
Submitted:
11 March 2024
Posted:
13 March 2024
You are already at the latest version
Abstract
Bacteriophages (phages) are potential alternatives to chemical antimicrobials against pathogens of public health significance. Understanding the diversity and host specificity of phages is important for developing effective phage biocontrol approaches. Here, we assessed the host range, morphology, and genetic diversity of eight Salmonella enterica phages isolated from a wastewater treatment plant. The host range analysis revealed that six out of eight phages lysed more than 81% of the 43 Salmonella enterica isolates tested. The genomic sequences of all phages were determined. Whole genome sequencing data (WGS) data revealed that phage genome sizes ranged from 41 to 114 kb with GC contents be-tween 39.9 and 50.0 %. Two of the phages SB13 and SB28 represent new species Epsep-timavirus SB13 and genera Macdonaldcampvirus, respectively as designated by the In-ternational Committee for the Taxonomy of Viruses (ICTV) as per genome based taxonomic classification. One phage (SB18) belonged to the Myoviridae morphotype while the re-maining phages belonged to the Siphoviridae morphotype. The gene content analyses showed that none of the phages possessed virulence, toxin, antibiotic resistance, type I-VI toxin-antitoxin modules or lysogeny genes. Three (SB3, SB15 and SB18) out of the eight phages possessed tailspike proteins. Whole genome-based phylogeny of the eight phages with their 113 homologs revealed three clusters A, B, C and seven subclusters (A1, A2, A3, B1, B2, C1 and C2). While cluster C1 phages were predominantly isolated from animal sources, cluster B contained phages from both wastewater and animal sources. The broad host range of these phages highlights their potential use to control the presence of S. enterica in foods.
Keywords:
Salmonella
; bacteriophage
; diversity
; host specificity
; comparative phylogenomics
; food safety.
1. Introduction
Nontyphoidal Salmonella (NTS) is a major cause of foodborne illness and hospitalizations in Canada [1]. Globally, NTS causes approximately 80 million foodborne-related illnesses and 155,000 deaths each year [2]. Non-typhoidal Salmonellosis is caused by the main species of the genus, Salmonella enterica, which consists of six subspecies (I, II, IIIa, IIIb, IV, and VI) with subspecies I containing more than 1,500 serotypes with high genetic diversity [3]. In Canada there have been several large national and international outbreaks of Salmonellosis linked to vegetables including whole onions, peaches, frozen corn, and cantaloupes since 2020 [4,5,6,7]. These outbreaks have been caused by several S. enterica serotypes (Enteritidis, Newport, Soahanina, Sundsvall, Oranienburg), and have highlighted the need for improved approaches to control presence and growth of diverse S. enterica in fresh and processed fruits and vegetables, which increasingly contribute to the burden of foodborne disease. Bacteriophages (phages) are increasingly being recognized as natural antimicrobials to reduce the growth and survival of foodborne pathogens (including Salmonella) during food production [8,9,10,11,12] because of their ability to kill their host bacteria [13,14], and also due to the fact that phages can exhibit broad host ranges [15] making them useful for controlling diverse bacterial species such as S. enterica [16]. The majority of phages described in the scientific literature appear to be generally host specific, infecting a subset of species, strains, or serotypes due to the specificity of their host receptors [17]. While some phages can infect a broad range of bacteria belonging to different serotypes, species, and/or genus [17,18], information on these phages are limited.
While Salmonella phages and their genomic sequences have been well documented [17,19,20,21,22,23,24,25], the phenotypic and genotypic diversity of Salmonella enterica means that there are likely many additional phages with unique features that remain to be characterized. Understanding the biological and genomic characteristics of these phages are essential to the development of phage-based antimicrobial methods to control this foodborne pathogen [13,26]. In this study, we determined the host range spectra, and performed comparative genomic and phylogenetic analyses, as well as morphological characterization of eight S. enterica phages isolated from wastewater obtained from the Jean R. Marcotte wastewater treatment plant in Montreal, Québec, Canada.
2. Materials and Methods
2.1. Bacterial Host Strains and Their Growth
Non-typhoidal S. enterica isolates (n = 43) representing 30 serotypes, and commonly associated with fresh produce outbreaks were obtained from the Salmonella foodborne Syst-OMICS database (SALFOS, Laval University, QC, Canada, https://salfos.ibis.ulaval.ca/) (Figure 1). Frozen stocks of the isolates were maintained at −80 ºC and were revived by streaking on Luria-Bertani (LB) agar plates followed by overnight incubation at 37 ºC. For all experiments, a single colony of a respective isolate from a fresh LB agar plate was inoculated into 10 mL of LB broth and incubated overnight at 37 ºC with shaking at 150 RPM.
2.2. Bacteriophage Isolation and Propagation
Bacteriophage isolation was carried out using pre-treated sewage sludge samples (1L) collected from the Jean-R. Marcotte Wastewater Treatment Plant (WWTP) in Montreal by adopting standard enrichment and agar overlay techniques [27] with a slight modification as described elsewhere [28]. Thirteen Salmonella isolates, originally isolated from food plants/fresh produce, and representing thirteen serotypes, were used for phage isolation (Figure 1).
A sewage sample (1 L) was transported to the laboratory on ice, where it was clarified by centrifugation at 10,000 × g for 30 min in 50 mL tubes and filtered through 0.22 µm syringe filter units (Pall Corporation, Port Washington, NY, United States). Thirty milliliters of filtrate was mixed with 9 mL of 5x Rappaport-Vassiliadis (RV), 1 mL of 1 M CaCl2 and 10 mL of an overnight culture of a Salmonella isolate, and the suspension was incubated in a 250 mL flask at 37 ºC for 48 h while shaking at 50 RPM. The culture was clarified by centrifugation at 10,000 × g for 10 min and the supernatant was filtered through a 0.22 µm filter to remove bacterial debris. An aliquot (10 µL) of filtrate was spot inoculated on a lawn of the Salmonella isolate used for isolation and the plate was incubated at 37 ºC overnight. The next day, the lawns were checked for zones of lysis or individual plaques. Plaque purification was carried out multiple times using a streak plating method, as previously described [29], to obtain a clonal phage isolate. Phage stocks were concentrated by centrifugation (40,000 × g for 2 h at 4 ºC) and stored at 4 ºC until further use. For routine use, phages were propagated by the liquid lysate method [28] where a mid-exponential phase bacterial host culture was infected with phage and incubated overnight. The next day, the phage and the bacterial host culture was centrifuged (10,000 × g for 10 min) and filtered through a 0.45 µm syringe filter, followed by determination of the phage titre.
2.3. Host Range Profiles
Host ranges for each of the isolated phages were determined using the agar overlay method. Following preparation of agar overlays (as described above), the lawns (prepared for each of the 43 Salmonella isolates to be tested) were spot inoculated with aliquots (10 μL) of each phage having titres of 108 plaque-forming units (PFUs)/mL. The spots were allowed to dry before incubating overnight at 37 ºC. The scoring for lysis was completed as reported elsewhere [30,31], where 0 indicated no lysis and +3 indicated complete clear lysis.
2.4. Phage DNA Isolation, Sequencing, and Annotation
Genomic DNA from the Salmonella phages was extracted using the Wizard DNA Clean-Up system (A7280; Promega) following the modified Promega Wizard method as described by the Center for Phage Technology, Texas A&M University, USA [32]. Extracted DNA was purified by ethanol precipitation [33], and whole genome sequencing (WGS) was performed on an Illumina MiSeq platform with 300-bp paired-end libraries and 30X coverage. Raw sequence reads were assembled using the A5 pipeline [34], and genome annotation was completed using the Bacterial and Viral Bioinformatics Resource Center (formerly PATRIC) [35,36]. Annotations were manually curated, and the coding sequences (CDS) were used to interrogate the NCBI database using BLASTP [37]. An HHpred search of the Pfams database was used to identify conserved protein motifs [38]. To assign a protein to a gene sequence, at least 90% identity was sought in BLASTP searches for protein motifs [39]. Based on the presence or absence of a gene encoding integrase, phages were putatively classified as temperate or virulent, respectively [26].
2.5. Phylogenetic and Comparative Genomic Analyses
The whole-genome alignments of the phages reported in this study and 113 homologs that were extracted from NCBI were generated using MAFFT v7.453 [40]. Maximum likelihood trees were constructed using IQtree v2.2 (https://github.com/Cibiv/IQ-TREE) [41] and visualized using Microreact [42]. Phages were assessed for genes encoding antimicrobial resistance, virulence, and type I-VI toxin-antitoxin modules using CARD [43], VFDB [44,45], and TADB [46], respectively. Likewise, phage genome sequences were screened for presence of tailspike proteins using an in-house manually curated custom database. Whole-genome comparisons and their visual representations were carried out using EasyFig (https://github.com/mjsull/Easyfig) [47].
2.6. Electron Microscopic Imaging of the Phages
Phages were purified for electron microscopy by equilibrium density gradient centrifugation through CsCl at ≈ 22,000 RCF for 24 h in a Beckman Ultra centrifuge (TL100) [48]. Post centrifugation, the residual CsCl was removed from the phage fraction by centrifuging 500 μL of the supernatant fluid through an Amicon Ultra-0.5, 30 kDa MWCO centrifugal filter unit (Millipore Ltd). Following purification, transmission electron microscopy was conducted at the Imaging - Microscopy Platform of the Institute of Integrative Biology and Systems (IBIS), Laval University, Quebec City, Canada.
3. Results
3.1. Phage Isolation and Biological Characterization by Host Range Profile
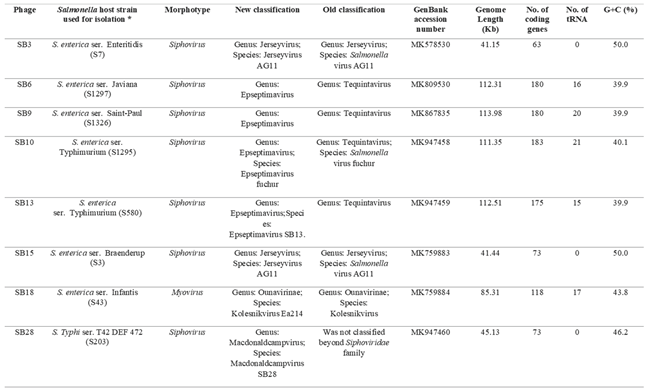
Eight phages (Table 1) were isolated using S. enterica isolates representing serotypes commonly associated with fresh produce outbreaks. The host range profile of the isolated phages was determined using 43 different Salmonella isolates from 30 serotypes (Figure 1&2). Salmonella isolates representing the top eight plant associated salmonellosis-causing serotypes were given importance due to their frequent implication in produce associated outbreaks, which account for the majority of foodborne outbreaks [49,50,51,52]. The top eight serotypes were identified as serotypes Newport, Javiana, Enteriditis, Typhimurium, Thompson, Heidelberg, Saintpaul, and Poona (Figure 2). One additional serotype, Litchfield, was also included because it has been implicated in numerous outbreaks associated with melons [53]. The broadest host range was exhibited by phages SB3 and SB6, followed by SB9. Phages SB3 and SB6 lysed 88.3% (n = 38/43) isolates, while phage SB9 lysed 86% (n = 37/43) isolates, phage SB10 lysed 76.7% (n = 33/43), phages SB13 and SB18 lysed 83.7% (n = 36/43) isolates, phage SB28 lysed 81.3% (n = 35/43) and phage SB15 showed the lowest lysis percentage of 67.4% (n = 29/43) isolates. Five of eight (62.5% (SB3, SB6, SB9, SB10, SB13)) phages isolated in this study lysed the top eight serotypes and Litchfield (Figure 2).
3.2. Comparative Phylogenomic Analysis of Phages Understudy
To ascertain the suitability of the isolated phages for biocontrol purposes, the phages were sequenced. Raw reads were assembled into draft genomes that were functionally annotated as described above (see Methods).
3.2.1. Comparative Gene Content Analysis of Phages under Study
The genome size of the phages ranged from 41 to 114 kb, while their GC contents were between 39.9 to 50.0% (Table 1). Whole genome-based phylogenetic analysis of the eight phages revealed two clusters (cluster A and B) and two singletons (Figure 3). Phages SB28, SB9, SB10 and SB13 were nested together in cluster A, SB15 and SB3 were clustered in cluster B, while SB6 and SB18 were singletons. Homology of phage genomes with previously identified phages were determined using a comparative genomic approach with BlastN. Phages in cluster A were heterogenous in terms of their genome size, number of tRNAs and coding sequences (Table 1). Phages SB9 and SB10 were similar to Salmonella phage 116 (accession number NC_048007.1, 99.58% identity, 88% coverage) and Salmonella phage fuchur (accession number: NC_048869.1, 97% identity and 96% coverage) that were both isolated from wastewater in Denmark [54] (Figure 4A). Phages SB13 and SB28 respectively had 99% (coverage 92%) and 96% (coverage 79%) nucleotide similarity to Escherichia phage Supergirl (accession number: MZ501105.1) that was isolated from a sewage plant in Switzerland (Figure 4B) [55] and Salmonella phage E1 (accession number: NC_010495.1) from an unknown source in the United Kingdom (Figure 4C) [56].
Phages SB3 and SB15 (Cluster B) had comparable genome size (41 kB), number of tRNAs and GC content, and shared 99% nucleotide similarity between them, and >95% nucleotide identity (coverage 97%) with Salmonella phage vB_SenS_AG11 (accession number NC_041991.1) that was isolated from sewage in Guelph, Canada in 2007 [57,58] (Figure 4D). The singletons: Phage SB6 and SB18 respectively had 99.35 and 98% nucleotide sequence similarity to Salmonella phage oselot (accession number: NC_048871.1, 95% coverage) recovered from wastewater in Denmark [59], and Erwinia phage phiEa21-4 (accession number NC_011811.1, 98% coverage) that was isolated in soil beneath a pear tree with active blight in Canada [60,61] (Figure 4E-F). Collectively, given the broad host ranges of the phages isolated in this study, these results confirm the utility of wastewater as a rich source from which to isolate diverse and broad host range phages [62,63].
The genomes of the phages isolated in this study were screened for the presence of lysogeny, antimicrobial resistance, virulence, or toxin-antitoxin related genes. Of note, genes encoding virulence, type I-VI toxin-antitoxin modules, antimicrobial resistance, or lysogeny were not detected in any of the phages. Given the broad host range of the isolated phages, an assessment of the genes encoding bacterial recognition proteins was conducted. Three (SB3, SB15 and SB18) out of the eight phages possessed tailspike proteins (Figure 3). The tailspike proteins from phages SB3 and SB15 were identical (100% nucleotide sequence similarity and coverage) and shared 93% nucleotide sequence similarity with Salmonella phage vB_SenS_AG11 tailspike protein (accession number: AF012431.1). Although functional annotation predicted these tailspike proteins to belong to Salmonella phage P22-like tailspike protein, comparative genomic analysis revealed that they shared only 66% nucleotide similarities with Salmonella phage P22 tailspike protein (accession number: NP_059644.1). Phage SB18 had a bulb like structure at the base of the tail typical of phages with tailspike proteins but there were no genes annotated as tailspike protein in its genome. More so, BLASTing phage SB18 genome against a manually curated custom database containing 8077 unduplicated tailspike proteins did not yield any significant hit. However, phage SB18 contained a gene annotated as a baseplate protein that had 97% nucleotide similarity to the baseplate spike protein of Erwinia phage phiEa21-4 (accession number: YP_004327040.1).
3.2.2. Assessing the Genetic Relatedness of Phages under Study with Other Salmonella Phage Genomes
Phylogenetic analysis of the phages showed high genetic relatedness with other phages. For example, based on genome comparisons with previously identified Salmonella phages deposited in public databases, The phages were clustered into three main clusters designated as A, B, which were further grouped into seven subclusters (A1, A2, B1, B2, C1, C2 and C3) (Figure 5). The singleton phages SB6 and SB18 (Figure 3) were placed in different subclusters of cluster C. Phage SB6 clustered with Bacteriophage T5-like in subcluster C1, whereas SB18 was found in subcluster C2 along with Erwinia phages phiEa21-4 and Salmonella phage ST-3 among others. Phage SB28 with Salmonella phage E1 formed a distinct subcluster (A1), while phages SB9, SB10, and SB13 were found in subcluster A2 with other Salmonella phages (Figure 5). Conversely, phages SB3 and SB15 were clustered in subcluster B1 together with Salmonella phage vB_SenS_AG11. In subcluster A2, phage SB9 was clustered with SB10 and SB13 along with Salmonella phages S116, fuchur, and Escherichia phage Supergirl, respectively (Figure 5).
Of note, phages that were isolated from animals/animal products were predominantly found in subcluster C1. Relative to other clusters, cluster B contained phages isolated from more diverse sources and were also associated with the presence of tailspike proteins. For the phages with Salmonella as host, serotype Typhi was enriched in cluster A (Figure 5). Overall, these results reiterate the high genetic diversity that has been observed among S. enterica phages [25].
3.3. Salmonella Phages under Study Are Morphologically and Taxonomically Different
Phage morphological characterization by Transmission Electron Microscopy (TEM) was performed. Based on the TEM imaging, all the phages belonged to the Siphoviridae morphotype except SB18, which belonged to the Myoviridae morphotype (Table 1). Siphoviridae morphotypes have long flexible tails while Myoviridae morphotypes are defined by a rigid contractile tail (ICTV [64,65]. Images of phages representative of cluster A, B and C are shown in Figure 6, Figure 7 and Figure 8, respectively.
In-silico analysis of the isolated phages and their comparison with existing homologs allowed for taxonomical associations. With recent changes to taxonomic classification of phages where major families such as Siphoviridae, Podoviridae, and Myoviridae were abolished by the International Committee on Taxonomy of Viruses (ICTV [64,65] (https://ictv.global/vmr/current). Phylogenomic analysis was employed to classify the phages into classes and genera based on the new ICTV classification schema [66]. All the phages were classified into class Caudoviricetes. The phage SB6, SB9. SB10 and SB13 were classified as family Demerecviridae; subfamily Markadamsvirinae; genus Epseptimavirus with species level classification only for SB10 and SB13 viz. Epseptimavirus fuchur and Epseptimavirus SB13, respectively. Phage SB18 belonged to the same family as phages SB6, SB10 and SB13 but was classified as subfamily Ounavirinae; genus Kolesnikvirus. Phage SB28 represent a novel virus that was recently classified by ICTV as Macdonaldcampvirus at genus level and as Macdonaldcampvirus SB28 at species level. Phages SB3 and SB15, were classified as subfamily Guernseyvirinae and Jerseyvirus AG11 at genus level (https://viralzone.expasy.org/6319).
4. Discussion
Salmonella enterica is a major foodborne pathogen of global importance, and its genomic diversity has been widely studied [67,68,69,70,71,72]. Subspecies I contains more than 1,500 of the total 2,600+ serotypes in the species and is of most importance with respect to human infections [73]. Many studies have reported on the isolation and characterization of Salmonella phages, but these studies report on isolation of phages from only a few of the 1,500 serotypes from subspecies I, with a specific focus on those serotypes most commonly implicated in human disease [74]. In this study, we isolated Salmonella enterica phages from wastewater and assessed their diversity and host specificity using a combination of microscopic, biological, and genomic approaches. Phage isolation was conducted on a panel of highly diverse S. enterica isolates (Figure 1&2) representing 30 serotypes that are commonly associated with fresh produce outbreaks [75,76,77]. There are only few reports of phages isolated from many of the serotypes chosen in this study, allowing for an assessment of Salmonella phage diversity from food plant associated serotypes.
Wastewater is reported to be a rich source of phages, containing a vast diversity of both temperate and virulent phages that infect a wide array of host bacteria, including Salmonella [63,78]. In this study, eight S. enterica phages were isolated from the Jean-R. Marcotte WWTP in Montreal, which is the third largest WWTP in North America and provides wastewater treatment for the entire island of Montreal [79]. Montreal (population 4.34 million) is a large urban and diverse city with inhabitants from more than 50 nationalities [80], and a rich history of gastronomy, meaning that a wide variety of North American and ethnic foods are consumed within the city as a whole. Therefore, the size of the Jean-R. Marcotte WWTP and the diversity of wastewater treated there made it an ideal location for isolation of a diverse group of S. enterica phages.
Our results indicate that the phages isolated in this study had broad host ranges, as the majority of the Salmonella isolates used in this study were lysed by the phages. The broadest host range phages (SB3, SB6 and SB9) lysed more than 85 % of the isolates used for lytic spectra testing, which suggests the presence of one or more conserved receptors used by these phages to infect S. enterica and indicates that these phages could be good candidates for phage-based control strategy for reducing microbial contamination of food plant produce. In an earlier study, we demonstrated that phages SB3 and SB6 successfully reduced S. enterica populations on lettuce and cantaloupe tissues [12]. Future studies will focus on elucidation of the bacterial receptor/s used by phages SB3, SB6 and SB9 to better assess their potential for use in controlling foodborne contamination due to common and rare Salmonella serotypes. In addition, further studies on the potential of other phages isolated in this study to reduce S. enterica on food matrices are required to fully assess their biocontrol efficacy.
Whole genome-based phylogeny of the eight phages in this study revealed the uniqueness and high genetic diversity among them and as well as previously sequenced phages. The genome size, GC and gene contents of the phages were heterogenous. Comparative genomic analysis showed that, when compared to 113 phages from public databases, the eight phages from this study were grouped into three different clusters: cluster A (SB28, SB9, SB10 and SB13), cluster B (SB3 and SB15), and cluster C (SB6, SB18) (Figure 5). Phages are known to be heterogenous and have been recognized as one of the major drivers of diversity, evolution and adaption of their hosts in different environmental matrices, including wastewater ([17,19,20,21,22,23,24,25,81,82]. Salmonella Typhi phages were enriched in cluster A, whereas phages that were isolated from animals/animal products were enriched in cluster C. This diversity is reflected in the host and/or source associated clustering of the phages in the phylogeny [20].
Taxonomic classification of phages are based on electron microscopy and whole-genome sequencing [83]. The great majority of the phages sequenced in this study belonged to the Siphoviridae morphotype, while one (SB18) belonged to the Myoviridae morphotype. Two phages (SB13 and SB28) differed significantly from the previously sequenced phage genomes and represent novel species. Based on this observation, the bacterial and archaeal viruses subcommittee (BAVS) of ICTV has created two new species viz. Epseptimavirus SB13 and Macdonaldcampvirus SB28 [84]. Novel genus Macdonaldcampvirus has one more species Macdonaldcampvirus ViIIE1 attributed to Salmonella phage Vi II-E1 (AM491472.1). The knowledge of classification aids in the design of phage cocktails for biocontrol purposes, as phages with different morphotypes use different host receptors [85] and help to overcome phage resistance [86]. Indeed, phage cocktails containing different morphotypes could provide more effective in reducing bacterial loads in food products. In a previous study, the inclusion of phages SB3 and SB6 (Siphoviridae morphotype) from this study with three other phages belonging to the Myoviridae morphotype in a five phage-cocktail was effective in reducing Salmonella enterica on lettuce and cantaloupe flesh sections [12].
One of the safety concerns of using phages as biocontrol in food products is its propensity to harbour and/or facilitate horizontal gene transfer of antimicrobial resistance determinants in the environment [87,88,89]. In this study, none of the phages carried genes encoding virulence or antimicrobial resistance. Another concern arises from the pharmacological limitations of using phages as antimicrobials. For example, there is a significant size disparity between phage particles and antibiotic and other antimicrobial compounds, with phages being millions of times larger and composed of multiple proteins. This size discrepancy restricts dosing options, diminishes uptake and transportation rates [90]. To address this limitation, interest is increasingly turning to utilizing phage components as antimicrobials. The majority of this work has been conducted using phage lysins that are active against Gram-positive bacteria [91]. These enzymes are not active against Gram-negative bacteria due to the protective nature of the outer-membrane protein. More recently, several groups have demonstrated the antimicrobial effects of phage tail-spike proteins against Gram negative bacteria. Phage tailspike proteins are highly thermostable and protease resistant [92]. They possess carbohydrate depolymerase activity and recognize and cleave components of the lipopolysaccharide (LPS) to position the phage towards a secondary membrane receptor during infection [93]. Ayariga et al. [92] demonstrated that the ɛ34 phage tail spike protein has enzymatic property as a LPS hydrolase and synergizes with Vero Cell culture supernatant in killing Salmonella Newington. Miletic and colleagues [94] expressed the receptor binding domain of the Phage P22 Gp9 tailspike protein in plant tissue (Nicotiana benthamiana), and demonstrated that, upon oral administration of lyophilized leaves expressing Gp9 tailspike protein to newly hatched chickens, Salmonella concentrations were reduced on average by approximately 0.75 log relative to controls. Other studies led to reduced Salmonella motility and colonization [94,95,96]. In this study, three phages possessed tailspike proteins viz. SB3 (GenBank: QBQ74073.1); SB15 (GenBank: QHI00505.1) and SB18 (GenBank: QHI00609.1). Future work will include isolation and purification of these tailspike proteins, and analysis as potential antimicrobials to control S. enterica in foods.
5. Conclusions
In order to study Salmonella enterica bacteriophage diversity and host specificity, we isolated and characterized eight bacteriophages using microscopic, biological, and genomic approaches. Biological characterization by host range profile revealed that all eight phages were broad host range phages; genomically none of the phages possessed virulence, toxin, antibiotic resistance or lysogeny genes and we could classify them using their physical/morphological characterization. Phages SB3 and SB6 had been previously identified and proved to be good biocontrol candidates owing to their desirable characteristics. Phages SB9, SB13, SB18 and SB28 had the broadest host range and could be promising candidates for phage-based biocontrol, either alone or in a cocktail. This study reported two new phage species recognised by ICTV i.e Epseptimavirus SB13 and Macdonaldcampvirus SB28. Most of the phage genomes have a significant number of hypothetical proteins and this lack of understanding or the unknown functions of these proteins could be a limitation to the use of these (and other) phages as biocontrol agents. Nonetheless, our attempt to understand the diversity and host specificity of the isolated phages could contribute constructively to our understanding of phage biology and to better utilize this understanding in the development of biocontrol strategies in controlling Salmonella worldwide, in various environmental settings.
Author Contributions
Conceptualization, Lawrence Goodridge; Data curation, Sudhakar Bhandare and Opeyemi Lawal; Formal analysis, Sudhakar Bhandare and Opeyemi Lawal; Funding acquisition, Lawrence Goodridge; Investigation, Sudhakar Bhandare and Opeyemi Lawal; Methodology, Sudhakar Bhandare, Opeyemi Lawal, Anna Colavecchio, Brigitte Cadieux, Yella Zahirovich-Jovich, Zeyan Zhong, Elizabeth Tompkins, Margot Amitrano, Irena Kukavica-Ibrulj, Brian Boyle, Roger Levesque, Pascal Delaquis and Lawrence Goodridge; Project administration, Lawrence Goodridge; Software, Opeyemi Lawal; Supervision, Lawrence Goodridge; Visualization, Sudhakar Bhandare and Opeyemi Lawal; Writing – original draft, Sudhakar Bhandare and Opeyemi Lawal; Writing – review & editing, Sudhakar Bhandare, Opeyemi Lawal, Siyun Wang, Roger Levesque, Pascal Delaquis, Michelle Danyluk and Lawrence Goodridge.
Funding
This research was funded by a Large Scale Applied Research Project (LSARP) from Genome Canada, grant number 8505.
Acknowledgments
We thank researchers who contributed to and/or maintained The Salmonella Foodborne Syst-OMICS Database and researchers at the Institute for Integrative and Systems Biology, Université Laval.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Thomas, M.K.; Murray, R.; Flockhart, L.; Pintar, K.; Fazil, A.; Nesbitt, A.; Marshall, B.; Tataryn, J.; Pollari, F. Estimates of foodborne illness-related hospitalizations and deaths in Canada for 30 specified pathogens and unspecified agents. Foodborne pathogens and disease 2015, 12, 820-827. [CrossRef]
- Majowicz, S.E.; Musto, J.; Scallan, E.; Angulo, F.J.; Kirk, M.; O'Brien, S.J.; Jones, T.F.; Fazil, A.; Hoekstra, R.M.; International Collaboration on Enteric Disease 'Burden of Illness, S. The global burden of nontyphoidal Salmonella gastroenteritis. Clinical infectious diseases: an official publication of the Infectious Diseases Society of America 2010, 50, 882-889. [CrossRef]
- Grimont, P.A.D., Weill, F-X. Antigenic formulae of the Salmonella serovars. 9th edition. http://www.scacm.org/free/Antigenic%20Formulae%20of%20the%20Salmonella%20Serovars%202007%209th%20edition.pdf (Accessed on 13-2-2023) 2007.
- PHAC. Public Health Notice by Public Health Agency of Canada: Outbreak of Salmonella infections linked to peaches imported from the United States http://tinyurl.com/44wz9p9v. 2020.
- CFIA. Food safety investigation by Canadian Food Inspection Agency: Outbreak of Salmonella infections linked to red onions imported from the United States http://tinyurl.com/bddaefyn. 2020.
- PHAC. Public Health Notice by Public Health Agency of Canada: Outbreak of Salmonella infections linked to frozen whole kernel corn http://tinyurl.com/ywywz5m7. 2022.
- PHAC. Public Health Notice by Public Health Agency of Canada: Outbreak of Salmonella infections linked to Malichita and Rudy brand cantaloupes http://tinyurl.com/dm94wt82. 2024.
- Beamud, B.; Garcia-Gonzalez, N.; Gomez-Ortega, M.; Gonzalez-Candelas, F.; Domingo-Calap, P.; Sanjuan, R. Genetic determinants of host tropism in Klebsiella phages. Cell reports 2023, 42, 112048. [CrossRef]
- Bhandare, S.; Colom, J.; Baig, A.; Ritchie, J.M.; Bukhari, H.; Shah, M.A.; Sarkar, B.L.; Su, J.; Wren, B.; Barrow, P.; et al. Reviving Phage Therapy for the Treatment of Cholera. The Journal of Infectious Diseases 2018, jiy563-jiy563. [CrossRef]
- Fong, K.; LaBossiere, B.; Switt, A.I.M.; Delaquis, P.; Goodridge, L.; Levesque, R.C.; Danyluk, M.D.; Wang, S. Characterization of Four Novel Bacteriophages Isolated from British Columbia for Control of Non-typhoidal Salmonella in Vitro and on Sprouting Alfalfa Seeds. Frontiers in microbiology 2017, 8. [CrossRef]
- Vaz, C.S.L.; Voss-Rech, D.; Alves, L.; Coldebella, A.; Brentano, L.; Trevisol, I.M. Effect of time of therapy with wild-type lytic bacteriophages on the reduction of Salmonella Enteritidis in broiler chickens. Veterinary microbiology 2020, 240, 108527. [CrossRef]
- Wong, C.; Delaquis, P.; Goodridge, L.D.; Levesque, R.; Fong, K.; Wang, S. Inactivation of Salmonella enterica on post-harvest cantaloupe and lettuce by a lytic bacteriophage cocktail. Current Research in Food Science 2019. [CrossRef]
- Goodridge, L.; Fong, K.; Wang, S.; Delaquis, P. Bacteriophage-based weapons for the war against foodborne pathogens. Current Opinion in Food Science 2018, 20, 69-75. [CrossRef]
- Endersen, L.; Coffey, A. The use of bacteriophages for food safety. Current Opinion in Food Science 2020, 36, 1-8. [CrossRef]
- Ross, A.; Ward, S.; Hyman, P. More Is Better: Selecting for Broad Host Range Bacteriophages. Frontiers in microbiology 2016, 7. [CrossRef]
- Gal-Mor, O.; Boyle, E.C.; Grassl, G.A. Same species, different diseases: how and why typhoidal and non-typhoidal Salmonella enterica serovars differ. Frontiers in microbiology 2014, 5. [CrossRef]
- Switt, A.I.; den Bakker, H.C.; Vongkamjan, K.; Hoelzer, K.; Warnick, L.D.; Cummings, K.J.; Wiedmann, M. Salmonella bacteriophage diversity reflects host diversity on dairy farms. Food microbiology 2013, 36, 275-285. [CrossRef]
- Yamaki, S.; Yamazaki, K.; Kawai, Y. Broad host range bacteriophage, EscoHU1, infecting Escherichia coli O157:H7 and Salmonella enterica: Characterization, comparative genomics, and applications in food safety. International journal of food microbiology 2022, 372, 109680. [CrossRef]
- Fong, K.; Lu, Y.T.; Brenner, T.; Falardeau, J.; Wang, S. Prophage Diversity Across Salmonella and Verotoxin-Producing Escherichia coli in Agricultural Niches of British Columbia, Canada. Frontiers in microbiology 2022, 13, 853703. [CrossRef]
- Bryan, D.W.; Hudson, L.K.; Wang, J.; Denes, T.G. Characterization of a Diverse Collection of Salmonella Phages Isolated from Tennessee Wastewater. Phage (New Rochelle) 2023, 4, 90-98. [CrossRef]
- Rivera, D.; Moreno-Switt, A.I.; Denes, T.G.; Hudson, L.K.; Peters, T.L.; Samir, R.; Aziz, R.K.; Noben, J.P.; Wagemans, J.; Duenas, F. Novel Salmonella Phage, vB_Sen_STGO-35-1, Characterization and Evaluation in Chicken Meat. Microorganisms 2022, 10. [CrossRef]
- Thanki, A.M.; Brown, N.; Millard, A.D.; Clokie, M.R.J. Genomic Characterization of Jumbo Salmonella Phages That Effectively Target United Kingdom Pig-Associated Salmonella Serotypes. Frontiers in microbiology 2019, 10, 1491. [CrossRef]
- Sritha, K.S.; Bhat, S.G. Genomics of Salmonella phage PhiStp1: candidate bacteriophage for biocontrol. Virus genes 2018, 54, 311-318. [CrossRef]
- Chen, L.; Guan, G.; Liu, Q.; Yuan, S.; Yan, T.; Tian, L.; Zhou, Y.; Zhao, Y.; Ma, Y.; Wei, T.; et al. Characterization and complete genomic analysis of two Salmonella phages, SenALZ1 and SenASZ3, new members of the genus Cba120virus. Archives of virology 2019, 164, 1475-1478. [CrossRef]
- Sevilla-Navarro, S.; Catala-Gregori, P.; Marin, C. Salmonella Bacteriophage Diversity According to Most Prevalent Salmonella Serovars in Layer and Broiler Poultry Farms from Eastern Spain. Animals (Basel) 2020, 10. [CrossRef]
- Fong, K.; Tremblay, D.M.; Delaquis, P.; Goodridge, L.; Levesque, R.C.; Moineau, S.; Suttle, C.A.; Wang, S. Diversity and Host Specificity Revealed by Biological Characterization and Whole Genome Sequencing of Bacteriophages Infecting Salmonella enterica. Viruses 2019, 11. [CrossRef]
- Van Twest, R.; Kropinski, A.M. Bacteriophage enrichment from water and soil. Methods in molecular biology (Clifton, N.J.) 2009, 501, 15-21. [CrossRef]
- Bhandare, S.G. Biocontrol of V. cholerae using bacteriophage. University of Nottingham, UK., Nottingham, UK, 2015.
- Phagesdb.org. Plaque Purification. Phage hunting protocols 2013.
- Khan Mirzaei, M.; Nilsson, A.S. Isolation of phages for phage therapy: a comparison of spot tests and efficiency of plating analyses for determination of host range and efficacy. Plos One 2015, 10, e0118557. [CrossRef]
- Kutter, E. Phage host range and efficiency of plating. Methods in molecular biology (Clifton, N.J.) 2009, 501, 141-149. [CrossRef]
- Summer, E.J. Preparation of a phage DNA fragment library for whole genome shotgun sequencing. Methods in molecular biology (Clifton, N.J.) 2009, 502, 27-46. [CrossRef]
- Green, M.; Sambrook, J. Molecular cloning: A Lab Manual, 4th Edition ed.; Cold Spring Harbour Lab Press: 2012; Volume 1.
- Coil, D.; Jospin, G.; Darling, A.E. A5-miseq: an updated pipeline to assemble microbial genomes from Illumina MiSeq data. Bioinformatics 2015, 31, 587-589. [CrossRef]
- Brettin, T.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Olsen, G.J.; Olson, R.; Overbeek, R.; Parrello, B.; Pusch, G.D.; et al. RASTtk: a modular and extensible implementation of the RAST algorithm for building custom annotation pipelines and annotating batches of genomes. Scientific reports 2015, 5, 8365. [CrossRef]
- Wattam, A.R.; Brettin, T.; Davis, J.J.; Gerdes, S.; Kenyon, R.; Machi, D.; Mao, C.; Olson, R.; Overbeek, R.; Pusch, G.D.; et al. Assembly, Annotation, and Comparative Genomics in PATRIC, the All Bacterial Bioinformatics Resource Center. Methods in molecular biology (Clifton, N.J.) 2018, 1704, 79-101. [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. Journal of molecular biology 1990, 215, 403-410. [CrossRef]
- Punta, M.; Coggill, P.C.; Eberhardt, R.Y.; Mistry, J.; Tate, J.; Boursnell, C.; Pang, N.; Forslund, K.; Ceric, G.; Clements, J.; et al. The Pfam protein families database. Nucleic acids research 2012, 40, D290-301. [CrossRef]
- Kropinski, A.M.; Borodovsky, M.; Carver, T.J.; Cerdeno-Tarraga, A.M.; Darling, A.; Lomsadze, A.; Mahadevan, P.; Stothard, P.; Seto, D.; Van Domselaar, G.; et al. In silico identification of genes in bacteriophage DNA. Methods in molecular biology (Clifton, N.J.) 2009, 502, 57-89. [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: a novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic acids research 2002, 30, 3059-3066. [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: a fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol Biol Evol 2015, 32, 268-274. [CrossRef]
- Argimon, S.; Abudahab, K.; Goater, R.J.E.; Fedosejev, A.; Bhai, J.; Glasner, C.; Feil, E.J.; Holden, M.T.G.; Yeats, C.A.; Grundmann, H.; et al. Microreact: visualizing and sharing data for genomic epidemiology and phylogeography. Microb Genom 2016, 2, e000093. [CrossRef]
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.Y.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.V.; Cheng, A.A.; Liu, S.; et al. CARD 2020: antibiotic resistome surveillance with the comprehensive antibiotic resistance database. Nucleic acids research 2020, 48, D517-D525. [CrossRef]
- Chen, L.; Zheng, D.; Liu, B.; Yang, J.; Jin, Q. VFDB 2016: hierarchical and refined dataset for big data analysis--10 years on. Nucleic acids research 2016, 44, D694-697. [CrossRef]
- Liu, B.; Zheng, D.; Zhou, S.; Chen, L.; Yang, J. VFDB 2022: a general classification scheme for bacterial virulence factors. Nucleic acids research 2022, 50, D912-D917. [CrossRef]
- Shao, Y.; Harrison, E.M.; Bi, D.; Tai, C.; He, X.; Ou, H.Y.; Rajakumar, K.; Deng, Z. TADB: a web-based resource for Type 2 toxin-antitoxin loci in bacteria and archaea. Nucleic acids research 2011, 39, D606-611. [CrossRef]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: a genome comparison visualizer. Bioinformatics 2011, 27, 1009-1010. [CrossRef]
- Sambrook, J.; Russell, D., (Eds.) Molecular Cloning: a Laboratory Manual. 3rd Edition ed.; Cold Spring Harbor Laboratory: Cold Spring Harbor, NY, 2001.
- Jackson, B.R.; Griffin, P.M.; Cole, D.; Walsh, K.A.; Chai, S.J. Outbreak-associated Salmonella enterica serotypes and food Commodities, United States, 1998-2008. Emerging infectious diseases 2013, 19, 1239-1244. [CrossRef]
- McCormic, Z.D.; Patel, K.; Higa, J.; Bancroft, J.; Donovan, D.; Edwards, L.; Cheng, J.; Adcock, B.; Bond, C.; Pereira, E.; et al. Bi-national outbreak of Salmonella Newport infections linked to onions: the United States experience. Epidemiol Infect 2022, 150, e199. [CrossRef]
- Centers for Disease, C.; Prevention. Outbreaks of Salmonella infections associated with eating Roma tomatoes--United States and Canada, 2004. MMWR Morb Mortal Wkly Rep 2005, 54, 325-328.
- Centers for Disease, C.; Prevention. Multistate outbreaks of Salmonella serotype Poona infections associated with eating cantaloupe from Mexico--United States and Canada, 2000-2002. MMWR Morb Mortal Wkly Rep 2002, 51, 1044-1047.
- Walsh, K.A.; Bennett, S.D.; Mahovic, M.; Gould, L.H. Outbreaks associated with cantaloupe, watermelon, and honeydew in the United States, 1973-2011. Foodborne pathogens and disease 2014, 11, 945-952. [CrossRef]
- Gencay, Y.E.; Gambino, M.; Prussing, T.F.; Brondsted, L. The genera of bacteriophages and their receptors are the major determinants of host range. Environmental microbiology 2019, 21, 2095-2111. [CrossRef]
- Maffei, E.; Shaidullina, A.; Burkolter, M.; Heyer, Y.; Estermann, F.; Druelle, V.; Sauer, P.; Willi, L.; Michaelis, S.; Hilbi, H.; et al. Systematic exploration of Escherichia coli phage-host interactions with the BASEL phage collection. PLoS biology 2021, 19, e3001424. [CrossRef]
- Pickard, D.; Thomson, N.R.; Baker, S.; Wain, J.; Pardo, M.; Goulding, D.; Hamlin, N.; Choudhary, J.; Threfall, J.; Dougan, G. Molecular characterization of the Salmonella enterica serovar Typhi Vi-typing bacteriophage E1. J Bacteriol 2008, 190, 2580-2587. [CrossRef]
- Anany, H.; Switt, A.I.; De Lappe, N.; Ackermann, H.W.; Reynolds, D.M.; Kropinski, A.M.; Wiedmann, M.; Griffiths, M.W.; Tremblay, D.; Moineau, S.; et al. A proposed new bacteriophage subfamily: "Jerseyvirinae". Archives of virology 2015, 160, 1021-1033. [CrossRef]
- Hany, A. Biocontrol of foodborne bacterial pathogens using immobilized bacteriophages. University of Guelph, 2010.
- Olsen, N.S.; Hendriksen, N.B.; Hansen, L.H.; Kot, W. A New High-throughput Screening (HiTS) Method for Phages – Enabling Crude Isolation and Fast Identification of Diverse Phages with Therapeutic Potential. bioRxiv 2020, 2020.2003.2027.011080. [CrossRef]
- Lehman, S.M.; Kropinski, A.M.; Castle, A.J.; Svircev, A.M. Complete genome of the broad-host-range Erwinia amylovora phage phiEa21-4 and its relationship to Salmonella phage felix O1. Appl Environ Microbiol 2009, 75, 2139-2147. [CrossRef]
- Gill, J.J.; Svircev, A.M.; Smith, R.; Castle, A.J. Bacteriophages of Erwinia amylovora. Appl Environ Microbiol 2003, 69, 2133-2138. [CrossRef]
- Runa, V.; Wenk, J.; Bengtsson, S.; Jones, B.V.; Lanham, A.B. Bacteriophages in Biological Wastewater Treatment Systems: Occurrence, Characterization, and Function. Frontiers in microbiology 2021, 12. [CrossRef]
- Alharbi, N.M.; Ziadi, M.M. Wastewater as a fertility source for novel bacteriophages against multi-drug resistant bacteria. Saudi Journal of Biological Sciences 2021, 28, 4358-4364. [CrossRef]
- Zhu, Y.; Shang, J.; Peng, C.; Sun, Y. Phage family classification under Caudoviricetes: A review of current tools using the latest ICTV classification framework. Frontiers in microbiology 2022, 13, 1032186. [CrossRef]
- Turner, D.; Kropinski, A.M.; Adriaenssens, E.M. A Roadmap for Genome-Based Phage Taxonomy. Viruses 2021, 13. [CrossRef]
- Zerbini, F.M.; Siddell, S.G.; Lefkowitz, E.J.; Mushegian, A.R.; Adriaenssens, E.M.; Alfenas-Zerbini, P.; Dempsey, D.M.; Dutilh, B.E.; García, M.L.; Hendrickson, R.C.; et al. Changes to virus taxonomy and the ICTV Statutes ratified by the International Committee on Taxonomy of Viruses (2023). Archives of virology 2023, 168, 175. [CrossRef]
- Colavecchio, A.; D'Souza, Y.; Tompkins, E.; Jeukens, J.; Freschi, L.; Emond-Rheault, J.G.; Kukavica-Ibrulj, I.; Boyle, B.; Bekal, S.; Tamber, S.; et al. Prophage Integrase Typing Is a Useful Indicator of Genomic Diversity in Salmonella enterica. Frontiers in microbiology 2017, 8, 1283. [CrossRef]
- Hayden, H.S.; Matamouros, S.; Hager, K.R.; Brittnacher, M.J.; Rohmer, L.; Radey, M.C.; Weiss, E.J.; Kim, K.B.; Jacobs, M.A.; Sims-Day, E.H.; et al. Genomic Analysis of Salmonella enterica Serovar Typhimurium Characterizes Strain Diversity for Recent U.S. Salmonellosis Cases and Identifies Mutations Linked to Loss of Fitness under Nitrosative and Oxidative Stress. mBio 2016, 7, e00154. [CrossRef]
- Lankau, E.W.; Cruz Bedon, L.; Mackie, R.I. Salmonella strains isolated from Galapagos iguanas show spatial structuring of serovar and genomic diversity. Plos One 2012, 7, e37302. [CrossRef]
- Pightling, A.W.; Pettengill, J.; Luo, Y.; Strain, E.; Rand, H. Genomic diversity of Salmonella enterica isolated from papaya samples collected during multiple outbreaks in 2017. Microbiology 2020. [CrossRef]
- Simpson, K.M.J.; Mor, S.M.; Ward, M.P.; Collins, J.; Flint, J.; Hill-Cawthorne, G.A.; Abd El Ghany, M. Genomic characterisation of Salmonella enterica serovar Wangata isolates obtained from different sources reveals low genomic diversity. Plos One 2020, 15, e0229697. [CrossRef]
- Sodagari, H.R.; Mohammed, A.B.; Wang, P.; O'Dea, M.; Abraham, S.; Robertson, I.; Habib, I. Non-typhoidal Salmonella contamination in egg shells and contents from retail in Western Australia: Serovar diversity, multilocus sequence types, and phenotypic and genomic characterizations of antimicrobial resistance. International journal of food microbiology 2019, 308, 108305. [CrossRef]
- Lamas, A.; Miranda, J.M.; Regal, P.; Vazquez, B.; Franco, C.M.; Cepeda, A. A comprehensive review of non-enterica subspecies of Salmonella enterica. Microbiological research 2018, 206, 60-73. [CrossRef]
- Majtanova, L.; Majtan, J.; Majtan, V. Trends in phage types of Salmonella enterica serovars Enteritidis and Typhimurium isolated in Slovakia from 1995 to 2009. Diagnostic microbiology and infectious disease 2011, 69, 454-456. [CrossRef]
- Bennett, S.D.; Sodha, S.V.; Ayers, T.L.; Lynch, M.F.; Gould, L.H.; Tauxe, R.V. Produce-associated foodborne disease outbreaks, USA, 1998–2013. Epidemiology & Infection 2018, 146, 1397-1406. [CrossRef]
- Aiyedun, S.O.; Onarinde, B.A.; Swainson, M.; Dixon, R.A. Foodborne outbreaks of microbial infection from fresh produce in Europe and North America: a systematic review of data from this millennium. International Journal of Food Science & Technology 2020, 56, 2215-2223. [CrossRef]
- Hanning, I.B.; Nutt, J.D.; Ricke, S.C. Salmonellosis outbreaks in the United States due to fresh produce: sources and potential intervention measures. Foodborne pathogens and disease 2009, 6, 635-648. [CrossRef]
- Ballesté, E.; Blanch, A.R.; Muniesa, M.; García-Aljaro, C.; Rodríguez-Rubio, L.; Martín-Díaz, J.; Pascual-Benito, M.; Jofre, J. Bacteriophages in sewage: abundance, roles, and applications. FEMS Microbes 2022, 3. [CrossRef]
- Olson, I. Retrofitting world's 3rd largest treatment plant with ozonation and improved incinerators will take 10 years. CBC News https://www.cbc.ca/news/canada/montreal/montreal-wastewater-treatment-plant-1.6711208 2023.
- GovtCanada. Census Profile, 2021 Census of Population for Montreal https://tinyurl.com/53fks2ad. 2021.
- Ramisetty, B.C.M.; Sudhakari, P.A. Bacterial ‘Grounded’ Prophages: Hotspots for Genetic Renovation and Innovation. Frontiers in genetics 2019, 10. [CrossRef]
- Colello, R.; Ruiz, M.J.; Padín, V.M.; Rogé, A.D.; Leotta, G.; Padola, N.L.; Etcheverría, A.I. Detection and Characterization of Salmonella Serotypes in the Production Chain of Two Pig Farms in Buenos Aires Province, Argentina. Frontiers in microbiology 2018, 9. [CrossRef]
- Simmonds, P.; Aiewsakun, P. Virus classification - where do you draw the line? Archives of virology 2018, 163, 2037-2046. [CrossRef]
- Adriaenssens, E.; Brister, J.R. How to Name and Classify Your Phage: An Informal Guide. Viruses 2017, 9. [CrossRef]
- Shin, H.; Lee, J.H.; Kim, H.; Choi, Y.; Heu, S.; Ryu, S. Receptor diversity and host interaction of bacteriophages infecting Salmonella enterica serovar Typhimurium. Plos One 2012, 7, e43392. [CrossRef]
- Rohde, C.; Resch, G.; Pirnay, J.P.; Blasdel, B.G.; Debarbieux, L.; Gelman, D.; Gorski, A.; Hazan, R.; Huys, I.; Kakabadze, E.; et al. Expert Opinion on Three Phage Therapy Related Topics: Bacterial Phage Resistance, Phage Training and Prophages in Bacterial Production Strains. Viruses 2018, 10. [CrossRef]
- Borodovich, T.; Shkoporov, A.N.; Ross, R.P.; Hill, C. Phage-mediated horizontal gene transfer and its implications for the human gut microbiome. Gastroenterology Report 2022, 10. [CrossRef]
- Garvey, M. Bacteriophages and Food Production: Biocontrol and Bio-Preservation Options for Food Safety. Antibiotics (Basel) 2022, 11. [CrossRef]
- Villa, T.G.; Feijoo-Siota, L.; Rama, J.R.; Sánchez-Pérez, A.; Viñas, M. Horizontal Gene Transfer Between Bacteriophages and Bacteria: Antibiotic Resistances and Toxin Production. In Horizontal Gene Transfer: Breaking Borders Between Living Kingdoms, Villa, T.G., Viñas, M., Eds.; Springer International Publishing: Cham, 2019; pp. 97-142.
- Matsuzaki, S.; Yasuda, M.; Nishikawa, H.; Kuroda, M.; Ujihara, T.; Shuin, T.; Shen, Y.; Jin, Z.; Fujimoto, S.; Nasimuzzaman, M.D.; et al. Experimental protection of mice against lethal Staphylococcus aureus infection by novel bacteriophage phi MR11. J Infect Dis 2003, 187, 613-624. [CrossRef]
- Fischetti, V.A. Bacteriophage lysins as effective antibacterials. Current opinion in microbiology 2008, 11, 393-400. [CrossRef]
- Joseph, A.A.; Logan, G.; Honghzuan, W.; Robert, V. The E34 Phage Tailspike Protein: An in vitro characterization, Structure Prediction, Potential Interaction with <em>S. newington</em> LPS and Cytotoxicity Assessment to Animal Cell Line. bioRxiv 2021, 2021.2009.2020.461090. [CrossRef]
- Schmidt, A.; Rabsch, W.; Broeker, N.K.; Barbirz, S. Bacteriophage tailspike protein based assay to monitor phase variable glucosylations in Salmonella O-antigens. BMC microbiology 2016, 16, 207. [CrossRef]
- Miletic, S.; Simpson, D.J.; Szymanski, C.M.; Deyholos, M.K.; Menassa, R. A Plant-Produced Bacteriophage Tailspike Protein for the Control of Salmonella. Frontiers in plant science 2016, 6. [CrossRef]
- Waseh, S.; Hanifi-Moghaddam, P.; Coleman, R.; Masotti, M.; Ryan, S.; Foss, M.; MacKenzie, R.; Henry, M.; Szymanski, C.M.; Tanha, J. Orally administered P22 phage tailspike protein reduces salmonella colonization in chickens: prospects of a novel therapy against bacterial infections. Plos One 2010, 5, e13904. [CrossRef]
- Ibrahim, I.; Ayariga, J.A.; Xu, J.; Adebanjo, A.; Samuel-Foo, M.; Ajayi, O. CBD resistant Salmonella strains are susceptible to Epsilon 34 phage tailspike protein. 2022. [CrossRef]
Figure 1.
Maximum likelihood tree of 55 Salmonella representing 30 serovars used for phage propagation and host specificity in the study. Salmonella strains used for propagating the phages in the study are indicated with red star. Core-genome sequence alignment was generated from draft genomes using MAFFT v7.453, and the tree that was bootstrapped with 1000 replicates for node support was constructed using FastTree v2.1.11. The scale bar at the bottom represents nucleotide substitution per site.
Figure 1.
Maximum likelihood tree of 55 Salmonella representing 30 serovars used for phage propagation and host specificity in the study. Salmonella strains used for propagating the phages in the study are indicated with red star. Core-genome sequence alignment was generated from draft genomes using MAFFT v7.453, and the tree that was bootstrapped with 1000 replicates for node support was constructed using FastTree v2.1.11. The scale bar at the bottom represents nucleotide substitution per site.
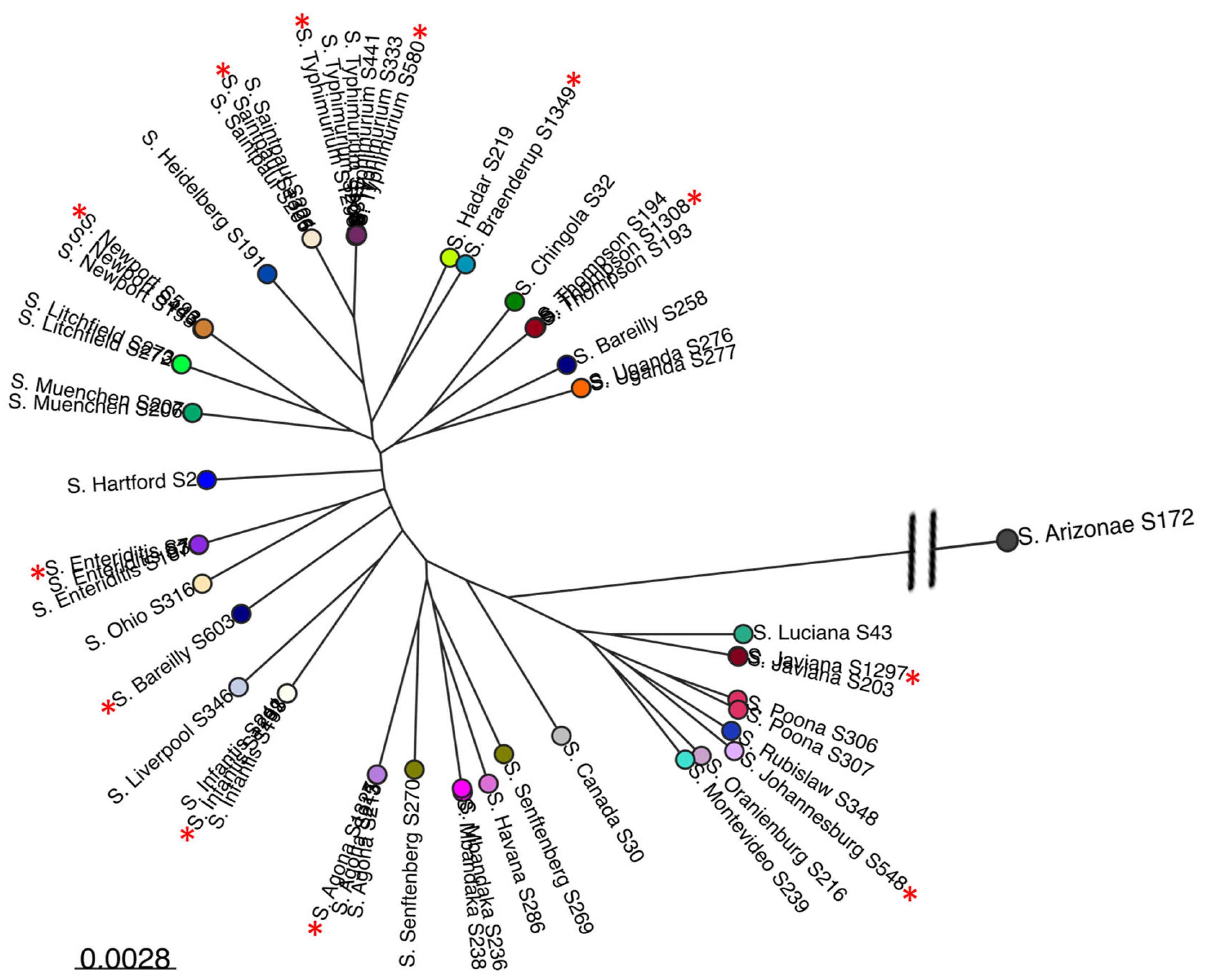
Figure 2.
Heat map of the lytic spectra of isolated phages with scoring of lysis, where 0 indicates no lysis and 1 to 3 indicates clearing (3 indicates complete clear lysis).
Figure 2.
Heat map of the lytic spectra of isolated phages with scoring of lysis, where 0 indicates no lysis and 1 to 3 indicates clearing (3 indicates complete clear lysis).

Figure 3.
Maximum likelihood phylogenetic tree showing the relatedness of phages under study. The phage genomes alignment was generated using MAFFT v7.453, and the tree that was bootstrapped with 1000 replicates for node support was constructed using IQtree v2.2. The scale bar at the bottom nucleotide substitution per site. Phages in light green rectangle belonged to cluster-A; those within blue rectangle are designated as cluster-B phages, while others were singletons. Grey and white box depicts presence or absence of tail spike proteins in the phages.
Figure 3.
Maximum likelihood phylogenetic tree showing the relatedness of phages under study. The phage genomes alignment was generated using MAFFT v7.453, and the tree that was bootstrapped with 1000 replicates for node support was constructed using IQtree v2.2. The scale bar at the bottom nucleotide substitution per site. Phages in light green rectangle belonged to cluster-A; those within blue rectangle are designated as cluster-B phages, while others were singletons. Grey and white box depicts presence or absence of tail spike proteins in the phages.
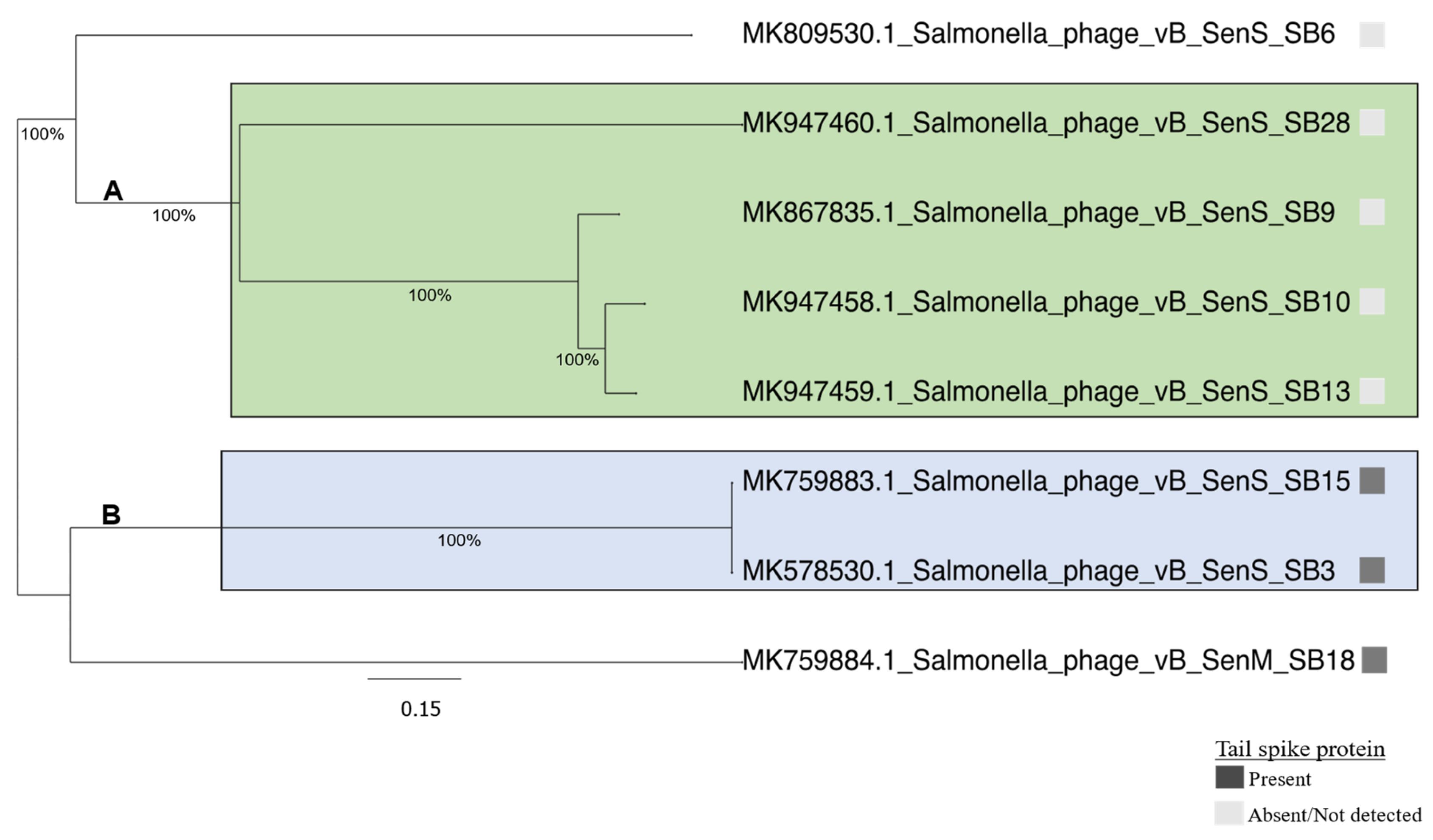
Figure 4.
Homology and gene synteny comparison of phages isolated in this study with previously sequenced phages. Blastn comparisons of the phages in this study with their closest references. (4A) SB9 and SB10 with Salmonella phage fuchur; (4B) SB13 with Escherichia phage Supergirl;. (4C) SB28 with the reference Salmonella phage E1; (4D) SB3 and SB15 with Salmonella phage vB_SenS_AG11; (4E) SB6 with Salmonella phage oselot and (4F) SB18 with Erwinia phage phiEa21-4. The direction of arrows indicates the DNA strand direction. Figures were generated using EasyFig.
Figure 4.
Homology and gene synteny comparison of phages isolated in this study with previously sequenced phages. Blastn comparisons of the phages in this study with their closest references. (4A) SB9 and SB10 with Salmonella phage fuchur; (4B) SB13 with Escherichia phage Supergirl;. (4C) SB28 with the reference Salmonella phage E1; (4D) SB3 and SB15 with Salmonella phage vB_SenS_AG11; (4E) SB6 with Salmonella phage oselot and (4F) SB18 with Erwinia phage phiEa21-4. The direction of arrows indicates the DNA strand direction. Figures were generated using EasyFig.
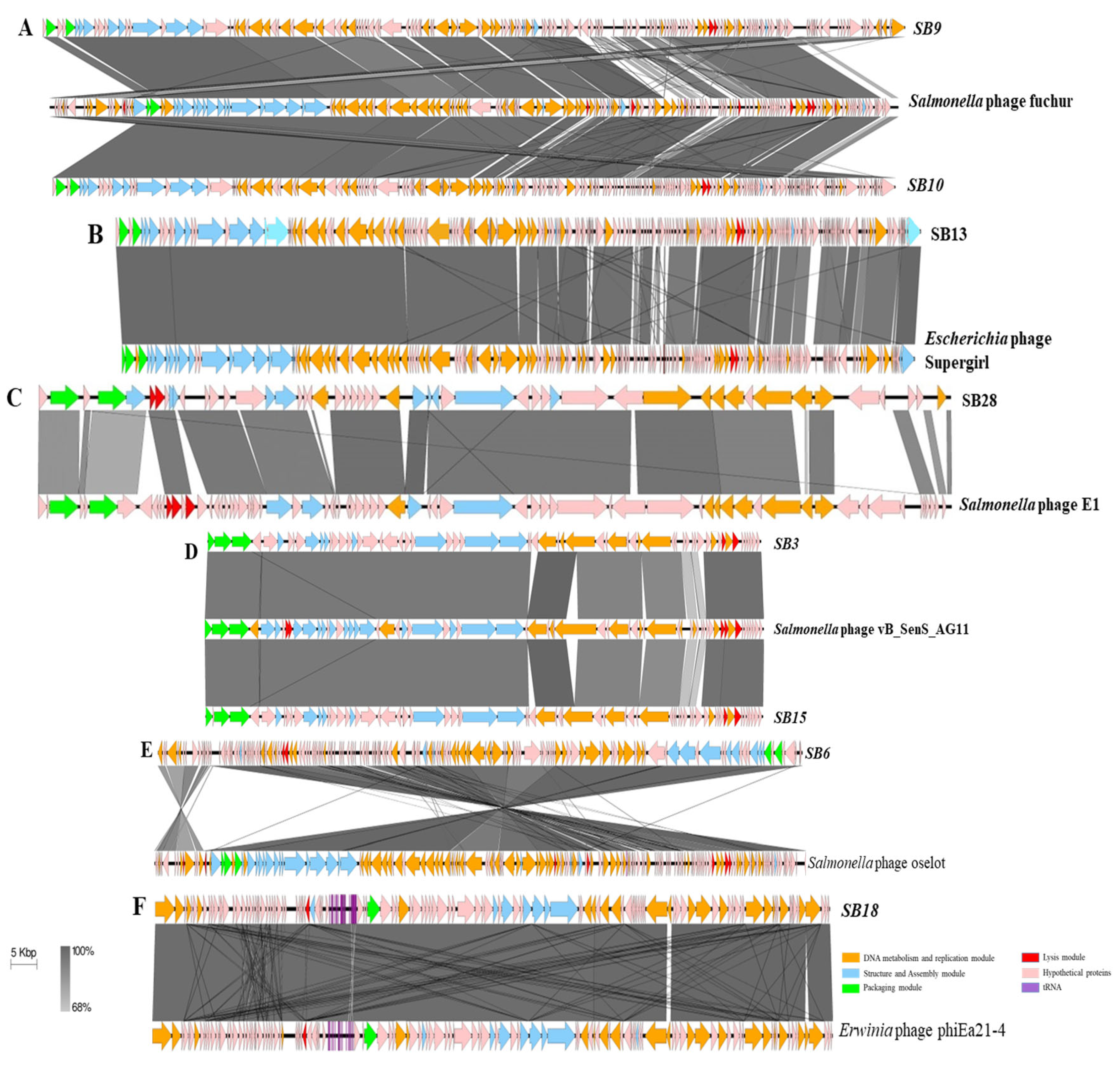
Figure 5.
Maximum likelihood phylogenetic tree showing the relatedness of phages under study with previously identified Salmonella phages having coverage of ≥80% and nucleotide homology of ≥ 95%. The phage genomes alignment was generated using MAFFT v7.453, and the maximum likelihood tree that was bootstrapped with 1000 replicates for node support was constructed using IQtree v2.2. The first three blocks showed the source, host, and serovars (for Salmonella host) of the phages, while the other blocks indicate the presence and absence of tail spike proteins. The scale bar at the bottom indicates nucleotide differences amongst them. The phages under study are indicated with red arrow.
Figure 5.
Maximum likelihood phylogenetic tree showing the relatedness of phages under study with previously identified Salmonella phages having coverage of ≥80% and nucleotide homology of ≥ 95%. The phage genomes alignment was generated using MAFFT v7.453, and the maximum likelihood tree that was bootstrapped with 1000 replicates for node support was constructed using IQtree v2.2. The first three blocks showed the source, host, and serovars (for Salmonella host) of the phages, while the other blocks indicate the presence and absence of tail spike proteins. The scale bar at the bottom indicates nucleotide differences amongst them. The phages under study are indicated with red arrow.

Figure 6.
Transmission Electron Microscopy (TEM) images of cluster A phages: subcluster A1: SB28 and subcluster A2: SB9, SB10 and SB13.
Figure 6.
Transmission Electron Microscopy (TEM) images of cluster A phages: subcluster A1: SB28 and subcluster A2: SB9, SB10 and SB13.
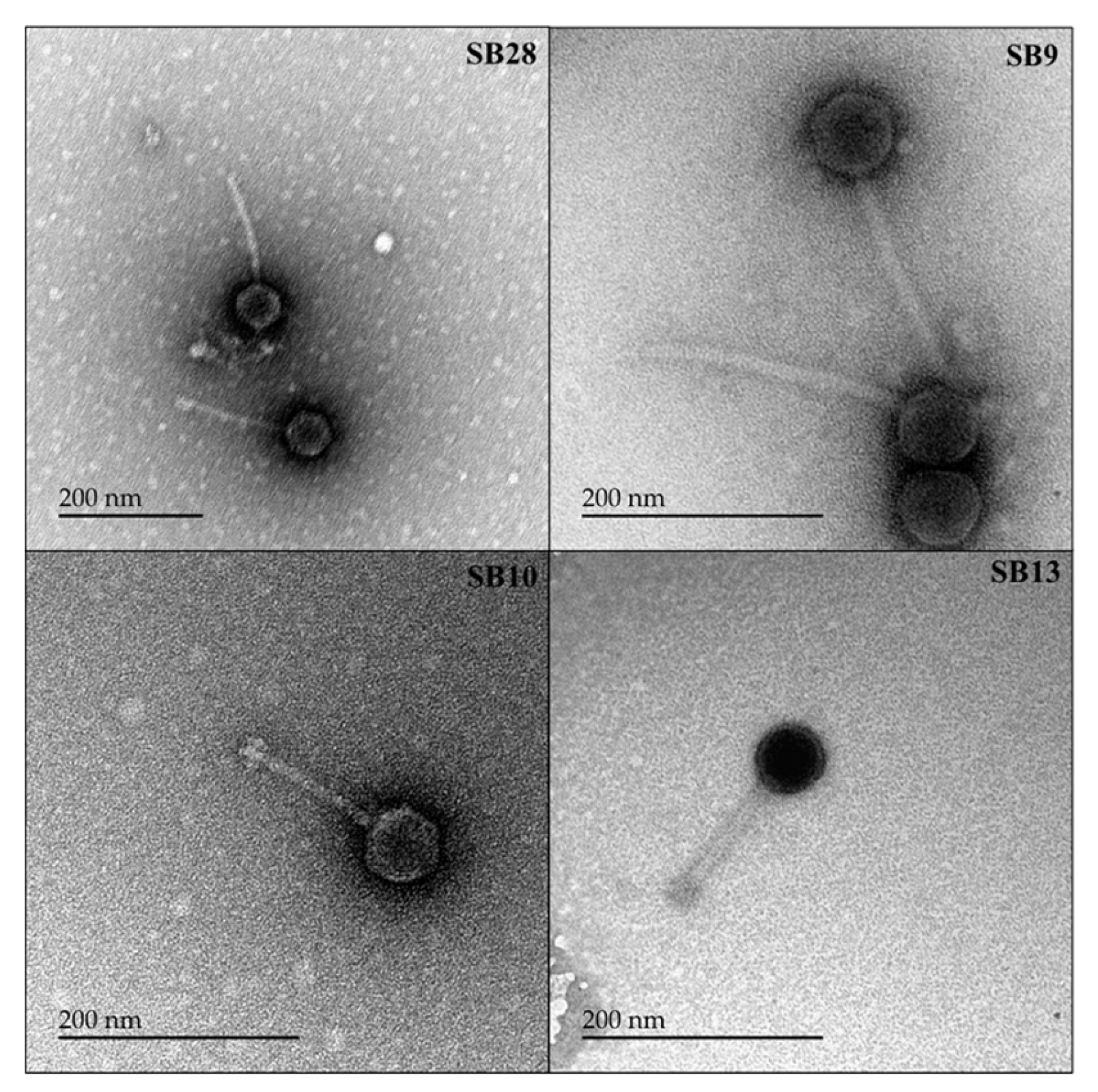
Figure 7.
Transmission Electron Microscopy (TEM) images of cluster B phages: SB3 SB15.
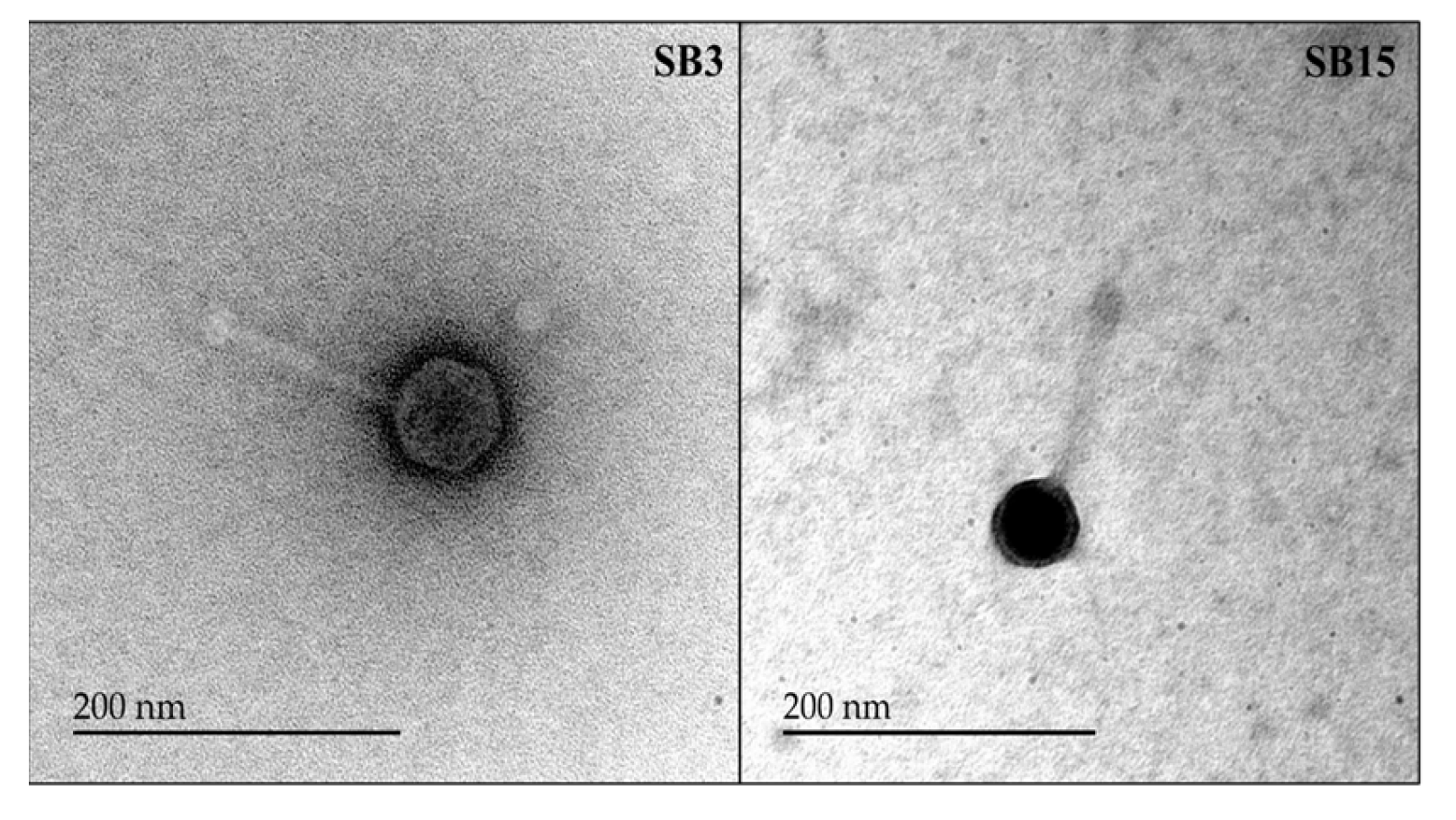
Figure 8.
Transmission Electron Microscopy (TEM) images of cluster C phages: subcluster C1: SB6 and subcluster C2: SB18.
Figure 8.
Transmission Electron Microscopy (TEM) images of cluster C phages: subcluster C1: SB6 and subcluster C2: SB18.
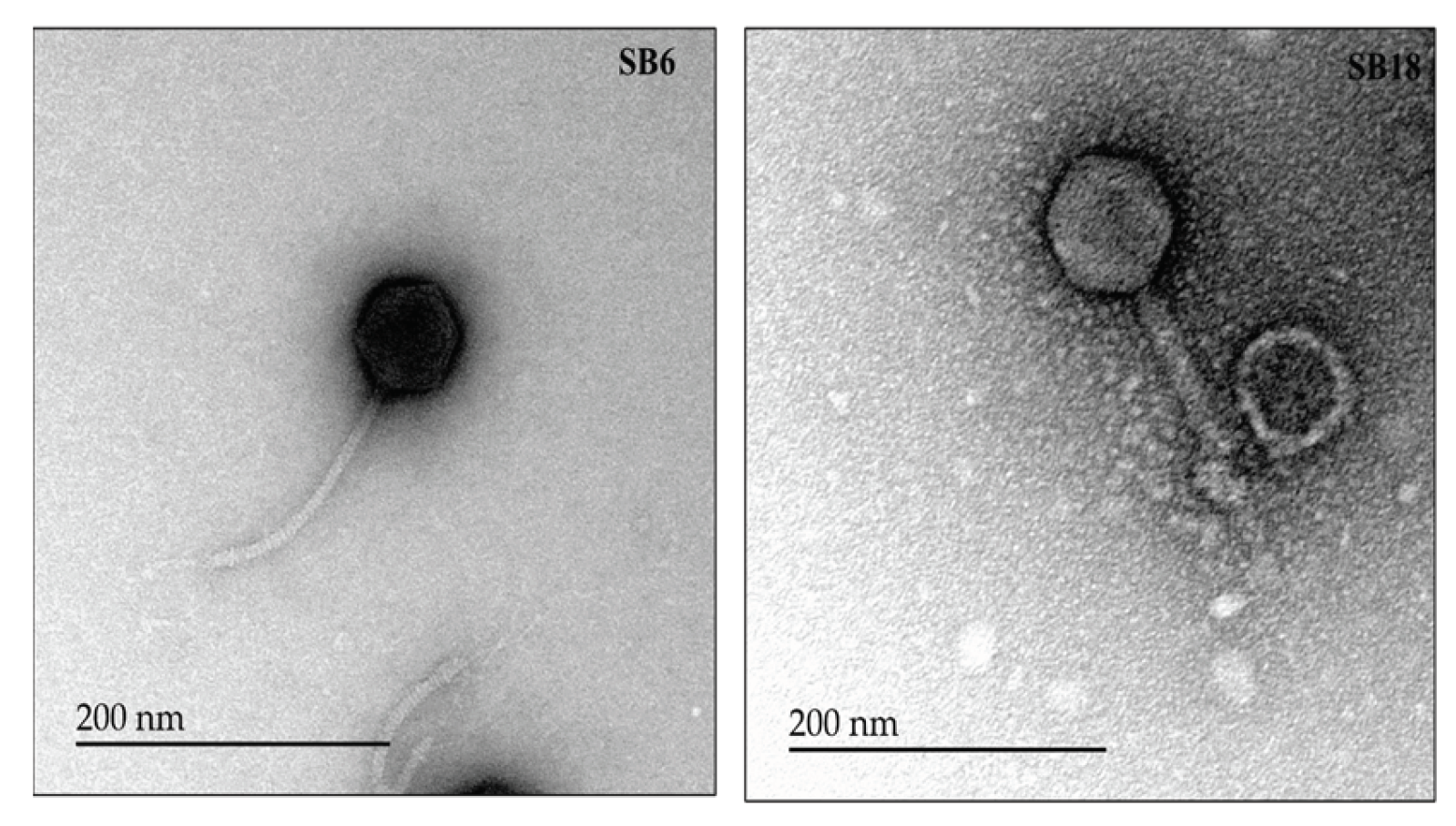

Table 1.
Summary of Salmonella enterica bacteriophages isolated in this study.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



