Submitted:
01 August 2024
Posted:
04 August 2024
You are already at the latest version
Abstract
The connection between the mother and the child has been considered one of the strongest bonds in nature. Though there are numerous factors that can influence the establishment of pregnancy, in its essence three are considered major: a good quality embryo, a receptive endometrium and a successful cross-talk between them. The placenta, that derives from the trophoblast of the embryo, develops when a successful implantation occurs. It is an ephemeral organ through which the turnover of nutrient, gas and waste molecules is realized. It serves as barrier and can provide the embryo with immune factors. Placental disorders are observed in some rare but life-threatening obstetric conditions like: preeclampsia (PE), fetal growth restriction (FGR), gestational tropho-blastic diseases (GTD) and gestational diabetes mellitus (GDM). The etiology and pathogenesis of some are still partially enigmatic. Our attention in this review was driven by the participation of small RNA molecules: miRNAs and piRNAs, as potential epigenetic modulators of genes that play pivotal role in placental functioning. The molecular approach for understanding of placental disorders may help new diagnostic and therapeutic solutions to be found.
Keywords:
miRNA
; piRNA
; placenta
; preeclampsia
; endometrial receptivity
1. Introduction
The placenta, from Greek πλακόεντα, with the meaning of "flat cake" named after its round, flat appearance in humans, is a temporary organ that begins developing from the trophectoderm of the implanting blastocyst [4,7]. It plays crucial roles for the embryo as it facilitates nutrients, gas and waste exchange between the actually separate maternal and fetal circulations, and is an important endocrine organ, producing hormones that regulate both maternal and fetal physiology during pregnancy. The placenta is connected to the fetus via the umbilical cord, and on the opposite side to the maternal uterus. In humans, a layer of maternal endometrial tissue comes away with the placenta when it is expelled from the uterus after the birth of the child. Placentas are a defining characteristic of placental mammals, but can also be found in marsupials and some non-mammals [6]. The placenta has evolved in evolution independently multiple times, starting in fish [3] and appearing in some reptiles as well [19].
The mammalian placenta occurred more than 100 million years ago and although all mammalian placentas have the same physiological meaning, there are differences in structure and function in different groups of mammals [6]. They are very different at both gross and microscopic view as well as in their function, for example the ability to supply the fetus with maternal immunoglobulins [26]. The protein syncytin, found in the outer barrier of the placenta - the syncytiotrophoblast, has a certain RNA signature in its genome thus leading to the hypothesis that the placenta originated from an ancient retrovirus - a virus that probably facilitated the transition from egg-laying to live-birth [1,2,5].
2. Physiology of the Placenta
Just after the ovulation occurs, the released oocyte is gently taken over by the fallopian tube. There it rests in the ampullary part for 36-48 hours waiting to be fertilized by the coming from the low genital tract spermatozoa [7]. If conception occurs then the newly formed zygote is starting a long 5-6 days journey in the tube advancing towards the uterine cavity. It enters the cavity as a free floating blastocyst surrounded by zona pellucida, consisting of an inner cell mass with a trophectoderm hull. Rolling on the carpet of the endometrium it hatches from zona pellucida. Appositioning of the polar hatched blastocyst happens to a putative site, probably orchestrated by some endometrial molecules, a cross-talk occurs between the embryo and the receptive endometrium.The blastocyst adheres to the endometrium and differentiation starts of the trophoblast cells into cytotrophoblast, an inner mass and syncytiotrophoblast - the outer part. The syncytiotrophoblast invades the luminal epithelium and a full implantation follows [7].
Macroscopically, at the moment of attachment, the trophoblast grows and forms a large syncytial cytoplasmic mass - syncytotrophoblast (STB). STB forms sheaths that, by lysing the underlying tissues, penetrates deep into the endometrium and paves the way for the blastocyst to sink. STB overcomes first the uterine epithelium, then the underlying decidua (loose connective tissue) and finally destroys the walls of the capillaries. Thus, the blastocyst finds itself in direct contact with the mother's blood, which freely pours out of the ruptured vessels into the spaces formed around it called - lacunae. Part of the trophoblast cells displace the cells of the endoglandular epithelium [27]. Implantation ends with complete sinking of the blastocyst into the endometrium - the entrance site is closed by regeneration of connective tissue and growth of a new covering epithelium [8].
The first schematic description of implantation was made by Elizabeth Ramsey, who later depicted the changes in uterine arteries and veins during pregnancy [8]. Until recently, it was assumed that trophoblastic invasion in the process of human placentation was limited to entering the connective tissue of the uterus and the uterine spiral arteries. It was believed that the spiral arteries connect with the intervillous space of the placenta and thus ensure a supply of maternal blood to the fetus. In recent years, this concept has changed significantly.
The invasion of the endoglandular trophoblast into the endometrial glands from the start of implantation creates the first nutrition for the fetus - the histiotrophic through the so-called "uterine milk" [9,10,11,12,13]. This is followed by entry of the endovenous trophoblast into the uterine veins, which provides outflow (drainage) of fluids from the placenta back into the maternal circulation [10,14,15]. Follows the entry of the endolymphatic trophoblast into the lymphatic vessels of the uterus. Only then does endoarterial trophoblast invade the spiral arteries, providing hemotropic nutrition, beginning in the second trimester of pregnancy [13].
The polarization of endometrial epithelial cells in the mid- secretory phase is considered to be an important factor in the occurrence of implantation [16].As the endometrium remodels in mid-secretory phase, its secretory epithelial cells increase their secretion, which coincides in time with the loss of polarity of the lumenal epithelial cells. Cell polarity refers to the uneven distribution of intracellular components [16]. Polarity is a fundamental characteristic of all cells and is differently expressed depending on the function of each cell type. Polarity occurs as a result of the action of molecular determinants of polarity, inside the cell, and thus determines both spatial relationships between cells and relationships with the extracellular space [17]. Epithelial cells have apico-basolateral polarity and planar polarity [16] Thanks to them, a functional barrier is formed and the orientation of cells in the tissue, the structure of the tissue, intercellular adhesions, as well as the transmission of signals between cells are regulated. All this also applies to the endometrial epithelium, in which cell polarity defines a repulsive apical surface, regulates the movement of molecules around the cell, directs the transmission of signaling molecules between cells, and maintains the integrity of the cytoskeleton [16].
The endometrial lumenal epithelial cells are the first point of contact with the polarized embryonic trophectoderm, and they would repel each other if the endometrial epithelium continued to maintain its polarity, rendering the endometrium unreceptive to the blastocyst attempting to implant [7,18]. The endometrial lumenal epithelium loses its polarity during the mid-secretory phase of MCs (menstrual cycles) [18]. This makes the endometrium receptive. The loss of polarity is due to morphological and molecular changes occurring in the epithelial cells. Changes occur in microvilli, cell-membrane surface markers, intercellular junctions, and cytoskeletal molecules. To provide opportunities for invasion, the lumenal epithelium weakens the lateral connections between cells by reducing surface epithelial adhesion proteins as well as focal adhesion to the basal membrane [18]. All these changes can be united under the concept of plasma membrane transformation (PMT) [16], an important event for implantation to occur. Abnormal endometrial receptivity is one of the main causes of embryo implantation failures and infertility. Any disturbance in the PMT affects endometrial receptivity and implantation. Parallels have been repeatedly drawn between PMT and epithelial-mesenchymal transformation (EMT) in tumorigenesis [16]. Plasma membrane transformation (PMT) and epithelial-mesenchymal transformation (EMT) are similar in the remodeling of the actin cytoskeleton in the attachment zone, integrin expression, and epithelial-stromal communication. Of course there are also many differences that are still being studied [16]
2.1. Maternal Placental Circulation and Immunity
In the process of implantation of the blastocyst, the endometrium undergoes decidualization. Spiral arteries become less convoluted, with increased diameter. This all increases maternal blood flow to the placenta [28] and the fetal villi are bathed by the maternal blood that pours into the intervillous space thus allowing exchange of gases, nutrients and waste molecules to happen . The maternal blood is into direct contact with the fetal chorion, without actual fluid exchange. With decreases of the pressure between pulses, the deoxygenated blood flows back through the endometrial veins [28].
Deoxygenated fetal blood from the fetal circulation passes through umbilical arteries to the placenta. As they enter the placenta, the umbilical arteries branch radially to form chorionic arteries. They, in turn, branch into cotyledon arteries. In the villi, the smallest vessels form an arterio-capillary-venous loops, that bring the fetal blood extremely close to the maternal blood without intermingling of fetal and maternal blood, thus forming the placental barrier [29]. The placenta plays the role of a selective barrier between maternal and fetal cells, preventing maternal blood cells, proteins and microbes (including bacteria and most viruses) from crossing the maternal-fetal barrier [30]. Deterioration in placental functioning, may be related to mother-to-child transmission of some infectious diseases [30]. A very small number of viruses including rubella virus, Zika virus and cytomegalovirus (CMV) can travel across the placental barrier, generally taking advantage of conditions at certain gestational periods as the placenta develops. CMV and Zika travel from the maternal bloodstream via placental cells to the fetal bloodstream [32].
IgG antibodies can pass through the human placenta after 13 weeks of gestation, with the largest transfer occurring in the third trimester, thus providing intrauterine protection to the fetus [33]. This passive immunity serves several months after birth, providing the newborn with the mother's long-term humoral immunity to protect the infant through the crucial first months of extrauterine life. IgM antibodies, because of their larger size, cannot cross the placenta [34]. For this reason infections acquired during pregnancy can be particularly hazardous for the fetus [34].
2.2. Hormonal Secretion
As an endocrine organ the placenta produces : hCG (human chorionic gonadotropin), progesterone, estrogen and hPL (human placental lactogene). The first hormone released by the placenta is the human chorionic gonadotropin (hCG). It stops the cyclicity of menstruation appearance after the corpus luteum ceases activity and atrophies. Its low secretion leads to spontaneous abortion. hCG stimulates the production of progesterone and estrogen by the corpus luteum [35]. hCG also helps male fetus by stimulating the testes to produce testosterone, necessary for the sex organs of the male to grow [35]. Progesterone, like hCG, is necessary to prevent spontaneous abortion because it prevents contractions of the uterus and is important for implantation [35]. Estrogen assists the enlargement of the uterus and breasts, assisting for growth of the fetus and production of milk. Estrogen is also responsible for increased blood supply in pregnancy through vasodilation [35]. Human placental lactogen (hPL) in pregnancy is important for fetal metabolism, general growth and development. Together with growth hormone, stimulates insulin-like growth factor production and regulates metabolism [35,36]. hPL modulates embryonic development and metabolism, and stimulates production of IGF, insulin, surfactant and adrenocortical hormones in the fetus.
2.3. Placental Barrier
The placenta and fetus may be accepted as a foreign body inside the mother and must be protected from the normal immune response of the mother that could cause their rejection. The placenta and fetus are thus treated as immune privileged, with immune tolerance. For this the placenta secretes neurokinin B-containing phosphocholine molecules. This same molecules are used by parasitic nematodes to avoid detection by the immune system of their host [37]. Small lymphocytic suppressor cells from the fetus can inhibit maternal cytotoxic T cells by inhibiting the response to interleukin 2 [38]. However, foreign fetal cells also persist in the maternal circulation, on the other side of the placental barrier [40].
3. Pathophysiology of the Placenta
The impairment of normal placental development is observed in several pregnancy pathologies, i.e., preeclampsia (PE), fetal growth restriction (FGR), gestational trophoblastic diseases (GTD) and gestational diabetes mellitus (GDM). Besides, external factors, such as microbial agents, and chemicals compounds, these can affect placental development and function, impairing pregnancy outcomes. These pregnancy complications result in increased maternal and fetal mortality and morbidity, leading to life-long health complications in both mother and child. Maternal diabetes or obesity, can change the levels of nutrient transporters in the placenta potentially resulting in overgrowth or restricted growth of the fetus [31].
3.1. Molecular Approach to Placental Disorders
Assessment of impairment of normal placental development on molecular level is a current approach in research in obstetrics. MiRNAs are small molecules with immense impact on the regulation of morphology and functioning of all tissues in the body [43]. Aberrant expression of miRNAs is associated with recurrent spontaneous abortions, preeclampsia (PE), fetal growth restriction (FGR), gestational diabetes mellitus (GDM), preterm birth (PTB), small-for-dates (SFD) [43,44].
MiRNAs are short non-coding single-stranded RNAs that modulate the expression of genes on post-transcriptional level [45]. Today over two thousand human miRNAs have been described and each of them can target hundreds of genes. Their number is increased adding their isoforms which can act cooperatively with the canonical form or have a quite different function [46,52]. Data have been gathered for the expression of miRNAs in human placenta in different gestational ages. The identified miRNAs are thought to be key regulators of placental development and maintenance of human pregnancy [43].
3.2. Biogenesis and Biological Function of miRNAs
Since their discovery in the early 1990s, large numbers of miRNAs have been sequenced and there are around 3000 known miRNAs [46]. MicroRNAs are encoded mainly by their own genes - MIR genes, as well as other types of genomic loci. MIR genes are transcribed as long primary transcripts (pre-microRNAs) by RNA polymerase II [51]. Pri-microRNA contains a partially self-complementary region that folds over to form a hairpin structure (fig. 1) in which a single-stranded loop and a double-stranded (ds) stem are found, terminating in a single-stranded 5' end and 3' -end. Pri-microRNA undergoes multistep processing to finally form mature microRNA (fig. 1). The hairpin structure is capped with 7-methylguanosine and is polyadenylated [50]. In the nucleus, pri-microRNA, which is approximately 65-80 nt in length, undergoes endonuclease cleavage, which is carried out by a microprocessor - a protein complex including the enzyme DROSHA (ribonuclease III) and the dsRNA-binding protein DGCR8 (DiGeorge syndrome critical region 8) [108,109,110]. DROSHA cleaves the double-stranded stem 11 nt from the junction with the single-stranded ends. The precision of this cutting is thought to be due to DGCR8, which serves as a kind of molecular meter, recognizing the pri-microRNA substrate and preparing the way for DROSHA [111]. The resulting double-stranded precursor (pre-microRNA) is transported across the nuclear membrane to the cytoplasm by the RanGTPase-dependent protein Exportin-5(fig.1). In the cytoplasm, pre-microRNA is further cleaved by the endoribonuclease enzyme DICER, assisted by the transactivation response element RNA-binding protein (TPBP) [111,112,113,114]. DICER (ribonuclease III) is an enzyme with multiple domains, including an amino-terminal PAZ helicase domain (Piwi/Argonaute/Zwille), and two RNase domains [115]. DICER recognizes and captures the free ends of the pre-microRNA and places it in a catalytic enzyme pocket. At this stage, the PAZ domain is important to measure the double-stranded stem from the 3′-end to produce a short dsRNA (microRNA duplex) of species-specific length and a typical two-nucleotide overhang at the 3′-ends [116] After cleavage of the pre-microRNA, a microRNA duplex of 20-24 nt in length is obtained, with each 3'-end 2 nt longer than the corresponding 5'-end (overhanging 3'-end). It is loaded onto the protein Argonaute2 (AGO2), which is a major component of an effector protein complex called miRISC (miRNA-induced silencing complex) [117,118] At this stage the microRNA duplex unfolds, the strand with lower internal stability at the 5'-end is retained in the complex (mature microRNA) and the other strand is discarded and degraded [119,120]. This canonical pathway occurs in most mammals, but alternative biosynthetic pathways have also been described.
Figure 1.
Model of canonical microRNA biogenesis, Created with BioRender.
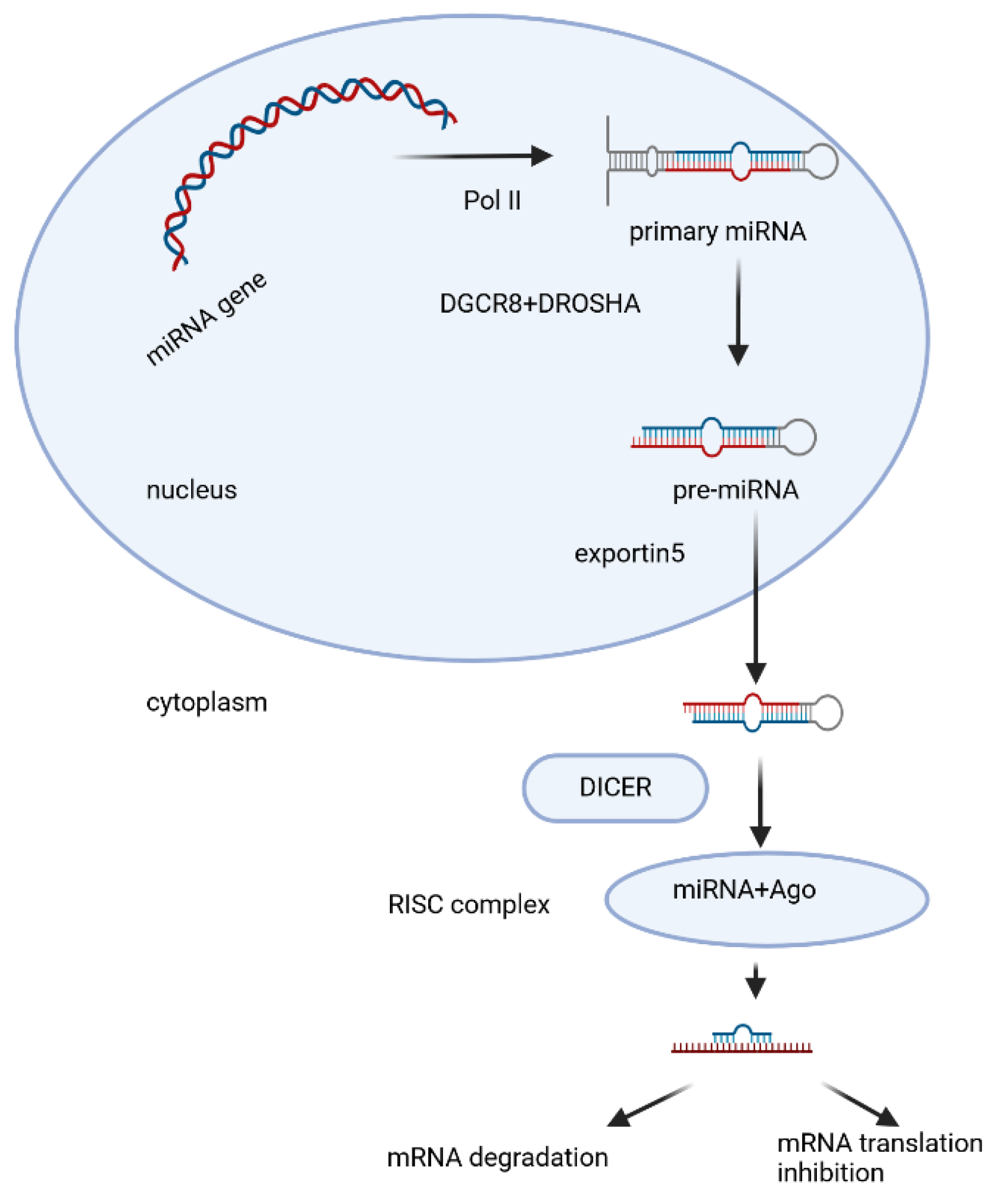
Changes in sequence or structure influences the maturation and turnover of miRNAs [46]. The mature miRNAs regulate gene expression on post-transcriptional level by translation suppression of target mRNAs resulting in their silencing or degradation and further in down regulation of target genes. MiRNA-binding sites are predominantly located in the 3′UTR of messenger RNAs (mRNAs), and the region at the 5′-end of miRNAs, from 2 to 7nt (nucleotide) is vital for their target recognition and has been termed the “seed sequence” [43,46,50]. Mature miRNAs mainly bind complementary with the “seed sequence” to target sites located at the 3′UTR of correspondingly targeted mRNA [46]. The nucleotide position of the seed sequence is important for proper target recognition. Imperfect binding of miRNA to their partially complementary sequences located at the 3′UTRs of the targeting mRNAs causes repression of subsequent protein translation; however, near-perfect binding of those miRNAs to their complementary sites within 3′UTR results in cleavage and further degradation of the targeting mRNAs [46]. Some miRNAs also bind to targeting sites at the 5′-untranslated region (5′-UTR). Recently, it has also been indicated that miRNAs negatively regulate gene expression in proliferating cells but up regulate gene expression in quiescent cells [50]. The biogenesis and function of miRNAs are precisely regulated and the dysregulation of miRNAs is associated with human diseases [43].
3.3. Expression of miRNAs in Human Placenta
762 miRNAs have been detected in human placenta [43]. It has been accepted that hypoxia, mainly resulting from impaired invasion of the trophoblast or some other oxidative stress (OS)-induced conditions, is a common risk factor that may deteriorate human placental development and function during pregnancy [53]. It is associated with increased incidence of gestational diseases. The expression of many miRNAs is controlled by hypoxia in human placenta [54]. For example, hypoxia up-regulates miRNA-210 (miR-210, well-known as a sensor of hypoxia) expression, which facilitates mitochondrial function in human trophoblast cells [55]. The expression of miR-141 is induced by hypoxia and the elevated expression of miR-141 affects trophoblast cell apoptosis, invasion and vascularization [56]. MiR-218 expression is enhanced by hypoxia to suppress trophoblast cell function [57]. In addition, other factors including environmental factors and epigenetic modification also exert effects on the expression of miRNAs during human pregnancy. Some chemical agents influence has been studied as well, like phthalates, mainly used as plastifiers, added to plastics to improve their flexibility, transparency and durability [106]. The maternal exposure to phthalates arouse oxidative stress and alterations in miRNAs expression in the placenta are observed, concerning: miR-17-5, miR155-5p, miR126-3p and miR16, that can finally trigger apoptosis of HTR8/SV neo cells (first trimester placental cell line) [107]. Exposure to phenol is associated with alterations of miRNA expression in human placenta, including miR-142-3p, miR15a-5p and miR-185 [58]. Similar phenomena can also be observed from metals and air toxics, which are closely associated with pregnancy-related complications [59].
Epigenetic regulation, involving heritable changes in expression of genes has been shown to participate in controlling the expression of human placental miRNAs. The DNA methylation has been proved to act as a key mediator in the expression of miRNAs [60]. The most popular example is the C19MC microRNA locus that is imprinted in human placenta [61]. Down-regulated C19MC due to methylation is associated with the increased maternal size, which in turn, gives rise to the larger body size of the child [62].
Recent studies have clarified that kinds of miRNAs exert effects on human trophoblast cell proliferation, apoptosis, migration and invasion, suggesting human placental development is controlled by various miRNAs. In an in vitro study using the first trimester placental explants, small interfering RNA (siRNA)-mediated knockdown of Dicer significantly promotes cytotrophoblast cell proliferation and also significantly induces the expression of two molecules in pro-mitogenic signaling within cytotrophoblasts, including ERK and SHP-2 [63], indicating the role of Dicer-dependent miRNAs in regulating human early placental development. Some other miRNAs have been found to affect trophoblast cell proliferation and apoptosis. MiR-376c elevates trophoblast cell proliferation by impairing transforming growth factor-β (TGF-β) [64], miR-210-3p regulates trophoblast cell proliferation by fibroblast growth factor1 (FGF1) [65], miR-133b boosts trophoblast cell proliferation by restricting SGK1 [66]. In contrast, miR-155 inhibits proliferation of human extravillous trophoblast cells via the down-regulation of cyclin D1 expression [67], miR-424 suppresses trophoblast cell proliferation through regulating ERRγ [68], miR-184 promotes apoptosis of human trophoblast cells by targeting WIG1 [69]. The movement (including migration and invasion) of EVTs (extravillous trophoblasts) towards the maternal decidua and myometrium are fundamental for effective placentation [71,72]. In EVTs (extravillous trophoblasts) of the human first trimester placenta, Dicer is observed to be abundantly expressed [70], suggesting miRNAs play a key role in trophoblast cell migration and invasion. A series of studies has uncovered a number of miRNAs that participate in regulating human trophoblast cell migration and invasion. MiR-34a targeting MYC inhibits human trophoblast cell invasion [73]; miR-27a suppresses trophoblast cell migration and invasion through mediating SMAD2 [74]; miR-320a negatively regulates trophoblast cell invasion by targeting estrogen-related receptor-gamma [75].
3.4. Micro-RNAs and Human Gestational Disorders
Aberrant expression levels of miRNAs have been both in vitro and in vivo detected in kinds of gestational complications, such as PE, FGR, GTD and GDM, and a number of studies have demonstrated the dysregulation of miRNA expression in compromised human pregnancies. We focus our attention on miRNAs’ role in preeclampsia as the data on the topic we found in literature was the most abundant and because it could be a life-threatening condition with high clinical importance for both mother and child.
3.5. Preeclampsia (PE) and miRNAs
Preeclampsia (PE) is a common disease of pregnancy occurring after 20 weeks of pregnancy, involving multiple organs and threatening the safety of the pregnant woman and the fetus; however, the etiology and pathogenesis of preeclampsia is still inconclusive. PE main symptoms are considered: maternal hypertension and proteinuria [76,77]. The incidence of preeclampsia is 2%–8% [102]. The clinical symptoms of most patients with preeclampsia are quickly relieved and improved after a pregnancy is terminated, indicating that the placenta plays a vital role in the pathogenesis of preeclampsia [103]. Recently many studies have been carried out to prove that miRNAs participate in the regulation of physiological function and pathological development of the placenta. The expression of miRNAs varies between the placentas from women with PE and normal placentas and the expression of a variety of miRNAs have been observed to be increased or decreased in placentas of PE [78,79]. Particularly, aberrant expression of those miRNAs in human trophoblast cells gives rise to their arrested proliferation and inadequate invasion, which further lead to the failure in sufficient remodeling of maternal spiral arteries and possibly consequent deficiency in angiogenesis, the so-called impaired invasion of the trophoblast [80,100]. The miRNAs with significantly up-regulated expression in PE placenta, including miR-17 [81], miR-155 [67], miR-431 [84], miR-30a-3p [85], miR-20a [86], miR-20b [87], miR-29b [88],miR-16 [89], miR-675 [90], miR-141 [91], miR-142-3p [92], miR-125b [93], miR-517a/b [94] and miR-517c [94], have been proven to negatively regulate trophoblast cell proliferation, migration, invasion and apoptosis. The effect of miRNAs on trophoblast cells is commonly dependent on the regulation of their corresponding target genes. For example, the upregulated expression of miR-17 is associated with PE, and miR-17 impedes trophoblast cell function by targeting EPHB4 and ephrin-B2, which have been shown to be critical for human early placental development [81]. On the other hand, miRNA-induced alterations in trophoblast cell function such as growth and mobility is mediated by low expression of related genes for example miR-125b (table1) inhibits migration and invasion of extra-villous trophoblastic cells by suppressing its target STAT3 expression, while STAT3 plays a key role in cell infiltration and vascular proliferation [82].

Table 1.
Expression levels (upregulated miRNAs) of miRNAs targeting trophoblast cell migration, proliferation, invasion, apoptosis and EV (extracellular vesicle) inclusion in PE modificated after Jin et al., 2022.
Table 1.
Expression levels (upregulated miRNAs) of miRNAs targeting trophoblast cell migration, proliferation, invasion, apoptosis and EV (extracellular vesicle) inclusion in PE modificated after Jin et al., 2022.
| Up-Regulated miRNAs | Affected Biological Process |
|---|---|
| miR-155 [128], miR-20a [129], miR-16 [130], miR-142-3p [131], miR-200p-3b [132], miR-137 [133], miR-146a [134] |
proliferation↓ |
| miR-155 [128], miR-431 [135], miR-20b [136], miR-16 [130], miR-125b [139], miR-200p-3b [129], miR-137 [133], miR-146a [140] |
migration↓ |
| miR-431 [135], miR-30a-3p [136], miR-20a [129], miR-20b [136], miR-141 [139], miR-142-3p [131], miR-125b [137], miR-146a [140], miR-517a/b [142], miR-517c [142] |
invasion↓ |
| miR-30a-3p [117], miR-29b [134], miR-16 [135], miR-200p-3b [140] | apoptosis↑ |
| miR-17-92 clusters [143], miR-106a0363 clusters [143] | differntiation↓ |
| 17 [144], miR-206 [145], miR-210 [146], miR-202-3p [147], miR-155p [149] |
other/targets↓ |
| miR-222-3p [149], miR-486-1-5p [150], miR-486-2-5p [150], miR-155 [151], miR-136 [152], miR-494 [152], miR-495 [152] |
in exosomes |
MiR-3935 expression that is significantly decreased in preeclamptic placental tissues, promotes human trophoblast cell EMT (epithelial-mesenchymal transition) through regulating TRAF6-RGS2 signaling [83](table2). MiRNAs are selectively packed into placenta-derived exosomes (a kind of extracellular vesicles) and miRNA species are differentially expressed in exosomes from women with PE and those from controls with normal pregnancy [95,96], indicating that placental exosomes may serve as mediators in the pathogenesis of PE and as early biomarkers of PE diagnosis.
Probably the placenta-derived exosomes participate in communication between maternal and fetal cells and thereby is hypothetised that those exosomal microRNAs act as mediators in cellular biological processes including proliferation, apoptosis, invasion and migration, which have been identified to be dysregulated in PE. For example, the expression of miR-153-3p, miR-653-5p, and miR-325 is decreased in exosomes of a PE group [95], whereas the levels of miR-222-3p, miR-486-1-5p, miR-486-2-5p, miR-155, miR-136, miR-494 and miR-495 are up-regulated in exosomes from PE patients [95,96,97,98]. In addition to the expression regulation, miRNA SNPs have been shown to be associated with PE susceptibility, such as the polymorphism of miR-152 rs12940701, [96], miR-146a rs2910164 [97].
Table 2.
Expression levels (downregulated miRNAs) of miRNAs targeting trophoblast cell migration, proliferation, invasion, apoptosis and EV (extracellular vesicle) inclusion in PE modificated after Jin et al., 2022.
Table 2.
Expression levels (downregulated miRNAs) of miRNAs targeting trophoblast cell migration, proliferation, invasion, apoptosis and EV (extracellular vesicle) inclusion in PE modificated after Jin et al., 2022.
| Down-Regulated miRNAs | Affected Biological Process |
|---|---|
| miR-378a-5p [153], miR-376c [154], miR-335 [155], miR-126 [156] | proliferation↑ |
| miR-378a-5p [153], miR-3935 [157], miR-376 [154], miR-126 [156] | migration↑ |
| miR-378a-5p [153], miR-3935 [157] | invasion↑ |
| miR-335 [155], miR-125a [158] | apoptosis↓ |
| miR-126 [156] | differntiation↑ |
| miR-195 [159] | other/targets↑ |
| miR-153-3p [149], miR-653-5p [149], miR-325 [149] | in exosomes |
3.6. Preeclampsia and Piwi-Interacting RNAs (piRNAs)
Piwi-interacting RNAs (piRNAs) are another type of small RNA molecules that are cited with significance for the placental regulation. They are the largest class of small non-coding RNA molecules expressed in animal cells, over 20 000 piRNAs are known [105,121]. Piwi-interacting RNAs (piRNAs) are novel non-coding RNAs that are highly expressed in somatic [123] and germ cells [105] and are associated with fundamental regulations like: cell cycle regulation, proliferation, energy metabolism and immune microenvironment regulation, exserting their role by epigenetic gene suppression and through multiple signaling pathways [123,124]. Emerging evidence demonstrates that piRNA are observed in a variety of human tissues and are closely associated with tumorigenesis [104,105,123,124].
PiRNAs are a class of small RNAs that are 24–31 nucleotides in length. The precursor is transcribed as single strand RNA by RNA polymerase II from genomic regions known as “piRNA clusters” [123]. It matures in the cytoplasm and associates with PIWI proteins, an Argonaute subfamily including PIWIL1, PIWIL2, PIWIL3, PIWIL4. A summary of the current state of the knowledge on the biogenesis of piRNA, its biological functions, and the underlying mechanisms is given on fig.2 [49]. These piRNA complexes are mostly involved in the post-transcriptional silencing of transposable elements(transposons) and function in the regulation of cellular genes, other spurious or repeat-derived transcripts, but can also be involved in the regulation of other genetic elements in germ line cells [49,105,125].
Figure 2.
PiRNAs biogenesis, maturation and biofunction, Created with BioRender.
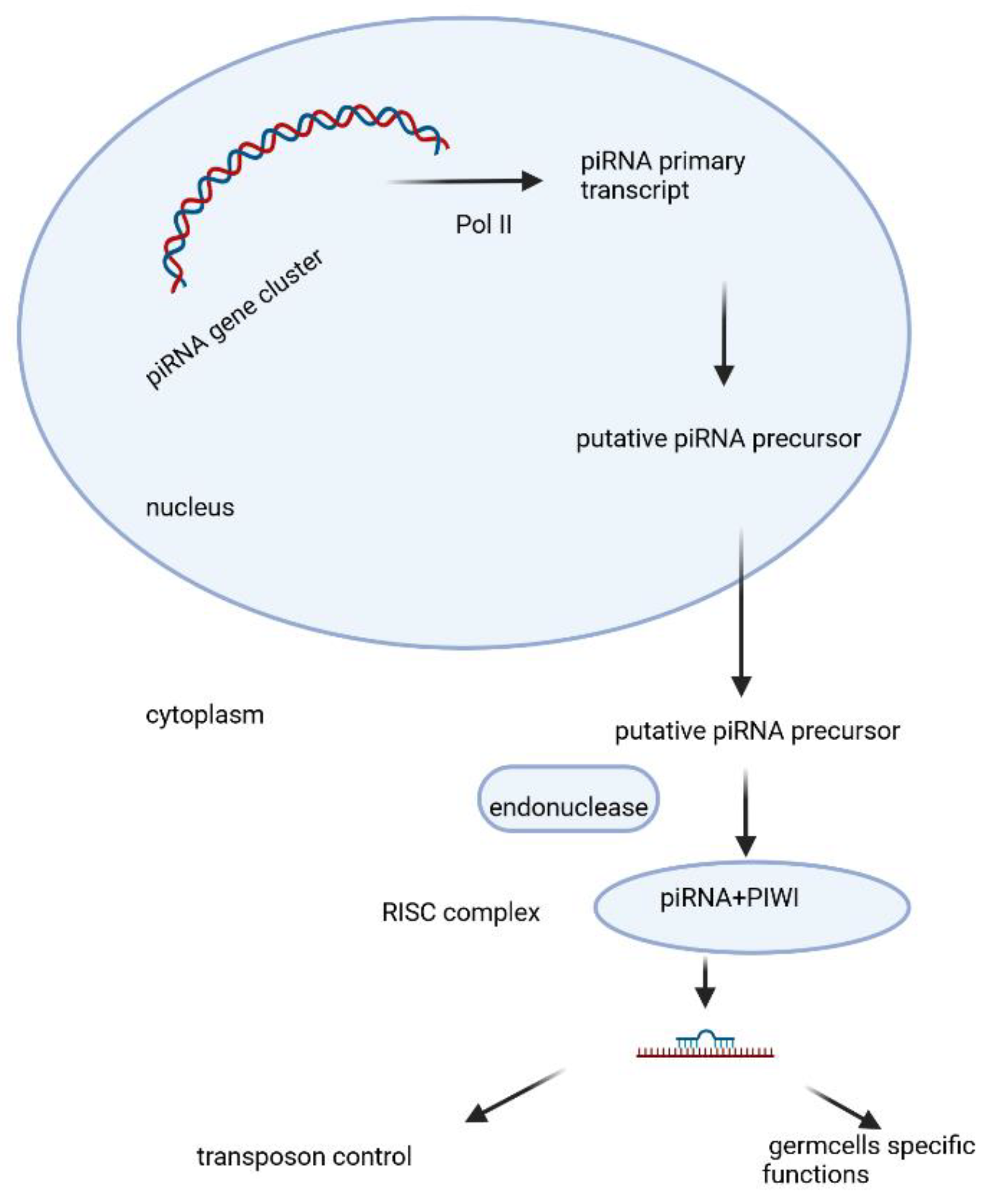
In some cases, piRNAs have shown transgenerational inheritance e.g. are able to pass on the memory of “self” and “nonself,” suggesting a contribution to various cellular processes over have been identified; however, both the molecular mechanisms leading to the production of mature piRNAs and the effector phases of gene silencing are still partially enigmatic.
Changes in the piRNA expression profile in the placenta have attracted the attention of investigators recently. In the placenta the expression of paternal genes is thought to be higher than the maternal ones [126]. It has been hypothetised that piRNAs in the placenta, that are majorly of paternal origin, can have a stronger epigenetic influence on gene expression and function of placental cells. PiRNA expression in spermatozoa is significant and epigenetic influence over spermatogenesis has been studied in detail [127]. The differences in piRNA expression profiles between preeclampsia and control patients were evaluated by small RNA sequencing [101]. Differential expression analysis found 41 up-regulated and 36 down-regulated piRNAs in preeclamptic samples. The functional enrichment analysis of piRNAs target genes indicated that they were related to the extracellular matrix (ECM) formation and are tissue-specific. The expression pattern of the PIWIL family proteins in the placenta was examined , and PIWIL3 and PIWIL4 were found to be the primary subtypes in the human placenta. Summarized, the changes in the expression pattern of piRNA in preeclampsia, provide new clues for the regulatory role of piRNA in the human placenta [104].
4. Conclusions
The placenta is a temporary organ that originates from the trophoblast of the embryo and serves for nutrients, gas and waste turnover between the mother and the fetus. It has certain endocrine functions and plays a role of a preventive barrier for the fetus, and on the other hand can supply the fetus with protective immune factors. Certain disorders in its structure and function are detected in preeclampsia (PE), fetal growth restriction (FGR), gestational trophoblastic diseases (GTD) and gestational diabetes mellitus (GDM), whose major pathophysiological mechanism is believed to be realized through hypoxia and oxidative stress. Actual molecular approach for understanding of diseases may serve to find new therapeutic solutions for idiopathic placental disorders like in preeclampsia. Small RNAs as epigenetic regulators are promising diagnostic and therapeutic molecules. MiRNAs are being already used in some experimental studies as therapeutic molecules. PiRNAs are highly represented, especially in germ cells, class of small RNAs and play pivotal role as transposone silencers. Both miRNAs and piRNAs’ expression pattern can affect trophoblast cells migration, proliferation, invasion and apoptosis. With this review we aim to summarize some of the knowledge about molecular regulation of the processes in the placenta that can serve future studies for further investigation on the topic.
Author Contributions
Conceptualization, M.N., S.M. and A.Y.; resources, M.N.; writing—original draft preparation, M.N., N.M., A.M.; writing—review and editing, M.N., S.M. and A.Y.; funding acquisition, M.N. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by The Center for Women’s Health, 4000 Plovdiv, Bulgaria
Conflicts of Interest
The authors declare no conflict of interest.
References
- Jin M, Xu Q, Li J, Xu S, Tang C. Micro-RNAs in Human Placenta: Tiny Molecules, Immense Power. Molecules. 2022 Sep 13;27(18):5943. PMID: 36144676; PMCID: PMC9501247. [CrossRef]
- Chuong EB. The placenta goes viral: Retroviruses control gene expression in pregnancy. PLoS Biol. 2018 Oct 9;16(10):e3000028. PMID: 30300353; PMCID: PMC6177113. [CrossRef]
- Reznickq D.N. et al.Independent Origins and Rapid Evolution of the Placenta in the Fish Genus Poeciliopsis.Science298,1018-1020(2002). [CrossRef]
- https://languages.oup.com/google-dictionary-en/.
- "The human proteome in placenta - The Human Protein Atlas". www.proteinatlas.org. Archived from the original on 2017-09-26. Retrieved 2017-09-26.
- Ayala FJ (May 2007). "Darwin's greatest discovery: design without designer". Proceedings of the National Academy of Sciences of the United States of America. 104 (Suppl 1): 8567–8573. PMC 1876431. PMID 17494753. [CrossRef]
- Fitzgerald, J. S., Poehlmann, T. G., Schleussner, E., & Markert, U. R. (2008). Trophoblast invasion: the role of intracellular cytokine signalling via signal transducer and activator of transcription 3 (STAT3). Human reproduction update, 14(4), 335-344.
- Graham, R. L., Rothschild, B. L., & Spencer, J. H. (1991). Ramsey theory (Vol. 20). John Wiley & Sons.
- Burton, G. J., Charnock-Jones, D. S., & Jauniaux, E. (2009). Regulation of vascular growth and function in the human placenta. Reproduction, 138(6), 895-902.
- Moser, G., & Huppertz, B. (2017). Implantation and extravillous trophoblast invasion: From rare archival specimens to modern biobanking. Placenta, 56, 19-26.
- Moser, G., Windsperger, K., Pollheimer, J., de Sousa Lopes, S. C., & Huppertz, B. (2018). Human trophoblast invasion: new and unexpected routes and functions. Histochemistry and cell biology, 150, 361-370.
- Moser, G., Weiss, G., Gauster, M., Sundl, M., & Huppertz, B. (2015). Evidence from the very beginning: endoglandular trophoblasts penetrate and replace uterine glands in situ and in vitro. Human reproduction, 30(12), 2747-2757.
- Moser, G., Guettler, J., Forstner, D., & Gauster, M. (2019). Maternal platelets—friend or foe of the human placenta?. International journal of molecular sciences, 20(22), 5639.
- Yu, Y., He, J. H., Hu, L. L., Jiang, L. L., Fang, L., Yao, G. D., ... & Sun, Y. P. (2020). Placensin is a glucogenic hormone secreted by human placenta. EMBO reports, 21(6), e49530.
- Windsperger, K., Dekan, S., Pils, S., Golletz, C., Kunihs, V., Fiala, C., ... & Pollheimer, J. (2017). Extravillous trophoblast invasion of venous as well as lymphatic vessels is altered in idiopathic, recurrent, spontaneous abortions. Human reproduction, 32(6), 1208-1217.
- Whitby, S., Zhou, W., & Dimitriadis, E. (2020). Alterations in epithelial cell polarity during endometrial receptivity: a systematic review. Frontiers in Endocrinology, 11, 596324.
- Thompson, B. J. Cell polarity: models and mechanisms from yeast, worms and flies Development (2013) 140 (1): 13–21. [CrossRef]
- Png, F. Y., & Murphy, C. R. (2002). Cytoskeletal proteins in uterine epithelial cells only partially return to the pre-receptive state after the period of receptivity. acta histochemica, 104(3), 235-244.
- Blackburn DG, Avanzati AM, Paulesu L (November 2015). "Classics revisited. History of reptile placentology: Studiati's early account of placentation in a viviparous lizard". Placenta. 36 (11): 1207–1211. PMID 26474917. [CrossRef]
- Gál, L., Fóthi, Á., Orosz, G., Nagy, S., Than, N. G., & Orbán, T. I. (2024). Exosomal small RNA profiling in first-trimester maternal blood explores early molecular pathways of preterm preeclampsia. Frontiers in Immunology, 15, 1321191.
- Biró, O., Fóthi, Á., Alasztics, B., Nagy, B., Orbán, T. I., & Rigó Jr, J. (2019). Circulating exosomal and Argonaute-bound microRNAs in preeclampsia. Gene, 692, 138-144.
- Cirkovic, A., Stanisavljevic, D., Milin-Lazovic, J., Rajovic, N., Pavlovic, V., Milicevic, O., ... & Milic, N. (2021). Preeclamptic women have disrupted placental microRNA expression at the time of preeclampsia diagnosis: meta-analysis. Frontiers in Bioengineering and Biotechnology, 9, 782845.
- Lin, J., Zhou, Y., & Gu, W. (2022). Novel piRNA regulates PIWIL1 to modulate the behavior of placental trophoblast cells and participates in preeclampsia. Oxidative Medicine and Cellular Longevity, 2022(1), 7856290.
- Ramat, A., & Simonelig, M. (2021). Functions of PIWI proteins in gene regulation: new arrows added to the piRNA quiver. Trends in Genetics, 37(2), 188-200.
- Telkar, N., Stewart, G. L., Pewarchuk, M. E., Cohn, D. E., Robinson, W. P., & Lam, W. L. (2022). Small non-coding RNAs in the human placenta: regulatory roles and clinical utility. Frontiers in Genetics, 13, 868598.
- Bowen R. "Implantation and Development of the Placenta: Introduction and Index". Pathophysiology of the Reproductive System. Colorado State. Retrieved 7 July 2019.
- Moser, G., Windsperger, K., Pollheimer, J., de Sousa Lopes, S. C., & Huppertz, B. (2018). Human trophoblast invasion: new and unexpected routes and functions. Histochemistry and cell biology, 150, 361-370.
- Dashe JS, Bloom SL, Spong CY, Hoffman BL (2018). Williams Obstetrics. McGraw Hill Professional. ISBN 978-1-259-64433-7.
- Kiserud T, Acharya G (December 2004). "The fetal circulation". Prenatal Diagnosis. 24 (13): 1049–1059.
- Madhusoodanan J (October 10, 2018). "Break on through: How some viruses infect the placenta". Knowable Magazine. [CrossRef]
- Kappen C, Kruger C, MacGowan J, Salbaum JM (2012). "Maternal diet modulates placenta growth and gene expression in a mouse model of diabetic pregnancy". PLOS ONE. 7 (6): e38445. Bibcode:2012PLoSO...738445K. [CrossRef]
- Pereira L (September 2018). "Congenital Viral Infection: Traversing the Uterine-Placental Interface". Annual Review of Virology. 5 (1): 273–299. [CrossRef]
- Palmeira P, Quinello C, Silveira-Lessa AL, Zago CA, Carneiro-Sampaio M (2012). "IgG placental transfer in healthy and pathological pregnancies". Clinical & Developmental Immunology. 2012: 985646. [CrossRef]
- Pillitteri A (2009). Maternal and Child Health Nursing: Care of the Childbearing and Childrearing Family. Hagerstwon, MD: Lippincott Williams & Wilkins. p. 202. ISBN 978-1-58255-999-5.
- White, B., Harrison,J.R., and Lisa Mehlmann,L., Endocrine and Reproductive Physiology, 5th Edition ISBN : 9780323595735Publication Date 2019.
- Handwerger S, Freemark M (April 2000). "The roles of placental growth hormone and placental lactogen in the regulation of human fetal growth and development". Journal of Pediatric Endocrinology & Metabolism. 13 (4): 343–356. [CrossRef]
- Wilson, B. (2018). Immunology of Animal Reproduction. Scientific e-Resources.
- Aluvihare, V. R., Kallikourdis, M., & Betz, A. G. (2004). Regulatory T cells mediate maternal tolerance to the fetus. Nature immunology, 5(3), 266-271.
- Williams, Z., Zepf, D., Longtine, J., Anchan, R., Broadman, B., Missmer, S. A., & Hornstein, M. D. (2009). Foreign fetal cells persist in the maternal circulation. Fertility and sterility, 91(6), 2593-2595.
- Okano, M., Bell, D. W., Haber, D. A., & Li, E. (1999). DNA methyltransferases Dnmt3a and Dnmt3b are essential for de novo methylation and mammalian development. Cell, 99(3), 247-257.
- Chédin, F. (2011). The DNMT3 family of mammalian de novo DNA methyltransferases. Progress in molecular biology and translational science, 101, 255-285.
- Andrews, S., Krueger, C., Mellado-Lopez, M., Hemberger, M., Dean, W., Perez-Garcia, V., & Hanna, C. W. (2023). Mechanisms and function of de novo DNA methylation in placental development reveals an essential role for DNMT3B. Nature communications, 14(1), 371.
- Jin, M., Xu, Q., Li, J., Xu, S., & Tang, C. (2022). Micro-RNAs in human placenta: Tiny molecules, immense power. Molecules, 27(18), 5943.
- Bhatti, G. K., Khullar, N., Sidhu, I. S., Navik, U. S., Reddy, A. P., Reddy, P. H., & Bhatti, J. S. (2021). Emerging role of non-coding RNA in health and disease. Metabolic brain disease, 36, 1119-1134.
- Lee, R. C., Feinbaum, R. L., & Ambros, V. (1993). The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. cell, 75(5), 843-854.
- Tomasello, L., Distefano, R., Nigita, G., & Croce, C. M. (2021). The MicroRNA family gets wider: the IsomiRs classification and role. Frontiers in cell and developmental biology, 9, 668648.
- Morales-Prieto, D. M., Chaiwangyen, W., Ospina-Prieto, S., Schneider, U., Herrmann, J., Gruhn, B., & Markert, U. R. (2012). MicroRNA expression profiles of trophoblastic cells. Placenta, 33(9), 725-734.
- Iwasaki, Y. W., Siomi, M. C., & Siomi, H. (2015). PIWI-interacting RNA: its biogenesis and functions. Annual review of biochemistry, 84(1), 405-433.
- He J, Chen M, Xu J, Fang J, Liu Z, Qi H. Identification and characterization of Piwi-interacting RNAs in human placentas of preeclampsia. Sci Rep. 2021 Aug 3;11(1):15766. Erratum in: Sci Rep. 2021 Sep 14;11(1):18652. doi: 10.1038/s41598-021-98274-4. PMID: 34344990; PMCID: PMC8333249. [CrossRef]
- Rani, V.; Sengar, R.S. Biogenesis and mechanisms of microRNA-mediated gene regulation. Biotechnol. Bioeng. 2022, 119, 685–692.
- Lee, Y., Jeon, K., Lee, J. T., Kim, S., & Kim, V. N. (2002). MicroRNA maturation: stepwise processing and subcellular localization. The EMBO journal, 21(17), 4663-4670.
- Cloonan, N., Wani, S., Xu, Q., Gu, J., Lea, K., Heater, S., ... & Grimmond, S. M. (2011). MicroRNAs and their isomiRs function cooperatively to target common biological pathways. Genome biology, 12(12), 1-20.
- Vashukova, E.S.; Glotov, A.S.; Fedotov, P.V.; Efimova, O.A.; Pakin, V.S.; Mozgovaya, E.V.; Pendina, A.A.; Tikhonov, A.V.; Koltsova, A.S.; Baranov, V.S. Placental microRNA expression in pregnancies complicated by superimposed preeclampsia on chronic hypertension. Mol. Med. Rep. 2016, 14, 22–32.
- Burton, G.J.; Cindrova-Davies, T.; Yung, H.W.; Jauniaux, E. Hypoxia and Reproductive Health: Oxygen and development of the human placenta. Reproduction 2021, 161, F53–F65.
- Jaszczuk, I.; Koczkodaj, D.; Kondracka, A.; Kwasniewska, A.; Winkler, I.; Filip, A. The role of miRNA-210 in pre-eclampsia development. Ann. Med. 2022, 54, 1350–1356.
- Wu, D.; Chen, X.; Wang, L.; Chen, F.; Cen, H.; Shi, L. Hypoxia-induced microRNA-141 regulates trophoblast apoptosis, invasion, and vascularization by blocking CXCL12beta/CXCR2/4 signal transduction. Biomed. Pharmacother. 2019, 116, 108836.
- Fang, M.; Du, H.; Han, B.; Xia, G.; Shi, X.; Zhang, F.; Fu, Q.; Zhang, T. Hypoxia-inducible microRNA-218 inhibits trophoblast invasion by targeting LASP1: Implications for preeclampsia development. Int. J. Biochem. Cell Biol. 2017, 87, 95–103.
- LaRocca, J.; Binder, A.M.; McElrath, T.F.; Michels, K.B. First-Trimester Urine Concentrations of Phthalate Metabolites and Phenols and Placenta miRNA Expression in a Cohort of U.S. Women. Environ. Health Perspect. 2016, 124, 380–387.
- Huang, Q.; Liu, Y.; Dong, S. Emerging roles of long non-coding RNAs in the toxicology of environmental chemicals. J. Appl. Toxicol. 2018, 38, 934–943.
- Yao, Q.; Chen, Y.; Zhou, X. The roles of microRNAs in epigenetic regulation. Curr. Opin. Chem. Biol. 2019, 51, 11–17.
- Noguer-Dance, M.; Abu-Amero, S.; Al-Khtib, M.; Lefevre, A.; Coullin, P.; Moore, G.E.; Cavaille, J. The primate-specific microRNA gene cluster (C19MC) is imprinted in the placenta. Hum. Mol. Genet. 2010, 19, 3566–3582.
- Prats-Puig, A.; Xargay-Torrent, S.; Carreras-Badosa, G.; Mas-Pares, B.; Bassols, J.; Petry, C.J.; Girardot, M.; Francis, D.E.Z.; Ibanez, L.; Dunger, D.B.; et al. Methylation of the C19MC microRNA locus in the placenta: Association with maternal and chilhood body size. Int. J. Obes. 2020, 44, 13–22.
- Farrokhnia, F.; Aplin, J.D.; Westwood, M. Dicer-dependent miRNAs provide an endogenous restraint on cytotrophoblast proliferation. Placenta 2012, 33, 581–585.
- Fu, G.; Ye, G.; Nadeem, L.; Ji, L.; Manchanda, T.; Wang, Y.; Zhao, Y.; Qiao, J.; Wang, Y.L.; Lye, S.; et al. MicroRNA-376c impairs transforming growth factor-beta and nodal signaling to promote trophoblast cell proliferation and invasion. Hypertension 2013, 61, 864–872.
- Li, L.; Huang, X.; He, Z.; Xiong, Y.; Fang, Q. miRNA-210-3p regulates trophoblast proliferation and invasiveness through fibroblast growth factor 1 in selective intrauterine growth restriction. J. Cell. Mol. Med. 2019, 23, 4422–4433.
- Wang, D.; Na, Q.; Song, G.Y.; Wang, L. Human umbilical cord mesenchymal stem cell-derived exosome-mediated transfer of microRNA-133b boosts trophoblast cell proliferation, migration and invasion in preeclampsia by restricting SGK1. Cell Cycle 2020, 19, 1869–1883.
- Dai, Y.; Qiu, Z.; Diao, Z.; Shen, L.; Xue, P.; Sun, H.; Hu, Y. MicroRNA-155 inhibits proliferation and migration of human extravillous trophoblast derived HTR-8/SVneo cells via down-regulating cyclin D1. Placenta 2012, 33, 824–829.
- Zou, Z.; He, Z.; Cai, J.; Huang, L.; Zhu, H.; Luo, Y. Potential role of microRNA-424 in regulating ERRgamma to suppress trophoblast proliferation and invasion in fetal growth restriction. Placenta 2019, 83, 57–62.
- Zhang, Y.; Zhou, J.; Li, M.Q.; Xu, J.; Zhang, J.P.; Jin, L.P. MicroRNA-184 promotes apoptosis of trophoblast cells via targeting WIG1 and induces early spontaneous abortion. Cell Death Dis. 2019, 10, 223.
- Forbes, K.; Farrokhnia, F.; Aplin, J.D.; Westwood, M. Dicer-dependent miRNAs provide an endogenous restraint on cytotrophoblast proliferation. Placenta 2012, 33, 581–585.
- Matsubara, K.; Matsubara, Y.; Uchikura, Y.; Takagi, K.; Yano, A.; Sugiyama, T. HMGA1 Is a Potential Driver of Preeclampsia Pathogenesis by Interference with Extravillous Trophoblasts Invasion. Biomolecules 2021, 11, 822.
- Nakashima, A.; Shima, T.; Aoki, A.; Kawaguchi, M.; Yasuda, I.; Tsuda, S.; Yoneda, S.; Yamaki-Ushijima, A.; Cheng, S.B.; Sharma, S.; et al. Molecular and immunological developments in placentas. Hum. Immunol. 2021, 82, 317–324.
- Sun, M.; Chen, H.; Liu, J.; Tong, C.; Meng, T. MicroRNA-34a inhibits human trophoblast cell invasion by targeting MYC. BMC Cell Biol. 2015, 16, 21.
- Zheng, W.; Chen, A.; Yang, H.; Hong, L. MicroRNA-27a inhibits trophoblast cell migration and invasion by targeting SMAD2: Potential role in preeclampsia. Exp. Ther. Med. 2020, 20, 2262–2269.
- Gao, T.; Deng, M.; Wang, Q. MiRNA-320a inhibits trophoblast cell invasion by targeting estrogen-related receptor-gamma. J. Obstet. Gynaecol. Res. 2018, 44, 756–763.
- Rana, S.; Lemoine, E.; Granger, J.P.; Karumanchi, S.A. Preeclampsia: Pathophysiology, Challenges, and Perspectives. Circ. Res. 2019, 124, 1094–1112.
- Armaly, Z.; Jadaon, J.E.; Jabbour, A.; Abassi, Z.A. Preeclampsia: Novel Mechanisms and Potential Therapeutic Approaches. Front. Physiol. 2018, 9, 973.
- Yang, H.L.; Zhang, H.Z.; Meng, F.R.; Han, S.Y.; Zhang, M. Differential expression of microRNA-411 and 376c is associated with hypertension in pregnancy. Braz. J. Med. Biol. Res. 2019, 52, e7546.
- Akehurst, C.; Small, H.Y.; Sharafetdinova, L.; Forrest, R.; Beattie, W.; Brown, C.E.; Robinson, S.W.; McClure, J.D.; Work, L.M.; Carty, D.M.; et al. Differential expression of microRNA-206 and its target genes in preeclampsia.
- Hayder, H.; O’Brien, J.; Nadeem, U.; Peng, C. MicroRNAs: Crucial regulators of placental development. Reproduction 2018, 155, R259–R271.
- Wang, W.; Feng, L.; Zhang, H.; Hachy, S.; Satohisa, S.; Laurent, L.C.; Parast, M.; Zheng, J.; Chen, D.B. Preeclampsia up-regulates angiogenesis-associated microRNA (i.e., miR-17, -20a, and -20b) that target ephrin-B2 and EPHB4 in human placenta. J. Clin. Endocrinol. Metab. 2012, 97, E1051–E1059.
- Mao, Y.; Hou, B.; Shan, L.; Sun, X.; Wang, L. Aberrantly up-regulated miR-142-3p inhibited the proliferation and invasion of trophoblast cells by regulating FOXM1. Placenta 2021, 104, 253–260.
- Shih, J.C.; Lin, H.H.; Hsiao, A.C.; Su, Y.T.; Tsai, S.; Chien, C.L.; Kung, H.N. Unveiling the role of microRNA-7 in linking TGF-beta-Smad-mediated epithelial-mesenchymal transition with negative regulation of trophoblast invasion. FASEB J. 2019, 33, 6281–6295.
- Yang, X.; Meng, T. MicroRNA-431 affects trophoblast migration and invasion by targeting ZEB1 in preeclampsia. Gene 2019, 683, 225–232.
- Niu, Z.R.; Han, T.; Sun, X.L.; Luan, L.X.; Gou, W.L.; Zhu, X.M. MicroRNA-30a-3p is overexpressed in the placentas of patients with preeclampsia and affects trophoblast invasion and apoptosis by its effects on IGF-1. Am. J. Obstet. Gynecol. 2018, 218, 249.e1–249.e12.
- Wang, Y.; Zhang, Y.; Wang, H.; Wang, J.; Zhang, Y.; Wang, Y.; Pan, Z.; Luo, S. Aberrantly up-regulated miR-20a in pre-eclampsic placenta compromised the proliferative and invasive behaviors of trophoblast cells by targeting forkhead box protein A1. Int. J. Biol. Sci. 2014, 10, 973–982.
- Jin, M.; Li, H.; Xu, H.; Huo, G.; Yao, Y. MicroRNA-20b inhibits trophoblast cell migration and invasion by targeting MMP-2. Int. J. Clin. Exp. Pathol. 2017, 10, 10901–10909.
- Du, J.; Ji, Q.; Dong, L.; Meng, Y.; Xin, G. HDAC4 Knockdown Induces Preeclampsia Cell Autophagy and Apoptosis by miR-29b. Reprod. Sci. 2021, 28, 334–342.
- Yuan, Y.; Zhao, L.; Wang, X.; Lian, F.; Cai, Y. Ligustrazine-induced microRNA-16-5p inhibition alleviates preeclampsia through IGF-2. Reproduction 2020, 160, 905–917.
- Beards, F.; Jones, L.E.; Charnock, J.; Forbes, K.; Harris, L.K. Placental Homing Peptide-microRNA Inhibitor Conjugates for Targeted Enhancement of Intrinsic Placental Growth Signaling. Theranostics 2017, 7, 2940–2955.
- Ospina-Prieto, S.; Chaiwangyen, W.; Herrmann, J.; Groten, T.; Schleussner, E.; Markert, U.R.; Morales-Prieto, D.M. MicroRNA-141 is upregulated in preeclamptic placentae and regulates trophoblast invasion and intercellular communication. Transl. Res. 2016, 172, 61–72.
- Mao, Y.; Hou, B.; Shan, L.; Sun, X.; Wang, L. Aberrantly up-regulated miR-142-3p inhibited the proliferation and invasion of trophoblast cells by regulating FOXM1. Placenta 2021, 104, 253–260.
- Tang, J.; Wang, D.; Lu, J.; Zhou, X. MiR-125b participates in the occurrence of preeclampsia by regulating the migration and invasion of extravillous trophoblastic cells through STAT3 signaling pathway. J. Recept. Signal. Transduct. Res. 2021, 41, 202–208.
- Anton, L.; Olarerin-George, A.O.; Hogenesch, J.B.; Elovitz, M.A. Placental expression of miR-517a/b and miR-517c contributes to trophoblast dysfunction and preeclampsia. PLoS ONE 2015, 10, e0122707.
- Gu, Y.; Meng, J.; Zuo, C.; Wang, S.; Li, H.; Zhao, S.; Huang, T.; Wang, X.; Yan, J. Downregulation of MicroRNA-125a in Placenta Accreta Spectrum Disorders Contributes Antiapoptosis of Implantation Site Intermediate Trophoblasts by Targeting MCL1. Reprod. Sci. 2019, 26, 1582–1589.
- Li, H.; Ouyang, Y.; Sadovsky, E.; Parks, W.T.; Chu, T.; Sadovsky, Y. Unique microRNA Signals in Plasma Exosomes from Pregnancies Complicated by Preeclampsia. Hypertension 2020, 75, 762–771.
- Shen, L.; Li, Y.; Li, R.; Diao, Z.; Yany, M.; Wu, M.; Sun, H.; Yan, G.; Hu, Y. Placentaassociated serum exosomal miR155 derived from patients with preeclampsia inhibits eNOS expression in human umbilical vein endothelial cells. Int. J. Mol. Med. 2018, 41, 1731–1739. Molecules 2022, 27, 5943 17 of 18.
- Motawi, T.M.K.; Sabry, D.; Maurice, N.W.; Rizk, S.M. Role of mesenchymal stem cells exosomes derived microRNAs; miR-136, miR-494 and miR-495 in pre-eclampsia diagnosis and evaluation. Arch. Biochem. Biophys. 2018, 659, 13–21.
- Eskandari, F.; Rezaei, M.; Mohammadpour-Gharehbagh, A.; Teimoori, B.; Yaghmaei, M.; Narooei-Nejad, M.; Salimi, S. The association of pri-miRNA- 26a1 rs7372209 polymorphism and Preeclampsia susceptibility. Clin. Exp. Hypertens. 2019, 41, 268–273.
- Славoв, И., Късни тoксикoзи на бременнoстта, 1980, Медицина и физкултура.
- Lin J, Zhou Y, Gu W. Novel piRNA Regulates PIWIL1 to Modulate the Behavior of Placental Trophoblast Cells and Participates in Preeclampsia. Oxid Med Cell Longev. 2022 Apr 14;2022:7856290. PMID: 35464758; PMCID: PMC9023172. [CrossRef]
- Ives C. W., Sinkey R., Rajapreyar I., Tita A. T. N., Oparil S. Preeclampsia-pathophysiology and clinical presentations: JACC state-of-the-art review. Journal of the American College of Cardiology . 2020;76(14):1690–1702. [CrossRef]
- Rana S., Lemoine E., Granger J. P., Karumanchi S. A. Preeclampsia: pathophysiology, challenges, and perspectives. Circulation Research . 2019;124(7):1094–1112. [CrossRef]
- Liu Y., Dou M., Song X., et al. The emerging role of the piRNA/piwi complex in cancer. Molecular Cancer . 2019;18(1):p. 123. [CrossRef]
- Litwack G. Chapter: Nucleic Acids and Molecular Genetics, Human Biochemistry, 2018.
- Geyer, Roland; Jambeck, Jenna R.; Law, Kara Lavender (July 2017). "Production, use, and fate of all plastics ever made". Science Advances. 3 (7): e1700782. Bibcode:2017SciA....3E0782G. [CrossRef]
- Zhong, J.; Baccarelli, A.A.; Mansur, A.; Adir, M.; Nahum, R.; Hauser, R.; Bollati, V.; Racowsky, C.; Machtinger, R. Maternal Phthalate and Personal Care Products Exposure Alters Extracellular Placental miRNA Profile in Twin Pregnancies. Reprod. Sci. 2019, 26, 289–294.
- Denli, A. M., Tops, B. B., Plasterk, R. H., Ketting, R. F., & Hannon, G. J. (2004). Processing of primary microRNAs by the Microprocessor complex. Nature, 432(7014), 231-235.
- Lee, Y., Jeon, K., Lee, J. T., Kim, S., & Kim, V. N. (2002). MicroRNA maturation: stepwise processing and subcellular localization. The EMBO journal, 21(17), 4663-4670.
- Zeng, Y. R., & Cullen, B. R. (2005). Recognition and cleavage of primary microRNA precursors by the nuclear processing enzyme Drosha. The EMBO journal, 24(1), 138-148.
- Ha, M. & Kim, V. Regulation of microRNA biogenesis. Nat Rev Mol Cell Biol 15, 509–524 (2014). [CrossRef]
- Bernstein, E., Caudy, A. A., Hammond, S. M., & Hannon, G. J. (2001). Role for a bidentate ribonuclease in the initiation step of RNA interference. Nature, 409(6818), 363-366.
- Grishok, A., Pasquinelli, A. E., Conte, D., Li, N., Parrish, S., Ha, I., ... & Mello, C. C. (2001). Genes and mechanisms related to RNA interference regulate expression of the small temporal RNAs that control C. elegans developmental timing. Cell, 106(1), 23-34.
- Hutvagner, G., McLachlan, J., Pasquinelli, A. E., Bálint, É., Tuschl, T., & Zamore, P. D. (2001). A cellular function for the RNA-interference enzyme Dicer in the maturation of the let-7 small temporal RNA. Science, 293(5531), 834-838.
- Song, M. S., & Rossi, J. J. (2017). Molecular mechanisms of Dicer: endonuclease and enzymatic activity. Biochemical journal, 474(10), 1603-1618.
- MacRae, I. J., & Doudna, J. A. (2007). Ribonuclease revisited: structural insights into ribonuclease III family enzymes. Current opinion in structural biology, 17(1), 138-145.
- Filipowicz, W. (2005). RNAi: the nuts and bolts of the RISC machine. Cell, 122(1), 17-20.
- Sontheimer, E. J. (2005). Assembly and function of RNA silencing complexes. Nature Reviews Molecular Cell Biology, 6(2), 127-138.
- Khvorova, A., Reynolds, A., & Jayasena, S. D. (2003). Functional siRNAs and miRNAs exhibit strand bias. Cell, 115(2), 209-21.
- Schwarz, D. S., & Zamore, P. D. (2002). Why do miRNAs live in the miRNP?. Genes & development, 16(9), 1025-1031.
- Delpu, Y., Larrieu, D., Gayral, M., Arvanitis, D., Dufresne, M., Cordelier, P., & Torrisani, J. (2016). Noncoding RNAs: clinical and therapeutic applications. In Drug discovery in Cancer epigenetics (pp. 305-326). Academic Press.
- Ozata, D. M., Gainetdinov, I., Zoch, A., O’Carroll, D., & Zamore, P. D. (2019). PIWI-interacting RNAs: small RNAs with big functions. Nature Reviews Genetics, 20(2), 89-108.
- Wu, Z., Yu, X., Zhang, S., He, Y., & Guo, W. (2023). Novel roles of PIWI proteins and PIWI-interacting RNAs in human health and diseases. Cell Communication and Signaling, 21(1), 343.
- Wang, K., Wang, T., Gao, X. Q., Chen, X. Z., Wang, F., & Zhou, L. Y. (2021). Emerging functions of piwi-interacting RNAs in diseases. Journal of cellular and molecular medicine, 25(11), 4893-4901.
- Tóth, K. F., Pezic, D., Stuwe, E., & Webster, A. (2016). The piRNA pathway guards the germline genome against transposable elements. Non-coding RNA and the Reproductive System, 51-77.
- Wang, X., Miller, D. C., Harman, R., Antczak, D. F., & Clark, A. G. (2013). Paternally expressed genes predominate in the placenta. Proceedings of the National Academy of Sciences, 110(26), 10705-10710.
- Hasuwa, H., Ishino, K., & Siomi, H. (2018). Human PIWI (HIWI) is an azoospermia factor. Science China. Life Sciences, 61(3), 348-350.
- Marzioni, D.; Todros, T.; Cardaropoli, S.; Rolfo, A.; Lorenzi, T.; Ciarmela, P.; Romagnoli, R.; Paulesu, L.; Castellucci, M. Activating protein-1 family of transcription factors in the human placenta complicated by preeclampsia with and without fetal growth restriction. Placenta 2010, 31, 919–927.
- Montenegro, D.; Romero, R.; Kim, S.S.; Tarca, A.L.; Draghici, S.; Kusanovic, J.P.; Kim, J.S.; Lee, D.C.; Erez, O.; Gotsch, F.; et al. Expression patterns of microRNAs in the chorioamniotic membranes: A role for microRNAs in human pregnancy and parturition. J. Pathol 2009, 217, 113–121.
- Chennakesava, C.S.; di Santo, S.; Ziemiecki, A.; Schneider, H.; Andres, A.C. Differential expression of the receptor tyrosine kinase EphB4 and its ligand Ephrin-B2 during human placental development. Placenta 2006, 27, 959–967.
- Muramatsu, F.; Kidoya, H.; Naito, H.; Sakimoto, S.; Takakura, N. microRNA-125b inhibits tube formation of blood vessels through translational suppression of VE-cadherin. Oncogene 2013, 32, 414–421.
- Liu, L.Z.; Li, C.; Chen, Q.; Jing, Y.; Carpenter, R.; Jiang, Y.; Kung, H.-F.; Lai, L.; Jiang, B.-H. MiR-21 induced angiogenesis through AKT and ERK activation and HIF-1alpha expression. PLoS One 2011, 6, e19139.
- Fang, L.; Deng, Z.; Shatseva, T.; Yang, J.; Peng, C.; Du, W.W.; Yee, A.J.; Ang, L.C.; He, C.; Shan, S.W.; et al. MicroRNA miR-93 promotes tumor growth and angiogenesis by targeting integrin-beta8. Oncogene 2011, 30, 806–821.
- Mayor-Lynn, K.; Toloubeydokhti, T.; Cruz, A.C.; Chegini, N. Expression profile of microRNAs and mRNAs in human placentas from pregnancies complicated by preeclampsia and preterm labor. Reprod. Sci 2011, 18, 46–56.
- Maccani, M.A.; Padbury, J.F.; Marsit, C.J. miR-16 and miR-21 expression in the placenta is associated with fetal growth. PLoS One 2011, 6, e21210.
- Yang, W.J.; Yang, D.D.; Na, S.; Sandusky, G.E.; Zhang, Q.; Zhao, G. Dicer is required for embryonic angiogenesis during mouse development. J. Biol. Chem 2005, 280, 9330–9335.
- Alpini, G.; Glaser, S.S.; Zhang, J.P.; Francis, H.; Han, Y.; Gong, J.; Stokes, A.; Francis, T.; Hughart, N.; Hubble, L.; et al. Regulation of placenta growth factor by microRNA-125b in hepatocellular cancer. J. Hepatol 2011, 55, 1339–1345.
- Zhang, Y.; Diao, Z.; Su, L.; Sun, H.; Li, R.; Cui, H.; Hu, Y. MicroRNA-155 contributes to preeclampsia by down-regulating CYR61. Am. J. Obstet. Gynecol 2010, 202, 466.e1–466.e7.
- Burton, G.J.; Charnock-Jones, D.S.; Jauniaux, E. Regulation of vascular growth and function in the human placenta. Reproduction 2009, 138.
- Babawale, M.O.; van Noorden, S.; Pignatelli, M.; Stamp, G.W.; Elder, M.G.; Sullivan, M.H. Morphological interactions of human first trimester placental villi co-cultured with decidual explants. Hum. Reprod 1996, 11, 444–450.
- Red-Horse, K.; Kapidzic, M.; Zhou, Y.; Feng, K.T.; Singh, H.; Fisher, S.J. EPHB4 regulates chemokine-evoked trophoblast responses: A mechanism for incorporating the human placenta into the maternal circulation. Development 2005, 132, 4097–4106.
- Zhao, C.; Dong, J.; Jiang, T.; Shi, Z.; Yu, B.; Zhu, Y.; Chen, D.; Xu, J.; Huo, R.; Dai, J. Early second-trimester serum miRNA profiling predicts gestational diabetes mellitus. PLoS One 2011, 6, e23925.
- Higashijima, A.; Miura, K.; Mishima, H.; Kinoshita, A.; Jo, O.; Abe, S.; Hasegawa, Y.; Miura, S.; Yamasaki, K.; Yoshida, A.; et al. Characterization of placenta-specific microRNAs in fetal growth restriction pregnancy. Prenat. Diagn 2013.
- He, L.; Hannon, G.J. MicroRNAs: Small RNAs with a big role in gene regulation. Nat. Rev. Genet 2004, 5, 522–531.
- Caniggia, I.; Mostachfi, H.; Winter, J.; Gassmann, M.; Lye, S.J.; Kuliszewski, M.; Post, M. Hypoxia-inducible factor-1 mediates the biological effects of oxygen on human trophoblast differentiation through TGFbeta(3). J. Clin. Invest 2000, 105, 577–587.
- Pang, R.T.; Leung, C.O.; Ye, T.M.; Liu, W.; Chiu, P.C.; Lam, K.K.; Lee, K.F.; Yeung, W.S. MicroRNA-34a suppresses invasion through downregulation of Notch1 and Jagged1 in cervical carcinoma and choriocarcinoma cells. Carcinogenesis 2010, 31, 1037–1044.
- Zhu, J.; Motejlek, K.; Wang, D.; Zang, K.; Schmidt, A.; Reichardt, L.F. β8 integrins are required for vascular morphogenesis in mouse embryos. Development 2002, 129, 2891–2903.
- Dunk, C.; Petkovic, L.; Baczyk, D.; Rossant, J.; Winterhager, E.; Lye, S. A novel in vitro model of trophoblast-mediated decidual blood vessel remodeling. Lab. Invest 2003, 83, 1821–1828.
- Kotlabova, K.; Doucha, J.; Hromadnikova, I. Placental-specific microRNA in maternal circulation—Identification of appropriate pregnancy-associated microRNAs with diagnostic potential. J. Reprod. Immunol 2011, 89, 185–191.
- Chim, S.S.; Shing, T.K.; Hung, E.C.; Leung, T.Y.; Lau, T.K.; Chiu, R.W.; Lo, Y.M. Detection and characterization of placental microRNAs in maternal plasma. Clin. Chem 2008, 54, 482–490.
- Miura, K.; Miura, S.; Yamasaki, K.; Higashijima, A.; Kinoshita, A.; Yoshiura, K.; Masuzaki, H. Identification of pregnancy-associated microRNAs in maternal plasma. Clin. Chem 2010, 56, 1767–1771.
- Pollheimer, J.; Knofler, M. Signalling pathways regulating the invasive differentiation of human trophoblasts: A review. Placenta 2005, 26, S21–S30.
- Camps, C.; Buffa, F.M.; Colella, S.; Moore, J.; Sotiriou, C.; Sheldon, H.; Harris, A.L.; Gleadle, J.M.; Ragoussis, J. hsa-miR-210 Is induced by hypoxia and is an independent prognostic factor in breast cancer. Clin. Cancer Res 2008, 14, 1340–1348.
- Sibai, B.; Dekker, G.; Kupferminc, M. Pre-eclampsia. Lancet 2005, 365, 785–799.
- Cui, Y.; Wang, W.; Dong, N.; Lou, J.; Srinivasan, D.K.; Cheng, W.; Huang, X.; Liu, M.; Fang, C.; Peng, J.; et al. Role of corin in trophoblast invasion and uterine spiral artery remodelling in pregnancy. Nature 2012, 484, 246–250.
- Morales Prieto, D.M.; Markert, U.R. MicroRNAs in pregnancy. J. Reprod. Immunol 2011, 88, 106–111.
- Keniry, A.; Oxley, D.; Monnier, P.; Kyba, M.; Dandolo, L.; Smits, G.; Reik, W. The H19 lincRNA is a developmental reservoir of miR-675 that suppresses growth and Igf1r. Nat. Cell Biol 2012, 14, 659–665.
- Gilad, S.; Meiri, E.; Yogev, Y.; Benjamin, S.; Lebanony, D.; Yerushalmi, N.; Benjamin, H.; Kushnir, M.; Cholakh, H.; Melamed, N.; et al. Serum microRNAs are promising novel biomarkers. PLoS One 2008, 3, e3148.
- Faye-Petersen, O.M. The placenta in preterm birth. J. Clin. Pathol 2008, 61, 1261–1275.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



