Submitted:
29 November 2024
Posted:
30 November 2024
You are already at the latest version
Abstract
DNA methylation, a highly conserved epigenetic modification, plays an essential role in regulating gene expression and maintaining genome stability. Aberrations in DNA methylation patterns can significantly disrupt plant developmental processes, often leading to abnormal phenotypes. These methylation states are governed by a dynamic interplay of de novo methylation, maintenance methylation, and active demethylation, mediated by a suite of specialized enzymes operating under distinct regulatory pathways. This review explores the multifaceted roles of DNA methylation in plants. We provide a comprehensive overview of methylases, demethylases, and regulatory factors, elucidating how these components interact within the broader methylation machinery. Key functions of DNA methylation, including its roles in transposon silencing, gene expression regulation, and chromatin interactions, are systematically examined. Additionally, the impact of DNA methylation on plant development is analyzed, with a focus on its involvement in responses to biotic and abiotic stressors. Finally, we discuss the potential applications of DNA methylation in crop improvement, highlighting its promise for advancing modern agricultural practices.
Keywords:
DNA methylation
; crop breeding
; stress response
1. Introduction
Epigenetic modifications represent heritable chemical changes that influence phenotypic traits without altering the underlying DNA nucleotide sequence [1,2]. In plants, the epigenome's dynamic and plastic nature is crucial for regulating development, environmental adaptation, and evolutionary processes [3]. Key epigenetic marks encompass DNA methylation and diverse post-translational modifications of histones-including acetylation, methylation, ubiquitination, SUMOylation, and phosphorylation-which predominantly occur on the histone N-terminal tails [4,5,6]. Additionally, non-coding RNAs and chromatin remodeling factors play integral roles in modulating gene expression and genome accessibility, thereby facilitating phenotypic plasticity [7]. Collectively, these epigenetic mechanisms enable plants to rapidly adapt to fluctuating environmental conditions, underscoring their evolutionary significance.
DNA methylation is a fundamental and extensively studied epigenetic mechanism in eukaryotes. Often referred to as 5-methylcytosine (5mC), it plays pivotal roles in transposon silencing, transcriptional regulation, and the maintenance of normal growth and development in numerous organisms [8,9,10]. Notably, plant genomes consistently exhibit extensive DNA methylation, with 5mC constituting up to 50% of total cytosine residues in some species [11]. This methylation occurs across all three sequence contexts: CG, CHG, and CHH (where H represents A, C, or T). Within plant genomes, heterochromatic regions, which are rich in transposable elements, are typically highly methylated across all sequence contexts. Conversely, dispersed transposons or repetitive elements in euchromatic arms are predominantly methylated and silenced. Furthermore, many genic regions in plant genomes are methylated at CG sites, although this methylation is not directly linked to transcriptional gene silencing [12]. Despite these insights, the broader functional implications of genomic methylation remain incompletely understood and warrant further investigation.
DNA methylation is a highly conserved epigenetic modification in both plants and mammals, playing a pivotal role in regulating development by maintaining precise genomic patterns. In both kingdoms, this modification is catalyzed by conserved DNA methyltransferases that utilize S-adenosyl-L-methionine as the methyl donor. Active demethylation, on the other hand, is mediated via the base excision repair pathway, underscoring the dynamic nature of DNA methylation [13,14,15,16]. In the model plant Arabidopsis thaliana, five functional DNA methyltransferases—MET1, CMT2, CMT3, DRM1, and DRM2—are critical for maintaining genomic methylation patterns [17] (Figure 1). MET1 is specifically involved in CG methylation, interacting with methyl-CG-binding proteins VIM1, VIM2, and VIM3 to facilitate semi-conservative methylation during DNA replication [18]. The maintenance of CHG methylation is primarily governed by CMT3, which operates through a positive feedback loop involving the mono- and dimethylation of histone H3 at lysine 9 (H3K9me1/2) [19]. Meanwhile, CMT2 plays a parallel role in catalyzing CHH methylation, predominantly in heterochromatic regions, through mechanisms closely resembling those employed by CMT3 [19]. DRM1 and DRM2, the latter being the major player, mediate CHH methylation through the RNA-directed DNA methylation (RdDM) pathway, a unique mechanism that employs non-coding RNAs to guide de novo methylation [20]. Beyond its involvement in RdDM, DRM2 serves as the primary enzyme responsible for de novo methylation across all sequence contexts, highlighting its centrality in epigenomic regulation.
This review examines recent advances and current perspectives on the regulation and function of DNA methylation in plants. The mechanisms that establish specific DNA methylation patterns are most comprehensively characterized in the model plant Arabidopsis thaliana, where mutations in DNA methylation and demethylation pathways, as well as alterations in regulatory factor composition, typically do not lead to plant lethality. In contrast, for plants with more complex genomes, DNA methylation is increasingly recognized as a critical regulator of development and adaptation to environmental stress. Emerging research has uncovered pivotal mechanisms governing plant DNA methylation. Notable examples include the initiation of de novo DNA methylation mediated by non-coding RNAs (ncRNAs), the activity of the novel protein complex IDM, which targets active DNA demethylation and paradoxically enhances DNA methylation, and the regulatory role of methylation-sensing genetic elements that dynamically balance methylation and demethylation processes. Additionally, this review highlights the dynamic roles of DNA methylation in regulating various biological processes, including transposon silencing, gene expression, chromosomal interactions, plant development, and responses to both biotic and abiotic environmental stimuli. Finally, the potential applications of DNA methylation dynamics in crop improvement and breeding are discussed, emphasizing its transformative potential in agricultural innovation.
2. DNA Methylation Kinetics
2.1. Establishment of DNA Methylation
In plants, de novo DNA methylation is facilitated by the RNA-directed DNA methylation (RdDM) pathway, which integrates small interfering RNAs (siRNAs), scaffold RNAs, and an array of associated proteins. Based on the current model of the canonical RdDM pathway in Arabidopsis thaliana [20,21,22], the biogenesis of 24-nucleotide siRNAs initiates with transcription by RNA polymerase IV (Pol IV). This process relies on RNA-dependent RNA polymerase 2 (RDR2), which converts the Pol IV transcript into double-stranded RNA (dsRNA). Subsequently, Dicer-like protein 3 (DCL3) cleaves the dsRNA into siRNAs. The siRNAs are then incorporated into ARGONAUTE (AGO) proteins, primarily AGO4 and AGO6, where they guide the recognition of complementary scaffold RNAs transcribed by RNA polymerase V (Pol V). AGO4 directly interacts with the DNA methyltransferase domain rearrangement methylase 2 (DRM2), which catalyzes de novo DNA methylation in a sequence-independent manner [23]. Additionally, this reaction may involve RNA-directed DNA methylation 1 (RDM1), which associates with both AGO4 and DRM2 and may also bind single-stranded methylated DNA [24].
Small interfering RNAs (siRNAs) originating from diverse sources can also associate with Argonaute (AGO) proteins to mediate DNA methylation [25]. Parasitic sequences, including active transposons and invading viruses, are identified by host cells based on distinctive RNA features. These foreign RNAs are subsequently converted into double-stranded RNA (dsRNA) by RNA-dependent RNA polymerases (e.g., RDR6) and processed into 21–24 nucleotide siRNAs [26]. These siRNAs are then incorporated into various AGO proteins to guide de novo DNA methylation. The mechanisms underpinning these pathways remain poorly defined and are collectively referred to as non-classical RNA-directed DNA methylation (RdDM) [27].
In RNA-directed DNA methylation (RdDM), sequence-specific pairing between small interfering RNAs (siRNAs) and scaffold RNAs is complemented by critical protein interactions. Specifically, ARGONAUTE 4 (AGO4) interacts with AGO hook-like proteins such as DNA-directed RNA polymerase V (Pol V) subunit 1 (NRPE1) and RDM3, which are essential for RdDM. Among these, NRPE1 serves as the largest subunit of Pol V, while RDM3 is hypothesized to function as a transcriptional elongation factor associated with Pol V activity [28,29]. For non-coding RNAs (ncRNAs) produced by Pol V transcription to act as scaffold RNAs, they must remain associated with chromatin. This retention appears to be mediated by RRP6-like 1 (RRP6L1), a homolog of the yeast and mammalian ribosomal RNA processing 6 (RRP6) protein, which facilitates RNA stabilization within chromatin [30]. Furthermore, the pairing of siRNAs with scaffold RNAs may be stabilized by the involvement of the IDN2-IDN2 PARALOGUE (IDP) complex. This complex not only binds RNA but also interacts with the SWI/SNF chromatin remodeling complex containing the SWI/SNF subunit SWI3B. Through this interaction, it participates in Pol V-mediated transcriptional silencing by modulating nucleosome positioning [31,32,33,34].
Histone modifications play a critical role in the recruitment of Pol IV to chromatin. The histone-binding protein SHH1 (also known as DTF1) preferentially binds to H3K9me2, thereby facilitating the recruitment of Pol IV to specific loci involved in RNA-directed DNA methylation (RdDM) [35,36]. ZMP interacts with NRPD1 to promote Pol IV binding in particular pericentromeric regions while simultaneously preventing its targeting of transposon elements (TEs) and certain euchromatic genes [37]. ZMP associates with H3K4me0 and chromatin regions near H3K4 methylation sites, suggesting that it plays a key role in monitoring H3K4 methylation levels to regulate Pol IV occupancy [37]. The recruitment of Pol IV to chromatin also requires the CLASSY family of chromatin remodeling factors, which directly interact with SHH1 and are essential for the production of Pol IV-dependent siRNAs [35,38,39]. Pol V, which binds chromatin to produce scaffold RNA, requires chromatin remodeling by the DDR complex. This complex includes DEFECTIVE IN RNA-DIRECTED DNA METHYLATION 1 (DRD1), the chromatin structure maintenance protein DEFECTIVE IN MERISTEM SILENCING 3 (DMS3), and RDM1 [40,41,42,43]. The DDR complex physically interacts with SUPPRESSOR OF VARIEGATION 3-9 HOMOLOG PROTEINS 2 (SUVH2) and SUVH9, members of the SUPPRESSOR OF VARIEGATION (SUV) 3-9 histone methyltransferase family, which lack histone methyltransferase activity [44,45]. SUVH2 and SUVH9, which recognize methylated cytosine through their SET and RING finger association (SRA) domains, are essential for the correct occupancy of Pol V at chromatin sites across the genome. It has been suggested that these proteins may recruit Pol V by pre-existing DNA methylation patterns [45]. Interestingly, SUVH9’s ability to bind zinc fingers to unmethylated DNA is sufficient to recruit Pol V and establish DNA methylation and gene silencing [45].
Given POL V's capacity to generate non-coding RNAs (ncRNAs) with distinct 5ʹ ends from the same locus, it appears that POL V can initiate transcription independently of a promoter [46]. Some of the ncRNAs transcribed by POL V possess 7-methylguanosine caps at their 5ʹ ends [46], indicating that transcripts produced by POL V may undergo RNA processing similar to that of mRNA transcribed by POL II. However, unlike mRNA, POL V transcripts lack polyadenylation at their 3ʹ ends [46]. The typical length of POL V transcripts is approximately 200 nucleotides. In contrast, POL IV-derived ncRNAs (P4 RNAs) are generally 26-50 nucleotides long. Until recently, small RNA deep sequencing of Arabidopsis thaliana dcl2, dcl3, and dcl4 triple mutants, as well as dcl1, dcl2, dcl3, and dcl4 quadruple mutants, revealed that the cleavage of POL IV- and RDR2-dependent double-stranded RNAs (dsRNAs) into 24-nucleotide siRNAs is often blocked [47,48,49,50,51]. In these mutants, P4 RNA accumulates and can be processed by exogenous DCL3 into 24-nucleotide siRNAs [49].
In addition to the classical POL IV-RDR2-DCL3 pathway, paralogous proteins can also generate small interfering RNAs (siRNAs) that trigger RNA-directed DNA methylation (RdDM) via non-canonical pathways [52]. For instance, RNA polymerase II (POL II) not only produces 24-nt siRNAs and scaffold RNAs but also recruits POL IV and POL V to regulate specific RdDM targets [53]. In certain genomic regions, RdDM is dependent on POL II and RDR6, rather than POL IV and RDR2. This alternative pathway generates 21- or 22-nt siRNAs through DCL2/DCL4, or 24-nt siRNAs through DCL3 [54,55]. Despite the substantial loss of siRNAs in the DCL quadruple mutant, approximately two-thirds of the RdDM-targeted DNA methylation remains intact, suggesting the potential existence of DCL-independent RdDM mechanisms [50,51]. Furthermore, other RNase III enzymes may collaborate with DCL to process transcripts and generate siRNAs. Additionally, some mRNA splicing factors are thought to interact with non-coding transcripts produced by POL IV and POL V, influencing their processing and stability, thereby regulating the abundance of siRNAs and scaffold RNAs. However, the precise mechanisms underlying these interactions remain unclear [56,57].
2.2. Maintenance of DNA Methylation
In Arabidopsis thaliana, the maintenance of DNA methylation patterns requires the action of all five DNA methyltransferases. Specifically, CHG and CHH methylation are maintained through self-reinforcing feedback loops. These loops involve DNA methyltransferases (CMT3, CMT2, DRM2, and DRM1), histone methyltransferases, and nucleosomes modified by repressive marks, such as H3K9me2 and H3K9me1 [58,59]. CG methylation is primarily preserved by the DNA methyltransferase MET1 in Arabidopsis, which is thought to methylate semi-methylated CG sites during DNA replication, thereby catalyzing the establishment of CG methylation on the newly synthesized DNA strand [59,60]. Both in vitro and in vivo studies have demonstrated that DNMT1, a mammalian homolog of MET1, preferentially acts on hemimethylated DNA substrates, supporting this semi-conserved model [61,62,63]. During DNA replication, DNMT1, along with the E3 ubiquitin ligase UHRF1, is localized to replication foci, a mechanism that is also essential for maintaining CG methylation in mammals [64,65]. In plants, the VIM genes encode homologs of UHRF1, and their function in CG methylation mirrors that of their mammalian counterparts. Specifically, Arabidopsis exhibits a complete loss of CG methylation when MET1 is mutated or in a triple mutant of VIM1/VIM2/VIM3 [66]. Both DNMT1 and UHRF1 are multi-domain proteins, with their catalytic domains located at the C-terminal and regulatory domains at the N-terminal [67]. Notably, the N-terminal domains of MET1 and VIM proteins differ from those of DNMT1 and UHRF1, suggesting that plants and mammals employ distinct regulatory mechanisms.
2.3. Demethylation of Active DNA
The failure to maintain DNA methylation following replication, due to the absence of either DNA methyltransferase activity or a methyl donor, results in passive DNA demethylation—a process distinct from enzymatic removal of methylation marks known as active DNA demethylation [68,69,70]. Unlike DNA methylation, which is catalyzed by a single DNA methyltransferase, active DNA demethylation requires the coordinated action of multiple enzymes, with the initial step carried out by a DNA demethylase. In plants, active DNA demethylation is initiated by a family of bifunctional 5-mC DNA glycosylase-purine/depyrimidine lyases, which excise 5-mC through the base excision repair (BER) pathway [71,72]. Similarly, active demethylation in mammals involves DNA glycosylases and the BER mechanism. However, a key distinction lies in substrate recognition: plant DNA glycosylases directly identify and excise 5-mC bases, whereas in mammals, 5-mC undergoes oxidation prior to glycosylase-mediated excision [73].
Arabidopsis thaliana possesses a family of four bi-functional 5-mC DNA glycosylases: REPRESSOR OF SILENCING 1 (ROS1), DEMETER (DME), and DEMETER-LIKE PROTEINS 2 and 3 (DML2 and DML3) [71,73]. These enzymes are capable of excising 5-mC from cytosines across all sequence contexts [72,74,75,76]. ROS1, DML2, and DML3 are broadly expressed across vegetative tissues, whereas DME exhibits a more specialized expression pattern, predominantly localized to the companion cells of female and male gametophytes. Specifically, DME is expressed in the central cells of female gametophytes and the trophic cells of male gametophytes [75,77].
During DNA demethylation, bifunctional enzymes initially function as DNA glycosylases, hydrolyzing the glycosyl bond between the base and deoxyribose. Subsequently, they act as apurinic/apyrimidinic (AP) lyases, cleaving the DNA backbone to generate abasic (base-free) sites. Following the excision of the 5-methylcytosine (5-mC) base, either a β-elimination or β, δ-elimination reaction occurs. These reactions create a gap in the DNA strand, terminating with either a 3ʹ-phosphate-α, β-unsaturated aldehyde or a 3ʹ-phosphate, respectively. In the downstream processing of these elimination reactions, AP endonuclease 1 (APE1) and the polynucleotide 3ʹ-phosphatase ZDP play crucial roles. APE1 operates following β-elimination, while ZDP acts after β, δ-elimination, both facilitating the generation of a 3ʹ-hydroxyl (3ʹ-OH) group. This modification prepares the DNA strand for subsequent repair by DNA polymerases and ligases, which restore the integrity of the DNA [78,79,80].
3. Molecular Functions of Plant DNA Methylation
3.1. Regulation of Gene Expression
In plants, gene-associated DNA methylation occurs within promoters or in transcribed regions of the genome (Figure 2). While promoter DNA methylation typically suppresses gene transcription, it can also activate transcription in certain contexts. For instance, this dual role has been observed in the regulation of the ROS1 gene in Arabidopsis and in hundreds of genes involved in inhibiting tomato fruit ripening [18,81,82,83]. Promoter DNA methylation suppresses transcription through both direct and indirect mechanisms. Directly, it can prevent the binding of transcriptional activators or facilitate the binding of transcriptional repressors. Indirectly, it operates by enhancing repressive histone modifications, such as H3K9me2, while simultaneously reducing permissive modifications, such as histone acetylation [84,85]. The mechanism by which promoter methylation activates transcription remains elusive. One hypothesis suggests that DNA methylation may increase the affinity of certain transcriptional activators or inhibit the binding of specific transcriptional repressors. Promoter DNA methylation often arises from the spread of methylation from nearby transposable elements or repetitive sequences. These transposons and repeats are frequently targeted by active DNA demethylation to shield adjacent genes from transcriptional silencing [86]. Interestingly, for genes whose expression depends on promoter DNA methylation, active demethylation can paradoxically lead to transcriptional repression. Such dynamic regulation highlights the intricate interplay between DNA methylation and transcriptional activity [82,87].
To achieve the silencing effect, promoter methylation requires the involvement of additional effector such as methyl-DNA-binding proteins, histone modifiers, chromatin remodeling factors, and molecular chaperones, resulting in a tight chromatin structure that makes transcription factors inaccessible. In Arabidopsis, an interesting role of chaperones in regulating chromatin state rich in mCG sequences, including promoter regions, has been revealed [88,89,90]. mCG is able to attract MBD proteins such as MBD7111, MBD5, and MBD6 [88,90], which have a C-terminal StkyC domain that interacts with the ACD family of chaperone proteins [88,90]. Subsequently, ACD proteins IDM2 and IDM3 recruit histone acetyltransferase IDM1 and related proteins HDP1 and HDP2 to form IDM complexes. This complex is necessary for DNA demethylation and transcriptional activity of the highly methylated viral 35S promoter [90,91]. In another locus containing a 35S promoter, the ACD proteins RDS1, RDS2, and IDM3 recruit other chaperone proteins, such as the DnaJ domain proteins SDJ4 and HSP70, to form molecular chaperone complexes, which, in different genetic contexts, can be used as chaperones. This complex can silence or express the locus [88]. However, the role of chaperone proteins in methylation loci remains to be further studied. MBD proteins have inherently disordered regions that may facilitate phase separation to form molecular agglomerates or aggregates. Acds are known as "maintainer" proteins because they bind to target proteins, preventing them from aggreging irreversibly [92]. DnaJ usually assists HSP70, which has an ATP-dependent chaperone activity that enables substrate proteins to regenerate or depolymerize [92]. These chaperone proteins may be able to regulate the aggregation state of MBD to control chromatin accessibility of McG-rich loci.
3.2. Transposon Silence
Transposons represent a significant challenge to genome stability, either by relocating DNA transposons or through the insertion of new copies of retrotransposons. In Arabidopsis thaliana, regions of heterochromatin surrounding the centromeres and certain euchromatic areas containing transposons or repetitive sequences are extensively methylated across all cytosine contexts [93] (Figure 2). RNA-directed DNA methylation (RdDM) plays a critical role in maintaining asymmetric CHH methylation at the edges of both short and long transposons. Within the internal regions of heterochromatic long transposons, asymmetric methylation is regulated by the chromatin remodeler DDM1 and catalyzed by CMT2 [19,94]. In the maize genome, active genes and inactive transposons are interspersed, often separated by RdDM-dependent CHH methylation "islands." These are short genomic regions characterized by elevated CHH methylation levels. The loss of CHH methylation islands frequently results in transcriptional activation, accompanied by hypomethylation of CG and CHG sites within adjacent transposons. This suggests that in maize, RdDM is indispensable for silencing transposons in euchromatic regions containing nearby active genes [95]. In sugar beet, CHH methylation appears more specifically involved in silencing DNA transposons, which exhibit higher CHH methylation levels compared to retrotransposons and genes [96]. Intriguingly, the pericentromeric regions of A. thaliana and tomato exhibit a context bias in CHH methylation, characterized by low levels of methylated CCG and enrichment in methylated CAA, CTA, or CAT contexts. In contrast, CHH methylation within euchromatic regions of dicot plants like Arabidopsis and tomato is not context-dependent [97]. Meanwhile, in monocot plants such as maize and rice, the context-specific CHH methylation bias is distributed more evenly across the chromosomes [97].
3.3. Chromosome Interaction
DNA methylation influences the epigenetic state of chromatin and plays a crucial role in mediating interactions between chromosomes. In the nucleus of Arabidopsis thaliana, all five chromosomes interact through a structure known as the KNOT [98]. This structure comprises interactive heterochromatin islands (IHIs), which are repressive chromatin regions embedded within euchromatin arms and are characterized by a high density of transposons and small RNAs [98,99]. Notably, IHI interactions remain unaffected in met1 and ddm1 mutants, which exhibit extensive DNA hypomethylation across all cytosine contexts. A similar observation is made in the suvh4-suvh5-suvh6 triple mutant, which lacks functional H3K9 methylation [99]. These findings suggest that both DNA methylation and H3K9me2 are essential for maintaining chromosomal interactions within IHIs.
Interestingly, mutants deficient in RNA-directed DNA methylation (RdDM) display increased frequencies of chromosomal interactions in specific RdDM-targeted regions. This indicates that, in wild-type plants, RdDM acts to suppress chromosome interactions in certain genomic regions [100]. Moreover, enhanced chromosomal interactions are observed between POL V-dependent DNA methylation sites and distal genes that are repressed by RdDM. These findings suggest that chromosomal interactions may also play a regulatory role in gene expression [100].
4. The Role of DNA Methylation in Plant Development
4.1. Genomic Imprinting and Seed Development
Arabidopsis plants employ a unique double fertilization strategy that relies on the multicellular organization of male and female gametophytes. During this process, each of the two sperm cells within a pollen grain fertilizes distinct female gametophyte cells—the egg cell and the central cell—giving rise to the embryo and the endosperm, respectively. In both rice and Arabidopsis thaliana, the endosperm exhibits global DNA hypomethylation compared to the embryo [101,102,103]. In A. thaliana, this hypomethylation is attributed in part to active demethylation mediated by DEMETER (DME) in pre-fertilization central cells, which are the partner cells of female gametes [101,103]. Interestingly, although MET1 transcription is repressed during female gametogenesis, this repression does not appear to drive extensive demethylation. Consistent with this observation, genome-wide CG hypomethylation, which would be expected under reduced MET1 activity, is not detected in wild-type endosperm. Moreover, DNA methylation is almost entirely restored in the endosperm of dme mutants, underscoring the critical role of DME-dependent pathways in establishing endosperm-specific epigenetic patterns [102,104].
DME-mediated demethylation occurs in androtrophic cells and is accompanied by the downregulation of DDM1. This process triggers the production of small interfering RNAs (siRNAs) derived from demethylated transposons, which are subsequently transported to sperm cells, where they enhance RNA-directed DNA methylation (RdDM). These transposon-derived siRNAs in sperm cells may also contribute to transposon silencing in egg cells after fertilization [105,106,107]. The levels of CHH methylation dynamically fluctuate during seed development and germination. Specifically, CHH methylation increases during seed development but decreases during germination, primarily due to passive demethylation, suggesting its potential role in regulating seed dormancy [108]. Notably, while male reproductive lineages exhibit lower overall CHH methylation than somatic cells, certain hypermethylated CHH sites are essential for meiosis [109]. In the endosperm, methylation levels of the maternal genome are lower than those of the paternal genome, particularly at CG sites, which are closely associated with gene imprinting. Maternal expression genes (MEGs) typically exhibit hypomethylation of maternal alleles and hypermethylation of paternal alleles. For instance, certain MEGs, such as MEDEA, are silenced not through DNA methylation but via histone modification by H3K27me3 [109,110,111,112]. Disruptions in the maternal function of DME or the paternal function of MET1 interfere with the imprinting of MEGs, underscoring the crucial role of DNA methylation in allele-specific regulation [109,113,114,115]. Conversely, the maternal alleles of paternally expressed genes (PEGs) are typically marked by H3K27me3, while the paternal alleles are expressed via H3K36me3. This coordinated regulation by histone modifications and DNA methylation establishes the distinct expression patterns of MEGs and PEGs in the endosperm [116,117].
4.2. Role of DNA Methylation in Plant Meristem and Leaf Epidermal Development
Plant meristem stem cells are fundamental to tissue and organ development, with RdDM factors exhibiting higher transcription levels in meristematic tissues compared to differentiated tissues. Notably, the highest DNA methylation levels are observed in the small columnar cells of the root meristem, potentially due to reduced exposure of pericentromeric chromatin to RdDM factors. While RdDM mutants in Arabidopsis do not exhibit pronounced meristem defects, corresponding mutants in rice and maize display severe developmental abnormalities, underscoring the critical role of RdDM in meristem development [118,119]. In rice, SDG711-mediated H3K27me3 deposition represses the expression of leaf development genes following shoot apical meristem development. This repression coincides with DRM2-catalyzed non-CG DNA methylation, suggesting a coordinated regulatory mechanism. Furthermore, physical interactions between SDG711 and DRM2 imply a synergistic effect in gene silencing [120]. In maize, differential regulation of maintenance DNA methyltransferases establishes distinct DNA methylation patterns across the division, transition, extension, and maturation zones of leaves. These methylation changes predominantly occur near genes involved in growth and cell cycle regulation, where promoter methylation is inversely correlated with gene expression [121]. DNA methylation also plays a pivotal role in leaf epidermal patterning in Arabidopsis. Dysfunction of ROS1 leads to hypermethylation of the EPF2 promoter, suppressing the expression of stomatal inhibitory factors and resulting in excessive stomatal production [122]. Similarly, IBM1 dysfunction enhances H3K9me2 and CHG methylation, inhibiting EPF2 receptor gene expression and disrupting stomatal patterning. Interestingly, ROS1 mutation-induced abnormalities can be alleviated by RdDM factor mutations, whereas IBM1 mutation-induced defects are mitigated by SUVH4 or CMT3 mutations. These findings suggest that DNA methylation governs leaf epidermal development through distinct regulatory pathways [123].
4.3. Role of DNA Methylation in Flower Development
DNA methylation has been shown to play a pivotal role in various aspects of flower development, including anthocyanidin deposition and floral patterning. For instance, DNA methylation influences anthocyanin accumulation in the orchid Oncidium. In Oncidium Gower Ramsey (GR), the lips of the flowers appear bright yellow due to the accumulation of carotenoid pigments. The genetic pathway responsible for this pigmentation is well characterized, and its variation accounts for the white coloration observed in the Oncidium variety known as 'White Jade' (WJ) [124]. The distinct color patterns observed between these orchid varieties arise from differential methylation levels of carotenoid-related genes. Specifically, in GR, the OgCCD1 gene is methylated and silenced. In contrast, in WJ tissues, the absence of methylation in the OgCCD1 promoter results in its active expression, leading to the degradation of carotenoids and the development of white flowers [125]. Another notable feature of the pigmentation pattern in GR flowers is the presence of red stripes in the perianth, which are absent in the Oncidium Honey Dollp (HD) cultivar. In this case, the contrasting flower colors are determined by the methylation status of genes involved in anthocyanin biosynthesis. In the HD variety, methylation of the OgCHS gene promoter prevents anthocyanin accumulation, whereas in GR, unmethylated genes are expressed, facilitating anthocyanin production [126]. Additionally, DNA methylation plays a significant role in floral color formation in many other ornamental plants. For example, in Malus halliana, the color of the petals changes from red to pale pink during development, which is linked to the downregulation of several genes. Notably, the promoter of the MhMYB10 gene, which is involved in anthocyanin biosynthesis, is highly methylated, leading to reduced expression of MhMYB10 and a consequent decrease in anthocyanin accumulation [127].
Furthermore, DNA methylation plays a crucial role in regulating floral fragrance production. In Chinese and Japanese plums (Prunus mume), several genes involved in eight floral biosynthetic pathways exhibit varying methylation levels during flower development [128]. These genes encode enzymes that are key regulators of floral scent production, including coniferol acetyltransferase (PmCFAT1a/1c) and benzyl acetyltransferase (PmBEAT36/3). The CFAT proteins, which belong to the acyltransferase family, are integral to the synthesis of eugenol, catalyzing the conversion of coniferol to coniferol acetate [116]. The BEAT protein facilitates the transfer of an acetyl group from acetyl-CoA to the carbonyl group of benzyl alcohol, resulting in the formation of benzyl acetate [117]. Whole-genome bisulfite sequencing of P. mume revealed that most of the differentially methylated genes are involved in various stages of the phenylpropanoid biosynthesis pathway, which produces more than 90% of the flower volatiles [128].
4.4. Role of DNA Methylation in Fruit Ripening
During the development of tomato fruit, about 1% of the DNA methylation group in the fruit peel is changed. Active DNA demethylation occurs on many fruit ripening genes whose promoter regions contain binding sites for ripening inhibitors (Rins), a major ripening transcription factor [129]. RIN binding to target promoters has been demonstrated in most known mature genes, and expression of these genes is inversely correlated with promoter DNA methylation levels. Treatment with chemical inhibitors of DNA methylation induces promoter hypomethylation and expression of genes encoding colorless immature (CNR), a key Rin-targeting gene for fruit ripening, which also induces premature ripening of tomato fruits [129]. The expression of DNA demethylase DME-LIKE 2 (DML2) in tomato (Solanum lycopersicum) is sharply increased in ripe fruits, which mediates progressive DNA demethylation that occurs during fruit ripening [130]. Tomato (S. lycopersicum) DML2 not only targets ripening induction genes, but also ripening suppressor genes, indicating that activation of ripening induction genes and inhibition of ripening suppressor genes both require demethylation of active DNA [131]. DNA methylation changes may be involved in the growth and ripening of other fruits. In apple fruit, anthocyanin accumulation was negatively correlated with DNA methylation levels in the apple MYB10 gene promoter [132,133]. At the genome-wide level, CHH hypermethylation was observed in developing apple fruits compared to leaves, and comparisons with genetically modified fruits also showed an association between lower DNA methylation levels and smaller fruit size [134].
5. The role of Plant DNA Methylation in Abiotic Stress
5.1. High Temperature Stress
High temperatures and drought are critical climate stressors that significantly impair global crop yields. Projections suggest that global temperatures may rise by approximately 4-5 °C by the end of the 21st century [135]. Prolonged heat stress coupled with insufficient rainfall often precipitates drought conditions. The synergistic effects of heat and drought typically result in more severe damage to crop production than either stressor alone [136]. Both environmental stresses have been shown to induce alterations in DNA methylation, including hypermethylation and demethylation, in both coding and intergenic regions. In response to heat stress, the heat-resistant rapeseed (Brassica napus) genotype Huyou2 exhibited predominantly hypomethylated DNA, in contrast to the more heat-sensitive genotype Fengyou1 [137]. Additionally, in the drought-tolerant crop millet (Setaria italica), methylation of CHG and CHH regions in the upstream and coding regions of heat shock protein (HSP) genes was enhanced under environmental stress conditions. In heat-tolerant varieties, heat stress induced hypomethylation of many SiHSP genes in CHG and CHH regions, which in turn enhanced their expression [138]. A MethylRAD library constructed from heat-treated maize inbred line B73 identified 325 differentially methylated genes (DMGs) [139]. Notably, DMGs associated with RNA splicing exhibited low methylation levels and were highly expressed, suggesting that spliceosome activity is augmented under heat stress. A nucleoproteome analysis of Pinus radiata subjected to high-temperature stress revealed a decrease in the abundance of S-adenosylmethionine (SAM) synthase and S-adenosyl-L-homocysteine hydrolase (SAHH), resulting in altered DNA methylation profiles [140]. SAM is a crucial cofactor and methyl group donor, while SAHH is essential for the regeneration of SAM during methylation-mediated gene silencing. Moreover, it was observed that DNA methylation regulates the activity of plant transit factors under heat stress.
5.2. Drought Stress
Drought stress typically induces dynamic alterations in DNA methylation across the plant genome, which are negatively correlated with gene expression. These modifications encompass methylation changes in promoter regions, genomic sequences, and transposable element regions. For instance, research on Polygonum persicaria has demonstrated that drought-treated parents enhance the ability of their offspring to develop deeper and faster-growing roots in arid environments by altering their DNA methylation patterns. This cross-generational stress memory suggests that DNA methylation serves as a molecular link between genetic and environmental signals [142]. Moreover, plant hormone regulation plays a significant role in modulating DNA methylation during drought stress. Studies on poplar have shown that hormone-responsive genes exhibit hypomethylation during stress recovery, which facilitates gene reactivation. Additionally, drought-induced abscisic acid (ABA) signaling pathways are closely associated with DNA methylation modifications [143]. In maize, drought-induced differential methylation was detected via AMP-PCR, with changes predominantly occurring in regions within genes responsible for plant survival functions, such as protein synthesis, DNA repair, and amino acid metabolism. These findings suggest that hormone signaling enhances stress tolerance by regulating dynamic DNA methylation changes. The cumulative impact of multiple drought events reflects stress memory in plants, which is intricately linked to DNA methylation. In alfalfa, a two-stage drought stress regime significantly enhanced drought tolerance compared to a single drought event, a phenomenon associated with methylation remodeling of relevant genes. Stress memory enables plants to respond more efficiently to repeated stress, indicating that DNA methylation underpins long-term adaptation to abiotic stress [144].
5.3. Salt Stress
Salt stress can cause changes in DNA methylation levels across the plant genome. This dynamic change usually involves three methylation patterns: CG, CHG, and CHH (C for cytosine and H for non-G bases). Studies have shown that in the initial stage of salt stress, demethylation occurs in the promoter region of many genes, which promotes the expression of related stress-resistance genes [145]. For example, Arabidopsis thaliana demethylates the promoter region of some salt-resistant genes (such as SOS1) under salt stress, thereby enhancing their expression levels and improving the plant's ion elimination ability. On the other hand, the level of methylation in genomic regions is often increased, which may be related to gene silencing and maintenance of genomic stability [145]. In rice, genome-wide methylation analysis showed that salt stress induced differential methylation of a large number of genes involved in osmoregulation, antioxidant response and signal transduction [146]. It has also been found that demethylation of transposition factors induced by salt stress may activate the mobility of these elements, thereby increasing genomic instability [147]. However, through the RdDM (RNA-dependent DNA methylation) pathway, these transposable factors are usually re-silenced to maintain genome integrity. Plant hormones and signaling molecules are closely related to DNA methylation regulation in response to salt stress. Abscisic acid (ABA) is an important signaling molecule in salt stress, and several genes involved in its signal transduction pathway (such as NCED3 and PYR1) exhibit significant changes in methylation levels under stress conditions [148]. For example, demethylation of genes associated with ABA synthesis under salt stress can promote ABA synthesis, thereby enhancing stomatal closure and antioxidant capacity [149]. In addition, ABA signaling pathway also participates in DNA methylation regulation by regulating transcription factors (such as ABI5), forming a positive feedback mechanism. Studies have also shown that there is an interaction between methylation regulation and ion homeostasis maintenance [145,149]. For example, some genes in the SOS signaling pathway are regulated by methylation states that help maintain intracellular Na /K ion balance under salt stress.
The epigenetic memory function of plants enables them to show higher resilience when subjected to repeated salt stress. Studies have shown that DNA methylation is one of the important molecular bases of salt stress memory. For example, in Arabidopsis thaliana, gene demethylation induced by initial salt stress can be partially left over after stress relief and rapidly activate related gene expression during subsequent stress [142,144,145]. Cross-generational genetic studies have shown that salt stress can transmit salt-resistant characteristics to the next generation by affecting the methylation patterns of gametes and embryos [146]. For example, in barley and rice, the offspring of salt-stressed parents showed greater salt tolerance, which was closely associated with changes in methylation patterns of some salt-resistant genes. The function of many salt-resistant genes is directly regulated by DNA methylation [149]. For example, the level of methylation in the promoter region of the OsDREB2A gene, which is involved in osmoregulation in rice, is reduced under salt stress, promoting gene expression. This demethylation may be achieved by activating binding sites of transcription factors such as MYB or bZIP. In addition, genes encoding antioxidant enzymes (such as SOD and CAT) in plants are also regulated by DNA methylation, and their expression is enhanced by demethylation under salt stress, thus alleviating stress-induced oxidative damage [150]. In addition, the methylation regulation of transposable factors and their adjacent genes also plays an important role in salinity resistance. The increased activity of transposable factors under salt stress may provide the potential for resistance to mutation by influencing the expression of neighboring genes. However, this instability is usually inhibited by the RdDM pathway to avoid unwanted genomic disturbances [151].
6. The Role of Plant DNA Methylation in Biological Stress
6.1. Dynamic Changes of DNA Methylation and Response to Biological Stress
In plants, DNA methylation patterns within the genome exhibit dynamic changes in response to biological stress. The methylation levels at CG, CHG, and CHH sites can be rapidly reconfigured to meet stress-related demands. For instance, infection of Arabidopsis thaliana with Plasmodiophora brassicae induces genome-wide DNA demethylation, thereby promoting the expression of genes involved in resistance responses [152]. Specifically, demethylation of promoter regions of genes associated with the biosynthesis and signaling of defense hormones, such as salicylic acid (SA), enhances plant immune responses. Similarly, in rice (Oryza sativa) infected by Xanthomonas campestris, promoter demethylation of numerous defense-related genes facilitates the expression of disease-resistance proteins. Insect herbivory also drives the reprogramming of DNA methylation patterns [153]. For example, in tomato plants (Solanum lycopersicum) subjected to bollworm (Helicoverpa armigera) feeding, demethylation of promoters of insect-responsive genes, including those encoding protease inhibitors, enhances the synthesis of insect-resistance proteins, thereby strengthening the plant’s defense against herbivory [154,155,156,157,158].
Advances in DNA methylation analysis techniques and extensive studies on Arabidopsis thaliana mutants deficient in various steps of DNA methylation, maintenance, and demethylation have significantly deepened our understanding of these processes in regulating plant responses to fungal infections [159] (Table 1). For instance, the highly methylated triple DNA demethylase mutant rdd (ros1dml2dml3) of A. thaliana exhibits markedly increased susceptibility to Fusarium oxysporum [160]. Most genes downregulated in rdd encode known defense-related functions, with their promoter regions enriched in transposable elements (TEs). Resistance to F. oxysporum requires DNA demethylase activity to target these TE sequences within defense gene promoters, thereby enhancing their expression. Interestingly, studies on Arabidopsis mutants defective in different stages of the RNA-directed DNA methylation (RdDM) pathway suggest contrasting effects on defense responses against biotrophic and necrotrophic pathogens. Generally, RdDM-deficient mutants display heightened sensitivity to necrotrophic pathogens but increased resistance to biotrophic ones. For example, as summarized in Table 1, hypomethylated Arabidopsis mutants exhibit increased susceptibility to the necrotrophic fungal pathogen Botrytis cinerea and the bacterial pathogen Plectosphaerella cucumerina [161]. Additionally, mutants such as rdr6 and dcl2/3/4 show heightened vulnerability to B. cinerea due to impaired small interfering RNA (siRNA) synthesis, which is critical for silencing fungal virulence genes [146]. Conversely, knocking out the RdDM component DRM2 homologs in the wheat ancestor Aegilops tauschii enhances resistance to wheat powdery mildew caused by the biotrophic pathogen Blumeria graminis f. sp. tritici. Moreover, the Arabidopsis trimutant ddc (drm1drm2cmt3), which lacks proper methylation maintenance, exhibits extreme sensitivity to the necrotrophic fungus Alternaria brassicicola [157,158,159].
6.2. DNA Methylation Regulation of Disease Resistance Genes
Resistance (R) genes are central to plant defense against pathogen invasion. DNA methylation plays a pivotal role in modulating plant resistance by regulating the expression of these genes. Under normal growth conditions, most R genes remain in a methylation-mediated silenced state, thereby conserving energy and minimizing the risk of autoimmune responses caused by their overexpression. For instance, in Arabidopsis thaliana, hypermethylation of the SUPPRESSOR OF npr1-1 CONSTITUTIVE 1 (SNC1) gene maintains its transcriptional silence [155]. However, during pathogen infection, DNA demethylation reactivates SNC1, bolstering the immune response. In addition to R gene regulation, transposable elements (TEs) in the vicinity of these genes are often controlled by DNA methylation. Pathogen infection can suppress TE activity via the RNA-dependent DNA methylation (RdDM) pathway, thereby preventing their insertion into R genes and the subsequent disruption of gene function. For example, in rice (Oryza sativa), many TEs located near R genes exhibit significantly increased methylation levels following infection with Magnaporthe oryzae (the causal agent of rice blast disease), contributing to genomic stability [156].
6.3. Synergistic Effect of DNA Methylation with Plant Hormone Signaling
Plant hormones play pivotal roles in mediating plant responses to biotic stress, with DNA methylation forming an intricate network that modulates plant defense mechanisms by regulating the expression of genes involved in hormone biosynthesis and signal transduction. Salicylic acid (SA), a key signaling molecule in plant immunity, exhibits significant changes in the methylation status of genes associated with its biosynthesis and signaling under biotic stress. For instance, the demethylation of ICS1 (a gene encoding an SA biosynthetic enzyme) and NPR1 (a central regulator of SA signaling) in Arabidopsis thaliana facilitates SA-mediated systemic acquired resistance (SAR) [154]. Jasmonic acid (JA), predominantly associated with defense against herbivores and necrotrophic pathogens, is similarly regulated. Demethylation of key genes in the JA signaling pathway, such as COI1 and MYC2, has been shown to activate these pathways following herbivore attack, thereby bolstering plant defenses. Ethylene also plays a critical role in coordinating broad-spectrum plant defenses against biotic stress. In rice (Oryza sativa) challenged by blast fungus, significant reductions in methylation levels within the promoter regions of ethylene signaling genes, such as ERF1, enhance the expression of defense-related genes. Conversely, the role of abscisic acid (ABA) in biotic stress responses appears more nuanced, with several genes in the ABA signaling pathway undergoing adaptive regulation via DNA methylation [150,152,157].
7. Prospects of DNA Methylation in Crop Breeding
Natural variations in DNA methylation are widespread in plants and hold immense potential for crop improvement. This potential is becoming increasingly realizable with advancements in gene-editing technologies. Numerous examples of natural epigenetic variation have been documented in plants. A classic case is the radially symmetrical flower mutant of Linaria vulgaris first described by Linnaeus, which results from a hypermethylated allele of the transcription factor CYCLOIDEA [158]. In Arabidopsis, epigenetic variation has elucidated key mechanisms governing DNA methylation regulation [159]. Beyond model plants, epialleles in various crops regulate critical agronomic traits, including plant architecture in rice [160], fruit ripening in tomatoes [161], photoperiodic sensitivity in cotton [162], and grain quality in maize [163]. Additionally, DNA methylation influences hybridization dynamics in Arabidopsis [164,165].
In addition to influencing qualitative traits, DNA methylation variation may also play a critical role in shaping quantitative traits. Naturally occurring epigenetic variations are believed to contribute to the phenomenon of "missing heritability" observed in studies of both animals and plants [166]. In Arabidopsis thaliana, DNA methylation variation is closely linked to environmental changes, particularly in immune-related genes [167]. Moreover, positive selection scans have identified methylated CG sites (mCGs) in promoter regions of genes associated with specific metabolites, indicating a potential role for CG methylation in the adaptive evolution of Arabidopsis [168].
Comparative methylation analyses in crops such as maize [169,170], soybean [171], rice [172], and wheat [173] have uncovered extensive differentially methylated regions (DMRs) across representative population varieties, with some linked to domestication or environmental adaptation. Genome-wide association studies (GWAS) targeting DNA methylation profiles in 263 maize inbred lines revealed that over 60% of DMRs were not marked by single nucleotide polymorphisms (SNPs). These studies also identified DMRs associated with 156 metabolic traits, suggesting that such traits may be regulated by methylation variation rather than traditional genetic variation [168]. Although genetic variation can influence DNA methylation patterns [174,175], current evidence suggests that the role of DNA methylation in phenotypic variation has been underestimated and warrants further quantification. In tomato and maize, hybridization-induced exalleles exhibit behaviors akin to somatic mutations, and these changes can persist through multiple generations of backcrossing and self-pollination [176,177]. This finding implies that hybridization-induced epigenetic changes may contribute to phenotypic diversity in conventional breeding programs. Epigenetic recombinant inbred lines (epiRILs) provide additional evidence for the potential of epigenetic variation in crop improvement. In Arabidopsis, epiRILs display pleiotropic phenotypic variation, some of which are comparable in magnitude to the phenotypic differences observed in recombinant inbred lines (RILs) [178,179]. Similarly, artificial selection for traits such as energy efficiency in genetically uniform populations of rapeseed and rice has yielded epigenetic lines with significantly enhanced field performance, demonstrating the practical value of epigenetic selection in boosting crop yields [180,181,182].
The introduction of epigenetic variation into traditional breeding programs is often constrained by extended breeding cycles and the phenomenon of epigenetic drag, wherein undesirable epigenetic mutations are inadvertently propagated. When specific target genes are identified, epigenome editing emerges as a precise and controllable strategy for enhancing agronomic traits. Numerous epigenome editing tools have been developed in plants, facilitating the study of DNA methylation regulation [183]. These tools generally comprise a targeting module that directs proteins to specific genomic loci, a modification module that alters epigenetic states, and, optionally, an auxiliary module to modulate the activity of the modification process.
Among these, CRISPR-dCas9 systems have gained prominence due to their simplicity in design and construction compared to traditional systems like TALE and artificial zinc finger domains [165]. Beyond the standard dCas9 usage, the nuclease activity of Cas9 can also be inhibited by shortening the spacer region of guide RNAs [184]. The modifier domains responsible for inducing epigenetic changes are highly versatile, ranging from the catalytic domain of the DRM gene to prokaryotic DNA methyltransferase MQ1, mammalian DNA demethylase TET1, and RdDM pathway components, all of which have been successfully employed for DNA methylation editing in Arabidopsis [62,185,186,187]. Innovative systems such as SunTag and MS2 can also be incorporated into the targeting module to amplify the local concentration of effector domains. Although still in its nascent stages, precise epigenome editing holds significant promise for efficiently modulating the epigenetic states of specific genes in crops. To realize its full potential in breeding programs, however, several challenges must be addressed: minimizing off-target effects, enhancing the cross-generational stability of edits, and accurately predicting optimal guide RNA target sites. In mammalian cells lacking DNMT activity, the dCas9-Dnmt3a modifier exhibits genome-wide activity, though ectopic methylation typically occurs at previously methylated sites and has minimal impact on gene expression [188]. Off-target effects can be mitigated by reducing overall effector levels while simultaneously increasing their local concentration [189]. These findings underscore the necessity for tighter regulation of DNA methyltransferase or demethylase activity compared to the endonuclease activity of Cas9. Furthermore, targeted de novo methylation of thousands of promoters in human cells has demonstrated environmentally contingent stability and transcriptional responses [190]. Given that chromatin state maintenance involves self-reinforcing loops formed by multiple components, disrupting these loops and establishing new ones may require simultaneous editing of multiple epigenetic marks [191]. Additionally, achieving precise transcriptional regulation of target loci demands a comprehensive understanding of their cis-regulatory elements and trans-regulatory factors.
7. Conclusion and Prospect
The escalating frequency and severity of extreme weather events driven by global climate change pose mounting challenges to agricultural production. Stresses such as elevated temperatures, drought, salinization, and pest infestations critically undermine crop yield and quality, highlighting the pressing need to develop crop varieties with enhanced adaptability and robust stress resistance. Recent advances have unveiled the pivotal role of DNA methylation in mediating plant responses to environmental stresses. Its inherent plasticity and heritable characteristics offer a promising theoretical foundation and a valuable tool for advancing crop breeding strategies.
DNA methylation plays a pivotal role in regulating plant responses to environmental stress and disease resistance by modulating gene expression. Leveraging this mechanism, modern molecular breeding techniques can be employed to enhance crop resilience through targeted adjustments of methylation patterns in specific genes. Traditional agriculture heavily relies on fertilizers and pesticides to sustain high yields, a practice that significantly pollutes the environment and poses risks to food safety. Investigations into the role of DNA methylation in nutrient uptake and stress responses have revealed its potential to enhance nutrient use efficiency in crops, thereby reducing reliance on chemical inputs. For instance, under nitrogen-deficient conditions, certain plants improve nitrogen absorption by modifying the methylation status of key genes, offering a novel strategy for breeding nutrient-efficient crop varieties. Moreover, regulating DNA methylation can bolster the natural disease resistance of crops, thereby decreasing pesticide requirements. This dual benefit—sustaining high yields while mitigating environmental harm—positions DNA methylation as a cornerstone in the development of sustainable green agriculture. Recent advancements in genomics, transcriptomics, and epigenetics have further enriched our understanding of crop trait improvement. By integrating multi-omics datasets, researchers can unravel the complex roles of DNA methylation in gene regulatory networks with greater precision. For example, analyses of stress-resistance traits frequently link dynamic methylation changes to the upregulation or downregulation of specific genes. Multi-omics approaches thus enable the identification of key regulatory genes and pathways, providing a robust scientific foundation for precision breeding.
The dynamic and multifaceted nature of DNA methylation offers immense potential for advancing crop breeding, yet it also presents significant challenges. Environmental factors influence DNA methylation patterns, raising critical questions about their stability and heritability that warrant further investigation. Moreover, the regulatory mechanisms of DNA methylation vary across species and genomic contexts, necessitating careful consideration when developing universal breeding strategies. The precision and safety of epigenetic editing technologies likewise require further refinement to enable their broad application in agriculture.Looking ahead, deeper insights into the role of DNA methylation in plant growth, development, and stress responses, combined with cutting-edge approaches such as genome and epigenetic editing, could revolutionize the breeding of high-yield, high-quality, and stress-resilient crops. International collaboration and interdisciplinary integration will be crucial for building comprehensive epigenetic data repositories spanning diverse crop species, thereby providing robust datasets and technical resources to advance breeding efforts.
In conclusion, harnessing DNA methylation in crop breeding has the potential to address pressing challenges in global agriculture while fostering the development of efficient, sustainable, and environmentally friendly farming practices. This field of research is poised to drive a transformative revolution in crop breeding, enhancing global food security and agricultural productivity.
Author Contributions
Conceptualization, S.Q. and W.S.; methodology, S.Q. and W.S.; software, S.T.; validation, A.L.; formal analysis,F.W.; data curation, W.H.; writing—original draft preparation, S.T. and T.W; writing—review and editing, S.T, S.Q. and T.W ; project administration, S.Y. and W.T.; funding acquisition, S.Y. and W.T. All authors have read and agreed to the published version of the manuscript.”.
Funding
This research was supported by the earmarked fund for China Agriculture Research System, grant number CARS10; Sichuan Science and Technology Program, grant number 2021YFYZ0019, 2021YFYZ0019 and 2024YFHZ0255.
Data Availability Statement
The original contributions presented in the study are included in the article, further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Berger, S.L.; Kouzarides, T.; Shiekhattar, R.; Shilatifard, A. An operational definition of epigenetics. Genes Dev. 2009, 23, 781–783.
- Henderson, I.R.; Jacobsen, S.E. Epigenetic inheritance in plants. Nature 2007, 447, 418–424.
- Maeji, H.; Nishimura, T. Epigenetic Mechanisms in Plants. Adv. Bot. Res. 2018, 88, 21–47.
- Fransz, P.F.; de Jong, J.H. Chromatin dynamics in plants. Curr. Opin. Plant Biol. 2002, 5, 560–567.
- Zilberman, D.; Gehring, M.; Tran, R.K.; Ballinger, T.; Henikoff, S. Genome-wide analysis of Arabidopsis thaliana DNA methylation uncovers an interdependence between methylation and transcription. Nat. Genet. 2007, 39, 61–69.
- Zhang, M.; Kimatu, J.N.; Xu, K.; Liu, B. DNA cytosine methylation in plant development. J. Genet. Genom. 2010, 37, 1–12.
- Ashapkin, V.V.; Kutueva, L.I.; Aleksandrushkina, N.I.; Vanyushin, B.F. Epigenetic Mechanisms of Plant Adaptation to Biotic and Abiotic Stresses. Int. J. Mol. Sci. 2020, 21, 7457.
- Li, E., Bestor, T. H. & Jaenisch, R. Targeted mutation of the DNA methyltransferase gene results in embryonic lethality. Cell 69, 915–926 (1992).
- Okano, M., Bell, D. W., Haber, D. A. & Li, E. DNA methyltransferases Dnmt3a and Dnmt3b are essential for de novo methylation and mammalian development. Cell 99, 247–257 (1999).
- Alleman, M. et al. An RNA-dependent RNA polymerase is required for paramutation in maize. Nature 442, 295–298 (2006).
- Wyatt, G. R. Occurrence of 5-methylcytosine in nucleic acids. Nature 166, 237–238 (1950).
- Vidalis, A. et al. Methylome evolution in plants. Genome Biol. 17, 264 (2016).
- Zhu, J. K. Active DNA demethylation mediated by DNA glycosylases. Annu. Rev. Genet. 43, 143–166 (2009).
- Wu, S. C. & Zhang, Y. Active DNA demethylation: many roads lead to Rome. Nat. Rev. Mol. Cell Biol. 11, 607–620 (2010).
- Law, J. A. & Jacobsen, S. E. Establishing, maintaining and modifying DNA methylation patterns in plants and animals. Nat. Rev. Genet. 11, 204–220 (2010).
- He, X. J., Chen, T. & Zhu, J. K. Regulation and function of DNA methylation in plants and animals. Cell Res. 21, 442–465 (2011).
- Law, J. A. & Jacobsen, S. E. Establishing, maintaining and modifying DNA methylation patterns in plants and animals. Nat. Rev. Genet. 11, 204–220 (2010).
- Stroud, H., Greenberg, M. V., Feng, S., Bernatavichute, Y. V. & Jacobsen, S. E. Comprehensive analysis of silencing mutants reveals complex regulation of the Arabidopsis methylome. Cell 152, 352–364 (2013).
- Stroud, H. et al. Non-CG methylation patterns shape the epigenetic landscape in Arabidopsis. Nat. Struct. Mol. Biol. 21, 64–72 (2014).
- Matzke, M. A. & Mosher, R. A. RNA-directed DNA methylation: an epigenetic pathway of increasing complexity. Nat. Rev. Genet. 15, 394–408 (2014).
- Zhang, H. & Zhu, J. K. RNA-directed DNA methylation. Curr. Opin. Plant Biol. 14, 142–147 (2011).
- Pikaard, C. S., Haag, J. R., Pontes, O. M., Blevins, T. & Cocklin, R. A transcription fork model for Pol IV and Pol V-dependent RNA-directed DNA methylation. Cold Spring Harb. Symp. Quant. Biol. 77, 205–212 (2012).
- Zhong, X. et al. Molecular mechanism of action of plant DRM de novo DNA methyltransferases. Cell 157, 1050–1060 (2014).
- Gao, Z. et al. An RNA polymerase II- and AGO4-associated protein acts in RNA-directed DNA methylation. Nature 465, 106–109 (2010).
- Vaucheret, H. & Voinnet, O. The plant siRNA landscape. Plant Cell 36, 246–275 (2023).
- Creasey, K. M. et al. miRNAs trigger widespread epigenetically activated siRNAs from transposons in Arabidopsis. Nature 508, 411–415 (2014).
- Cuerda-Gil, D. & Slotkin, R. K. Non-canonical RNA-directed DNA methylation. Nat. Plants 2, 16163 (2016).
- He, X. J. et al. An effector of RNA-directed DNA methylation in Arabidopsis is an ARGONAUTE 4-and RNA-binding protein. Cell 137, 498–508 (2009).
- Bies-Etheve, N. et al. RNA-directed DNA methylation requires an AGO4-interacting member of the SPT5 elongation factor family. EMBO Rep. 10, 649–654 (2009).
- Zhang, H. et al. An Rrp6-like protein positively regulates noncoding RNA levels and DNA methylation in Arabidopsis. Mol. Cell 54, 418–430 (2014).
- Ausin, I., Mockler, T. C., Chory, J. & Jacobsen, S. E. IDN1 and IDN2 are required for de novo DNA methylation in Arabidopsis thaliana. Nat. Struct. Mol. Biol. 16, 1325–1327 (2009).
- Zheng, Z. et al. An SGS3-like protein functions in RNA-directed DNA methylation and transcriptional gene silencing in Arabidopsis. Plant J. 62, 92–99 (2010).
- Ausin, I. et al. INVOLVED IN DE NOVO 2-containing complex involved in RNA-directed DNA methylation in Arabidopsis. Proc. Natl Acad. Sci. USA 109, 8374–8381 (2012).
- Zhang, C. J. et al. IDN2 and its paralogs form a complex required for RNA-directed DNA methylation. PLoS Genet. 8, e1002693 (2012).
- Zhang, H. et al. DTF1 is a core component of RNA-directed DNA methylation and may assist in the recruitment of Pol IV. Proc. Natl Acad. Sci. USA 110, 8290–8295 (2013).
- Law, J. A. et al. Polymerase IV occupancy at RNA-directed DNA methylation sites requires SHH1. Nature 498, 385–389 (2013).
- Wang, Y. et al. ZMP recruits and excludes Pol IV-mediated DNA methylation in a site-specific manner. Sci. Adv. 8, eadc9454 (2022).
- Zhou, M. et al. The CLASSY family controls tissue-specific DNA methylation patterns in Arabidopsis. Nat. Commun. 13, 244 (2022).
- Zhou, M., Palanca, A. M. S. & Law, J. A. Locus-specific control of the de novo DNA methylation pathway in Arabidopsis by the CLASSY family. Nat. Genet. 50, 865–873 (2018).
- Kanno, T. et al. Involvement of putative SNF2 chromatin remodeling protein DRD1 in RNA-directed DNA methylation. Curr. Biol. 14, 801–805 (2004).
- Kanno, T. et al. A structural-maintenance-of-chromosomes hinge domain-containing protein is required for RNA-directed DNA methylation. Nat. Genet. 40, 670–675 (2008).
- Law, J. A. et al. A protein complex required for polymerase V transcripts and RNA- directed DNA methylation in Arabidopsis. Curr. Biol. 20, 951–956 (2010).
- Zhong, X. H. et al. DDR complex facilitates global association of RNA polymerase V to promoters and evolutionarily young transposons. Nat. Struct. Mol. Biol. 19, 870–875 (2012).
- Liu, Z. W. et al. The SET domain proteins SUVH2 and SUVH9 are required for Pol V occupancy at RNA-directed DNA methylation loci. PLoS Genet. 10, e1003948 (2014).
- Johnson, L. M. et al. SRA- and SET-domain-containing proteins link RNA polymerase V occupancy to DNA methylation. Nature 507, 124–128 (2014). References 35 and 37 reveal how POL V may be recruited to some RdDM loci.
- Wierzbicki, A. T., Haag, J. R. & Pikaard, C. S. Noncoding transcription by RNA polymerase Pol IVb/Pol V mediates transcriptional silencing of overlapping and adjacent genes. Cell 135, 635–648 (2008).
- levins, T. et al. Identification of Pol IV and RDR2-dependent precursors of 24 nt siRNAs guiding de novo DNA methylation in Arabidopsis. eLife 4, e09591 (2015).
- Li, S. F. et al. Detection of Pol IV/RDR2-dependent transcripts at the genomic scale in Arabidopsis reveals features and regulation of siRNA biogenesis. Genome Res. 25, 235–245 (2015).
- Zhai, J. et al. A one precursor one siRNA model for Pol IV-dependent siRNA biogenesis. Cell 163, 445–455 (2015).
- Yang, D. L. et al. Dicer-independent RNA-directed DNA methylation in Arabidopsis. Cell Res. 26, 1264 (2016).
- Ye, R. Q. et al. A Dicer-independent route for biogenesis of siRNAs that direct DNA methylation in Arabidopsis. Mol. Cell 61, 222–235 (2016).
- Zheng, B. et al. Intergenic transcription by RNA polymerase II coordinates Pol IV and Pol V in siRNA-directed transcriptional gene silencing in Arabidopsis. Genes Dev. 23, 2850–2860 (2009).
- Duan, C. G. et al. Specific but interdependent functions for Arabidopsis AGO4 and AGO6 in RNA-directed DNA methylation. EMBO J. 34, 581–592 (2015).
- McCue, A. D. et al. ARGONAUTE 6 bridges transposable element mRNA-derived siRNAs to the establishment of DNA methylation. EMBO J. 34, 20–35 (2015).
- Mari-Ordonez, A. et al. Reconstructing de novo silencing of an active plant retrotransposon. Nat. Genet. 45, 1029–1039 (2013).
- Huang, C. F. et al. A pre-mRNA-splicing factor is required for RNA-directed DNA methylation in Arabidopsis. PLoS Genet. 9, e1003779 (2013).
- Dou, K. et al. The PRP6-like splicing factor STA1 is involved in RNA-directed DNA methylation by facilitating the production of Pol V-dependent scaffold RNAs. Nucleic Acids Res. 41, 8489–8502 (2013).
- Du, J., Johnson, L. M., Jacobsen, S. E. & Patel, D. J. DNA methylation pathways and their crosstalk with histone methylation. Nat. Rev. Mol. Cell Biol. 16, 519–532 (2015).
- Riggs, A. D. X inactivation, differentiation, and DNA methylation. Cytogenet. Cell Genet. 14, 9–25 (1975).
- Holliday, R. & Pugh, J. E. DNA modification mechanisms and gene activity during development. Science 187, 226–232 (1975).
- Bashtrykov, P. et al. Specificity of Dnmt1 for methylation of hemimethylated CpG sites resides in its catalytic domain. Chem. Biol. 19, 572–578 (2012).
- Song, J., Teplova, M., Ishibe-Murakami, S. & Patel, D. J. Structure-based mechanistic insights into DNMT1-mediated maintenance DNA methylation. Science 335, 709–712 (2012).
- Adam, S., Klingel, V., Radde, N. E., Bashtrykov, P. & Jeltsch, A. On the accuracy of the epigenetic copy machine: comprehensive specificity analysis of the DNMT1 DNA methyltransferase. Nucleic Acids Res. 51, 6622–6633 (2023).
- Bostick, M. et al. UHRF1 plays a role in maintaining DNA methylation in mammalian cells. Science 317, 1760–1764 (2007).
- Sharif, J. et al. The SRA protein Np95 mediates epigenetic inheritance by recruiting Dnmt1 to methylated DNA. Nature 450, 908–912 (2007).
- Woo, H. R., Dittmer, T. A. & Richards, E. J. Three SRA-domain methylcytosine-binding proteins cooperate to maintain global CpG methylation and epigenetic silencing in Arabidopsis. PLoS Genet. 4, e1000156 (2008).
- Petryk, N., Bultmann, S., Bartke, T. & Defossez, P. A. Staying true to yourself: mechanisms of DNA methylation maintenance in mammals. Nucleic Acids Res. 49, 3020–3032 (2021).
- Rocha, P. S. et al. The Arabidopsis HOMOLOGY-DEPENDENT GENE SILENCING1 gene codes for an S-adenosyl-L-homocysteine hydrolase required for DNA methylation-dependent gene silencing. Plant Cell 17, 404–417 (2005).
- Zhang, H. et al. Sulfamethazine suppresses epigenetic silencing in Arabidopsis by impairing folate synthesis. Plant Cell 24, 1230–1241 (2012).
- Zhou, H. R. et al. Folate polyglutamylation is involved in chromatin silencing by maintaining global DNA methylation and histone H3K9 dimethylation in Arabidopsis. Plant Cell 25, 2545–2559 (2013).
- Gong, Z. et al. ROS1, a repressor of transcriptional gene silencing in Arabidopsis, encodes a DNA glycosylase/lyase. Cell 111, 803–814 (2002). This study provides a genetic evidence for base excision repair-mediated active DNA demethylation.
- Gehring, M. et al. DEMETER DNA glycosylase establishes MEDEA polycomb gene self-imprinting by allele-specific demethylation. Cell 124, 495–506 (2006).
- Wu, X. & Zhang, Y. TET-mediated active DNA demethylation: mechanism, function and beyond. Nat. Rev. Genet. 18, 517–534 (2017).
- Agius, F., Kapoor, A. & Zhu, J. K. Role of the Arabidopsis DNA glycosylase/lyase ROS1 in active DNA demethylation. Proc. Natl Acad. Sci. USA 103, 11796–11801 (2006).
- Morales-Ruiz, T. et al. DEMETER and REPRESSOR OF SILENCING 1 encode 5-methylcytosine DNA glycosylases. Proc. Natl Acad. Sci. USA 103, 6853–6858 (2006).
- Penterman, J. et al. DNA demethylation in the Arabidopsis genome. Proc. Natl Acad. Sci. USA 104, 6752–6757 (2007).
- Huh, J. H., Bauer, M. J., Hsieh, T. F. & Fischer, R. L. Cellular programming of plant gene imprinting. Cell 132, 735–744 (2008).
- Martinez-Macias, M. I. et al. A DNA 3 ‘ phosphatase functions in active DNA demethylation in Arabidopsis. Mol. Cell 45, 357–370 (2012).
- Lee, J. et al. AP endonucleases process 5-methylcytosine excision intermediates during active DNA demethylation in Arabidopsis. Nucleic Acids Res. 42, 11408–11418 (2014).
- Raja P, Sanville BC, Buchmann RC, Bisaro DM (2008) Viral genome methylation as an epigenetic defense against geminiviruses. J Virol 82:8997–9007.
- Zhang, H. & Zhu, J. K. Active DNA demethylation in plants and animals. Cold Spring Harb. Symp. Quant. Biol. 77, 161–173 (2012).
- Lei, M. G. et al. Regulatory link between DNA methylation and active demethylation in Arabidopsis. Proc. Natl Acad. Sci. USA 112, 3553–3557 (2015).
- Williams, B. P., Pignatta, D., Henikoff, S. & Gehring, M. Methylation-sensitive expression of a DNA demethylase gene serves as an epigenetic rheostat. PLOS Genet. 11, e1005142 (2015).
- Domcke, S. et al. Competition between DNA methylation and transcription factors determines binding of NRF1. Nature 528, 575–579 (2015).
- Zhu, H., Wang, G. H. & Qian, J. Transcription factors as readers and effectors of DNA methylation. Nat. Rev. Genet. 17, 551–565 (2016).
- Tang, K., Lang, Z., Zhang, H. & Zhu, J. K. The DNA demethylase ROS1 targets genomic regions with distinct chromatin modifications. Nat. Plants 2, 16169 (2016).
- Lei, M. G. et al. Regulatory link between DNA methylation and active demethylation in Arabidopsis. Proc. Natl Acad. Sci. USA 112, 3553–3557 (2015).
- Feng, Z. et al. Genetic analysis implicates a molecular chaperone complex in regulating epigenetic silencing of methylated genomic regions. J. Integr. Plant Biol. 63, 1451–1461 (2021).
- Qian, W. et al. Regulation of active DNA demethylation by an alpha-crystallin domain protein in Arabidopsis. Mol. Cell 55, 361–371 (2014).
- Boone, B. A. et al. ACD15, ACD21, and SLN regulate the accumulation and mobility of MBD6 to silence genes and transposable elements. Sci. Adv. 9, eadi9036 (2023).
- Duan, C. G. et al. A pair of transposon-derived proteins function in a histone acetyltransferase complex for active DNA demethylation. Cell Res. 27, 226–240 (2017).
- Haslbeck, M., Weinkauf, S. & Buchner, J. Small heat shock proteins: simplicity meets complexity. J. Biol. Chem. 294, 2121–2132 (2019).
- Cokus, S. J. et al. Shotgun bisulphite sequencing of the Arabidopsis genome reveals DNA methylation patterning. Nature 452, 215–219 (2008).
- Du, J. M. et al. Dual binding of chromomethylase domains to H3K9me2-containing nucleosomes directs DNA methylation in plants. Cell 151, 167–180 (2012).
- Li, Q. et al. RNA-directed DNA methylation enforces boundaries between heterochromatin and euchromatin in the maize genome. Proc. Natl Acad. Sci. USA 112, 14728–14733 (2015).
- Zakrzewski, F., Schmidt, M., Van Lijsebettens, M. & Schmidt, T. DNA methylation of retrotransposons, DNA transposons and genes in sugar beet (Beta vulgaris L.). Plant J. 90, 1156–1175 (2017).
- Gouil, Q. & Baulcombe, D. C. DNA methylation signatures of the plant chromomethyltransferases. PLoS Genet. 12, e1006526 (2016).
- Grob, S., Schmid, M. W. & Grossniklaus, U. Hi-C analysis in Arabidopsis identifies the KNOT, a structure with similarities to the flamenco Locus of Drosophila. Mol. Cell 55, 678–693 (2014).
- Feng, S. et al. Genome-wide Hi-C analyses in wild-type and mutants reveal high-resolution chromatin interactions in Arabidopsis. Mol. Cell 55, 694–707 (2014).
- Rowley, M. J., Rothi, M. H., Bohmdorfer, G., Kucinski, J. & Wierzbicki, A. T. Long-range control of gene expression via RNA-directed DNA methylation. PLoS Genet. 13, e1006749 (2017).
- Gehring, M., Bubb, K. L. & Henikoff, S. Extensive demethylation of repetitive elements during seed development underlies gene imprinting. Science 324, 1447–1451 (2009).
- Hsieh, T. F. et al. Genome-wide demethylation of Arabidopsis endosperm. Science 324, 1451–1454 (2009).
- Ibarra, C. A. et al. Active DNA demethylation in plant companion cells reinforces transposon methylation in gametes. Science 337, 1360–1364 (2012).
- Park, K. et al. DNA demethylation is initiated in the central cells of Arabidopsis and rice. Proc. Natl Acad. Sci. USA 113, 15138–15143 (2016).
- Slotkin, R. K. et al. Epigenetic reprogramming and small RNA silencing of transposable elements in pollen. Cell 136, 461–472 (2009).
- Martínez, G., Panda, K., Köhler, C. & Slotkin, R. K. Silencing in sperm cells is directed by RNA movement from the surrounding nurse cell. Nat. Plants. 2, 16030 (2016).
- Ingouff, M. Live-cell analysis of DNA methylation during sexual reproduction in Arabidopsis reveals context and sex-specific dynamics controlled by noncanonical RdDM. Genes Dev. 31, 72–83 (2017).
- Walker, J. Sexual-lineage-specific DNA methylation regulates meiosis in Arabidopsis. Nat. Genet. 50, 130–137 (2018).
- Zhang, M. et al. Genome-wide high resolution parental-specific DNA and histone methylation maps uncover patterns of imprinting regulation in maize. Genome Res. 24, 167–176 (2014).
- Klosinska, M., Picard, C. L. & Gehring, M. Conserved imprinting associated with unique epigenetic signatures in the Arabidopsis genus. Nat. Plants 2, 16145 (2016).
- Rodrigues, J. A. et al. Imprinted expression of genes and small RNA is associated with localized hypomethylation of the maternal genome in rice endosperm. Proc. Natl Acad. Sci. USA 110, 7934–7939 (2013).
- Kinoshita, T. et al. One-way control of FWA imprinting in Arabidopsis endosperm by DNA methylation. Science 303, 521–523 (2004).
- Hsieh, T. F. et al. Regulation of imprinted gene expression in Arabidopsis endosperm. Proc. Natl Acad. Sci. USA 108, 1755–1762 (2011).
- Kinoshita, T. et al. One-way control of FWA imprinting in Arabidopsis endosperm by DNA methylation. Science 303, 521–523 (2004).
- Bratzel, F. et al. Regulation of the new Arabidopsis imprinted gene AtBMI1C requires the interplay of different epigenetic mechanisms. Mol. Plant 5, 260–269 (2012).
- Moreno-Romero, J., Jiang, H., Santos-Gonzalez, J. & Kohler, C. Parental epigenetic asymmetry of PRC2-mediated histone modifications in the Arabidopsis endosperm. EMBO J. 35, 1298–1311 (2016).
- Dong, X. M. et al. Dynamic and antagonistic allele-Specific epigenetic modifications controlling the expression of imprinted genes in maize endosperm. Mol. Plant 10, 442–455 (2017).
- Baubec, T., Finke, A., Scheid, O. M. & Pecinka, A. Meristem-specific expression of epigenetic regulators safeguards transposon silencing in Arabidopsis. EMBO Rep. 15, 446–452 (2014).
- Kawakatsu, T. et al. Unique cell-type-specific patterns of DNA methylation in the root meristem. Nat. Plants 2, 16058 (2016). This study demonstrates diversified epigenomes in different somatic cell types in A. thaliana roots.
- Zhou, S. L. et al. Cooperation between the H3K27me3 chromatin mark and non-CG methylation in epigenetic regulation. Plant Physiol. 172, 1131–1141 (2016).
- Candaele, J. et al. Differential methylation during maize leaf growth targets developmentally regulated genes. Plant Physiol. 164, 1350–1364 (2014).
- Yamamuro, C. et al. Overproduction of stomatal lineage cells in Arabidopsis mutants defective in active DNA demethylation. Nat. Commun. 5, 4062 (2014).
- Wang, Y. H., Xue, X. Y., Zhu, J. K. & Dong, J. Demethylation of ERECTA receptor genes by IBM1 histone demethylase affects stomatal development. Development 143, 4452–4461 (2016).
- Hieber, A.D.; Mudalige-Jayawickrama, R.G.; Kuehnle, A.R. Color genes in the orchid Oncidium Gower Ramsey: Identification, expression, and potential genetic instability in an interspecific cross. Planta 2006, 223, 521–531.
- Chiou, C.Y.; Pan, H.A.; Chuang, Y.N.; Yeh, K.W. Differential expression of carotenoid-related genes determines diversified carotenoid coloration in floral tissues of Oncidium cultivars. Planta 2010, 232, 937–948.
- Liu, X.J.; Chuang, Y.N.; Chiou, C.Y.; Chin, D.C.; Shen, F.Q.; Yeh, K.W. Methylation effect on chalcone synthase gene expression determines anthocyanin pigmentation in floral tissues of two Oncidium orchid cultivars. Planta 2012, 236, 401–409.
- Han, M.L.; Yin, J.; Zhao, Y.H.; Sun, X.W.; Meng, J.X.; Zhou, J.; Shen, T.; Li, H.H.; Zhang, F. How the Color Fades from Malus halliana Flowers: Transcriptome Sequencing and DNA Methylation Analysis. Front. Plant Sci. 2020, 11, 576054.
- Yuan, X.; Ma, K.; Zhang, M.; Wang, J.; Zhang, Q. Integration of Transcriptome and Methylome Analyses Provides Insight Into the Pathway of Floral Scent Biosynthesis in Prunus mume. Front. Genet. 2021, 12, 779557.
- Zhong, S. et al. Single-base resolution methylomes of tomato fruit development reveal epigenome modifications associated with ripening. Nat. Biotechnol. 31, 154–159 (2013).
- Liu, R. et al. A DEMETER-like DNA demethylase governs tomato fruit ripening. Proc. Natl Acad. Sci. USA 112, 10804–10809 (2015).
- Lang, Z. et al. Critical roles of DNA demethylation in the activation of ripening-induced genes and inhibition of ripening-repressed genes in tomato fruit. Proc. Natl Acad. Sci. USA 114, E4511–E4519 (2017).
- Telias, A. et al. Apple skin patterning is associated with differential expression of MYB10. BMC Plant Biol. 11, 93 (2011).
- El-Sharkawy, I., Liang, D. & Xu, K. N. Transcriptome analysis of an apple (Malus x domestica) yellow fruit somatic mutation identifies a gene network module highly associated with anthocyanin and epigenetic regulation. J. Exp. Bot. 66, 7359–7376 (2015).
- Daccord, N. et al. High-quality de novo assembly of the apple genome and methylome dynamics of early fruit development. Nat. Genet. 49, 1099–1106 (2017).
- Tollefson J (2020) How hot will earth get by 2100? Nature 580:443–445.
- Zandalinas SI, Mittler R, Balfagón D, Arbona V, Gómez-Cadenas A (2018) Plant adaptations to the combination of drought and high temperatures. Physiol Plant 162:2–12.
- Gao G, Li J, Li H et al (2014) Comparison of the heat stress induced variations in DNA methylation between heat-tolerant and heat-sensitive rapeseed seedlings. Breed Sci 64:125–133.
- Singh RK, Jaishankar J, Muthamilarasan M, Shweta S, Dangi A, Prasad M (2016) Genome-wide analysis of heat shock proteins in C4 model, foxtail millet identifies potential candidates for crop improvement under abiotic stress. Sci Rep 6:32641.
- Qian Y, Hu W, Liao J, Zhang J, Ren Q (2019) The dynamics of DNA methylation in the maize (Zea mays L.) inbred line B73 response to heat stress at the seedling stage. Biochem Biophys Res Commun 512:742–749.
- Lamelas L, Valledor L, Escandón M, Pinto G, Cañal MJ, Meijón M (2020) Integrative analysis of the nuclear proteome in Pinus radiata reveals thermopriming coupled to epigenetic regulation. J Exp Bot 71:2040–2057.
- Liu H, Able AJ, Able JA (2021a) Priming crops for the future: rewiring stress memory. Trends Plant Sci 11:1360–1385.
- Yu A, Lepère G, Jay F, Wang J, Bapaume L, Wang Y, Abraham AL, Penterman J, Fischer RL, Voinnet O, Navarro L (2013) Dynamics and biological relevance of DNA demethylation in Arabidopsis antibacterial defense. Proc Natl Acad Sci USA 110:2389–2394.
- Lafon-Placette C, Le Gac AL, Chauveau D et al (2018) Changes in the epigenome and transcriptome of the poplar shoot apical meristem in response to water availability affect preferentially hormone pathways. J Exp Bot 69:537–551.
- Arora H, Singh RK, Sharma S, Sharma N, Panchal A, Das T, Prasad A, Prasad M. DNA methylation dynamics in response to abiotic and pathogen stress in plants. Plant Cell Rep. 2022 Oct;41(10):1931-1944.
- Herman, J. J., & Sultan, S. E. (2016). DNA methylation mediates stress-induced transgenerational plasticity in plants. Proceedings of the National Academy of Sciences, 113(32), 8804–8809.
- Cai Q, Qiao L, Wang M, He B, Lin FM, Palmquist J, Huang SD, Jin H (2018) Plants send small RNAs in extracellular vesicles to fungal pathogen to silence virulence genes. Science 360:1126–1129.
- Ventouris, Y., et al. (2020). Adaptive epigenetic memory in alfalfa subjected to recurrent drought. Environmental and Experimental Botany, 179, 104218.
- Sallam, H., & Moussa, M. (2021). Amplified polymorphic DNA-based insights into drought tolerance in maize. Molecular Biology Reports, 48, 5647–5658.
- Wang, M., et al. (2015). Epigenetic basis of plant responses to salinity stress. Journal of Experimental Botany, 66(3), 809–820.
- Kumar, S., et al. (2020). DNA methylation dynamics under salt stress in crop plants. Plant Physiology and Biochemistry, 157, 192–202.
- Song, Q., et al. (2019). Role of epigenetics in plant stress responses and adaptation. Plant, Cell & Environment, 42(3), 583–589.
- Dowen, R. H., Pelizzola, M., Schmitz, R. J., Lister, R., Dowen, J. M., Nery, J. R., Dixon, J. E., & Ecker, J. R. (2012). Widespread dynamic DNA methylation in response to biotic stress. Proceedings of the National Academy of Sciences, 109(32), E2183–E2191.
- Yu, A., Lepère, G., Jay, F., Wang, J., Bapaume, L., Wang, Y., ... & Navarro, L. (2013). Dynamics and biological relevance of DNA demethylation in Arabidopsis antibacterial defense. Proceedings of the National Academy of Sciences, 110(6), 2389–2394.
- López A, Ramírez V, García-Andrade J, Flors V, Vera P (2011) The RNA silencing enzyme RNA polymerase V is required for plant immunity. PLoS Genet 7:e1002434.
- Wang, W., Pan, Y., Zhao, X., Zhu, L., Fu, B., & Li, Z. (2016). DNA methylation is critical for defense against the fungal pathogen Verticillium dahliae in Arabidopsis. The Plant Cell, 28(5), 1237–1251.
- Kou, H. P., Li, Y., Song, X. X., Ou, X. F., Xing, S. C., Ma, J., ... & Liu, B. (2011). Heritable alteration in DNA methylation induced by nitrogen-deficiency stress accompanies enhanced tolerance in rice (Oryza sativa L.). The Plant Journal, 65(5), 655–664.
- Boyko, A., & Kovalchuk, I. (2010). Transgenerational response to stress in plants and its application for breeding. Journal of Experimental Botany, 62(3), 633–644.
- Cubas, P., Vincent, C. & Coen, E. An epigenetic mutation responsible for natural variation in floral symmetry. Nature 401, 157–161 (1999).
- Zhang, H., Lang, Z. & Zhu, J. K. Dynamics and function of DNA methylation in plants. Nat. Rev. Mol. Cell Biol. 19, 489–506 (2018).
- Le TN, Schumann U, Smith NA, Tiwari S, Au PC, Zhu QH, Taylor JM, Kazan K, Llewellyn DJ, Zhang R, Dennis ES, Wang MB (2014) DNA demethylases target promoter transposable elements to positively regulate stress responsive genes in Arabidopsis. Genome Biol 15:1–18.
- Manning, K. et al. A naturally occurring epigenetic mutation in a gene encoding an SBP-box transcription factor inhibits tomato fruit ripening. Nat. Genet. 38, 948–952 (2006).
- Song, Q., Zhang, T., Stelly, D. M. & Chen, Z. J. Epigenomic and functional analyses reveal roles of epialleles in the loss of photoperiod sensitivity during domestication of allotetraploid cottons. Genome Biol. 18, 99 (2017).
- Pilu, R. et al. A paramutation phenomenon is involved in the genetics of maize low phytic acid1-241 (lpa1-241) trait. Heredity 102, 236–245 (2009).
- Kawanabe, T. et al. Role of DNA methylation in hybrid vigor in Arabidopsis thaliana. Proc. Natl Acad. Sci. USA 113, E6704–E6711 (2016).
- Zhang, Q. et al. The chromatin remodeler DDM1 promotes hybrid vigor by regulating salicylic acid metabolism. Cell Discov. 2, 16027 (2016).
- Brachi, B., Morris, G. P. & Borevitz, J. O. Genome-wide association studies in plants: the missing heritability is in the field. Genome Biol. 12, 232 (2011).
- Kawakatsu, T. et al. Epigenomic diversity in a global collection of Arabidopsis thaliana accessions. Cell 166, 492–505 (2016).
- Kawakatsu, T. et al. Epigenomic diversity in a global collection of Arabidopsis thaliana accessions. Cell 166, 492–505 (2016).
- Eichten, S. R. et al. Epigenetic and genetic influences on DNA methylation variation in maize populations. Plant Cell 25, 2783–2797 (2013).
- Xu, G. et al. Evolutionary and functional genomics of DNA methylation in maize domestication and improvement. Nat. Commun. 11, 5539 (2020).
- Shen, Y. et al. DNA methylation footprints during soybean domestication and improvement. Genome Biol. 19, 128 (2018).
- Zhao, L. et al. Integrative analysis of reference epigenomes in 20 rice varieties. Nat. Commun. 11, 2658 (2020).
- Gardiner, L. J. et al. Hidden variation in polyploid wheat drives local adaptation. Genome Res. 28, 1319–1332 (2018).
- Xu, J. et al. Population-level analysis reveals the widespread occurrence and phenotypic consequence of DNA methylation variation not tagged by genetic variation in maize. Genome Biol. 20, 243 (2019).
- Schmitz, R. J. et al. Patterns of population epigenomic diversity. Nature 495, 193–198 (2013).
- Gohlke J, Scholz CJ, Kneitz S, Weber D, Fuchs J, Hedrich R, Deeken R (2013) DNA methylation mediated control of gene expression is critical for development of crown gall tumors. PLoS Genet 9:e1003267.
- Gouil, Q. & Baulcombe, D. C. Paramutation-like features of multiple natural epialleles in tomato. BMC Genomics 19, 203 (2018).
- Cao, S. et al. Small RNAs mediate transgenerational inheritance of genome-wide trans-acting epialleles in maize. Genome Biol. 23, 53 (2022).
- Cortijo, S. et al. Mapping the epigenetic basis of complex traits. Science 343, 1145–1148 (2014).
- Zhang, Y. Y., Latzel, V., Fischer, M. & Bossdorf, O. Understanding the evolutionary potential of epigenetic variation: a comparison of heritable phenotypic variation in epiRILs, RILs, and natural ecotypes of Arabidopsis thaliana. Heredity 121, 257–265 (2018).
- Schmidt, M. et al. Methylome and epialleles in rice epilines selected for energy use efficiency. Agronomy 8, 163 (2018).
- Hauben, M. et al. Energy use efficiency is characterized by an epigenetic component that can be directed through artificial selection to increase yield. Proc. Natl Acad. Sci. USA 106, 20109–20114 (2009).
- Zhan, X., Lu, Y., Zhu, J. K. & Botella, J. R. Genome editing for plant research and crop improvement. J. Integr. Plant Biol. 63, 3–33 (2021).
- Pan, C. et al. Boosting plant genome editing with a versatile CRISPR-Combo system. Nat. Plants 8, 513–525 (2022).
- Gallego-Bartolome, J. et al. Targeted DNA demethylation of the Arabidopsis genome using the human TET1 catalytic domain. Proc. Natl Acad. Sci. USA 115, E2125–E2134 (2018).
- Ji, L. et al. TET-mediated epimutagenesis of the Arabidopsis thaliana methylome. Nat. Commun. 9, 895 (2018).
- Papikian, A., Liu, W., Gallego-Bartolome, J. & Jacobsen, S. E. Site-specific manipulation of Arabidopsis loci using CRISPR-Cas9 SunTag systems. Nat. Commun. 10, 729 (2019).
- Ghoshal, B., Picard, C. L., Vong, B., Feng, S. & Jacobsen, S. E. CRISPR-based targeting of DNA methylation in Arabidopsis thaliana by a bacterial CG-specific DNA methyltransferase. Proc. Natl Acad. Sci. USA 118, e2125016118 (2021).
- Galonska, C. et al. Genome-wide tracking of dCas9-methyltransferase footprints. Nat. Commun. 9, 597 (2018).
- Pflueger, C. et al. A modular dCas9-SunTag DNMT3A epigenome editing system overcomes pervasive off-target activity of direct fusion dCas9-DNMT3A constructs. Genome Res. 28, 1193–1206 (2018).
- Chan, W. F. et al. Activation of stably silenced genes by recruitment of a synthetic de-methylating module. Nat. Commun. 13, 5582 (2022).
- Swain, T. et al. A modular dCas9-based recruitment platform for combinatorial epigenome editing. Nucleic Acids Res. 52, 474–491 (2024).
Figure 1.
Specific DNA methyltransferases and demethylases mediate cytosine methylation in different sequence contexts. CG, CHG, and CHH methylation are carried out by MET1, CMT3, and CMT2, respectively. DRM2, involved in the RdDM pathway, regulates all sequence context methylation. ROS1, DME, DML2, and DML3 act as demethylases.
Figure 1.
Specific DNA methyltransferases and demethylases mediate cytosine methylation in different sequence contexts. CG, CHG, and CHH methylation are carried out by MET1, CMT3, and CMT2, respectively. DRM2, involved in the RdDM pathway, regulates all sequence context methylation. ROS1, DME, DML2, and DML3 act as demethylases.
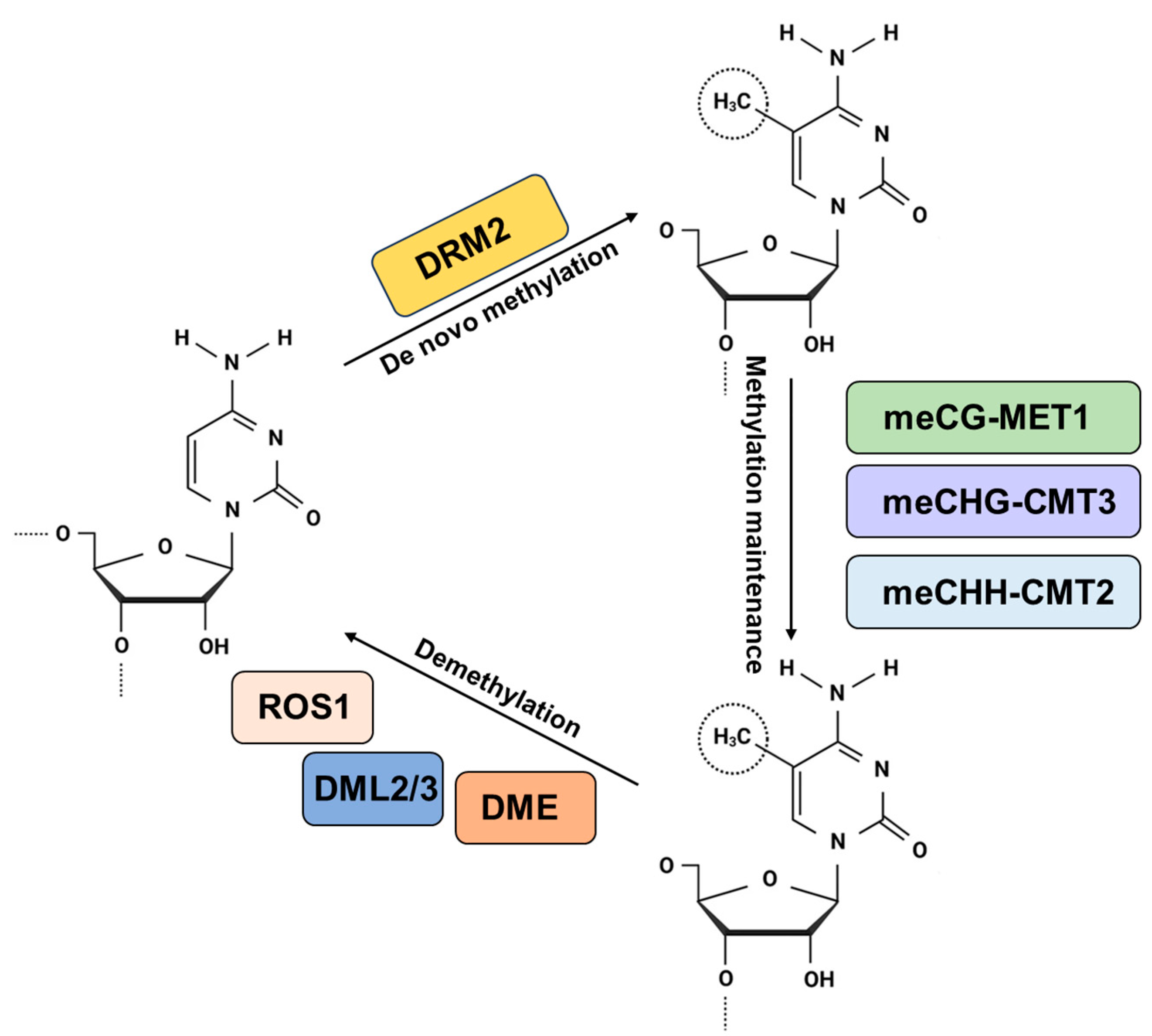
Figure 2.
Different roles of DNA methylation in plant cells. (A) DNA methylation in a transcription factor (TF) recognition motif may directly affect transcription factor binding. Most transcription factor s, including transcription activators and repressors, are sensitive to DNA methylation. When DNA methylation occurs on a cis-element bound by a transcription activator, it represses transcription. When DNA methylation occurs on a cis element bound by a transcription repressor, it derepresses transcription. A W-box motif is presented for illustration but any transcription factor motif containing C or G may apply. (B) methylation of the coding regions generates an inaccessible chromatin structure suppressing the aberrant transcription start site. (C) DNA methylation affects genome stability by silencing transposons and other DNA repeated sequences. (D) chromosome interactions through pericentromeric regions or heterochromatin islands depend on the methylation of these regions; (E) DNA methylation could be involved in the biogenesis of circRNAs.
Figure 2.
Different roles of DNA methylation in plant cells. (A) DNA methylation in a transcription factor (TF) recognition motif may directly affect transcription factor binding. Most transcription factor s, including transcription activators and repressors, are sensitive to DNA methylation. When DNA methylation occurs on a cis-element bound by a transcription activator, it represses transcription. When DNA methylation occurs on a cis element bound by a transcription repressor, it derepresses transcription. A W-box motif is presented for illustration but any transcription factor motif containing C or G may apply. (B) methylation of the coding regions generates an inaccessible chromatin structure suppressing the aberrant transcription start site. (C) DNA methylation affects genome stability by silencing transposons and other DNA repeated sequences. (D) chromosome interactions through pericentromeric regions or heterochromatin islands depend on the methylation of these regions; (E) DNA methylation could be involved in the biogenesis of circRNAs.
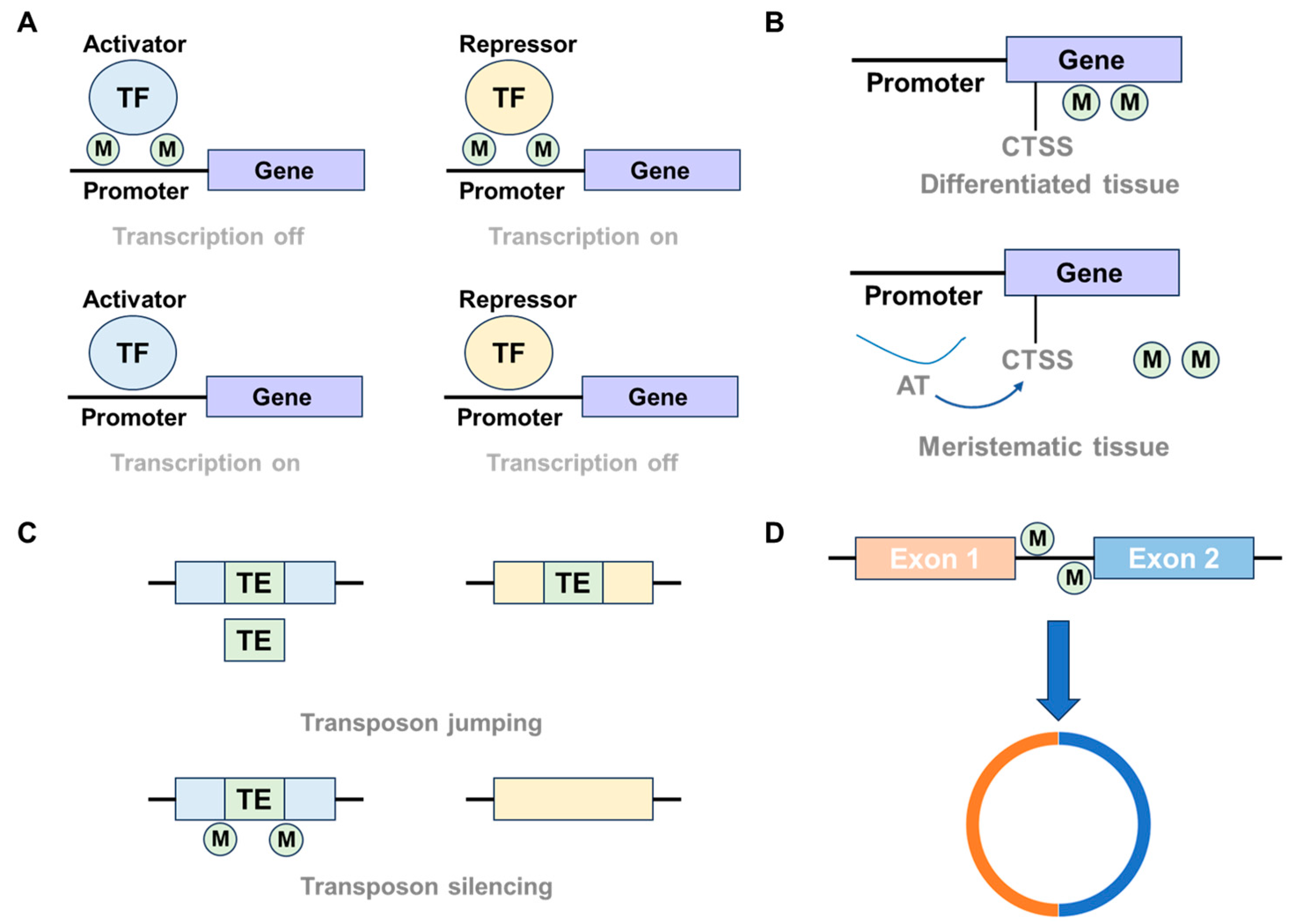

Table 1.
The defense responses of the Arabidopsis mutants with hypomethylated and hypermethylated DNA to different bacterial, viral and fungal pathogens.
Table 1.
The defense responses of the Arabidopsis mutants with hypomethylated and hypermethylated DNA to different bacterial, viral and fungal pathogens.
| Arabidopsis mutants | Pathogen | Phenotype | Defense response | References |
|---|---|---|---|---|
| DNA Hypomethylation | ||||
| drd1 | P. syringae pv. tomato DC3000 (Pst) | Resistant | Enhancement of SA-dependent defense | Dowen et al. [152] |
| P. cucumerina | Susceptible | Suppression of JA-dependent defense | López et al. [154] | |
| ago4 | B. cinerea | Susceptible | Suppression of JA-dependent defense | López et al. [154] |
| rdr2 | P. cucumerina | Susceptible | Suppression of JA-dependent defense | López et al. [154] |
| Pst | Resistant | Enhancement of SA-dependent defense | Dowen et al. [152] | |
| rdr6 | B. cinerea | Susceptible | Loss of transfer siRNAs that target pathogen genes | Cai et al. [146] |
| Pst | Resistant | − | Dowen et al. [152] | |
| nrpd1 | Pst | Resistant | Enhancement of SA-dependent defense | Dowen et al. [152] |
| nrpe1 | P. cucumerina | Susceptible | Suppression of JA-dependent defense | López et al. [154] |
| B. cinerea | Susceptible | Suppression of JA-dependent defense | López et al. [154] | |
| Pst | Resistant | Enhancement of SA-dependent defense | López et al. [154] | |
| nrpd2 | P. cucumerina | Susceptible | Suppression of JA-dependent defense | López et al. [154] |
| B. cinerea | Susceptible | Suppression of JA-dependent defense | López et al. [154] | |
| Pst | Resistant | Enhancement of SA-dependent defense | López et al. [154] | |
| nrpd1/nrpe1 | P. cucumerina | Susceptible | Suppression of JA-dependent defense | López et al. [154] |
| Pst | Resistant | Enhancement of SA-dependent defense | López et al. [154] | |
| drm1/drm2 | P. cucumerina | Susceptible | Suppression of JA-dependent defense | López et al. [154] |
| Pst | Resistant | Enhancement of SA-dependent defensePrimed state of defense response | Yu et al. [142] | |
| CaLCuV | Susceptible | − | Raja et al. [80] | |
| BCTV | Susceptible | − | Raja et al. [80] | |
| drm1/drm2/cmt3 (ddc) | A. tumefaciens | Susceptible | Enhancement of ABA-dependent response | Gohlke et al. [176] |
| Pst | Resistant | Enhancement of SA-dependent defense | Dowen et al. [152] | |
| dcl2/3/4 | B. cinerea | Susceptible | Loss of siRNAs that move into fungal cells and suppress virulence genes | Cai et al. [146] |
| Pst | Resistant | Enhancement of SA-dependent defense | Dowen et al. [152] | |
| CaLCuV | Susceptible | − | Raja et al. [80] | |
| BCTV | Susceptible | − | Raja et al. [80] | |
| DNA Hypermethylation | ||||
| ros1 | Pst | Susceptible | Methylation at the promoter of RMG1 and RLP43 | Yu et al. [142] |
| ros1/dml2/dml3 (rdd) | F. oxysporum | Susceptible | Suppression of defense-related genes | Le et al. [160] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



