Submitted:
14 October 2024
Posted:
16 October 2024
You are already at the latest version
Abstract
Alphaviruses are known for being model viruses for studying cellular functions related to viral infection but also for causing epidemics in different parts of the world. More recently, alphavirus-based expression systems have demonstrated efficacy as vaccines against infectious diseases and as therapeutic applications for different cancers. The self-replicating feature of alphaviruses has provided the advantage of extremely high transgene expression of vaccine-related antigens and therapeutic anti-tumor and immunostimulatory genes, which has also permitted significantly reduced doses for prophylactic and therapeutic applications potentially reducing adverse events. Furthermore, alphaviruses have shown favorable flexibility as they can be delivered as recombinant viral particles, RNA replicons or DNA replicon-based plasmids. In the context of infectious diseases robust immune responses against the surface proteins of target agents have been observed and protection against challenges with lethal doses of infectious agents in rodents and primates. Similarly, expression of anti-tumor genes and immunostimulatory genes from alphavirus vectors has provided tumor growth inhibition, tumor regression and cure in animal cancer models. Moreover, protection against tumor challenges has been observed. In clinical settings, clinical benefits have been reported. Alphaviruses have also been considered for the treatment of neurological disorders due to their neurotrophic preference. Attention has been paid to alphavirus vector and expression system improvement. Point mutations in the non-structural alphaviral replicase genes have generated enhanced transgene expression and created temperature-sensitive expression vectors. The recently engineered trans-amplifying RNA system has facilitated vector production and enhanced the expression capacity.
Keywords:
alphaviruses
; viral particles
; RNA replicons: DNA delivery
; infectious diseases
; cancer
; vaccines
; neurological disorders
; therapy
; clinical trials
1. Introduction
Alphaviruses are single-stranded RNA (ssRNA) viruses with a genome of positive polarity [1]. They are enveloped viruses possessing a structure consisting of trimeric spike proteins. Already in the 1980s, the life cycle of alphaviruses was described including the major steps of host cell recognition, RNA delivery to host cells, RNA replication involving the viral replicase complex, translation of viral structural proteins, nucleocapsid formation, post-translational modifications and transport to the plasma membrane and finally release of mature viral particles by budding [1]. The alphaviruses have commonly been divided into old and new-world viruses. The most commonly investigated alphaviruses are the Semliki Forest virus (SFV) [2]. Sindbis virus (SIN) [3], and Venezuelan equine encephalitis virus (VEE) [4]. The breakthrough in genetic engineering technologies made it possible to develop efficient expression systems for alphaviruses, especially for SFV [5], SIN [6], and VEE [7]. This development made great contributions to gene delivery in vitro and in vivo applied for both prophylactic and therapeutic interventions in animal models and humans in clinical trials. Moreover, the neurovirulent nature of alphaviruses has made them attractive as delivery tools in basic neuroscience and potentially for therapeutic applications in the fields of neurodegeneration and neurological disorders [8].
In this review, the application of alphaviruses in basic research on the viral life cycle and cellular functions related to post-translational modifications and transport of cellular and viral proteins is summarized. Moreover, the engineering of various alphaviral vectors, attenuated alphavirus strains, and identification of natural or engineered oncolytic alphaviruses are described. Additionally, vector and expression system development are discussed. The pathogenicity of alphaviruses resulting in epidemics is also presented. Finally, prophylactic and therapeutic interventions using alphaviruses are described.
2. The alphavirus Life Cycle
The life cycle of alphaviruses has been investigated since the 1980s as a model for RNA viruses [9] (Figure 1). In addition to determining the crystal structure of alphavirus particles [10], the virus-host interaction including host cell recognition [11], virus entry [12], RNA replication [13], nucleocapsid assembly [14], post-translational modifications and transport of viral glycoproteins [15], and the release of mature viral particles [16] have been investigated and described in detail [1]. Briefly, host cell recognition has been identified as mediated by many different types of receptors explaining the broad host range including insects, vertebrates, and amphibians. It has also been demonstrated that, for example, different neurovirulent alphaviruses use different receptors [16]. Laminin [17] and heparan sulfate [18] have been identified as common receptors on mammalian cells for alphaviruses. The entry of viral particles involves fusion with the cellular membrane [19], the delivery of nucleocapsids to the cytoplasm [20], and the release of the RNA genome [21]. Alphavirus entry has also been described to occur through endocytosis in clathrin-coated vesicles followed by transfer to endosomes [22,23]. Furthermore, it has been demonstrated that a low pH in the endosomes is essential but also a voltage potential is of importance [24]. Once the RNA genome is released, RNA replicase complexes, composed of the nonstructural proteins (nsP1-4), are assembled in modified endosomal and lysosomal membranes [25]. The RNA replicase complexes remain stable throughout the infection cycle and are responsible for the extensive viral RNA replication in host cells from a negative-strand RNA template [26]. Due to the strong subgenomic promoter, alphavirus structural proteins are expressed at high levels leading to the assembly of RNA into C protein forming nucleocapsids [14]. Simultaneously, the viral envelope proteins are glycosylated and subjected to post-translational modifications in the endoplasmic reticulum (ER) and the Golgi [15]. The process of viral glycoprotein folding is initiated immediately after entry into the ER and requires molecular chaperons, folding enzymes, and the formation of disulfide bonds [27]. Subsequently, the envelope proteins are transported through the Golgi complex to the cell membrane [28]. Finally, during the maturation of nucleocapsids surrounded by envelope proteins mature alphavirus particles are released by budding [29]. A multitude of findings has not only substantially enhanced our understanding of the molecular and cellular aspects of alphaviruses but has also enhanced our knowledge of cellular biology in general. The information on the pathogenesis and epidemiology of alphaviruses has also been enriched as described below.
3. Pathogenesis and Epidemiology of Alphaviruses
Different alphaviruses have been identified as pathogens in both animals and humans [30]. It is well-documented that several epidemics have been caused by alphaviruses. For example, VEE has been responsible for outbreaks in South America resulting in a few fatal cases in horses and fever epidemics in humans [16]. Moreover, SFV and SIN have been associated with outbreaks in Africa [31,32] and painful polyarthritis in Northern Europe [33]. Chikungunya virus (CHIKV) has caused epidemics in the Republic of Congo [34], the island of Reunion [35], and Brazil [36] more recently. Taking this information into account, avirulent and attenuated alphavirus strains have been used for the engineering of expression systems.
On the molecular level, some of the pathogenic effects have been related to the host cell killing induced by apoptosis of alphaviruses [37]. It has been shown that apoptosis is associated with the loss of membrane potential in mitochondria followed by the activation of caspases-3, -8, and -9 [38]. Another issue relates to the shut-off of the host cell protein synthesis by alphaviruses in infected cells [39]. Additionally, it has been demonstrated that nsP2 and nsP3 associated with the shut-off specifically suppress antiviral pathways [39]. The pathogenicity of alphaviruses, especially related to their use as expression systems, has been addressed by engineering less cytotoxic vectors as described below.
4. Alphavirus Vector Development
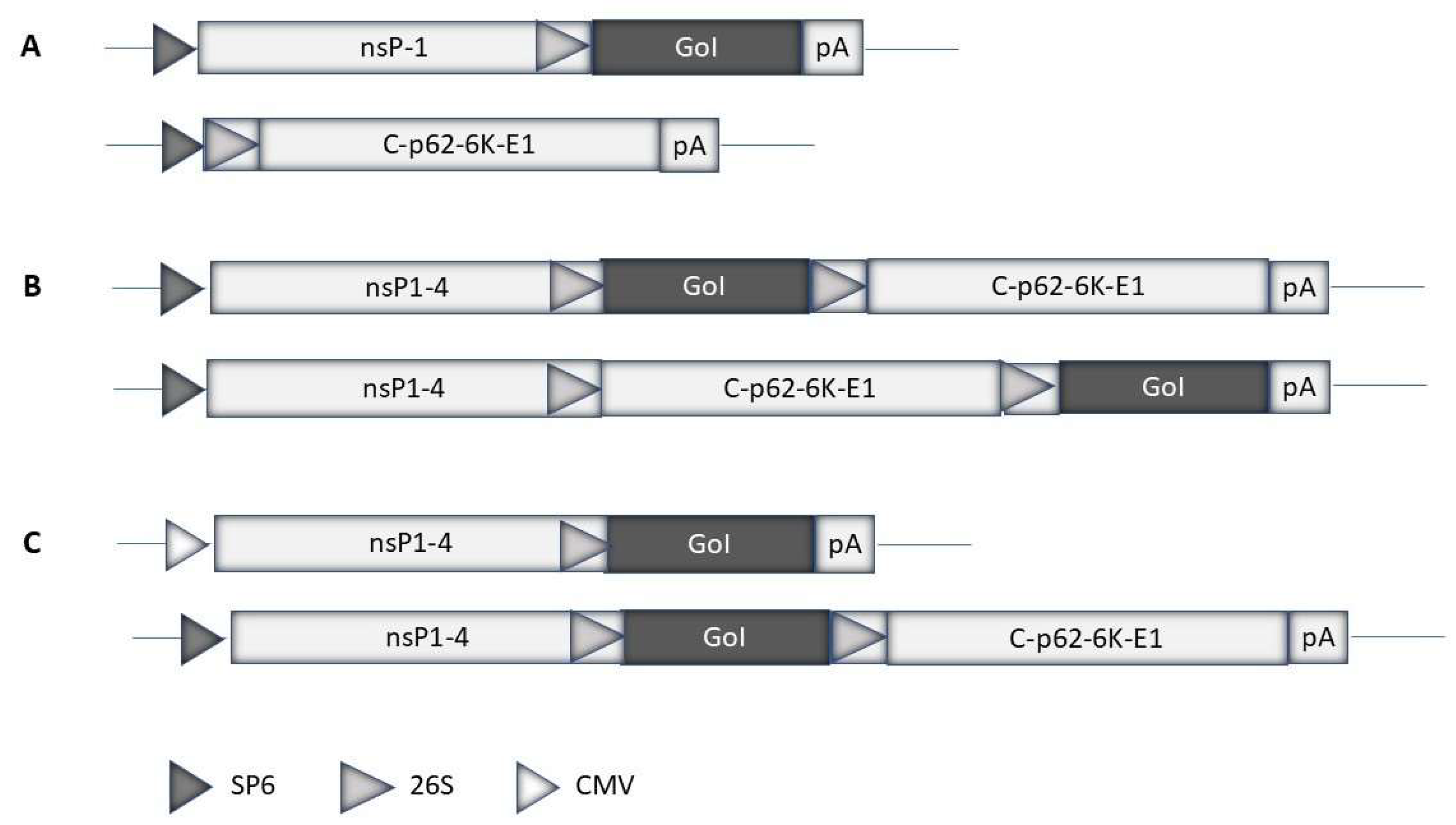
Expression systems have been engineered for different alphaviruses. The most commonly used alphavirus expression systems are based on SFV [5], SIN [6], and VEE [7]. Both replication-deficient and -competent expression systems have been engineered (Figure 2a). Generally, replication-deficient alphavirus particles are obtained by transfecting mammalian host cells with in vitro transcribed RNA from an expression vector containing the alphavirus nsP1-4 genes and the gene of interest (GoI) and a helper vector carrying the alphavirus structural protein genes [5]. Only RNA from the expression vector will be packaged in viral particles as the RNA packaging signal is located either in the nsP1 or nsP2 gene [40]. For this reason, the generated viral particles do not carry RNA coding for structural proteins and new viral particles cannot be produced. However, these “suicide particles” can infect host cells and express the GoI. Alternatively, the GoI can be introduced into the alphavirus full-length genome either downstream of the replicase genes or the structural genes (Figure 2b). In this case, in vitro transcribed RNA from a plasmid DNA template is transfected/electroporated into mammalian host cells for the production of replication-proficient recombinant particles. These particles provide expression of the GoI in infected host cells as well as the production of new virus progeny. The advantage of using replication-proficient alphaviruses is the potential spread to adjacent cells, especially in vivo, and the prolongation of transgene expression. On the other hand, safety issues must be considered to prevent any uncontrolled spread of the virus. The application of alphavirus-based expression systems is flexible as in addition to recombinant particles also RNA replicons can be used. In this case, in vitro transcribed RNA from replication-deficient and -proficient expression vectors can be directly used for delivery. However, due to the single-stranded nature of the RNA, it is highly sensitive to degradation, which strongly affects the efficacy of delivery [41]. For this reason, RNA replicons have been encapsulated in lipid nanoparticles (LNPs), which prevent RNA degradation and improve delivery [42]. Alternatively, the direct use of alphavirus DNA replicons as plasmid vectors has been possible by replacing the SP6 or T7 RNA polymerase promoter with a CMV promoter [43] (Figure 2c).
Independent of whether expression systems based on viral particles, RNA replicons, or DNA plasmids are applied, the efficient RNA amplification in host cells combined with the strong 26S subgenomic generates extremely high levels of transgene expression [44].
Despite the encouraging experience of using alphavirus expression systems during the last twenty years, many improvements have been made. For example, introductions of mutations into the replicase genes of SFV [45] and SIN [46] vectors have resulted in less cytopathogenic vectors providing enhanced and prolonged transgene expression. Moreover, temperature-sensitive mutants have been proven useful for neuroscience studies, where expression is mainly neuron-specific at 37°C while it is glial-specific at 33°C in primary neurons and hippocampal slice cultures [47]. Another approach has been to introduce the translation enhancement signal of the alphavirus capsid protein into the expression vector, which has resulted in a 5-10-fold increase in transgene expression levels [48]. Moreover, a bipartite trans-amplifying RNA (taRNA) system has been engineered, where the replicase genes have been replaced by the GoI in the expression vector, and the replicase genes are provided from an optimized non-replicating RNA (nrRNA) vector in trans [49]. This also substantially reduces the size of the expression vector and has generated a 10-100-fold increase in transgene expression [49].
5. Prophylactics and therapeutics
Applying alphavirus expression systems for vaccine and drug development has received much attention lately. As alphaviruses are known for their transient expression profile due to their relatively rapid degradation of their RNA, the application range is restricted mainly to acute diseases. Other options should be considered for chronic diseases. The main areas of interest are infectious diseases and different cancers, for which both prophylactic and therapeutic approaches have been taken. In the case of neurological disorders, the main area of applications of alphaviruses comprises the establishment of animal models for the initial evaluation of therapeutic interventions before conducting clinical trials in humans. As alphavirus-based prophylactic and therapeutic interventions have been reviewed in more detail elsewhere [50], selected examples are described below and summarized in Table 1, Table 2 and Table 3.
5.1. Infectious Diseases
In the context of infectious diseases, VEE particles expressing the Lassa virus (LASV) glycoprotein (GPC) or nucleoprotein (NP) showed protection against LASV challenges in immunized guinea pigs [51] (Table 1). Moreover, immunization of guinea pigs with VEE particles expressing the Junin virus (JUNV) or Machupo virus (MACV) were protected against challenges with JUNV and MACV, respectively [52]. In another approach targeting VEE, VEE DNA replicons expressing the VEE capsid gene protected immunized mice [53] and macaques [54] against challenges with VEE.
Prime-boost immunizations with VEE particles expressing the Dengue virus (DENV) pre-membrane protein and envelope proteins (prME) and a vaccine based on prME expression from a conventional DNA plasmid have been conducted [55]. Although three doses of plasmid DNA (DDD) or VEE particles (VVV) reduced viremia in macaques, only two doses of plasmid DNA and one dose of VEE particles (DDV) provided complete protection against DENV challenges [55]. In the case of Zika virus (ZIKV), VEE RNA replicons expressing the ZIKV prME encapsulated in nanocarrier lipids (NCLs) protected mice against ZIKV challenges after a single immunization with 10 ng of RNA [56]. SFV particles have been employed for the expression of the louping ill virus (LIV) prME and NS1 proteins [57]. In contrast to a commercial inactivated whole virus vaccine SFV-LIV-prME/NS1 particles protected 100% of mice against challenges with LIV. Moreover, SFV particles expressing the HIV-1 envelope protein (Env) showed superior antibody responses compared to mice immunized with a conventional plasmid DNA or recombinant Env protein [58]. VEE DNA replicons have also been used for the expression of Env gp160 showing similar humoral immune responses in mice to those obtained with a conventional plasmid DNA vaccine but with 10-100-fold lower doses [59]. Furthermore, VEE-Gag particles (AVX001) were subjected to a phase I trial but due to vaccine instability, the trial was prematurely terminated [60].
Due to the seasonal influenza virus epidemics, alphavirus vectors have also been applied for influenza A virus (IFVA) vaccine development. For example, only 10 μg of SFV RNA replicons expressing the IFVA HA gene protected 90% of immunized mice against IFVA challenges [61]. Moreover, approximately 100 times lower doses of VEE RNA replicons were required for the protection of mice against IFVA challenges compared to synthetic mRNA [62]. As the bipartite taRNA system generates 10-100-fold enhanced transgene expression, 50 ng of SFV-HA taRNA was sufficient to protect mice against IFVA challenges [49].
In the context of COVID-19 vaccines, VEE RNA replicons containing the prefusion-stabilized SARS-CoV-2 spike (S) protein encapsulated in LNPs elicited strong immune responses in BALB/c mice [61]. The VEE-SARS-CoV-2 S LNPs (LNP-nCoV saRNA) showed good safety and tolerability in the first-in-human phase I clinical trial and neutralization of SARS-CoV-2 was achieved in 15-48% of vaccinated individuals [64]. In a phase II study, the LNP-nCoV saRNA vaccine was administered at a prime dose of 1 μg followed by a booster vaccination with 10 μg, which resulted in higher seroconversion rates than in the phase I dose-ranging study [65]. Another RNA replicon-based vaccine candidate comprising the VEE-SARS-CoV-2 S RNA encapsulated in lipid inorganic nanoparticles (LION) elicited Th1-biased immune responses in macaques [66]. Furthermore, the safety, tolerability, and immunogenicity of the repRNA-COV2S-LION vaccine candidate were demonstrated in phase II/III [67] supporting the approval of emergency use authorization (EUA) of the vaccine in India [68].
Among non-viral diseases, anthrax caused by Bacillus anthracis has been targeted by expression of the B. anthracis protective antigen (PA) from SIN particles, which elicited PA-specific IgG and neutralizing antibodies and some protection in mice [69]. In another study, it was demonstrated that SFV DNA replicons expressing the PA gene elicited superior immune responses than those obtained after the administration of conventional plasmid DNA [66]. Furthermore, SFV-PA DNA replicons protected mice against challenges with the B. anthracis A16R strain [70]. Brucellosis has been targeted by the expression of the Brucella abortus translation initiation factor 3 (IF3) from SFV particles, which resulted in the protection of mice from challenges with B. abortus [71]. In the context of malaria, protection was achieved in mice after immunization with SIN particles expressing a cytotoxic T-lymphocyte CTL) epitope (SYVPSAEQI) of the malaria parasite [72].

Table 1.
Examples of prophylactic/therapeutic interventions with alphavirus vectors for infectious diseases.
Table 1.
Examples of prophylactic/therapeutic interventions with alphavirus vectors for infectious diseases.
| Disease | Vector / Target gene(s) | Findings |
|---|---|---|
| Infections | ||
| LASV JUNV MACV VEE DENV ZIKV LIV HIV/AIDS HIV/AIDS HIV/AIDS IFVA IFVA IFVA COVID-19 COVID-19 COVID-19 COVID-19 COVID-19 COVID-19 Anthrax Anthrax Brucellosis Malaria |
VEE RP / LASV-GPC(NP VEE RP / JUNV-GPC VEE RP / MACV-GPC VEE DNA / VEE-Capsid VEE RP / DENV-prME + pDNA VEE RNA / ZIKV-prME-NCLs VEE RP / LIV-prME/NS1 SFV RP / HIV-1 Env VEE DNA / HIV-Env gp160 VEE RP / HIV-Gag SFV RNA / IFVA-HA VEE RNA / IFVA-HA SFV taRNA / IFVA-HA VEE RNA-LNPs / SARS-CoV-2 S VEE RNA-LNPs / SARS-CoV-2 S VEE RNA-LNPs / SARS-CoV-2 S VEE RNA-LION / SARS-CoV-2 S VEE RNA-LION / SARS-CoV-2 S VEE RNA-LION / SARS-CoV-2 S SIN RP / B. anthracis PA SFV RP/DNA / B. anthracis PA SFV RP / B. abortus IF3 SIN RP / P. yoelii CTL epitope |
Protection against LASV in guinea pigs [51] Protection against JUNV in guinea pigs [52] Protection against MACV in guinea pigs [52] Protection in mice [36] and macaques [54] Complete protection after prime boost in macaques [55] Protection against ZIKV challenges in mice [56] 100% protection against LIV in mice [57] Superior Ab response to pDNA and recombinant Env [58] 10-100-fold lower doses needed than pDNA [59] Phase I terminated due to vaccine instability [60] 90% protection with 10 μg RNA replicon in mice [61] 100-fold lower doses of RNA replicons [62] 50 ng of taRNA sufficient for protection in mice [49] Robust S-specific immune response in mice [63] Good safety, immune responses in phase I [64] Higher seroconversion rates in phase II than in phase I [65] Th1-biased immunity in macaques [66] Safe, tolerable, and immune responses in phase II/III [67] EUA in India [68] Immune responses, some protection in mice [69] Protection against B. anthracis A16R strain in mice [70] Protection against B. abortus challenges in mice [71] Protection against malaria in mice [72] |
DENV, Dengue virus; Env, envelope protein; EUA, emergency use authorization; GPC, glycoprotein; IF3, translation initiation factor 3; JUNV, Junin virus; LASV, Lassa virus; LION, lipid inorganic nanoparticles; LIV, louping ill virus; LNPs, lipid nanoparticles; MACV, Machupo virus; NCLs, nanocarrier lipids; NP, nucleoprotein; pDNA, conventional plasmid DNA; PA, B. anthracis protective antigen; prME, pre-membrane and envelope proteins; RP, recombinant particles; S, spike protein; SFV, Semliki Forest virus; SIN, Sindbis virus; taRNA, trans-amplifying RNA; VEE, Venezuelan equine encephalitis virus; ZIKV, Zika virus.
5.2. Cancers
Alphavirus vectors have been applied for both prophylactic and therapeutic evaluations for different cancers (Table 2). Due to the apoptotic activity of alphaviruses, SFV particles expressing the enhanced green fluorescent protein (EGFP) effectively killed human H358a non-small cell lung cancer (NSCLC) cells in vitro and showed strong tumor regression in nu/nu mice with implanted H358a tumors [73]. Similarly, SFV-LacZ RNA elicited robust immune responses in mice [74]. Furthermore, a single administration of 0.1 μg of SFV-LacZ RNA protected mice from challenges with CT26 colon tumor cells [74].
Alphaviruses have targeted breast cancer in several studies. For instance, VEE particles expressing the extracellular domain (ECD) and the transmembrane (TM) domains of HER2 completely protected mice challenged with HER2/neu tumors [75]. In phase I, the VEE-HER2-ECD/TM particles showed good tolerability, partial response (PR), and stable disease (SD) in stage IV HER2 overexpressing breast cancer patients [75]. Moreover, SIN DNA replicons expressing the HER2/neu gene inhibited tumor growth in mice challenged with A2L2 breast tumor cells [76]. In another study, in comparison to conventional plasmid DNA delivery, SIN-HER2/neu DNA replicons required 80% less DNA for eliciting similar immune responses and protecting BALB/c mice against tumor challenges [77].
Human papillomavirus (HPV) has been the target for prevention and therapy of cervix cancer. For example, immunization with VEE particles expressing the HPV E7 protein protected C57BL/6 mice against tumor challenges [78]. Furthermore, immunization of mice carrying cervical tumors with SFV particles expressing the HPV E6-E7 fusion protein resulted in complete tumor eradication [79]. SFV-HPV E6-E7 particles were administered to patients with cervical intraepithelial neoplasia in a phase I trial, which showed good safety and elicited specific immune responses in all 12 tested patients [80]. In another approach, VEE particles expressing the carcinoembryonic antigen (CEA) induced antigen-specific immunogenicity and prolonged the survival of patients with stages III and IV colorectal cancer in phase I [81].
Combination therapy with alphavirus vectors, antibodies, and drugs has turned out to be successful. For example, the combination of VEE particles with antagonist and agonist monoclonal antibodies have proven a promising alternative. For example, VEE particles expressing the tyrosine-related protein-2 (TRP-2) combined with the antagonist anti-CTL antigen-4 (CTLA-4) monoclonal antibody (mAb) generated complete regression of B16F10 melanoma tumors in 50% of immunized mice [82]. However, VEE-TRP-2 particles combined with the agonist anti-glucocorticoid-induced tumor necrosis factor receptor (GITR) mAb showed complete regression in 90% of mice [82]. In another study, the SFV DNA replicon co-expressing the vascular growth factor receptor-2 (VEGFR-2) and interleukin-12 (IL-12) was co-administered with the SFV DNA replicon co-expressing survivin and the β-hCG antigen to a B16 mouse melanoma model [83]. Co-administration resulted in superior tumor growth inhibition and survival compared to treatment with either SFV-VEGFR-2/IL-12 DNA replicons or SFV-survivin-β-hCG Ag alone. Moreover, although VEE particles expressing the prostate-specific membrane antigen (PSMA) elicited strong immune responses in mice [84], only weak PSMA-specific immunogenicity was discovered in castration-resistant metastatic prostate cancer (CRPC) patients in phase I [85].
Oncolytic alphaviruses have proven to be an attractive alternative for cancer therapy, due to their specific killing of tumor cells. For example, the oncolytic M1 alphavirus effectively killed triple-negative breast cancer (TNBC) cells [86]. Superior eff8icacy was observed after co-administration of M1 and doxorubicin, significantly reducing mouse tumor growth [86]. In another study, intravenous administration of the oncolytic SIN AR339 strain to mice with implanted cervical tumors resulted in substantial tumor regression [87].
Table 2.
Examples of prophylactic/therapeutic interventions with alphavirus vectors for cancers.
| Cancer | Vector / Target gene(s) | Findings |
|---|---|---|
| NSCLC Colon Breast Breast Breast Cervix Cervix Cervix Colorectal Melanoma Melanoma Melanoma Prostate Prostate Breast Cervical |
SFV RP / EGFP SFV RNA / LacZ VEE RP / HER2 EDM/TM SIN DNA /HER2/neu SIN DNA / HER2/neu VEE RP / HPV E7 SFV RP / HPV E6-E7 SFV RP / HPV E6-E7 VEE RP / CEA VEE RP + CTLA-4 / TRP-2 VEE RP + GITR / TRP-2 SFV DNA / VEGFR-2/IL-12 + survivin/β-hGC VEE RP / PSMA VEE RP / PSMA Oncolytic M1 + doxorubicin Oncolytic SIN AR339 |
Killing of H358a cells, tumor regression in mice [73] Protection against CT26 tumor challenges in mice [74] Protection in mice, clinic benefits in phase I [75]. Protection in mice against A2L2 challenges [76] Protection with 80% less DNA compared to pDNA [77] Protection against tumor challenges in mice [78] Complete tumor eradication in mice [79] Good safety, immunogenicity in all patients in phase I [80] Safe, prolonged survival in phase I [81] Complete tumor regression in 50% of mice [82] Complete tumor regression in 90% of mice [82] Superior tumor growth inhibition and survival after co-administration of SFV DNA replicons in mice [83] Strong immune response in TRAMP mice [84] Weak immunogenicity in CRPC patients in phase I [85] Strong TNBC tumor regression in mice [86] Tumor regression in mice [87] |
CEA, carcinoembryonic antigen; CTLA-4, anti-CTL antigen-4 mAb; EGFP, enhanced green fluorescent protein; GITR, glucocorticoid-induced tumor necrosis factor mAb; HPV, human papillomavirus; NSCLC; non-small cell lung cancer; LNPs, lipid nanoparticles; pDNA, conventional plasmid DNA; RP, recombinant particles; SFV, Semliki Forest virus; SIN, Sindbis virus; taRNA, trans-amplifying RNA; TNBC, triple-negative breast cancer; TRAMP, transgenic adenocarcinoma of the mouse prostate; TRP-2, tyrosine-related protein-2; VEE, Venezuelan equine encephalitis virus; VEGFR-2, vascular endothelial growth factor receptor-2.
5.3. Neurological Disorders
Alphaviruses have demonstrated a strong neurotropism making them potential delivery vectors for neurological disorders. As the demand for long-term expression is necessary for the therapeutic efficacy of chronic diseases, the transient nature of alphavirus expression might not be ideal, but certainly, alphavirus vectors have demonstrated their feasibility in animal models for human diseases as described below and summarized in Table 3. For example. SFV particles expressing IL-10 have been evaluated in an experimental autoimmune encephalitis (EAE) model for multiple sclerosis (MS) [88]. Intranasal delivery of SFV-IL-10 particles resulted in therapeutic benefits in the EAE model in BALB/c mice. Moreover, SFV-based expression of the tissue inhibitor of metalloproteinase 2 (TIMP-2) inhibited EAE development in mouse CNS [89]. In another study, SFV particles expressing the transforming growth factor β1 (TGF-β1) showed significant inhibition of EAE in BALB/c mice [90]. In another approach, the peripheral administration of SFV particles and myelin basic protein (MBP) elicited superior immune responses in the splenocytes of EAE-susceptible SJL mice than in control mice [91].
Table 3.
Examples of alphavirus-based disease models for neurological disorders.
| Disease | Vector / Target gene(s) | Findings |
|---|---|---|
| EAE EAE EAE EAE |
SFV RP / IL-10 SFV RP / TIMP-2 SFV RP / TGF-β1 SFV RP + MBP |
Therapeutic benefits in EAE mouse model [88] Inhibition of EAE development in mice [89] Inhibition of EAE in BALB/c mice [90] Superior immune responses in EAE-susceptible mice [91] |
EAE, experimental autoimmune encephalitis; IL-10, interleukin-10; MBP, myelin basic protein; RP, recombinant particles; SFV, Semliki Forest virus; TGF-β1; transforming growth factor β1; TIMP-2, tissue inhibitor of metalloproteinase 2.
6. Conclusions
This review aims to describe the research on the lifecycle of alphaviruses as models for viruses in general and the cellular aspects of protein translation, post-translational modifications, and transport. Moreover, as several alphaviruses have been associated with epidemics in domestic animals and humans, their spread and pathogenicity have been the target of thorough investigations. Due to a breakthrough in molecular biology genetic engineering in the 1990s, applying alphavirus expression systems represented an attractive alternative for recombinant protein expression in vitro and in vivo. In this context, necessary attention was dedicated to reducing the cytopathogenic effects caused on host cells by alphaviruses. The application of attenuated and avirulent strains showed a substantial reduction in cytopathogenicity. However, extensive mutagenesis of the replicase genes provided extended survival of host cells and enhanced recombinant protein expression levels. Moreover, temperature-sensitive mutant vectors have shown feasibility in neuroscience allowing neuron-specific or glial-specific expression by a simple switch of the culture temperature for primary neurons or hippocampal slice cultures.
Alphaviruses have been subjected to prophylactic and therapeutic interventions in the areas of infectious diseases, cancers, and neurological disorders as summarized in Table 1, Table 2 and Table 3. One asset of alphaviruses is the flexibility to use recombinant viral particles, RNA replicons, and DNA replicon-based plasmids. Immunization with alphaviruses has provided protection against challenges with LASV, JUNV, MACV, DENV, ZIKV, IFVA, and SARS-CoV-2 in rodent models. Moreover, protection was also achieved against challenges with B. anthracis, B. abortus, and malaria in mice. Interestingly, in comparison to synthetic mRNA delivery, the administration of RNA replicons required 10-100-fold lower doses to obtain similar immune responses. Likewise, the doses used for DNA replicons can be 10-100-fold lower compared to conventional plasmid DNA administration. These findings are important as RNA and DNA replicons can provide better efficacy and potentially fewer adverse events than their corresponding counterparts. In the context of clinical trials, compared to other viral vectors the number of human studies with alphaviruses are much fewer, which to some extent relates to the more recent engineering and lesser use of alphavirus expression systems. However, the recent COVID-19 pandemic has given a boost for alphavirus-based systems, especially those applying LNP encapsulated RNA replicons. Although less advanced than vaccines based on synthetic mRNA, some encouraging results have been obtained in phase I and II trials resulting the granting EUA for the VEE RNA-LION vaccine in India.
In the context of cancer prevention and therapy, alphavirus vectors have elicited strong immune responses, tumor regression, and even total cure in rodent models. Moreover, clinical benefits have been obtained in clinical trials in cancer patients. Interestingly, administration of alphavirus vectors expressing HPV E6 and E7 proteins resulted in immune responses in all tested cervix cancer patients in phase I, which bodes well for future prophylactic applications.
The transient nature of alphavirus-based expression systems have made them suitable for acute diseases such as infectious diseases and cancers. In the case of both prevention and therapy short-term high-level expression is preferable, whereas in the case of chronic diseases it is advantageous to be able to supply long-term expression. However, in the context of neurological disorders, alphaviruses have proven useful for demonstrating the proof-of-concept in mouse EAE models for MS.
Overall, the success with using alphaviruses for prophylactic and therapeutic interventions has encouraged further investment in these delivery systems. Despite some setbacks in re producing findings from preclinical studies in humans, further vector engineering and dose optimization need to be addressed to further expand the application range of alphaviruses in the future.
Funding
This research received no external funding. ding.
Conflicts of Interest
The author declares no conflicts of interest.
References
- Strauss, J.H.; Strauss, E.G. The alphaviruses: Gene expression, replication and evolution. Microbiol. Rev. 1994, 58, 491–562. [Google Scholar] [CrossRef]
- Garoff H, Wilschut J, Liljeström P, Wahlberg JM, Bron R, Suomalainen M, Smyth J, Salminen A, Barth BU, Zhao H, et al. Assembly and entry mechanisms of Semliki Forest virus. Arch Virol. Suppl. 1994, 9, 329-338. [CrossRef]
- Birdwell, C.R.; Strauss, E.G.; Strauss, J.H. Replication of Sindbis virus. III. An electron microscopic study of virus maturation using the surface replica technique. Virology 1973, 56, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Davis, N.L.; Powell, N.; Greenwald, G.F.; Willis, L.V.; Johnson, B.J.B.; Smith, J.; Johnston, R.E. 1991. Attenuating mutations in the E2 glycoprotein gene of Venezuelan equine encephalitis virus: construction of single and multiple mutations in a full-length cDNA clone. Virology 1991. 183, 20-31.
- Liljestrom, P.; Garoff, H. A new generation of animal cell expression vectors based on the Semliki Forest virus replicon. Biotechnology 1991, 9, 1356–1361. [Google Scholar] [CrossRef] [PubMed]
- Xiong, C.; Levis, R.; Shen, P.; Schlesinger, S.; Rice, C.M.; Huang, H.V. Sindbis virus: An efficient, broad host range vector for gene expression in animal cells. Science 1989, 243, 1188–1191. [Google Scholar] [CrossRef] [PubMed]
- Davis, N.L.; Willis, L.V.; Smith, J.F.; Johnston, R.F. In vitro synthesis of infectious Venezuelan equine encephalitis virus RNA from a cDNA clone: Analysis of a viable deletion mutant. Virology 1989, 171, 189–204. [Google Scholar] [CrossRef]
- Atkins, G.J.; Sheahan, B.J.; Liljestrom, P. The molecular pathogenesis of Semliki Forest virus: a model virus made useful? J. Gen. Virol. 1999, 80, 2287–2297. [Google Scholar] [CrossRef]
- Brown, D.T.; Condreay, L.D. Replication of alphaviruses in mosquito cells. In The Togaviridae and Flaviviridae. Schlesinger, S.; Schlesinger, M.J., Eds.; Plenum Publishing Corp., New York, 1986 pp. 171-203.
- Cheng, R.H.; Kuhn, R.J.; Olson, N.H.; Rossmann, M.G.; Choi, H.K.; Smith, T.J.; Baker, T.S. Nucleocapsid and glycoprotein organization in an enveloped virus. Cell 1995, 80, 621–630. [Google Scholar] [CrossRef]
- Birdwell, C.R.; Strauss, J.H. Distribution of the receptor sites for Sindbis virus on the surface of chicken and BHK cells. J. Virol. 1974, 14, 672–678. [Google Scholar] [CrossRef]
- Fan, D.; Sefton, B.M. The entry into host cells of Sindbis virus, vesicular stomatitis virus and Sendai virus. Cell 1978, 15, 985–992. [Google Scholar] [CrossRef]
- Froshauer, S.; Kartenbeck, J.; Helenius, A. Alphavirus RNA replicase is located on the cytoplasmic surface of endosomes and lysosomes. J. Cell Biol. 1988, 107, 2075–2086. [Google Scholar] [CrossRef]
- Doms, R.W.; Lamb, R.A.; Rose, J.K.; Helenius, A. Folding and assembly of viral membrane glycoproteins. Virology 1993, 193, 545–562. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Garoff, H. Role of cell surface spikes in alphavirus budding. J. Virol. 1992, 66, 7089–7095. [Google Scholar] [CrossRef]
- Smith, A.L.; Tignor, G.H. Host cell receptors for two strains of Sindbis virus. Arch. Virol. 1980, 66, 11–26. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.-S.; Kuhn, R.J.; Strauss, E.G.; Ou, S.; Strauss, J.H. 1992. High-affinity laminin receptor is a receptor for Sindbis virus in mammalian cells. J. Virol. 1992, 66, 4992–5001. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W., Wang, L.; Yang, Y.; Jia, J.; Fu, S.; Feng, Y.; He, Y.; Li, J.P.; Liang, G. Interaction of E2 glycoprotein with heparan sulfate is crucial for cellular infection of Sindbis virus. PLoS One 2010, 5, e9656. [CrossRef]
- Hoekstra, D.; Kok, J.W. Entry mechanisms of enveloped viruses. Implications for fusion of intracellular membranes. Biosci. Rep. 1989, 9, 273–305. [Google Scholar] [CrossRef]
- Marsh, M.; Bolzau, E.; Helenius, A. Penetration of Semliki Forest virus from acidic prelysosomal vacuoles, Cell 1983, 32, 931-940. [CrossRef]
- Kääriäinen, L.; Keränen, S.; Lachmi, B.; Söderlund, H.; Tuomi, K.; Ulmanen, I. Replication of Semliki Forest virus. Med. Biol. 1975, 53, 342–351. [Google Scholar]
- Marsh, M. 1984. The entry of enveloped viruses into cells by endocytosis. Biochem. J. 1984, 218, 1–10. [Google Scholar] [CrossRef]
- Stegmann, T.; R.; Doms, R.W.; Helenius, A. 1989. Protein mediated membrane fusion. Annu. Rev. Biophys. Biophys. Chem. 1989, 18, 187-211.
- Helenius, A.; Kielian, M.; Wellsteed, J.; Mellman, I.; Rudnick, G. Effects of monovalent cations on Semliki Forest virus entry into BHK-21 cells. J. Biol. Chem. 1985, 260, 5691–5697. [Google Scholar] [CrossRef]
- Froshauer, S.; Kartenbeck, J.; Helenius, A. 1988. Alphavirus RNA replicase is located on the cytoplasmic surface of endosomes and lysosomes. J. Cell Biol. 1998, 107, 2075–2086. [Google Scholar] [CrossRef]
- Pietilä, M.K.; Hellström, K.; Ahola, T. Alphavirus polymerase and RNA replication. Virus Res. 2017, 234, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Garoff, H.; Simons, K.; Dobberstein. B. 1978. Assembly of the Semliki Forest virus membrane glycoproteins in the membrane of the endoplasmic reticulum in vitro. J. Mol. Biol. 1978, 124, 587-600.
- de Curtis, I.; Simons, K. 1988. Dissection of Semliki Forest virus glycoprotein delivery from the trans-Golgi network to the cell surface in permeabilized BHK cells. Proc. Natl. Acad. Sci. USA 1988, 85, 8052–8056. [Google Scholar] [CrossRef] [PubMed]
- Cadd, T.L.; Skoging, U.; Liljeström, P. Budding of enveloped viruses from the plasma membrane. Bioessays 1997, 19, 993–1000. [Google Scholar] [CrossRef]
- Weaver, S.; Salas, R.; Rico-Hesse, R.; Ludwig, G.V.; Oberste, M.S.; Boshell, J.; Tesh, J.B. Re-emergence of epidemic Venezuelan equine encephalomyelitis in South America. VEE Study Group. Lancet 1996, 348, 43640. [Google Scholar] [CrossRef]
- Mathiot C, Grimaud G, Garry P.; Bouquety, J.C.; Mada, A.; Daguisy, A.M.; Georges, A.J. An outbreak of human Semliki Forest virus infection in Central African Republic. Am. J. Trop. Med. Hyg. 1990, 42, 386-393.
- Niklasson, B. Sindbis and Sindbis-like viruses. In The arboviruses: epidemiology and ecology. Monath, T.P., Ed., CRC Press Inc., Boca Raton, 1988, pp.167-176.
- Niklasson, B.; Aspmark, A.; LeDuc, J.; Gargan, T.P.; Ennis, W.A.; Tesh, J.B.; Main, A.J. Jr. Association of a Sindbis-like virus with Ockelbo disease in Sweden. Am. J. Trop. Med. Hyg. 1984, 33, 1212–1217. [Google Scholar] [CrossRef]
- Kelvin, A. Outbreak of Chikungunya in the Republic of Congo and the global picture. J. Infect. Dev. Ctries 20ll, 5. 4414.
- Jansen, K. The 20A5-2N7 Chikungunya epidemic in Reunion: ambiguous etiologies, memories, and meaning-making. Med. Anthropol. 20l3, 32, fr4-89.
- Rodrigues Faria, N.; Lourenco, J.; Marques de Cerqueira, E.; et al. Epidemiology of Chikungunya virus in Bahia, Brazil, 2014-2015. PLoS Curr. 2016, 8, ecurrents.outbreaks.c97507e3e48efb946401755d468c28b2. [Google Scholar]
- Levine, B.; Huang, Q.; Isaacs, J.T.; Reed, J.C.; Griffin, D.E.; Hardwick, J.M. Conversion of lytic to persistent alphavirus infection by the bcl-2 cellular oncogene. Nature 1993, 361, 739–742. [Google Scholar] [CrossRef]
- Barry, G.; Fragkoudis, R.; Ferguson, M.C.; Lulla, A.; Merits, A.; Kohl, A.; Fazakerley, J.K.J. Semliki Forest virus-induced endoplasmic reticulum stress accelerates apoptotic death of mammalian cells. Virol. 2010, 84, 7369–7377. [Google Scholar] [CrossRef]
- Fros, J.J.; Pijlman, G.P. Alphavirus Infection: Host Cell Shut-Off and Inhibition of Antiviral Responses. Viruses 2016, 8, 166. [Google Scholar] [CrossRef]
- Frolova, E.; Frolov, I.; Schlesinger, S. Packaging signals in alphaviruses. J. Virol. 1997, 71, 248–258. [Google Scholar] [CrossRef] [PubMed]
- Burgess, D.J. RNA stability: remember your driver. Nat. Rev. Genet. 2012, 13, 72. [Google Scholar] [CrossRef] [PubMed]
- Blakney, A.K.; McKay, P.F.; Yus, B.I.; Aldon, Y.; Shattock, R. J. Inside out: optimization of lipid nanoparticle formulations for exterior complexation and in vivo delivery of saRNA. Gene Ther. 2019; 26, 363–372. [CrossRef]
- DiCiommo, P.D.; Bremner, R. (1998). Rapid, high-level protein production using DNA based Semliki Forest virus vectors. J. Biol. Chem. 1998, 273, 18060–18066. [Google Scholar] [CrossRef]
- Lundstrom, K. Semliki Forest virus vectors for rapid and high-level expression of integral membrane protein. Biochim. Biophys. Acta 2003, 1610, 90–96. [Google Scholar] [CrossRef]
- Lundstrom, K.; Abenavoli, A.; Malgaroli, A.; Ehrengruber, M.U. Novel Semliki Forest virus vectors with reduced cytotoxicity and temperature sensitivity for long-term enhancement of transgene expression. Mol Ther. 2003, 7, 202–209. [Google Scholar] [CrossRef]
- Agapov, E.; Frolov, I.; Lindenbach, B.; Pragai, B.M.; Schlesinger, S.; Tice, C.M. Noncytopathic Sindbis virus RNA vector for heterologous gene expression. Proc. Natl. Acad. Sci. USA 1998, 95, 12989–12994. [Google Scholar] [CrossRef]
- Lundstrom, K.; Rotrnann, D.; Hermann, D. Novel mutant Semliki Forest virus vectors: gene expression and localization studies in neuronal cells. Histochem. Cell Biol. 2001, 115, 83–91. [Google Scholar] [CrossRef]
- Rodriguez-Madoz. J.; Prieto, J.; Smerdou, C. Semliki Forest virus vectors engineered to express higher IL-12 levels induce efficient elimination of murine colon adenocarcinomas. Mol. Ther. 2005, 12, 153-163.
- Beissert, T.; Perkovic, M.; Vogel, A.; Erbar, S.; Walzer, K.C.; Hempel, T.; Brill, S.; Haefner, E.; Becker, R.; Türeci, Ö.; et al. A Trans-amplifying RNA Vaccine Strategy for Induction of Potent Protective Immunity. Mol. Ther. 2020, 28, 119–128. [Google Scholar] [CrossRef]
- Lundstrom, K. Self-Amplifying RNA Virus Vectors for Drug Delivery. Exp. Opin. Drug Deliv. 2024, in press. in press.
- Wang, M.; Jokinen, J.; Tretyakova, I.; Pushko, P.; Lukashevich, I.S. Alphavirus-based replicon particles expressing multivalent cross-protective Lassa virus glycoproteins. Vaccine 2018, 36, 683–690. [Google Scholar] [CrossRef]
- Johnson, D.M.; Jokinen, J.D.; Wang, M.; Pfeffer, T.; Tretaykova, I.; Carrion, R. Jr.; Griffiths, A.; Pushko, P.; Lukashevich, I.S. Bivalent Junin and Machupo experimental vaccine based on alphavirus RNA replicon vector. Vaccine 2020, 38, 2949–2959. [Google Scholar] [CrossRef]
- Tretyakova, I.; Tibbens, A.; Jokinen, J.D.; Johnson, D.M.; Lukashevich, I.S.; Pushko, P. Novel DNA-launched Venezuelan equine encephalitis virus vaccine with rearranged genome. Vaccine 2019, 37, 3317–3325. [Google Scholar] [CrossRef] [PubMed]
- Tretyakova, I.; Plante, K.S.; Rossi, S.L.; Lawrence, W.S.; Peel, J.E.; Gudjohnsen, S.; Wang, E.; Mirchandani, D.; Tibbens, A.; Lamichhane, T.N.; et al. Venezuelan equine encephalitis vaccine with rearranged genome resists reversion and protects non-human primates from viremia after aerosol challenge. Vaccine 2020, 38, 3378–3386. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Ewing, D.; Subramanian, H.; Block, K.; Rayner, J.; Alterson, K.D.; Sedegah, M.; Hayes, C.; Porter, K.; Raviprakash, K. A heterologous DNA prime-Venezuelan equine encephalitis virus replicon particle boost dengue vaccine regimen affords complete protection from virus challenge in cynomolgus macaques. J. Virol. 2007, 81, 11634–11639. [Google Scholar] [CrossRef]
- Erasmus, J.H.; Khandhar, A.P.; Guderian, J.; Granger, B.; Archer, J.; Archer, M.; Gage, E.; Fuerte-Stone, J.; Larson, E.; Lin, S.; et al. A nanostructured lipid carrier for delivery of a replicating viral RNA provides single, low-dose protection against Zika. Mol. Ther. 2018, 26, 2507–2522. [Google Scholar] [CrossRef]
- Morris-Downes, M.M.; Sheahan, B.J.; Fleeton, M.N.; Liljeström, P.; Reid, H.W.; Atkins, G.J. A recombinant Semliki Forest virus particle vaccine encoding the prME and NS1 proteins of louping ill virus is effective in a sheep challenge model. Vaccine 2001, 19, 3877–3884. [Google Scholar] [CrossRef]
- Brand, D.; Lemiale, F.; Turbica, I.; Buzelay, L.; Brunet, S.; Barin, F. Comparative analysis of humoral immune responses to HIV type 1 envelope glycoproteins in mice immunized with a DNA vaccine, recombinant Semliki Forest virus RNA, or recombinant Semliki Forest virus particles. AIDS Res. Hum. Retrovir. 1998, 14, 1369–1377. [Google Scholar] [CrossRef]
- Ljungberg, K.; Whitmore, A.C.; Fluet, M.E.; Moran, T.P.; Shabman, R.S.; Collier, M.L.; Kraus, A.A.; Thompson, J.M.; Montefiori, D.C.; Beard, C.; et al. Increased immunogenicity of a DNA-launched Venezuelan equine encephalitis virus-based replicon DNA vaccine. Virology 2007, 81, 13412–13423. [Google Scholar] [CrossRef]
- Wecker, M.; Gilbert, P.; Russell, N.; Hural, J.; Allen, M.; Pensiero, M.; Chulay, J.; Chiu, Y.-L.; Abdool Karim, S.S.; Burke, D.S.; et al. Phase I safety and immunogenicity evaluations of an alphavirus replicon HIV-1 subtype C gag vaccine in healthy HIV-1-uninfected adults. Clin. Vaccine Immunol. 2020, 19, 1651–1660. [Google Scholar] [CrossRef]
- Fleeton, M.N.; Chen, M.; Berglund, P.; Rhodes, G.; Parker, S.E.; Murphy, M.; Atkins, G.J.; Liljeström, P. Self-replicative RNA vaccines elicit protection against influenza A virus, respiratory syncytial virus, and a tickborne encephalitis virus. J. Infect. Dis. 2001, 183, 1395–1398. [Google Scholar] [CrossRef]
- Vogel, A.B.; Lambert, L.; Kinnear, E.; Busse, D.; Erbar, S.; Reuter, K.C.; Wicke, L.; Perkovic, M.; Beisset, T.; Haas, H.; et al. Self-amplifying RNA vaccines give equivalent protection against influenza to mRNA vaccines but at much lower doses. Mol. Ther. 2018, 26, 446–455. [Google Scholar] [CrossRef]
- McKay, P.F.; Hu, K.; Blakney, A.K.; Samnuan, K.; Brown, J.C.; Penn, R.; Zhou, J.; Bouton, C.R.; Rogers, P.; Polra, K.; et al. Self-amplifying RNA SARS-CoV-2 lipid nanoparticle vaccine candidate induces high neutralizing antibodies in mice. Nat. Commun. 2020, 11, 3523. [Google Scholar] [CrossRef] [PubMed]
- Pollock, K.M.; Cheeseman, H.M.; Szubert, A.J.; Libri, V.; Boffito, M.; Owen, D.; Bern, H.; O'Hara, J.; McFarlane, L.R.; Lemm, N.M.; et al. Safety and Immunogenicity of a Self-Amplifying RNA Vaccine Against COVID-19: COVAC1, a Phase I, Dose-Ranging Trial. EClinicalMedicine 2022, 44, 101262. [Google Scholar] [CrossRef] [PubMed]
- Szubert, A.J.; Pollock, K.M.; Cheeseman, H.M.; Alagaratnam, J.; Bern, H.; Bird, O.; Boffito, M.; Byrne, R.; Cole, T.; Cosgrove, C.A.; et al. COVAC1 phase 2a expanded safety and immunogenicity study of a self-amplifying RNA vaccine against SARS-CoV-2. EClinicalMedicine 2023, 56, 101823. [Google Scholar] [CrossRef]
- Erasmus, J.H.; Khandhar, A.P.; Walls, A.C.; Hemann, E.A.; O'Connor, M.A.; Murapa, P.; Archer, J.; Leventhal, S.; Fuller, J.; Lewis, T.; et al. Single-dose replicating RNA vaccine induces neutralizing antibodies against SARS-CoV-2 in nonhuman primates. bioRxiv 2020, 2020.05.28.121640. [CrossRef]
- Safety, Tolerability and Immunogenicity of the candidate GEMCOVAC-19 (COVID-19) vaccine in healthy pediatric subjects of 5 to less than 18 years. www.ctri.nic.in/Clinicaltrials/pmaindet2.php?trialid=66638.
- HDT Bio receives Emergency Use Approval for COVID-19 vaccine in India. www.pharmaceutical-technology.com/news/hdt-bio-vaccine-india-approval/.
- Thomas, J.M.; Moen, S.T.; Gnade, B.T.; Vargas-Inchaustegui, D.A.; Foltz, S.M.; Suarez, G.; Heidner, H.W.; König, R.; Chopra, A.K.; Peterson, J.W. Recombinant Sindbis virus vectors designed to express protective antigen of Bacillus anthracis protect animals from anthrax and display synergy with ciprofloxacin. Clin. Vaccine Immunol. 2009, 16, 1696–1699. [Google Scholar] [CrossRef]
- Wang, H.C.; An, H.J.; Yu, Y.Z.; Xu, Q. Potentiation of anthrax vaccines using protective antigen-expressing viral replicon vectors. Immunol. Lett. 2015, 163, 206–213. [Google Scholar] [CrossRef]
- Cabrera, A.; Saez, D.; Cespedes, S.; Andrews, E.; Onate, A. Vaccination with recombinant Semliki Forest virus particles expressing translation initiation factor 3 of Brucella abortus induces protective immunity in BALB/c mice. Immunobiology 2009, 214, 467–474. [Google Scholar] [CrossRef]
- Tsuji, M.; Bergmann, C.C.; Takita-Sonoda, Y.; Murata, K.; Rodrigues, E.G.; Nussenzweig, R.S.; Zavala, F. Recombinant Sindbis viruses expressing a cytotoxic T-lymphocyte epitope of a malaria parasite or of influenza virus elicit protection against the corresponding pathogen in mice. J. Virol. 1998, 72, 6907–6910. [Google Scholar] [CrossRef]
- Murphy, A.M.; Morris-Downes, M.M.; Sheahan, B.J.; Atkins, G.J. Inhibition of human lung carcinoma cell growth by apoptosis induction using Semliki Forest virus recombinant particles. Gene Ther. 2000, 7, 1477–1482. [Google Scholar] [CrossRef]
- Ying, H.; Zaks, T.Z.; Wang, R.-F.; Irvine, K.R.; Kammula, U.S.; Marincola, F.M.; Leitner, W.W.; Restifo, NP. Cancer therapy using a self-replicating RNA vaccine. Nat. Med. 1999, 5, 823–827. [Google Scholar] [CrossRef]
- Crosby, E.J.; Gwin, W.; Blackwell, K.; Marcom, P.K.; Chang, S.; MAecker, H.T.; Broadwater, G.; Hyslop, T.; Kim, S.; Rogatko, A.; et al. Vaccine-induced memory CD8+ T cells provide clinical benefit in HER2 expressing breast cancer: a mouse to human translational study. Clin. Cancer Res. 2019, 25, 2723–2736. [Google Scholar] [CrossRef]
- Lachman, L.B.; Rao, X.M.; Kremer, R.H.; Ozpolat, B.; Kiriakova, G.; Price, J.E. DNA vaccination against neu reduces breast cancer incidence and metastasis in mice. Cancer Gene Ther. 2001, 8, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, J.-P.; Maughan, M.F.; Lachman, L.B. Alphavirus replicon particles containing the gene for HER2/neu inhibit breast cancer growth and tumorigenesis. Breast Cancer Res. 2005, 7, R145–R155. [Google Scholar] [CrossRef] [PubMed]
- Velders, M.P.; McElhiney, S.; Cassetti, M.C.; Eiben, G.L.; Higgins, T.; Kovacs, G.R.; Elmishad, A.G.; Kast, W.M.; Smith, L.R. Eradication of established tumors by vaccination with Venezuelan equine encephalitis virus replicon particles delivering human papillomavirus 16 E7 RNA. Cancer Res. 2001, 61, 7861–7867. [Google Scholar] [PubMed]
- Daemen, T.; Riezebos-Brilman, A.; Bungener, L.; Regts, J.; Dontje, B.; Wilschut, J. Eradication of established HPV16-transformed tumours after immunisation with recombinant Semliki Forest virus expressing a fusion protein of E6 and E7. Vaccine 2003, 21, 1082–1088. [Google Scholar] [CrossRef]
- Komdeur, F.L.; Singh, A.; Van de Wall, S.; Meulenberg, J.J.M.; Boerma, A.; Hoogeboom, B.N.; Paijens, S.T.; Ovarce, C.; de Bruyn, M.; Schuuring, E.; et al. First-in-human phase I clinical trial of an SFV-based RNA replicon cancer vaccine against HPV-induced cancers. Mol. Ther. 2021, 29, 611–625. [Google Scholar] [CrossRef]
- Crosby, E.J.; Hobeika, A.C.; Niedzweicki, D.; Rushing, C.; Hsu, D.; Berglund, P.; Smith, J.; Osada, T.; Gwin lii, W.R.; Hartman, Z.C.; et al. Long-term survival of patients with stage III colon cancer treated with VRP-CEA(6D), an alphavirus vector that increases the CD8+ effector memory T cell to Treg ratio. J. Immunother. Cancer 2020, 8, e001662. [Google Scholar] [CrossRef]
- Avogadri, F.; Zappasodi, R.; Yang, A.; Budhu, S.; Malandro, N.; Hirschhorn-Cymerman, D.; Tiwari, S.; Maughan, M.F.; Olmsted, R.; Wolchok, J.D.; et al. Combination of alphavirus replicon particle-based vaccination with immunomodulatory antibodies: Therapeutic activity in the B16 melanoma mouse model and immune correlates. Cancer Immunol. Res. 2014, 2, 448–458. [Google Scholar] [CrossRef]
- Yin, X.; Wang, W.; Zhu, X.; Wang, Y.; Wu, S.; Wang, Z.; Wang, L.; Du, Z.; Gao, J.; Yu, J. Synergistic antitumor efficacy of combined DNA vaccines targeting tumor cells and angiogenesis. Biochem. Biophys. Res. Comm. 2015, 465, 239–244. [Google Scholar] [CrossRef]
- Durso, R.J.; Andjelic, S.; Gardner, J.P.; Margitich, D.J.; Donovan, G.P.; Arrigale, R.R.; Wang, X.; Maughan, M.F.; Talarico, T.L.; Olmsted, R.A.; et al. A novel alphavirus vaccine encoding prostate-specific membrane antigen elicits potent cellular and humoral immune responses. Clin. Cancer Res. 2007, 13, 3999–4008. [Google Scholar] [CrossRef]
- Slovin, S.F.; Kehoe, M.; Durso, R.; Fernandez, C.; Olson, W.; Gao, J.P.; Israel, R.; Scher, H.I.; Morris, S. A phase I dose escalation trial of vaccine replicon particles (VRP) expressing prostate-specific membrane antigen (PSMA) in subjects with prostate cancer. Vaccine 2013, 31, 943–949. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, Y.; Tan, J.; Zhang, Y.; Wong, C.-W.; Lin, Z.; Liu, X.; Sander, M.; Yang, X.; Liang, L.; et al. Necroptotic virotherapy of oncolytic alphavirus M1 cooperated with Doxorubicin displays promising therapeutic efficacy in TNBC. Oncogene 2021, 40, 4783–4795. [Google Scholar] [CrossRef] [PubMed]
- Unno, Y.; Shino, Y.; Kondo, F.; Igarashi, N.; Wang, G.; Shimura, R.; Yamaguchi, T.; Asano, T.; Saisho, H.; Sekiya, S.; et al. Oncolytic virotherapy for cervical and ovarian cancer cells by Sindbis virus strain AR339. Clin. Cancer Res. 2005, 11, 4553–4560. [Google Scholar] [CrossRef] [PubMed]
- Jerusalmi, A.; Morris-Downes, M.M.; Sheahan, B.J.; Atkins, G.J. Effect of intranasal administration of Semliki Forest virus recombinant particles expressing reporter and cytokine genes on the progression of experimental autoimmune encephalomyelitis. Mol. Ther. 2003, 8, 886–894. [Google Scholar] [CrossRef]
- Nygårdas, P.T.; Grönberg, S.A.; Heikkilä, J.; Joronen, K.; Sorsa, T.; Hinkkanen, A. Treatment of experimental autoimmune encephalomyelitis with a neurotropic alphavirus vector expressing tissue inhibitor of metalloproteinase-2. Scand. J. Immunol. 2004, 60, 372–381. [Google Scholar] [CrossRef]
- Vähä-Koskela, M.J.; Kuusinen, T.I.; Holmlund-Hampf, J.C.; Furu, P.T.; Heikkilä, J.E.; Hinkkanen, A.E. Semliki Forest virus vectors expressing transforming growth factor beta inhibit experimental autoimmune encephalomyelitis in Balb/c mice. Biochem. Biophys. Res. Commun. 2007, 355, 776–781. [Google Scholar] [CrossRef]
- Mokhtarian, F.; Shi, Y.; Zhu, P.F.; Grob, D. Immune responses, and autoimmune outcome, during virus infection of the central nervous system. Cell Immunol. 1994, 157, 195–210. [Google Scholar] [CrossRef]
Figure 1.
Schematic illustration of the life cycle of alphaviruses. Alphavirus particles deliver single-stranded RNA where the ssRNA+ strand is transcribed and amplified. Among structural alphavirus proteins, the capsid (C) protein forms nucleocapsids (NCs) with the RNA genome and the envelope (E) proteins are transported through the endoplasmic reticulum and Golgi for assembly with the NCs for release by budding of mature alphavirus particles.
Figure 1.
Schematic illustration of the life cycle of alphaviruses. Alphavirus particles deliver single-stranded RNA where the ssRNA+ strand is transcribed and amplified. Among structural alphavirus proteins, the capsid (C) protein forms nucleocapsids (NCs) with the RNA genome and the envelope (E) proteins are transported through the endoplasmic reticulum and Golgi for assembly with the NCs for release by budding of mature alphavirus particles.

Figure 2.
Schematic illustration of alphavirus expression systems. A. Replication-deficient system: The alphavirus expression vector contains the non-structural protein (nsP1-4) replicase genes and the gene of interest (GoI). The helper vector carries the structural protein genes (capsid (c), p62, precursor of the E2 and E3 envelope protein, and E1 envelope protein). B. Replication-proficient systems: The GoI can be inserted downstream of the nsP1-4 genes or alternatively downstream of the structural protein genes. C. DNA replicon-based systems: Replication-deficient and -proficient vectors used for direct DNA plasmid transfer have been engineered by replacing the SP6 RNA polymerase promoter with the cytomegalovirus (CMV) promoter.
Figure 2.
Schematic illustration of alphavirus expression systems. A. Replication-deficient system: The alphavirus expression vector contains the non-structural protein (nsP1-4) replicase genes and the gene of interest (GoI). The helper vector carries the structural protein genes (capsid (c), p62, precursor of the E2 and E3 envelope protein, and E1 envelope protein). B. Replication-proficient systems: The GoI can be inserted downstream of the nsP1-4 genes or alternatively downstream of the structural protein genes. C. DNA replicon-based systems: Replication-deficient and -proficient vectors used for direct DNA plasmid transfer have been engineered by replacing the SP6 RNA polymerase promoter with the cytomegalovirus (CMV) promoter.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



