
Submitted:
17 June 2024
Posted:
20 June 2024
You are already at the latest version
Abstract
Amyloid self-assembled from amyloid peptides βΑ40 or βΑ42 is notorious for its neurotoxic effects in plaque and neurofibrillary tangle formations leading to neuron dysfunction and diseases of cognitive decline (e.g. Alzheimer’s, Parkinson’s and Huntington's disease). This contrasts with so-called functional amyloids which are non-toxic ordered template structures amenable to applications in tissue engineering. Amyloid fibrils of variable morphology including long and hollow fibres and flattened tube and spiral ribbon-like structures have been used in engineering applications in nano-biology. Protein assemblies based on amyloid core structures display diverse biological functionalities could be applied in futuristic self-assembling biomaterials in nano-electronics. These possibilities have revolutionized the development of next generation computers and biosensors, ultracapacitors, memristors, actuators, molecular switches and could also be used to develop artificial synapses. Amyloid fibril assemblies have also been used in photoelectric and photon capture light harvesting technologies and been applied in innovative nano-photoelectronics and photovoltaics. Hybrid Aβ(16–22)-α-synuclein amyloid fibrils also exhibit light-harvesting and electron-transfer properties. Engineered amyloid assemblies are thus facilitating innovative futuristic advances in nano-technology. Furthermore, with a better understanding of amyloid fibril assembly processes it may be possible to develop therapeutic methods that prevent the toxic build up of this polymer in brain tissues that leads to diseases of cognitive decline. With the ever-expanding prevalence of these diseases in the ageing general global population, there certainly is a clear and present need to find a remedy for these debilitating conditions.
Keywords:
amyloid fibrils
; nano-electronics
; memristors
; bio-sensors
; actuators
; molecular switches
; photovoltaics
; artificial synapses
; functional amyloids
; neuromorphic computing.
1. Introduction
The aim of this study was to review the roles of amyloid protein aggregates in neurodegeneration and contrast this with beneficial aspects of its structural organization which have found application in nanobiology in several innovative tissue engineering applications.
1.1. Definition of Amyloid
Amyloid is a historic generic term for insoluble misfolded protein aggregates with defining -pleated sheet content that can be assembled from 36 unrelated proteins. Amyloid can be stained with Congo red, exhibiting a characteristic green-yellow birefringence under polarized light [1,2,3,4,5,6]. These misfolded proteins exhibit specific features that facilitate assembly of ordered repeat structures, resulting in formation of insoluble amyloid deposits [2,3,4]. The A peptides specifically released from appican (APP) in brain tissues are assembled into dimers, trimers, oligomers and distinctive fibrillar structures. Generic amyloid deposits can occur in many tissues in the human body other than the brain, leading to organ and tissue dysfunction [7,8]. Hence, ‘Aβ’ the molecule and ‘amyloid’ the fibrillar deposit are not synonymous terms and should be distinguished. Aβ refers exclusively to the protein that, when aggregated into distinctive fibrils, constitutes the specific type of amyloid that most commonly accumulates in the aging brain in diseases of cognitive decline such as Alzheimer’s (AD) and Parkinson’s (PD). In pathology, amyloid refers to ‘mainly extracellular deposits of protein fibrils, recognized by certain properties, such as green-yellow birefringence after staining with Congo red’ [7] (for historical considerations of amyloid, see [2,3,4,5,6], and for more on the definition of amyloid see [2,9,10]). Amyloid can arise from over 36 different proteins in various parts of the body in different human diseases [7,8,11]. Amyloid deposits in the brain display distinctive morphologies (Figure 1). Amyloid plaques are visible with the light microscope using a variety of staining procedures, including silver stains, Congo red, Thioflavin, cresyl violet and periodic acid-Schiff (PAS) stains [7,12]. These methods stain different components of amyloid plaques, with variable sensitivity. Immunolocalization of amyloid plaques with a range of specific antibodies to Aβ epitopes and to other amyloid-associated components has also been utilized to visualize amyloid formations [13]. Examination of human autopsy samples and experimental models of AD have identified the biochemical, cytological, and inflammatory processes involved in the generation of these amyloid plaques [7]. Amyloid fibrils are diverse molecular structures [14], ENTAIL [15] and PARROT [16] are two systems that have been developed for the classification of amyloid fibril biodiversity. Ultrasensitive, new generation amyloid biosensors have also been developed for the detection of amyloid peptides in tissues, plasma and cerebrospinal fluid [17,18,19,20,21,22,23,24]. Amyloid plaque formation may be linked to trauma of the brain microvascular system [25]. Chronic brain inflammation and immune dysfunction may also contribute to amyloid deposition [26,27].
Aβ40 and Aβ42 oligomers of amyloid-β (Aβ) peptide associated with AD only differ by two C-terminal residues, however Aβ42 aggregates much faster than Aβ40 and is more toxic, forming pore-like structures in cell membranes [28,29,30]. The additional C-terminal residues in Aβ42 allow electrostatic interactions which stabilize the β-hairpin and promote dimer formation and oligomerization through intermolecular β-bridge formation [31,32]. A combination of hydrophobic and charged amino acids in Aβ peptides also contribute to aggregate formation; glycosaminoglycans (GAGs) also act at the earliest stage of fibril formation, namely during amyloid-beta nucleation [33]. The amyloid plaques in Alzheimer's brains consist mainly of Aβ42 and some plaques contain only Aβ42, even though Aβ40 concentrations in cerebrospinal fluids is several-fold higher than Aβ42 in the brain. The aggregative process in vivo thus favours Aβ42 interactions and, in this respect, differs from aggregative processes investigated in vitro [34,35]. By better understanding these aggregative processes in vivo we may be in a better position to develop therapeutic interventions which more effectively inhibit such aggregative processes and the deposition of Aβ42 plaques in AD [36,37].
1.2. Historical Amyloid Studies
Amyloidosis [38,39] was first identified as a disease in 1639 by Nicolaus Fontanus [40,41]. The visualized protein aggregates were called “amyloids” in 1838 by Matthias Scheiden, since they stained blue with an iodine stain, and thus were mistakenly believed to be some form of starch related to glycogen. The name persisted even though amyloid was later found to have a different chemical structure to starches. Amyloid deposition in the brain leads to AD and is a leading cause of dementia affecting individuals over the age of 60, with its incidence increasing in the ageing global population [41,42].
1.3. Amyloid Toxic Protein Aggregates Promote Neurodegeneration, Cognitive Decline, and Motor Dysfunction, However Functional Amyloids Have Beneficial Properties
The amyloid cascade hypothesis proposes that dysfunctional metabolism of amyloid precursor protein (APP) initiates the pathogenesis of AD [43]. This results in aggregation of Aβ and formation of neuritic plaques which cause pathological changes through formation of neurofibrillary tangles that disrupt normal synaptic connections. This results in compromised neuronal synaptic activity, neuronal cell death, cognition, and memory problems and development of dementia [44]. Natural amyloids are toxic proteins notorious for their roles in the pathogenesis of cognitive decline in AD and PD [43,45,46,47]. On the other hand, not all amyloids are toxic [48] nor do they all disrupt normal organ functions. So-called functional amyloids have beneficial properties in their own right through unique structural features and have been receiving considerable attention in tissue engineering applications in highly innovative areas of nanobiology [49]. From an engineering perspective, the self-assembling regular and tight packing structure of amyloids provides strength, and these are useful molecular templates amenable to structural modification and application in tissue engineering. Such self-assembled networks are highly suited to biomaterial scaffold developments [50,51,52,53]. Furthermore, amyloids are some of the strongest protein structures ever identified in nature [54,55]. For example, spider-web drag-line silk is composed of repeat peptide modules with a high -sheet content; on a weight for weight basis web silk has a strength exceeding that of high quality structural steel [56,57,58].
1.4. The Morphology of Toxic Amyloid Deposits in Brain Tissues
Insoluble amyloid deposits in brain tissues display characteristic morphologies as shown in the amyloid immunolocalizations depicted in Figure 1. Diffuse “cotton-wool like deposits” (Fig 4d) as well as prominent core deposits surrounded by a peripheral radial deposition pattern (Fig 4b, g, h) and diffuse deposits with internal granular punctate deposits (Figure 4c,e) are all evident [7,59]
2. Amyloid Precursor Protein and Its Bioactive Fragments
Appican is a transmembrane CS-proteoglycan precursor form of amyloid, containing an embedded amyloid precursor protein (APP) module within its core protein [61,62]. APP is a 110-130kDa type 1 transmembrane glycoprotein. APP is also a component of the related APP-like proteins 1 and 2 (APLP-1, 2). APP, APLP1, and APLP2 form a family of mammalian membrane proteins. Importantly, unlike APP, the APLPs lack the Aβ sequence, thus they do not give rise to the AD Aβ peptide [63]. APP and APLP regulate synaptic transmission, plasticity, and calcium homeostasis, and are therefore important cell messengers and regulators of nerve activity during neural transduction, nerve network formation and tissue development, thus having neuroprotective effects [64,65,66,67]. The soluble secreted fragment sAPPα counteracts the deleterious effects of the small APP-derived Aβ peptide that leads to amyloid deposition. APP, has a key role in pathological changes in AD, and has roles in synaptic transmission, synaptic plasticity, memory formation, and maintenance of neurons.
2.1. Proteolytic Processing of APP
APP is cleaved by γ-secretase, α-secretases, ADAM 9, 10, and 17, and β-secretases 1 and 2 (BACE1, BACE2) [68]. Newly identified secretases which cleave APP include η-secretase [63,69], meprin-β and δ-secretase [70]. These enzymes generate several biologically active fragments of APP [70]; some of these fragments are associated with the pathogenesis of AD and may provide opportunities as clinical targets in the treatment or prevention of AD [71]. APP can be cleaved to produce a 37 to 49 amino acid peptide, Aβ [67], that is central to amyloid formation [72]. Amyloid fibrils represent primary components of the neurotoxic plaques found in AD [73]. While specific proteins have been identified as components of amyloid aggregates, this is a generic term that includes any small protein that can undergo abnormal folding to form a -sheet that can then stack and propagate to form insoluble amyloid fibrils [74]. In addition, besides SAP, APP and A peptides, other misfolded proteins such as atrial natriuretic factor, prion protein, tau protein, immunoglobulin light and heavy chains, transthyretin, 2-microglobulin, synuclein, lysozyme and human islet protein can be incorporated into amyloid aggregates. Brain amyloid, however, is predominantly composed of Aβ40 and Aβ42 peptides.
2.2. The Enigmatic and Perplexing Story of APP Processing in Brain Tissues
APP was first described as a cell-surface receptor and was highlighted as a cell adhesion molecule [75]. APP is a type I transmembrane glycoprotein notorious for its involvement in the pathogenesis of AD through A40 and A42 peptides generated from APP in an amyloidogenic protease pathway in the brain involving β-secretase (BACE-1, -site APP cleaving enzyme, mepapsin 2) and γ-secretase [76]. These amyloid peptides self-assemble to form insoluble plaques and neurofibrillary tangles which detrimentally impact on synaptic function, neuron viability and cognitive processes in the brain, leading to dementia [77,78,79]. APP, however, is also processed by -secretase via a non-amyloidogenic pathway; this generates the soluble amyloid precursor sAPPα peptide [76]. sAPPα promotes neuroprotection, synaptic plasticity, memory formation, neurogenesis, neuritogenesis, and reduces amyloid and tau pathology [80,81,82]. Several studies also suggest that sAPPα regulates the trafficking of APP and its processing by proteases via both amyloidogenic or non-amyloidogenic pathways, which may decrease the risk of developing AD [83]. Aβ monomers share similar properties to sAPPα however, when self-assembled into Aβ oligomers, they become neurotoxic [82]. The precise physiological functions of full length APP and its proteolytic fragments is thus a complex story [75,84,85], Aβ production is not deleterious per se but when assembled into oligomers A has toxic consequences [84,85,86]. While the exact role of A in the pathophysiology of AD is still unresolved it has proposed that A has neuroprotective properties in multiple sclerosis (MS) and potential therapeutic benefits[87]. MS is an autoimmune disorder propagated by immune cells of the peripheral circulation which have damaging effects on the CNS gray and white matter. Demyelination of focal regions of the CNS occur due to infiltration of lymphocytes and macrophages which cause axonal damage [88,89] and these infiltrating cells have been proposed as immunohistochemical biomarkers of this degenerative process along with associated A, also found in these regions of tissue damage[90,91]. A is a key component of amyloid plaques in AD and associated with activated microglia[92] and astrogliotic astrocytes[93] in neuroinflammation. However in an animal model of experimental autoimmune encephalomyelitis (EAE), peripheral injection of A peptides had a significant protective effects against progressive degenerative tissue changes in EAE[89], a perplexing observation. Thus A can have both beneficial and detrimental effects depending on tissue context.
2.3. Amyloid Deposition Can Impact on Tissue Pathology
Progressive deterioration of cognitive functions induced by amyloid deposition include progressive loss of neuron metabolic and synaptic function in specific brain regions, deposition of tissue disruptive amyloid plaques and neurofibrillary tangles, neuroinflammation, blood-brain barrier breakdown, mitochondrial dysfunction, and ER and oxidative stress. Dementia (now termed major neurocognitive disorder) and AD effect more than 24 million people worldwide. This can occur with ageing and has an increasing prevalence due to extended global lifespans with an estimated 10 million new cases occurring annually [94], affecting over 10% of the total world general population aged 65 years and older. Model projections indicate such cases may increase three-fold by 2050 to 48.98 million [94]. The World Alzheimer´s Report indicated a prevalence of 46.8 million people affected by AD worldwide and a projected tripling of this condition by 2050. Alzheimers Disease International (https://www.alzint.org) reported an incidence of 55 million AD individuals in 2020.
2.4. Determining the Impact of Amyloid Fibrils on Normal Tissue Physiology Using Quantum Mechanics
Quantum mechanics has been used to compute the impact of misfolded amyloid fibril structures on neurodegenerative disorders [95] and in cardiac mechanobiology [96]. The impact of an increase in highly reactive hydrophobic centre interactive -sheet structures in amyloid deposits on the propagation of amyloid fibrils can be accurately computed using quantum mechanics. This technique is reported to be more accurate than molecular dynamics measurements in the prediction of amyloid fibril assembly processes. Quantum mechanics has opened up new approaches for the accurate non-invasive mapping of the heart’s conductivity with the aid of quantum sensors [96]. Transfer learning techniques using a smart machine learning algorithm has been developed to analyse important attributes to cardiac activity. An electric heart model has been developed where different amyloid deposition levels can be simulated to assess its impact on cardiac function. Atrial fibrillation is affected by cardiac amyloid levels and cardiac function can be computationally assessed using AI algorithms and quantum mechanics.
3. Identification of Proteins with Amyloid Aggregative Potential
The propensity of a peptide to form an amyloid fibril is dependent on factors such as polypeptide charge, sequence, hydrophobicity and peptide secondary structure and not by a specific “amyloid” amino-acid sequence per se [97,98,99,100,101,102,103]. Amyloidogenic peptides self-assemble into repetitive stacked structures that attract further proteins to propagate fibril formation [74]. Predictive algorithms have been developed to assess peptides that display a propensity to form amyloid fibrils in web-based software that predicts aggregation-prone protein sequences [104,105,106].
AMYLPRED2 (http://biophysics.biol.uoa.gr/AMYLPRED2) is a public web tool for the prediction of amyloidogenic determinants in 'aggregation-prone' peptide sequences within proteins [105].
3.1. Determination of Amyloid Forming Peptide Sequences in Proteins
Development of methods to produce controlled amyloid fibrils in vitro is an important innovation and opens the door for the design of new biomaterials exploiting the superior structural properties of amyloid fibrils and the inherent diversity of peptide sequences [107]. A number of bioinformatics [108,109] and computational studies [110,111,112,113,114,115,116,117,118,119] have examined the peptide sequences of amyloid proteins [120,121,122,123,124,125,126] to better understand the aggregation process and to determine peptide sequences that initiate and propagate assembly of fibrillar material. Software developed for the prediction of peptide sequences with inherent amyloid fibril forming capability should be very useful in the design of specific amyloid nano-components for specific tissue engineering applications [127,128,129] and in therapeutics development of compounds that prevent amyloid assembly.
3.2. The Impact of Misfolded Proteins on Normal Tissue Functional Properties
The increasing presence of misfolded mutant or damaged proteins in cells sets in motion protective cellular stress responses against this accumulation which, if not resolved, can lead to cell death [130,131,132]. Neurodegenerative disorders such as AD, PD, HD, ALS and prion diseases share a characteristic accumulation of aggregated misfolded proteins [133,134,135]. These elicit an ER stress response that may protect cells against the toxic buildup of these misfolded proteins [135,136,137,138,139]. If these proteins accumulate to an excessive extent, this can overwhelm the cells ‘quality control’ system, impairing protective mechanisms designed to promote correct protein folding and elimination of faulty proteins. This can ultimately lead to organelle dysfunction and cell death [135,136,137,138,139,140,141,142,143,144].
Proteins have an intrinsic propensity to undergo misfolding interactions influenced by amino acid composition and certain mutations accelerate this process [134]. Environmental conditions such as increased temperature, high or low pH, agitation, elevated glucose or oxidative agents can also promote a loss of native protein structure, protein denaturation and unfolding and the production of non-functional proteins [133]. The unfolded state is thermodynamically unstable and unfolded proteins tend to undergo aggregative processes. This leads to the formation of more stable intermediate oligomers and to highly ordered fibrils enriched in cross- structure. Amyloid is a stable aggregated protein conformation of unbranched linear peptide fibrils; these bind fluorescent Congo red and thioflavin derivatives aiding in their identification.
4. The Attributes of Functional Amyloids
In nature, amyloids have a range of functions across diverse organisms, including mammals, bacteria, fungi and marine organisms [38,39,145,146,147,148]. Functional amyloids participate in an array of beneficial physiological processes such as regulation of pigment formation, storage and controlled release of peptide hormones, memory, fertilization of oocytes by sperm, antimicrobial responses, regulated necrosis, cellular responses to stress and powerful adhesive properties. Amyloid as a biological glue has potential application in highly specialized surgical procedures [140,147,148,149,150,151,152,153,154,155,156]. Naturally occurring amyloids are also associated with a number of disease processes (Table 1A). Amyloid fibrils are also found as components in marine bioadhesives such as those which provide adhesion of barnacles and mussels to substrata. These have powerful adhesive properties that have inspired the development of tissue adhesives of potential application in highly specialized surgical procedures [157]. The unique architectural assembly processes and exceptional mechanical strength of amyloid fibrils makes these structures of interest in innovative applications in organic micro-circuitry in nano-electronics, in the development of actuators, molecular switches, memristors and microcomputing [158,159,160,161,162,163,164,165,166,167]. The memristor is a resistor with memory that behaves similarly to biological synapses [168]. The low power requirements and ultra-high speed signal transmittance capability of memristors is revolutionising development of neuromorphic circuits that are used in synthetic neural networks, switching devices and low-power sensors in microcomputing [169,170,171]. Nano-wires cast using hollow amyloid fibrils as casting templates have been used in bio-sensing, optoelectronics and photovoltaics and show potential in the development of synthetic synapses in highly innovative bio-nanotechnological applications [172,173,174]. Furthermore, carbon-nanomaterial-amyloid fibril hybrids have potential uses in organic micro-electronics and bio-sensing in biomedicine and in structural nano-biomaterials [172,173,174,175]. Amyloid fibrils have found a number of applications in tissue engineering (Table 1) [176]. Photobiomodulation therapy, using near infra-red 700-1400 nm low-level laser phototherapy, reduces the deposition of beta-amyloid in the AD brain, ameliorating neuroinflammation and oxidant stress, and supporting mitochondrial homeostasis to elicit a healing or regenerative response [177]. The surface chemistry of engineered amyloid fibrils can be modified depending on the amino acids used and bacterial expression systems used in their assembly; fibrils can also coated with chemicals that modify their responsiveness to specific chemical microenvironments [178,179,180]. Fibrils can be coated with gold nanomaterials modified with peptides or other chemicals. Gold itself is chemically inert but highly conductive and, when modified with additional components, can fine-tune the surface interactivity of amyloid fibrils. Some chemicals can improve the light harvesting and electron transfer properties of fibrils which can improve the efficiency of photobiomodulation therapy [181]. Other amyloid fibrils can display catalytic enzymatic properties of hydrolases, esterases, lipases that can be harnessed for specific purposes or it can be used as a nanoscaffold for enzyme immobilisation with enzymatic activities that disassemble insoluble amyloid deposits [181,182,183,184].
4.1. Bacterial Amyloids
Amyloid fibrils have roles in the formation of fimbriae in some bacteria and in transmission of epigenetic traits in fungi. Fimbriae are small adherent 3–10 nm diameter appendages several micrometers in length that attach many Gram-negative and some Gram-positive bacteria to one another, to animal cells or to abiotic surfaces resulting in biological fouling. Gram-negative bacteria assemble surface fibres called Curli in E. coli, Salmonella, and members of the Enterobacteriales [107,196] (Figure 2).
In fruit flies, Orb2, a polyadenylation RNA binding amyloid-like protein, has roles in long-term memory formation and retention [200,201]. Orb2 fibrils in Drosophila display a remarkably similar structure to mammalian amyloid fibrils, forming ~75-nanometer-long threefold-symmetric amyloid filaments. Interestingly, these fruit fly proteins are non-toxic (Figure 3). Amyloid precursor-like protein also has roles in the assembly of structures involved in memory formation in Drosophila [196,202,203].
Cryo-EM has provided detailed structural information on heterogeneous nuclear ribonucleoproteins with a modular structure that facilitates their assembly into pathological or functional amyloids [205]. Mutations in these assembly domains are associated with hereditary neurodegenerative conditions, including amyotropic lateral sclerosis (ALS), dementia, AD and PD [206]. Unlike pathological amyloids functional amyloid assembly is predominantly stabilised by hydrophobic centres and hydrogen bonding. Nuclear ribonucleoproteins are a large family of RNA-binding proteins that control the biogenesis of RNA in a tightly controlled process where ribonucleoproteins are further processed to give rise to mature mRNAs [207]. Many of the ribonuclear proteins have prion-like sequences that drive the assembly of fibrillar assemblies in functional amyloids, however when missense mutations are present, formation of pathological amyloids may lead to neurodegenerative conditions or cancer [206]. Amyloidogenic proteins from the cytoplasmic polyadenylation element-binding protein family play a crucial role in long-term memory formation in humans [208,209]. These proteins are well-characterized ribonuclear binding proteins (RBPs) that stabilize activity-dependent changes in synaptic connections [209]. RBPs regulate the flow of genetic information between the genome and proteome [207].
4.2. Marine Amyloids
Amyloid fibrils have also been observed in adherent marine organisms and contribute to their astonishing strength, tenacity and very rapid adhesive properties under adverse environmental biological conditions [54,107] contributing to the high performance of such adhesives and the tenacity of binding displayed by marine organisms.[145,210]. Such polymers in barnacles and mussels have been de-engineered and new polymers created, inspiring the development of a new generation of high-performance surgical adhesives. These may obviate the need for sutures in demanding surgical procedures in very soft tissues where sutures may not hold adequately and may be a potential site where tearing of suture sites can increase the likelihood of microbial infection [157,211,212]. Surgical bioadhesives based on amyloid are non-immunogenic and may obviate the need for sutures altogether, providing improved healing responses in cardiac surgery and can even be used directly on the beating heart where their rapid, exceptional tissue adhesive properties are an important innovative surgical application [213].
4.3. Insect Amyloids
Amyloid fibrils also occur in high performance structural insect proteins. Spider-web drag-line and aquatic silk fibroin copolymers in silk-moth (Bombyx mori), caddis-fly larvae (order Trichoptera) and sandcastle worms (Phragmatopoma californica) have assembly properties similar to those in brain amyloid fibril formation based on modular silk-homology repeat motifs and -sheet formations which promote formation of block polymers [56,57]. Silk proteins have found application as suturing material and the preparation of engineered composite constructs used in biomedicine [157]. Functional amyloid fibril formation in insects is controlled by specific chaperone proteins and regulatory domains [48,156] which prevent uncontrolled aggregation of protein assemblies that could threaten control of cell and tissue function. This occurs in protein aggregation protein-misfolding diseases in humans such as AD, PD, Huntington's disease, Creutzfeldt-Jakob disorder, cystic fibrosis, and Gaucher's disease [156,214].
4.4. Human Amyloids
In humans, amyloids are insoluble self-assembling protein aggregates that spontaneously occur within tissues. These proteins are folded into a form that allows many copies of that protein or an unrelated one to stick to such assemblies to form a stacked aggregate structure, facilitating microscopic fibril formation [40,41]. The macromolecular structure of such aggregates may be visualised in tissues through interactive properties with the azo-dye Congo red. Intercalation of Congo red with amyloid fibrils generates a characteristic apple-green birefringence when viewed under polarised light. Amyloid formation is linked to the development of a number of human diseases. Amyloid macromolecular protein aggregates disrupt normal tissue function by a process known as amyloidosis [200]. While amyloid deposits can form from many dissimilar proteins a feature in common held by all of these proteins is that they are of small molecular weight and have an ability to form sheet structures that assemble into long unbranched fibres [40]. Pathogenic amyloid aggregates occur when previously healthy proteins lose their normal physiological functions, attain a misfolded form and form fibrous deposits in so called plaque formations around cells that disrupt normal cellular functions.
4.5. Mammalian Amyloid-Related Proteins
Serum amyloid P (SAP, Pentraxin-2) [215] is a 25kDa pentameric protein of the pentraxin protein family [216] which becomes incorporated into insoluble amyloid protein aggregates [217]. Deletion of SAP delays formation of amyloid in tissues [218]; SAP prevents the proteolysis and clearance of amyloid deposits [141]. C-reactive protein (CRP) is a 120kDa protein that shares ~50% homology in amino acid sequence with SAP and is released during inflammation. CRP is used as a disease marker; its levels may increase 1000-fold to regulate inflammation and infection [219].
Amyloid fibrils are ordered nanostructures that undergo self-assembly [74,176]. Charged and hydrophobic amino acid sequences (Figure 4) have been shown to have roles in these aggregative processes, GAGs can also aid in amyloid fibril assembly in the early nucleation stages of fibril formation [220,221,222]. Cooperative hydrogen bonding promotes amyloid formation [223]. Amyloid fibrils can disrupt normal cellular activity and tissue architecture affecting tissue functional properties. In neurodegenerative disorders such as AD and PD this results in dysfunctional neuronal synaptic and neural network activity and feed-on effects on cognitive processes, memory and motor functions affecting neuromuscular control of normal bodily movement.
4.6. Amyloids that Display Metallo-Enzyme Catalytic Activities
Amyloid fibrils are remarkable interactive structures; short prion peptides have been observed to assemble into fibrillar structures that display properties of metallo-enzymes [224]. Enzymes folded into defined 3D protein structures exhibit a high catalytic efficiency and selectivity [224]. From an evolutionary standpoint, it has been proposed that the earliest enzymes may have arisen from self-assembly of such short peptides into supramolecular amyloid-like structures. Amyloid fibrils are ordered nanostructures that are self-assembled in vivo. Several artificially-produced amyloids display catalytic activity and offer advantages over natural enzymes due to their regular modular structure, flexibility, stability, and reusability [164]. Hydrolases and esterases, are common amyloid-like nanozymes; some of these act as carbonic anhydrases [225]. Carbonic anhydrase (carbonate dehydratase EC 4,2.1.1) is a very ancient metalloenzyme present in red blood cells (RBCs) that catalyses the bidirectional interconversion of CO2 and water (H2O) into bicarbonate (HCO3−) ions and protons (H+) controlling acid-base levels, pH and fluid balance, impacting on numerous physiological processes within and across numerous tissue compartments throughout the human body. When RBCs reach the lungs, haemoglobin binds oxygen and delivers this via the bloodstream to all cells. This is required to fuel the breakdown of sugars and fat in cellular metabolism for energy production and generates CO2 that needs to be removed from the body. Approximately 20% of the CO2 produced by cellular metabolism binds to hemoglobin, while ~70% is converted to carbonic acid and bicarbonate ions by carbonic anhydrase and transported in the blood then converted back to CO2 to be carried out of the lungs during expiration. While this interconversion of CO2 can occur in the absence of carbonic anhydrase this enzyme assists rapid inter-conversion of CO2 and water into carbonic acid, protons and bicarbonate ions by approximately one-million-fold. This is a fundamental evolutionary adaptation essential to life in mammals. Carbonic anhydrase inhibitors have been shown to reduce cerebral, vascular, and glial Aβ accumulation and amyloid fibril deposition and improved cognitive processes in mice. Carbonic anhydrase inhibitors may be useful in the treatment of AD and cerebral amyloid angiopathy [226]. Carbonic anhydrase occurs as 14 isozyme zinc metalloenzymes that catalyze the reversible hydration of carbon dioxide to bicarbonate and have major roles in controlling acid-base balances and pH in tissues [227]. This is particularly important in the regulation of the astrocyte [228] in brain tissues which controls ion profiles in niche environments that control neuronal activation and neurotransduction and signaling activity in neural networks [229]. Recent studies highlight the involvement of activators of carbonic anhydrase in brain processes essential for the transmission of neuronal signals [230]. Carbonic anhydrase activation may be a potential therapeutic option in the treatment of AD and neurodegenerative conditions characterized by memory loss and cognitive impairment. Furthermore, CO2 has important roles to play in the regulation of mitochondrial metabolism [231]. Carbonic anhydrase in mitochondria regulates CO2 fixation assisted by enzymes of the tricarboxylic acid (TCA) and Embden Meyerhof Parnas (EMP) aerobic glycolysis and gluconeogenesis pathways respectively which are responsible for ATP generation and energy production [232]. This is important in neuronal energy production, the neuron is an energy demanding cell type [233].
5. Amyloid Fibrils as Building Blocks for Innovative Biomaterials
Protein assemblies based on amyloid core structures display diverse biological functions with potential application in futuristic self-assembling biomaterials [52,181,185,234,235,236,237]. These roles contrast with the stigmatized picture of amyloid fibrils as pathological toxic structures in a range of human diseases of cognitive decline [74,238], however, not all amyloids are toxic. So-called functional amyloids have low toxicity and beneficial properties in a range of physiologic processes (eg, long-term memory formation, gradual release of stored peptide hormones). Interestingly, amyloid fibrils formed by HET-s, a fungal protein [239,240], can confer functional prion-like fibril assembly behavior. The assembled structures have a much higher Young's modulus and tensile strength than those of PrP, i.e., they are much stiffer and stronger due to a tighter packing of the fibril structure [241]. The HET-s fungal cytoplasmic prion protein assembles into amyloid-like nucleation structures which accelerate formation of amyloid-like fibrils [242]. These stain with Congo red and are birefringent under polarised light. HET-s fibrils resist compressive and tensile forces better than PrP and are stiffer and stronger than A aggregates.
Functional amyloids play a role in cell fate and control cell-signalling cascades in mammals, bacteria and fungi. Amyloids have a range of functions inside and outside cells such as hormone and toxin storage and release, formation of cell surface structures in microorganisms and scaffolding associated with pigment synthesis [243,244,245]. The csgA gene encodes curlin - the major subunit protein of curli, a thin, coiled surface bacterial structure which binds to a variety of extracellular matrix and serum proteins and has roles in E coli biofilm formation [246]. Functional amyloids, such as CsgA and FapC are highly stable, and can be used to assemble monopolymeric amyloids [247]. CsgA has been used to prepare hydrogels suitable for gastrointestinal colonization and regenerative tissue engineering, cell-specific drug release, water-purification filters, and biosensors [248]. FapC shares many features with CsgA but has yet to be used to develop functional scaffolds.
5.1. Functional Amyloids and Their Application in Tissue Engineering
Functional amyloids display many features that make them attractive biomaterial candidates for tissue engineering applications [56,176,249]. Amyloid fibrils undergo self-assembly, forming regularly organised structures with impressive biophysical properties and a range of morphologies including extended straight filaments, tapes, twisted ribbons, and hollow tubes. Fibrils are typically 5–20 nm in diameter with a length in the micron size range. This is irrespective of the many different proteins from which amyloid structures are assembled; currently 36 proteins have been demonstrated to be capable of forming amyloid deposits. Fibrils can be assembled from a diverse range of small proteins and polypeptides over a wide range of assembly conditions influenced by temperature, pH and solvent conditions. This provides flexibility in the procedures that may be utilised in engineering applications to assemble fibrils at the nanoscale level. Furthermore, since fibrils are assembled from arrangements of amino acids these are amenable to genetic manipulation and the use of a range of bacterial expression systems for their production. Moreover, introduction of point mutations in fibril structures can potentially be used to design structures with variable chemical surface and electrostatic characteristics to produce fibrils of variable binding properties and unique environmental responsiveness [237,250].
Amyloid fibrils possess a Young’s Modulus in the GPa scale and a strength comparable to steel. High-resolution data gathered from X-ray diffraction and NMR experiments, demonstrate the presence of extensive cross β-sheet structure within the core of the fibril which forms an expansive hydrogen-bonding network throughout the polypeptide backbone spanning the length of the fibril. It is this cooperative intermolecular hydrogen-bonding network which confers stability and unique material properties to amyloid fibrils [54,251]. Amyloid fibrils have similar mechanical properties to dragline spider silk, which is one of the strongest and most rigid biomaterials in nature [252]. Silk and amyloid fibrils both contain expansive hydrogen-bonded β-sheet networks.
5.2. Engineered Amyloid Polymers
Several classes of engineered amyloid polymers have so far been developed with impressive credentials as biomaterials for engineering applications. These include (i) templates for nanowires and nanoelectronics, (ii) hydrogels applied to cell scaffolds and stem cell and drug delivery, (iii) light harvesting electron transport biomaterials for biophotonics, (iv) biosensors, actuators and molecular switches [183,253].
Hollow ~100 nm nanotubes have been developed using self-assembly of amyloid-like fibrillar structures to form templates within which silver nanowires can be cast of 10-nm width and lengths ranging from 60-100 microns [162,163,254]. The central nanowire is recovered by proteolytic dissolution of the peptide shell. Multi-layered co-axial nano-wire assemblies have also been prepared decorated on the exterior of the nanowire with metallic gold [161]. The nanowire structures were confirmed by TEM and energy dispersive X-ray analysis. Gold or silver nanowires with widths of ~100 nm have demonstrated high conductivities and low resistances (~80 Ω). Nanowires can be applied to microcircuitry in nano-electronics and have revolutionised the development of next generation computers and biosensors [255]. Ultracapacitors have also been developed using an external magnetic field to orient horizontal and vertically aligned arrangements of nanotubes. These amyloid assemblies display enhanced capacitance relative to carbon and carbon nanotube–modified electrodes [160].
5.3. Amyloid Cell Attachment Matrices and Hydrogels for Cell Delivery in Tissue Repair Strategies
The ECM has important instructive properties over cells, communication between the cells and ECM is encoded in peptide epitopes of structural and signaling components that act as molecular directors of cellular activity [256]. Peptide epitopes can be incorporated into synthetic biomatrices to mimic the specific communication that occurs between cells and tissues to control cell adhesion, differentiation, immunomodulation and ECM turnover to achieve tissue homeostasis. Self-assembled amyloid peptide biomatrices have been prepared with RGD motifs to improve cellular attachment [257]. Lysozyme amyloid micronetworks have been shown to support chondrocyte viability and ECM deposition, while α-synuclein and β-lactoglobulin matrices maintained cell viability [256]. Synthetic amyloid biomatrices with specific added proteins hold promise in cartilage regeneration [258].
Amyloids are novel cell-adhesive matrices [259]; cell adhesion on amyloid fibrils occurs despite a lack of integrin recognition motifs [260]. Amyloid biomatrices have also been prepared containing RGD cell attachment motifs [257]. Amyloid scaffolds represent versatile biomaterials for cell adhesion and tissue engineering applications [52]. Amyloid fibril hydrogels have also been prepared and shown to be suitable 3D macrostructures for cell culture and as cell delivery vehicles for cultured stem cells which have undergone cellular differentiation [261,262]. In tissues, amyloid fibrils also promote neural proliferation and neurite outgrowth through interaction with ECM components [75,263]. Amyloid fibril 3D scaffolds have been used to culture neurons and used to develop a model of AD [264]. Amyloid fibril bioscaffolds have been used to culture neural progenitor cells to assess if an A sheet environment guided differentiation of cultured neural progenitor cells simulating conditions found in brain tissues in which amyloid deposition was evident. A sheet bioscaffolds produced neural progenitor cells of a phenotype similar to that induced by amyloidosis in AD tissues [265].
Amyloid hydrogels are highly hydrated 3D polymer networks with cell adhesive properties that are suitable as a scaffold for tissue engineering, regeneration, and repair [266]. Such hydrogels are smart materials capable of transitioning to altered rheological properties in response to spatiotemporal changes in temperature, pH, ionic strength, and external stresses and are also applicable for drug delivery and stem cells in regenerative procedures [186,267].
5.4. Application of Amyloid Fibrils in Nanobiology and Organic Microelectronics
Amyloid fibrils have found application in photoelectric and light harvesting technologies [268]. A peptide fragment of transerythritin (TTR 105-115) spontaneously forms highly aligned uniform fibrils in which fluors can be incorporated with little disruption in fibril structure to provide an amyloid fibril with the ability to capture photons and harvest light [269]. Aβ(16–22)-α-synuclein hybridised amyloid fibrils also exhibit light-harvesting and electron-transfer properties [270].
While naturally occurring amyloids are robust self-assembled nanometer-sized fibril biomaterials, it is now possible using in-vitro methods to assemble large 10-20 μm diameter amyloid fibers several mm in length [271]. Using a short, hydrophobic director α-helical template peptide and mixtures of peptides it is possible to self-assemble large amyloid fibers [272], encoded by micron-sized self-assembled structures at the genetic level, with tailored rectangular or cylindrical cross-sectional morphologies and robust material properties (modulus 0.1-2.5 GPa) [273]. A template peptide expressed in E. coli has been used to grow customized 200-500 nm amyloid fibrils and 7-20 μm wide fibers for tissue engineering.
5.5. Amyloid Fibrillar Assemblies and Neuromorphic Computing
Quantum computers offer the computational power required to drive neuromorphic hardware in neural network dynamic simulations [274]. Machine learning and artificial intelligence algorithms running on neuromorphic hardware by quantum computers are being developed to assist in data analysis in artificial synapse modular supercomputing developments. Self-assembling amyloid fibrillar structures can be modeled to provide neuromorphic hardware due to varied fibril surface chemistry and their responsiveness to specific electrochemical microenvironments. This may be useful in the development of electrochemical random-access memory using ionic neurotransistors, leading to neuromorphic computing networks that drive sensory intelligent perception systems [275,276].
6. Concluding Remarks
This review has illustrated the toxic features of amyloid plaques and neurotangles in brain tissues which lead to neurodegeneration, impaired neuron synaptic function and consequential diseases of cognitive decline. The beneficial properties of functional amyloids are also reviewed and engineering applications that have been developed using the amyloid fibril as a molecular template which can be manipulated in a number of innovative applications in significant advancements in nano-technology. These have made important contributions to neuro-signaling in neural repair strategies and in areas of bio-regulation using nano-electronics. Development of biosensors, actuators, ultracapacitors, memristors, molecular switches and next generation micro-computing technologies are examples of these innovative applications that have been applied in bioregulation. The high conductance of nano-wires cast in hollow amyloid fibril molecular templates, low resistance and ultra-high signaling capability of such structures makes them particularly suitable in nano-electronic applications. Furthermore engineered amyloid fibril assemblies have also found application in photoelectric and photon capture light harvesting technologies of application in innovative nano-photoelectronics and photovoltaic applications which promise to revolutionise specific areas of optical neuro-regulatory processes. Furthermore, the application of AI methodology in brain-interface technologies offers particularly exciting possibilities in the improvement of real-time bidirectional control systems between living brains and actuators in motor and sensory neuromorphic applications and have had notable clinical success in the treatment of paralyzed patients’ and expanded the mobility of disabled patients [277].
7. Conclusions
This review has shown that functional amyloids are useful cell attachment matrices and hydrogels for cell delivery in tissue repair strategies and in innovative nano-electronics, bio-sensors, memristors and light harvesting technologies in photovoltaics. The stigma associated with amyloid fibrils that disrupt normal brain function impacting on cognition and memory processes thus needs to be balanced with the innovative properties of functional amyloids employed in the development of futuristic applications in nanobiology, organic nano-electronics and neuromorphic computing.
Author Contributions
JM conceptualized the review, MMS had intellectual input into the content of the review. Both authors shared in the writing and editorial review of the manuscript and agreed on its final content. JM prepared the figures used in this review.
Disclosures
JM has received consultancy fees from Arthropharm/Sylvan Scientific Pty Ltd. MMS is clinical director of Arthropharm/Sylvan Scientific Pty Ltd. This company had no input into the writing or interpretation of this review or the reason to publish. The authors have no conflicts to report.
Acknowledgments
This study was funded by the Melrose Personal Research Fund, Sydney, Australia.
References
- Benson:, M. : et al.,. (2018) Amyloid nomenclature 2018: recommendations by the International Society of Amyloidosis (ISA) nomenclature committee. Amyloid. 25, 215-219.
- Ke, P. , et al. (2020) Half a century of amyloids: past, present and future.. Chem Soc Rev. 5473; 49. [Google Scholar]
- Buxbaum, J.N.; Linke, R.P. A Molecular History of the Amyloidoses. J. Mol. Biol. 2012, 421, 142–159. [Google Scholar] [CrossRef]
- Puchtler, H.; Sweat, F. A REVIEW OF EARLY CONCEPTS OF AMYLOID IN CONTEXT WITH CONTEMPORARY CHEMICAL LITERATURE FROM 1839 TO 1859. J. Histochem. Cytochem. 1966, 14, 123–134. [Google Scholar] [CrossRef]
- Sipe, J. , Cohen, AS (2000) Review: history of the amyloid fibril.. J Struct Biol.
- Yakupova, E.I.; Bobyleva, L.G.; Vikhlyantsev, I.M.; Bobylev, A.G. Congo Red and amyloids: history and relationship. Biosci. Rep. 2019, 39, 1495–1506. [Google Scholar] [CrossRef]
- Walker, L. (2020) Aβ plaques. 1.
- Westermark, G. e. a. (2018) Noncerebral Amyloidoses: Aspects on Seeding, Cross-Seeding, and Transmission.. Cold Spring Harb Perspect Med. 0243; 8. [Google Scholar]
- Chiti, F. , Dobson, CM. . ( and human disease.. Annu Rev Biochem. 75, 333–366.
- Fändrich, M. On the structural definition of amyloid fibrils and other polypeptide aggregates. Cell. Mol. Life Sci. 2007, 64, 2066–2078. [Google Scholar] [CrossRef]
- Fowler, D.M.; Koulov, A.V.; Alory-Jost, C.; Marks, M.S.; E Balch, W.; Kelly, J.W. Functional Amyloid Formation within Mammalian Tissue. PLOS Biol. 2005, 4, e6. [Google Scholar] [CrossRef]
- Lamy, C.; Duyckaerts, C.; Delaere, P.; Payan, C.; Fermanian, J.; Poulain, V.; Hauw, J.J. COMPARISON OF SEVEN STAINING METHODS FOR SENILE PLAQUES AND NEUROFIBRILLARY TANGLES IN A PROSPECTIVE SERIES OF 15 ELDERLY PATIENTS. Neuropathol. Appl. Neurobiol. 1989, 15, 563–578. [Google Scholar] [CrossRef]
- Menter, T.; Bachmann, M.; Grieshaber, S.; Tzankov, A. A More Accurate Approach to Amyloid Detection and Subtyping: Combining in situ Congo Red Staining and Immunohistochemistry. Pathobiology 2016, 84, 49–55. [Google Scholar] [CrossRef]
- Li, D.; Liu, C. Structural Diversity of Amyloid Fibrils and Advances in Their Structure Determination. Biochemistry 2020, 59, 639–646. [Google Scholar] [CrossRef] [PubMed]
- Auriemma Citarella, A. , Di Biasi, L, De Marco, F, Tortora, G.. (2022) ENTAIL: yEt aNoTher amyloid fIbrils cLassifier.. BMC Bioinformatics. 23, 517. [CrossRef]
- Griffith, D.; Holehouse, A.S. PARROT is a flexible recurrent neural network framework for analysis of large protein datasets. eLife 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ren, B.; Zhang, D.; Liu, Y.; Zhang, M.; Zhao, C.; Zheng, J. Design principles and fundamental understanding of biosensors for amyloid-β detection. J. Mater. Chem. B 2020, 8, 6179–6196. [Google Scholar] [CrossRef]
- Kaushik, A. , Jayant, RD, Tiwari, S, Vashist, A, Nair, M.. (2016) Nano-biosensors to detect beta-amyloid for Alzheimer's disease management.. Biosens Bioelectron. 80, 273-287. [CrossRef]
- Gao, H. , Chen, J, Huang, Y, Zhao, R.. (2024) Advances in targeted tracking and detection of soluble amyloid-β aggregates as a biomarker of Alzheimer's disease.. Talanta. 268, 125311. [CrossRef]
- Sharma, P. , Kim, ES, Mishra, S, Ganbold, E, Seong, RS, Kim, YM, Jahng, GH, Rhee, HY, Han, HS, Kim, DH, Kim, ST, Kim, NY.. (2022) Ultrasensitive probeless capacitive biosensor for amyloid beta (Aβ1-42) detection in human plasma using interdigitated electrodes.. Biosens Bioelectron. 212, 114365. [CrossRef]
- Zheng, Y.; Zhang, L.; Zhao, J.; Li, L.; Wang, M.; Gao, P.; Wang, Q.; Zhang, X.; Wang, W. Advances in aptamers against Aβ and applications in Aβ detection and regulation for Alzheimer's disease. Theranostics 2022, 12, 2095–2114. [Google Scholar] [CrossRef]
- Jeong, D.; Kim, J.; Chae, M.-S.; Lee, W.; Yang, S.-H.; Kim, Y.; Kim, S.M.; Lee, J.S.; Lee, J.H.; Choi, J.; et al. Multifunctionalized Reduced Graphene Oxide Biosensors for Simultaneous Monitoring of Structural Changes in Amyloid-β 40. Sensors 2018, 18, 1738. [Google Scholar] [CrossRef]
- Wang, X. , Li, L, Gu, X, Yu, B, Jiang, M.. (2021) Switchable electrochemical aptasensor for amyloid-β oligomers detection based on triple helix switch coupling with AuNPs@CuMOF labeled signaling displaced-probe.. Mikrochim Acta. 188, 49. [CrossRef]
- Xing, Y. , Xia, N.. (2015) Biosensors for the Determination of Amyloid-Beta Peptides and their Aggregates with Application to Alzheimer's Disease. 48.
- Bu, X.-L. , Xiang, Y, Jin, W.-S.et al.. (2018) Blood-derived amyloid-β protein induces Alzheimer's disease pathologies. Molecular Psychiatry. 1948; 23. [Google Scholar]
- Heppner, F. , Ransohoff, RM, Becher, B (2015) Immune attack: the role of inflammation in Alzheimer disease Nature Reviews Neuroscience. 16.
- Jorfi, M. , Maaser-Hecker, A, Tanzi, RE.. (2023) The neuroimmune axis of Alzheimer's disease. Genome Medicine. 15.
- Brody, D. , Magnoni, S, Schwetye, KE, Spinner, ML, Esparza, TJ, Stocchetti, N, Zipfel, GJ, Holtzman, DM.. (2008) Amyloid-beta dynamics correlate with neurological status in the injured human brain.. Science. 1221. [Google Scholar]
- Serra-Batiste, M. , Ninot-Pedrosa, M, Bayoumi, M, Gairí, M, Maglia, G, Carulla, N.. (2016) Aβ42 assembles into specific β-barrel pore-forming oligomers in membrane-mimicking environments.. Proc Natl Acad Sci U S A. 1086. [Google Scholar]
- Kim, W. , Hecht, MH. . ( 2005) Sequence determinants of enhanced amyloidogenicity of Alzheimer A beta 42 peptide relative to A beta 40.. J Biol Chem. 280, 35069–35076.
- Itoh, S. , Yagi-Utsumi, M, Kato, K, Okumura, H.. (2022) Key Residue for Aggregation of Amyloid-β Peptides. 3139; 13. [Google Scholar]
- Hsu, F. , Park, G, Guo, Z.. (2018) Key Residues for the Formation of Aβ42 Amyloid Fibrils.. ACS Omega. 8401; 3. [Google Scholar]
- McLaurin, J. , Franklin, T, Zhang, X, Deng, J, Fraser, PE.. (1999) Interactions of Alzheimer amyloid-beta peptides with glycosaminoglycans effects on fibril nucleation and growth.. Eur J Biochem. 1101. [Google Scholar]
- Gu, L. , Guo, Z.. (2013) Alzheimer's Aβ42 and Aβ40 peptides form interlaced amyloid fibrils.. J Neurochem.
- Iwatsubo, T. , Mann, DMA, Odaka, A, Suzuki, N, Ihara, Y.. (1995) Amyloid β protein (Aβ) deposition: Aβ42(43) precedes Aβ40 in Down syndrome.. Ann Neurol. 37.
- Chen, G.F.; Xu, T.H.; Yan, Y.; Zhou, Y.R.; Jiang, Y.; Melcher, K.; Xu, H.E. Amyloid beta: structure, biology and structure-based therapeutic development. Acta Pharmacol. Sin. 2017, 38, 1205–1235. [Google Scholar] [CrossRef]
- Zhang, Y. , Chen, H, Li, R, Sterling, K, Song, W.. (2023) Amyloid β-based therapy for Alzheimer's disease: challenges, successes and future.. Signal Transduct Target Ther. 8.
- Dovidchenko, N.V.; Leonova, E.I.; Galzitskaya, O.V. Mechanisms of amyloid fibril formation. Biochem. (Moscow) 2014, 79, 1515–1527. [Google Scholar] [CrossRef]
- Siddiqi, M. , Majid, N, Malik, S, Alam, P, Khan, RH. . ( Protofibrils and Fibrils.. Subcell Biochem. 93, 471–503.
- Kyle, R. (2001) Amyloidosis: a convoluted story.. British J Haematol.
- Ow, S. , Dunstan, DE.. (2014) A brief overview of amyloids and Alzheimer's disease.. Protein Sci. 1315; 23. [Google Scholar]
- Citron, M. Strategies for disease modification in Alzheimer's disease. Nat. Rev. Neurosci. 2004, 5, 677–685. [Google Scholar] [CrossRef]
- Hardy, J.A.; Higgins, G.A. Alzheimer's disease: The amyloid cascade hypothesis. Science 1992, 256, 184–185. [Google Scholar] [CrossRef]
- Hardy, J. , Allsop, D.. (1991) Amyloid deposition as the central event in the aetiology of Alzheimer's disease. Trends Pharmacol Sci. 12.
- O'Brien, R.J.; Wong, P.C. Amyloid Precursor Protein Processing and Alzheimer's Disease. Annu. Rev. Neurosci. 2011, 34, 185–204. [Google Scholar] [CrossRef]
- Chiellini, G. Understanding Amyloid Structures and Disease: A Continuing Challenge in Health Research. Int. J. Mol. Sci. 2021, 22, 6620. [Google Scholar] [CrossRef]
- dos Santos, H.M.; Bertollo, A.G.; Mingoti, M.E.D.; Grolli, R.E.; Kreuz, K.M.; Ignácio, Z.M. Dementia and depression: Biological connections with amyloid β protein. Basic Clin. Pharmacol. Toxicol. 2024, 134, 563–573. [Google Scholar] [CrossRef]
- Jackson, M.P.; Hewitt, E.W. Why are Functional Amyloids Non-Toxic in Humans? Biomolecules 2017, 7, 71. [Google Scholar] [CrossRef]
- Rubel, M.S.; Fedotov, S.A.; Grizel, A.V.; Sopova, J.V.; Malikova, O.A.; Chernoff, Y.O.; Rubel, A.A. Functional Mammalian Amyloids and Amyloid-Like Proteins. Life 2020, 10, 156. [Google Scholar] [CrossRef]
- Reynolds, N.P. Amyloid-like peptide nanofibrils as scaffolds for tissue engineering: Progress and challenges (Review). Biointerphases 2019, 14, 040801. [Google Scholar] [CrossRef]
- Mankar, S.; Anoop, A.; Sen, S.; Maji, S.K. Nanomaterials: amyloids reflect their brighter side. Nano Rev. 2011, 2. [Google Scholar] [CrossRef]
- Das, S.; Jacob, R.S.; Patel, K.; Singh, N.; Maji, S.K. Amyloid Fibrils: Versatile Biomaterials for Cell Adhesion and Tissue Engineering Applications. Biomacromolecules 2018, 19, 1826–1839. [Google Scholar] [CrossRef]
- Binaymotlagh, R.; Chronopoulou, L.; Palocci, C. Peptide-Based Hydrogels: Template Materials for Tissue Engineering. J. Funct. Biomater. 2023, 14, 233. [Google Scholar] [CrossRef]
- Knowles, T.P.J.; Buehler, M.J. Nanomechanics of functional and pathological amyloid materials. Nat. Nanotechnol. 2011, 6, 469–479. [Google Scholar] [CrossRef]
- Sweers, K.K.M.; Bennink, M.L.; Subramaniam, V. Nanomechanical properties of single amyloid fibrils. J. Physics: Condens. Matter 2012, 24, 243101. [Google Scholar] [CrossRef]
- Li, J. , Zhang, F.. (2021) Amyloids as Building Blocks for Macroscopic Functional Materials: Designs, Applications and Challenges.. Int J Mol Sci. 1096; 22. [Google Scholar]
- Fukuma, T.; Mostaert, A.S.; Jarvis, S.P. Explanation for the mechanical strength of amyloid fibrils. Tribol. Lett. 2006, 22, 233–237. [Google Scholar] [CrossRef]
- Sawaya, M. , Hughes, MP, Rodriguez, JA, Riek, R, Eisenberg, DS.. (2021) The expanding amyloid family: Structure, stability, function, and pathogenesis.. Cell. 4857. [Google Scholar]
- Boon, B. , Bulk, M, Jonker, AJ, Morrema, THJ, van den Berg, E, Popovic, M, Walter, J, Kumar, S, van der Lee, SJ, Holstege, H, Zhu, X, Van Nostrand, WE, Natté, R, van der Weerd, L, Bouwman, FH, van de Berg, WDJ, Rozemuller, AJM, Hoozemans, JJM.. (2020) The coarse-grained plaque: a divergent Aβ plaque-type in early-onset Alzheimer's disease.. Acta Neuropathol.
- Kim, K. , et al. ( 1988) Production and characterization of monoclonal antibodies reactive to synthetic cerebrovascular amyloid peptide.. Neurosci Res Commun. 2, 121–130.
- Tsuchida, K.; Shioi, J.; Yamada, S.; Boghosian, G.; Wu, A.; Cai, H.; Sugahara, K.; Robakis, N.K. Appican, the Proteoglycan Form of the Amyloid Precursor Protein, Contains Chondroitin Sulfate E in the Repeating Disaccharide Region and 4-O-Sulfated Galactose in the Linkage Region. J. Biol. Chem. 2001, 276, 37155–37160. [Google Scholar] [CrossRef]
- Pangalos, M.N.; Shioi, J.; Efthimiopoulos, S.; Wu, A.; Robakis, N.K. Characterization of Appican, the Chondroitin Sulfate Proteoglycan Form of the Alzheimer Amyloid Precursor Protein. Neurodegeneration 1996, 5, 445–451. [Google Scholar] [CrossRef]
- Müller, U.C.; Deller, T.; Korte, M. Not just amyloid: physiological functions of the amyloid precursor protein family. Nat. Rev. Neurosci. 2017, 18, 281–298. [Google Scholar] [CrossRef]
- Hefter, D. , Ludewig, S, Draguhn, A, Korte, M. . ( and Electrical Activity of the Brain.. Neuroscientist. 26, 231–251.
- Turner, P. , O'Connor, K, Tate, WP, Abraham, WC. . ( plasticity and memory.. Prog Neurobiol. 70, 1–32.
- Korte, M. , Herrmann, U, Zhang, X, Draguhn, A.. (2012) The role of APP and APLP for synaptic transmission, plasticity, and network function: lessons from genetic mouse models.. Exp Brain Res.
- Russell, C.L.; Semerdjieva, S.; Empson, R.M.; Austen, B.M.; Beesley, P.W.; Alifragis, P. Amyloid-β Acts as a Regulator of Neurotransmitter Release Disrupting the Interaction between Synaptophysin and VAMP2. PLOS ONE 2012, 7, e43201. [Google Scholar] [CrossRef]
- Allinson, T. , Parkin, ET, Turner, AJ, Hooper, NM.. (2003) ADAMs family members as amyloid precursor proteinα-secretases. J Neurosci Res. 74.
- Willem, M.; Tahirovic, S.; Busche, M.A.; Ovsepian, S.V.; Chafai, M.; Kootar, S.; Hornburg, D.; Evans, L.D.B.; Moore, S.C.; Daria, A.; et al. η-Secretase processing of APP inhibits neuronal activity in the hippocampus. Nature 2015, 526, 443–447. [Google Scholar] [CrossRef]
- Andrew, R.J.; Kellett, K.A.; Thinakaran, G.; Hooper, N.M. A Greek Tragedy: The Growing Complexity of Alzheimer Amyloid Precursor Protein Proteolysis. J. Biol. Chem. 2016, 291, 19235–19244. [Google Scholar] [CrossRef]
- Gabriele, R.M.C.; Abel, E.; Fox, N.C.; Wray, S.; Arber, C. Knockdown of Amyloid Precursor Protein: Biological Consequences and Clinical Opportunities. Front. Neurosci. 2022, 16, 835645. [Google Scholar] [CrossRef]
- Hampel, H. , Hu, Y, Hardy, J, Blennow, K, Chen, C, Perry, G, Kim, SH, Villemagne, VL, Aisen, P, Vendruscolo, M, Iwatsubo, T, Masters, CL, Cho, M, Lannfelt,L, Cummings, JL, Vergallo, A. T. (2023) he amyloid-β pathway in Alzheimer's disease: a plain language summary.. Neurodegener Dis Manag. 13.
- Chen, G. , Xu, TH, Yan, Y, Zhou, YR, Jiang, Y, Melcher, K, Xu, HE. (2017) Amyloid beta: structure, biology y and structure-based therapeutic development.. Acta Pharmacol Sin. 1205; 38. [Google Scholar]
- Rambaran, R. , Serpell, LC.. (2008) Amyloid fibrils: abnormal protein assembly.. Prion. 2.
- Sosa, L.J.; Cáceres, A.; Dupraz, S.; Oksdath, M.; Quiroga, S.; Lorenzo, A. The physiological role of the amyloid precursor protein as an adhesion molecule in the developing nervous system. J. Neurochem. 2017, 143, 11–29. [Google Scholar] [CrossRef]
- Al-Kuraishy, H. , Jabir,MS, Al-Gareeb, AI, Albuhadily, AK, Albukhaty, S, Sulaiman, GM, Batiha, GE.. (2023) Evaluation and targeting of amyloid precursor protein (APP)/amyloid beta (Aβ) axis in amyloidogenic and non-amyloidogenic pathways: A time outside the tunnel.. Ageing Res Rev. 92, 102119. [CrossRef]
- Orobets, K. , Karamyshev, AL.. (2023) Amyloid Precursor Protein and Alzheimer's Disease.. Int J Mol Sci. 24, 14794. [CrossRef]
- Kuo, C.-C.; Chiang, A.W.T.; Baghdassarian, H.M.; Lewis, N.E. Dysregulation of the secretory pathway connects Alzheimer’s disease genetics to aggregate formation. Cell Syst. 2021. [Google Scholar] [CrossRef]
- Sun, J.; Roy, S. The physical approximation of APP and BACE-1: A key event in alzheimer's disease pathogenesis. Dev. Neurobiol. 2017, 78, 340–347. [Google Scholar] [CrossRef]
- Tackenberg, C. , Nitsch, RM.. (2019) The secreted APP ectodomain sAPPα, but not sAPPβ, protects neurons against Aβ oligomer-induced dendritic spine loss and increased tau phosphorylation.. Mol Brain. 12, 27. [CrossRef]
- Baratchi, S.; Evans, J.; Tate, W.P.; Abraham, W.C.; Connor, B. Secreted amyloid precursor proteins promote proliferation and glial differentiation of adult hippocampal neural progenitor cells. Hippocampus 2011, 22, 1517–1527. [Google Scholar] [CrossRef]
- Dar, N.J.; Glazner, G.W. Deciphering the neuroprotective and neurogenic potential of soluble amyloid precursor protein alpha (sAPPα). Cell. Mol. Life Sci. 2020, 77, 2315–2330. [Google Scholar] [CrossRef]
- Habib, A.; Sawmiller, D.; Tan, J. Restoring Soluble Amyloid Precursor Protein α Functions as a Potential Treatment for Alzheimer's Disease. J. Neurosci. Res. 2016, 95, 973–991. [Google Scholar] [CrossRef]
- Castro, M.A.; Hadziselimovic, A.; Sanders, C.R. The vexing complexity of the amyloidogenic pathway. Protein Sci. 2019, 28, 1177–1193. [Google Scholar] [CrossRef]
- Nalivaeva, N. , Turner, AJ.. (2013) The amyloid precursor protein: a biochemical enigma in brain development, function and disease.. FEBS Lett. 587, 2046-2054. [CrossRef]
- Chasseigneaux, S.; Allinquant, B. Functions of Aβ, sAPPα and sAPPβ: similarities and differences. J. Neurochem. 2011, 120, 99–108. [Google Scholar] [CrossRef]
- Proft, J. , Weiss, N.. (2012) Jekyll and Hide, The two faces of amyloid β. 5.
- Frohman, E.M.; Racke, M.K.; Raine, C.S. Multiple Sclerosis — The Plaque and Its Pathogenesis. New Engl. J. Med. 2006, 354, 942–955. [Google Scholar] [CrossRef]
- Grant, J. , Ghosn, EE, Axtell, RC, Herges, K, Kuipers, HF, Woodling, NS, et al.. (2012) Reversal of paralysis and reduced inflammation from peripheral administration of beta-amyloid in TH1 and TH17 versions of experimental autoimmune encephalomyelitis. 4.
- Trapp, B.D.; Peterson, J.; Ransohoff, R.M.; Rudick, R.; Mörk, S.; Bö, L. Axonal Transection in the Lesions of Multiple Sclerosis. New Engl. J. Med. 1998, 338, 278–285. [Google Scholar] [CrossRef]
- Ferguson, B.; Matyszak, M.K.; Esiri, M.M.; Perry, V.H. Axonal damage in acute multiple sclerosis lesions. Brain 1997, 120, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Simard, A.R.; Soulet, D.; Gowing, G.; Julien, J.-P.; Rivest, S. Bone Marrow-Derived Microglia Play a Critical Role in Restricting Senile Plaque Formation in Alzheimer's Disease. Neuron 2006, 49, 489–502. [Google Scholar] [CrossRef] [PubMed]
- Itagaki, S.; McGeer, P.; Akiyama, H.; Zhu, S.; Selkoe, D. Relationship of microglia and astrocytes to amyloid deposits of Alzheimer disease. J. Neuroimmunol. 1989, 24, 173–182. [Google Scholar] [CrossRef]
- Li, X. , Feng, X, Sun, X, Hou, N, Han, F, Liu, Y.. (2022) Global, regional, and national burden of Alzheimer's disease and other dementias, 1990-2019. Front Aging Neurosci. 9: 14, 9374; 14. [Google Scholar]
- Gillet, J.-N. From molecular dynamics to quantum mechanics of misfolded proteins and amyloid-like macroaggregates applied to neurodegenerative diseases. J. Mol. Graph. Model. 2021, 110, 108046. [Google Scholar] [CrossRef]
- Sohail, A.; Ashiq, U. Quantum inspired improved AI computing for the sensors of cardiac mechano-biology. Sensors Int. 2023, 4. [Google Scholar] [CrossRef]
- De la Paz, M. , Serrano, L. . ( 2004) Sequence deteminants of amyloid fibril formation.. Proc Natl Acad Sci USA.. 101, 87–92.
- DuBay, K.F.; Pawar, A.P.; Chiti, F.; Zurdo, J.; Dobson, C.M.; Vendruscolo, M. Prediction of the Absolute Aggregation Rates of Amyloidogenic Polypeptide Chains. J. Mol. Biol. 2004, 341, 1317–1326. [Google Scholar] [CrossRef]
- Pawar, A. , Dubay, KF, Zurdo, J, et al.. (2005) Prediction of “aggregation-prone” and “aggregation-susceptible” regions in proteins associated with neurodegenerative disease.. J Mol Biol.
- Zibaee, S. , Makin, OS, Goedert, M, et al.. (2007) A simple algorithm locates beta-strands in the amyloid fibril core of alpha-synuclein, Abeta and tau using the amino acid sequence alone. 16.
- Fernandez-Escamilla, A.-M.; Rousseau, F.; Schymkowitz, J.; Serrano, L. Prediction of sequence-dependent and mutational effects on the aggregation of peptides and proteins. Nat. Biotechnol. 2004, 22, 1302–1306. [Google Scholar] [CrossRef]
- Palato, L.M.; Pilcher, S.; Oakes, A.; Lamba, A.; Torres, J.; Monjaraz, L.I.L.; Munoz, C.; Njoo, E.; Rinauro, D.J.; Menefee, K.A.; et al. Amyloidogenicity of naturally occurring full-length animal IAPP variants. J. Pept. Sci. 2019, 25, e3199–e3199. [Google Scholar] [CrossRef]
- Conchillo-Solé, O.; de Groot, N.S.; Avilés, F.X.; Vendrell, J.; Daura, X.; Ventura, S. AGGRESCAN: a server for the prediction and evaluation of "hot spots" of aggregation in polypeptides. BMC Bioinform. 2007, 8, 65–65. [Google Scholar] [CrossRef] [PubMed]
- Castillo, V.; Graña-Montes, R.; Sabate, R.; Ventura, S. Prediction of the aggregation propensity of proteins from the primary sequence: Aggregation properties of proteomes. Biotechnol. J. 2011, 6, 674–685. [Google Scholar] [CrossRef] [PubMed]
- de Groot, N. , Castillo, V, Graña-Montes, R, Ventura, S.. (2012) AGGRESCAN: method, application, and perspectives for drug design.. Methods Mol Biol.
- Tsolis, A. , Papandreou, NC, Iconomidou, VA, Hamodrakas, SJ.. (2013) A consensus method for the prediction of 'aggregation-prone' peptides in globular proteins.. PLoS One. 5417; 8. [Google Scholar]
- Cherny, I.; Gazit, E. Amyloids: Not Only Pathological Agents but Also Ordered Nanomaterials. Angew. Chem. Int. Ed. 2008, 47, 4062–4069. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Chen, H.; Lai, L. Identification of amyloid fibril-forming segments based on structure and residue-based statistical potential. Bioinformatics 2007, 23, 2218–2225. [Google Scholar] [CrossRef] [PubMed]
- Bondarev, S.A.; Uspenskaya, M.V.; Leclercq, J.; Falgarone, T.; Zhouravleva, G.A.; Kajava, A.V. AmyloComp: a bioinformatic tool for prediction of amyloid co-aggregation. J. Mol. Biol. 2024, 168437. [Google Scholar] [CrossRef] [PubMed]
- Wojciechowski, J.W.; Szczurek, W.; Szulc, N.; Szefczyk, M.; Kotulska, M. PACT - Prediction of amyloid cross-interaction by threading. Sci. Rep. 2023, 13, 1–10. [Google Scholar] [CrossRef]
- Emily, M.; Talvas, A.; Delamarche, C. MetAmyl: A METa-Predictor for AMYLoid Proteins. PLOS ONE 2013, 8, e79722. [Google Scholar] [CrossRef] [PubMed]
- Charoenkwan, P. , Kanthawong, S, Nantasenamat, C, Hasan, MM, Shoombuatong, W.. (2021) iAMY-SCM: Improved prediction and analysis of amyloid proteins using a scoring card method with propensity scores of dipeptides.. Genomics.
- Niu, M.; Li, Y.; Wang, C.; Han, K. RFAmyloid: A Web Server for Predicting Amyloid Proteins. Int. J. Mol. Sci. 2018, 19, 2071. [Google Scholar] [CrossRef] [PubMed]
- Conchillo-Solé, O.; de Groot, N.S.; Avilés, F.X.; Vendrell, J.; Daura, X.; Ventura, S. AGGRESCAN: a server for the prediction and evaluation of "hot spots" of aggregation in polypeptides. BMC Bioinform. 2007, 8, 65–65. [Google Scholar] [CrossRef]
- Hamodrakas, S.J. Protein aggregation and amyloid fibril formation prediction software from primary sequence: towards controlling the formation of bacterial inclusion bodies. FEBS J. 2011, 278, 2428–2435. [Google Scholar] [CrossRef]
- Charoenkwan, P. , Ahmed, S, Nantasenamat, C, Quinn, JMW, Moni, MA, Lio,' P, Shoombuatong, W.. (2022) AMYPred-FRL is a novel approach for accurate prediction of amyloid proteins by using feature representation learning.. Sci Rep. 7697; 12. [Google Scholar]
- Nair, S.S.K.; Reddy, N.S.; Hareesha, K. AmylPepPred: Amyloidogenic Peptide Prediction tool. Bioinformation 2012, 8, 994–995. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Wu, N.; Guo, J.; Fan, Y. Prediction of amyloid fibril-forming segments based on a support vector machine. BMC Bioinform. 2009, 10, S45–S45. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Robang, A.S.; Sarma, S.; Le, J.V.; E Helmicki, M.; Lambert, M.J.; Guerrero-Ferreira, R.; Arboleda-Echavarria, J.; Paravastu, A.K.; Hall, C.K. Sequence patterns and signatures: Computational and experimental discovery of amyloid-forming peptides. PNAS Nexus 2022, 1, pgac263. [Google Scholar] [CrossRef]
- Morris, K. , Rodger, A, Hicks, MR, Debulpaep, M, Schymkowitz, J, Rousseau, F, Serpel, LC.. (2013) Exploring the sequence-structure relationship for amyloid peptides.
- López de la Paz, M. , Serrano, L. . ( 2004) Sequence determinants of amyloid fibril formation.. Proc Natl Acad Sci U S A. 101, 87–92.
- Al-Garawi, Z.S.; Morris, K.L.; Marshall, K.E.; Eichler, J.; Serpell, L.C. The diversity and utility of amyloid fibrils formed by short amyloidogenic peptides. Interface Focus 2017, 7, 20170027–20170027. [Google Scholar] [CrossRef] [PubMed]
- Thompson, M.J.; Sievers, S.A.; Karanicolas, J.; Ivanova, M.I.; Baker, D.; Eisenberg, D. The 3D profile method for identifying fibril-forming segments of proteins. Proc. Natl. Acad. Sci. 2006, 103, 4074–4078. [Google Scholar] [CrossRef]
- Galzitskaya, O. , Garbuzynskiy, SO, Lobanov, MY. . ( 2006) Prediction of amyloidogenic and disordered regions in protein chains.. PLoS Comput Biol. 2, e177.
- Família, C. , Dennison, SR, Quintas, A, Phoenix, DA. . ( 2015) Prediction of Peptide and Protein Propensity for Amyloid Formation.. PLoS One. 10, e0134679.
- Belli, M. , Ramazzotti, M, Chiti, F. . ( 2011) Prediction of amyloid aggregation in vivo.. EMBO Rep. 12, 657–663.
- Abdelrahman, S.; Alghrably, M.; Lachowicz, J.I.; Emwas, A.-H.; Hauser, C.A.E.; Jaremko, M. “What Doesn’t Kill You Makes You Stronger”: Future Applications of Amyloid Aggregates in Biomedicine. Molecules 2020, 25, 5245. [Google Scholar] [CrossRef]
- Xiao, X. , Wang, Y, Seroski, DT, Wong, KM, Liu, R, Paravastu, AK, Hudalla, GA, Hall, CK.. (2021) De novo design of peptides that coassemble into β sheet-based nanofibrils.. Sci Adv. 7668; 7. [Google Scholar]
- Cherny, I.; Gazit, E. Amyloids: Not Only Pathological Agents but Also Ordered Nanomaterials. Angew. Chem. Int. Ed. 2008, 47, 4062–4069. [Google Scholar] [CrossRef] [PubMed]
- Soto, C.; Pritzkow, S. Protein misfolding, aggregation, and conformational strains in neurodegenerative diseases. Nat. Neurosci. 2018, 21, 1332–1340. [Google Scholar] [CrossRef] [PubMed]
- Herczenik, E.; Gebbink, M.F.B.G. Molecular and cellular aspects of protein misfolding and disease. FASEB J. 2008, 22, 2115–2133. [Google Scholar] [CrossRef] [PubMed]
- Rao, R.V.; E Bredesen, D. Misfolded proteins, endoplasmic reticulum stress and neurodegeneration. Curr. Opin. Cell Biol. 2004, 16, 653–662. [Google Scholar] [CrossRef] [PubMed]
- Selkoe, D. (2003) Folding proteins in fatal ways.. Nature.
- Taylor, J.P.; Hardy, J.; Fischbeck, K.H. Toxic Proteins in Neurodegenerative Disease. Science 2002, 296, 1991–1995. [Google Scholar] [CrossRef] [PubMed]
- Kopito, R. , Ron, D. . ( 2000) Conformational disease.. Nat Cell Biol. 2, E207–E209.
- Kopito, R.R. Aggresomes, inclusion bodies and protein aggregation. Trends Cell Biol. 2000, 10, 524–530. [Google Scholar] [CrossRef] [PubMed]
- Dobson, C.M.; Ellis, R.J. Protein folding and misfolding inside and outside the cell. EMBO J. 1998, 17, 5251–5254. [Google Scholar] [CrossRef] [PubMed]
- Dobson, C. (2003) Protein folding and misfolding.. Nature.
- Kaufman, R.J. Stress signaling from the lumen of the endoplasmic reticulum: coordination of gene transcriptional and translational controls. Genes Dev. 1999, 13, 1211–1233. [Google Scholar] [CrossRef]
- Roan, N.R.; Müller, J.A.; Liu, H.; Chu, S.; Arnold, F.; Stürzel, C.M.; Walther, P.; Dong, M.; Witkowska, H.E.; Kirchhoff, F.; et al. Peptides Released by Physiological Cleavage of Semen Coagulum Proteins Form Amyloids that Enhance HIV Infection. Cell Host Microbe 2011, 10, 541–550. [Google Scholar] [CrossRef]
- Tennent, G. , Lovat, LB, Pepys, MB (1995) Serum amyloid P component prevents proteolysis of the amyloid fibrils of Alzheimer disease and systemic amyloidosis.. PNAS USA. 4299; 92. [Google Scholar]
- Goldberg, A.L. Protein degradation and protection against misfolded or damaged proteins. Nature 2003, 426, 895–899. [Google Scholar] [CrossRef] [PubMed]
- Sitia, R.; Braakman, I. Quality control in the endoplasmic reticulum protein factory. Nature 2003, 426, 891–894. [Google Scholar] [CrossRef] [PubMed]
- Sherman, M. , Goldberg, AL.. (2001) Cellular defenses against unfolded proteins: a cell biologist thinks about neurodegenerative diseases.. Neuron. 29.
- Mostaert, A.S.; Crockett, R.; Kearn, G.; Cherny, I.; Gazit, E.; Serpell, L.C.; Jarvis, S.P. Mechanically functional amyloid fibrils in the adhesive of a marine invertebrate as revealed by Raman spectroscopy and atomic force microscopy. Arch. Histol. Cytol. 2009, 72, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Guyonnet, B.; Egge, N.; Cornwall, G.A.; Ghule, P.N.; Xie, R.-L.; Medina, R.; Colby, J.L.; Jones, S.N.; Lian, J.B.; Stein, J.L.; et al. Functional Amyloids in the Mouse Sperm Acrosome. Mol. Cell. Biol. 2014, 34, 2624–2634. [Google Scholar] [CrossRef] [PubMed]
- Whelly, S.; Johnson, S.; Powell, J.; Borchardt, C.; Hastert, M.C.; Cornwall, G.A. Nonpathological Extracellular Amyloid Is Present during Normal Epididymal Sperm Maturation. PLOS ONE 2012, 7, e36394. [Google Scholar] [CrossRef] [PubMed]
- Whelly, S.; Muthusubramanian, A.; Powell, J.; Johnson, S.; Hastert, M.C.; Cornwall, G.A. Cystatin-related epididymal spermatogenic subgroup members are part of an amyloid matrix and associated with extracellular vesicles in the mouse epididymal lumen. Mol. Hum. Reprod. 2016, 22, 729–744. [Google Scholar] [CrossRef] [PubMed]
- Guyonnet, B.; Egge, N.; Cornwall, G.A.; Ghule, P.N.; Xie, R.-L.; Medina, R.; Colby, J.L.; Jones, S.N.; Lian, J.B.; Stein, J.L.; et al. Functional Amyloids in the Mouse Sperm Acrosome. Mol. Cell. Biol. 2014, 34, 2624–2634. [Google Scholar] [CrossRef] [PubMed]
- Sood, R. , Domanov, Y, Pietiainen, M, Kontinen, VP, Kinnunen, PKJ.. (2008) Binding of LL-37 to model biomembranes: Insight into target vs. host cell recognition.. Biochem Biophys Acta. 1778. [Google Scholar]
- Maji, S.K.; Perrin, M.H.; Sawaya, M.R.; Jessberger, S.; Vadodaria, K.; Rissman, R.A.; Singru, P.S.; Nilsson, K.P.R.; Simon, R.; Schubert, D.; et al. Functional Amyloids As Natural Storage of Peptide Hormones in Pituitary Secretory Granules. Science 2009, 325, 328–332. [Google Scholar] [CrossRef] [PubMed]
- Fowler, D.M.; Koulov, A.V.; Alory-Jost, C.; Marks, M.S.; E Balch, W.; Kelly, J.W. Functional Amyloid Formation within Mammalian Tissue. PLOS Biol. 2005, 4, e6. [Google Scholar] [CrossRef]
- Münch, J.; Rücker, E.; Ständker, L.; Adermann, K.; Goffinet, C.; Schindler, M.; Wildum, S.; Chinnadurai, R.; Rajan, D.; Specht, A.; et al. Semen-Derived Amyloid Fibrils Drastically Enhance HIV Infection. Cell 2007, 131, 1059–1071. [Google Scholar] [CrossRef]
- Jang, H.; Arce, F.T.; Mustata, M.; Ramachandran, S.; Capone, R.; Nussinov, R.; Lal, R. Antimicrobial Protegrin-1 Forms Amyloid-Like Fibrils with Rapid Kinetics Suggesting a Functional Link. Biophys. J. 2011, 100, 1775–1783. [Google Scholar] [CrossRef] [PubMed]
- Li, J. , McQuade, T, Siemer, AB, Napetschnig, J, Moriwaki, K, Hsiao, YS, Damko, E, Moquin, D, Walz, T, McDermott, A, et al.. (2012) The RIP1/RIP3 necrosome forms a functional amyloid signaling complex required for programmed necrosis.. Cell.
- Audas, T.E.; Audas, D.E.; Jacob, M.D.; Ho, J.D.; Khacho, M.; Wang, M.; Perera, J.K.; Gardiner, C.; Bennett, C.A.; Head, T.; et al. Adaptation to Stressors by Systemic Protein Amyloidogenesis. Dev. Cell 2016, 39, 155–168. [Google Scholar] [CrossRef] [PubMed]
- Melrose, J. High Performance Marine and Terrestrial Bioadhesives and the Biomedical Applications They Have Inspired. Molecules 2022, 27, 8982. [Google Scholar] [CrossRef] [PubMed]
- Buchanan, J.A.; Varghese, N.R.; Johnston, C.L.; Sunde, M. Functional Amyloids: Where Supramolecular Amyloid Assembly Controls Biological Activity or Generates New Functionality. J. Mol. Biol. 2023, 435, 167919. [Google Scholar] [CrossRef] [PubMed]
- Reches, M.; Gazit, E. Molecular Self-Assembly of Peptide Nanostructures: Mechanism of Association and Potential Uses. Curr. Nanosci. 2006, 2, 105–111. [Google Scholar] [CrossRef]
- Adler-Abramovich, L.; Aronov, D.; Beker, P.; Yevnin, M.; Stempler, S.; Buzhansky, L.; Rosenman, G.; Gazit, E. Self-assembled arrays of peptide nanotubes by vapour deposition. Nat. Nanotechnol. 2009, 4, 849–854. [Google Scholar] [CrossRef]
- Carny, O.; Shalev, D.E.; Gazit, E. Fabrication of Coaxial Metal Nanocables Using a Self-Assembled Peptide Nanotube Scaffold. Nano Lett. 2006, 6, 1594–1597. [Google Scholar] [CrossRef] [PubMed]
- Reches, M.; Gazit, E. Casting Metal Nanowires Within Discrete Self-Assembled Peptide Nanotubes. Science 2003, 300, 625–627. [Google Scholar] [CrossRef]
- Reches, M.; Gazit, E. Controlled patterning of aligned self-assembled peptide nanotubes. Nat. Nanotechnol. 2006, 1, 195–200. [Google Scholar] [CrossRef]
- Tiwari, O. , Gazit, E.. (2024) Characterization of amyloid-like metal-amino acid assemblies with remarkable catalytic activity.. Methods Enzymol. 697, 181-209. [CrossRef]
- Gazit, E. Self-assembled peptide nanostructures: the design of molecular building blocks and their technological utilization. Chem. Soc. Rev. 2007, 36, 1263–1269. [Google Scholar] [CrossRef]
- Hauser, C.A.E.; Maurer-Stroh, S.; Martins, I.C. Amyloid-based nanosensors and nanodevices. Chem. Soc. Rev. 2014, 43, 5326–5345. [Google Scholar] [CrossRef] [PubMed]
- Taheri, R.A.; Akhtari, Y.; Moghadam, T.T.; Ranjbar, B. Assembly of Gold Nanorods on HSA Amyloid Fibrils to Develop a Conductive Nanoscaffold for Potential Biomedical and Biosensing Applications. Sci. Rep. 2018, 8, 9333. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Joshi, S.; Savel’ev, S.E.; Jiang, H.; Midya, R.; Lin, P.; Hu, M.; Ge, N.; Strachan, J.P.; Li, Z.; et al. Memristors with diffusive dynamics as synaptic emulators for neuromorphic computing. Nat. Mater. 2016, 16, 101–108. [Google Scholar] [CrossRef]
- Miranda, E.; Suñé, J. Memristors for Neuromorphic Circuits and Artificial Intelligence Applications. Materials 2020, 13, 938. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Wang, J.; He, X.; Ming, J.; Wang, L.; Wang, Y.; Shao, H.; Zheng, C.; Xie, L.; Ling, H. Pseudo-transistors for emerging neuromorphic electronics. Sci. Technol. Adv. Mater. 2023, 24, 2180286. [Google Scholar] [CrossRef]
- Barraj, I.; Mestiri, H.; Masmoudi, M. Overview of Memristor-Based Design for Analog Applications. Micromachines 2024, 15, 505. [Google Scholar] [CrossRef] [PubMed]
- Jariwala, D. , Sangwan, VK, Lauhon, LJ, Marks, TJ, Hersam, MC. . ( and sensing.. Chem Soc Rev. 42, 2824–2860.
- Li, C.; Mezzenga, R. The interplay between carbon nanomaterials and amyloid fibrils in bio-nanotechnology. Nanoscale 2013, 5, 6207–6218. [Google Scholar] [CrossRef]
- Terrones, H.; Terrones, M.; López–Urías, F.; Rodríguez–Manzo, J.A.; Mackay, A.L. Shape and complexity at the atomic scale: the case of layered nanomaterials. Philos. Trans. R. Soc. A: Math. Phys. Eng. Sci. 2004, 362, 2039–2063. [Google Scholar] [CrossRef]
- Li, C.; Mezzenga, R. The interplay between carbon nanomaterials and amyloid fibrils in bio-nanotechnology. Nanoscale 2013, 5, 6207–6218. [Google Scholar] [CrossRef]
- Mankar, S.; Anoop, A.; Sen, S.; Maji, S.K. Nanomaterials: amyloids reflect their brighter side. Nano Rev. 2011, 2. [Google Scholar] [CrossRef]
- Ramanishankar, A.; S, A.S.; Begum, R.F.; Jayasankar, N.; Nayeem, A.; Prajapati, B.G.; Nirenjen, S. Unleashing light's healing power: an overview of photobiomodulation for Alzheimer's treatment. Futur. Sci. OA 2024, 10, FSO922. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Chang, Y.; Liu, L.; Wang, J. Nanomaterials for Modulating the Aggregation of β-Amyloid Peptides. Molecules 2021, 26, 4301. [Google Scholar] [CrossRef] [PubMed]
- Ender, A.M.; Kaygisiz, K.; Räder, H.-J.; Mayer, F.J.; Synatschke, C.V.; Weil, T. Cell-Instructive Surface Gradients of Photoresponsive Amyloid-like Fibrils. ACS Biomater. Sci. Eng. 2021, 7, 4798–4808. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, M.R.; Dobson, C.M. Chemical modification of insulin in amyloid fibrils. Protein Sci. 2003, 12, 2637–2641. [Google Scholar] [CrossRef] [PubMed]
- Morris, R. , MacPhee, C. (2013) Amyloid Protein Biomaterials. In: Roberts, G.C.K. (eds) Encyclopedia of Biophysics. Springer, Berlin, Heidelberg. [CrossRef]
- Vaziri, S.; Fazilati, M.; Arasteh, A.; Nazem, H. Amyloid Nano-biofibrils as a New Nano-Scaffold for Lipase Immobilization. Protein Pept. Lett. 2018, 25, 862–870. [Google Scholar] [CrossRef] [PubMed]
- Pilkington, S.M.; Roberts, S.J.; Meade, S.J.; Gerrard, J.A. Amyloid fibrils as a nanoscaffold for enzyme immobilization. Biotechnol. Prog. 2009, 26, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Wittung-Stafshede, P. Chemical catalysis by biological amyloids. Biochem. Soc. Trans. 2023, 51, 1967–1974. [Google Scholar] [CrossRef]
- Fowler, D. , Koulov, AV, Balch, WE, Kelly, JW. . ( 2007) Functional amyloid from bacteria to humans.. Trends Biochem Sci. 32, 217–224.
- Gras, S.L.; Tickler, A.K.; Squires, A.M.; Devlin, G.L.; Horton, M.A.; Dobson, C.M.; MacPhee, C.E. Functionalised amyloid fibrils for roles in cell adhesion. Biomaterials 2007, 29, 1553–1562. [Google Scholar] [CrossRef]
- Maji, S.K.; Schubert, D.; Rivier, C.; Lee, S.; Rivier, J.E.; Riek, R. Amyloid as a Depot for the Formulation of Long-Acting Drugs. PLoS Biol. 2008, 6, e17. [Google Scholar] [CrossRef]
- Pertinhez, T.A.; Conti, S.; Ferrari, E.; Magliani, W.; Spisni, A.; Polonelli, L. Reversible Self-Assembly: A Key Feature for a New Class of Autodelivering Therapeutic Peptides. Mol. Pharm. 2009, 6, 1036–1039. [Google Scholar] [CrossRef]
- Scheibel, T.; Parthasarathy, R.; Sawicki, G.; Lin, X.-M.; Jaeger, H.; Lindquist, S.L. Conducting nanowires built by controlled self-assembly of amyloid fibers and selective metal deposition. Proc. Natl. Acad. Sci. USA 2003, 100, 4527–4532. [Google Scholar] [CrossRef] [PubMed]
- Bhak, G. , Lee, S, Park, JW, Cho, S, Paik, SR.. (2010) Amyloid hydrogel derived from curly protein fibrils of alpha-synuclein.. Biomaterials. 5986; 31. [Google Scholar]
- Herland, A.; Björk, P.; Nilsson, K.P.R.; Olsson, J.D.M.; Åsberg, P.; Konradsson, P.; Hammarström, P.; Inganäs, O. Electroactive Luminescent Self-Assembled Bio-organic Nanowires: Integration of Semiconducting Oligoelectrolytes within Amyloidogenic Proteins. Adv. Mater. 2005, 17, 1466–1471. [Google Scholar] [CrossRef]
- Tanaka, H.; Herland, A.; Lindgren, L.J.; Tsutsui, T.; Andersson, M.R.; Inganäs, O. Enhanced Current Efficiency from Bio-Organic Light-Emitting Diodes Using Decorated Amyloid Fibrils with Conjugated Polymer. Nano Lett. 2008, 8, 2858–2861. [Google Scholar] [CrossRef]
- Herland, A.; Thomsson, D.; Mirzov, O.; Scheblykin, I.G.; Inganäs, O. Decoration of amyloid fibrils with luminescent conjugated polymers. J. Mater. Chem. 2007, 18, 126–132. [Google Scholar] [CrossRef]
- Hamedi, M.; Herland, A.; Karlsson, R.H.; Inganäs, O. Electrochemical Devices Made from Conducting Nanowire Networks Self-Assembled from Amyloid Fibrils and Alkoxysulfonate PEDOT. Nano Lett. 2008, 8, 1736–1740. [Google Scholar] [CrossRef] [PubMed]
- Ahn, M. , Kang, S, Koo, HJ, Lee, JH, Lee, YS, Paik, SR.. (2010) Nanoporous protein matrix made of amyloid fibrils of beta2-microglobulin.. Biotechnol Prog. 1759; 26. [Google Scholar]
- Preat, T.; Goguel, V. Role of Drosophila Amyloid Precursor Protein in Memory Formation. Front. Mol. Neurosci. 2016, 9, 142. [Google Scholar] [CrossRef]
- Evans, M.L.; Chapman, M.R. Curli biogenesis: Order out of disorder. Biochim. et Biophys. Acta (BBA) - Mol. Cell Res. 2014, 1843, 1551–1558. [Google Scholar] [CrossRef]
- Bhoite, S.; van Gerven, N.; Chapman, M.R.; Remaut, H. Curli Biogenesis: Bacterial Amyloid Assembly by the Type VIII Secretion Pathway. EcoSal Plus 2019, 8. [Google Scholar] [CrossRef]
- Klauck, G. , Serra, DO, Possling, A, Hengge, R.. (2018) Spatial Organization of Different Sigma Factor Activities and C-Di-GMP Signalling within the Three-Dimensional Landscape of a Bacterial Biofilm.. Open Biol. 8. [CrossRef]
- Ulamec, S.M.; Radford, S.E. Spot the Difference: Function versus Toxicity in Amyloid Fibrils. Trends Biochem. Sci. 2020, 45, 635–636. [Google Scholar] [CrossRef] [PubMed]
- Hervas, R.; Rau, M.J.; Park, Y.; Zhang, W.; Murzin, A.G.; Fitzpatrick, J.A.J.; Scheres, S.H.W.; Si, K. Cryo-EM structure of a neuronal functional amyloid implicated in memory persistence in Drosophila. Science 2020, 367, 1230–1234. [Google Scholar] [CrossRef]
- Goguel, V.; Belair, A.-L.; Ayaz, D.; Lampin-Saint-Amaux, A.; Scaplehorn, N.; Hassan, B.A.; Preat, T. DrosophilaAmyloid Precursor Protein-Like Is Required for Long-Term Memory. J. Neurosci. 2011, 31, 1032–1037. [Google Scholar] [CrossRef]
- Hervás, R. , Li, L, Majumdar, A, Fernández-Ramírez Mdel, C, Unruh, JR, Slaughter, BD, Galera-Prat, A, Santana, E, Suzuki, M, Nagai, Y, Bruix, M, Casas-Tintó, S, Menéndez, M, Laurents, DV, Si, K, Carrión-Vázquez, M. . ( 2016) Molecular Basis of Orb2 Amyloidogenesis and Blockade of Memory Consolidation.. PLoS Biol. 14, e1002361.
- Hervás, R.; Murzin, A.G.; Si, K. Implications of the Orb2 Amyloid Structure in Huntington’s Disease. Int. J. Mol. Sci. 2020, 21, 6910. [Google Scholar] [CrossRef]
- Garcia-Pardo, J.; Ventura, S. Cryo-EM structures of functional and pathological amyloid ribonucleoprotein assemblies. Trends Biochem. Sci. 2024, 49, 119–133. [Google Scholar] [CrossRef] [PubMed]
- Bartolomé-Nafría, A.; García-Pardo, J.; Ventura, S. Mutations in human prion-like domains: pathogenic but not always amyloidogenic. Prion 2024, 18, 28–39. [Google Scholar] [CrossRef] [PubMed]
- Keene, J.D. Ribonucleoprotein infrastructure regulating the flow of genetic information between the genome and the proteome. Proc. Natl. Acad. Sci. 2001, 98, 7018–7024. [Google Scholar] [CrossRef]
- Si, K.; Kandel, E.R. The Role of Functional Prion-Like Proteins in the Persistence of Memory. Cold Spring Harb. Perspect. Biol. 2016, 8, a021774. [Google Scholar] [CrossRef]
- Kozlov, E.; Shidlovskii, Y.V.; Gilmutdinov, R.; Schedl, P.; Zhukova, M. The role of CPEB family proteins in the nervous system function in the norm and pathology. Cell Biosci. 2021, 11, 1–14. [Google Scholar] [CrossRef]
- Li, X.; Li, S.; Huang, X.; Chen, Y.; Cheng, J.; Zhan, A. Protein-mediated bioadhesion in marine organisms: A review. Mar. Environ. Res. 2021, 170, 105409. [Google Scholar] [CrossRef] [PubMed]
- Balkenende, D.W.; Winkler, S.M.; Messersmith, P.B. Marine-inspired polymers in medical adhesion. Eur. Polym. J. 2019, 116, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Ninan, L.; Monahan, J.; Stroshine, R.L.; Wilker, J.J.; Shi, R. Adhesive strength of marine mussel extracts on porcine skin. Biomaterials 2003, 24, 4091–4099. [Google Scholar] [CrossRef] [PubMed]
- Lang, N.; Pereira, M.J.; Lee, Y.; Friehs, I.; Vasilyev, N.V.; Feins, E.N.; Ablasser, K.; O’cearbhaill, E.D.; Xu, C.; Fabozzo, A.; et al. A Blood-Resistant Surgical Glue for Minimally Invasive Repair of Vessels and Heart Defects. Sci. Transl. Med. 2014, 6, 218ra6–218ra6. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, T. , Paul, S.. (2006) Protein-misfolding diseases and chaperone-based therapeutic approaches.. FEBS J. 1331. [Google Scholar]
- Cathcart, E.S.; Shirahama, T.; Cohen, A.S. Isolation and identification of a plasma component of amyloid. Biochim. et Biophys. Acta (BBA) - Protein Struct. 1967, 147, 392–393. [Google Scholar] [CrossRef]
- Emsley, J. , White, HE, O'Hara, BP, Oliva, G, Srinivasan, N, Tickle, IJ, Blundell, TL, Pepys, MB, Wood (1994) Structure of pentameric human serum amyloid P component.. Nature.
- Pepys, M. , Booth, DR, Hutchinson, WL, Gallimore, JR, Collins, PM, Hohenester, E (1997) Amyloid P component. A critical review.. Amyloid. 4.
- Botto, M.; Hawkins, P.N.; Bickerstaff, M.C.; Herbert, J.; Bygrave, A.E.; Mcbride, A.; Hutchinson, W.L.; Tennent, G.A.; Walportz, M.J.; Pepys, M.B. Amyloid deposition is delayed in mice with targeted deletion of the serum amyloid P component gene. Nat. Med. 1997, 3, 855–859. [Google Scholar] [CrossRef] [PubMed]
- Sproston, N.R.; Ashworth, J.J. Role of C-Reactive Protein at Sites of Inflammation and Infection. Front. Immunol. 2018, 9, 754. [Google Scholar] [CrossRef] [PubMed]
- Iannuzzi, C.; Irace, G.; Sirangelo, I. The Effect of Glycosaminoglycans (GAGs) on Amyloid Aggregation and Toxicity. Molecules 2015, 20, 2510–2528. [Google Scholar] [CrossRef] [PubMed]
- Quittot, N.; Sebastiao, M.; Bourgault, S. Modulation of amyloid assembly by glycosaminoglycans: from mechanism to biological significance. Biochem. Cell Biol. 2017, 95, 329–337. [Google Scholar] [CrossRef]
- Sebastiao, M.; Quittot, N.; Marcotte, I.; Bourgault, S. Glycosaminoglycans Induce Amyloid Self-Assembly of a Peptide Hormone by Concerted Secondary and Quaternary Conformational Transitions. Biochemistry 2019, 58, 1214–1225. [Google Scholar] [CrossRef]
- Tsemekhman, K.; Goldschmidt, L.; Eisenberg, D.; Baker, D. Cooperative hydrogen bonding in amyloid formation. Protein Sci. 2007, 16, 761–764. [Google Scholar] [CrossRef]
- Navarro, S.; Díaz-Caballero, M.; Peccati, F.; Roldán-Martín, L.; Sodupe, M.; Ventura, S. Amyloid Fibrils Formed by Short Prion-Inspired Peptides Are Metalloenzymes. ACS Nano 2023, 17, 16968–16979. [Google Scholar] [CrossRef]
- Occhipinti, R.; Boron, W.F. Role of Carbonic Anhydrases and Inhibitors in Acid–Base Physiology: Insights from Mathematical Modeling. Int. J. Mol. Sci. 2019, 20, 3841. [Google Scholar] [CrossRef]
- Canepa, E. , Parodi-Rullan, R, Vazquez-Torres, R, Gamallo-Lana, B, Guzman-Hernandez, R, Lemon, NL, Angiulli, F, Debure, L, Ilies, MA, Østergaard, L, Wisniewski, T, Gutiérrez-Jiménez, E, Mar, AC, Fossati, S.. (2023) FDA-approved carbonic anhydrase inhibitors reduce amyloid β pathology and improve cognition, by ameliorating cerebrovascular health and glial fitness.. Alzheimers Dement. 5048; 19. [Google Scholar]
- Sapirstein, V.S.; Strocchi, P.; Gilbert, J.M. Properties and Function of Brain Carbonic Anhydrase. Ann. New York Acad. Sci. 1984, 429, 481–493. [Google Scholar] [CrossRef]
- Theparambil, S.M.; Begum, G.; Rose, C.R. pH regulating mechanisms of astrocytes: A critical component in physiology and disease of the brain. Cell Calcium 2024, 120, 102882. [Google Scholar] [CrossRef] [PubMed]
- Ruusuvuori, E. , Kaila, K.. (2014) Carbonic Anhydrases and Brain pH in the Control of Neuronal Excitability. In: Frost, S, McKenna, R. (eds) Subcellular Biochemistry. 75, Springer, Dordrecht. [CrossRef]
- Poggetti, V.; Salerno, S.; Baglini, E.; Barresi, E.; Da Settimo, F.; Taliani, S. Carbonic Anhydrase Activators for Neurodegeneration: An Overview. Molecules 2022, 27, 2544. [Google Scholar] [CrossRef]
- Wood, H.G.; Utter, M.F. The role of CO2 fixation in metabolism. . 1965, 1, 1–27. [Google Scholar] [PubMed]
- Gamble, J. , Lehninger, AL. ( NewYork, 611–622.
- Roosterman, D.; Meyerhof, W.; Cottrell, G.S. Proton Transport Chains in Glucose Metabolism: Mind the Proton. Front. Neurosci. 2018, 12, 404. [Google Scholar] [CrossRef]
- Reynolds, N.P. Amyloid-like peptide nanofibrils as scaffolds for tissue engineering: Progress and challenges (Review). Biointerphases 2019, 14, 040801. [Google Scholar] [CrossRef]
- Peña-Díaz, S.; Olsen, W.P.; Wang, H.; Otzen, D.E. Functional Amyloids: The Biomaterials of Tomorrow? Adv. Mater. 2024, 36, e2312823. [Google Scholar] [CrossRef] [PubMed]
- Aggeli, A. , Boden, N, Zhang, S (editors). . ( medicine and engineering.. Springer, Dordrecht.
- Zhang, S. (2003) More than just bare scaffolds: towards multi-component and decorated fibrous biomaterials.. Nat Biotechnol. 1171; 21. [Google Scholar]
- Eisenberg, D.; Jucker, M. The Amyloid State of Proteins in Human Diseases. Cell 2012, 148, 1188–1203. [Google Scholar] [CrossRef] [PubMed]
- Daskalov, A.; Dyrka, W.; Saupe, S.J. Theme and variations: evolutionary diversification of the HET-s functional amyloid motif. Sci. Rep. 2015, 5, srep12494. [Google Scholar] [CrossRef] [PubMed]
- Riek, R.; Saupe, S.J. The HET-S/s Prion Motif in the Control of Programmed Cell Death. Cold Spring Harb. Perspect. Biol. 2016, 8, a023515. [Google Scholar] [CrossRef] [PubMed]
- Lamour, G.; Nassar, R.; Chan, P.H.; Bozkurt, G.; Li, J.; Bui, J.M.; Yip, C.K.; Mayor, T.; Li, H.; Wu, H.; et al. Mapping the Broad Structural and Mechanical Properties of Amyloid Fibrils. Biophys. J. 2017, 112, 584–594. [Google Scholar] [CrossRef] [PubMed]
- Dos Reis, S.; Coulary-Salin, B.; Forge, V.; Lascu, I.; Bégueret, J.; Saupe, S.J. The HET-s Prion Protein of the Filamentous Fungus Podospora anserina Aggregates in Vitro into Amyloid-like Fibrils. J. Biol. Chem. 2002, 277, 5703–5706. [Google Scholar] [CrossRef] [PubMed]
- Greenwald, J. , Riek, R.. (2010) Biology of amyloid: structure, function and regulation. 1244; 18. [Google Scholar]
- Pham, C. , Kwan, AH, Sunde, M.. (2014) Functional amyloid: Widespread in nature, diverse in purpose. Essays Biochem. 56.
- Syed, A. , Boles, BR (2014) Fold modulating function: bacterial toxins to functional amyloids.. Frontiers in microbiology. 5.
- Barnhart, M. , Chapman, MR. . ( 2006) Curli biogenesis and function. Annu Rev Microbiol. 60, 131–147.
- Sønderby, T.V.; Zou, Y.; Wang, P.; Wang, C.; Otzen, D.E. Molecular-level insights into the surface-induced assembly of functional bacterial amyloid. Biophys. J. 2022, 121, 3422–3434. [Google Scholar] [CrossRef]
- Xuan, Q.; Wang, Y.; Chen, C.; Wang, P. Rational Biological Interface Engineering: Amyloidal Supramolecular Microstructure-Inspired Hydrogel. Front. Bioeng. Biotechnol. 2021, 9. [Google Scholar] [CrossRef]
- Morris, R. , MacPhee, C.. (2013) Amyloid Protein Biomaterials. In: Roberts, GCK (ed) Encyclopedia of Biophysics.. Springer, Berlin, Heidelberg. [CrossRef]
- Collier, J. , Messersmith, PB.. (2004) Self-assembling polymer peptide conjugates: nanostructural tailoring.. Adv Mater. 16.
- Knowles, T.P.; Fitzpatrick, A.W.; Meehan, S.; Mott, H.R.; Vendruscolo, M.; Dobson, C.M.; Welland, M.E. Role of Intermolecular Forces in Defining Material Properties of Protein Nanofibrils. Science 2007, 318, 1900–1903. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Lewis, R.V. Structure of a protein superfiber: spider dragline silk. Proc. Natl. Acad. Sci. 1990, 87, 7120–7124. [Google Scholar] [CrossRef]
- Yemini, M.; Reches, M.; Gazit, E.; Rishpon, J. Peptide Nanotube-Modified Electrodes for Enzyme−Biosensor Applications. Anal. Chem. 2005, 77, 5155–5159. [Google Scholar] [CrossRef]
- Scheibel, T.; Parthasarathy, R.; Sawicki, G.; Lin, X.-M.; Jaeger, H.; Lindquist, S.L. Conducting nanowires built by controlled self-assembly of amyloid fibers and selective metal deposition. Proc. Natl. Acad. Sci. USA 2003, 100, 4527–4532. [Google Scholar] [CrossRef] [PubMed]
- Willner, I. , Katz, E, editors.. (2005) Bioelectronics: from theory to applications. Weinheim: Wiley-VCH, GmBH & Co.KGaA.
- Ligorio, C.; Mata, A. Synthetic extracellular matrices with function-encoding peptides. Nat. Rev. Bioeng. 2023, 1, 518–536. [Google Scholar] [CrossRef] [PubMed]
- Deidda, G.; Jonnalagadda, S.V.R.; Spies, J.W.; Ranella, A.; Mossou, E.; Forsyth, V.T.; Mitchell, E.P.; Bowler, M.W.; Tamamis, P.; Mitraki, A. Self-Assembled Amyloid Peptides with Arg-Gly-Asp (RGD) Motifs As Scaffolds for Tissue Engineering. ACS Biomater. Sci. Eng. 2016, 3, 1404–1416. [Google Scholar] [CrossRef] [PubMed]
- van Dalen, M.C.; Karperien, M.; Claessens, M.M.; Post, J.N. Choice of Protein, Not Its Amyloid-Fold, Determines the Success of Amyloid-Based Scaffolds for Cartilage Tissue Regeneration. ACS Omega 2023, 8, 24198–24209. [Google Scholar] [CrossRef] [PubMed]
- Jacob, R. , Das, S, Singh, N, Patel, K, Datta, D, Sen, S, Maji, SK.. (2018) Amyloids Are Novel Cell-Adhesive Matrices.. Adv Exp Med Biol. 1112, 79-97. [CrossRef]
- Jacob, R.S.; George, E.; Singh, P.K.; Salot, S.; Anoop, A.; Jha, N.N.; Sen, S.; Maji, S.K. Cell Adhesion on Amyloid Fibrils Lacking Integrin Recognition Motif. J. Biol. Chem. 2016, 291, 5278–5298. [Google Scholar] [CrossRef] [PubMed]
- Jacob, R.S.; Ghosh, D.; Singh, P.K.; Basu, S.K.; Jha, N.N.; Das, S.; Sukul, P.K.; Patil, S.; Sathaye, S.; Kumar, A.; et al. Self healing hydrogels composed of amyloid nano fibrils for cell culture and stem cell differentiation. Biomaterials 2015, 54, 97–105. [Google Scholar] [CrossRef]
- Xuan, Q.; Wang, Y.; Chen, C.; Wang, P. Rational Biological Interface Engineering: Amyloidal Supramolecular Microstructure-Inspired Hydrogel. Front. Bioeng. Biotechnol. 2021, 9. [Google Scholar] [CrossRef]
- Koo, E.H.; Park, L.; Selkoe, D.J. Amyloid beta-protein as a substrate interacts with extracellular matrix to promote neurite outgrowth. Proc. Natl. Acad. Sci. 1993, 90, 4748–4752. [Google Scholar] [CrossRef]
- Ranjan, V. , Qiu, L, Lee, JW, Chen, X, Jang, SE, Chai, C, Lim, KL, Tan, EK, Zhang, Y, Huang, WM, Zeng, L.. (2020) A microfiber scaffold-based 3D in vitro human neuronal culture model of Alzheimer's disease.. Biomater Sci. 8, 4861-4874. [CrossRef]
- Mathes, T.G.; Monirizad, M.; Ermis, M.; de Barros, N.R.; Rodriguez, M.; Kraatz, H.-B.; Jucaud, V.; Khademhosseini, A.; Falcone, N. Effects of amyloid-β-mimicking peptide hydrogel matrix on neuronal progenitor cell phenotype. Acta Biomater. 2024. [Google Scholar] [CrossRef] [PubMed]
- Barbucci, R. , editor.. (2009) Hydrogels: biological properties and applications. Milan/New York: Springer.
- Yanlian, Y.; Ulung, K.; Xiumei, W.; Horii, A.; Yokoi, H.; Shuguang, Z. Designer self-assembling peptide nanomaterials. Nano Today 2009, 4, 193–210. [Google Scholar] [CrossRef]
- Liang, Y.; Guo, P.; Pingali, S.V.; Pabit, S.; Thiyagarajan, P.; Berland, K.M.; Lynn, D.G. Light harvesting antenna on an amyloid scaffold. Chem. Commun. 2008, 6522–6524. [Google Scholar] [CrossRef]
- Channon, K.J.; Devlin, G.L.; MacPhee, C.E. Efficient Energy Transfer within Self-Assembling Peptide Fibers: A Route to Light-Harvesting Nanomaterials. J. Am. Chem. Soc. 2009, 131, 12520–12521. [Google Scholar] [CrossRef]
- Choi, Y.S.; Kim, J.; Bhak, G.; Lee, D.; Paik, S.R. Photoelectric Protein Nanofibrils of α-Synuclein with Embedded Iron and Phthalocyanine Tetrasulfonate. Angew. Chem. 2011, 123, 6194–6198. [Google Scholar] [CrossRef]
- Ridgley, D.M.; Ebanks, K.C.; Barone, J.R. Peptide Mixtures Can Self-Assemble into Large Amyloid Fibers of Varying Size and Morphology. Biomacromolecules 2011, 12, 3770–3779. [Google Scholar] [CrossRef]
- Ridgley, D.M.; Barone, J.R. Evolution of the Amyloid Fiber over Multiple Length Scales. ACS Nano 2012, 7, 1006–1015. [Google Scholar] [CrossRef] [PubMed]
- Ridgley, D. , Claunch, EC, Barone, JR. (2012) The effect of processing on large, self-assembled amyloid fibers. 1029; 8. [Google Scholar]
- Diaz-Pier, S.; Carloni, P. Impact of quantum and neuromorphic computing on biomolecular simulations: Current status and perspectives. Curr. Opin. Struct. Biol. 2024, 87, 102817. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Liu, T.; Liu, W.; Luo, B.; Li, Y.; Fan, X.; Zhang, X.; Cui, W.; Teng, Y. SemiSynBio: A new era for neuromorphic computing. Synth. Syst. Biotechnol. 2024, 9, 594–599. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, H.; Xie, D. Recent Advance in Synaptic Plasticity Modulation Techniques for Neuromorphic Applications. Nano-Micro Lett. 2024, 16, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Ma, Z.; Zheng, H.; Li, T.; Chen, K.; Wang, X.; Liu, C.; Xu, L.; Wu, X.; Lin, D.; et al. The combination of brain-computer interfaces and artificial intelligence: applications and challenges. Ann. Transl. Med. 2020, 8, 712–712. [Google Scholar] [CrossRef] [PubMed]
Figure 2.
The morphology and structural organization of Curli fimbriae amyloid fibres produced by E. coli k-12 strain BW25113 grown on a low salt agar plate shown in a TEM image (a). Curli fibres produced in-vitro from purified CsgA monomers visualized in a TEM image (b). Schematic depiction of the structural organization of Curli fibres as proposed by Evans and Chapman [197] showing CsgA as a major component in this assembly (c). Some features of the repeat structures in CsgA (d) showing the five-sheet pseudo-repeat stacked structures (N22, R1-R5) which are prominent features of curlin (e). Localisation of amyloid like structures in an E.Coli K-12 strain AR3110 macro-colony biofilm visualized using Thioflavine S (f). Images in a, b reproduced from [198]. Images in c, d, e modified from [197], image f reproduced from [199]. All images obtained through open access with permission.
Figure 2.
The morphology and structural organization of Curli fimbriae amyloid fibres produced by E. coli k-12 strain BW25113 grown on a low salt agar plate shown in a TEM image (a). Curli fibres produced in-vitro from purified CsgA monomers visualized in a TEM image (b). Schematic depiction of the structural organization of Curli fibres as proposed by Evans and Chapman [197] showing CsgA as a major component in this assembly (c). Some features of the repeat structures in CsgA (d) showing the five-sheet pseudo-repeat stacked structures (N22, R1-R5) which are prominent features of curlin (e). Localisation of amyloid like structures in an E.Coli K-12 strain AR3110 macro-colony biofilm visualized using Thioflavine S (f). Images in a, b reproduced from [198]. Images in c, d, e modified from [197], image f reproduced from [199]. All images obtained through open access with permission.
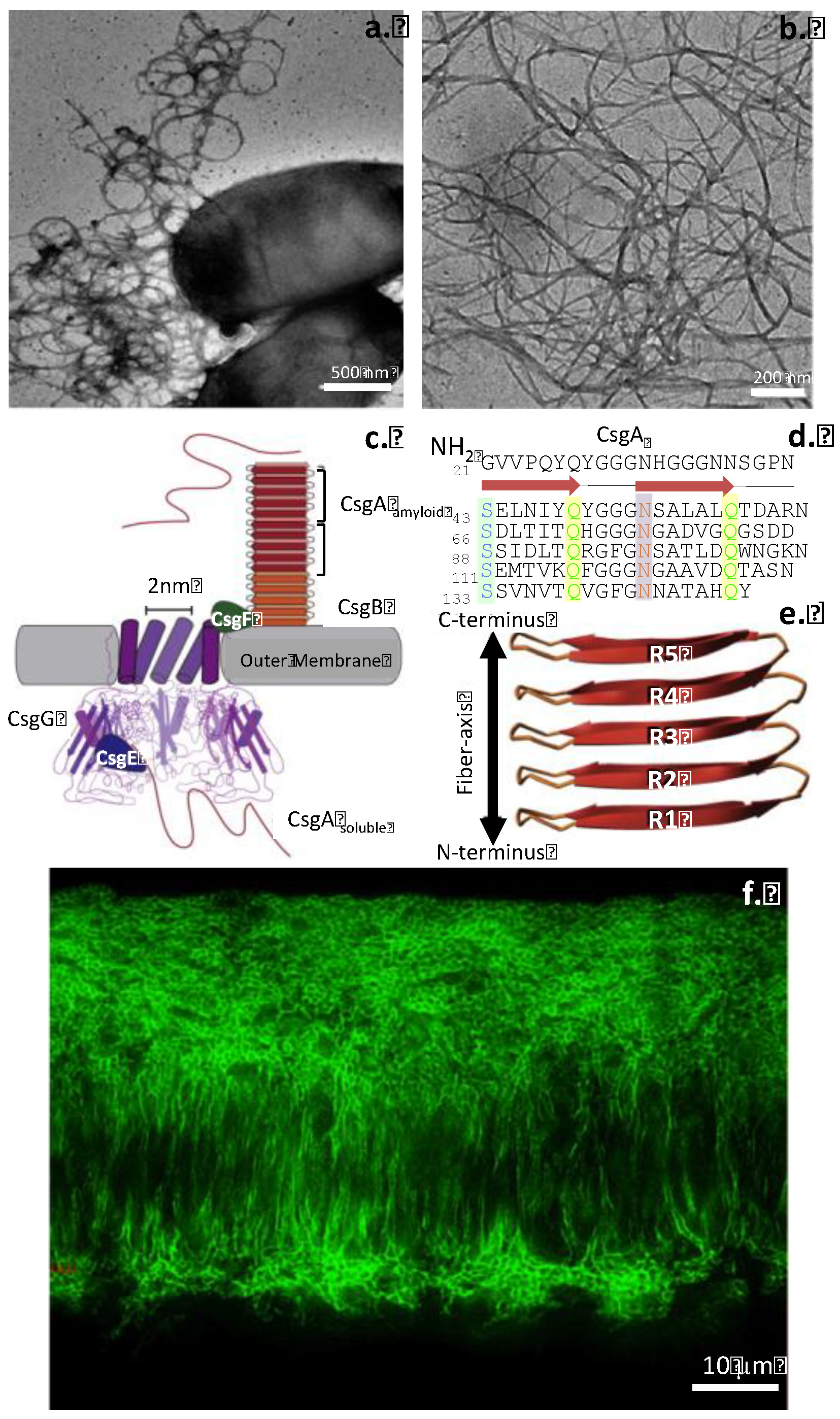
Figure 3.
Comparison of the structural organization of Orb2 and Huntington amyloids (a, b) showing the amyloid-forming sequences and high -sheet content. Schematic of in vivo anti-parallel hairpin fold and -sheets in Orb2 filaments derived from Cryo-EM analysis showing arrangements of three Orb2 molecule in-register multilayer packing of parallel β-sheets (c). Anti-parallel -hairpin and -sheet structure of in vitro assembled Huntington amyloid visualized by solid state nuclear magnetic resonance (d). Figure modified from [204].
Figure 3.
Comparison of the structural organization of Orb2 and Huntington amyloids (a, b) showing the amyloid-forming sequences and high -sheet content. Schematic of in vivo anti-parallel hairpin fold and -sheets in Orb2 filaments derived from Cryo-EM analysis showing arrangements of three Orb2 molecule in-register multilayer packing of parallel β-sheets (c). Anti-parallel -hairpin and -sheet structure of in vitro assembled Huntington amyloid visualized by solid state nuclear magnetic resonance (d). Figure modified from [204].
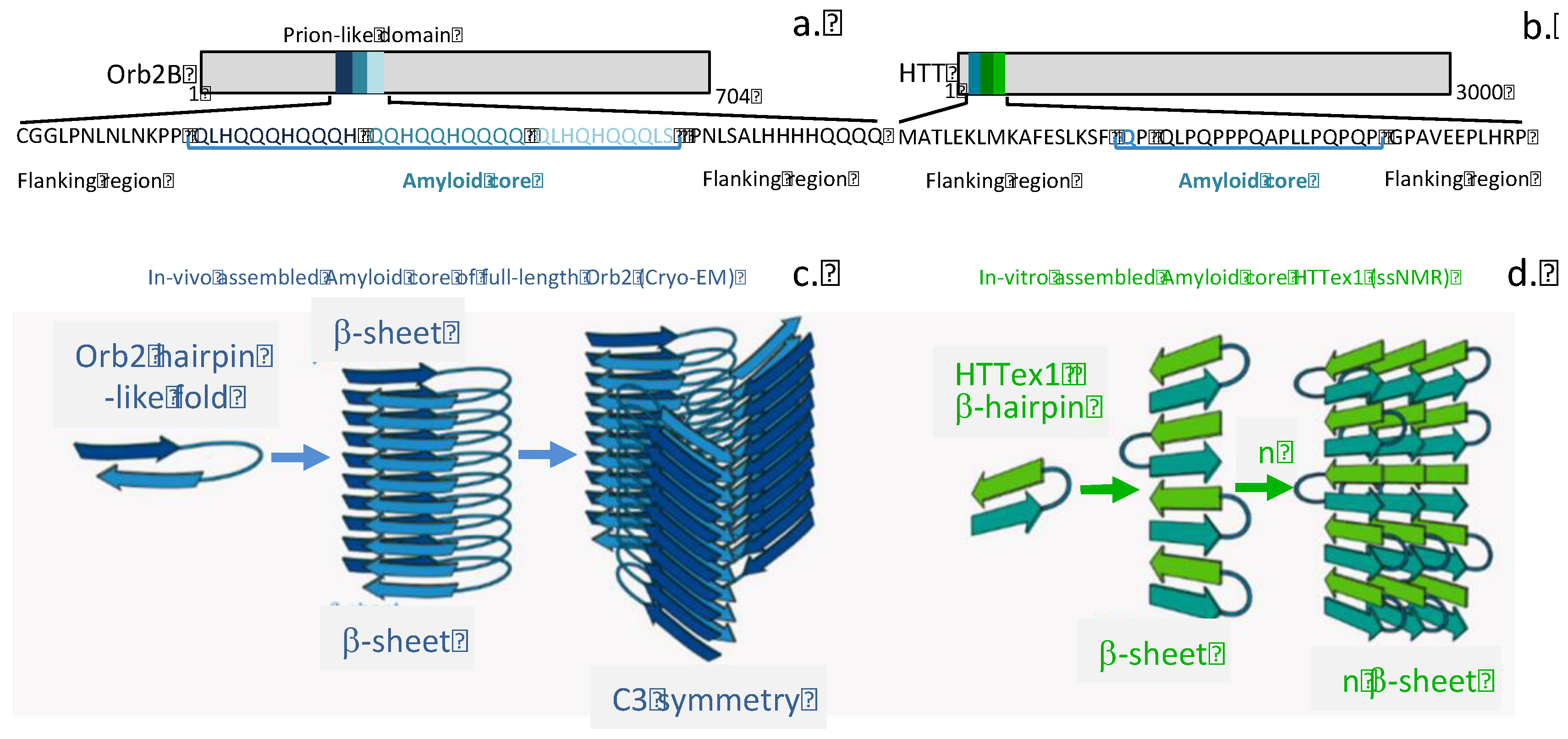
Figure 4.
Characteristic senile A plaques stained with silver or immunolocalised within A antibodies in tissue sections from the cortex of patients who died with AD. a. Naoumenko-Feigen silver-stained plaques counterstained with Periodic acid Schiff; an amyloid core (dark pink) is surrounded by profuse abnormal neurites (black). b.Brown-stained A plaques visualized using anti-A Mab 4G8 with a Nissl counterstain (blue). Glial nuclei are visible in the plaque core and the surrounding stained peripheral region. Brown A plaques stained with anti-A40 monoclonal antibody (MAb) (c-e) and A42 MAb (f-h) showing coarse punctate (c, d), diffuse (e,f) and prominent core staining (g, h). Scale bars 20 m. Mouse MAb 4G8 from Covance, Labcorp Drug Development (Princeton, NJ) raised against residues 17-24 of Aβ peptide, with a 4G8 epitope at residues 18-22 [60]. Images reproduced from [7].
Figure 4.
Characteristic senile A plaques stained with silver or immunolocalised within A antibodies in tissue sections from the cortex of patients who died with AD. a. Naoumenko-Feigen silver-stained plaques counterstained with Periodic acid Schiff; an amyloid core (dark pink) is surrounded by profuse abnormal neurites (black). b.Brown-stained A plaques visualized using anti-A Mab 4G8 with a Nissl counterstain (blue). Glial nuclei are visible in the plaque core and the surrounding stained peripheral region. Brown A plaques stained with anti-A40 monoclonal antibody (MAb) (c-e) and A42 MAb (f-h) showing coarse punctate (c, d), diffuse (e,f) and prominent core staining (g, h). Scale bars 20 m. Mouse MAb 4G8 from Covance, Labcorp Drug Development (Princeton, NJ) raised against residues 17-24 of Aβ peptide, with a 4G8 epitope at residues 18-22 [60]. Images reproduced from [7].
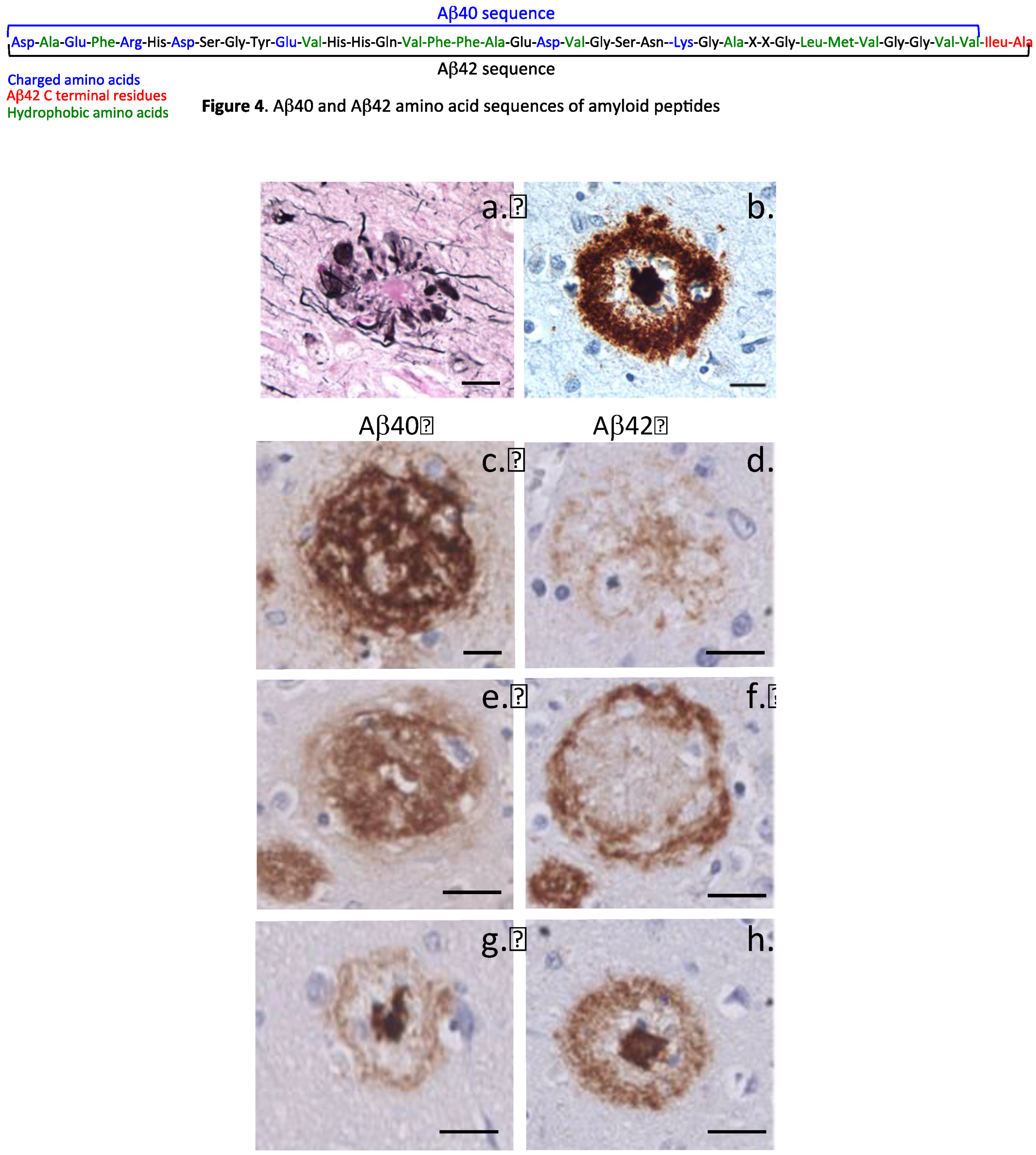

| Disease process | Amyloid precursor protein | Amyloid monomer |
|---|---|---|
| AD | APP | A |
| Atrial amyloidosis | Atrial natriuritic protein | Amyloid ATF |
| Spongiform encephalopothy | Prion protein PrPc | PrPsc |
| Primary systemic amyloidosis | Ig L and H chains | AL, AH |
| Secondary systemic amyloidosis | Apo serum amyloid A | SAA |
| Familial amyloid polyneuropathy I | Transthyretin | ATTR |
| Familial amyloid polyneuropathy II | ApoA | AApoA |
| Haemodialysis amyloidosis | 2-microglobulin | A2M |
| Hereditary systemic amyloidosis | Lysozyme | ALys |
| Diabetes type II | ProIAPP | APP/amylin |
| Insulin injection amyloidosis | Insulin | AIns |
| Cerebral amyloid angiopathy | Cystatin C | ACys |
| Finnish hereditary systemic amyloidosis | Gelsolin | AGel |
| Age associated pituitary prolactinomas | Prolactin | APro |
| Familial amyloidosis | Fibrinogen A chain | AFib |
Table 2.
Functional Amyloids used in bio-nanotechnology applications.
| Amyloid | Applications | Reference |
|---|---|---|
| Transthyretin peptide (105-115) | Cell adhesive properties | [186] |
| Gonadotropin releasing hormone | Long acting peptide/protein drug depot/delivery | [187] |
| Candida albicans Killer decapeptide | Auto-delivery of therapeutic peptides | [188] |
| Yeast Sup35p NM prion domain | Nanowire development | [189] |
| -synuclein fibrils | Enzyme entrapment hydrogel | [190] |
| insulin fibril semi-conductor oligoelectrolyte |
Optoelectronic nanowire assembly | [191] |
| PPF coated Insulin fibrils | Polymer light emitting diode fibrils | [192] |
| APFO-12 coated Insulin fibrils | Optical nanowires | [193] |
| PEDOT-S coated Insulin fibrils | Conductive nanowires | [194] |
| 2-microglobulin | Nanoporous cell support matrix | [195] |
Abbreviations: PEDOT-S, Poly(3,4-ethylenedioxythiophene) polystyrene sulfonate; PPF, luminescent polyfluorene ;APFO, Ammonium pentadecafluorooctanoate.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



