Submitted:
02 May 2024
Posted:
03 May 2024
You are already at the latest version
Abstract
Primary ciliary dyskinesia (PCD) is caused by pathologic variants in over 50 different genes that affect the structure and function of motile cilia. Low nasal NO (nNO) is a biomarker for diagnosis in patients with features of PCD; but the reason why it is low is unclear. At the cellular level, a characteristic feature shared by most PCD patients is that antigens and other particles are not cleared from the epithelial surface. Poor antigen clearance results in pro-oxidant pathway activation and airway epithelial damage and may predispose PCD patients to DUOX1- and IL33- mediated asthma. Secondary ciliary dysfunction, such as that caused by viruses or by smoking, can also contribute to asthma development. Variants in genes underlying the function of cilia, can be associated with poor lung function, even in the absence of PCD. Single nucleotide polymorphisms in PCD – related genes can be associated with increased asthma severity. The role of cilia and antigen stasis is the pathophysiology of asthma is an important area for research, because specific airway clearance techniques and other therapeutic interventions, such as antioxidants, could be of value in preventing the development of asthma and asthma-like symptoms.
Keywords:
primary ciliary dyskinesia 1
; asthma 2
; nitric oxide 3
; oxidative stress 4
1. Introduction
1.1. Antigen Stasis in Cilia as Predisposition for Asthma
Primary ciliary dyskinesia (PCD) is an inherited condition, commonly diagnosed in childhood, that is caused by pathologic variants in over 50 many different genes that affect the structure, and function of motile cilia1. PCD may be more common than was previously thought, with a world-wide incidence estimated to be about half that of cystic fibrosis: at least one in 7,600 [2]. The diagnosis of PCD is challenging, and measurement of nasal NO (nNO) has been widely adopted as a biomarker for disease diagnosis in individuals with features typical of PCD. These clinical attributes include newborn respiratory distress, chronic nasal congestion; daily cough, bronchitis, and bronchiectasis; and disorders of left-right symmetry (both situs inversus totalis and heterotaxy)[1,2,3,4,5]. Most patients with a confirmed diagnosis of PCD have low nNO3-5, however the reason for this is unknown.
At the cellular level, a characteristic feature shared by most PCD patients is that antigens and other particles are not well mobilized from the epithelial surface [6]. We have shown that PCD cells have decreased clearance of the antigen Dermatophagoides pteronyssinus (Derp-1) from the cell surface compared to normal airway epithelium [7]. Derp-1 is a house dust mite (HDM) allergen belongs to the group-1 allergens, which induce allergy commonly. We and others have shown that antigen stasis on airway epithelial surface results in upregulation of pro-oxidant enzymes that promote airway inflammation [8,9,10,11,12]. It is important to note here that these studies have been done with primary cell cultures grown at air-liquid interface from healthy control subjects and PCD patients. This is the most carefully controlled method for studying the redox biochemistry of airway epithelial cells: direct sampling of airway epithelial cells from the nose can give rise to a number of artifacts, making the redox data difficult to interpret Here, I propose that antigen stasis, particularly in children, leads to airway inflammation and to asthma-like symptoms in patients with PCD.
Our evidence suggests that PCD can cause airway injury and low airway levels of NO through upregulation of oxidant enzymes in the epithelium. In PCD, nNO levels are so low that measurement has become a component of the consensus ATS approach to PCD diagnosis [3]. Expression of NO synthase isoforms is not generally decreased in in PCD airways [13,14,15]. Our data suggest that nNO levels are low because NO is oxidized before leaving the airways. Antigens and irritants stuck on the epithelial surface upregulate activity of pro-oxidant epithelial enzymes that produce O2-, and NO is depleted by forming cytotoxic ONOO-/ONOOH. Additional NO oxidation reactions can deplete NO and form NO2, HNO2/NO2- [16]. These reactions cause airway injury and can lead bronchospasm [17].
Additionally, antigen stasis upregulates DUOX1 [8,9,18], injuring the airway through formation of H2O2. One pro-oxidant enzyme is dual oxidase 1 (DUOX1), upregulation of which during antigen stasis is mediated by P2Y and PAR receptors, leading to activation of type 2 alarmins [9]. DUOX1 upregulation may be directly associated with cilia movement18. P2Y and PAR activation have been linked with activation of DUOX1, but not necessarily with upregulation of DUOX1 protein levels. This oxidative stress, in turn, can both decrease gas phase NO concentrations and contribute to airway nitrosative stress and airway epithelium injury. While airway NO concentrations are low in children with PCD [1,3,4,19], our evidence demonstrates that the oxidation products of NO in the PCD airway are normal or high. In addition to depleting NO and injuring the airway, each of these reactions has the potential to cause bronchoconstriction and inflammation [17,18,20,21]. Further, when antigen stasis leads to DUOX1 upregulation there is epithelial release of the local alarmin, interleukin (IL)33 [22]. Downstream, IL33 can interact with its receptor (ST2) to initiate a cascade of mediators leading to asthma, including T-helper 2 (T2) cell mediated asthma, non-T2 asthma and mixed asthma [23]. Our data to date suggest that PCD-associated asthma will be characterized primarily by non-T2, low FeNO asthmatic airway inflammation; though it could also cause a so-called “mixed” asthma pathophysiology. These possibilities will require further study. Either way, I will argue that PCD, causing antigen stasis, leads commonly to bronchial hyper-reactivity in patients with PCD.
2. The Majority of PCD Patients Have Asthma
Many children with PCD require asthma treatment, such as inhaled long-acting β2 agonists (LABAs) and inhaled corticosteroids (ICS), to prevent episodes of acute bronchoconstriction. Indeed, a recent survey-based review of PCD management in the 22 PCD centers in Italy revealed that all centers prescribed ICS for their patients [24]. Some centers pre-scribed them to all patients, while others only if there were a positive test for bronchial hyper-reactivity (BHR) a sine qua non for asthma diagnosis [25]. Two studies have already studied BHR retrospectively, and my group is currently performing a formal, prospective study. An Israeli retrospective review demonstrated that 56% of PCD patients studied had significant reversal of obstruction following albuterol (BHR) [26], Table 1. A second, Canadian retrospective study of serial clinic visits found that most of their PCD patients had an improvement in forced expiratory volume at 1 second (FEV1) with the β2 agonist, albuterol, but only 18% had a 12% improvement [27], Table 1. There are several weaknesses in these two retrospective studies that help to explain the differences. Neither study used the formal criteria for assessing BHR as recommended by the American Thoracic Society (ATS). For the diagnosis of asthma, large networks like the Severe Asthma Research Program (SARP) [28] and Precision Interventions for Severe and Exacerbation-Prone Asthma (PrecISE) [29] use the ATS standards. These studies first require that the subject stop chronic controller medicines like ICS and LABA for at least a day be-fore coming in for BHR testing. Second, a subject can sequentially receive increasing doses of albuterol until there is no further bronchodilatation. Third, if a subject does not fully bronchodilate with albuterol, another drug withhold is done before a second visit, and the subject undergoes methacholine challenge test. If a subject is determined to have BHR by either test (bronchodilatation with albuterol or bronchoconstriction with methacholine – or other bronchoconstricting agents), a diagnosis of asthma is given. None of these requirements were met in either of the retrospective studies. Further, in the Canadian study, the older criteria of 12% improvement in FEV1 was used: the Israeli study used the more recently accepted norm, 10% [27]. Nevertheless, strikingly, most of the PCD subjects in both studies experienced improvement in FEV1 with albuterol. These studies are summarized in Table 1. Note that the Indiana study, though reported in abstract form, is still enrolling; its results will likely be the most conclusive, as it is a prospective study using best practice guidelines for diagnosis. The other studies to date have been retrospective reviews that did not rely on the forma assessment of bronchial hyper-reactivity.
In my research group, we have also addressed this question by two additional methods. First, we have queried large informatic databases to assess whether asthma is common among children with PCD. Though there is not an International Classification of Diseases (ICD)10 code for PCD itself, we interrogated a commonly used ICD10 code (Q89.3), as well as established codes for both bronchiectasis and situs inversus totalis, in the Indiana Network for Patient Care (INPCR) electronic health record [HER] database. We also studied a validation cohort using the IBM Explorys database (over 80 million individual EHRs). We matched patients who had the relevant ICD10 codes to age, sex and race-matched controls who did not have those codes at a ratio of 1 PCD subject: 3 controls. In these groups, we compared the proportion of subjects who did and did not have any ICD10 code for asthma using chi-squared tests. Informatic analysis of 18 million individual EHRs showed that subjects who definitively had PCD (both bronchiectasis and situs inversus totalis) PCD had ~ a 20-fold increased risk of also having an asthma diagnosis.
Secondly, we are performing a prospective study of BHR, using formal criteria as de-scribed above, among our PCD patient population. Our preliminary results prospective validation study (published in abstract form) shows that ~ 80% of confirmed PCD patients studied to date have a BHR test diagnostic for asthma. These data are consistent with the Israeli and Italian studies noted above (see Table 1) [26]. Indeed, PCD is often treated like asthma with routine use of inhaled steroids and β-agonists [31]. Taken together, these data suggest that the majority of patients with inherited ciliopathies also have asthma, and that antigen stasis on the epithelial surface contributes to the association between ciliopathy and asthma.
3. In Epithelial Cells, Cilia Can also Serve as Sensory Cilia, and Loss of Epithelia Can Affect Epithelial Signaling and Airway Smooth Muscle Relaxation
As noted above, NO oxidation is likely to be an important reason for the relatively pathognomonic low nNO observed in PCD. However, there may well be other contributing factors as well, factors that may be of relevance to the relationship between inherited ciliary dysfunction and asthma. Decreased expression of NO synthase (NOS) isoforms does not appear to account for the profoundly low nNO levels observed in PCD: expression is not much different in PCD airways compared to healthy individuals [13]. However, decreased NOS activity is a possibility, and we have previously reported that decreased ciliary motion prevents airway epithelial endothelial NOS (eNOS) activity [32], and this difference in activity could theoretically contribute to low airway NO values, though there is no difference in expression. A similar effect in endothelial cell signaling has been reported by Straub and coworkers [33], whereby hemoglobin expression and regulation of the heme iron oxidation state determines the bioactivity of colocalized eNOS. Indeed, NOS makes bronchodilator S-nitrosothiols in addition to NO [17,34]. We have identified and studied for the first-time hemoglobin (Hb) present in human airway pseudostratified epithelium [32]. Our data suggest that it is expressed, and that it could have at least two functions. First, Hbβ mRNA appears to increase during maturation and ciliation of airway epithelium. On blinded analysis, Hbβ knockdown (maximum achieved, 50%) caused a modest decrease in ciliation, and overexpression caused complete loss of ciliation and loss of pseudostratified architecture. Though mechanisms underlying these effects may involve cellular redox chemistry, more detailed analysis will be required in the future.
Secondly, somatic cell Hb in the airway epithelial cell may affect nitrogen oxide biology in the ciliated airway epithelium. Apical eNOS, which can signal through passive ciliary motion (airflow) as it does in other ciliated organs [35], is co-localized with Hbβ. Air pressure and flow activate eNOS by allowing calcium entry into the cell apex, producing NO and S-nitrosothiols [32]. CO pre-treatment almost completely ablated constitutive (NOS-dependent) [14,17,35] NO3- and NO2- accumulation in epithelial medium, consistent with its actions in the endothelium. Thus, we consider that the paradigm demonstrated by Angelo and coworkers [36] is relevant in the ciliated epithelium as it is at the myoendothelial junction: eNOS ultimately produces NO and S-nitrosothiols if the heme iron is oxidized, and it produces inert oxidation products if the heme iron is reduced. Though we have shown that exogenous nitrogen oxides increase cGMP accumulation in human air-way smooth muscle [37], we did not find a significant cGMP increase in our system [32]. We have shown [38] that Vest treatment increase NOS activity in vivo. Ciliary motion increases NOS activity independently of its expression. In this sense, airway cilia can function as sensory cilia, and loss of ciliary function could predispose to S-nitrosothiol deficiency and asthma [32].
Note also that ciliary paralysis and dysfunction can affect airway metabolism. Indeed, end-tidal oxygen tension is much higher in patients with PCD than in patients with CF or asthma, likely reflecting decreased effective respiration, and potentially contributing to oxidative loss of NO, as well as nitrosative stress [39].
4. Secondary Ciliopathy
The findings discussed above suggest that PCD is a risk factor for asthma. A corollary to consider is this: can secondary ciliary dysfunction – such as that caused by common childhood respiratory viruses, also contribute to asthma? Respiratory syncytial virus (RSV) is an example. It is a ubiquitous, seasonal respiratory viral pathogen and a major cause of morbidity and mortality in infants (children younger than 12 months) world-wide. Many years of observational studies have consistently shown an association be-tween RSV bronchiolitis and the development of childhood asthma [40,41,42]. A recent meta-analysis has supported this strong association [41].
It has been published recently that healthy children who were not infected with respiratory syncytial virus (RSV) during their first year of life were 26% less likely to have asthma by age 5 years than those who had been infected, according to results from an observational study that included 1946 participants. The researchers calculated that preventing RSV infections during infancy could avoid 15% of asthma cases in 5-year-olds. Moreover, a child’s risk of developing asthma was linked to the severity of their RSV infection. Children with milder RSV infections had a lower risk of asthma at age 5 years than did those who had more severe RSV infections [43].
RSV infects the respiratory epithelium, leading to chronic inflammation [44], and induces loss of cilia activity, two factors that determine mucus clearance and the increase in sputum volume. Other changes caused by RSV includes ultrastructural abnormalities con-fined to ciliated cells, such as increased cilia loss and mitochondrial damage [44,45]. I hypothesize that secondary ciliary dysmotility caused by viral infections like RSV could, as in PCD, cause antigen stasis, contributing to an asthma diathesis46.
RSV infects the respiratory epithelium, leading to chronic inflammation and induces the release of mucins and the loss of cilia activity, two factors that determine mucus clearance and the increase in sputum volume. Other changes caused by RSV includes ultrastructural abnormalities confined to ciliated cells, such as increased cilia loss and mitochondrial damage [44]. These alterations involve reactive oxygen species-dependent mechanisms. ROS overproduction plays an important role in the pathogenesis of severe RSV infection and serves as a major factor in pulmonary inflammation and tissue damage. Thus, antioxidants seem to be an effective treatment for severe RSV infection [47]. The antioxidant N-acetylcysteine (NAC) has proven useful in the management of COPD, reducing symptoms, exacerbations, and accelerated lung function decline in RSV [46]. Thus, antioxidants prescribed for RSV treatment, may be useful to manage secondary ciliopathy as well.
Other pathogens, such as Pseudomonas aeruginosa, can affect respiratory cilia by inhibiting its beating [48], as well as Streptococcus pneumoniae [49] by producing oxidant H202 [18]. Mycotoxins released by the opportunistic fungi such as Aspergillus flavus can reduce ciliary motility [50]. The effects of RSV and bacterial airway infections thus can create a vicious cycle. They decrease ciliary function, causing antigen stasis and oxidative stress; this, in turn, worsens ciliary function. I would argue that the downstream result is an asthma diathesis.
A toxic chemical exposure, such as smoking, can also result in secondary ciliopathies. Heavy smokers have shorter cilia in the large and small airways compared to nonsmokers, with further shortening observed in smokers with COPD [51,52]. Electron microscopic assessment of cilia ultrastructure demonstrates that smokers with chronic bronchitis have greater numbers of ciliary abnormalities compared to nonsmokers, including compound cilia and giant cilia, and other abnormalities in the microtubules and the axonemal pattern of organization and in cilia orientation [53,54,55]. These abnormalities may persist after smoking cessation [56]. In this case as well, loss of ciliary function could lead to antigen stasis, augmenting asthma risk.
Motile ciliary disorder in patients with bronchiectasis. Motile ciliary disorder (MCD), such as defective ciliogenesis, abnormal ciliary ultrastructure, or defective ciliary beating, were present in 89.8% of patients with bronchiectasis, 67.6% showed secondary MCD, and 16.2% showed primary plus secondary MCD. Compared with healthy control participants, patients with bronchiectasis yielded abnormal staining patterns of DNAH5, DNAI1, and RSPH9 (but not Ezrin) that were more prominent in moderate to severe bronchiectasis. MCD pattern scores largely were consistent between upper and lower airways and between large-to-medium and small airways in bronchiectasis. Coexisting nasal diseases and asthma did not confound nasal ciliary ultrastructural marker expression significantly. The propensity of MCD was unaffected by the airway or systemic inflammatory endotypes. MCD, particularly an ultrastructural abnormality, was notable in patients with mild bronchiectasis who showed blood or sputum eosinophilia [57].
Note also that abnormal expression patterns of RSPH1, RSPH4A, RSPH9, and DNAH5 were significantly more common in patients with nasal polyps, a chronic upper airway inflammatory disease that is frequently triggered by defective host-defense [58], even in the absence of classic PCD.
5. Antigen Stasis Leading to Oxidant Generation Is Increased in the PCD Airway Epithelium and May Contribute to both Ciliopathies and Asthma Pathogenesis. Antioxidant Treatment May Be Beneficial for PCD Management
Low nasal nitric oxide is now considered to be diagnostic for PCD in children over five years old [4,19,59]. If decreased ciliary function increases oxidative stress in the airway, NO could be consumed, and the airway injured. Specifically, NO reacts with oxidants, including H2O2, O2-, OH● and O2 itself [17,20,60,61]. Kinetics vary widely and in some cases are higher order. Products include NO2, HNO2-/ NO2- , NO3-, ONOO-/ONOOH and other aqueous nitrogen oxides [17,20,60,61]. All these oxidative reactions deplete the reactant, NO. In addition to contributing to low NO concentrations in the PCD airway, oxidative stress can contribute to airway injury. Superoxide reacts rapidly with NO to form ONOO-/ONOOH (pKa ~6.5), and ONOOH reacts with protein tyrosines to cause protein injury through tyrosine nitration [20,61]. Depending on conditions - such as –H - the products can have cytotoxicities: oxidative stress in general, and products of NO oxidation in particular, can injure the airway epithelium [17,20,60]. One pro-oxidant enzyme is dual oxidase 1 (DUOX1), upregulation of which during antigen stasis is mediated by P2Y and PAR receptors, leading to activation of type 2 alarmins [8,9]. DUOX1 upregulation may be directly associated with cilia movement [18]. P2Y and PAR activation have been linked with activation of DUOX1, but not necessarily with upregulation of DUOX1 protein levels. We hypothesized that PCD airway cells might not efficiently clear antigens from the epithelial surface, resulting in increased oxidative stress. This possibility is supported by recent evidence that antigens on the airway epithelial surface increase ex-pression of pro-oxidant enzymes in airway epithelial cells [61,62]. This oxidative stress, in turn, could both decrease gas phase NO concentrations and contribute to airway nitrosative stress and airway epithelium injury. While airway NO concentrations are low in PCD [1,3,4,5,31,59], our evidence demonstrates that the oxidation products of NO in the PCD airway are normal or high. In healthy airways, ciliary function is normal. Antigens and irritants are rapidly cleared, and NO enters the gas phase normally to be exhaled. In the PCD airways, however, our preliminary data show that antigens, such as Dermatophagoides pteronyssinus (Derp)1, and irritants in the airways are not as well cleared [7]. This defect leads to oxidative stress, marked by increased DUOX1 expression and decreased superoxide dismutase [SOD] activity. H2O2, in high concentrations in the PCD airway, injures the airway. NO is oxidized under these circumstances, primarily by O2-, rather than being exhaled. Tissue injury, at least in part through tyrosine nitrosation chemistry, may predispose PCD patients to asthma; additionally, the immune response to antigens that are not cleared from the epithelial surface may promote the asthmatic airway response.
Thus, we hypothesized that PCD patients may benefit from airway clearance [63] and antioxidant therapy. Antioxidants decrease production of different oxidants. For example, Apocynin and SOD decrease O2-, [64] and, in the presence of NO, ONOOH; catalase de-creases levels of H2O2 [65]. Glutathione and vitamins A, C, E and β-carotenes can have more generalized antioxidant effects [66,67,68]. We have shown that treatment with either apocynin or SOD treatment increases headspace NO over DNAH11 PCD cells [7]. Vitamin D may be beneficial for PCD patients with bronchiectasis and vitamin D deficiency [69]. Mucous retention, antigen stasis and bacterial or viral infection, due to cilia immotility or inefficient motility in PCD patients, promotes infections and inflammation, which in turn can lead to pulmonary exacerbations and bronchiectasis. As a result, quality of life, lung function and structure are relatively poor in PCD patients. Airway clearance (both pharmacological – b-agonists, pulmozymee (DNA-ase), hypertonic saline, antibiotics (tobramycine) and mechanical (Vest, Chest PT) or physical exercise can be beneficial as well [70].
Exhaled Nitric Oxide fraction measurement can be used routinely to evaluate the response and adherence to treatments [71].
6. PCD Genes and Asthma Risk
To date, pathogenic variants in over 50 genes are known to cause primary ciliary dyskinesia (PCD). Most of these causes of PCD are inherited in an autosomal recessive fashion. Some data argue that being a carrier for a recessive PCD gene could be a risk factor for asthma. Moving forward, as more genes and variants are discovered, it will be important to continue to track whether impaired ciliary function represented by heterozygosity could represent a risk factor for poor lung function and for asthma.
A few examples suggesting an impact of PCD gene heterozygosity and increased risk for obstructive lung disease are as follows. Variants in genes underlying the development or function of cilia, such as DNAH14 and DNAAF3, were associated with poor lung function in cystic fibrosis, whereas variants in DNAH6 were associated with preserved lung function in cystic fibrosis. Associations between DNAH14 and lung function were replicated in disease-related phenotypes characterized by obstructive lung disease in adults [72]. One study has shown that expression of PCD – related genes, DNAH5, KIFC2 and KIF3A are downregulated in asthma, and that SNPs in these genes correlate with asthma and disease severity [73]. Several single nucleotide polymorphisms (SNPs) in PCDH1 have been linked to asthma and BHR, however, the functional consequences of this mutation have not been explored [74].
An association of RSPH3 (the gene is related to primary ciliary dyskinesia) is observed in non-obese children with asthma [68]
7. Conclusions
Inherited ciliopathy leading to antigen stasis may predispose to asthma development in children. Asthma management should be considered in children with PCDs. On the other hand, severe asthmatics should be evaluated for the evidence of ciliopathies, Further, strategies should likely be considered to help clear antigens and irritants after acute airway infections such as RSV, for the purpose of preventing antigen stasis-induced asthma. Antigen stasis in PCD upregulates pro-oxidant generation, leading to decrease nNO and airway epithelial injury as well as increased IL33 production. In developing antioxidant treatment regimens, biomarkers such as nNO and breath H2O2 might be considered as surrogates for the effectiveness of the therapy. Indeed, compelling clinical evidence is beginning to emerge that the majority of PCD patients also have asthma. This association leads to a broader set of questions regarding the development of asthma. Specifically, do secondary ciliopathies predispose to asthma through similar mechanisms, and could partially dysfunctional cilia in some heterozygous patients also be a risk factor for asthma? Either way, the role of cilia and antigen stasis is the pathophysiology of asthma is an important area for research, because specific airway clearance techniques and other therapeutic interventions, such as antioxidants, could be of value in preventing the development of asthma and asthma-like symptoms. This proposed scheme is summarized in Figure 1.
Author Contributions
NM – wrote the paper.
Funding
This research was funded P01 HL158507 and P01 HL128192.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing is not applicable to this article.
Acknowledgments
Virginia Ferrante made the figure for the paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Leigh, M.W.; Horani, A.; Kinghorn, B.; O’Connor, M.G.; Zariwala, M.A.; Knowles, M.R. Primary Ciliary Dyskinesia (PCD): A genetic disorder of motile cilia. Transl Sci Rare Dis 2019, 4, 51–75. [Google Scholar] [CrossRef] [PubMed]
- Hannah, W.B.; Seifert, B.A.; Truty, R.; Zariwala, M.A.; Ameel, K.; Zhao, Y.; Nykamp, K.; Gaston, B. The global prevalence and ethnic heterogeneity of primary ciliary dyskinesia gene variants: a genetic database analysis. Lancet Respir Med 2022, 10, 459–468. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, A.J.; Zariwala, M.A.; Ferkol, T.; Davis, S.D.; Sagel, S.D.; Dell, S.D.; Rosenfeld, M.; Olivier, K.N.; Milla, C.; Daniel, S.J.; et al. Diagnosis, monitoring, and treatment of primary ciliary dyskinesia: PCD foundation consensus recommendations based on state of the art review. Pediatr Pulmonol 2016, 51, 115–132. [Google Scholar] [CrossRef] [PubMed]
- Noone, P.G.; Leigh, M.W.; Sannuti, A.; Minnix, S.L.; Carson, J.L.; Hazucha, M.; Zariwala, M.A.; Knowles, M.R. Primary ciliary dyskinesia: diagnostic and phenotypic features. Am J Respir Crit Care Med 2004, 169, 459–467. [Google Scholar] [CrossRef]
- Walker, W.T.; Liew, A.; Harris, A.; Cole, J.; Lucas, J.S. Upper and lower airway nitric oxide levels in primary ciliary dyskinesia, cystic fibrosis and asthma. Respir Med 107, 380-386.
- Wallmeier, J.; Al-Mutairi, D.A.; Chen, C.T.; Loges, N.T.; Pennekamp, P.; Menchen, T.; Ma, L.; Shamseldin, H.E.; Olbrich, H.; Dougherty, G.W. , et al. Mutations in CCNO result in congenital mucociliary clearance disorder with reduced generation of multiple motile cilia. Nat Genet 2014, 46, 646–651. [Google Scholar] [CrossRef] [PubMed]
- Gaston, B.; Smith, L.A.; Davis, M.D.; Saunders, J.; Daniels, I.; Horani, A.; Brody, S.L.; Giddings, O.; Zhao, Y.; Marozkina, N. Antigen stasis and airway nitrosative stress in human primary ciliary dyskinesia. Am J Physiol Lung Cell Mol Physiol 2024, 326, L468–l476. [Google Scholar] [CrossRef] [PubMed]
- Hristova, M.; Habibovic, A.; Veith, C.; Janssen-Heininger, Y.M.; Dixon, A.E.; Geiszt, M.; van der Vliet, A. Airway epithelial dual oxidase 1 mediates allergen-induced IL-33 secretion and activation of type 2 immune responses. J Allergy Clin Immunol 2016, 137, 1545–1556.e1511. [Google Scholar] [CrossRef]
- van der Vliet, A.; Danyal, K.; Heppner, D.E. Dual oxidase: a novel therapeutic target in allergic disease. Br J Pharmacol 2018, 175, 1401–1418. [Google Scholar] [CrossRef]
- Habibovic, A.; Hristova, M.; Heppner, D.E.; Danyal, K.; Ather, J.L.; Janssen-Heininger, Y.M.; Irvin, C.G.; Poynter, M.E.; Lundblad, L.K.; Dixon, A.E.; et al. DUOX1 mediates persistent epithelial EGFR activation, mucous cell metaplasia, and airway remodeling during allergic asthma. JCI Insight 2016, 1, e88811. [Google Scholar] [CrossRef]
- Dweik, R.A.; Comhair, S.A.; Gaston, B.; Thunnissen, F.B.; Farver, C.; Thomassen, M.J.; Kavuru, M.; Hammel, J.; Abu-Soud, H.M.; Erzurum, S.C. NO chemical events in the human airway during the immediate and late antigen-induced asthmatic response. Proc Natl Acad Sci U S A 2001, 98, 2622–2627. [Google Scholar] [CrossRef]
- Herbert, C.A.; King, C.M.; Ring, P.C.; Holgate, S.T.; Stewart, G.A.; Thompson, P.J.; Robinson, C. Augmentation of permeability in the bronchial epithelium by the house dust mite allergen Der p1. Am J Respir Cell Mol Biol 1995, 12, 369–378. [Google Scholar] [CrossRef]
- Degano, B.; Valmary, S.; Serrano, E.; Brousset, P.; Arnal, J.F. Expression of nitric oxide synthases in primary ciliary dyskinesia. Hum Pathol 2011, 42, 1855–1861. [Google Scholar] [CrossRef] [PubMed]
- Asano, K.; Chee, C.B.; Gaston, B.; Lilly, C.M.; Gerard, C.; Drazen, J.M.; Stamler, J.S. Constitutive and inducible nitric oxide synthase gene expression, regulation, and activity in human lung epithelial cells. Proc Natl Acad Sci U S A 1994, 91, 10089–10093. [Google Scholar] [CrossRef]
- Smith, C.M.; Fadaee-Shohada, M.J.; Sawhney, R.; Baker, N.; Williams, G.; Hirst, R.A.; Andrew, P.W.; O’Callaghan, C. Ciliated cultures from patients with primary ciliary dyskinesia do not produce nitric oxide or inducible nitric oxide synthase during early infection. Chest 2013, 144, 1671–1676. [Google Scholar] [CrossRef] [PubMed]
- Jourd’heuil, D.; Jourd’heuil, F.L.; Kutchukian, P.S.; Musah, R.A.; Wink, D.A.; Grisham, M.B. Reaction of Superoxide and Nitric Oxide with Peroxynitrite: IMPLICATIONS FOR PEROXYNITRITE-MEDIATED OXIDATION REACTIONSIN VIVO *. Journal of Biological Chemistry 2001, 276, 28799–28805. [Google Scholar] [CrossRef] [PubMed]
- Marozkina, N.V.; Gaston, B. Nitrogen chemistry and lung physiology. Annu Rev Physiol 2015, 77, 431–452. [Google Scholar] [CrossRef]
- Price, M.E.; Sisson, J.H. Redox regulation of motile cilia in airway disease. Redox Biol 2019, 27, 101146. [Google Scholar] [CrossRef]
- Piacentini, G.L.; Bodini, A.; Peroni, D.; Rigotti, E.; Pigozzi, R.; Pradal, U.; Boner, A.L. Nasal nitric oxide for early diagnosis of primary ciliary dyskinesia: practical issues in children. Respir Med 2008, 102, 541–547. [Google Scholar] [CrossRef]
- van der Vliet, A.; Eiserich, J.P.; Shigenaga, M.K.; Cross, C.E. Reactive nitrogen species and tyrosine nitration in the respiratory tract: epiphenomena or a pathobiologic mechanism of disease? Am J Respir Crit Care Med 1999, 160, 1–9. [Google Scholar] [CrossRef]
- Wan, W.Y.; Hollins, F.; Haste, L.; Woodman, L.; Hirst, R.A.; Bolton, S.; Gomez, E.; Sutcliffe, A.; Desai, D.; Chachi, L.; et al. NADPH Oxidase-4 Overexpression Is Associated With Epithelial Ciliary Dysfunction in Neutrophilic Asthma. Chest 2016, 149, 1445–1459. [Google Scholar] [CrossRef]
- Cook, D.P.; Thomas, C.M.; Wu, A.Y.; Rusznak, M.; Zhang, J.; Zhou, W.; Cephus, J.-Y.; Gibson-Corley, K.N.; Polosukhin, V.V.; Norlander, A.E.; et al. Cystic Fibrosis Reprograms Airway Epithelial IL-33 Release and Licenses IL-33-Dependent Inflammation. Am J Respir Crit Care Med 2023, 207, 1486–1497. [Google Scholar] [CrossRef] [PubMed]
- Gauvreau, G.M.; Bergeron, C.; Boulet, L.P.; Cockcroft, D.W.; Côté, A.; Davis, B.E.; Leigh, R.; Myers, I.; O’Byrne, P.M.; Sehmi, R. Sounding the alarmins-The role of alarmin cytokines in asthma. Allergy 2023, 78, 402–417. [Google Scholar] [CrossRef] [PubMed]
- Ullmann, N.; Santamaria, F.; Allegorico, A.; Fainardi, V.; Borrelli, M.; Ferraro, V.A.; Proietti, E.; Parisi, G.F.; Romagnoli, V.; Lucca, F.; et al. Primary ciliary dyskinesia: A multicenter survey on clinical practice and patient management in Italy. Pediatr Pulmonol 2023, 58, 1127–1135. [Google Scholar] [CrossRef]
- Graham, B.L.; Steenbruggen, I.; Miller, M.R.; Barjaktarevic, I.Z.; Cooper, B.G.; Hall, G.L.; Hallstrand, T.S.; Kaminsky, D.A.; McCarthy, K.; McCormack, M.C.; et al. Standardization of Spirometry 2019 Update. An Official American Thoracic Society and European Respiratory Society Technical Statement. Am J Respir Crit Care Med 2019, 200, e70–e88. [Google Scholar] [CrossRef] [PubMed]
- Levine, H.; Bar-On, O.; Nir, V.; West, N.; Dizitzer, Y.; Mussaffi, H.; Prais, D. Reversible Bronchial Obstruction in Primary Ciliary Dyskinesia. J Clin Med 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Seidl, E.; Gatt, D.; Wee, W.B.; Wilson, D.; Ratjen, F.; Grasemann, H. Bronchodilator responsiveness in children with primary ciliary dyskinesia. ERJ Open Res 2024, 10. [Google Scholar] [CrossRef] [PubMed]
- Jarjour, N.N.; Erzurum, S.C.; Bleecker, E.R.; Calhoun, W.J.; Castro, M.; Comhair, S.A.; Chung, K.F.; Curran-Everett, D.; Dweik, R.A.; Fain, S.B.; et al. Severe asthma: lessons learned from the National Heart, Lung, and Blood Institute Severe Asthma Research Program. Am J Respir Crit Care Med 2012, 185, 356–362. [Google Scholar] [CrossRef] [PubMed]
- Georas, S.N.; Wright, R.J.; Ivanova, A.; Israel, E.; LaVange, L.M.; Akuthota, P.; Carr, T.F.; Denlinger, L.C.; Fajt, M.L.; Kumar, R.; et al. The Precision Interventions for Severe and/or Exacerbation-Prone (PrecISE) Asthma Network: An overview of Network organization, procedures, and interventions. J Allergy Clin Immunol 2022, 149, 488–516.e489. [Google Scholar] [CrossRef]
- Joseph, P. Castlen, E.M.M.J.Z., Beth Eubanks Julia Meyer,Charles Clem, Benjamin Gaston, Nadzeya Marozkina. Prevalence of Bronchial Hyperreactivity and Asthma in Patients with Primary Ciliary Dyskinesia: A Preliminary Study. PCD on the move: abstract 2024.
- Strippoli, M.P.; Frischer, T.; Barbato, A.; Snijders, D.; Maurer, E.; Lucas, J.S.; Eber, E.; Karadag, B.; Pohunek, P.; Zivkovic, Z.; et al. Management of primary ciliary dyskinesia in European children: recommendations and clinical practice. Eur Respir J 2012, 39, 1482–1491. [Google Scholar] [CrossRef]
- Marozkina, N.; Smith, L.; Zhao, Y.; Zein, J.; Chmiel, J.F.; Kim, J.; Kiselar, J.; Davis, M.D.; Cunningham, R.S.; Randell, S.H.; Gaston, B. Somatic cell hemoglobin modulates nitrogen oxide metabolism in the human airway epithelium. Sci Rep 2021, 11, 15498. [Google Scholar] [CrossRef]
- Straub, A.C.; Lohman, A.W.; Billaud, M.; Johnstone, S.R.; Dwyer, S.T.; Lee, M.Y.; Bortz, P.S.; Best, A.K.; Columbus, L.; Gaston, B.; Isakson, B.E. Endothelial cell expression of haemoglobin alpha regulates nitric oxide signalling. Nature 2012, 491, 473–477. [Google Scholar] [CrossRef] [PubMed]
- Gaston, B.; Reilly, J.; Drazen, J.M.; Fackler, J.; Ramdev, P.; Arnelle, D.; Mullins, M.E.; Sugarbaker, D.J.; Chee, C.; Singel, D.J.; et al. Endogenous nitrogen oxides and bronchodilator S-nitrosothiols in human airways. Proc Natl Acad Sci U S A 1993, 90, 10957–10961. [Google Scholar] [CrossRef] [PubMed]
- Kobzik, L.; Bredt, D.S.; Lowenstein, C.J.; Drazen, J.; Gaston, B.; Sugarbaker, D.; Stamler, J.S. Nitric oxide synthase in human and rat lung: immunocytochemical and histochemical localization. Am J Respir Cell Mol Biol 1993, 9, 371–377. [Google Scholar] [CrossRef] [PubMed]
- Angelo, M.; Singel, D.J.; Stamler, J.S. An S-nitrosothiol (SNO) synthase function of hemoglobin that utilizes nitrite as a substrate. Proc Natl Acad Sci U S A 2006, 103, 8366–8371. [Google Scholar] [CrossRef] [PubMed]
- Gaston, B.; Drazen, J.M.; Jansen, A.; Sugarbaker, D.A.; Loscalzo, J.; Richards, W.; Stamler, J.S. Relaxation of human bronchial smooth muscle by S-nitrosothiols in vitro. J Pharmacol Exp Ther 1994, 268, 978–984. [Google Scholar] [PubMed]
- Marozkina, N.; Bosch, J.; Cotton, C.; Smith, L.; Seckler, J.; Zaman, K.; Rehman, S.; Periasamy, A.; Gaston, H.; Altawallbeh, G.; et al. Cyclic compression increases F508 Del CFTR expression in ciliated human airway epithelium. American Journal of Physiology-Lung Cellular and Molecular Physiology 2019, 317, L247–L258. [Google Scholar] [CrossRef] [PubMed]
- Mendelsohn, L.; Wijers, C.; Gupta, R.; Marozkina, N.; Li, C.; Gaston, B. A novel, noninvasive assay shows that distal airway oxygen tension is low in cystic fibrosis, but not in primary ciliary dyskinesia. Pediatr Pulmonol 2019, 54, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Driscoll, A.J.; Arshad, S.H.; Bont, L.; Brunwasser, S.M.; Cherian, T.; Englund, J.A.; Fell, D.B.; Hammitt, L.L.; Hartert, T.V.; Innis, B.L.; et al. Does respiratory syncytial virus lower respiratory illness in early life cause recurrent wheeze of early childhood and asthma? Critical review of the evidence and guidance for future studies from a World Health Organization-sponsored meeting. Vaccine 2020, 38, 2435–2448. [Google Scholar] [CrossRef] [PubMed]
- Hartert, T.V.; Wu, P.; Brunwasser, S.M. Respiratory syncytial virus and asthma: untying the Gordian knot. Lancet Respir Med 2021, 9, 1092–1094. [Google Scholar] [CrossRef]
- Wittig, H.J.; Glaser, J. The relationship between bronchiolitis and childhood asthma; a follow-up study of 100 cases of bronchiolitis. J Allergy 1959, 30, 19–23. [Google Scholar] [CrossRef]
- Harris, E. RSV Infection During Infancy Tied to Asthma Later. JAMA 2023, 329, 1731–1731. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.M.; Kulkarni, H.; Radhakrishnan, P.; Rutman, A.; Bankart, M.J.; Williams, G.; Hirst, R.A.; Easton, A.J.; Andrew, P.W.; O’Callaghan, C. Ciliary dyskinesia is an early feature of respiratory syncytial virus infection. Eur Respir J 2014, 43, 485–496. [Google Scholar] [CrossRef] [PubMed]
- Lara-Hernandez, I.; Muñoz-Escalante, J.C.; Bernal-Silva, S.; Noyola, D.E.; Wong-Chew, R.M.; Comas-García, A.; Comas-Garcia, M. Ultrastructural and Functional Characterization of Mitochondrial Dynamics Induced by Human Respiratory Syncytial Virus Infection in HEp-2 Cells. Viruses 2023, 15. [Google Scholar] [CrossRef] [PubMed]
- Mata, M.; Sarrion, I.; Armengot, M.; Carda, C.; Martinez, I.; Melero, J.A.; Cortijo, J. Respiratory syncytial virus inhibits ciliagenesis in differentiated normal human bronchial epithelial cells: effectiveness of N-acetylcysteine. PLoS One 2012, 7, e48037. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Liu, X.; Nie, Y.; Zhan, F.; Zhu, B. Oxidative stress and ROS-mediated cellular events in RSV infection: potential protective roles of antioxidants. Virology Journal 2023, 20, 224. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.; Pitt, T.; Taylor, G.; Watson, D.; MacDermot, J.; Sykes, D.; Roberts, D.; Cole, P. Pyocyanin and 1-hydroxyphenazine produced by Pseudomonas aeruginosa inhibit the beating of human respiratory cilia in vitro. J Clin Invest 1987, 79, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Steinfort, C.; Wilson, R.; Mitchell, T.; Feldman, C.; Rutman, A.; Todd, H.; Sykes, D.; Walker, J.; Saunders, K.; Andrew, P.W.; et al. Effect of Streptococcus pneumoniae on human respiratory epithelium in vitro. Infect Immun 1989, 57, 2006–2013. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.J.; Workman, A.D.; Carey, R.M.; Chen, B.; Rosen, P.L.; Doghramji, L.; Adappa, N.D.; Palmer, J.N.; Kennedy, D.W.; Cohen, N.A. Fungal Aflatoxins Reduce Respiratory Mucosal Ciliary Function. Sci Rep 2016, 6, 33221. [Google Scholar] [CrossRef] [PubMed]
- Hessel, J.; Heldrich, J.; Fuller, J.; Staudt, M.R.; Radisch, S.; Hollmann, C.; Harvey, B.G.; Kaner, R.J.; Salit, J.; Yee-Levin, J.; et al. Intraflagellar transport gene expression associated with short cilia in smoking and COPD. PLoS One 2014, 9, e85453. [Google Scholar] [CrossRef]
- Leopold, P.L.; O’Mahony, M.J.; Lian, X.J.; Tilley, A.E.; Harvey, B.G.; Crystal, R.G. Smoking is associated with shortened airway cilia. PLoS One 2009, 4, e8157. [Google Scholar] [CrossRef]
- Fox, B.; Bull, T.B.; Oliver, T.N. The distribution and assessment of electron-microscopic abnormalities of human cilia. Eur J Respir Dis Suppl 1983, 127, 11–18. [Google Scholar] [PubMed]
- Lungarella, G.; Fonzi, L.; Ermini, G. Abnormalities of bronchial cilia in patients with chronic bronchitis. An ultrastructural and quantitative analysis. Lung 1983, 161, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Wedes, S.H.; Wu, W.; Comhair, S.A.; McDowell, K.M.; DiDonato, J.A.; Erzurum, S.C.; Hazen, S.L. Urinary bromotyrosine measures asthma control and predicts asthma exacerbations in children. J Pediatr 2011, 159, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Verra, F.; Escudier, E.; Lebargy, F.; Bernaudin, J.F.; De Crémoux, H.; Bignon, J. Ciliary abnormalities in bronchial epithelium of smokers, ex-smokers, and nonsmokers. Am J Respir Crit Care Med 1995, 151, 630–634. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.L.; Pan, C.X.; Tang, C.L.; Cen, L.J.; Zhang, X.X.; Huang, Y.; Lin, Z.H.; Li, H.M.; Zhang, X.F.; Wang, L.; et al. Motile Ciliary Disorders of the Nasal Epithelium in Adults With Bronchiectasis. Chest 2023, 163, 1038–1050. [Google Scholar] [CrossRef]
- Zi, X.X.; Guan, W.J.; Peng, Y.; Tan, K.S.; Liu, J.; He, T.T.; Ong, Y.K.; Thong, M.; Shi, L.; Wang, D.Y. An Integrated Analysis of Radial Spoke Head and Outer Dynein Arm Protein Defects and Ciliogenesis Abnormality in Nasal Polyps. Front Genet 2019, 10, 1083. [Google Scholar] [CrossRef] [PubMed]
- Pifferi, M.; Bush, A.; Maggi, F.; Michelucci, A.; Ricci, V.; Conidi, M.E.; Cangiotti, A.M.; Bodini, A.; Simi, P.; Macchia, P.; Boner, A.L. Nasal nitric oxide and nitric oxide synthase expression in primary ciliary dyskinesia. Eur Respir J 2011, 37, 572–577. [Google Scholar] [CrossRef]
- Gray, D., Lissi, E. , and Heicklen, J. Reaction of hydrogen peroxide with nitrogen dioxide and nitric oxide. J Phys Chem 1972, 1919-1924.
- Haddad, I.Y.; Pataki, G.; Hu, P.; Galliani, C.; Beckman, J.S.; Matalon, S. Quantitation of nitrotyrosine levels in lung sections of patients and animals with acute lung injury. J Clin Invest 1994, 94, 2407–2413. [Google Scholar] [CrossRef] [PubMed]
- Chan, T.K.; Tan, W.S.D.; Peh, H.Y.; Wong, W.S.F. Aeroallergens Induce Reactive Oxygen Species Production and DNA Damage and Dampen Antioxidant Responses in Bronchial Epithelial Cells. The Journal of Immunology 2017, 199, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Bonser, L.R.; Zlock, L.; Finkbeiner, W.; Erle, D.J. Epithelial tethering of MUC5AC-rich mucus impairs mucociliary transport in asthma. J Clin Invest 2016, 126, 2367–2371. [Google Scholar] [CrossRef]
- Stefanska, J.; Pawliczak, R. Apocynin: molecular aptitudes. Mediators Inflamm 2008, 2008, 106507. [Google Scholar] [CrossRef]
- Miles, A.M.; Bohle, D.S.; Glassbrenner, P.A.; Hansert, B.; Wink, D.A.; Grisham, M.B. Modulation of Superoxide-dependent Oxidation and Hydroxylation Reactions by Nitric Oxide (∗). Journal of Biological Chemistry 1996, 271, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Barrera-Mendoza, C.C.; Ayala-Mata, F.; Cortés-Rojo, C.; García-Pérez, M.E.; Rodríguez-Orozco, A.R. [Antioxidant vitamins in asthma]. Rev Alerg Mex 2018, 65, 61–77. [Google Scholar] [CrossRef] [PubMed]
- Tesoriere, L.; Bongiorno, A.; Pintaudi, A.M.; D’Anna, R.; D’Arpa, D.; Livrea, M.A. Synergistic interactions between vitamin A and vitamin E against lipid peroxidation in phosphatidylcholine liposomes. Arch Biochem Biophys 1996, 326, 57–63. [Google Scholar] [CrossRef]
- Buettner, G.R. The pecking order of free radicals and antioxidants: lipid peroxidation, alpha-tocopherol, and ascorbate. Arch Biochem Biophys 1993, 300, 535–543. [Google Scholar] [CrossRef] [PubMed]
- Mirra, V.; Caffarelli, C.; Maglione, M.; Valentino, R.; Perruolo, G.; Mazzarella, C.; Di Micco, L.L.; Montella, S.; Santamaria, F. Hypovitaminosis D: a novel finding in primary ciliary dyskinesia. Ital J Pediatr 2015, 41, 14. [Google Scholar] [CrossRef]
- Paff, T.; Omran, H.; Nielsen, K.G.; Haarman, E.G. Current and Future Treatments in Primary Ciliary Dyskinesia. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef]
- Miranda, L.; Guerrero, J. [Measurement of exhaled nitric oxide fraction in lung diseases]. Rev Med Chil 2021, 149, 1173–1181. [Google Scholar] [CrossRef] [PubMed]
- Blue, E.; Louie, T.L.; Chong, J.X.; Hebbring, S.J.; Barnes, K.C.; Rafaels, N.M.; Knowles, M.R.; Gibson, R.L.; Bamshad, M.J.; Emond, M.J. Variation in Cilia Protein Genes and Progression of Lung Disease in Cystic Fibrosis. Ann Am Thorac Soc 2018, 15, 440–448. [Google Scholar] [CrossRef]
- Sivaprasad, U.; Gibson, A.M.; Wang, N.; Khurana Hershey, G.K. Expression of Cilia Structural Genes is Downregulated in Asthma and Single Nucleotide Polymorphisms in These Genes Correlate with Asthma. Journal of Allergy and Clinical Immunology 2009, 123, S81. [Google Scholar] [CrossRef]
- Koppelman, G.H.; Meyers, D.A.; Howard, T.D.; Zheng, S.L.; Hawkins, G.A.; Ampleford, E.J.; Xu, J.; Koning, H.; Bruinenberg, M.; Nolte, I.M.; et al. Identification of PCDH1 as a novel susceptibility gene for bronchial hyperresponsiveness. Am J Respir Crit Care Med 2009, 180, 929–935. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Antigen stasis in PCD increases the risk of asthma. Antigens, such as cockroaches, mites, RSV and chemical irritants, such as produced by smoking, cannot be removed easily from PCD lungs as from healthy lungs due to ciliary immotility. This brings activation of pro-oxidant enzymes, such as DUOX1 and increases IL33 production, and, ultimately, airway epithelial injury, predisposing PCD lungs for asthma development. Additional genetic factors, such as genetic variations in some PCD – causing genes (DNAH14, DNAAF3, DNAH6, DNAH5, KIFC2, KIF3A) are associated with increased risk of asthma development.
Figure 1.
Antigen stasis in PCD increases the risk of asthma. Antigens, such as cockroaches, mites, RSV and chemical irritants, such as produced by smoking, cannot be removed easily from PCD lungs as from healthy lungs due to ciliary immotility. This brings activation of pro-oxidant enzymes, such as DUOX1 and increases IL33 production, and, ultimately, airway epithelial injury, predisposing PCD lungs for asthma development. Additional genetic factors, such as genetic variations in some PCD – causing genes (DNAH14, DNAAF3, DNAH6, DNAH5, KIFC2, KIF3A) are associated with increased risk of asthma development.
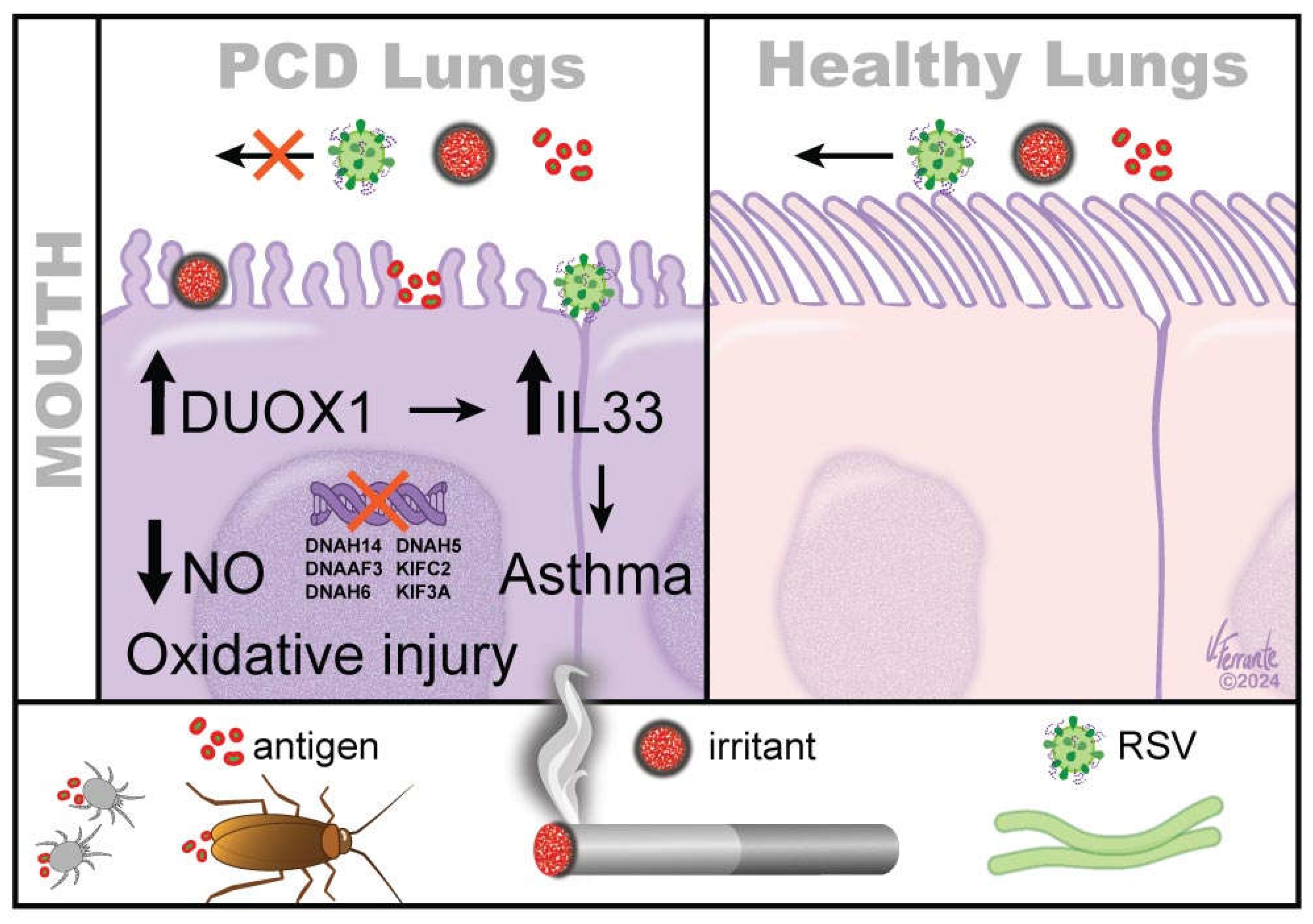

Table 1.
PCD patients with positive bronchodilatator response.
| Study | Asthma drug withhold before the study | Maximum bronchodilator | Methacholine if failed bronchodilator | Prospective | Number | Percent of PCD patients with positive bronchodilatator response (BD) | Reference |
|---|---|---|---|---|---|---|---|
| Indiana | Yes | Yes | Yes | Yes | 9 | 81 | [30] |
| Israel | No | No | No | No | 46 | 56 | [26] |
| Toronto | No | No | No | No | 474 | 18 | [27] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



