
Submitted:
03 January 2025
Posted:
04 January 2025
You are already at the latest version
Abstract
Serum amyloid A (SAA) is one of the most abundant acute phase response proteins and has been extensively studied in vertebrates for its role in modulation of the inflammatory response and marker of disease diagnosis. By comparison, SAA is rarely identified in aquatic species and its physical functions are also not well studied. The present study identified the only one gene encoding SAA protein in oyster Crassostrea gigas. The open reading frame (ORF) of CgSAA was of 417 bp, encoding a putative polypeptide of 138 amino acid residues with predicted molecular weight of 15.66 kDa. CgSAA was composed of a signal peptide (residues 1-22) and a conserved SAA domain (residues 36-138). The mRNA expression of CgSAA in normal individuals was detectable but in low level, with the lowest expression level in tissue of labial palp and slightly higher expression level in haemocytes. The mRNA expression level of CgSAA was significantly up-regulated at 6 h (2.76-fold of that in control group, p < 0.01) post V. splendidus stimulation. It was also significantly induced under environmental stress of high temperature (34 ℃) or low permeability (15‰ salinity). The recombinant protein rCgSAA was expressed in Escherichia coli and purified by affinity chromatography. After rCgSAA was injected into oysters or incubated with culture primary haemocytes, the mRNA expressions of cytokine CgIL17-1, CgIL17-5 and CgTNF were all significantly un-regulated. The results collectively suggested that CgSAA, as a conserved acute phase response protein in oyster, was quickly induced under environmental stress and promoted the expressions of cytokines, which provided fresh ideas for understanding the roles of SAA proteins in aquatic invertebrates.
Keywords:
acute phase response
; serum amyloid A (SAA)
; cytokines production
; oyster Crassostrea gigas
1. Introduction
The acute phase response (APR) is a well-documented phenomenon in vertebrates, characterized by the rapid synthesis and release of acute-phase proteins (APPs) in response to infection, inflammation, or tissue injury [1]. It has been widely used in the early diagnosis of diseases. Serum amyloid A (SAA), one of the most abundant APPs, has been extensively studied in vertebrates for its role in opsonization, phagocytosis, and modulation of the inflammatory response [2,3]. In vertebrates, SAA is primarily synthesized in the liver during the APR and is secreted into the bloodstream. It binds to high-density lipoprotein (HDL) and accumulates in the tissues, particularly at sites of inflammation. SAA exhibits multiple functions, including opsonization of gram-negative bacteria, modulation of cytokine production, and stimulation of the innate immune system [4,5]. Its role in innate immunity is further underscored by its ability to induce the production of other immune mediators, such as complement components and reactive oxygen species [6].
Despite the well-documented roles of SAA in vertebrates, the existence and function of SAA-like proteins in invertebrates remain largely unknown. The primary challenge lies in the fact that invertebrates lack a liver, the primary site of SAA synthesis in vertebrates [7]. Analyze using transcriptome and genome data and in several protostome taxa made up of 21 marine bivalve species and basal metazoans, 51 SAA-like proteins were retrieved [7]. These SAA-like proteins share certain structural features with vertebrate SAA, including a conserved N-terminal region and a variable C-terminal region[7,8,9,10,11]. They are typically small, with molecular weights ranging from 10 to 20 kDa. Recently, proteins with structural and functional similarities to SAA have been identified in some invertebrate species. They are often synthesized in response to immune challenges, such as bacterial or viral infections, and are secreted into the hemolymph or other body fluids [8,9]. For example, in oyster Crassostrea hongkongensis, SAA was found highestly expressed in the mantle, and was significantly up-regulated in haemocytes following bacterial challenge [9]. In Tridacna crocea, the recombinant protein rTcSAA was found to bind bacteria Vibrio coralliilyticus and Vibrio alginolyticus and reduce the lethality rate of the clams after bacterial challenge [10]. Currently, the information of SAA in invertebrates is still limited.
The Pacific oyster Crassostrea gigas is filter-feeding bivalve and globally important economically farmed specie. However, infectious diseases have become the main obstacle to the development of the oyster farming industry, and a variety of diseases have seriously threatened the farming output of Crassostrea gigas [12]. Given that the rearing environments of oysters are usually coastal and estuarine areas, it is a big challenging for environmental management and control when dealing with the outbreak of infectious diseases. [13,14]. The early diagnosis of oyster infection is critical to prevent large-scale mortality, and the acute phase response related SAA might provide a fresh idea [15]. However, the molecular features and potential functions of SAA protein in Crassostrea gigas is still not clear. The present study identified the only saa gene (CgSAA) from oyster C. gigas, and its molecular characteristics, expression patter as well as possible functions were further investigated, with the aim to explore the evolution of SAAs and the possible role of CgSAA in the immune response.
2. Materials and Methods
2.1. Oysters, Treatments and Tissues Collection
Collect adult Crassostrea gigas with an average shell length of about 7 - 10 cm from a commercial aquaculture farm in Zhuanghe, Liaoning Province, and then place them in filtered seawater at 20 ± 2 ℃ for temporary rearing.
One week after temporary rearing, tissue samples were taken from 6 adult oysters for mRNA and protein extraction. The tissues included the labial palp, hepatopancreas, muscle, gill, ganglion, mantle and haemocytes. The immune stimulation protocol was modified according to previous reports [16]. The oysters were randomly divided into two groups with 36 individuals in each group. The oysters stimulated with 100 µL of Vibrio splendidus were used as the V. splendidus group, while the oysters in SW group received an injection of 100 μL sterilized sea water (SW). Subsequently, the haemolymph of the Pacific oysters Crassostrea gigas in each group was collected at 0, 6, 12, and 24 h after stimulation respectively. Then, under the conditions of 4 °C and 800 × g, it was centrifuged for 15 minutes to obtain haemocytes, and 1 mL of Trizol reagent was added for RNA extraction.
The environmental stress experiment in oysters was conducted according to previous report [17]. Briefly, thirty-six oysters were evenly divided into three groups. The oysters cultured in seawater of 34 ℃, 30‰ salinity (high temperature stress), seawater of 18 °C, 15‰ salinity (hypotonic stress) and normal seawater (18 °C, 30‰ salinity) were used as the hyperthermia group, low salinity group, and SW group, respectively. Then, haemocytes were collected at 0, 6, 12, and 24 h after stimulation for RNA extraction according to the above method.
2.2. cDNA synthesis, Cloning and Sequence Analysis of the CgSAA
After total mRNA extraction using Trizol reagent, the integrity and concentration of mRNA were analyzed by agarose gel electrophoresis and Nanodrop 2000. The cDNA was synthesized by reverse transcription kit (TransGen Biotech, China), and reaction conditions were 42 °C for 15 min and 85 °C for 5 sec, and the obtained cDNA was stored at -80 °C.
Based on the sequence information of CgSAA (XM_066077310.1) in the NCBI database (https://www.ncbi.nlm.nih.gov/), specific primers CgSAA-F and CgSAA-R were designed (Table 1). The obtained PCR products were transferred into competent cells of Trans 5α for massive amplification and then confirmed by DNA sequencing. The amino acid sequence was predicted using the Expert Protein Analysis System (http://www.expasy.org). The SAA domain was predicted with SMART (http://smart.embl-heidelberg.de/), and the neighbor-joining phylogenetic tree was constructed using MEGA-X software. GeneDoc and DNAMAN10.0 were utilized to conduct multiple sequence alignments for CgSAA.
2.3. Recombinant Expression and Purification of CgSAA Protein
The recombinant expression and purification of CgSAA protein were carried out according to the previous report[18]. The simplified process was as follows: PCR amplification was performed using primers CgSAA-EXF and CgSAA-EXR (Table 1). The PCR product was inserted into pET-30a (+) from Novagen in the USA. Then, pET-30a-CgSAA was transformed into E. coli (DE3) cells (TransGen Biotech, CN) and 0.5 mM IPTG was added for rCgSAA expression. The purification of rCgSAA was carried out as described in the previous report.
2.4. rCgSAA Treatment and Sample Collection
The in vivo and in vitro rCgSAA treatment experiments in oysters were conducted according to previous report [18,19]. To explore how CgSAA induce immune response in oyster, a total of 30 oysters were divided into two groups. In one group, each oyster was injected with 100 μL of rCgSAA at a concentration of 500 μg/mL. In another group, each oyster was injected with 100 μL of PBS as a negative control, and the individuals without injection served as the blank control. Random samples were taken from each group at 6 h and 12 h after injection respectively. The haemocytes of C. gigas were collected, and then RNA was extracted according to the subsequent methods to detect the gene expression levels.
The in vitro impaction of rCgSAA on gene expression of haemocyte were also conducted. The collected haemocytes were washed twice with M-L15, they were resuspended in the M-L15 and inoculated into 12-wells microplate (1 mL per well). The inoculated cells were then cultured at 18 °C for 6 h, then 0 μg, 5 μg, 10 μg or 50 μg of the recombinant protein rCgSAA was added to each well. The 12-wells microplate were taken out after an additional 6 h of incubation at 18 °C. Then the haemocytes were collected from the 12-well plates with three parallels for each treatment to extract mRNA, and the mRNA expression levels of CgIL17-1 [17], CgIL17-5 [20] and CgTNF[21] genes were detected as former mentioned.
2.5. Real-Time PCR Analysis of mRNA Expression
The expression levels of CgSAA in the tissues, embryos and larvae of Crassostrea gigas, as well as those in response to Vibrio splendidus, high temperature stress and low salt stress were detected by qRT-PCR. The primers used have been listed (table 1), the mRNA expression of candidate genes in hemocytes treated with rCgSAA was also detected. Taking CgEF (CGI_10012474) as an internal control, the analysis was carried out using the 2−ΔΔCT method.
2.7. Statistical Analysis
Data as mean ± SD, two-sample Student's t-test for group comps. Multiple groups by one-way ANOVA, then Tukey's test in SPSS 19.0. significant at p < 0.05.
3. Results
3.1. The Molecular Characters of CgSAA Gene
The open reading frame (ORF) of CgSAA measured 417 bp, which was responsible for encoding a presumed polypeptide composed of 138 amino acid residues. This polypeptide had an estimated molecular weight of 15.66 kDa and a theoretical isoelectric point (pI) of 9.84 (Figure 1A). The structural prediction of the amino acid sequence indicates that the oyster CgSAA is composed of two parts. The signal peptide covers the amino acid residues from residues 1-22, the CgSAA domain extends from residues 36-138 (Figure 1B).
Upon conducting multiple sequence comparisons, it was revealed that the SAA domain of the CgSAA gene exhibited a significant degree of similarity to those found in both chordates and invertebrates. Notably, the similarity was most pronounced (99%) with respect to that of Crassostrea virginica (Figure 2A). For the purpose of phylogenetic analysis, ten SAA-5 sequences from diverse species of vertebrates and invertebrates were chosen, CgSAA-5 was clustered with CvSAA-5 from C. virginica (Figure 2B).
A: The sequence of ORF and encoded amino acids. The nucleotides and amino acids are numbered along the left margin. The symbol (-) represents a stop codon. The SAA domain is shadowed.
B: Prediction of protein domains by SMART analysis.
A: Multiple sequence alignment analysis of putative domains of CgSAA with other SAA deposited in GenBank, the following are the proteins analyzed: Strongylocentrotus purpuratus (XP_030837042.1), Crassostrea virginica (XP_022321638.1), Branchiostoma belcheri (XP_019623558.1), Poecilia reticulata (XP_008433017.1), Poecilia latipinna (XP_014890824.1), Scleropages Formosus (XP_018618982.1), Microtus ochrogaster (XP_005366816.1), Alligator sinensis (XP_025057901.1), Crocodylus porosus (XP_019407521.1). The identical amino acid residues are shaded in black and the similar amino acid are shaded in grey.
B: Use the MEGA-X software to conduct phylogenetic analysis on CgSAA and other SAA proteins from multiple different species.
3.2. The Distribution of CgSAA mRNA Transcripts
Analysis of the expression of CgSAA in different tissues of Crassostrea gigas. The mRNA expression of CgSAA in normal individuals was detectable in all the sampled tissues but in low level, including hepatopancreas, mantle, gill, haemocytes, ganglion, adductor muscle and labial palp. In contrastive analysis, the expression level of CgSAA was detected to be the least in the labial palp. Conversely, haemocytes exhibited a relatively greater expression level of CgSAA, which was 3.85-fold higher than that in the labial palp (p < 0.01). It was also higher expressed in ganglions and adductor muscles, which were 3.42-fold (p < 0.05) and 2.99-fold (p < 0.05) of that in labial palp, respectively. While relatively lower in gill and mantle, which were 2.14-fold (p < 0.05), 1.89-fold (p < 0.05) of labial palp, respectively (Figure 3A).
During the early developmental process of the oyster Crassostrea gigas, the expression level of CgSAA in the larval stage was higher than that in the embryonic stage, and the expression level of CgSAA was the lowest at the blastula stage. When the embryos of the oyster developed to the trochophore stage, the expression level began to rise, which was 19.11-fold (p < 0.05) that of the blastula stage. The expression level was the highest at the Pediveliger stage, which was 275.32-fold (p < 0.05) that of the blastula stage (Figure 3B).
A: Expression of CgSAA mRNA in different tissues of oyster detected by real-time PCR.
B: The expression levels of CgSAA mRNA in the embryonic and larval stages of the Crassostrea gigas. Data was shown as mean ± S.D (N = 3). Statistically significant differences were designated at p < 0.05. Labial palp, Hepatopancreas, Muscle, Gill, Ganglion, Mantle, Haemocytes.
3.3. The Response of CgSAA in Oyster Under Immune Stimulation or Environmental Stress
The mRNA expressions of CgSAA in haemocytes were detected at 0, 6, 12 and 24 h after V. splendidus stimulation. The mRNA expression level of CgSAA was up-regulated at 6 h (2.76-fold, p < 0.01), surged at 12 h (7.88-fold, p < 0.05), and ascended to the peak level at 24 h, which was 21.85-fold (p < 0.05) higher than that in control group post V. splendidus stimulation, respectively (Figure 4A).
After being subjected to high temperature stress (34 ℃), the expression level of CgSAA in the Crassostrea gigas remained relatively unchanged in the first 6 h. Then it was significantly unregulated and peaked at 12 h, reaching 21.77-fold (p < 0.01) of that in the control group, and kept in high level at 24 h (14.6-fold of that in the control group, p < 0.05) (Figure 4B). Under hypotonic stress, the expression of CgSAA in haemocytes was quickly induced, with the highest expression level at 6 h in low salinity group, which was 3.58-fold (p < 0.01) that of the control group. Then the expression level of CgSAA decreased sharply with the prolongation of treatment time, and reached the lowest at 24 h (Figure 4C).
A: The mRNA expression of CgSAA in haemocytes following the stimulation by Vibrio splendidus.
B: The mRNA expression of CgSAA in haemocytes after 34 ℃ seawater treatment.
C: 15‰ salinity seawater treatment. Haemocytes were collected at 0, 6, 12 and 24 h after treatment respectively. Vertical bars represent the mean ± S.D (N = 4). The asterisks (*p < 0.05, **p < 0.01) indicate the significant differences.
3.4. The Expressions of Cytokines Following Injection of rCgSAA in Oysters
The recombinant protein of CgSAA (rCgSAA) containing SAA domain was expressed and purified by the CBP-tag Protein Fusion and Purification System. The band with molecular weight of 19.7 kDa was observed (CBP-tag is about 4 kDa), which were consistent with the predicted molecular weight (15.66 kDa) of CgSAA (Figure 5). Then the purified rCgSAA protein was used for oyster injection.
The mRNA expression of cytokine CgIL17-1, CgIL17-5 and CgTNF was detected after rCgSAA injection. The expression level of CgIL17-1 increased at 6 h after rCgSAA stimulation and peaked at 12 h, which was 10.29-fold (p < 0.01) that of that in the control group (Figure 6A). The relative expression level of CgIL17-5 was significantly induced at 12 h and 24 h, which were 1.5- and 22.18-fold of that in the control group, respectively, p < 0.05 (Figure 6B). Within 12 h after PBS injection, the expression levels of CgIL17-1 and CgIL17-5 did not change significantly. The relative expression level of CgTNF was up-regulated at 6 h and 12 h after rCgSAA injection, which was 1.64-fold (p > 0.05) and 2.48-fold (p < 0.01) higher than that of blank group, respectively (Figure 6C).
A-C: The expression levels of CgIL17-1, CgIL17-5 and CgTNF in the Crassostrea gigas after treatment with rCgSAA.
D: The expression levels of CgIL17-1, CgIL17-5 and CgTNF in haemocytes after rCgSAA treatment. Vertical bars represent the mean ± S.D. (N = 3). Statistically significant differences were designated at p < 0.05.
3.5. The Expressions of Cytokines in Haemocytes Following Incubation with rCgSAA
To explore how rCgSAA induce immune response in oyster, the cultured primary haemocytes were incubated with rCgSAA for 6 h, and expression of CgIL17-1, CgIL17-5 and CgTNF mRNA in haemocytes was validated. When the concentration of rCgSAA reached 50 μg/mL, it could significantly (p < 0.01) induce the expression of CgIL17-1 and CgIL17-5 in haemocytes, which were 2.48-fold or 2.55-fold that of 0 μg/mL group, respectively, (Figure 6D). In particular, when the concentration of rCgSAA reaches 10 μg/mL, the expression level of CgTNF in haemocytes is significantly enhanced, which is 1.77-fold (p < 0.05) that of the 0 μg/mL group.
4. Discussion
The acute phase serum amyloid A (SAA), a major acute-phase protein in vertebrates, plays a crucial role in the innate immune response and inflammation. It was isolated and named over 50 years ago and has a striking relationship to the acute phase response with serum levels rising as much as 1000-fold in 24 hours [8,15]. SAA proteins are encoded in a family of closely-related genes and have been remarkably conserved throughout vertebrate evolution[3]. However, the study of SAA in invertebrates is relatively nascent. Thought more and more components of the invertebrate immune system resembling vertebrate counterparts have been discovered, but the presence of SAA has been reported so far in a very limited number of invertebrates, especially in aquatic species. The present study identified the only saa gene (CgSAA) from oyster C. gigas, and its molecular characteristics, expression patter as well as possible functions were further investigated, with the aim to explore the evolution of SAAs and the possible role of CgSAA in the immune response.
There are varied number of SAA genes in different organisms, and SAA-related proteins and genes constitute a closely-related family. In human, there are 4 SAA genes identified and all of them are clustered on a single chromosome 11p [2]. In mice, there are 4 functional SAA genes and one pseudogene, which are located in chromosome 7. By genomic and transcriptomic analysis, 51 SAA-like proteins were retrieved in several protostome taxa comprising 21 marine bivalve species and basal metazoans [7]. Most of these organisms only harbor only one SAA gene. In oyster C. gigas, only one SAA gene was screened, which was consistent with that in its sister specie Crassostrea hongkongensis. Invertebrate SAA-like proteins share certain structural features with vertebrate SAA, including a conserved N-terminal region and a variable C-terminal region. These proteins are typically small, with molecular weights ranging from 10 to 20 kDa [4,7]. In oyster C. gigas, the open reading frame of CgSAA encoded a putative polypeptide of 138 amino acid residues with predicted molecular weight of 15.66 kDa. By structural prediction, CgSAA was found to contain a signal peptide (residues 1-22) and a structural SAA domain (residues 36-138). Multiple sequence comparisons showed that the SAA domain of the CgSAA gene shared high similarity with those of chordates and invertebrates, with the highest similarity (99%) to that of Crassostrea virginica [9]. Ten SAA-5 from various species of vertebrates and invertebrates were selected for phylogenetic analysis, CgSAA was clustered with CvSAA-5 from C. virginica. These results indicated that CgSAA was a conserved homolog of SAA family in oyster.
Functional studies have shown that invertebrate SAA-like proteins exhibit similar immune-modulatory activities to vertebrate SAA. In vertebrates, SAA proteins are mainly synthesized in the liver in response to immune challenges, such as bacterial or viral infections and are secreted into the hemolymph or other body fluids [4]. In invertebrates no specific liver tissue was evolved and developed, and the synthesis of SAA protein was mainly focused on immune-related tissues [22]. In the present study, CgSAA was detectable in all the sampled tissues of normal individuals but in low level, with relateive higher level in haemocytes, ganglion and mantle. While in oyster C. hongkongensis, ChSAA was lowest expressed in haemocytes but higher expressed in mantle and labial palps [9]. Though not highly expressed in normal condition, the expression of CgSAA was quickly un-regulated after immune stimulation, which was similar with that in vertebrates. Living in intertidal environment, oysters always face environmental stresses, such as high temperature and low oxygen [23]. The mass death caused by high temperature (summary mortality) and continuous rainfall (hypotonic death) has severely dampened the oyster industry, but the mechanism is still not clear [24]. In the present study, the expression of CgSAA was quickly un-regulated in oysters cultured under high temperature (34℃) stress or hypotonic stress (15‰). As an acute phase response protein, SAA in human can rise 1000-fold in 24–36 h following the initiating stimulus, which is commonly used as a diagnosing marker of infection or inflammation [25]. The prompt expression changes of CgSAA under infection and environmental stresses inferred that it could be used as a sensitive diagnosing marker of oyster disease.
The role of SAA proteins in immune responses is multifaceted. In vertebrates, they can bind to lipoproteins, enhance phagocytosis, and stimulate the production of immune mediators in mammalian [26]. Some SAA proteins have been shown to exhibit antimicrobial and antiviral activity, directly inhibiting the growth of pathogens. They can also modulate the production of cytokines and other immune mediators, thereby regulating the inflammatory response. While in invertebrates, the role of SAA protein in mediating immune response has not been fully uncovered, and only limited studies reported its expression changes under immune stimulation. In echinoderm, for example, SAA proteins have been shown to be induced in response to LPS stimulation or during intestinal regeneration [8,27]. These proteins are secreted into the hemolymph and bind to lipoproteins, forming complexes that are recognized by immune cells. In mollusks Tridacna crocea, SAA protein has been found to exhibit binding and antimicrobial activity against a range of bacteria pathogens, and could reduce the lethality rate of infected individuals [10]. In the present study, to explore the potential role of SAA in immune response, the recombinant rCgSAA was prepared and injected into oysters or incubated with primary cultured haemocytes. The expressions of cytokines including IL17-1, IL17-1 and TNF were all significantly induced after rCgSAA treatment, indicating the important role of CgSAA in inflammation. In oyster C. hongkongensis, the over-expression of ChSAA via transfection with a ChSAA expression vector led to significantly increased NF-kB activity in HEK293T cells [9]. In mammalian, the SAA was also found to display pro-inflammatory effect and could promote synovial macrophage activation and chronic arthritis via NFAT5.
Though the detail mechanism of CgSAA induced inflammation is still missing, it could be inferred that the NF-kB signal might play a role just as that in oyster C. hongkongensis. In oyster, the inflammation caused by cytokines including IL17-1, IL17-1 and TNF [20,28], plays an important role in mass mortality under stressful environment. Considering the high expression of CgSAA and serious mortality in oyster under high temperature stress or hypotonic stress, the CgSAA induced severe inflammation might be the main cause of summary mortality in oyster industry. The discovery of SAA function in oyster has opened up new possibilities for the development of disease-resistant strains and biotechnology applications.
5. Conclusions
In this study, the only gene CgSAA encoding SAA protein was identified in Crassostrea gigas and the results collectively suggested that CgSAA is a conserved acute phase response protein in oyster. In normal conditions, the mRNA expression level of CgSAA in individual Crassostrea gigas can be detected but remains at a low level. The expression level is the lowest in the tissue of the labial palp and slightly higher in haemocytes and was quickly induced under immune stimulation or environmental stress. By regulating the production of cytokines, CgSAA played an important role in severe inflammation induced under high temperature or hypotonic stress, which provided fresh ideas for understanding the roles of SAA proteins in disease occurrence of aquatic species.
Author Contributions
Junlei Cheng, Weilin Wang, Lingling Wang conceived and designed the experiment; Junlei Cheng, Xuemei Cheng to carry out experiments; Xuesong Li, Luyi Lun, Xingye Lian analysis data; Junlei Cheng, Weilin Wang wrote the manuscript; Weilin Wang, Lingling Wang, Linsheng Song revised the manuscript. All the authors approved the final publication of the manuscript.
Funding
The authors thank all laboratory members for their valuable discussions. This research was supported by grants from Dalian High Level Talent Innovation Support Program (2021RQ091), National Science Foundation of China (Nos. 32230110), earmarked fund (CARS-49), the Fund for Outstanding Talents and Innovative Team of Agricultural Scientific Research in MARA, the innovation team of Aquaculture Environment Safety from Liaoning Province (LT202009), and grants from Natural Science Foundation of Liaoning Province (Nos. 2023-MS-286).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original data supporting the conclusion of this paper will be provided by the author without reservation.
Acknowledgments
We are very grateful to all MBBL laboratory members for their technical advice and helpful discussions.
Conflicts of Interest
All the authors are affiliated with Dalian Ocean University. The authors declare that this research was conducted without any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Baumann, H.; Gauldie, J. The Acute Phase Response. Immunol Today 1994, 15, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Sack, G.H. Serum Amyloid A (SAA) Proteins. Subcell Biochem 2020, 94, 421–436. [Google Scholar] [CrossRef] [PubMed]
- Sack, G.H. Serum Amyloid A - a Review. Mol Med 2018, 24, 46. [Google Scholar] [CrossRef] [PubMed]
- Malle, E.; De Beer, F.C. Human Serum Amyloid A (SAA) Protein: A Prominent Acute-Phase Reactant for Clinical Practice. Eur J Clin Invest 1996, 26, 427–435. [Google Scholar] [CrossRef]
- Malle, E.; Steinmetz, A.; Raynes, J.G. Serum Amyloid A (SAA): An Acute Phase Protein and Apolipoprotein. Atherosclerosis 1993, 102, 131–146. [Google Scholar] [CrossRef] [PubMed]
- Cray, C.; Zaias, J.; Altman, N.H. Acute Phase Response in Animals: A Review. Comp Med 2009, 59, 517–526. [Google Scholar]
- Rosani, U.; Domeneghetti, S.; Gerdol, M.; Franzoi, M.; Pallavicini, A.; Venier, P. Serum Amyloid A in Marine Bivalves: An Acute Phase and Innate Immunity Protein. Dev Comp Immunol 2016, 59, 136–144. [Google Scholar] [CrossRef]
- Santiago-Cardona, P.G.; Berríos, C.A.; Ramírez, F.; García-Arrarás, J.E. Lipopolysaccharides Induce Intestinal Serum Amyloid A Expression in the Sea Cucumber Holothuria Glaberrima. Dev Comp Immunol 2003, 27, 105–110. [Google Scholar] [CrossRef]
- Qu, F.; Xiang, Z.; Yu, Z. The First Molluscan Acute Phase Serum Amyloid A (A-SAA) Identified from Oyster Crassostrea Hongkongensis: Molecular Cloning and Functional Characterization. Fish Shellfish Immunol 2014, 39, 145–151. [Google Scholar] [CrossRef]
- Wang, J.; Yang, Y.; Zhang, A.; Zeng, L.; Xiao, S.; Ma, H.; Li, J.; Mao, F.; Zhang, Y.; Zhang, Y.; et al. Serum Amyloid Protein (SAA) as a Healthy Marker for Immune Function in Tridacna Crocea. Fish Shellfish Immunol 2022, 122, 495–500. [Google Scholar] [CrossRef]
- Eckhardt, E.R.M.; Witta, J.; Zhong, J.; Arsenescu, R.; Arsenescu, V.; Wang, Y.; Ghoshal, S.; de Beer, M.C.; de Beer, F.C.; de Villiers, W.J.S. Intestinal Epithelial Serum Amyloid A Modulates Bacterial Growth in Vitro and Pro-Inflammatory Responses in Mouse Experimental Colitis. BMC Gastroenterol 2010, 10, 133. [Google Scholar] [CrossRef] [PubMed]
- King, W.L.; Jenkins, C.; Seymour, J.R.; Labbate, M. Oyster Disease in a Changing Environment: Decrypting the Link between Pathogen, Microbiome and Environment. Mar Environ Res 2019, 143, 124–140. [Google Scholar] [CrossRef] [PubMed]
- Petton, B.; Destoumieux-Garzón, D.; Pernet, F.; Toulza, E.; de Lorgeril, J.; Degremont, L.; Mitta, G. The Pacific Oyster Mortality Syndrome, a Polymicrobial and Multifactorial Disease: State of Knowledge and Future Directions. Front Immunol 2021, 12, 630343. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.-Z.; Fang, D.-Z.; Chen, F.-F.; Zhao, Q.-F.; Cai, C.-M.; Cheng, M.-G. Utilization of Recombinase Polymerase Amplification Method Combined with Lateral Flow Dipstick for Visual Detection of Respiratory Syncytial Virus. Mol Cell Probes 2020, 49, 101473. [Google Scholar] [CrossRef]
- Lavie, M.; Voisset, C.; Vu-Dac, N.; Zurawski, V.; Duverlie, G.; Wychowski, C.; Dubuisson, J. Serum Amyloid A Has Antiviral Activity against Hepatitis C Virus by Inhibiting Virus Entry in a Cell Culture System. Hepatology 2006, 44, 1626–1634. [Google Scholar] [CrossRef]
- Trevisan, R.; Mello, D.F.; Delapedra, G.; Silva, D.G.H.; Arl, M.; Danielli, N.M.; Metian, M.; Almeida, E.A.; Dafre, A.L. Gills as a Glutathione-Dependent Metabolic Barrier in Pacific Oysters Crassostrea Gigas: Absorption, Metabolism and Excretion of a Model Electrophile. Aquat Toxicol 2016, 173, 105–119. [Google Scholar] [CrossRef]
- Arredondo-Espinoza, R.; Ibarra, A.M.; Roberts, S.B.; Sicard-González, M.T.; Escobedo-Fregoso, C. Transcriptome Profile in Heat Resilient Pacific Oyster Crassostrea Gigas Families under Thermal Challenge. Comp Biochem Physiol Part D Genomics Proteomics 2023, 47, 101089. [Google Scholar] [CrossRef]
- Ribeiro, V.T.; Asevedo, E.A.; de Paiva Vasconcelos, L.T.C.; Filho, M.A.O.; de Araújo, J.S.; de Macedo, G.R.; de Sousa Júnior, F.C.; Dos Santos, E.S. Evaluation of Induction Conditions for Plasmid pQE-30 Stability and 503 Antigen of Leishmania i. Chagasi Expression in E. Coli M15. Appl Microbiol Biotechnol 2019, 103, 6495–6504. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.A.; Henderson, S.; Miller-Ezzy, P.; Li, X.X.; Qin, J.G. Immune Response to Temperature Stress in Three Bivalve Species: Pacific Oyster Crassostrea Gigas, Mediterranean Mussel Mytilus Galloprovincialis and Mud Cockle Katelysia Rhytiphora. Fish Shellfish Immunol 2019, 86, 868–874. [Google Scholar] [CrossRef]
- Wang, L.; Sun, J.; Wu, Z.; Lian, X.; Han, S.; Huang, S.; Yang, C.; Wang, L.; Song, L. AP-1 Regulates the Expression of IL17-4 and IL17-5 in the Pacific Oyster Crassostrea Gigas. Fish Shellfish Immunol 2020, 97, 554–563. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Liu, X.; Liu, D.; Yin, Z.; Yang, X.; Zeng, M. Oyster (Crassostrea Gigas) Polysaccharide Ameliorates High-Fat-Diet-Induced Oxidative Stress and Inflammation in the Liver via the Bile Acid-FXR-AMPKα Pathway. J Agric Food Chem 2022, 70, 8662–8671. [Google Scholar] [CrossRef]
- Zhang, Y.; Ni, M.; Bai, Y.; Shi, Q.; Zheng, J.; Cui, Z. Full-Length Transcriptome Analysis Provides New Insights Into the Diversity of Immune-Related Genes in Portunus Trituberculatus. Front Immunol 2022, 13, 843347. [Google Scholar] [CrossRef] [PubMed]
- Trevisan, R.; Mello, D.F. Redox Control of Antioxidants, Metabolism, Immunity, and Development at the Core of Stress Adaptation of the Oyster Crassostrea Gigas to the Dynamic Intertidal Environment. Free Radic Biol Med 2024, 210, 85–106. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Shi, C.; Yang, B.; Li, Q.; Liu, S. High Temperature Aggravates Mortalities of the Pacific Oyster (Crassostrea Gigas) Infected with Vibrio: A Perspective from Homeostasis of Digestive Microbiota and Immune Response. Aquaculture 2023, 568, 739309. [Google Scholar] [CrossRef]
- Abdullahi, A.; Stanojcic, M.; Yu, N.; Samadi, O.; Sadri, A.-R.; Vinaik, R.; Coburn, N.; Jeschke, M.G. Serum Amlyoid A: An Inflammatory Adipokine Mediating Postburn Outcomes. Clin Transl Med 2021, 11, e412. [Google Scholar] [CrossRef] [PubMed]
- Abouelasrar Salama, S.; Gouwy, M.; Van Damme, J.; Struyf, S. The Turning Away of Serum Amyloid A Biological Activities and Receptor Usage. Immunology 2021, 163, 115–127. [Google Scholar] [CrossRef]
- Lee, H.J.; Kim, J.H.; Kim, S.W.; Joo, H.A.; Lee, H.W.; Kim, Y.S.; Park, S.J.; Hong, S.P.; Kim, T.I.; Kim, W.H.; et al. Proteomic Analysis of Serum Amyloid A as a Potential Marker in Intestinal Behçet’s Disease. Dig Dis Sci 2017, 62, 1953–1962. [Google Scholar] [CrossRef] [PubMed]
- Kuret, T.; Peskar, D.; Kreft, M.E.; Erman, A.; Veranič, P. Comprehensive Transcriptome Profiling of Urothelial Cells Following TNFα Stimulation in an in Vitro Interstitial Cystitis/Bladder Pain Syndrome Model. Front Immunol 2022, 13, 960667. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Sequence features of CgSAA.
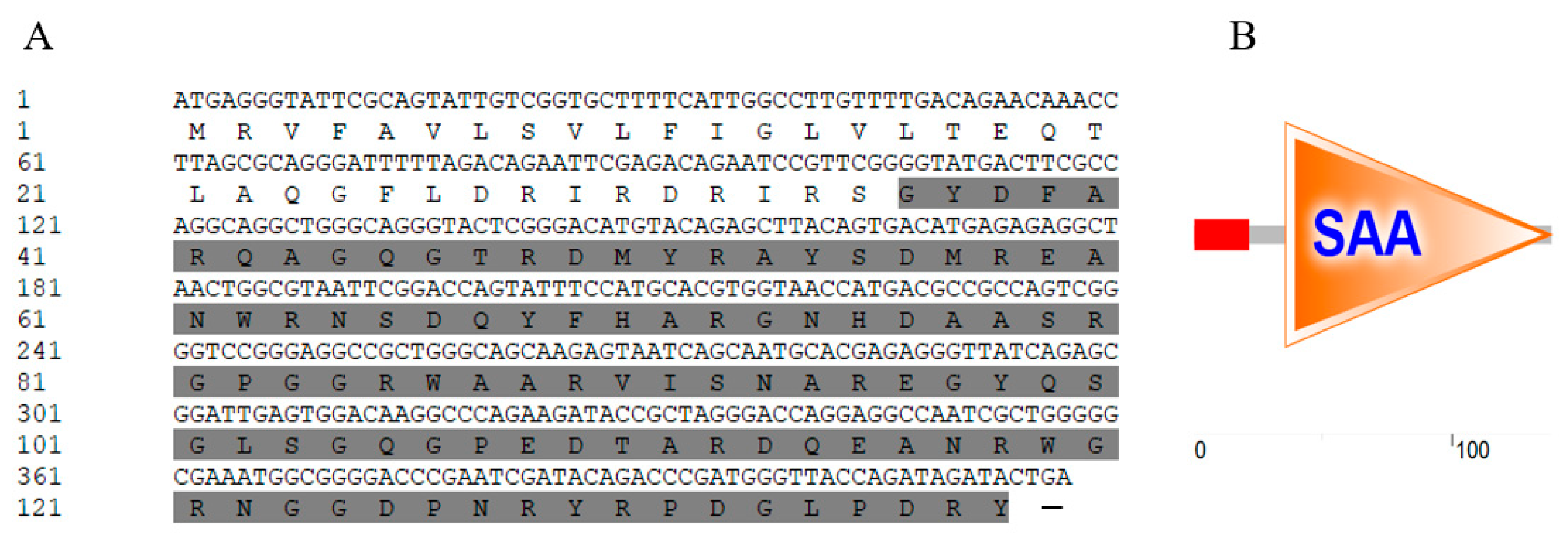
Figure 2.
Regarding CgSAA and its homologues, both the multiple sequence alignment and the phylogenetic tree were constructed.
Figure 2.
Regarding CgSAA and its homologues, both the multiple sequence alignment and the phylogenetic tree were constructed.
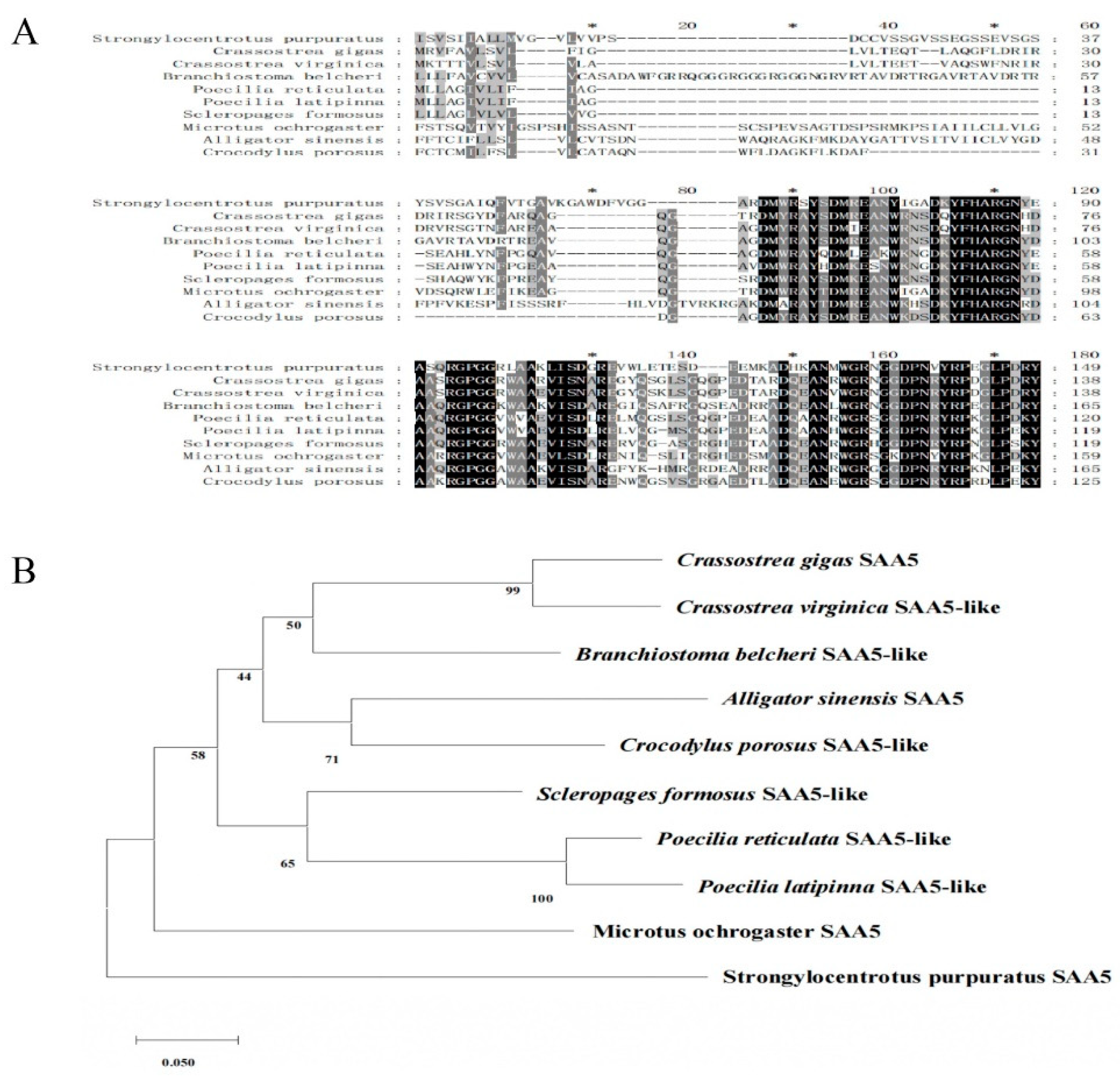
Figure 3.
The spatiotemporal expression pattern of CgSAA mRNA.
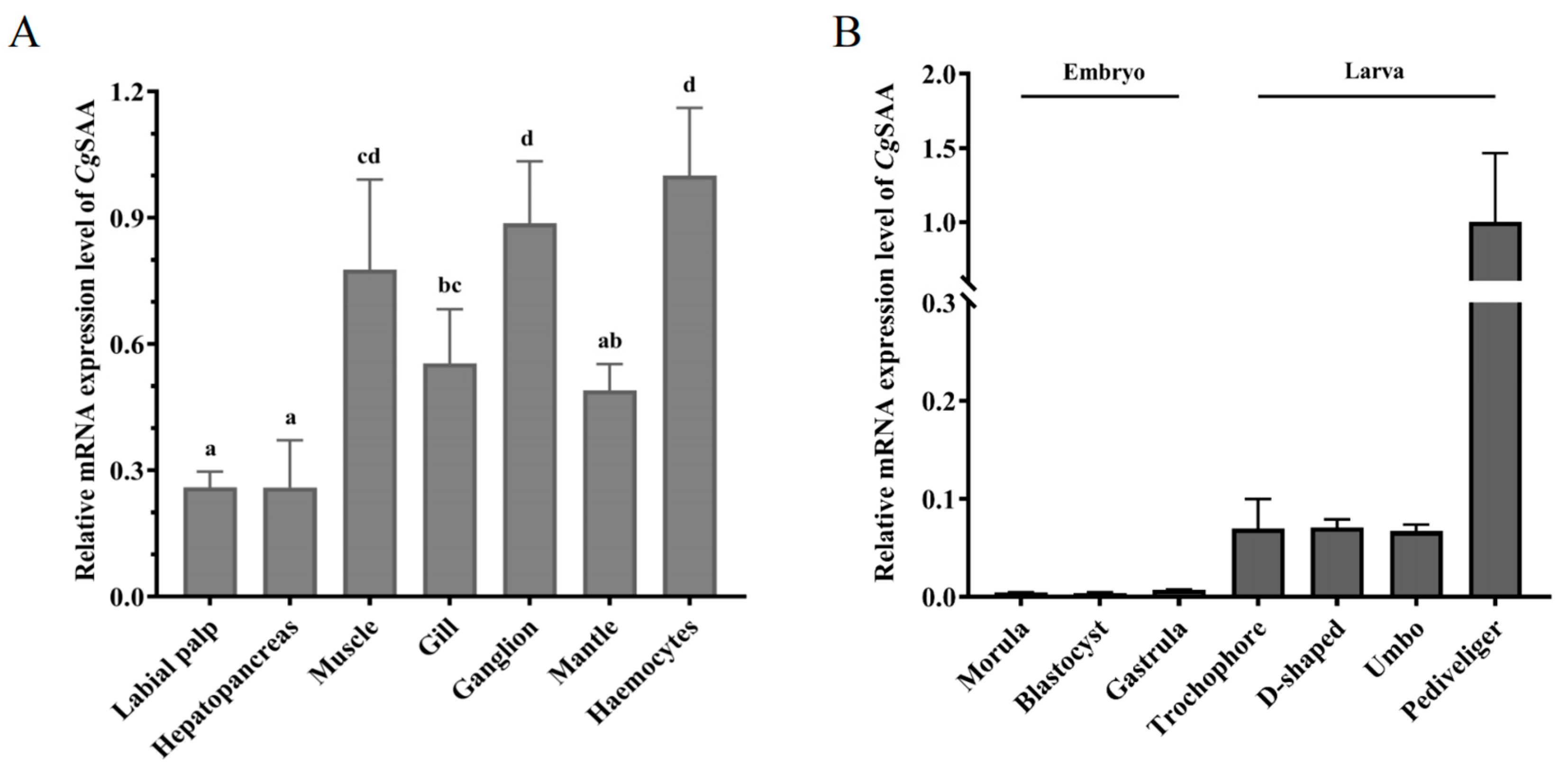
Figure 4.
The expression levels of CgSAA in the Crassostrea gigas after being stimulated by different environmental factors.
Figure 4.
The expression levels of CgSAA in the Crassostrea gigas after being stimulated by different environmental factors.

Figure 5.
SDS-PAGE analysis of rCgSAA. Lane M: standard protein molecular mass marker; Lane 1: negative control (without induction); Lane 2: induced rCgSAA; Lane 3: purified rCgSAA.
Figure 5.
SDS-PAGE analysis of rCgSAA. Lane M: standard protein molecular mass marker; Lane 1: negative control (without induction); Lane 2: induced rCgSAA; Lane 3: purified rCgSAA.
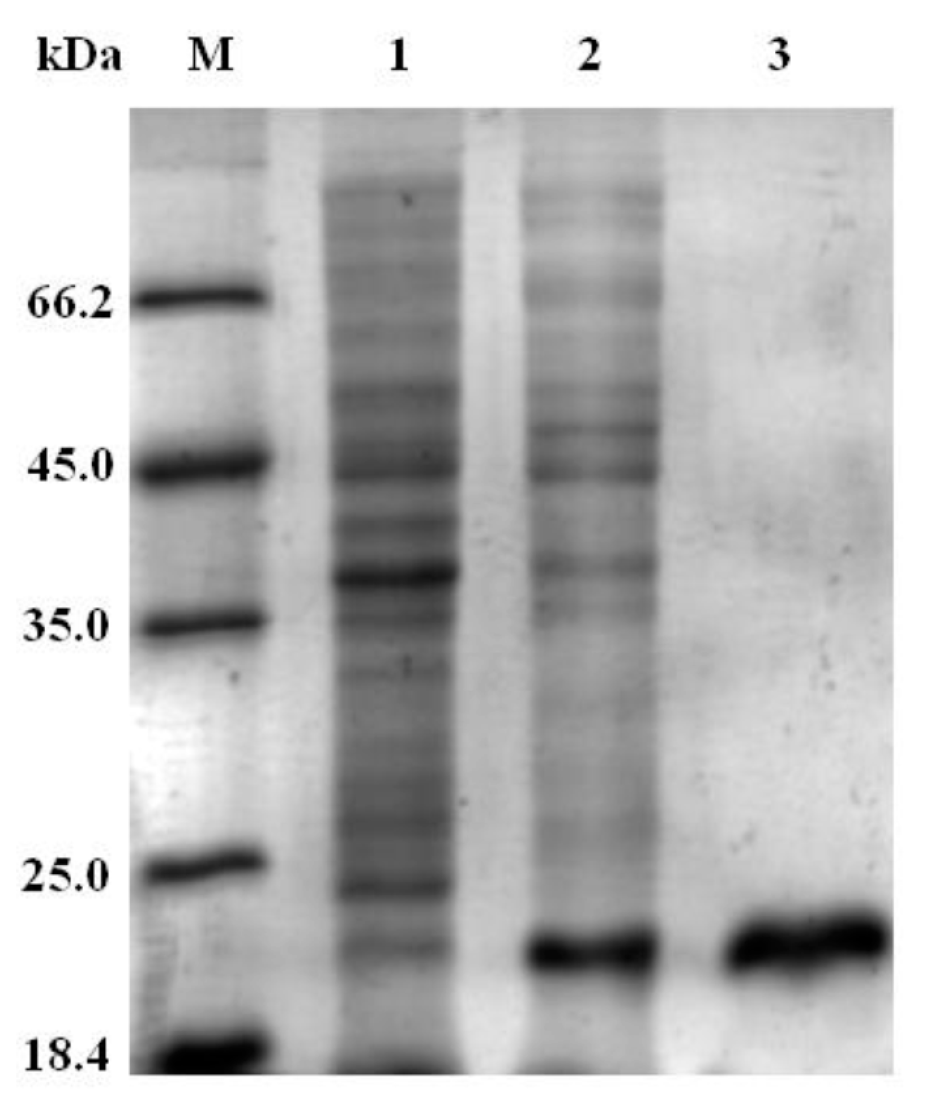
Figure 6.
The promoting effect of rCgSAA on cytokine gene expression.
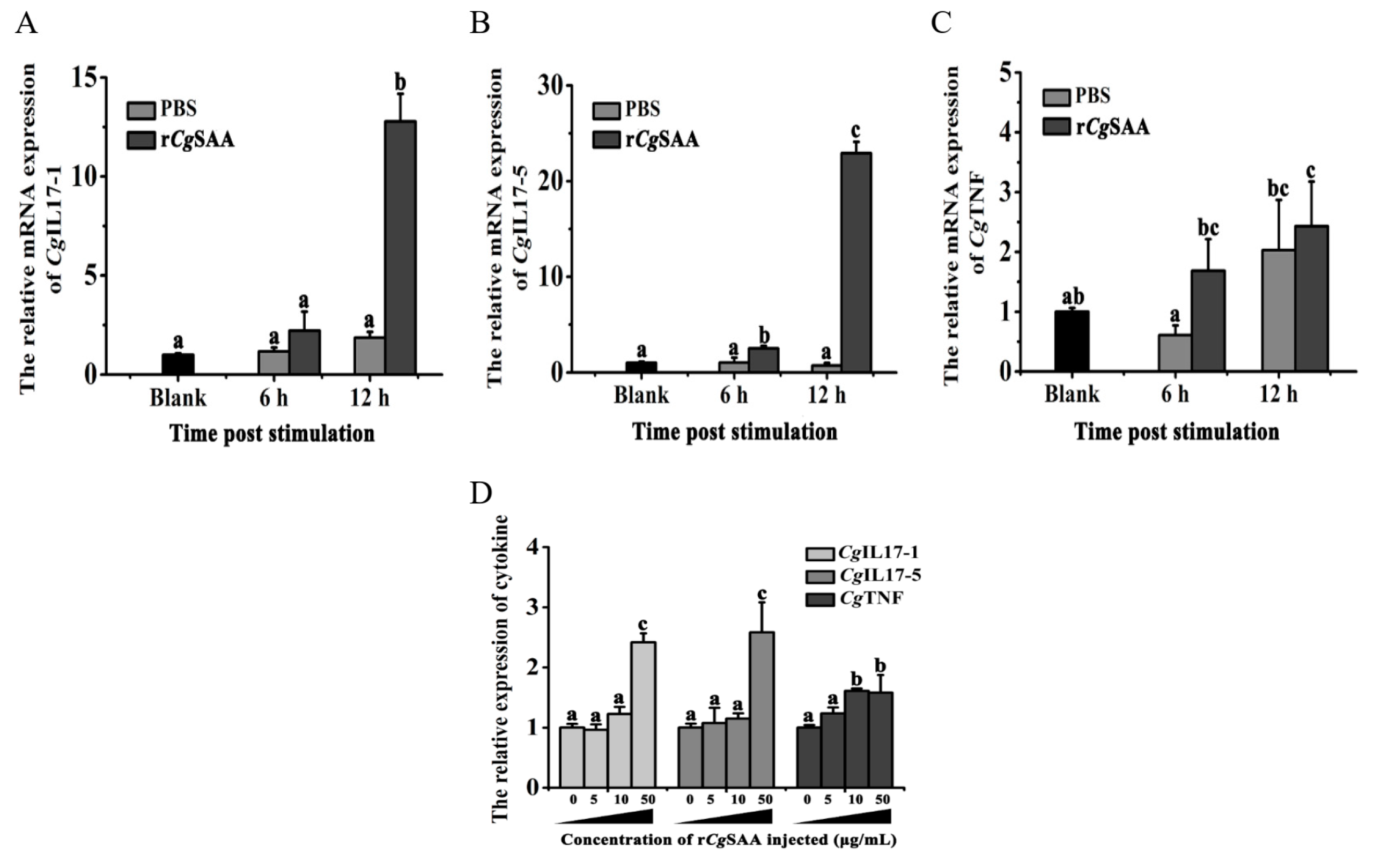

Table 1.
Primers used for RT-PCR in this study.
| Gene name | Primer Sequence (5'-3') |
|---|---|
| Clone primers | |
| CgSAA-F | ATGAGGGTATTCGCAGTA |
| CgSAA-R | CAGTATCTATCTGGTAACCC |
| Primers for recombinant expression | |
| CgSAA-EXF | CGCGGATCCCAGGGATTTTTAGACAGAATTCGAGAC |
| CgSAA-EXR | CCGCTCGAGGTATCTATCTGGTAACCCATCGGGTCT |
| RT-PCR primers | |
| CgIL17-1-RT-F | GCGAACGCCACAGTGTCAAA |
| CgIL17-1-RT-R | GACGCTACGAGGAAATACGGAC |
| CgIL17-5-RT-F | TCTGGCTGACTCTCGTCCTTG |
| CgIL17-5-RT-R | GACCCTGTCGTTGTCCTCTACC |
| CgILTNF-RT-F | CTTCTCGTCTGCGGCTTCTTT |
| CgILTNF-RT-R | CAGGGCTGCGGTCTTTCC |
| CgEF-RT-F | AGTCACCAAGGCTGCACAGAAAG |
| CgEF-RT-R | ACGACGATCGCATTTCTCTT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



