005, 120(1), 15-20. [CrossRef]
- Ouellet, D.L.; Perron, M.P.; Gobeil, L.A.; Plante, P.; Provost, P. MicroRNAs in gene regulation: when the smallest governs it all. J Biomed Biotechnol. 2006, 2006(4), 69616. [Google Scholar] [CrossRef]
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nat Rev Mol Cell Biol. 2014, 15(8), 509–524. [Google Scholar] [CrossRef] [PubMed]
- Leonov, G. , Shah, K., Yee, D., Timmis, J., Sharp, T.V., Lagos, D. Suppression of AGO2 by miR-132 as a determinant of miRNA-mediated silencing in human primary endothelial cells. Int J Biochem Cell Biol, 2015, 69, 75-84. [CrossRef]
- Zhang, B.; Pan, X.; Cobb, G.P.; Anderson, T.A. microRNAs as oncogenes and tumor suppressors. Dev Biol. 2007, 302(1),1-12. [Google Scholar] [CrossRef]
- Sempere, L.F.; Azmi, A.S.; Moore, A. microRNA-based diagnostic and therapeutic applications in cancer medicine. Wiley Interdiscip Rev RNA, 2021; 12(6), e1662. [Google Scholar] [CrossRef]
- Zheng, Q.; Hou, W. Regulation of angiogenesis by microRNAs in cancer. Mol Med Rep. 2021, 24(2), 583. [Google Scholar] [CrossRef]
- Uzuner, E.; Ulu, G.T.; Gürler, S.B.; Baran, Y. The Role of MiRNA in Cancer: Pathogenesis, Diagnosis, and Treatment. Methods Mol Biol. 2022, 2257, 375–422. [Google Scholar] [CrossRef]
- Calin, G.A.; Dumitru, C.D.; Shimizu, M.; Bichi, R.; Zupo, S.; Noch, E.; Aldler, H.; Rattan, S.; Keating, M.; Rai, K.; et al. Frequent deletions and down-regulation of micro- RNA genes miR15 and miR16 at 13q14 in chronic lymphocytic leukemia. Proc Natl Acad Sci U S A. 2002, 99(24), 15524–15529. [Google Scholar] [CrossRef]
- Hosseini, S.A.; Horton, S.; Saldivar, J.C.; Miuma, S.; Stampfer, M. R.; Heerema, N. A.; & Huebner, K. Common chromosome fragile sites in human and murine epithelial cells and FHIT/FRA3B loss-induced global genome instability [published correction appears in Genes Chromosomes Cancer. 2019 Nov;58(11):824. https://doi.org/10.1002/gcc.22797]. . Genes Chromosomes Cancer. 2013, 52(11), 1017-1029. 10.1002/gcc.22097. [CrossRef]
- Behrooz, A.B.; Latifi-Navid, H.; Nezhadi, A. , Świat, M.; Los, M.; Jamalpoor, Z.; & Ghavami, S. Molecular mechanisms of microRNAs in glioblastoma pathogenesis. Biochim Biophys Acta Mol Cell Res. 2023, 1870(6), 119482. [Google Scholar] [CrossRef]
- Makowska, M.; Smolarz, B.; Romanowicz, H. microRNAs (miRNAs) in Glioblastoma Multiforme (GBM)-Recent Literature Review. Int J Mol Sci. 2023, 24(4), 3521. [Google Scholar] [CrossRef]
- Barciszewska A., M. MicroRNAs as efficient biomarkers in high-grade gliomas. Folia neuropathologica. 2016, 54(4), 369–374. [Google Scholar] [CrossRef]
- Coolen, M. , Katz, S., & Bally-Cuif, L. miR-9: a versatile regulator of neurogenesis. Frontiers in cellular neuroscience. 2013, 7, 220. [Google Scholar] [CrossRef]
- Dong, Z. , & Cui, H. Epigenetic modulation of metabolism in glioblastoma. Seminars in cancer biology. 2019, 57, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Munoz, J. L. , Rodriguez-Cruz, V., & Rameshwar, P. High expression of miR-9 in CD133+ glioblastoma cells in chemoresistance to temozolomide. Journal of cancer stem cell research, 2015, 3, e1003. [Google Scholar] [CrossRef]
- Yuan, G. Q. , Wei, N. L., Mu, L. Y., Wang, X. Q., Zhang, Y. N., Zhou, W. N., & Pan, Y. W.. A 4-miRNAs signature predicts survival in glioblastoma multiforme patients. Cancer biomarkers : section A of Disease markers. 2017, , 20(4), 443–452. [CrossRef]
- Rezaei, O. , Honarmand, K., Nateghinia, S., Taheri, M., & Ghafouri-Fard, S. miRNA signature in glioblastoma: Potential biomarkers and therapeutic targets. Experimental and molecular pathology, . 2020, 117, 104550. [Google Scholar] [CrossRef]
- Santolini, E. , Puri, C., Salcini, A. E., Gagliani, M. C., Pelicci, P. G., Tacchetti, C., & Di Fiore, P. P. Numb is an endocytic protein. The Journal of cell biology.. 2000, 151(6), 1345–1352. [Google Scholar] [CrossRef] [PubMed]
- de Menezes, M. R. , Acioli, M. E. A., da Trindade, A. C. L., da Silva, S. P., de Lima, R. E., da Silva Teixeira, V. G., & Vasconcelos, L. R. S. Potential role of microRNAs as biomarkers in human glioblastoma: a mini systematic review from 2015 to 2020. Molecular biology reports, 2021, 48(5), 4647–4658. [Google Scholar] [CrossRef]
- Junior, L. G. D. , Baroni, M., Lira, R. C. P., Teixeira, S., Fedatto, P. F., Silveira, V. S., Suazo, V. K., Veronez, L. C., Panepucci, R. A., Antônio, D. S. M. et al. High-throughput microRNA profile in adult and pediatric primary glioblastomas: the role of miR-10b-5p and miR-630 in the tumor aggressiveness. Molecular biology reports, 2020, 47(9), 6949–6959. [Google Scholar] [CrossRef]
- Gabriely, G. , Yi, M., Narayan, R. S., Niers, J. M., Wurdinger, T., Imitola, J., Ligon, K. L., Kesari, S., Esau, C., Stephens, R. M., Tannous, B. A., & Krichevsky, A. M. Human glioma growth is controlled by microRNA-10b. Cancer research, 2011, 71(10), 3563–3572. [Google Scholar] [CrossRef]
- Sun, L. , Yan, W., Wang, Y., Sun, G., Luo, H., Zhang, J., Wang, X., You, Y., Yang, Z., & Liu, N. MicroRNA-10b induces glioma cell invasion by modulating MMP-14 and uPAR expression via HOXD10. Brain research. 2011, 1389, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Sun, B. , Zhao, X., Ming, J., Liu, X., Liu, D., & Jiang, C. Stepwise detection and evaluation reveal miR-10b and miR-222 as a remarkable prognostic pair for glioblastoma. Oncogene, . 2019, 38(33), 6142–6157. [Google Scholar] [CrossRef]
- Cai, Q. , Zhu, A., & Gong, L. Exosomes of glioma cells deliver miR-148a to promote proliferation and metastasis of glioblastoma via targeting CADM1. Bulletin du cancer 2018, 105(7-8), 643–651. [CrossRef]
- Srinivasan, S. , Patric, I. R., & Somasundaram, K. A ten-microRNA expression signature predicts survival in glioblastoma. PloS one, . 2011, 6(3), e17438. [Google Scholar] [CrossRef]
- Vo, D. T. , Qiao, M., Smith, A. D., Burns, S. C., Brenner, A. J., & Penalva, L. O. The oncogenic RNA-binding protein Musashi1 is regulated by tumor suppressor miRNAs. RNA biology 2011, 8(5), 817–828. [CrossRef]
- Shea, A. , Harish, V., Afzal, Z., Chijioke, J., Kedir, H., Dusmatova, S., Roy, A., Ramalinga, M., Harris, B., Blancato, J., et al. MicroRNAs in glioblastoma multiforme pathogenesis and therapeutics. Cancer medicine. 2016, 5(8), 1917–1946. [Google Scholar] [CrossRef]
- Dambal, S. , Shah, M., Mihelich, B., & Nonn, L. The microRNA-183 cluster: the family that plays together stays together. Nucleic acids research. 2015, 43(15), 7173–7188. [CrossRef]
- Schneider, B. , William, D., Lamp, N., Zimpfer, A., Henker, C., Classen, C. F., & Erbersdobler, A. The miR-183/96/182 cluster is upregulated in glioblastoma carrying EGFR amplification. Molecular and cellular biochemistry, 2022, 477(9), 2297–2307. [Google Scholar] [CrossRef]
- Vilar, J. B. , Christmann, M., & Tomicic, M. T. Alterations in Molecular Profiles Affecting Glioblastoma Resistance to Radiochemotherapy: Where Does the Good Go? Cancers. 2022, 14(10), 2416. [Google Scholar] [CrossRef]
- Kouri, F. M. , Ritner, C., & Stegh, A. H. miRNA-182 and the regulation of the glioblastoma phenotype - toward miRNA-based precision therapeutics. Cell cycle (Georgetown, Tex.), 2015, 14(24), 3794–3800. [Google Scholar] [CrossRef]
- Zhao, H. , Shen, J., Hodges, T. R., Song, R., Fuller, G. N., & Heimberger, A. B. Serum microRNA profiling in patients with glioblastoma: a survival analysis. Molecular cancer. 2017, 16(1), 59. [CrossRef]
- Xiao, Y. , Zhang, L., Song, Z., Guo, C., Zhu, J., Li, Z., & Zhu, S. Potential Diagnostic and Prognostic Value of Plasma Circulating MicroRNA-182 in Human Glioma. Medical science monitor: international medical journal of experimental and clinical research. [CrossRef]
- Jiang, L. , Mao, P. , Song, L., Wu, J., Huang, J., Lin, C., Yuan, J., Qu, L., Cheng, S. Y., & Li, J. miR-182 as a prognostic marker for glioma progression and patient survival. The American journal of pathology. 2010, 177(1), 29–38. [Google Scholar] [CrossRef]
- Chen, C, Zhang, Y, Zhang, L, Weakley, S.M., Yao, Q. MicroRNA-196: critical roles and clinical applications in development and cancer. J Cell Mol Med. 2011, 15(1), 14-23. [CrossRef]
- Hassan, A. , Mosley, J., Singh, S., & Zinn, P. O. A Comprehensive Review of Genomics and Noncoding RNA in Gliomas. Topics in magnetic resonance imaging. 2017, 26(1), 3–14. [Google Scholar] [CrossRef]
- Lakomy, R. , Sana, J., Hankeova, S., Fadrus, P., Kren, L., Lzicarova, E., Svoboda, M., Dolezelova, H., Smrcka, M., Vyzula, R., et al. MiR-195, miR-196b, miR-181c, miR-21 expression levels and O-6-methylguanine-DNA methyltransferase methylation status are associated with clinical outcome in glioblastoma patients. Cancer science. ( 2011, 102(12), 2186–2190. [CrossRef]
- Cardia A, Epistolio S, Zaed I, Sahnane N, Cerutti R, Cipriani D, Barizzi J, Spina P, Stefanini FM, Cerati M et al. Identification of MGMT Downregulation Induced by miRNA in Glioblastoma and Possible Effect on Temozolomide Sensitivity. J Clin Med. 2023, 12(5), 2061. [Google Scholar] [CrossRef]
- Huse, J. T. , Brennan, C., Hambardzumyan, D., Wee, B., Pena, J., Rouhanifard, S. H., Sohn-Lee, C., le Sage, C., Agami, R., Tuschl, T. et al. The PTEN-regulating microRNA miR-26a is amplified in high-grade glioma and facilitates gliomagenesis in vivo. Genes & development, 1327. [Google Scholar] [CrossRef]
- Guo, P. , Lan, J., Ge, J., Nie, Q., Guo, L., Qiu, Y., & Mao, Q. MiR-26a enhances the radiosensitivity of glioblastoma multiforme cells through targeting of ataxia-telangiectasia mutated. Experimental cell research. 2014, 320(2), 200–208. [Google Scholar] [CrossRef]
- ParvizHamidi, M. , Haddad, G., Ostadrahimi, S., Ostadrahimi, N., Sadeghi, S., Fayaz, S., & Fard-Esfahani, P. Circulating miR-26a and miR-21 as biomarkers for glioblastoma multiform. Biotechnology and applied biochemistry 2019, 66(2), 261–265. [CrossRef]
- Siegel, S. R. , Mackenzie, J., Chaplin, G., Jablonski, N. G., & Griffiths, L. Circulating microRNAs involved in multiple sclerosis. Molecular biology reports. 2012, 39(5), 6219–6225. [Google Scholar] [CrossRef] [PubMed]
- Epistolio S, Dazio G, Zaed I, Sahnane N, Cipriani D, Polinelli F, Barizzi J, Spina P, Stefanini FM, Cerati M, Balbi S, Mazzucchelli L, Sessa F, Pesce GA, Reinert M, Cardia A, Marchi F, Frattini M. Clinical Relevance and Interplay between miRNAs in Influencing Glioblastoma Multiforme Prognosis. Cells. 2024, 13(3), 276. [Google Scholar] [CrossRef]
- Kreth, S. , Limbeck, E., Hinske, L. C., Schütz, S. V., Thon, N., Hoefig, K., Egensperger, R., & Kreth, F. W. In human glioblastomas transcript elongation by alternative polyadenylation and miRNA targeting is a potent mechanism of MGMT silencing. Acta neuropathological. 2013, 125(5), 671–681. [Google Scholar] [CrossRef]
- Kalkan, R. , & Atli, E. İ. The Impacts of miRNAs in Glioblastoma Progression. Critical reviews in eukaryotic gene expression. 2016, 26(2), 137–142. [Google Scholar] [CrossRef]
- Yang, C. H. , Wang, Y., Sims, M., Cai, C., & Pfeffer, L. M. MicroRNA-1 suppresses glioblastoma in preclinical models by targeting fibronectin. Cancer letters. 2019, 465, 59–67. [CrossRef]
- Marumoto, T. , & Saya, H. Molecular biology of glioma. Advances in experimental medicine and biology. 2012, 746, 2–11. [Google Scholar] [CrossRef]
- Cardoso, A. M. , Morais, C. M., Pena, F., Marante, T., Cunha, P. P., Jurado, A. S., & Pedroso de Lima, M. C. Differentiation of glioblastoma stem cells promoted by miR-128 or miR-302a overexpression enhances senescence-associated cytotoxicity of axitinib. Human molecular genetics 2021, 30(3-4), 160–171. [CrossRef]
- Balandeh, E. , Mohammadshafie, K., Mahmoudi, Y., Hossein Pourhanifeh, M., Rajabi, A., Bahabadi, Z. R., Mohammadi, A. H., Rahimian, N., Hamblin, M. R., & Mirzaei, H. Roles of Non-coding RNAs and Angiogenesis in Glioblastoma. Frontiers in cell and developmental biology, 2021, 9, 716462. [Google Scholar] [CrossRef]
- Sun, J. , Zheng, G., Gu, Z., & Guo, Z. (2015). MiR-137 inhibits proliferation and angiogenesis of human glioblastoma cells by targeting EZH2. Journal of neuro-oncology 2015. 122(3), 481–489. [CrossRef]
- Li, Y. , Fan, S., Xia, W., Qiao, B., Huang, K., Zhou, J., & Liang, M. MiR-181b suppresses angiogenesis by directly targeting cellular communication network factor 1. Laboratory investigation; a journal of technical methods and pathology, 2021, 101(8), 1026–1035. [Google Scholar] [CrossRef]
- Nadaradjane, A. , Briand, J., Bougras-Cartron, G., Disdero, V., Vallette, F. M., Frenel, J. S., & Cartron, P. F. miR-370-3p Is a Therapeutic Tool in Anti-glioblastoma Therapy but Is Not an Intratumoral or Cell-free Circulating Biomarker. Molecular therapy Nucleic acids. [CrossRef]
- Kirstein, A. , Schmid, T. E., & Combs, S. E. The Role of miRNA for the Treatment of MGMT Unmethylated Glioblastoma Multiforme. Cancers. 2020, 12(5), 1099. [Google Scholar] [CrossRef]
- Morales-Martínez, M. , & Vega, M. I. Role of MicroRNA-7 (MiR-7) in Cancer Physiopathology. International journal of molecular sciences, 2022, 23(16), 9091. [Google Scholar] [CrossRef]
- Alamdari-Palangi, V. , Amini, R., & Karami, H. MiRNA-7 enhances erlotinib sensitivity of glioblastoma cells by blocking the IRS-1 and IRS-2 expression. The Journal of pharmacy and pharmacology. 2020, 72(4), 531–538. [Google Scholar] [CrossRef]
- Li, Y. , Guessous, F., Zhang, Y., Dipierro, C., Kefas, B., Johnson, E., Marcinkiewicz, L., Jiang, J., Yang, Y., Schmittgen, T. D., et al. MicroRNA-34a inhibits glioblastoma growth by targeting multiple oncogenes. Cancer research, 2009, 69(19), 7569–7576. [Google Scholar] [CrossRef]
- Bazzoni, R. , & Bentivegna, A. Role of Notch Signaling Pathway in Glioblastoma Pathogenesis. Cancers. 2019, 11(3), 292. [Google Scholar] [CrossRef]
- Janaki Ramaiah, M. , Divyapriya, K., Kartik Kumar, S., & Rajesh, Y. B. R. D. Drug-induced modifications and modulations of microRNAs and long non-coding RNAs for future therapy against Glioblastoma Multiforme. Gene, 2020, 723, 144126. [Google Scholar] [CrossRef]
- Francipane, M. G. , Douradinha, B., Chinnici, C. M., Russelli, G., Conaldi, P. G., & Iannolo, G. Zika Virus: A New Therapeutic Candidate for Glioblastoma Treatment. International journal of molecular sciences, 2021, 22(20), 1099. [Google Scholar] [CrossRef]
- Zhu, Z.; Gorman, M.J.; McKenzie, L.D.; Chai, J.N.; Hubert, C.G.; Prager, B.C.; Fernandez, E.; Richner, J.M.; Zhang, R.; Shan, C.; et al. Zika virus has oncolytic activity against glioblastoma stem cells. J. Exp. Med. 2017, 214, 2843–2857. [Google Scholar] [CrossRef]
- Lubin, J.; Zhang, R.R.; Kuo, J. Zika Virus has Oncolytic Activity Against Glioblastoma Stem Cells. Neurosurgery. 2018, 82, E113–E114. [Google Scholar] [CrossRef]
- Kaid, C.; Goulart, E.; Caires-Júnior, L.C.; Araujo, B.H.S.; Schanoski, A.S.; Bueno, H.; Silva, K.A.T.; Astray, R.M.; Assoni, A.F.; Júnior, A.F.R.; et al. Zika Virus Selectively Kills Aggressive Human Embryonal CNS Tumor Cells In Vitro and In Vivo. Cancer Res. 2018, 78, 3363–3374. [Google Scholar] [CrossRef] [PubMed]
- Baronti, C.; Piorkowski, G.; Charrel, R.N.; Boubis, L.; Leparc-Goffart, I.; de Lamballerie, X. Complete Coding Sequence of Zika Virus from a French Polynesia Outbreak in 2013. Genome Announc. 2014, 2, e00500–14. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Pathway of miRNA biogenesis and action. The figure has been created applying images reported at https://smart.servier.com/.
Figure 1.
Pathway of miRNA biogenesis and action. The figure has been created applying images reported at https://smart.servier.com/.
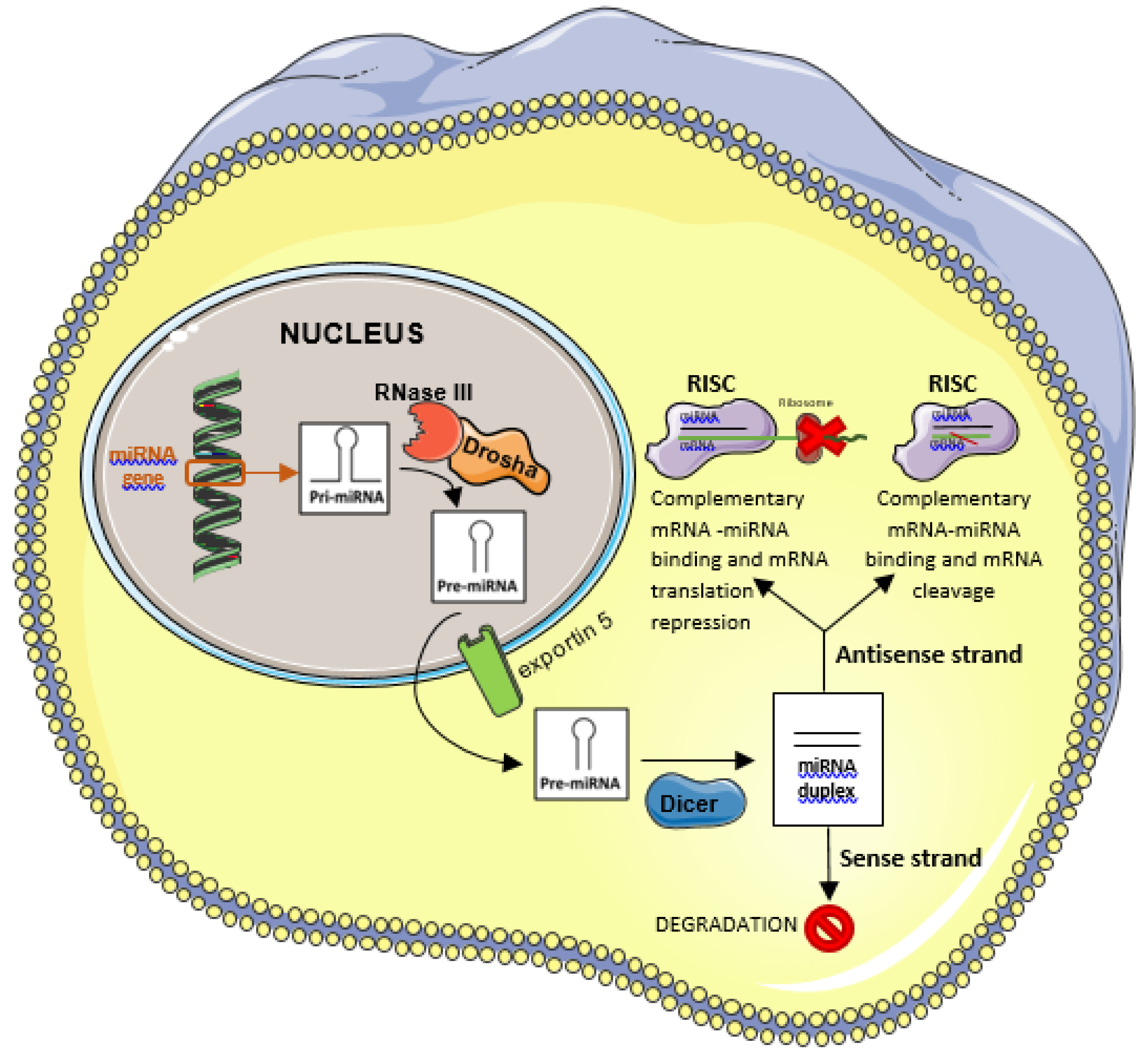
Figure 2.
Targets of OncomiR in glioblastoma. MiRNAs that block a specific pathway are shown in red, those that activate a pathway in green. The figure has been created applying images reported at https://smart.servier.com/.
Figure 2.
Targets of OncomiR in glioblastoma. MiRNAs that block a specific pathway are shown in red, those that activate a pathway in green. The figure has been created applying images reported at https://smart.servier.com/.
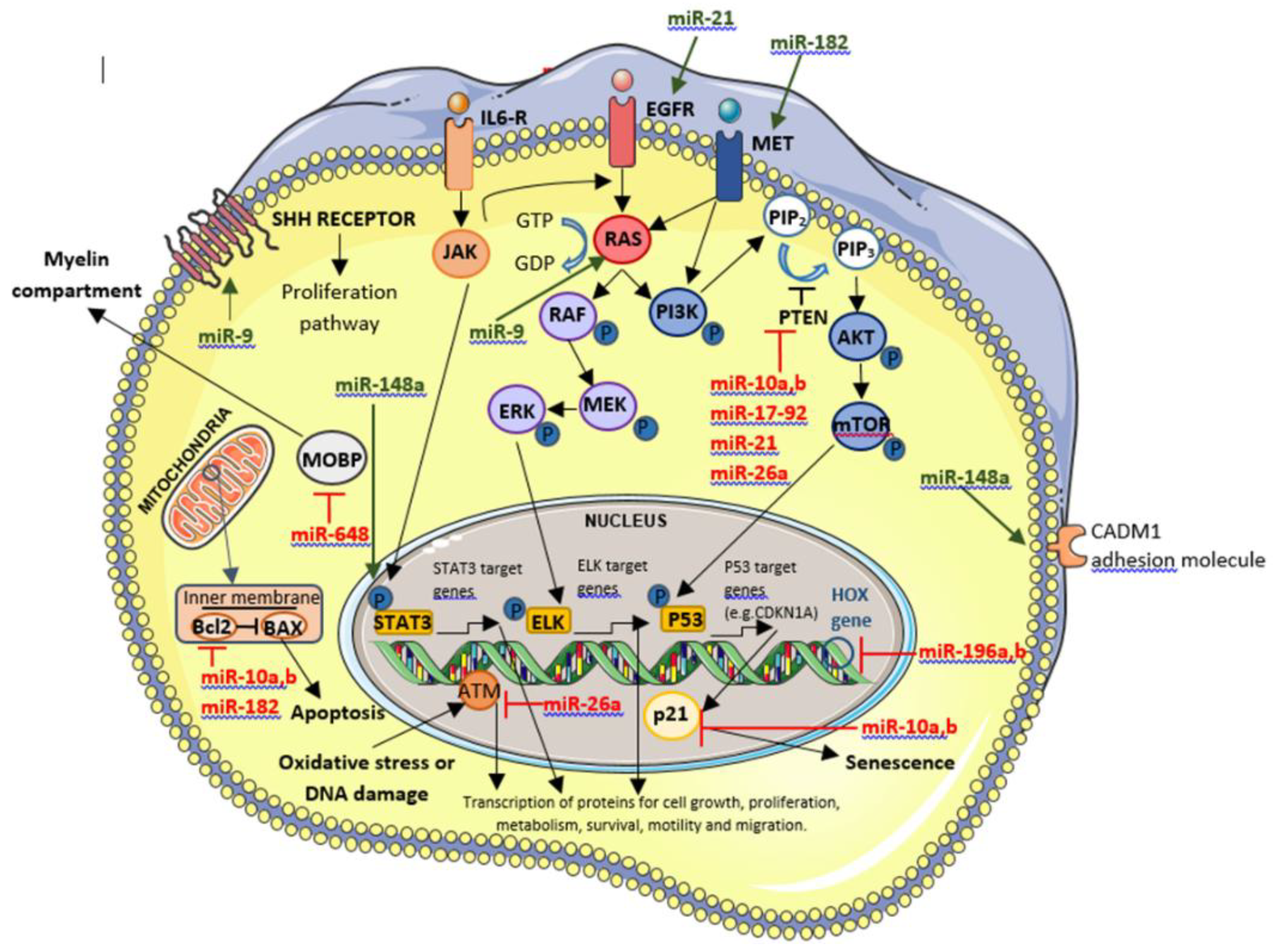
Figure 3.
Targets of tumor suppressor miRNAs in glioblastoma. MiRNAs that block a specific pathway are shown in red, those that activate a pathway in green. The figure has been created applying images reported at https://smart.servier.com/.
Figure 3.
Targets of tumor suppressor miRNAs in glioblastoma. MiRNAs that block a specific pathway are shown in red, those that activate a pathway in green. The figure has been created applying images reported at https://smart.servier.com/.
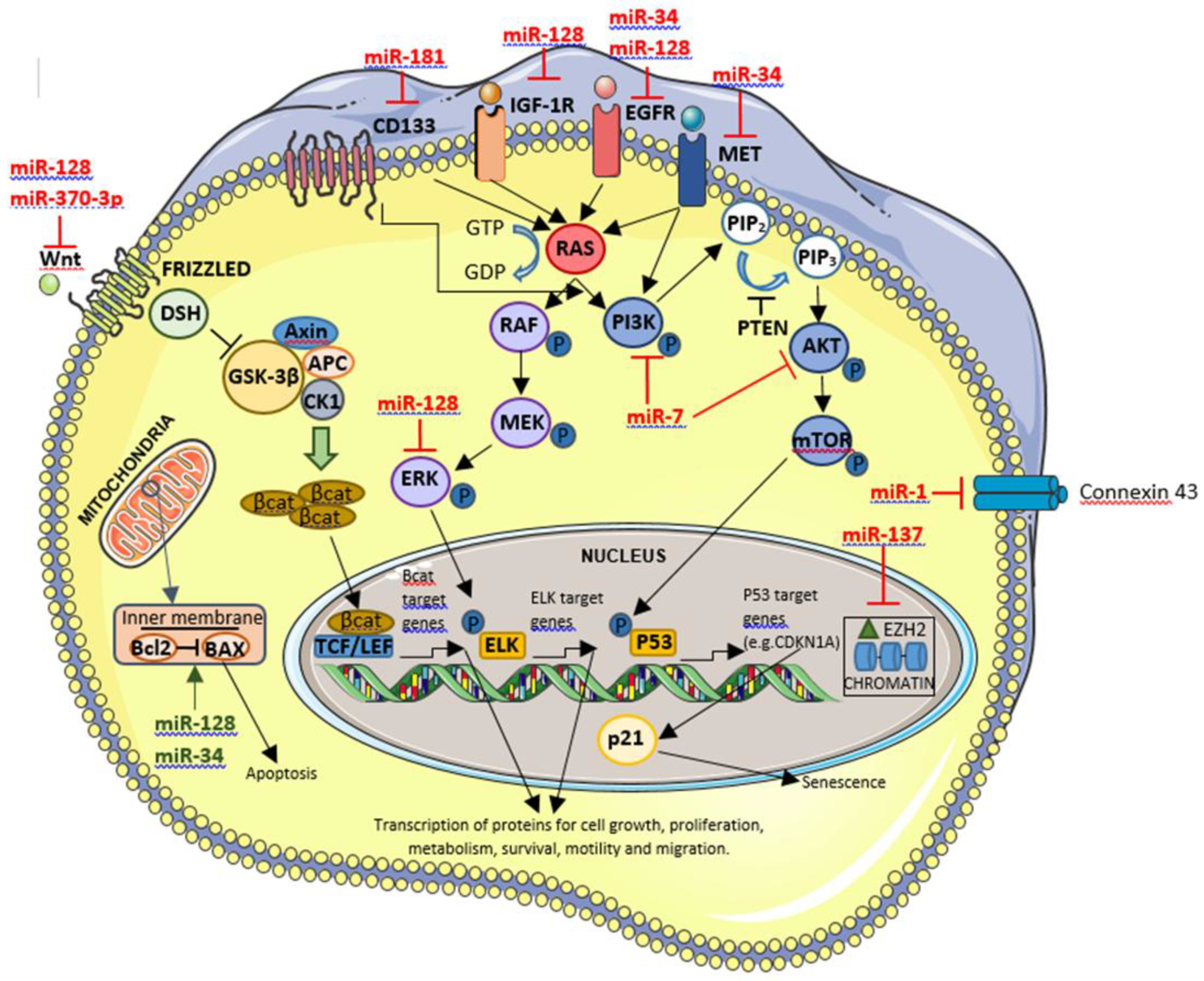

Table 1.
List and characteristics of OncomiR in GBM.
| miRNA | Target | Expression in GBM | Function/role in GBM if the miRNA is overexpressed | Clinical application | References |
|---|---|---|---|---|---|
| miR-9 | -RAS and MYC -PTCH1 |
↑ | Cancer cells proliferation↑ Tumor cells transformation↑ Inflammation↑ Angiogenesis↑ Apoptosis↓ |
In vitro: overexpression can enhance the resistance to TMZ in GBM cells. In patients: associated to short term survivors. |
[49,50,51,52] |
| miR-10a, b | -CDKN1A, BIM, BCL2, TEAP2C and PTEN -HOXD10 |
↑ | Cancer cells proliferation↑ Tumor cells migration↑ Invasion↑ EMT promotion↑ Apoptosis↓ GSC differentiation↑ |
In patients: the expression is higher in GBM than in other gliomas. |
[53,54,55,56,57,58] |
|
miR-17-92 cluster |
-Some cell-cycle inhibitors such as PTEN and CDKN1A | ↑ | Cancer cells proliferation↑ Apoptosis↓ GSC differentiation↑ |
In vitro: are associated with high aggressiveness, higher invasion and replication capability. |
[48] |
| miR-21 | -PTEN, p53 -EGFR, Cyclin D1 and AKT2 -SPOCK1 -RECK and TIMP3 |
↑ | Cancer cells proliferation↑ Tumor cells migration↑ Invasion↑ |
In patients: hypothesis that silencing of this miRNA can be used, in the future, as a therapy in the treatment of GBM. |
[35,48,53,55] |
| miR-26a | -PTEN, ATM | ↑ | Cancer cells proliferation↑ Invasion↑ |
In vitro: overexpression of this miRNA, reducing DNA repair ability, enhances radio sensitivity to radiotherapy. |
[75,76,77] |
| miR-148a | -CADM1 -FIH1 |
↑ | Angiogenesis↑ Invasion↑ |
In patients: -overexpression in the plasma from the serum of GBM patients if compared to healthy cases. -has been found to be expressed in the high risk group (i.e. patients characterized by low survival). |
[53,60,61,62] |
| miR-182 | -BCL2L12 -HIF2A -MET -CYLD -LRRC4 |
↑ | Uncontrolled cells proliferation↑ Apoptosis↑ GSC differentiation↑ |
In patients: -miR-182 correlates with better response to TMZ based chemotherapy and with better survival. |
[64,65,66] |
| miR-196a, b | -HOXB8, HOXC8, HOXD8, HOXA7, HOXB7 -ERG -HMGA2 -ANXA1 |
↑ | Cancer cells proliferation↑ Apoptosis↓ |
In vitro: -overexpression of this miRNA favours cells proliferation. In patients: -miR-196b expression correlated with OS. |
[71,72,73,74] |
| miR-648 | -MOBP | ↑ | Cancer cells proliferation↑ Invasion↑ |
In vitro: - the expression by transfection of miR-648 enhanced responsivity of TMZ in MGMT expressing T98G glioma cells. In patients: -correlation between OS and miR-648 expression. |
[74,78,79,80] |
Table 2.
List and characteristics of Tumor suppressor miRNAs in GBM.
| miRNA | Target | Expression in GBM | Function/role in GBM if the miRNA expression is inhibited | Clinical application | References |
|---|---|---|---|---|---|
| miR-1 | -Connexin-43 -G6PD | ↓ | Cancer cells proliferation↑ Tumor cells migration↑ Apoptosis↓ |
In vitro: inhibition can enhance the cells proliferation and the sensitivity of GBM cells towards TMZ. |
[53,81,82] |
|
miR-7 |
-EGFR, AKT/ PI3K pathway -PKM2 |
↓ | Cancer cells proliferation↑ GSC differentiation↑ |
In vitro: transfection of this miRNA in U373-MG GBM cell line resulted in significant suppression of EGFR mRNA and protein bringing to the inhibition of cells duplication. |
[35,90,91] |
|
miR-34 |
-Bcl2, NOTCH, NUMB -CDK6 -EGFR -c-Met |
↓ | Cancer cells proliferation↑ Apoptosis↓ Invasion↑ |
In vitro: -in G[92–99SC cultures the infection by ZIKV induced miR-34 expression inhibiting the anti-apoptotic protein Bcl-2 and Numb, involved in GSC invasion. - in mouse models ZIKV reduced brain tumor size and metastasis. |
[92,93,94,95,96,97,98,99] |
| miR-128 | -WNT -ERK -EGFR -IGF1R -Bcl2 -PDGFRA -caspase |
↓ | Cancer cells proliferation↑ Apoptosis↓ |
In vitro: its low expression can be associated to high-grade glioma cell lines and, consequently, to a worst prognosis. |
[53,83,84] |
|
miR-137 |
- EZH2 | ↓ | Cancer cells proliferation↑ Apoptosis↓ Angiogenesis↑ |
In vitro: expression level of miR-137 was down-regulated in GBM cells. In patients: the low level of this miRNA was related to poor prognosis in GBM patients. |
[53,85,86] |
| miR-181 family | -CD133 and BMI1 CCN1 |
↓ | Cancer cells proliferation↑ GSC differentiation↑ Apoptosis↓ Invasion↑ |
In patients: -the low level of this miRNA was related to poor prognosis in GBM patients. - low expression of miR-181c or low expression of miR-181d in combination with expression of miR-648, predict the worst prognosis. |
[73,79,81,87] |
| miR-370-3p | -WNT - FOX01, FOXM1 and TGFβ. |
↓ | Cancer cells proliferation↑ Invasion↑ |
In vitro: when miR-370-3p is upregulated, GBM growth is inhibited |
[88,89] |
Abbreviations: GBM, Glioblastoma; GSC, GBM stem cells; OS, Overall survival; TMZ, Temozolomide.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



