
Submitted:
30 January 2024
Posted:
31 January 2024
You are already at the latest version
Abstract
The presence of valuable bioactive compounds in European eel (Anguilla anguilla) skin was studied. For it, proximate and lipid class compositions and analysis of the fatty acid (FA) profile (individual FAs; FA groups, i.e., saturated, monounsaturated, and polyunsaturated; FA ratios, i.e., polyunsaturated/saturated, ω3/ω6) were determined and compared to the composition of the eel muscle. As a result, higher (p<0.05) levels of proteins (271.6 g·kg-1), lipids (38.0 g·kg-1), ash (27.7 g·kg-1) and ω6 FAs were observed in the skin tissue. Contrary, the muscle tissue showed higher (p<0.05) moisture, ω3 FA and ω3/ω6 values. Regarding lipid classes, a higher (p<0.05) proportion of phospholipids (111.1 g·kg-1 lipids), sterols (104.7 g·kg-1 lipids), α-tocopherol (274.0 mg·kg-1 lipids), and free fatty acids (43.6 g·kg-1 lipids) was observed in the skin tissue. No differences (p>0.05) between both tissues could be detected for triacylglycerol and FA group (saturated, monounsaturated, and polyunsaturated) values, as well as for the polyunsaturated/saturated ratio. It is concluded that eel skin, a by-product resulting from commercial processing, can be considered a valuable source to provide the food and pharmaceutical industries with useful value-added constituents such as proteins, lipids, ω3 FAs, phospholipids, and α-tocopherol.
Keywords:
Anguilla anguilla
; skin
; muscle
; proteins
; phospholipids
; sterols
; α-tocopherol
; ω3 fatty acids
; ω3/ω6 ratio.
1. Introduction
A wide range of studies have recognised the fish fatty acid (FA) profile and the lipid class composition as responsible for the health benefits of fish-enriched diets [1,2]. Among polyunsaturated FAs (PUFAs), eicosapentaenoic (EPA) and docosahexaenoic (DHA) acid consumption has been associated with low prevalence of several human diseases such as cardiovascular and neurodegenerative concerns [3,4]. According to their amphiphilic character, phospholipid (PL) compounds have shown to be valuable drug delivery systems for their high bioavailability and protecting effects on different kinds of diseases [5,6]. Regarding tocopherol compounds, fishery products have shown to represent an important source of this effective lipid-soluble antioxidant system [7,8].
As a result of fish processing, a large and considerable volume of undesired by-products is obtained, constituting an important environmental contamination source unless efforts for their recovery are attained [9,10] and their commercial value can be enhanced via extraction of valuable constituents [11,12]. Remarkably, such by-products have been reported to include a relevant presence of highly nutritional constituents. Although the traditional by-products have just included fish meal and fish oil, by-products are reported to also contain valuable and profitable components such as amino acids, enzymes, collagen, pigments, chitin, vitamins, minerals, and other bioactive compounds which may be beneficial for the human health [13,14].
The European eel (Anguilla anguilla), belonging to the Anguillidae family, is a commercially valuable species in Europe (namely, Spain, Portugal, Italy and The Netherlands) and Asia (namely, Japan, China, Korea and Taiwan). As a result of recent fishery overfishing on coasts and several biological concerns [15], a great attention has been accorded to its development as a farmed product [16,17]. Previous studies account for the analysis of the proximate composition and FA profile [18,19] and essential and toxic elements in the muscle [20]. Additionally, the evolution of the eel muscle quality has been studied during different processing conditions such as refrigeration [21,22], cooking [23] and canning [24].
However, previous research on European eel by-products can be considered very scarce. Thus, Sila et al. [25] carried out the extraction and characterisation of sulphated glycosaminoglycans, Taktak et al. [26] developed novel eco-friendly gelatine-based microfibers from eel skin for fish encapsulation, and Teng et al. [27] prepared peptide-chelated calcium from European eel bones. Regarding eel skin, it is considered a thick substrate that is commonly treated as a waste material during eel muscle processing and is normally converted into low-value products or discarded. The unemployment of this by-product not only results in the loss of a large amount of bioactive-rich components, but also leads to environmental problems.
The current study focused on the presence of valuable bioactive compounds in European eel (A. anguilla) skin. Determination of proximate and lipid class compositions and analysis of the FA profile, i.e., individual FAs, FA groups (saturated, STFAs; monounsaturated, MUFAs; PUFAs) and FA ratios (PUFAs/STFAs and ω3 FAs/ω6 FAs) were carried out. This composition study was performed in parallel with the muscle tissue.
2. Results and Discussion
2.1. Determination of the Proximate Composition
Values obtained for the proximate composition are included in Table 1. Moisture showed to be the most abundant constituent in both eel tissues, a higher (p < 0.05) value being detected in the muscle. Protein levels higher than 160 g·kg-1 tissue were observed in both tissues; notably, values obtained in the skin tissue (ca. 272 g·kg-1) were higher (p < 0.05) than in the muscle. The lipid content of the present eel samples depicted values included in the 28-38 g·kg-1 tissue range. As for protein content, lipid values were found higher (p < 0.05) in the skin tissue. Regarding the ash content, skin samples (27.7 g·kg-1) showed higher values (p < 0.05) than their counterparts corresponding to the muscle tissue (9.9 g·kg-1).
Protein content obtained in the present study is higher than the one found in the muscle of most commercial fish species [28,29,30]. Therefore, this by-product can be considered a protein-rich substrate. Regarding the current lipid content of European eel skin, this substrate maybe ranked as a medium-fat substrate [28] and could be considered a valuable source of lipid components.
To the best of our knowledge, no previous research has focused on the proximate composition of European eel (A. anguilla) skin. However, previous research provides information regarding the muscle tissue of this fish species. Thus, higher lipid contents (5.0%) than in the present study were obtained by Özogul et al. [18] in individuals caught in the North-eastern Mediterranean. Additionally, higher protein (19.2-19.6%), lipid (5.0-10.21%), and ash (1.23-1.50%) levels were detected in European eel (A. anguilla) muscle when studying freshwater individuals corresponding to several sizes [19].
Previous studies regarding the proximate composition of related eel species have been carried out. Thus, Park et al. [31] obtained a protein content included in the 11.0-40.9% range for Conger myriaster skin by employing green extracting technologies. Regarding the edible tissue, Oku et al. [32] carried out a comparative study on wild and cultured Japanese eel (Anguilla japonica) muscle; as a result, higher protein (19-0 and 18.9%, respectively) and lipid (11.6 and 13.1%, respectively) values than in the present study were obtained although moisture values were lower (69.1 and 67.4%, respectively). A higher protein content (ca. 18.1%) than in the current study was also detected in farmed and freshwater eel (Monopterus albus) muscle [33]. A varying lipid content (3.6-20.4%) resulting from the catching season and location was proved for freshwater eel (A. japonica) muscle [34] as well as by comparing A. japonica individuals in the initial and terminal stages of spawning migration (0.3-20.2%) [35].
2.2. Analysis of the FA Composition
The FA profile of both eel tissues is depicted in Table 2. From a qualitative point of view, both tissues revealed a similar composition. Thus, the two major FAs were C16:0 and C:18:1ω9. Additionally, relatively abundant FAs were: C18:0, C16:1ω7, C18:1ω7, C20:4ω6, C20:5ω3, C22:5ω3, and C22:6ω3. However, comparison of the FA profile of both tissues showed some differences from a quantitative point of view. Thus, a higher content (p < 0.05) on C17:0, C20:1ω9, C22:1ω9, and C20:2ω6 was detected in the skin tissue. Contrary, C14:0, C16:1ω7, C20:4ω6, C22:4ω6, C20:5ω3, C22:5ω3, and C22:6ω3 revealed a higher presence (p < 0.05) in the muscle samples.
Contents on STFA, MUFA and PUFA groups did not show significant differences (p > 0.05) by comparing both kinds of tissues (Figure 1). According to the individual FA profile, the MUFA group showed to be the most abundant (p < 0.05) in both tissues, while the PUFA group depicted the lowest (p < 0.05) presence. In agreement with this similar composition for the FA groups, no differences (p > 0.05) between both tissues could be outlined for the PUFA/STFA ratio (Figure 2).
Contents of total ω3 and total ω6 FAs revealed remarkable differences between both tissues (Figure 1). Thus, samples corresponding to the skin tissue showed a lower content (p < 0.05) of ω3 FAs but higher (p < 0.05) of ω6 FAs. As a result, a higher ω3/ω6 ratio (p < 0.05) was proved in samples corresponding to the muscle tissue (Figure 2).
A great interest has been attributed to the presence of ω3 PUFAs according to their beneficial health effects [36,37]. Based on epidemiological and clinical studies, EPA consumption has been related to circulatory, inflammatory, and coronary diseases [38], while DHA has been associated with prevention of neurodegenerative diseases, foetal development, and correct functioning of the nervous system and visual organs in the foetus [39]. Meantime, a relevant interest has also been given to the ω3/ω6 FA ratio [40,41]. Remarkably, recent studies have proved that Western populations do not include appropriate levels of ω3 FAs in the diet through natural dietary sources. In an attempt to avoid cardiovascular, neurological, and inflammatory concerns, the World Health Organization (WHO) recommends a higher ratio than 1:10 in the human diet [42]. Present results have shown lower levels for EPA, DHA, and ω3 FAs than those present in the muscle of marine fish and invertebrate species [28,29,30]. However, results can be considered notably higher than in non-aquatic food such as poultry and egg [43], milk [44] and meat [45]. Notably, ω3/ω6 ratio showed a higher value than 1/10 as recommended by the WHO [42].
No previous research is available regarding the FA composition of European eel (A. anguilla) skin. However, previous studies have focused on the FA composition of European eel muscle. According to the present results, a decreasing sequence for FA groups in muscle samples was described, i.e., MUFAs > STFAs > PUFAs, for individuals obtained in the North-eastern Mediterranean Sea [18], for wild and cultivated fish from Tunisian Mediterranean coasts [19] and for freshwater individuals from the River Ulla (Galicia, NW Spain) [46]. In such studies, and also in agreement with the current results, C18:1ω9 and C16:0 were the most abundant FAs.
When FA ratios are concerned, lower PUFA/STFA ratios than in the present case were obtained for European eel (A. anguilla) muscle from the Mediterranean Sea (0.37) [18] and from wild (0.46) and cultivated (0.52) individuals caught in the Tunisian Mediterranean coasts [46]. Regarding the ω3/ω6 ratio, higher values were detected in the muscle of freshwater individuals from the Ulla River (1.66-2.07) [19]. Additionally, Achouri et al. [46] found higher (3.28) and lower (1.31) ω3/ω6 ratio values in the muscle of cultivated and wild individuals, respectively, obtained from Tunisian Mediterranean coasts.
Previous studies also account for the FA composition of related eel species. Thus, the same FA group distribution (MUFAs > STFAs > PUFAs) as in the current study was detected in skin from C. myriaster eel from South Korea [31], in freshwater eel A. japonica muscle (Lee et al., 2020), in wild and cultivated Japanese eel (A. japonica) muscle [32], and in Japanese freshwater eel (A. japonica) muscle [35]. Regarding FA ratios, a higher ω3/ω6 ratio (4.48-5.41) than in the present work was detected by Park et al. [31] in C. myriaster eel skin from South Korea. Contrary, Lee et al. [34] obtained a similar ω3/ω6 ratio (0.90-1.67) for A. japonica muscle than in the current study.
2.3. Lipid Class Composition
Results obtained for the lipid class composition are described in Table 3. Triacylglycerols (TAGs) showed to be the most abundant lipid class in both tissues, values being included in the 400-412 g·kg-1 lipid range. A higher average content was observed in the muscle tissue; however, differences were not found significant (p > 0.05) between both tissues.
Free fatty acids (FFAs), compounds resulting from the hydrolysis of higher-molecular weight compounds (i.e., TAGs and PLs) [47,48,49], provided values included in the 30-44 g·kg-1 lipids; notably, higher values (p < 0.05) were detected in the skin tissue than in its counterpart edible fraction.
A remarkable presence of structured lipid classes (PLs and sterols, STs) was detected in the skin tissue (ca. 111 and 105 g·kg-1 lipids, respectively). Values were higher (p < 0.05) than those obtained in their counterparts corresponding to the muscle tissue, especially for the ST compounds.
The analysis of the tocopherol composition of the current substrates revealed that the only tocopherol compound present was α-tocopherol. Its content was found notably higher (p < 0.05) in the skin tissue (274 mg·kg-1 lipids) than in the muscle (178 mg·kg-1 lipids).
Table 3 also indicates the lipid class content expressed on tissue basis. As for the previously mentioned results on lipid basis, higher (p < 0.05) FFA, PL, ST, and α-tocopherol values were detected in the skin tissue than in its counterpart muscle substrate. Notably, a higher average value of TAGs was observed in skin samples although differences with their corresponding muscle tissues were not found significant (p > 0.05).
Based on their amphiphilic character, remarkable attention has been given to PL compounds present in fish [5,6]. Thus, remarkable functions of PL compounds have been related to food production and pharmaceutical industries [50,51], these including valuable antioxidant properties during food processing [52,53]. PL contents observed in the current eel by-product can be considered as valuable and corresponding to a medium-fat fish substrate [28,54] and lower than in a lean fish species [29,30]. Based on their important role as lipid-soluble chain-breaking antioxidants, tocopherol compounds have received a great attention from marine technologists for their important role as lipophilic antioxidants [7,8]. Among them, α- tocopherol has been found to be the most abundant in fish species [28]. As for PL content, α-tocopherol levels found in European eel skin can be considered as valuable and corresponding to a medium-fat substrate [28] and lower than in the case of a lean fish species [29,30].
Previous research regarding the analysis of lipid classes of European eel (A. anguilla) samples (skin or muscle) can be considered very scarce. According to the present results, TAGs showed to be the most abundant lipid class in the muscle tissue from pre-migrant and migrant eel individuals [55]. In such study, phosphatidylcholine (PC) showed to be the most abundant PL class.
Previous research has focused on the lipid class analysis of related eel species. Thus, Park et al. [31] detected α-, β+γ-, and δ-tocopherol in eel C. myriaster skin from South Korea; as in the present case, α-tocopherol showed to be the most abundant, with a content included in the 31-100 mg/100 g skin range. Saito et al. [35] compared the lipid class composition of initial and terminal stages of spawining migration of wild Japanese freshwater eel (A. japonica) muscle; as a result, TAGs were the major component in the initial–phase eels, but presented a remarkable content decrease in individuals corresponding to the terminal phase. A comparative study of the lipid class profile in wild and cultivated individuals of Japanese eel (A. japonica) was carried out by Oku et al. [22]; both in wild and cultivated individuals, TAGs showed to be the most abundant lipid class of muscle (67.9-68.2%), other lipid classes determined being sterylesters (9.5-10.2%), FFAs (9.9-11.2%), STs (4.5%), PC (2.3-2.4%), and phosphatidylethanolamine (1.2-1.5%).
3. Materials and Methods
3.1. Fish Material and Sampling
Fresh European eel (A. anguila) (weight: 53-83 g; length: 32-39 cm) were purchased at a local market (Mariscos Vivos del Grove, Quintela de Canedo, Ourense, Spain). The eels were transported to the laboratory in insulated boxes on ice (0-1 ºC). The fish (60 individuals) were distributed into four groups (fifteen individuals per group), which were considered independently for the statistical analysis (n = 4). As a first processing step, the fish were eviscerated and washed with running water. In each individual fish, the skin and muscle were excised and considered separately. Inside each of the four groups, portions corresponding to the same tissue (skin or muscle) were pooled together and subjected to the different chemical analyses.
Solvents and chemical reagents used in this study were of reagent grade (Merck, Darmstadt, Germany). In the case of tocopherol analysis, solvents used were liquid chromatographic grade.
3.2. Proximate Composition Analysis
Moisture content was determined as the weight difference in the homogenised tissue (1-2 g) before and after 4 h at 105 ºC [56]. Results were calculated as g water·kg-1 tissue.
Protein content was measured using the Kjeldahl method [56] with a conversion factor of 6.25. Results were calculated as g protein·kg-1 tissue.
The total lipid fraction was extracted using the Bligh and Dyer [57] method, which employs a single-phase solubilisation of the lipids using a chloroform-methanol (1:1) mixture. Results were calculated as g total lipids·kg-1 tissue.
Ash content was measured according to the AOAC [56] method by heating the fish tissue at 550 ºC. Results were calculated as g ash·kg-1 tissue.
3.3. FA Analysis
Lipid extracts were converted into fatty acid methyl esters (FAMEs) by using acetylchloride in methanol and then analysed by gas-liquid chromatography (Perkin-Elmer 8700 chromatograph, Madrid, Spain) according to an established procedure [58]. For it, a fused silica capillary column SP-2330 (0.25 mm i.d. x 30 m, Supelco, Inc., Bellefonte, PA, USA) was employed and the temperature program was as follows: increased from 145 to 190 ºC at 1.0 ºC min-1 and from 190 ºC to 210 ºC at 5.0 ºC min-1; held for 13.5 min at 210 ºC. The carrier gas was nitrogen at 10 psig and detection was performed with a flame ionisation detector at 250 ºC. A programmed temperature vaporiser injector was employed in the split mode (150:1) and was heated from 45 to 275 ºC at 15 ºC min-1.
Peaks corresponding to FAMEs were identified by comparing their retention times with those of standard mixtures (Qualmix Fish, Larodan, Malmo, Sweden; FAME Mix, Supelco, Inc.). Peak areas were automatically integrated; C19:0 fatty acid was used as internal standard for quantitative purposes. Content of each FA was expressed as g·100 g-1 total FAs.
Results concerning FA groups (STFAs, MUFAs, PUFAs; ω3 and ω6 FAs) and FA ratios (total ω3 FAs/total ω6 FAs and total PUFAs/total STFAs) were calculated taking into account the results obtained in individual FAs.
3.4. Analysis of Lipid Classes
To measure the TAG content, the total lipid extracts were first purified on 20 x 20 cm thin-layer chromatography plates coated with a 0.5 mm-layer of silica gel G from Merck (Darmstadt, Germany) using a mixture of hexane-ethyl ether-acetic acid (90/10/1, v/v/v; two developments) as eluent [59]. Once the TAG fraction was purified, the method of Vioque and Holman [60] was used to measure the ester linkage content according to the conversion of the esters into hydroxamic acids and subsequent complexion with Fe (III). Results were calculated as g TAGs·kg-1 lipids and as g TAGs·kg-1 tissue.
FFA content of the total lipid extracts was determined following the Lowry and Tinsley [61] method, which is based on the formation of a complex with cupric acetate-pyridine. In this study, benzene was replaced by toluene as organic solvent. Results were calculated as g FFAs·kg-1 lipids and as g FFAs·kg-1 tissue.
PLs were quantified by measuring the organic phosphorus in the total lipid extracts according to the Raheja et al. [62] method, which is based on a complex formation with ammonium molybdate. Results were calculated as g PLs·kg-1 lipids and as g PLs·kg-1 tissue.
STs were determined on total lipid extracts by the method of Huang et al. [63] based on the Liebermann-Buchardt reaction. Results were calculated as g STs·kg-1 lipids and as g STs·kg-1 tissue.
The content of tocopherol compounds was determined in both tissues according to the method of Cabrini et al. [64] with some modifications. For this purpose, a lipid fraction was carried out to dryness under nitrogen flux, dissolved in isopropanol and analysed by HPLC (ODS column, 15 cm × 0.46 cm i.d.). The column was fluxed with methanol for 2 min; then, a gradient from 0 to 50% of isopropanol in 10 min was applied. A 1.5-mL·min-1 flow rate was employed and detection was carried out at 280 nm. The possible presence of α-, β-, γ-, and δ-tocopherol molecules was checked. For quantitative purposes, the content of each tocopherol compound present in the lipid extract was calculated with calibration curves prepared from the corresponding commercial tocopherol molecule and calculated as mg·kg-1 lipids and mg·kg-1 tissue.
3.5. Statistical Analysis
Data (n = 4) obtained from the different chemical analyses (proximate composition, individual FAs, FA groups and ratios, and lipid classes) were subjected to the ANOVA method to investigate differences between both tissues, i.e., skin and muscle (Statistica version 6.0, 200; Statsoft Inc., Chicago, Il, USA). Comparison of means was performed using a least-squares difference (LSD) method. The 95% confidence intervals of each chemical parameter were calculated; for it, the standard deviation of each sample and the number of replicates were considered.
4. Conclusions
The present study provides a first approach on the chemical composition of European eel (A. anguilla) skin. In it, a comparative study between skin and muscle focused on the presence of valuable bioactive compounds was carried out. Thus, higher (p < 0.05) levels of proteins (271.6 g·kg-1), lipids (38.0 g·kg-1), ash (27.7 g·kg-1) and ω6 FAs were observed in the skin tissues. Contrary, the muscle tissue showed higher (p < 0.05) moisture, ω3 FA and ω3/ω6 values. Regarding lipid classes, higher proportions of PLs (111.1 g·kg-1 lipids), STs (104.7 g·kg-1 lipids), α-tocopherol (274.0 mg·kg-1 lipids), and FFAs (43.6 g·kg-1 lipids) were observed in the skin tissue. No differences (p > 0.05) between both tissues could be detected for TAG and FA group (STFAs, MUFAs and PUFAs) values, as well as for the total PUFAs/total STFAs ratio.
It is concluded that eel skin, a by-product resulting from the commercial processing, can be considered a valuable source to provide the food and pharmaceutical industries with useful value-added constituents such as proteins, lipids, ω3 FAs, PLs, and α-tocopherol. The study agrees with the current search for alternative sources of healthy and nutritious compounds from waste substrates. As for edible parts of fish, convenient handling and storage during the skin processing ought to be carried out to avoid the development of damage mechanisms such as autolysis, microbial activity and lipid oxidation.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, S.M. and S.P.A.; methodology, A.B. and M.T.; data curation, A.B. and M.T.; writing—original draft preparation, S.P.A.; writing—review and editing, S.M. and S.P.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Tilami, S.K.; Sampels, S. Nutritional Value of Fish: lipids, proteins, vitamins, and minerals. Rev. Fish. Sci. 2018, 26, 242–253. [Google Scholar] [CrossRef]
- Xu, H.; Turchini, G.M.; Francis, D.S.; Liang, M.; Mock, T.S.; Rombenso, A.; Ai, Q. Are fish what they eat? A fatty acid’s perspective. Prog. Lipid Res. 2020, 80, 101064. [Google Scholar] [CrossRef] [PubMed]
- Minihane, A.; Armah, C.; Miles, E.; Madden, J.; Clark, A.; Caslake, M.; Calder, P. Consumption of fish oil providing amounts of eicosapentaenoic acid and docosahexaenoic acid that can be obtained from the diet reduces blood pressure in adults with systolic hypertension: A retrospective analysis. J. Nutr. 2016, 146, 516–523. [Google Scholar] [CrossRef] [PubMed]
- Schunck, W.; Konkel, A.; Fischer, R.; Weylandt, K. Therapeutic potential of omega-3 fatty acid-derived epoxy eicosanoids in cardiovascular and inflammatory diseases. Pharmacol. Ther. 2018, 183, 177–204. [Google Scholar] [CrossRef] [PubMed]
- Küllenberg, D.; Taylor, L.A.; Schneider, M.; Massing, U. Health effects of dietary phospholipids. Lipids Health Dis. 2012, 11, Article 3. [Google Scholar] [CrossRef]
- Li, J.; Wang, X.; Zhang, T.; Huang, Z.; Luo, X.; Deng, Y. A review on phospholipids and their main applications in drug delivery systems. Asian J. Pharm. Sci. 2015, 10, 81–98. [Google Scholar] [CrossRef]
- Kalogeropoulos, N.; Chiou, A. Antioxidants. In Handbook of Seafood and Seafood Products Analysis; Nollet, L.M., Toldrá, F., Eds.; CRC Press, Taylor and Francis Group: Boca Raton, FL, USA, 2010; pp. 309–326. [Google Scholar] [CrossRef]
- Kim, Y.N. Vitamins. In Handbook of Seafood and Seafood Products Analysis; Nollet, L.M., Toldrá, F., Eds.; CRC Press, Francis and Taylor Group: Boca Raton, FL, USA, 2010; pp. 327–350. [Google Scholar] [CrossRef]
- Arvanitoyannis, I.S.; Kassaveti, A. Fish industry waste: treatments, environmental impacts, current and potential uses. Int. J. Food Sci. Technol. 2008, 43, 726–745. [Google Scholar] [CrossRef]
- Özyurt, G.; Özkütük, S. Advances in discard and by-product processing. In Innovative Technologies in Seafood Processing; Özogul, Y., Ed.; CRC Press, Taylor and Francis Group: Boca Raton, FL, USA, 2020; pp. 323–350. [Google Scholar] [CrossRef]
- Rustad, T.; Storro, I.; Slizyte, R. Possibilities for the utilisation of marine by-products. Int. J. Food Sci. Technol. 2011, 46, 2001–2014. [Google Scholar] [CrossRef]
- Olsen, R.L.; Toppe, J.; Karunasagar, I. Challenges and realistic opportunities in the use of by-products from processing of fish and shellfish. Trends Food Sci. Technol. 2014, 36, 144–152. [Google Scholar] [CrossRef]
- Atef, M.; Ojagh, M. Health benefits and food applications of bioactive compounds from fish byproducts: A review. J. Funct. Foods 2017, 35, 673–681. [Google Scholar] [CrossRef]
- Vázquez, J.A.; Meduiña, A.; Durán, A.I.; Nogueira, M.; Fernández-Compás, A.; Pérez-Martín, R.I.; Rodríguez-Amado, I. Production of valuable compounds and bioactive metabolites from by-products of fish discards using chemical processing, enzymatic hydrolysis, and bacterial fermentation. Mar. Drugs 2019, 17, 139. [Google Scholar] [CrossRef]
- Goymer, A.; Steele, K.; Jenkins, F.; Burgess, G.; Andrews, L.; Baumgartner, N.; Gubili, C.; Griffiths, A.M. For R-eel ?! Investigating international sales of critically endangered species in freshwater eel products with DNA barcoding. Food Cont. 2023, 150, 109752. [Google Scholar] [CrossRef]
- Heinsbroek, L.T.N.; Støttrup, J.G.; Jacobsen, C.; Corraze, G.; Kraiem, M.M.; Holst, L.K.; Tomkiewicz, J.; Kaushik, S.J. A review on broodstock nutrition of marine pelagic spawners: the curious case of the freshwater eels (Anguilla spp.). Aquacult. Nutr. 2013, 19, 1–24. [Google Scholar] [CrossRef]
- Støttrup, J.G.; Tomkiewicz, J.; Jacobsen, C.; Butts, I.A.E.; Holst, L.K.; Krüger-Johnsen, M.; Graver, C.; Lauesen, P.; Fontagne-Dicharry, S.; Heinsbroek, L.T.N.; Geneviève, C.; Kaushik, S. Development of a broodstock diet to improve developmental competence of embryos in European eel, Anguilla anguilla. Aquacult. Nutr. 2016, 22, 725–737. [Google Scholar] [CrossRef]
- Özogul, Y.; Ucar, Y.; Takadaş, F.; Durmus, M.; Köşker, A.R.; Polat, A. Comparison of green and conventional extraction methods on lipid yield and fatty acid profiles of fish species. Eur. J. Lipid Sci. Technol. 2018, 120, 1800107. [Google Scholar] [CrossRef]
- Gómez-Limia, L.; Cobas, N.; Martínez, S. Proximate composition, fatty acid profile and total amino acid contents in samples of the European eel (Anguilla anguilla) of different weights. Int. J. Gastr. Food Sci. 2021, 25, 100364. [Google Scholar] [CrossRef]
- Rudovica, V.; Bartkevics, V. Chemical elements in the muscle tissues of European eel (Anguilla anguilla) from selected lakes in Latvia. Environm. Monit. Assess. 2015, 87, 608. [Google Scholar] [CrossRef] [PubMed]
- Kucukgulmez, A.; Yanar, Y.; Gerçek, G.; Gülnaz, O.; Celik, M. Effects of chitosan on color, sensory and microbiological properties of European eel (Anguilla anguilla) fillets during refrigerated storage. J. Food Proc. Preserv. 2013, 37, 766–771. [Google Scholar] [CrossRef]
- Özogul, I.; Polat, A.; Özogul, Y.; Boga, E.K.; Özogul, F.; Ayas, D. Effects of laurel and myrtle extracts on the sensory, chemical and microbiological properties of vacuum-packed and refrigerated European eel (Anguilla anguilla) fillets. Int. J. Food Sci. Technol. 2014, 49, 847–853. [Google Scholar] [CrossRef]
- Ersoy, B. Effects of cooking methods on the proximate, mineral and fatty acid composition of European eel (Anguilla anguilla). Int. J. Food Sci. Technol. 2011, 46, 522–527. [Google Scholar] [CrossRef]
- Gómez-Limia, L.; Carballo, J.; Rodríguez-González, M.; Martínez, S. Impact of the filling medium on the colour and sensory characteristics of canned European eels (Anguilla anguilla L.). Foods 2022, 11, 1115. [Google Scholar] [CrossRef]
- Sila, A.; Bougatef, H.; Capitani, F.; Krichen, F.; Mantovani, V.; Ben Amor, I.; Galeotti, F.; Maccari, F.; Nedjar, N.; Volpi, N.; Bougatef, A. Studies on European eel skin sulfated glycosaminoglycans: recovery, structural characterization and anticoagulant activity. Int. J. Biol. Macrom. 2018, 115, 891–899. [Google Scholar] [CrossRef]
- Taktak, W.; Nasri, R.; López-Rubio, A.; Chentir, I.; Gómez-Mascaraque, L.G.; Boughriba, S.; Nasri, M.; Karra-Chaabouni, M. Design and characterization of novel ecofriendly European fish eel gelatin-based electrospun microfibers applied for fish oil encapsulation. Process Biochem. 2021, 106, 10–19. [Google Scholar] [CrossRef]
- Teng, H.; Qian, Y.; Fan, X.; Cao, H.; Tian, Y.; Chen, L. Nutritional properties of Europen eel (Anguilla anguilla) bone peptide-calcium and its apoptosis effect on Caco-2 cells. Food Sci and Human Wellness 2022, 11, 1482–1490. [Google Scholar] [CrossRef]
- Piclet, G. Le poisson aliment. Composition-Intérêt nutritionnel. Cah. Nutr. Diét. 1987, XXII, 317–335. [Google Scholar]
- Prego, R.; Pazos, M.; Medina, I.; Aubourg, S.P. Comparative chemical composition of different muscle zones in angler (Lophius piscatorius). J. Food Compos. Anal. 2012, 28, 81–87. [Google Scholar] [CrossRef]
- Barbosa, R.G.; Trigo, M.; Prego, R.; Fett, R.; Aubourg, S.P. The chemical composition of different edible locations (central and edge muscles) of flat fish (Lepidorhombus whiffiagonis). Int. J. Food Sci. Technol. 2018, 53, 271–281. [Google Scholar] [CrossRef]
- Park, J.S.; Chandra Roy, V.; Kim, S.Y.; Lee, S.C.; Chun, B.S. Extraction of edible oils and amino acids from eel by-products using clean compressed solvents: an approach of complete valorization. Food Chem. 2022, 38, 132949. [Google Scholar] [CrossRef]
- Oku, T.; Sugawara, A.; Choudhury, M.; Komatsu, M.; Yamada, S.; Ando, S. Lipid and fatty acid compositions differentiate between wild and cultured Japanese eel (Anguilla japonica). Food Chem. 2009, 115, 436–440. [Google Scholar] [CrossRef]
- Nurfaidah; Metusalach; Sukarno; Mahendradatta, M. Protein and albumin contents in several freshwater fish species of Makassar, South Sulawesi, Indonesia. Int. Food Res. J. 2021, 28, 745–751. [CrossRef]
- Lee, K.; Kim, Y.J.; Hong, Y.K.; Song, M.Y.; Lee, W.O.; Hwang, K.T. Lipid content and fatty acid composition of freshwater eels Anguilla japonica caught in different seasons and locations in South Korea. Fish. Sci. 2020, 86, 573–580. [Google Scholar] [CrossRef]
- Saito, H.; Kurogi, H.; Chow, S.; Mochioka, N. Variation of lipids and fatty acids of the Japanese eel, Anguilla japonica, during spawning migration. J. Oleo Sci. 2015, 64, 603–616. [Google Scholar] [CrossRef]
- Magalhães, J.P.; Müller, M.; Rainger, G.; Steegenga, W. Fish oil supplements, longevity and aging. Aging 2016, 8, 1578–1582. [Google Scholar] [CrossRef]
- Devassy, J.G.; Leng, S.; Gabbs, M.; Monirujjaman, M.; Aukema, H.M. Omega-3 polyunsaturated fatty acids and oxylipins in neuroinflammation and management of Alzheimer disease. Adv. Nutr. 2016, 7, 905–916. [Google Scholar] [CrossRef]
- Swanson, S.; Block, R.; Mousa, S. Omega-3 fatty acids EPA and DHA: Health benefits throughout life. Adv. Nutr. 2012, 3, 1–7. [Google Scholar] [CrossRef]
- Ofosu, F.K.; Daliri, E.B.M.; Lee, B.H.; Yu, X. Current trends and future perspectives on omega-3 fatty acids. Res. J. Biol. 2017, 5, 11–20. [Google Scholar]
- Uauy, R.; Valenzuela, A. Marine oils: The health benefits of n-3 fatty acids. Nutrition 2000, 16, 680–684. [Google Scholar] [CrossRef]
- Šimat, V.; Vlahović, J.; Soldo, B.; Mekinić, I.G.; Čagalj, M.; Hamed, I.; Skroza, D. Production and characterization of crude oils from seafood processing by-products. Food Biosci. 2020, 33, 100484. [Google Scholar] [CrossRef]
- Kumari, P.; Kumar, M.; Reddy, C.R.; Jha, B. Algal lipids, fatty acids and sterols. In Functional Ingredients from Algae for Foods and Nutraceuticals; Domínguez, H., Ed.; Woodhead Publishing: Cambridge, UK, 2013; pp. 87–134. [Google Scholar]
- Cantor, A.H.; Decker, E.A.; Collins, V.P. Fatty acids in poultry and egg products. In Fatty Acids in Foods and their Health Implications; Chow, C.K., Ed.; 3rd Ed., CRC Press, Boca Raton, FL, USA, 2008; pp. 127–154. [CrossRef]
- Palmquist, D.L.; Jensen, R.G. Fatty acids in milk fat. In Fatty Acids in Foods and their Health Implications; Chow, C.K., Ed.; 3rd Ed., CRC Press, Boca Raton, FL, USA, 2008; pp. 109–125.
- Wood, J.D.; Enser, M.; Richardson, R.I.; Whittington, F.M. Fatty acids in meat and meat products. In Fatty Acids in Foods and their Health Implications; Chow, C.K., Ed.; 3rd Ed., CRC Press, Boca Raton, FL, USA, 2008; pp. 87–107. [CrossRef]
- Achouri, A.; Kharrat, N.; Smichi, N.; Miled, N.; Gargouri, Y.; Fendri, A. Nutritional properties, oxidative stability, and in vitro digestibility of oils extracted from muscles of wild and breeding eels (Anguilla anguilla). J. Food Process. Preserv. 2018, 42, e13519. [Google Scholar] [CrossRef]
- Medina, I.; Sacchi, R.; Aubourg, S.P. A 13C-NMR study of lipid alterations during fish canning: Effect of filling medium. J. Sci. Food Agric. 1995, 69, 445–450. [Google Scholar] [CrossRef]
- Aubourg, S.P; Quitral, V.; Larraín, M.A.; Rodríguez, A.; Gómez, J.; Maier, L.; Vinagre, J. Autolytic degradation and microbiological activity in farmed Coho salmon (Oncorhynchus kisutch) during chilled storage. Food Chem. 2007, 104, 369–375. [Google Scholar] [CrossRef]
- Vázquez, M.; Torres, J.A.; Gallardo, J.M.; Saraiva, J.A.; Aubourg, S.P. Lipid hydrolysis and oxidation development in frozen mackerel (Scomber scombrus): Effect of a high hydrostatic pressure pre-treatment. Innov. Food Sci. Emer. Technol. 2013, 18, 24–30. [Google Scholar] [CrossRef]
- Takahashi, K.; Inoue, Y. Marine by-product phospholipids as booster of medicinal compounds. Adv. Food Nutr. Res. 2012, 65, 31–46. [Google Scholar] [PubMed]
- Köhler, A.; Sarkinnen, E.; Tapola, N.; Niskanen, T.; Bruheim, I. Bioavailability of fatty acids from krill oil, krill meal and fish oil in healthy subjects–a randomized, single-dose, cross-over trial. Lipids Health Dis. 2015, 14, 19. [Google Scholar] [CrossRef] [PubMed]
- Bandarra, N.M.; Campos, R.M.; Batista, I.; Nunes, M.L.; Empis, J.M. Antioxidant synergy of alpha-tocopherol and phospholipids. J. Am Oil Chem. Soc. 1999, 76, 905–913. [Google Scholar] [CrossRef]
- Hidalgo, F.J.; Leon, M.M.; Zamora, R. Antioxidative activity of amino phospholipids and phospholipid/amino acid mixtures in edible oils as determined by the rancimat method. J. Agric. Food Chem. 2006, 54, 5461–5467. [Google Scholar] [CrossRef]
- Medina, I.; Aubourg, S.P.; Pérez-Martín, R. Composition of phospholipids of white muscle of six tuna species. Lipids 1995, 30, 1127–1135. [Google Scholar] [CrossRef] [PubMed]
- Van Ginneken, V.; Hekman, M.; Verheij, E. The lipid composition and biochemistry of the migrating European eel (Anguilla anguilla L.): A LCMS-study following a lipidomics based systems biology approach. Adv. Biochem Biotechnol. 2018, 3, 165. [Google Scholar]
- AOAC. Official Methods for Analysis of the Association of Analytical Chemistry, 15th ed.; Association of Official Chemists, Inc.: Arlington, VA, USA, 1990; pp. 931–937. [Google Scholar]
- Bligh, E.; Dyer, W. A rapid method of total extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Aubourg, S.P.; Medina, I.; Pérez-Martín, R. Polyunsaturated fatty acids in tuna phospholipids: Distribution in the sn-2 location and changes during cooking. J. Agric. Food Chem. 1996, 44, 585–589. [Google Scholar] [CrossRef]
- Álvarez, V.; Medina, I.; Prego, R.; Aubourg, S.P. Lipid and mineral distribution in different zones of farmed and wild blackspot seabream (Pagellus bogaraveo). Eur. J. Lipid Sci. Technol. 2009, 111, 957–966. [Google Scholar] [CrossRef]
- Vioque, E.; Holman, R. Quantitative estimation of esters by thin-layer chromatography. J. Am. Oil Chem. Soc. 1962, 39, 63–66. [Google Scholar] [CrossRef]
- Lowry, R.; Tinsley, I. Rapid colorimetric determination of free fatty acids. J. Am. Oil Chem. Soc. 1976, 53, 470–472. [Google Scholar] [CrossRef]
- Raheja, R.; Kaur, C.; Singh, A.; Bhatia, A. New colorimetric method for the quantitative determination of phospholipids without acid digestion. J. Lipid Res. 1973, 14, 695–697. [Google Scholar] [CrossRef]
- Huang, T.; Chen, C.; Wefler, V.; Raftery, A. A stable reagent for the Liebermann-Buchardt reaction. Anal. Chem. 1961, 33, 1405–1407. [Google Scholar] [CrossRef]
- Cabrini, L.; Landi, L.; Stefanelli, C.; Barzanti, V.; Sechi, A. Extraction of lipid and lipophilic antioxidants from fish tissues: A comparison among different methods. Comp. Biochem. Physiol. Biochem. Molec. Biol. 1992, 101, 383–386. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Profile of fatty acid (FA) groups of eel skin and muscle (g·100 g-1 total FAs). Average values of four (n = 4) replicates; standard deviations are indicated by bars. For each FA group, different letters (a,b) denote significant differences (p < 0.05). Abbreviations: STFAs (saturated fatty acids), MUFAs (monounsaturated fatty acids), and PUFAs (polyunsaturated fatty acids).
Figure 1.
Profile of fatty acid (FA) groups of eel skin and muscle (g·100 g-1 total FAs). Average values of four (n = 4) replicates; standard deviations are indicated by bars. For each FA group, different letters (a,b) denote significant differences (p < 0.05). Abbreviations: STFAs (saturated fatty acids), MUFAs (monounsaturated fatty acids), and PUFAs (polyunsaturated fatty acids).
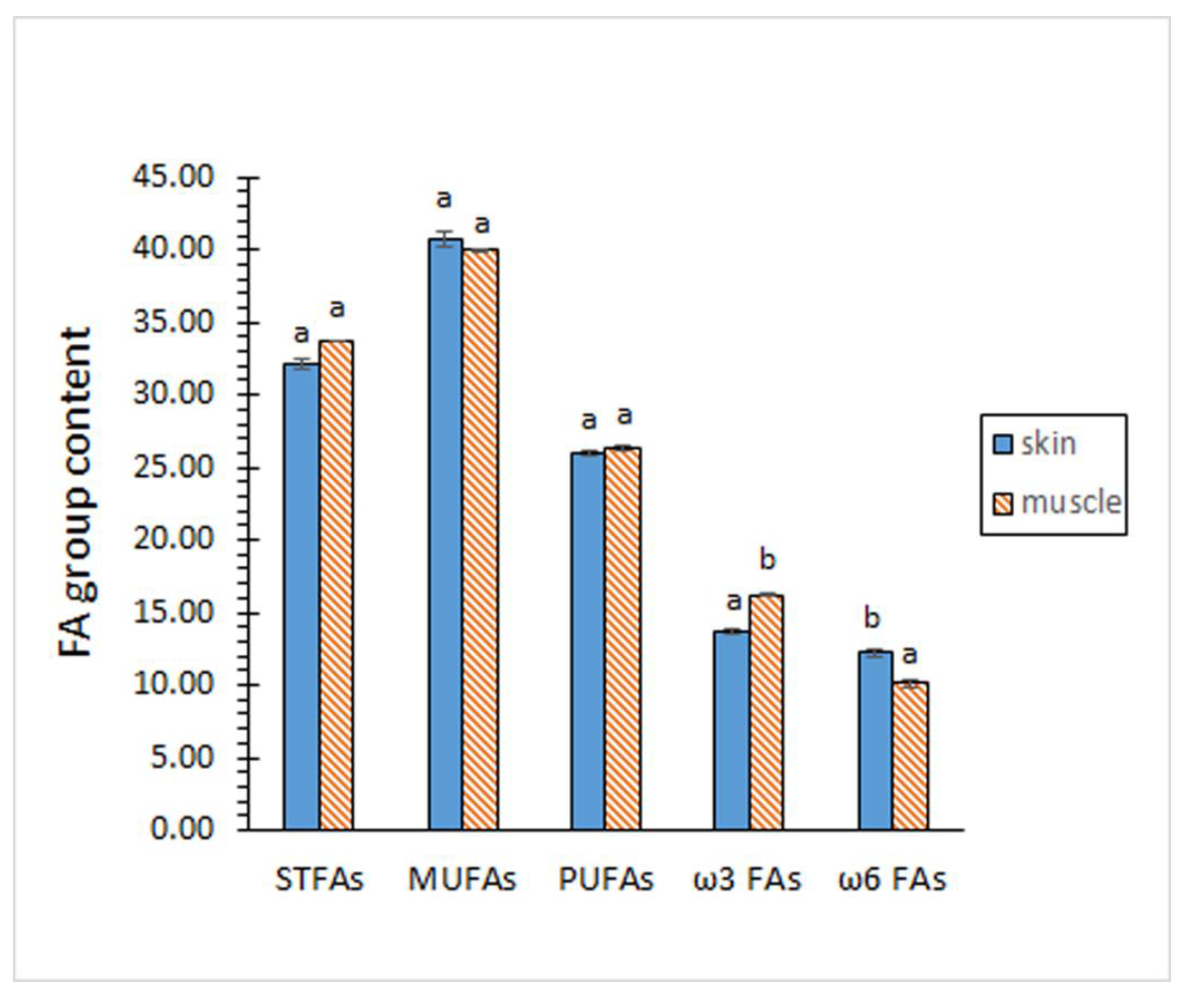
Figure 2.
Fatty acid (FA) ratios of the eel skin and muscle. Average values of four (n = 4) replicates; standard deviations are indicated by bars. For each FA ratio, different letters (a,b) denote significant differences (p < 0.05). Abbreviations as expressed in Figure 1.
Figure 2.
Fatty acid (FA) ratios of the eel skin and muscle. Average values of four (n = 4) replicates; standard deviations are indicated by bars. For each FA ratio, different letters (a,b) denote significant differences (p < 0.05). Abbreviations as expressed in Figure 1.
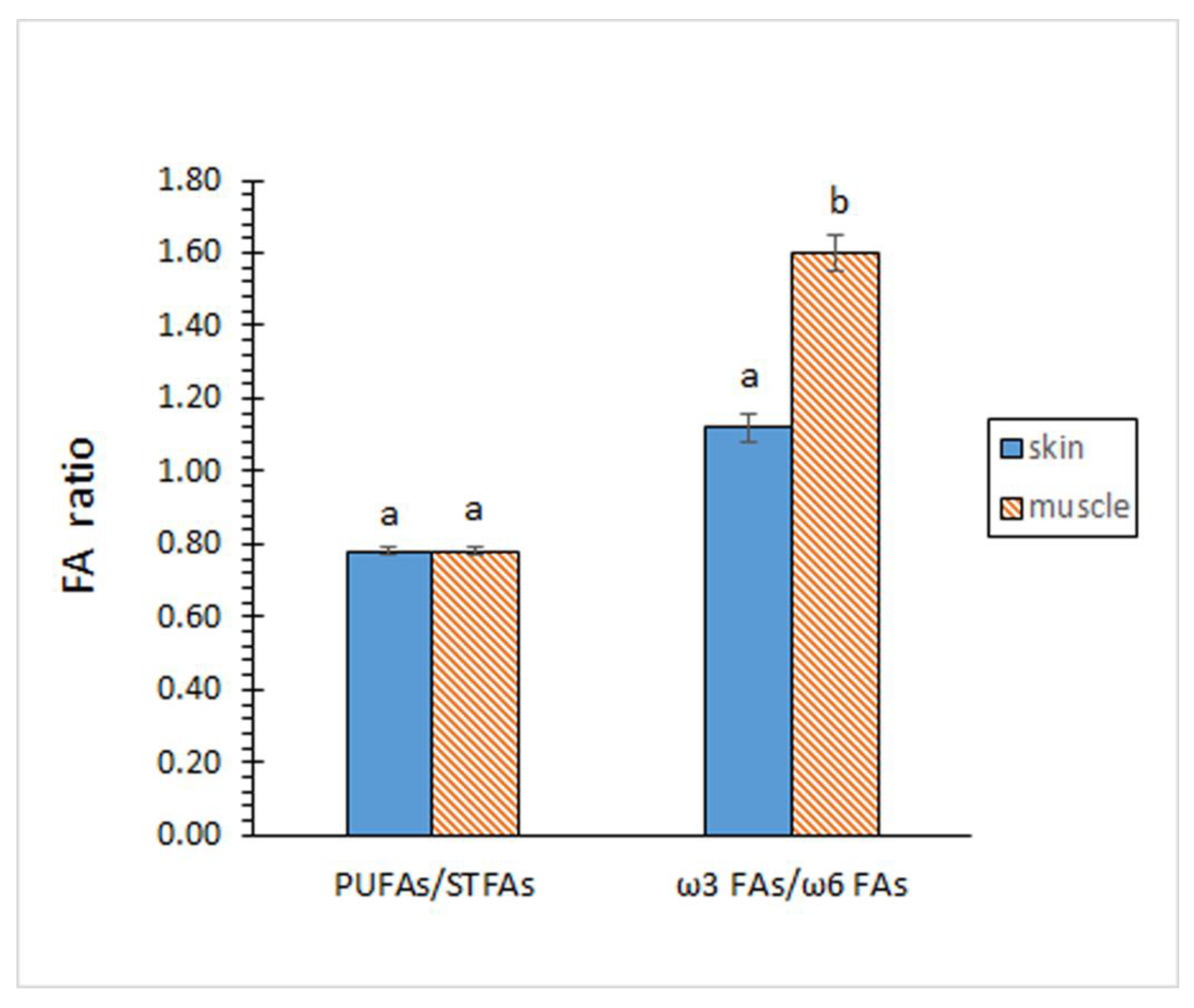

Table 1.
Proximate composition (g·kg-1 tissue) of eel skin and muscle*.
| Chemical constituent | Tissue | |
|---|---|---|
| Skin | Muscle | |
| Moisture | 677.0 ± 7.0 a | 783.3 ± 3.5 b |
| Proteins | 271.6 ± 7.2 b | 166.4 ± 1.8 a |
| Lipids | 38.0 ± 0.9 b | 28.6 ± 1.6 a |
| Ash | 27.7 ± 2.1 b | 9.9 ± 1.0 a |
* Average values ± standard deviations of four (n = 4) replicates. In each row, different letters (a,b) denote significant differences (p < 0.05).
Table 2.
Fatty acid (FA) profile (g·100 g-1 total FAs) of eel skin and muscle.
| FA | Tissue | |
|---|---|---|
| Skin | Muscle | |
| 14:0 | 2.70 ± 0.03 a | 3.09 ± 0.04 b |
| 15:0 | 0.70 ± 0.06 a | 0.65 ± 0.01 a |
| 16:0 | 22.37 ± 0.15 a | 22.39 ± 0.09 a |
| 17:0 | 1.48 ± 0.05 b | 1.05 ± 0.02 a |
| 18:0 | 5.97 ± 0.13 a | 5.91 ± 0.20 a |
| 16:1 ω7 | 6.63 ± 0.12 a | 6.92 ± 0.15 b |
| 18:1 ω7 | 6.93 ± 0.06 a | 6.84 ± 0.16 a |
| 18:1 ω9 | 25.34 ± 0.40 a | 25.09 ± 0.47 a |
| 20:1 ω9 | 1.25 ± 002 b | 1.12 ± 0.01 a |
| 22:1 ω9 | 0.16 ± 0.01 b | 0.13 ± 0.01 a |
| 24:1 ω9 | 0.45 ± 0.03 b | 0.15 ± 0.02 a |
| 18:2 ω6 | 2.35 ± 0.05 a | 2.25 ± 0.14 a |
| 20:2 ω6 | 1.25 ± 0.05 b | 1.05 ± 0.08 a |
| 20:4 ω6 | 5.45 ± 0.09 a | 4.25 ± 0.13 b |
| 22:4 ω6 | 3.23 ± 0.14 a | 2.44 ± 0.10 b |
| 20:5 ω3 | 5.83 ± 0.14 a | 6.74 ± 0.07 b |
| 22:5 ω3 | 4.82 ± 0.18 a | 5.14 ± 0.08 b |
| 22:6 ω3 | 3.06 ± 0.08 a | 4.23 ± 0.16 b |
* Average values ± standard deviations of four (n = 4) replicates. In each row, different letters (a,b) denote significant differences (p < 0.05).
Table 3.
Lipid class composition of eel skin and muscle*.
| Lipid class | Tissue | |
|---|---|---|
| Skin | Muscle | |
| Triacylglycerols | 400.6 ± 28.0 a (15.2 ± 3.0 a) |
411.6 ± 7.3 a (11.8 ± 0.6 a) |
| Free fatty acids | 43.6 ± 1.4 b (1.7 ± 0.4 b) |
30.9 ± 0.8 a (0.9 ± 0.1 a) |
| Phospholipids | 111.1 ± 5.5 b (4.2 ± 0.6 b) |
93.4 ± 5.5 a (2.7 ± 0.6 a) |
| Sterols | 104.7 ± 5.8 b (4.0 ± 0.6 b) |
24.2 ± 1.1 a (0.7 ± 0.2 a) |
| Alpha-tocopherol | 274.0 ± 14.7 b (10.4 ± 1.8 b) |
178.0 ± 38.8 a (5.1 ± 1.9 a) |
* Average values ± standard deviations of four (n = 4) replicates. Data expressed as g·kg-1 lipids, except for alpha-tocopherol (mg·kg-1 lipids). Data in brackets indicate the content in tissue basis expressed as g·kg-1 tissue, except for α-tocopherol (mg·kg-1 tissue). In each row, different letters (a,b) denote significant differences (p < 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



