Submitted:
29 July 2024
Posted:
30 July 2024
You are already at the latest version
Abstract
Cryptococcosis is a disease that originates in the lungs and is prevalent among immunosup-pressed individuals. In severe cases, it can lead to meningoencephalitis and even death. Bio-chemical studies have revealed that the capsule of this fungus is primarily composed of glucu-ronoxylomannan (GXM), which constitutes the majority of the capsule, while glucuronoxylo-mannogalactan (GXMGal) is present in smaller amounts. Previous studies have demonstrated that polysaccharide components exhibit different immunomodulatory activities. While GXM primarily exerts an anti-inflammatory effect, GXMGal conversely demonstrates greater pro-inflammatory activity. Myeloid-derived suppressor cells (MDSCs) encompass a heterogeneous population of immature myeloid cells, comprising myeloid progenitor cells and precursors of macrophages, granulocytes, and dendritic cells at various stages. It has been observed that MDSC populations contribute to negatively regulating the immune system in numerous pathol-ogies, such as bacterial and fungal infections, through mechanisms such as the inhibition of T lymphocyte proliferation. Here, we demonstrate that infection with either B3501 or CAP67 strains induced the accumulation of granulocytic precursors of myeloid-derived suppressor cells (MDSCs) in peritoneal and bronchoalveolar cavities. While the MDSCs recruited by the B3501 strain exhibit suppressive action on T lymphocytes, those recruited by the CAP67 strain could not suppress T lymphocyte proliferation. Additionally, we observed the presence of the pro-grammed death ligand PD-L1 in recruited MDSCs, indicating that this may represent a form of immunosuppression in this infection. These findings uncover a mechanism by which capsule polysaccharides from C. neoformans might compromise host immune responses.
Keywords:
Cryptococcus neoformans
; Glucuronoxylomannan (GXM)
; Glucuronoxylomannogalactan (GXMGal)
; Myeloid-Derived Suppressor Cells (MDSC)
; Infection
; Cellular Microbiology
; Microbial Pathogens
1. Introduction
Cryptococcus neoformans is an opportunistic fungus that, during infection, grows in the form of yeast and exhibits tropism for the nervous system [1,2]. Cryptococcosis affects both animals and humans, predominantly immunosuppressed individuals, although there are reports of certain genotypes of C. neoformans infecting immunocompetent patients [3,4]. In humans, C. neoformans is known to cause CNS infections, resulting in meningoencephalitis, a severe complication of the disease [1,5]. The worldwide incidence of cryptococcal meningitis was recently estimated at approximately 220,000 cases annually, with the majority occurring in immunosuppressed individuals [6].
The primary virulence factor among fungi is the polysaccharide capsule, primarily composed of glucuronoxylomannan (approximately 88%), glucuronoxylomannogalactan (around 10%), and mannoproteins (about 2%) [7,8,9]. Once within the host organism, the fungus continually releases the capsule and its constituents, significantly influencing the host's defense mechanisms. Research indicates that the capsule influences leukocyte migration, fungal phagocytosis by macrophages, and neutrophil chemotaxis [7,10,11], consequently compromising the maturation of dendritic cells and activation of T lymphocytes [10,12]. The effects of isolated C. neoformans capsular polysaccharides GXM and GXMGal fractions on the immune system are multifaceted. GXM generally exhibits anti-inflammatory properties, inducing the production of IL-10 [13,14,15], reducing the expression of cell activation molecules such as MHC-II and CD80 in macrophages [9], and inhibiting neutrophil extracellular traps (NET) release [16]. Conversely, GXMGal predominantly exerts pro-inflammatory effects, including increased expression of TNF-α and iNOS [15,17], as well as heightened expression of MHC-II and CD80 molecules, alongside the production of IL-17 and IL-23 [17,18].
Infections caused by other pathogenic fungi such as Aspergillus fumigatus and Candida albicans similarly manipulate the immune cell profile during infection. For instance, signaling through Dectin-1/CARD9 leads to T lymphocyte regulation by a cell population known as myeloid-derived suppressor cells (MDSCs) [19]. Furthermore, it has been described that A. fumigatus inhibits the cytotoxic effect of natural killer (NK) cells through MDSCs [20].
MDSCs represent a group of cells of medullary origin capable of suppressing the immune response, first described in a lung carcinoma model in the 1980s and extensively studied in several cancer models [21,22,23,24,25]. These cells are generated due to the strong stimulus of myelopoiesis, and hematopoiesis given by factors secreted by tumor cells or infectious agents, such as growth factors (VEGF, M-CSF, GM-CSF, G-CSF), cytokines (IFN-γ, IL-1β, IL-10), and lipopolysaccharide (LPS) [26,27]. In this environment of chronic inflammation, cells constantly produced and recruited from the bone marrow exhibit immature morphology, high production of anti-inflammatory cytokines, and reactive oxygen species (ROS) [28].
MDSCs consist of a heterogeneous cell population found in both mice and humans [27]. Until the late 1990s, there were no typical cell markers for MDSCs, but today it is known that, in murine models, they are represented by CD11b+Ly6C+Ly6G- cells (MDSCs of monocytic origin, M-MDSC) and CD11b+Ly6C+Ly6G+ (MDSCs of granulocytic origin, PMN-MDSC) [29,30]. The most important feature of MDSCs is immunosuppression, which is exerted through the inhibition of the immune response and its cellular components. Several studies have demonstrated their ability to inhibit T lymphocytes [31,32,33], alter the homeostasis of regulatory T lymphocytes [34,35], and influence the regulation of NK cells, dendritic cells, and B lymphocytes [27,36,37]. These processes occur through the production of ROS, the up-regulation of inducible nitric oxide synthase (iNOS) the metabolism of L-arginine by arginase, and the production of cytokines that negatively modulate the immune system [38,39,40].
Beyond their role in cancer, MDSCs have emerged as key players in autoimmune diseases [41,42,43] and infections of viral, parasitic, and fungal origin [19,44,45]. Their precise immunosuppressive mechanisms remain elusive but appear to be intricately linked to infection establishment [46]. MDSCs accumulate in tissues during chronic infections [47] and contribute to local immunosuppression, for instance, by producing IL-10 and TGF-β, aiding in M2 macrophage differentiation [48]. Furthermore, these cells are recognized as inducers of immunosuppressive response and local tissue remodeling during cryptococcosis [49].
Additionally, during cryptococcosis, the migration profile of MDSCs to the infection site can be modulated in different ways. For instance, PMN-MDSCs accumulate at the infection site via binding of the fungus to Lectin type C receptors through the Dectin-1 and CARD9 pathways [19]. However, it is also described that β-glucans purified from fungi such as Aspergillus sp. bind to Dectin-1 and restrict the expansion of MDSCs [50]. Therefore, the molecular mechanisms by which pathogenic fungi promote the recruitment of MDSCs still need further characterization.
In this context, the spotlight is on opportunistic diseases like cryptococcosis, where the causative agent C. neoformans can exacerbate clinical conditions of immunosuppression stemming from medical debilitation or pre-existing diseases such as neoplasms, immunodeficiencies, and infections [51,52,53]. Furthermore, it is described that the GXM polysaccharide of C. neoformans tends to reduce the expression of iNOS, an important enzyme in oxidative stress [17]. Cellular oxidative stress reduction by iNOS inactivation exacerbates the clinical manifestations of Cryptococcus infections [49,54]. This oxidative environment is known to counter act the immunosuppressive effect of MDSCs [55], thus, the loss of performance of these cells can indicates possible relationship with the decreased severity of damage in the disease. However, the role of MDSCs is diverse and poorly elucidated during infections, so this work aimed to investigate the participation of myeloid suppressor MDSCs cells during infection caused by the opportunistic fungus C. neoformans.
2. Material and Methods
2.1. Cryptococcus Strains
Cryptoccocus neoformans wild-type (B3501 serotype D) [56] and the GXM deficient (Cap67 serotype D) [57] strains were provided by Dr. Tamara Doering (Department of Molecular Microbiology, Washington University School of Medicine, St Louis, MO, USA) and Dr. Robert Cherniak (Georgia State University, Atlanta, GA, USA), respectively. The cells were cultured in a liquid defined medium (Sabouraud’s medium) at 30 °C with continuous shaking (100 rpm) for 4 days, followed by an additional of 5 days in minimal medium.
2.2. Animals
Female or male Balb/C mice aged between 8 and 10 weeks, weighing between 25g and 30g were used in this study. The Balb/C mice were provided by the Instituto de Veterinária, UFRRJ, RJ, Brasil. The mice were maintained in sterile, grouped cages under standardized conditions of temperature (22-23 ºC) and a 12h light/dark cycle, with ad libitium access to commercial feed and water. The use of the animals in this study was approved by the Ethics Committee on the Use of Animals (CEUA) at UFRJ (Approval No.: 092/21). The mice were sacrificed according to CEUA- approved criteria. All animal procedures were performed in accordance with Animal Research: Reporting of In Vivo Experiments (ARRIVE) guidelines and regulations.
2.3. Anesthesia
A regimen of Ketamine (100 mg/Kg) combined with Xylazine (10 mg/Kg) was used to induce sedation in mice prior to infection. A total of 100 μL of the anesthetic combination was administered to each mouse via peritoneal injection.
2.4. Infection
Lung infection was induced through a small skin incision over the trachea using a 30G insulin syringe, injecting 5x106 fungal cells. Peritoneal infection was induced by injecting 5x105 fungal cells with a 29G syringe. The entire procedure was performed aseptically, and after injection, the skin cut was sutured with a surgical thread.
2.5. Cell Washes
Bronchoalveolar lavage (BAL) was performed by instilling 1 mL of phosphate-buffered saline through an intratracheal catheter. The skin and tissue over the trachea were removed, and a small incision was realized for catheter insertion. The volume was injected and removed with a gentle chest massage five times before complete removal. Peritoneal lavage was performed by injecting 5 ml of DMEM medium (Sigma, Saint Louis, USA) without Fetal Bovine Serum (FBS) into the peritoneum through an abdominal incision. Both lavage fluids were kept on ice until further use.
2.6. Morphological Analysis
Approximately 100 – 200 µL of BAL cells were centrifuged at 800 rpm for 4 minutes using a cytocentrifuge. The slides were then stained using the Panoptic method (Laborcli, Rio de Janeiro, Brazil) for further morphological analysis under optical microscopy.
2.7. Flow Cytometry
Peritoneal lavage and/or BAL samples were washed once with FACS buffer (1x PBS, 0.02% Azide, and 3% FBS). The supernatant was discarded, and the cells resuspended in 300 µl of blocking solution with Fc Block (BD Pharmingen, New Jersey, USA), then incubated on ice for 20 minutes. After centrifugation, the cells were incubated on ice for 30 minutes with anti-Siglec F, anti-CD11c, anti-CD11b, anti-Ly6G, anti-Ly6C and anti-CD274 antibodies. After washing, the cells were analyzed in a flow cytometer (BD LSRFortessa).
2.8. Cell Sorting
Peritoneal lavage from infected animals was performed with DMEM medium supplemented with 0.5% FBS and 1 µg/ml of Amphotericin B. Cells were treated to block nonspecific epitopes and labeled with anti-Ly6G antibody as described in the flow cytometry methodology (Section 2.7). After three steps of washes, cells were resuspended in DMEM medium supplemented with 0.5% FBS and 1 µg/ml of Amphotericin B. The sorting of Ly6G+ cells was realized in a MoFlo High Performance Cell Sorter flow cytometer (Dako Cytomation) obtaining cells with more than 95% purity. Sorted cells were collected in tubes containing DMEM medium supplemented with 10% FBS and kept on ice until further use.
2.9. Cell Proliferation Assay
Ly6G+ cells obtained after cell separation and co-cultured with total T lymphocytes enriched in a nylon wool column. A flat-bottom 96-well polystyrene plate was coated overnight with 1 µg/ml of anti-CD3, followed by plating of T lymphocytes (2x105) and then Ly6G+ cells in different ratios. In some conditions, L-NIL (50 µM), DPI (2 µM) or NAC (100 µM) inhibitors were added to the culture. The cells were cultured at 37°C for 72 hours after adding all components in their respective ratios and conditions. At 48 hours, 5 µCi of 3 H-methyl-thymidine was added to the cell culture. Cell proliferation was monitored by quantifying the radioactivity incorporated into the cellular DNA, measured in a scintillator counter and expressed as counts per minute (cpm).
2.10. Statistical Analysis
Data were analyzed using the analysis of variance (ANOVA) for unpaired samples and by Dunnett's post-test for individual comparisons with the control group. The t-test was applied for paired samples between different groups. All analysis were performed using GraphPad Prism 8.4 software.
3. Results
3.1. Observation of Precursor Cells in Peritoneal and Bronchoalveolar Lavage after Infections with C. neoformans
In experimental settings, cryptococcal infection often initiates primarily within the lungs. However, systemic dissemination can be easily achieved by inoculating the pathogen intraperitoneally in the test animals. This method allows for a swifter and more controlled spread of the infection throughout the organism, facilitating the investigation of systemic effects and the assessment of cell migration profiles. To evaluate the morphological characteristics of cell populations, present in these sites, we performed cytospin and staining with eosin and hematoxylin. Under an optical microscope, we observed that cells from the peritoneal and bronchoalveolar lavage of uninfected mice, treated with phosphate buffer solution (PBS) to mimic the infection, consisted mostly of monocytes (Figure 1). In contrast, cells obtained from infected animals exhibited heterogeneous appearances, varying in maturation stages. Particularly, we observed neutrophils with immature characteristics, characterized by poorly condensed chromatin, circular ring-shaped nuclei, and poorly granulated cytoplasm (Figure 1).
The cellular infiltration observed in both sites can be modulated by either the infection or by the capsular polysaccharides present in the pathogen. However, there are inconsistent descriptions of the immunomodulatory properties of these polysaccharides during experimental in vivo treatment. Therefore, to investigate whether infection with the acapsulated mutant strain CAP67, primarily consisting of GXMGal and mannoproteins, induces a similar recruitment of immature cell profile as the wild-type strain, we infected mice via both the peritoneal and tracheal routes and performed the morphological analyses. We observed that, similar to the wild-type strain, the mutant CAP67 strain induced the recruitment of cells morphologically similar to those recruited by the wild-type strain of C. neoformans (Figure 1).
3.2. Characterization of Granulocytic MDSCs after Infection with C. neoformans
The predominance of immature cells with morphology resembling granulocytes raised the possibility of granulocytic lineage MDSCs cells participating in C. neoformans infection. Therefore, to assess the phenotype of the recruited neutrophil population, we analyzed the cells present at infection sites by flow cytometry, considering that MDSCs have the primary CD11b+GR1+ phenotype [60]. We observed that cells recruited by both the B3501 and CAP67 strains were positive for the CD11b+Ly6G+Ly6C+ phenotype in both bronchoalveolar lavage and peritoneal lavage (Figure 2)
Our data show that cells recruited by both wild-type and mutant strains of C. neoformans exhibit morphological and phenotypic characteristics with MDSCs [40]. However, the definitive evidence for classifying myeloid-derived suppressor cells lies in their ability to suppress the immune system [61]. Thus, to evaluate the effect of granulocytes recruited during infection on regulating T lymphocytes responses, we performed a proliferation assay using total T lymphocytes isolated from the spleen of naive mice stimulated with anti-CD3 antibody. Granulocytes recruited following peritoneal infection with both strains were purified and co-cultured with T lymphocytes in different ratios. The proliferative rate was quantified after 72 hours of culture, using radioactive thymidine incorporated into the cellular DNA. We observed that immature cells recruited by infection with the wild-type B3501 strain exhibited a potent suppressive action on T lymphocytes (Figure 3A), even at low ratios such as 1:40. In contrast, cells recruited by infection with the mutant CAP67 strain were not able to suppress the proliferation of T lymphocytes (Figure 3B). We can, therefore, infer that infection with wild-type C. neoformans was able to recruit MDSCs, while on the other hand, infection by unencapsulated mutant C. neoformans was able to recruit cells like MDSCs.
3.3. The Suppressive Effect Is Not Dependent on the Production of Reactive Oxygen Species
The morphological features of the recruited cell population include a ring-shaped or segmented nucleus. Additionally, the suppressive effect induced by the B3501 strain suggests that C. neoformans is capable of inducing PMN-MDSCs. This subtype of myeloid suppressor cells depresses the immune system preferentially through the production of reactive oxygen species, while the monocytic subtype acts via the nitric oxide pathway [40]. Although our data suggest the predominance of the granulocytic subtype of MDSCs, given the heterogeneous nature of this suppressor population, we used inhibitors targeting both the ROS pathway and the NO pathway in the lymphocyte proliferation assay. The presence of the NADPH oxidase inhibitor Diphenyleneiodonium chloride (DPI) (Figure 4B) and the antioxidant N-acetyl-cysteine (NAC) (Figure 4C), and inducible nitric oxide synthase (iNOS) inhibitor N6-(1-iminoethyl)-L-lysine dihydrochloride (L-NIL) (Figure 4D), did not reverse the suppression of T lymphocytes caused by the recruited myeloid cells. This suggests that the suppression mechanism exerted by PMN-MDSCs is independent of the oxidative burst.
3.4. Recruited MDSCs Show PD-L1 Expression in Bronchoalveolar and Peritoneal Infection
Studies have shown that MDSCs can induce suppression through PD-L1 expression, which serves as a distinguishing marker for this cell population [62]. Therefore, we investigated the presence of PD-L1 on these cells using flow cytometry. Our data show an increased presence of the PD-L1 in cells recruited following intratracheal infection with the B3501 strain. Furthermore, infection with the CAP67 strain also demonstrated PD-L1 expression, suggesting that despite not inhibiting T lymphocytes proliferation, the capsular polysaccharide GXMGal may still exert suppressive effects. Overall, our findings suggest that the suppression pathway utilized by MDSCs recruited during Cryptococcus neoformans infection may involve PD-L1 action (Figure 5).
4. Discussion
Cryptococcus neoformans is an opportunistic fungus and one of the causes of cryptococcosis, a disease characterized as an initial pneumonia that can progress to fatal meningoencephalitis. In recent years, it has become the most commonly diagnosed symptomatic fungal infection and the leading cause of death among immunocompromised patients [12,63]. C. neoformans possesses virulence factors that protect it from hostile environment in its natural habitat and facilitate the establishment of infection.
Among these factors, the capsule is the most extensively studied due to its properties and the significant influence it exerts on the host's immune system [16,17]. The capsule of C. neoformans is a polysaccharide with a heterogeneous structure, primarily composed of glucuronoxylomannan (GXM) and glucuronoxylomannogalactan (GXMGal) as its main components. Polysaccharides form layers around the cell wall, providing increased fluidity and permeability to the outer layers of the capsule, while the inner layers shield the fungal cell from antibodies and components of the immune system [64]. Fungal infections trigger various mechanisms of both innate and adaptive immune responses. It has been observed that the polysaccharide components of the cryptococcal capsule possess a diverse mechanism aimed at modulating the host's immune response. The data indicate that GXM can inhibit phagocytosis, likely due to its polyanionic nature, which results in electrostatic repulsion with other cells. Additionally, GXM can induce the production and secretion of anti-inflammatory cytokines such as IL-10 and TGF-β in macrophages. Monari and colleagues demonstrated that capsular GXM can induce macrophage apoptosis through Fas and Fas-L. Another capsular component, GXMGal, has also been shown to possess immunomodulatory functions, including the ability to induce the production of pro-inflammatory cytokines such as IL-6 and TNF-α, wich in turn leads to the expression of iNOS and subsequent production of NO. These results suggest that GXMGal may have more immunoprotective actions [9,13,65,66]. Although initially studied in cancer, the immunosuppressive functions performed by MDSCs suggest that this cell population may have similar roles in diseases caused by pathogens. While this phenomenon has been observed in bacterial and viral diseases; there are few cases described in fungal diseases [46]. Nevertheless, it has been demonstrated that in diseases caused by the fungus C. albicans, for example, the presence of these cells has a protective effect [19]. Similarly, in other pathological conditions such as autoimmune diseases, MDSCs have shown a beneficial effect, preventing damage caused by an exaggerated immune response [67].
In this study, we observed the recruitment of myeloid-origin suppressor cells during experimental infection with C. neoformans. Cells displaying features such as poorly condensed chromatin, a circular ring-shaped nucleus, and poorly granular cytoplasm were observed in the bronchoalveolar and peritoneal lavage samples from mice infected with both the wild-type strain (B3501) and the hypovirulent strain (CAP67). These cells were later identified by flow cytometry to exhibit a phenotype corresponding to the granulocytic lineage of MDSCs, a population previously observed in infections caused by Aspergillus fumigatus, Candida albicans [19], and Pneumocystis jirovecii [53]. This recruitment occurs rapidly after infection, indicating that the suppressive effects of this cell population may contribute to infection’s progression. This suggests that, like other fungal infections, C. neoformans can recruit these cells to infection sites, where they potentially influence the infection outcome. Since their discovery MDSCs are recognized for their immunosuppressive activities in cancer, infections, and autoimmune diseases, primarily targeting T cells.
The main factors associated with immunosuppression include iNOS, TGF-β, IL-10, among others. However, in recent years, it has been observed that the mechanisms vary according to the subpopulation of MDSCs [68]. Monocytic MDSCs have their function associated via nitric oxide (NO) and cytokines, thereby inhibiting T cell response. On the other hand, granulocytic MDSCs have their function via reactive oxygen species (ROS) and Arginase-1 (Arg-1) and these suppressive activities involve the induction of tolerance to specific antigens of T lymphocytes [69]. In models of C. albicans infection, for example, it has been observed that recruited MDSCs have influence the maturation of NK cells and induction of Th17 responses, leading to host protection. [19]. Conversely, in infection with the opportunistic fungus Pneumocystis jirovecii, T cell inhibition has been observed [53]. Our data demonstrate that MDSCs recruited by peritoneal infection with the B3501 strain can inhibit the proliferation of T cells purified from mice and activated with anti-CD3. However, the addition of NADPH inhibitors (DPI), antioxidant N-acetyl-cysteine (NAC) and iNOS inhibitor (L-NIL), did not reverse this proliferation inhibition, indicating a mechanism independent of ROS. On the other hand, when the infection occurs with the acapsulated strain CAP67, inhibition of T lymphocytes is not observed, suggesting that the capsular component GalXM alone is not able to induce the recruitment of this cell population. It was also observed that, although both strains and capsular components can induce the recruitment of MDSCs with a granulocytic phenotype to the infection site, only the presence of GXM is able of activate the immunosuppressive functions of this cell. Therefore, it is necessary to investigate other mechanisms that induce immunosuppression in this model of infection.
Studies have indicated that during experimental infection with P. jirovecii, alveolar macrophages start expressing the programmed death receptor PD-1, while MDSCs express its ligand, PD-L1. This recently identified suppression mechanism contributes to the diminished immune response and facilitates the progression of the disease caused by this fungus [62]. Additionally, we observed the presence of the PD-L1 ligand in cells recruited during infection with both the B3501 and CAP67 strains, suggesting that this may be the suppression mechanism utilized in this instance.
It is important to mention the small discrepancies found in the error bars of the results presented. However, this is easily explained by the fact that we are evaluating the results in animal models, where variation between results can occur.
Our data suggest that the presence of MDSCs may be a critical factor in inhibiting the immune response, resulting in inadequate disease control and the subsequent spread of fungal cells throughout the body.
In the context of cancer immunotherapy, several treatments are currently being studied and applied to patients, including those based on type I interferon [70]. Evidence indicates that activation of this pathway can induce changes in the tumor microenvironment, reducing immunosuppression and enhancing the efficacy of anti-cancer drugs. Studies in both human and murine models have demonstrated that treatment with anti-colony-stimulating factor-1 receptor (anti-CSF-1R) can deplete tumor-associated macrophages, thereby preventing the accumulation of MDSCs [71]. Therefore, it is plausible that targeting these cells during infection could serve as an effective therapeutic strategy for cryptococcosis.
5. Conclusions
In this study, we observed that C. neoformans can induce the recruitment and accumulation of immature cells known as MDSCs. These cells, when recruited by virulent strains containing the capsular polysaccharide GXM, exhibit immunosuppressive activities on T lymphocytes activation. The success of infection by this fungus depends not only on the immunomodulatory and immunosuppressive properties of its polysaccharide capsule but also on the suppressive capacity of the individual's cellular components.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Gate strategy used for MDSCs cells in intratracheal infection. Figure S2: Gate strategy used for MDSCs cells in peritoneal infection.
Author Contributions
Conceptualization, J.C.G.O, I.D.L, I.M.F.S and E.B.S.J.; methodology, J.C.G.O, L.M.F and L.F.L.; formal analysis, J.C.G.O, H.L.M.G. L.M.P and J.O.P.; investigation, C.G.F.L, D.D.R and J.C.G.O.; resources, C.G.F.L, LP.C. and A.M.; writing—original draft preparation, J.C.G.O, L.P.C, D.D.R and C.G.F.L.; writing—review and editing, C.G.F.L, L.M.F and D.D.R.; supervision, L.F.L, H.L.M.G, A.M, D.D.R and C.G.F.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Brazilian National Research Council (CNPq), Rio de Janeiro State Science Foundation (FAPERJ), and Fundação Oswaldo Cruz (FIOCRUZ).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data are contained within this article.
Acknowledgments
We thank Renata Sant’Anna Rezende Ribeiro and Lindomar Miranda for their helpful technical assistance.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Husain, S.; Wagener, M.M.; Singh, N. Cryptococcus neoformans Infection in Organ Transplant Recipients: Variables Influencing Clinical Characteristics and Outcome. Emerg Infect Dis. 2001, 7, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Heitman, J. The biology of the Cryptococcus neoformans species complex. Annu Rev Microbiol. 2006, 60, 69–105. [Google Scholar] [CrossRef] [PubMed]
- Chau, T.T.; Mai, N.H.; Phu, N.H.; Nghia, H.D.; Chuong, L.V.; Sinh, D.X.; Duong, V.A.; Diep, P.T.; Campbell, J.I.; Baker, S.; et al. A prospective descriptive study of cryptococcal meningitis in HIV uninfected patients in Vietnam - high prevalence of Cryptococcus neoformans var grubii in the absence of underlying disease. BMC Infect Dis 2010, 10. [Google Scholar] [CrossRef] [PubMed]
- Kwon-Chung, K.J.; Fraser, J.A.; Doering, T.L.; Wang, Z.; Janbon, G.; Idnurm, A.; Bahn, Y.S. Cryptococcus neoformans and Cryptococcus gattii, the etiologic agents of cryptococcosis. Cold Spring Harb Perspect Med. 2015, 4, 7. [Google Scholar] [CrossRef]
- Firacative, C.; Lizarazo, J.; Illnait-Zaragozí, M.T.; Castañeda, E. The status of cryptococcosis in latin America. Mem Inst Oswaldo Cruz. 2018, 113, 7. [Google Scholar] [CrossRef] [PubMed]
- Rajasingham, R.; Smith, R.M.; Park, B.J.; Jarvis, J.N.; Govender, N.P.; Chiller, T.M.; Denning, D.W.; Loyse, A.; Boulware, D.R. Global burden of disease of HIV-associated cryptococcal meningitis: an updated analysis. Lancet Infect Dis. 2017, 17, 873–881. [Google Scholar] [CrossRef] [PubMed]
- Cherniak, R.; Sundstrom, J.B. Polysaccharide antigens of the capsule of Cryptococcus neoformans. Infect Immun. 1994, 62, 1507–1512. [Google Scholar] [CrossRef] [PubMed]
- Decote-Ricardo, D.; LaRocque-de-Freitas, I.F.; Rocha, J.D.B.; Nascimento, D.O.; Nunes, M.P.; Morrot, A.; Freire-de-Lima, L.; Previato, J.O.; Mendonça-Previato, L.; Freire-de-Lima, C.G. Immunomodulatory Role of Capsular Polysaccharides Constituents of Cryptococcus neoformans. Front Med 2019, 6. [Google Scholar] [CrossRef] [PubMed]
- Zaragoza, O.; Rodrigues, M.L.; De Jesus, M.; Frases, S.; Dadachova, E.; Casadevall, A. The Capsule of the Fungal Pathogen Cryptococcus neoformans. Adv Appl Microbiol. 2009, 68, 133–216. [Google Scholar]
- Maziarz, E.K.; Perfect, J.R. Cryptococcosis. Infect Dis Clin North Am. 2016, 30, 179–206. [Google Scholar] [CrossRef]
- Zhao, M.D.; Murphy, J.W. Effects of the two varieties of Cryptococcus neoformans cells and culture filtrate antigens on neutrophil locomotion. Infect Immun. 1995, 63, 2632–2644. [Google Scholar]
- Li, S.S.; Mody, C.H. Cryptococcus. Proc Am Thorac Soc. 2010, 7, 186–196. [Google Scholar] [CrossRef] [PubMed]
- Retini, C.; Vecchiarelli, A.; Monari, C.; Bistoni, F.; Kozel, T.R. Encapsulation of Cryptococcus neoformans with glucuronoxylomannan inhibits the antigen-presenting capacity of monocytes. Infect Immun. 1998, 66, 664–669. [Google Scholar] [CrossRef] [PubMed]
- Syme, R.M.; Bruno, T.F.; Kozel, T.R.; Mody, C.H. The capsule of Cryptococcus neoformans reduces T-lymphocyte proliferation by reducing phagocytosis, which can be restored with anticapsular antibody. Infect Immun. 1999, 67, 4620–4627. [Google Scholar] [CrossRef] [PubMed]
- Walenkamp, A.M.; Chaka, W.S.; Verheul, A.F.; Vaishnav, V.V.; Cherniak, R.; Coenjaerts, F.E.; Hoepelman, I.M. Cryptococcus neoformans and its cell wall components induce similar cytokine profiles in human peripheral blood mononuclear cells despite differences in structure. FEMS Immunol Med Microbiol. 1999, 26, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Rocha, J.D.; Nascimento, M.T.; Decote-Ricardo, D.; Côrte-Real, S.; Morrot, A.; Heise, N.; Nunes, M.P.; Previato, J.O.; Mendonça-Previato, L.; DosReis, G.A. , et al. Capsular polysaccharides from Cryptococcus neoformans modulate production of neutrophil extracellular traps (NETs) by human neutrophils. Sci Rep. 2015, 5, 8008. [Google Scholar] [CrossRef] [PubMed]
- Villena, S.N.; Pinheiro, R.O.; Pinheiro, C.S.; Nunes, M.P.; Takiya, C.M.; DosReis, G.A.; Previato, J.O.; Mendonça-Previato, L.; Freire-de-Lima, C.G. Capsular polysaccharides galactoxylomannan and glucuronoxylomannan from Cryptococcus neoformans induce macrophage apoptosis mediated by Fas ligand. Cell Microbiol. 2008, 10, 1274–1285. [Google Scholar] [CrossRef] [PubMed]
- LaRocque-de-Freitas, I.F.; Rocha, J.D.B.; Nunes, M.P.; Oliveira, P.A.V.; Nascimento, D.O.; Freire-de-Lima, L.; Takiya, C.M.; Morrot, A.; Decote-Ricardo, D.; Previato, J.O. , et al. Involvement of the capsular GalXM-induced IL-17 cytokine in the control of Cryptococcus neoformans infection. Sci Rep. 2018, 8, 1. [Google Scholar]
- Rieber, N.; Singh, A.; Öz, H.; Carevic, M.; Bouzani, M.; Amich, J.; Ost, M.; Ye, Z.; Ballbach, M.; Schäfer, I. , et al. Pathogenic fungi regulate immunity by inducing neutrophilic myeloid-derived suppressor cells. Cell Host Microbe. 2015, 17, 507–514. [Google Scholar] [CrossRef]
- Mueller-Leisse, J.; Brueggemann, S.; Bouzani, M.; Schmitt, A.L.; Einsele, H.; Loeffler, J. Polymorphonuclear neutrophils and granulocytic myeloid-derived suppressor cells inhibit natural killer cell activity toward Aspergillus fumigatus. Med Mycol. 2015, 53, 622–629. [Google Scholar] [CrossRef]
- Chen, M.F.; Kuan, F.C.; Yen, T.C.; Lu, M.S.; Lin, P.Y.; Chung, Y.H.; Chen, W.C.; Lee, K.D. IL-6-stimulated CD11b+CD14+HLA-DR- myeloid-derived suppressor cells, are associated with progression and poor prognosis in squamous cell carcinoma of the esophagus. Oncotarget. 2014, 5, 8716–8728. [Google Scholar] [CrossRef] [PubMed]
- Gabrilovich, D.I.; Bronte, V.; Chen, S.H.; Colombo, M.P.; Ochoa, A.; Ostrand-Rosenberg, S.; Schreiber, H. The terminology issue for myeloid-derived suppressor cells [1]. Cancer Res. 2007, 67, 425. [Google Scholar] [CrossRef] [PubMed]
- Jessup, J.M.; Le Grue, S.J.; Kahan, B.D.; Pellis, N.R. Induction of suppressor cells by a tumor-derived suppressor factor. Cell Immunol. 1985, 93, 9–25. [Google Scholar] [CrossRef] [PubMed]
- Young, M.R.; Endicott, R.A.; Duffie, G.P.; Wepsic, H.T. Suppressor alveolar macrophages in mice bearing metastatic Lewis lung carcinoma tumors. J Leukoc Biol. 1987, 42, 682–688. [Google Scholar] [CrossRef] [PubMed]
- Young, M.R.; Wheeler, E.; Newby, M. Macrophage-mediated suppression of natural killer cell activity in mice bearing Lewis lung carcinoma. J Natl Cancer Inst. 1986, 76, 745–750. [Google Scholar] [CrossRef]
- Talmadge, J.E.; Gabrilovich, D.I. History of myeloid-derived suppressor cells. Nat Rev Cancer. 2013, 13, 739–752. [Google Scholar] [CrossRef]
- Zhao, Y.Y.; Wu, T.; Shao, S.; Shi, B.; Zhao, Y.Y. Phenotype, development, and biological function of myeloid-derived suppressor cells. Oncoimmunology. 2016, 5, 2. [Google Scholar] [CrossRef] [PubMed]
- Bronte, V.; Brandau, S.; Chen, S.H.; Colombo, M.P.; Frey, A.B.; Greten, T.F.; Mandruzzato, S.; Murray, P.J.; Ochoa, A.; Ostrand-Rosenberg, S.; et al. Recommendations for myeloid-derived suppressor cell nomenclature and characterization standards. Nat Commun 2016, 7. [Google Scholar] [CrossRef]
- Bronte, V.; Wang, M.; Overwijk, W.W.; Surman, D.R.; Pericle, F.; Rosenberg, S.A.; Restifo, N.P. Apoptotic death of CD8+ T lymphocytes after immunization: induction of a suppressive population of Mac-1+/Gr-1+ cells. J Immunol. 1998, 161, 5313–5320. [Google Scholar] [CrossRef]
- Youn, J.-I.; Nagaraj, S.; Collazo, M.; Gabrilovich, D.I. Subsets of Myeloid-Derived Suppressor Cells in Tumor-Bearing Mice. J Immunol. 2008, 181, 5791–5802. [Google Scholar] [CrossRef]
- Dugast, A.S.; Haudebourg, T.; Coulon, F.; Heslan, M.; Haspot, F.; Poirier, N.; Vuillefroy de Silly, R.; Usal, C.; Smit, H.; Martinet, B.; et al. Myeloid-Derived Suppressor Cells Accumulate in Kidney Allograft Tolerance and Specifically Suppress Effector T Cell Expansion. J Immunol. 2008, 180, 7898–7906. [Google Scholar] [CrossRef]
- Guan, Q.; Blankstein, A.R.; Anjos, K.; Synova, O.; Tulloch, M.; Giftakis, A.; Yang, B.; Lambert, P.; Peng, Z.; Cuvelier, G.D. , et al. Functional Myeloid-Derived Suppressor Cell Subsets Recover Rapidly after Allogeneic Hematopoietic Stem/Progenitor Cell Transplantation. Biol Blood Marrow Transplant. 2015, 21, 1205–1214. [Google Scholar] [CrossRef] [PubMed]
- Movahedi, K.; Guilliams, M.; Van den Bossche, J.; Van den Bergh, R.; Gysemans, C.; Beschin, A.; De Baetselier, P.; Van Ginderachter, J.A. Identification of discrete tumor-induced myeloid-derived suppressor cell subpopulations with distinct T cell suppressive activity. Blood. 2008, 111, 4233–4244. [Google Scholar] [CrossRef]
- Deshane, J.; Zmijewski, J.W.; Luther, R.; Gaggar, A.; Deshane, R.; Lai, J.F.; Xu, X.; Spell, M.; Estell, K.; Weaver, C.T.; et al. Free radical-producing myeloid-derived regulatory cells: Potent activators and suppressors of lung inflammation and airway hyperresponsiveness. Mucosal Immunol. 2011, 4, 503–518. [Google Scholar] [CrossRef]
- Soler, D.C.; Young, A.B.; Fiessinger, L.; Galimberti, F.; Debanne, S.; Groft, S.; McCormick, T.S.; Cooper, K.D. Increased, but Functionally Impaired, CD14+ HLA-DR–/low Myeloid-Derived Suppressor Cells in Psoriasis: A Mechanism of Dysregulated T Cells. J Invest Dermatol. 2016, 136, 798–808. [Google Scholar] [CrossRef]
- Greifenberg, V.; Ribechini, E.; Rößner, S.; Lutz, M.B. Myeloid-derived suppressor cell activation by combined LPS and IFN-γ treatment impairs DC development. Eur J Immunol. 2009, 39, 2865–2876. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Han, Y.; Guo, Q.; Zhang, M.; Cao, X. Cancer-Expanded Myeloid-Derived Suppressor Cells Induce Anergy of NK Cells through Membrane-Bound TGF-β1. J Immunol. 2009, 182, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Bruchard, M.; Ghiringhelli, F. Microenvironnement tumoral. Medecine/Sciences. 2014, 30, 429–435. [Google Scholar] [CrossRef]
- Lim, H.X.; Hong, H.-J.; Cho, D.; Kim, T.S. IL-18 Enhances Immunosuppressive Responses by Promoting Differentiation into Monocytic Myeloid-Derived Suppressor Cells. J Immunol. 2014, 193, 5453–5460. [Google Scholar] [CrossRef]
- Youn, J.I.; Gabrilovich, D.I. The biology of myeloid-derived suppressor cells: The blessing and the curse of morphological and functional heterogeneity. Eur J Immunol. 2010, 40, 2969–2975. [Google Scholar] [CrossRef]
- Egelston, C.; Kurkó, J.; Besenyei, T.; Tryniszewska, B.; Rauch, T.A.; Glant, T.T.; Mikecz, K. Suppression of dendritic cell maturation and T cell proliferation by synovial fluid myeloid cells from mice with autoimmune arthritis. Arthritis Rheum. 2012, 64, 3179–3188. [Google Scholar] [CrossRef] [PubMed]
- Kurkó, J.; Vida, A.; Glant, T.T.; Scanzello, C.R.; Katz, R.S.; Nair, A.; Szekanecz, Z.; Mikecz, K. Identification of myeloid-derived suppressor cells in the synovial fluid of patients with rheumatoid arthritis: A pilot study. BMC Musculoskelet Disord. 2014, 15, 1. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.L.; Luan, B.; Wang, X.F.; Qiao, J.Y.; Song, L.; Lei, R.R.; Gao, W.X.; Liu, Y. Peripheral Blood MDSCs, IL-10 and IL-12 in Children with Asthma and Their Importance in Asthma Development. PLoS One. 2013, 8, 5. [Google Scholar] [CrossRef]
- Pan, W.; Zhou, H.J.; Shen, Y.J.; Wang, Y.; Xu, Y.X.; Hu, Y.; Jiang, Y.Y.; Yuan, Z.Y.; Ugwu, C.E.; Cao, J.P. Surveillance on the Status of Immune Cells after Echinnococcus granulosus Protoscoleces Infection in Balb/c Mice. PLoS One. 2013, 8, 3. [Google Scholar] [CrossRef] [PubMed]
- Vollbrecht, T.; Stirner, R.; Tufman, A.; Roider, J.; Huber, R.M.; Bogner, J.R.; Lechner, A.; Bourquin, C.; Draenert, R. Chronic progressive HIV-1 infection is associated with elevated levels of myeloid-derived suppressor cells. AIDS. 2012, 26, 12. [Google Scholar] [CrossRef] [PubMed]
- Tamadaho, R.S.E.; Hoerauf, A.; Layland, L.E. Immunomodulatory effects of myeloid-derived suppressor cells in diseases: Role in cancer and infections. Immunobiology. 2018, 223, 432–442 [PubMed]. [Google Scholar] [CrossRef] [PubMed]
- Dorhoi, A.; Plessis, N.D. . Monocytic myeloid-derived suppressor cells in chronic infections. Front Immunol. 2018, 8, 1895. [Google Scholar] [CrossRef] [PubMed]
- Salminen, A.; Kaarniranta, K.; Kauppinen, A. The role of myeloid-derived suppressor cells (MDSC) in the inflammaging process. Ageing Res Rev. 2018, 48, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Oliveira-Brito, P.K.M.; Rezende, C.P.; Almeida, F.; Roque-Barreira, M.C.; da Silva, T.A. iNOS/Arginase-1 expression in the pulmonary tissue over time during Cryptococcus gattii infection. Innate Immun. 2020, 26, 117–129. [Google Scholar] [CrossRef]
- Sander, L.E.; Sackett, S.D.; Dierssen, U.; Beraza, N.; Linke, R.P.; Müller, M.; Blander, J.M.; Tacke, F.; Trautwein, C. Hepatic acute-phase proteins control innate immune responses during infection by promoting myeloid-derived suppressor cell function. J Exp Med. 2010, 207, 1453–1464. [Google Scholar] [CrossRef]
- Perfect, J.R.; Dismukes, W.E.; Dromer, F.; Goldman, D.L.; Graybill, J.R.; Hamill, R.J.; Harrison, T.S.; Larsen, R.A.; Lortholary, O.; Nguyen, M.H. , et al. Clinical practice guidelines for the management of cryptococcal disease: 2010 update by the infectious diseases society of America. Clin Infect Dis. 2010, 50, 291–322. [Google Scholar] [CrossRef] [PubMed]
- Perfect, J.R.; Casadevall, A. Cryptococcosis. Infect Dis Clin North Am. 2002, 16, 837–874. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Lei, G.S.; Shao, S.; Jung, H.W.; Durant, P.J.; Lee, C.H. Accumulation of myeloid-derived suppressor cells in the lungs during Pneumocystis pneumonia. Infect Immun. 2012, 80, 3634–3641. [Google Scholar] [CrossRef] [PubMed]
- Rossi, G.R.; Cervi, L.A.; García, M.M.; Chiapello, L.S.; Sastre, D.A.; Masih, D.T. Involvement of nitric oxide in protecting mechanism during experimental cryptococcosis. Clin Immunol. 1999, 90, 256–265. [Google Scholar] [CrossRef] [PubMed]
- Ohl, K.; Tenbrock, K. Reactive Oxygen Species as Regulators of MDSC-Mediated Immune Suppression. Front Immunol. 2018, 9, 2499 [PubMed]. [Google Scholar] [CrossRef]
- Kwon-Chung, K.J.; Wickes, B.L.; Stockman, L.; Roberts, G.D.; Ellis, D.; Howard, D.H. Virulence, serotype, and molecular characteristics of environmental strains of Cryptococcus neoformans var. gattii. Infect Immun. 1992, 60, 1869–1874. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, E.S.; Ayers, D.J.; Harrell, A.C.; Nicholas, C.C. Genetic and phenotypic characterization of capsule mutants of Cryptococcus neoformans. J Bacteriol. 1982, 150, 1292–1296. [Google Scholar] [CrossRef] [PubMed]
- Chaka, W.; Verheul, A.F.; Vaishnav, V.V.; Cherniak, R.; Scharringa, J.; Verhoef, J.; Snippe, H.; Hoepelman, I.M. Cryptococcus neoformans and cryptococcal glucuronoxylomannan, galactoxylomannan, and mannoprotein induce different levels of tumor necrosis factor alpha in human peripheral blood mononuclear cells. Infect Immun. 1997, 65, 272–278. [Google Scholar] [CrossRef]
- James, P.G.; Cherniak, R. Galactoxylomannans of Cryptococcus neoformans. Infect Immun. 1992, 60, 1084–1088. [Google Scholar] [CrossRef]
- Ribechini, E.; Greifenberg, V.; Sandwick, S.; Lutz, M.B. Subsets, expansion and activation of myeloid-derived suppressor cells. Med Microbiol Immunol. 2010, 199, 273–281. [Google Scholar] [CrossRef]
- Gabrilovich, D.I.; Nagaraj, S. Myeloid-derived suppressor cells as regulators of the immune system. Nat Rev Immunol. 2009, 9, 162–174. [Google Scholar] [CrossRef] [PubMed]
- Yaseen, M.M.; Abuharfeil, N.M.; Darmani, H.; Daoud, A. Mechanisms of immune suppression by myeloid-derived suppressor cells: The role of interleukin-10 as a key immunoregulatory cytokine. Open Biol. 2020, 10, 9. [Google Scholar] [CrossRef] [PubMed]
- Lester, S.J.; Malik, R.; Bartlett, K.H.; Duncan, C.G. Cryptococcosis: Update and emergence of Cryptococcus gattii. Vet Clin Pathol. 2011, 40, 4–17. [Google Scholar] [CrossRef] [PubMed]
- O'Meara, T.R.; Alspaugh, J.A. The Cryptococcus neoformans capsule: a sword and a shield. Clin Microbiol Rev. 2012, 25, 387–408. [Google Scholar] [CrossRef] [PubMed]
- Bürgel, P.H.; Marina, C.L.; Saavedra, P.H.V.; Albuquerque, P.; de Oliveira, S.A.M.; Veloso Janior, P.H.H.; de Castro, R.A.; Heyman, H.M.; Coelho, C.; Cordero, R.J.B. , et al. Cryptococcus neoformans Secretes Small Molecules That Inhibit IL-1 β Inflammasome-Dependent Secretion. Mediators Inflamm. 2020, 2020, 3412763 [PubMed]. [Google Scholar] [CrossRef]
- Srikanta, D.; Santiago-Tirado, F.H.; Doering, T.L. Cryptococcus neoformans: Historical curiosity to modern pathogen. Yeast. 2014, 31, 47–60. [Google Scholar] [CrossRef] [PubMed]
- Pawelec, G.; Verschoor, C.P.; Ostrand-Rosenberg, S. Myeloid-derived suppressor cells: Not only in tumor immunity. Front Immunol. 2019, 10, 1099. [Google Scholar] [CrossRef]
- Gabrilovich, D.I. Myeloid-derived suppressor cells. Cancer Immunol Res. 2017, 5, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Raber, P.L.; Thevenot, P.; Sierra, R.; Wyczechowska, D.; Halle, D.; Ramirez, M.E.; Ochoa, A.C.; Fletcher, M.; Velasco, C.; Wilk, A. , et al. Subpopulations of myeloid-derived suppressor cells impair T cell responses through independent nitric oxide-related pathways. Int J Cancer. 2014, 134, 2853–2864. [Google Scholar] [CrossRef]
- Li, K.; Shi, H.; Zhang, B.; Xuejin, O.; Qizhi, M.; Yue, C.; Pei, S.; Dan, L.; Yongsheng, W. Myeloid-derived suppressor cells as immunosuppressive regulators and therapeutic targets in cancer. Sig Transduct Target Ther 6. 2021, 6, 1–6. [Google Scholar] [CrossRef]
- Yu, R.; Bo, Z.; Degao, C. Type I Interferon-Mediated Tumor Immunity and Its Role in Immunotherapy. Cellular and Molecular Life Sciences 2022. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Morphology of peritoneal and bronchoalveolar lavage cells after infection with C. neoformans. Animals were infected with the strain of C. neoformans B3501, CAP67 (5x106) or received only PBS. After a period of (A) 4h or (B) 8h, the animals were euthanized, and the peritoneal lavage was collected. (C) Bronchoalveolar lavage was collected after a period of 48h. Both washes were centrifuged using a cytocentrifuge and later stained by the Panoptic method. The morphology of the washed cells was observed under an optical microscope at 400x magnification. Representative results from three independent experiments.
Figure 1.
Morphology of peritoneal and bronchoalveolar lavage cells after infection with C. neoformans. Animals were infected with the strain of C. neoformans B3501, CAP67 (5x106) or received only PBS. After a period of (A) 4h or (B) 8h, the animals were euthanized, and the peritoneal lavage was collected. (C) Bronchoalveolar lavage was collected after a period of 48h. Both washes were centrifuged using a cytocentrifuge and later stained by the Panoptic method. The morphology of the washed cells was observed under an optical microscope at 400x magnification. Representative results from three independent experiments.
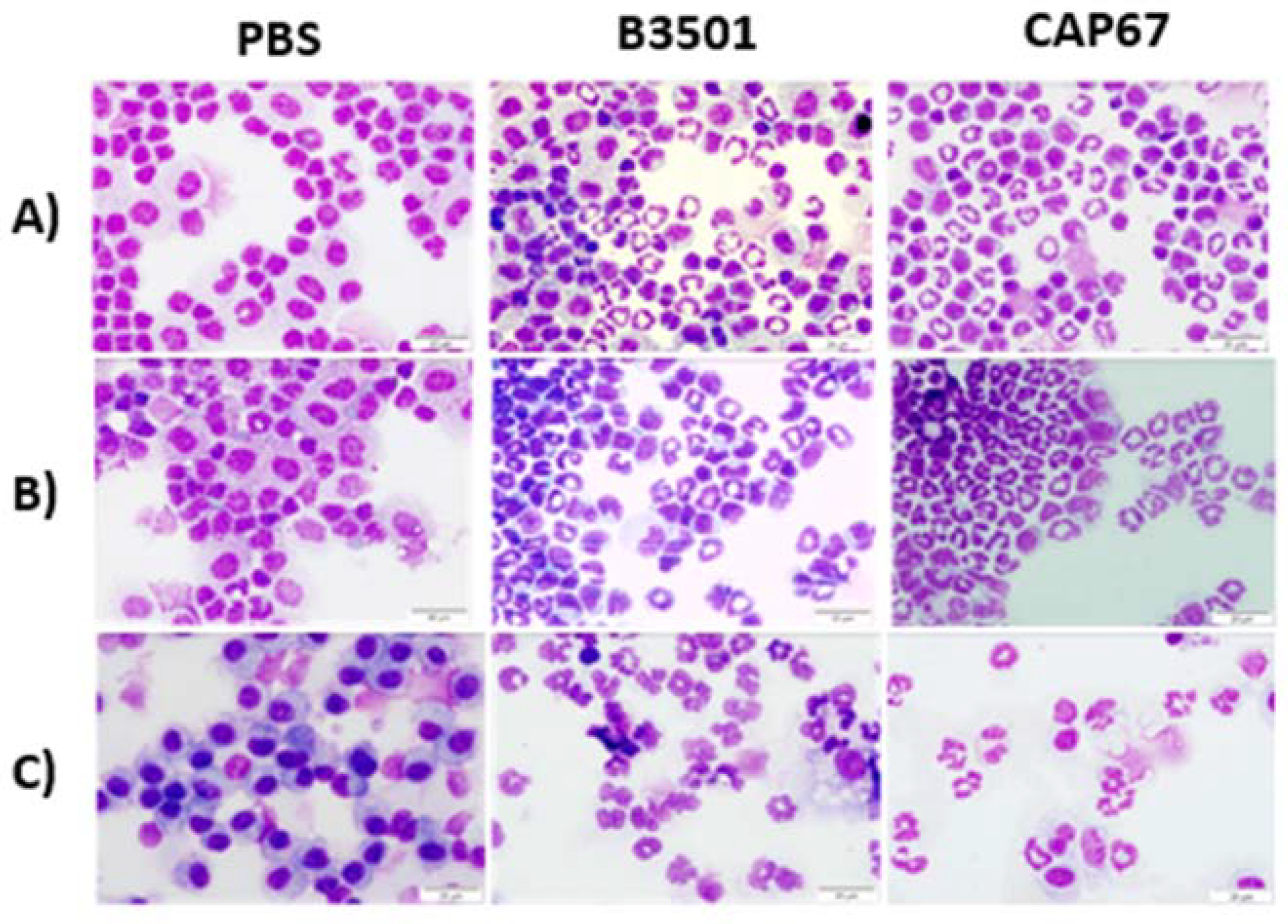
Figure 2.
Frequency of immature neutrophils in bronchoalveolar and peritoneal lavage. Percentage of neu-trophils in the bronchoalveolar lavage sample after 48 hours (A) or 21 days (B) of sublethal infec-tion of 5 x 105 fungal B3501 or CAP67 cells in the lung. Percentage of neutrophils in the peritoneal lavage sample after 4 hours (C) or 8 hours (D) of sublethal infection of 5 x 105 fungal B3501 or CAP67 cells in the peritoneum. Results for each group are represented as mean ± standard error. The results presented are representative of two experiments performed separately. (*) p>0.1, (***) p>0.001, (****) p>0.0001.
Figure 2.
Frequency of immature neutrophils in bronchoalveolar and peritoneal lavage. Percentage of neu-trophils in the bronchoalveolar lavage sample after 48 hours (A) or 21 days (B) of sublethal infec-tion of 5 x 105 fungal B3501 or CAP67 cells in the lung. Percentage of neutrophils in the peritoneal lavage sample after 4 hours (C) or 8 hours (D) of sublethal infection of 5 x 105 fungal B3501 or CAP67 cells in the peritoneum. Results for each group are represented as mean ± standard error. The results presented are representative of two experiments performed separately. (*) p>0.1, (***) p>0.001, (****) p>0.0001.
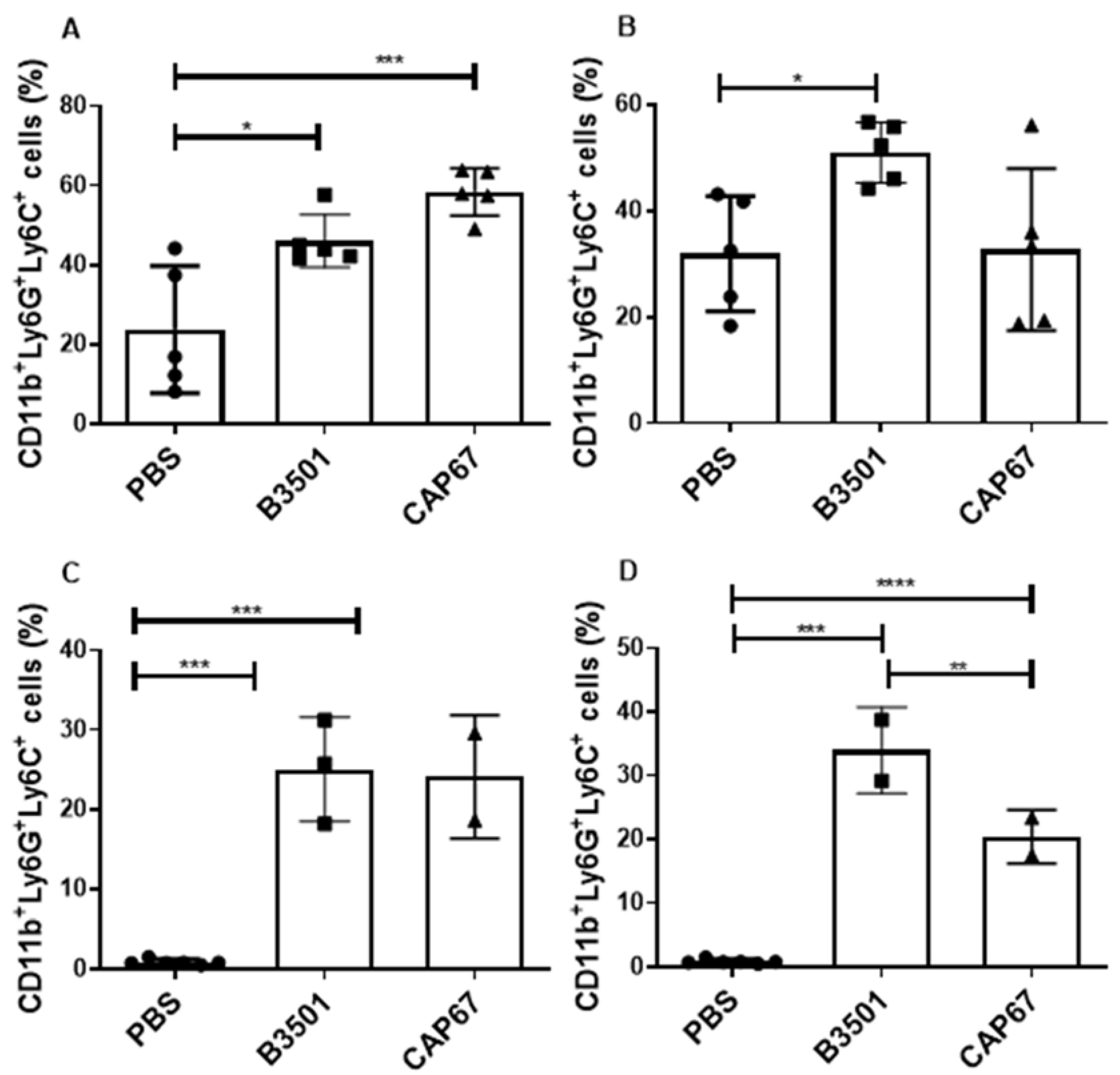
Figure 3.
Functional characterization of recruited neutrophils. Nylon wool column-enriched T lymphocytes were cultured with neutrophils purified from the peritoneal lavage of animals infected with strain B3501 and strain CAP67 at 4-6 hours of infection. The ratio between neutrophils and lymphocytes in culture was 1 neutrophil to 5 lymphocytes (1N:5T), 1 neutrophil to 10 lymphocytes (1N:10T), 1 neutrophil to 20 lymphocytes (1N:20T), 1 neutrophil to 40 lymphocytes (1N :40T) and 1 neutro-phil for 80 lymphocytes (1N:80T). The control column refers to lymphocytes stimulated with a-CD3 in the absence of neutrophils. (A) neutrophils recruited by strain B3501 show a potent sup-pressive action on T lymphocytes. (B) granulocytes recruited by strain CAP67 do not suppress T lymphocytes. The results presented are representative of two experiments performed separately (**) p>0.01, (***) p>0.001.
Figure 3.
Functional characterization of recruited neutrophils. Nylon wool column-enriched T lymphocytes were cultured with neutrophils purified from the peritoneal lavage of animals infected with strain B3501 and strain CAP67 at 4-6 hours of infection. The ratio between neutrophils and lymphocytes in culture was 1 neutrophil to 5 lymphocytes (1N:5T), 1 neutrophil to 10 lymphocytes (1N:10T), 1 neutrophil to 20 lymphocytes (1N:20T), 1 neutrophil to 40 lymphocytes (1N :40T) and 1 neutro-phil for 80 lymphocytes (1N:80T). The control column refers to lymphocytes stimulated with a-CD3 in the absence of neutrophils. (A) neutrophils recruited by strain B3501 show a potent sup-pressive action on T lymphocytes. (B) granulocytes recruited by strain CAP67 do not suppress T lymphocytes. The results presented are representative of two experiments performed separately (**) p>0.01, (***) p>0.001.
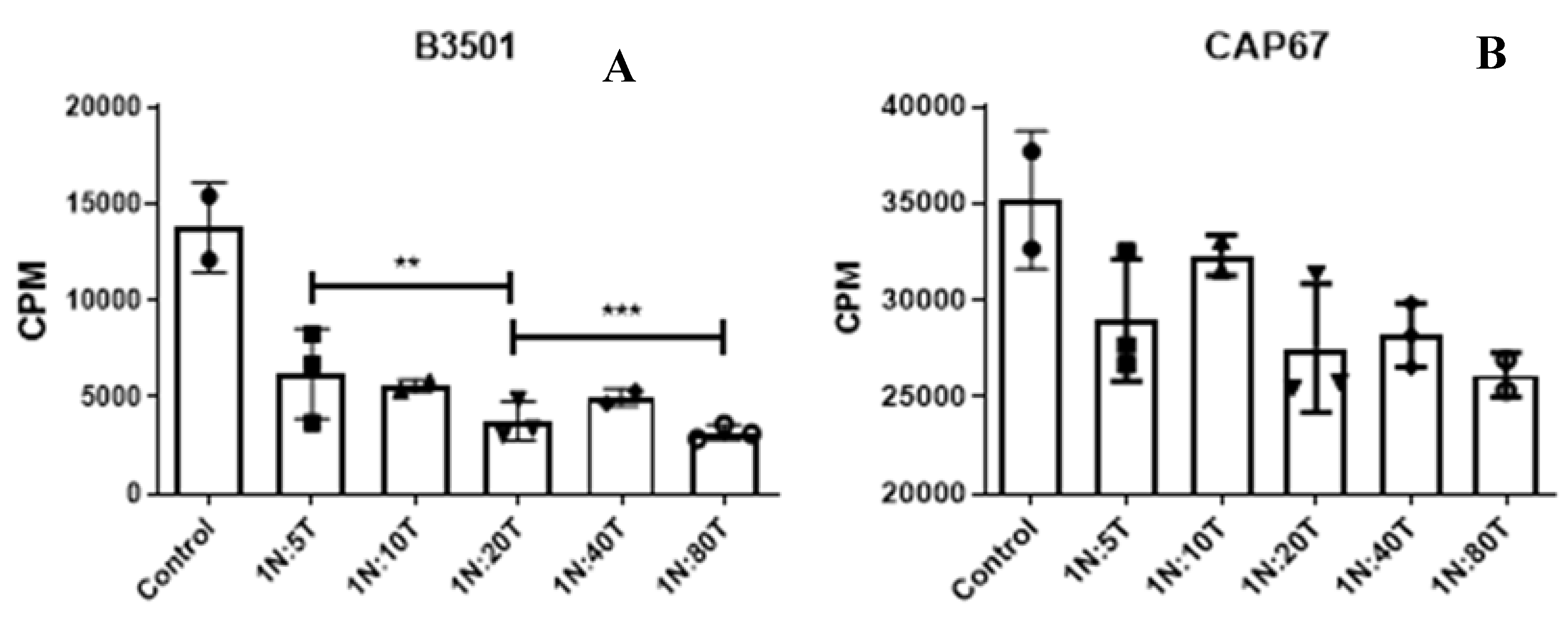
Figure 4.
α-CD3 stimulated T lymphocyte proliferation assay. Nylon wool column-enriched T lymphocytes were cultured with neutrophils purified from the peritoneal lavage of animals infected with the B3501 strain (A,B,C,D). The ratio between neutrophils and lymphocytes in culture was 1 neutro-phil to 5 lymphocytes (N1:5T), 1 neutrophil to 10 lymphocytes (N1:10T), 1 neutrophil to 20 lym-phocytes (N1:20T) and 1 neutrophil to 40 lymphocytes (N1 :40T). The control column refers to lymphocytes stimulated with a-CD3 in the absence of neutrophils. The DPI Control, NAC Control and LNIL Control columns refer to lymphocytes stimulated with α-CD3, treated with DPI, NAC and LNIL respectively, in the absence of neutrophils. After 48 hours, tritium-labeled thymidine was added to the culture (5uCi). After another 24 hours of culture, monitoring of cell proliferation was carried out by quantifying the radioactivity incorporated into the cellular DNA, expressed in counts per minute (cpm). The results presented are representative of two experiments performed separately. Data were compared relative to control. (***) p<0.0001.
Figure 4.
α-CD3 stimulated T lymphocyte proliferation assay. Nylon wool column-enriched T lymphocytes were cultured with neutrophils purified from the peritoneal lavage of animals infected with the B3501 strain (A,B,C,D). The ratio between neutrophils and lymphocytes in culture was 1 neutro-phil to 5 lymphocytes (N1:5T), 1 neutrophil to 10 lymphocytes (N1:10T), 1 neutrophil to 20 lym-phocytes (N1:20T) and 1 neutrophil to 40 lymphocytes (N1 :40T). The control column refers to lymphocytes stimulated with a-CD3 in the absence of neutrophils. The DPI Control, NAC Control and LNIL Control columns refer to lymphocytes stimulated with α-CD3, treated with DPI, NAC and LNIL respectively, in the absence of neutrophils. After 48 hours, tritium-labeled thymidine was added to the culture (5uCi). After another 24 hours of culture, monitoring of cell proliferation was carried out by quantifying the radioactivity incorporated into the cellular DNA, expressed in counts per minute (cpm). The results presented are representative of two experiments performed separately. Data were compared relative to control. (***) p<0.0001.
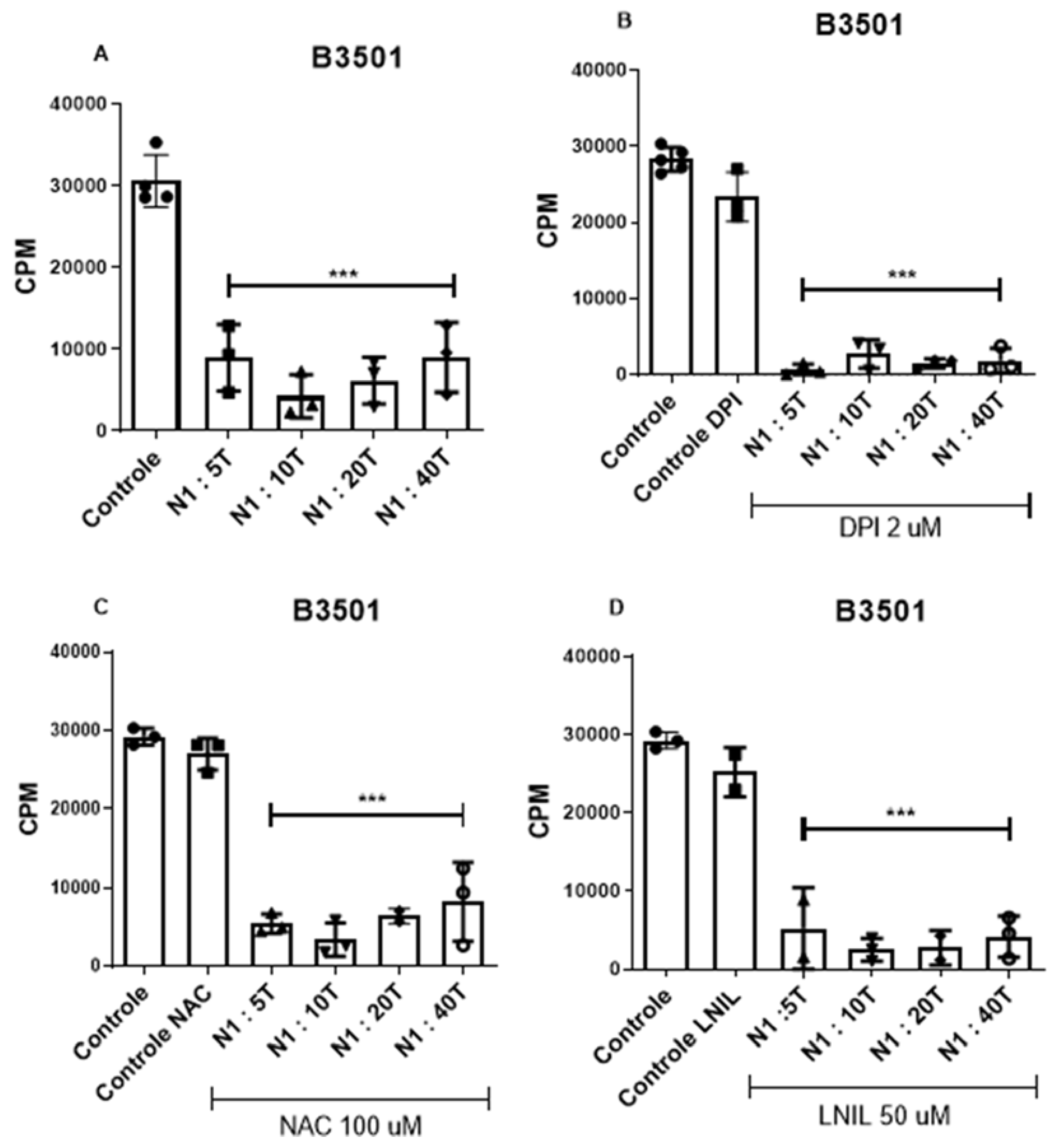
Figure 5.
Frequency of CD11b+ Ly6G+Ly6C-PD-L1+ cells in bronchoalveolar and peritoneal lavage. Per-centage of MDSCs PD-L1+ in the bronchoalveolar lavage sample after 48 hours (A) or 21 days (B) of sublethal infection of 5 x 105 fungal B3501 or CAP67 cells in the lung. Results for each group are represented as mean ± standard error. The results presented are representative of two experiments performed separately (*) p> 0.1, (**) p>0.01.
Figure 5.
Frequency of CD11b+ Ly6G+Ly6C-PD-L1+ cells in bronchoalveolar and peritoneal lavage. Per-centage of MDSCs PD-L1+ in the bronchoalveolar lavage sample after 48 hours (A) or 21 days (B) of sublethal infection of 5 x 105 fungal B3501 or CAP67 cells in the lung. Results for each group are represented as mean ± standard error. The results presented are representative of two experiments performed separately (*) p> 0.1, (**) p>0.01.
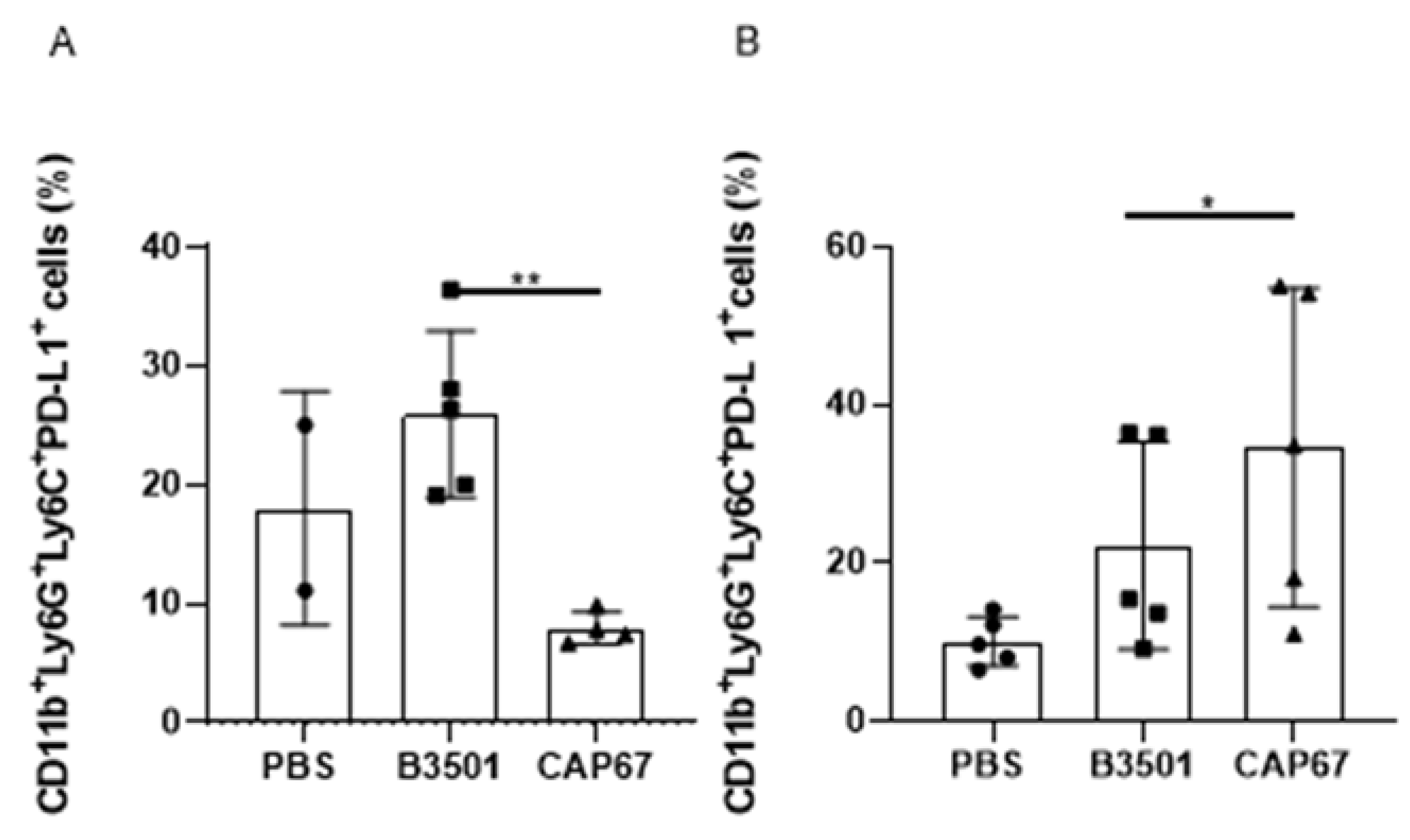
Figure 6.
Cryptococcus neoformans infection induces the recruitment of MDSCs, leading to T cell inhibition that compromises the protective host immune response, thereby facilitating systemic spread of the disease. These recruited cells exhibit high levels of PD-L1, contributing to immune responses suppression.
Figure 6.
Cryptococcus neoformans infection induces the recruitment of MDSCs, leading to T cell inhibition that compromises the protective host immune response, thereby facilitating systemic spread of the disease. These recruited cells exhibit high levels of PD-L1, contributing to immune responses suppression.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



