
Submitted:
15 February 2024
Posted:
16 February 2024
You are already at the latest version
Abstract
Diopatra neapolitana (Delle Chiaje, 1841) is one of the most exploited polychaete species in European waters, particularly in Ria de Aveiro, a coastal lagoon in mainland Portugal, where the overexploitation of this resource has led to a generalized decline of local populations. In an attempt to reduce the impact of harvesting, several management actions were implemented, but, illegal poaching still fuels a parallel economy that threats the sustainable use of this marine resource. The present study evaluated the combination of fatty acid profiles and elemental fingerprints of the whole body and jaws, respectively, of D. neapolitana collected from four harvesting locations within Ria de Aveiro in order to determine if their geographic origin could be correctly assigned post-harvesting. Results showed that both fatty acid profiles and elemental fingerprints differ significantly among locations, discriminating the geographic origin with higher accuracy when combining these two natural barcodes than when employing each individually. The present work can therefore contribute for the implementation of an effective management plan for the sustainable use of this marine resource, making possible to detect if D. neapolitana was sourced from no take zones and if it was collected from the place of origin claimed by live baits traders.
Keywords:
bait fishing
; harvesting
; polychaetes
; resource management
; Ria de Aveiro
; sustainability
1. Introduction
The collection of polychaetes in subtidal and intertidal mudflat habitats is a widespread activity worldwide, playing a key role in coastal economies [1,2,3]. Live polychaetes are used for multiple proposes, from maturation diets for crustacean and finfish broodstock in aquaculture facilities [4], to perform bioremediation processes of marine fish farms effluents [5,6], and as live baits for recreational and commercial fishing [2]. Several polychaetes species are exploited for these purposes, such as Arenicola marina, Atilla virens, Diopatra neapolitana, Glycera dibranchiata, Halla parthenopeia, Hediste diversicolor, Marphysa sanguinea, Namalycastis rhodochorde, Ophelia neglecta, Perinereis linea, Scoletoma impatiens and Sigalion squamosus [1,2]. Over the past decades, D. neapolitana has been one of the main polychaete species harvested in Ria de Aveiro, a coastal lagoon in mainland Portugal, as well as in other estuaries along the Portuguese coast line (e.g. Tagus estuary, Sado estuary and the coastal lagoon Ria Formosa) [2,7,8,9,10]. The overexploitation of this resource has prompted a decline in local populations [1] and, in an attempt to reduce the impact of the harvesting activity, several measures have subsequently been implemented. According to the Portuguese legislation (Portaria nº 1228/2010) bait gatherers can only operate with a personal license and are only allowed to work using hand gathering or a restricted gear. A more recent Ordinance was published in January 2014 (Portaria nº 14/2014) in an attempt to define the maximum daily catch limit. The daily limit is assumed to reflect the Maximum Sustainable Yield (MSY) of digging, ensuring a sustainable income to diggers.
According to this Ordinance, the daily catch limit for annelids should be 0.5 L day−1 per digger, excluding the tube [11]. In 1999, a commercial bait harvesting value of around 200 million euros was estimated for Europe but, the existing gaps in the supervision and regulation of this activity, could result in an underestimation of the real value [12]. In 2001/2002, the harvesting of D. neapolitana was quantified for the first time in Ria de Aveiro, with an estimate annual catch volume of around 45 tons and a commercial value of approximately 350 thousand euros [13]. Nevertheless, most commercial diggers, at a national and international level, do not have valid licenses, neither properly report their harvesting, as such, they contribute to foster to a parallel economy by not declaring their sales for tax purposes and impair any management plan ruling this activity to be successfully implemented [12,14]. The commercial potential of live fishing bait is so high that several attempts were made to intensively culture D. neapolitana [15,16]. Nonetheless, several constraints still hamper this approach and the harvesting of specimens from wild populations remains the sole supply source of this highly priced polychaete species.
The effects of bait harvesting on species density and population structure has a direct impact on the community (neglecting non-target species) and, consequently, on ecosystem functioning and processes [17,18]. The polychaetes population response to harvesting is influenced by the nature and extent of bait digging pressure and by the demographic characteristics of the population being exploited [14]. The current status of this species and the sustainability of its harvest are issues of growing concern [11]. As such, reliably tracing the geographic origin of polychaetes harvested from the wild could be paramount for a sustainable management plan for bait harvesting, that could foresee no take periods and areas, as well as expose illegal poaching activities by less scrupulous bait diggers.
At present, several tools have been applied to confirm the geographic origin of marine organisms [19,20]. Fatty acid (FA) profiles recorded in soft tissues (e.g. adductor muscle of bivalves), have been successfully used for this purpose in common cockles (Cerastoderma edule) [21,22] and manila clams (Ruditapes decussatus) [23]. The particular physicochemical conditions of each ecosystem shape FA composition of marine organisms in the sense that salinity and temperature are known to modulate the structure, fluidity and thus the composition of cell membranes [24]. Higher salinity fluctuations and/or lower water temperatures promote a decrease in the levels of saturated FA (SFA) and an increase in the concentration of polyunsaturated FA (PUFA), responsible for the stabilization of the bilayer structure [24]. Elemental fingerprints (EF) use the elemental profile recorded in hard biogenic structures, such as shells of common cockles (C. edule) [25,26], otoliths of California halibut (Paralichthys californicus) and Garibaldi (Hypsypops rubicundus) [27] and bony plates of Long-Snouted Seahorse (Hippocampus guttulatus) [28]. Considering that elements are influenced by the environmental and chemical features of each ecosystem [29] and that these mineral structures grow throughout the year, EF have already been successfully used to discriminate specimens originating from geographically close locations [30,31,32].
The combination of two different traceability tools can be more effective to confirm the geographic origin of marine organisms, as already confirmed by Zhang, et al. [33] and Perez, et al. [34] that showed that the use of stable isotope ratios in combination with FA profiles could successfully discriminate scallop species (Patinopecten yessoensis, Chlamys farreri, and Argopecten irradians) and warty venus (Venus vecurrosa), respectively, from different geographic locations. This combination of tools was also able to trace the geographic origin and seasonality of the whitemouth croaker (Micropogonias furnieri) [35]. Moreover, Matos, et al. [36], using stable isotopes and elemental fingerprints effectively traced the geographic origin of eastern oysters (Crassostrea virginica).
In an attempt to contribute to a better management of D. neapolitana stocks, the present study tested, for the first time, if the combination of FA profiles of the whole polychaete body and the EF of its jaws differs between specimens originanting from different locations in Ria de Aveiro a coastal lagoon in mainland Portugal where the capture of this polychaete being used as live bait for sports fishing is an important economic and social activity.
2. Materials and Methods
2.1. Sample collection
A total of forty specimens of D. neaplitana were collected using a shovel, mimicking the method used by professional bait collectors, in Espinheiro (E), Ílhavo (I) and Mira (M1 and M2) channels located in Ria de Aveiro, mainland Portugal (Figure 1); this coastal lagoon is one of the most important locations for the harvesting of D. neapolitana for recreational and commercial fishing in mainland Portugal [2,37]. Ten specimens of D. neapolitana were sourced from each location described above, stored in aseptic plastic boxes and kept refrigerated during sampling and transport to the laboratory. Upon arrival at the laboratory, specimens were left to depurate during 24 h in containers with artificial seawater (prepared by mixing Tropic Marin Pro Reef salt (Tropic Marin, Wartenberg, Germany) and freshwater purified by a reverse osmosis unit). All specimens were split up in two sub-groups: whole body for FA analysis and jaw for elemental analysis (4 locations X 2 methods X 10 replicates = 80 samples). The jaws from each specimen were dissected using a scalpel with ceramic-coated blades. Subsequently, the whole body and jaws were individually homogenized using a mortar grinder (RM 200, Retsch, Hann, Germany) and stored at -80 ºC until further analysis.
2.2. Fatty acid analysis
Methyl esters of fatty acids (FAME) of D. neapolitana bioamass (~50 mg) were prepared following the method described by Aued-Pimentel, et al. [38] (through transmetylation of FA using a mixture of methanolic solution KOH (2 M) and saturated NaCl). The resulting FAME were analyzed in a QP2010 Ultra, Shimadzu gas chromatography-mass spectrometry, equipped with an auto-sampler, a DB-FFAP column with 30 m length, 0.32 mm internal diameter and 0.25 µm film thickness (J&W Scientific, Folsom, CA). The column was initially programed to 80 °C, increasing 25 °C min-1 until 160 °C, 2 °C min-1 from 160 to 220 °C and 30 °C min-1 from 220 to 250 °C, using helium as the carrier gas, at a flow of 1.8 mL min-1. FAME identification was accomplished through comparison of retention times with those mixed FAME standards (C4-C24, Supelco 37 Component Fame Mix) and by comparison of the mass spectrum of each relative to standard spectra from the library “AOCS Lipid Library” (http://lipidlibrary.aocs.org/).
2.3. ICP-MS
The jaws of D. neapolitana (~50 mg) were weighed in digestion tubes (DigiTUBEs). Digestion tubes were soaked with high-purity concentrated HNO3, HCl (37%) and H2O2 (30% w/v) on a digestion block (DigiPrep, SCP Science, Canada) at 85 °C and over 15 minutes. Subsequently, solutions were diluted with Milli – Q (Millipore) water to a final HNO3 concentration of 1-2% to reduce acid concentration and prevent damage to the equipment. Finally, total concentrations of aluminum (Al), barium (Ba), calcium (Ca), cerium (Ce), cobalt (Co), iron (Fe), potassium (K), lanthanum (La), magnesium (Mg), manganese (Mn), sodium (Na), nickel (Ni), phosphorus (P), strontium (Sr), and yttrium (Y) were analysed using an Agilent 7700 ICP-MS equipped with an octopole reaction system (ORS) collision/reaction cell technology to minimize spectral interferences. Blanks and certified reference materials BCS-CRM-513 (SGT Limestone 1) were treated using the same method.
2.4. Data and statistical analysis
2.4.1. Analysis of fatty acid profiles
The FA profiles was represented by the relative abundance of the total pool of FA per replicate for each location. The FA were grouped by classes as saturated FA (SFA), monounsaturated FA (MUFA) and polyunsaturated FA (PUFA). A Boruta analysis was performed to select the most relevant FA to discriminate specimens originating from different sampling locations. Statistical differences (p < 0.05) in FA profiles among locations were tested using vegan adonis() function for permutational multivariate analysis of variance (PERMANOVA) [39] using the Bray-Curtis similarity coefficient. To test differences among locations for each FA individually, a one-way analysis of variance (ANOVA) was performed. A Radom Forest (RF) classification was used to evaluate the possibility of successfully discriminating the geographic origin of specimens through the FA profile. To test for normality and the assumption of homogenous variances, the Shapiro.test() and bartlett.test() functions were used, respectively, and data was transformed (log x+1). All statistical analyses were performed using R [40].
2.4.2. Analysis of elemental fingerprint
The concentration of elements present in the jaws of D. neapolitana was expressed as a ratio relatively to Ca (mg/mg) in order to minimize mass effects [25,28,41]. For a better understanding of our results, a Boruta analysis was used to determine the most important elements to discriminate between different sampling locations. A resemblance matrix using the ratio of each element per sample was prepared using Euclidean distance with the vegdist() function after performing a scale () transformation. A PERMANOVA was performed to detect significant differences in the EF of jaws from D. neapolitana originating from different locations. A one-way ANOVA was used to assess differences among locations for each individual element after confirming normality with the Shapiro test and homogeneity of variance with the Bartlett test. A RF classification was performed to evaluate the potential use of elements present in the jaws of this polychaete to discriminate between different locations. All statistical analyses were performed using R [40].
2.4.3. Combination of fatty acid profiles and elemental fingerprint
In order to increase the number of predictor variables, a combination of FA profiles and EF was used. This combination underwent a Boruta analysis to select the best subset of variables that may explain potential differences in specimens of D. neapolitana originating from different sampling locations [42]. PERMANOVA was performed to assess differences among locations and a RF classification was applied to test if the combination of FA and EF could be used to predict the geographic origin from which each polychaete was collected. All data were scale() transformed and used to produce a matrix using Euclidean distances with the vegdist() function in the vegan package. All statistical analyses were performed using R [40].
3. Results
3.1. Fatty acid profiles
The average FA profiles recorded for the whole body of D. neapolitana at different locations is presented in Table S1 (as supplementary data). A total of twenty-four FA was identified, comprising eight SFA, six MUFA and ten PUFA. SFA comprised 40% to 50% (location M2 and I, respectively; Table S1) of all FA identified in the whole body of D. neapolitana from different locations. The main SFA were palmitic acid (16:0) and stearic acid (18:0), corresponding to more than 76% of all SFA. MUFA represented 14% to 20% (locations M1 and E, respectively; Table S1) of all FA. Dominant MUFA were vaccenic (18:1n-7) and eicosenoic (20:1n-9) acid, representing over 50% of all MUFA. PUFA comprised 32% to 43% (locations E and M2, respectively; Table S1) of all FA. The most abundant PUFA were eicosapentaenoic (20:5n-3; EPA) and docosadienoic (22:2n-9; DHA) acid, representing over 42% of all PUFA.
The Boruta analysis performed showed that the FA 18:1n-7, 20:0, 18:2n-6, 16:0, 15:0, 22:4n-6, 17:0, 16:1n-7, 20:4n-6, 18:3n-3 and 18:0 (Figure 2a) were the ones that most contributed for the differences recorded among locations. The PERMANOVA performed revealed the existence of significant differences among locations, apart from those within the Mira Channel (M1 and M2) (p < 0.05; Table S2 on supplementary data). Considering each FA individually, FA 22:0 and 22:3n-6 did not display any significant difference among locations (Table S2). Comparatively to the other locations, specimens from location E recorded significantly higher levels of 15:0, 16:0, 17:0, 18:1n-7 and 18:2n-6 (Table S2). Specimens from location M2 presented the highest levels of 16:1n-7, 20:4n-6 and 22:4n-6, being significantly different from other locations (Table S2). Location I presented higher levels of FA 18:0, with significant differences between this location and the others, and the sole exception was with M1.
The RF classification showed an overall accuracy of 75% (Table 1 and Figure 3a). Specimens from location E exhibited the highest percentage of correct classification (100%). Specimens of D. neapolitana originating from Mira Channel (M1 and M2) registered three misclassified replicates, resulting in a correct classification of 70%. Most misclassifications were associated with specimens sourced from location I, with only 60% of correct classification (Figure 3a and Table 1).
3.2. Elemental fingerprints
The concentration of the fourteen elements recorded on the EF determined for the jaws of D. neapolitana in the different locations of Ria de Aveiro is shown in Figure S1 (supplementary data). The Boruta analysis revealed that Ba, Mn, K, Na and Ni were the elements that most contributed for the differences recorded among locations (Figure 2b). The PERMANOVA revealed significant differences among locations, with the exception for specimens collected in locations E vs I and M1 vs I (Table S2 on supplementary data). Concerning individual elements, the highest concentration of Ba was registered in location M2, with significant differences occurring between locations M2 vs E, M2 vs I and M1 vs E (Figure S1). The jaws of D. neapolitana presented the highest levels of Mn also in location M2 with significant differences being recorded between this location and location E. Specimens from M1 registered significantly higher concentrations of Na, comparatively with locations E and I (Figure S1). Concerning, the Ni, the highest concentrations were recorded in specimens from locations M2 and I, whereas K was more concentrated in specimens from location M1, with no significant differences being recorded among locations for both of these elements (Figure S1).
The RF classification revealed an accuracy of 72.5%, when using EF to allocate D. neapolitana to their geographical origin. Specimens from location I showed the highest percentage of correct allocation (90%; one replicate misclassified), followed by those originating from location M1, with an accuracy of 80% (two replicates were misclassified). Most misclassifications were associated with D. neapolitana collected in locations E and M2, with 40% of collected specimens being erroneously assigned to other locations (Figure 3b and Table 1).
3.3. Elemental fingerprints
The Boruta analysis showed that the most relevant combination of predictive variables when simultaneously considering FA profiles and EF were Ba, 18:1n-7, 16:0, 18:2n-6, 20:0, 15:0, 17:0, 22:4n-6, 16:1n-7, 20:4n-6, 18:3n-3 and 18:0 (Figure 2c). The PERMANOVA revealed the existence of significant differences among all locations (Table S2). The RF classification resulted in an overall accuracy of 85% (Figure 3c and Table 1). Location E had the highest percentage of correct allocation (100%), whereas two replicates from locations I, M1 and M2 were misclassified, resulting in an overall correct allocation of 80%.
4. Discussion
The use of FA profiles of soft tissues and EF of mineral structures in marine species has been optimized in order to put forward faster, more accurate and environmentally safer methods to discriminate the geographic origin of these organisms, namely those that feature an important commercial value [23,28,43,44,45,46,47,48]. Most of available studies to date using these methods, either when these are applied individually or combining more than one approach (e.g., FA profiles and stable isotopes [34], elemental fingerprint and stable isotopes [36]), are mostly focused on food safety issues, rather than in the implementation of effective management plans for endogenous marine resources that may be vulnerable to poaching. The present study showed, for the first time, that the combination of biogeochemical tools (FA profiles and EF) using the whole body and the jaws of D. neapolitana (respectively) can successfully be used to confirm the geographic origin of bait digging with a high accuracy level (Table 1). It is therefore legitimate to say that these natural barcodes can be successfully used towards the implementation of more effective fishery management plans, allowing the enforcement of no take zones.
The FA profiles displayed by the whole body of D. neapolitana revealed as the most dominant FA the 16:0, followed by the PUFA 20:5n-3. In general, this trend was similar to that found for other polychaetes by Fernandes, et al. [49] for Hediste diversicolor and by Jerónimo, et al. [50] for H. diversicolor, D. neapolitana, Sabella cf. pavonina and Terebella lapidaria, all collected in the same location in Ria de Aveiro. The same was found for Alvinella pompejana [51], Arenicola marina [52], Nepthys hombergii and Lanice conchilega [53] sampled in others locations.
The FA belonging to the SFA and PUFA classes were responsible for most of the differences recorded among locations (p < 0.05; Table S1 on supplementary data). These dissimilarities in FA profiles of polychaetes among locations were likely associated with a differential physiological response to changes in environmental conditions (e.g. salinity and temperature) that shape the environment on their sampling locations [22,49,54,55]. At higher temperatures, the reorganization of the membrane structure is needed to maintain membrane fluidity and homeostasis, leading to an increase in FA saturation or the prevalence of shorter-chain FA [56]. Salinity is responsible for changes in FA profiles involved in the osmoregulation process, inducing changes in membrane permeability [57]. Higher salinities, like those registered at M1, I and E are associated with a decrease in PUFA to reduce membrane permeability [58].
The biogenic carbonate hard parts of marine species, such as shells, otoliths, plates, fish scales and fish bones, incorporate and retain elements from the surrounding environment throughout their lifetimes [28,47,59,60,61,62]. It is important to note that this study represents the first dataset reporting the EF of polychaete jaws. Similar to other mineral structures, (e.g. Cerastoderma edule shells [26,63]), the levels of elements recorded in aragonitic jaws of D. neapolitana [64] differed among locations within Ria de Aveiro. These discrepancies are associated with both local physical conditions and elements availability in the environment. Considering that location M2 is more upstream than M1 (within the same channel and closer to the inlet), hydrological conditions differ, which result in a substantial enrichment of Ba and Mn upstream due to freshwater inputs and nutrient runoff, as suggested by Ricardo et al., [26]. In fact, the presence of high levels of Ba and Mn in location M2 had already been previously reported for these authors in C. edule shells [26]. The highest concentrations of K and Na observed in M1 could be related with its geographical proximity to the inlet of the coastal lagoon, suggesting a potential association with the physicochemical properties of oceanic seawater (e.g. temperature, salinity [65,66]). Locations I and M2 registered the highest levels of Ni in D. neapolitana jaws. This trend was previously reported for the body of D. neapolitana by Pires, et al. [67] exactly on those same locations. High levels of Ni could be associated with anthropogenic impacts [68,69], such as the presence of an important shipyard, commercial harbor, and industrial activities in location I [70], along with agriculture runoff in location M2 [71].
The use of Random Forest classifications based on FA profiles and EF enhanced the discriminations of geographic origin between specimens of D. neapolitana (Table 1). Indeed, the use of each tool individually was associated with some constrains. When FA profiles were used alone to determine the geographic origin of collected specimens, location E was well discriminated, contrarily to the other locations. When using EF, locations I and M1 were discriminated with a high level of accuracy, contrarily to locations E and M2 (Table 1). Thus, the combination of FA profiles and EF showed to be a more efficient approach to successfully allocate sampled specimens of D. neapolitana to their true geographic origin, thus increasing the success rate (Table 1). The combination of different fingerprinting methods (e.g. FA profiles with stable isotopes or multi-elements with stable isotope) had already been successfully employed to discriminate the geographic origin of different species. When using FA profiles combined with stable isotope analysis, the geographic origin of sea cucumbers (Apostichopus japonicus) [72], jumbo squids (Dosidicus gigas) and scallops (Patinopecten yessoensis, Chlamys farreri, and Argopecten irradians) [33] was determined with high success rate of correct allocations. Employing a multi-elements and stable isotope approach for different shrimp species (Penaeus indicus, P. merguiensis, P. monodon, P. notialis, P vannamei, Pleoticus muelleri, and Pandalus borealis) was also highly successful when aiming to allocate their geographic origin [73]. These approaches were also used to distinguish production methods (farmed vs. wild), namely by using FA profiles and stable isotopes on European eel (Anguilla anguilla) [74], and Atlantic salmon (Salmo salar) [75], as well as for shrimps when using multi-elements and stable isotope analysis (see above; [73]).
5. Conclusions
The present study showed that the combination of FA profiles of the body of D. neapolitana and EF of its jaws is an accurate and reliable traceability tool that can be used to confirm the geographic origin of live specimens of this species sourced from different locations within a coastal lagoon with a resolution <2 Km. Despite being an initial study, the results here presented can play a key role to support the implementation of conservation/management plans for bait harvesting activities in this coastal lagoon and elsewhere, namely estuarine systems where bait harvesting is more intense and relevant from an economic, social and ecological perspective. To improve the potential use of this tool, further insights are required on the existence of temporal variability (both seasonal and interannual), which pinpoints to how frequently the predictive model needs to be calibrated to secure an accurate classification of the place of origin of the specimens being surveyed. Extending the application of this methodology to other polychaete species with commercial interest could also be important to contribute to a more a sustainable use of these important marine living resources.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Ratios of elements to Calcium (Ca) concentrations (mg to mg) (average ± SD; n = 10) of Diopatra neapolitana jaws from four locations within the coastal lagoon Ria de Aveiro, mainland Portugal: Espinheiro Channel (E:), Ílhavo Channel (I) and Mira Channel (M1 and M2). Different letters indicate significant differences among sampling locations at p < 0.05.; Table S1: Fatty acid profiles (data presented as percentage of relative abundances) of the whole body of Diopatra neapolitana (values are means of 10 replicates ± SD) from Espinheiro (E), Ílhavo (I) and Mira (M1 and M2) channels in the coastal lagoon Ria de Aveiro, mainland Portugal. SFAs – saturated fatty acids; MUFAs – monounsaturated fatty acids; and PUFAs – polyunsaturated fatty acids. Different superscript letters indicate significant differences among sampling locations at p < 0.05.; Table S2: Permutational multivariate analysis of variance (PERMANOVA) among fatty acid profiles of whole body, elemental ratios of jaws and combination of fatty acid profiles of whole body and elemental ratios of jaws of the polychaete Diopatra neapolitana sourced from Espinheiro Channel (E:), Ílhavo Channel (I) and Mira Channel (M1 and M2). Significant differences p < 0.05.
Author Contributions
Conceptualization, F.R. and R.C; methodology, F.R., M.L.L., and R.M.; validation, M.R.D., C.P. and R.C.; formal analysis, F.R. and R.C.; investigation, F.R., M.R.D., C.P. and R.C.; resources, M.R.D., C.P. and R.C.; writing—original draft preparation, F.R. and R.C.; writing—review and editing, F.R., M.L.L., R.M., M.R.D., E.F.S., C.P. and R.C.; visualization, F.R.; supervision, M.R.D., E.F.S., C.P. and R.C.; project administration, R.C.; funding acquisition, C.P. and R.C. All authors have read and agreed to the published version of the manuscript.
Funding
This work was financially supported by project CITAQUA, “Desenvolvimento do Projeto de Reforço do Polo de Aveiro (H4)”, framed within Measure 10 of Investment TC-C10-i01 - Hub Azul - Rede de Infraestruturas para a Economia Azul, financed by the Recovery and Resilience Plan (PRR) and supported by Fundo Azul of the Portuguese Government. The authors acknowledge the University of Aveiro, Fundação para a Ciência e a Tecnologia (FCT, Portugal) and Ministério da Ciência e Tecnologia (MCT) for the financial support for the research units CESAM (UIDP/50017/2020 + UIDB/50017/2020 + LA/P/0094/2020) and LAQV-REQUIMTE (FCT UIDB/50006/2020).
Data Availability Statement
Acknowledgments
We thank Andreia Santos for their technical support.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Olive, P. Management of the exploitation of the lugworm Arenicola marina and the ragworm Nereis virens (Polychaeta) in conservation areas. Aquat. Conserv. 1993, 3, 1–24. [Google Scholar] [CrossRef]
- Cunha, T.; Hall, A.; Queiroga, H. Estimation of the Diopatra neapolitana annual harvest resulting from digging activity in Canal de Mira, Ria de Aveiro. Fish. Res. 2005, 76, 56–66. [Google Scholar] [CrossRef]
- Olive, P. Polychaeta as a world resource: a review of pattern of exploitation as sea angling baits and the potential for aquaculture based production. Mem. Mus. Natl. Hist. Nat. 1994, 162, 603–610. [Google Scholar]
- Meunpol, O.; Meejing, P.; Piyatiratitivorakul, S. Maturation diet based on fatty acid content for male Penaeus monodon (Fabricius) broodstock. Aquac. Res. 2005, 36, 1216–1225. [Google Scholar] [CrossRef]
- Marques, B.; Lillebø, A.I.; Ricardo, F.; Nunes, C.; Coimbra, M.A.; Calado, R. Adding value to ragworms (Hediste diversicolor) through the bioremediation of a super-intensive marine fish farm. Aquacult. Env. Interac. 2018, 10, 79–88. [Google Scholar] [CrossRef]
- Jerónimo, D.; Lillebø, A.I.; Santos, A.; Cremades, J.; Calado, R. Performance of polychaete assisted sand filters under contrasting nutrient loads in an integrated multi-trophic aquaculture (IMTA) system. Sci. Rep. 2020, 10, 20871. [Google Scholar] [CrossRef] [PubMed]
- Pires, A.; Gentil, F.; Quintino, V.; Rodrigues, A.M. Reproductive biology of Diopatra neapolitana (Annelida, Onuphidae), an exploited natural resource in Ria de Aveiro (Northwestern Portugal). Mar. Ecol. 2012, 33, 56–65. [Google Scholar] [CrossRef]
- Pires, A.; Freitas, R.; Quintino, V.; Rodrigues, A.M. Can Diopatra neapolitana (Annelida: Onuphidae) regenerate body damage caused by bait digging or predation? Estuar. Coast. Shelf S. 2012, 110, 36–42. [Google Scholar] [CrossRef]
- Aleixo, A.; Queiroga, H.; Xenarios, S.; Lillebø, A. Catch estimates and bioecomomic analysis of bait digging: The case of the tube worm Diopatra neapolitana. Bioforsk Rapport 2014, 9, 136. [Google Scholar]
- Costa, F.E.; Sá, E.; Alves, A.S.; Cabral, S.; Castro, N.; Picard, D.; Castro, J.J.; Cancela Da Fonseca, L.; Chainho, P.; Canning-Clode, J. Anelídeos poliquetas como isco vivo: caracterização da atividade de apanha em ambientes salobros costeiros Portugueses. Entre Rios E Mares Um Património De Ambientes História E Sabers – Tomo V Da Rede BrasPor 2016, 33–43.
- Xenarios, S.; Queiroga, H.; Lillebø, A.I.; Aleixo, A. Introducing a regulatory policy framework of bait fishing in European coastal lagoons: The case of Ria de Aveiro in Portugal. Fishes 2018, 3, 2. [Google Scholar] [CrossRef]
- Olive, P.J. Polychaete aquaculture and polychaete science: a mutual synergism. In Reproductive Strategies and Developmental Patterns in Annelids; Springer: Dordrecht, The Netherlands, 1999; pp. 175–183. [Google Scholar]
- Freitas, F.; Cunha, T.; Hall, A.; Queiroga, H. Diopatra neapolitana, Importância Sócio-económica e Sustentabilidade das Capturas, no Canal de Mira, Ria de Aveiro. In Jornadas da Ria de Aveiro, Universidade de Aveiro, Portugal; 2011; pp. 60–66.
- De Carvalho, A.N.; Vaz, A.S.L.; Sérgio, T.I.B.; dos Santos, P.J.T. Sustainability of bait fishing harvesting in estuarine ecosystems–Case study in the Local Natural Reserve of Douro Estuary, Portugal. Rev. De Gestão Costeira Integr.-J. Integr. Coast. Zone Manag. 2013, 13, 157–168. [Google Scholar] [CrossRef]
- Conti, G.; Massa, F. Experienze de allavamento del polichete Diopatra neapolitana Delle Chiaje, 1841 nella Laguna di S. Gilla (Sardegna Meridionale). Biol. Mar. Mediterr. 1998, 5, 1473–1480. [Google Scholar]
- Safarik, M.; Redden, A.M.; Schreider, M.J. Density-dependent growth of the polychaete Diopatra aciculata. Sci. Mar. 2006, 70, 337–341. [Google Scholar] [CrossRef]
- Kraan, C.; Piersma, T.; Dekinga, A.; Koolhaas, A.; Van der Meer, J. Dredging for edible cockles (Cerastoderma edule) on intertidal flats: short-term consequences of fisher patch-choice decisions for target and non-target benthic fauna. ICES J. Mar. Sci. 2007, 64, 1735–1742. [Google Scholar] [CrossRef]
- Fonseca, L.; Costa, P.; Fidalgo, E. Poliquetas: sua obtenção, impactos e medidas de gestão. In Proceedings of the 14º Congresso da Associação Portuguesa para o Desenvolvimento Regional-Ambiente e Conservação da Natureza; 2008; p. 851. [Google Scholar]
- Leal, M.C.; Pimentel, T.; Ricardo, F.; Rosa, R.; Calado, R. Seafood traceability: current needs, available tools, and biotechnological challenges for origin certification. Trends Biotechnol. 2015, 33, 331–336. [Google Scholar] [CrossRef]
- Santos, A.; Ricardo, F.; Domingues, M.R.M.; Patinha, C.; Calado, R. Current trends in the traceability of geographic origin and detection of species-mislabeling in marine bivalves. Food Control 2023, 152, 109840. [Google Scholar] [CrossRef]
- Ricardo, F.; Maciel, E.; Domingues, M.R.; Calado, R. Spatio-temporal variability in the fatty acid profile of the adductor muscle of the common cockle Cerastoderma edule and its relevance for tracing geographic origin. Food Control 2017, 81, 173–180. [Google Scholar] [CrossRef]
- Ricardo, F.; Pimentel, T.; Moreira, A.S.P.; Rey, F.; Coimbra, M.A.; Rosário Domingues, M.; Domingues, P.; Costa Leal, M.; Calado, R. Potential use of fatty acid profiles of the adductor muscle of cockles (Cerastoderma edule) for traceability of collection site. Sci. Rep. 2015, 5, 11125. [Google Scholar] [CrossRef]
- Mamede, R.; Ricardo, F.; Santos, A.; Díaz, S.; Santos, S.A.O.; Bispo, R.; Domingues, M.R.M.; Calado, R. Revealing the illegal harvesting of Manila clams (Ruditapes philippinarum) using fatty acid profiles of the adductor muscle. Food Control 2020, 107368. [Google Scholar] [CrossRef]
- Nemova, N.N.; Fokina, N.N.; Nefedova, Z.A.; Ruokolainen, T.R.; Bakhmet, I.N. Modifications of gill lipid composition in littoral and cultured blue mussels Mytilus edulis L. under the influence of ambient salinity. Polar Rec. 2013, 49, 272–277. [Google Scholar] [CrossRef]
- Ricardo, F.; Mamede, R.; Bruzos, A.L.; Díaz, S.; Thébault, J.; da Silva, E.F.; Patinha, C.; Calado, R. Assessing the elemental fingerprints of cockle shells (Cerastoderma edule) to confirm their geographic origin from regional to international spatial scales. Sci. Total Environ. 2022, 814, 152304. [Google Scholar] [CrossRef]
- Ricardo, F.; Génio, L.; Costa Leal, M.; Albuquerque, R.; Queiroga, H.; Rosa, R.; Calado, R. Trace element fingerprinting of cockle (Cerastoderma edule) shells can reveal harvesting location in adjacent areas. Sci. Rep. 2015, 5, 11932. [Google Scholar] [CrossRef]
- Carson, H.S.; López-Duarte, P.C.; Cook, G.S.; Fodrie, F.J.; Becker, B.J.; DiBacco, C.; Levin, L.A. Temporal, spatial, and interspecific variation in geochemical signatures within fish otoliths, bivalve larval shells, and crustacean larvae. Mar. Ecol. Prog. Ser. 2013, 473, 133–148. [Google Scholar] [CrossRef]
- Cabral, A.E.; Ricardo, F.; Patinha, C.; Silva, E.F.d.; Correia, M.; Palma, J.; Planas, M.; Calado, R. Successful Use of Geochemical Tools to Trace the Geographic Origin of Long-Snouted Seahorse Hippocampus guttulatus Raised in Captivity. Animals 2021, 11, 1534. [Google Scholar] [CrossRef] [PubMed]
- Takesue, R.K.; Bacon, C.R.; Thompson, J.K. Influences of organic matter and calcification rate on trace elements in aragonitic estuarine bivalve shells. Geochim. Cosmochim. Ac. 2008, 72, 5431–5445. [Google Scholar] [CrossRef]
- Becker, B.J.; Fodrie, F.J.; McMillan, P.; Levin, L.A. Spatial and temporal variation in trace elemental fingerprints of mytilid mussel shells: A precursor to invertebrate larval tracking. Limnol. Oceanogr. 2004, 50, 48–61. [Google Scholar] [CrossRef]
- Sorte, C.J.; Etter, R.J.; Spackman, R.; Boyle, E.E.; Hannigan, R.E. Elemental Fingerprinting of Mussel Shells to Predict Population Sources and Redistribution Potential in the Gulf of Maine. PloS ONE 2013, 8, e80868. [Google Scholar] [CrossRef]
- Zacherl, D.C. Spatial and temporal variation in statolith and protoconch trace elements as natural tags to track larval dispersal. Mar. Ecol. Prog. Ser. 2005, 290, 145–163. [Google Scholar] [CrossRef]
- Zhang, X.; Han, D.; Chen, X.; Zhao, X.; Cheng, J.; Liu, Y. Combined use of fatty acid profile and fatty acid δ13C fingerprinting for origin traceability of scallops (Patinopecten yessoensis, Chlamys farreri, and Argopecten irradians). Food Chem. 2019, 298, 124966. [Google Scholar] [CrossRef]
- Perez, V.; Olivier, F.; Tremblay, R.; Neumeier, U.; Thébault, J.; Chauvaud, L.; Meziane, T. Trophic resources of the bivalve, Venus verrucosa, in the Chausey archipelago (Normandy, France) determined by stable isotopes and fatty acids. Aquat. Living Resour. 2013, 26, 229–239. [Google Scholar] [CrossRef]
- Chaguri, M.P.; Maulvault, A.L.; Nunes, M.L.; Santiago, D.A.; Denadai, J.C.; Fogaça, F.H.; Sant’Ana, L.S.; Ducatti, C.; Bandarra, N.; Carvalho, M.L. Different tools to trace geographic origin and seasonality of croaker (Micropogonias furnieri). LWT-Food Sci. Tech. 2015, 61, 194–200. [Google Scholar] [CrossRef]
- Matos, M.P.; Engel, M.E.; Mangrum, J.B.; Jackson, G.P. Origin determination of the Eastern oyster (Crassostrea virginica) using a combination of whole-body compound-specific isotope analysis and heavy metal analysis. Anal. Methods 2021, 13, 3493–3503. [Google Scholar] [CrossRef]
- Cabral, S.; Alves, A.S.; Castro, N.; Chainho, P.; Sá, E.; da Fonseca, L.C.; e Costa, P.F.; Castro, J.; Canning-Clode, J.; Pombo, A. Polychaete annelids as live bait in Portugal: Harvesting activity in brackish water systems. Ocean Coast. Manage. 2019, 181, 104890. [Google Scholar] [CrossRef]
- Aued-Pimentel, S.; Lago, J.H.G.; Chaves, M.H.; Kumagai, E.E. Evaluation of a methylation procedure to determine cyclopropenoids fatty acids from Sterculia striata St. Hil. Et Nauds seed oil. J. Chromatogra A 2004, 1054, 235–239. [Google Scholar] [CrossRef]
- Anderson, M.; Clarke, K.; Gorley, R. PERMANOVA+ for Primer. Guide to Software and Statistical Methods; University of Auckland and PRIMER-E Ltd: Plymouth, UK, 2008. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. R Found. Stat Comput Vienna Austria 2021. [Google Scholar]
- Ricardo, F.; Mamede, R.; Bispo, R.; Santos, A.; Ferreira da Silva, E.; Patinha, C.; Calado, R. Cost-efficiency improvement of bivalves shells preparation when tracing their geographic origin through ICP-MS analysis of elemental fingerprints. Food Control 2020, 118, 107383. [Google Scholar] [CrossRef]
- Kursa, M.B.; Jankowski, A.; Rudnicki, W.R. Boruta–a system for feature selection. Fundam. Informaticae 2010, 101, 271–285. [Google Scholar] [CrossRef]
- Ricardo, F.; Gonçalves, D.; Pimentel, T.; Mamede, R.; Rosário, M. Domingues, M.; Lillebø, A.I.; Calado, R. Prevalence of phylogenetic over environmental drivers on the fatty acid profiles of the adductor muscle of marine bivalves and its relevance for traceability. Ecol. Indic. 2021, 129, 108017. [Google Scholar] [CrossRef]
- da Costa, E.; Ricardo, F.; Melo, T.; Mamede, R.; Abreu, M.H.; Domingues, P.; Domingues, M.R.; Calado, R. Site-Specific Lipidomic Signatures of Sea Lettuce (Ulva spp., Chlorophyta) Hold the Potential to Trace Their Geographic Origin. Biomolecules 2020, 10, 489. [Google Scholar] [CrossRef] [PubMed]
- Mamede, R.; Ricardo, F.; Gonçalves, D.; Ferreira da Silva, E.; Patinha, C.; Calado, R. Assessing the use of surrogate species for a more cost-effective traceability of geographic origin using elemental fingerprints of bivalve shells. Ecol. Indic. 2021, 130, 108065. [Google Scholar] [CrossRef]
- Mamede, R.; Ricardo, F.; Abreu, M.H.; da Silva, E.F.; Patinha, C.; Calado, R. Spatial variability of elemental fingerprints of sea lettuce (Ulva spp.) and its potential use to trace geographic origin. Algal Res. 2021, 59, 102451. [Google Scholar] [CrossRef]
- Bennion, M.; Morrison, L.; Shelley, R.; Graham, C. Trace elemental fingerprinting of shells and soft tissues can identify the time of blue mussel (Mytilus edulis) harvesting. Food Control 2020, 121, 107515. [Google Scholar] [CrossRef]
- Bennion, M.; Morrison, L.; Brophy, D.; Carlsson, J.; Abrahantes, J.C.; Graham, C.T. Trace element fingerprinting of blue mussel (Mytilus edulis) shells and soft tissues successfully reveals harvesting locations. Sci. Total Environ. 2019, 121, 107515. [Google Scholar] [CrossRef]
- Fernandes, J.F.; Ricardo, F.; Jerónimo, D.; Santos, A.; Domingues, M.R.; Calado, R.; Madeira, D. Modulation of fatty acid profiles by global and local ocean change drivers in the ragworm Hediste diversicolor: implications for aquaculture production. Aquaculture 2021, 542, 736871. [Google Scholar] [CrossRef]
- Jerónimo, D.; Lillebø, A.I.; Maciel, E.; Domingues, M.R.M.; Cremades, J.; Calado, R. Unravelling the fatty acid profiles of different polychaete species cultured under integrated multi-trophic aquaculture (IMTA). Sci. Rep. 2021, 11, 1–14. [Google Scholar] [CrossRef]
- Phleger, C.F.; Nelson, M.M.; Groce, A.K.; Cary, S.C.; Coyne, K.; Gibson, J.A.; Nichols, P.D. Lipid biomarkers of deep-sea hydrothermal vent polychaetes—Alvinella pompejana, A. caudata, Paralvinella grasslei and Hesiolyra bergii. Deep Sea Res. Part I: Oceanogra. Res. 2005, 52, 2333–2352. [Google Scholar] [CrossRef]
- Olive, P.J.; Duangchinda, T.; Ashforth, E.; Craig, S.; Ward, A.C.; Davies, S.J. Net gain of long-chain polyunsaturated fatty acids (PUFA) in a lugworm Arenicola marina bioturbated mesocosm. Mar. Ecol. Prog. Ser. 2009, 387, 223–239. [Google Scholar] [CrossRef]
- Braeckman, U.; Provoost, P.; Sabbe, K.; Soetaert, K.; Middelburg, J.J.; Vincx, M.; Vanaverbeke, J. Temporal dynamics in the diet of two marine polychaetes as inferred from fatty acid biomarkers. J. Sea Res. 2012, 68, 6–19. [Google Scholar] [CrossRef]
- Dalsgaard, J.; St John, M.; Kattner, G.; Müller-Navarra, D.; Hagen, W. Fatty acid trophic markers in the pelagic marine environment. Adv. Mar. Biol. 2003, 46, 225–340. [Google Scholar]
- Madeira, D.; Fernandes, J.F.; Jerónimo, D.; Martins, P.; Ricardo, F.; Santos, A.; Domingues, M.R.; Diniz, M.S.; Calado, R. Salinity shapes the stress responses and energy reserves of marine polychaetes exposed to warming: From molecular to functional phenotypes. Sci. Total Environ. 2021, 795, 148634. [Google Scholar] [CrossRef]
- Török, Z.; Crul, T.; Maresca, B.; Schütz, G.J.; Viana, F.; Dindia, L.; Piotto, S.; Brameshuber, M.; Balogh, G.; Péter, M.; et al. Plasma membranes as heat stress sensors: From lipid-controlled molecular switches to therapeutic applications. Biochim. Biophys. Acta–Biomembr. 2014, 1838, 1594–1618. [Google Scholar] [CrossRef]
- Holmstrup, M.; Hovvang, M.H.; Slotsbo, S. Salinity of the growth medium is important for production potential and nutritional value of white worms (Enchytraeus albidus Henle). Aquac. Res. 2020, 51, 2885–2892. [Google Scholar] [CrossRef]
- Frolov, A.V.; Pankov, S.L.; Geradze, K.N.; Pankova, S.A. Influence of salinity on the biochemical composition of the rotifer Brachionus plicatilis (Muller). aspects of adaptation. Comp. Biochem. Physi. A 1991, 99, 541–550. [Google Scholar] [CrossRef]
- Lavaud, R.; Thébault, J.; Lorrain, A.; van der Geest, M.; Chauvaud, L. Senilia senilis (Linnaeus, 1758), a biogenic archive of environmental conditions on the Banc d'Arguin (Mauritania). J. Sea Res. 2013, 76, 61–72. [Google Scholar] [CrossRef]
- Albuquerque, R.; Queiroga, H.; Swearer, S.E.; Calado, R.; Leandro, S.M. Harvest locations of goose barnacles can be successfully discriminated using trace elemental signatures. Sci. Rep 2016, 6, 27787. [Google Scholar] [CrossRef] [PubMed]
- Campana, S.E.; Thorrold, S.R. Otoliths, increments, and elements: keys to a comprehensive understanding of fish populations? Can. J. Fish. Aquat. Sci. 2001, 58, 30–38. [Google Scholar] [CrossRef]
- Morán, P.; Cal, L.; Cobelo-García, A.; Almécija, C.; Caballero, P.; Garcia de Leaniz, C. Historical legacies of river pollution reconstructed from fish scales. Environ. Pollut. 2018, 234, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Ricardo, F.; Pimentel, T.; Génio, L.; Calado, R. Spatio-temporal variability of trace elements fingerprints in cockle (Cerastoderma edule) shells and its relevance for tracing geographic origin. Sci. Rep. 2017, 7, 3475. [Google Scholar] [CrossRef] [PubMed]
- Colbath, G.K. Jaw mineralogy in eunicean polychaetes (Annelida). Micropaleontol. 1986, 186–189. [Google Scholar] [CrossRef]
- Elegbede, I.; Lawal-Are, A.; Favour, O.; Jolaosho, T.; Goussanou, A. Chemical compositions of bivalves shells: Anadara senilis, Crassostrea gasar, and Mytilus edulis and their potential for a sustainable circular economy. SN Appl. Sci. 2022, 5. [Google Scholar] [CrossRef]
- Nambiar, R.; Hauzer, H.; Gray, W.R.; Henehan, M.J.; Cotton, L.; Erez, J.; Rosenthal, Y.; Renema, W.; Müller, W.; Evans, D. Controls on potassium incorporation in foraminifera and other marine calcifying organisms. Geochim. Cosmochim. Ac. 2023, 351, 125–138. [Google Scholar] [CrossRef]
- Pires, A.; Velez, C.; Figueira, E.; Soares, A.M.V.M.; Freitas, R. Effects of sediment contamination on physiological and biochemical responses of the polychaete Diopatra neapolitana, an exploited natural resource. Mar. Pollut. Bull. 2017, 119, 119–131. [Google Scholar] [CrossRef]
- Sprovieri, M.; Feo, M.L.; Prevedello, L.; Manta, D.S.; Sammartino, S.; Tamburrino, S.; Marsella, E. Heavy metals, polycyclic aromatic hydrocarbons and polychlorinated biphenyls in surface sediments of the Naples harbour (southern Italy). Chemosphere 2007, 67, 998–1009. [Google Scholar] [CrossRef] [PubMed]
- Iyaka, Y.A. Nickel in soils: A review of its distribution and impacts. Sci. Res. Essays 2011, 6, 6774–6777. [Google Scholar] [CrossRef]
- Lopes, J.; Dias, J.; Cardoso, A.; Silva, C. The water quality of the Ria de Aveiro lagoon, Portugal: From the observations to the implementation of a numerical model. Mar. Environ. Res. 2005, 60, 594–628. [Google Scholar] [CrossRef]
- Pereira, F.; Picado, A.; Pereira, H.; Pinheiro, J.P.; Lopes, C.L.; Dias, J.M. Impact of Extreme Wind and Freshwater Runoff on the Salinity Patterns of a Mesotidal Coastal Lagoon. J. Mar. Sci. Eng. 2023, 11, 1338. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, Y.; Li, Y.; Zhao, X. Identification of the geographical origins of sea cucumber (Apostichopus japonicus) in northern China by using stable isotope ratios and fatty acid profiles. Food Chem. 2017, 218, 269–276. [Google Scholar] [CrossRef]
- Ortea, I.; Gallardo, J.M. Investigation of production method, geographical origin and species authentication in commercially relevant shrimps using stable isotope ratio and/or multi-element analyses combined with chemometrics: An exploratory analysis. Food Chem. 2015, 170, 145–153. [Google Scholar] [CrossRef]
- Vasconi, M.; Lopez, A.; Galimberti, C.; Moreno Rojas, J.M.; Muñoz Redondo, J.M.; Bellagamba, F.; Moretti, V.M. Authentication of farmed and wild european eel (Anguilla anguilla) by fatty acid profile and carbon and nitrogen isotopic analyses. Food Control 2019, 102, 112–121. [Google Scholar] [CrossRef]
- Thomas, F.; Jamin, E.; Wietzerbin, K.; Guérin, R.; Lees, M.; Morvan, E.; Billault, I.; Derrien, S.; Moreno Rojas, J.M.; Serra, F.; et al. Determination of Origin of Atlantic Salmon (Salmo salar): The Use of Multiprobe and Multielement Isotopic Analyses in Combination with Fatty Acid Composition to Assess Wild or Farmed Origin. J. Agric. Food Chem. 2008, 56, 989–997. [Google Scholar] [CrossRef]
Figure 1.
Sampling locations of Diopatra neapolitana in the coastal lagoon Ria de Aveiro, mainland Portugal: Espinheiro Channel (E: 40°39'48.50"N, 8°41'45.03"W), Ílhavo Channel (I: 40°38'35.40"N, 8°41'35.40"W) and Mira Channel (M1: 40°38'26.30"N, 8°43'58.90"W and M2: 40°35'58.30"N, 8°44'47.80"W). Top left corner: picture of a Diopatra neapolitana.
Figure 1.
Sampling locations of Diopatra neapolitana in the coastal lagoon Ria de Aveiro, mainland Portugal: Espinheiro Channel (E: 40°39'48.50"N, 8°41'45.03"W), Ílhavo Channel (I: 40°38'35.40"N, 8°41'35.40"W) and Mira Channel (M1: 40°38'26.30"N, 8°43'58.90"W and M2: 40°35'58.30"N, 8°44'47.80"W). Top left corner: picture of a Diopatra neapolitana.
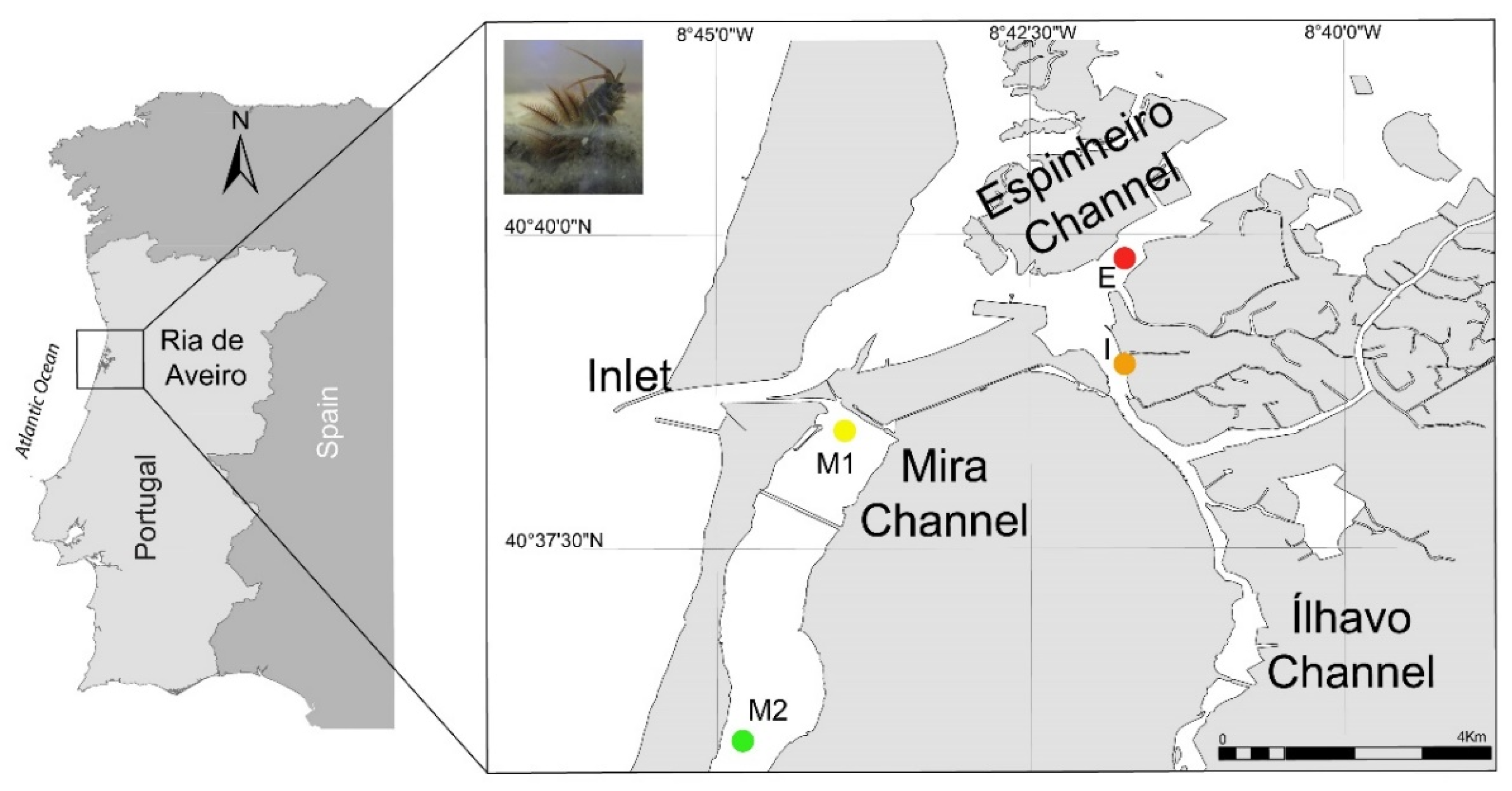
Figure 2.
Box plots of Z-scores registered by the Boruta algorithm used to determine the most relevant (a) fatty acids present on the of whole body of Diopatra neapolitana, (b) elemental ratios of the jaws of these polychaetes and (c) a combination of fatty acids from the whole body and the elemental ratios of jaws of these organisms sourced from different locations within the coastal lagoon Ria de Aveiro, mainland Portugal. Shadow variables are in blue, variables confirmed as unimportant in red, tentative in yellow, and most relevant in green.
Figure 2.
Box plots of Z-scores registered by the Boruta algorithm used to determine the most relevant (a) fatty acids present on the of whole body of Diopatra neapolitana, (b) elemental ratios of the jaws of these polychaetes and (c) a combination of fatty acids from the whole body and the elemental ratios of jaws of these organisms sourced from different locations within the coastal lagoon Ria de Aveiro, mainland Portugal. Shadow variables are in blue, variables confirmed as unimportant in red, tentative in yellow, and most relevant in green.

Figure 3.
Multidimensional scaling (MDS) ordinations of proximity scores from random forest classifications based on a) fatty acids present on the whole body of Diopatra neapolitan, b) elemental ratios of the jaws of these polychaetes and c) a combination of fatty acids from the whole body and the elemental ratios of jaws of these organisms sourced from different locations within the coastal lagoon Ria de Aveiro, mainland Portugal. Espinheiro Channel (E), Ílhavo Channel (I) and Mira Channel (M1 and M2).
Figure 3.
Multidimensional scaling (MDS) ordinations of proximity scores from random forest classifications based on a) fatty acids present on the whole body of Diopatra neapolitan, b) elemental ratios of the jaws of these polychaetes and c) a combination of fatty acids from the whole body and the elemental ratios of jaws of these organisms sourced from different locations within the coastal lagoon Ria de Aveiro, mainland Portugal. Espinheiro Channel (E), Ílhavo Channel (I) and Mira Channel (M1 and M2).
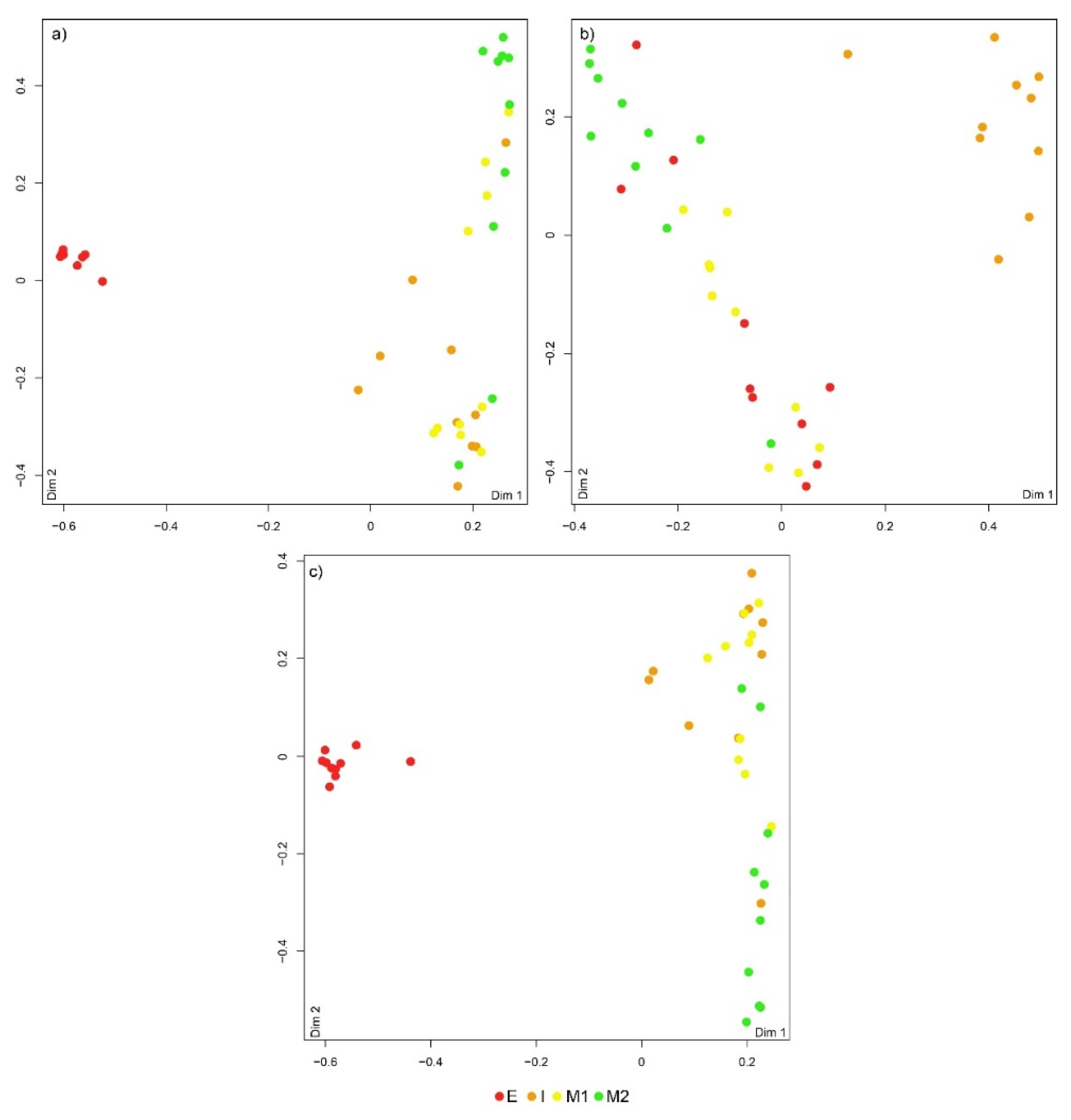

Table 1.
This Classification success (by location and structure(s)) of Random Forest model when using fatty acid profiles of the whole body, b) elemental ratios of jaws and c) a combination of fatty acid profiles of the whole body and elemental ratios of jaws of the polychaete Diopatra neapolitana. Espinheiro Channel (E:), Ílhavo Channel (I) and Mira Channel (M1 and M2). is a table. Tables should be placed in the main text near to the first time they are cited.
Table 1.
This Classification success (by location and structure(s)) of Random Forest model when using fatty acid profiles of the whole body, b) elemental ratios of jaws and c) a combination of fatty acid profiles of the whole body and elemental ratios of jaws of the polychaete Diopatra neapolitana. Espinheiro Channel (E:), Ílhavo Channel (I) and Mira Channel (M1 and M2). is a table. Tables should be placed in the main text near to the first time they are cited.
| Structure (s) Fingerprint (s) | Original Location | Predicted Location | Total per location | % correct | % correct (location) | |||
| E | I | M1 | M2 | |||||
| Whole body - FA profile | E | 10 | 0 | 0 | 0 | 10 | 100 | 75 |
| I | 0 | 6 | 3 | 1 | 10 | 60 | ||
| M1 | 0 | 2 | 7 | 1 | 10 | 70 | ||
| M2 | 0 | 1 | 2 | 7 | 10 | 70 | ||
| Jaws - EF | E | 6 | 1 | 0 | 3 | 10 | 60 | 72.5 |
| I | 0 | 9 | 0 | 1 | 10 | 90 | ||
| M1 | 2 | 0 | 8 | 0 | 10 | 80 | ||
| M2 | 3 | 0 | 1 | 6 | 10 | 60 | ||
| Whole body & Jaws - FA profile & EF | E | 10 | 0 | 0 | 0 | 10 | 100 | 85 |
| I | 0 | 8 | 1 | 1 | 10 | 80 | ||
| M1 | 0 | 1 | 8 | 1 | 10 | 80 | ||
| M2 | 0 | 1 | 1 | 8 | 10 | 80 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



