Submitted:
07 February 2024
Posted:
08 February 2024
You are already at the latest version
Abstract
The bioindicator potential of seagrass ecosystems in coastal trace metal monitoring is low for tropical island ecosystems. This study evaluated the bioindicator potential of six seagrass species exposed to anthropogenic trace metal pollution in the Andaman and Nicobar Islands (ANI) of India. Sediment and seagrass biomass, samples were analyzed for trace metals (Co, Cr, Cu, Fe, Mg, Mn, Ni, Pb, Zn) from four locations of ANI exposed to anthropogenic contamination. Geo accumulation Index values indicated moderate trace metal contamination in seagrass sediment. Seagrass trace metal accumulation from sediment was both species-specific and location- specific for ANI. Small seagrass species such as H. ovalis, H. beccarii and H. uninervis accumulated the maximum concentration of six trace metals (Cr, Cu, Fe, Ni, Pb, Zn) in their tissues compared to big seagrass species like T. hemprichii and E. acoroides of ANI. This study indicates leaves of small seagrass and roots of big seagrass species can serve as short- and long-term bioindicators respectively, of coastal trace metal contamination in tropical islands of ANI.
Keywords:
Coastal Pollution
; Trace metals
; Accumulation
; Andaman and Nicobar Islands
; Seagrass
1. Introduction
Seagrasses are marine macrophytes that inhabit intertidal areas to deep waters (Esteban et al., 2018; Short et al., 2007). These ecosystems are highly productive and play an important role in providing various ecosystem services, such as habitat and nursery for various marine fish, invertebrates and mammals, blue carbon sequestration, coastal nutrient, metals and organic matter cycling, shoreline protection from erosion (Barbier et al., 2011; M. Stankovic et al., 2021; Nordlund et al., 2016; Unsworth et al., 2022; zu Ermgassen et al., 2021). These ecosystem services support millions of coastal communities by providing food and nutritional security, livelihood options and contribute towards well-being of the communities (Ho et al., 2018; Joseph et al., 2018; McKenzie et al., 2021). However, globally seagrass ecosystems are under decline due to various anthropogenic activities related to coastal pollution and land-use changes derived from various human activities (Hu et al., 2021; Lu et al., 2018; Nazneen et al., 2022; Stockbridge et al., 2020). These anthropogenic pollution activities include nutrient run-offs from agricultural activities, industrial and domestic wastewater discharges, oil spills, etc., and act as source of various anthropogenic contaminants. These contaminants concentrate in seagrass ecosystems, as they are filtered from the water column by seagrass leaves and shoots and deposited in the sediment where these contaminates gets adsorbed into fine grain sediments and increase their concentration(Bonanno & Borg, 2018a; Gao et al., 2016; A. K. Mishra et al., 2022; A. K. Mishra & Farooq, 2022a; Sungur & Özcan, 2015). Once concentrated these contaminants (such as trace metals) gets accumulated in seagrass tissues (e.g., roots and rhizomes) and cause severe adverse effects on the seagrass plant physiology, associated biota and the coastal communities that utilize seagrass associated biodiversity as source of food and nutrition (Lewis & Richard, 2009; Y. Li et al., 2023; Oreska et al., 2018).
Trace metals are one of the globally concerned ubiquitous anthropogenic contaminants that impart adverse impacts on the seagrass plants and their associated biodiversity (Govers et al., 2014; Y. Li et al., 2023; Nishitha et al., 2022; Thorne-Bazarra et al., 2023). Despite trace metals occurring in very low concentration in the marine environment, they play a critical role in the marine ecosystem functioning (Avelar et al., 2013; Coclet et al., 2021; A. K. Mishra et al., 2022; A. K. Mishra & Farooq, 2022a). Some of the trace metals are categorized as essential (e.g., Co, Cu, Mn, Zn), while others are considered as toxic (e.g., As, Cd, Cr, Hg, Ni and Pb) for marine plants and associated biodiversity(Schneider et al., 2018a). However, the essential trace metals can also impart adverse impacts or become toxic once their threshold levels are breached (Dong et al., 2016; Stockdale et al., 2016; Ward, 1984). Concentrations above threshold level posses a serious threat to seagrass eco-physiology because of their non-biodegradable and persistent nature in the marine sediment and consequent accumulation of these metals into seagrass roots, rhizomes and leaves (Aljahdali & Alhassan, 2020; A. K. Mishra et al., 2022; A. K. Mishra & Farooq, 2022a; Nikalje & Suprasanna, 2018; Rainbow, 2007). Once trace metals are accumulated in seagrass tissues, they are transferred to seagrass associated herbivores (e.g., fish and turtles), detritus grazers, leaf epiphytes and other organisms and gets biomagnified in trophic food chains and webs (Jiang et al., 2023; Schneider et al., 2018a; Suheryanto & Ismarti, 2018; Wilkinson et al., 2022).
Seagrasses of 16 different species are observed along the entire coast of India including the islands of Andaman and Nicobar (ANI) and the Lakshadweep to a depth of 21 m (Bayyana et al., 2020; Geevarghese et al., 2018). These ecosystems being present at the land and sea interface are subjected to various levels of trace metal contamination due to anthropogenic activities along the coast. A recent review on trace metal bioindicator potential of India’s marine macrophytes (e.g., seagrasses and saltmarshes), have suggested that seagrass are better bioindicators of trace metal accumulations than saltmarsh plants that inhabit similar intertidal zones (Mishra and Farooq, 2022; Nazneen et al., 2022). Small seagrass species like Halophila ovalis, Halophila beccarii and Halodule uninervis were found to be better bioindicators of various trace metals due to their higher growth rates and gorilla way of meadow expansion (Arisekar et al., 2021; Govindasamy & Azariah, 1999; Jagtap, 1983; Nazneen et al., 2022; Ragupathi Raja Kannan et al., 2011; Sachithanandam et al., 2020a) . However, bioindicator potential of large seagrass species like Enhalus acoroides and Syringodium isoetifolium may be higher than the small seagrass species, but data on trace metal accumulations in these big seagrasses that are present below 5m depth are very low in India (Arisekar et al., 2021; Nobi et al., 2010; Thangaradjou et al., 2010) .
This study quantified the concentration of trace metals (Co, Cr, Cu, Fe, Mg, Mn, Ni, Pb and Zn) in water, sediment and tissues (above-and below-ground) of six seagrasses that have a combination of both small and large species (i.e., H. ovalis, H. beccarii, H. uninervis, Thalassia hemprichii, Cymodocea rotundata and E. acoroides) across four locations of ANI of India. This study also assessed the species-specific accumulation and bioindicator potential of these six seagrass species to various trace metals. This study hypothesizes that different seagrass tissues accumulate varying concentration of metals and can be utilized as metal-specific bioindicators and this bioindicator potential is influenced by local anthropogenic influx concentration.
Four locations were selected that have seagrass ecosystems and received anthropogenic influxes mainly due to short-term tourism, local waste water release of untreated domestic sewage and industrial waste from ship yards. Trace metal studies on seagrass ecosystems of ANI is very limited (n=3). But these island ecosystems inhabit,10 out of 16 seagrass species found on the Indian coast (Gole et al., 2023; A. K. Mishra et al., 2023; A. K. Mishra & Farooq, 2022a) and trace metal concentration in the vulnerable seagrass H. beccarii has never been quantified from these islands, which has been recently recorded in these islands (Nobi et al., 2010; Sachithanandam et al., 2020a; Thangaradjou et al., 2010). Being vulnerable and present at the land and sea interface H. beccarii is subjected to maximum exposure towards anthropogenic contamination and habitat disturbances that can lead towards local extirpation of this vulnerable species. (Jagtap, 1983; A. Kumar. Mishra & Apte, 2021).
2. Materials and Methods
2.1. Study Site
The four study locations are part of the ANI of India (Figure 1). The ANI is located in the southeast coast of India in the Andaman Sea of the Indian Ocean Region (A. K. Mishra et al., 2023; A. K. Mishra & Farooq, 2023). The human population of these islands is 0.4 million as of 2023 (India Census, 2023), which have increased in the last decade resulting in increased anthropogenic impacts on the coastal ecosystems (VishnuRadhan et al., 2015). These islands have a tropical setting, and coral reef barriers resulting in a high diversity of seagrass and mangrove species which are part of the Tropical Indo-Pacific bioregion (Gole et al., 2023; A. Mishra & Apte, 2020; A. K. Mishra et al., 2023; A. K. Mishra, Narayana, et al., 2021)). Being an island ecosystem, the human populations have settled within few kilometers of the coastal zones which leads to the direct release of domestic and anthropogenic untreated waste into the coastal ecosystems (A. K. Mishra & Kumar, 2020; Nobi et al., 2010; Sachithanandam et al., 2020a). This study surveyed four locations of the ANI; Saheed Dweep (hereafter called as Neil Island Island), Swaraj Dweep (hereafter called as Havelock Island Island), Burmanallah and Haddo Bay situated on the southeast coast of ANI (Figure 1). These locations (except Haddo Bay) are tourism locations where in the dry season (October-April) a lot of touristic activities and the associated waste water discharge directly in to costal systems (Sachithanandam et al., 2022; VishnuRadhan et al., 2015). The Burmanallah location receives local untreated domestic waste water from the surrounding village (Mishra & Apte, 2020; Mishra & Kumar, 2020; Mishra & Mohanraju, 2018) The Haddo Bay receives anthropogenic input from the Portblair city along with waste discharges from the small industries (boat making, coloring, and dismantling industries) present along the bay (Sahu et al., 2013). The sampling was carried out in April 2023, usually considered as the dry season period (October-April) of the year.
Neil Island & Havelock Island
The Neil Island and Havelock Island are part of Ritche’s Archipelago and are situated in the southeast of ANI (Figure 1). Both islands are famous tourist destinations of the ANI. The study locations of both Neil Island and Havelock Island are situated next to the beach where touristic hotels and resorts are situated and the islands untreated domestic waste water is released (VishnuRadhan et al., 2015). The Havelock Island has a large fleet of small boats used for water sports, and the assembly, painting and recycling activities of these boats also takes places in the island. In Neil Island we sampled the Ram Nagar beach, which receives domestic waste water and the seagrass T. hemprichii is present as mono-specific meadows in shallow depths of 0.5 m. In Havelock Island we sampled the Vijay Nagar beach, where in shallow depths of 0.5 m mixed meadows of five seagrass species, i.e., H. ovalis, H. uninervis, T. hemprichii, Cymodocea rotundata and E. acoroides are present.
Burmanallah
Burmanallah is situated in the southeast of the ANI (Figure 1). This island has rocky intertidal beaches and have manmade concrete structures. It is next to a fishing village and few hotels and resorts and aquaculture facilities are located close to the beach (Nobi et al., 2010; Sachithanandam et al., 2020; Vishnu Radhan et al., 2015). The study location is also connected by a mangrove creek which is daily flooded by tidal inundations. We sampled recorded two species of seagrasses (T. hemprichii and C. rotundata) in shallow depths of 0.3 m at the sampling location
Haddo Bay
The Haddo Bay is located in the center of Portblair city of ANI (Figure 1). All the major shipping ports and associated boat industries of ANI are situated here. This area has been significantly modified by anthropogenic activities and the majority of the waste water draining channels of the city are drained in the surroundings of this study location (Sahu et al., 2023). The study location inhabits four species of seagrass, i.e., H. ovalis, H. uninervis, T. hemprichii, H. beccarii that are present in shallow water depths of 0.5m.
2.2. Water and Sediment Sampling and Analysis
Surface water (n=5/location) and sediment (n=5/species) samples were collected from the same areas across the four locations of ANI, where seagrass samples were collected. Physical parameters (pH, salinity and temperature) were measured in-situ for the surface water above the seagrass meadows during sampling. pH and temperature (HI991301P, Hannah Instruments) and salinity (HI98203, Hannah Instruments) were measured using hand held probes. Collected water samples were filtered using 0.45-micron polycarbonate filter and acidified in 14 M ultrapure HNO3 in 250 ml high density polyethylene bottles and brought to the laboratory for trace metal analysis. The sediment cores (diameter of 5 cm) of the top 10 cm of the sediment were recovered for each seagrass species from each study location with the help of a plastic corer. After collection, the sediment samples were stored in zip locked plastic bags and transported to the laboratory. In the laboratory these sediment samples were oven dried at 60 °C for 48 h in a hot air oven. After drying the sediment samples were analyzed for grain size fractions. A subset of the sediment samples was homogenized using a disc mill (Retsch, RS200, USA) and stored for further analysis. Loss on Ignition (LOI) method was used to quantify sediment organic matter (OM%) content (Howard et al., 2014), where 5 gm of homogenized sediment was combusted at 500° C for 4 h in a muffle furnace. The LOI was calculated following Eq.1
Geoaccumulation Index (Igeo)
The surface sediment trace metal contamination level of the four location of ANI was assessed using the Geoaccumulation Index (Igeo) values, which was computed using Eq. (2)
Where Cn is the trace metal concentration measured in the sediment, Bn is the geochemical background value derived from the average shale value of each element in earth’s upper crust (Turekian & Wedepohl, 1961). The natural fluctuation for each element concentration is represented by the constant 1.5. Our results were compared to the below mentioned six Igeo classes, that defines the degree of contamination.
| Class | Value | Sediment Quality |
| 0 | Igeo <0 | Uncontaminated |
| 1 | 0 < Igeo <1 | Uncontaminated to moderate contaminated |
| 2 | 1<Igeo < 2 | Moderately contaminated |
| 3 | 2 <Igeo <3 | Moderately to heavily contaminated |
| 4 | 3 <Igeo <4 | Heavily contaminated |
| 5 | 4 <Igeo <5 | Heavily to extremely contaminated |
| 6 | 5 <Igeo <6 | Extremely contaminated |
The Bio Sediment Accumulation Factor (BSAF) was estimated based on Eq.3
Where Mp is the concentration of the metal in the seagrass roots and Ms is the concentration of metals in the sediment. The BSAF showcases the efficiency of seagrass species to absorb elements from sediment and accumulate in its tissues. Higher BCF (>1) values implies an increase ability of the plant species to accumulate metals from the sediment (Bonanno & Raccuia, 2018; (EPA), 2009).
2.3. Seagrass Sampling and Analysis
From each location for each species (n=5) quadrats of 20 x 20 cm were collected at least 1 m apart. The seagrass samples were collected using a hand shovel till 10 cm, where most of the below-ground root biomass were present. The seagrass plants were washed in the field with seawater to remove most of the algal growth (see Figure 2) and stored in zip locked plastic bags and brought to the laboratory. In the laboratory they were washed again with deionized water and any presence of epiphytic algae was removed using glass slides. Then the plants were separated into above-ground (AG: Leaves) and below-ground (BG: roots/rhizomes) tissues and dried at 60°C for 48 h in a hot air oven. After drying the plant tissues were homogenized and powdered using a ball mill (Retsch, RS200, USA) and stored for further analysis.
2.4. Analytical Methods for Water, Sediment and Seagrass
The concentration of trace metals (Co, Cr, Cu, Fe, Mg, Mn, Ni, Pb and Zn) in the surface water samples were determined by a dual view Inductively Coupled Plasma Optical Emission Spectroscopy (ICP-OES; 5110, Agilent).
The sediment and plant (AG and BG-tissues) samples (n=5/species) were digested in a microwave digestion system (Multiwave Pro, Antan Paar). For this purpose, 0.30 mg of homogenized sediment samples along the certified reference material (CRM) for sediment (HISS-1) and blanks were weighed into the digestion vessels for each batch (n=8). The sediment and standard samples were then treated with a mixture of acids, i.e., 4.5 ml of hydrochloric acid (HCl) and 4.5 ml of nitric acid (HNO3) of ultra-pure grade and kept in microwave system. The microwave sample digestion was carried out by setting the power of the system to 1300 watt for 20 minutes ramp, followed by 60 minutes hold time. For plant, 0.30 mg of samples were weighed and treated with 9 ml of ultrapure HNO3 acid along with reference material (ERM-CD281) and blanks for each batch. The digestion procedure was same as that of the sediment samples. After the digestion both sediment and plant samples were transferred to 50 ml volumetric flask and the volume was made up to 50 ml using 10% HNO3. Then the samples were centrifuged at 1000 rpm for 1 minute, filtered (using 0.45-micron polycarbonate filter) and analyzed using the ICP-OES. The percentage recovery of the trace metals was checked by comparing the derived trace metal concentrations from this study with the concentration of CRMs and the recovery was between 92-100% (Table 1). Standard precautions for trace metal analysis were followed through the analytical procedures.
2.4. Statistics
A two-way ANOVA was used to test the significant differences between trace element concentration among locations and compartment (biomass and sediment) for each seagrass species except E. acoroides and H. beccarii. Holm-Sidak multiple comparison tests was performed to test differences when interaction was present between the factors. For the remaining two seagrass species (E. acoroides and H. beccarii) a one-way ANOVA was used using only compartment as fixed factor. Statistical significance was also tested surface water abiotic parameters (pH, temperature and salinity) and sediment OM and sediment grain size. All data were pre-checked for homogeneity of variance and normality using Levene’s and Shapiro-Wilk test respectively. Pearson correlation was used to test the corelationship between water (pH, salinity, temperature) and sediment abiotic parameters (i.e., sediment grain size and OM) with sediment trace metal concentrations for each seagrass species across the study locations. All statistical tests were conducted at a significance level of p<0.05 and data is presented as mean ± standard deviation (SD). GraphPad Prism (ver. 10.1.2) software was used for all statistical analyses and graph making.
3. Results
3.1. Variation in Abiotic Parameters in Water and Sediment
Significant differences were observed between the surface water pH (one way ANOVA, F 3,20 = 10.50, p=0.0002) and temperature (F 3,16= 16.96, p<0.0001) above seagrass meadows among the four study locations, except for salinity (Table 2). Among the four locations, the surface water pH was lower at Haddo Bay (8.00 ± 0.08), and similar between Neil Island, Havelock Island and Burmanallah locations. The highest temperature of surface water was recorded at Havelock Island (34 ± 1.22 °C) and the lowest at Haddo Bay (30 ± 0.46 °C) (Table 2). Between the sediment abiotic parameters, significant difference was observed for the sediment OM (F 3,36=18.21) between locations and no significance was observed for sediment grain size fractions (Table 2). The highest sediment OM was observed at Burmanallah (38.70 ±5.07%), which was 1.5-fold higher than the sediment OM content of Haddo Bay (25.44 ± 3.01%) (Table 2).
3.2. Trace Metal Concentrations in Water, Sediment and Seagrass Biomass
The trace metal concentration (Co, Cr, Cu, Fe, Mg, Mn, Ni, Pb and Zn) in surface water samples above the seagrass ecosystems were below detection limit for all trace metals except Mg. The range of Mg surface water ranged from 25.49–26.34 µg L-1 across the four study locations.
The seagrass sediment Igeo values across the four locations of ANI showed moderate contamination of few trace metals (Co, Cr, and Zn) in sediment, i.e., Co at Burmanallah, Co and Cr at Havelock Island and Co and Zn at Haddo Bay (Figure 3).
The BSAF values indicated different accumulation patterns of trace metals in seagrass species from sediment across the four locations of ANI (Figure 4). In T. hemprichii, BSAF>1 was observed for all trace metals except for Cr at Havelock Island, Burmanallah and Haddo Bay. Similarly, BSAF>1 was observed for Cu and Mn at Haddo Bay and Neil Island respectively. BSAF>1 for Pb was observed at all four locations, whereas BSAF>1 for Zn in T. hemprichii meadows was only observed at Burmanallah and Haddo Bay (Figure 4a). In H. uninervis BSAF>1 was observed for all trace metals at both Havelock Island and Haddo Bay, except for Cu, Pb and Zn in Haddo Bay (Figure 4b). In C. rotundata, BSAF>1 was observed for all trace metals at Havelock Island, except Mn, whereas at Burmanallah none of the trace metals showed BSAF>1, except Co (Figure 4c). In H. ovalis, the BSAF>1 was only observed Co for in Havelock Island and Haddo Bay, where BSAF >1 for Cr, Mg and Ni was observed only at Haddo Bay (Fig.4d). In E. acoroides, the BSAF> observed for five trace metals, except Cr, Cu, Mn and Pb at Havelock Island (Figure 4e). In H. beccarii, the BSAF>1 was observed for all trace metals, except Cr and Zn at Haddo Bay (Figure 4f).
The concentration of trace metals in seagrasses was significantly different across the four locations and between compartments (Figure 5, Figure 6 and Figure 7). Among the four locations, at Haddo Bay the highest concentration of six trace metals (Co, Cr, Cu, Ni, Pb and Zn) was observed, whereas at Burmanallah the highest concentration of Fe, Mg and Mn was observed (Figure 5, Figure 6 and Figure 7). Between the locations and among the various seagrass species, the highest concentration of Co was observed in the AG-biomass of T. hemprichii that accumulated 26.3-fold higher concentration of Co (97.85 ± 0.5 mg Kg-1) at Haddo Bay than the lowest concentration of Co (3.72 ± 2.2 mg Kg-1) observed in the sediment of C. rotundata at Havelock Island (Figure 5a). The BG-biomass of H. ovalis at Haddo Bay accumulated 41-fold higher concentration of Cr (135.56 ± 0.5 mg Kg-1) than the lowest concentration of Cr observed in AG-biomass (3.28 ± 0.9 mg Kg-1) of H. uninervis at Havelock Island (Figure 5c). The concentration of Cu in the sediment (71.44 ± 1.4 mg Kg-1) of H. uninervis at Haddo Bay was 35-fold higher compared to the Cu concentration in AG-biomass (2.03 mg Kg-1) of E. acoroides at Havelock Island (Figure 5c).
The highest concentration of Fe in the sediment (18400.99 ± 14.1 mg Kg-1) of C. rotundata meadows at Burmanallah was 34-fold higher than the lowest concentration of Fe in the BG-biomass (543 ± 116 mg Kg-1) of E. acoroides at Havelock Island (Figure 6a). Similarly, the highest concentration of Mg observed in the AG-biomass (9190.79 ± 5.2 mg Kg-1) of T. hemprichii at Burmanallah was 6-fold higher than the lowest concentration of Mg in the AG-biomass (1428.36 ± 2.7 mg Kg-1) of H. ovalis at Havelock Island (Figure 6b). The highest concentration of Mn in the sediment (251.24 ± 38.2 mg Kg-1) of T. hemprichii meadows at Burmanallah was 16-fold higher than the lowest concentration of Mn in the AG-biomass (15.41 ± 5.4 mg Kg-1) of T. hemprichii at Neil Island (Figure 6c).
The highest concentration of Ni recorded in the AG-biomass (39.06 ± 0.7 mg Kg-1) of H. uninervis at Haddo Bay was 72-fold higher than the lowest concentration of Ni observed in BG-biomass (0.54 ±0.02 mg Kg-1) of C. rotundata at Burmanallah (Figure 7a). The highest concentration of Pb observed in the sediment (15.48 ±1.2 mg Kg-1) of C. rotundata at Haddo Bay was 33-fold higher than the lowest concentration of Pb observed in the BG-biomass (0.47± 0.15 mg Kg-1) of E. acoroides at Havelock Island (Figure 7b). The highest concentration of Zn observed in the BG-biomass (70.4 ± 1.0 mg Kg-1) of H. beccarii at Haddo Bay was 116-fold higher than the lowest concentration of Zn observed in the AG-biomass (0.60 mg Kg-1) of T. hemprichii at Burmanallah (Figure 7c).
3.3. Correlation between Surface Water and Sediment Abiotic Parameters on Trace Metals in Seagrass Sediment
Among the locations and seagrass species, the correlation between sediment and surface water abiotic parameters and sediment trace metal concentration were location-species-specific (Figure 8 and Figure 9 and Supplementary S2).
3.3.1. Correlations between Seagrass Species at Havelock Island
In T. hemprichii meadows, the sediment Cu, Fe, Ni and Zn concentration showed positive correlation with surface water pH, whereas Mg concentration showed negative correlation with pH at Havelock Island (8b). Similarly in H. uninervis meadows, the sediment Fe and Mn concentration showed positive correlation with surface water pH of Havelock Island (Figure 9a). In contrast, the sediment Fe and Mg concentration in H. ovalis meadows showed negative correlation with surface water pH of Havelock Island (Figure 9c). The sediment Mn concentration showed negative correlation with sediment clay content, whereas Ni and Pb showed positive correlation with silt and OM respectively within T. hemprichii meadows at Havelock Island (Figure 8a). In H. uninervis meadows, both Cu and Mn concentration and in H. ovalis meadows only Mn concentration showed positive correlation with sediment OM content (Figure 9a & c). In H. ovalis meadows, the sediment Ni concentration showed negative correlation with sediment OM and sand content, whereas positive correlation was observed with sediment silt content (Figure 9c). The Pb concentration in H. ovalis sediment showed positive and negative correlation with sand and silt respectively (Figure 9c). Similarly, Pb and Zn concentration showed positive correlation with sediment OM, and silt content respectively in C. rotundata meadows of Havelock Island Island (Supplementary S2).
3.3.2. Correlations between Seagrass Species at Burmanallah
In the sediment of T. hemprichii meadows, sediment Co concentration showed positive correlation with surface water pH at Burmanallah (Figure 8c).
3.3.3. Correlations between Seagrass Species at Haddo Bay
The Pb concentration in H. beccarii meadows showed negative correlation with surface water pH (Supplementary S2). In H. uninervis sediment, Co concentration showed positive correlation with both sediment OM and clay content and in H. ovalis meadows Co showed positive correlation with sediment silt content (Figure 9b & d). In T. hemprichii meadows, Cr concentration in sediment showed negative correlation with sediment OM and positive correlation with sediment sand fraction (Figure 8d). In contrast, sediment Cr concentration within H. uninervis meadows, showed negative correlation with sand content and positive correlation with sediment OM, silt and clay content (Figure 9b). In T. hemprichii meadows, the sediment Cu concentration showed negative correlation with sediment OM and clay fraction and positive correlation with sediment sand fraction (Figure 8d). In contrast, the sediment Cu concentration in H. uninervis meadows showed positive correlation with sediment OM and silt content (Figure 9b). In H. ovalis meadows, the Cu showed positive correlation with sediment sand content, but negative correlation with silt and clay fractions (Figure 9d). In T. hemprichii, Fe showed negative correlation with sand and positive correlation with sediment silt fraction at Haddo Bay (Figure 8d). In H. uninervis meadows, the Fe concentration showed positive correlation with sediment OM at Haddo Bay (Figure 9b). In T. hemprichii and H. uninervis meadows, the sediment Mn concentration showed positive correlation with both sediment OM and clay content at Haddo Bay (Figure 8d, Figure 9b). In T. hemprichii meadows, the sediment Pb concentration showed negative correlation with sediment OM, silt and clay fraction, and positive correlation with sediment sand content at Haddo Bay (Figure 8d). In contrast, the concentration of Pb in H. uninervis meadows showed positive and negative correlation with sand and silt respectively (Figure 9b). The sediment Zn concentration of T. hemprichii meadows showed positive correlation with sediment OM, silt and clay fraction and negative correlation with sand content of Haddo Bay (Figure 8d). In contrast, the Zn concentration in the sediment of H. beccarii meadows showed positive correlation with sand and negative correlation with clay content (Supplementary S2).
In H. uninervis meadows, sediment Mg concentration showed positive correlation with sediment OM, silt and clay fractions and negative correlation with sand (Figure 9b). In H. uninervis meadows, the sediment Ni concentration showed positive correlation with sediment OM, silt and clay content and negative correlation with sediment sand content (Figure 9b). In H. ovalis and H. beccarii meadows, Pb concentration showed positive correlation with silt and clay, but negative correlation with sand (Figure 9d).
4. Discussion
Anthropogenic activities are a major source of coastal trace metal contamination and the coastal ecosystems at the land -sea interface and are the storehouse of these contaminants (Jiang et al., 2023; Lu et al., 2018; Mishra et al., 2022; Mishra & Farooq, 2022; Schneider et al., 2018b; Thorne-Bazarra et al., 2023). Seagrass ecosystems that inhabit the intertidal areas interacts with these anthropogenic contaminants and can be utilized as suitable bioindicators of these trace metals accumulation. However, till date their utilization for monitoring of trace metal contamination in coastal ecosystems has been limited (Govers et al., 2014; H. Lee et al., 2023). In this study six seagrass species that inhabited four intertidal locations of ANI, India subjected to anthropogenic contamination has been utilized, to assess their bioindicator potential to nine trace metals (Co, Cr, Cu, Fe, Mg, Mn, Ni, Pb and Zn) and observed that seagrass bioindicator potential to trace metals is both location and species-specific (Figure 5, Figure 6 and Figure 7 and Supplementary S1). This study also observed that within the same location of ANI, different seagrass species showed variation in trace metal accumulation. To our knowledge this study presents the trace metal accumulation of the vulnerable seagrass H. beccarii from ANI, India for the first time (Mishra & Farooq, 2022; Mishra & Apte, 2021; Nazneen et al., 2022). This study also highlights that abiotic parameter of sediment (such as OM, and sediment grain size fractions) and pH of surface water above seagrass meadows positively/negatively interact with the available trace metals in the sediment and play an important role in their availability for uptake in seagrass ecosystems of ANI (Figure 8 and Figure 9 and Supplementary S2).
4.1. Effect of Sediment Traits (Organic Matter Content) on Metal Accumulation in Seagrass Sediment
The anthropogenic source of trace metals to coastal marine ecosystems is mostly through riverine input of land derived contaminants and direct disposal of industrial or domestic waste water into the coastal ecosystems that accumulates in the sediment of seagrass ecosystems (H. Lee et al., 2023; Naik et al., 2023; Nazneen et al., 2022). This riverine input also contributes significant amount of land derived OM, silt and clay fractions into the seagrass sediment that plays an important role in trace metal adsorption onto the fine fraction of the sediment (Sahu et al., 2023; Mishra et al., 2020; Nishitha et al., 2022; Sadanandan et al., 2023). However, in our study locations of ANI, there are no direct riverine input as ANI is an island ecosystem and lacks any well-developed riverine system, but our study locations like Burmanallah has a small mangrove creek, whereas Havelock Island and Haddo Bay has permanent waste water drains that provide a continuous input of land derived OM and industrial and domestic waste water rich in OM and silt into these coastal seagrass ecosystems (Sahu et al., 2023; Mishra et al., 2023; Mishra & Kumar, 2020; VishnuRadhan et al., 2015). The increase in sediment OM and silt content by such inputs has been previously documented for seagrass ecosystems at the locations of ANI, considered for this study (Gole et al., 2022; Mishra & Apte, 2020; Mishra & Farooq, 2023; Mishra & Kumar, 2020).
Seagrasses are considered as ‘ecosystem engineers’ because of their ability to modify the surrounding environment to accumulate allochthonous nutrients and OM that can benefit their growth and productivity (De Boer, 2007; Duarte & Krause-Jensen, 2017; Haviland et al., 2022). This interaction between seagrass and their surrounding environment resulting in increase in sediment OM content has been observed for T. hemprichii and H. ovalis meadows from our study locations at Burmanallah, Neil Island and Havelock Island previously (Gole et al., 2022; Mishra et al., 2021; Mishra & Farooq, 2023; Savurirajan et al., 2018), and other locations of ANI, where seagrass ecosystems are present within the intertidal regions dominated by coral reefs and mangrove ecosystems (Nobi et al., 2010; Sachithanandam et al., 2020a). Similar interactions have also been observed for seagrass ecosystems of the Lakshadweep Islands (Thangaradjou et al., 2014) and from the coast of Tamil Nadu at Palk Bay region ( Baby et al., 2017). The range of sediment OM (25.44– 38.70 %) observed in this study is 10-fold higher than the sediment OM previously reported from seagrass and coral reef meadows of ANI (Sachithanandam et al., 2020a) and 2-fold lower than previously reported for Neil Island, Havelock Island and Burmanallah T. hemprichii meadows (A. K. Mishra et al., 2023). These differences in our study and previous authors are a result of anthropogenic input at these locations, which varies yearly, as each summer season is followed by a wet season of monsoon and high wave actions, that either increases or decreases the sediment OM matter by sediment erosion or deposition, as observed from the coast of India (Pradhan et al., 2014) and ANI (Sahu et al., 2023).
The variation of sediment OM between locations and the surface water pH plays an important role in bioavailability of trace metals in sediment for seagrasses (Bonanno & Borg, 2018; Lee et al., 2023; Mishra & Farooq, 2022) . This interplay between sediment OM and surface water pH has been observed for various seagrass species globally (Mishra et al., 2020; Olivé et al., 2017; Renzi et al., 2011), where low pH helps in release of trace metals bound to sediment OM and increases their availability for uptake (Basallote et al., 2014). This interaction is observed in our study mostly at the Haddo Bay location, where the surface water pH and sediment OM is lower compared to the other three locations, but the trace metals concentrations are the highest among the locations due to the effects of low pH releasing the sediment OM and silt bound trace metals (Table 4). Further, at this location, most of the trace metals showed positive or negative corelationship with sediment OM compared to other locations (Figure 7 and Figure 8 and Supplementary S2). In this study, low pH helped in increase of Pb concentration in the sediment of H. beccarii meadows at Haddo Bay, as a result of significant negative correlation with pH (Supplementary S2). Similarly, low pH (7.9-7.5) leading to increase in concentration of Pb in sediment and seagrass tissues has been observed for other seagrass species like Posidonia oceanica and Cymodocea nodosa meadows in the Mediterranean Sea (Mishra et al., 2020; Vizzini et al., 2013). Furthermore, this inverse relationship between low pH and sediment OM and trace metals (like As, Pb and Co) have been observed in the Bay of Bengal region and southeast coast of India (Jayaprakash et al., 2016; Naik et al., 2023; Sadanandan et al., 2023).
4.2. Species-Specific Accumulation
The sediment of seagrass ecosystems acts as sink of anthropogenic trace metals (Lee et al., 2023; Mishra et al., 2022; Mishra & Farooq, 2022b). The sediment trace metal concentrations range observed in this study were lower than trace metals concentration range observed for seagrass ecosystems across the coast of India, except for Mg in the sediment of T. hemprichii meadows of Burmanallah, ANI (Table 3). These higher range of Mg (2206.07 – 8474.93 mg Kg-1) in seagrass meadows of ANI are within the range of previous results, where sediment of seagrass ecosystems of ANI accumulated higher concentration of Mg compared to dead coral reef and mangrove ecosystems (Nobi et al., 2010). The 1.3-fold higher range of Mg in our study compared to the coast of India, is a result of lack of research and monitoring of trace metals in the sediment of seagrass ecosystems across India, including the ANI, as the last assessment of Mg in the sediment of seagrass meadows was quantified more than two decades ago (Jagtap, 1983; Nobi et al., 2010). However, during this period both coastal communities and anthropogenic pollution have increased along the coast of ANI, which is reflected in the sediment of seagrass meadows. This increased anthropogenic pollution has resulted in increase of the minimum concentration (min. values) of all trace metals in this study compared to the seagrass sediments from the coast of India (Table 3).
This increase in minimum concentration of trace metals in sediment of seagrass ecosystems of ANI also resulted in significant increase of trace metal concentrations in the seagrass biomass (Table 4). The minimum and maximum Co concentration of in H. uninervis and T. hemprichii biomass in this study is 48-fold and 8.78-fold higher than the Co concentration observed in S. isoetifolium and H. uninervis biomass from Lakshadweep Islands (Thangaradjou et al., 2013). Similarly, Cr concentration in biomass of H. uninervis in this study was 33-fold higher than the Cr concentration observed in S. isoetifolium biomass from Gulf of Mannar, Tamil Nadu (Arisekar et al., 2021; Immaculate et al., 2018; Pasumpon & Vasudevan, 2021). The minimum concentrations of Cu and Fe observed in this study in the tissues of H. uninervis are also 5-fold and 2468-fold higher than the minimum concentration of Cu and Fe observed in S. isoetifolium biomass from the Palk Bay region of Tamil Nadu (Gopi et al., 2020; Immaculate et al., 2018; Pasumpon & Vasudevan, 2021). Similarly, the concentration of Ni, Zn and Pb in S. isoetifolium at Palk Bay, Tamil Nadu and Lakshadweep Islands are 5-fold, 4-fold and 4.7-fold lower than the minimum concentration observed for these trace metals in this study in the biomass of C. rotundata (Ni), E. acoroides (Pb) and T. hemprichii (Zn) (Table 4). This pattern of trace metals accumulation in this study and across India suggests that big seagrass species such as S. isoetifolium and E. acoroides tends to accumulate low concentration of trace metals in their tissues compared to their surrounding environment due to their unique growth patterns and presence at increased depths, presence of low organic matter content compared to the small seagrass species, which generally inhabits the intertidal regions that are continuously exposed to land derived anthropogenic contaminants with high sediment organic matter (Bonanno & Borg, 2018a; Bonanno & Raccuia, 2018; A. K. Mishra & Farooq, 2022b; Nazneen et al., 2022). The big seagrass species generally follow guerrilla growth strategy, i.e., low horizontal growth and high vertical growth rates (with wider leaves), that allows them to fulfill their minimum trace metal needs from the water column. Contrastingly small seagrass species follow a phalanx growth strategy, where these species grow faster horizontally (with high leaf turnover rates) getting exposed to high levels of trace metals that are found in the top 10 cm of the seagrass sediment (Govers et al., 2014; G. Lee et al., 2019; Y. Li et al., 2023; Vieira et al., 2022). These differences between small and big seagrass species trace metals accumulation capacity have been observed between Posidonia oceanica, Cymodocea nodosa and Halophila stipulacea in the Mediterranean Sea (Bonanno & Raccuia, 2018), between E. acoroides, T. hemprichii and H. ovalis and H. beccarii in the South China Sea (Lin et al., 2016; Zhang et al., 2021a, 2021b) and from the coast of India (Arulkumar et al., 2019; Gopi et al., 2020; Pasumpon & Vasudevan, 2021). Other factors such as interspecific differences in metal accumulation between seagrass species due to internal factors (such as uptake capacity) and external factors such as pH, OMcontent, water temperature and presence of fine grain fractions (<65 µm) also helps in determining the metal-species-specific accumulations as observed in this study (Table 4).
Despite small seagrass species accumulated higher concentration of trace metals than the minimum concentration observed for various seagrass species of India, this did not result in breaching the maximum concentration recorded for trace metals in various seagrass species from India’s coast. Exceptions were trace metals like Co, Ni and Zn for which the seagrasses of this study in ANI accumulated maximum concentration (Table 4). The Co concentration of in tissues of T. hemprichii in this study was 8.7-fold higher than the maximum concentration of Co observed in the tissues of H. uninervis from Lakshadweep Islands (Jagtap and Untawale, 1984; Thangaradjou et al., 2013). Similarly, the concentration of Ni and Zn in H. uninervis and H. beccarii was 2-fold and 1-fold higher than maximum concentration of these metals observed in the tissues of S. isoetifolium from Palk Bay, Tamil Nadu. These differences between our study and other studies from the coast of India are probably due to the presence of riverine and mangrove input at the locations from the coast of India, whereas in ANI there are no source of riverine input of trace metals (A. K. Mishra & Kumar, 2020; Nobi et al., 2010; Sachithanandam et al., 2020b). Additionally, being an island ecosystem the short-term spikes in trace metals input during dry season are balanced during the wet season, where large influx of freshwater and wave actions results in washing of the surface sediment and OM, resulting in transport of these metals to deeper waters, that does not allow a long-term accumulation of these metals. We sampled during the dry season, where trace metal concentration and anthropogenic input (due to tourism activities) are higher, that resulted in high concentration of metals in the sediment. However, these anthropogenic activities also cause short-term spikes in nutrient enrichment in these seagrass meadows, leading to growth of various macroalgae (Figure 2), that utilizes the available nutrients in the water column and available trace metals, thus reducing the load in the sediment (Gopinath et al., 2011; Nobi et al., 2010; Sachithanandam et al., 2020b; Schneider et al., 2018b; Tupan & Azrianingsih, 2016). This is one of the reasons why trace metals in the water column were in very low levels in this study.
4.3. Bioindicator Potential
Seagrass plants are considered as efficient bioindicators of coastal trace metal contamination, as seagrasses possess trace metal accumulation capacity both from the water column and sediment (Aljahdali & Alhassan, 2022; Bonanno & Borg, 2018b; H. Lee et al., 2023; Zhang et al., 2021b). Additionally, seagrasses contribute significantly to the coastal primary productivity and plays an important role in trace metal cycling (Nazneen et al., 2022; Sanz-Lázaro et al., 2012). Furthermore, seagrasses also regenerate and shed their leaves, as a result it is important to understand which part of the seagrass tissues are suitable as long-and short-term indicators (Y. Li et al., 2023; A. K. Mishra et al., 2022). From this study, it is evident that seagrass AG and BG-tissues are suitable indicators for different trace metals. For examples, the AG-tissues (i.e., leaves) of T. hemprichii were suitable bioindicators of trace metals like Co and Mg, whereas the BG-tissues (i.e., roots) of T. hemprichii are suitable bioindicators of Mn. Similarly, the AG-tissues of H. uninervis are better indicators of metals like Fe, Ni and Pb and BG-tissues of Cu. For trace metals like Cr and Zn, BG-tissues of H. ovalis and H. beccarii are better indicators (Figure 5, Figure 6 and Figure 7). This indicates small seagrass leaves of ANI can serve as short-term bioindicators, whereas roots can serve as long-term bioindicators of coastal trace metal contamination as observed for seagrasses worldwide (Aljahdali & Alhassan, 2022; H. Lee et al., 2023; A. K. Mishra & Farooq, 2022b; Zhang et al., 2021a).
4.3. Toxic Effects of Trace Metals on Seagrasses
In India, there are no trace metal toxicity studies on seagrasses ( Mishra et al., 2022; Mishra & Farooq, 2022b). However, globally various seagrass species have been used to assess the toxic effects of trace metals (Aljahdali & Alhassan, 2022; de los Santos et al., 2019; Gu et al., 2021; Lin et al., 2016; Mishra et al., 2021 and references therein). Among these seagrass species studied, few are observed in India, i.e., C. serrulata, H. ovalis, H. uninervis, H. stipulacea and T. hemprichii and species like T. hemprichii, H. ovalis, H. uninervis and Cymodocea sp., are part of this study in ANI. Globally, it has been observed that the concentration range to exert trace metal toxicity in seagrass tissues is both metal and species -specific (Ambo-rappe et al., 2011; L. Li & Huang, 2012; Malea & Kevrekidis, 2013). In T. hemprichii, the trace metal concentration observed in this study in AG-and BG-tissues are above the concentration levels that can exert toxic effects (i.e., reduction in photosynthetic pigments) for Cu (0.1 mg L-1), and Zn (10 mg L-1) (Lei et al., 2012), except the Zn concentration in leaves of T. hemprichii at Burmanallah. This suggests, that except for Burmanallah location this species may be experiencing toxic effects of Cu and Zn along the study locations of ANI, as a result, its growth rates are declining as observed for T. hemprichii (A. Mishra & Apte, 2020). Similarly, for H. ovalis the Cu (0.1 mg L-1) and Pb (10 mg L-1) levels are toxic to photosystem -II and leaf growth rates (Ambo-rappe et al., 2011; Prange & Dennison, 2000), and the concentration of Cu and Pb in H. ovalis tissues were significantly above these concertation levels in our study. This indicates that H. ovalis leaves experiencing the toxic effects of these metals, as a result they have the shorter leaves from the coast of ANI compared to the other areas of India (Mishra et al., 2021; Mishra et al., 2021).In H. uninervis, the concentration of Cu (1 mg L-1) is toxic to photosystem -II (Prange & Dennison, 2000) and the H. uninervis leaf tissues in our study has multifold higher concentration, possibly reducing its growth rates and resulting in lower canopy height in ANI compared to the other coastal areas of India (Dilipan et al., 2020; Mishra et al., 2021). However, for H. beccarii, E. acoroides and C. rotundata there have been no trace metal toxicity studies, which needs attention. However, as H. beccarii is a vulnerable seagrass species and inhabits intertidal areas or close to mangroves areas that are exposed to high anthropogenic contamination of trace metals, assessing the toxic effects of trace metals on H. beccarii needs to be prioritized in India (Mishra & Apte, 2021; Zhang et al., 2021c). However, it is important to note here that trace metal toxicity does not always depend on the total accumulated metal concentration in seagrass tissues, but on the threshold concentration of internal metabolically available concentration of the trace metals, (Rainbow, 2007), which needs further investigation for various seagrass species of ANI, India.
5. Conclusion
The influx of anthropogenic nutrients, trace metals, and persistent contaminants into coastal ecosystems is increasing rapidly. For effective assessment of the costal ecosystem health, it is necessary to identify various bioindicator species that may effectively demonstrate contaminant trends in both short-term and long-term. The present study highlights that seagrass ecosystems of ANI, India are exposed to increased anthropogenic trace metal influxes due to increase in human population and unregulated anthropogenic activities in these tropical islands. However, the input of anthropogenic trace metals to the seagrass ecosystems are location specific (Haddo Bay> Burmanallah> Havelock Island> Neil Island). The trace metal accumulation and indicator capacity of the six seagrass species assessed in this study indicates that small seagrass species like H. uninervis, H. ovalis and H. beccarii are better bioindicators of six trace metals (Cr, Cu, Fe, Ni, Pb, Zn) in their leaves or roots compared to big seagrass species like T. hemprichii and E. acoroides. Due to high growth rate and spatial coverage, small seagrass species can be utilized as short-term bioindicators and the big seagrass species as long-term bioindicators of trace metals. Thus, the integration of these seagrass species is recommended into the coastal monitoring programs of ANI and India. However, a crucial aspect that needs consideration is the trace metal toxicity on these various seagrass species. This comprehensive evaluation is essential to gain insights into the toxic effects of trace metals on seagrass physiology and trophic transfer of metals to seagrass consumers, such as endangered Dugong dugon that feeds on the dugong grass H. ovalis. Future studies in this direction will not only enhance our understanding of the potential toxic effects on human health through the consumption of fish associated with seagrass ecosystems but also contribute valuable information to coastal management strategies.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Funding
This project was supported through a Rauf Ali Fellowship 1/2021 supported through Rauf Ali Foundation, India.
Data availability
The data for this article is available as supplementary information and in the results section of the article.
Acknowledgements
We are thankful to IIT Bhubaneswar for their laboratory support.
Declaration of competing interest
The authors declare they have no conflict of interest to declare.
References
- Aljahdali, M.O.; Alhassan, A.B. Heavy metal accumulation and anti-oxidative feedback as a biomarker in Seagrass Cymodocea Serrulata. Sustainability (Switzerland) 2020, 12(7). [Google Scholar] [CrossRef]
- Aljahdali, M.O.; Alhassan, A.B. The efficiency of trace element uptake by seagrass Cymodocea serrulata in Rabigh lagoon, Red Sea. Environmental Science and Pollution Research 2022, 29(10), 14948–14960. [Google Scholar] [CrossRef]
- Ambo-rappe, R.; Lajus, D.L.; Schreider, M.J. Heavy metal impact on growth and leaf asymmetry of seagrass, Halophila ovalis. Environmental Chemistry 2011, 3(June), 149–159. [Google Scholar]
- Arisekar, U.; Jeya Shakila, R.; Shalini, R.; Jeyasekaran, G.; Sivaraman, B.; Surya, T. Heavy metal concentrations in the macroalgae, seagrasses, mangroves, and crabs collected from the Tuticorin coast (Hare Island), Gulf of Mannar, South India. Marine Pollution Bulletin 2021, 163(November 2020), 111971. [Google Scholar] [CrossRef]
- Arulkumar, A.; Nigariga, P.; Paramasivam, S.; Rajaram, R. Metals accumulation in edible marine algae collected from Thondi coast of Palk Bay, Southeastern India. Chemosphere 2019, 221, 856–862. [Google Scholar] [CrossRef]
- Avelar, M.; Bonilla-Heredia, B.; Merino-Ibarra, M.; Herrera-Silveira, J.A.; Ramirez, J.; Rosas, H.; Valdespino, J.; Carricart-Ganivet, J.P.; Martínez, A. Iron, cadmium, and chromium in seagrass (Thalassia testudinum) from a coastal nature reserve in karstic Yucatán. Environmental Monitoring and Assessment 2013, 185(9), 7591–7603. [Google Scholar] [CrossRef] [PubMed]
- Barbier, E.B.; Hacker, S.D.; Kennedy, C.; Koch, E.W.; Stier, A.C.; Silliman, B.R. The value of estuarine and coastal ecosystem services. Ecological Monographs 2011, 81(2), 169–193. [Google Scholar] [CrossRef]
- Basallote, M.D.; De Orte, M.R.; DelValls, T.Á.; Riba, I. Studying the Effect of CO2 -Induced Acidification on Sediment Toxicity Using Acute Amphipod Toxicity Test. Environmental Science & Technology 2014, 48(15), 8864–8872. [Google Scholar] [CrossRef]
- Sahu, B.K.; Goswami, P.; Begum, M.; Jha, D.K.; Vinithkumar, N.V.; Dharani, G. A comparative investigation of physicochemical and biological variables of Aerial & Port Blair Bays, Andaman Islands with focus on the anthropogenic influence. Indian Journal of Geo-Marine Sciences 2023, 52(02), 79–90. [Google Scholar] [CrossRef]
- Bayyana, S.; Pawar, S.; Gole, S.; Dudhat, S.; Pande, A.; Mitra, D.; Johnson, J.A.; Sivakumar, K. Detection and mapping of seagrass meadows at Ritchie’s archipelago using Sentinel 2A satellite imagery. Current Science 2020, 118(8), 1275–1282. [Google Scholar] [CrossRef]
- Bonanno, G.; Borg, J.A. Comparative analysis of trace element accumulation in seagrasses Posidonia oceanica and Cymodocea nodosa: Biomonitoring applications and legislative issues. Marine Pollution Bulletin 2018, 128(December 2017), 24–31. [Google Scholar] [CrossRef]
- Bonanno, G.; Borg, J.A. Comparative analysis of trace element accumulation in seagrasses Posidonia oceanica and Cymodocea nodosa: Biomonitoring applications and legislative issues. Marine Pollution Bulletin 2018, 128(January), 24–31. [Google Scholar] [CrossRef]
- Bonanno, G.; Orlando-Bonaca, M. Trace elements in Mediterranean seagrasses and macroalgae. A review. Science of the Total Environment 2018, 618, 1152–1159. [Google Scholar] [CrossRef]
- Bonanno, G.; Raccuia, S.A. Comparative assessment of trace element accumulation and bioindication in seagrasses Posidonia oceanica, Cymodocea nodosa and Halophila stipulacea. Marine Pollution Bulletin 2018, 131(February), 260–266. [Google Scholar] [CrossRef]
- Coclet, C.; Garnier, C.; D’Onofrio, S.; Durrieu, G.; Pasero, E.; Le Poupon, C.; Omanović, D.; Mullot, J.U.; Misson, B.; Briand, J.F. Trace Metal Contamination Impacts Predicted Functions More Than Structure of Marine Prokaryotic Biofilm Communities in an Anthropized Coastal Area. Frontiers in Microbiology 2021, 12(February), 1–16. [Google Scholar] [CrossRef]
- De Boer, W.F. Seagrass-sediment interactions, positive feedbacks and critical thresholds for occurrence: A review. Hydrobiologia 2007, 591(1), 5–24. [Google Scholar] [CrossRef]
- de los Santos, C.B.; Arenas, F.; Neuparth, T.; Santos, M.M. Interaction of short-term copper pollution and ocean acidification in seagrass ecosystems: Toxicity, bioconcentration and dietary transfer. Marine Pollution Bulletin 2019, 142(July 2018), 155–163. [Google Scholar] [CrossRef]
- Dilipan, E.; Nobi, E.P.; Change, C.; Thangaradjou, T. Spatial variability in productivity and biomass of seagrasses in the Andaman group of Islands, India. Ecology 2020, 26(4), 1693–1701. [Google Scholar]
- Dong, Y.; Rosenbaum, R.K.; Hauschild, M.Z. Assessment of Metal Toxicity in Marine Ecosystems: Comparative Toxicity Potentials for Nine Cationic Metals in Coastal Seawater. Environmental Science and Technology 2016, 50(1), 269–278. [Google Scholar] [CrossRef] [PubMed]
- Duarte, C.M.; Krause-Jensen, D. Export from seagrass meadows contributes to marine carbon sequestration. Frontiers in Marine Science 2017, 4(JAN), 1–7. [Google Scholar] [CrossRef]
- (EPA), E.P.A. (2009). Occurrence of Contaminants of Emerging Concern in Wastewater From Nine Publi c ly Owned Treatment Works August 2009. Epa, August.
- Esteban, N.; Unsworth, R.K.F.; Gourlay, J.B.Q.; Hays, G.C. The discovery of deep-water seagrass meadows in a pristine Indian Ocean wilderness revealed by tracking green turtles. Marine Pollution Bulletin 2018, 134(April 2017), 99–105. [Google Scholar] [CrossRef]
- Gao, W.; Du, Y.; Gao, S.; Ingels, J.; Wang, D. Heavy metal accumulation reflecting natural sedimentary processes and anthropogenic activities in two contrasting coastal wetland ecosystems, eastern China. Journal of Soils and Sediments 2016, 16(3), 1093–1108. [Google Scholar] [CrossRef]
- Geevarghese, G.A.; Akhil, B.; Magesh, G.; Krishnan, P.; Purvaja, R.; Ramesh, R. A comprehensive geospatial assessment of seagrass distribution in India. Ocean and Coastal Management 2018, 159(May 2017), 16–25. [Google Scholar] [CrossRef]
- Gole, S.; Mohammed, P.I.; Apte, D.; Marimuthu, N. Holothurian spatial variability and substratum preference in the intertidal habitats of the Andaman Sea. Regional Studies in Marine Science 2022, 56, 102633. [Google Scholar] [CrossRef]
- Gole, S.; Prajapati, S.; Prabakaran, N.; Das, H.; Kuppusamy, S.; Johnson, J.A. Spatial diversity and habitat characteristics of seagrass meadows with management recommendations in the Andaman and Nicobar Islands, India. Frontiers in Marine Science 2023, 10(December), 1–17. [Google Scholar] [CrossRef]
- Gopi, S.; Arulkumar, A.; Ganeshkumar, A.; Rajaram, R.; Miranda, J.M.; Paramasivam, S. Heavy metals accumulation in seagrasses collected from Palk Bay, South-eastern India. Marine Pollution Bulletin 2020, 157(June), 111305. [Google Scholar] [CrossRef]
- Gopinath, A.; Muraleedharan, N.S.; Chandramohanakumar, N.; Jayalakshmi, K.V. Statistical significance of biomonitoring of marine algae for trace metal levels in a coral environment. Environmental Forensics 2011, 12(1), 98–105. [Google Scholar] [CrossRef]
- Govers, L. L., Lamers, L. P. M., Bouma, T. J., Eygensteyn, J., de Brouwer, J. H. F., Hendriks, A. J., Huijbers, C. M., & van Katwijk, M. M. (2014). Seagrasses as indicators for coastal trace metal pollution: a global meta-analysis serving as a benchmark, and a Caribbean case study. Environmental Pollution (Barking, Essex : 1987), 195, 210–217. [CrossRef]
- Govindasamy, C.; Azariah, J. Seasonal variation of heavy metals in coastal water of the Coromandel coast, Bay of Bengal, India. Indian Journal of Marine Sciences 1999, 28(3), 249–256. [Google Scholar]
- Gu, R.; Lin, H.; Zhou, Y.; Song, X.; Xu, S.; Yue, S.; Zhang, Y.; Xu, S.; Zhang, X. Programmed responses of different life-stages of the seagrass Ruppia sinensis to copper and cadmium exposure. Journal of Hazardous Materials 2021, 403(February 2020), 123875. [Google Scholar] [CrossRef]
- Haviland, K.A.; Howarth, R.W.; Marino, R.; Hayn, M. Variation in sediment and seagrass characteristics reflect multiple stressors along a nitrogen-enrichment gradient in a New England lagoon. Limnology and Oceanography 2022, 67(3), 660–672. [Google Scholar] [CrossRef]
- Ho, N.A.J.; Ooi, J.L.S.; Affendi, Y.A.; Chong, V.C. Influence of habitat complexity on fish density and species richness in structurally simple forereef seagrass meadows. Botanica Marina 2018, 61(6), 547–557. [Google Scholar] [CrossRef]
- Howard, J.; Hoyt, S.; Isensee, K.; Pidgeon, E.; Telszewski, M. Coastal Blue Carbon. National Wetlands Newsletter 2014, 36(1), 5–7. [Google Scholar]
- Hu, W.; Zhang, D.; Chen, B.; Liu, X.; Ye, X.; Jiang, Q.; Zheng, X.; Du, J.; Chen, S. Mapping the seagrass conservation and restoration priorities : Coupling habitat suitability and anthropogenic pressures. Ecological Indicators 2021, 129, 107960. [Google Scholar] [CrossRef]
- Immaculate, TT, L., & Patterson, J. (2018). Macro and micro nutrients of seagrass species from Gulf of Mannar, India. MOJ Food Processing & Technology, 6(4), 391–398. [CrossRef]
- Jagtap, T.G. Metal Distribution in Halophila beccarii (Aschers) and sourrounding environment along the central west coast of India.pdf. Oceanography 1983, 16(4), 429–434. [Google Scholar]
- Jagtap,TG and Untawale, A. (1984). Chemical composition of marine macrophytes and their surrounding water and sediments, from Minicoy, Lakshadweep. Indian Journal of Marine Sciences, 13(September), 123–125.
- Jayaprakash, M.; Gopal, V.; Anandasabari, K.; Kalaivanan, R.; Sujitha, S.B.; Jonathan, M.P. Enrichment and toxicity of trace metals in near-shore bottom sediments of Cuddalore, SE coast of India. Environmental Earth Sciences 2016, 75(19), 1303. [Google Scholar] [CrossRef]
- Jiang, R.; Wu, Y.; Huang, X.; Liu, S.; Chen, Q. Heavy metal enrichment characteristics and risk assessment of typical fishes in tropical seagrass beds. South China Fisheries Science 2023, 19(1), 48–57. [Google Scholar] [CrossRef]
- Joseph, L.; Singh, P.; Singh, A.A.; Raj, K.; Maharaj, A. Implications of Seagrass Ecosystem Degradation on Marine Resources and People’s Livelihood: A Case Study from Komave Village, Fiji. Asian Journal of Fisheries and Aquatic Research 2018, February, 1–13. [Google Scholar] [CrossRef]
- Lee, G.; Suonan, Z.; Kim, S.H.; Hwang, D.-W.; Lee, K.-S. Heavy metal accumulation and phytoremediation potential by transplants of the seagrass Zostera marina in the polluted bay systems. Marine Pollution Bulletin 2019, 149(July), 110509. [Google Scholar] [CrossRef]
- Lee, H.; Morrison, C.; Doriean, N.J.C.; Welsh, D.T.; Bennett, W.W. Metals in coastal seagrass habitats: A systematic quantitative literature review. Critical Reviews in Environmental Science and Technology 2023, 53(17), 1568–1585. [Google Scholar] [CrossRef]
- Lei, L.I.; Xiaoping, H.; Borthakur, D.; Hui, N.I. Photosynthetic activity and antioxidative response of seagrass T halassia hemprichii to trace metal stress. Acta Oceanologica Sinica 2012, 31(3), 98–108. [Google Scholar] [CrossRef]
- Lewis, M.A.; Richard, D. Nonnutrient anthropogenic chemicals in seagrass ecosystems: Fate and effects. Environmental Toxicology and Chemistry 2009, 28(3), 644–661. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Huang, X. Three tropical seagrasses as potential bio-indicators to trace metals in Xincun Bay, Hainan Island, South China. Chinese Journal of Oceanology and Limnology 2012, 30(2), 212–224. [Google Scholar] [CrossRef]
- Li, Y.; Chen, F.; Zhou, R.; Zheng, X.; Pan, K.; Qiu, G.; Wu, Z.; Chen, S.; Wang, D. A review of metal contamination in seagrasses with an emphasis on metal kinetics and detoxification. Journal of Hazardous Materials 2023, 454(April), 131500. [Google Scholar] [CrossRef] [PubMed]
- Libin Baby, Gireesh Kumar TR, Remyakumari KR, J. V., & Sankar TV and Chandramohanakumar N. (2017). Comparison of hydrographic and sediment characteristics of seagrass meadows of Gulf of Mannar and Palk Bay, South West Coast of India. International Journal of Fisheries and Aquatic Studies, 5(2), 80–84. http://www.fisheriesjournal.com/archives/2017/vol5issue2/PartB/4-6-91-582.pdf.
- Lin, H.; Sun, T.; Zhou, Y.; Zhang, X. Anti-oxidative feedback and biomarkers in the intertidal seagrass Zostera japonica induced by exposure to copper, lead and cadmium. Marine Pollution Bulletin 2016, 109(1), 325–333. [Google Scholar] [CrossRef]
- Lu, Y.; Yuan, J.; Lu, X.; Su, C.; Zhang, Y.; Wang, C.; Cao, X.; Li, Q.; Su, J.; Ittekkot, V.; Garbutt, R.A.; Bush, S.; Fletcher, S.; Wagey, T.; Kachur, A.; Sweijd, N. Major threats of pollution and climate change to global coastal ecosystems and enhanced management for sustainability. Environmental Pollution 2018, 239, 670–680. [Google Scholar] [CrossRef] [PubMed]
- M. Stankovic, R. Ambo-Rappe, F. Carly, et al. (2021). Quantification ofblue carbon in seagrass ecosystems ofSoutheast Asia and their potential for climate change mitigation. Science of The Total Environment, 146858. [CrossRef]
- Malea, P., & Kevrekidis, T. (2013). Trace element ( Al , As , B , Ba , Cr , Mo , Ni , Se , Sr , Tl , U and V ) distribution and seasonality in compartments of the seagrass Cymodocea nodosa. Science of the Total Environment, The, 463–464, 611–623. [CrossRef]
- McKenzie, L.; Yoshida, R.; Aini, J.; Andréfouët, S.; Colin, P.; Cullen-Unsworth, L.; Hughes, A.; Payri, C.; Rota, M.; Shaw, C.; Tsuda, R.; Vuki, V.; Unsworth, R. Seagrass ecosystem contributions to people’s quality of life in the Pacific Island Countries and Territories. Marine Pollution Bulletin 2021, 167. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.; Apte, D. Ecological connectivity with mangroves influences tropical seagrass population longevity and meadow traits within an island ecosystem. Marine Ecology Progress Series 2020, 644, 47–63. [Google Scholar] [CrossRef]
- Mishra, A.K.; Acharya, P.; Apte, D.; Farooq, S.H. Seagrass ecosystem adjacent to mangroves store higher amount of organic carbon of Andaman and Nicobar Islands, Andaman Sea. Marine Pollution Bulletin 2023, 193(June), 115135. [Google Scholar] [CrossRef]
- Mishra, A.K.; Cabaço, S.; de los Santos, C.B.; Apostolaki, E.T.; Vizzini, S.; Santos, R. Long-term effects of elevated CO2 on the population dynamics of the seagrass Cymodocea nodosa: Evidence from volcanic seeps. Marine Pollution Bulletin 2021, 162(July 2020), 111824. [Google Scholar] [CrossRef]
- Mishra, A.K.; Farooq, S.H. Trace metal accumulation in seagrass and saltmarsh ecosystems of India: comparative assessment and bioindicator potential. Marine Pollution Bulletin 2022, 174, 113251. [Google Scholar] [CrossRef]
- Mishra, A.K.; Farooq, S.H. Trace metal accumulation in seagrass and saltmarsh ecosystems of India: comparative assessment and bioindicator potential. Marine Pollution Bulletin 2022, 174, 113251. [Google Scholar] [CrossRef]
- Mishra, A.K.; Farooq, S.H. Sediment organic matter content drives bivalve density in tropical oligotrophic seagrass ecosystem. Indian Journal of Geo-Marine Sciences 2023, 52(02), 91–100. [Google Scholar] [CrossRef]
- Mishra, A. K., Khadanga, M. K., Patro, S., Apte, D., & Farooq, S. H. (2021). Population structure of a newly recorded ( Halodule uninervis ) and native seagrass ( Halophila ovalis ) species from an intertidal creek ecosystem. Lakes & Reservoirs: Science, Policy and Management for Sustainable Use, 26(3), 1–12. [CrossRef]
- Mishra, A.K.; Kumar, M. Andaman mangrove sediments : source of nutrients and sink of heavy metals. Indian Journal of Geo Marine Sciences 2020, 49(January), 156–166. [Google Scholar] [CrossRef]
- Mishra, A.K.; Mohanraju, R. Epiphytic Bacterial Communities in Seagrass Meadows of Oligotrophic Waters of Andaman Sea. OALib 2018, 05(03), 1–12. [Google Scholar] [CrossRef]
- Mishra, A.K.; Narayana, S.; Apte, D. Loss of Dugong Grass [Halophila Ovalis (R. Brown)] Population Structure Due to Habitat Disturbance in an Island Ecosystem. Indian Journal of Geo-Marine Sciences 2021, 50(02), 115–121. http://nopr.niscair.res.in/handle/123456789/56320. [CrossRef]
- Mishra, A. K., Sahoo, R., Samantaray, S. S., & Apte, D. (2022). Seagrass Ecosystems of India as Bioindicators of Trace Elements. In S. P. Madhav S., Nazneen S. (Ed.), Coastal Ecosystems (1st ed., pp. 45–65). Springer Nature. [CrossRef]
- 65. Mishra, A. K., Santos, R., & Hall -Spencer, J. M. (2020). Elevated trace elements in sediments and seagrasses at CO2 seeps. Marine Environmental Research, 153(September 2019), 104810. [CrossRef]
- Mishra, A. Kumar., & Apte, Deepak. (2021). The current status of Halophila beccarii: An ecologically significant, yet vulnerable seagrass of India. Ocean & Coastal Management, 200(August 2020), 105484. [CrossRef]
- Naik, S., Pradhan, U., Karthikeyan, P., Bandyopadhyay, D., Sahoo, R. K., Panda, U. S., Mishra, P., & Murthy, M. V. R. (2023). Ecological risk assessment of heavy metals in the coastal sediment in the South-western Bay of Bengal. Frontiers in Marine Science, 10(October). [CrossRef]
- Nazneen, S.; Mishra, A.K.; Raju, N.J.; Mehmood, G. Coastal macrophytes as bioindicators of trace metals in the Asia’s largest lagoon ecosystem. Marine Pollution Bulletin 2022, 178, 113576. [Google Scholar] [CrossRef] [PubMed]
- Nikalje, G.C.; Suprasanna, P. Coping with metal toxicity – cues from halophytes. Frontiers in Plant Science 2018, 9(June), 1–11. [Google Scholar] [CrossRef]
- Nishitha, D.; Amrish, V.N.; Arun, K.; Warrier, A.K.; Udayashankar, H.N.; Balakrishna, K. Study of trace metal contamination and ecological risk assessment in the sediments of a tropical river estuary, Southwestern India. Environmental Monitoring and Assessment 2022, 194(2), 94. [Google Scholar] [CrossRef] [PubMed]
- Nobi, E.P.; Dilipan, E.; Thangaradjou, T.; Sivakumar, K.; Kannan, L. Geochemical and geo-statistical assessment of heavy metal concentration in the sediments of different coastal ecosystems of Andaman Islands, India. Estuarine, Coastal and Shelf Science 2010, 87(2), 253–264. [Google Scholar] [CrossRef]
- Nordlund, L.M.; Koch, E.W.; Barbier, E.B.; Creed, J.C. Seagrass Ecosystem Services and Their Variability across Genera and Geographical Regions. PLoS ONE 2016, October. [Google Scholar] [CrossRef]
- Olivé, I.; Silva, J.; Lauritano, C.; Costa, M.M.; Ruocco, M.; Procaccini, G.; Santos, R. Linking gene expression to productivity to unravel long- and short-term responses of seagrasses exposed to CO2 in volcanic vents. Scientific Reports 2017, 7(February), 42278. [Google Scholar] [CrossRef] [PubMed]
- Oreska, M.P.J.; Wilkinson, G.M.; McGlathery, K.J.; Bost, M.; McKee, B.A. Non-seagrass carbon contributions to seagrass sediment blue carbon. Limnology and Oceanography 2018, 63, S3–S18. [Google Scholar] [CrossRef]
- Pasumpon, N.; Vasudevan, S. Seasonal variation of heavy metals in seagrasses along Thondi coast, Palk Bay, India. Environmental Science and Pollution Research 2021, 28(21), 26849–26857. [Google Scholar] [CrossRef]
- Pradhan, U.K.; Wu, Y.; Shirodkar, P.V.; Zhang, J.; Zhang, G. Sources and distribution of organic matter in thirty five tropical estuaries along the west coast of India-a preliminary assessment. Estuarine, Coastal and Shelf Science 2014, 151, 21–33. [Google Scholar] [CrossRef]
- Prange, J. a., & Dennison, W. C. (2000). Physiological responses of five seagrass species to trace metals. Marine Pollution Bulletin, 41(7–12), 327–336. [CrossRef]
- Ragupathi Raja Kannan, R.; Arumugam, R.; Anantharaman, P. Chemometric studies of multielemental composition of few seagrasses from Gulf of Mannar, India. Biological Trace Element Research 2011, 143(2), 1149–1158. [Google Scholar] [CrossRef] [PubMed]
- Rainbow, P.S. Trace metal bioaccumulation: Models, metabolic availability and toxicity. Environment International 2007, 33(4), 576–582. [Google Scholar] [CrossRef] [PubMed]
- Renzi, M.; Romeo, T.; Guerranti, C.; Perra, G.; Italiano, F.; Focardi, S.E.; Esposito, V.; Andaloro, F. Temporal trends and matrix-dependent behaviors of trace elements closed to a geothermal hot-spot source (Aeolian Archipelago, Italy). Procedia Earth and Planetary Science 2011, 4(1), 10–28. [Google Scholar] [CrossRef]
- Sachithanandam, V.; Bonthu, S.; Mageswaran, T.; Singh, K.S.; Vimala, J.; Sridhar, R.; Purvaja, R.; Ramesh, R. Effect of hydrodynamic conditions on seagrass ecosystems during Cyclone Lehar in the South Andaman Islands, India. Ecohydrology and Hydrobiology 2022, 22(4), 640–659. [Google Scholar] [CrossRef]
- Sachithanandam, V., Parthasarathy, P., Sai Elangovan, S., Kasilingam, K., Dhivya, P., Mageswaran, T., & Mohan, P. M. (2020a). A baseline study on trace metals concentration and its ecological risk assessment from the coast of South Andaman Island, India. Regional Studies in Marine Science, 36, 101242. [CrossRef]
- Sachithanandam, V.; Parthasarathy, P.; Sai Elangovan, S.; Kasilingam, K.; Dhivya, P.; Mageswaran, T.; Mohan, P.M. A baseline study on trace metals concentration and its ecological risk assessment from the coast of South Andaman Island, India. Regional Studies in Marine Science 2020, 36, 101242. [Google Scholar] [CrossRef]
- Sadanandan, H.; Dharmalingam, S.N.; Sridharan, M.; Agarwal, N.; Anbuselvan, N. Assessment of Trace Metal Pollution in Surface Sediments from Southwestern Part of Bay of Bengal, India. Journal of the Geological Society of India 2023, 99(3), 383–389. [Google Scholar] [CrossRef]
- Sahu, B.K.; Begum, M.; Khadanga, M.K.; Jha, D.K.; Vinithkumar, N.V.; Kirubagaran, R. Evaluation of significant sources influencing the variation of physico-chemical parameters in Port Blair Bay, South Andaman, India by using multivariate statistics. Marine Pollution Bulletin 2013, 66(1–2), 246–251. [Google Scholar] [CrossRef]
- Sanz-Lázaro, C.; Malea, P.; Apostolaki, E.T.; Kalantzi, I.; Marín, a.; Karakassis, I. The role of the seagrass Posidonia oceanica in the cycling of trace elements. Biogeosciences 2012, 9(7), 2497–2507. [Google Scholar] [CrossRef]
- Savurirajan, M.; Lakra, R.K.; Equbal, J.; Ganesh, K.S.T. A note on morphometric, shoot density and biomass of Thalassia hemprichii from the South Andaman coast, Andaman and Nicobar Islands, India. Indian Journal of Geo-Marine Sciences 2018, 47(6), 1222–1227. [Google Scholar]
- Schneider, L., Maher, W. A., Potts, J., Taylor, A. M., Batley, G. E., Krikowa, F., Adamack, A., Chariton, A. A., & Gruber, B. (2018a). Trophic transfer of metals in a seagrass food web: Bioaccumulation of essential and non-essential metals. Marine Pollution Bulletin, 131(December 2017), 468–480. 20 December. [CrossRef]
- Schneider, L., Maher, W. A., Potts, J., Taylor, A. M., Batley, G. E., Krikowa, F., Adamack, A., Chariton, A. A., & Gruber, B. (2018b). Trophic transfer of metals in a seagrass food web: Bioaccumulation of essential and non-essential metals. Marine Pollution Bulletin, 131(December 2017), 468–480. 20 December. [CrossRef]
- Short, F., Carruthers, T., Dennison, W., & Waycott, M. (2007). Global seagrass distribution and diversity: A bioregional model. Journal of Experimental Marine Biology and Ecology, 350(1–2), 3–20. [CrossRef]
- Stockbridge, J.; Jones, A.R.; Gillanders, B.M. A meta-analysis of multiple stressors on seagrasses in the context of marine spatial cumulative impacts assessment. Scientific Reports 2020, 10(1), 1–11. [Google Scholar] [CrossRef]
- Stockdale, A.; Tipping, E.; Lofts, S.; Mortimer, R.J.G. Effect of Ocean Acidification on Organic and Inorganic Speciation of Trace Metals. Environmental Science and Technology 2016, 50(4), 1906–1913. [Google Scholar] [CrossRef] [PubMed]
- Suheryanto, S., & Ismarti, I. (2018). Bio-concentration factors of copper (Cu) and lead (Pb) in seagrass and some fish from coast Batam, Riau Islands, Indonesia. Journal of Physics: Conference Series, 1095(1), 0–5. [CrossRef]
- Sungur, A.; Özcan, H. Chemometric and geochemical study of the heavy metal accumulation in the soils of a salt marsh area (Kavak Delta, NW Turkey). Journal of Soils and Sediments 2015, 15(2), 323–331. [Google Scholar] [CrossRef]
- Thangaradjou, T.; Nobi, E.P.; Dilipan, E.; Sivakumar, K.; Susila, S. Heavy metal enrichment in seagrasses of Andaman Islands and its implication to the health of the coastal ecosystem. Indian Journal of Marine Sciences 2010, 39(March), 85–91. [Google Scholar]
- Thangaradjou, T.; Raja, S.; Subhashini, P.; Nobi, E.P.; Dilipan, E. Heavy metal enrichment in the seagrasses of Lakshadweep group of islands - A multivariate statistical analysis. Environmental Monitoring and Assessment 2013, 185(1), 673–685. [Google Scholar] [CrossRef]
- Thangaradjou, T.; Subhashini, P.; Raja, S.; Dilipan, E.; Nobi, E.P. Evidences for heavy metal contamination in surface sediments of seagrass ecosystem of Lakshadweep archipelago, India. Environmental Earth Sciences 2014, 71(3), 1135–1146. [Google Scholar] [CrossRef]
- Thorne-Bazarra, T.; Lozano-Bilbao, E.; Hardisson, A.; González-Weller, D.; Rubio, C.; Paz, S.; Gutiérrez, Á.J. Seagrass meadows serve as buffers for metal concentrations in the fish species Sparisoma cretense in the Canary Islands (Atlantic EC, Spain). Regional Studies in Marine Science 2023, 67, 103192. [Google Scholar] [CrossRef]
- Tupan, C.I.; Azrianingsih, R. Accumulation and deposition of lead heavy metal in the tissues of roots, rhizomes and leaves of seagrass Thalassia hemprichii (Monocotyledoneae, Hydrocharitaceae). AACL Bioflux 2016, 9(3), 580–589. [Google Scholar]
- Turekian, K.K., and Wedepohl,K.H., T. (1961). Distribution of the Elements in Some Major Units of the Earth’s Crust. Geological Society of America Bulletin, 72(February), 175–192. [CrossRef]
- Unsworth, Richard K F., C.-U. (2022). The planetary role of seagrass conservation. Science, August, 0–6. [CrossRef]
- Vieira, V.M.N.C.S.; Lobo-Arteaga, J.; Santos, R.; Leitão-Silva, D.; Veronez, A.; Neves, J.M.; Nogueira, M.; Creed, J.C.; Bertelli, C.M.; Samper-Villarreal, J.; Pettersen, M.R.S. Seagrasses benefit from mild anthropogenic nutrient additions. Frontiers in Marine Science 2022, 9(November), 1–14. [Google Scholar] [CrossRef]
- VishnuRadhan, R.; Thresyamma, D.D.; Sarma, K.; George, G.; Shirodkar, P.; Vethamony, P. Influence of natural and anthropogenic factors on the water quality of the coastal waters around the South Andaman in the Bay of Bengal. Natural Hazards 2015, 78(1), 309–331. [Google Scholar] [CrossRef]
- Vizzini, S.; Costa, V.; Tramati, C.; Gianguzza, P.; Mazzola, A. Trophic transfer of trace elements in an isotopically constructed food chain from a semi-enclosed marine coastal area (Stagnone di Marsala, Sicily, Mediterranean). Archives of Environmental Contamination and Toxicology 2013, 65(4), 642–653. [Google Scholar] [CrossRef]
- Ward, T. J. (1984). Role of acute metal toxicity in structuring seagrass fauna near a lead smelter. 17, 117–124.
- Wilkinson, A.; Ariel, E.; van de Merwe, J.; Brodie, J. Trace element concentrations in forage seagrass species of Chelonia mydas along the Great Barrier Reef. PLOS ONE 2022, 17(6), e0269806. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L., Ni, Z., Cui, L., Li, J., He, J., Jiang, Z., & Huang, X. (2021a). Heavy metal accumulation and ecological risk on four seagrass species in South China. Marine Pollution Bulletin, 173(PB), 113153. [CrossRef]
- Zhang, L., Ni, Z., Cui, L., Li, J., He, J., Jiang, Z., & Huang, X. (2021b). Heavy metal accumulation and ecological risk on four seagrass species in South China. Marine Pollution Bulletin, 173(PB), 113153. [CrossRef]
- Zhang, L., Ni, Z., Cui, L., Li, J., He, J., Jiang, Z., & Huang, X. (2021c). Heavy metal accumulation and ecological risk on four seagrass species in South China. Marine Pollution Bulletin, 173(PB), 113153. [CrossRef]
- zu Ermgassen, P. S. E., DeAngelis, B., Gair, J. R., Ermgassen, S. zu, Baker, R., Daniels, A., MacDonald, T. C., Meckley, K., Powers, S., Ribera, M., Rozas, L. P., & Grabowski, J. H. (2021). Estimating and Applying Fish and Invertebrate Density and Production Enhancement from Seagrass, Salt Marsh Edge, and Oyster Reef Nursery Habitats in the Gulf of Mexico. Estuaries and Coasts, 1588–1603. [CrossRef]
Figure 1.
Map showing the four study locations (Neil, Havelock, Burmanallah and Haddo Bay) in the Andaman and Nicobar Islands (ANI) of India.
Figure 1.
Map showing the four study locations (Neil, Havelock, Burmanallah and Haddo Bay) in the Andaman and Nicobar Islands (ANI) of India.
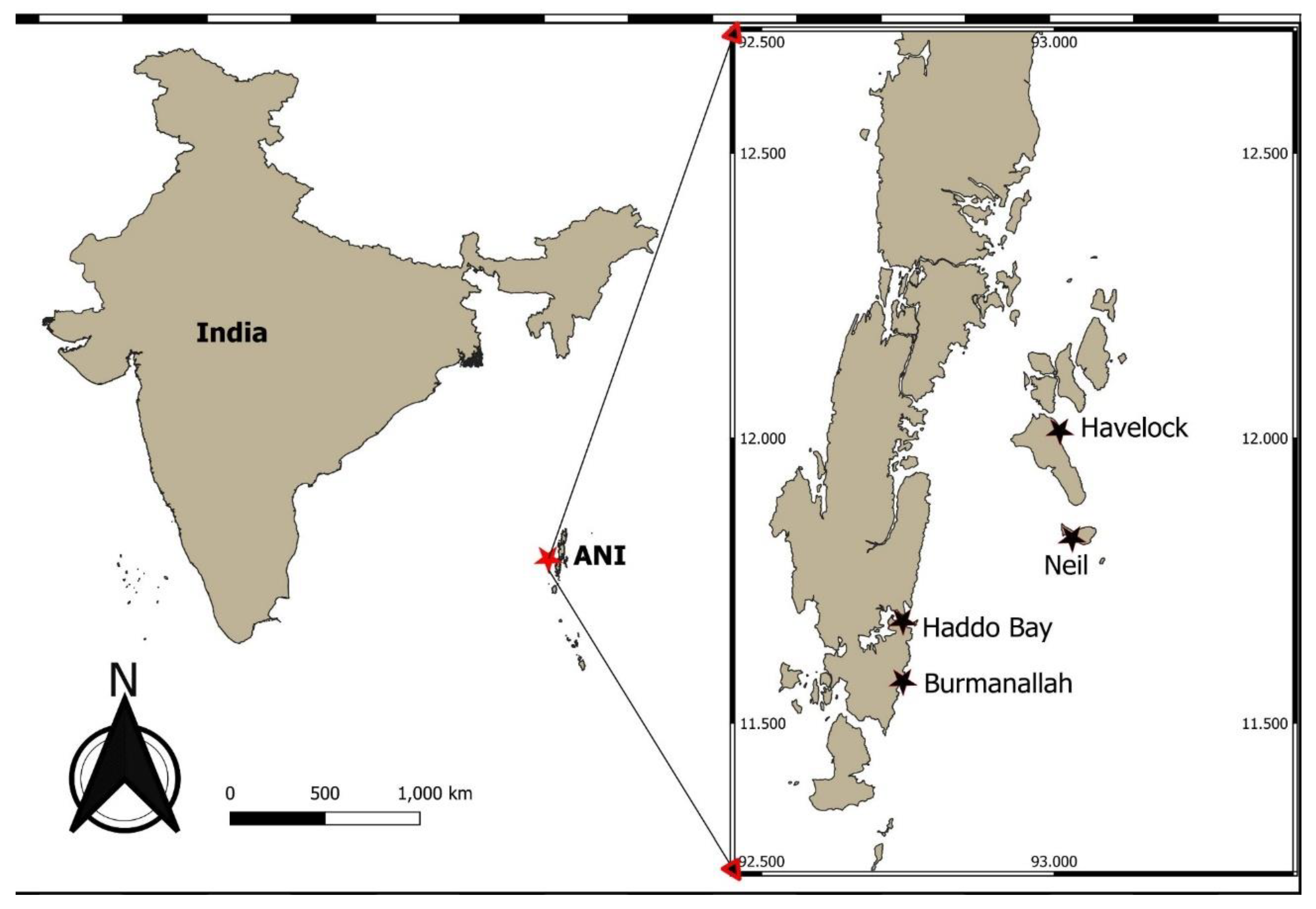
Figure 2.
Pictures of all six seagrass species a) T. hemprichii, b) C. rotundata, c) H. uninervis, d) H. ovalis, e) H. beccarii and f) E. acoroides across the four locations of ANI.
Figure 2.
Pictures of all six seagrass species a) T. hemprichii, b) C. rotundata, c) H. uninervis, d) H. ovalis, e) H. beccarii and f) E. acoroides across the four locations of ANI.



Figure 3.
Geoaccumulation Index (Igeo) of various trace metals in the sediment of seagrass meadows across the four study locations of ANI.
Figure 3.
Geoaccumulation Index (Igeo) of various trace metals in the sediment of seagrass meadows across the four study locations of ANI.
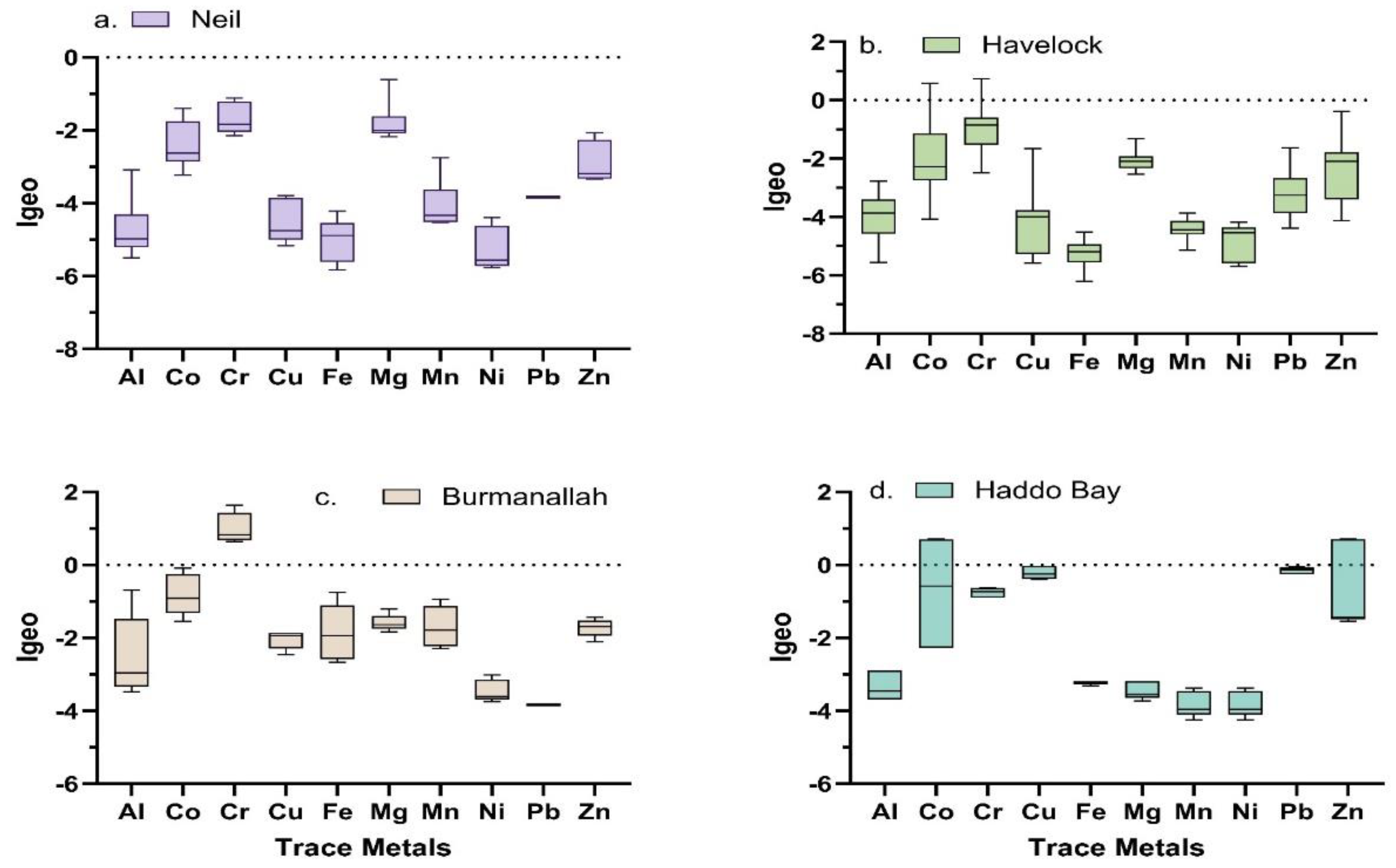
Figure 4.
Bio-Sediment Accumulation Factor (BSAF) values of various trace metals between sediment and below-ground tissues of seagrass meadows across the four study locations of ANI. BSAF>1 indicates higher accumulation capacity for the specific trace metals.
Figure 4.
Bio-Sediment Accumulation Factor (BSAF) values of various trace metals between sediment and below-ground tissues of seagrass meadows across the four study locations of ANI. BSAF>1 indicates higher accumulation capacity for the specific trace metals.
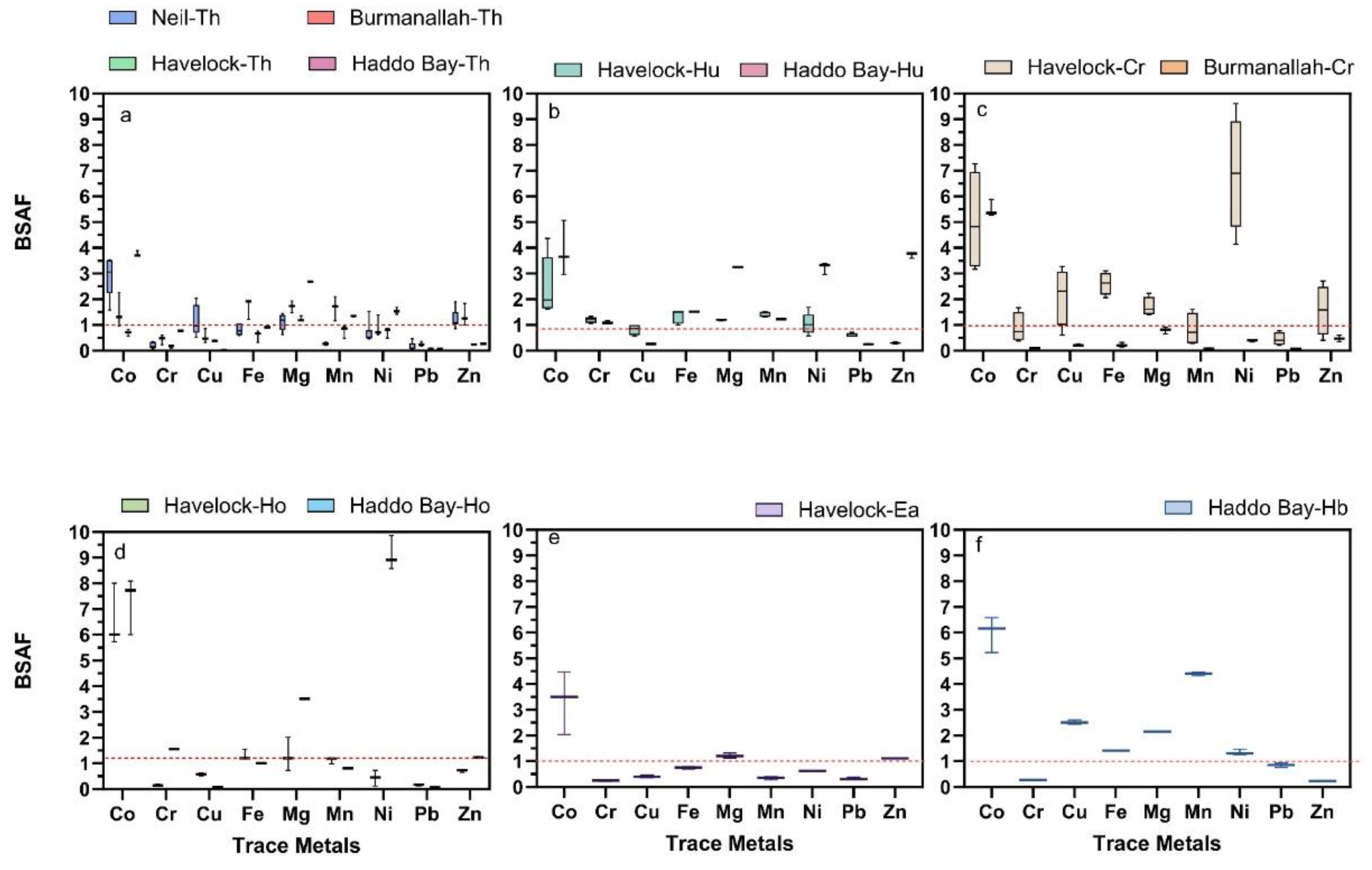
Figure 5.
Trace metals concentration of a) Co, b) Cr and c) Cu in sediment (Sd) and above- ground (AG) and below-ground (BG) tissues of T. hemprichii (Th), C. rotundata (Cr), H. uninervis (Hu) and H. ovalis (Ho) across the four study locations of ANI. Significant differences were derived from two-way ANOVA analysis using location and compartments (Sd, AG, BG) as fixed factors. (p<0.0001***, p<0.001**).
Figure 5.
Trace metals concentration of a) Co, b) Cr and c) Cu in sediment (Sd) and above- ground (AG) and below-ground (BG) tissues of T. hemprichii (Th), C. rotundata (Cr), H. uninervis (Hu) and H. ovalis (Ho) across the four study locations of ANI. Significant differences were derived from two-way ANOVA analysis using location and compartments (Sd, AG, BG) as fixed factors. (p<0.0001***, p<0.001**).
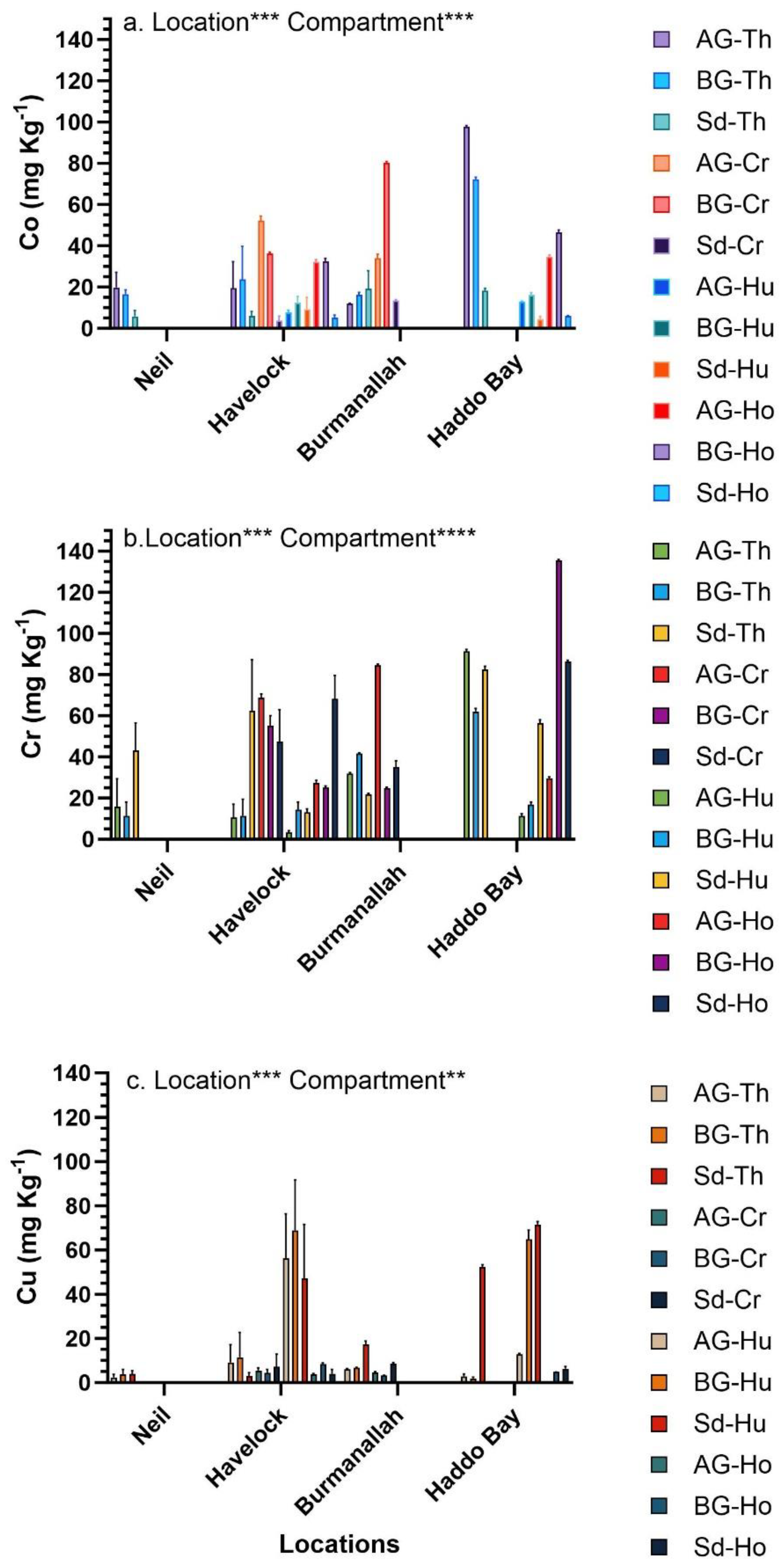
Figure 6.
Trace metals concentration of a) Fe, b) Mg and c) Mn in sediment (Sd) and above- ground (AG) and below-ground (BG) tissues of T. hemprichii (Th), C. rotundata (Cr), H. uninervis (Hu) and H. ovalis (Ho) across the four study locations of ANI. Significant differences were derived from two-way ANOVA analysis using location and compartments (Sd, AG, BG) as fixed factors. (p<0.0001***, p<0.001**).
Figure 6.
Trace metals concentration of a) Fe, b) Mg and c) Mn in sediment (Sd) and above- ground (AG) and below-ground (BG) tissues of T. hemprichii (Th), C. rotundata (Cr), H. uninervis (Hu) and H. ovalis (Ho) across the four study locations of ANI. Significant differences were derived from two-way ANOVA analysis using location and compartments (Sd, AG, BG) as fixed factors. (p<0.0001***, p<0.001**).
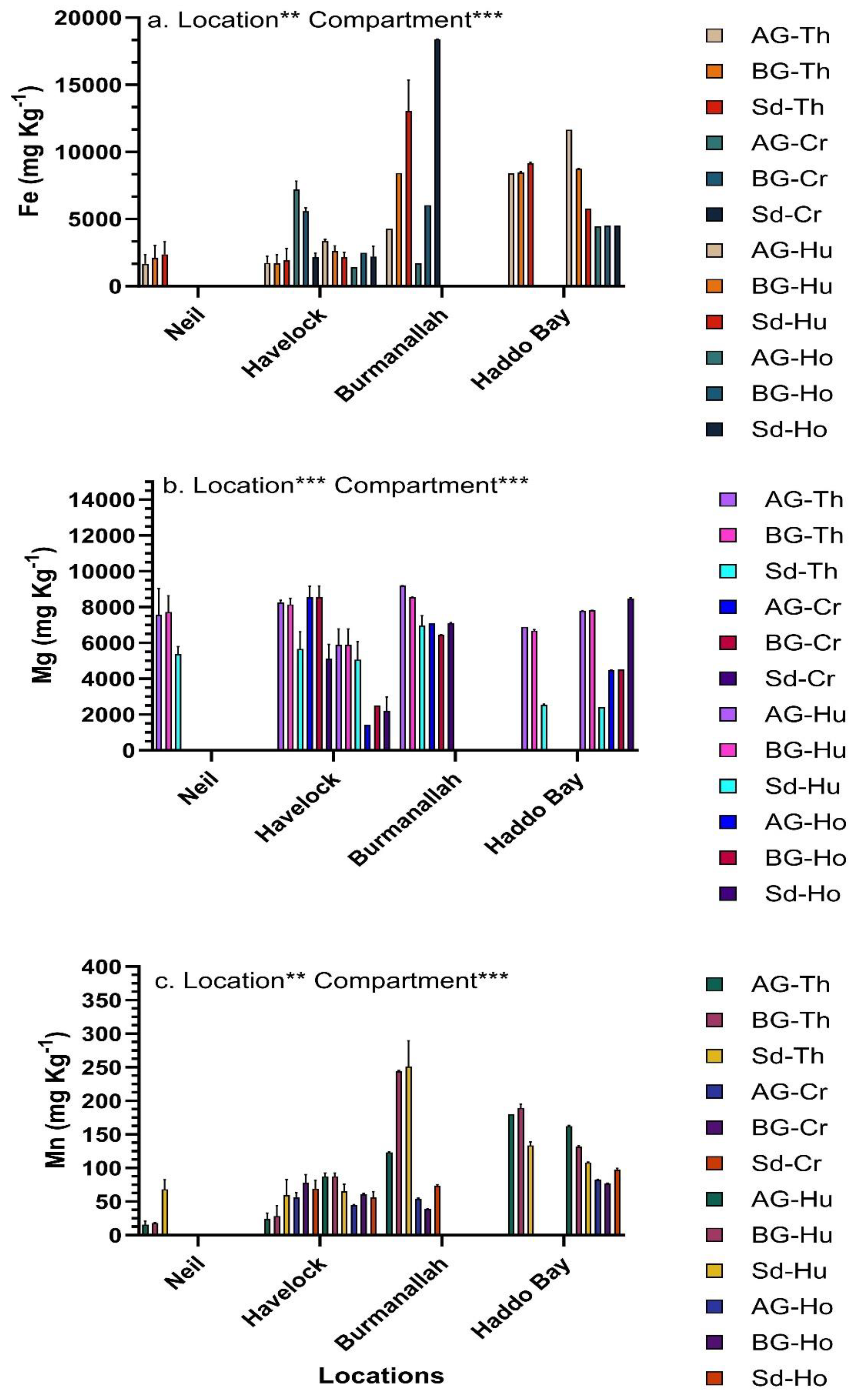
Figure 7.
Trace metals concentration of a) Ni b) Pb and c) Zn in sediment (Sd) and above- ground (AG) and below-ground (BG) tissues of T. hemprichii (Th), C. rotundata (Cr), H. uninervis (Hu) and H. ovalis (Ho) across the four study locations of ANI. Significant differences were derived from two-way ANOVA analysis using location and compartments (Sd, AG, BG) as fixed factors. (p<0.0001***, p<0.001**, p<0.01*).
Figure 7.
Trace metals concentration of a) Ni b) Pb and c) Zn in sediment (Sd) and above- ground (AG) and below-ground (BG) tissues of T. hemprichii (Th), C. rotundata (Cr), H. uninervis (Hu) and H. ovalis (Ho) across the four study locations of ANI. Significant differences were derived from two-way ANOVA analysis using location and compartments (Sd, AG, BG) as fixed factors. (p<0.0001***, p<0.001**, p<0.01*).
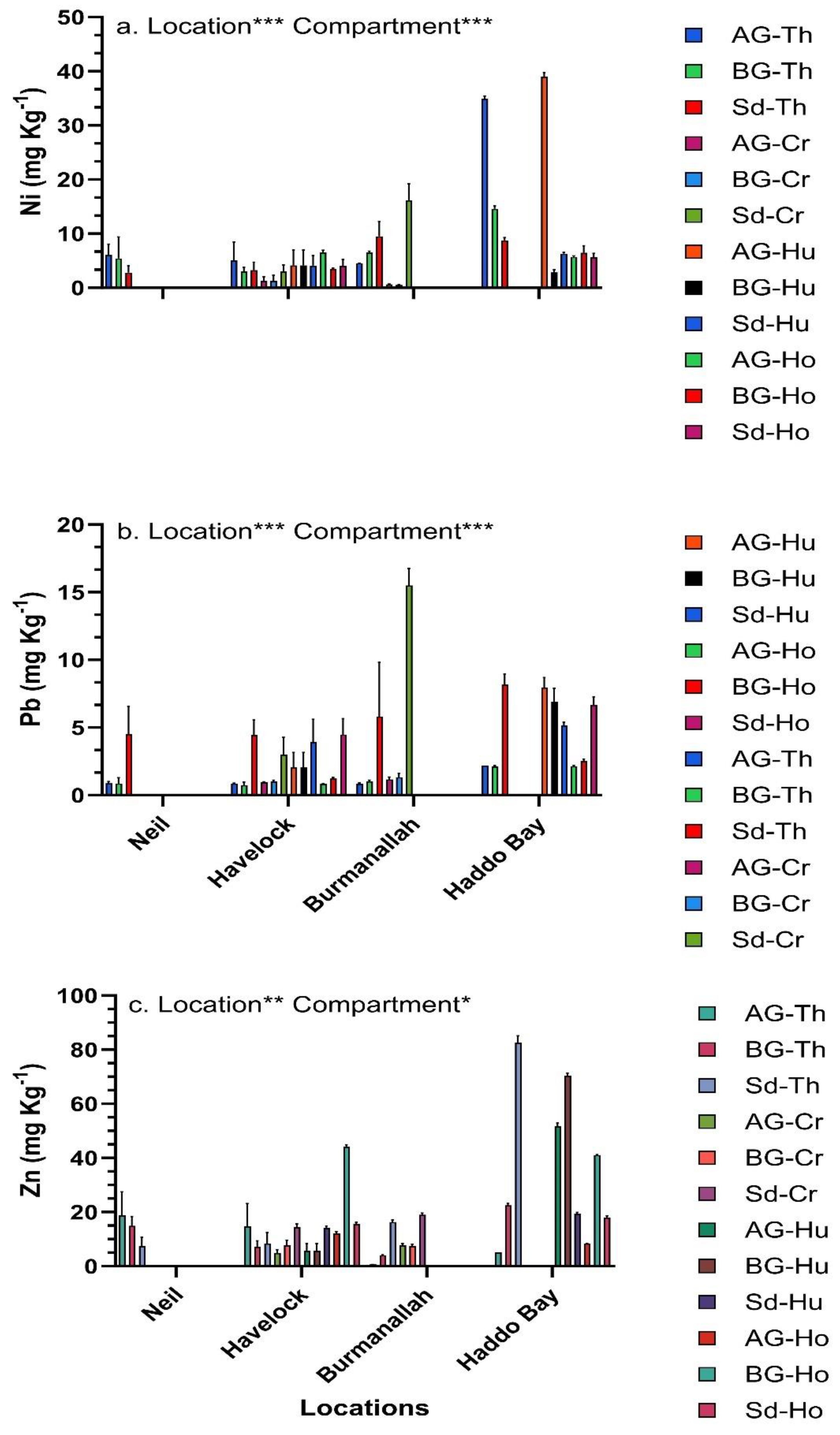
Figure 8.
Pearson correlation between the trace metals in sediment and abiotic parameters of sediment (OM and sediment grain size) and surface water (pH) across the four locations of ANI for T. hemprichii.
Figure 8.
Pearson correlation between the trace metals in sediment and abiotic parameters of sediment (OM and sediment grain size) and surface water (pH) across the four locations of ANI for T. hemprichii.
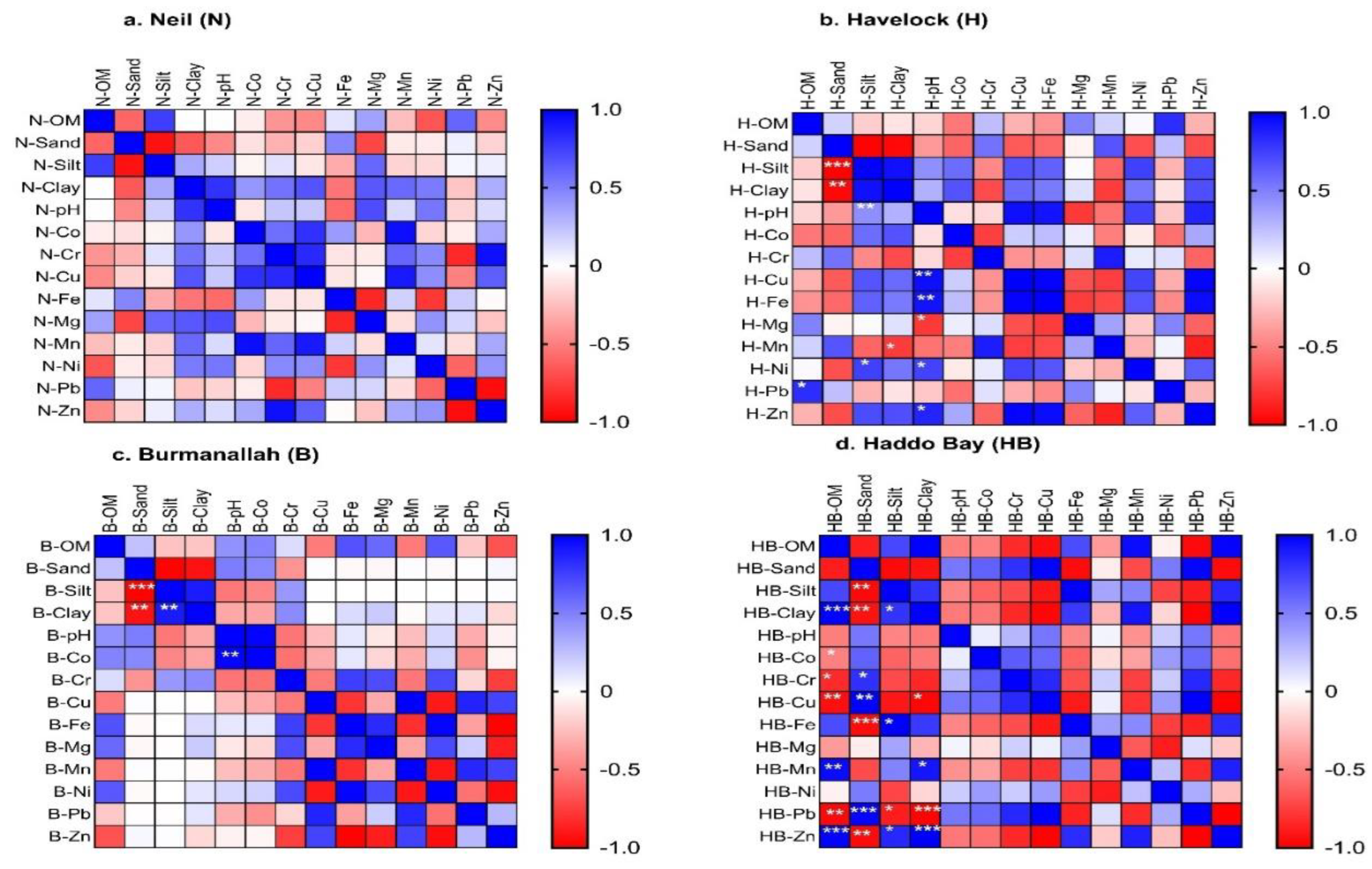
Figure 9.
Pearson correlation between the trace metals in sediment and abiotic parameters of sediment (OM and sediment grain size) and surface water (pH) across the four locations of ANI for H. uninervis (a &b) and H. ovalis (c & d).
Figure 9.
Pearson correlation between the trace metals in sediment and abiotic parameters of sediment (OM and sediment grain size) and surface water (pH) across the four locations of ANI for H. uninervis (a &b) and H. ovalis (c & d).
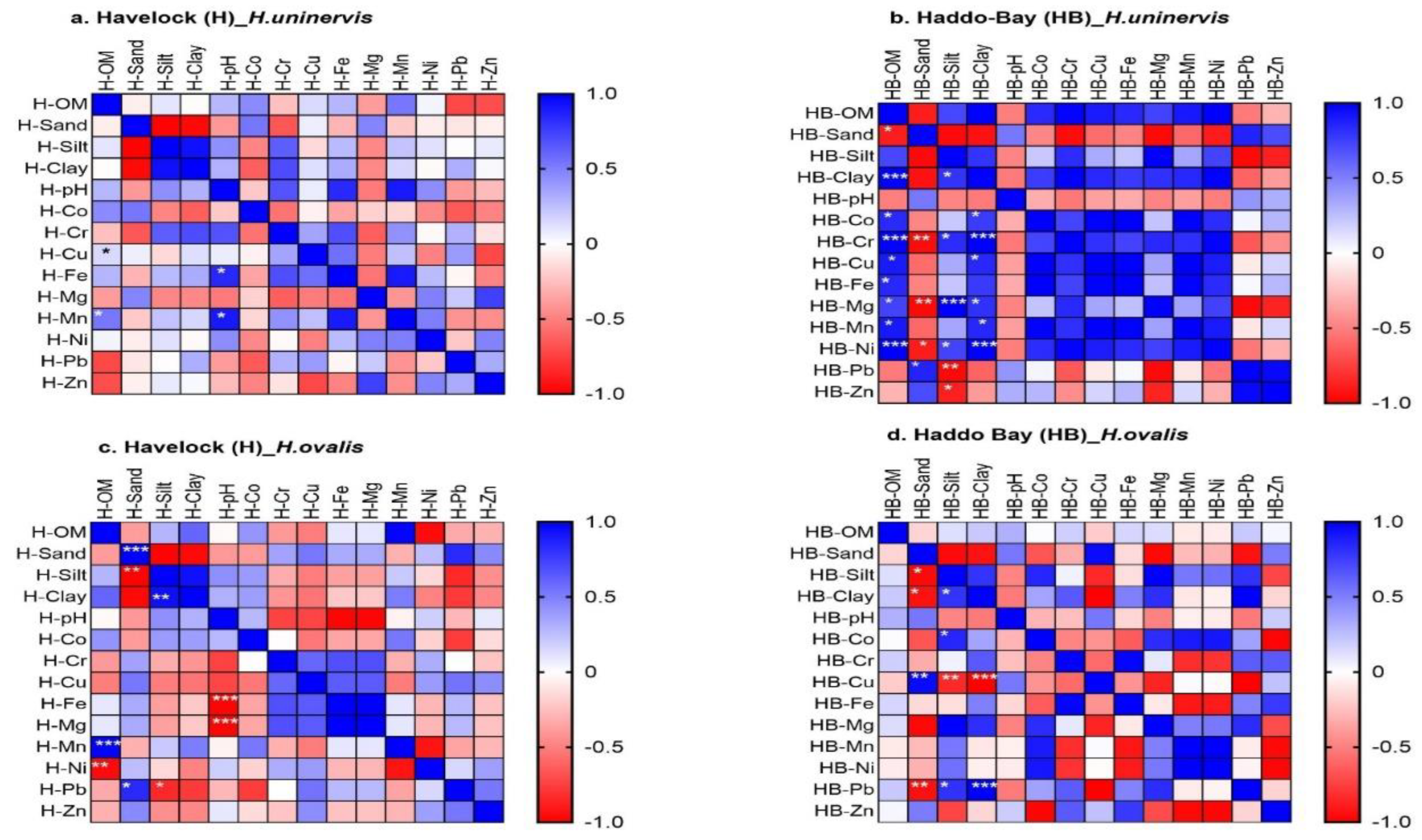

Table 1.
Percentage recovery of various trace metals in standards for plant biomass (ERM-CD281) and sediment (Hiss-1).
Table 1.
Percentage recovery of various trace metals in standards for plant biomass (ERM-CD281) and sediment (Hiss-1).
| Plant biomass | Sediment | |||||
|---|---|---|---|---|---|---|
| Trace metals | Certified value for ERM-CD281 (mg/Kg) | Recovered value (mg/Kg) | Percentage recovery (%) | Certified value for Hiss-1 (mg/Kg) | Recovered value (mg/Kg) | Percentage recovery (%) |
| Co | - | - | - | 0.65 | 0.60 | 93.77 |
| Cr | 24.8 | 23.01 | 92.79 | 30 | 27.16 | 90.54 |
| Cu | 10.2 | 9.71 | 95.26 | 2.29 | 2.11 | 92.55 |
| Fe | 180 | 172.85 | 96.03 | 2460 | 2342.90 | 95.24 |
| Mg | 1600 | 1441 | 90.10 | |||
| Mn | 82 | 74.17 | 90.46 | 66.10 | 61.60 | 93.20 |
| Ni | 15.2 | 14.33 | 94.28 | 2.16 | 2.02 | 93.84 |
| Pb | 1.67 | 1.57 | 94.38 | 3.13 | 3.06 | 97.91 |
| Zn | 30.5 | 30.06 | 98.57 | 4.94 | 4.89 | 99 |
Table 2.
Mean ± SD of abiotic parameters of surface water (pH, temperature and salinity) and sediment [organic matter (OM%) and sediment grain size] of seagrass ecosystems from the four locations of ANI. One way-ANOVA was used to test the statistical significance (p< 0.05) using location as fixed factors.
Table 2.
Mean ± SD of abiotic parameters of surface water (pH, temperature and salinity) and sediment [organic matter (OM%) and sediment grain size] of seagrass ecosystems from the four locations of ANI. One way-ANOVA was used to test the statistical significance (p< 0.05) using location as fixed factors.
| Variables | Locations | One-way ANOVA | ||||||
|---|---|---|---|---|---|---|---|---|
| Neil | Havelock | Burmanallah | Haddo Bay | DF | MS | F (DFn, DFd) |
P value | |
| pH | 8.10 ± 0.01 | 8.14 ± 0.05 | 8.14 ± 0.08 | 8.00 ± 0.08 | 3 | 0.025 | F (3,20) = 10.50 | = 0.002 |
| Temp.°C | 33.20 ± 1.30 | 34 ± 1.22 | 31.00 ±0.89 | 30 ± 0.46 | 3 | 17.87 | F (3,16) = 16.96 | <0.001 |
| Salinity (‰) | 32.67 ± 0.57 | 32 ± 1.01 | 33 ± 0.01 | 32.33 ± 1.15 | 3 | 0.55 | F (3,8) = 0.83 | = 0.512 |
| OM (%) | 38.17 ± 4.32 | 27.52 ± 7.86 | 38.70 ±5.07 | 25.44 ± 3.01 | 3 | 634.0 | F (3,66) = 18.21 | <0.0001 |
| Grain size (Sand%) | 43.70 ± 15.13 | 45.23 ± 9.50 | 33.89 ± 9.98 | 37.94 ± 13.63 | 3 | 202.8 | F (3,24) = 0.26 | = 0.85 |
| Grain size (Silt%) | 46.88 ± 12.11 | 39.33 ± 2.09 | 51.34 ± 3.16 | 49.60 ±9.37 | 3 | 232.2 | F (3,25) = 0.51 | =0.67 |
| Grain size (Clay%) | 9.41 ± 5.89 | 15.44 ± 8.46 | 14.76 ± 8.94 | 12.85 ± 6.25 | 3 | 65.28 | F (3,24) = 1.01 | =0.36 |
Table 3.
Comparison of ranges of trace metal levels in surface water and sediments of seagrass ecosystems of India adopted from Mishra and Farooq (2022), with our study. Below Detection Limit (BDL).
Table 3.
Comparison of ranges of trace metal levels in surface water and sediments of seagrass ecosystems of India adopted from Mishra and Farooq (2022), with our study. Below Detection Limit (BDL).
| Metals | Seagrass ecosystems | This study | References | ||
|---|---|---|---|---|---|
| Water (µg L-1) |
Sediment (mg kg-1) |
Water (µg L-1) |
Sediment (mg kg-1) |
||
| Co | - | 0.16–100 | BDL | 3.72 – 18.35 | Nobi et al., 2010; Thangaradjou et al., 2014; Sachithanandam et al., 2020 |
| Cr | 0.26-2.03 | 2.32–887 | BDL | 13.09 – 86.92 | Baby et al., 2017; Govindaswamy et al., 2011; Thangaradjou et al., 2014; Nobi et al., 2010; Jagtap and Untawale, 1984; Sachithanandam et al., 2020 |
| Cu | 0.11-1.02 | 1.58–130 | BDL | 2.97 – 71.44 | Baby et al., 2017; Govindaswamy et al., 2011; Thangaradjou et al., 2014; Nobi et al., 2010; Jagtap,1983; Jagtap and Untawale, 1984; Kumaresan et al., 1998 Sachithanandam et al., 2020 |
| Fe | 0.12-7.04 | 16.5–75500 | BDL | 1864 – 18400.99 | Jagtap, 1983; Jagtap and Untawale, 1984; Govindaswamy et al., 2011; Thangaradjou et al., 2014; Baby et al., 2017; Nobi et al., 2010; Kumarsen et al., 1998; Sachithanandam et al., 2020 |
| Mg | 0.16-18338 | 42–6204 | 25.49–26.34 | 2206.07 –8474.93 | Jagtap,1983; Jagtap and Untawale, 1984; Baby et al., 2017; Nobi et al., 2010 |
| Mn | 0.35-0.89 | 4–940 | BDL | 56.23 – 251.24 | Jagtap,1983; Govindaswamy et al., 2011; Thangaradjou et al., 2014; Baby et al., 2017; Nobi et al., 2010; Kumarsen et al., 1998; Sachithanandam et al., 2020; |
| Ni | 0.19-0.56 | 0.64–607 | BDL | 2.83 – 16.18 | Jagtap,1983; Jagtap and Untawale, 1984; Govindaswamy et al., 2011; Baby et al., 2017; Thangaradjou et al., 2014; Nobi et al., 2010; Sachithanandam et al., 2020 |
| Pb | 0.01-0.12 | 0.54–29 | BDL | 2.25 – 15.48 | Jagtap and Untawale, 1984; Baby et al., 2017; Sachithanandam et al., 2020 |
| Zn | 0.1-11.61 | 2–127.2 | BDL | 7.49 – 25.93 | Jagtap and Untawale, 1984; Govindaswamy et al., 2011; Baby et al., 2017; Thangaradjou et al., 2014; Nobi et al., 2010; Kumarsen et al., 1998; Sachithanandam et al., 2020 |
Table 4.
Minimum and maximum concentration of trace metals (mg Kg-1) in the tissues of various seagrass species of India adopted from Mishra and Farooq, (2022) and compared with this study. Palk Bay, TN; Gulf of Mannar (GOM), Lakshadweep Island (LK), Andaman and Nicobar Islands (ANI); Neil (N; ANI), Havelock (HV; ANI), Burmanallah (B; ANI), Haddo Bay (HB; ANI).
Table 4.
Minimum and maximum concentration of trace metals (mg Kg-1) in the tissues of various seagrass species of India adopted from Mishra and Farooq, (2022) and compared with this study. Palk Bay, TN; Gulf of Mannar (GOM), Lakshadweep Island (LK), Andaman and Nicobar Islands (ANI); Neil (N; ANI), Havelock (HV; ANI), Burmanallah (B; ANI), Haddo Bay (HB; ANI).
| Seagrass | Seagrass tissues; This study | References | |||
|---|---|---|---|---|---|
| Min (mg kg-1) | Max (mg kg-1) | Min (mg kg-1) | Max (mg kg-1) | ||
| Species | S. isoetifolium (LK) | H. uninervis (LK) | H. uninervis (H) | T. hemprichii (HB) | Nobi et al., 2010; Kannan et al., 2011; Thangaradjou et al., 2013; Arisekar et al., 2021 |
| Co | 0.16 | 11.21 | 7.74 | 97.85 | |
| Species | S. isoetifolium (GOM) | Seagrass (ANI) | H. uninervis (N) | H. ovalis (HB) | Nobi et al., 2010; Kannan et al., 2011; Thangaradjou et al., 2013; Immaculate et al., 2018; Arisekar et al., 2021; Pasumpon and Vasudevan, 2021 |
| Cr | 0.1 | 138.2 | 3.28 | 135.56 | |
| Species | S. isoetifolium (PB) | Seagrass (ANI) | H. ovalis (HB) | H. uninervis (HV) | Jagtap, 1983; Mathevan, 1990; Kannan et al., 1992; Nobi et al., 2010; Kannan et al., 2011; Govindaswamy et al., 2011; Gopinath et al., 2011; Sudharsan et al., 2012; Thangaradjou et al., 2013; Immaculate et al., 2018; Gopi et al., 2020; Arisekar et al., 2021; Pasumpon and Vasudevan, 2021 |
| Cu | 0.05 | 86.75 | 0.28 | 68.85 | |
| Species | S. isoetifolium (PB) | H. beccarii (GO) | E. acoroides (HV) | H. uninervis (HB) | Jagtap, 1983; Mathevan, 1990; Kannan et al., 1992; Nobi et al., 2010; Kannan et al., 2011; Govindaswamy et al., 2011; Gopinath et al., 2011; Sudharsan et al., 2012; Thangaradjou et al., 2013; Immaculate et al., 2018; Arisekar et al., 2021 |
| Fe | 0.22 | 32562 | 543 | 11655.49 | |
| Species | C. rotundata (GOM) | S. isoetifolium (LK) | H. ovalis (HV) | T. hemprichii (B) | Jagtap, 1983; Nobi et al., 2010; Thangaradjou et al., 2010; Kannan et al., 2011; Thangaradjou et al., 2013; Immaculate et al., 2018 |
| Mg | 91.54 | 80,050 | 1428.36 | 9190.79 | |
| Species | C. serrulata (PB) | H. ovalis (PB) | T. hemprichii (N) | T. hemprichii (B) | Jagtap, 1983; Mathevan, 1990; Kannan et al., 1992; Nobi et al., 2010; Govindaswamy et al., 2011; Kannan et al., 2011; Sudharsan et al., 2012; Thangaradjou et al., 2013; Arisekar et al., 2021; Pasumpon and Vasudevan, 2021 |
| Mn | 0.24 | 2250 | 15.41 | 244.10 | |
| Species | S. isoetifolium (PB) | H. decipens (LK) | C. rotundata (B) | H. uninervis (HB) | Jagtap, 1983; Nobi et al., 2010; Kannan et al., 2011; Thangaradjou et al., 2013; Sudharsan et al., 2012; Immaculate et al., 2018 |
| Ni | 0.1 | 19.49 | 0.54 | 39 | |
| Species | S. isoetifolium (LK) | H. uninervis (LK) | E. acoroides (HV) | H. uninervis (HB) | Nobi et al., 2010; Kannan et al., 2011; Sudharsan et al., 2012; Thangaradjou et al., 2013; Immaculate et al., 2018; Gopi et al., 2020; Arisekar et al., 2021; Pasumpon and Vasudevan, 2021 |
| Pb | 0.1 | 23.12 | 0.47 | 7.95 | |
| Species | S. isoetifolium (PB) | H. pinifolia (PB) | T. hemprichii (B) | H. beccarii (HB) | Mathevan, 1990; Kannan et al., 1992; Nobi et al., 2010; Kannan et al., 2011; Gopinath et al., 2011; Sudharsan et al., 2012; Govindaswamy et al., 2012; Thangaradjou et al., 2013; Immaculate et al., 2018; Gopi et al., 2020; Arisekar et al., 2021; Pasumpon and Vasudevan, 2021 |
| Zn | 0.15 | 69.17 | 0.60 | 70.4 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



