
Submitted:
28 November 2024
Posted:
29 November 2024
You are already at the latest version
Abstract
The world's production of butter generates 6.5.106 tons per year of buttermilk. This byproduct has important nutrients such as proteins, lactose, and salts. Proteins can be broken to produce peptides, which have biological activity as antioxidant and antimicrobial. This study aimed to obtain peptides of low molar weight from buttermilk by a combined process using enzymatic hydrolysis and ultrafiltration (UF). Different enzymes showed different hydrolysis degrees (HD), as the sequential hydrolysis with Alcalase® and Neutrase® reached a HD of 34 %. UF separated the peptides of low molar weight by applying 100 kPa (1 bar) of pressure and under 40 ºC. The UF membrane (with the molar weight cut off - MWCO in a range of 30 – 80 kDa) rejected 38 % of proteins from hydrolysed buttermilk. The ultrafiltered hydrolysed buttermilk has higher antioxidant activity when compared to the buttermilk. The results have shown that it is possible to obtain peptides with antioxidant activity allied to the valorization of a dairy byproduct from the dairy industry.
Keywords:
cleavage
; protein
; bioactivity
; peptide
; membranes
1. Introduction
Buttermilk, a by-product of butter production, is a liquid of yellow color released from the churning of cream milk and it contains proteins, fats, oligosaccharides, phospholipids, glycolipids and other constituents with biological activity [1]. Proteins can be submitted to hydrolysis to produce biopeptides [2], including proteins from buttermilk. Sakkas et al. [3] reported that the annual world production of buttermilk is 6.5.106 ton, and this byproduct is a good opportunity for valorization in different food markets.
Proteins in buttermilk contain encrypted peptides with bioactivity in their structure. These peptides are released in the digestion process, and after absorption and transport, they can participate in several metabolic routes and generate healthy benefits. However, as the peptides are trapped in the main structure of the protein as they have lesser biological activity. In addition, globular proteins in the buttermilk can be allergenic for some people because the enzymes of digestive system can’t reach the specific sites to break the protein chain. The enzymatic hydrolysis releases the peptides and reduces the allergenicity by the exposure of the chains and makes the digestion easier [4], and the HD and the molar weight of the peptides obtained in the hydrolysis are directly related with the reduction of allergenic effect [5]. The size of the peptides in a narrow range between 500 Daltons (Da) to 3000 Da is a specific factor that promotes antioxidant activity (AA) [6]. In contrast, small peptides can be a bitter taste [7] and the consumption can be limited. Furthermore, apart from the hydrolysis, we also can separate the peptides to obtain a higher biological activity. In fact, the downstream can be a barrier for a competitive production of any bioprocess because they could require expensive methods such as chromatography [8].
The membrane separation processes are quite suitable to separate the hydrolysed peptides [9]. UF has been used to purify the hydrolysed peptides from waste meat [10], milk [11], spent brewer yeast [8], microalgae biomass [12], soybean milk [13] and whey protein [14,15]. The hydrolysis and separation can be combined in a Membrane Bioreactor (MBR), where the transformation and the separation can be operated in an integrated process. The hydrolysis breaks the proteins in low peptides and the ultrafiltration membrane separates the high molar weight proteins (in retentate stream) from the low molar weight peptides (in permeate stream) [16]. Thus, MBR allows the control of the molar weight of products as the most suitable MWCO of the membrane was chosen [17,18].
The literature brings some works considered to valorization of buttermilk as the application of supercritical fluid to separate the constituents [19]. Additionally, buttermilk has phospholipids and hydrolysis has been tested to separate the protein from the milk fat globule membrane (MFGM). In fact, the literature has paid attention in the nutraceutical properties of MFGM of buttermilk [20,21,22,23,24,25,26] and recovery of the fat fraction of the buttermilk. Calvo et al. [27] studied a broad characterization of neutral and polar lipids from the MFGM in the buttermilk. Jahkar et al. [28] evaluated the development of fruit juices using buttermilk as beverages. Spitsberg et al. [26] recovery MFGM from buttermilk and evaluated the effect of salts and calcium bonds in the membrane globule. Barry et al. [29] search the production of phospholipids from buttermilk by the enzymatic hydrolysis and membrane separation. However, few works have focused on the recovery and valorization of the protein from buttermilk, and this paper aims to fulfill this scientific gap.
This work aimed to obtain peptides with AA of low molar weight from buttermilk in a membrane bioreactor, using ultrafiltration following enzymatic hydrolysis by the combination of two sequential enzymes.
2. Materials and Methods
The research followed three steps:
- Sampling and characterization of buttermilk;
- Batch experiments of protein hydrolysis using different enzymes to select the best enzyme for the operation of the MBR;
- Protein hydrolysis and separation of peptides in the MBR.
2.1. Sampling and Characterization of Buttermilk
Buttermilk was donated by Randon Company (Vacaria – Brazil). The buttermilk (30 L) was collected and transported in a cooled plastic vessel under 5 ºC. The physical-chemical characterization of the buttermilk was performed by a Fourier Transform Infrared (FT-IR) Lactoscope from Delta Instruments following the protocol 141C of the International Dairy Federation (IDF). The concentration (w/v) of the protein, fat and lactose in buttermilk was, respectively: 2.07±0.02%, 2.35±0.03%, 1.75±0.02%; pH was 6.35±0.10.
2.2. Batch Experiments of Protein Hydrolysis Using Different Enzymes
The tests to select the best enzymes to hydrolysate the protein in buttermilk were performed in reactors (beakers of 2 L in volume) in a stirred hot plate (model 501 from Fisatom – Brazil) to control of temperature. 1 L of buttermilk was inserted into a beaker and different concentrations for each enzyme (according to the suppliers’ instruction). NaOH 0.1 M (or lactic acid 0.5 M) were added to correction of pH under constant stirring.
The reaction was performed in the course of time at different conditions following the suppliers’ instruction. As amino acids are released in the reaction medium, the pH tends to reduce. However, the pH was kept under the ideal condition by adding 0.1 M NaOH. The reaction was finished by heating at 90 ºC.
The HD was performed by the pH-stat method suggested by Adler-Nissen [30] (Equation 1).
Where B is the volume of NaOH used to maintain the constant pH; N is the gram-equivalent of NaOH; Mp is the mass of protein (g); α is the dissociation degree of the amino acids; H is the number of total peptide bonds in the beginning;
The dissociation degree was predicted by Equation 2.
Where α is the dissociation degree; pK changes according to the temperature and pH is constant; pK was calculated by Equation 3, where T is the temperature (K).
Three enzymes were tested to hydrolyze the proteins in the buttermilk: the endopeptidase Alcalase® (EC3.4.21.62) and metalloprotease Neutrase® (EC3.4.22) from Novozymes (Bagsværd, Denmark) were tested. Both enzymes were gently donated by the company LNF Latin America (Brazil). The endopeptidase Prolyve (EC 3.4.21.62) from Soufflet Biotechnologies SAS (Colombelles, France) and donated by WGM Systems (Brazil) were also tested. The end of the reaction was noted by the constant pH in the course of time. Table 1 shows the reaction’s conditions of each tested enzyme.
After testing the hydrolysis by individual enzymes, we tested a sequence of two enzymes to improve the hydrolysis of proteins.
2.3. Enzymatic Hydrolysis and Separation of Peptides in the Membrane Bioreactor
2.3.1. MBR Operation
The MBR worked in batches. First, the hydrolysate was produced, and in sequence, the hydrolysate liquor was fed to the UF system as shown in Figure 1. The rig equipment was supplied by WGM Systems and it has a UF membrane (Koch Membrane Systems) made on polyacrylonitrile in tubular configuration, with an effective area of 0.1 m² and MWCO in range of 30 kDa – 80 kDa.
A chemical cleaning was performed before each experiment with (NaOH, pH 11) and rinses until neutral pH with Mili-Q® water. 10 L of hydrolysed buttermilk was fed into the stainless-steel tank (20 L in volume) and temperature were kept at 40 ºC to increase the permeate flux as temperature has a positive effect on permeation. A pneumatic pump forces the feed to the membrane and a valve was used to set the pressure on 100 kPa (1 bar) and checked in a manometer. The permeate (with the small peptides) was removed and the retentate (with the bigger proteins) returned to the feed tank. The time of separation of peptides was in range between 2 h and 4 h, and the total time (reaction + separation) was 6 h – 8 h.
The membrane rejection (MR) was calculated by Equation 4:
2.3.2. Peptide Characterization
The peptides were characterized by the prediction of the average length of the chain (ALC), the average molar weight (AVW) and by electrophoresis. In addition, the bioactivity of AA was also evaluated.
ALC was predicted by the concept of an amino acid has a molar weight of 120 Da, as reported by Perea et al. [31] in Equations 5 and 6.
The electrophoresis was performed by the Tris-Tricine-SDS-PAGE system as described by Jiang et al. (2016). The Mini-Protean II Cell from Bio-Rad with glasses plates with 6 cm x 8 cm and width of 1.0 mm was used.
The AA of the buttermilk and the hydrolysed was evaluated by the ABTS - 2,2′-Azino-bis(3-ethylbenzthiazoline-6-sulfonic acid) solution as method proposed by Re et al. [32]. 100 mL of solution 7 mM of ABTS was mixed with 100 mL of potassium persulfate 2.45 Mm to generate the ABTS•+ radical. This solution was then kept in dark for 16 h before use. Then, the solution was diluted with ethyl alcohol until reaching the absorbance of 0.7 ±0.05 in 734 nm of wavelength.
2.3.3. Data Analysis
All experiments were performed in two replicates. Analysis of variance (ANOVA) was used to determine significant differences (p <0.05) between experimental conditions and Tukey’s test to check differences between means.
3. Results
The HD obtained by enzymes Alcalase ®, Prolyve ®, Neutrase were 25.8%, 25.2%, 5.89%, respectively. Figure 2 shows the advance of the HD in the course of time for each enzyme.
Alcalase and Prolyve exhibit similar behavior to both endoproteases. Therefore, Alcalase is a serine-protease, with a differential in its sequence of amino acids and three-dimensional structure, presenting the catalytic triad Asparagine, Histidine and Serine at the active site. The substrates are linked to the serine-protease through the insertion of the amino acid adjacent to the cleavage site in a cavity at the active site of the enzyme. The exopeptidase, neutralized, is most active at neutral pH and has regions of α-helix (38%) and preheated β-sheet (17%) [33,34].
In order to increase HD, the Alcalase and Neutrase enzymes were used in sequence, and the HD is presented in Figure 3. The combination of the two enzymes with different action mechanisms promotes a higher HD (34.02 %) than only one enzyme.
The hydrolysis performed in a sequence of the Alcalase and Neutrase enzymes resulted in the best hydrolysis degree. Then, we decided to operate the MBR by using this sequence of enzymes. A new experiment (in two replicates) was performed under these conditions for the operation of the MBR. Figure 4 shows the permeate flux of the operation of the MBR. For comparison, the MBR also ran only with buttermilk.
The initial permeate flux of the hydrolysed buttermilk was higher (187 L/h.m²) and a strong decrease was observed until 15 L/h.m² after 120 min of operation. On the other hand, the operation of the membrane bioreactor with buttermilk showed a lower initial permeate flux, but a decrease was also observed, and after 120 min of operation the permeate flux was 23 L/h.m². Rejection of proteins by MBR was also different for the two situations. The protein rejection of the buttermilk by the UF membrane was 55.28%, while the rejection of the hydrolysed buttermilk was lower (38.46%). This fact indicates that hydrolysis reduces the size of the buttermilk proteins and a higher passage of proteins through the membrane pores were observed.
The concentration of protein, lactose, and fat in the permeate and retentate obtained by MBR, before and after the enzymatic hydrolysis process, is shown in Table 2.
The mass of protein in the feed was distributed into permeate and retentate streams. The concentration of proteins is higher in the permeate obtained with hydrolysed buttermilk. This phenomenon has been expected as its molecular size is small and pass through the membrane. The concentration of lactose and fat in both cases are similar and do not depends on the hydrolysis of protein.
Table 3 shows hydrolysis promoted an increase on Antioxidant Activity.
Hydrolysed samples with higher HD have shown an increase in AA when compared to non-hydrolysed buttermilk. Permeate (it has more small peptides) has shown the highest AA (956 μmol TE/g of protein) and retentate has the same AA than the hydrolysed buttermilk. The breakdown of the protein structure exposed amino acids and free peptides, previously unavailable in the native protein structure, results in a general increase in AA [35].
To evaluate the molecular mass of the peptides, the samples of buttermilk, buttermilk hydrolysed by Alcalase + Neutrase, the permeate and retentate were submitted to electrophoresis (Figure 5).
We can observe a strong mark in the electrophoretic profile of buttermilk (B) in 5 kDa and lighter marks in a range between 5 kDa and 100 kDa. On the other hand, hydrolysed streams (R, HB, P) have no marks above 5 kDa. Further discussion is presented in Section 4.
Table 4 shows the average package of the estimated rates for each sample of hydrolysed buttermilk.
We have progressive decrease in the molecular length chains and a molecular mass media as the HD increases. The final ALC using the Alcalase + Neutrase enzymes was 2.939.
4. Discussion
The different enzymes shown different levels of hydrolysis of the buttermilk proteins. Assessed separately, the Alcalase enzyme showed a maximum HD of 25.8% in 2 h of reaction. It is known that this enzyme mainly hydrolyzes peptide bonds containing hydrophobic residues on the carboxylic side [30]. Adamson and Reynolds [36] verified it in the hydrolysis of casein, where Alcalase® hydrolyzes preferentially peptide bonds containing residues of Glu, Met, Leu, Tyr, Lys and Gln on the carboxylic side. Pessato [37] reported an HD from 18.1% to 23.9% in the hydrolysis of whey. Perea et al. [31] used Alcalase to hydrolyze whey protein, and obtained 16% and 20% HD for the purified fractions of α-La and β-Lg (respectively) using the same concentration of enzyme:substrate of this work. However, buttermilk also contains casein, which seems to have an affinity with the enzyme Alcalase, according to Adamson and Reynolds [36].
The highest HD found for the Neutrase enzyme was 5.89% in 1.8 h of reaction. After this period, there was no further changes of the pH and we consider that there was no further hydrolysis. Neutrase is removing amino acids or pairs of amino acids from two ends of the polypeptide chain and is not efficient in the cleavage of intact proteins. Neutrase is indicated for joint application with an endoprotease [38]
The combination of enzymes (Alcalase + Neutrase) has different action mechanisms and specificities and presents a positive effect in the production of protein hydrolysed containing peptides with higher AA when compared to hydrolysates produced with enzymes in isolation [40]. The sequential use of Alcalse e Neutrase enzymes was chosen, with the premise that Alcalase activates the internal regions of the polypeptide chains, breaking the bonds with residues of aromatic, sulfurized, aliphatic, hydroxyl and basic amino acids on the carboxylic side. As it neutralizes the external regions, there are cleavage of the remaining bonds of carboxy-terminus (aromatic amino acids) and also affecting hydrophobic and aliphatic amino acids. The exopeptidases catalyze the cleavage of peptide bonds in the N or C terminal end of polypeptide chains and endopeptidases act preferentially in the internal regions of polypeptide chains. Otte et al. [41] tested several proteases in the hydrolysis of β-Lg, including: bromelain, trypsin, endoproteinase, aminopeptidase and carboxypeptidase. The standard hydrolysis conditions are 37 ºC and pH 8, except for carboxypeptidase (pH 6.5). β-Lg was rapidly hydrolysed into medium-sized peptides (1-5 kDa) and 49% HD. However, the proteins in buttermilk are a mixture of whey and casein proteins, and they are not a single protein specie. Thus, there will be different enzyme-substrate affinities, and an intermediate HD will be reached. Sabadin, Villas-Boas, Zollner et al. [57] reported MM peptides smaller than 6.5 kDa in β-Lg hydrolysed (produced with free Alcalase) contain approximately 12% of HD. Doucet, Otter, Gauthier [41] reports the formation of peptides smaller than 2 kDa from hydrolysis with Alcalase for 5 h.
Colbert and Decker [42] hydrolysed buttermilk with the enzyme Alcalase, and the antioxidant activity of hydrolysed buttermilk, expressed in the percentage of inhibition was 55.96 % and 60.64%. Therefore, hydrolysis increases the AA of buttermilk, and this is a cross-check of our results. Additionally, the operation of MBR with the hydrolysed buttermilk separated a permeate constituted of peptides with a lower molecular chain and an increased AA (Table 3). This process increased the biological activity of the proteins from this by-product of the dairy industry.
Mendis et al. [43] and Ferreira et al. [44] reported that the hydrolysis of peptides and peptides with lower average length chain have higher AA. Mendis et al. [43] suggested that the hydrophobicity and specific position of two residues of amino acids in the sequence of the peptide chain represent an important role in AA. Ferreira et al. [44] indicates that the reducing power (and, thus, the AA) of hydrolysed protein suggesting a direct relationship with the capacity to give electrons to free radicals and convert them into more stable products, interrupting reactions in the chain.
Regarding the operation of the MBR, we observed a high initial permeate flux followed by a strong decay. This behavior is characteristic of protein ultrafiltration processes [48]. Cheison, Wang and Xu [17] state that high recovery rates of non-permeated protein are closely related to its transport through membrane pores, and are also favored by the enzyme concentration, since hydrolysis increases permeation of protein. Guadix [49] reported that the soluble enzyme in MBR is not attached on the membrane surface and do not improve the fouling. The soluble enzyme remains in contact with the substrate, while the smaller products than the pore size of membrane permeate through the membrane and also contribute to membrane internal fouling. Leindecker [50] found that a UF membrane with a MWCO of 10 kDa presents higher rejection of milk proteins when compared to membranes of 30 and 50 kDa, but the authors do not reported the effect of hydrolysed proteins on this separation. Atra et al. [51] reported a range of 92% - 98% protein rejection with a membrane with 6-8 kDa of MWCO. We used an UF membrane with MWCO in a range of 30 – 80 kDa, and a lower protein rejection, but the hydrolysis reduced the length of the proteins and we observed a higher passage of them through the membrane.
Rombaut et al. [52] used UF (30 kDa) and MF (0.1 μm and 0.2 μm) membranes for separation of buttermilk. The protein rejection (82.5%) was similar to our work (87.55%). The researchers observed a rapid decrease in permeation due to the formation of a layer of polarization concentration, followed by a slow reduction of permeate flux caused by an increase of phospholipids in this layer. As a consequence, there was a strong fouling on the membrane surface.
Morin et al. [53] microfiltered buttermilk and reported that fouling agents as residual casein and milk protein, small fat globules, ionic calcium and MFGM material push the casein aggregates on the surface of the membrane, creating a porous layer that limits the permeation. We also observed a strong decay in the permeate flux in the UF of buttermilk. In the UF of hydrolysed buttermilk, the permeate flux was only 7% of the initial value and in the native buttermilk this percentage was 18%. We theorized that small peptides pass through the asymmetric layer of the UF membrane and fouled into membrane pores. This fact drives a stronger decay in the permeation.
Roesch and Corredig [54] recovered lipids from buttermilk by UF, and obtained an increase of total lipid from 8% to 16% in the retenated. Barry et al. [29] hydrolysed buttermilk by the combination of Neutrase and Alcalase and just Alcalase aiming to determine the best proteolytic enzyme to separate the phospholipids. The authors conclude that a HD of 19% digests proteins buttermilk sufficiently to allow permeation through a 50 kDa UF membrane, and thus recover the removed fats to use the phospholipids. There was fat passage to the permeate, because there is free fats in the buttermilk [55]. The free fats cannot be separated by skimming and the molecules are probably too small to be removed by the UF membranes, and they will be into the permeate. These works aimed to recovery the lipids, and the protein was not the aim of the studies. However, we observe that the combination of Alcalase + Neutrase + UF in MBR configuration represents a compact way to recovery peptides from buttermilk, and both streams retentate (with lipids) and permeate (with biopeptides) can be used with improved biological value.
Small peptides have higher antioxidant activity. In Table 4 we predicted the size of peptides using theoretical equations and we used the electrophoresis to help us to confirm the molar mass of the peptides. Abdelmoneim et al. [56] performed electrophoresis in buttermilk and explained the molecular markings in the following: α-lactoblobulin marks between 10 and 15 kDa; β-lactoblobulin is in range between 15 and 20 kDa; Caseins (β, α and ĸ) are between 20 and 40 kDa. In addition, above 50 kDa there are the MFGM proteins (butyrophilin, xanthine oxidase and dehydrogenase). Butyrophilin is a glycoprotein that represents more than 40% of the total protein mass of MFGM. Xanthine oxidase represents 12% of the total protein mass. Electrophoretic band B in Figure 5 shows that only the buttermilk protein still contains marks above 50 kDa and, thus, MFGM proteins with mainly phospholipids. After enzymatic hydrolysis, proteins and caseins derived from MFGM are hydrolysed in smaller peptides than 5 kDa and they are too small and, thus, cannot be retained in the gel. Furthermore, maybe the smaller peptides than 5 kDa (Table 4) elute out of the gel during electrophoresis, as also reported by Önay-Uçar et al. [45].
Electrophoresis showed that buttermilk contains proteins with different molecular masses, and a stronger labeling of MFGM proteins (phospholipids). Britten et al. [46] reported that MFGM proteins are responsible for 19% of the total protein in buttermilk, and Wong et al. [47] suggested that there is a strong contribution of MFGM proteins to the antioxidant capacity of buttermilk. In addition, native peptides from milk protein (α-lactalbumin and β-lactoglobulin) are also responsible for the AA of buttermilk. Caseins are more antioxidants than whey proteins as they have ability to capture iron and phosphoseril residues on the surface of casein micelles [25]. However, the reduction of molar mass of the proteins releases free chains to capture radicals, and, thus, increases the AA.
The ALC varied according to the HD. Hydrolysis with Alcalase + Neutrase showed an ALC of 2.939. This value suggests that the peptides formed are bi and tri-peptides. Guadix et al. [49] reported similar results: an ALC of 4.3 for a maximum DH of 23%, with 19.7% of the two peptides with a molar mass between 1 kDa and 5 kDa. In addition, 79% of peptides have molar mass lower than 1 kDa. Perea and Ugalde [58] also hydrolysed whey and they found 0.635 kDa and 4.2 for MMM and ALC, respectively. Furthermore, 27% of two peptides have molar mass between 1 kDa and 3 kDa and 73% were lower than 1 kDa. Therefore, our estimates are that the permeate from MBR has a low molar mass, and, a higher biological activity than the native buttermilk protein.
A last observation is that the hydrolysis of the proteins from buttermilk and their separation in a membrane bioreactor allows the use of the total content of protein from buttermilk (permeate and retentate) as both streams have higher bioactivity (AA) than the native buttermilk. Additionally, the retentate can be used for the valorization phospholipids, and total recovery of buttermilk in two high biological value products.
5. Conclusions
Peptides of low molar mass and higher bioactive properties were obtained from buttermilk. The permeate from the Membrane Bioreactor working with a combination of enzymes Alcalase and Neutrase has shown antioxidant activity of 956 μmol TE/g protein whilst native buttermilk has 356 μmol TE/g protein of antioxidant activity.
The use of buttermilk for the production of bioactive hydrolysed peptides can be an alternative for the valorisation of this by-product. This can expand the dairy products with health benefits to the consumer in the Latin industry.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, V.B.B. Methodology, V.B.B. and S.S.F.; Validation, V.B.B., S.S.F., S.G., J.S., I.A.F.; Formal analysis, V.B.B., S.S.F., J.S., I.A.F.; Investigation, V.B.B., S.S.F., S.G., F.V.T., G.H.T., D.T.N.; Resources, V.B.B., J.S., I.A.F.; Data curation, V.B.B., S.S.F; Writing—original draft preparation, S.S.F., V.B.B.; Writing—review and editing, V.B.B., S.G.; Visualization, V.B.B., S.G.; Supervision, V.B.B.; Project administration, V.B.B.; Funding acquisition, V.B.B. All authors have read and agreed to the published version of the manuscript.” Please turn to the CRediT taxonomy for the term explanation. Authorship must be limited to those who have contributed substantially to the work reported.
Funding
This research was partially funded by CAPES – Coordenação de Aperfeiçoamento de Pessoal de Nível Superior - Brazil, financial code 001.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study were derived from the following resources available in the public repository of the Brazilian thesis: http://tede.upf.br:8080/jspui/handle/tede/2063.
Acknowledgments
Author S.S.F. thanks CAPES for the scholarship.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Pereira, C.; Gomes, D.; Dias, S.; Santos, S.; Pires, A.; Viegas, J. Impact of Probiotic and Bioprotective Cultures on the Quality and Shelf Life of Butter and Buttermilk. Dairy 2024, Vol. 5, Pages 625-643 2024, 5, 625–643. [Google Scholar] [CrossRef]
- Astaire, J.C.; Ward, R.; German, J.B.; Jiménez-Flores, R. Concentration of Polar MFGM Lipids from Buttermilk by Microfiltration and Supercritical Fluid Extraction. J Dairy Sci 2003, 86, 2297–2307. [Google Scholar] [CrossRef] [PubMed]
- Sakkas, L.; Evageliou, V.; Igoumenidis, P.; Moatsou, G. Properties of Sweet Buttermilk Released from the Churning of Cream Separated from Sheep or Cow Milk or Sheep Cheese Whey: Effect of Heat Treatment and Storage of Cream. Foods 2022, Vol. 11, Page 465 2022, 11, 465. [Google Scholar] [CrossRef] [PubMed]
- Golkar, A.; Milani, J.M.; Vasiljevic, T. Altering Allergenicity of Cow’s Milk by Food Processing for Applications in Infant Formula. Crit Rev Food Sci Nutr 2019, 59, 159–172. [Google Scholar] [CrossRef]
- Xu, D.; Yuan, F.; Jiang, J.; Wang, X.; Hou, Z.; Gao, Y. Structural and Conformational Modification of Whey Proteins Induced by Supercritical Carbon Dioxide. Innovative Food Science and Emerging Technologies 2011, 12, 32–37. [Google Scholar] [CrossRef]
- Cheung, I.W.Y.; Cheung, L.K.Y.; Tan, N.Y.; Li-Chan, E.C.Y. The Role of Molecular Size in Antioxidant Activity of Peptide Fractions from Pacific Hake (Merluccius Productus) Hydrolysates. Food Chem 2012, 134, 1297–1306. [Google Scholar] [CrossRef]
- Zhou, P.; Yang, C.; Ren, Y.; Wang, C.; Tian, F. What Are the Ideal Properties for Functional Food Peptides with Antihypertensive Effect? A Computational Peptidology Approach. Food Chem 2013, 141, 2967–2973. [Google Scholar] [CrossRef]
- Marson, G.V.; Belleville, M.P.; Lacour, S.; Hubinger, M.D. Membrane Fractionation of Protein Hydrolysates from By-Products: Recovery of Valuable Compounds from Spent Yeasts. Membranes 2021, Vol. 11, Page 23 2020, 11, 23. [Google Scholar] [CrossRef]
- Faucher, M.; Geoffroy, T.R.; Thibodeau, J.; Gaaloul, S.; Bazinet, L. Semi-Industrial Production of a DPP-IV and ACE Inhibitory Peptide Fraction from Whey Protein Concentrate Hydrolysate by Electrodialysis with Ultrafiltration Membrane. Membranes 2022, Vol. 12, Page 409 2022, 12, 409. [Google Scholar] [CrossRef]
- Sbeghen, A.L.; Lira, A.L.; Fernandes, I.A.; Steffens, C.; Brião, V.B.; Zeni, J.; Steffens, J. Use of Ultrafiltration in the Separation of Hydrolysates from Mechanically Separated Chicken Meat and Evaluation of Antioxidant Activity. J Food Process Eng 2022, 45, e14151. [Google Scholar] [CrossRef]
- Nath, A.; Csighy, A.; Eren, B.A.; Nugraha, D.T.; Pásztorné-Huszár, K.; Tóth, A.; Takács, K.; Szerdahelyi, E.; Kiskó, G.; Kovács, Z.; et al. Bioactive Peptides from Liquid Milk Protein Concentrate by Sequential Tryptic and Microbial Hydrolysis. Processes 2021, 9. [Google Scholar] [CrossRef]
- Sossella, F.; Rempel, A.; Monroe Araújo Nunes, J.; Biolchi, G.; Migliavaca, R.; Farezin Antunes, A.C.; Vieira Costa, J.A.; Hemkemeier, M.; Colla, L.M. Effects of Harvesting Spirulina Platensis Biomass Using Coagulants and Electrocoagulation–Flotation on Enzymatic Hydrolysis. Bioresour Technol 2020, 311, 123526. [Google Scholar] [CrossRef]
- Nath, A.; Kailo, G.G.; Mednyánszky, Z.; Kiskó, G.; Csehi, B.; Pásztorné-Huszár, K.; Gerencsér-Berta, R.; Galambos, I.; Pozsgai, E.; Bánvölgyi, S.; et al. Antioxidant and Antibacterial Peptides from Soybean Milk through Enzymatic- and Membrane-Based Technologies. Bioengineering 2020, Vol. 7, Page 5 2019, 7, 5. [Google Scholar] [CrossRef] [PubMed]
- O’Loughlin, I.B.; Murray, B.A.; Brodkorb, A.; FitzGerald, R.J.; Robinson, A.A.; Holton, T.A.; Kelly, P.M. Whey Protein Isolate Polydispersity Affects Enzymatic Hydrolysis Outcomes. Food Chem 2013, 141, 2334–2342. [Google Scholar] [CrossRef]
- Yu, X.X.; Liang, W.Y.; Yin, J.Y.; Zhou, Q.; Chen, D.M.; Zhang, Y.H. Combining Experimental Techniques with Molecular Dynamics to Investigate the Impact of Different Enzymatic Hydrolysis of β-Lactoglobulin on the Antigenicity Reduction. Food Chem 2021, 350. [Google Scholar] [CrossRef]
- Swaminathan, A.V.; Molitor, M.S.; Burrington, K.J.; Otter, D.; Lucey, J.A. Partial Enrichment of Phospholipids by Enzymatic Hydrolysis and Membrane Filtration of Whey Protein Phospholipid Concentrate. JDS Communications 2023, 4, 175–180. [Google Scholar] [CrossRef]
- Cheison, S.C.; Wang, Z.; Xu, S.Y. Hydrolysis of Whey Protein Isolate in a Tangential Flow Filter Membrane Reactor. II. Characterisation for the Fate of the Enzyme by Multivariate Data Analysis. J Memb Sci 2006, 286, 322–332. [Google Scholar] [CrossRef]
- Prieto, C.A.; Guadix, A.; González-Tello, P.; Guadix, E.M. A Cyclic Batch Membrane Reactor for the Hydrolysis of Whey Protein. J Food Eng 2007, 78, 257–265. [Google Scholar] [CrossRef]
- García-Cano, I.; Yeh, P.W.; Rocha-Mendoza, D.; Jiménez-Flores, R. Supercritical CO2 Treatment Reduces the Antigenicity of Buttermilk β-Lactoglobulin and Its Inflammatory Response in Caco-2 Cells. JDS Communications 2021, 2, 1–6. [Google Scholar] [CrossRef]
- Anto, L.; Warykas, S.W.; Torres-Gonzalez, M.; Blesso, C.N. Milk Polar Lipids: Underappreciated Lipids with Emerging Health Benefits. Nutrients 2020, 12. [Google Scholar] [CrossRef]
- Bourlieu, C.; Michalski, M.C. Structure-Function Relationship of the Milk Fat Globule. Curr Opin Clin Nutr Metab Care 2015, 18, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Contarini, G.; Povolo, M. Phospholipids in Milk Fat: Composition, Biological and Technological Significance, and Analytical Strategies. Int J Mol Sci 2013, 14, 2808. [Google Scholar] [CrossRef] [PubMed]
- Rombaut, R.; Dejonckheere, V.; Dewettinck, K. Filtration of Milk Fat Globule Membrane Fragments from Acid Buttermilk Cheese Whey. J Dairy Sci 2007, 90, 1662–1673. [Google Scholar] [CrossRef] [PubMed]
- Fontecha, J.; Brink, L.; Wu, S.; Pouliot, Y.; Visioli, F.; Jiménez-Flores, R. Sources, Production, and Clinical Treatments of Milk Fat Globule Membrane for Infant Nutrition and Well-Being. Nutrients 2020, 12. [Google Scholar] [CrossRef]
- Ries, D.; Ye, A.; Haisman, D.; Singh, H. Antioxidant Properties of Caseins and Whey Proteins in Model Oil-in-Water Emulsions. Int Dairy J 2010, 20, 72–78. [Google Scholar] [CrossRef]
- Spitsberg, V.L.; Ivanov, L.; Shritz, V. Recovery of Milk Fat Globule Membrane (MFGM) from Buttermilk: Effect of Ca-Binding Salts. J Dairy Res 2019, 86, 374–376. [Google Scholar] [CrossRef]
- Calvo, M.V.; Martín-Hernández, M.C.; García-Serrano, A.; Castro-Gómez, M.P.; Alonso-Miravalles, L.; García-Martín, R.; Megino-Tello, J.; Alonso, L.; Fontecha, J. Comprehensive Characterization of Neutral and Polar Lipids of Buttermilk from Different Sources and Its Milk Fat Globule Membrane Isolates. Journal of Food Composition and Analysis 2020, 86, 103386. [Google Scholar] [CrossRef]
- Jakhar, M.; Jain, M. Development of Buttermilk with Fruit Juices and Appraisal of Their Nutritional Qualities. The Pharma Innovation Journal 2019, 8, 404–407. [Google Scholar]
- Barry, K.M.; Dinan, T.G.; Kelly, P.M. Selective Enrichment of Dairy Phospholipids in a Buttermilk Substrate through Investigation of Enzymatic Hydrolysis of Milk Proteins in Conjunction with Ultrafiltration. Int Dairy J 2017, 68, 80–87. [Google Scholar] [CrossRef]
- Adler-Nissen, J. Methods in Food Protein Hydrolysis. In: Enzymic Hydrolysis of Food Protein. Enzymic hydrolysis of food proteins 1986, 110–130. [Google Scholar]
- Perea, A.; Ugalde, U.; Rodriguez, I.; Serra, J.L. Preparation and Characterization of Whey Protein Hydrolysates: Applications in Industrial Whey Bioconversion Processes. Enzyme Microb Technol 1993, 15, 418–423. [Google Scholar] [CrossRef] [PubMed]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant Activity Applying an Improved ABTS Radical Cation Decolorization Assay. Free Radic Biol Med 1999, 26, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Whitaker, J.R. Principles of Enzymology for the Food Sciences. Principles of Enzymology for the Food Sciences 2018. [Google Scholar] [CrossRef]
- Beynon, R.J.; Bond, J.S. Proteolytic Enzymes: A Practical Approach. 1989, 259.
- Conway, V.; Gauthier, S.F.; Pouliot, Y. Antioxidant Activities of Buttermilk Proteins, Whey Proteins, and Their Enzymatic Hydrolysates. J Agric Food Chem 2013, 61, 364–372. [Google Scholar] [CrossRef]
- Adamson, N.J.; Reynolds, E.C. Characterization of Casein Phosphopeptides Prepared Using Alcalase: Determination of Enzyme Specificity. Enzyme Microb Technol 1996, 19, 202–207. [Google Scholar] [CrossRef]
- Pessato, T.B.; de Carvalho, N.C.; Tavano, O.L.; Fernandes, L.G.R.; Zollner, R. de L.; Netto, F.M. Whey Protein Isolate Hydrolysates Obtained with Free and Immobilized Alcalase: Characterization and Detection of Residual Allergens. Food Research International 2016, 83, 112–120. [Google Scholar] [CrossRef]
- Saha, B.C.; Hayashi, K. Debittering of Protein Hydrolyzates. Biotechnol Adv 2001, 19, 355–370. [Google Scholar] [CrossRef]
- Motta, J.F.G.; de FREITAS, B.C.B.; de ALMEIDA, A.F.; Martins, G.A. de S.; Borges, S.V. Use of Enzymes in the Food Industry: A Review. Food Science and Technology 2023, 43, e106222. [CrossRef]
- Ceylan, F.D.; Adrar, N.; Günal-Köroǧlu, D.; Subas, B.G.; Capanoglu, E. Combined Neutrase–Alcalase Protein Hydrolysates from Hazelnut Meal, a Potential Functional Food Ingredient. ACS Omega 2022, 8, 1618. [Google Scholar] [CrossRef]
- Doucet, D.; Gauthier, S.F.; Otter, D.E.; Foegeding, E.A. Enzyme-Induced Gelation of Extensively Hydrolysed Whey Proteins by Alcalase: Comparison with the Plastein Reaction and Characterization of Interactions. J Agric Food Chem 2003, 51, 6036–6042. [Google Scholar] [CrossRef]
- Colbert, L.B.; Decker, E.A. Antioxidant Activity of an Ultrafiltration Permeate from Acid Whey. J Food Sci 1991, 56, 1248–1250. [Google Scholar] [CrossRef]
- Mendis, E.; Rajapakse, N.; Kim, S.K. Antioxidant Properties of a Radical-Scavenging Peptide Purified from Enzymatically Prepared Fish Skin Gelatin Hydrolysate. J Agric Food Chem 2005, 53, 581–587. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, I.M.P.L.V.O.; Pinho, O.; Mota, M.V.; Tavares, P.; Pereira, A.; Gonçalves, M.P.; Torres, D.; Rocha, C.; Teixeira, J.A. Preparation of Ingredients Containing an ACE-Inhibitory Peptide by Tryptic Hydrolysis of Whey Protein Concentrates. Int Dairy J 2007, 17, 481–487. [Google Scholar] [CrossRef]
- Önay-Uçar, E.; Arda, N.; Pekmez, M.; Yilmaz, A.M.; Böke-Sarikahya, N.; Kirmizigül, S.; Yalçin, A.S. Comparison of Antioxidant Capacity, Protein Profile and Carbohydrate Content of Whey Protein Fractions. Food Chem 2014, 150, 34–40. [Google Scholar] [CrossRef]
- Britten, M.; Lamothe, S.; Robitaille, G. Effect of Cream Treatment on Phospholipids and Protein Recovery in Butter-Making Process. Int J Food Sci Technol 2008, 43, 651–657. [Google Scholar] [CrossRef]
- Wong, P.Y.Y.; Kitts, D.D. Chemistry of Buttermilk Solid Antioxidant Activity. J Dairy Sci 2003, 86, 1541–1547. [Google Scholar] [CrossRef]
- Cheryan, M. Ultrafiltration and Microfiltration Handbook. Ultrafiltration and Microfiltration Handbook 1998. [Google Scholar] [CrossRef]
- Guadix, A.; Camacho, F.; Guadix, E.M. Production of Whey Protein Hydrolysates with Reduced Allergenicity in a Stable Membrane Reactor. J Food Eng 2006, 72, 398–405. [Google Scholar] [CrossRef]
- Leindecker, G.C. Separação Das Proteínas Do Soro Do Leite in Natura Por Ultrafiltração. 2011.
- Atra, R.; Vatai, G.; Bekassy-Molnar, E.; Balint, A. Investigation of Ultra- and Nanofiltration for Utilization of Whey Protein and Lactose. J Food Eng 2005, 67, 325–332. [Google Scholar] [CrossRef]
- Rombaut, R.; Dejonckheere, V.; Dewettinck, K. Filtration of Milk Fat Globule Membrane Fragments from Acid Buttermilk Cheese Whey. J Dairy Sci 2007, 90, 1662–1673. [Google Scholar] [CrossRef]
- Morin, P.; Britten, M.; Jiménez-Flores, R.; Pouliot, Y. Microfiltration of Buttermilk and Washed Cream Buttermilk for Concentration of Milk Fat Globule Membrane Components. J Dairy Sci 2007, 90, 2132–2140. [Google Scholar] [CrossRef] [PubMed]
- Corredig, M.; Roesch, R.R.; Dalgleish, D.G. Production of a Novel Ingredient from Buttermilk. J Dairy Sci 2003, 86, 2744–2750. [Google Scholar] [CrossRef] [PubMed]
- Mulder, H.; Walstra, P. The Milk Fat Globule Emulsion Science as Applied to Milk Products and Comparable Foods; 4th ed.; Commonwealth Agricultural Bureaux: Farnham Royal, 1974; Vol. 19;
- Ali, A.H. Current Knowledge of Buttermilk: Composition, Applications in the Food Industry, Nutritional and Beneficial Health Characteristics. Int J Dairy Technol 2019, 72, 169–182. [Google Scholar] [CrossRef]
- Sabadin, I.S.; Villas-Boas, M.B.; de Lima Zollner, R.; Netto, F.M. Effect of Combined Treatment of Hydrolysis and Polymerization with Transglutaminase on Beta-Lactoglobulin Antigenicity. EUROPEAN FOOD RESEARCH AND TECHNOLOGY 2012, 235, 801. [Google Scholar] [CrossRef]
- Perea, A.; Ugalde, U. Continuous Hydrolysis of Whey Proteins in a Membrane Recycle Reactor. Enzyme Microb Technol 1996, 18, 29–34. [Google Scholar] [CrossRef]
Figure 1.
Scheme of the Membrane Bioreactor for production of peptides from proteins in buttermilk.
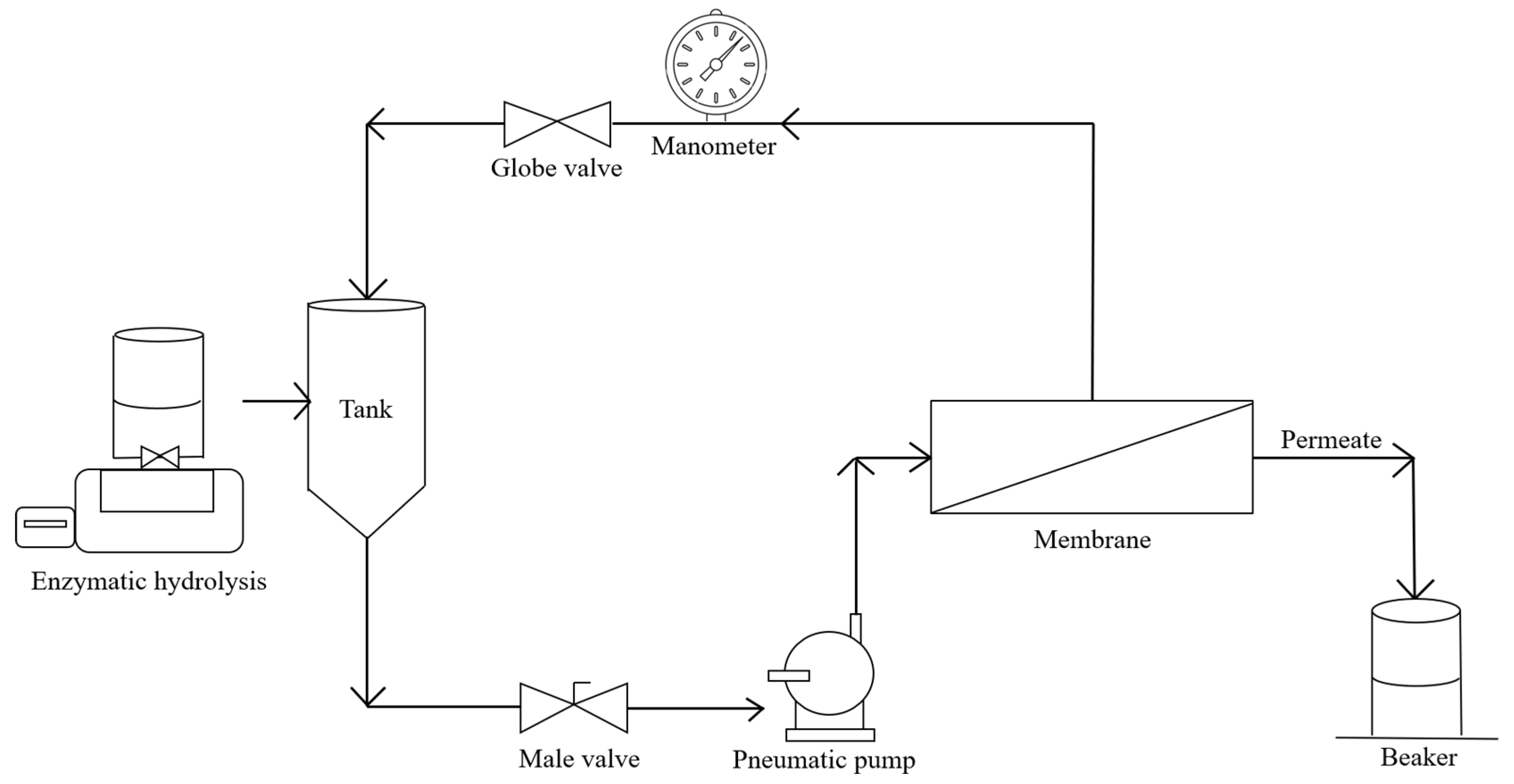
Figure 2.
Hydrolysis Degree (HD) of buttermilk protein using different enzymes: (a) Alcalase®; (b) Prolyve®; (c) Neutrase.
Figure 2.
Hydrolysis Degree (HD) of buttermilk protein using different enzymes: (a) Alcalase®; (b) Prolyve®; (c) Neutrase.
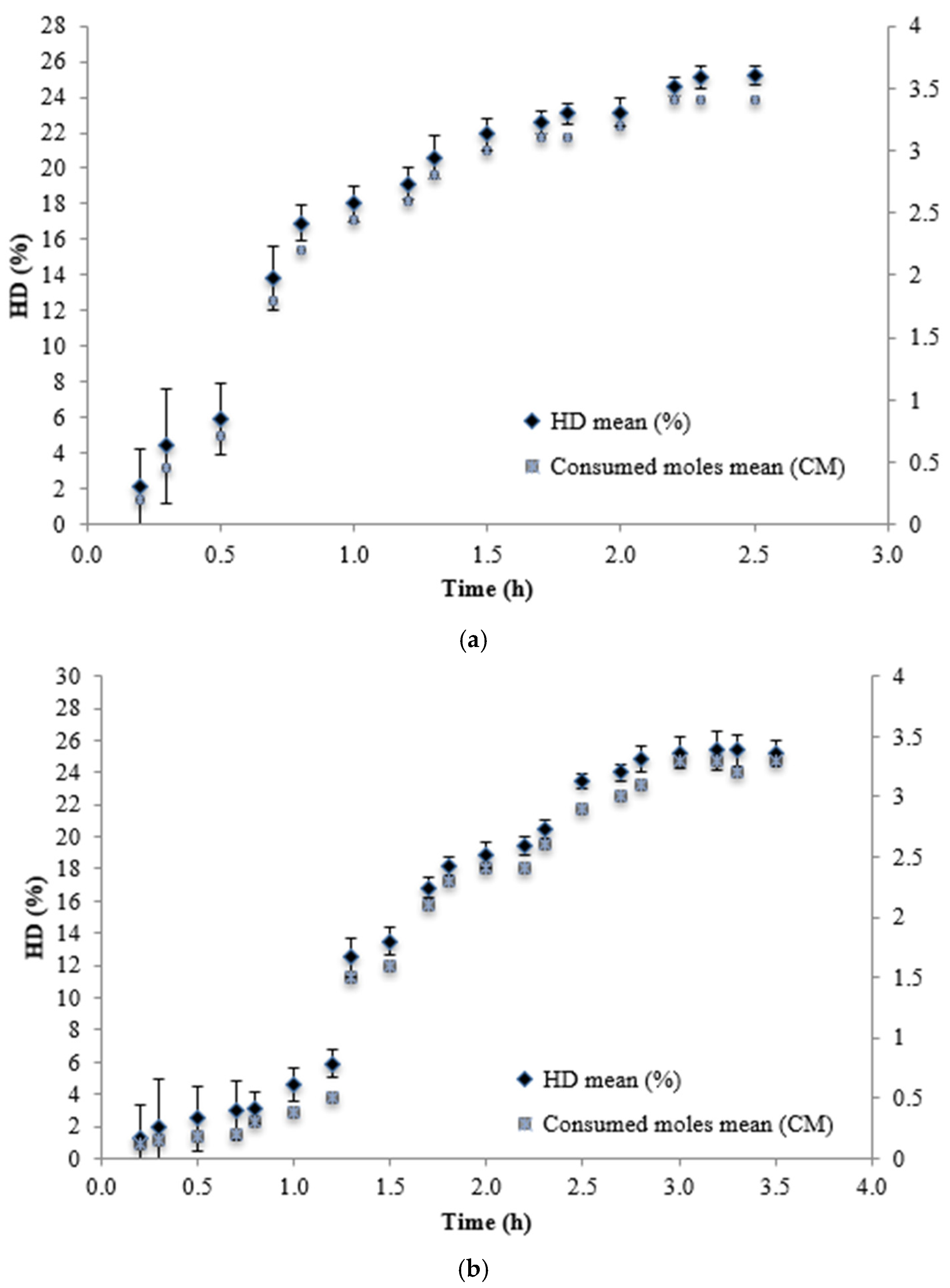
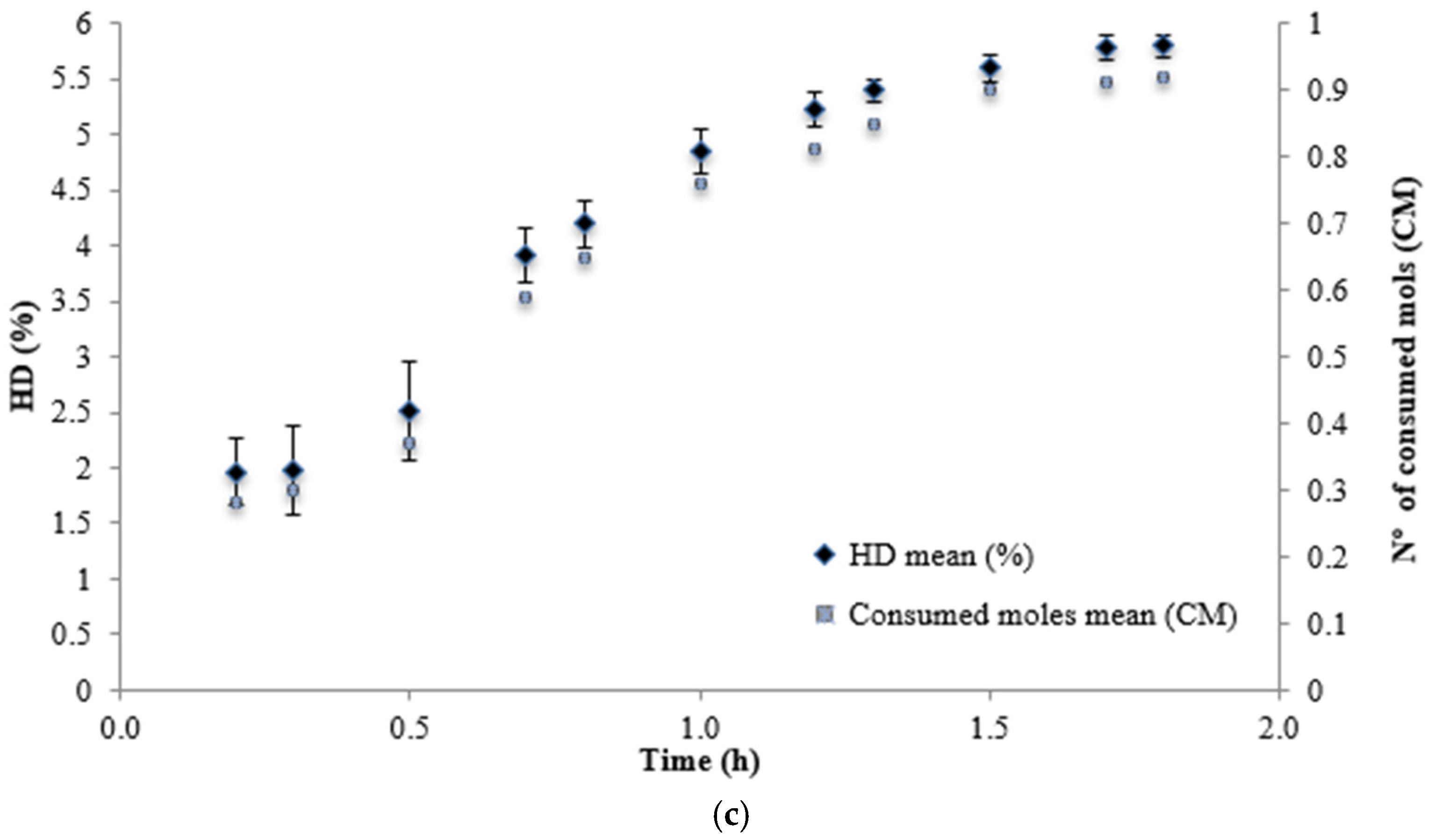
Figure 3.
Hydrolysis Degree (HD) of buttermilk protein using sequential: Alcalase® until 2.5 h followed by Neutrase after 2,5 h.
Figure 3.
Hydrolysis Degree (HD) of buttermilk protein using sequential: Alcalase® until 2.5 h followed by Neutrase after 2,5 h.
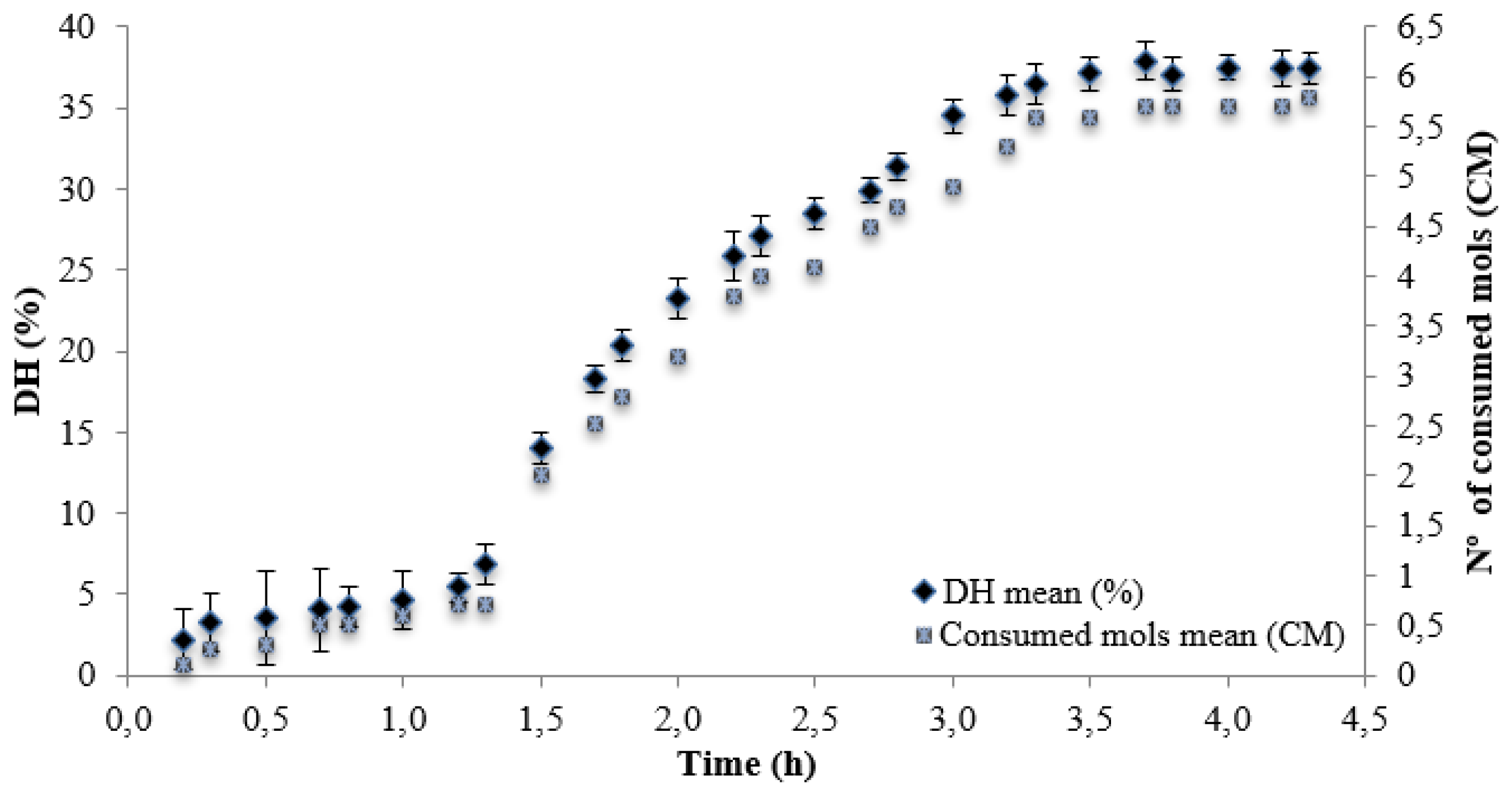
Figure 4.
Permeate flux operating on membrane bioreactor with: (a) Buttermilk without hydrolysis; (b) Hydrolysed Buttermilk sequentially with the enzymes Alcalase and Neutrase. Pressure: 100 kPa, T = 40 ºC.
Figure 4.
Permeate flux operating on membrane bioreactor with: (a) Buttermilk without hydrolysis; (b) Hydrolysed Buttermilk sequentially with the enzymes Alcalase and Neutrase. Pressure: 100 kPa, T = 40 ºC.
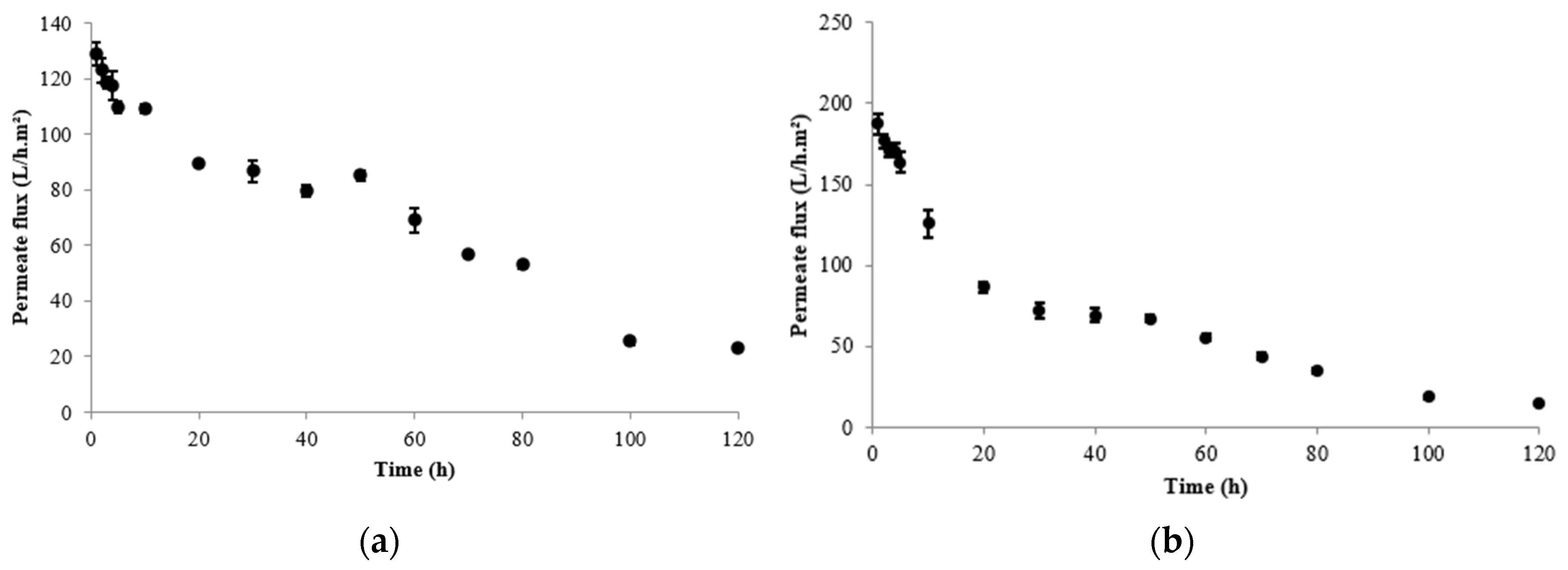
Figure 5.
Gel electrophoresis analysis of buttermilk samples. Subtitle: M: Master markers (5 to 250 kDa) R: Retentate; HB: hydrolysed buttermilk; P: Permeate; B: non-hydrolysed buttermilk.
Figure 5.
Gel electrophoresis analysis of buttermilk samples. Subtitle: M: Master markers (5 to 250 kDa) R: Retentate; HB: hydrolysed buttermilk; P: Permeate; B: non-hydrolysed buttermilk.
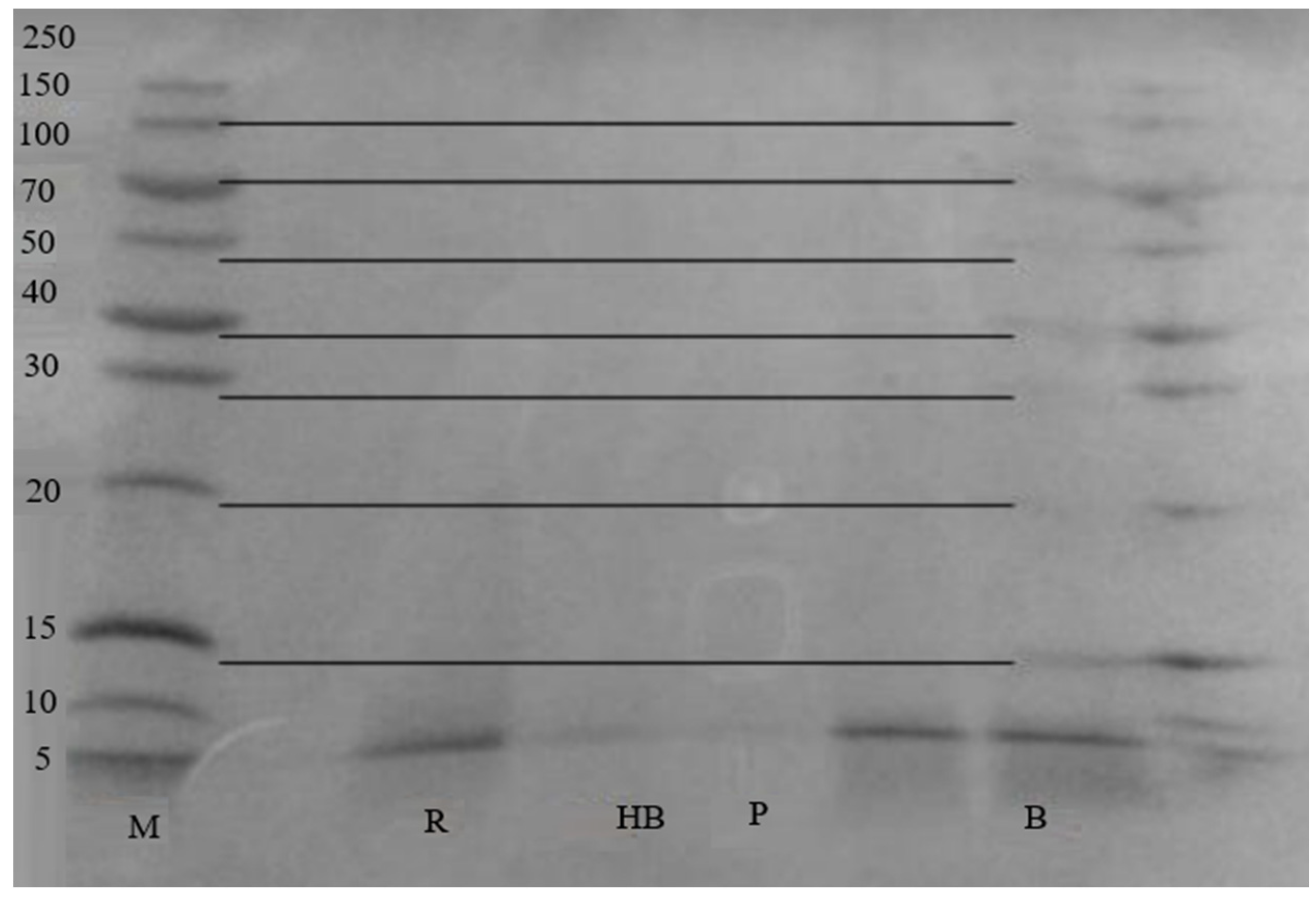

Table 1.
Conditions of reaction for each enzyme tested for the hydrolysis of protein in buttermilk.
| Enzyme | Action | pH | Temperature (ºC) |
Enzyme concentration (g 100 mL -1) |
Inactivation |
|---|---|---|---|---|---|
| Alcalase® | Endopeptidase | 8.0 | 55 | 0.5 | 90 ºC / 20 min |
| Neutrase® | Metallopeptidase | 7.0 | 55 | 1.25 | 90 ºC / 20 min |
| Prolyve® | Endopeptidase | 8.0 | 55 | 0.5 | 90 ºC / 5 min |
Table 2.
Concentration of protein, lactose, and fat in the buttermilk, hydrolysed buttermilk by Alcalase + Neutrase, permeate and retentate from membrane bioreactor.
Table 2.
Concentration of protein, lactose, and fat in the buttermilk, hydrolysed buttermilk by Alcalase + Neutrase, permeate and retentate from membrane bioreactor.
| Sample | Stream | Protein (%) | Lactose (%) | Fats (%) |
|---|---|---|---|---|
|
Buttermilk |
Feed | 2.08±0.03 | 1.76±0.04 | 2.43±0.09 |
| Permeate | 0.93±0.08 a | 0.91±0.09 a | 0.23±1.03 a | |
| Retentate | 1.08±1.08 a | 0.62±1.12 a | 2.12±0.98 a | |
|
Hydrolysed buttermilk |
Feed | 2.08±0.03 | 1.76±0.04 | 2.43±0.09 |
| Permeate | 1.28±0.09 b | 0.94±0.08 a | 0.24±0.08 a | |
| Retentate | 0.78±1.02 b | 0.59±0.09 a | 2.18±0.05 a |
* Mean results ± standard deviation; different letters in the same column correspond to a significant difference (p < 0.05) according to the Tukey test.
Table 3.
Antioxidant Activity (AA) of buttermilk, hydrolysed buttermilk by different enzymes, permeate and retentate from the Membrane Bioreactor (MBR).
Table 3.
Antioxidant Activity (AA) of buttermilk, hydrolysed buttermilk by different enzymes, permeate and retentate from the Membrane Bioreactor (MBR).
| Enzyme | Time (h) |
Degree of hydrolysis (DH) |
Antioxidant activity ( μmol TE/g protein) |
|---|---|---|---|
| Buttermilk | - | - | 396.81±3.95ª |
| Alcalase | 2.5 | 25.8 | 803.58±1.57 b |
| Prolyve | 3.5 | 25.2 | 795.73±5.66 b |
| Neutrase | 1.8 | 5.8 | 418.82±4.16ª |
| Alcalase + Neutrase | 4.3 | 34.02 | 834.21±2.40 b |
| Permeate | - | - | 956.87±1.57 c |
| Retentate | - | - | 825.48±3.12 b |
AA expressed by mean ± standard deviation; different letters in the same column correspond to a significant difference (p < 0.05) according to the Tukey test.
Table 4.
Average lenght of protein chains and estimated average molar mass of buttermilk and hydrolysed buttermilk with different enzymes.
Table 4.
Average lenght of protein chains and estimated average molar mass of buttermilk and hydrolysed buttermilk with different enzymes.
| Enzyme | Time (h) | Hydrolysis Degree (HD) | Average lenght of the chains (No. of residues of aminoacids) |
Average molar mass (kDa) |
|---|---|---|---|---|
| Buttermilk | - | - | 100 | 12,000.00 |
| Alcalase | 2.5 | 25.8 | 4.0 | 483.22 |
| Prolyve | 3.5 | 25.2 | 4.0 | 476.19 |
| Neutrase | 1.8 | 5.8 | 17.2 | 2068.97 |
| Alcalase + Neutrase | 4.3 | 34.02 | 2.9 | 0.35 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



